
New bradoriid arthropods from the Giles Creek Dolostone (Cambrian Series 3, Stage 5, Templetonian),...
16
New bradoriid arthropods from the Giles Creek Dolostone (Cambrian Series 3, Stage 5; Templetonian), Amadeus Basin, central Australia PATRICK M. SMITH, GLENN A. BROCK, JOHN R. PATERSON & TIMOTHY P. TOPPER BRADORIIDS are an important group of Cambrian to early Ordovician (Tremadocian) marine bivalved arthropods. Increasing knowledge of bradoriid systematics, biogeography and biostratigraphy has brought the group to greater prominence in Cambrian faunal investigations (Gozalo & Hinz-Schallreuter 2002; Hinz-Schallreuter et al. 2007; Topper et al. 2011; Williams et al. 2007; Williams et al. 2011). Recent documentation of bradoriids from Cambrian Series 2 (Stages 3-4) successions in South Australia (broadly equivalent to the Atdabanian-Toyonian using the Russian stages) has revealed Australia as a diversity hotspot for the group (Skovsted et al. 2006; Topper et al. 2007, 2011; Betts et al. 2014), though this may be an artifact of recent increased sampling and documentation of faunas. A recent review by Jones & Kruse (2009) reported increasing diversity of bradoriids from stratigraphic successions straddling Cambrian Series 2-3, Stages 4-5 boundary (approximately equivalent to the traditional Lower-Middle Cambrian boundary) from localities across central and northern Australia. This interval approximates the Ordian to Mindyallan stages using the Australian stage regional subdivisions (Peng et al. 2012). Most Ordian (approximately equivalent to the Series 2-3 boundary) bradoriids from central Australia have previously been reported to belong to three families, the Bradoriidae Matthew 1902, Comptalutidae Öpik 1968a and Hipponicharionidae Sylvester-Bradley 1961 (Jones & Laurie 2006) and are derived from sequence 1 successions (Southgate & Shergold 1991) in the Georgina, Wiso and Daly basins. The succeeding Templetonian (Series 3, Stage 5) formations from Georgina Basin sequence 2 (Southgate & Shergold 1991) contain some of the richest bradoriid faunas, in particular from the Beetle Creek Formation (Monastery Creek Phosphorite Member) in the Georgina Basin. At least four families, possibly a fifth, are represented within this formation. These include: Svealutidae Öpik 1968a, Comptalutidae Öpik 1968a, Monasteriidae Jones & McKenzie 1980, Hipponicharionidae Ulrich & Bassler 1931 and possibly Beyrichonidae Ulrich & Bassler 1931, which may be represented by the species Galeopsis glabra Hinz-Schallreuter 1993a. Younger formations across central Australia have, by comparison, much lower bradoriid diversity (Jones & Kruse 2009). Despite numerous reports of fossils from stratigraphic units within the Pertaoorrta Group in the Amadeus Basin (see overviews by Shergold 1986; Shergold et al. 1991), there is a surprising dearth of published palaeontological data, especially biostratigraphic data, from this sedimentary succession. Given the ubiquitous presence of Cambrian bradoriids in almost all other central Australian basins, the occurrence of bradoriids in the Amadeus Basin has long been suspected, and recent work by Smith (2012) has demonstrated their presence. The main aim of this contribution is to provide the first systematic analysis and biostratigraphic range data for bradoriid assemblages from the Cambrian (Series 3, Stage 5; Templetonian) Giles Creek Dolostone in the Ross River Gorge area in the NE Amadeus SMITH, P.M., BROCK, G.A., PATERSON, J.R. & TOPPER, T.P., 2014:04:25. New bradoriid arthropods from the Giles Creek Dolostone (Cambrian Series 3, Stage 5; Templetonian), Amadeus Basin, central Australia. Memoirs of the Association of Australasian Palaeontologists 45, 233-248. ISSN 0810-8889. A new assemblage of bradoriids is reported from the Giles Creek Dolostone (Cambrian Series 3, Stage 5; Templetonian) in the Ross River Gorge area, NE Amadeus Basin, central Australia. One new genus and four new species are described: Annge iperte gen. et sp. nov., Flemingopsis anteriospinata sp. nov., Monasterium ferox sp. nov. and Mongolitubulus tunpere sp. nov. Only M. tunpere sp. nov. may have been described previously from adjacent basins. This study provides the first systematic description of a Cambrian bradoriid assemblage from the Amadeus Basin. P.M. Smith ([email protected]) and G.A. Brock ([email protected]) Department of Biological Sciences, Macquarie University, Sydney, New South Wales, 2109, Australia. J.R. Paterson ([email protected]) School of Environmental and Rural Science, University of New England, Armidale, New South Wales, 2351, Australia. T.P. Topper (timothy. [email protected]) Geological Museum, Natural History Museum of Denmark, University of Copenhagen, Øster Voldgade 5-7, DK-1350 Copenhagen K, Denmark and Department of Earth Sciences, Palaeobiology, Uppsala University, Villavägen 16, SE- 752 36 Uppsala, Sweden. Received 4 October 2013. Keywords: Bradoriida, bivalved arthropod, new taxa, Annge, Flemingopsis, Monasterium, Mongolitubulus, Northern Territory, Gondwana.
-
Upload
australianmuseum -
Category
Documents
-
view
0 -
download
0
Transcript of New bradoriid arthropods from the Giles Creek Dolostone (Cambrian Series 3, Stage 5, Templetonian),...

New bradoriid arthropods from the Giles Creek Dolostone (Cambrian Series 3, Stage 5; Templetonian), Amadeus Basin, central Australia
PATRICK M. SMITH, GLENN A. BROCK, JOHN R. PATERSON &TIMOTHY P. TOPPER
BRADORIIDS are an important group of Cambrian to early Ordovician (Tremadocian) marine bivalved arthropods. Increasing knowledge of bradoriid systematics, biogeography and biostratigraphy has brought the group to greater prominence in Cambrian faunal investigations (Gozalo & Hinz-Schallreuter 2002; Hinz-Schallreuter et al. 2007; Topper et al. 2011; Williams et al. 2007; Williams et al. 2011). Recent documentation of bradoriids from Cambrian Series 2 (Stages 3-4) successions in South Australia (broadly equivalent to the Atdabanian-Toyonian using the Russian stages) has revealed Australia as a diversity hotspot for the group (Skovsted et al. 2006; Topper et al. 2007, 2011; Betts et al. 2014), though this may be an artifact of recent increased sampling and documentation of faunas. A recent review by Jones & Kruse (2009) reported increasing diversity of bradoriids from stratigraphic successions straddling Cambrian Series 2-3, Stages 4-5 boundary (approximately equivalent to the traditional Lower-Middle Cambrian boundary) from localities across central and northern Australia. This interval approximates the Ordian to Mindyallan stages using the Australian stage regional subdivisions (Peng et al. 2012). Most Ordian (approximately equivalent to the Series 2-3 boundary) bradoriids from central Australia have previously been reported to belong to three families, the Bradoriidae Matthew 1902, Comptalutidae Öpik 1968a and Hipponicharionidae Sylvester-Bradley 1961 (Jones & Laurie 2006) and are derived from sequence 1 successions (Southgate & Shergold 1991) in the Georgina,
Wiso and Daly basins. The succeeding Templetonian (Series 3, Stage 5) formations from Georgina Basin sequence 2 (Southgate & Shergold 1991) contain some of the richest bradoriid faunas, in particular from the Beetle Creek Formation (Monastery Creek Phosphorite Member) in the Georgina Basin. At least four families, possibly a fifth, are represented within this formation. These include: Svealutidae Öpik 1968a, Comptalutidae Öpik 1968a, Monasteriidae Jones & McKenzie 1980, Hipponicharionidae Ulrich & Bassler 1931 and possibly Beyrichonidae Ulrich & Bassler 1931, which may be represented by the species Galeopsis glabra Hinz-Schallreuter 1993a. Younger formations across central Australia have, by comparison, much lower bradoriid diversity (Jones & Kruse 2009).
Despite numerous reports of fossils from stratigraphic units within the Pertaoorrta Group in the Amadeus Basin (see overviews by Shergold 1986; Shergold et al. 1991), there is a surprising dearth of published palaeontological data, especially biostratigraphic data, from this sedimentary succession. Given the ubiquitous presence of Cambrian bradoriids in almost all other central Australian basins, the occurrence of bradoriids in the Amadeus Basin has long been suspected, and recent work by Smith (2012) has demonstrated their presence. The main aim of this contribution is to provide the first systematic analysis and biostratigraphic range data for bradoriid assemblages from the Cambrian (Series 3, Stage 5; Templetonian) Giles Creek Dolostone in the Ross River Gorge area in the NE Amadeus
SMITH, P.M., BROCK, G.A., PATERSON, J.R. & TOPPER, T.P., 2014:04:25. New bradoriid arthropods from the Giles Creek Dolostone (Cambrian Series 3, Stage 5; Templetonian), Amadeus Basin, central Australia. Memoirs of the Association of Australasian Palaeontologists 45, 233-248. ISSN 0810-8889.
A new assemblage of bradoriids is reported from the Giles Creek Dolostone (Cambrian Series 3, Stage 5; Templetonian) in the Ross River Gorge area, NE Amadeus Basin, central Australia. One new genus and four new species are described: Annge iperte gen. et sp. nov., Flemingopsis anteriospinata sp. nov., Monasterium ferox sp. nov. and Mongolitubulus tunpere sp. nov. Only M. tunpere sp. nov. may have been described previously from adjacent basins. This study provides the first systematic description of a Cambrian bradoriid assemblage from the Amadeus Basin.
P.M. Smith ([email protected]) and G.A. Brock ([email protected]) Department of Biological Sciences, Macquarie University, Sydney, New South Wales, 2109, Australia. J.R. Paterson ([email protected]) School of Environmental and Rural Science, University of New England, Armidale, New South Wales, 2351, Australia. T.P. Topper ([email protected]) Geological Museum, Natural History Museum of Denmark, University of Copenhagen, Øster Voldgade 5-7, DK-1350 Copenhagen K, Denmark and Department of Earth Sciences, Palaeobiology, Uppsala University, Villavägen 16, SE-752 36 Uppsala, Sweden. Received 4 October 2013.
Keywords: Bradoriida, bivalved arthropod, new taxa, Annge, Flemingopsis, Monasterium, Mongolitubulus, Northern Territory, Gondwana.

AAP Memoir 45 (2014)234
Figure 1. A, Map of Australia showing location of the Amadeus Basin and position of RRG section within the basin. B, Map of the local area around Ross River Gorge, Northern Territory, showing the position of the RRG section. C, Geological map of the Ross River Syncline with highlighted study area.

AAP Memoir 45 (2014) 235
Basin, Northern Territory.
GEOLOGICAL SETTING AND LOCALITYThe Amadeus Basin is a large intracratonic depocentre of Neoproterozoic, Cambrian and Ordovician marine sedimentary rocks located in central Australia (Fig. 1A). The eastern and western boundaries of the basin are buried under younger sedimentary deposits, and the northern and southern limits are structural and/or erosional rather than depositional (Edgoose 2012). The Ross River Gorge area is located 64 km east of Alice Springs within the Ooraminna sub-basin and provides perhaps the best surface exposure of the Pertaoorrta Group (Fig. 1B, C). The Pertaoorrta Group within the Ooraminna sub-basin comprises 2100 m of mixed siliciclastic and carbonate sedimentary packages ranging in age from Neoproterozoic to Furongian (Wells et al. 1967, 1970; Kennard & Lindsay 1991). Kennard & Lindsay (1991) divided the Pertaoorrta Group into five sequence stratigraphic packages (Є1- Є5). The oldest sequence package (Є1) is represented by units 1 and 2 of the Arumbera Sandstone which is largely Ediacaran in age. Sequence package Є2 is represented by lower Cambrian units 3 and 4 of the Arumbera Sandstone and the conformably overlying carbonate package of the Todd River Dolostone. Sequence package Є3 is represented by the Giles Creek Dolostone, deposited after an hiatus during the early Cambrian Botoman – early Toyonian stage. The final two sequence packages, Є4-5 are represented by carbonate-dominated units, the Shannon and Goyder Formations.
The Giles Creek Dolostone was originally described by Wells et al. (1967) based on the type section (ASW1) in Ross River Gorge. The formation is generally resistant to weathering and forms distinctive strike ridges with steep gorges in the Ooraminna sub-basin. It crops out as far west as the Alice Springs railway line, and as far south as latitude 24° 30’ (Wells et al. 1967).
At the type section in Ross River Gorge (ASW1), the formation reaches a thickness of 253 m (Wells et al. 1967),
though it reaches a maximum thickness of 402 m in the Ooraminna Anticline. Complete drill cores through the formation have been obtained from Alice 1, Bluebush 1, Wallaby 1, Dingo 1 and 2, and Orange 1 and 2 drillholes. The maximum thickness of the formation in these cores occurs in Wallaby 1 well at 383 m (Deckelman & Oaks 1986).
The fossiliferous material reported herein comes from the RRG section (23° 36′ 14″ S, 134° 28′ 56″ E) measured by GAB, JRP and TPT through 302.3 m (true thickness) of the lower Cambrian (Series 2) Todd River Dolostone and disconformably overlying earliest Templetonian (Cambrian Series 3, Stage 5) Giles Creek Dolostone (Figs 1-3). The Giles Creek Dolostone at this section (base at 23° 36’ 10” S, 134° 28’ 57” E) is represented by a total of 187 m of outcrop, of which the lower 60% is the most fossiliferous (Fig. 3).
The RRG section intercepts five distinct lithofacies of the Giles Creek Dolostone (Fig. 3). The basal 22 m is dominated by dolomitic to grey micritic limestone with black chert nodules containing large (1-3 cm), organophosphatic (botsfordiid) and calcareous brachiopods. This is overlain by a 13 m thick package of interbedded green shale, siltstone and limestone containing carbonate nodules that host well preserved Pagetia Walcott 1916, Xystridura Whitehouse 1936, dolichometopid, solenopleurid, alokistocarid and nepeid trilobites, as well as, agnostid arthropods and linguliformean brachiopods. These taxa are currently under investigation and will be described separately. The green calcilutites transition into a thick succession (84 m) of parasequences consisting of grey micrites, sparites and interbedded shales. These layers contain similar fossil faunas to those of the green calcilutites in addition to numerous shelly fragments readily apparent in thin sections. This package is sharply overlain by 18 m of dolomitic carbonate, which passes into massive, highly resistant, unfossiliferous buff dolostone, forming the massive east-west strike ridges in the region. The top of the RRG section is capped by a distinctive oolitic and oncoidal horizon at 136 m above the base of the Giles Creek Dolostone.
Figure 2. Detailed map of the RRG section within the Ross River Gorge, Northern Territory.

AAP Memoir 45 (2014)236
The deposition of the Giles Creek Dolostone occurred during a broad transgressive event across a prograding epeiric carbonate ramp. Deckelman & Oaks (1986) recognised five different depositional environments in the Giles Creek Dolostone in the Ross River Gorge area. The RRG section passes through the lower three of these. The earliest of the depositional environments is interpreted as a lagoon, tidal flat and mixed shoal setting. Lithofacies from the RRG section support this interpretation with micrites hosting microbially generated laminated carbonates dominant for 12 m true thickness (Fig. 3) in the basal part of the formation. There is a change to a shallow open shelf environment in the RGG section represented by an overlying 105 m thick shale and micrite-rich package (Fig. 3), accompanied by an increasing abundance of trilobites. This lithology gradually transitions into a package of grey micrites and sparites with interbedded shale parasequences (Fig. 3), suggesting a gradual shallowing during a sea level highstand (Kennard & Lindsay 1991). The increase in lithoclasts and presence of oolites in the upper parts of the unit (118 m above the base) marks the initiation of high energy, shallow ooid shoal facies (Deckelman & Oaks 1986).
AGE AND BIOSTRATIGRAPHYThe Giles Creek Dolostone disconformably overlies the lower Cambrian (Series 2, Stage 3) Todd River Dolostone in the RRG section (Figs 1-3). Regionally, the disconformity surface is cryptic, with little change in lithology or strike and dip of the two formations. The only obvious indication of chronostratigraphic change is the presence of a distinctive Templetonian fauna in the Giles Creek Dolostone, including an abundant, but low diversity trilobite assemblage (discussed below), and acrotretoid and large (1-3 cm) acrotheloid brachiopods.
Whilst trilobite faunas from the Giles Creek Dolostone are not the focus of this investigation, they are relatively abundant in the unit and are key for constraining the age of this formation. The Giles Creek Dolostone was originally assigned to the Australian early Middle Cambrian (Ordian/early Templetonian stage) by Joyce Gilbert-Tomlinson (in Wells et al. 1967) and Öpik (1968b, 1970, 1975) based on the co-occurrence of the trilobites Redlichia Cossmann 1902, Onaraspis Öpik 1968b and Xystridura, in addition to the hyolith Guduguwan hardmani (Etheridge in Foord 1890). Shergold (1986), however, noted that the presence of unidentified species belonging to Dinesus Etheridge 1896, Kootenia Walcott 1888, Nepea Whitehouse 1939 and ptychoparioid trilobites, found in the formation, may indicate a later Templetonian age. Shergold (1986) and Gravestock & Shergold (2001) also suggested that the Ordian/early Templetonian hiatus documented in the Georgina Basin (Shergold et al. 1985; Southgate & Shergold 1991; Smith et al. in press) may be present within the formation.
Intensive sampling of the RRG section has revealed an abundant, but relatively low diversity trilobite fauna. This fauna is currently under investigation and preliminary identifications have shown some species to be age-diagnostic, especially Xystridura filifera Öpik 1975 and
Figure 3. Lithostratigraphic column of the Giles Creek Dolostone through the RRG section. All sampled horizons are indicated. Stratigraphic ranges of bradoriid taxa, are plotted next to the column. Measurements are in metres true thickness from the base of the Giles Creek Dolostone. TRD is an abbreviation of Todd River Dolostone.

AAP Memoir 45 (2014) 237
Pagetia silicunda Jell 1975.Öpik (1975) originally described Xystridura filifera
from the Sandover Beds in the Elkedra area of the Northern Territory, but did not provide detailed age information except to say that the occurrence was ‘early Templetonian’. However, in Öpik’s (1979) subsequent monograph on agnostids, he noted the occurrence of Itagnostus elkedraensis (Etheridge 1902) from one of the localities where X. filifera is abundant (NT63). According to Laurie’s (2004, 2006) recent redefinition and zonal subdivision of the Australian Ordian and Templetonian stages, I. elkedraensis is indicative of the Pentagnostus anabarensis Zone of the earliest Templetonian. Although Laurie (2004) has questioned Öpik’s (1979) identification of the illustrated specimens of I. elkedraensis from Australia and thus reassigned them to a new species of Itagnostus Öpik 1979, I. oepiki Laurie 2004, this latter taxon is also indicative of a Pentagnostus anabarensis Zone age (Laurie 2006).
Pagetia silicunda was first described by Jell (1975) from the Coonigan Formation of western New South Wales. The diverse trilobite fauna from the Coonigan Formation (including the First Discovery Limestone) has yet to be described in detail, but Roberts & Jell (1990, p. 260) reported that Pentagnostus anabarensis Lermontova in Vologin 1940 and Pentagnostus veles Öpik 1979 co-occur with P. silicunda. Laurie (2004) has questioned the occurrence of Pentagnostus anabarensis in the Coonigan Formation and considers Pentagnostus veles to be a junior synonym of Pentagnostus praecurrens (Westergård 1936) which would imply an age within the eponymous zone immediately above the P. anabarensis Zone (cf. Laurie 2006, fig. 3).
Pending the description of trilobites and other faunal elements from the RRG section, the Giles Creek Dolostone in the Ross River Gorge area should tentatively be considered early Templetonian in age, possibly within the Pentagnostus anabarensis or Pentagnostus praecurrens Zones (sensu Laurie 2006). This is supported not only by the presence of Xystridura filifera and Pagetia silicunda (discussed above), but the co-occurrence of nepeids (Paterson 2005) and dolichometopids (Öpik 1982), the bradoriid Flemingopsis Jones & McKenzie 1981 (see Jones & Kruse 2009), and the apparent absence of Redlichia and Onaraspis, genera that are typical of Ordian or older successions (Laurie 2006; Paterson & Brock 2007). Previous reports of Redlichia and Onaraspis from the Giles Creek Dolostone may indicate that the formation is diachronous across the Amadeus Basin.
SYSTEMATIC PALAEONTOLOGYTerminology used to describe bradoriids follows, for the most part, that of Siveter & Williams (1997) and Williams & Siveter (1998). Abbreviations for lobation patterns follow those of ostracods, i.e., L1 = anterior lobe, L2 = anterior dorsal lobe and L3 = posterior lobe and S1 = sulcus between L1 and L2. However, this does not imply phylogenetic relationships between the bradoriids and ostracods, but is used for ease of discussion when referring to the position of the lobes in terms of valve area. Use of the term valve (‘bivalved head shield’ or just ‘head-shield’ of Siveter et al. 2003, p.13) is also used in a strictly descriptive sense and has no phylogenetic connotation. All measurements are in millimeters unless otherwise stated. All illustrated and described specimens are provided with SAM P numbers and are held at the South Australian Museum. Stratigraphic ranges for all taxa are given in terms of true thickness from the base of the Giles Creek Dolostone.
Phylum ARTHROPODA Siebold & Stannius 1845 Order BRADORIIDA Raymond 1935 Family COMPTALUTIDAE Öpik 1968a
Annge gen. nov. Type species. Annge iperte gen. et sp. nov.
Etymology. From Arrernte aboriginal language annge = seed, referring to the seed-like shape of the closed bradoriid shields. Pronounced “uh-ng”.
Diagnosis. Valve postplete, anterior margin slightly convex, posterior margin highly convex. Anterodorsal lobe (L1) indistinct, dorsal lobe (L2) well developed. Deep furrow separates anterior lobe (L1) from dorsal lode (L2). Marginal rim well defined. Valve covered with punctate ornament of evenly spaced circular indentations.
Remarks. The presence of a straight hinge line, distinct lateroadmarginal ridge, posterodorsal curve and more than a single dorsal lobe (L2) separated by a sulcus (S1) (Fig. 4A, D, F, H) suggests an affinity with the Comptalutidae Öpik 1968a. There are no genera within this family that have the unique lobation present in the new genus. The subtriangular outline of the valves is reminiscent of taxa in the Hipponicharionidae Sylvester-Bradley 1961 such as Albrunnicola bengtsoni Hinz-Schallreuter 1993b, but the position of the lobes within the anterior half of the valve argues against such an assignment. The anterior lobe (L1) is similar to species of Bradoria Matthew 1899 and Walcottella Ulrich & Bassler 1931 – both assigned to the Bradoriidae Matthew 1902 – although the presence of the secondary dorsal lobe (L2) and a sulcus (S1) between these lobes is not characteristic of this family.
Four subfamilies within the Comptalutidae are currently recognised: Comptalutinae Öpik 1968a, Oepikalutinae Jones & McKenzie 1980, Phasoiinae Hinz-Schallreuter 1999 and Quetopsinae Hinz-Schallreuter 1999. Annge gen. nov. cannot be easily assigned to any of these subfamilies because it does not possess the full set of characteristics that define them. The Quetopsinae is probably the most similar, but the presence of a distinct dorsal lobe (L2) excludes the genus from this subfamily. The lobation of Annge gen. nov. is comparable to that of Wutingella binodosa Zhang 1974, a species within the Comptalutinae, with both genera possessing two subovate, anterodorsal lobes separated by a sulcus. However, in Annge gen. nov., the anterior lobe (L1) is placed more dorsally and lacks a connecting ridge to the dorsal lobe (L2). The lack of a connecting ridge makes assignment of Annge gen. nov. to the Comptalutinae difficult.
Annge gen. nov. is similar to Aristaluta Öpik 1961, a taxon that is also difficult to assign to an existing subfamily; both taxa have postplete valves with two distinct anterior (L1) and dorsal lobes (L2) separated by a sulcus (S1), and punctate ornament. Aristaluta, however, differs from Annge gen. nov. in having a distinct sulcus (S2) behind the dorsal lobe (L2), a more convex anterior margin and greater inflation of the valves. Both taxa may belong to a new comptalutid subfamily but specimens of Aristaluta from the Devoncourt Limestone, eastern Georgina Basin, are damaged and effaced in “crack out” specimens (Öpik 1961, pl. 24, figs 2a-c, 3a-d, 4). Given the indifferent nature of the preservation of the type material of Aristaluta, we refrain from creating a new subfamily until better preserved material can be discovered.

AAP Memoir 45 (2014)238
Figure 4. Annge iperte gen. et sp. nov. from the early Templetonian Giles Creek Dolostone at the RRG section. A, C; holotype, SAMP48892, sample RRG/280.3, 51.52 m above the base of the section; A, lateral view of eroded left valve; C, internal view of left valve; B; SAMP48893, sample RRG/232.8, 10.58 m above the base of the section; highly eroded left valve; D, I; SAMP48894, sample RRG/280.3, 51.52 m above the base of the section; D, lateral view of fragment from left valve anterior; (continued opposite)

AAP Memoir 45 (2014) 239
Annge gen. nov. is thus left under open nomenclature at subfamily level. Annge iperte sp. nov. (Fig. 4)
Etymology. From Arrernte aboriginal language iperte = pit/ditch, referring to the pitted ornament covering the surface of the shields. Pronounced “ee-per-tee”.
Diagnosis. As for genus.
Type material. Holotype: SAMP48892 (Fig. 4A, C) from sample RRG/280.3, 51.52 m above the base of the section. Paratypes: SAMP48894 (Fig. 4D, I); SAM P48895 (Fig. 4E, G, H, J) and SAMP48896 (Fig. 4F, K) from sample RRG/280.3, 51.52 m above the base of the section. SAMP48893 (Fig. 4B) from sample RRG/232.8, 10.58 m above the base of the section. All type specimens were recovered from the RRG section, Giles Creek Dolostone, Ross River Syncline.
Description. Valve postplete, small (average 0.65 mm in length), subtriangular in lateral outline (Fig. 4A). Anterior margin slightly convex (Fig. 4D); posterior margin highly convex (Fig. 4A, C, D), posterodorsal curve present (Fig. 4A, D); hinge line straight (Fig. 4A, C, D, F). Valve slightly inflated, more so posteroventrally. Lateroadmarginal ridge present (Fig. 4D, F) and well defined around entire valve. Anterodorsal lobe (L1) indistinct, dorsal lobe (L2) well developed and very rounded in shape (Fig. 4A, D, E, G, H). Deep furrow separates anterodorsal lobe (L1) from dorsal lode (L2). Dorsal lobe (L1) at valve mid-length; lobe dorsoventrally flattened, forming small, triangular projection with lobe apex directed anteriorly (Fig. 4H). External surface of valve covered with punctate ornament consisting of regular (7 μm diameter), evenly spaced (20 μm apart), circular indentations (Fig. 4I).
Remarks. The external ornament of this taxon appears to be unique (Fig. 4D, I, F). The ornament of Liangshanella circumbolina Topper, Skovsted, Brock & Paterson 2011 is similar to that of Annge iperte gen. et sp. nov. in possessing evenly spaced circular pits (Topper et al. 2011, fig. 5C-E), but in L. circumbolina there are raised circular structures in the centre of each pit, rather than circular depressions. The ornament of Phasoia armini Jones & Kruse 2009 is also somewhat similar to A. iperte gen. et sp. nov. (Jones & Kruse 2009, fig. 11J), although in P. armini, indentations are adjacent and joined to one another and not widely spaced (Fig. 4I).
Distribution. Giles Creek Dolostone, Amadeus Basin, Northern Territory. Cambrian Series 3, Stage 5; early Templetonian.
Material. 17 valves in total, including 5 specimens figured (1 conjoined, 4 left, 1 right and 11 fragmented) from 10.58 m to 51.52 m (Fig. 3) above the base of the Giles Creek Dolostone.
Subfamily OEPIKALUTINAE Jones & McKenzie 1980
Flemingopsis Jones & McKenzie 1981
Type species. Flemingia duo Jones & McKenzie 1980.
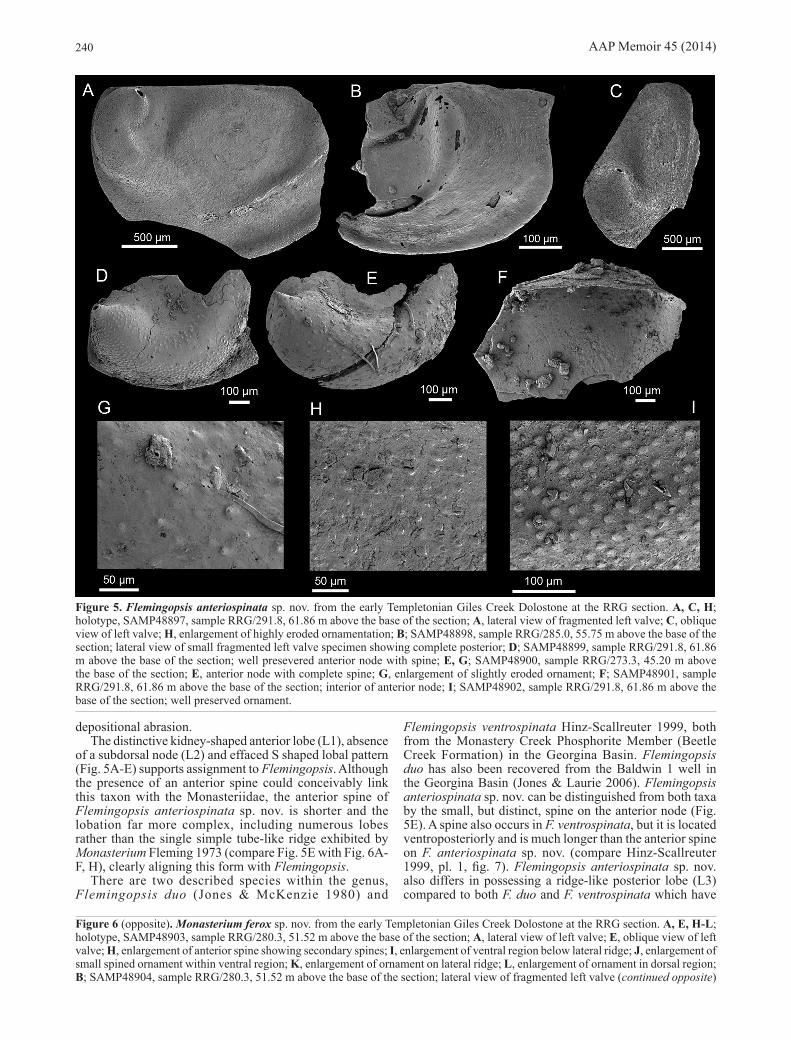
Flemingopsis anteriospinata sp. nov. (Fig. 5)
Etymology. Derived from the Latin anterior = front or foremost, and spina = spine or thorn. In reference to the unique, small spine on the anterior of valve.
Type material. Holotype: SAMP48897 (Fig. 5A, C, H) from sample RRG/291.8, 61.86 m above the base of the section. Paratypes: SAMP48898 (Fig. 5B) from sample RRG/285.0, 55.75 m above the base of the section. SAMP48900 (Fig. 5E, G) from sample RRG/273.3, 45.20 m above the base of the section. SAMP48899 (Fig. 5D); SAMP48901 (Fig. 5F) and SAMP48902 (Fig. 5I) from sample RRG/291.8, 61.86 m above the base of the section. All type specimens were recovered from the RRG section, Giles Creek Dolostone, Ross River Syncline.
Diagnosis. Valve amplete, subrectangular in lateral outline, anterior margin convex, posterior margin slightly concave, hinge line straight. Valve moderately inflated. Lateroadmarginal ridge present. Anterodorsal lobe (L1) well defined and kidney-shaped, sharp, conical spine on lobe (L1) apex. Sulcus (S1) separates anterior lobe from indistinct anterior dorsal swelling. Long, curved posterior lobe (L3) separated from other lobes, parallels the valve margin, forming a distinct ridge towards posterior. Ornament of evenly spaced pustules.
Description. Valve amplete, subrectangular in lateral outline, anterior margin convex (Fig. 5A), posterior margin slightly concave (Fig. 5B), hinge line straight (Fig. 5A, B). Valve moderately inflated (Fig. 5C). Lateroadmarginal ridge separated from main valve by a shallow furrow extending around most of margin. Anterodorsal lobe (L1) well defined, kidney-shaped, adorned with a small (120 μm high, 98 μm diameter), sharp, conical spine projecting from lobe apex in a posterodorsal direction (Fig. 5A, D, E). Sulcus (S1) separates anterior lobe from distinct anterior dorsal swelling (Fig. 5A). Long, curved posterior lobe (L3) separated from other lobes, appears to parallel the valve margin extending over mid to posterior valve, forming swelling slightly towards posterior forming a ridge (Fig. 5A, B). L1, the anterior dorsal swelling and L3 appear to have been effaced from a continuous S shaped lobal pattern (Fig. 5A). External surface covered with ornament of small (12 um in size), evenly spaced pustules (Fig. 5G-I).
Remarks. This taxon was recovered from four sample horizons (Fig. 3), mainly as fragmentary valves with the anterior lobe (L1) preserved (Fig. 5D, E). The holotype is the most complete specimen (Fig. 5A, C, H) and shows the key morphological features of the new taxon. The distinctive pustulose ornament of this species (Fig. 5G-I) is variably expressed, mainly due to moderate levels of post-
I, enlargement of ornamentation; E, G, H, J; SAMP48895, sample RRG/280.3, 51.52 m above the base of the section; E, oblique view of left valve co-joined to right valve; G, oblique view of right valve co-joined to left valve ; H, dorsal view with anterior positioned left; J, enlargement of left valve showing broken region with possible secondary shell layer below; F, K; SAMP48896, sample RRG/280.3, 51.52 m above the base of the section; F, lateral view of fragment from right valve anterior; K, cross section showing two shell layers.
AAP Memoir 45 (2014)240
depositional abrasion. The distinctive kidney-shaped anterior lobe (L1), absence
of a subdorsal node (L2) and effaced S shaped lobal pattern (Fig. 5A-E) supports assignment to Flemingopsis. Although the presence of an anterior spine could conceivably link this taxon with the Monasteriidae, the anterior spine of Flemingopsis anteriospinata sp. nov. is shorter and the lobation far more complex, including numerous lobes rather than the single simple tube-like ridge exhibited by Monasterium Fleming 1973 (compare Fig. 5E with Fig. 6A-F, H), clearly aligning this form with Flemingopsis.
There are two described species within the genus, Flemingopsis duo (Jones & McKenzie 1980) and
Flemingopsis ventrospinata Hinz-Scallreuter 1999, both from the Monastery Creek Phosphorite Member (Beetle Creek Formation) in the Georgina Basin. Flemingopsis duo has also been recovered from the Baldwin 1 well in the Georgina Basin (Jones & Laurie 2006). Flemingopsis anteriospinata sp. nov. can be distinguished from both taxa by the small, but distinct, spine on the anterior node (Fig. 5E). A spine also occurs in F. ventrospinata, but it is located ventroposteriorly and is much longer than the anterior spine on F. anteriospinata sp. nov. (compare Hinz-Scallreuter 1999, pl. 1, fig. 7). Flemingopsis anteriospinata sp. nov. also differs in possessing a ridge-like posterior lobe (L3) compared to both F. duo and F. ventrospinata which have
Figure 5. Flemingopsis anteriospinata sp. nov. from the early Templetonian Giles Creek Dolostone at the RRG section. A, C, H; holotype, SAMP48897, sample RRG/291.8, 61.86 m above the base of the section; A, lateral view of fragmented left valve; C, oblique view of left valve; H, enlargement of highly eroded ornamentation; B; SAMP48898, sample RRG/285.0, 55.75 m above the base of the section; lateral view of small fragmented left valve specimen showing complete posterior; D; SAMP48899, sample RRG/291.8, 61.86 m above the base of the section; well presevered anterior node with spine; E, G; SAMP48900, sample RRG/273.3, 45.20 m above the base of the section; E, anterior node with complete spine; G, enlargement of slightly eroded ornament; F; SAMP48901, sample RRG/291.8, 61.86 m above the base of the section; interior of anterior node; I; SAMP48902, sample RRG/291.8, 61.86 m above the base of the section; well preserved ornament.
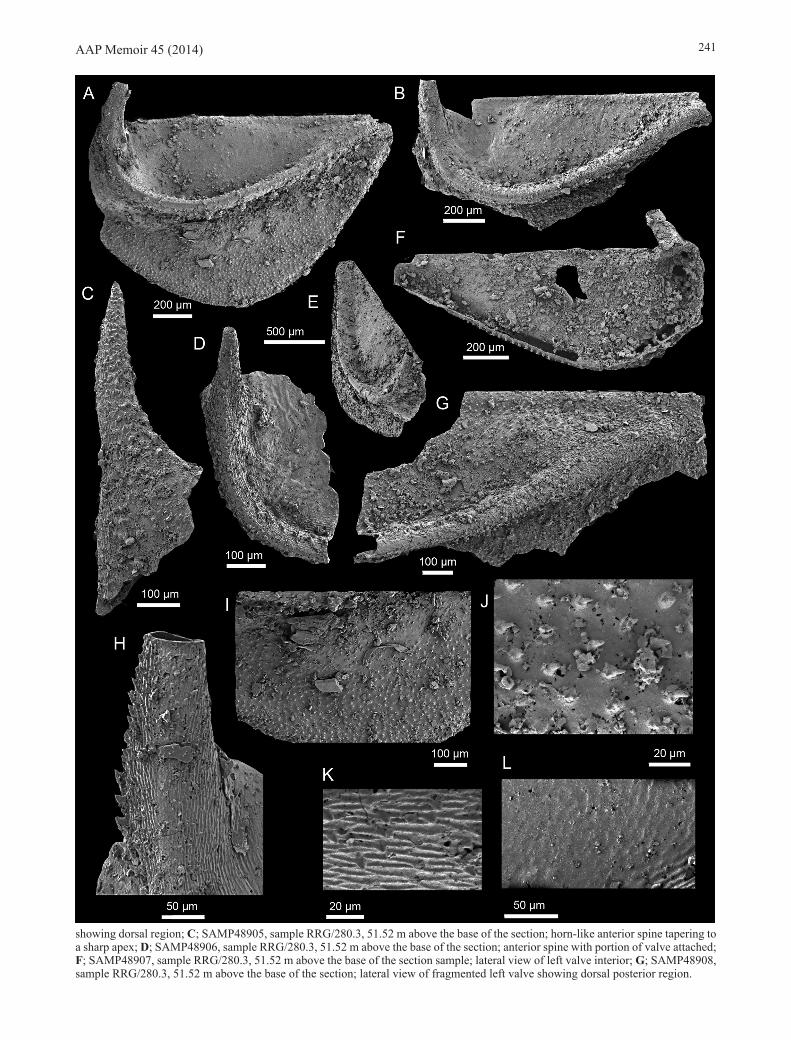
Figure 6 (opposite). Monasterium ferox sp. nov. from the early Templetonian Giles Creek Dolostone at the RRG section. A, E, H-L; holotype, SAMP48903, sample RRG/280.3, 51.52 m above the base of the section; A, lateral view of left valve; E, oblique view of left valve; H, enlargement of anterior spine showing secondary spines; I, enlargement of ventral region below lateral ridge; J, enlargement of small spined ornament within ventral region; K, enlargement of ornament on lateral ridge; L, enlargement of ornament in dorsal region; B; SAMP48904, sample RRG/280.3, 51.52 m above the base of the section; lateral view of fragmented left valve (continued opposite)
AAP Memoir 45 (2014) 241
showing dorsal region; C; SAMP48905, sample RRG/280.3, 51.52 m above the base of the section; horn-like anterior spine tapering to a sharp apex; D; SAMP48906, sample RRG/280.3, 51.52 m above the base of the section; anterior spine with portion of valve attached; F; SAMP48907, sample RRG/280.3, 51.52 m above the base of the section sample; lateral view of left valve interior; G; SAMP48908, sample RRG/280.3, 51.52 m above the base of the section; lateral view of fragmented left valve showing dorsal posterior region.

AAP Memoir 45 (2014)242
a much less distinct and well rounded posterior lobe (L3).
Distribution. Giles Creek Dolostone, Amadeus Basin, Northern Territory. Cambrian Series 3, Stage 5; early Templetonian.
Material. 14 valves in total, including 6 specimens figured (12 left and 2 right) from 36.07 m to 63.12 m (Fig. 3) above the base of the Giles Creek Dolostone.
Family MONASTERIIDAE Jones & McKenzie 1980
Monasterium Fleming 1973
Type species. Monasterium oepiki Fleming 1973.
Monasterium ferox sp. nov. (Fig. 6)
Etymology. Derived from the Latin word ferox, savage. In reference to the numerous spines and large horn-like spine on the valve resembling those adorning savage beasts of fantasy and folklore.
Type material. Holotype: SAMP48903 (Fig. 6A, E, H-L) from sample RRG/280.3, 51.52 m above the base of the section. Paratypes: SAMP48904 (Fig. 6B); SAMP48905 (Fig. 6C); SAMP48906 (Fig. 6D); SAMP48907 (Fig. 6F) and SAMP48908 (Fig. 6G) from sample RRG/280.3, 51.52 m above the base of the section. All type specimens were recovered from the RRG section, Giles Creek Dolostone, Ross River Syncline.
Diagnosis. Subcircular amplete to postplete outline. Lateral tube-like ridge transverses midvalve surface. Anterior lobe (L1) forming a thick horn-like spine projecting dorsally from valve surface. Dorsal surface above lateral ridge with weakly defined, raised polygonal texture. Lateral ridge covered with elongate, squamose reticulate network forming distinct transverse, closely packed micro ridges. Small spinules interspersed between secondary ridges on ventral and dorsal surface. Ventral valve surface below the lateral ridge covered with small, irregular, evenly dispersed, recurved spines.
Description. Small (1.8 mm average length) subcircular amplete to postplete valve with a straight hinge line (Fig. 6A, B, F, G). Valve very slightly convex in the transverse plane, reaching maximum convexity near midlength (Fig. 6E). Lateral tube-like ridge traverses the mid-valve surface originating at anterodorsal margin, extending ventrally to 33% of valve length, then recurving anteriorly before terminating at posterodorsal margin (Fig. 6A, B). Ridge gradually increasing in height to 33% of valve length (reaching a maximum of 200 μm above valve surface) after which height decreases towards ridge termination. Anterior lobe (L1) forming a thick horn-like spine which projects dorsally from valve surface and recurves towards the posterior (Fig. 6A, B, D, H). Apex of spine tapering to sharp point (Fig. 6C). Largest complete spine 0.5 mm long and 0.3 mm in width.
Surface ornament consists of three distinct types of sculpture located on different regions of valve (Fig. 6A). Dorsal valve surface above lateral ridge bears a weakly defined, raised polygonal texture (Fig. 6L). Lateral ridge covered with elongate, squamose reticulate network forming distinct transverse, closely packed microridges (Fig. 6K).
Small spinules interspersed between secondary ridges on ventral and dorsal surface (Fig. 6H). Ventral region of valve below the lateral ridge covered by small, irregular, evenly dispersed, recurved spines (Fig. 6I, J). The ornament types are sharply differentiated at lateral ridge boundaries.
Remarks. All valves come from a single sample horizon (RRG/280.3; Fig. 3). Absence of a marginal rim and presence of a single pair of very well developed horn-like spines (Fig. 6A-E) permit confident assignment to Monasterium.
There are five described species of Monasterium, two from Kazakhstan, one from China and the remaining two from the Beetle Creek Formation (Monastery Creek Phosphorite Member), Georgina Basin. The Chinese taxon, Monasterium bucerum Zhang 1987, and the two species from Kazakhstan, Monasterium ivshini Melnikova 1990, and Monasterium tchizhovi Melnikova 2003 all lack a transverse lateral ridge. Monasterium dorium Fleming 1973 from the Monastery Creek Phosphorite Member also lacks this distinctive feature. Instead, these species possess either a distinct rib, as in M. tchizhovi, or the valve surface is essentially smooth, as in M. bucerum, M. dorium and M. ivshini. The type species, Monasterium oepiki Fleming 1973, has a lateral ridge which is similar in shape and size to Monasterium ferox sp. nov., but is far less distinct (compare, for example, Fig. 6A, B, D, E, G to Fleming 1973, pl. 2, figs 7-10). The prominence of the lateral ridge in M. ferox sp. nov. may partially be the result of differential accommodation in a thin, flexible valve to post-depositional compaction, as has been suggested for some specimens of M. oepiki by Fleming (1973, p. 9, pl. 2, figs 6-10, pl. 4, figs 1-3). However, the ubiquitous presence of this prominent ridge in all specimens of M. ferox sp. nov. (Fig. 6A, B, D, E, G) indicate it is a stable diagnostic feature of this new taxon.
The ornament on the surface of the spine and valve of M. ferox sp. nov. also differs dramatically from descriptions in the other species (Fig. 6 H-L). Monasterium oepiki, and possibly M. bucerum, share the same raised polygonal texture on the upper surface of the valve, but lack the smaller spinules on the lower surface, instead, possessing smaller and smoother pustules or a continuation of the polygonal texture (Fleming 1973, pl. 4, fig. 7; Zhang 1987, fig. 9A). Similarly, the ornament on the spine of M. ferox sp. nov. resembles that of M. oepiki in that both have raised ridges with a reticulate network, but M. oepiki lacks the interdispersed secondary spines of M. ferox sp. nov. (Fig. 6H).
Distribution. Giles Creek Dolostone, Amadeus Basin, Northern Territory. Cambrian Series 3, Stage 5; early Templetonian.
Material. 10 valves in total, including 6 specimens figured (8 left and 2 right) from 51.52 m (Fig. 3) above the base of the Giles Creek Dolostone.
Family MONGOLITUBULIDAE Topper, Skovsted, Harper & Ahlberg 2013
Mongolitubulus Missarzhevsky 1977
Type species. Mongolitubulus squamifer Missarzhevsky 1977.
Remarks. Isolated, ornamented phosphatic spines are common constituents in Cambrian faunal assemblages. The

AAP Memoir 45 (2014) 243
majority of these isolated spines are associated with the generic name Mongolitubulus and have been interpreted to have affinities with protoconodonts, tubular hyolithelminths, lobopods, trilobites and bradoriids (see Topper et al. 2013 for review). Isolated spines are of course inevitably imperfectly preserved, exhibiting broken basal margins and hence obscuring their biological affinity. A number of bradoriid arthropod species from around the globe (Skovsted & Peel 2001; Skovsted 2005; Zhang 2007; Topper et al. 2007; Kouchinsky et al. 2011) have now been documented with one or more ornamented spines attached to their valves, providing compelling evidence that bradoriids possessed Mongolitubulus-like spines. However, documentation of
ornamented spines on Cambrian trilobites (Li et al. 2012) and lobopodians (Caron et al. 2013) demonstrate that ornamented spines probably arose numerous times across many animal lineages.
The type species Mongolitubulus squamifer Missarzhevsky 1977 was erected based on four tapering, ornamented phosphatic spines from the lower Cambrian of Mongolia. These spines were isolated and to date no spine that explicitly matches the holotype specimen (Missarzhevsky 1977, pl. 1, figs 1-2) has been found attached to the valve of a bradoriid arthropod. It is thus conceivable that Mongolitubulus (sensu stricto) may represent spines from an unknown scleritome-bearing animal. There are some morphological differences
Figure 7. Mongolitubulus tunpere sp. nov. valve material from the early Templetonian Giles Creek Dolostone at the RRG section. A; SAMP48909, sample RRG/280.3, 51.52 m above the base of the section; valve with two large attached spines; B, E, F; holotype, SAMP48910, sample RRG/280.3, 51.52 m above the base of the section; B, oblique view of valve surface showing features of effaced hinge line and large attached spine; E, close up on effaced hinge line region; F, oblique view of valve surface showing features of effaced hinge line and large attached spine; C, I; SAMP48911, sample RRG/280.3, 51.52 m above the base of the section; C, effaced valve fragment showing dorsal hinge line; I, top down view showing the three layered valve microstructure; D; SAMP48912, sample RRG/280.3, 51.52 m above the base of the section; enlargement of valve ornamentation; G, H; SAMP48913, sample RRG/280.3, 51.52 m above the base of the section; G, enlargement of ventral valve margin showing numerous small spine bases (indicated with white arrows); H, oblique view showing the three layered valve microstructure.

AAP Memoir 45 (2014)244
between spines assigned to M. squamifer and those spines documented to be directly attached to bradoriid valves, especially the lack of an expanded base and slight differences in ornament but, the absence of an expanded base and slight variation in ornament does not exclude the possibility that M. squamifer has bradoriid affinities (see Topper et al. 2013). Specimens assigned to M. squamifer and associated bradoriid valves from the early Cambrian of Antarctica
do show similar microstructures (Wrona 2004, 2009), however the spines are not directly attached to bradoriid valves. Similarities in documented microstructure, general size and spatial and temporal distribution of M. squamifer do provide persuasive evidence for a bradoriid arthropod affinity, and consequently supports assignment of the spine-bearing bradoriid documented here to Mongolitubulus. However, we acknowledge that the type species has not yet
Figure 8. Mongolitubulus tunpere sp. nov. spine material from the early Templetonian Giles Creek Dolostone at the RRG section. A, D; SAMP48914, sample RRG/280.3, 51.52 m above the base of the section; A, enlargement of large spine showing typical Mongolitubulus ornament; D, section from a large spine; B; SAMP48915, sample RRG/280.3, 51.52 m above the base of the section; large spine showing flaring at base; C; SAMP48916, sample RRG/280.3, 51.52 m above the base of the section; spine showing flaring at base; E; SAMP48917, sample RRG/280.3, 51.52 m above the base of the section; long spine with ornament showing hooked tip; F; SAMP48918, sample RRG/280.3, 51.52 m above the base of the section; specimen showing distal end of spine with hooked tip; G; SAMP48919, sample RRG/280.3, 51.52 m above the base of the section; cross section through spine showing three layers; H; SAMP48920, sample RRG/280.3, 51.52 m above the base of the section; enlargement of hooked spine tip; I; SAMP48921, sample RRG/280.3, 51.52 m above the base of the section; hooked tipped spine showing folding; J; SAMP49120, sample RRG/280.3, 51.52 m above the base of the section; enlargement of transitional valve to spine ornament; K; SAMP49121, sample RRG/280.3, 51.52 m above the base of the section; spine attached to valve showing transition ornament from valve to spine.

AAP Memoir 45 (2014) 245
been documented directly attached to a bradoriid valve and consequently there remains some doubt over the taxonomic affinity of the type species.
Mongolitubulus tunpere sp. nov. (Figs 7, 8)
?1998 ?Silicunculus sp.; Kruse, p. 56, fig.48C-G. ?2009 Monasterium oepiki Fleming; Jones & Kruse, p. 77,
fig. 13A-B.
Etymology. From Arrernte aboriginal language tunpere = hook. In reference to the large hooked tip that crown the spines of this bradoriid. Pronounced “toon-per”. Type material. Holotype: SAMP48910 (Fig. 7B, E, F) from sample RRG/280.3, 51.52 m above the base of the section. Paratypes: SAMP48909 (Fig. 7A); SAMP48911 (Fig. 7C, I); SAMP48912 (Fig. 7D); SAMP48913 (Fig. 7G, H); SAMP48914 (Fig. 8A, D); SAMP48915 (Fig. 8B); SAMP48916 (Fig. 8C); SAMP48917 (Fig. 8E); SAMP48918 (Fig. 8F); SAMP48919 (Fig. 8G); SAMP48920 (Fig. 8H); SAMP48921 (Fig. 8I); SAMP49120 (Fig. 8J) and SAMP49121 (Fig. 8K) from sample RRG/280.3, 51.52 m above the base of the section. All type specimens were recovered from the RRG section, Giles Creek Dolostone, Ross River Syncline.
Diagnosis. Valve incompletely known; hinge line curved dorsally. Valve with at least two larger spines on the surface, numerous smaller spines along ventral margin. Ornament consists of small, mushroom-shaped pustules and a smooth hinge area. Valve and spine share three layered microstructure. Spine apex curved, with sharp, recurved tip similar to a hook in appearance. Spine ornament transitions from valve mushroom-shaped pustules to small, semi-regularly arranged rhomboid scale-like protuberances on outer surface.
Description. Valve outline incompletely known, originally possibly postplete (Fig. 7C), extensively folded. Valves hinge line curve dorsally (Fig. 7B). Valve fragments frequently covered by spine bases (Fig. 7A-C, F). More than one large spine present on single valve (Fig. 7A) and numerous smaller spines along ventral margin of a separate valve (Fig. 7G). Surface covered with small, mushroom-shaped pustules (26 μm high, 18 μm wide), distinctly different from ornament on the spine (Fig. 7D). Spines relatively long (max 2.5 mm length), slender, straight to very gently curved with a circular to slightly oval cross-section (450-560 μm diameter). Most spines detached from valves and broken into fragments. Spines with apical end closed, flattened in place of curvature, with a sharp, recurved tip (0.1 mm in average length); hook-like in appearance (Fig. 8E, F, H, I). Abapical region expands rapidly to a flared base for attachment to valve (Fig. 8K). Spine ornament transitions from valve mushroom-shaped pustules to small, semi-regularly arranged rhomboid scale-like protuberances on outer surface (Fig. 8A). Individual protuberances 10-20 μm in diameter. Spine and valve made up of three distinct layers (Figs 7H, I, 8G), a thick (8 μm) outer and intermediate layer, with a thinner (4 μm) internal layer.
Remarks. This taxon is largely represented by broken, detached spines. Most valves and spines consist of fragments that are highly folded and compressed (Figs 7A-C, F, 8I),
suggesting the valves were weakly mineralised, typical of bradoriids (Topper et al. 2013). Valves with attached spine bases bearing the same ornament as isolated spines demonstrate unequivocally that the spines were attached to the valve.
Spines ornamented with rhomboid scales (Fig. 8A-F, H-K) are typical of the bradoriid genus Mongolitubulus (Wrona 2009; Topper et al. 2011, 2013). Although, the holotype of M. squamifer (Missarzhevsky 1977, pl. 1, figs 1-2) possesses a more diamond-shaped scale ornament, assemblages of M. squamifer typically show a high degree of surface ornament variation with diamond-shaped and rounded scales intergrading with the rhomboidal protuberances (Skovsted & Peel 2001). Recent work by Li et al. (2012) demonstrates that similarly ornamented spines also adorned some eodiscoid trilobites (e.g., Hupeidiscus orientalis Zhang 1953 from South China). Topper et al. (2013) showed that in almost all known species of Mongolitubulus, the size of the spines are approximately twice or greater than those of H. orientalis spines; with the spine of Mongolitubulus unispinosa Topper, Skovsted, Brock & Paterson 2007, approaching 1.5 times the size of H. orientalis. The large spines of M. tunpere sp. nov. are 5.6 times greater in diameter (Fig. 8A, D) than those of the largest specimens of H. orientalis, discounting an affinity with H. orientalis for the isolated spines in the Giles Creek Dolostone assemblage. Mongolitubulus tunpere sp. nov. spines have the second largest recorded diameter (560 μm), with only “M. squamifer” spines from the early Cambrian Shabakhty Formation, Kazakhstan (Dzik 2003, fig. 2A) being larger. Topper et al. (2013) raised doubt about the bradoriid affinity of “M. squamifer” spines recorded by Dzik (2003). This would make the spines of Mongolitubulus tunpere sp. nov. the largest known spines of Mongolitubulus (sensu stricto).
Similar long, recurved, hooked spines (Fig. 8E, F, H, I) have been described from the Gum Ridge Formation, Thorntonia Limestone (Georgina Basin) and Montejinni Limestone (Wiso Basin) by Kruse (1998); and Jones & Kruse (2009). Kruse (1998) initially referred a few specimens to ?Silicunculus sp., a type of sponge spicule, but later Jones & Kruse (2009) referred them to Monasterium oepiki. Jones & Kruse (2009) argued that the hooked tip and apparent lack of ornament was characteristic of Monasterium rather than Mongolitublus spines. Although Monasterium is present in the Giles Creek Dolostone, the spines discussed here bear no resemblance to Monasterium ferox sp. nov. spines, which terminate in a straight, pointed apex and are ornamented with a distinctive reticulate network bearing secondary spinules. The hooked spines from the Georgina and Wiso Basins thus may be conspecific with M. tunpere sp. nov., being of a similar age. However, hooked spines are known from all Cambrian Series 3 and Furongian species of Mongolitubulus, including Mongolitubulus reticulatus Kouchinsky, Bengtson, Clausen, Gubanov, Malinky & Peel 2011 and M. aspermachaera Topper, Skovsted, Harper & Ahlberg 2013. Hence, without any Monogolitubulus valve material from the Georgina and Wiso Basins to examine, only tentative connections can be made with the Giles Creek Dolostone species.
Mongolitubulus tunpere sp. nov. differs from other Mongolitubulus species in its spinosity and valve ornamentation. Possession of more than a single spine on the valve (Fig. 7A) of M. tunpere sp. nov. differentiates this species from both M. unispinosa and M. unialata Zhang 2007. Mongolitubulus anthelios Betts, Topper, Valentine,

AAP Memoir 45 (2014)246
Cossmann, m., 1902. Revue Critique de Paldozoologie. Rectifications de la nomenclature 16, 52 p.
DeCkelman, J.a. & oaks, R.Q., 1986. Giles Creek Dolostone. 12th International Sedimentological congress, Field excursion 25B, 34 - 40.
Dzik, J., 2003. Early Cambrian lobopodian sclerites and associated fossils from Kazakhstan. Palaeontology 46, 93-112.
eDgoose, C.J., 2012. The Amadeus Basin, Central Australia. Episodes 35, 256-263.
etheRiDge, R.J., 1896. Evidence of the existence of a Cambrian fauna in Victoria. Proceedings of the Royal Society of Victoria 8, 52-64.
etheRiDge, R.J., 1902. Official Contributions to the Palaeontology of South Australia. No. 13. Evidence of further Cambrian trilobites. South Australian Parliamentary Papers 127, 13-14, pl.11.
Fleming, P.J.g., 1973. Bradoriids Form the Xystridura Zone of the Georgina Basin, Queensland. Geological Survey of Queensland, Palaeontology Papers 31, 1-17.
FooRD, a.h., 1890. Description of fossils from the Kimberley district, Western Australia. Geological Magazine (Decade III) 7, 98-106.
gozalo, R. & hinz-sChallReuteR, i., 2002. Biostratigraphy and palaeobiogeography of the Cambrian genus Hipponicharion (Ostracoda). Paläontologische Zeitschrift 76, 65-74.
gRavestoCk, D.i. & sheRgolD, J.h., 2001. Australian Early and Middle Cambrian sequence biostratigraphy with implications for species diversity and correlation. 107-136 in Zhuravlev, A.Y. & Riding, R. (eds.), The Ecology of the Cambrian Radiation. Columbia University Press, New York.
hinz-sChallReuteR, i., 1993a. Ostracodes from the Middle Cambrian of Australia. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 188, 305-326.
hinz-sChallReuteR, i., 1993b. Cambrian ostracods mainly from Baltoscandia and Morocco. Archiv für Geschiebekunde 1, 385–448.
hinz-sChallReuteR, i., 1999. Systematics and phylogeny of the Oepikalutidae (Cambrian Ostracoda). Greifswalder Geowissenschaftliche Beiträge 6, 23-53.
hinz-sChallReuteR, i., gozalo, R. & liñán, e., 2007. New Bradorid Arthropods from the Lower Cambrian of Spain. Micropaleontology 53, 497-509.
Jell, P.a., 1975. Australian Middle Cambrian eodiscoids with a review of the superfamily. Palaeontographica Abteilung A 150, 1-97.
Jones, P.J. & kRuse, P.D., 2009. New Middle Cambrian bradoriids (Arthropoda) from the Georgina Basin. Memoirs of the Association of Australasian Palaeontologists 37, 55-86.
Jones, P.J. & lauRie, J.R., 2006. Bradoriida and Phosphatocopida (Arthropoda) from the Arthur Creek Formation (Middle Cambrian), Georgina Basin, central Australia. Memoirs of the Association of Australasian Palaeontologists 32, 205-224.
Jones, P.J. & mCkenzie, k.g., 1980. Queensland Middle Cambrian Bradoriida (Crustacea): new taxa, palaeobiogeography and biological affinities. Alcheringa 4, 203-225.
Jones, P.J. & mCkenzie, k.g., 1981. Flemingopsis, a new name for the Cambrian phosphatocopine ostracode genus Flemingia Jones & McKenzie 1980. Alcheringa 5, 310-310.
kennaRD, J.m. & linDsay, J.F., 1991. Sequence stratigraphy of the latest Proterozoic-Cambrian Pertaoorrta Group, northern Amadeus Basin, central Australia. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 236, 171-194.
kouChinsky, a., Bengtson, s., Clausen, s., guBanov, a., malinky, J.m. & Peel, J.s., 2011. A middle Cambrian fauna of skeletal fossils from the Kuonamka Formation, northern Siberia.
Skovsted, Paterson & Brock 2014 and M. aspermachaera are also likely to have had only a single spine and hence are also distinguishable from this new taxon. The remaining species, M. henrikseni Skovsted & Peel 2001 and M. reticulatus Kouchinsky, Bengtson, Clausen, Gubanov, Malinky & Peel 2011, are known to have possessed multiple spines on a single valve. Mongolitubulus henrikseni is the most similar as it has larger spines on the valve surface, and numerous smaller spines arranged along the valve margin (compare Fig. 7A, B, F, G with fig. 1A-D of Skovsted & Peel 2001). However, both M. henrikseni and M. reticulatus have no valve ornamentation, a distinctive feature present on all valve surfaces of M. tunpere sp. nov. Previous authors (Skovsted & Peel 2001; Topper et al. 2007, 2013) have noted that the spines of Mongolitubulus species frequently display numerous types of ornamentation, suggesting this is not a taxonomically useful character. On the other hand, valve ornament is more consistent and therefore of greater taxonomic significance than the ornament adorning the spine. The ornamentation on the valve surface of M. tunpere sp. nov. is similar to M. aspermachaera in comprising raised subcircular structures, though in M. tunpere sp. nov. the nodes on top of these structures are much higher and rounded, giving them their mushroom-like appearance (compare Fig. 7B, D-F with fig. 4.1-4.20 of Topper et al. 2013). Interestingly, the ornament on the base of the spines shows a transition from the valve ornamentation, which slowly leans away from the spine’s apex to form the more typical Mongolitubulus scale-like ornamentation on the spine (Fig. 7J). No other species of Mongolitubulus shows this type of ornament transition.
Distribution. Giles Creek Dolostone, Amadeus Basin, Northern Territory; Cambrian Series 3, Stage 5; early Templetonian. Possibly the Gum Ridge Formation and Thorntonia Limestone in the Georgina Basin, and the Montejinni Limestone in the Wiso Basin; Series 2, Stage 4; Ordian.
Material. More than 100 specimens in total, including 14 specimens figured (9 valve fragments and over 100 spines) from 27.08 m to 71.70 m (Fig. 3) above the base of the Giles Creek Dolostone.
ACKNOWLEDGEMENTS We are indebted to the aboriginal land owners of Loves Creek Station for access to localities in the vicinity of Ross River Gorge. Field assistance was provided by Seung-bae Lee and Peter Gilham. We are also indebted to Dean Oliver (Dean Oliver Graphics) for the original drafting of Figs 1-3. This work has been supported by grants from the Macquarie University Postgraduate Fund (to PMS) and the Australian Research Council Discovery Project #129104251 (to GAB and JRP). We are grateful to Mark Williams and Ryszard Wrona for their constructive reviews of the manuscript.
REFERENCESBetts, m.J., toPPeR, t.P., valentine, J.l., skovsteD, C.B.,
PateRson, J.R. & BRoCk, g.a., 2014. A new early Cambrian bradoriid (Arthropoda) assemblage from the northern Flinders Ranges, South Australia. Gondwana Research 25, 420-437.
CaRon, J-B., smith, m.R. & haRvey, t.h.P., 2013. Beyond the Burgess Shale: Cambrian microfossils track the rise and fall of hallucigeniid lobopodians. Proceedings of the Royal Society B, 280 20131613, http://dx.doi.org/10.1098/rspb.201301613

AAP Memoir 45 (2014) 247
Alcheringa 35, 123-189.kRuse, P.D., 1998. Cambrian palaeontology of the eastern Wiso
and western Georgina Basins. Northern Territory Geological Survey Report 9, 68 p.
lauRie, J.R., 2004. Early Middle Cambrian trilobite faunas from NTGS Elkedra 3 corehole, southern Georgina Basin, Northern Territory. Memoirs of the Association of Australasian Palaeontologists 30, 221-260.
lauRie, J.R., 2006. Early Middle Cambrian trilobite faunas from Pacific Oil & Bas Baldwin 1 well, southern Georgina Basin, Northern Territory. Memoirs of the Association of Australasian Palaeontologists 32, 127-204.
lauRie, J.R. & sheRgolD, J.h., 1985. Phosphatic organisms and the correlation of Early Cambrian carbonate formations in central Australia. BMR Journal of Australian Geology & Geophysics 9, 83-89.
li, g., steineR, m., zhu, m. & zhao, X., 2012. Early Cambrian eodiscoid trilobite Hupeidiscus orientalis from South China: ontogeny and implications for affinities of Mongolitubulus-like sclerites. Bulletin of Geosciences 87, 159-169.
matthew, g.F., 1899. Preliminary notice of the Etcheminian fauna of Cape Breton. Bulletin of the New Brunswick Natural History Society 4, 198-208.
matthew, g.F., 1902. Ostracoda of the basal Cambrian rocks in Cape Breton. Canadian Record of Science 8, 437-470.
melnikova, l.m., 1990. Ranne- i Pozdnekembriyskie Bradoriida (ostrakody) severo-vostoka tsentralnogo Kazakhstana. Trudy Instituta Geologii i Geofiziki 765, 170-176.
melnikova, l.m., 2003. Cambrian Bradoriida (Arthropoda) of the Malyi Karatau (Kyr-Shabakty Section). Paleontological Journal 37, 57-61.
missaRzhevsky, v., 1977. Konodonty (?) I fosfatnye problematiki kembriya Mongolii I Sibiri. 10-19 in Tatarinov, L.P. (ed.), Bespozvonochnye Paleozoya Mongolii. Nauka, Moscow.
ÖPik, a.a., 1961. The geology and palaeontology of the headwaters of the Burke River, Queensland. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 53, 249 p.
ÖPik, a.a., 1968a. Ordian (Cambrian) Crustacea Bradoriida of Australia. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 103, 46 p.
ÖPik, a.a., 1968b. The Ordian Stage of the Cambrian and its Australian Metadoxididae. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 92, 133-170.
ÖPik, a.a., 1970. Redlichia of the Ordian (Cambrian) of Northern Australia and New South Wales. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 114, 67 p.
ÖPik, a.a., 1975. Templetonian and Ordian xystridurid trilobites of Australia. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 121, 84 p.
ÖPik, a.a., 1979. Middle Cambrian agnostids: systematics and biostratigraphy. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 172, v.1, 188 pp., v.2, 67 pls.
ÖPik, a.a., 1982. Dolichometopid trilobites of Queensland, Northern Territory and New South Wales. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 175, 85 p.
PateRson, J.R., 2005. Systematics of the Cambrian trilobite family Nepeidae, with revision of Australian species. Palaeontology 48, 479-517.
PateRson, J.R. & BRoCk, g.a., 2007. Early Cambrian trilobites from Angorichina, Flinders Ranges, South Australia, with a new assemblage from the Pararaia bunyerooensis Zone. Journal of Paleontology 81, 116-142.
Peng, s., BaBCoCk, l.e. & CooPeR, R.a., 2012. The Cambrian Period, 437-488 in Gradstein, F.M., Ogg, J.G., Schmitz, M.D. & Ogg, G.M. (eds), The Geologic Time Scale 2012, volume 2,
Elsevier, Oxford.RaymonD, P.e., 1935. Leanchoilia and other mid-Cambrian
arthropods. Bulletin of the Museum of Comparative Zoology 46, 203-230.
RoBeRts, J. & Jell, P.a., 1990. Early Middle Cambrian (Ordian) brachiopods of the Coonigan Formation, western New South Wales. Alcheringa 14, 257-309.
sheRgolD, J.h., 1986. Review of the Cambrian and Ordovician palaeontology of the Amadeus Basin, central Australia. Bureau of Mineral Resources, Geology and Geophysics, Report 276, 21 p.
sheRgolD, J.h., elPhinstone, R., lauRie, J.R., niColl, R.s., walteR, m.R., young, g.C. & zang w.l., 1991. Late Proterozoic and Early Palaeozoic palaeontology and biostratigraphy of the Amadeus Basin Bureau of Mineral Resources, Geology and Geophysics, Bulletin 236, 97-112.
sheRgolD, J.h., Jago, J.B., CooPeR, R.a. & lauRie, J.R., 1985. The Cambrian system in Australia, Antarctica, and New Zealand. International Union of Geological Sciences Publication 19, 85 p.
sieBolD, C.t.e. & stannius, h., 1845. Lehrbuch ver Vergleichenden Anatomie. Von Viet, Berlin, 679 p.
siveteR, D.J., waloszek, D. & williams, m., 2003. An early Cambrian phosphatocopid crustacean with three-dimensionally preserved soft parts from Shropshire, England. Special Papers in Palaeontology 70, 9-30.
siveteR, D.J. & williams, m., 1997. Cambrian bradoriid and phosphatocopid arthropods of North America. Special Papers in Palaeontology 57, 1-67.
skovsteD, C.B., 2005. Carapace of the bradoriid arthropod Mongolitubulus from the Early Cambrian of Greenland. GFF 127, 217-220.
skovsteD, C.B., BRoCk, g.a. & PateRson, J.R., 2006. Bivalved arthropods from the Lower Cambrian Mernmerna Formation, Arrowie Basin, South Australia and their implications for identification of Cambrian ‘small shelly fossils’. Memoirs of the Association of Australasian Palaeontologists 32, 7-41.
skovsteD, C.B. & Peel, J.s., 2001. The problematic fossil Mongolitubulus from the Lower Cambrian of Greenland. Bulletin of the Geological Society of Denmark 48, 135-147.
smith, P.m., 2012. Faunal dynamics and palaeoecology in relation to isotope evolution from the lower-middle Cambrian Pertaoorrta Group, Amadeus Basin, Northern Territory, Unpublished Honours Dissertation. Macquarie University, Sydney.
smith, t.e., kelman, a., lauRie, J.R., niColl, R.s., CaRR, l.k., hall, l. & eDwaRDs, D., in press. An updated stratigraphic framework for the Georgina Basin, Northern Territory and Queensland. APPEA Journal 53.
southgate, P.n. & sheRgolD, J.h., 1991. Application of sequence stratigraphic concepts to Middle Cambrian phosphogenesis, Georgina Basin, Australia. BMR Journal of Australian Geology & Geophysics 12, 119-144.
sylvesteR-BRaDley, P.C., 1961. Archaeocopida. 100-105 in MooRe, R.C. (ed.), Treatise on Invertebrate Paleontology Pt. Q, Arthropoda 3. Geological Society of America, Boulder and University of Kansas Press, Lawrence.
toPPeR, t.P., skovsteD, C.B., BRoCk, g.a. & PateRson, J.R., 2007. New bradoriids from the lower Cambrian Mernmerna Formation, South Australia: systematics, biostratigraphy and biogeography. Memoirs of the Association of Australasian Palaeontologists 33, 67-100.
toPPeR, t.P., skovsteD, C.B., BRoCk, g.a. & PateRson, J.R., 2011. The oldest bivalved arthropods from the early Cambrian of East Gondwana: Systematics, biostratigraphy and biogeography. Gondwana Research 19, 310-326.

AAP Memoir 45 (2014)248
toPPeR, t.P., skovsteD, C.B., haRPeR, D.a.t. & ahlBeRg, P., 2013. A bradoriid and brachiopod dominated shelly fauna from the Furongian (Cambrian) of Västergötland, Sweden. Journal of Paleontology 87, 69-83.
ulRiCh, e.o. & BassleR, R.s., 1931. Cambrian bivalved Crustacea of the Order Conchostraca. Proceedings of the US National Museum 78, 1-130.
vologin, a., leRmontova, e., yavoRsky, v. & yanishevsky, m., 1940. Atlas rukovodyashchikh form iskopaemykh faun SSSR. 1, Kembriy. Gosgeolizdat, Moscow. 194 p.
walCott, C.D., 1888. Cambrian fossils from Mount Stephen, northwest territory of Canada. American Journal of Science 36, 163-166.
walCott, C.D., 1916. Cambrian geology and paleontology III, No.5.- Cambrian trilobites. Smithsonian Miscellaneous Collections 64, 303-488.
wells, a.t., FoRman, D.J., RanFoRD, l.C. & Cook, P.J., 1970. Geology of the Amadeus Basin, central Australia. Bureau of Mineral Resources, Geology and Geophysics, Bulletin 100, 222 p.
wells, a.t., RanFoRD, l.C., stewaRt, a.J., Cook, P.J. & shaw, R.D., 1967. Geology pf the north-eastern part of the Amadeus Basin, Northern Territory. Bureau of Mineral Resources, Geology and Geophysics, Report 113, 93 p.
westeRgåRD, a.h., 1936. Paradoxides oelandicus beds of Öland. Sveriges Geologiska Undersokning Series C 394, 1-66.
whitehouse, F.w., 1936. The Cambrian Faunas of North-Eastern Australia. Part I, Stratigraphic outline. Part II, Trilobita (Miomera). Memoirs of the Queensland Museum 11, 59-112.
whitehouse, F.w., 1939. The Cambrian faunas of north-eastern
Australia. Part 3: the polymerid trilobites (with supplement no. 1). Memoirs of the Queensland Museum 11, 179-282.
williams, m. & siveteR, D.J., 1998. British Cambrian and Tremadoc bradoriid and phosphatocopid arthropods. Monograph of the Palaeontographical Society London 153, 49 p.
williams, m., siveteR, D.J., PoPov, l.e., vannieR, J.m.C., 2007. Biogeography and affinities of the bradoriid arthropods: Cosmopolitan microbenthos of the Cambrian seas. Palaeogeography, Palaeoclimatology, Palaeoecology 248, 202–232.
williams, m., vannieR, J., CoRBaRi, l. & massaBuau, J.-C., 2011. Oxygen as a driver of early arthropod micro-benthos evolution. PLoS ONE 6(12), e28183.
wRona, R., 2004. Cambrian microfossils from glacial erratics of King George Island, Antarctica. Acta Palaeontologica Polonica 49, 13-56.
wRona, R., 2009. Early Cambrian bradoriide and phosphatocopide arthropods from King George Island, West Antarctica: biogeographic implications. Polish Polar Research 30, 347-377.
zhang, w.t., 1953. Some Lower Cambrian trilobites from western Hubei. Acta Palaeontologica Sinica 1, 121-149.
zhang, w.t., 1974. Bradoriida. 107-111 in Nanjing Institute of Geology and Palaeontology (ed.), Handbook of stratigraphy and palaeontology of southwest China. Science Press, Beijing.
zhang, X.g., 1987. Moult stages and dimorphism of Early Cambrian bradoriids from Xichuan, Henan, China. Alcheringa 11, 1-19.
zhang, X.g., 2007. Phosphatized bradoriids (Arthropoda) from the Cambrian of China. Palaeontographica Abteilung A 281, 93-173.




















