
Arthropods, 2014, 3(1)
99
Arthropods Vol. 3, No. 1, 1 March 2014 International Academy of Ecology and Environmental Sciences
Transcript of Arthropods, 2014, 3(1)

Arthropods
Vol. 3, No. 1, 1 March 2014
International Academy of Ecology and Environmental Sciences

Arthropods ISSN 2224-4255 Volume 3, Number 1, 1 March 2014 Editor-in-Chief WenJun Zhang Sun Yat-sen University, China International Academy of Ecology and Environmental Sciences, Hong Kong E-mail: [email protected], [email protected] Editorial Board Andre Bianconi (Sao Paulo State University (Unesp), Brazil) Anton Brancelj (National Institute of Biology, Slovenia) Hans-Uwe Dahms (Sangmyung University, Korea) A. K. Dhawan (Punjab Agricultural University, India) John A. Fornshell (Northern Virginia Community College, USA) Xin Li (Northwest A&F University, China) Oscar E. Liburd (University of Florida, USA) Ivana Karanovic (Hanyang University, Korea) Enoch A Osekre (KN University of Science and Technology, Ghana) Rajinder Peshin (Sher-e-Kashmir University of Agricultural Sciences and Technology of Jammu, India) Michael Stout (Louisiana State University Agricultural Center, USA) Eugeny S. Sugonyaev (Russian Academy of Sciences, Russia)
Editorial Office: [email protected] Publisher: International Academy of Ecology and Environmental Sciences Address: Flat C, 23/F, Lucky Plaza, 315-321 Lockhart Road, Wanchai, Hong Kong Tel: 00852-6555 7188 Fax: 00852-3177 9906 Website: http://www.iaees.org/ E-mail: [email protected]

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Article
Epigeus macroinvertebrates species assemblages along a
successional gradient in Hailuotu Island (Bothnia Bay), Finland
Adolfo A. Del Rio Mora Instituto de Investigaciones Agropecuarias y Forestales, Universidad Michoacana de San Nicolás de Hidalgo., Unidad San
Juanito Itzícuaro, C.P. 58330, Morelia, Michoacán., México
E-mail: [email protected]
Received 22 May 2013; Accepted 26 June 2013; Published online 1 March 2014
Abstract
Epigeus macroinvertebrates were collected during summer time in 2007, by using pitfall traps in different
sites representing vegetation patches situated on land uplift area on successional gradients in the dune shore of
Bothnian on the island of Hailuoto, Northern part of the Gulf of Bothnia, Finland. The sites were divided into
six vegetation patches types or open sands, all of them localized on early, deflation zone and late successional
stages or ecological subgroups: 1) Empetrum patches or microsites (small-scale element distribution in soil-
plant-systems in patches of Empetrum nigrum, in early succession; 2) Empetrum nigrum patches in deflation
zone; 3) open sand in early succession; 4) open sand in deflation zone; 5) Empetrum nigrum patches in late
succession, and 6) open sand in late succession. A total of 19034 specimens belonging to 14 species of Insecta
and only one group to Aranea species were caught and identified. Afterwards they were grouped by trophic
groups as follows: herbivores, predators and detritivores and calculated their richness, abundance, diversity
and evenness for each vegetation type. The data obtained were analyzed by different analytical methods and
relevant between them as MRPP for the purpose of identifying the possible differences between groups and
habitats, which denoted no statistically significant between the 6 environmental types, but if for the case of
composition or populations general diversity as abundance, richness, evenness, diversity. It is enclosed too
Correspondence Analysis (CA) and cluster analysis for epigeus invertebrates species assemblages. As a
support to analysis of results we added on ended the species-accumulation curve and estimation curves Chao1
and Jacknife2 for all ecological types.
Keywords epigeus invertebrates; patches; succession; diversity; richness; Empetrum nigrum ssp.
hermaphroditum (Hagerup) Böche.
1 Introduction
Soil invertebrates are fundamentally in essential processes for the habitat in which they are developed. They
can alter primary production, the structure of the ground, the landlords of microbial activity, the dynamics of
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
the organic matter and the cycle of nutrients (Berg and Staaf, 1981; Petersen and Lurton, 1982; Slansky and
Rodriguez, 1987; McClaugherty et al., 1982).
Spatio-temporal variability of the community in turn has important implications for the study system,
which is a coastal forest strip located on the Hailotu Island-Bothnia bay -with a strong successional gradient,
which changes in species composition and trophic structure of soil invertebrate communities should affect
species interactions and food web dynamics (Doblas, 2007; Doblas et al., 2008, 2009a, b, c; Niemelä, 1997;
1999; Similä et al., 2002).
Longer-term studies regarding macroinvertebrates associated with vegetation patches, with emphasis on
Carabids species- have argued that the overall change in the species distribution is caused mostly by climatic
factors and that effects of human influences seem to be secondary (Hengeveld, 1985).
Microhabitats are common on island ecosystems where the flora and fauna which is grouped in patches,
which are potentially maintained by complex interrelationships involving their own soil invertebrates that live
there, depending on that vegetation grouped for protection and life cycles and in turn serve to disperse the
owner plant community in the ecosystem(Borges et al., 2004; Koricheva et al., 2000; Kouki et al., 2001).
Different from aboveground invertebrates, soil invertebrate species, their interrelationships and spatial
distribution has been little studied until today (Zhang 2008, 2011a, b). Between the Arthropod group in coastal
habits, the spiders are known to respond sensitively to environmental and structural changes, which makes
them suitable to study organism–habitat relationships (Wise, 1993; Bell et al., 2001; Oxbrough et al., 2005;
Hendrickx et al., 2007), in coastal habitats in particular.
Spiders constitute one of the most abundant and species–rich arthropod orders. They range among the most
numerous arthropods in all kinds of habitat types (Basset, 1991; Coddington et al., 1991; Borges and Brown,
2004). Spider species occupy a wide array of spatial and temporal niches. Their occurrence is frequently
related to environmental factors (Hatley and MacMahon, 1980; Schmidt et al., 2005; Entling et al., 2007;
Finch et al., 2008)
Poor establishment and reduced seedling growth of Scots pine (Pinus silvestris L.) in northern Sweden is
related to an allelopathic inhibition by the dwarf shrub Empetrum hermaphroditum Hagerup. Indoor bioassays
with green and brown leaves of Empetrum have strong negative effects on rooting ability, radicle elongation,
and growth of Scots pine seedlings. Bioassays with soil samples show that phytotoxic substances leached from
Empetrum foliage accumulate in the soil (Rautio and Markkola, 2006).
Known to as alone that Mycelial fungal biomass in the soil in the vicinity of the seedling roots is higher in
Empetrum than in empty patches and increased along the succession. In Scots pine roots both the diversity of
ectomycorrhizal morphotypes and proportion of root tips colonized by suilloid morphotypes with abundant
external mycelia were in mid succession higher and in late succession lower in seedlings grown in Empetrum
patches compared to patches without Empetrum. In the harsh conditions of the dune shore in the early and mid
succession Empetrum is suggested to promote Scots pine seedling establishment by providing mechanical and
physical shelter whereas in late succession negative interactions (competition and allelopathy) between the
shrub and the Scots pine are dominating. This fact was the first findings/results that an ericoid mycorrhizal
shrub could enhance the performance of both the ECM host and its fungal symbionts (Rautio and Markkola,
2006).
On the same, relative importance of positive and negative interactions between plant species and their
epigeus arthropofauna associated with them may change along disturbance and resource gradients, so positive
interactions are suggested to prevail in low resource, low productivity (high stress) conditions and negative
interactions in high resource availability. Mountain crowberry (Empetrum nigrum ssp. hermaphroditum) is
known to have allelopathic impacts on both Scots pine (Pinus sylvestris) and its ectomycorrhizal symbionts.
2

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
On primary succession gradients in the dune shore of Bothnian Bay, however, Scots pine seedlings are
founded to occur more abundantly in Empetrum patches in early and mid succession stages, whereas patches
without Empetrum are preferred in late succession, however, as it is seem to, this is not reflected on the
number of seedlings or natural regeneration at all these places, which could be due to high pine seed
depredation by carabids and what before has been widely researched (Lindroth, 1985, 1986; Nystrand and
Granström, 2002), although for the study area little details are knew about epigeus invertebrate assemblages
and their abundance at the different environmental types along the successional gradient at Hailotu Island.
The main focus of this study was determine the actual structure and diversity of the soil macroinvertebrates
community under conditions of a successional gradient in Hailotu Island (Bothnia Bay) where is common the
growing of native vegetation and pioneers distributed in patches or microsites, since is not clear even the real
impact of the macroarthropod assemblages on the native vegetation on study area.
2 Methodology
2.1 Study area and caught insects
The study was carried out in northern Finland on the island of Hailuoto 65º03´N, 24º36´E) on two separate
sites around two03´N, 24 (65 kilometres from each others. Both sites are situating on land uplift area on a
deflation basin behind the dune zone on poor, sandy soil without podsol formation. Vegetation is patchy,
consisting of dwarf shrubs (Empetrum nigrum, Vaccinium uliginosum) lichens (Cladonia spp., Cladina spp.,
Stereocaulon sp.) and mosses (Racomitrium canescens, Polytrichum piliferum, P. juniperinum). Sparsely
distributed young Scots pines are the dominant tree species. Humus layer is patchy and very thin (0.5–1.0 cm),
if present. The mineral soil is acidic (pH 4.8), and organic matter and total nitrogen content are low (0.2% and
< 0.01% of soil dry mass, respectively).
2.2 Trials
Epigeus macroinvertebrates were collected during summer time 2007 year, by using pitfall traps(total 120), for
it the trial sites were divided into six vegetation patches types or open sands, all them localized on early,
deflation zone and late successional stages or ecological subgropups: 1) Empetrum patches in early succession
(Eesu); 2) Empetrum patches in deflation zone (Edefbas); 3) open sand in early succession (OpSesu); 4) open
sand in deflation zone (OpSdefb); 5) Empetrum patches in late succession (Elatsu), and 6) open sand in late
succession (OpSlatsu). All pitfalls trials of medial size (9 cm diameter of 11 cm in height) and made of plastic
were baited with a solution of water mixed with NaCl. Later they were dried, mounted, identified and grouped
by trophic groups as follows: herbivores, predators and detritivores and calculated their richness, abundance,
diversity and evenness for each vegetation type.
All the specimens collected during the study are in the collection of the Zoological Oulu University
Museum, Oulu, Finland.
2.3 Analysis of the results
We assume the pitfalls were equally efficient in catching soil invertebrates in the different 6 succesional
gradient in the forest type described before and growing in coast zone, because all the sample points of them
were homogenous in general structure. Therefore it is suggested that the differences in the catch among sites
should reflect real differences in the abundances of epigeus individuals, specie groups and habitats. For the
composition analysis species were selected according their abundance as minimum 2 specimens,
To evaluate the consistency of the differences in species composition of the soil arthropofauna under
successional gradient, we used the nonparametric statistical test multiple response permutation (MRPP) with
Blossom w2008.04.02; and the structure of epigeus invertebrate assemblages by detrended correspondence
analysis (DCA) with Multivariate Statistical Package 3.1 (Kovach, 1999).
3

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
The values of abundance, richness, diversity and evenness were analyzed by two ways ANOVA with
Minitab15.
Insect caught by pitfall traps were pooled for each site and species richness was estimated for each
successional type (6), as well as for the regional data set using the nonparametric estimators Chao 1 and
Jacknife 2. Accumulation curves were generated after 100 randomizations using Estimate S 8.0 (Colwell,
2006).
Chao1 gives an estimate of the absolute number of species in an assemblage based on the number of rare
species (singletons and doubletons) in a sample, and it is the ratio between observed and estimated
richness(Sorensen et al., 2002; Scharff et al., 2003).
Jacknife2 has been found to perform quite well in extrapolation of species richness with greater precision,
less bias, and less dependence on sample size than other estimators (Petersen et al., 2003; Chiarucci et al.,
2003; Finch et al., 2008.).
The average abundance was measured in number of individuals by trap and the richness as species number
of traps per group.
To calculate the Diversity was used the Shannon Index: H = - ∑ pi log pi; this index considers
simultaneously Richness and Evenness, and their values vary between zero, when a single species, and the
logarithm of Richness (S), where all species are represented with the same number of individuals (Magurran,
1988). The Evenness was calculated using Pielou index (Magurran, 1988): J´= H´/Hmax=H´/lnS which
measures the proportion of the diversity observed in relation to the expected maximum diversity and its values
vary between 0 and 1, the last value corresponds to situations where all species are equally abundant (Moreno,
2001). 3 Results and Discussion 3.1 Species caught and their distribution by successional stages
A total of 19034 specimens belong to 14 species of Insecta and only one group to Araneae species. As shown
in Fig. 1, the largest number of individuals (soil invertebrates) collected during the study corresponded to the
patches OpSlatsu, followed by Elatsu, while in the other vegetable strata had similar amounts of catch
specimens (Eesu, Edefbas, OpSesu and OpSdefb patches). This greater number of specimens collected in the
first two layers correspond to patches mentioned are the preferential microhabitats Lassius niger ant
(Hymenoptera: Formicidae) and also can be seen in Fig. 2. Fig. 1 Total number of epigeus invertebrates specimens caught (Mean with SEM). Hailuoto Island (Bothnia Bay, Summer 2007). Empetrum patches in early succession (Eesu), Empetrum patches in deflation zone (Edefbas), open sand in early succession (OpSesu), open sand in deflation zone (OpSdefb), Empetrum patches in late succession (Elatsu) and open sand in late succession (OpSlatsu).
4

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
The arthropods collected and according to their habits and abundance were classified in: herbivores (8),
predators (5) and detritivores (2), taking for it the antecedents registered for each species according to
literature review (Slansky and Rodriguez, 1987). We listed below the habit groups, scientific names and their
corresponding abbreviations (left) are referenced along the manuscript:
Predators
Carabidae
(cer ) Calathus erratus
(cme) Calathus melanocephalus
(aqu) Amara quenseli
( cma) Cymindis macularis (hru) Harpalus rufipes Detritivores (lhi) Lagria hirta (Lagriidae) (bopul)Bolitochara pulchra (Staphylinidae)
Herbivores
(ln) Lasius niger (Formicidae) (mma) Myrmeleotettix maculatus (Acrididae)
(aspp) Aphrodes spp (Cicadellidae) (ptr) Planaphrodes trifasciata(Cicadellidae)
(ar)Araneae (soil spider) (ono) Otiorhynchus nodosus(Curculionidae) (oov) Otiorhynchus ovatus(Curculionidae) (hab) Hylobius abietis (Curculionidae)
The results (Fig. 2) reveal the logical dominance of herbivore species on other types of feeding consumers
representatives of ecosystem unstable dunes and near seashore, where the accumulation of organic matter
could be limited, which explain the few presence of invertebrate detritivores, although many of the soil spiders
(Araneae), for example, could be grouped as herbivores-detritivores and more even as predators; Some species
of them have been reported in coastal grassy vegetation and detritus, in sandy sites with low vegetation and in
clay shores and stones (Heimer and Nentwig, 1991).
Moreover, seem to be the population model is simple and continue on seaboard coast: cycle beginning
richness of herbivores and predators from Empetrum early succession patches and keeping stable on next stage
(Edefbas) and lowing for the case of the predators; on Empetrum late succession there a apparent equilibrium
between richness of herbivores and predator but significantly low of the two first successional stages. When
open sand early succession newly is raised the herbivores and predator tough no as in Eesu and Edefb patches
and ending decrease both fully on open late successional stage, where the ant colonies are prevalent. Occurs,
this system is not close and part of these populations come of the natural emigration process, mostly from the
develop forests near to the coastal zone (see fauna composition and species richness).
5

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Fig. 2 15 Invertebrate species caught in 6 ecological vegetation types (patches) (number of specimens was transformed in Log10).
The soil macroinvertebrate assemblage at the study sites is dominated by arthropods and the dominant
group in number under all the successional gradient is Formicidae (Lasius niger), mostly on patches of Elatsu
and Oslatsu, where evidently its colonies have at the same time more space and less competence for carrying
off and store great amounts of food.
After of ants, Leafhopper Aphrodes spp. (Cicadellidae) and soil spider (Araneae) were the
macroinvertebrates more abundant on all microhabitats monitored. This possibly due the decomposition
process is driven by litter quality, physicochemical environment and composition of the decomposer
community and this last process as a result of carbon and nitrogen utilization by heterotrophic microbial and
animal populations (Hättenschwiler et al., 2005; Berg and Staff, 1981; Berg et al., 1984; McClaugherty et al.,
1982).
Carabids more abundant represented by Calathus erratus are actives as predators, moving on Empetrum
patches and open sand early succession habitats. The others species of this family are more restricted for a
habitat in particular so Calathus melanocephalus have preference only by Empetrum deflation basin and
Cymindis macularis moves inside Empetrum early succession patches; Amara quenseli and Harpalus rufipes
are not common for the study area.
Other carabid species no included here and collected on Empetrum patches by us on late summer (mid-
September 2006) in the study area were: Bembidion nigricorne, Pterostichus niger and Carabus hortensis.
Snout beetles (Curculionidae) Otiorhynchus ovatus and Hylobius abietis (This species is regarded as the most
important pest of conifer seedlings) are occasional on the primary vegetation in the coastal area of the bay,
although periods of high populations of this last specie could be regarding with Pinus seedlings died growing
on Empetrum patches, as we have observed in other years. On the same, relative importance of positive and
negative interactions between plant species and their epigeus arthropods associated with them may change
along disturbance and resource gradients, so positive interactions are suggested to prevail in low resource, low
productivity (high stress) conditions and negative interactions in high resource availability. Mountain
crowberry (Empetrum nigrum ssp. hermaphroditum) is known to have allelopathic impacts on both Scots pine
(Pinus sylvestris) and its ectomycorrhizal symbionts. On primary succession gradients in the dune shore of
Bothnian Bay, however, Scots pine seedlings are founded to occur more abundantly in Empetrum patches in
6
Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
early and mid succession stages, whereas patches without Empetrum are preferred in late succession, however,
as it is seem to, this is not reflected on the number of seedlings or natural regeneration at all these places,
which could be due too to high pine seed depredation by carabids and what before has been widely
documented (Ovenistrand and Granstroem, 2000), but none of carabid species reported and cited above are not
Pine seed consumers (Lindroth, 1985, 1986; Nystrand and Granström, 2000).
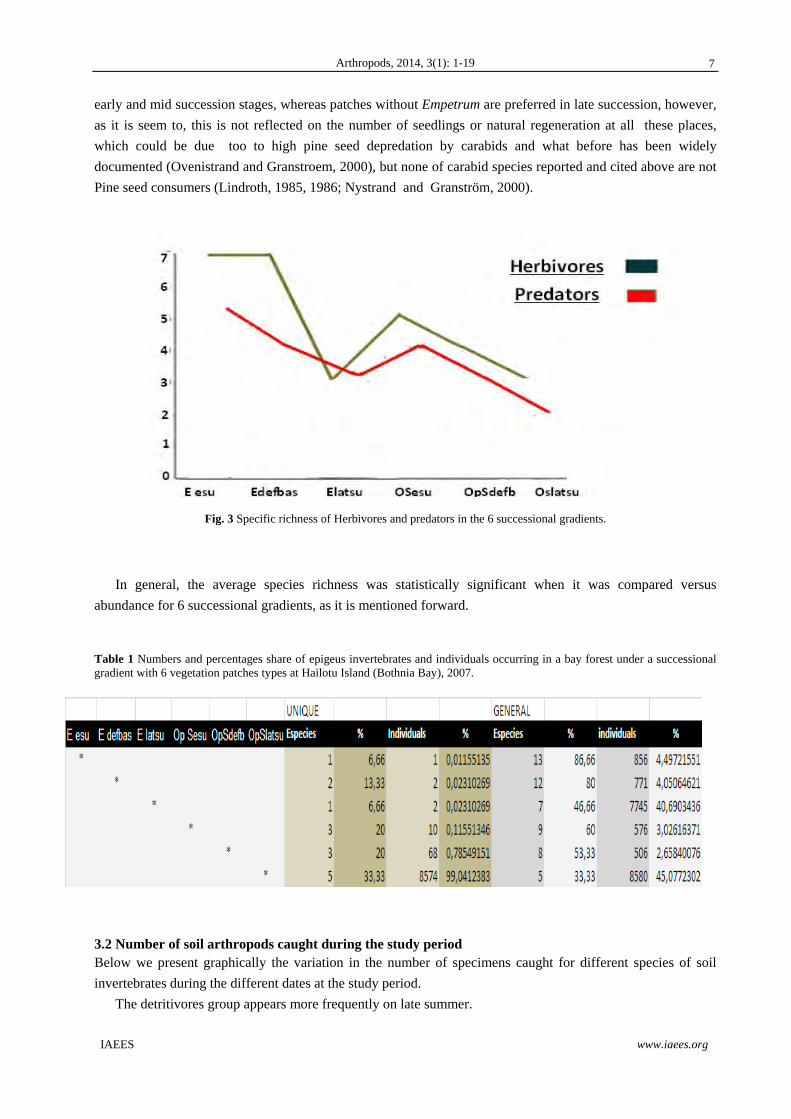
Fig. 3 Specific richness of Herbivores and predators in the 6 successional gradients.
In general, the average species richness was statistically significant when it was compared versus
abundance for 6 successional gradients, as it is mentioned forward.
Table 1 Numbers and percentages share of epigeus invertebrates and individuals occurring in a bay forest under a successional gradient with 6 vegetation patches types at Hailotu Island (Bothnia Bay), 2007.
3.2 Number of soil arthropods caught during the study period Below we present graphically the variation in the number of specimens caught for different species of soil
invertebrates during the different dates at the study period.
The detritivores group appears more frequently on late summer.
7

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Fig. 4 Number of invertebrates caught during first sampling period (June 28-July 4/2007).
Fig. 5 Number of invertebrates caught during second sampling period (July 4-August 7/2007).
8
Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
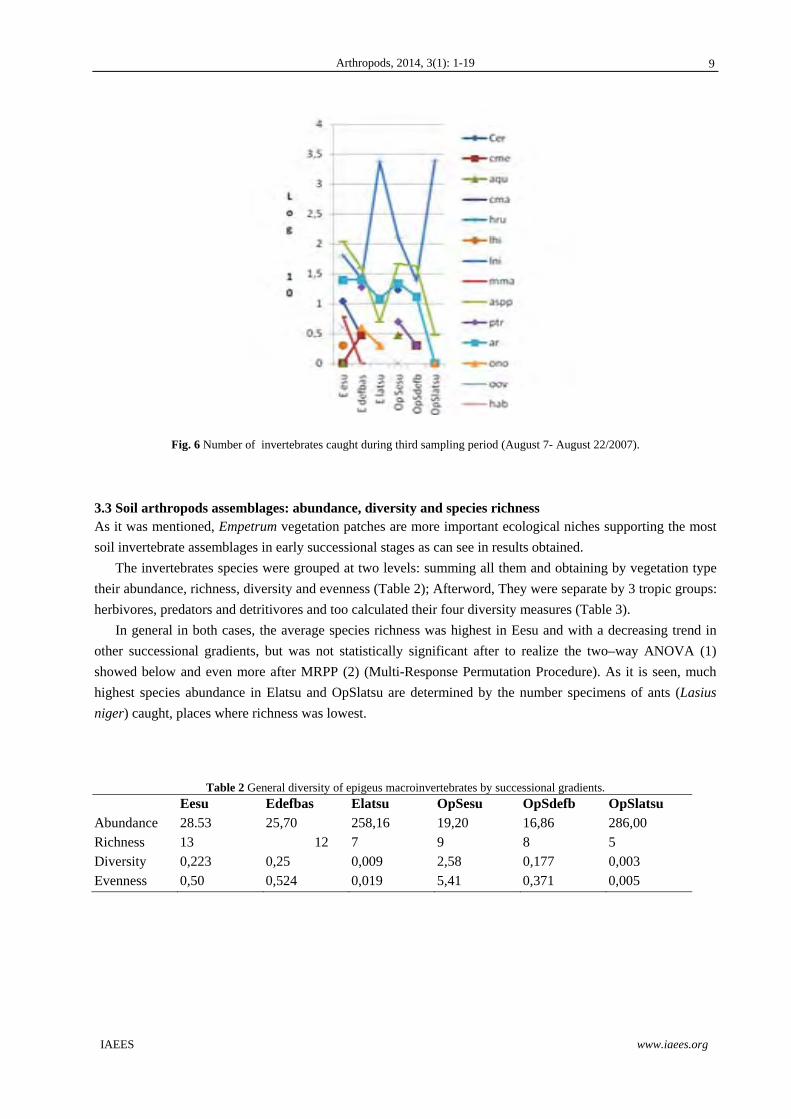
Fig. 6 Number of invertebrates caught during third sampling period (August 7- August 22/2007).
3.3 Soil arthropods assemblages: abundance, diversity and species richness As it was mentioned, Empetrum vegetation patches are more important ecological niches supporting the most
soil invertebrate assemblages in early successional stages as can see in results obtained.
The invertebrates species were grouped at two levels: summing all them and obtaining by vegetation type
their abundance, richness, diversity and evenness (Table 2); Afterword, They were separate by 3 tropic groups:
herbivores, predators and detritivores and too calculated their four diversity measures (Table 3).
In general in both cases, the average species richness was highest in Eesu and with a decreasing trend in
other successional gradients, but was not statistically significant after to realize the two–way ANOVA (1)
showed below and even more after MRPP (2) (Multi-Response Permutation Procedure). As it is seen, much
highest species abundance in Elatsu and OpSlatsu are determined by the number specimens of ants (Lasius
niger) caught, places where richness was lowest.
Table 2 General diversity of epigeus macroinvertebrates by successional gradients.
Eesu Edefbas Elatsu OpSesu OpSdefb OpSlatsu Abundance 28.53 25,70 258,16 19,20 16,86 286,00 Richness 13 12 7 9 8 5 Diversity 0,223 0,25 0,009 2,58 0,177 0,003 Evenness 0,50 0,524 0,019 5,41 0,371 0,005
9

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Table 3 Specific diversity of epigeus macroinvertebrates by trophic groups in 6 ecological succesional gradients.
Herbivorous Eesu Edefbas Elatsu OpSesu OpSdefb OpSlatsuAbundance 22,9 19,13 257,43 16,4 14,7 285,8 Richness 7 7 3 5 4 3 Diversity 0,314 0,343 0,006 0,315 0,295 0,11 Evenness 0,371 0,405 0,014 0,451 0,49 0,23 Predators Eesu Edefbas Elatsu OpSesu OpSdefb OpSlatsuAbundance 5,53 0,53 0,66 2,8 2,13 0,2 Richness 5 4 3 4 3 2 Diversity 0,33 0,303 0,171 0,351 0,21 0,195 Evenness 0,472 0,503 0,358 0,583 0,44 0,647 Detritivoros Eesu Edefbas Elatsu OpSesu OpSdefb OpSlatsuAbundance 0,1 0,03 0,06 0 0,03 0 Richness 1 1 1 0 1 0
In general, the more high values of species richness, diversity and evenness are observed in Eesu, Edefbas
and OpSesu, and lower of them are for Elatsu and OpSlatsu.
The herbivores abundance in OpSlatsu is regarding the ant colonies, Lassius niger. For that, the
community of epigeus invertebrates living on Empetrum patches plays an important factor for the stability of
those forest ecosystems. In the case of detritivores, the values corresponding to diversity and evenness of the
specific diversity were zero for 6 habitats, due to lower number of species belongs to these consumers group.
ANOVA test results coincide with the MRPP analysis, although this last analysis give seen significance
between the general diversity between factors (abundance, richness, diversity and evenness obtained between
the invertebrate species caught at the 6 environmental types as shown in the Tables 4 and 5.
10

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Similarly when all factors were compared under MRPP test, have the successional gradient (vegetation
types), presence of herbivores, predators and detritivores under general and specific diversity as response
variables are not significant. Otherwise, general diversity between factors represented by abundance, richness,
diversity and evenness have differences and this is particularly clear for the case of abundance versus richness.
Table 5 MRPP analysis: comparing grouping variable(s) between response variables.
(-)NS, not significance; *, P<0.05, **, P<0.01
Fig. 7 Species-accumulation curve and estimation curves Chao 1 and Jackknife 2 in ecological subgroup: Eesu (all samples pooled) dataset. Curves are generated from 100 randomizations.
Grouping variable(s)
Response variables
Number of observations
Number of groups
Significance(-)
Successional Gradient
General diversity
12 6 NS
Herbivores Specific diversity
12 6 NS
Predators ----------- 12 6 NS
Detritivores ------------ 12 6 NS
Abundance Richness Diversity Evenness
General diversity between factors
12 4 **
Abundance vs Richness ----------- 6 2 *
Abundance vs Diversity ----------- 6 2 NS Abundance vs Evenness ----------- 6 2 NS
Richness vs Diversity ----------- 6 2 NS Richness vs Evenness ------------ 6 2 NS
Diversity vs Evenness ------------- 6 2 NS
11
Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
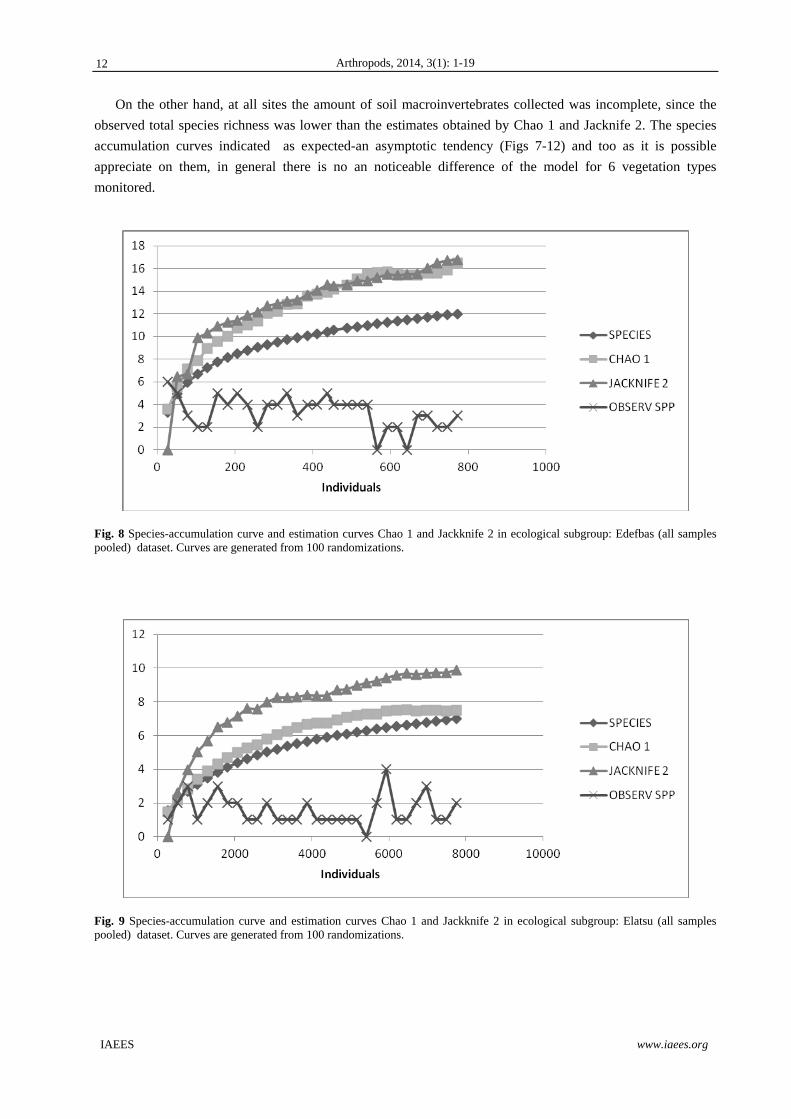
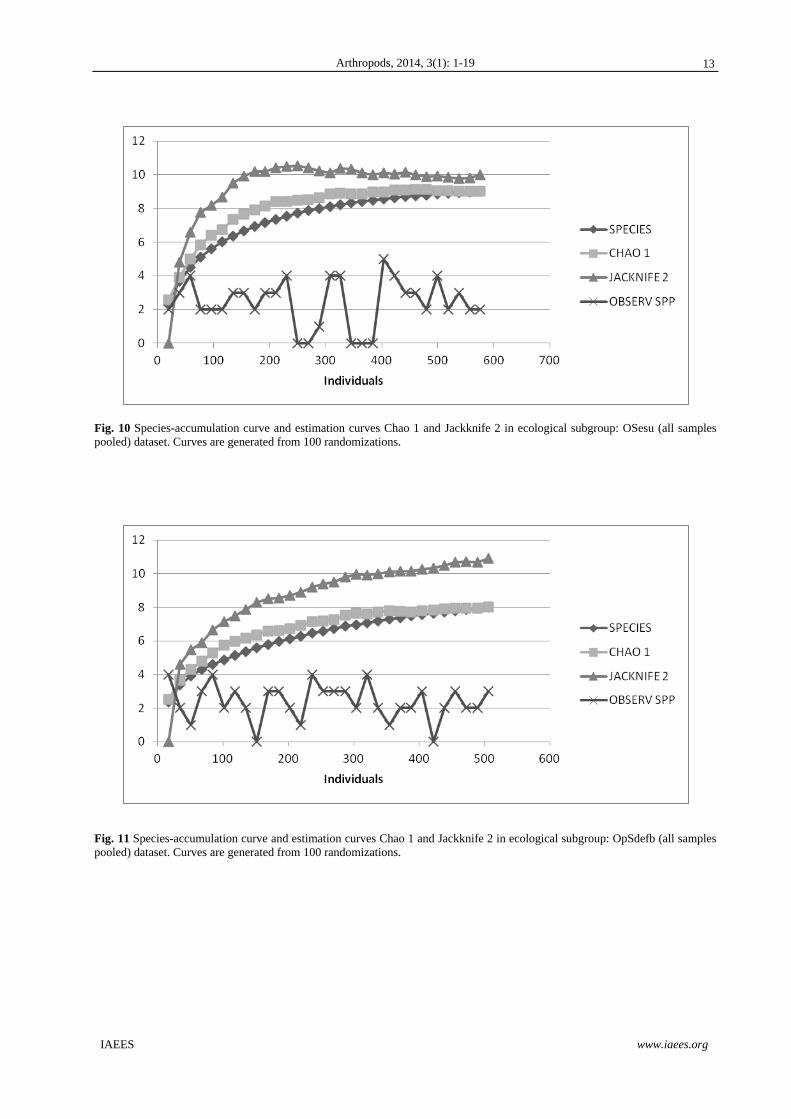
On the other hand, at all sites the amount of soil macroinvertebrates collected was incomplete, since the
observed total species richness was lower than the estimates obtained by Chao 1 and Jacknife 2. The species
accumulation curves indicated as expected-an asymptotic tendency (Figs 7-12) and too as it is possible
appreciate on them, in general there is no an noticeable difference of the model for 6 vegetation types
monitored.
Fig. 8 Species-accumulation curve and estimation curves Chao 1 and Jackknife 2 in ecological subgroup: Edefbas (all samples pooled) dataset. Curves are generated from 100 randomizations.
Fig. 9 Species-accumulation curve and estimation curves Chao 1 and Jackknife 2 in ecological subgroup: Elatsu (all samples pooled) dataset. Curves are generated from 100 randomizations.
12
Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Fig. 10 Species-accumulation curve and estimation curves Chao 1 and Jackknife 2 in ecological subgroup: OSesu (all samples pooled) dataset. Curves are generated from 100 randomizations.
Fig. 11 Species-accumulation curve and estimation curves Chao 1 and Jackknife 2 in ecological subgroup: OpSdefb (all samples pooled) dataset. Curves are generated from 100 randomizations.
13

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Fig. 12 Species-accumulation curve and estimation curves Chao 1 and Jackknife 2 in ecological subgroup: Oslatsu (all samples pooled) dataset. Curves are generated from 100 randomizations.
3.4 Correspondence analysis (CA) For the canonical analysis (CA) options were taken the next steps: it was given a common species weighting
and data transformation chosen with square root and calculated the similarity/correlations matrices using cyclic
Jacobi.
The results of similarity matrix and eigenvalues are detailed as follows:
14

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
The complete matrix was displayed in a scatter plot (joint plot), creating CA graph with the main purpose
to visualize more clearly the affinities between different soil invertebrates species and their relation with the
vegetation patches or habitats.
Fig. 13 Correspondence Analysis (CA) for 11 epigeus invertebrates at 6 microhabitats in Hailotu Island (Summer 2007).
In correspondence analysis(CA) and obtained in this case only for 11 macroinvertebrates species (analysis
were removed from those species collected in numbers 1), can be differentiated 3 invertebrate groups relevant
to their respective microhabitats and are preferably for ant ln more open space and less competition between
species, and greater availability of pasture and moving mostly on microhabitats as OpSlatsu, Elatsu; the next
group is composed by soil spiders ar and leafhoppers ptr occupying microhabitats of intermediate or transition:
Edefbas and OpSdebas and finally, as the leafhopper species aspp preferably in OpSesu and carabids cer and
aqu together here along with the more abundant grasshopper mma caught in the microhabitat-Eesu.
Other species such as carabid cme and weevil ono showed no preference for a specific microhabitat (axis 1,
15
Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
North, Fig. 13), and a detritivores , the long-jointed beetle lhi (axis 1, South, Fig. 13) although this may be
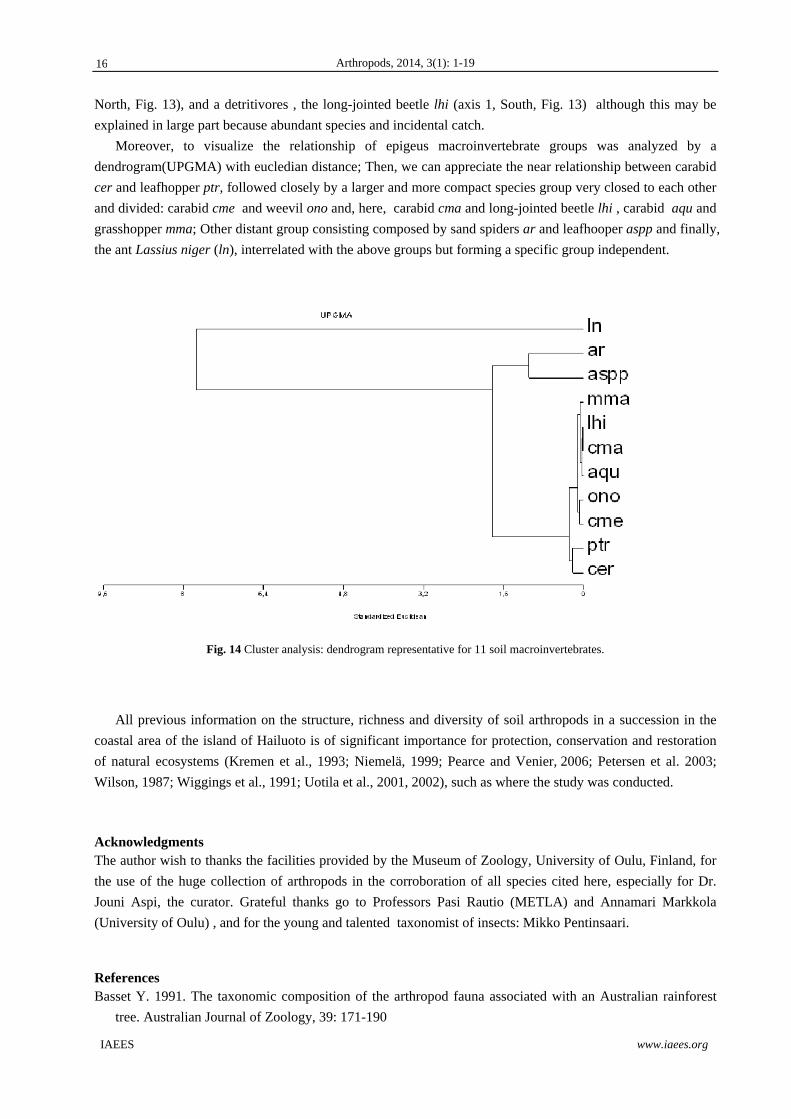
explained in large part because abundant species and incidental catch. Moreover, to visualize the relationship of epigeus macroinvertebrate groups was analyzed by a
dendrogram(UPGMA) with eucledian distance; Then, we can appreciate the near relationship between carabid
cer and leafhopper ptr, followed closely by a larger and more compact species group very closed to each other
and divided: carabid cme and weevil ono and, here, carabid cma and long-jointed beetle lhi , carabid aqu and
grasshopper mma; Other distant group consisting composed by sand spiders ar and leafhooper aspp and finally,
the ant Lassius niger (ln), interrelated with the above groups but forming a specific group independent.
Fig. 14 Cluster analysis: dendrogram representative for 11 soil macroinvertebrates.
All previous information on the structure, richness and diversity of soil arthropods in a succession in the
coastal area of the island of Hailuoto is of significant importance for protection, conservation and restoration
of natural ecosystems (Kremen et al., 1993; Niemelä, 1999; Pearce and Venier, 2006; Petersen et al. 2003;
Wilson, 1987; Wiggings et al., 1991; Uotila et al., 2001, 2002), such as where the study was conducted.
Acknowledgments The author wish to thanks the facilities provided by the Museum of Zoology, University of Oulu, Finland, for
the use of the huge collection of arthropods in the corroboration of all species cited here, especially for Dr.
Jouni Aspi, the curator. Grateful thanks go to Professors Pasi Rautio (METLA) and Annamari Markkola
(University of Oulu) , and for the young and talented taxonomist of insects: Mikko Pentinsaari.
References Basset Y. 1991. The taxonomic composition of the arthropod fauna associated with an Australian rainforest
tree. Australian Journal of Zoology, 39: 171-190
16

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Bell JR, Wheater CP, Cullen WR. 2001. The implications of grassland and heathland management for the
conservation of spider communities: a review. Journal of Zoology, 255: 377-387
Berg C, Christensen J, Ressel P. 1984. Harmonic Analysis on Semigroups: Theory of Positive Definite and
Related Functions. Springer, Netherlands
Berg B, Staaf H. 1981. Leaching, accumulation and release of nitrogen in decomposing forest floor litter. In:
Terrestrial Nitrogen Cycles (Clark FE, Rosswall T, eds). Ecological Bulletin, 33: 163-178
Blossom w2008.04.02. Blossom Statistical Software
Borges PAV, Brown VK. 2004. Arthropod community structure in pastures of an island archipelago (Azores):
looking for local–regional species richness patterns at fine scales. Bulletin of Entomological Research, 94:
111-121
Brooks AJ, et al. 2005. Hydraulic microhabitats and the distribution of macroinvertebrates assemblages in
riffles. Fresh Water Biology, 50: 331-344
Chiarucci A, Enright NJ, Perry GLW, et al. 2003. Performance of nonparametric species richness estimators in
a high diversity plant community. Diversity and Distributions, 9: 283-295
Chiarucci A, Enright NJ, Perry GLW, et al. 2003. Performance of nonparametric species richness estimators in
a high diversity plant community. Diversity and Distributions, 9: 283-295
Coddington JA, Grisworld CE, Dávila DS, et al. 1991. Designing and testing sampling protocols to estimate
biodiversity in tropical ecosystems. In: Proceedings of the Fourth International Congress of Systematic and
Evolutionary Biology. Dioscorides Press, Portland, USA
Colwell RK. 2000. EstimateS-Statistical Estimation of Species Richness and Shared Species from Samples.
University of Connecticut, Connecticut, USA
Colwell RK. 2006. EstimateS: statistical estimation of species richness and shared species from samples.
Version 8.0. User's Guide and Application.
Doblas E. 2007. Ecology of soil macroinvertebrates in a Mediterranean arid ecosystem. PhD thesis. Univerity
of Granada, Spain
Doblas-Miranda E, Wardle DA, Peltzer DA, et al. 2008. Changes in the community structure and diversity of
soil invertebrates across the Franz Josef Glacier chronosequence. Soil Biology and Biochemistry, 40:
1069-1081
Doblas-Miranda E, Sánchez-Piñero F, González-Megías A. 2009a. Different structuring factors but connected
dynamics shape litter and belowground soil macrofaunal food webs. Soil Biology and Biochemistry, 41:
2543-2550
Doblas-Miranda E, Sánchez-Piñero F, González-Megías A. 2009b. Different microhabitats affect soil
macroinvertebrate assemblages in a Mediterranean arid ecosystem. Applied Soil Ecology, 41: 329-335
Doblas-Miranda E, Sánchez-Piñero F, González-Megías A. 2009c. Vertical distribution of soil macrofauna in
an arid ecosystem: are litter and belowground compartmentalized habitats? Pedobiologia, 52: 361-373
Entling W, Schmidt MH, Bacher S, et al. 2007. Niche properties of Central European spiders: shading,
moisture and the evolution of the habitat niche. Global Ecology and Biogeography, 16: 440-448
Finch OD, Krummen H, Plaisier F, et al. 2007. Zonation of spiders (Araneae) and carabid beetles (Coleoptera:
Carabidae) in island salt marshes at the North Sea coast. Wetlands Ecology and Management, 15: 207-228
Finch OD, Blick T, Schuldt A. 2008. patterns of spider richness across Europe. Biodiversity and Conservation,
17: 2849-2868
Hatley CL, MacMahon JA. 1980. Spider community organization: seasonal variation and the role of vegetation
architecture. Environmental Entomology, 9: 632-639
17

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Hättenschwiler S, Tiunov A,Scheu S. 2005. Biodiversity and litter decomposition in terrestrial ecosystems ;
Annual Review of Ecology, Evolution and Systematics, 36: 191-218
Heimer S, Nentwig W. 1991. Spinnen mitteleuropas. Paul Parey, Berlin, Hamburg, Germany
Hendrickx F, Maeilfait JP, Wingerden WV, et al. 2007. How landscape structure, land–use intensity and
habitat diversity affect components of total arthropod diversity in agricultural landscapes. Journal of
Applied Ecology, 44: 340-351
Hengeveld R. 1985. Dynamics of Dutch ground beetle species during the twentieth century (Col;
Carabidae).Journal of Biogeography, 12: 389-411
Kouki A, Hyvärinen E, Lappalainen H, et al. 2012. Landscape context affects the success of habitat restoration:
large-scale colonization patterns of saproxylic and fire-associated species in boreal forests. Diversity and
Distributions, 18(4): 348-355
Koricheva J, Mulder CPH, Schmid B, et al. 2000. Numerical responses of different trophic groups of
invertebrates to manipulations of plant diversity in grasslands. Oecologia, 125: 271-282
Kouki J, Löfman S, Martikainen P, et al. 2001. Forest fragmentation in Fennoscandia: linking habitat
requirements of wood-associated threatened species to landscape and habitat changes. Scandinavian
Journal of Forest Research, Suppl. 3: 27-37
Kovach WL. 1999. MVSP - A Multivariate Statistical Package for Windows (Ver.3.1). Kovach Computing
Services, Pentraeth, Wales, UK
Kremen C, Colwell RK, Erwin TL, et al. 1993. Terrestrial arthropod assemblages: their use in conservation
planning. Conservation Biology, 7(4): 796-808
Lindroth CH. 1985. The Carabid (Coleoptera) of fennoscandia and Denmark; Fauna entomologica
scandinavica (Vol.15, part 1). E. J. Brill/Scandinavian Science Press, Denmark
Lindroth CH. 1986. The Carabid(Coleoptera) of fennoscandia and Denmark; Fauna entomologica scandinavica,
vol.15, part 2. E. J. Brill/Scandinavian Science Press, Denmark
Magurran AE. 1988. Ecological Diversity and Its Measurement. University Press, Cambridge, UK
McClaugherty CA, Aber JD, Melillo JM. 1982. The role of fine roots in the organic matter and nitrogen
budgets of two forested ecosystems. Ecology, 63: 1481-1490
Marshall SA, Anderson RS, Roughley RE, et al. 1994. Terrestrial arthropod biodiversity: planning a study and
recommended sampling techniques. Bulletin of the Entomological Society of Canada 26(1): 33
Martikainen P, Siitonen J, Punttila P, et al. 2000. Species richness of Coleoptera in mature managed and old-
growth boreal forests in southern Finland. Biological Conservation, 94: 199-209
Minitab@15 software. www.minitab.com
Moreno CE. 2001. Métodos para Medir la Biodiversidad. Volumen I. Manuales y Tesis. Sociedad
Entomológica Aragonesa. Zaragoza, España
Niemelä J. 1997. Invertebrates and boreal forest management. Conservation Biology, 11: 601-610
Niemelä J. 1999. Management in relation to disturbance in the boreal forest. Forest Ecology and Management,
115: 127-134
Nystrand O, Granström A. 2000. Predation on Pinus sylvestris seeds and juvenile seedlings in Swedish boreal
forest in relation to stand disturbance by logging. Journal of Applied Ecology, 37(3): 449-463
Oxbrough AG, Gittings T, O‘Halloran J, et al. 2005. Structural indicators of spider communities across the
forest plantation cycle. Forest Ecology and Management, 212: 171-183
Pearce JL, Venier LA. 2006. The use of ground beetles (Coleoptera: Carabidae) and spiders (Araneae) as
bioindicators of sustainable forest management: A review. Ecological Indicators, 6(4): 780-793
18

Arthropods, 2014, 3(1): 1-19
IAEES www.iaees.org
Petersen WJ, Lurton H. 1982. A comparative analysis of soil fauna populations and their role in decomposition
processes. Oikos, 39: 288-388
Petersen GD, Cumming GS, Carpenter SR. 2003. Scenario Planning: a Tool for Conservation in an Uncertain
World. Conservation Biology, 17(2): 358-366
Rautio P, Markkola A. 2006. Competition or facilities: effects on Empetrum nigrum mycorrhizal colonization
on seedlings of Pinus sylvestris in a successional gradient (Manuscript unpublished)
Scharff N, Coddington JA, Griswold CE, et al. 2003. When to quit? Estimating spider species richness in a
northern European deciduous forest. Journal of Arachnology, 31: 246-273
Schmidt MH, Roschewitz I, Thies C, et al. 2005. Differential effects of landscape and management on
diversity and density of ground–dwelling farmland spiders. Journal of Applied Ecology, 42: 281-287
Similä M. 2002. Patterns of Beetle Species Diversity in Fennoscandian Boreal Forests: Effects of Forest Age,
Naturalness and Fertility, and Co-variation with Other Forest-dwelling Taxa Volumen 141 de Tiedonantoja
/ Joensuun yliopisto, metsätieteellinen tiedekunta. University of Joensuu, Finland
Similä M, Kouki J, Mönkkönen M. 2002. Ecography: Pattern and diversity in ecology. Diversity and
Distributions, 25(1): 42-52
Slansky F, Rodriguez JG. 1987. Nutritional Ecology of Insects, Mites, Spiders, and Related Invertebrates.
Wiley, USA
Sorensen LL, Coddington JA, et al. 2002 .Inventorying and estimating subcanopy spiders diversity using
semiquantitative sampling methods in anafromontane forest. Environmental Entomology, 31: 319-330
Uotila A, Maltamo M, Uuttera J, et al. 2001. Stand structure in seminatural and managed forests in eastern
Finland and Russian Karelia. Ecological Bulletins, 49: 149-158
Uotila A, Kouki J, Kontkanen H, et al. 2002. Assessing the naturalness of eastern Fennoscandian boreal forests.
Forest Ecology and Management, 161: 257-277
Wiggins GB, Marshall SA, Downes JA. 1991. The importance of research collections of terrestrial arthropods.
A brief prepared by the Biological Survey of Canada (Terrestrial Arthropods). Bulletin of the
Entomological Society of Canada, 23(2): 16
Wilson EO. 1987. The little things that run the world (the importance and conservation of invertebrates).
Conservation Biology, 1: 344-346
Wise DH. 1993. Spiders in Ecological Webs. Cambridge University Press, Cambridge, UK
Zhang WJ. 2011a. Constructing ecological interaction networks by correlation analysis: hints from community
sampling. Network Biology, 1(2): 81-98
Zhang WJ. 2011b. Simulation of arthropod abundance from plant composition. Computational Ecology and
Software, 1(1): 37-48
Zhang WJ, Zhong XQ, Liu GH. 2008. Recognizing spatial distribution patterns of grassland insects: neural
network approaches. Stochastic Environmental Research and Risk Assessment, 22(2): 207-216
19

Arthropods, 2014, 3(1): 20-26
IAEES www.iaees.org
Article
Mineral composition of edible crab Podophthalmus vigil Fabricius
(Crustacea: Decapoda)
P. Soundarapandian1, D. Varadharajan1, S. Ravichandran2 1Faculty of Marine Sciences, Centre of Advanced Study in Marine Biology, Annamalai University, Parangipettai-608 502. Tamil
Nadu, India 2Department of Zoology, Government Arts College, Kumbakonam, India
E-mail: [email protected]
Received 21 February 2013; Accepted 25 March 2013; Published online 1 March 2014
Abstract
Totally 7 minerals were reported in the present study. For the individual contribution 5
(Sodium>Calcium>Potassium>Iron>Magnesium),7(Sodium>Calcium>Iron>Potassium>Phosphorus>Magnesi
um>Zinc) and 4(Calcium>Sodium>Iron>Magnesium) minerals were reported in males, females and berried
females respectively. In all sexes sodium and calcium were maximum and magnesium was minimum.
Comparatively females contain maximum amount of minerals than males and berried females. Phosphorous
and zinc were absent in males whereas potassium was absent in addition to phosphorous and zinc in berried
females. Among different sexes females contain maximum amount of minerals (61.56 mg) followed by males
(39.92 mg) and berried females (35.11 mg). From the study females contain maximum amount of minerals
than berried females and males. So it is recommended to consume females to get maximum minerals.
Keywords minerals; males; females; berried females; Podophthalmus vigil.
1 Introduction
Minerals are called as micronutrients and necessary for physiological and biochemical processes by which the
human body acquires assimilates and utilized food to maintain health and activity but also ensuring adequate
immune-competence and cognitive development. So exploring minerals from marine organisms is important
especially in crabs. It becomes a worldwide delicacy amongst seafood aficionados and high in essential
nutrients and is extremely beneficial for health (Soundarapandian and Ananthan, 2008; Soundarapandian et al.,
2010). Thus, the meats are also prominent sources of minerals, mainly iron, calcium, potassium phosphorus
and zinc, which aids in reducing oxidative damage to cells and tissues and acts as an antioxidant by cancelling
out the carcinogenic effects. Mineral values of different crab species have been previously investigated in
various parts of the world. Davis (1996) studied marine crustaceans; Küçükgülmez et al. (2006) studied
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences
Arthropods, 2014, 3(1): 20-26
IAEES www.iaees.org
Callinectes sapidus; Moronkola et al. (2011) reported Callinectes amnicola. Jimmy and Arazu (2012) studied
two edible Crabs Callinectes amnicola and Uca tangeri. However, the study of the mineral content in
Podophthalmus vigil Fabricius is very limited. Since crab is consumed by local population on regular basis. In
the present investigation mineral content was studied in all sexes of the crab, P. vigil.
2 Materials and Methods
The male, female and berried females of P. vigil were procured from Parangipettai
(Lat. 11021’ N; Long. 790 46’ E) landing centres. The carapace of the crabs was opened and the edible parts of
muscle tissues were removed with sharp forceps. The removed muscle tissues were homogenized with pestle
and mortar. The grounded muscles were then freeze dried and powdered and eventually stored in refrigerator
for further analysis. To the 5g of crab tissue samples, mixture of hydrochloric acid, nitric acid and perchloric
acid at a ratio of 10:5:1 was added for digestion at 30 ºC. The digests were filtered suitably and aspirated in
digital flame photometer (Modal No.CL 22D, Elico pvt, India), the values obtained were expressed in mg/100g
(Guzman and Jimeneza, 1992).
The data were subjected to One-way Analysis of Variance (ANOVA) and difference between means were
determined by Duncan’s multiple range tests (P<0.05) using SPSS version 17.0.
3 Results
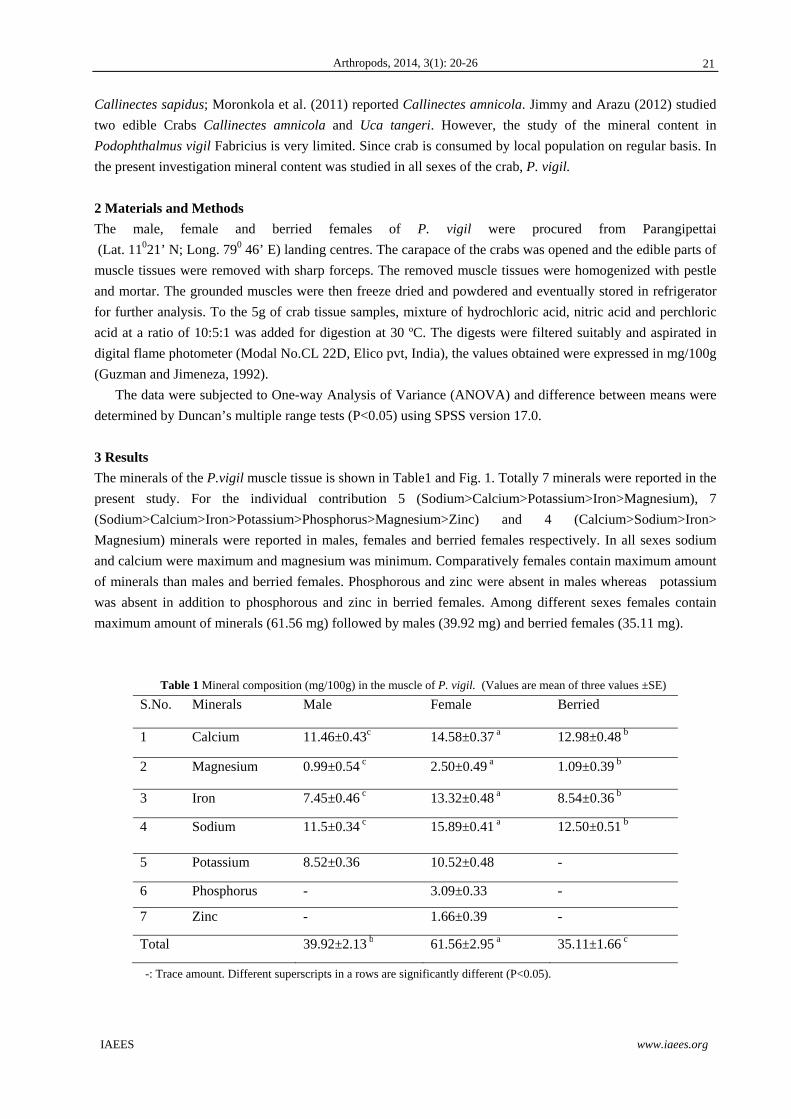
The minerals of the P.vigil muscle tissue is shown in Table1 and Fig. 1. Totally 7 minerals were reported in the
present study. For the individual contribution 5 (Sodium>Calcium>Potassium>Iron>Magnesium), 7
(Sodium>Calcium>Iron>Potassium>Phosphorus>Magnesium>Zinc) and 4 (Calcium>Sodium>Iron>
Magnesium) minerals were reported in males, females and berried females respectively. In all sexes sodium
and calcium were maximum and magnesium was minimum. Comparatively females contain maximum amount
of minerals than males and berried females. Phosphorous and zinc were absent in males whereas potassium
was absent in addition to phosphorous and zinc in berried females. Among different sexes females contain
maximum amount of minerals (61.56 mg) followed by males (39.92 mg) and berried females (35.11 mg).
Table 1 Mineral composition (mg/100g) in the muscle of P. vigil. (Values are mean of three values ±SE)
S.No. Minerals Male Female Berried
1 Calcium 11.46±0.43c 14.58±0.37 a 12.98±0.48 b
2 Magnesium 0.99±0.54 c 2.50±0.49 a 1.09±0.39 b
3 Iron 7.45±0.46 c 13.32±0.48 a 8.54±0.36 b
4 Sodium 11.5±0.34 c 15.89±0.41 a 12.50±0.51 b
5 Potassium 8.52±0.36 10.52±0.48 -
6 Phosphorus - 3.09±0.33 -
7 Zinc - 1.66±0.39 -
Total 39.92±2.13 b 61.56±2.95 a 35.11±1.66 c
-: Trace amount. Different superscripts in a rows are significantly different (P<0.05).
21

Arthropods, 2014, 3(1): 20-26
IAEES www.iaees.org
Fig. 1 Mineral composition (mg/100g) in the muscle of P. vigil.
4 Discussion
Marine foods are very rich sources of mineral components. The total content of minerals in the raw flesh of
marine fish and invertebrates is in the range of 0.6 – 1.5% wet weight. Mineral components such as sodium,
potassium, magnesium, calcium, iron, phosphorus and iodine are important for human nutrition (Sikorski et al.,
1990). Crustaceans are also good sources of various minerals and high quality protein. Crab meat is an
excellent source of minerals, particularly calcium, iron, zinc, potassium and phosphours (Sifa et al., 2000;
Adeyeye, 2002; Gokoglu and Yerlikaya, 2003; Naczk et al., 2007). Living organisms require trace amounts of
some heavy metals including iron, cobalt, copper, manganese, molybdenum, strontium, vanadium and zinc.
Excessive levels of these metals, however, can be detrimental to living organisms (Prajapati et al., 2012).
Other heavy metals such as cadmium, lead and mercury have no known beneficial effect on organisms and
their accumulation over time in the bodies of mammals can cause serious illness (Hawkes, 1997). The fish and
shellfish can absorb minerals directly from the aquatic environment through gills and body surfaces. Almost all
the elements that occur in seawater are found to some extent in aquatic animals and these includes Na, K, Ca,
P, Al, Ba, Cd, I, Cr, Pb, Li, Hg, Ag, St and Va. Eyo (2001) reported that the mineral content of fish makes
unavoidable in the diet, as it is a source of different minerals that contribute greatly to good health.
The minerals are serving as components of bones, soft tissues (Sulfur amino acids, metalloproteins) co-
factors and co-activators of various enzymes important in human nutrition. Calcium, phosphorus, magnesium
and electrolytes (sodium and potassium) are considered to be as macro elements and iron, copper, zinc, iodine,
chromium, cobalt, manganese, molybdenum, selenium are considered as trace elements that are required for
normal functioning, for instance the more soluble minerals such as Ca, P, Na, K and Cl are involved in the
maintenance of acid-base balance and membrane potential. The main functions of essential minerals include
skeletal structure, maintenance of colloidal system and regulation of acid-base equilibrium. Minerals also
constitute important component of hormones, enzymes and enzyme activators (Belitz and Grosch, 2001). It is
known that variations in the mineral composition of marine foods are closely related to seasonal and biological
differences (species, size, dark/white muscle, age, sex and sexual maturity), area of catch, processing method,
food source and environmental conditions (water chemistry, salinity, temperature and contaminant).
22

Arthropods, 2014, 3(1): 20-26
IAEES www.iaees.org
Considering the elemental composition of common food items (dairy products, meat, fish, cereals and
fruits), C. pagurus hepatopancreas is a good source of Ca, Fe, Cu, Zn and Se (FAO/WHO, 2002; Martins,
2006). The more soluble minerals such as Ca, P, Na, K and Cl also have osmoregulatary function and the
maintenance of acid-base balance and membrane potentials (Davis et al., 1992). Some elements such as Mg,
Al, Ca, Fe, Co, Cu and Zn are necessary for maintenance of optimum health thus are important from nutrition
point of view. Metals such as Pb, Cd, As and Hg are detrimental to optimum health and have toxicological
effect and the tissue samples are also used as a bio-indicator to assess bioavailability of contaminant
concentrations in coastal water in environmental studies (Mohapatra et al., 2007).
The aquatic environmental/ecosystem and their inhabitants are exposed and sensitive to effects of
environmental pollution from heavy metal contamination. Aquatic animals accumulate large quantities of these
xenobiotics and the accumulation depends upon the intake and elimination from their body (Karadede et al.,
2004). Among different aquatic organisms; oysters, crab and mussels, accumulate large quantities of heavy
metals due to their habitat and feeding nature. Many metals (Co, Cu, Mn, Fe and Zn) are essential trace
elements for aquatic organisms and are involved in biochemical processes such as enzyme activation (Lall,
1989).
In the present study, sodium and calcium was maximum and Magnesium was minimum irrespective of the
sex. In individual contribution 5(Sodium > Calcium > Potassium > Iron > Magnesium), 7 (Sodium >
Calcium > Iron> Potassium > Phosphorus > Magnesium > Zinc) and 4(Calcium> Sodium > Iron > Magnesium)
minerals are reported in males, females and berried females respectively. These are very much comparable
with studies of Hagashi et al. (1979), Anon (1999), Thirunavukkarasu (2005) and Sudhakar (2009). Gokoglu
and Yerlikaya (2003) investigated the mineral contents of blue crab, C. sapidus and swimming crab P.
pelagicus and suggested that Na, Ca, Zn, Cu values for blue crab and swimming crab were not significantly
different. Trace elements content in haemolymph of normal and red sternum mud crab were observed by
Salaenoi et al (2006). The average Ca contents of green tiger shrimp and speckled shrimp were 60.28mg/10 g
and 60.44 mg/10 g, respectively (Yannar and Celik, 2006). Chen et al. (2007) reported the concentration of
nine elements (Zn, Fe, K, Na, Mn, Cu, Mg, Ca, and P) in different tissues of crab meat and edible viscera of
Chinese mitten crab, E. sinensis. Mohapatra et al. (2009) studied the concentration of 10 elements (ppm) (K,
Ca, Mn, Fe, Cu, Zn, Se, Br, Sr and Pb) in S. serata, S. tranquebarica, P. monodon, P. indicus and
M.rosenbergii. Sudhakar et al. (2009) assessed the minerals content of hard and soft shell crabs P.
sangiunolentus. Mohapatra et al. (2009) recorded the concentration of nine elements (K, Ca, Mn, Fe, Cu, Zn,
Se, Br and Pb) in different tissues of mud crab S. serrata.
The calcium and phosphorus together account for 70 to 80% of the minerals in the skeleton of fish (Nair
and Mathew, 2000). In the present study calcium and megnesium alone contribute 50%. Calcium is maximum
in females than males of P. vigil. Similar results were reported in P. sanguinolentus (Sudhakar et al. 2009), S.
tranquebarica (Thirunavukkarasu, 2005) and E. sinesnsis (Chen et al., 2007). Ca has an essential role in blood
clotting, muscle contraction and nerve transmission. Calcium is nutritionally very important (up to 1.9% Ca is
available in human body) and provides rigidity to the skeleton and plays a role in many metabolic processes
(FAO/WHO, 2002). It is also essential for hard tissue structure, blood clotting, muscle contraction, nerve
transmission and osmoregulation and as a cofactor for enzymatic procession (Lovel, 1989). The higher Ca
content in male crabs are likely because this species has a sexual dimorphism, in which males have bigger
claws and harder exoskeletons (composed by calcium phosphate). Particularly during the premoult period of C.
pagurus, hepatopancreas accumulates Ca that is likely used in the exoskeleton calcification (Luquet and Marin,
2004).
23

Arthropods, 2014, 3(1): 20-26
IAEES www.iaees.org
Magnesium is maximum in females than berried females and males. Magnesium was already reported in P.
sanguinolentus (Sudhakar et al., 2009), S. tranquebarica (Thirunavukkarasu, 2005) and E. sinesnsis (Chen et
al., 2007). Magnesium is important for human nutrition and it is required for body’s enzyme system. In
addition to maintain bone health, magnesium acts in all cells of soft tissues, where it forms part of the protein-
making machinery and necessary for energy metabolism. Mg is cofactor for enzyme systems (Food and
Nutrition Board, National Research council, 1989).
Iron is maximum in females than berried females and males. Iron was already reported in E.sinesnsis
(Chen et al., 2007). Iron is one of the very important essential trace elements since it has several vital
functions in human system. It serves as a carrier of oxygen to tissues from the lungs by red blood cell.
Adequate Fe in the diet is very important for avoiding some major health problems (Belitz and Grosch, 2001;
Camara et al., 2005). Adequate iron in the diet is very important for decreasing the incidence of anaemia,
which is considered a major health problem, especially in young children. Iron deficiency occurs when the
demand for iron is high, e.g., in growth, high menstrual loss, and pregnancy (Belitz and Grosch, 2001; Camara
et al., 2005). Transition metal ions, particularly Cu and Fe, have been known as the major catalysts for
oxidation (Thanonkaew et al., 2006). Sodium contribution is maximum irrespective of the sex. In individual contribution sodium is the highest
in females followed by berried females and males of P. vigil. Sodium was already reported in P.
sanguinolentus (Sudhakar et al., 2009), S. tranquebarica (Thirunavukkarasu, 2005) and E. sinesnsis (Chen et
al., 2007). Sodium is the principal cation of the extra cellular fluid and regulator of its volume. Sodium also
helps to maintain acid-base balance and is essential for nerve system.
Potassium is maximum in females than males and totally absent in berried females. Potassium was already
reported in P. sanguinolentus (Sudhakar et al., 2009), S. tranquebarica (Thirunavukkarasu, 2005) and E.
sinesnsis (Chen et al., 2007). Potassium is important to maintain the pH, storage and transfer of energy and
nucleotide synthesis. Phosphorous is available only in females and totally absent in males and berried females.
Phosphorous was already reported in E. sinesnsis (Chen et al., 2007). The phosphorous (adenosine
polyphosphate) act as a key substance for energy release and present in phospholipids (Decker and Tuczek,
2000). Ca and P are necessary to maintain an optimal bone development, with more of both minerals being
required during childhood and growing stages to prevent rickets and steomalacia. The calcium and
phosphorous together account for 70 to 80% of the minerals in the skeleton of fish (Nair and Mathew, 2000).
Zinc is available only in females and totally absent in males and berried females. Zinc was already reported
in P. sanguinolentus (Sudhakar et al., 2009), S.tranquebarica (Thirunavukkarasu, 2005) and E. sinesnsis
(Chen et al., 2007). Zinc is an essential trace element for all living species, since is an important component of
several enzymes (Chen et al., 2007) and plays an essential role in a number of biological processes involved in
growth and development (FAO/WHO, 2002). MacFarlane et al. (2000) in semaphore crab also reported higher
Cu and Zn accumulation in females than to males. From the study female contains maximum amount of
minerals than berried females and males. So it is recommended to consume females to get maximum minerals.
References
Adeyeye EI. 2002. Determination of chemical composition of the nutritionally valuable parts of male female
common West African freshwater crab. Sudanautes africanus africanus. International Journal of Food
Science Nutrition, 52(3): 189-196
Anon 1999. Results from the USDA, nutrient database for standard reference crustaceans, Crab blue, cooked,
moist head. Blue crab-Nutrition html. 1-3.
24

Arthropods, 2014, 3(1): 20-26
IAEES www.iaees.org
Belitz HD, Grosch W. 2001. Schieberle, P.Lehrbuch der Lehbensmittelchemie, Aufl. Springer-Verlag, Berlin,
Heidelberg, New York
Camara F, Amaro MA, Barbera R, Clemente G, et al. 2005. Bioaccesibility of minerals in school meals;
comparision between dialysis and solubility methods. Food Chemistry, 92: 481-489
Chen HP, Zhang M, Shrestha S, et al. 2007. Compositional characteristics and nutritional quality of Chinese
mitten crab (Eriocheir sinensis ). Food Chemistry, 103: 1343-1349
Davis A. 1996. Dietary mineral requirements of fish and marine crustaceans. Reviews in Fisheries Science,
4(1): 75-99
Davis DA, Lawrence AL, Garlin DM, et al. 1992. Mineral requirements of Penaeus vannamei, a preliminary
examination of the dietary essentially of thirteen minerals. Journal of World Aquaculture Society, 23: 8-14
Decker H, Tuczek F. 2000. Tyrosinase/catecholoxidase activty of hemocyanins: Structural basis and molecular
mechanism. Trends in biochemical Sciences, 25: 392-397
Eyo AA. 2001. Fish Processing Technology in the Tropics. 66-70, National Institute for freshwater, Fisheries
Research, University of Ilorin Press, Nigeria
FAO/WHO. 2002. Human vitamines and mineral requirements. Report of joint food and Agricultural
Organizations of the United Nations/World Health Organization Expert Consultation. Bankok, Thailand
Food and Nutrition Board, National Research Council, 1989. Recommended Dietary Allowances (10th edn).
National Academy Press, Washington DC, USA
Gokoglu N, Yerlikaya P. 2003. Determination of proximate composition and mineral contents of blue crab
(Callinectus sapidus) and swim crab (Portuns pelagicus) caught off the Gulf of the Antalya. Food
Chemistry, 80: 495-498
Guzman HM, Jimeneza CE. 1992. Concentration of coral reefs by heavy metals along the Carribean cost of
Central America (Costarica and Panama). Marine Pollution Bulletin, 24(11): 554-561
Hagashi T, Asakawa A, Yamaguchi K, Konoso S. 1979. Studies on flavour components in boiled crabs.
Bulletin Japanese Society of Scientific Fisheries, 45(10): 1325-1329
Hawkes SJ. 1997. What is a “heavy metal”? Journal of Chemical Education, 74: 1374
Jimmy UP, Arazu VN. 2012. The Proximate and Mineral Composition of Two Edible Crabs Callinectes
amnicola and Uca tangeri (Crustecea: Decapoda) of The Cross River, Nigeria. Pakistan Journal of
Nutrition, 11: 78-82
Karadede H, Oymak SA, Unlu E, et al. 2004. Heavy metals in mullet, Liza abu and catfish, Silurus triostegus
from the Ataturk Dam Lake (Euphrates), Turkey. Environment International, 30: 183-188
Küçükgülmez A, Mehmet Ç, Yasemen Y, et al. 2006. Proximate composition and mineral contents of the blue
crab (Callinectes sapidus) breast meat, claw meat and hepatopancreas. International Journal of Food
Science and Technology, 41(9): 1023-1026
Lall SP. 1989. The mineral. In: Fish Nutrition (2nd edn) (Halver JE, ed). 219-257, Academic press, New York,
USA
Lovel RT. 1989. Nutrition and Feeding of Fish. Van Nostrand Reinhold, New York, USA
Luquet G, Marin F. 2004. Biomeralization in crustaceans: storage strategies. Comptes Rendus Palevol, 3(6-7):
515-534
MacFarlane GR, Booth DJ, Brown KR. 2000. The Semaphore crab, Helioecius cordiformis: Bio-indication
potential for heavy metals in estuarine systems. Aquatic Toxicology, 50: 153-166
Martins I. 2006. Food Composition Table. Centre of Nutrition and Food Safety of Health Ministry, Lisbon,
Portugal
25

Arthropods, 2014, 3(1): 20-26
IAEES www.iaees.org
Mohapatra A, Rautray TR, Vijan V, et al. 2007. Trace elements characterization of some food crustacean
tissue samples by EDXRF technique. Aquaculture, 270: 552-558
Mohapatra A, Rautray TR, Patra AK, et al. 2009. Elemental composition in mud crab, Scylla serrata from
Mahanadi estuary India. Food and chemistry Toxicology, 47: 119-123
Moronkola BA, Olowu RA, Tovidea OO, et al. 2011. Determination of proximate and mineral contents of crab
(Callinectes Amnicola) living on the shore of Ojo River, Lagos. Nigeria Scientific Reviews and Chemical
Communications, 1(1): 1-6
Naczk M, Williams J, Brennam K, et al. 2007. Compositional characteristics of green crab (Carcinus sapidus).
Food Chemistry, 88: 429-434
Nair PG, Mathew S. 2000. Biochemical composition of fish and shellfish. CIFT Technology Advisory Series.
CIFT, India
Prajapati SK, Meravi N, Singh S. 2012. Phytoremediation of Chromium and Cobalt using Pistia stratiotes: A
sustainable approach. Proceedings of the International Academy of Ecology and Environmental Sciences,
2(2): 136-138
Salaenoi J, Sancharoen A, Thongpan A, et al. 2006. Morphology and hemolymph composition changes in red
sternum mud crab (Scylla serrata). Kasetsart Journal-Natural Sciences, 40: 158-166
Sifa L, Wanqi C, Shuming Chenhong ZL, et al. 2000. Quality analysis of Chinese mitten crab Eriocheir
sinensis in Yangchenghu Lake. Journal of Fisheries of China, 7(3): 71-74
Sikorski ZE, Lolakowska A, Pan BS. 1990. The nutrition composition of the major groups of marine food
oraganisms. In: Resources Nutritional Compositon and Preservation (Sikorski ZE, ed). 30-52, CRC, Boca
Raton, Florida, USA
Soundarapandian P, Ananthan G. 2008. Effect of unilateral eyestalk ablation and diets on the biochemical
composition of commercially important juveniles of Macrobrachium malcomsonii (Edwards). International
Journal of Zoological Research, 4(2): 106-112
Soundarapandian P, Dinakaran GK, Mrinmoy G, et al. 2010. Effect of diets on the biochemical changes of
fattened commercially important crab Portuns sanguinolentus (Herbst). Current Research Journal of
Biological Sciences, 2(2): 107-113
Sudhakar M, Manivannan K, Soundarapandian P, et al. 2009. Nutritive value of hard and soft shell crabs of
Portunus sanguinolentus (Herbst). Internatational Journal of Animal and Veterinary Advance, 1(2): 44-48
Thanonkaew A, Benjakul S, Visessanguan W, et al. 2006. The effect of metal ions on lipid oxidation, colour
and physiological properties of cuttlefish (Sepia pharaonis) subjected to multiple freeze thaw cycles. Food
Chemistry, 94: 210-219
Thirunavukkarasu N. 2005. Biology, nutritional evaluation and utilization of mud crab Scylla tranquebarica
(Fabricius). Ph.D Thesis. Annamalai University, India
Yannar Y, Celik M. 2006. Seasonal amino acid profiles and mineral contents of green tiger shrimp (Penaeus
seminsulcatus De Haan, 1844) and speckled shrimp (Metapenaeus monoceros Fabricius, 1789) from the
eastern Mediterrranean. Food Chemistry, 94: 33-36
26

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
Article
The effects of some domestic pollutants on the cumacean (Crustacea)
community structure at the coastal waters of the Dardanelles, Turkey
A.Suat Ateş1, Tuncer Katağan2, Murat Sezgin3, Hasan G. Özdilek4, Selçuk Berber1, Musa Bulut1 1Çanakkale Onsekiz Mart University, Faculty of Marine Sciences and Technology, TR- 17100 Çanakkale, Turkey 2Ege University, Fisheries Faculty, TR- 35100 Bornova-İzmir, Turkey 3Sinop University, Fisheries Faculty, TR- 57000 Sinop, Turkey 4Çanakkale Onsekiz Mart University, Faculty of Engineering and Architecture, TR- 17100 Çanakkale, Turkey
E-mail: [email protected]
Received 2 September 2013; Accepted 8 October 2013; Published online 1 March 2014
Abstract
This study was carried out to determine the effects of sewage pollution on the cumacean assemblages found in
the coastal waters of the Dardanelles. The samples were collected by a SCUBA diver between July 2008 and
April 2009 and a total of 102 specimens belong to 5 cumacea species, Bodotria arenosa mediterranea,
Cumopsis goodsir, Cumella limicola, Iphinoe maeotica and Pseudocuma longicorne was recorded. The
dominant species, Iphinoe maeotica has the highest dominance value (36.66%). Multiregression approach
resulted in statistically insignificant relationship between physical, chemical and biochemical variables of
water and sediment and Bodotria arenosa mediterranea, Cumopsis goodsir, Cumella limicola, and Iphinoe
maeotica. Based on multiple regression test, a significant relationship with R2 = 92.2%, F= 7.876 and p= 0.000
was found between six water and sediment quality constituents and numbers of Pseudocuma longicornis at the
stations studied of the Dardanelles. On the other hand, water temperature (β= -0.114; t= -2.811, p= 0.016);
sediment organic matter (β= -0.011; t= -2.406; p= 0.033) and water phosphorus (PO4) (β= 0.323; t= 3.444; p=
0.005) were found to be the most important water and sediment parameters that affect Pseudocuma longicornis.
Keywords Cumacea; crustacean; community; sewage pollution; the Dardanelles; Turkey.
1 Introduction
Every human activity generates wastes in all forms, solid, gaseous, and liquid (Metcalf and Eddy, 2003).
Domestic water pollution from industrial, agricultural and urban areas end up in freshwater or brackish water
sources, affecting water resources if not properly treated before discharge (Botkin and Keller, 2003). Coastal
water pollution is among one of the most crucial water pollution problems since community use such sources
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
for fishing, swimming and other recreational activities. In Turkey, wastewater treatment is a major problem
since 45% of domestic wastewater is discharged into seas (TSI, 2010). Despite the fact that industrial water
pollution is not a major problem in Turkey, quantity of domestic wastewater has increased dramatically over
the past few decades due to expansion of urbanization in Turkey. Turkish seas are connected via two sea straits,
named Bosporus and Dardanelles. Dardanelles connects the Aegean Sea and the Sea of Marmara. Dominant
water flow is from Black Sea to the Sea of Marmara and then to the Aegean. However, coastal urban areas are
main concern due to their untreated wastewater discharges followed inputs from ships. It is reported that
approximately 50000 ships pass through the Dardanelles every year. Coastal zone, however, are more polluted
with wastewaters disposal from small communities followed by ship activities. The effect of untreated
wastewater disposal on coastal zone is in general observed as eutrophication. With the effect of dry period and
ambient air temperature, eutrophication is observed especially in August. However, dilution of water pollution
with respect to higher precipitation and therefore runoff do not cause eutrophication in the winter months,
especially in January.
Crustaceans are mostly used as bioindicators and biomonitors in different aquatic ecosystems and they are
known as bioindicators in polluted areas (Rinderhagen et al., 2000). Sewage pollution impacts benthic
organisms. Changes due to pollution are determined by community structure of benthic fauna primarily
(Arasaki et al., 2004). Although some temporary changes in organic matter are observed due to sewage
pollution at initial stages, sensitive species soon leave the polluted zone if pollution is a persistent problem
(Bat et al., 2001). To determine the distribution of indicator species is crucial in maping of pollution gradients
(Corbera and Cardell, 1995). Many study on the subject were carried out on polychaetes and molluscs (Bellan,
1967).
Crustaceans are the first of sensitive livings among benthic assemblages affected by quantity of sewage
pollution (Bat et al., 2001; Del Vals et al., 1998; Guerra-García and García-Gómez, 2004). Physical
environmental factors as well as pollutant concentrations are important in determination of benthic community
and low biodiversity is a fact on sewage pollution affected areas in coastal zones (Morrisey et al., 2003).
Sewage pollution’s effects on crustaceans in the Mediterranean ecosystem are found in the following studies.
Del Val et al. (1998) and Guerra-García and García-Gómez (2004) studied the effects of sewage pollution on
crustaceans found in soft bottoms of two different littoral systems of Cadiz Bay (the eastern Atlantic) and of
Ceuta Harbour (the Gibraltar Strait) respectively. Additionally, García Raso and Manjón Cabeza (2002)
determined the effects of sewage pollution on decapod crustaceans in the upper-infralittoral zone of Barbate
Coast (the southern Spain). Recently, Hamouda and Abdel-Salam (2010) studied the distribution models of
macrobenthic communities including cumaceans found in Abu-Qir Bay (Alexandria, Egypty) in where organic
pollution occurs. Extreme organic pollution causes to disappear of sensitive species in the environment
(Hamouda and Abdel-Salam, 2010).
Bat et al. (2001) mentioned effects of domestic wastewater discharge on several crustacean species at the
depths of 0.5-1 m on the coast of Sinop Harbour (the southern Black Sea, Turkey). Albayrak et al. (2006)
sampled four different locations in the northern Marmara Sea in order to determine levels of pollution due to
organic material discharges. Tuğrul-İçemer and Koşun (2003) carried out a study underlining that benthic
community structure under sewage pollution in Antalya Bay (the eastern Mediterranean). Similar to our study,
Corbera and Cardell (1995) studied indicative cumaceans affected by eutrophication in the soft bottoms of
Barcelona coast (the southern Spain).
Despite of several studies (Tuğrul-İçemer and Koşun, 2003 for the Mediterranean Sea, Kocataş et al., 1988
for the Aegean Sea, Albayrak et al., 2006 for the Sea of Marmara, Okuş et al., 1996 for the Bosporus, Bat et al.,
2001 for the Black Sea) regarding the effects of pollution on benthic communities of the Turkish coast
28

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
occurred, there is no research indicated the effects sewage pollutants on the structure of macrozoobenthic
communities in relation to environmental conditions in the Dardanelles. The Dardanelles is a long, narrow, and
shallow strait. Saros Bay is located in the North part of the Dardanelles is known as an important fisheries
activity area and it is expressed that water coming from the Black Sea is discharged into the northern Aegean
and the Saros after passing the Dardanelles. Therefore, the end point of the water flow from the Black Sea is
this region. Any type of pollution is highly expected to accumulate in the region.
The aim of this study is to determine the effects of sewage pollution on the cumacean assemblages on soft
bottoms of the Dardanelles.
2 Material and Methods
2.1 Sample collection, study area and field measurements.
Study area includes 8 sampling points, 4 of them are located on the Anatolian Coast and other 4 are located on
the European Side. GPS coordinates of the sampling stations are: 1. Gelibolu; 40°40΄617˝N 26°66΄692˝E, 2.
Lapseki; 40°34΄661˝N 26°67΄985˝E, 3. Çanakkale; 40°15΄474˝ N 26°40΄879˝ E, 4. Kilya Inlet (reference site);
40°20΄472˝N 26°36΄117˝E, 5. Eceabat; 40°18΄253˝N 26°36΄046˝E, 6. Kilitbahir; 40°15΄048˝N 26°37΄878˝E, 7.
Kepez Harbour (reference site); 40°10΄360˝N 26°37΄339˝E, 8. Dardanos; 40°07΄493˝N 26°35΄806˝E (Fig. 1).
Samples of cumacean at the depths of 0 to 5 m were collected from three different transects (depth counters of
0.5, 2 and 4 m) using a quadrate system of 30x30 cm by a Scuba diver and preserved in buffered formalin.
Samplings were carried out on July 17, 2008, November 12, 2008, February 16, 2009 and April 29, 2009,
seasonally.
Fig. 1 Map of the study area showing the sampling stations (1: Gelibolu, 2: Lapseki, 3: Çanakkale, 4: Kilya Inlet, 5: Eceabat, 6: Kilitbahir 7: Kepez Harbour, 8: Dardanos).
2.2 Sediment type of sampling sites
Sampling depths were selected in localities that pouring down domestic waste to the marine environment. For
the reference stations, the same sampling depths were adopted. Hence, the depth of each sample showed a
different structure of the bottom. The bottom of 2-5 m of Kilitbahir sampling station is partly covered by
Mytilus galloprovincialis. Kilya station is silty. Eceabat station (2-4 m) is sandy bottom with meadows,
29

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
Zostera marina. While Lapseki sampling site has fine sand, Çanakkale sampling site is partly covered by algae
and coarse sand with rocks. Kepez Harbour is dominated by mytilid bivalve, Mytilus galloprovincialis at depth
of 4 m. Dardanos sampling site is covered with fine sand of meadows, Zostera marina. Sediment organic
matter was determined by 550 °C incineration of washed known weight of sediment samples (generally 50
grams) for two hours. The difference between incinerated and raw sediment weight provided percent organic
matter. Sediment organic matter was determined by the high temperature oxidation method adopted by Craft et
al., 1991. In addition to loss of ignition at 550 °C using furnace, organic carbon and total nitrogen were also
determined by the high temperature oxidation method. Loss of ignition method requires a weighed dry
material and slowly increasing furnace set up 550 °C and waiting generally 1 to 2 hours at this temperature.
TOC analyzers are gaining in favour because of short test periods. Clean sand sample was also tested to
compare with results obtained from sediment samples.
2.3 Sea water quality parameters
Dissolved oxygen, water temperature, electrical conductivity, pH and salinity were measured using YSI 556
model MPS on site. Water samples were collected using dark color 1 liter clean glass bottles. Ammonia,
nitrate, nitrite, Biological Oxygen Demand, Chemical Oxigen Demand, and anionic detergents were
determined using Standard Methods (Apha Awwa, 2005). Moreover, water organic constituents were
determined using Hach-5000 spectrophotometer. There replicates were tested for all parameters and
statistically no difference were found after data evaluation. Quality control was ensured when testing
parameters in laboratory.
2.4 Ecological data analyses
Triocular stereo microscope was used to determine of specimens belonging to cumaceans found from the study
area. The specimens of cumacean were defined based on the study of Ledoyer (1965). To elucidate the
community structure, Soyer (1970)’s frequency index (¦f%), Bellan-Santini (1969)’s quantitative dominance
index (Di%), Shannon and Weaver (1949)’s diversity index (Hˊ) as well as its evenness component (Jˊ) and
Bray and Curtis (1957)’s similarity index were calculated. The community structure was investigated by
group-averaging cluster analysis based on the Bray and Curtis (1957). In calculating of (Shannon and Weaver,
1949)’s diversity index (Hˊ) as well as its evenness component (Jˊ) primer program 6.0 was used.
The frequency index of a particular species was estimated by
f= (m/M)×100
where, m is number of stations where species of concern exists, and M is number of all stations.
The dominance index of a certain species was estimated by Di = (m/M)×100, where m = individual number
of a species in the stations and, M = total individual numbers of all species.
The Shannon-Weaver diversity index was estimated by
H´= - Σ pi (log2pi),
where S = total individual number of a species, and N =total individual numbers of all species.
The Pielou evenness index was estimated by
J´= H´/ log2S
where H’= Shannon index value; S = species number.
Bray and Curtis (1957)’s similarity index is
Bray-Curtis Sjk=100 {1- |Σ|yij - yik|/Σ|yij + yik||}
2.5 Statistical data analyses
Variance analyses were completed to see whether there are statistical differences between sampling sites and
sampling time (seasons). These analyses were completed using SPSS 10.0. Moreover, physical and chemical
water quality parameters were tested in order to see which ones are correlated. Friedmann test was adopted to
30

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
see cumacea species distribution with respect to seasonal changes
FD = [12 / (b×t×(t+1)] × Σ[(Sj)2-(3×b×(t+1)]
where b is number of sampling stations (8) and t is number of seasons (4), and Sj is standart deviation of the
data set.
All collected parameters were evaluated using MANOVA (multiple analysis of variance). The correlation
between species compositioin and polluants was evaluated using a Spearman Correlation. The relationship
between the physico-chemical parameters and domestic pollutants in sea water was identified by the method of
canonical discriminant analysis.
3 Results
3.1 Environmental conditions
Seasonal mean dissolved oxygen (DO) concentration was computed to be 7.13 (±0.59) mg L-1. Çanakkale was
found to have the highest DO concentration (9.79 mg L-1) in April 2009. Lapseki discharge site was found to
have the lowest DO value (3.68 mg L-1) in July 2008. Seasonal average value of salinity was computed to be
25.52 (±1.30) ‰ for all sampling sites and times. Dardanos discharge sampling site was found to have the
highest salinity value (30.5‰) in November 2008. Lapseki discharge sampling site was found to have the
lowest salinity value (23.6‰) in April 2009. Mean sea water temperature among all stations and sampling
times was determined to be 16.02 (±5.82) °C for sampling area. The highest sea water temperature (26.77 °C)
belongs to Kilya reference sampling site in July 2008 despite the fact the lowest (8.87 °C) was recorded at
Gelibolu discharge site in February 2009 (Table 1). Table 1 includes all the physical and chemical data
measured in the sampling area. Based on two-way ANOVA, only sampling times was found to create
differences in DO, pH and sea water temperature statistically. However, both sampling times and sampling
sites were found to create statistical differences in salinity and electrical conductivity according to two-way
analysis of variance (MANOVA).
3.2 Water pollutants (NH4+, NO3
-, NO2-, PO4
-3, sediment organic matter and anionic detergent)
Ammonia (NH4+) concentration of all sampling times and locations was computed as 35.94 (±50.26) mg L-1 on
average. The highest ammonia levels belong to Çanakkale and Lapseki discharges as 168 and 166 mg L-1 in
July 2008 respectively. Kilya and Kepez harbour sites were detected to have the very low ammonia
concentration as 0.06 mg L-1 (both) in April 2009. Moreover, Kilya reference site was found to be the cleanest
in terms of NH4+ as 0.05 mg L-1 in February 2009 (3.68 mg L-1) in July 2008. Nitrate (NO3
-) concentration of
all sampling times and locations was computed as 0.30 (±0.22) mg L-1 on average. The highest nitrate level
(0.85 mg L-1) was detected at the Lapseki discharge in November 2008. Çanakkale and Eceabat discharges
were found to have the lowest nitrate concentration as 0.01 mg L-1 (both) in February 2009. Mean nitrite
(NO2-) level of all sampling times and locations was computed as 0.057 (±0.04) mg L-1. The maximum nitrite
level was measured at the Gelibolu discharge as 0.158 mg L-1 in February 2009. Interestingly Kepez harbour
reference station was found to have a high level of nitrite as 0.151 mg L-1 in February 2009. Dardanos
discharge site was found to have the lowest nitrite concentration of 0.004 mg L-1 in July 2008.
Mean sediment organic matter (percent SOM) of all sampling times and locations was computed as 2.85
(±2.16) %. The maximum SOM level was found at Lapseki discharge as 9.86% in April 2009 possibly due to
less variation in temperature, continuous sewage discharge and structure of the site since it is less affected by
flow in the strait. The lowest SOM (0.55%) was measured at Kilitbahir discharge because of the fact that the
highest flow in the strait is near this site. As expected. Kilitbahir site was recorded with the lowest SOM as
1.75 %. Mean anionic detergent of all sampling times and locations was computed as 0.04 (±0.02) mg L-1. The
highest detergent value. 0.105 mg L-1. was measured at Gelibolu discharge site in July 2008. The lowest
31

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
amount of detergent (0.011 mg L-1) was found at Kepez harbour probably due to the fact that there is a
wastewater treatment plant. Kepez and Kilya reference sites. Average anionic detergent concentration was
computed to be 0.63 and 0.62 mg L-1 at Gelibolu and Lapseki discharge sites to be the highest among all
sampling stations taking into account only of individual sampling sites not the individual sampling time (Table
2).
Table 1 Dissolved oxygen, seawater temperature, seawater salinity, pH, electrical conductivity values measured at study site.
Sampling
Period
July 2008 November 2008 February 2009 April 2009
Stations O2
mg L-1
T
(°C)
S
(‰)
pH O2
mg L-1
T
(°C)
S
(‰)
pH O2
mg L-1
T
(°C)
S
(‰)
pH O2
mg L-1
T
(°C)
S(‰) pH
Çanakk
ale (D) 4.19 23.7 23.3 8.21 5.00 15.25 25.6 8.32 9.63 9.18 27.8 5.30 9.79 14.26 24.4 7.07
Lapseki
(D) 3.68 24.57 22.6 8.15 3.34 15.70 24.6 8.25 9.65 9.31 27.4 6.40 8.72 13.68 23.6 6.85
Gelibol
u (D) 5.58 25.03 22.8 8.33 5.56 16.17 25.5 8.51 9.61 8.87 27.6 7.48 8.13 13.10 24.3 6.50
Kilya
Inlet
(R)
8.46 26.77 23.1 8.53 5.90 16.30 25.7 8.55 9.25 9.24 26.5 7.55 7.95 13.50 23.3 6.48
Eceabat
(D) 7.40 25.60 22.9 8.39 6.01 16.01 25.5 8.46 9.56 9.24 27.4 8.09 8.90 13.31 24.2 6.52
Kilitbah
ir (D) 5.16 25.1 23.1 8.31 5.68 16.37 25.6 8.33 9.20 9.12 27.6 8.79 8.90 13.23 24.3 7.05
Kepez
Harbou
r (R)
5.14 24.39 23.5 8.30 5.28 16.22 26.1 8.45 5.68 9.65 28.3 5.44 8.65 14.10 24.8 6.74
Dardan
os (D) 6.49 24.36 28.1 8.44 5.83 16.07 30.5 8.70 7.94 9.61 28.3 5.13 8.04 15.75 28.6 6.88
DO (mg L-1): Dissolved oxygen. T (°C): Seawater temperature. T (‰): Seawater salinity. pH: Activity of Hidrojen ion. EC (mS/cm): Conductivity.
32
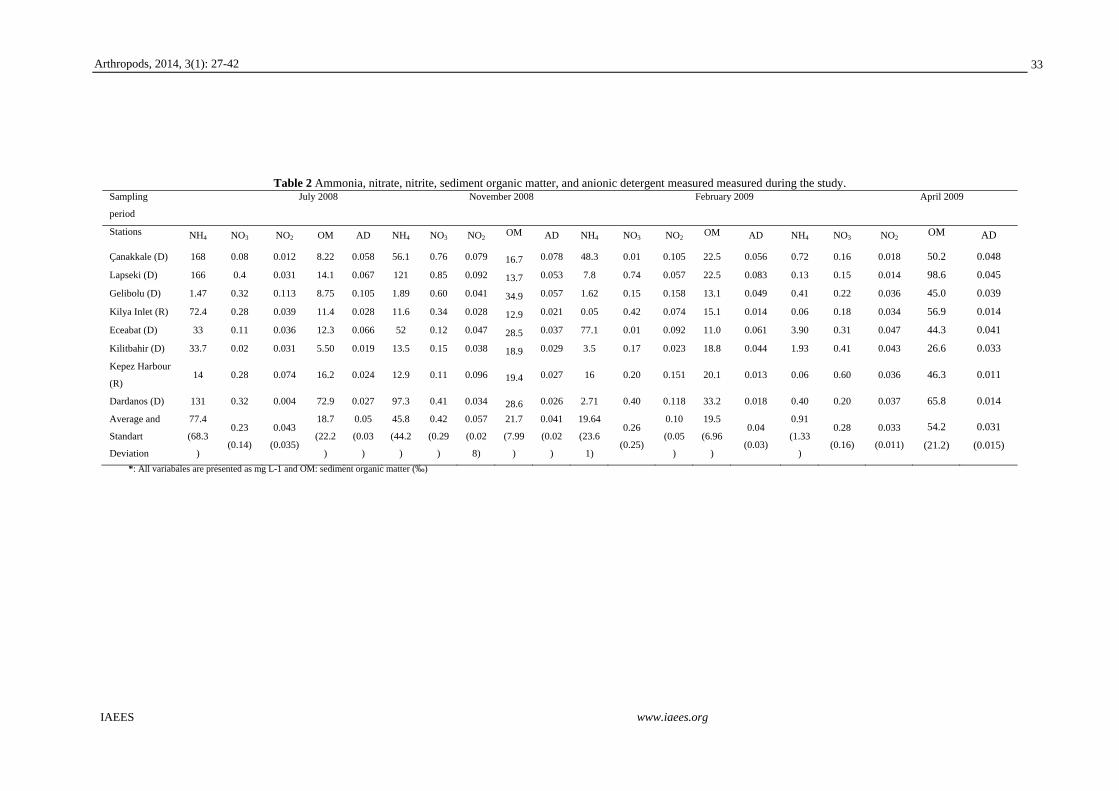
Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
Table 2 Ammonia, nitrate, nitrite, sediment organic matter, and anionic detergent measured measured during the study. Sampling
period
July 2008 November 2008 February 2009 April 2009
Stations NH4 NO3 NO2 OM AD NH4 NO3 NO2 OM AD NH4 NO3 NO2 OM AD NH4 NO3 NO2 OM AD
Çanakkale (D) 168 0.08 0.012 8.22 0.058 56.1 0.76 0.079 16.7 0.078 48.3 0.01 0.105 22.5 0.056 0.72 0.16 0.018 50.2 0.048
Lapseki (D) 166 0.4 0.031 14.1 0.067 121 0.85 0.092 13.7 0.053 7.8 0.74 0.057 22.5 0.083 0.13 0.15 0.014 98.6 0.045
Gelibolu (D) 1.47 0.32 0.113 8.75 0.105 1.89 0.60 0.041 34.9 0.057 1.62 0.15 0.158 13.1 0.049 0.41 0.22 0.036 45.0 0.039
Kilya Inlet (R) 72.4 0.28 0.039 11.4 0.028 11.6 0.34 0.028 12.9 0.021 0.05 0.42 0.074 15.1 0.014 0.06 0.18 0.034 56.9 0.014
Eceabat (D) 33 0.11 0.036 12.3 0.066 52 0.12 0.047 28.5 0.037 77.1 0.01 0.092 11.0 0.061 3.90 0.31 0.047 44.3 0.041
Kilitbahir (D) 33.7 0.02 0.031 5.50 0.019 13.5 0.15 0.038 18.9 0.029 3.5 0.17 0.023 18.8 0.044 1.93 0.41 0.043 26.6 0.033
Kepez Harbour
(R) 14 0.28 0.074 16.2 0.024 12.9 0.11 0.096 19.4 0.027 16 0.20 0.151 20.1 0.013 0.06 0.60 0.036 46.3 0.011
Dardanos (D) 131 0.32 0.004 72.9 0.027 97.3 0.41 0.034 28.6 0.026 2.71 0.40 0.118 33.2 0.018 0.40 0.20 0.037 65.8 0.014
Average and
Standart
Deviation
77.4
(68.3
)
0.23
(0.14)
0.043
(0.035)
18.7
(22.2
)
0.05
(0.03
)
45.8
(44.2
)
0.42
(0.29
)
0.057
(0.02
8)
21.7
(7.99
)
0.041
(0.02
)
19.64
(23.6
1)
0.26
(0.25)
0.10
(0.05
)
19.5
(6.96
)
0.04
(0.03)
0.91
(1.33
)
0.28
(0.16)
0.033
(0.011)
54.2
(21.2)
0.031
(0.015)
*: All variabales are presented as mg L-1 and OM: sediment organic matter (‰)
33
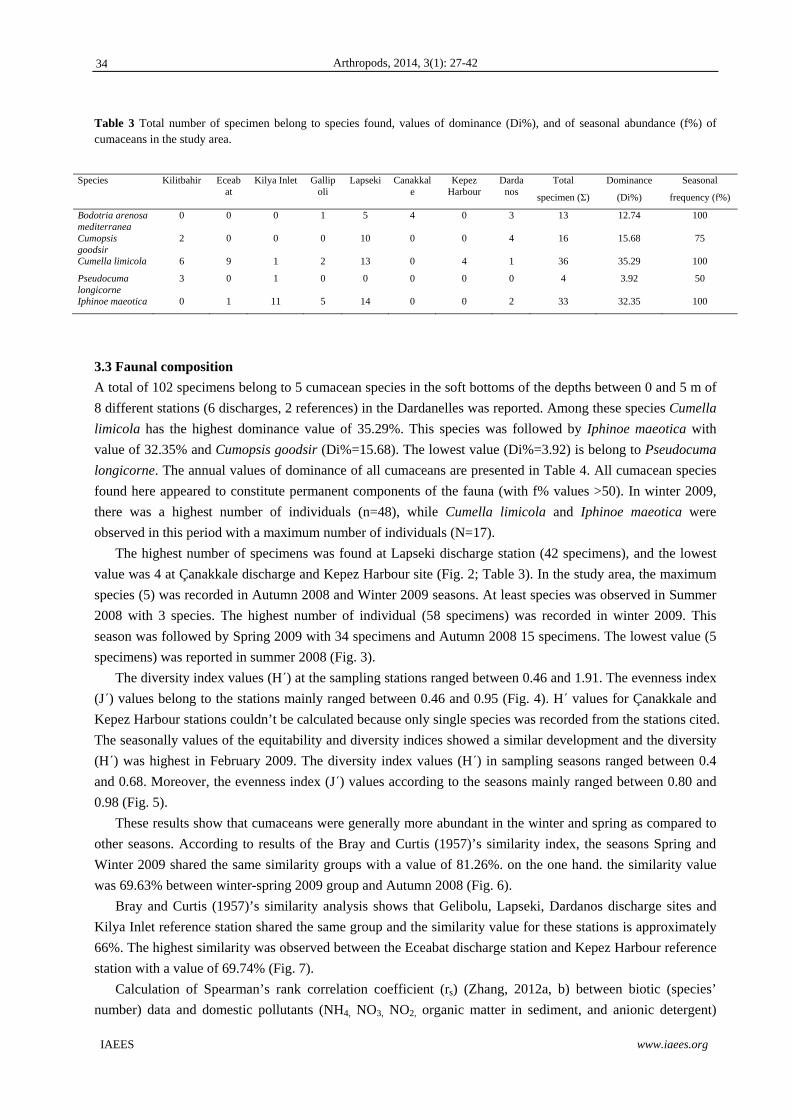
Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
Table 3 Total number of specimen belong to species found, values of dominance (Di%), and of seasonal abundance (f%) of cumaceans in the study area.
3.3 Faunal composition
A total of 102 specimens belong to 5 cumacean species in the soft bottoms of the depths between 0 and 5 m of
8 different stations (6 discharges, 2 references) in the Dardanelles was reported. Among these species Cumella
limicola has the highest dominance value of 35.29%. This species was followed by Iphinoe maeotica with
value of 32.35% and Cumopsis goodsir (Di%=15.68). The lowest value (Di%=3.92) is belong to Pseudocuma
longicorne. The annual values of dominance of all cumaceans are presented in Table 4. All cumacean species
found here appeared to constitute permanent components of the fauna (with f% values >50). In winter 2009,
there was a highest number of individuals (n=48), while Cumella limicola and Iphinoe maeotica were
observed in this period with a maximum number of individuals (N=17).
The highest number of specimens was found at Lapseki discharge station (42 specimens), and the lowest
value was 4 at Çanakkale discharge and Kepez Harbour site (Fig. 2; Table 3). In the study area, the maximum
species (5) was recorded in Autumn 2008 and Winter 2009 seasons. At least species was observed in Summer
2008 with 3 species. The highest number of individual (58 specimens) was recorded in winter 2009. This
season was followed by Spring 2009 with 34 specimens and Autumn 2008 15 specimens. The lowest value (5
specimens) was reported in summer 2008 (Fig. 3).
The diversity index values (Hˊ) at the sampling stations ranged between 0.46 and 1.91. The evenness index
(Jˊ) values belong to the stations mainly ranged between 0.46 and 0.95 (Fig. 4). Hˊ values for Çanakkale and
Kepez Harbour stations couldn’t be calculated because only single species was recorded from the stations cited.
The seasonally values of the equitability and diversity indices showed a similar development and the diversity
(Hˊ) was highest in February 2009. The diversity index values (Hˊ) in sampling seasons ranged between 0.4
and 0.68. Moreover, the evenness index (Jˊ) values according to the seasons mainly ranged between 0.80 and
0.98 (Fig. 5).
These results show that cumaceans were generally more abundant in the winter and spring as compared to
other seasons. According to results of the Bray and Curtis (1957)’s similarity index, the seasons Spring and
Winter 2009 shared the same similarity groups with a value of 81.26%. on the one hand. the similarity value
was 69.63% between winter-spring 2009 group and Autumn 2008 (Fig. 6).
Bray and Curtis (1957)’s similarity analysis shows that Gelibolu, Lapseki, Dardanos discharge sites and
Kilya Inlet reference station shared the same group and the similarity value for these stations is approximately
66%. The highest similarity was observed between the Eceabat discharge station and Kepez Harbour reference
station with a value of 69.74% (Fig. 7).
Calculation of Spearman’s rank correlation coefficient (rs) (Zhang, 2012a, b) between biotic (species’
number) data and domestic pollutants (NH4, NO3, NO2, organic matter in sediment, and anionic detergent)
Species Kilitbahir Eceabat
Kilya Inlet Gallipoli
Lapseki Canakkale
Kepez Harbour
Dardanos
Total
specimen (Σ)
Dominance
(Di%)
Seasonal
frequency (f%)
Bodotria arenosa mediterranea
0 0 0 1 5 4 0 3 13 12.74 100
Cumopsis goodsir
2 0 0 0 10 0 0 4 16 15.68 75
Cumella limicola 6 9 1 2 13 0 4 1 36 35.29 100
Pseudocuma longicorne
3 0 1 0 0 0 0 0 4 3.92 50
Iphinoe maeotica 0 1 11 5 14 0 0 2 33 32.35 100
34
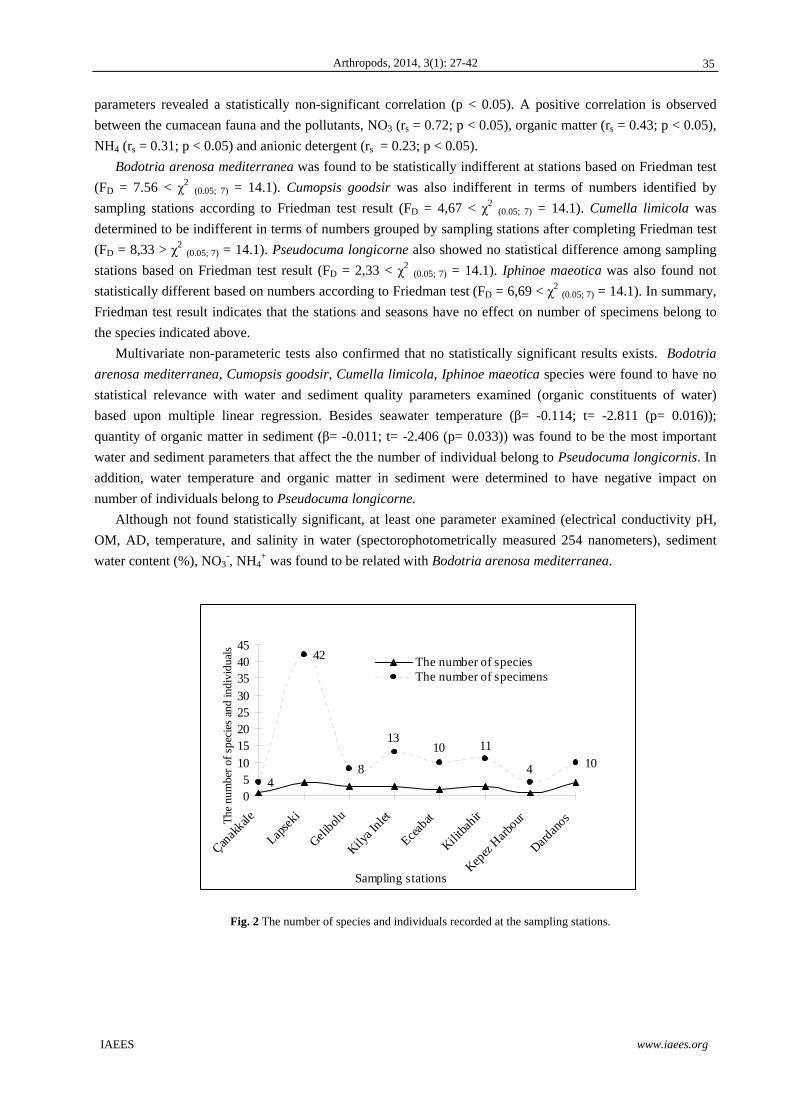
Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
parameters revealed a statistically non-significant correlation (p < 0.05). A positive correlation is observed
between the cumacean fauna and the pollutants, NO3 (rs = 0.72; p < 0.05), organic matter (rs = 0.43; p < 0.05),
NH4 (rs = 0.31; p < 0.05) and anionic detergent (rs = 0.23; p < 0.05).
Bodotria arenosa mediterranea was found to be statistically indifferent at stations based on Friedman test
(FD = 7.56 < χ2 (0.05; 7) = 14.1). Cumopsis goodsir was also indifferent in terms of numbers identified by
sampling stations according to Friedman test result (FD = 4,67 < χ2 (0.05; 7) = 14.1). Cumella limicola was
determined to be indifferent in terms of numbers grouped by sampling stations after completing Friedman test
(FD = 8,33 > χ2 (0.05; 7) = 14.1). Pseudocuma longicorne also showed no statistical difference among sampling
stations based on Friedman test result (FD = 2,33 < χ2 (0.05; 7) = 14.1). Iphinoe maeotica was also found not
statistically different based on numbers according to Friedman test (FD = 6,69 < χ2 (0.05; 7) = 14.1). In summary,
Friedman test result indicates that the stations and seasons have no effect on number of specimens belong to
the species indicated above.
Multivariate non-parameteric tests also confirmed that no statistically significant results exists. Bodotria
arenosa mediterranea, Cumopsis goodsir, Cumella limicola, Iphinoe maeotica species were found to have no
statistical relevance with water and sediment quality parameters examined (organic constituents of water)
based upon multiple linear regression. Besides seawater temperature (β= -0.114; t= -2.811 (p= 0.016));
quantity of organic matter in sediment (β= -0.011; t= -2.406 (p= 0.033)) was found to be the most important
water and sediment parameters that affect the the number of individual belong to Pseudocuma longicornis. In
addition, water temperature and organic matter in sediment were determined to have negative impact on
number of individuals belong to Pseudocuma longicorne.
Although not found statistically significant, at least one parameter examined (electrical conductivity pH,
OM, AD, temperature, and salinity in water (spectorophotometrically measured 254 nanometers), sediment
water content (%), NO3-, NH4
+ was found to be related with Bodotria arenosa mediterranea.
Fig. 2 The number of species and individuals recorded at the sampling stations.
4
42
8 104
111013
05
1015202530
354045
Çanak
kale
Lapse
ki
Gelibolu
Kilya I
nlet
Eceab
at
Kilitba
hir
Kepez
Harb
our
Dardan
os
Sampling stations
The
num
ber
of s
peci
es a
nd in
divi
dual
s
The number of speciesThe number of specimens
35

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
15
34
3
5 5 45
48
05
1015202530354045505560
Summer 2008 Autumn 2008 Winter 2009 Spring 2009
Seasons
The
num
ber o
f spe
cies
and
indi
vidu
als
The number of species
The number of specimens
Fig. 3 The number of species and individual found during the sampling seasons.
1,91
0,00 0,00
1,84
1,29
0,770,46
1,43
0,900,95
0,000,00
0,92
0,81
0,480,46
0,00
0,50
1,00
1,50
2,00
2,50
Kilitba
hir
Eceab
at
Kilya I
nlet
Gelibo
lu
Lapse
ki
Çanak
kale
Kepez
Har
bour
Dardan
os
Sampling sites
H
0,000,100,200,300,400,50
0,600,700,800,901,00
JH´ J´
Fig. 4 Values of the diversity (Hˊ), evenness (Jˊ) at the sampling points.
36
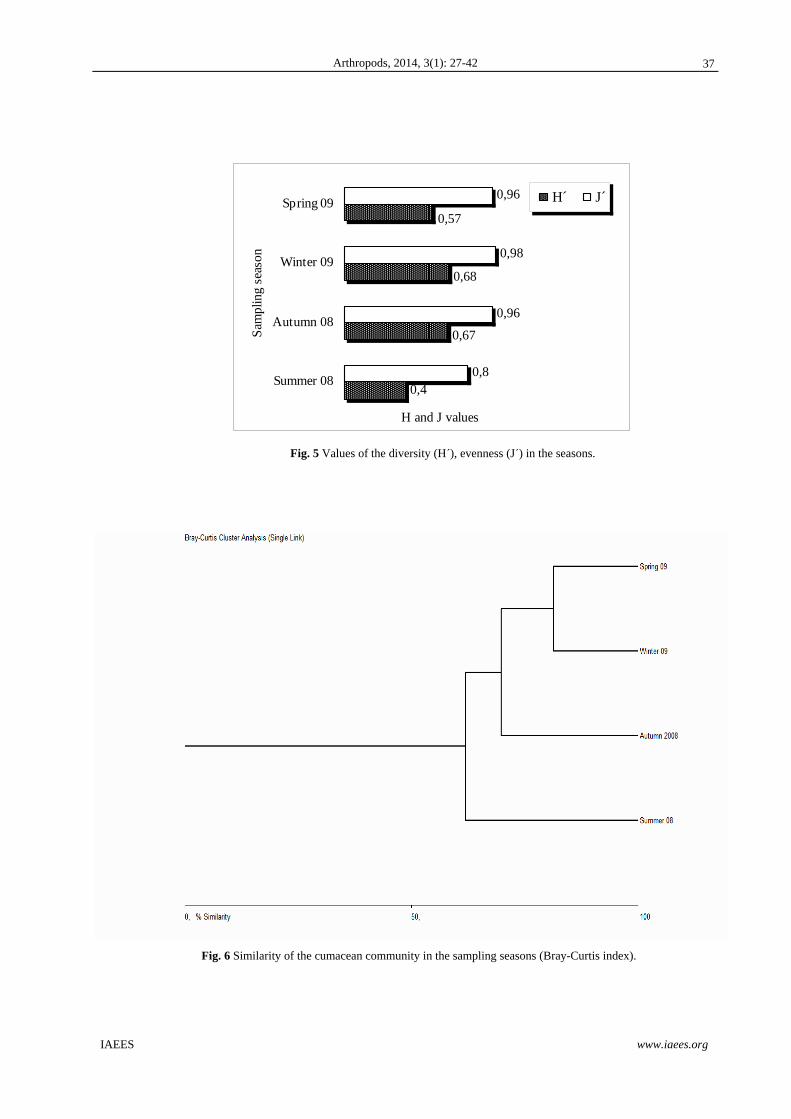
Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
0,40,8
0,96
0,98
0,96
0,68
0,67
0,57
Summer 08
Autumn 08
Winter 09
Spring 09
Sam
plin
g se
ason
H and J values
H´ J´
Fig. 5 Values of the diversity (Hˊ), evenness (Jˊ) in the seasons.
Fig. 6 Similarity of the cumacean community in the sampling seasons (Bray-Curtis index).
37

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
Fig. 7 Similarity of the cumacean community at the sampling stations (Bray-Curtis index).
4 Discussion
Benthic assemblages are widely used as indicators and for the detection of human impacts on marine
environments (Del Pilar Ruso et al., 2007). In this study, the consequences of domestic sewage discharge from
different localities of the Dardanelles, which connects the Black Sea to the Mediterranean Sea and a part of
Turkish straits system, and reference sites, which have natural back sides and have less pollution were
examined. The effects of raw sewage discharged at different communities situated on the strait on coastal
macrobenthic communities were not specifically studied. With this detailed study, the effects of sewage
disposals released from communities (namely Eceabat (population 5,498 according to 2007 census). Gallipoli
(population 31,746 according to 2007 census), Lapseki (population 10,612 according to 2007 census) and
Çanakkale (population 86,544 based on 2007 census), which is the capital of Çanakkale Province and one
municipality (Kepez population is approximately 10,000) and one village (Kilitbahir, population is
approximately 1,000)) and one natural (Kilya Inlet) were examined. In addition to these, Dardanos is a
summer resort and domestic sewage is also discharged there. Kepez is the only community that has been
served with a wastewater treatment plant since 2007. The sampling site at Kepez is also next to the harbour
and is affected by strong surface current. Effects of pollution of coastal waters (0-5 m depth) from domestic
wastewater on cumacea species were studied. As known, shallow coastal waters and estuarines are dynamic
zones because of strong waves and mixing of freshwater and salty water. This natural variation may cause
main reason of organism stress. Yet, nutrients, organic matter and pollutant entrance in such environments
might change environmental circumstances (Venturini et al., 2004).
Present literature about sewage pollution on seas underline that environmental factors (water mass, benthic
38

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
and pelagic communities sediment, trophic chain) are affected differently with diverse discharges (Echavarri-
Erasun et al., 2007). Sewage pollution can change structural integrity of biodiversity (Terlizzi Et al., 2002).
Several benthic groups such as macroalgae, tunicates, poriferas, bivalves, and crustaceans impact in where
domestic pollution is a reality. Moreover, the present populations of these organisms decline with respect to
time (Chapman et al., 1995), since domestic wastes bring nitrogen, organic carbon, and phosphorus into
aquatic systems. In other words, sewage pollution is a mixture of many types of toxic matter and its harmfully
effects the biota. Eutrophication has an important role in destruction of biodiversity in aquatic environments
(Elmanama et al., 2006).
With the effect of pollution, dominant cumacean species are reported between 5 and 13 m as Bodotroia
pulchella and Pseudocuma longicorne and Pseudocuma simile and Iphinoe armata below 20 m by Corbera
and Cardell (1995). In addition, sediment organic matter was found to be between 0.6 and 1.1% in Barcelona,
Spain (Corbera and Cardell, 1995) also reported that Iphinoe rhodaniensis has the highest biomass value and
its existence is triggered with the increase of pollution. In our study, reported cumaceans are Bodotria arenosa
mediterranea. Iphinoe maeotica, and Pseudocuma longicorne. These species were found in winter season
when sediment organic matter is 1.95±0.69%. Moreover, these species were found dominant at the depths
between 0 and 2 m. The most dominant species were found to be Cumella limicola (Di=35.29%) and Iphinoe
maeotica (Di=%32.35).
Hamouda and Abdel-Salam (2010) reported two cumaceans, Bodotria scorpioides and Iphinoe serrata on
the coasts of Abu-Qir Bay (Alexandria, Egypt) in where organic matter is observed. Iphinoe serrata was the
dominant species with its dominance value of 97.75%. The structure of cumacea community in this study are
affected more by nitrite content in seawater.
The physical environmental factors and the concentrations of pollutants in seawater are important in
determining of benthic community structure and in polluted areas the community has a low species diversity
(Morrisey et al., 2003). This status is same for Lapseki, Gallipoli, and Çanakkale discharges. While no species
was recorded for Lapseki discharge in summer sampling, the same station has 21 cumacean specimens for
spring season. Discharge points for Çanakkale and Gallipoli, while these values for summer, spring is 0 to 0
and 3 and 5 respectively. This can be explained with the increase in the number of specimens in the stations
cited owning to excessive precipitation during spring period. Due to precipitations observed during spring the
input of freshwater carried by stream constituted the pollution in Lapseki sampling point caused the dilution of
local contaminants.
The Dardanelles is rarely polluted by urbanization and industrial facilities where are located its around
because of Çanakkale has the lowest population density in Marmara Region up to date. Also, an important
amount of wastewater originated with all eastern Europe, a part of the middle Europe, and the group of
independent states are unpurifiedly moved by upper current to the Black Sea, the Bosporus, the Sea of
Marmara, and the Dardanelles. Even if the pollutant concentrations are low in the areas such as bay sheltered
(example of anionic detergent as Kilya Inlet, the average annual; 0.019±0.005 mg/L), accumulation of some
pollution is reality during no fresh water input (in summer months especially) if discharges of domestic
pollution happens (e.g. Lapseki).
Deep discharge of sewage of Çanakkale province center (in the Dardanelles undercurrent Aegean Sea to
the Marmara Sea), domestic pollution, especially the tide was low at times (summer and autumn), the inlets
and small bays in areas where pollutant accumulation is possible. Wastewater is less dense than sea water
(have lower specific gravity), but without any particular stream impacts directly related to the horizontal
surface layer can be transported away from the discharge point. Denitrification via nitrite and nitrate in water
and nitrous oxide are converted (Sawyer et al., 2003).
39

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
Lapseki discharge points throughout the year than other stations in the NH4+ and NO3
- the reason why the
highest value of the accumulated sediments in the long term can be connected to the organic debris. The most
important element of livings on the basis of only the nitrogen and phosphorus in terms of nitrate nitrogen,
which is a derivative of the Dardanelles, nor seasonal sampling locations differ in terms of what can be said.
The organic matter content of water in the Black Sea and the Marmara and Aegean probably varies with the
convection dynamics. North-east wind is generally strong, mistral wind and south due to the strong currents of
change in systems locality has led to removal of pollutants from the study area. Based on our observations we
can say that, especially in summer time when parts of the residential area of the strait at night discharges are
intense. The impact of these discharges under the influence of wind and currents in the Strait will be gone in
the morning as possible, but it was a part of sunlight during the discharge of pollution that cannot be intense.
Canonical Discriminant Analysis showed that anionic detergent concentration is the main factor among all
physical and chemical factors examined in the study. Wilks’ Lambda was found to be 4.981 (p=0.018) for
anionic detergent. Discriminant function analysis showed that physical and chemical constituents measured in
this study differ notably in winter compared to other seasons. It is hard to tell, however, that whether
temperature or fresh water inputs to the system or both together, equally, cause differences among the
parameters examined. It could be concluded that change in some physical and chemical constituents differ due
to the fact that summer resorts, seasonal runoff difference, although not examined, and most importantly
temperature in the study area. With the expected population change, increase in some physical and chemical
water quality parameters will probably be experienced in the future if pollution control measures would not be
taken beforehand.
However, study in discharge points in recorded number of species than the expected due to the reason that
the pollutant factors of sediment in water and strong currents and wind action because of its intensity
negatively affecting absence is considered.
Acknowledgements
This study was supported by the Scientific and Technological Research Council of Turkey CAYDAG
107Y332 code project.
References
Albayrak S, Balkıs H, Zenetos A, et al. 2006. Ecological quality status of coastal benthic ecosystems in the Sea
of Marmara. Marine Pollution Bulletin, 52: 790-799
Apha Awwa 2005. 21st Edition of Standard Methods for the Examination of Water and Wastewater (Eaton A
D, Clesceri, LS, Rice EW, Greenberg AE, eds). American Public Health Association, Washington, USA
Arasaki E, Muniz P, Pires-Vanin AMS. 2004. A Functional Analysis of the Benthic Macrofauna of the São
Sebastião Channel (Southeastern Brazil). Marine Ecology, 25 (4): 249-263
Bat L, Akbulut M, Sezgin M, Çulha M. 2001. Effects of sewage pollution on the structure of the community of
Ulva lactuca, Enteremorpha linza and rocky macrofauna in Dışliman of Sinop. Turkish Journal of Biology,
25: 93-102
Bellan G. 1967. Pollution et peuplements benthiques sur substrat meuble dans la région de Marseille Première
partie. Rev. Int. d’Océa. Méd, 6-7: 53-87
Bellan-Santini D. 1969. Etudefloristuque et faunistuque de quelquespeuplements infralittoraux de substrat
rocheus. Recueil des Travaux de la Station Marine d'Endoume, 41: 237-298
Botkin DB, Keller EA. 2003. Environmental Science: Earth as a Living Planet (4th Edition). John Wiley and
40

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
Sons, Inc, Hoboken, NJ, USA
Bray JR, Curtis JT. 1957. An ordination of the upland forest communities of South Wisconsin. Ecological
Monographs, 27: 325-347
Chapman MG, Underwood AJ, Skilleter GA. 1995. Variability at different spatial scales between a subtidal
assemblage exposed to the discharge of sewage and two control assemblages. Journal of Experimental
Marine Biology and Ecology, 189(1-2): 103-122
Corbera J, Cardell MJ. 1995. Cumaceans as indicators of eutrophication on soft bottoms. Scientia Marina, 59
(1): 63-69
Craft CB, Seneca ED, Broome SW. 1991. Loss on ignition and Kjeldahl digestion for estimating organic
carbon and total nitrogen in estuarine marsh soils: calibration with dry combustion. Estuaries, 14: 175-179
Del Pilar Ruso Y, De La Ossa Carretero JA, Giménez Casalduero F, et al. 2007. Spatial and temporal changes
in infaunal communities inhabiting soft-bottoms affected by brine discharge. Marine Environmental
Research, 64: 492-503
Del Vals TA, Conradi M, Garcia-Adiego E, et al. 1998. Analysis of macrobenthic community structure in
relation to different environmental sources of contamination in two littoral ecosystems from the Gulf of
Cádiz (SW Spain). Hydrobiologia, 385: 59-70
Echavarri-Erasun B, Juanes JA, García-Castrillo G, et al. 2007. Medium-term responses of rocky bottoms to
sewage discharges from a deepwater outfall in the NE Atlantic. Marine Pollution Bulletin, 54: 941-954
Elmanama AA, Afifi S, Bahr S. 2006. Seasonal and spatial variation in the monitoring parameters of Gaza
Beach during 2002–2003. Environmental Research, 101: 25-33
García-Raso JE, Manjón-Cabeza ME. 2002. An infralittoral decapod crustacean community of southern Spain
affected by anthropogenic disturbances. Journal of Crustacean Biology, 22: 83-90
Guerra-García JM, García-Gómez JC. 2004. Crustacean assemblages and sediment pollution in an exceptional
case study: A harbour with two opposing entrances. Crustaceana, 77(3): 353-370
Hamouda AZ, Abdel-Salam KM. 2010. Acoustic seabed classification of marine habitats: Studies in the Abu-
Qir Bay. Journal of Oceanography Marine Sciences, 1 (1): 11-22
Kocataş A, Ergen Z, Katağan T, et al. 1988. Evolution a long terme (1974-1987) des peuplements benthiques
sur substrat meuble du Golfe d' İzmir scumis a des multiples pollutions.- Rapp. Comm. Int. Mer. Medit., 31:
2
Ledoyer M. 1965. Sur Quelques Especes Nouvelles D’Iphinoe (Crustacea Cumacea) Discussion et Description
Comparative des Especes Europeennes Deja Connues. Recueil des Travaux de la Station Marine
d'Endoume, 39(55): 253-295
Metcalf, Eddy Inc. 2003. Wastewater Engineering: Treatment and Reuse (revised by Tchobanoglous G,
Burton FL, Stensel HD), McGraw-Hill, Boston, USA
Morrisey DJ, Turner SJ, Mills GN, et al. 2003. Factors affecting the distribution of benthic macrofauna in
estuaries contaminated by urban runoff. Marine Environmental Research, 55: 113-136
Okuş E, Uysal A, Yüksek A, et al. 1996. Biological aspects of the rehabilitation of Golden Horn”. İstanbul
Water and Sewage Administration. Istanbul University, Institute of Marine Sciences and Management,
Vefa, Istanbul (in Turkish)
Rinderhagen M, Ritterhoff, J, Zauke GP. 2000. Crustaceans as bioindicator. In: Biomonitoring of Polluted
Water-Reviews on Actual Topics. Trans Tech Publications - Scitech Publications (Gerhardt A, ed).
Environmental Research Forum 2000 9, Uetikon-Zuerich, 161-194
Sawyer CN, McCarty PL, Parkin GF. 2003. Chemistry for Environmental Engineering and Science (5th
edition). McGraw-Hill, Boston, USA
41

Arthropods, 2014, 3(1): 27-42
IAEES www.iaees.org
Shannon CE, Weaver V. 1949. A mathematical theory of communication. 101-117, Princeton University Press
Illinois, USA
Soyer J. 1970. Contribution a l'etude des copepodes harpactioides de Mediterranee occidentale”, 2. Tachidiidae
Sars, Lang. Vie Milieu, 21 (2-A): 261-278
Terlizzi A, Fraschetti S, Guidetti, P, et al. 2002. The effects of sewage discharge on shallow hard substrate
sessile assemblages. Marine Pollution Bulletin, 4: 544-550
Tuğrul İçemer G, Koşun E. 2003. The effects of Sewage on Benthic Community in Antalya Bay, Turkey. The
Sixth International Conference on The Mediterranean Coastal Environment (MEDCOAST 03). 753-758,
Ravenna, Italy
Turkish Statistical Institute (TSI). 2010. Belediye Atıksu İstatistikleri Veri Tabanı.- data set of municipal
wastewater statistics, retrieved from the internet from
http://tuikrapor.tuik.gov.tr/reports/rwservlet?cevredb2=&report=CEVAT13.RDF&p_pkod=0&p_yil1=200
6&desformat=html&p_dil=1&ENVID=cevredb2Env, 2006. Accessed on April 20 2010
Venturini N, Muniz P, Rodríguez M. 2004. Macrobenthic subtidal communities in relation to sediment
pollution: the phylum-level meta-analysis approach in a south-eastern coastal region of South America.
Marine Biology, 144: 119-126
Zhang WJ. 2012a. Computational Ecology: Graphs, Networks and Agent-based Modeling. World Scientific,
Singapore
Zhang WJ. 2012b. How to construct the statistic network? An association network of herbaceous plants
constructed from field sampling. Network Biology, 2(2): 57-68
42

Arthropods, 2014, 3(1): 43-47
IAEES www.iaees.org
Article
Redescription and new distributional records of Matuta planipes
(Fabricius, 1798) (Crustacea; Decapoda; Matutidae) from Chennai
Coast, Tamil Nadu K. Silambarasan, K. Velmurugan, E. Rajalakshmi, A. Anithajoice
P.G. and Research Department of Zoology, Sir Theagaraya College, Chennai-600021, India
E-mail: [email protected]
Received 18 November 2013; Accepted 25 December 2013; Published online 1 March 2014
Abstract
Matuta planipes is reported for the first occurrence from Chennai coast, Tamil Nadu. Four female and two
male specimens was caught in trawl net near Kasimedu fish landing center, on September 2013. The
morphological characters of Matuta planipes, is having on surface regions of male chela a single spine, frontal
lobes and carapace covered with reticulated loops as compared with bispinose chela, rounded lobes and
minutely spotted carapace of the latter, these characters mostly differs from Matuta victor. The specimen has
been compared with the earlier reports and other similar species.
Keywords Matuta planipes; trawl net; Kasimedu; Chennai coast.
1 Introduction
Crabs constitute an important crustacean resource in trawl catches and they also from an important ecological
entity in their habitat. Although the commercial fishery supported by species belonging to the family
Portunidae, there are numerous brachyuran crab species from other families which are also caught accidentally
in trawl nets and landed as low value by catch. Brachyuran crabs comprise about 700 genera and 5000 to
10,000 species worldwide (Ng, 1998; Ng et al., 2008). Crabs of the family Calappidae and Matutidae known as
box crabs, shamefaced or moon crabs are one of the most fascinating crabs in the tropical and subtropical seas
of the world ocean (Galil and Clark, 1995).The moon crabs belonging to family Calapidae, it was containing
16 genera, of which 7 in fossils. The Callapidae crabs have been studied from Indian waters was represented
by (Dana, 1852). In India 705 brachyuran crab species, 28 families, 270 genera heave been reported
(Venkataraman and Wafar, 2005). A total of 991 species of brachyuran crabs have been recorded from the
Indian waters (Lakshmi Pillai and Thirumullu, 2008). Tamil Nadu coast one of the states in India has 404
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2014, 3(1): 43-47
IAEES www.iaees.org
species of crabs belonging to 26 families and 152 genera were recorded by Kathirvel (2008). Though relatively
more taxonomic works on the brachyuran crabs have been done on the fauna of Indian regions, our
information on the fauna of Indian sub-continent is very scanty. Among the species reported from India many
need detailed taxonomic revision and redescription. One such species is Matuta planipes first described by
Fabricius 1798. In the present study, this species is redescribed and newly recorded from the Kasimedu,
Chennai coast, Tamil Nadu.
2 Materials and Methods
During the survey of brachyuran fauna of Matuta planipes (Fabricius, 1798) has four adult female and two
male species were collected in Chennai coast using trawl net. Trawl net hauled from depth of 40 m to 60 m.
The specimen was preserved in 10% formalin and its taxonomy was confirmed to species level using various
literatures (Sethuramalingam and Ajmalkhan, 1991; Jayabaskaran et al., 2000). The specimen was deposited in
the Department of Zoology, Sir Theagaraya College, Chennai, Tamil Nadu, India.
3 Results
This species is available in various parts of the world. In earlier periods Alcock, 1896 was recorded in Indian
water, after a long period recorded in this species at Kasimedu, Bay of Bengal, Chennai Coast. So, this species
consider as the redescription and new distributional records of Chennai coast (Fig .1 a & b). The systematic
position and diagnostic features, coloration and distribution are as follows:
Systematic Position
Phylum - Arthropoda
Class - Crustacea
Order - Decapoda
Sub order - Pleocymata
Infra order - heterotremata
Super family - Leucosioidea
Family - Matutidae
Genus - Matuta
Species - planipes
(a). Matuta planipes – dorsal view
44

Arthropods, 2014, 3(1): 43-47
IAEES www.iaees.org
(b). Matuta planipes- ventral view
Fig. 1 a & b Matuta planipes.
Diagnostic features
Surface of carapace minutely granulate, small tubercles clustering around six dorsal tubercles, largest cluster
surrounding mesogastric tubercle, front with straight horizontal lobes laterally and slightly emarginated
rostrum medially. Ischium of third maxilliped tuberculate. Anterolateral margins nearly uniformly crenulate,
tubercles somewhat larger posteriorly. Lateral spine 0.2 carapace width. Postero lateral margin oblique, with
granulate carina extending to base of lateral spine. Carpus of cheliped with two obtuse tubercles on outer
surface, its upper margin carinate, granulate, internal anterior angle produced. Upper margin of palm cut in to
three teeth, proximal tooth tuberculate. Upper external surface with two rows of granulate low tubercles,
proximal most in lower row largest. Mid palm, in male, rounded ridges extending to tip of lower finger,
proximally with granulate tubercle followed by a prominent, acuminate spine. Mid palm, in female, five
tubercles, second tubercle spine like. At lower proximal angle of palm a small granulate tubercle. Lower
margins with row of tubercles terminating at base of dactylus. In female, an additional row of obtuse granules
parallel to lower margin. Distinctly milled ridge on outer surface of dactylus in male, absent in female.
Plastron finely granular.
Colouration
Carapace with reticulating brown lines forming small rings anteriorly and larger, elongate loops posteriorly.
Type Locality
Bay of Bengal (Chennai Coast).
Distribution
Indian Ocean (Fabricius, 1798, Galil and Clark, 1995); Persian Gulf (Stephensen, 1945); Strait of Hormuz
45

Arthropods, 2014, 3(1): 43-47
IAEES www.iaees.org
(Stephensen, 1945); Pakistan- Karachi (Alocock, 1896, Tirmizi and Kazmi, 1991); India – Sundarbans and
Gangetic Delta, Mahanadhi Delta, Madras and Bombay (Alocock, 1896); Srilanka (Galil and Clark, 1995);
Burma (Alocock, 1896);and near Tavoy (Chopra and Das, 1937); Distribution map for Matuta planipes was
given in Fig. 2.
Fig. 2 Distribution map for Matuta planipes (courtesy: Ocean Biogeographic Information System).
4 Discussion
The present record matches well with the earlier descriptions and illustrations of Matuta planipes was reported
by Alocock (1896) from Indian Ocean. Dana (1852) was reported 16 genera of Calappidae family from Indian
waters. Sankarankutty (1965) were reported the species from Andaman and Nicobar islands. Ng et al (2008)
has been reported the species from Singapore coastal waters. Meher Fatima (2003) has recorded two species of
Matutidae family from Karachi in Pakistan. Recently Varadharajan et al (2012) have reported the species from
Pondicherry coast, South east coast of India. Although, Matuta planipes and Matuta victor are very similar
morphologically certain distinctive characters have been described in having a single spine on external surface
of male chela, straight lateral frontal lobes (Galil and Mendelson, 2013). Hence, this species report is an
extended distribution and a new record from the Bay of Bengal at Indian water.
References
Alcock A. 1896. Materials for a carcinological fauna of India. No.2. the Brachyura oxystoma. Journal of
Asiatic society of Bengal, 65(2): 134-296
Chopra BN, Das KN. 1937. Further notes on crustacea decapoda in the Indian museum. IX. On three
collections of crabs from Tavoy and Margay archipelago. Record of the Indian Museum, Calcutta, 39(4):
377-434
Dana JD. 1852. Conspectus Crusacerorum Quae in Orbis Terrarum circumnavigation, Carole Wilkes e classe
Reipublicae Foederatae e Duce, and lexit et descripts. Proceedings of the Academy of National Sciences of
Philadelphia, 10- 28, USA
46

Arthropods, 2014, 3(1): 43-47
IAEES www.iaees.org
Galil BS, Clark PF. 1995. A Revision of the genus Matuta weber, 1795 (Crustacea: Brachyura: Calappidae).
Zoologische Verhandelingen, Leiden, 294: 1-55
Galil BS, Mendelson M. 2013. A record of the moon crab Matuta victor from the Mediterranean of Israel. Bio
Invasion Records, 2(1): 69-71
Jeyabaskaran R, Ajmalkhan S, Ramaiyan V. 2000. Biodiversity project on Gulf of Mannar biosphere reserve.
Parangipettai: Centre of Advanced Study of Marine Biology, Annamalai University, India
Kathirvel M. 2008. Biodiversity of Indian marine brachyuran crabs. Rajivghandhi Chair Special Publications,
7: 67-78
Lakshmipillai S, Thirumillu P. 2008. New records of Brachyuran crabs from the Chennai coast. Journal of
Marine Biological Association of India, 50(2): 238-240
MeherFathima. 2003. Length weight study of two species of crabs Matuta planipes and Matuta lunaris from
Karachi, Pakistan. Pakistan Journal of Biological Sciences, 6(4): 397-398
Ng PKL. 1998. Crabs. In: FAO Special Guide for Fishery Purposes. The Living Marine Resources of The
Western Central Pacific. Volume 2: Cephalopods, Crustaceans, Holothurians and Sharks (Carpenter KE,
Niem VH, eds). 1045-1155, Food and Agricultural Organization, Rome, Italy
Ng PKL, Guinot D, Davies PJF. 2008. Systema brachyuran. Part- I. An annotated checklist of extent
brachyuran crabs of the world. Raffles Bulletin Zoology, 17: 1-286
Sankarankutty C. 1965. On Decapoda brachyuran from the Gulf of Mannar and Palks bay. Proceedings
symposium of crustaceans. Marine Biological Association of India, I: 347-362
Sethuramalingam SA, Ajmalkhan S. 1991. Brachyuran Crabs of Parangipettai. Tamil Nadu: Centre of
Advanced Study in Marine Biology, Annamalai University, India
Stephensen K, 1945. The brachyuran of the Iranian Gulf with an appendix: The male pleopod of the Brachyura.
In: Danish Scientific Investigations in Iran, Part-4. 57-237, Copenhagen, Denmark
Tirmizi NM, Kazmi QB. 1991. Marine fauna of Pakistan: crustacean: Brachyura (Dromiacea,
Archaeobrachyura, Oxystomata, Oxyrhyncha). University of Karachi BCCI (Bank Credit Common Int)
Foundation Chair Publication, 1: 1-244
Varadharajan D, Pushparajan N, Soundarapandian P. 2012. New record of portunid crabs from Pondicherry
coast, south east coast of India. International Journal of Pharmaceutical & Biological Archives, 3(5): 1255-
1257
Venkataramana K, Waffer M. 2005. Coastal and marine biodiversity of India. Indian Journal of Marine
Sciences, 34(1): 57-75
47

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
Article
Checklist of the subfamilies Mirinae and Orthotylinae (Hemiptera:
Heteroptera: Miridae) in western parts of Kerman Province, Iran Mohsen Shamsi1, Reza Hosseini1, Asghar Shirvani2
1Department of Plant Protection, Faculty of Agricultural Sciences, University of Guilan, Rasht, Iran 2Department of Plant Protection, Faculty of Agriculture, Kerman University, Kerman, Iran
E-mail: [email protected]
Received 1 September 2013; Accepted 5 October 2013; Published online 1 March 2014
Abstract
A faunal study was carried out on the subfamilies Mirinae and Orthotylinae (Heteroptera: Miridae) from
different parts of western Kerman Province on various host plants. In total 16 species belonging to 14 genera
were collected and identified from different host plants and localities.
Keywords Fauna; Miridae; plant bugs; Kerman Province; Iran.
1 Introduction
The Plant bugs (Hemiptera: Miridae) are the most populated family of Hemiptera order, with approximately
11,020 described species (Cassis and Schuh, 2012). Size variation in Mirid bugs is from 1 to 15 mm. In term
of food behavior, Plant bugs are Phytophagous, Carnivorous, and Omnivorous. This family comprising eight
subfamilies which among them subfamilies Mirinae and Orthotylinae are the most diverse. The Mirinae is the
largest subfamily of Miridae with 6 tribe and more than 4000 described species (Cassis & Schuh, 2012). This
subfamily is defined by pretarsal and genitalic characters (Schwartz, 2008). Orthotylinae is another subfamily
that is comprised with six recognized tribes and more than 2000 described species (Schuh, 2002-2013). The
outstanding features of this subfamily that can be noted, are the greatly enlarged male parameres and
exaggerated endosomal spicules (Asquith, 1994).The existing species of these two subfamily have a wide
range of hosts including those which are plant feeder or predacious (Slater & Baranowski, 1978). Many of
plant feeder in Mirinae subfamily like Lygus Hahn and Adelphocoris Reuter directly damage the organs of
plants by feeding from plants sap and also indirectly damages them by potentially transferring plant pathogens.
Also species in Phytocoris Fallen genus are predaceous that nymphs and adults prey on mites, mite eggs,
aphids, and other small arthropods. In Orthotylinae subfamily species Ceratocapsus Reuter genus are
predaceous that nymphs and adults prey on mites and aphids (Braimah et al., 1982).
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
Recently much attention has been focused on Iranian Plant bugs fauna. There are numbers of publications have
been published on Iranian mirid bugs in different regions (Hosseini, 1997; Hosseini and Linnavuori, 2000;
Hosseini et al., 2000, 2002a, b; Hosseini, 2013a, b, c; Hosseini, 2014; Linnavuori and Hosseini, 1998, 1999,
2000; Lashkari and Hosseini, 2012; Yarmand et al., 2004; Linnavuori, 2006, 2007, 2009; Mirab-Baloo, 2008;
Arkani; 2009, Ebrahimi et al, 2012). As a part of an extensive research on Plant bugs in Kerman, the aim of
this study was to collect and identify the species of the subfamilies Mirinae and Orthotylinae in Kerman
Province.
The Kerman Province, geographically is located on the south of Iran, neighbors with Khorasan and Yazd
Province in the north, Hormozgan Province in the South, Sistan-Baluchistan Province in the East and Fars
province in the West. Kerman is one of the Iran's largest provinces and occupied about 11% of this country.
The precipitation rate in this province is about 120 mm per year. Kerman province has a great variety of
climates. Extreme differences in elevation, latitude, and located in the vicinity of the world's driest deserts are
the reasons for this diversity (Society and Department of Geography Kerman Province, 2012).
Fig. 1 Map of Kerman province, its adjacent provinces and position in Iran. where shows the position of three cities of Baft, Sirjan and Bardsir among other cities of Kerman.
2 Materials and Methods
The research was conducted in the west of Kerman Province from different locations by collecting adult mirids
during summer 2012. Sampling was conducted in three cities in the western regions of the province including;
Baft (2283m, 29˚13'59''N, 56˚36'08''E) that is located on the southern east of province, neighbors with Jiroft in
49

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
the east, Sirjan in the west, Bardsir in the north and Hormozgan Province in the south). Sirjan (1746m,
29˚27'07''N, 55˚40'52''E) that is neighbor with Baft from the east, Shahrebabak from the north, Fars province
from the west and Hormozgan Province from the south) and Bardsir (2046m, 29˚55'39''N, 56˚34'19''E) that is
adjacent to Baft from the south, Kerman from the east and north, and Sirjan from the west (Fig. 1). Specimens
were collected by sweep net and light trap. The sweeping method was used to collect living bugs on flowering
plants, shrubs, and foliage of trees. A sweep net (45cm diameter and 75cm length) was used for sweeping in
vegetation, and bushnet was used for tree foliages. The collected specimens were killed promptly in a small
tube containing Ethyl acetate, then they were transferred to the laboratory and were prepared for identification
under stereomicroscope (Olympus SZX 12). The genitalia was separated from males and mounted on slide by
using glycerin. Identification was done by relevant taxonomic keys (Wagner and Weber, 1964; Wagner, 1971;
1973) and compared with type species available in the Natural History Museum of University of Guilan. All
the specimens were deposited in the department of Plant Protection, Faculty of Agriculture, University of
Guilan, Rasht, Iran.
3 Results
In this study, a total of 60 specimens belonging to16 species and 14 genera were collected. The collected
species are as follow:
Subfamily: Mirinae
Stenodema turanica Reuter, 1904
Material examined
Iran, Kerman, Baft: Torang, August 2012, (2145m, 28˚45'21''N, 56˚48'52''E), Medicago sativa (Fabaceae), (M.
Shamsi), Gogher, July 2012, (2625m, 29˚28'42''N, 56˚24'37''E), (Weed), (M. Shamsi), Bardsir: Bardsir,
September 2012, (2046m, 29˚55'39''N, 56˚34'19''E), (Weeds), (M. Shamsi), Negar, August 2012, (2094m,
29˚51'34''N, 56˚47'57''E), Glycyrrhiza sp. (Fabaceae), (M. Shamsi).
Comments
Europe, Asia (Aukema & Rieger, 1999), Irano-Turanian (Linnavuori, 2009).
Lygus gemellatus (Herrich-Schaeffer, 1835)
Material examined
Iran, Kerman, Baft: Torang, Agust 2012, (2145m, 28˚45'21''N, 56˚48'52''E), Teriticum sativum (Poaceae), (M.
Shamsi), Bezenjan, August 2012, (2358m, 29˚14'48''N, 56˚41'50''E), (M. Shamsi), Kiskan, August 2012,
(2611m, 29˚22'46''N, 56˚38'14''E), Medicago sativa (Fabaceae), (M. Shamsi), Rabor, August 2012, (2330m,
29˚17'29''N, 56˚54'45''E), (Weed), (M. Shamsi), Gogher, August 2012, (2625m, 29˚28'42''N, 56˚24'37''E),
Teriticum sativum (Poaceae), (M. Shamsi), Baft, August 2012, (2283m, 29˚13'59''N, 56˚36'08''E), M. sativa
(Fabaceae), (M. Shamsi), khabr, August 2012, (2140m 28˚48'59''N, 56˚20'49''E), T. sativum (Poaceae), (M.
Shamsi), Sirjan: Zeydabad, July 2012, (1726m, 29˚36'55''N, 55˚32'12''E), Trifolium sp. (Leguminosae), (M.
Shamsi), Bardsir: Bardsir, August 2012, (2046m, 29˚55'39''N, 56˚34'19''E), M. sativa (Fabaceae), (M. Shamsi),
Negar, September 2012, (2094m, 29˚51'34''N, 56˚47'57''E), T. sativum (Poaceae), (M. Shamsi).
Comments
Holopalaearctic (Linnavuori 2007), Europe, Asia , North Africa, North India, Nepal, Pakistan (Aukema &
Rieger, 1999).
Lygus pratensis (Linnaeus, 1758)
50

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
Material examined
Iran, Kerman, Baft:Torang, August 2012, (2145m, 28˚45'21''N, 56˚48'52''E), Medicago sativa (Fabaceae), (M.
Shamsi), Dashtab, July 2012, (2026m, 29˚59'45''N, 56˚38'12''E), M. sativa (Fabaceae), (M. Shamsi), Bezenjan,
September 2012, (2358m, 29˚14'48''N, 56˚41'50''E), M. sativa (Fabaceae), (M. Shamsi), Kiskan, August 2012,
(2611m, 29˚22'46''N, 56˚38'14''E), Trifolium resupinatum (Fabaceae), (M. Shamsi), Rabor, July 2012, (2330m,
29˚17'29''N, 56˚54'45''E), T. resupinatum (Fabaceae), (M. Shamsi), Orzooiieh, July 2012, (1169m, 28˚22'42''N,
56˚29'10''E), M. sativa (Fabaceae), (M. Shamsi), Sirjan: Sirjan, August 2012, (1746m, 29˚27'07''N,
55˚40'52''E), T. resupinatum (Fabaceae), (M. Shamsi), Bardsir: Negar, July 2012, (2094m, 29˚51'34''N,
56˚47'57''E), (M. Shamsi), T. resupinatum (Fabaceae), Lalezar, July 2012, (2844m, 29˚31'11''N, 56˚49'09''E),
M. sativa (Fabaceae), (M. Shamsi), Ghaleaskar, September 2012, (2659m, 29˚30'44''N, 56˚41'07''E), M. sativa
(Fabaceae), (M. Shamsi).
Comments
Holopalaearctic (Linnavuori 2007).
Eurystylus bellevoyei (Reuter, 1879)
Material examined
Iran, Kerman, Baft: Baft, July 2012, (2283m, 29˚13'59''N, 56˚36'08''E), Pirus malus (Rosaceae), (M. Shamsi),
Dashtab, September 2012, (2026m, 29˚59'45''N, 56˚38'12''E), (Weed), (M. Shamsi), Kiskan, September 2012,
(2611m, 29˚22'46''N, 56˚38'14''E), Medicago sativa (Fabaceae), (M. Shamsi), Rabor, September 2012, (2330m,
29˚17'29''N, 56˚54'45''E), Onobrychis sativa (Leguminosae), (M. Shamsi), Orzooiieh, July 2012, (1169m,
28˚22'42''N, 56˚29'10''E), Solanum tuberosum (Solanaceae), (M. Shamsi), August 2012, (2844m, 29˚31'11''N,
56˚49'09''E), M. sativa (Fabaceae), (M. Shamsi), Gogher, August 2012, (2625m, 29˚28'42''N, 56˚24'37''E), M.
sativa (Fabaceae), (M. Shamsi), Sirjan: Sirjan, August 2012, (1746m, 29˚27'07''N, 55˚40'52''E), M. sativa
(Fabaceae), (M. Shamsi), Zeydabad, July 2012, (1726m, 29˚36'55''N, 55˚32'12''E), (Weeds), (M. Shamsi),
Nosratabad, September 2012, (1724m, 29˚30'31''N, 55˚35'46''E), (Weed), (M. Shamsi), Bardsir: Bardsir,
August 2012, (2046m, 29˚55'39''N, 56˚34'19''E), S. tuberosum (Solanaceae), (M. Shamsi), Lalezar, July 2012,
(2844m, 29˚31'11''N, 56˚49'09''E), (Weed), (M. Shamsi), Ghaleaskar, August 2012, (2659m, 29˚30'44''N,
56˚41'07''E), Pirus malus (Rosaceae),(M. Shamsi), Negar, September 2012, (2094m, 29˚51'34''N, 56˚47'57''E),
(M. Shamsi),
Comments
Eremian with a wide distributional range in the Holomediterranean and sudanes subregions (Linnavuori, 2009),
Europe, Asia, Afrotropical regions and Orietal (Aukema & Rieger, 1999).
Adelphocoris lineolatus (Goeze, 1778)
Material examined
Iran, Kerman, Baft: Baft, July 2012, (2283m, 29˚13'59''N, 56˚36'08''E), Medicago sativa (Fabaceae), (M.
Shamsi), Dashtab, July 2012, (2026m, 29˚59'45''N, 56˚38'12''E), M. sativa (Fabaceae), (M. Shamsi), Gogher,
August 2012, (2625m, 29˚28'42''N, 56˚24'37''E), Glycyrrhiza sp. (Fabaceae), (M. Shamsi), Kiskan, August
2012, (2611m, 29˚22'46''N, 56˚38'14''E), M. sativa (Fabaceae), (M. Shamsi), Sirjan: Nosratabad, September
2012, (1724m, 29˚30'31''N, 55˚35'46''E), Glycyrrhiza sp. (Fabaceae), (M. Shamsi), Mahmoodabad, September
2012, (1733m, 29˚31'40''N, 55˚36'23''E), M. sativa (Fabaceae), (M. Shamsi), Sirjan, September 2012, (1746m,
29˚27'07''N, 55˚40'52''E), Onobrychis sativa (Leguminosae), (M. Shamsi), Bardsir: Ghaleaskar, September
2012, (2659m, 29˚30'44''N, 56˚41'07''E), M. sativa (Fabaceae), (M. Shamsi), Negar, August 2012, (2094m,
29˚51'34''N, 56˚47'57''E), M. sativa (Fabaceae), (M. Shamsi).
51

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
Comments
Holopalaearctic (Linnavuori, 2007), Europe, Asia, North Africa, North India, Nepal, Pakistan, Kashmir and
North America (Aukema & Rieger, 1999).
Agnocoris reclairei (Wagner, 1949)
Material examined
Iran, Kerman, Baft: khabr, August 2012, (2140m, 28˚48'59''N, 56˚20'49''E), Salix sp. (Salicaceae), (M.
Shamsi), Bongan, September 2012, (2615m, 29˚18'31''N, 56˚43'30''E), S. pendula (Salicaceae), (M. Shamsi),
Orzooiieh, July 2012, (1169m, 28˚22'42''N, 56˚29'10''E), S. Purpurea sp. (Salicaceae), (M. Shamsi), Torang,
August 2012, (2145m, 28˚45'21''N, 56˚48'52''E), Salix sp. (Salicaceae), (M. Shamsi), Dashtab, september 2012,
(2026m, 29˚59'45''N, 56˚38'12''E), Salix sp. (Salicaceae), (M. Shamsi), Sirjan: Sirjan, August 2012, (1746m,
29˚27'07''N, 55˚40'52''E), S. sp. (Salicaceae), (M. shamsi), Balvard, July 2012, (1949m, 29˚24'26''N,
56˚00'41''E), S. sp. (Salicaceae), (M. Shamsi), Mahmoodabad, September 2012, (1733m, 29˚31'40''N,
55˚36'23''E), S. sp. (Salicaceae), (M. Shamsi), Bardsir: Ghaleaskar, August 2012, (2659m, 29˚30'44''N,
56˚41'07''E), S. sp. (Salicaceae), (M. Shamsi).
Comments
The species was collected by light trap in gardens and deciduous forests. Euro-Siberian (Linnavuori, 2007).
Dichrooscytus persicus Josifov, 1974
Material examined
Iran, Kerman, Baft: Torang, August 2012, (2145m, 28˚45'21''N, 56˚48'52''E), Cupressus sempervirens
(Cupressaceae), (M. Shamsi), khabr, july 2012, (2140m, 28˚48'59''N, 56˚20'49''E), C. sempervirens
(Cupressaceae), (M. Shamsi), Rabor, September 2012, (2330m, 29˚17'29''N, 56˚54'45''E), C. sempervirens
(Cupressaceae), (M. Shamsi).
Comments
Irano-Turanian (Linnavuori, 2007).
Liocoris tripustulatus (Fabricus, 1781)
Material examined
Iran, Kerman, Bardsir: Lalezar, July 2012, (2844m, 29˚31'11''N, 56˚49'09''E), Salix sp. (Salicaceae), (M.
Shamsi).
Comments
Holomediterranean, extending to Turkey, Azerbaijan, Iran, and Iraq (Linnavuori, 2007).
Charagochilus gyllenhali (Fabricus, 1807)
Material examined
Iran, Kerman, Baft: Baft, July 2012, (2283m, 29˚13'59''N, 56˚36'08''E), Galium sp. (Rubiaceae), (M. Shamsi),
Khabr, August 2012, (2140m, 28˚48'59''N, 56˚20'49''E), (Weed), (M. Shamsi), Gogher, August 2012, (2625m,
29˚28'42''N, 56˚24'37''E), M. sativa (Fabaceae), (M. Shamsi), Bongan, July 2012, (2615m, 29˚18'31''N,
56˚43'30''E), Galium sp. (Rubiaceae), (M. Shamsi), Bardsir: Lalezar, July 2012, (2844m, 29˚31'11''N,
56˚49'09''E), Onobrychis sp. (Leguminosae), (M. Shamsi), Ghaleaskar, September 2012, (2659m, 29˚30'44''N,
56˚41'07''E), (Weed), (M. Shamsi).
Comments
Holopalaearctic (Linnavuori, 2007).
52

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
Megacoelum hormozganicum Linnavuori, 2004
Material examined
Iran, Kerman, Baft: Torang, July 2012, (2145m, 28˚45'21''N, 56˚48'52''E), Glycyrrhiza sp. (Fabaceae), (M.
Shamsi), Khabr, July 2012, (2140m, 28˚48'59''N, 56˚20'49''E), (Weed), (M. Shamsi), Dashtab, September 2012,
(2026m, 29˚59'45''N, 56˚38'12''E), Glycyrrhiza sp. (Fabaceae), (M. Shamsi).
Comments
Endemic to Iran (Linnavuori, 2009).
Orthops frenatus (Horvath, 1894)
Material examined
Iran, Kerman, Baft: Khabr, August 2012, (2140m, 28˚48'59''N, 56˚20'49''E), Medicago sativa (Fabaceae), (M.
Shamsi), Gogher, July 2012, (2625m, 29˚28'42''N, 56˚24'37''E), M. sativa (Fabaceae), (M. Shamsi), Dashtab,
July 2012, (2026m, 29˚59'45''N, 56˚38'12''E), Onobrychis sp. (Leguminosae), (M. Shamsi), Torang, July 2012,
(2145m, 28˚45'21''N, 56˚48'52''E), (M. Shamsi), Bardsir: Lalezar, July 2012, (2844m, 29˚31'11''N, 56˚49'09''E),
M. sativa (Fabaceae), (M. Shamsi), Ghaleaskar, September 2012, (2659m, 29˚30'44''N, 56˚41'07''E), M. sativa
(Fabaceae), (M. Shamsi).
Comments
The species were collected by light trap. Irano-Turanian, recorded from Armenia, Iran, Afghanistan, and
Middle Asia (Linnavuori, 2007).
Orthops pilosulus (Jakovlev, 1877)
Material examined
Iran, Kerman, Baft: Gogher, July 2012, (2625m, 29˚28'42''N, 56˚24'37''E), Amaranthus sp. (Amaranthaceae),
(M. Shamsi), Torang, August 2012, (2145m, 28˚45'21''N, 56˚48'52''E), (Weed), (M. Shamsi), Kiskan, August
2012, (2611m, 29˚22'46''N, 56˚38'14''E), Onobrychis sp. (Leguminosae), (M. Shamsi), Rabor, september 2012,
(2330m, 29˚17'29''N, 56˚54'45''E), (Weed), (M. Shamsi), Bardsir: Lalezar, september 2012, (2844m,
29˚31'11''N, 56˚49'09''E), Onobrychis sp. (Leguminosae), (M. Shamsi), Ghaleaskar, August 2012, (2659m,
29˚30'44''N, 56˚41'07''E), Amaranthus sp. (Amaranthaceae), (M. Shamsi), Torang, September 2012, (2145m,
28˚45'21''N, 56˚48'52''E), Amaranthus sp. (Amaranthaceae), (M. Shamsi).
Comments
Irano-Turanian (Linnavuori, 2007).
Creontiades pallidus (Rambur, 1839)
Material examined
Iran, Kerman, Baft: Orzooiieh, July 2012, (1169m, 28˚22'42''N, 56˚29'10''E), Gossypium arboreum
(Malvaceae), (M. Shamsi), Torang, August 2012, (2145m, 28˚45'21''N, 56˚48'52''E), Onobrychis sp.
(Leguminosae), (M. Shamsi), Khabr, August 2012, (2140m, 28˚48'59''N, 56˚20'49''E), Onobrychis sp.
(Leguminosae), (M. Shamsi).
Comments
Holomediterranean, widely distributed in the Middle East and the Ethiopian Region (Linnavuori, 2007).
Taylorilygus apicalis (Fieber, 1861)
Material examined
Iran, Kerman, Baft:Torang, september 2012, (2145m, 28˚45'21''N, 56˚48'52''E), Artemisia aucheri
53

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
(Asteraceae), (M.Shamsi), Khabr, August 2012, (2140m, 28˚48'59''N, 56˚20'49''E), A. aucheri (Asteraceae),
(M.Shamsi), Dashtab, July 2012, (2026m, 29˚59'45''N, 56˚38'12''E), (Weed), (M. Shamsi).
Comments
The species were collected by light trap. Cosmopolitan in tropical and subtropical regions (Linnavuori, 1999).
Subfamily: Orthotylinae
Orthotylus flavosparsus Sahlberg, 1841
Material examined
Iran, Kerman, Baft: Bezenjan, July 2012, (2358m, 29˚14'48''N, 56˚41'50''E), (Weed), (M. Shamsi), Khabr,
August 2012, (2140m, 28˚48'59''N, 56˚20'49''E), Rosa damascene (Rosaceae), (M. Shamsi), Gogher, July 2012,
(2625m, 29˚28'42''N, 56˚24'37''E), (M. Shamsi), Sirjan: Balvard, August 2012, (1949m, 29˚24'26''N,
56˚00'41''E), (Weed), (M. Shams), Bardsir: Lalezar, september 2012, (2844m, 29˚31'11''N, 56˚49'09''E),
(Weed), (M. Shamsi), Ghaleaskar, September 2012, (2659m, 29˚30'44''N, 56˚41'07''E), R. damascene
(Rosaceae), (M. Shamsi),
Comments
Holarctic (Linnavuori, 2007).
Globiceps fulvicollis Jakovlev, 1877
Material examined
Iran, Kerman, Bardsir: Lalezar, July 2012, (2844m, 29˚31'11''N, 56˚49'09''E), Onobrychis sp. (Leguminosae),
(M. Shamsi).
Comments
West-Palaearctic (Linnavuori, 2007).
References
Arkani T. 2009. Biodiversity and faunal study of plant bugs (Miridae) in crops and fruit trees of Arak and
suburb. Msc Thesis, Islamic Azad University Arak Branch, Iran
Asquith A. 1994. Revision of the endemic Hawaiian genus Sarona Kirkaldy (Heteroptera: Miridae:
Orthotylinae). Bishop Museum Occas Paper, 40: 1-81
Aukema B, Chr Rieger. 1999. Catalogue of the Heteroptera of the Palaeratic Region Vol 3. The Netherlands
Entomological Society, Netherlands
Blatchley WS. 2010. Heteroptera or True Bugs of Eastern North America. The Nature Publishing Company,
Indianapolis, USA
Brimah SA, Kelton LA, Stevart RK. 1982. The predaceous and Phytophagus Plant bugs (Heteroptera: Miridae)
found on apple trees in Quebec. Le Naturaliste Canadien, 109(2): 154-180
Cassis G, Schuh RT. 2012. Systematic, biodiversity, biogeography, and host associations of the Miridae
(Insecta: Hemiptera: Heteroptera: Cimicomorpha). The Annual Review of Entomology, 57: 377-404
Ebrahimi A, Hosseini R, Shoushtari RV. 2012. A faunal study of plant bugs (Hemiptera: Miridae) in Ghorveh
and its counties (Kurdistan province, Iran). Entomofauna, 33(4): 25-40
Hosseini R. 1997. A faunal study of Miridae (Heteroptera) in Guilan province. M.Sc thesis, Guilan University,
Iran
Hosseini R. 2013a. On the genus Pilophorus HAHN (Hemiptera: Miridae) in Guilan province and adjacent
areas. Entomofauna, 34: 105-116
54

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
Hosseini R. 2013b. On the tribe Dicyphini (Hemiptera: Heteroptera: Miridae: Bryocorinae) in Guilan province
and adjacent area (Iran). Entomofauna, 34: 157-158
Hosseini R. 2013c. On the tribe Stenodemini (Hemiptera: Miridae: Mirinae) in Guilan province and adjacent
areas (Iran). Entomofauna, 34: 377-396
Hosseini R. 2014. On the genus Adelphocoris (Hemiptera: Miridae) in Guilan province (Iran) and its adjacent
areas. Entomofauna, 35: 413-421
Hosseini R, Linnavuori, R. 2000. A faunal study on the mirids of Guilan province (Het.: Miridae,
Orthotylinae). Proceeding of the 14th Iranian Plant Protection Congress. Isfahan University of Technology,
Iran
Hosseini R, Linnavouri R, Sahragard A. et al. 2000. Taxonomic study on the Miridae (Heteroptera) of Guilan
province (subfamily: Orthotylinae). Proceeding of the 14th Iranian Plant Protection Congress. Isfahan
University of Technology, Iran
Hosseini R, Sahragard A, Hajizadeh J. et al. 2002. Taxonomic study of Mirid bugs in Guilan province-Tribe
Phylini. Proceeding of the 15th Iranian Plant Protection Congress. Razi University of Kermanshah, Iran
Hosseini R, Sahragard A, Hajizadeh J. et al. 2002b. Taxonomic study on the Miridae (Heteroptera) of Guilan
province. Proceeding of the 15th Iranian Plant Protection Congress, 307, Razi University of Kermanshah,
Iran
The Society and Department of Geography Kerman province. 2013. http://geography.kermanedu.ir/toc.htm.
Access on 2013
Lashkari M, Hosseini R. 2012. A revised identification key to the Lygus-species in Iran (Hemiptera: Miridae).
Entomofauna, 33: 81-92
Linnavouri RE. 2006. Studies on the Miridae (Heteroptera) of guilan and the adjacent provinces in northern
Iran.I. Description of new species. Acta Universittatis Carolinae Biologica, 49: 219-243
Linnavouri RE. 2007. Studies on the Miridae (Heteroptera) of guilan and the adjacent provinces in northern
Iran.II. List of species. Acta Entomologyca Musei Nationalis Pragae, 47: 17-56
Linnavouri RE. 2009. Studies on the Nepomorpha, Geromorpha, Leptopodomorpha and Miridae excluding
Phylini (Hemiptera: Heteroptera) of Khuzestan and adjacent province of Iran. Acta Entomologyca Musei
Nationalis Prage, 49(1): 1-32
Linnavouri RE, Hosseini R. 1998. New species of the Miridae (Heteroptera) from Iran. Acta Universitis
Carolinae, Biologica, 42: 3-15
Linnavouri RE, Hosseini R. 1999. On the genus Dicyphus (Heteroptera, Miridae, Dicyphinae) in Iran. Acta
Universitis Carolinae, Biologica, 43: 155-162
Linnavouri RE, Hosseini R. 2000. On the Polymerus subgenus Poeciloscytus FIEBER (Heteroptera, Miridae,
Mirinae) in Iran. Acta Universitis Carolinae, Biologica, 44: 189-194
Mirab-Balou M, Rasoulian GH, Khanjani R, et al. 2008. Study on Taxonomy of Phytophagus bugs of the
family Miridae and introducing insects natural enemies of the Alfalfa tarnished plant bug in Hamadan
Alfalfa farms (west of Iran). Pakistan Entomology Journal, 30(1)
Schuh RT. 2002-2013. On-line Systematic Catalog of Plant Bugs (Insecta: Heteroptera: Miridae).
http://research.amnh.org/pbi/catalog.
Schwartz MD. 2008. Revision of the Stenodemini with a review of the included genera (Hemiptera:
Heteroptera: Miridae: Mirinae). Proceedings of the Entomological Society of Washington, 110(4): 1111-
1201
Slater JA, Baranowski RM. 1978. How to Know the True Bugs (Hemiptera- Heteroptera). Brown Company
Publisher, Dubuque, Iowa, USA
55

Arthropods, 2014, 3(1): 48-56
IAEES www.iaees.org
Wagner E,Weber H. 1964. Heteroptera Miridae. Faune de France 67. Federation Francaise des Societies de
Sciences Naturelles, Paris, France
Wagner E. 1971. Drei neun Heteroptera anus Iran (Heteroptera, Miridae). Reichenbachia, 14: 31- 37
Wagner E. 1974. Die Miriden Hahn, 1831. Des Mittelmeerrranmes und der Markaronescischen Inseln. Teil 1.
Entomologische Abhandlungen Herausgegeben von Stoatl. Mus. Fur Naturkunde Dresden 37,
Supplementary 1-2: 484
Yarmand H, Sadeghi E, Asgari H, et al. 2004. Diversity of some Miridae (Heteroptera) species associated with
forests and rangelands of Iran. Proceeding of 16th Iranian Plant Protection Congress, Iran
56

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Article
A study on the genus Orthops FIEBER (Hemiptera: Miridae: Mirinae)
in Iran Reza Hosseini Department of Plant Protection, Faculty of Agricultural Sciences, University of Guilan, Rasht, Iran
E-mail: [email protected]
Received 10 September 2013; Accepted 1 October 2013; Published online 1 March 2014
Abstract
This paper is the extension of a series of synoptic taxonomic treatments on the Miridae known from Guilan
and other provinces in Iran. In the genus Orthops FIEBER five species are known from Iran, including
Orthops (Montanorthops) pilosulus (Jakovlev, 1877), Orthops (Orthops) frenatus (Horváth, 1894), Orthops
(Orthops) basalis (Costa, 1853), Orthops (Orthops) campestris (Linnaeus, 1758) and Orthops (Orthops) kalmii
(Linnaeus, 1758). Pinalitus cervinus (Herrich-Schaeffer, 1841) as a similar species to Orthops group is
included in this study. In this paper diagnoses, host-plant information, distribution data, and illustrated keys to
the genera and species are provided. For all species, illustrations of the adults and selected morphological
characters are provided to facilitate identification.
Keywords Miridae, Orthops; taxonomy; Guilan province.
1 Introduction
Mirid bugs (Hemiptera: Heteroptera) are one of the most species rich families of insects, with approximately
11020 described species. This family comprising eight subfamilies which among them Mirinae subfamily has
six tribes, including Herdoniini, Hyalopeplini, Mecistoscelini, Mirini, Resthenini and Stenodemini (Cassis and
Schuh, 2012), however Schuh (2013) has added Scutelliferini tribe to the above list.
Species in Orthops genus belong to the tribe Mirini. So far, 35 species has been recognized in this genus
belong to two subgenus Orthops and Montanorthops (Schuh, 2013). They have 3-6 mm body length. Vertex
provided with a margin or border. Genitalia of the males and females distinctly small. Body color variable, but
very seldom green, in that case the rostrum does not exceed the intermediate coxae (Wagner and Weber, 1964)
So far a few taxonomic works has been done on Miridae bugs in Iran (Hosseini, 1997; Hosseini and
Linnavuori, 2000; Hosseini et al., 2000, 2002a, b; Hosseini, 2013a, b, c; Hosseini, 2014; Linnavuori and
Hosseini, 1998, 1999, 2000; Linnavuori and Modarres, 1999; Linnavuori, 1999, 2006, 2007, 2009, 2010;
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Arkani et al., 2011; Lashkari et al., 2011; Lashkari and Hosseini, 2012; Ebrahimi et al., 2012).
In this genus five species are known from Guilan and other provinces in Iran, including, Orthops
(Montanorthops) pilosulus (Jakovlev, 1877), Orthops (Orthops) frenatus (Horváth, 1894), Orthops (Orthops)
basalis (Costa, 1853), Orthops (Orthops) campestris (Linnaeus, 1758) and Orthops (Orthops) kalmii
(Linnaeus, 1758) (Linnavuori, 2007).
There is a lack of information for identification of Miridae species in Iran specially in Guilan province.
This work is an extension of a series of synoptic taxonomic treatments and re-description on the Miridae
known from Guilan and other provinces of Iran in order to fill this information gap. In this paper following
information including diagnoses, host-plant information, distribution data, and illustrated keys for five species
of Orthops and also Pinalitus cervinus (Herrich-Schaeffer, 1841) as a similar species to Orthops group are
provided. For most species, illustrations of selected morphological characters are provided to facilitate
identification.
2 Materials and Methods
2.1 Collection of specimens
The sweep net, bush net and light trap was used for collecting mirids on vegetation and trees. The bugs felt on
the net were quickly picked off by an aspirator. Then collected specimens were killed promptly in a small tube
contains Ethyl acetate. Specimens after transferring to the laboratory were mounted on rectangular cards. All
specimens were examined using an Olympus SZX 12 stereomicroscope. A part of specimens examined in this
study were used from the insects’ collection available at the Natural History Museum of University of Guilan.
Illustrations of genitalia were prepared using a drawing tube attached to the stereomicroscope. Photographs of
specimens were taken using a Canon EOS 500D (Digital Rebel/Kiss X3 Digital) camera equipped by a Canon
EF 100mm f/2.8 USM Macro Lens. Identification was done by relevant taxonomic keys (Wagner and Weber,
1964; Wagner, 1971). Identified species were confirmed by mirid specialist Dr. R.E. Linnavuori (Finland). All
species are kept in the insect collection of the Natural History Museum of University of Guilan. For species
collected from Guilan province information including altitude, latitude and longitude were provided for each
collection site. The system and nomenclature follows the catalogue of Aukema and Rieger (1999).
3 Results
The following is the key to the species of Orthops.
1- Species with shallow punctures, collar hair much longer than width of collar, spines on tibia dark
(Subgenus Orthops) …………………………………………………………………………....…….……….2
- Species with deep punctures, suppressed hairs on pronotal collar, spines on tibia yellow, color golden
yellow with reddish splashes and black markings (Subgenus Motanorthops) …………………..……..……. 5
2- A brown line on external side of each tibia from its base to the middle ……. O. (Orthops) frenatus
- Rarely brown line is present, but if present, interrupted near the knee …………….. ………............ 3
3- Side edge of the corium pale, concolorous. Apophysis of right paramere in the form of a
punch ..............................................................………………………………….. O. (Orthops) campestris
- Side edge of corium narrowly black. Apophysis of right paramere in the form of a hook ............... 4
4- Length of 3rd antennal segment < 0.55 mm, Elongated species ……… O. (Orthops) basalis
- Length of 3rd antennal segment > 0.5 mm, Scutellum usually with a triangular basal spot. Oval
elongated species…………..………………………………………………….... O. (Orthops) kalmii
5- Species entirely orange-red ………………………………………..…. O. (Montaorthops) pilosulus
58

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Orthops (Orthops) frenatus (Horváth, 1894) (Fig. 1A-C, 7A)
Material examined
Guilan province: Deylaman (36°53'05'' N, 49°54'26'' E, elev. 141 m), 16.-20.vii.1996, 18.-20.vii.1998; Barasar
(36°46'43'' N, 49°45'31'' E, elev. 1171 m), 23.vii.1996; Jirandeh (36°42'00" N, 49°47'28" E, 1343 m), 6.-
8.vii.1996.
This species has been reported from Korasan province: (70 km W of Darreh Gaz, , 14.VI.1994; Khalkanlod
30 kmE of Quchan, 7.VI. 1994; Khargh 70 km SW of Quchan, 8-9.VI. 1994; Mashhad, V-VII.1994) –
collected from mountain meadows (Linnavuori and Modarres 1999); Ardabil province: (Khalkhal, 30.vii.1996,
20.vii.1998; 20-30 km E of Khalkhal, 1.-21.vii.1996; Majareh – Khalkhal, 28.vii.1996), Tehran province:
(Azad Bar, 8.-10.vii.1995) and Alborz province: (Karaj, 12.-13.vii.2002) Linnavuori (2007); Khuzestan
province: (Bagh Malek, 9.–11.v.2007); Esfahan province: (Shahreza – Semirom, 11.vi.2003); Fars province:
(Dasht-e-Arzhan, 12.–13.vi.2003; Shul – Sangar, 17.–18.vi.2003); Ilam province: ( Ilam, 8.–9.vi.2005); West
Azerbaijan province (Kitkeh 50 km S of Mahabad, 19.–20.vii.2004, Marangalu near Urumiyeh, 15.–
17.vii.2004) (Linnavuori 2009)
Diagnosis
O. fernatus has a different morphology from O. kalmii (Linnaeus, 1758), O. campestris (Linnaeus, 1758) and
O. basalis (Costa, 1853). In this species there is a brown line on external side of each tibia from its base to the
middle (Fig. 1B) while in other three species external side of tibia is entirely pale or with a brown spot at the
knee and rarely the brown line is present but interrupted near the knee (Kerzhner, 1996). Apophysis of left
paramere in the form a sharp curved hook. Sensory lobe with a strong hunch (Fig. 1C).
Specific taxonomical diagnostic characters are shown in Table 1.
Fig. 1 Orthops (Orthops) frenatus (Horváth, 1894). (A): Adult; (B): Leg; (C): Left paramere.
Comments
Orthops frenatus Horvath, 1894 has been considered as a junior synonym of Orthops kalmii Linnaeus, 1758 in
Schuh (2013). This species usually is found in mountain meadows and at light in gardens at riversides and in
59
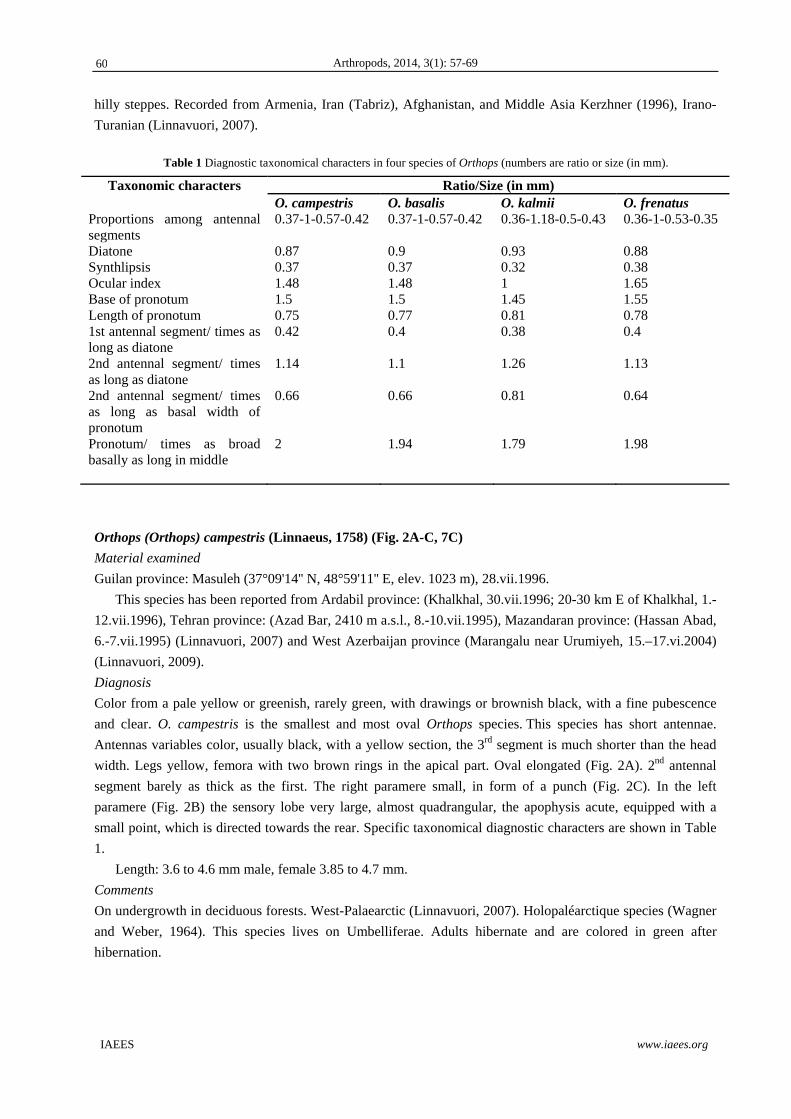
Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
hilly steppes. Recorded from Armenia, Iran (Tabriz), Afghanistan, and Middle Asia Kerzhner (1996), Irano-
Turanian (Linnavuori, 2007).
Table 1 Diagnostic taxonomical characters in four species of Orthops (numbers are ratio or size (in mm).
Orthops (Orthops) campestris (Linnaeus, 1758) (Fig. 2A-C, 7C)
Material examined
Guilan province: Masuleh (37°09'14'' N, 48°59'11'' E, elev. 1023 m), 28.vii.1996.
This species has been reported from Ardabil province: (Khalkhal, 30.vii.1996; 20-30 km E of Khalkhal, 1.-
12.vii.1996), Tehran province: (Azad Bar, 2410 m a.s.l., 8.-10.vii.1995), Mazandaran province: (Hassan Abad,
6.-7.vii.1995) (Linnavuori, 2007) and West Azerbaijan province (Marangalu near Urumiyeh, 15.–17.vi.2004)
(Linnavuori, 2009).
Diagnosis
Color from a pale yellow or greenish, rarely green, with drawings or brownish black, with a fine pubescence
and clear. O. campestris is the smallest and most oval Orthops species. This species has short antennae.
Antennas variables color, usually black, with a yellow section, the 3rd segment is much shorter than the head
width. Legs yellow, femora with two brown rings in the apical part. Oval elongated (Fig. 2A). 2nd antennal
segment barely as thick as the first. The right paramere small, in form of a punch (Fig. 2C). In the left
paramere (Fig. 2B) the sensory lobe very large, almost quadrangular, the apophysis acute, equipped with a
small point, which is directed towards the rear. Specific taxonomical diagnostic characters are shown in Table
1.
Length: 3.6 to 4.6 mm male, female 3.85 to 4.7 mm.
Comments
On undergrowth in deciduous forests. West-Palaearctic (Linnavuori, 2007). Holopaléarctique species (Wagner
and Weber, 1964). This species lives on Umbelliferae. Adults hibernate and are colored in green after
hibernation.
Taxonomic characters Ratio/Size (in mm) O. campestris O. basalis O. kalmii O. frenatus
Proportions among antennal segments
0.37-1-0.57-0.42 0.37-1-0.57-0.42 0.36-1.18-0.5-0.43 0.36-1-0.53-0.35
Diatone 0.87 0.9 0.93 0.88 Synthlipsis 0.37 0.37 0.32 0.38 Ocular index 1.48 1.48 1 1.65 Base of pronotum 1.5 1.5 1.45 1.55 Length of pronotum 0.75 0.77 0.81 0.78 1st antennal segment/ times as long as diatone
0.42 0.4 0.38 0.4
2nd antennal segment/ times as long as diatone
1.14 1.1 1.26 1.13
2nd antennal segment/ times as long as basal width of pronotum
0.66 0.66 0.81 0.64
Pronotum/ times as broad basally as long in middle
2 1.94 1.79 1.98
60

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Fig. 2 Orthops (Orthops) campestris (Linnaeus, 1758). (A): Adult; (B): Left paramere; (C): Right paramere.
Orthops (Orthops) basalis (Costa, 1853) (Fig. 3A-C, 7B)
Material examined
Guilan province: Deylaman (36°53'05'' N, 49°54'26'' E, elev. 141 m), 16.-20.vii.1996; Barasar (36°46'43'' N,
49°45'31'' E, elev. 1171 m), 23.vii.1996; Ganjeh (36°44'25" N, 49°24'03" E, elev. 302 m), 14.v.-13.vi.1995,
Jirandeh (36°42'00" N, 49°47'28" E, elev. 1343 m), 6.-8.vii.1996; Masuleh (37°09'14'' N, 48°59'11'' E, elev.
1023 m), 6.-26.vi.1995, 3.-8.vii.1996.
This species has been reported from Ardabil province: (Khalkhal, 30.vii.1996; Khalkhal – Kivi, 4.viii.1996;
Majareh – Kolur, 21.vii.1996), Zanjan province: (Gilankesh 15 km NW of Gilvan, 26.-27.vi.2004) (Linnavuori
2007), West Azerbaijan province: (50 km S of Mahabad, 19.–20.vii.2004), Markazi province (Tafresh, 7.–
8.vi.2006) (Linnavuori, 2009).
Diagnosis
O. basalis is always dark in color and never green like O. campestris and is larger, more elongate. Sometimes
in males three pale spots are seen on the scutellum.
Although this species is very similar to O. kalmii, the 3rd antennal segment is relatively long. The outer
edge of the corium very narrowly black, forehead usually with two black spots. Body shape longer and
narrower (Fig. 3A). Right paramere with a hook-shaped apophysis (Fig. 2C). Left paramere small, rounded
sensory lobe, provided with small teeth, the curved apophysis, very wide at its apex (Fig. 2B). Specific
taxonomical diagnostic characters are shown in Table 1.
Length: 4.1 to 5.2 mm male, female 4.2 to 5.3 mm.
Comments
On undergrowth in gardens and deciduous forests. Holomediterranean with a wide range in Europe. Usually
reported on Apiaceae (Linnavouri, 2007).
61

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Fig. 3 Orthops (Orthops) basalis (Costa, 1853). (A): Adult; (B): Left paramere; (C): Right paramere.
Orthops (Orthops) kalmii (Linnaeus, 1758) (Fig. 4A-C, 7D)
Material examined
Guilan province: Darreh Dasht (36°48'10'' N, 49°38'24'' E, elev. 1144 m), 18.-21.viii.1998; Dasht-e-Veel
(36°50'48" N , 49°35'32" E, elev. 293 m), 20.viii.-11.ix.1998, 8.-10.ix.2000, 27.-29.vii.2002; Ganjeh
(36°51'14" N, 49°28'09" E, elev. 212 m), 14.v.-13.vi.1995; Lowshan (36°38'09" N, 49°29'26" E, elev. 323 m),
18.-20.viii. 2002; Manjil (36°44'25" N, 49°24'03" E, elev. 302 m), 15.-17.ix.2000; Nasir Masaleh (37°05'41" N,
49°18'56" E, elev. 128 m), 14.-15.viii.2002; Sang Rud (36°39'59" N, 49°42'06" E, elev. 1338 m), 19.-
20.viii.2002.
This species has been reported from Khorasan province: (Golestan Park 150 km W of Bojnurd,
14.VII.1994; Khargh 70 km SW of Quchan, 8-9.VI.1994; Lotfabad, 15.VI.1994; Mashhad, V-VII.1994; Nodeh
30-40 km ESE of Bojnurd, 1 1.VH.1994; Zaman Soofi 65 km W of Bojnurd, 12-13.VII.1994) (Linnavuori and
Modarres 1999); Ardabil province: (20-30 km E of Khalkhal, 1.-21.vii.1996; Kivi, 9.-11.viii.2002); Zanjan
province: (Gilankesh, 5.-6.vii.2004, Abbhar, 29.viii.-9.x.2000); Mazandaran province: (Qaehm Shahr, 1.-
2.vii.2004), Golestan province: (Gorgan Mian Dareh, 13.-14.vii.2003; Talulestan, 15.-16.vii.2004) Linnavuori
(2007) and West Azerbaijan province (Marangalu near Urumiyeh, 15.–17.vii.2004) (Linnavuori, 2009)
Diagnosis
O. kalmii is more elongate and often more extensively dark-marked. This species is very similar to O. basalis,
however the 3rd antennal segment is slightly shorter.
Color like the preceding species. Lateral edge of the corium narrowly black. The drawings of the dorsal
sharper (Fig. 4A). Smaller size. The right paramer higher, its apophysis shorter (Fig. 4B). Left paramer (Fig.
4C) more robust, the sensory lobe rounded, provided with small teeth, the smallest apophysis. Specific
taxonomical diagnostic characters are shown in Table 1.
Length: male 4.1 to 4.7 mm, female 3.9 to 4.6 mm.
Comments
On undergrowth in deciduous forests and gardens and at light in gardens near salt marshes. Holopalaearctic
(Linnavuori, 2007). This species is often found on Apiaceae.
62

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Fig. 4 Orthops (Orthops) kalmii (Linnaeus, 1758). (A): Adult; (B): Right paramere (different views); (C): Left paramere (different views).
Orthops (Montanorthops) pilosulus (Jakovlev, 1877) (Fig. 5A-C, 7E)
Material examined
Guilan province: Manjil (36°44'25" N, 49°24'03" E, elev. 302 m), 8.-10.v.2000; Sang Rud (36°39'59" N,
49°42'06" E, elev. 1338 m), 21.ix.2000.
This species has been reported from Khorasan province: (Chan Chiroc 50 km S of Tabas, 17.V.1994; Deh
Shor 25 km N of Tabas, 17.IV.1994; Parvand 70-80 km W of Sabzevar, 14-15.V.1996; Sorond and Modar 70
km S of Tabas, 18.V.1994; Tabas, 16-17.IV.1994) (Linnavuori and Modarres 1999); Zanjan province: (Abbhar,
28.ix.-9.x.2000; Gilankesh 9.-13.x.2000); Tehran province: (Evin near Tehran, 14.-16.vii.1995; 25 km NE of
Firuzkuh, 9.vii.2003); Semnan province: (20 km SW of Biyarjomand, 13.-14.v.1996) Linnavuori (2007), and
Kerman province (Caravanseray-e-Farhali, 20.v.1996; Raviz Shahabieh, 1800 m a.s.l., 24.v.1996) (Linnavuori,
2009).
Diagnosis
This species is entirely orange-red. Body size about 4 mm. Finely hairy with whitish densely punctured. Head
smooth, shiny, collar thick, white. Rostrum reddish at the base, and brown at the end. Antenna orange yellow,
where the 3rd and 4th segments fine brownish. Clavus, cuneus reddish-orange in the center.
Underneath of body white, orange-red. Clavus, cuneus in the center and a wide part at the end of the
corium orange-red. Membrane pale, transparent. Legs yellowish, hind leg end red, yellow tibial spines.
Specific taxonomical diagnostic characters are shown in Table 2.
Comments
On Pteropyrum aucheri in hilly steppes. Irano-Turanian (Linnavuori, 2007).
In the insect collection of University of Guilan, R.E. Linnavuori has labeled another species of Orthops in
the subgenus Montanorthops, under title O. sanguinolentus (Reuter), however examining of this specimen did
not show any significant differences with O. pilosulus. He never mentioned the presence of O. sanguinolentus
63
Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
in Iran in any of his papers, a possible explanation is that he regarded the two species as synonyms.
Fig. 5 Orthops (Montanorthops) pilosulus (Jakovlev, 1877). (A): Adult; (B): right paramere (different views); (C): left paramere
(different views).
Table 2 Diagnostic taxonomical characters in Orthops (Montanorthops) pilosulus.
Taxonomic characters Ratio/Size (in mm) Proportions between 1st and 2nd antennal segments 0.22-1.06 Diatone 0.9 Synthlipsis 0.35 Ocular index 1.29 Base of pronotum 1.375 Lenght of pronotum 0.75 1nd antennal segment/ times as long as diatone 0.24 2nd antennal segment/ times as long as diatone 1.17 2nd antennal segment/ times as long as basal width of pronotum 0.77 Pronotum/ times as broad basally as long in middle 1.83
Pinalitus cervinus (Herrich-Schaeffer, 1841) (Fig. 6A-C, 7F)
Syn: Orthops cervinus (Herrich-Schaeffer, 1841)
Material examined
Guilan province: Darreh Dasht (36°48'10'' N, 49°38'24'' E, elev. 1144 m), 27.v.-28.vi.1995, 20.-25.viii.1998;
Dasht-e-Veel (36°50'48" N, 49°35'32" E, elev. 293 m), 20.-25.viii.1998; 40-50 km E of Khalkhal (37°37'57'' N,
48°30'33'' E, elev. 1765 m), 21.vii.1996; Manjil (36°44'25" N, 49°24'03" E, elev. 302 m), 16.v.-14.vi.1995,
30.x.-1.xi.2000; Nasir Mahaleh (Shaft) (37°05'41" N, 49°18'56" E, elev. 128 m), 14.-15.viii.2002; Rasht
(37°15'37'' N, 49°35'25'' E, elev. 1.5 m), vi.-viii.1996; Salan Sar (36°54'36'' N, 49°26'20'' E, elev. 741 m),
23.viii.1998; Sang Rud (36°39'59" N, 49°42'06" E, elev. 1338 m), 19.-20.viii.2002; Sang Rud – Jirandeh
(36°42'00" N, 49°47'28" E, elev. 1343 m), 31.v.1995 ; Tutkabon – Rudbar (36°53'30'' N, 49°31'43'' E, elev.
181 m), 6.-8.vi.2002.
64

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
This species has been reported from Ardabil province (20-30 km E of Khalkhal, 21.vii.1996; Majareh –
Khalkhal, 21.vii.1996) (Linnavuori, 2007).
Diagnosis
Elongated oval species (Table 3). Color from a yellowish brown, reddish or grayish pubescence, thin and clear.
Color of antenna yellow, the apical portion of 2nd , 3rd and 4th segments of antenna blackish brown. Posterior
margin of pronotum brown or black. Cuneus at its top brownish red or black. Length of species 2.5 to 3 times
longer than the width of the pronotum. Second segment of antennae as long or only slightly shorter than the
width of pronotum. Right paramere (Fig. 6C), the mastoid-shaped punch. Left paramere (Fig. 6B) small,
sensory lobe very small.
Length: male 3.8 to 4.2 mm, female 3.9 to 4.4 mm.
Comments
On deciduous trees. West-Palaearctic (Linnavuori, 2007). This species lives on trees with foliage (Tilia,
Fraxinus, Sorbus). Two types of color have been reported for this species: one color property grayish or
grayish brown and the ventral surface greenish, the other one yellowish brown or reddish and yellowish
ventrally (Wagner and Weber, 1964).
Table 3 Diagnostic taxonomical characters in Pinalitus cervinus.
Taxonomic characters Ratio/Size (in mm) Proportions among antennal segments 0.38-1.52-0.56-0.48 Diatone 0.88 Synthlipsis 0.27 Ocular index 0.84 Base of pronotum 1.37 Lenght of pronotum 0.63 1nd antennal segment/ times as long as diatone 0.43 2nd antennal segment/ times as long as diatone 1.72 2nd antennal segment/ times as long as basal width of pronotum 1.1 Pronotum/ times as broad basally as long in middle 2.17
Fig. 6 Pinalitus cervinus (Herrich-Schaeffer, 1841), , (A), Adult, (B), Left paramere, (C), Right paramere.
65

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Fig. 7 (A-C): (A): Orthops (Orthops) frenatus (Horváth, 1894), (B): Orthops (Orthops) basalis (Costa, 1853), (C): Orthops (Orthops) campestris (Linnaeus, 1758).
66

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Fig. 7 (D-F): (D): Orthops (Orthops) kalmii (Linnaeus, 1758), (E): Orthops (Montanorthops) pilosulus (Jakovlev, 1877), (F): Pinalitus cervinus (Herrich-Schaeffer, 1841).
67

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Acknowledgments
The author wishes to thank Dr. Rauno E. Linnavuori (Finland) for confirmation of the identified species.
References
Arkani T, Hosseini R, Vafaei Shoushtari R. 2011. Faunistic study of plant bugs (Miridae) and determination
dominant species in the agricultural farmlands and gardens of Arak and suburbs. Journal of Entomological
Research, 3: 85-93
Aukema B, Rieger Chr. 1999. Catalogue of the Heteroptera of the Palaeractic Region Vol 3. The Netherlands
Entomological Society, Netherlands
Cassis G, Schuh RT. 2012. Systemetic, biodiversity, biogeography, and host associations of the Miridae
(Insecta: Hemiptera: Heteroptera: Cimicomorpha). The Annual review of Entomology, 57: 377-404
Ebrahimi A, Hosseini R, Vafaei Shoushtari R. 2012. A faunal study of plant bugs (Hemiptera: Miridae) in
Ghorveh and its counties (Kurdistan province, Iran). Entomofauna, 33: 25-40
Hosseini R. 1997. A faunal study of Miridae (Herteroptera) in Guilan province. MSc thesis, University of
Guilan, Iran
Hosseini R. 2013a. On the genus Pilophorus hahn (Hemiptera: Miridae) in Guilan province and adjacent areas.
Entomofauna 34: 105-116.
Hosseini R. 2013b. On the tribe Dicyphini (Hemiptera: Heteroptera: Miridae: Bryocorinae) in Guilan province
and adjacent area (Iran). Entomofauna, 34: 157-158
Hosseini R. 2013c. On the tribe Stenodemini (Hemiptera: Miridae: Mirinae) in Guilan province and adjacent
areas (Iran). Entomofauna, 34: 377-396
Hosseini R. 2014. On the genus Adelphocoris (Hemiptera: Miridae) in Guilan province (Iran) and its adjacent
areas. Entomofauna, 35: 413-421
Hosseini R, Linnavuori, R. 2000. A faunal study on the mirids of Guilan province (Het.: Miridae,
Orthotylinae). Proceeding of the 14th Iranian Plant Protection Congress. Isfahan University of Technology,
Iran
Hosseini R, Linnavouri R, Sahragard A. et al. 2000. Taxonomic study on the Miridae (Heteroptera) of Guilan
province (sub family: Orthotylinae). Proceeding of the 14th Iranian Plant Protection Congress. Isfahan
University of Technology, Iran
Hosseini R, Sahragard A, Hajizadeh J. et al. 2002a. Taxonomic study of Mirid bugs in Guilan province-Tribe
Phylini. Proceeding of the 15th Iranian Plant Protection Congress. Razi University of Kermanshah, Iran
Hosseini R, Sahragard A, Hajizadeh J. et al. 2002b. Taxonomic study on the Miridae (Heteroptera) of Guilan
province. Proceeding of the 15th Iranian Plant Protection Congress, 307, Razi University of Kermanshah,
Iran
Kerzhner IM. 1996. On type specimens of some Palaearctic Miridae in the Hungarian Museum of Natural
History (Heteroptera). Zoosytematica Rossica, 5(1): 99-102
Kerzhner IM, Josifov M. 1999. Cimicomorpha II: Miridae Vol 3. In: Catalogue of the Heteroptera of the
Palaearctic Region (Aukema B, Rieger C, eds). Wageningen, Netherlands
Lashkari M, Hosseini R, Shahbazvar, N. 2011. A preliminary study on the Miridae (Hemiptera) fauna in
Mazandaran province in Northern Iran. Entomofauna, 32: 421-428
Lashkari M, Hosseini R. 2012. A revised identification key to the Lygus-species in Iran (Hemiptera: Miridae).
Entomofauna, 33: 81-92
Linnavuori RE, Modarres, M. 1999. Studies on the Heteroptera of the Khorasan province in N.E. Iran. II.
68

Arthropods, 2014, 3(1): 57-69
IAEES www.iaees.org
Cimicomorpha: Miridae. Entomologica Fennica, 10: 215-231
Linnavuori RE. 2006. Studies on the Miridae (Heteroptera) of Gilan and the adjacent provinces in northern
Iran. I. Description of new species. Acta Universitatis Carolinae Biologica, 49: 219-243
Linnavuori RE. 2007. Studies on the Miridae (Heteroptera) of Gilan and the adjacent provinces in northern
Iran. II. List of species. Acta Entomologica Musei Nationalis Pragae, 47: 17-56
Linnavuori RE. 2009. Studies on the Nepomorpha, Gerromorpha, Leptopodomorpha and Miridae excluding
Phylini (Hemiptera: Heteroptera) of Khuzestan and the adjacent provinces of Iran. Acta Entomologica
Musei Nationalis Pragae, 49(1): 1-32
Linnavuori RE. 2010. Studies on the Miridae (Phylinae, addenda to Deraeocorinae and Orthotylinae) of
Khuzestan and the adjacent provinces of Iran. Acta Entomologica Musei Nationalis Pragae, 50(2): 469-414
Linnavuori RE, Hosseini R. 1998. New species of the Miridae (Heteroptera) from Iran. Acta Universitatis
Carolinae, Biologica, 42: 3-15
Linnavuori RE, Hosseini R.1999. On the genus Dicyphus (Heteroptera, Miridae, Dicyphinae) in Iran. – Acta
Universitatis Carolinae, Biologica, 43: 155-162
Linnavuori RE, Hosseini R. 2000. On the Polymerus subgenus Poeciloscytus fieber (Heteroptera, Miridae,
Mirinae) in Iran. Acta Universitatis Carolinae, Biologica, 44:189-194
Schuh RT. 2013. On-line Systematic Catalog of Plant Bugs (Insecta: Heteroptera: Miridae). Retrieved April
2013 from http://research.amnh.org/pbi/catalog/
Wagner E, Weber H. 1964. Heteroptera Miridae. Faune de France 67. – Federation Francaise des societies de
Sciences Naturelles, Paris, France
Wagner E. 1971. Die Miriden Hahn, 1831. des Mittelmeerraumes und der Markaronesischen Inseln. Teil 1. –
Entomologische Abhandlungen Dresden 37, Supplementary 1
69

Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
Article
Predatory habits of Lutzia (Metalutzia) fuscana (Wiedmann) (Diptera:
Culicidae) in the arid environments of Jodhpur, western Rajasthan,
India Himmat Singh, Robin Marwal, Anusha Mishra, Karam Vir Singh Desert Medicine Research Centre, New Pali Road, Jodhpur, India -342005
E-mail: [email protected]
Received 30 August 2013; Accepted 13 October 2013; Published online 1 March 2014
Abstract
The stable breeding of Lutzia (Metalutzia) fuscana was recorded form different locations of Indian Desert the
“Thar” for the first time. The species being predatory in its larval form was investigated for evaluation of its
biological control aspect in the desert setup where breeding sites and prey species are limited. Though its
predatory habit is established yet using it as biological controlling agent was not found promising due to
untargeted approach due to unlimited outdoor breeding places in sub-humid climatic conditions in rest of India.
Whereas in desert due to limited water sources, mosquito vectors share the available breeding niche this
increases possibility of targeted biological control using predatory species. Laboratory experiments on
predatory habit of Lutzia (Metalutzia) fuscana showed that it preferred Aedes aegypti larvae most (88.5%),
Anopheles stephensi (47.5%) and Culex quinquefasciatus larvae (39.0%). Average consumption of daily larvae
is 18.89 larvae/day. If colonized properly and released in controlled conditions they can be useful in
controlling of socially protected and unattended breeding containers resulting reduction in mosquito population.
Keywords Lutzia (Metalutzia) fuscana; predatory; biological control; desert.
1 Introduction
Vetorborne disease burden has increased considerably worldwide in recent decades, globally cases have
increased from few millions to several billions per year (WHO Report, 2010). During the year 2010 - 2011 in
India alone the vector-borne disease cases were about 781603 including 1390 deaths which included 745599
malaria cases with 233 deaths, 14047 cases with 93 deaths of dengue, 14820 cases of chickungunya and 7137
cases with 1064 deaths reported due to Japanese encephalitis (National Vectorborne Disease Control Program
Report, 2011).
One of the effective methods of control over vector-borne transmission is through reduction in density of
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
vector population through various chemical means but now it is known that continuous use of chemicals for
control is associated with the emergence of insecticide resistance, this has necessitated exploring and opting
for new alternate methods.
Biological control is most suitable in this context as few attempt for introduction of biological agents like
larvivorus fishes i.e. gambusia and guppy have been successful (Chandra et al., 2008) bacteria (Dua et al.,
1993; Kumar et al., 1995; Indranil et al., 1997, Shukla et al., 1997; Biswas et al., 1997), fungi (Chandrahas
and Rajagopalan, 1979; Roberts and Strand, 1977, Poopathi and Tyagi, 2006) and predatory mosquitoes of
subgenera Mucidus (Mattingly, 1961), Culex fuscanus (Ikeshoji, 1966; Panicker et al., 1982) Toxorhynchites
spp. to control Aedes aegypti and Culex quinquefasciatus larvae (Gerberg and Visser, 1978; Focks et al., 1982;
Sempala, 1983; Ramalingam and Ramakrishnan, 1971; Mogi and Chan, 1996). Introduction of predatory
mosquito species is one of the targeted approaches for control of immature forms of mosquitoes. Lutzia
(Metalutzia) fuscana is one of such species of mosquito whose larvae is reported to feed upon vector species
larvae i.e Anopheles, Aedes and Culex species in several parts of India (Geetha, et al. 1982; Panicker et al.,
1982). Its predatory habit was found to be excellent yet non-targeted in the mesic environmental conditions
due to availability of several outdoor breeding habitats. Stable breeding of Lutzia (Metalutzia) fuscana has
been first time reported from this arid zone (Singh et al., 2013).
The genus of predatory mosquito Lutzia fuscana has been elevated from Culex to Lutzia (earlier Lutzia was
subgenus) earlier Culex (Lutzia) fuscanus (Wiedmann, 1820) is now Lutzia (Metalutzia) fuscana (Wiedmann)
(Tanaka, 2003). We have also used the same in present communication.
The present study is a laboratory-based study conducted in Vector Biology Laboratory of Desert Medicine
Research Centre, Jodhpur, Rajasthan. We envisage to explore the predatory species preference and potential of
Lutzia (Metalutzia) fuscana and possibility of its being utilized in mosquito control in the limited outdoor
breeding habitats and in community containers which are socio-culturally protected for cattle drinking, looking
into the predatory habits of Lutzia (Metalutzia) fuscana larvae, laboratory experiments were carried out on
feeding capacity and preference on immature stages of Culex, Anopheles and Aedes species. The results are
presented through this communication.
2 Material and Methods
Larval collection were carried out at seven selected study areas of urban and peri-urban localities of Jodhpur
city (Jhalamand, Khema ka Kunwa, Mandore, Nagori Gate, Salawas and Nehru Park) during 2010-11 on
monthly basis. Collections were carried out at various domestic and peri-domestic water storing containers,
cemented tanks, ponds, ditches, gardens and green complexes with parks, stored/ stagnated water, community
tanks for cattle drinking and artificial fountains in the parks.
Larvae were collected using white enamel bowls (4 cm diameter) with classical dipping method. The
larvae collections were kept in white enamel tray (20 x 15 x 3 cm) in laboratory at 26- 27Co
Three main breeding species of mosquito vectors in the region i.e. Anopheles stephensi, Aedes aegypti and
Culex quinquefasciatus, were used for determining the feeding preference of Lutzia (Metalutzia) fuscana
(other sympatric mosquitoes species of seasonal occurrence and of less significance as disease vectors were
not included in the study). However, for the daily consumption of larvae in predatory activity and feeding
preference on different stages of mosquito larvae, laboratory reared Aedes aegypti larvae were used. Only early
IVth stage larvae of predatory species were included in the present experiments to avoid any bias of the
different size and age of larvae.
For estimating species preference, equal number (50 larvae of IVth Instar) of larvae each of Aedes, Culex
and Anopheles were provided to larvae of Lutzia (Metalutzia) fuscana larvae by keeping them in one container
71
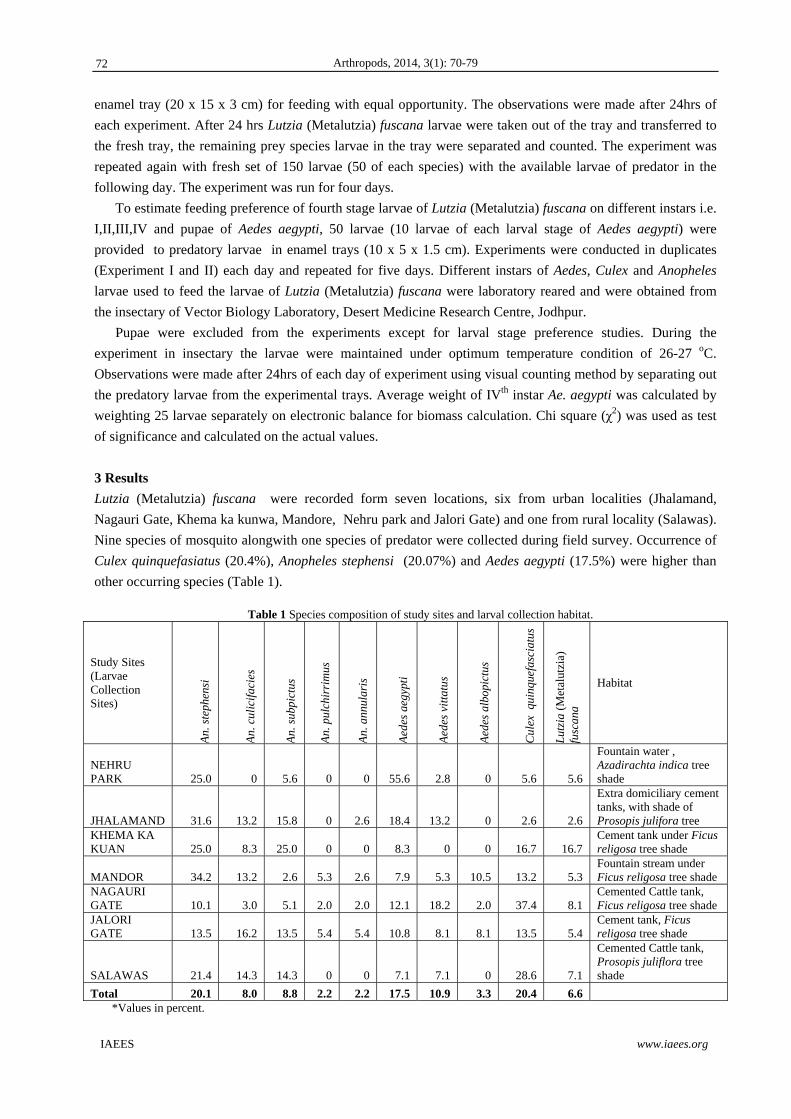
Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
enamel tray (20 x 15 x 3 cm) for feeding with equal opportunity. The observations were made after 24hrs of
each experiment. After 24 hrs Lutzia (Metalutzia) fuscana larvae were taken out of the tray and transferred to
the fresh tray, the remaining prey species larvae in the tray were separated and counted. The experiment was
repeated again with fresh set of 150 larvae (50 of each species) with the available larvae of predator in the
following day. The experiment was run for four days.
To estimate feeding preference of fourth stage larvae of Lutzia (Metalutzia) fuscana on different instars i.e.
I,II,III,IV and pupae of Aedes aegypti, 50 larvae (10 larvae of each larval stage of Aedes aegypti) were
provided to predatory larvae in enamel trays (10 x 5 x 1.5 cm). Experiments were conducted in duplicates
(Experiment I and II) each day and repeated for five days. Different instars of Aedes, Culex and Anopheles
larvae used to feed the larvae of Lutzia (Metalutzia) fuscana were laboratory reared and were obtained from
the insectary of Vector Biology Laboratory, Desert Medicine Research Centre, Jodhpur.
Pupae were excluded from the experiments except for larval stage preference studies. During the
experiment in insectary the larvae were maintained under optimum temperature condition of 26-27 oC.
Observations were made after 24hrs of each day of experiment using visual counting method by separating out
the predatory larvae from the experimental trays. Average weight of IVth instar Ae. aegypti was calculated by
weighting 25 larvae separately on electronic balance for biomass calculation. Chi square (χ2) was used as test
of significance and calculated on the actual values.
3 Results
Lutzia (Metalutzia) fuscana were recorded form seven locations, six from urban localities (Jhalamand,
Nagauri Gate, Khema ka kunwa, Mandore, Nehru park and Jalori Gate) and one from rural locality (Salawas).
Nine species of mosquito alongwith one species of predator were collected during field survey. Occurrence of
Culex quinquefasiatus (20.4%), Anopheles stephensi (20.07%) and Aedes aegypti (17.5%) were higher than
other occurring species (Table 1).
Table 1 Species composition of study sites and larval collection habitat.
Study Sites (Larvae Collection Sites)
An.
ste
phen
si
An.
cul
icifa
cies
An.
sub
pict
us
An.
pul
chir
rim
us
An.
ann
ular
is
Aed
es a
egyp
ti
Aed
es v
ittat
us
Aed
es a
lbop
ictu
s
Cul
ex q
uinq
uefa
scia
tus
Lutz
ia (
Met
alut
zia)
fu
scan
a
Habitat
NEHRU PARK 25.0 0 5.6 0 0 55.6 2.8 0 5.6 5.6
Fountain water , Azadirachta indica tree shade
JHALAMAND 31.6 13.2 15.8 0 2.6 18.4 13.2 0 2.6 2.6
Extra domiciliary cement tanks, with shade of Prosopis julifora tree
KHEMA KA KUAN 25.0 8.3 25.0 0 0 8.3 0 0 16.7 16.7
Cement tank under Ficus religosa tree shade
MANDOR 34.2 13.2 2.6 5.3 2.6 7.9 5.3 10.5 13.2 5.3 Fountain stream under Ficus religosa tree shade
NAGAURI GATE 10.1 3.0 5.1 2.0 2.0 12.1 18.2 2.0 37.4 8.1
Cemented Cattle tank, Ficus religosa tree shade
JALORI GATE 13.5 16.2 13.5 5.4 5.4 10.8 8.1 8.1 13.5 5.4
Cement tank, Ficus religosa tree shade
SALAWAS 21.4 14.3 14.3 0 0 7.1 7.1 0 28.6 7.1
Cemented Cattle tank, Prosopis juliflora tree shade
Total 20.1 8.0 8.8 2.2 2.2 17.5 10.9 3.3 20.4 6.6 *Values in percent.
72

Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
The breeding of Lutzia (Metalutzia) fuscana were found maximum in Cemented tanks which were found
under shades of trees like Ficus religosa and Prosopis juliflora (Table 1).
The predatory larvae of Lutzia (Metalutzia) fuscana were found from July to March except three months
from April to June in these locations (Fig. 1). It is observed that high temperature and low RH of summer did
not support outdoor breeding and in addition to this nearly drying of the water bodies and other water
collection results in reduction of prey community during these months. After onset of monsoon the predatory
mosquito larvae population were observed to appear again and its density increases with increase in the density
of prey mosquito species belonging to genera Aedes, Anopheles and Culex (Fig. 1).
Fig. 1 Monthly occurrence of Lutzia (Metalutzia) fuscana with respect to rainfall and temperature (Singh et al., 2013).
Fig. 2 Monthly occurrence of Lutzia (Metalutzia) fuscana with other prey species.
73

Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
The occurrence of Lutzia (Metalutzia) fuscana is also associated with density of prey population in
breeding sites, frequency of occurrence of prey species during the hotter months of June and July was found
very low (Fig. 2).
The larvae of predator shared niche with Ae. aegypti, Ae. albopictus, Ae. vittatus, An. stephensi, An.
culicifaces, An. subpictus and Cx. quinquefaciatus larvae. The predatory larvae mainly found in peri-domestic
cemented containers which were under shades of large trees like Peepal (Ficus religosa), Neem (Azadirachta
indica) and Kikar (Prosopis juliflora).
Occurrence of predatory larvae species was found high with Cx. quinquefaciatus (11time out of 13 times
when predator was found in the cement tanks) followed by Ae. aegypti (10 times out of 13) and Ae. vittatus (9
out of 13 times in cement tanks) though breeding of An. stephensi was high in cement tanks but the occurrence
with Lutzia (Metalutzia) fuscana was very low (4 out of 13 observations of predator species) (Table 2).
Table 2 Showing details of occurrence of different species larvae with Lutzia (Metalutzia) fuscana.
Breeding sites
An. stephens
i
An. culicifaci
es
An. subpictu
s
An. pulchirrimu
s
An. annulari
s
Ae.aegypti
Ae. vittatus
Ae. albopictus
Cx quniquefasi
atus
Lutzia fuscanu
s
No. of
Obs.
cement tanks
22 (4)*
3 (0)
10 (0)
0 (0)
1 (0)
28 (10)
21 (9)
0 (0)
29 (11) 13 47
Fountains 24 (3)
7 (3)
9 (1)
1 (0)
5 (0)
2 (1)
1 (0)
3 (0)
11 (2) 3 32
River bed 2 (0)
2 (0)
0 (0)
0 (0)
1 (0)
2 (0)
0 (0)
0 (0)
2 (0) 0 3
Stone quarry
5 (0)
4 (0)
3 (0)
0 (0)
0 (0)
2 (0)
2 (0)
0 (0)
3 (0) 0 7
Pond 0 (0.0)
0 (0)
0 (0)
0 (0)
1 (0)
1 (1)
0 (0)
0 (0)
1 (1) 1 1
Water pit 1 (0.0)
0 (0)
1 (0)
0 (0)
0 (0)
1 (0)
0 (0)
0 (0)
1 (0) 0 2
*Figures in parenthesis are occurrence of species with Lutzia (Metalutzia) fuscana larvae in same breeding spot
The result showed that Lutzia (Metalutzia) fuscana preferred Ae. aegypti larvae the most (50.6%, χ2=
31.20 , p≥ 0.001 at df =2) which was found significantly higher than An. stephensi larvae (27.1%, χ2 = 4.02 at
df =2) and Cx. quinquefasciatus (22.3%, χ2= 12.8 , p≥ 0.001 at df =2) (Table 3).
When single predator (IVth stage) larvae was given the choice to feed on different stages of larvae of Ae.
aegypti (50 larvae, 10 of each instar i.e. I,II, III, IV and pupae). The preference of feeding was on fourth instar
larvae (45.5%, χ2= 54.33, p≥ 0.001 at df = 4), followed by third instar (33.5%, χ2= 15.29, p≥ 0.01) and the least
preferred larvae were Ist instar (1.8%, χ2 = 27.66, p≥ 0.01 at df = 4). Although pupae were not consumed by
predator larvae but were found dead may be due to attack by predator causing mortality 2.99 % (Table 4).
Average consumption of larvae by predator (IVth Instar) per day was found to be about 18.4 larvae (using
IVth instar larvae of Ae. aegypti as they were most preferred) (Table 5). On the other hand using mixture of
equal number of three species larvae (Aedes, Anopheles and Culex) the average consumption per day was
about 18.4 larvae (Table 3) which was almost similar to the consumption using single species larvae feeding
experiment (Table 5).
Predatory larvae consumed about 37.21 mg biomass per day (calculated using mean daily consumption
multiplied by the average weight of live Ae. aegypti (n=25, mean weight = 1.96±0.22).
74
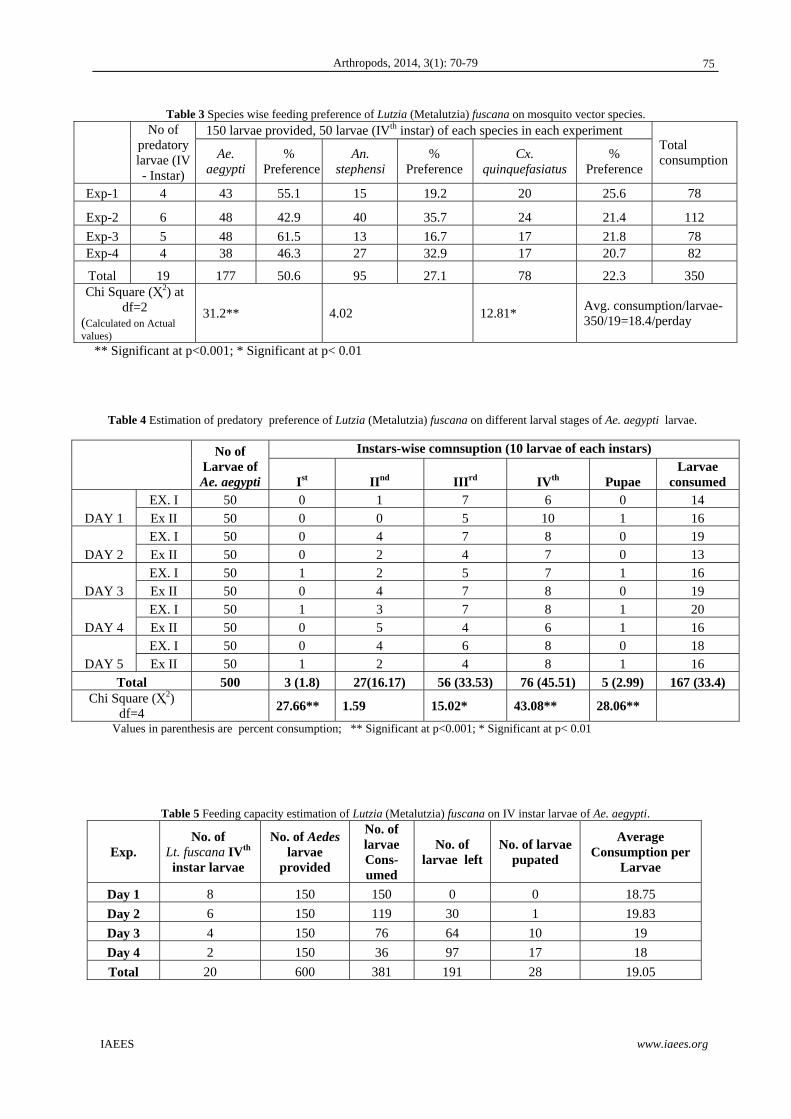
Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
Table 3 Species wise feeding preference of Lutzia (Metalutzia) fuscana on mosquito vector species.
No of predatory larvae (IV
- Instar)
150 larvae provided, 50 larvae (IVth instar) of each species in each experiment Total consumptionAe.
aegypti %
Preference An.
stephensi %
PreferenceCx.
quinquefasiatus %
Preference
Exp-1 4 43 55.1 15 19.2 20 25.6 78
Exp-2 6 48 42.9 40 35.7 24 21.4 112
Exp-3 5 48 61.5 13 16.7 17 21.8 78 Exp-4 4 38 46.3 27 32.9 17 20.7 82
Total 19 177 50.6 95 27.1 78 22.3 350 Chi Square (Ҳ2) at
df=2 (Calculated on Actual values)
31.2** 4.02 12.81* Avg. consumption/larvae- 350/19=18.4/perday
** Significant at p<0.001; * Significant at p< 0.01
Table 4 Estimation of predatory preference of Lutzia (Metalutzia) fuscana on different larval stages of Ae. aegypti larvae.
Values in parenthesis are percent consumption; ** Significant at p<0.001; * Significant at p< 0.01
Table 5 Feeding capacity estimation of Lutzia (Metalutzia) fuscana on IV instar larvae of Ae. aegypti.
Exp. No. of
Lt. fuscana IVth instar larvae
No. of Aedes larvae
provided
No. of larvae Cons-umed
No. of larvae left
No. of larvae pupated
Average Consumption per
Larvae
Day 1 8 150 150 0 0 18.75
Day 2 6 150 119 30 1 19.83
Day 3 4 150 76 64 10 19
Day 4 2 150 36 97 17 18
Total 20 600 381 191 28 19.05
No of Larvae of Ae. aegypti
Instars-wise comnsuption (10 larvae of each instars)
Ist IInd IIIrd IVth Pupae Larvae
consumed
DAY 1 EX. I 50 0 1 7 6 0 14 Ex II 50 0 0 5 10 1 16
DAY 2 EX. I 50 0 4 7 8 0 19 Ex II 50 0 2 4 7 0 13
DAY 3 EX. I 50 1 2 5 7 1 16 Ex II 50 0 4 7 8 0 19
DAY 4 EX. I 50 1 3 7 8 1 20 Ex II 50 0 5 4 6 1 16
DAY 5 EX. I 50 0 4 6 8 0 18 Ex II 50 1 2 4 8 1 16
Total 500 3 (1.8) 27(16.17) 56 (33.53) 76 (45.51) 5 (2.99) 167 (33.4) Chi Square (Ҳ2)
df=4 27.66** 1.59 15.02* 43.08** 28.06**
75

Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
4 Discussion
There have been several reports on its feeding behavior (Jin et al., 2006). The predatory activity of these larvae
was observed during entomological surveys in rest of the places other than the desert part of India. This is also
a first report of presence of this species in the desert scenario. Presence of stable population of Lutzia
(Metalutzia) fuscana is indicator of stability of vector species (Aedes, Anopheles and Culex species) on which
its larvae prey upon in this desert climate.
Except for three months (April to June) it is found throughout the year with stable reemerging population
which also signifies the stability of this predatory mosquito in this fragile and hostile climate of desert.
It was interesting to note that Lutzia (Metalutzia) fuscana preferred Ae. aegypti over Cx. quinquefasciatus
as reported in other part of India (Ikeshoji, 1966; Panicker et al. 1982; Singh et al. 1984; Thangam and
Kathiresan, 1996; Mariappan et al., 1997; Yanovisk, 2001), This changed preference of feeding of this
predator becomes very important for bio-control of Ae. Aegypti in this part of western Rajasthan as Ae aegypti
is major vector for Dengue and DHF. The possible change in the feeding preference might be due to niche
sharing by both prey and predator species (Aedes aegypti and Lutzia (Metalutzia) fuscana). A study by Shrama
et al. (2008) on surveillance of Aedes aegypti in Jodhpur showed cement tanks as the most important breeding
containers for Aedes breeding. Our study revealed that Lutzia (Metalutzia) fuscana also prefer to breed in the
same types of cement containers which are pri-domestic placed under shades of big trees for cattle drinking
The second reason may be due to limited availability of conducive outdoor breeding containers might bound
the mosquito species to share the common niches. Similar observations on the Aedes preference is recently
reported in Sri-lanka (Sinnathamby et al., 2013).
Lutzia (Metalutzia) fuscana is container and tree hole breeder (Belkin, 1962; Jackson, 1953, Tanaka, 2003;
Rattanarithikul et al., 2005) and breeds in the containers where other mosquitoes are breeding as they serves
food for this mosquito larvae, though cannibalism is obligatory (Hopkins, 1952; Edwards, 1941). Our
preliminary observation seems to be promising for biological control in important mosquito breeding
containers in part of western Rajasthan.
The study by Jackson (1953) in west Nigeria showed that the density of Lutzia (Metalutzia) fuscana larvae
is dependent on the density of other larvae available i. e. one larvae of Lutzia (Metalutzia) fuscana per 60 to 90
other larvae of either species of Aedes, Anopheles or Culex and this was found to be true in this part of desert
also.
Results on feeding behavior shows that single IVth instar predator larvae of Lutzia (Metalutzia) fuscana can eat
an average of 18.8 larvae per day of IVth instar larvae of Aedes aegypti and the average biomass consumption
was 37.21 mg/day which was found considerably lower in contrast to the consumption in sub humid areas of
south east coast of India where biomass consumption was reported 76.0 mg/day (Thangam and Kathiresan,
1996). Present study have shown that in case of western Rajasthan where water resources are limited but socio
cultural practices of water storing have stabilize mosquito breeding of vectors of dengue and malaria. In
addition to this in recent past increased supply of water through Indira Gandhi Canal water to city have
increased overall ambience of Jodhpur city and has resulted in creation of gardens and increased larger trees
might have attracted this tree hole / container breeder predatory mosquitoes to this part of desert, on the other
hand due to limited breeding sources available, the sharing of species might have changed the preference of
Lutzia (Metalutzia) fuscana from Cx. quinquefasciatus (Thangam and Kathiresan, 1996 ) to Ae. aegypti in this
part of desert.
The dynamics of species occurrence in the desert of Lutzia (Metalutzia) fuscana is correlated with other
species occurrence during the year it can be a tool for focused biological control if colonized properly and
released in controlled conditions. They can cover a lot of unattended key breeding containers and resulting
76

Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
reduction in mosquito population (Hopkins, 1952; Edwards, 1941) this biological control method seems more
suitable and focused in this setting of desert condition . Although some reports show that there may be lack of
interaction between larvae of mosquito vectors and their natural enemies and/or lower predator survivorship in
certain habitats, particularly shallow water pools and cement tanks (Das et al., 2006) and urban environments
such as temporal habitats (Carlson et al., 2004). A study by Jin et al. (2006) through gut content analysis of
Lutzia (Metalutzia) fuscana showed chironomidae larvae 78.6% and only 2.5% larvae of mosquitoes in
province of China whereas in this arid part of India where chironomids are not common in appearance, as arid
climate do not support much to them, the possibility of prey choice increases towards mosquito larvae in the
desert part due to less habitat availability the species interaction may occur more frequently
It was observed that ability of larvae consumption increases as larvae grew up and passes stages up to IVth
instar larvae (Appawu et al., 2000). It was also interesting to note that predatory larvae attacks on mostly on
its equal sized prey larvae by attacking them at the joint of head and thorax (Jin et al., 2006).
Ikeshoji (1966) used larvae of Lutzia (Metalutzia) fuscana to control Cx quinquefasciatus larvae in small
ditches in simulated field conditions.
Introduction of such predatory species to those public tanks which are filled for cattle and other animals to
drink water and community do not accept use of insecticide in these key can prove to be very useful. Secondly
in desert there are limited water resources and very less available breeding containers as a result of that species
of mosquito show niche sharing which might have changed this predatory species feeding preference from
Culex to Aedes species. Therefore the predatory species becomes more targeted in absence of large outdoor
breeding sources.
Environmental methods and biological control are alternatives to chemical control and are key components
of the integrated strategy which may go hand in hand to the National Vector Control Program. The successful
implementation of these organisms depends on an in-depth understanding of the ecology of both the targeted
species and the biological controlling predators.
Introduction of prey comes under environmental management strategies that can reduce or eliminate vector
breeding through use of biological controls that target and reduce vector larvae without generating the
ecological impacts of chemical use. The approach is cost-effective, ecological balanced and sustainable for
vector control if used in this type of climatic condition where mostly outdoor breeder larvae are restricted in
pockets.
Acknowledgements
Authors are thankful to Director, Desert Medicine Research Centre, Jodhpur for providing facilities of
Laboratory. We are also thankful to staff of Vector Biology Laboratory for providing assistance in laboratory
as well as field work related to present communication
References
Appawu MA, Dadzie SK, Quartey SQ. 2000. Studies on the feeding behaviour of larvae of the predaceous
mosquito Culex (Lutzia) tigripes Grandpre and Chamoy (Diptera: Culicidae). Insect Science and its
Application, 20(4): 245-250
Belkin JN. 1962. The Mosquitoes of the South Pacific (Diptera, Culicidae) Vol 2. University of California
Press, Berkeley, CA, USA
Biswas D, Ghosh SK, Dutta RN, Mukhopadhyay AK. 1997. Field trial of bacticide on larval population of
two species of vector mosquitoes in Calcutta. Indian Journal of Malariology, 34(1): 37-41
77

Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
Carlson J, Keating J, Mbogo CM, et al. 2004. Ecological limitations of aquatic predator colonization in the
urban environment. Journal of Vector Ecology, 29(2): 331-339
Chandra G, Bhattacharjee I, Chatterjee SN, Ghosh A. 2008. Mosquito control by larvivorous fish. Indian
Journal of Medical Research, 127: 13-27
Chandrahas RK, Rajagopalan PK. 1979. Observations on mosquito breeding and the natural parasitism of
larvae by a fungus Coelomomyces and a mermithid nematode Romanomermis in paddy fields in
Pondicherry. Indian Journal of Medical Research, 69: 63-67
Das PK, Sivagnaname N, Amalraj DD. 2006. Populations interactions between Culex vishnui mosquitoes and
their natural enemies in Pondicherry, India. Journal of Vector Ecology, 31(1): 84-88
Dua VK, Sharma SK, Sharma VK. 1993. Application of Bactoculicide (Bacillus thuringiensis H-14) for
controlling mosquito breeding in industrial scrap at BHEL, Hardwar (U P). Indian of Journal Malariology,
30: 29-35
Edwards FW. 1941. Mosquitoes of the Ethiopian Region. III.-Culicine adults and pupae. Mosquitoes of the
Ethiopian Region. III.-Culicine Adults and Pupae.
Focks DA, Sackett SR, Bailey DL. 1982. Field experiments on the control of Aedes aegypti and Culex
quinquefasciatus by Toxorhynchites rutilus rutilus (Diptera: Culicidae). Journal of Medical Entomology
19: 336-339
Geetha Bai, Viswam MK, Panick KN. 1982. Culex (Lutzia) fuscanus (Diptera: Culicidae) - a predator of
mosquito. Indian Journal of Medical Research, 76: 837-839
Gerberg EJ, Visser WM. 1978. Preliminary field trial for the biological control of Aedes aegypti by means
of Toxorhynchites brevipalpis, a predatory mosquito larva. Mosquito New, 38: 197-200
Hopkins GHF. 1952. Mosquitoes of the Ethopian Region (2nd edn)Vol.1. B.M. (N.H.), London, UK
Ikeshoji T. 1966. Bionomics of Culex( Lutzia) fuscanus. Japanese Journal of Experimental Medicine, 36: 321-
334
Indranil K, Alex Eapen, Ravindran KJ, Chandrahas RK, et al 1997. Field evaluation of Bacillus sphericus
H5a5b and B. thuringiensis var. israelensis, H-14 against the Bancroftian filariasis vector Culex
quinquefasciatus, Say in Chennai, India. Indian of Journal Malariology, 34: 25-36
Jackson N. 1953. Observation on the feeding habits of predacious mosquito larvae, Culex
trigripes. Proceedings of Royal Entomological Society London, 28: 153-159
Jin LQ, Luo JM, Fu YC, et al. 2006. Prey and feeding behavior of larval Culex (Lutzia) fuscanus (Diptera:
Culicidae) in Shantou, Guangdong Province, China. Journal of Medical Entomology, 43(4): 785-786
Kumar A, Sharma VP, Thavaselvam D, Sumodan PK. 1995. Control of An. stephensi breeding in construction
sites and abandoned overhead tanks with Bacillus thuringiensis var isrealensis. Journal of American
Mosquito Control Association, 11(1): 86-89
Mariappan P, Narayanan M, Balasundaram C. 1997. Occurrence of mosquito larvae and its predators in and
around Palayankottai, Tamil Nadu. Environment and Ecology, 15(3): 678-682
Mattingly PF. 1961. The Culicine Mosquitoes of the Indomalayan Area: Genus Aëdes Meigen, subgenera
Mucidus Theobald, Ochlerotatus Lynch arribalzaga, and Neomelaniconion Newstead. Printed by Order of
the Trustees of the British Museum, UK
Mogi M, Chan KL. 1996. Predatory habits of dipteran larvae inhabiting Nepenthes pitchers. Raffles Bulletin
of Zoology,44 : 233-245
National Vector Borne Disease Control Programme Directorate General of Health Services Ministry of Health
& Family Welfare. Available on http://nvbdcp.gov.in/
Panicker KN, Bai MG, Sabesan S. 1982. A note on laboratory colonization of Culex (Lutzia) fuscanus
78

Arthropods, 2014, 3(1): 70-79
IAEES www.iaees.org
Wiedemann, 1820 (Diptera : Culicidae). Indian Journal of Medical Research, 75: 45-46
Poopathi S, Tyagi BK. 2006. The Challenge of Mosquito Control Strategies: from Primordial to Molecular
Approaches. Biotechnology and Molecular Biology Review, 1(2): 51-65
Ramalingam S, Ramakrishnan K. 1971. Re-description of Aedes (Alanstonea) Brevitibia (Edwards) from
Brunei, Borneo. Proceedings of Entomological Society Washington, 73: 231-238
Rattanarithikul R, Harrison BA, Panthusiri P, et al. 2005. Illustrated keys to the mosquitoes of Thailand 1.
Background; geographic distribution; lists of genera, subgenera, and species; and a key to the genera.
Southeast Asian Journal of Tropical Medicine and Public Health, 36: 1-80
Roberts DW, Strand A. 1977. Pathogens of medically important arthropods. WHO Bulletin, 55 (Suppl I): 419
Sempala SDK. 1983. Interactions between immature Aedes africanus (Theobald) and larvae of two predatory
Toxorhynchites (Diptera: Culicidae) in Zika forest, Uganda. Bulletin of Entomological Research, 73: 19-
24
Sharma K, Angel B, Singh H, et al. 2008. Entomological studies for surveillance and prevention of dengue in
Arid and Semi-Arid District of Rajasthan India. Journal of Vector Borne Disease, 45: 140-149
Shukla RP, Kohli VK, Ojha VP. 1997. Larvicidal efficacy of Bacullus sphaericus H5a, 5b and B. thuringiensis
var israelensis H-14 against malaria vectors in Bhabar area, District Nainital, UP. Indian of Journal
Malariology, 34(4): 208-212
Singh H, Marwal R, Mishra A, Singh KV. 2013. Invasion of the predatory mosquito Culex (Lutzia)fuscanus in
the western desert parts of Rajasthan, India. Poland Journal of Entomology, 82: 49-58
Singh K, Kumar K, Biswas S. 1984. Laboratory studies on the predacious efficacy of Culex (Lutzia) fuscanus
larvae against other mosquito larvae. Journal of Communicable Disease, 16(4): 320-322
Sinnathamby NS, Pavilupillai JJ, Arulanantham CT, et al. 2013. Predatory Efficacy of Culex (Lutzia) fuscanus
on Mosquito Vectors of Human Diseases in Sri Lanka. Journal of American Mosquito Control Association,
29(2): 168-170
Tanaka K. 2003. Studies on the pupal mosquitoes of Japan (9). Genus Lutzia, with establishment of two new
subgenera, Metalutzia and Insulalutzia (Diptera, Culicidae). Japanese Journal of Systematic Entomology,
159-169
Thangam TS, Kathiresan K. 1996. The prey consumption and prey preference of the larvae of the mosquito
Culex (Lutzia) raptor on the larvae of Culex quinquefasciatus. Experientia, 52(4): 380-382
World Health Organization. Available on http://www.who.int/topics/en/
Yanovisk SP. 2001. The macrofauna of water-filled tree holes on Barro Colorado Island, Panama. Biotropica,
33(1): 110-120
79

Arthropods, 2014, 3(1): 80-87
IAEES www.iaees.org
Article
Diversity of damselflies (Zygoptera) in Gorewada International Bio-
Park, Nagpur, Central India
Patil Kishor Gopal1, Shende Virendra Abaji2, Uke Shrikant Bhimrao3 1Department of Zoology, Institute of Science, R. T. Marg, Nagpur (M.S.) India 2K. Z. S. Science College, Bramhani-Kalmeshwar, Dist- Nagpur (M.S.) India 3Office of Superintendent of Police, Railways, Nagpur (M.S.) India
E-mail: [email protected]
Received 26 September 2013; Accepted 30 November 2013; Published online 1 March 2014
Abstract
Gorewada International Bio-Park consists of a lake as a major water source, marshy shore and heterogeneity in
vegetation. Its geographical location is 21°11′N 79°2′E. Observations are made through walking line transects
along the lake border to determine the diversity of damselfly. Total 21 species of damselflies belonging to nine
genera (Aciagrion, Agriocnemis, Ceriagrion, Enallagma, Ischnura, Pseudagrion, Rhodischnura, Copera and
Lestes) and three families (Coenagrionidae, Lestidae and Platycnemididae) have been recorded. Out of total
damselflies examined, 52.38% are common, 19.05% are occasional and 28.57% are rare species. The present
study encourages the conservation of a wide range of indigenous damselfly species in this area.
Keywords Damselfly; Zygoptera; Odonata; Insects; Gorewada.
1 Introduction
The earth’s (approximately 3.8 billion year) history of life is representing just 0.1% of all the species of flora
and fauna that have presently lived on earth. Thus 99.9% or virtually all of life that has existed on earth has
gone extinct (Raup, 1991).
Damselflies and dragonflies are among the most attractive creatures on earth belonging to the most popular
insect order- Odonata. These are observed near the ponds, lakes, rivers, ditches and all over the marshy places.
Damselflies (suborder- Zygoptera) have front and hind wings are similar in shape with narrowed base. The
wings of the two sexes are similar in shape. At resting condition wings are held either together above the body
or slightly divergent. The head is elongate transversely and in dorsal view is usually wider than the thorax.
Damselflies and dragonflies can be traced back to the Carboniferous and Permian periods of the Paleozoic
Era (500-200 million years ago). However, modern families of these insects date from the upper Jurassic and
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2014, 3(1): 80-87
IAEES www.iaees.org
Cretaceous periods (150-60 million years ago) (Westfall and May, 1996). Silsby (2001) described about 6000
species of dragonflies in all over the world. India is also highly diverse with more than 500 species of Odonata
(Subramanian, 2005). Total almost 1,100 species, Coenagrionidae is the largest family of damselflies, forming
a major part of the odonate fauna in all continents. With Lestidae, it is the only damselfly family of which
many species inhabit standing waters (Dijkstra and Kalkman, 2012). Subramanian (2009) recorded 267 species
diversity of damselfly belonging to 87 genera and 8 families in India.
Gorewada is developing International Bio-Park situated at North-West of Nagpur city and its geographical
location is 21°11′N 79°2′E (Fig. 1 and 2). It is basically divided into African Safari, Bio-Park, Energy Plaza,
Trails, Indian Safari, Height Safari, Rescue Safari and Gorewada Reservoir. Gorewada Reservoir is bordered
by thick forest on three sides. Reservoirs catchment area is approx. 11 sq. mile (17,702.74 sq.mt.). Gorewada
is a good habitat for biodiversity of damselflies. In spite of its global significance, studies of damselflies
diversity of Gorewada International Park have been least undertaken.
Since, the main objective of this study has been conduct preliminary observation of damselflies and carried
out the checklist, occurrence and richness inhibiting the Gorewada International Bio-Park.
Fig. 1 Line transects along the Lake of Gorewada International Bio-Park (Courtesy- Google Map).
Fig. 2 Photograph showing ecology of Gorewada Lake.
81

Arthropods, 2014, 3(1): 80-87
IAEES www.iaees.org
2 Material and Methods
Watching and recording of damselflies has been done during Sunday and holidays in such a way that there
should be least one visit in each line transect during a week for a period of two years from March 2011 to
February 2013. Observations were made through walking transects of 0.7 km to 1.0 km length with 2 m to 5 m
on either side of the lake border. The present study is based on 4 line transects (Fig. 1) to study the damselflies
population. The sites were visited to note maximum species of damselflies and record its activities. The
observations were recorded with the aid of binocular and digital cameras and the species are identified with the
help of photographs, reference books and publications.
3 Results
Gorewada International Bio-Park provides heterogeneity in vegetation. The importance of the lake is as a
major water source and marshy shore. This lake is rich in aquatic fauna which includes micro and macro-
organisms, shrimps, fishes and protein-rich invertebrates. This region is suitable for feeding and resting for
many odonates due to the abundance of food throughout the year.
Table 1 Damselflies of Gorewada International Bio-Park, Nagpur, Central India.
S. N.
Species Common Name Status
Family: Coenagrionidae 1 Aciagrion hisopa(Selys, 1876) Violet-Striped Slender Dartlet O 2 Aciagrion occidentale (Laidlaw, 1919) Green-Striped Slender Dartlet O 3 Agriocnemis lacteola(Selys, 1877) Milky Dartlet R 4 Agriocnemis pygmaea (Rambur, 1842) Pigmy Dartlet C 5 Agriocnemis splendidissima (Laidlaw, 1919) Splendid Dartlet O 6 Ceriagrion coromandelianum (Fabricius, 1798) Coromandel Marsh Dart C 7 Ceriagrion olivaceum (Laidlaw, 1914) Rusty Marsh Dart R 8 Ceriagrion rubiae (Laidlaw, 1916) Orange Marsh Dart O 9 Enallagma parvum (Selys, 1876) Azure Dartlet C 10 Ischnura aurora (Brauer, 1865) Golden Dartlet C 11 Ischnura senegalensis (Rambur, 1842) Senegal Golden Dartlet C 12 Pseudagrion decorum (Rambur, 1842) Three-Lined Dart C 13 Pseudagrion indicum (Fraser, 1924) Yellow-Striped Blue Dart R 14 Pseudagrion microcephalum (Rambur, 1842) Blue Grass Dartlet R 15 Pseudagrion rubriceps (Selys, 1876) Saffron-Faced Blue Dart C 16 Rhodischnura nursei (Morton, 1907) Pixie Dartlet C Family: Platycnemididae 17 Copera marginipes (Rambur, 1842) Yellow Bush Dart C 18 Copera vittata (Selys, 1917) Blue Bush Dart C Family: Lestidae 19 Lestes elatus (Selys, 1862) Emerald Spreadwing R 20 Lestes umbrinus (Selys, 1891) Brown Spreadwing C 21 Lestes viridulus (Rambur, 1842) Emerald-Striped Spreadwing R
Abbreviations- C- Common; O- Occasional and R- Rare
Total 21 species of damselflies belonging to 9 genera and 3 families (Coenagrionidae, Platycnemididae,
Lestidae) have been recorded (Table 1). Coenagrionids are medium to small sized damselflies with transparent
wings which are rounded attips. The hindwings are shorter than abdomen with non-metalliccolors. Family-
82
Arthropods, 2014, 3(1): 80-87
IAEES www.iaees.org
Coenagrionidae consists of 16 species belong to 7 genera (Aciagrion, Agriocnemis, Ceriagrion, Enallagma,
Ischnura, Pseudagrion and Rhodischnura). Platycnemidids are black colored medium-sized damselflies with
yellow markings and wings are transparent with round-tips. Hindwings are shorter than the abdomen.
Platycnemididae family is consisting of 2 species belonging to single genus (Copera). Lestids are medium-
sized damselflies shows predominantly green or brown ground colored with iridescent markings. Family
Lestidaeis consists of 3 species belonging to single genus (Lestes) (Plate 1 and 2).
Family- Coenagrionidaeis the largest family carries the maximum number of genera and species followed
by Lestidae and Platycnemididae. Out of total 21 damselflies species examined, 11 (52.38%) are common, 4
(19.05%) are occasional and 6 (28.57%) are rare species (Table 2 and Fig. 3).
Table 2 Status of Damselflies of Gorewada International Bio-Park.
S.N. Status No. of species % of species 1. Common 11 52.38 2. Occasional 04 19.05 3. Rare 06 28.57 21 100
Fig. 3 Status of damselflies.
83

Arthropods, 2014, 3(1): 80-87
IAEES www.iaees.org
Plate 1
Figures showing diversity of damselflies. 1. Aciagrionhisopa (Violet-Striped Slender Dartlet); 2. Agriocnemis pygmaea (Pigmy Dartlet); 3.Ceriagrion coromandelianum (Coromandel Marsh Dart); 4. Ceriagrion rubiae (Orange Marsh Dart); 5. Enallagma parvum (Azure Dartlet); 6. Ischnura aurora (Golden Dartlet); 7. Pseudagrion decorum (Three-Lined Dart); 8. Pseudagrion indicum (Yellow-Striped Blue Dart).
84

Arthropods, 2014, 3(1): 80-87
IAEES www.iaees.org
Plate 2
Figures showing diversity of damselflies. 9. Pseudagrion rubriceps (Saffron-Faced Blue Dart); 10. Rhodischnura nursei (Pixie Dartlet); 11. Copera marginipes (Yellow Bush Dart); 12. Copera vittata (Blue Bush Dart), and 13. Lestes umbrinus (Brown Spreadwing).
85
Arthropods, 2014, 3(1): 80-87
IAEES www.iaees.org
Fig. 4 Distribution of genera and species of damselflies in respective families.
4 Discussion
Indian Agricultural Research Institute, New Delhi India, had a collection of 273 species of odonata, out of
which 125 species of damselflies were belonging to 10 families. Family- Coenagrionidae reported 38 species
while Lestidae and Platycnemididae reported 10 species each (Sharma et al., 2009). Subramanian et al. (2011)
had examined 8 families of damselflies belonging to 29 genera and 67 species, of which 25 were endemic. The
maximum number of species were reported in Family- Coenagrionidae (25 species) followed by protoneuridae
(15 species), Platystictidae and Lestidae (8 species), Euphaeidae (4 species), Chlorocyphidae and
Calopterygidae (3 species) and only two species were reported in Family-Platycnemididae of damselflies in the
Western Ghats. Manwar et al. (2012) had reported total 9 species of damselflies of which 8 species were
belonging to the family Coenagrionidae and single species from family Platycnemididae in Chatri Lake Region
of Pohara–Malkhed Reserve Forest, Amravati, Maharashtra (India).
However, during the months July 2010 to June 2011, Tijare and Patil (2012) were observed 8 species of
damselflies in and around Gorewada National Park, Nagpur; of which 5 species belonging to Family-
Coenagrionidae, 2 species from Lestidae and single species from Platycnemididae. These above observations
are similar with the present observations.
This study encourages the conservation of a wide range of indigenous damselfly species in an area.
References
Dijkstra DB, Kalkman VJ. 2012. Phylogeny, classification & taxonomy of European dragonflies &
damselflies (Odonata): A review. Organisms Diversity and Evolution, 12(3): 209-227
Manwar NA, Rathod PP, Raja IA. 2012. Diversity & abundance of dragonflies & damselflies of Chatri Lake
86

Arthropods, 2014, 3(1): 80-87
IAEES www.iaees.org
Region, in Pohara–Malkhed Reserve Forest, Amravati, Maharashtra (India). International Journal of
Engineering Research & Applications, 2(5): 521-523
Raup DM. 1991. Extinction: Bad Genes or Bad Luck? Life on Earth An Encyclopedia of Biodiversity,
Ecology & Evolution Volume-1, A-G. W.W. Norton, New York, USA
Sharma G, Ramamurthy VV, Kumar R. 2009. Collection of damselflies & dragonflies (Odonata :Insecta) in
National Pusa Collection, Division of Entomology, Indian Agricultural Research Institute, New Delhi,
India. Biological Forum, 1(2): 47-50
Silsby J. 2001. Dragonflies of the World. Natural History Museum in association with CSIRO Publishing, UK
Subramanian KA. 2005. Dragonflies & damselflies of Penisular India - A field guide. Project Lifescape. Indian
Academy of Sciences, Banglore, India
Subramanian KA. 2009. A Checklist of Odonata (Insecta) of India.
http://zsi.gov.in/checklist/Odonata_Indica_151209.pdf
Subramanian KA, Kakkassery F, Nair MV. 2011. The status & distribution of dragonflies & damselflies
(Odonata) of the Western Ghats. In: The Status & Distribution of Freshwater Biodiversity in the Western
Ghats, India (Molur, et al., eds). 63-71, IUCN, Cambridge, UK
Tijare RV, Patil KG. 2012. Diversity of Odonets in & around Gorewada National Park, Nagpur, M.S. (India).
Bionano Frontier, Special Issue 9:182-183
Westfall MJ, May ML. 1996. Damselflies of North America. Scientific Publishers, Gainesville, Florida, USA
87

Arthropods, 2014, 3(1): 88-95
IAEES www.iaees.org
Article
New indices for measuring some quality control parameters of the
Mediterranean fruit fly, Ceratitis capitata (Wied.) M. F. Mahmoud Plant Protection Department, Faculty of Agriculture, Suez Canal University, 41522, Ismailia, Egypt
E-mail: [email protected]
Received 1 September 2013; Accepted 5 October 2013; Published online 1 March 2014
Abstract
Even though the existence of interspecific competition and competitive displacement between the
Mediterranean fruit fly Ceratitis capitata (Wied.) and peach fruit fly Bactrocera zonata (Saunders) in the last
two decades in Egypt, Mediterranean fruit fly still occurs and threats many kinds of fruits and vegetables in
Egypt. The objective of this study was to estimate the sexual compatibility, mating performance and relative
sterility between laboratory and wild flies of the Mediterranean fruit fly, C. capitata by new indices (relative
mating index, RMI; relative isolation index, RII; isolation index, ISI; male relative performance index, MRPI;
female relative performance index, FRPI and relative sterility index, RSI). The results revealed that different
doses of gamma radiation 10, 30, 50, 70, and 90 Gy had no effect on the various parameters of mating
compatibility, performance and competitiveness of lab strain males of medflies when mated with wild males.
Moreover, no significant assortative or disassortative mating was observed. Therefore, we suggest that the lab
strain males of medfly are compatible of mating with the wild males, at least under the laboratory conditions
employed here.
Keywords sexual compatibility; mating performance; relative sterility; Ceratitis capitata.
1 Introduction
The Mediterranean fruit fly or medfly, Ceratitis capitata (Wiedemann) a colorful insect of the dipteran family
Tephritidae (Trypetidae), is considered one of the most destructive and important pests of edible fruits
worldwide (Weems, 1981; Liquido et al., 1991, 1998; Copeland et al., 2002). It was described by Wiedmann
in 1824 from the type specimen collected a board a ship in the Indian Ocean (1817).
In 1998, peach fruit fly or PFF, Bactrocera. zonata (Saunders) was recorded for the first time in Egypt
(Agamy and Sbahia, near Alexandria). Today, it is well established in most Egyptian provinces and it causes a
severe damage to a wide range of fruits (El-Minshawy et al., 1999). C. capitata and B. zonata causing a
Arthropods ISSN 22244255 URL: http://www.iaees.org/publications/journals/arthropods/onlineversion.asp RSS: http://www.iaees.org/publications/journals/arthropods/rss.xml Email: [email protected] EditorinChief: WenJun Zhang Publisher: International Academy of Ecology and Environmental Sciences

Arthropods, 2014, 3(1): 88-95
IAEES www.iaees.org
considerable damage in many fruit species (Ghanium, 2009). The existence of medfly and PFF in the same
hosts caused interspecific competition and competitive displacement between them. Most studies of
competition are descriptive: an increase in the population density of one species co-occurs with a decrease in
another (Connell, 1983; Schoener, 1983, 1985). Therefore, attention of both researches and farmers tend to
control the peach fruit fly only, although sticky traps and infested fruits recorded numbers of medfly in
different regions of Egypt.
Over the last decades, aerial malathion-based bait-spray application has been the most common and
effective control tool against exotic fruit flies (USDA, 1993). The spray consists of the organophosphorous
insecticide malathion mixed with a protein hydrolysate acting as an attractant and feeding stimulant. Malathion
is highly toxic to terrestrial and aquatic non target invertebrates. In fact, a malathion bait spray may disrupt a
substantial portion of natural biological control agents (USDA, 2001). In Egypt, medfly and PPF populations
are typically managed by trapping, partial or complete spraying, bait spraying/collection and deep burial of
fallen fruits. These methods reduce but not sufficiently eliminate its populations.
The Sterile Insect technique (SIT) is a promising, environmentally friendly, methodology for control or
eradication of insect pests. The effectiveness of (SIT) depends on the quality and the ability of sterile
laboratory males to search for females, mating compatibility and effective competitiveness with wild males
(Orozco et al., 2007).
In the present study, sexual compatibility, mating performance and relative sterility between sterile
laboratory males and wild flies of the Mediterranean fruit fly, C. capitata was measured by new indices.
2 Materials and Methods
2.1 Wild strain
Wild flies were collected from infested mandarin fruits in El-Sharkia Governorate. Infested fruits were
collected from the host plant and the ground under the trees then distributed in plastic trays (40 cm diameter)
furnished with fine sand to let the larvae mature. The full grown larvae raised out "popping" to pupate in the
fine sand. The pupae were daily collected and pupae in the same age were kept in separate Petri-dishs (10 cm
diameter). Then pupae were kept in adult cages till eclosion. The eclosed peach fruit flies were removed from
cages by aspiration.
2.2 Laboratory strain
The old strain of Medfly was maintained under laboratory conditions 25 ± 2oC, 65 – 75% RH and a
photoperiod of 12/12h (L/D) in the Plant Protection Department, Faculty of Agriculture, Suez Canal
University. The adults were maintained in a cage (75 x 28 x 28 cm) coated with muslin fabric and placed over
a plastic dish (40 cm in a diameter) filled with tap water (1 liter). Enclosed adults were provided with a diet
consisted of 1 part of protein hydrolysate and 3 parts of sugar by weight. A water soaked cotton clump in a
small cup served as a water source. Larvae were reared in plastic trays (15 × 5 × 3 cm) half-filled with an
artificial diet. The larval diet consisted of wheat bran (1000 g), brewer’s yeast (250 g), sugar (250 g), sodium
benzoate (10 g), hydrochloric acid (10 ml) and water (2000 ml) (Mahmoud et al., 2013).
2.3 Sexual compatibility
Samples of pupae from lab strain were irradiated 2 days before adult emergence with doses of 10, 30, 50, 70
and 90 Gy using a 60Co source (Gammacell irradiator, model 4000A) located at the Atomic Energy Authority,
Gamma Irradiation Unit, Naser City, Cairo, Egypt. The gamma irradiation was carried out at a dose rate of 66
Gy/min. Gamma cell calibrated by using alanine reference dosimeter to verify the dose delivered to the pupae.
5 males, 5 females of the tested wild flies and 5 sterile males, 5 sterile females of lab strain were caged in
small cage measuring 20 × 20 × 20 cm (each group in a separate cage). In order to identify the sterile males
89

Arthropods, 2014, 3(1): 88-95
IAEES www.iaees.org
and sterile females, few amount of fluorescent dye was applied onto the surface of the pupae, which is then
transferred to the teneral adult upon emergence. Throughout the observation period the numbers of matings
were recorded as wild male and female (WW), sterile male and female (SS), wild male and sterile female (WS),
and sterile male and wild female (SW). The tests were conducted with 5 replicates for each dose of irradiation.
Each cage contained sugar and protein hydrolysate at the ratio of 3:1 by weight and source of water.
2.4 Sexual competitiveness
To measure the relative sterility index (sexual competitiveness) between the previously prepared sterile lab
males at 10, 30, 50, 70 and 90 Gy and wild males toward wild female: five competitiveness cages (25 × 25 ×
30 cm) were prepared into each cage 15 sterile lab males: 5 wild males: 5 wild females were put. Each cage
contained the previous adult diet. The number of matings was recorded as sterile lab male and wild female
(SW) and wild male and wild female (WW).
2.5 Statistical analysis
To estimate sexual compatibility, the index of sexual isolation (ISI), male and female relative performance
indices MRPI and FRPI, and the relative sterility index (RSI) (Calkins and Parker 2005) were calculated. We
used the 0.25 value as variance limit for equal mating propensity in ISI, MRPI, and FRPI, and for equal
competitiveness in the RSI. Data obtained in all presented experiments were subjected to an analysis of
variance (ANOVA) with the honestly significant difference value calculated as Tukey’s statistic at α = 0.05
(SAS Institute, 2002).
3 Results
The results obtained from the mating compatibility test are shown in Table 1. The relative mating index (RMI)
indicates the overall percentage of the couples that mated. If the competitiveness of the males were equal then
0.25 of the mating pairs would be from each of the treatment groups. All the RMI values were larger than 0.25,
indicating good mating performance (FAO/IAEA/USDA, 2003). In this experiment RMI was 0.82 at 50 Gy,
0.80 at 10 and 70 Gy, 0.78 at 90 Gy and 0.68 at 30 Gy. The relative isolation index (RII) gives an indication of
mating compatibility between wild and sterile laboratory strain. A value of 1 indicates random mating, with
larger values indicating assortative mating. The value of RII can be interpreted as the number of sterile males
that have to be employed to be equivalent to one wild male. Values normally vary between 1.5 and 5, and
values consistently above 3 are a cause for concern. Results in Table 1 show values of RII range from 1.68 to
3.65. This values indicate that satisfactory levels and there was no significant difference among populations in
the experiment (F = 1.6212; P = 0.2079). The index of sexual isolation (ISI) is a measure of mating
compatibility between populations. The index considers the number of couples obtained for each possible
mating combination, with values range from -1 (complete negative assortative mating, that is, all mating are
with members of the opposite population) through 0 (random mating) to +1 (complete positive assortative
mating, that is, total mating isolation of the two populations) (Fig. 1). The ISI values (Fig. 2) show good levels
of compatibility between the sterile flies and the different wild populations, and there was no significant
difference among populations (F = 0.6036; P = 0.6645). The relative sterility index (RSI) indicates the sexual
competitiveness between two strains. Values range between 0 and +1. Zero means that wild females mate only
with wild males; a value of +0.5 indicate that wild females mate indiscriminately with wild or sterile males; a
value of +1 indicate that wild females mate only with sterile males. The RSI in most cases reflected the
preference of wild females for wild males over sterile males. Data shows no significant difference between lab
and wild flies (F = 0.2494; P = 0.9066). The male relative performance index (MRPI) is a measure of the
propensity of sterile males to mate with wild females, with values ranking from -1 to +1. A value of -1
indicates that all matings were carried out by wild males, while a value of +1 indicates that all matings were
90

Arthropods, 2014, 3(1): 88-95
IAEES www.iaees.org
carried out by sterile males. Zero indicates that males from both populations participated equally in matings.
Fig. 3 shows that the sterile males participated equally at obtaining mates with wild males and there was no
differences between the populations (F = 1.0005; P = 0.4304). The female relative performance index (FRPI)
is a measure of the propensity of sterile females to mate with wild males, with values ranking from -1 to +1. A
value of -1 indicates that all matings were carried out by wild females, while a value of +1 indicates that all
matings were carried out by sterile females. Zero indicates that females from both populations participated
equally in mating. Fig. 4 shows that the sterile females were equal at obtaining mates with wild females and
there was no overall differences between the populations (F = 1.1416; P = 0.3656). Data in Fig. 3 and Fig. 4
suggests that the sterile males and sterile females were participated equally in matings when competing against
wild flies of medfly populations
Table 1 Relative mating index (RMI), relative isolation index (RII), isolation index (ISI), relative sterile index (RSI), male relative performance index (MRPI) and female relative performance index (FRPI) obtained from irradiated lab strain mated with wild flies.
Radiation dose(Gy)
RMI RII ISI RSI MRPI FRPI
10 0.80 3.65 ± 2.147 a 0.281 ± 0.189 a 0.383 ± 0.370 a -0.124 ± 0.208 a -0.024 ± 0.143 a30 0.68 1.68 ± 0.535 a 0.174 ± 0.089 a 0.499 ± 0.227 a -0.231 ± 0.245 a -0.028 ± 0.100 a50 0.82 2.28 ± 1.156 a 0.168 ± 0.123 a 0.449 ± 0.182 a -0.084 ± 0.268 a -0.044 ± 0.060 a70 0.80 2.30 ± 1.254 a 0.223 ± 0.080 a 0.316 ± 0.123 a -0.162 ± 0.221 a -0.192 ± 0.050 a90 0.78 2.06 ± 0.855 a 0.187 ± 0.157 a 0.488 ± 0.197 a 0.040 ± 0.178 a -0.053 ± 0.266 aF 1.6212 0.6036 0.2494 1.0005 1.1416 P < 0.1 < 0.1 < 0.1 < 0.1 < 0.1 RMI (Relative mating index) = No. of pairs collected/No. of females released. RII (Relative isolation index) = (SS×WW) / (SW×WS). ISI (Isolation index) = (SS+WW)-(SW+WS) / (SS+WW+SW+WS). RSI (Relative sterile index) = (SW) / (SW+WW). MRPI (Male relative performance index) = (SS+SW)-(WS+WW) / (SS+WW+SW+WS). FRPI (Female relative performance index) = (SS+WS)-(SW+WW) / (SS+SW+WS+WW). Data (± SD) followed by the same letter do not differ significantly according to Tukey’s HSD test.
Fig. 1 Index of sexual isolation comparing the compatibility of the sterile flies with the wild flies.
91

Arthropods, 2014, 3(1): 88-95
IAEES www.iaees.org
Fig. 2 Male mating competitiveness between sterile flies and wild flies.
Fig. 3 Male relative performance index between wild flies with the sterile flies.
Fig. 4 Female relative performance index between wild flies with the sterile flies.
92

Arthropods, 2014, 3(1): 88-95
IAEES www.iaees.org
4 Discussion
The sterile insect technique (SIT) is a specific control method that may be applied in the area-wide integrated
pest management of insect pests of medical, veterinary, and agricultural importance. Contrary to chemical and
biological products, sterile insects are non-invasive agents rather than intrusive toxic, pathogenic or otherwise
destructive entities. Therefore, the SIT alone poses a priori an exceptionally low risk to the environment
(Hendrichs, 2001). SIT is widely used to suppress infestations of the Mediterranean fruit fly (medfly), C.
capitata (Wied.) (Hendrichs et al., 2002). The success of an SIT program depends largely on the ability of
sterile lab males to obtain matings with wild males.
Mating compatibility, mating performance and relative sterility are important quality control parameters
that affect the performance of released sterile insects. The present study showed the indices of mating
compatibility and mating competitiveness between wild and sterile population of medfly at different doses of
gamma radiation from 10 to 90 Gy. Wild flies may expose constantly to the natural environmental conditions,
while laboratory flies may expose to fairly stable environmental conditions. This differences cause some
changes in the behavior of laboratory flies. Attempts to obtain 100% sterility in males usually reduce quality,
and it will often be better to reduce the dose so as to obtain a better induction of sterility in the field females by
giving a more competitive male (Toledo et al., 2004; Dyck et al., 2005).
Laboratory tests of mating compatibility and competitiveness may not reliably indicate the situation in the
field (Katsoyannos et al., 1999). A much better measure can be obtained by using a field cage. Standered
procedure for field-cage operation have been defined (Cayol et al., 2002; FAO/IAEA/USDA 2003). A field
cage consists of a mesh cage, 2 m high and 3 m wide, erected over a host plant or other suitable vegetation.
But, it is usually not possible to detect mating pairs directly in the field. Therefore, in this present study, all
experiments of compatibility and competitiveness carried out in lab-cage similar to the field cage but smaller
in size (20 cm high and 20 cm wide) under laboratory conditions.
Data of mating compatibility and competitiveness indices (mating index, RMI; relative isolation index, RII;
isolation index, ISI; male relative performance index, MRPI; female relative performance index, FRPI and
relative sterility index, RSI) demonstrated good sexual compatibility between wild flies and mass reared lab
flies. Results of RMI revealed satisfactory mating propensity between wild and sterile lab flies, because of all
values were larger than 0.25 of observed matings. The RII showed good indication of mating compatibility
between wild and sterile lab flies. Values at all doses of gamma radiation range from 1.68 to 3.65 and indicate
assortative mating. The ISI showed good mating compatibility between wild and sterile lab populations at all
doses of gamma radiation (10, 30, 50, 70 and 90 Gy). Values of MRPI revealed that the sterile lab males which
produced from pupae irradiated with different of gamma doses (10, 30, 50 and 70 Gy) are almost equal mating
propensity with wild males. However, sterile lab males produced from pupae irradiated with 90 Gy were as
effective in copulating with wild females as the wild males. Values of FRPI showed that the sterile lab females
are almost equal mating propensity with wild males and the wild females.
Data in the present study indicate that doses of gamma radiation had no effect on the various parameters of
mating compatibility, performance and competitiveness of lab males of medflies when mated with wild males.
These results can be interpreted in two ways. First, irradiation generally may have no effect on the mating
competitiveness of male medflies. Alternately, irradiation may negatively affect mating ability, but its impact
may vary with the general ‘vigor’ of the flies irradiated, being greater for individuals of low quality.
The satisfactory levels of compatibility and the good competitiveness of irradiated lab males and wild
males observed in this study may encourage the application of the SIT to control C. capitata populations.
93

Arthropods, 2014, 3(1): 88-95
IAEES www.iaees.org
References
Cayol JP, Coronado P, Taher M. 2002. Sexual compatibility in Medfly (Diptera: Tephritidae) from different
origins. Florida Entomologist, 85: 51-57
Calkins CO, Parker AG. 2005. Sterile insect quality. In: Sterile Insect Technique: Principles and Practice in
Area-wide Integrated Pest Management (Dyck VA, Hendrichs J, Robinson AS, eds). 269-296, Springer,
Dordrecht, Netherlands
Connell JH. 1983. On the prevalence and relative importance of interspecific competition: Evidence from field
experiments. American Naturalist, Ill: 661-696
Copeland R, Wharton R, Luke Q, et al. 2002. Indigenous Hosts of Ceratitis capitata (Diptera:Tephritidae) in
Kenya. Annals of the Entomological Society of America, 95(6): 672-694
Dyck VA, Hendrichs J, Robinson AS. 2005. The Sterile Insect Technique: Principles and Practice in Area-
Wide Integrated Pest Management. Springer, Dordrecht , Netherlands
El–Minshawy AM, El–Eryan MA, Awad AI. 1999. Biological and morphological studies on the guava fruit fly
Bactrocera zonata (Saunders) (Diptera: Tephritidae) found recently in Egypt. 8th National Conference of
Pests and Diseases of Vegetables and Fruits in Ismailia. 71-82, Egypt
FAO/IAEA/USDA, 2003. Manual for Product Quality Control and Shipping Procedures for Sterile Mass-
Reared Tephritid Fruit Flies. Version 5.0. International Atomic Energy Agency, Vienna, Austria
Ghanim NM. 2009. Studies on the peach fruit fly, Bactrocera zonata (Saunders) (Tephritidae, Diptera). Ph. D.
Thesis, Mansoura University, Egypt
Hendrichs J. 2001. The sterile insect technique worldwide, In: Proceedings, Seminar, Madeira Med: Sterile
Insect Technique as an Environmentally Friendly and Effective Insect Control System. 25-53, Madeira
Regional Direction of Agriculture, Portugal
Hendrichs J, Robinson AS, Cayol JP, et al. 2002. Medfly area-wide sterile insect technique programmes for
prevention, suppression or eradication: the importance of mating behavior studies. Florida Entomologist,
85: 1-13
Katsoyannos BI, Papadopoulos NT, Hendrichs J, et al. 1999. Comparative response to citrus foliage and citrus
fruit odour by wild and mass reared sterile Mediterranean fruit fly males of a genetic sexing strain. Journal
of Applied Entomology, 123: 139-143
Liquido NJ, Barr PG, Cunningham RT. 1998. An encyclopedic bibliography of the host plants of the
Mediterranean fruit fly, Ceratitis capitata (Wiedemann). USDA-ARS-144, version 1, cd database, USDA,
USA
Liquido NJ, Shinoda LA, Cunningham RT. 1991. Host Plants of the Mediterranean Fruit Fly (Diptera:
Tephritidae): An Annotated World Review. Miscellaneous Publications of the Entomological Society of
America, MPPEAL 77: 1-52
Mahmoud MF, Shoukry A, Khalil D. 2013 Behavioral and Ecological Studies Mediterranean Fruit Fly.
Lambert Academic Publishing, Germany
Orozco-Davila D, Hernandez R, Meza S. et al. 2007. Sexual competitiveness and compatibility between mass-
reared sterile flies and wild populations Anastrepha ludens (Diptera: Tephritidae) from different regions in
Mexico. Florida Entomologist, 90(1): 19-26
SAS Institute Inc. 2002. SAS/STAT User’s guide, version 9.1. SAS Institute Inc. Cary, North Carolina, USA.
Schoener TW. 1983. Field experiments on interspecific competition. American Naturalist, 122: 240-285
Schoener TW. 1985. Some comments on Connell's and my reviews of field experiments on interspecific
competition. American Naturalist, 125: 730-740
Toledo J, Rull J, Oropeza A, et al. 2004. Irradiation of Anastrepha oblique (Diptera: Tephritidae) revisited:
94

Arthropods, 2014, 3(1): 88-95
IAEES www.iaees.org
optimizing sterility induction. Journal of Economic Entomology, 97: 383-389
United States Department of Agriculture (USDA). 1993. Medfly cooperative eradication program final
environmental impact statement. United States Department of Agriculture, Marketing and Regulatory
Programs, Animal and Plant Health Inspection Service, Riverdale, MD, USA
United States Department of Agriculture (USDA). 2001. Fruit fly cooperative control program final
environmental impact statement. United States Department of Agriculture, Marketing and Regulatory
Programs, Animal and Plant Health Inspection Service, Riverdale, MD, USA
Weems HVJr. 1981. Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera; Tephritidae).
Entomology Circular No. 230. Florida Department of Agriculture & Consumer Services, USA
95

Arthropods
Arthropods account for more than 65% of global species and 85% of animal species. On a temperate grassland,
arthropods hold a huge biomass (1,000kg/ha), seconded to plant (20,000kg/ha) and microorganisms (7,000kg/ha)
but much higher than mammals (1.2kg/ha), birds (0.3kg/ha), and nemantodes (120kg/ha). Arthropods play the
role of both pests and beneficial organisms. Some arthropods are important crop pests but others are natural
enemies. Some arthropods are important health pests but many crustaceans are important food sources of
humankinds. Arthropods govern the structures and functions of natural ecosystems, but are always ignored by
researchers. On the global scale, the surveys of mammals, birds and vascular plants were relatively perfect
because they were economically important and easily surveyed. However, arthropods, despite their ecological
and economical importance, have not yet been fully surveyed and recorded due to their difficulties to be
sampled. The research on arthropods must be further promoted. The journal, Arthropods, is inaugurated to
provide a public and appropriate platform for the publication of studies and reports on arthropods.
ARTHROPODS (ISSN 2224-4255) is an international journal devoted to the publication of articles on various
aspects of arthropods, e.g., ecology, biogeography, systematics, biodiversity (species diversity, genetic diversity,
et al.), conservation, control, etc. The journal provides a forum for examining the importance of arthropods in
biosphere (both terrestrial and marine ecosystems) and human life in such fields as agriculture, forestry, fishery,
environmental management and human health. The scope of Arthropods is wide and embraces all
arthropods-insects, arachnids, crustaceans, centipedes, millipedes, and other arthropods. Articles/short
communications on new taxa (species, genus, families, orders, etc.) and new records of arthropods are
particularly welcome.
Authors can submit their works to the email box of this journal, [email protected]. All manuscripts
submitted to Arthropods must be previously unpublished and may not be considered for publication elsewhere
at any time during review period of this journal.
In addition to free submissions from authors around the world, special issues are also accepted. The organizer of
a special issue can collect submissions (yielded from a research project, a research group, etc.) on a specific
topic, or submissions of a conference for publication of special issue.
Editorial Office: [email protected]
Publisher: International Academy of Ecology and Environmental Sciences
Address: Flat C, 23/F, Lucky Plaza, 315-321 Lockhart Road, Wanchai, Hong Kong
Tel: 00852-6555 7188
Fax: 00852-3177 9906
E-mail: [email protected]

Arthropods ISSN 2224-4255 Volume 3, Number 1, 1 March 2014 Articles Epigeus macroinvertebrates species assemblages along a successional gradient in Hailuotu
Island (Bothnia Bay), Finland
Adolfo A. Del Rio Mora 1-19
Mineral composition of edible crab Podophthalmus vigil Fabricius (Crustacea: Decapoda)
P. Soundarapandian, D. Varadharajan, S. Ravichandran 20-26
The effects of some domestic pollutants on the cumacean (Crustacea) community structure
at the coastal waters of the Dardanelles, Turkey
A.Suat Ates, Tuncer Katagan, Murat Sezgin, et al. 27-42
Redescription and new distributional records of Matuta planipes (Fabricius, 1798)
(Crustacea; Decapoda; Matutidae) from Chennai Coast, Tamil Nadu
K. Silambarasan, K. Velmurugan, E. Rajalakshmi, et al. 43-47
Checklist of the subfamilies Mirinae and Orthotylinae (Hemiptera: Heteroptera: Miridae)
in western parts of Kerman Province, Iran
Mohsen Shamsi, Reza Hosseini, Asghar Shirvani 48-56
A study on the genus Orthops FIEBER (Hemiptera: Miridae: Mirinae) in Iran
Reza Hosseini 57-69
Predatory habits of Lutzia (Metalutzia) fuscana (Wiedmann) (Diptera: Culicidae) in the arid
environments of Jodhpur, western Rajasthan, India
Himmat Singh, Robin Marwal, Anusha Mishra, et al. 70-79
Diversity of damselflies (Zygoptera) in Gorewada International Bio-Park, Nagpur, Central
India
Patil Kishor Gopal, Shende Virendra Abaji, Uke Shrikant Bhimrao 80-87
New indices for measuring some quality control parameters of the Mediterranean fruit fly,
Ceratitis capitata (Wied.)
M. F. Mahmoud 88-95
IAEES http://www.iaees.org/




















