
transylvania county, north carolina farmland protection plan
Upload
independentCategory
view
3download
0
This article was downloaded by: [Lincoln University Library]On: 28 September 2014, At: 14:17Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
New Zealand Journal of ZoologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnzz20
Effect of boundary type and season onpredatory arthropods associated withfield margins on New Zealand farmlandMH Bowiea, J Klimaszewskib, CJ Vinkc, S Hodgea & SD Wrattend
a Agriculture and Life Sciences Division, Lincoln University,Lincoln, New Zealandb Natural Resources Canada, Canadian Forest Service, LaurentianForestry Centre, Québec, Canadac Canterbury Museum, Christchurch, New Zealandd Bio-Protection Research Centre, Lincoln University, Lincoln, NewZealandPublished online: 24 Sep 2014.
To cite this article: MH Bowie, J Klimaszewski, CJ Vink, S Hodge & SD Wratten (2014): Effect ofboundary type and season on predatory arthropods associated with field margins on New Zealandfarmland, New Zealand Journal of Zoology, DOI: 10.1080/03014223.2014.953552
To link to this article: http://dx.doi.org/10.1080/03014223.2014.953552
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

RESEARCH ARTICLE
Effect of boundary type and season on predatory arthropods associated with fieldmargins on New Zealand farmland
MH Bowiea*, J Klimaszewskib, CJ Vinkc, S Hodgea and SD Wrattend
aAgriculture and Life Sciences Division, Lincoln University, Lincoln, New Zealand; bNatural Resources Canada,Canadian Forest Service, Laurentian Forestry Centre, Québec, Canada; cCanterbury Museum, Christchurch, NewZealand; dBio-Protection Research Centre, Lincoln University, Lincoln, New Zealand
(Received 6 December 2013; accepted 31 July 2014)
Pitfall traps were used to monitor predatory arthropod numbers along two types of field boundary, apost and wire fence line and a Cupressus macrocarpa hedge, along the same paddock margin inCanterbury, New Zealand, over 24 months. The seven most abundant predator groups recorded were:Araneae > Phalangiidae > Staphylinidae > Coccinellidae > Chilopoda > Hemerobiidae > Carabidae.Araneae, Phalangiidae, Staphylinidae, Chilopoda and Hemerobiidae were found in larger numbers atthe wire fence than at the hedge site, whereas the numbers of Carabidae and Coccinellidae adultsexhibited no field margin preference. However, more species of Araneae and Staphylinidae werecaught at the hedge site, whereas species richness of carabid beetles was greatest at the wire fence.Principal component analysis clearly separated the samples collected from the two habitats based onthe assemblages of Araneae, Staphylinidae and Carabidae, and certain species in each of thesetaxonomic groups appeared to be particularly associated with one boundary type or the other. All themain taxonomic groups exhibited clear seasonal patterns, with distinct peaks in abundance occurring atcertain times of the year. The results of the study reinforce the idea that management of fieldboundaries can be used to manipulate the type and abundance of particular groups of predatoryarthropods, and that seasonal patterns should be taken into account in schemes of integrated pestmanagement so that any adverse effects of biocide application on these beneficial species may beminimised.
Keywords: Araneae; boundaries; Carabidae; Chilopoda; Coccinellidae; Hemerobiidae; Phalangiidae;phenology; predators; Staphylinidae
Introduction
The potential of field boundaries to make substan-tial contributions to the total biodiversity of agro-ecosystems is widely recognised (Dennis et al.1994; Wratten et al. 1998; Thomas et al. 2001;Tscharntke et al. 2005; Pollard & Holland 2006;Eyre & Leifert 2011; Fukuda et al. 2011). Hedge-rows can provide ‘conservation corridors’, provid-ing a network of links with other hedgerows, and ameans of connectivity between remnant patches ofwoodland or restoration sites (Fischer et al. 2013).The diversity of resident invertebrates within field
boundaries can be influenced by many factors, suchas adjacent vegetation (Juen & Traugott 2004; Eyre& Leifert 2011), species composition (Fukuda et al.2011), vegetation structure (Wu&Cheng 2012) andage (Clements & Alexander 2009). Florally andstructurally diverse field boundaries can increasethe diversity of many invertebrate groups, such aspollinators (Boutin et al. 2011; Minarro & Prida2013), spiders and beetles (Fukuda et al. 2011; Wu& Cheng 2012). In a 2-year study, 830 species ofDiptera (almost 17% of the total British fauna) wererecorded in an 85 m length of mixed species
*Corresponding author. Email: [email protected]
New Zealand Journal of Zoology, 2014http://dx.doi.org/10.1080/03014223.2014.953552
© 2014 The Royal Society of New Zealand
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

hedgerow in southwest England (Wolton et al.2014).
The importance of management and manipula-tion of these field margins to support populations ofnatural enemies to enhance pest control has beenwell documented (Gurr et al. 1998; Landis et al.2000; Ponti et al. 2005; Pywell et al. 2005; Wrattenet al. 2013). Because field boundaries tend to beless subjected to disturbance from tillage and thedirect application of pesticides, they provide astable microhabitat for overwintering predatoryinvertebrates. Predatory beetles and spiders canbe abundant in weed strips and ‘beetle banks’(Thomas et al. 1992; Lys & Nentwig 1994; Frank& Reichhart 2004; Holland 2012), although someresearch has indicated that margins consisting ofdiverse woody vegetation such as hedgerows mayoffer a better refuge than grass or herb borders(Sotherton 1985; Dennis et al. 1994; Morandinet al. 2011).
In the northern hemisphere, the main inverteb-rate predators living in these field boundariesinclude ground beetles (Carabidae) (Thomas et al.2001; Millán de la Peña et al. 2003; Griffiths et al.2007), spiders (Araneae) (Wu & Cheng 2012;Fischer et al. 2013), rove beetles (Staphylinidae)(Meek et al. 2002; Griffiths et al. 2007), lacewings(Neuroptera) (Kiss et al. 1997) and harvestmen(Opiliones) (Meek et al. 2002). These groups alsooccur in New Zealand field boundaries, but muchless is known about the details of abundance,diversity and seasonality of these predatory inver-tebrates. On farmland in Canterbury, New Zealand,large numbers of spiders have been found alongshelter belts (McLachlan & Wratten 2003) andvideo monitoring has revealed a complex assem-blage of predatory invertebrates active under fieldboundaries (Merfield et al. 2004). In the NorthIsland of New Zealand, Fukuda et al. (2011) re-ported diverse assemblages of spiders and beetlesin field boundaries surrounding dairy pasture,and described how these assemblages could besignificantly affected by farm management and thecomposition of the boundaries in terms of exotic ornative plants.
In the Canterbury region of South Island, NewZealand, shelterbelts or windbreaks of Cupressus
macrocarpa Hartw. ex Gordon hedging (locallyreferred to as ‘macrocarpa’) are a common featureof the agricultural landscape. The aim of the presentwork was to compare the assemblages of predatoryinvertebrate species below a macrocarpa hedge withthat occurring under a post and wire fence along thesame area of pasture. Abundance, species richnessand seasonality of a number of arthropod groups(e.g. carabids, staphylinids, spiders) were continu-ally monitored over 2 years using pitfall traps, andthe structure of the assemblages in each habitatcompared using principal component analysis.
Methods
Field site
The study site was on the Lincoln University MixedCropping Farm (43.642°S, 172.466°E) in Canter-bury in central South Island, New Zealand. Inverte-brates were collected using pitfall traps placed intwo habitats: (1) 1 m recessed in from edge of a10 m wide, 7 m high Cupressus macrocarpa hedgeorientated east-west; and (2) along a post and wirefence line of the same paddock orientated north-south. The litter below the hedge was approxima-tely 10 cm deep and contained a few young nativeseedlings and exotic weeds. The wire fence wassprayed with herbicide in a strip approximately 3 mwide at approximately 3-month intervals and there-fore contained only sparse coverings of grasses andsmall amounts of leaf litter.
Sampling regime
Five 350 mL honey pots (6628NA, Stowers,Christchurch; diameter of opening 8 cm) wereused as traps along the two sites at intervals ofabout 10 m at each habitat. Each trap was seated in agalvanised sleeve to maintain a good trap/soilinterface and a galvanised iron roof was used toreduce rain and vegetative debris entering the trapsand minimise the removal of trapped invertebratesby birds. A 10% solution of sodium benzoate con-taining 2% surfactant (detergent) was used in thepitfalls as a preservative. The traps were emptiedevery 1–2 weeks and the contents sieved using a teastrainer (0.35 × 0.35 mm mesh), labelled and stored
2 MH Bowie et al.
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

in 70% ethanol until further processing. Trappingwas carried out over a 2-year period from 17September 1996 to 2 October 1998; a total of 7520trap days.
The total number of individuals in each of themain taxonomic groups was counted for the full24 months, apart from coccinellid larvae, whichwere recorded from 21 November 1997. Only thefirst year’s catch of spiders, carabids and staphy-linids were identified to species due to the largenumber of specimens obtained and the specialisedexpertise required for identification. Spiders wereidentified using the following taxonomic refer-ences: Forster & Wilton (1968), Forster (1970),Forster & Blest (1979), Forster et al. (1988), Vink(2002), Paquin et al. (2010), Vink et al. (2009)and Smith et al. (2012). Juvenile specimens ofErigone prominens Bösenberg & Strand, 1906 andErigone wiltoni Locket, 1973 could not be identi-fied to species and adults of the two speciescan sometimes be confused with each other; there-fore these two species were recorded as Erigonespp. Staphylinidae identifications were based onKlimaszewski et al. (1996) and Klimaszewski andWatt (1997). The carabid and coccinellid beetles,and other predators, were identified by direct com-parison to reference specimens in the collectionsof the Lincoln University Entomology ResearchMuseum.
Temperature and soil water
Tinytag and Tinytalk temperature data loggers (RSComponents Ltd, Auckland, New Zealand) wereplaced in each of the habitats beside a pitfall trapto record the temperature on the soil surface. Thedata loggers were used in each boundary type overthe period from 16 July 1998 to 14 September1998 for winter data, and from 1 November 1997to 31 January 1998 for summer data.
Soil water was measured beside each trap (i.e.five replicates for each of the wire fence and hedgesites) using time domain reflectometry moisturemeasuring equipment (Soil Moisture EquipmentCorp, California, USA) with 20 cm probes on 6April 1998 and 13 August 1998.
Statistical analysis
All statistical analyses were carried out usingMinitab v16 (Minitab Ltd, Pennsylvania, USA).Differences in abundance between the two habitatsfor the major taxonomic groups were analysedby a simple χ2 test based on total counts in eachhabitat over the whole monitoring period.
When examining seasonal patterns, abundancedata were converted to ‘specimens collected perday’ to correct for pitfall traps being in the fieldfor different durations.
To compare the structure of the assemblages ofthe three main taxonomic groups (i.e. spiders,carabid ground beetles and staphylinids) in eachhabitat, principal component analysis (PCA) wasperformed, using correlation matrices on untrans-formed data. The sample data used for these an-alyses consisted of the total catches of animals inindividual pitfall traps. The results of the PCAclearly separated the samples collected in each hab-itat along PCA axis 1. Thus, to establish whethercertain species had strong associations with a parti-cular habitat type, Pearson correlation coefficientswere calculated between the abundance of eachspecies in the pitfall samples and the respectivePCA axis 1 scores.
Differences in temperature and soil water con-tent between the two habitats were assessed using apaired t-test based on the data collected at each timepoint.
Results
The seven most abundant invertebrate predatorgroups recorded were: spiders (Araneae) > har-vestmen (Phalangiidae) > rove beetles (Staphylini-dae) > ladybirds (Coccinellidae) > centipedes(Chilopoda) > lacewings (Hemerobiidae) > groundbeetles (Carabidae) (Table 1). Greater numbers ofmost of these groups were collected at the postand wire site than at the hedgerow site (Table 1).Abundances of Carabidae and coccinellid adultswere not significantly different between the habi-tats, whereas coccinellid larvae were significantlymore abundant under the macrocarpa hedge(Table 1).
Effect of boundary type and season on predatory arthropods 3
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14
Spiders
Spiders were over four times more abundant thanother predator taxa counted along the wire fenceand the second most abundant taxon after rovebeetles under the hedge. The wire fence pitfallscaught almost twice as many individual spidersthan the hedge site. However, the hedgerow hadgreater overall species richness: 24 spider specieswere collected in total, with 22 of these found in thehedgerow and 15 along the wire fence (Tables 1, 2).Pitfall catches at the wire fence were dominatedby Anoteropsis hilaris (L. Koch, 1877), Erigone spp.and Tenuiphantes tenuis (Blackwall, 1852), whereasCantuaria n. sp. were predominantly (89%) found inpitfalls under the hedge.
The phenology of these species varied greatly.Anoteropsis hilaris abundance peaked in Februaryand was dominated by juveniles (Fig. 1A). ForErigone spp., abundance peaked in August and wasstrongly dominated bymales (Fig. 1B). Tenuiphantestenuis, although not as common as the other threespecies, had a more even distribution throughout theyear, with a small peak in May (Fig. 1C). Cantuarian. sp. was found between May and Septemberwith the exception of one juvenile found in March(Fig. 1D). All adult Cantuaria n. sp. were males.
Harvestmen
Only one species of Phalangiidae was recorded,Phalangium opilio Linnaeus, 1758, the common
European harvestman. More than twice as manyindividuals of this species were captured underthe wire fence than under the hedge, with thegreatest disparity between sites occurring in num-bers from the first year (Table 1; Fig. 2). Althoughpresent for most of the year, the greatest catchesof P. opilio occurred between January and April(Fig. 2).
Centipedes
Lamyctes emarginatus (Newport, 1844) was theonly chilopod caught in the traps and was found ingreater numbers under the wire fence (Table 1).There was a clear peak in abundance of this speciesbetween December 1996 and February 1997, witha similar but less dramatic peak during the samemonths the following year (Fig. 3).
Staphylinidae
Staphylinids were the most common predators inthe hedge (Tables 1, 3) and their abundance underthe hedge was more than three times as high asat the fence (Table 1; Fig. 4). Thirty-two species ofStaphylinidae were caught under the hedge com-pared with only 23 along the wire fence. There weredistinct peaks in abundance of staphylinids over thesummer months (December to February) in both ofthe sampling years, this being most prominent in1997–98 season (Fig. 4). The dominant subfamily
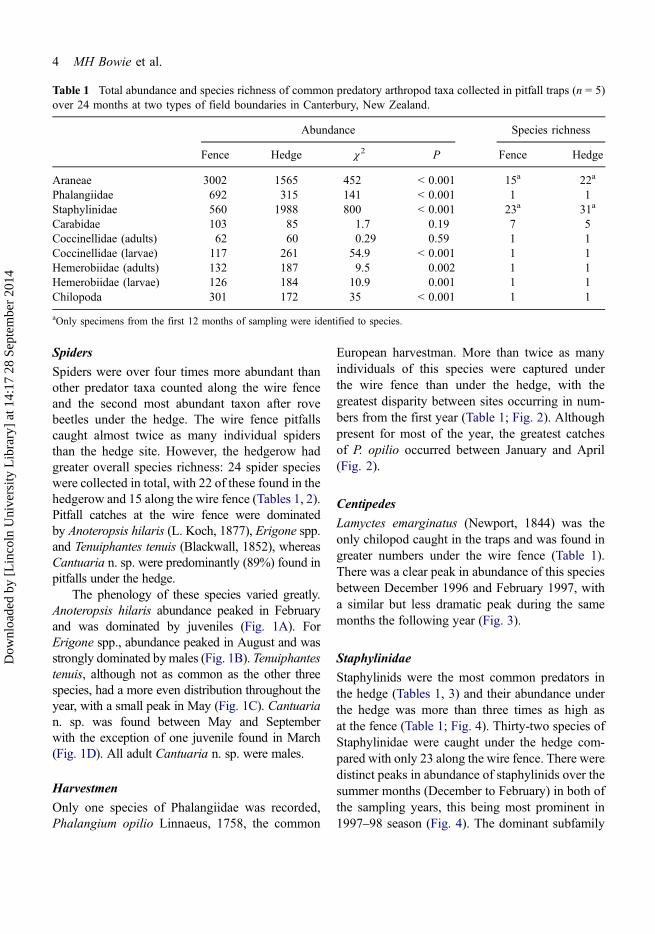
Table 1 Total abundance and species richness of common predatory arthropod taxa collected in pitfall traps (n = 5)over 24 months at two types of field boundaries in Canterbury, New Zealand.
Abundance Species richness
Fence Hedge χ2 P Fence Hedge
Araneae 3002 1565 452 < 0.001 15a 22a
Phalangiidae 692 315 141 < 0.001 1 1Staphylinidae 560 1988 800 < 0.001 23a 31a
Carabidae 103 85 1.7 0.19 7 5Coccinellidae (adults) 62 60 0.29 0.59 1 1Coccinellidae (larvae) 117 261 54.9 < 0.001 1 1Hemerobiidae (adults) 132 187 9.5 0.002 1 1Hemerobiidae (larvae) 126 184 10.9 0.001 1 1Chilopoda 301 172 35 < 0.001 1 1
aOnly specimens from the first 12 months of sampling were identified to species.
4 MH Bowie et al.
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

was Aleocharinae, which was represented by atleast 17 species, most of which were found underthe hedge (Table 3). Xantholininae were found ingreater numbers along the wire fence and were mostabundant between November and February (whichalso coincided with the presence of their larvae).Another common species, Tachyporus nitidulus(Fabricius, 1781), was found mainly under thehedge and its abundance peaked in December–January (Table 3).
Coccinellidae
Only one species of coccinellid was captured, the11-spotted ladybird, Coccinella undecimpunctata(Linnaeus, 1758), with similar numbers beingrecorded under the hedge and wire fence (Table 1;Fig. 5). The phenology of this species was notconsistent, for example a peak in adult abundance
seen in December 1997 was not observed theprevious year (Fig. 5). Larvae were more thantwice as abundant along the wire fence as underthe hedge (Table 1) and were found in a narrowperiod between November and January, peakingin December.
Carabidae
Seven carabid species were collected of which onlythree are native, Metaglymma moniliferum Bates,1867, Megadromus antarcticus (Chaudoir, 1865)and Hypharpax antarcticus (Laporte de Castelnau,1867) (Table 3). Metaglymma moniliferum was themost common carabid species found, with 26 and32 caught along the wire fence and hedge, respect-ively (Table 3). The Australian species, Hypharpaxaustralis, was the second most abundant carabid,with 50 of the 52 individuals being found along the
Table 2 Spiders collected in pitfall traps over 12 months at two types of field boundaries in Canterbury, NewZealand.
Family Species Wire fence Hedge
Corinnidae Supunna picta (L. Koch, 1873) 1 1Gnaphosidae Anzacia gemmea (Dalmas, 1917) 0 4
Nauhea tapa Forster, 1979a 1 1Zelanda kaituna (Forster, 1979)a 0 9Zelanda miranda (Forster, 1979)a 0 1
Hahniidae Rinawa cantuaria Forster (1970)a 2 13Idiopidae Cantuaria n. sp.a 24 189Linyphiidae Araeoncus humilis (Blackwall, 1841) 0 1
Erigone spp. 826 4Diplocephalus cristatus (Blackwall, 1833) 1 3Haplinis fucatina (Urquhart, 1894)a 35 2Tenuiphantes tenuis (Blackwall, 1852) 206 87Ostearius melanopygius (Pickard-Cambridge, 1879) 4 3
Lycosidae Anoteropsis hilaris (L. Koch, 1877)a 1069 49Micropholcommatidae unidentified sp.a 3 0Zoropsidae Uliodon albopunctatus L. Koch, 1873a 0 3Nemesiidae Stanwellia n. sp.a 0 9Salticidae unidentified sp. 1a 0 3
unidentified sp. 2a 0 6Theridiidae Cryptachaea veruculata (Urquhart, 1886) 0 2
Cryptachaea blattea (Urquhart, 1886) 5 26unidentified sp. 1a 4 28unidentified sp. 2a 1 0
aDenotes endemic species.
Effect of boundary type and season on predatory arthropods 5
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

Figure 1 Seasonal patterns in abundance of adult male, adult female and juveniles of most common spider taxa intheir ‘preferred’ habitat. Abundance is expressed as individuals per trap day. A, Anoteropsis hilaris under the wirefence. B, Erigone spp. under the wire fence. C, Tenuiphantes tenuis under the wire fence. D, Cantuaria n. sp. underthe macrocarpa hedge.
6 MH Bowie et al.
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

wire fence. Haplanister crypticusMoore, 1996 wassix times more frequent in the hedge site comparedwith along the wire fence site (Table 3). Only fourspecimens of Megadromus antarcticus were foundin total (Table 3).
As a group, carabids were present throughoutthe year but never very abundant, although thereappeared to be a slight increase in captures at thefence line in the winter months (May–September)(Fig. 6).
Figure 2 Seasonal patterns in abundance of harvestmen (Phalangium opilio) collected by pitfall trapping under awire fence and macrocarpa hedge in Canterbury, New Zealand. Abundance is expressed as mean individuals per trapday over each calendar month.
Figure 3 Seasonal patterns in abundance of centipedes (Lamyctes emarginatus) collected by pitfall trapping under awire fence and macrocarpa hedge in Canterbury, New Zealand. Abundance is expressed as mean individuals per trapday over each calendar month.
Effect of boundary type and season on predatory arthropods 7
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

Table 3 Coleoptera collected in pitfall traps located under a hedge and along a wire fence over 12 months(September 1996 to September 1997) in Canterbury, New Zealand.
Family Subfamily Species Wire fence Hedge
Carabidae Broscinae Metaglymma moniliferum Bates, 1867 31 43Harpalinae Anisodactylus binotatus (Fabricius, 1787) 1 1
Haplanister crypticus Moore, 1996 5 30Hypharpax antarcticus (Laporte de Castelnau, 1867) 4 0Hypharpax australis (Dejean, 1829) 50 2Megadromus antarcticus (Chaudoir, 1965) 1 3
Scaritinae Clivina vegans Putzeys, 1866 15 0Coccinellidae Coccinella undecimpunctata L. 1758 60 62
Coccinella undecimpunctata (larvae) 261 117Staphylinidae Aleocharinae Amischa analis (Gravenhorst, 1802) 0 45
Amischa decipiens (Sharp, 1869) 2 32Amischa nigrofusca (Stephens, 1832) 0 28Amischa sp.1 7 87Atheta amicula (Stephens, 1832) 9 29Atheta coriaria (Kraatz, 1856) 1 0Atheta fungi (Gravenhorst, 1806) 1 41Atheta (Liogluta) sp.1 2 0Atheta nigra (Kraatz, 1856) 0 15Atheta sordida Marsham, 1802 1 6Atheta sp.1 2 26Colle sp.? 0 1Dasytricheta sp.? 0 1Oligota pumilio Kiesenwetter, 1858 0 2Oligota sp.1 0 9Oligota sp.1 6 74Oxpoda haemorrhoa (Mannerheim, 1830) 16 25
Omaliinae Omaliomimus sp.? 0 1Oxytelinae Anotylus vinsoni (Cameron, 1936) 5 1
Anotylus sp.1 1 19Anotylus sp.2 5 2Carpelimus sp.1 7 17Carpelimus sp.2 0 1Carpelimus sp.3 3 4Carpelimus sp.4 0 1
Paedarinae Lithocharis sp.1 1 1Sunius sp.1 2 1
Pselaphinae Unknown gen. sp. 1 0Staphylininae Gabrius nigritulus (Gravenhorst, 1802) 0 1
Grabius sp.1 1 1Ocypus ater (Gravenhorst, 1802) 1 1Philonthus rectangulus Sharp, 1874 0 1
Tachyporinae Tachyporus nitidulus (Fabricius, 1781) 16 51Xantholininae Xantholinus linearis (Olivier, 1795) 24 6
Xantholinus sp.1 72 1Xantholinus (larvae) 19 0
8 MH Bowie et al.
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

Hemerobiidae
Only one species was recorded, the Tasmanianlacewing Micromus tasmaniae (Walker, 1860).Both adults and larvae were found in largernumbers in the hedge than in the wire fence pitfall
traps (Table 1). There was a conspicuous peak inabundance of adults (and larvae) from November1997 to February 1998 (Fig. 7). However, a similarseasonal peak was not observed the previous year(Fig. 7).
Figure 4 Seasonal patterns in abundance of staphylinid rove beetles collected by pitfall trapping under a wire fenceand macrocarpa hedge in Canterbury, New Zealand. Abundance is expressed as mean individuals per trap day overeach calendar month.
Figure 5 Seasonal patterns in abundance of Coccinellidae (Coccinella undecimpunctata) adults collected by pitfalltrapping under a wire fence and macrocarpa hedge in Canterbury, New Zealand. Abundance is expressed as meanindividuals per trap day over each calendar month.
Effect of boundary type and season on predatory arthropods 9
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

Multivariate analysis
The principal component analysis used the totalcatches of invertebrates in each individual pitfalltrap as the sample unit. The collections of spiders,
carabids and staphylinids from the two boundaryhabitats were all clearly separated along the firstPCA axis (Fig. 8). For each taxonomic group,certain species were determined as having strong
Figure 6 Seasonal patterns in abundance of Carabidae collected by pitfall trapping under a wire fence andmacrocarpa hedge in Canterbury, New Zealand. Abundance is expressed as mean individuals per trap day over eachcalendar month.
Figure 7 Seasonal patterns in abundance of lacewing adults (Micromus tasmaniae) collected by pitfall trappingunder a wire fence and macrocarpa hedge in Canterbury, New Zealand. Abundance is expressed as mean individualsper trap day over each calendar month.
10 MH Bowie et al.
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

associations with habitat by calculation of correla-tion coefficients between their abundance in eachtrap and the PCA axis 1 score for that pitfall trap(Table 4). Some species, such as the salticidspiders and staphylinids such as Amischa analis(Gravenhorst, 1802), were exclusive to the hedge.Other species, such as the wolf spider Anoteropsishilaris and the carabid beetle Haplanister crypti-cus, occurred in both habitats, but were signifi-cantly more abundant, and found consistentlyamong the samples, in one particular boundarytype (Tables 2, 3).
Environmental conditions
In the summer, the wire fence daily minimumtemperatures and, especially, daily maximum tem-peratures at ground level were significantly higher(P < 0.01) than those under the hedge (Fig. 9A).In contrast, with the exception of the maximumtemperatures in September 1998, the winter meandaily minimum temperatures were significantlyhigher under the hedge (P < 0.001) comparedwith the wire fence line (Fig. 9B). Thus, the hedgeprovides a less variable habitat, having a coolingeffect in summer and a warming effect in winter.The wire fence line experienced 11 frosts during themonitoring period, compared with none occurringunder the hedge.
In the two measurements that were taken,moisture levels were higher at the wire fence sitethan under the hedge. Volumetric soil moisturecontent (θv, cm3 cm–3) at a 0–15 cm depth at thewire fence and the hedge sites on 6 April 1998were 0.259 (±0.003 95% CI) and 0.106 (0.00395% CI), respectively. On 13 August 1998, thesoil moisture in the fence line and under the hedgewere 0.345 (±0.07 95% CI) and 0.166 (±0.05 95%CI), respectively.
Discussion
The results of this survey indicate that the assem-blages of predatory invertebrates under a wire fenceand macrocarpa hedge differed both in terms ofabundance and structure. Some groups, such as
Figure 8 Plots of principal component analysisscores (PCA axis 2 vs PCA axis 1) obtained fromanalysis of the total catches of A, spiders, B,carabids and C, staphylinids in pitfall traps set undera wire fence or macrocarpa hedge over a 12-monthperiod.
Effect of boundary type and season on predatory arthropods 11
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

spiders, harvestmen and carabids were more abund-ant under the wire fence, whereas other taxa, suchas Staphylinidae and lacewings, were more abund-ant under the hedge. Various species had clearassociations with one boundary type or the otherand in some cases were recorded in only one of thehabitats. This effect of field boundary type onmodifying invertebrate assemblages has beenreported by many other researchers. For example,Wu & Cheng (2012) found in a Chinese study thatthe floral composition of the hedge affected com-munities of spiders; Morandin et al. (2011) reportedthat woody hedgerows contained more beneficialinsects than did weedy field boundaries; andGriffiths et al. (2007) established that many gener-alist predators were at highest densities under wirefences compared with hedgerows on farms in theUK. In New Zealand, the fact that native species ofspiders and beetles were found to be more abundantwhen field boundaries consisted of native species asopposed to exotic plants, and that fencing off theboundary from grazing animals increased spiderabundance, is possibly as a consequence of thetaller ground vegetation (Fukuda et al. 2011).
The deep litter layer and herbs below the hedge,compared with the herbicide-induced sparsity ofplants along the wire fence, may have contributedto the differences in invertebrate fauna betweenthe two habitats (Fukuda et al. 2011). In terms ofenvironmental influences, in this study, soil mois-ture levels and temperatures differed markedlybetween the two boundary types, with conditionsunder the hedge appearing to be less extreme thanthose under the wire fence. The differences indistribution of some taxa between the boundarytypes may be associated with these environmental
differences if taxa have different tolerance thresh-olds, or respond differently to extreme events suchas frosts. Many seasonal patterns in invertebrateabundance are linked to the trends in daily tem-peratures and day length, and Holland et al. (2007)reported that abundance and emergence patterns ofpredatory beetles in arable fields in the UK werelinked to soil moisture.
An indirect indication that responses to envir-onmental cues are affecting the activity of inverte-brates in these boundary habitats is provided bythe clear seasonality observed for most of themajor groups. Whereas Carabidae adults wereobserved throughout the sampling period (Hutch-ison 2007), the staphylinid beetles and coccinel-lids were much more seasonal, with clear peaks inabundance. The spiders also showed distinct peaksin abundance over the sampling period, althoughthese tended to be different times of the year forthe different species (Fig. 1; Miliczky et al. 2008).Seasonality of spiders in agro-ecological settings,often with a single sharp peak in abundance, hasbeen described by many previous researchers(e.g. Marshall et al. 2002; Reed & Nicholas2008) and has been explained by responses toenvironmental factors such as temperature and daylength (Cardoso et al. 2007) and also externalfactors such as time of crop harvest, whenindividuals seek refuge in relatively undisturbedfield boundaries (Wu & Cheng 2012). For Can-tuaria n. sp., the obvious peak in abundance isrelated to mating behaviour, as this species norm-ally resides in its lidded burrow except whenmales seek mates between May and September(Forster & Wilton 1968; C.J. Vink, pers. obs).
Table 4 Species exhibiting strong associations with either boundary habitat (rp; correlation coefficient betweenabundance and PCA axis 1 score; Fig. 8).
Group Hedge species Wire fence species
Araneae Taieria kaituna (0.99) Tenuiphantes tenuis (–0.74)Carabidae Metaglymma moniliferum (0.66) Hypharpax antarcticus (–0.72)Staphylinidae Amischa decipiens (0.96) Xantholinus sp. 2 (–0.88)
Amischa nigrofusca (0.89) Xantholinus sp. 3 (–0.81)
12 MH Bowie et al.
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

Although it is generally accepted that theapplication of biocides should be avoided whenpollinators such as honeybees and bumble bees areactive, less consideration tends to be given tominimising the adverse effects on predatory speciesby applying insecticides when they are least exposed(Horton et al. 2012). The seasonal trends recorded inthis study reinforce the need to not only account forspatial differences in abundance of predatory speciesbut also temporal differences when implementingintegrated pest management strategies.
The collections of invertebrates in terms ofabundance and species richness were dominatedby spiders and staphylinid beetles, with thecarabid beetles the only other major group playinga major role in overall diversity. These taxonomicgroups are typical of field boundaries in agricul-tural settings and can also play a significant role inthe natural control of crop pests.
Of the spiders, Tenuiphantes tenuis, Anoter-opsis hilaris and Erigone spp. comprised morethan 85% of the total individuals collected in this
Figure 9 Mean daily maximum and minimum temperatures (°C) at ground level at wire fence and hedgeboundaries A, in summer (November 1997 and January 1998) and B, in winter (July and September 1998) (mean +se) in Canterbury, New Zealand. Comparisons were made by paired t-test. ***P < 0.001; **P < 0.01; ns, notsignificant.
Effect of boundary type and season on predatory arthropods 13
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

study, and all three were also found in the NorthIsland shelter belts studied by Fukuda et al.(2011). Other studies in pasture and arable cropsin Canterbury have similarly found that T. tenuisand the native wolf spider A. hilaris occurfrequently in samples and are often abundant(Sivasubramaniam et al. 1997; Hodge & Vink2000; Vink et al. 2004). As generalist predators,A. hilaris, T. tenuis and Erigone spp. would tendto feed on a range of pasture arthropods includingsome pest species. For example, a recent studyusing PCR gut analysis demonstrated that T. tenuisis a potentially significant predator of the pestweevil, Listronotus bonariensis in Canterburypastures (Vink & Kean 2013). The trapdoor spider(Cantuaria n. sp.) was caught in large numbers inthe hedge pitfalls in the current study and, likemost spiders, Cantuaria n. sp. feeds on a widerange of invertebrate species (such as adults ofgrass grub Costelytra zealandica [White, 1846])and has been suggested to be an effective predatorin undisturbed pasture (Irish 2001).
In a similar study in the UK, Griffiths et al.(2007) found 23 staphylinid species in fence linesand 24 species under hedges, very similar to the 23and 32 species found under the wire fence andhedge, respectively, in this study. In terms ofbiocontrol, the predatory role of Staphylinidae canbe spatially and temporally complex. Aphid preda-tion by Philonthus and Tachyporus, for example,can be dependent on their location on the wheatplant, timing of dispersal flights and the presence ofalternative foods such as fungi (Dennis & Sotherton1994). Members of the genera belonging to thesubtribe Philonthina, particularly those of the genusPhilonthus, are considered general predators, orpredators favouring Diptera larvae (e.g. Smetana1958, 1995). In our system, Xantholinus mayprovide the greatest biological control benefits dueto their relatively large body size and high densitiesat the wire boundaries. Xantholinus linearis (Oliv-ier, 1795) is known to be a predator of eggs andlarvae of the pest grass grub Costelytra zealandicain New Zealand (East et al. 1981).
Little appears to have been written on thecarabids collected in the study. Megadromus ant-arcticus is a predator of slugs and lepidopteran
larvae under laboratory conditions (Chapman et al.1997; Hodge et al. 1999), although only a fewindividuals of this species were caught in this studyand their impact on pest populations would be slightat these densities. The introduced Anisodactylusbinotatus (Fabricius, 1787) feeds on molluscs andwas reported by Pilgrim (1963) to be already fairlywidespread in Canterbury in the early 1960s. Theomnivorous Hypharpax australis (Dejean, 1829) iswidespread in New Zealand open country includingagro-ecosystems (Larochelle & Larivière 2001).
Both adults and larvae of the 11-spotted lady‐bird, Coccinella undecimpunctata and the Tasma-nian lacewing Micromus tasmaniae are consideredvoracious predators of many crop pests, especiallyaphids, and are often implicated in biocontrol andintegrated pest management programmes in NewZealand (e.g. Fagan et al. 2010; Jacometti et al.2010; Wearing et al. 2010).
Conclusions
In general, the predatory arthropods of fieldboundaries in New Zealand are poorly investi-gated and it is often only speculated as to whatextent the diverse arthropod fauna of field bound-aries actually contributes to pest management.This survey of beneficial arthropods inhabitingtypical wire fence and hedge field margins inCanterbury has identified which taxa appearsufficiently abundant to potentially influence pestpopulations, highlighted key differences in thefauna between the two habitats and described clearseasonality in the activity of some groups. Untilrecently, hedges in New Zealand were dominatedby non-native, woody species such as macrocarpaand gorse (Ulex europaeus L.) and it is unsurpris-ing that in agricultural systems dominated byexotic plant species the fauna in these fieldboundaries contains many introduced or adventivespecies. In other regions, such as Europe, theconservation, restoration and management ofdiverse, sometimes ancient, hedgerows consistingof native woody plants is considered of majorimportance in agricultural landscapes, and theirrole in preserving native faunal and floral diversityhas long been recognised (e.g. Millán de la Peña
14 MH Bowie et al.
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

et al. 2003; Pywell et al. 2005). Currently, the useof native plantings as field margins and alongriparian borders is gaining momentum, a practicethat appears to be at least partially linked to theconversion of farmland to intensive dairying andthe subsequent need to manage nutrient leaching(Fukuda et al. 2011; Hahner et al. 2014). Theeffects of replacing fences or exotic hedges withnative plant species on the resident invertebratecommunities warrants further investigation, espe-cially in terms of the enhancement of nativeinvertebrate species and those exotic species thatare beneficial to the agricultural systems of whichthey are part.
Acknowledgements
We thank Rowan Emberson for his assistance withCarabidae identification and Roger Cresswell in the Soiland Physical Sciences Department at Lincoln Universityfor loaning the Time Domain Reflectometer and instruc-tion on its use. We also thank Anthony White (HeinzWattie’s) for his support of the research on KowhaiFarm. We also thank the referees for their valuablesuggestions.
References
Boutin C, Baril A, McCabe SK, Martin PA, Guy M2011. The value of woody hedgerows for mothdiversity on organic and conventional farms.Environmental Entomology 40: 560–569.
Cardoso P, Silva I, De Oliveira NG, Serrano ARM2007. Seasonality of spiders (Araneae) in Mediter-ranean ecosystems and its implications in theoptimum sampling period. Ecological Entomology32: 516–526.
Chapman RB, Simeonidis AS, Smith JT 1997. Evaluationof metallic green ground beetles as a predator ofslugs. Proceedings of the 50th NZ Plant ProtectionConference. Lincoln, New Zealand Plant ProtectionSociety. Pp. 51–55.
Clements DK, Alexander KNA 2009. A comparativestudy of the invertebrate faunas of hedgerows ofdiffering ages, with particular reference to indica-tors of ancient woodland and ‘old growth’. TheJournal of Practical Ecology and Conservation 8:7–27.
Dennis P, Sotherton NW 1994. Behavioural aspects ofstaphylinid beetles that limit their aphid feedingpotential in cereal crops. Pedobiologia 38: 222–237.
Dennis P, Thomas MB, Sotherton N 1994. Structuralfeatures of field boundaries which influence theoverwintering densities of beneficial arthropodpredators. The Journal of Applied Ecology 31:361–370.
East R, King PD, Watson RN 1981. Population studiesof grass grub (Costelytra zealandica) and blackbeetle (Heteronychus arator) (Coleoptera: Scara-baeidae). New Zealand Journal of Ecology 4:56–64.
Eyre AU, Leifert C 2011. Crop and field boundaryinfluences on the activity of a wide range ofbeneficial invertebrate groups on a split conven-tional/organic farm in northern England. Bulletinof Entomological Research 101: 135–144.
Fagan LL, McLachlan A, Till CM, Walker MK 2010.Synergy between chemical and biological controlin the IPM of currant-lettuce aphid (Nasonoviaribisnigri) in Canterbury, New Zealand. Bulletin ofEntomological Research 100: 217–223.
Fischer C, Schlinkert H, Ludwig M, Holzschuh A, GalleR, Tscharntke T et al. 2013. The impact of hedge-forest connectivity and microhabitat conditions onspider and carabid beetle assemblages in agricul-tural landscapes. Journal of Insect Conservation17: 1027–1038.
Forster RR 1970. The spiders of New Zealand. Part III.Otago Museum Bulletin 3: 1–184.
Forster RR, Blest AD 1979. The spiders of NewZealand. Part V. Otago Museum Bulletin 5: 1–173.
Forster RR, Millidge AF, Court DJ 1988. The spiders ofNew Zealand. Part VI. Otago Museum Bulletin 6:1–124.
Forster RR, Wilton CL 1968. The spiders of NewZealand. Part II. Otago Museum Bulletin 2: 1–180.
Frank T, Reichhart B 2004. Staphylinidae and Carabi-dae overwintering in wheat and sown wildflowerareas of different age. Bulletin of EntomologicalResearch 94: 209–217.
Fukuda Y, Moller H, Burns B 2011. Effects of organicfarming, fencing and vegetation origin on spidersand beetles within shelterbelts on dairy farms. NewZealand Journal of Agricultural Research 54:155–176.
Griffiths GJK, Winder L, Holland JM, Thomas CFG,Williams E 2007. The representation and functionalcomposition of carabid and staphylinid beetles indifferent field boundary types at a farm-scale.Biological Conservation 135: 145–152.
Gurr GM, van Emden HF, Wratten SD 1998. Habitatmanipulation and natural enemy efficiency: impli-cations for the control of pests. In: Barbosa P ed.Conservation biological control. San Diego, Aca-demic Press. Pp. 155–183.
Hahner JL, Robinson BH, Hong-Tao Z, Dickinson NM2014. The phytoremediation potential of native
Effect of boundary type and season on predatory arthropods 15
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

plants on New Zealand dairy farms. InternationalJournal of Phytoremediation 16: 719–734.
Hodge S, Vink CJ 2000. An evaluation of Lycosahilaris as a bioindicator of organophosphate insect-icide contamination. New Zealand Plant Protection53: 226–229.
Hodge S, Wratten S, Smith J, Thomas M, Frampton C1999. The role of leaf wounding and an epigealpredator on caterpillar damage to tomato plants.Annals of Applied Biology 134: 137–141.
Holland J 2012. Rediscovering beetle banks. Game &Wildlife Review 44: 48–51.
Holland JM, Thomas CFG, Birkett T, Southway S 2007.Spatio-temporal distribution and emergence ofbeetles in arable fields in relation to soil moisture.Bulletin of Entomological Research 97: 89–100.
Horton DR, Miliczky ER, Jones VP, Baker CC, UnruhTR 2012. Diversity and phenology of the generalistpredator community in apple orchards of CentralWashington State (Insecta, Araneae). CanadianEntomologist 144: 691–710.
Hutchison MAS 2007. Seasonality and life histories oftwo endemic New Zealand carabid beetles(Coleoptera: Carabidae): Mecodema oconnoriBroun and Megadromus capito (White). NewZealand Journal of Zoology 34: 79–89.
Irish LL 2001. The minefield spiders. Dunedin, Biodi-versity Solutions. 232 p.
Jacometti M, Jorgensen N, Wratten S 2010. Enhancingbiological control by an omnivorous lacewing:floral resources reduce aphid numbers at low aphiddensities. Biological Control 55: 159–165.
Juen A, Traugott M 2004. Spatial distribution ofepigaeic predators in a small field in relation toseason and surrounding crops. Agriculture Ecosys-tems and Environment 103: 613–620.
Kiss J, Penksza K, Toth F, Kader F 1997. Evaluation offields and field margins in nature productioncapacity with special regard to plant protection.Agriculture, Ecosystems & Environment 63:227–232.
Klimaszewski J, Newton AF, Thayer MK 1996. Areview of the New Zealand rove beetles (Coleop-tera: Staphylinidae). New Zealand Journal ofZoology 23: 143–160.
Klimaszewski J, Watt JC 1997. Coleoptera: family-group review and keys to identification. Fauna ofNew Zealand 37: 1–199.
Landis DA, Wratten SD, Gurr GM 2000. Habitatmanagement to conserve natural enemies of arth-ropod pests in agriculture. Annual Review ofEntomology 45: 175–201.
Larochelle A, Larivière MC 2001. Carabidae (Insecta:Coleoptera): catalogue. Fauna of New Zealand 43:1–285.
Lys J-A, Nentwig W 1994. Improvement of the over-wintering sites for Carabidae, Staphylinidae andAraneae by strip management in a cereal field.Pedobiologia 38: 238–242.
Marshall SD, Pavuk DM, Rypstra AL 2002. A compar-ative study of phenology and daily activity patternsin the wolf spiders Pardosa milvina and Hognahelluo in soybean agroecosystems in southwesternOhio (Araneae, Lycosidae). Journal of Arachnol-ogy 30: 503–510.
McLachlan ARG, Wratten SD 2003. Abundance andspecies richness of field-margin and pasture spiders(Araneae) in Canterbury, New Zealand. NewZealand Journal of Zoology 30: 57–67.
Meek B, Loxton D, Sparks T, Pywell R, Pickett H,Nowakowski M 2002. The effect of arable fieldmargin composition on invertebrate biodiversity.Biological Conservation 106: 259–271.
Merfield CN, Wratten SD, Navntoft S 2004. Videoanalysis of predation by polyphagous invertebratepredators in the laboratory and field. BiologicalControl 29: 5–13.
Miliczky ER, Horton DR, Calkins C 2008. Observationson phenology and overwintering of spiders asso-ciated with apple and pear orchards in south-central Washington. Journal of Arachnology 36:565–573.
Millán de la Peña N, Butet A, Delettre Y, Morant P,Burel F 2003. Landscape context and carabidbeetles (Coleoptera: Carabidae) communities ofhedgerows in Western France. Agricultural Eco-systems & Environment 94: 59–72.
Minarro M, Prida E 2013. Hedgerows surroundingorganic apple orchards in north-west Spain: poten-tial to conserve beneficial insects. Agricultural andForest Entomology 15: 382–390.
Morandin L, Long RF, Pease C, Kremen C 2011.Hedgerows enhance beneficial insects on farms inCalifornia's Central Valley. California Agriculture65: 197–201.
Paquin P, Vink CJ, Dupérré N 2010. Spiders of NewZealand: annotated family key & species list.Lincoln, Manaaki Whenia Press. vii + 118 p.
Pilgrim RLC 1963. Anisodactylus binotatus Fabr, acarabid beetle new to New Zealand, and a reviewof the exotic carabid fauna. Pacific Insects 5:837–847.
Pollard KA, Holland JM 2006. Arthropods within thewoody element of hedgerows and their distributionpattern. Agricultural and Forest Entomology 8:203–211.
Ponti L, Ricci C, Veronesi F, Torricelli R 2005. Naturalhedges as an element of functional biodiversity inagroecosystems: the case of a Central Italy vine-yard. Bulletin of Insectology 58: 19–23.
16 MH Bowie et al.
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14

Pywell RF, James KL, Herbert I, Meek WR, Carvell C,Bell D et al. 2005. Determinants of overwinteringhabitat quality for beetles and spiders on arablefarmland. Biological Conservation 123: 79–90.
Reed DH, Nicholas AC 2008. Spatial and temporalvariation in a suite of life-history traits in twospecies of wolf spider. Ecological Entomology33: 488–496.
Sivasubramaniam W, Wratten SD, Klimaszewski J.1997. Species composition, abundance, and activ-ity of predatory arthropods in carrot fields, Canter-bury, New Zealand. New Zealand Journal ofZoology 24: 205–212.
Smetana, A. 1958. Die Larve von Philonthus politus L.(Col., Staphylinidae). Norsk Entomologisk Tids-skrift 10: 202–206.
Smetana, A. 1995. Rove beetles of the subtribePhilonthina of America north of Mexico (Coleop-tera: Staphylinidae). Classification, phylogeny andtaxonomic revision. Memoirs on Entomology,International 3. x + 946 p.
Smith HM, Vink CJ, Fitzgerald BM, Sirvid PJ 2012.Redescription and generic placement of the spiderCryptachaea gigantipes (Keyserling, 1890) (Ara-neae: Theridiidae) and notes on related synanthro-pic species in Australasia. Zootaxa 3507: 38–56.
Sotherton, NW 1985. The distribution and abundance ofpredatory coleoptera overwintering in field bound-aries. Annals of Applied Biology 106: 17–21.
Thomas CFG, Parkinson L, Griffiths GJK, Fernandez-Garcia A, Marshall EJP 2001. Aggregation andtemporal stability of carabid beetle distributions infield and hedgerow habitats. Journal of AppliedEcology 38: 100–116.
Thomas MB, Wratten SD, Sotherton NW 1992. Cre-ation of ‘Island’ habitats in farmland to manipulatepopulations of beneficial arthropods: predatordensities and species composition. The Journal ofApplied Ecology 29: 524–531.
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I,Thies C 2005. Landscape perspectives on agricul-tural intensification and biodiversity – ecosystemservice management. Ecology Letters 8: 857–874.
Vink CJ 2002. Lycosidae (Arachnida: Araneae). Faunaof New Zealand 44: 1–94.
Vink CJ, Dupérré N, Paquin P, Fitzgerald BM, Sirvid PJ2009. The cosmopolitan spider Cryptachaea blat-tea (Urquhart 1886) (Araneae: Theridiidae): rede-scription, including COI sequence, and newsynonymy. Zootaxa 2133: 55–63.
Vink CJ, Kean JM 2013. PCR gut analysis reveals thatTenuiphantes tenuis (Araneae: Linyphiidae) is apotentially significant predator of Argentine stemweevil, Listronotus bonariensis (Coleoptera: Cur-culionidae), in New Zealand pastures. New Zeal-and Journal of Zoology 40: 304–313.
Vink CJ, Teulon DAJ, McLachlan ARG, StufkensMAW 2004. Spiders (Araneae) and harvestmen(Opiliones) in arable crops and grasses in Canter-bury, New Zealand. New Zealand Journal ofZoology 31: 149–159.
Wearing CH, Attfield BA, Colhoun K 2010. Biologicalcontrol of woolly apple aphid, Eriosoma lanigerum(Hausmann), during transition to integrated fruitproduction for pipfruit in Central Otago, NewZealand. New Zealand Journal of Crop and Horti-cultural Science 38: 255–273.
Wolton RJ, Bentley H, Chandler PJ, Drake CM, KramerJ, Plant AR et al. 2014. The diversity of Dipteraassociated with a British hedge. Dipterists Digest21: 1–36.
Wratten SD, van Emden HF, Thomas MB 1998. Within-field and border refugia for the enhancement ofnatural enemies. In: Pickett CH, Bugg RL eds.Enhancing biological control. Berkeley, Universityof California Press. Pp. 375–403.
Wratten SD, Sandhu H, Cullen R, Costanza R, eds.2013. Ecosystem services in agricultural and urbanlandscapes. Chichester, Wiley-Blackwell.
Wu YH, Cheng X. 2012. Spider community structuresin different types of hedgerows in the purple soilhilly areas of Sichuan Basin. Journal of Ecologyand Rural Environment 28: 661–668.
Effect of boundary type and season on predatory arthropods 17
Dow
nloa
ded
by [
Lin
coln
Uni
vers
ity L
ibra
ry]
at 1
4:17
28
Sept
embe
r 20
14
Copyright © 2022 FDOKUMEN




















