
Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes. Effects of Exogenous Fatty Acids on the...
11
Eur. J. Biochem. 54, 373-383 (1975) Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes Effects of Exogenous Fatty Acids on the Content, Synthesis and Degradation of the Enzyme Kaoru KITAJIMA, Shin-ichi TASHIRO, and Shosaku NUMA Department of Medical Chemistry, Kyoto University Faculty of Medicine (Received December 9, 1974/February 13, 1975) Studies were made on the content, synthesis and degradation of acetyl-coenzyme-A carboxylase in JTC-25 . P3 cells, hepatocytes which can be maintained in a protein-free and lipid-free chemically defined medium. The addition of corn oil or fatty acid to the medium resulted in a decrease in the activity level of the enzyme without impairing the viability of cells. All the fatty acids tested exhibited this effect, although linoleic acid and oleic acid were more effective than palmitic acid, stearic acid and arachidonic acid. Immunochemical titration and Ouchterlony double-diffusion analysis indicated that the decrease in the activity level of the enzyme observed in cells incubated in medium supple- mented with fatty acid can be ascribed to a reduction of the quantity of the enzyme. Isotopic leucine incorporation studies with the use of immunochemical techniques demonstrated that this reduction of the enzyme content is due to a decrease in the rate of synthesis of the enzyme. The rate of degrada- tion of the enzyme was essentially unaffected, the half-life being 25 and 28 h, respectively, in cells incubated in the presence and absence of fatty acid. It was shown that most of the isotopic fatty acid added to the medium was incorporated into cellular phospholipids, while a very small portion of it was recovered in triglyceride and nonesterified fatty acid. Acetyl-coenzyme-A carboxylase catalyzes the first step leading specifically to long-chain fatty acid synthesis and plays a critical role in the regulation of this synthetic process (see [l]). The activity level of this enzyme in mammalian tissues varies in accord with the rate of fatty acid synthesis under various dietary, hormonal, developmental and genetic condi- tions of the animal (see [2,3]). Particular emphasis has recently been placed on the questions, whether the variations in the level of acetyl-CoA carboxylase activity in tissue extracts observed under the various conditions are actually determined by changes in the quantity of the enzyme, and if so, whether these changes are due to alterations in the rate of synthesis or in that of degradation of the enzyme. Immuno- chemical studies with rats subjected to different Enzymes. Acetyl-CoA carboxylase or acetyl-CoA : carbon- dioxide ligase (ADP-forming) (EC 6.4.1.2) ; lactate dehydrogenase or L-lactate : NAD' oxidoreductase (EC 1.1.1.27); ATP-citrate lyase or ATP : citrate oxaloacetate-lyase (pro-3S-CH2COO- + acetyl-CoA; ATP-dephosphorylating) (EC 4.1.3.8); malic enzyme or L-malate : NADP' oxidoreductase (oxaloacetate-decarboxylat- ing) (EC 1.1.1.40). dietary manipulations [4- 61 and alloxan-diabetes [6] as well as with genetically obese mice [7] demon- strated that the fluctuations of the level of acetyl-CoA carboxylase activity in the liver of these animals reflect changes in the quantity of the enzyme. Further- more, it was shown with the use of combined immuno- chemical and isotopic techniques that the increase in the acetyl-CoA carboxylase content of the liver of rats fasted and subsequently refed a fat-free diet can be ascribed exclusively to a rise in the rate of synthesis of the enzyme, while the decrease in the enzyme content of the liver of fasted rats is due both to retarded synthesis and to accelerated degradation of the enzyme [5,6]. Analogous studies revealed that the subnormal hepatic enzyme content of alloxan- diabetic rats [6] as well as the elevated enzyme content of genetically obese mice [7] can be attributed prin- cipally to corresponding changes in the rate of synthe- sis of the enzyme. Thus, it appears of great importance to know which factor(s) is more directly responsible for determining the rates of synthesis and degrada- tion of acetyl-CoA carboxylase in the liver. Eur. J. Biochem. 54 (1975)
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes. Effects of Exogenous Fatty Acids on the...

Eur. J. Biochem. 54, 373-383 (1975)
Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes Effects of Exogenous Fatty Acids on the Content, Synthesis and Degradation of the Enzyme
Kaoru KITAJIMA, Shin-ichi TASHIRO, and Shosaku NUMA
Department of Medical Chemistry, Kyoto University Faculty of Medicine
(Received December 9, 1974/February 13, 1975)
Studies were made on the content, synthesis and degradation of acetyl-coenzyme-A carboxylase in JTC-25 . P3 cells, hepatocytes which can be maintained in a protein-free and lipid-free chemically defined medium. The addition of corn oil or fatty acid to the medium resulted in a decrease in the activity level of the enzyme without impairing the viability of cells. All the fatty acids tested exhibited this effect, although linoleic acid and oleic acid were more effective than palmitic acid, stearic acid and arachidonic acid. Immunochemical titration and Ouchterlony double-diffusion analysis indicated that the decrease in the activity level of the enzyme observed in cells incubated in medium supple- mented with fatty acid can be ascribed to a reduction of the quantity of the enzyme. Isotopic leucine incorporation studies with the use of immunochemical techniques demonstrated that this reduction of the enzyme content is due to a decrease in the rate of synthesis of the enzyme. The rate of degrada- tion of the enzyme was essentially unaffected, the half-life being 25 and 28 h, respectively, in cells incubated in the presence and absence of fatty acid. It was shown that most of the isotopic fatty acid added to the medium was incorporated into cellular phospholipids, while a very small portion of it was recovered in triglyceride and nonesterified fatty acid.
Acetyl-coenzyme-A carboxylase catalyzes the first step leading specifically to long-chain fatty acid synthesis and plays a critical role in the regulation of this synthetic process (see [l]). The activity level of this enzyme in mammalian tissues varies in accord with the rate of fatty acid synthesis under various dietary, hormonal, developmental and genetic condi- tions of the animal (see [2,3]). Particular emphasis has recently been placed on the questions, whether the variations in the level of acetyl-CoA carboxylase activity in tissue extracts observed under the various conditions are actually determined by changes in the quantity of the enzyme, and if so, whether these changes are due to alterations in the rate of synthesis or in that of degradation of the enzyme. Immuno- chemical studies with rats subjected to different
Enzymes. Acetyl-CoA carboxylase or acetyl-CoA : carbon- dioxide ligase (ADP-forming) (EC 6.4.1.2) ; lactate dehydrogenase or L-lactate : NAD' oxidoreductase (EC 1.1.1.27); ATP-citrate lyase or ATP : citrate oxaloacetate-lyase (pro-3S-CH2COO- + acetyl-CoA; ATP-dephosphorylating) (EC 4.1.3.8); malic enzyme or L-malate : NADP' oxidoreductase (oxaloacetate-decarboxylat- ing) (EC 1.1.1.40).
dietary manipulations [4- 61 and alloxan-diabetes [6] as well as with genetically obese mice [7] demon- strated that the fluctuations of the level of acetyl-CoA carboxylase activity in the liver of these animals reflect changes in the quantity of the enzyme. Further- more, it was shown with the use of combined immuno- chemical and isotopic techniques that the increase in the acetyl-CoA carboxylase content of the liver of rats fasted and subsequently refed a fat-free diet can be ascribed exclusively to a rise in the rate of synthesis of the enzyme, while the decrease in the enzyme content of the liver of fasted rats is due both to retarded synthesis and to accelerated degradation of the enzyme [5,6]. Analogous studies revealed that the subnormal hepatic enzyme content of alloxan- diabetic rats [6] as well as the elevated enzyme content of genetically obese mice [7] can be attributed prin- cipally to corresponding changes in the rate of synthe- sis of the enzyme. Thus, it appears of great importance to know which factor(s) is more directly responsible for determining the rates of synthesis and degrada- tion of acetyl-CoA carboxylase in the liver.
Eur. J. Biochem. 54 (1975)

314 Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes
In the present investigation, we have attempted to understand the mechanisms involved in the regula- tion of synthesis and degradation of acetyl-CoA carboxylase with the use of cultured hepatocytes, thus avoiding the complex physiological interactions encountered in the whole animal. The hepatocytes employed were JTC-25 . P3 cells, which can be maintained in a protein-free and lipid-free chemically defined medium (see [8]). The advantage of using these cells is evident, since the effect of adding lipids to the culture medium can be assessed more readily. By means of immunochemical methods, we demon- strated that the addition of fatty acid to the medium results in a decrease in the acetyl-CoA carboxylase content of the cells. Furthermore, it was shown by combined immunochemical and isotopic techniques that the decrease in the enzyme content brought about by exogenous fatty acid is due to retarded synthesis of the enzyme. Preliminary accounts pertinent to a portion of this work have been presented [9- 111.
MATERIALS AND METHODS
Reagents, Preparations and Determinations
Corn oil was obtained from Nagai Company (Tokyo, Japan). Palmitic acid, stearic acid, oleic acid and linoleic acid were purchased from Applied Science Laboratories (State College, U.S.A.), and arachidonic acid from Sigma (St. Louis, U.S.A.). Fatty-acid-free bovine serum albumin was a product of Miles Laboratories (Kankakee, U.S.A.). The cul- ture medium DM-120 was obtained from Kyokuto Pharmaceutical Industries Company (Tokyo, Japan), and millipore filters from Sartorius (Gottingen, Ger- many). Sephadex G-50 was purchased from Pharniacia (Uppsala, Sweden), and silica gel H from Merck (Darmstadt, Germany). [14C]NaHC03, [U-14C]pal- mitic acid, [ l-14C]linoleic acid and ~-[4,5-~H]leucine were products of the Radiochemical Centre (Amer- sham, England). Other chemicals were as described previously [6].
Acetyl-CoA was prepared by the method of Simon and Shemin [I21 and was determined as described by Buckel and Eggerer [13]. Phosphatidylcholine and phosphatidylethanolamine were prepared from egg yolk according to the procedure of Rhodes and Lea [14], and trigljceride from corn oil as described by Hirsch and Ahrens [15]. Protein was determined by the method of Lowry et al. [16] with bovine serum albumin as the standard. The quantitation of fatty acid bound to albumin was conducted spectrophoto- metrically according to the procedure described by Itaya and Ui [17].
Cell Culture
JTC-25 ' P3 cells were derived from liver paren- chymal cells of a JAR-1 strain rat [18] and have been maintained since 1968 in DM-120, a lipid-free and protein-free chemically defined medium (see [8]). This cell line was kindly supplied by Professor H. Kat- suta (Tokyo University Institute of Medical Science). Normally, cells were grown at 37 "C in stationary culture in TD-40-type flasks [I91 containing 10 ml DM-120. For Ouchterlony double-diffusion analyses as well as for [3H]leucine incorporation studies, large-scale culture was conducted at 37 "C in roller bottles containing 50 ml DM-120, which were rotated at a speed of one revolution per 8 min. The medium was renewed twice a week. Cells were subcultured at weekly intervals when they reached confluence. For subculture, cells were removed from the glass surface by the aid of a rubber policeman without trypsin treatment and were seeded at a dilution of 1 :3 to 1:5. The growth of cells was examined by counting the cell number, and the dye-excluding ability of cells by the erythrosin B exclusion test according to the procedures described by Katsuta et al. [20].
Preparation of Cell Extracl
Cells in a confluent state were washed at room temperature three times with 20 ml (per TD-40-type flask) or 50 ml (per roller bottle) D-solution (see [S]), a buffered balanced salt solution. The washed cells were suspended in 40ml (per TD-40-type flask) or 80ml (per roller bottle) D-solution. An aliquot (0.2- 1.0 ml) of the cell suspension was tested for cell number, and another aliquot (0.2 ml) for dye- excluding ability as described above. The following operations were carried out at 0-4 "C. The rest of the cell suspension was centrifuged at 500xg for 10 min, and the resulting cell pellet was suspended in 0.3 ml (per TD-40-type flask) or 2.5 ml (per roller bottle) 0.25 M sucrose. The cells derived from small- scale culture in TD-40-type flasks were disrupted by sonication for 8 s at minimum output with a Branson W185D sonifier, while the cells obtained by large- scale culture in roller bottles were homogenized in a Potter-Elvehjem-type Teflon-glass homogenizer with 10-20 strokes. Both methods of cell disruption resulted in essentially the same recovery of protein and acetyl-CoA carboxylase activity. Less enzyme activity was recovered when cells were sonicated for less than 5 s or for more than 10 s, or were subjected to repeated freezing and thawing. The homogenate was centrifuged at 105000 x g for 60 min, and the resulting supernatant fluid (cell extract) was collected.
Eur. J . Biochem. 54 (1975)

K. Kitajima, S. Tashiro, and S. Numa 315
Assay of Enzymes
The cell extract, after being passed through a Sephadex G-50 column, was assayed for the activity of acetyl-CoA carboxylase at 37 "C by the Hi4CO;- fixation method as reported previously [6]. Three different protein concentrations were used for the assay to ensure the proportionality between the initial reaction velocity and the amount of enzyme added. The radioactive reaction product was identified by paper chromatography as malonyl-CoA according to the procedure described by Matsuhashi et al. [21]. The activity of lactate dehydrogenase was assayed at 24 "C spectrophotometrically as described by Nissel- baum and Bodansky [22]. For this purpose, the cell extract was used without gel-filtration. One unit of an enzyme is defined as that amount which catalyzes the transformation of 1 pmol substrate per min under the assay conditions used.
Addition of Lipids
Corn oil was added to the medium as an emulsion, which was prepared by sonicating a mixture of 3.0 mg/ ml corn oil and 20 mg/ml fatty-acid-free bovine serum albumin in DM-120. Prior to sonication, corn oil and the albumin solution were sterilized by being passed through a millipore filter (0.45 pm). Fatty acids were added to the medium as their bovine serum albumin complexes, which were prepared by the method of Spector and Hoak [23]. The fatty acid/ albumin complexes were sterilized as just described.
l4C-Labelled Fatty Acid Incorporation Studies
After cells were incubated in the presence of 14C-labelled fatty acid, lipids were extracted from the homogenate of the cells according to the principle of Folch et al. [24]. The washed chloroform phase was concentrated under reduced pressure with the use of a rotary evaporator, and aliquots were subjected to thin-layer chromatography on a silica gel H plate impregnated with 10 mM Na2C03 in n-hexaneldiethy1 ether/acetic acid (50: 50: 1, by vol.) and in chloroform/ methanol/acetone/acetic aciqwater (200 : 40 : 80 : 40 : 20, by vol.) as the developing solvent. Authentic samples of linoleic acid, triglyceride, phosphatidylcholine and phosphatidylethanolamine were chromatographed simultaneously. After chromatography, the lipids were visualized by exposure to iodine vapor. The area containing each lipid species was scraped into a scintillation vial and assayed for radioactivity in 5 ml of Bray's solution [25] with a liquid scintillation spectrometer. The fatty acid constituents of the cellular lipids extracted were examined as follows.
The lipids were saponified with 2 M methanolic KOH, and the fatty acids extracted with petroleum ether (b.p. range, 30- 50 "C) were esterified with diazo- methane. The methyl esters were analyzed by radio- active gas-liquid chromatography under the condi- tions described elsewhere [26].
Purification of Rat Liver Acetyl-CoA Carboxylase
Homogeneous acetyl-CoA carboxylase was prepar- ed from rat liver according to the method of Nakanishi and Numa [6] as modified by Hashimoto and Numa [27]. The homogeneity of the final enzyme preparation, the specific activity of which was 8 - 9 U/mg of pro- tein, was ascertained by the following criteria. Firstly, the enzyme sedimented in an analytical ultracentrifuge as a single, sharp, symmetrical boundary with no evidence of impurity ; secondly, the enzyme migrated as a single band upon electrophoresis on a 1 % agar plate in 25 mM sodium diethylbarbiturate buffer pH 8.6 ; thirdly, Ouchterlony double-diffusion analysis [28] with the use of antibody prepared against the enzyme yielded a single precipitation band (see Fig. 5); finally, immunoelectrophoresis with the use of the same antibody conducted on a 1 % agar plate in 50 mM sodium diethylbarbiturate buffer pH 8.6 [29] exhibited a single precipitation arc. As carrier enzyme for [3H]leucine incorporation studies (see below), partially purified rat liver acetyl-CoA carboxylase (the second ammonium sulfate fraction [6,27]) with a specific activity of 0.3-0.35 U/mg was used after being passed through a 10-fold volume of SephadexG-50 equilibrated with 20 mM Tris-C1 buffer pH 7.5 con- taining 0.15 M NaC1, 5 mM 2-mercaptoethanol and 1 mM EDTA.
Preparation of Antibody
Rabbit antiserum was prepared against homo- geneous rat liver acetyl-CoA carboxylase, and the y-globulin fraction was isolated as described previ- ously [6].
r3H]Leucine Incorporation Studies
Large-scale culture was carried out in roller bottles as described above. ~-[4,5-~H]Leucine was added to the medium, and cells were incubated and harvested as specified for each experiment. The cell extract (2.5-3 ml) was prepared, and a 0.2-ml ali- quot of it was passed through SephadexG-50 and assayed for acetyl-CoA carboxylase activity as de- scribed above. Another aliquot (0.05-0.1 ml) of the cell extract was used for the determination of the radioactivity incorporated into soluble protein ; the
Eur. J. Biochem. 54 (1975)

376 Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes
procedure was as reported previously [6]. A third aliquot of the cell extract was subjected to DEAE- cellulose chromatography for partial purification of acetyl-CoA carboxylase. 2 ml of the cell extract was applied to a DEAE-cellulose column (0.8 x 4 cm) equilibrated previously with 50 mM Tris-C1 buffer pH 7.5 containing 5 mM 2-mercaptoethanol and 1 mM EDTA. After the column was washed with 2 ml of the same solution, the enzyme was eluted with 3 ml of the same solution containing 0.2 M NaC1. This procedure yielded more than 90% of the initial enzyme activity present in the cell extract and 2 - 3-fold purification. The partially purified preparation was employed to determine the radioactivity incorporated into the protein precipitated by anti(acety1-CoA carboxylase) ?-globulin. This determination, together with the correction for nonspecific precipitation of radioactive material, was carried out as described previously [6]. except that four of the seven aliquots (0.5 - 0.55 ml) contained 0.13 U carrier enzyme (see above) and the remaining aliquots 0.26 U carrier enzyme. The extent of nonspecific precipitation was approximately 20 "/, of the total radioactivity precip- itated by the antibody.
Sodium Dodecylsulfate - Polyacrylamide Gel Electrophoresis
The radioactive immunoprecipitate was analyzed by sodium dodecylsulfate - polyacrylamide gel electro- phoresis according to the method of Weber and Osborn [30] with minor modifications. Samples containing approximately 200 pg protein were run in cylindrical gels (0.65 x 8 cm) at 25 volts and room temperature for 6 h. After electrophoresis, the protein was visualized with a Coomassie brilliant blue solution. The gels were cut into slices of l-mm thickness, and the indi- vidual slices, which were placed into scintillation vials, were solubilized by incubation with 0.1 ml hydrogen peroxide at 60 "C for 1 - 2 h. After solubili- zation of the gels, 0.4 ml water and 5 ml of the scintil- lator solution of Patterson and Greene [31] were added to each vial, and the radioactivity was determin- ed with a liquid scintillation spectrometer.
RESULTS
For all the following experiments, JTC-25 P3 cells which had just reached a confluent state in 7 - 10 days after subculture were used. The number of cells in a confluent culture was usually 1.0- 1.2 x i07 per TD-40-type flask (small-scale culture) or 6- 7 x lo7 per roller bottle (large-scale culture). The extract from lo7 cells contained approximately 1 mg protein.
" 0 1 2 3 4 5 Experimental period (days)
Fig. 1. Periodic changes in the level of aceiyl-CoA carboxylase activity in JTC-25 P3 cells. Confluent small-scale cultures were used. At the times indicated, cells were collected to assay the enzyme activity. Results are given as mean values & S.E.M. for three cultures. (0) Cells that received fresh medium at zero time and, when indicated, also after three days; (0) cells that received fresh medium at zero time but not after three days
Fluctuation of the Level of Acetyl-CoA Carboxylase Activity Following Renewal of Culture Medium
Initially, studies were made to examine whether the level of acetyl-CoA carboxylase activity in JTC-25 P3 cells remains constant when cells in a confluent
state are incubated in DM-120. In the experiment presented in Fig. 1, the culture medium was renewed at zero time and after three days. The activity level of the enzyme fluctuated periodically with renewal of the medium. One day after the medium was renewed, the activity attained a maximal level, which was about twice as high as the initial level, and returned then gradually to the original level during the following two days. Such a fluctuation of the enzyme activity did not occur when the medium was not renewed after three days. In subsequent studies, therefore, substances to be tested were added to the medium when it was renewed, and their effects were studied for the following three days without change of the medium. The mechanism underlying the fluctuation of the enzyme activity associated with renewal of the medium remains to be elucidated.
Effect of Corn Oil on the Level of Acetyl-CoA Carboxylase Activity
In Fig. 2 are shown studies on the effect of corn oil added to the medium on the level of acetyl-CoA carboxylase activity in JTC-25 . P3 cells. The addition of corn oil resulted in a marked decrease in the enzyme activity. The activity level in cells incubated in the presence of 90 pg/ml corn oil for three days was
Eur. J. Biochem. 54 (1975)

K. Kitajima, S. Tashiro, and S. Numa 311
" 0 1 2 3 Experimental period (days)
Fig. 2. Effect of corn oil on the level of acetyl-CoA carboxylase activity in JTC-25 . P3 cells. Confluent small-scale cultures were incubated in medium containing 90 pg/ml corn oil (0) or in medium containing no corn oil but fatty-acid-free bovine serum albumin at an equivalent concentration (600 pg/ml) (0). At the times indicat- ed, cells were harvested to assay the enzyme activity. Results are given as mean values f S.E.M. for three cultures
Table 1. Effects of varying concentrations of corn oil on the level of acetyl-CoA carboxylase activity in JTC-25 P3 cells Confluent small-scale cultures were incubated with the additions indicated. The media without addition of corn oil contained, when indicated, fatty acid-free bovine serum albumin at equivalent concentrations. After incubation for three days, cells were harvested to assay the enzyme activity. Results are given as mean values f S.E.M. for three cultures
Addition
None Fatty-acid-free albumin alone, 6 pg/ml Corn oil, 0.9 pg/ml Fatty-acid-free albumin alone, 60 pg/ml Corn oil, 9 pg/ml Fatty-acid-free albumin alone, 600 pg/ml Corn oil, 90 pg/ml
Specific activity
mU/mg 2.90 f 0.21 2.97 f 0.07 1.60 f 0.06 2.90 f 0.06 0.93 rt 0.09 2.90 f 0.10 0.60 f 0.06
approximately one-fifth that in control cells. As is evident from Table 1, the enzyme activity decreased with increasing concentrations of corn oil in the medium. The enzyme activities in cells incubated for three days in media containing 0.9,9 and 90 yg/ml corn oil were 54, 32 and 21% of the control value, respectively. Corn oil was added to the medium as an emulsion in a fatty-acid-free bovine serum albumin solution (see Materials and Methods), but the addition of albumin alone exerted no influence on the activity level of the enzyme.
" 0 1 2 3 Experimental period (days)
I I I
0 1 2 3 0 '
Experimental period (days) Fig. 3. Eflects of linoleic acid ( A ) and oleic acid ( B ) on the level of acetyl-CoA carboxylase activity in JTC-25 . P3 cells. Confluent small-scale cultures were incubated in medium containing 10 pM linoleic acid or 10 pM oleic acid (0) or in medium containing no fatty acid but fatty-acid-free bovine serum albumin at an equivalent concentration (85 pg/ml) (0). At the times indicated, cells were harvested to assay the enzyme activity. Results are given as mean values f S.E.M. for three cultures
Effects of Fatty Acids on the Level of Acetyl-CoA Carboxylase Activity
Since the fatty acids that occur most frequently in corn oil are linoleic acid and oleic acid [32], we next studied the effects of these fatty acids on the level of acetyl-CoA carboxylase activity in JTC-25 - P3 cells. As shown in Fig. 3, the enzyme activity was definitely diminished by the addition of each of these fatty acids. After incubation in the presence of 10 yM linoleic acid for three days, the enzyme activity was reduced to 55 % of the activity in control cells (Fig. 3A), while 62 % of the control value was observed in cells incubat- ed in the presence of 10 yM oleic acid for three days (Fig. 3B). Table 2 shows the effects of some other fatty acids. Palmitic acid, stearic acid or arachidonic acid added at a concentration of 10 pM gave rise to a reduction of the enzyme activity to 69- 78 % of the
Eur. J. Biochem. 54 (1975)
378 Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes
Table 2. Effects of palmitic acid, stearic acid and arachidonic acid on the level of aceryl-CoA carboxylase activity in JTC-25 P3 cells Confluent small-scale cultures were incubated with the additions indicated. After incubation for three days, cells were harvested to assay the enzyme activity. Results are given as mean values f S.E.M. for three cultures
Addition Specific activity
mU/mg Fatty-acid-free albumin alone, 85 pg/ml Palmitic acid, 10 pM Stearic acid, 10 pM
3.20 & 0.12 2.50 f 0.06 2.33 f 0.14 2.20 * 0.20 Arachidonic acid, 10 pM
control value, thus being less effective than linoleic acid and oleic acid added at the same concentration. Although all these fatty acids were added to the medium as their albumin complexes (see Materials and Methods), the activity level of the enzyme was not affected by albumin itself as described above. The effect of the fatty acid addition on the enzyme activity was reversible. When cells that had been incubated in the presence of linoleic acid for three days were switched to medium containing no fatty acid, the enzyme activity was restored in three days completely to the level observed in cells that had not been exposed to fatty acid (see the legend to Fig. 6).
Viability of Cells
In order to exclude the possibility that the reduced level of acetyl-CoA carboxylase activity in JTC-25 . P3 cells observed after the addition of corn oil or fatty acid is due to cytotoxic action of these substances, we tested the growth and the dye-excluding ability of cells incubated in their presence and absence. Neither of these tests were affected appreciably by the addition of corn oil or any of the fatty acids examined. Moreover, the rate of incorporation of t3H]leucine into total soluble protein of cells was hardly influenced by exogenous fatty acids (see Table 3). Finally, the activity level of lactate dehydrogenase, an enzyme that is not directly involved in fatty acid synthesis, was not altered by the addition of linoleic acid, being 2.4 and 2.2 U/mg of protein, respectively, in the extracts derived from cells incubated in the presence and absence of the fatty acid. All these results, together with the above-mentioned reversibility of the fatty acid effect, support the view that the observed depres- sion of the level of acetyl-CoA carboxylase activity is not due to nonspecific effects of corn oil and fatty acids.
T 7 Q 1 2 3
Experimental period (days)
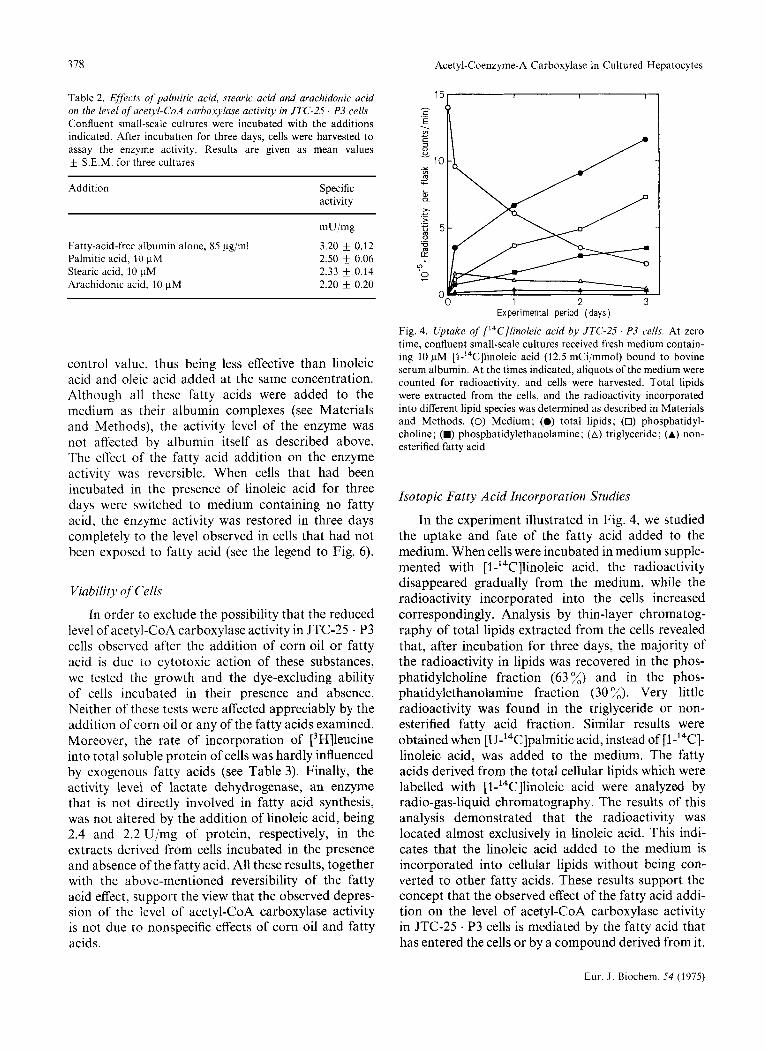
Fig. 4. Uptake of ['4C]linoleic acid by JTC-2.5 ' P3 cells. At zero time, confluent small-scale cultures received fresh medium contain- ing 10 pM [l-'4C]linoleic acid (12.5 mCipmol) bound to bovine serum albumin. At the times indicated, aliquots of the medium were counted for radioactivity, and cells were harvested. Total lipids were extracted from the cells, and the radioactivity incorporated into different lipid species was determined as described in Materials and Methods. (0) Medium; (0) total lipids; (0) phosphatidyl- choline; (W) phosphatidylethanolamine; (A) triglyceride; (A) non- esterified fatty acid
Isotopic Fatty Acid Incorporation Studies
In the experiment illustrated in Fig. 4, we studied the uptake and fate of the fatty acid added to the medium. When cells were incubated in medium supple- mented with [ l-'4C]linoleic acid, the radioactivity disappeared gradually from the medium, while the radioactivity incorporated into the cells increased correspondingly. Analysis by thin-layer chromatog- raphy of total lipids extracted from the cells revealed that, after incubation for three days, the majority of the radioactivity in lipids was recovered in the phos- phatidylcholine fraction (63 %) and in the phos- phatidylethanolamine fraction (30 x). Very little radioactivity was found in the triglyceride or non- esterified fatty acid fraction. Similar results were obtained when [U-'4C]palmitic acid, instead of [1-I4C]- linoleic acid, was added to the medium. The fatty acids derived from the total cellular lipids which were labelled with [l-'4C]linoleic acid were analyzed by radio-gas-liquid chromatography. The results of this analysis demonstrated that the radioactivity was located almost exclusively in linoleic acid. This indi- cates that the linoleic acid added to the medium is incorporated into cellular lipids without being con- verted to other fatty acids. These results support the concept that the observed effect of the fatty acid addi- tion on the level of acetyl-CoA carboxylase activity in JTC-25 . P3 cells is mediated by the fatty acid that has entered the cells or by a compound derived from it.
Eur. J . Biochem. 54 (1975)

K . Kitajima, S. Tashiro, and S. Numa 379
Ouchterlony Double-Diffusion Analysis
Immunochemical techniques were utilized in stud- ies designed to examine whether the decrease in the level of acetyl-CoA carboxylase activity observed in JTC-25 . P3 cells that were incubated in the presence of fatty acid is due to a diminished content of the enzyme or to a reduced catalytic efficiency per enzyme molecule. For this purpose, antibody against homo- geneous rat liver acetyl-CoA carboxylase was used, because it is difficult to purify the enzyme from cultur- ed cells to homogeneity. In Fig. 5 is presented a pattern of Ouchterlony double-diffusion analysis [28] obtained with this antibody and enzyme preparations which were derived from JTC-25 . P3 cells incubated in the presence and absence of linoleic acid and from rat liver. Since the enzyme concentration in the cell extracts was too low to yield a visible precipitation band, the enzyme was concentrated by ammonium sulfate fractionation (0 - 30 % saturation) as described previously [6]. The double diffusion analysis demon- strated the completeness of connections of the preci- pitation bands, showing that antibody against the rat liver enzyme crossreacts with the enzyme from JTC-25 . P3 cells, and that the enzyme molecules derived from cells incubated with and without addi- tion of the fatty acid are immunologically similar to each other. Furthermore, the appearance of a single precipitation band indicated that the rat liver enzyme preparation used as antigen was homogeneous.
Immunochemical Titration
If the extract from cells incubated in medium supplemented with fatty acid contained enzyme which was immunologically reactive and enzymatically in- active, and the extract from cells incubated in fatty- acid-free medium contained enzyme which was both immunologically reactive and enzymatically active, the amount of enzyme activity which can be neu- tralized by a fixed amount of antibody would be smal- ler when the former cell extract is titrated than when the latter cell extract is titrated. Fig. 6 represents the results of immunochemical titrations with the extracts from cells incubated in the presence of linoleic acid, cells incubated in its absence and cells incubated in its presence and subsequently switched to fatty-acid- free medium. Despite the fact that the specific activity of acetyl-CoA carboxylase varied distinctly among the three groups of cells, the equivalence point, i.e. the point at which the enzyme activity first appeared in the supernatant fluid, was the same for all the three cell extracts when based on the amount of enzyme activity added. Similar results were also obtained when the medium was supplemented with 10 pM
Fig. 5. Ouchterlony doublediffusion analysis of acetyl-CoA car- boxylase from JTC-25 . P3 cells and rat liver. Confluent large-scale cultures were incubated in the presence and absence of 10 pM linoleic acid for three days (for details of the media, see the legend to Fig. 3). Double diffusion analysis was conducted as described previously [6]. The plate was developed at 4 "C for 48 h before the photograph was taken. Wells 1 and 6, 0.18 mg control y-globulin; wells 2-5, 0.15 mg anti(acety1-CoA carboxylase) y-globulin; wells 7 and 10, 1.92 mg enzyme from JTC-25 . P3 cells incubated in the presence of linoleic acid (specific activity, 4.1 mU/mg); wells 8 and 11, 0.99 mg enzyme from JTC-25 . P3 cells incubated in the absence of linoleic acid (specific activity, 8.1 mU/mg); well 9, 60 pg partially purified rat liver enzyme (the second ammonium sulfate fraction [6,27], specific activity 300 mujmg)
/ O/ 0 5 10 15
Activi ty added (mu) Fig. 6. Immunochemical titration of' acetyl-CoA carboxylase from JTC-25 . P3 cells. Confluent large-scale cultures were incubated as follows: (0) cells incubated in the presence of 10 pM linoleic acid for three days (specific activity of acetyl-CoA carboxylase, 2.0 mU/mg); (0) cells incubated in the absence of linoleic acid for three days (specific activity, 3.8 mU/mg); (A) cells incubated in the presence of 10 pM linoleic acid for three days and subsequently in the absence of the fatty acid for three more days (specific activity, 4.2 mU/mg); for details of the media, see the legend to Fig. 3. Experimental procedures were as described previously [6 ] ; the amount of anti(acety1-CoA carboxylase) y-globulin added was 4.0 Pg
oleic acid. These results demonstrated that the ob- served changes in the activity level of the enzyme were accompanied by proportionate changes in the quantity of immunologically reactive protein. Thus, it is indicated that the decrease in the level of acetyl- CoA carboxylase activity in JTC-25 . P3 cells brought
Eur. J. Biochem. 54 (1975)
380 Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes
Table 3. Effect5 of linoleic acid and oleic acid on the rate of acetyl- CoA carboxylase synthesis in JTC-25 . P3 cells Incubation conditions were the same as described for Fig. 3, except that confluent large-scale cultures were used. After incuba- tion for three days, 200 pCi ~-[4,5-~H]leucine (55 Cijmmol) was added to the medium. 1.5h later, cells were harvested to determine the enzyme activity, the radioactivity in the enzyme and the radio- activity in total soluble protein as described in Materials and Methods. The data for [3H]leucine incorporation are expressed as radioactivity per milligram of protein in the cell extract
Expt Addition Specific [3H]Leucine incorporation activity
(4 (b) 1 o2 acetyl- total x a/b CoA soluble carboxyl- protein ase
1 Fatty-acid-free albumin. 85 pg/ml
Oleic acid. 10 pM Linoleic dcid.
10 pM
albumin, 85 pg ml
Oleic acid, 10 pM Linoleic dcid,
2 Fatty-acid-free
10 pM
mU/mg counts x min-' x mg-'
3.0 249 17100 1.46 1.6 131 16000 0.82
1.5 126 15100 0.83
3.6 260 20800 1.25 1.6 151 20100 0.75
1.8 141 19000 0.74
about by exogenous fatty acid can be ascribed to a diminished content of the enzyme.
Isotopic Leucine Incorporation Studies
The tissue content of an enzyme is generally determined by a balance between the rate of synthesis and that of degradation of the enzyme [33]. In order to investigate whether the reduction of the acetyl-CoA carboxylase content of JTC-25 . P3 cells following the supplementation of the medium with fatty acid is due to retarded synthesis or to accelerated degrada- tion of the enzyme, we undertook studies on the incorporation of isotopic leucine into the enzyme. As a measure of the rate of enzyme synthesis, the amount of [j Hlleucine incorporation into the protein precipitated by anti(acety1-CoA carboxylase) was determined after pulse labelling. The amount of radio- activity in the enzyme increased linearly with time for 2 h after the addition of [3H]leucine to the medium. In the experiments shown in Table 3, therefore, cells were incubated with [3H]leucine for 1.5 h. Since the extent of labelling of total soluble protein is reflected in that of the enzyme, the ratio of the radioactivity
I 0 20 40
Experimental period ( h )
5L
0 20 40 Experimental period (h )
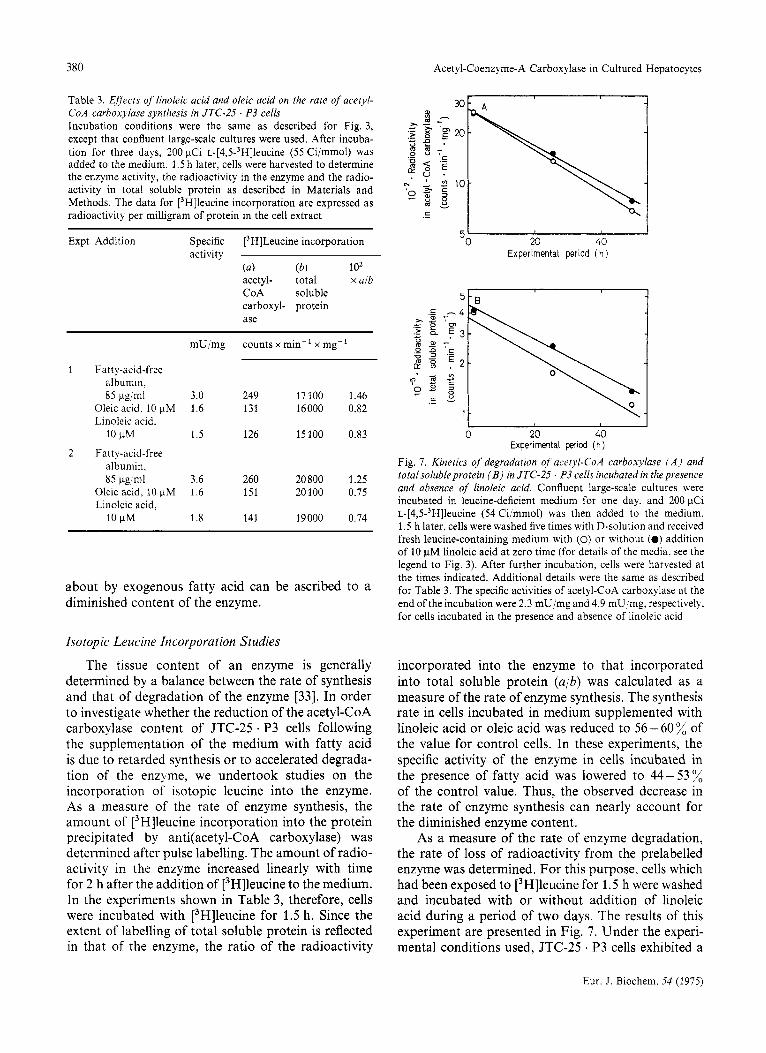
Fig. 7. Kinetics of degradation of' acetyl-CoA carboxylase ( A ) and totalsolubleprotein ( B ) in JTC-25 . P3 cells incubated in the presence and absence of linoleic acid. Confluent large-scale cultures were incubated in leucine-deficient medium for one day, and 200 pCi ~-[4,5-~H]leucine (54 Ci/mmol) was then added to the medium. 1.5 h later, cells were washed five times with D-solution and received fresh leucine-containing medium with (0) or without (0) addition of 10 pM linoleic acid at zero time (for details of the media, see the legend to Fig. 3). After further incubation, cells were harvested at the times indicated. Additional details were the same as described for Table 3. The specific activities of acetyl-CoA carboxylase at the end of the incubation were 2.3 mU/mg and 4.9 mU/mg, respectively. for cells incubated in the presence and absence of linoleic acid
incorporated into the enzyme to that incorporated into total soluble protein (a/b) was calculated as a measure of the rate of enzyme synthesis. The synthesis rate in cells incubated in medium supplemented with linoleic acid or oleic acid was reduced to 56- 60 % of the value for control cells. In these experiments, the specific activity of the enzyme in cells incubated in the presence of fatty acid was lowered to 44- 53 % of the control value. Thus, the observed decrease in the rate of enzyme synthesis can nearly account for the diminished enzyme content.
As a measure of the rate of enzyme degradation, the rate of loss of radioactivity from the prelabelled enzyme was determined. For this purpose, cells which had been exposed to [3H]leucine for 1.5 h were washed and incubated with or without addition of linoleic acid during a period of two days. The results of this experiment are presented in Fig. 7. Under the experi- mental conditions used, JTC-25 * P3 cells exhibited a
Eur. J. Biochem. 54 (1975)

K. Kitajima, S. Tashiro, and S. Numa
E D
.1 5
381
0 0 1 0 x ) 3 0 4 0 5 0 6 0 7 0
Distance from origin (mm) Fig. 8. Sodium dodecylsulfate-polyacrylamide gel electrophoresis of labelled immunoprecipitate. 200 pCi ~-[4,5-~H]leucine (55 Ci/ mmol) was added to a confluent large-scale culture. 1.5 h later, cells were harvested. The labelled immunoprecipitate was collected, and an aliquot of it was analyzed by sodium dodecylsulfate-poly- acrylamide gel electrophoresis as described in Materials and Meth- ods. E and D indicate the positions to which subunits of homo- geneous rat liver acetyl-CoA carboxylase and the tracking dye (bromphenol blue) migrated, respectively
fluctuation of the level of acetyl-CoA carboxylase activity following renewal of the medium (see Fig. 1). The rate of degradation of an enzyme under conditions of changing contents is given by the change in total radioactivity in the enzyme, not by the change in radioactivity per unit amount of the enzyme [34]. As seen in Fig. 7A, the decay of the radioactivity in acetyl-CoA carboxylase per milligram of soluble cellular protein followed first-order kinetics, the half- life being 25 h and 28 h, respectively, in cells incubated in the presence and absence of linoleic acid. Thus, there was essentially no difference in the rate of enzyme degradation between the two groups of cells. Fig. 7B shows that the half-life of total soluble protein was also practically the same in the two groups of cells, being 26 h in the presence of linoleic acid and 29 h in its absence.
Analysis of Immunoprecipitate by Sodium Dodecylsulfate - Polyacrylamide Gel Electrophoresis
In order to confirm that the protein precipitated by the antibody used actually represented acetyl-CoA carboxylase, we analyzed the labelled immunoprecip- itate by sodium dodecylsulfate - polyacrylamide gel electrophoresis. As illustrated in Fig. 8, allmost all of the radioactivity in the immunoprecipitate migrated as a single band to the same position as subunits of rat liver acetyl-CoA carboxylase. These data indicate that the antibody used specifically precipitated acetyl- CoA carboxylase.
DISCUSSION
The results of the present investigation demon- strated that the level of acetyl-CoA carboxylase activity in JTC-25 . P3 cells is reduced by the addition of corn oil or fatty acid to the medium. This effect was shown not to be attributable to impaired viability of cells. Among the fatty acids tested, linoleic acid and oleic acid were the most effective. Immunochemical analyses indicated that the reduction of the enzyme activity observed after the addition of these fatty acids is due to a decrease in the cellular content of the enzyme. Palmitic acid, stearic acid and arachidonic acid ex- hibited similar effects on the enzyme level, although they were less effective than linoleic acid and oleic acid. On the other hand, corn oil added at a concentra- tion of 9 pg/ml or 90 pg/ml lowered the enzyme level more markedly than linoleic acid and oleic acid, which were added at a concentration of 10 pM. The difference in effectiveness between corn oil and the fatty acids may be due to the different concentrations of these substances used. On the assumption that corn oil is composed of oleodilinolein 1321, the concentrations 9 pg/ml and 90 pg/ml would correspond to 10.2 pM and 102 pM, respectively. Hence, the corn oil added would provide 3 or 30-fold more fatty acids than when linoleic acid or oleic acid itself was added. Attempts to examine the effect of adding fatty acids at a concen- tration higher than 10 pM were unsuccessful, because the viability of cells was then impaired.
Studies of Bortz et al. [35] indicated that the administration of corn oil to rats previously fed a fat-free diet results in a depression of the level of acetyl- CoA carboxylase activity in the liver. Likewise, Gibson’s group 136,371 observed a progressive diminu- tion of the activity levels of hepatic lipogenic enzymes including acetyl-CoA carboxylase when animals main- tained on a fat-free diet received corn oil or methyl esters of polyunsaturated fatty acids, such as linoleic acid, linolenic acid and arachidonic acid. Methyl esters of saturated and monounsaturated fatty acids were hardly effective. However, subsequent studies in vivo yielded controversial results with respect to the question, whether or not the depression of the activity levels of hepatic lipogenic enzymes is due to a specific action of polyunsaturated fatty acids [38 - 401.
Some recent studies have dealt with the effects of fatty acids or serum on the levels of lipogenic enzymes in cultured cells. Work of Jacobs et al. [41] with cultur- ed human skin fibroblasts demonstrated that cells grown in the presence of fetal calf serum contain less acetyl-CoA carboxylase than cells grown with lipid- deficient serum. The effect of the lipid-deficient medium was reversed by the addition of normal serum or any of the fatty acids tested including saturat-
Eur. J. Biochem. 54 (1975)

382 Acetyl-Coenzyme-A Carboxylase in Cultured Hepatocytes
ed, monounsaturated and polyunsaturated fatty acids. An analogous effect of serum on the levels of acetyl- CoA carboxylase and fatty acid synthetase was shown by Alberts et al. [42] with several other cell lines in culture. Furthermore, Goodridge et al. [43] reported that the increase in the activity levels of fatty acid synthetase, ATP-citrate lyase and malic enzyme ob- served in hepatocytes from chick embryos during culture is blocked by the addition of stearic acid to the medium. In studies with Saccharomyces cere- visiae, another eukaryotic organism, Kamiryo and Numa [44] demonstrated that various fatty acids including saturated, monounsaturated and polyun- saturated fatty acids depress the cellular content of acetyl-CoA carboxylase. Thus, these findings in vitro, together with our present results, support the view that the reduction of the cellular content of lipogenic enzymes can not be ascribed to a specific effect of polyunsaturated fatty acids.
The decrease in the cellular content of acetyl-CoA carboxylase could be due either to retarded synthesis or to accelerated degradation of the enzyme or to both. In previous studies with cultured cells, no at- tempts were made to measure the rates of synthesis and degradation of the enzyme. The present experi- ments demonstrated that the reduction of the acetyl- CoA carboxylase content of JTC-25 . P3 cells brought about by exogenous fatty acid is due to retarded synthesis of the enzyme, while the rate of degradation of the enzyme is essentially unaffected. The half-life for degradation of acetyl-CoA carboxylase in these cells was shown to be 25 - 28 h. This value is consider- ably shorter than the half-life for breakdown of the enzyme in rat and mouse liver found in vivo under normal conditions, i.e. 50-67 h [5-71. This may reflect a difference in the extent of reutilization of [3H]leucine, which would lead to an overestimation of the half-life [34].
It is to be noted that most of the fatty acid added to the medium is incorporated into cellular phospho- lipids, while a very small portion of it is recovered in triglyceride and nonesterified fatty acid. This finding supports the concept that the fatty acid that has entered the cells or a compound metabolically related to it is involved in the regulation of the rate of acetyl- CoA carboxylase synthesis.
JTC-25 . P3 cells, which were used in the present investigation, can be cultured in the protein-free and lipid-free chemically defined medium DM-120. Other cultured cells usually require serum or some lipid components for their growth. Worthy of note in this context is the finding of Kagawa et al. [45] that L . P3 cells, which originated from mouse fibroblasts and have likewise been maintained in DM-120, exhibit normal cellular activities with respect to growth and
mitochondria1 function, despite the fact that essential fatty acids are replaced by monounsaturated fatty acids in the lipids of these cells.
We are indebted to Professor H. Katsuta and Dr T. Takaoka for their kind help and advice in conducting cell culture. This in- vestigation was supported in part by Research Grants from the Ministry of Education of Japan, the Mitsubishi Foundation, the Japanese Foundation of Metabolism and Diseases, the Iyakushigen Foundation and the Japanese Medical Association. K. Kitajima was a recipient of the Iue Foundation Fellowship.
REFERENCES 1. Numa, S., Nakanishi, S., Hashimoto, T., Iritani, N. & Okazaki,
2. Numa, S. (1974) Ergeb. Physiol. Biol. Chem. Exp. Pharmakol.
3. Numa, S . & Yamashita, S. (1974) Curr. Top. Cell. Regul. 8,
4. Numa, S., Nakanishi, S. & Iritani, N. (1969) Proc. Jap. Conf
5. Majerus, P. W. & Kilburn, E. (1969) J . Bid . Chem. 244,6254-
6. Nakanishi, S. & Numa, S. (1970) Eur. J. Biochem. 16,161 - 173. 7. Nakanishi, S. & Numa, S. (1971) Proc. Nail Acad. Sci. U.S.A.
8. Katsuta, H. & Takaoka, T. (1973) Methods Cell Bid . 6 , 1-42. 9. Kitajima, K. & Numa, S. (1973) Seikaguku, 45, 532.
T. (1970) Vitam. Horm. 28, 213-243.
69,53 - 96.
197-246.
Biochem. Lipids, 11,235 - 240.
6262.
68,2288 - 2292.
10. Numa, S. (1975) Naturwissenschaften, 62, 80- 86. 11. Numa, S., Kitajima, K., Teraoka, H. & Nakanishi, S. (1975)
in lntracellular Protein Turnover (Schimke, R. T. & Katsu- numa, N., eds) Academic Press, New York, in press.
12. Simon, E. J. & Shemin, D. (1953) J . Am. Chem. Soc. 75, 2520. 13. Buckel, W. & Eggerer, H. (1965) Biochem. 2. 343, 29-43. 14. Rhodes, D. N. &Lea, C. H. (1957) Biochemistry, 65,526-533. 15. Hirsch, J. & Ahrens, E. H. (1958) J. Bid . Chem. 233,311 -320. 16. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J.
17. Itaya, K. & Ui, M. (1965) J. Lipid Res. 6, 16-20. 18. Takaoka, T. & Katsuta, H. (1973) Jap. J . Exp. Med. 43,403-
411. 19. Katsuta, H., Takaoka, T., Hori, M., Okumura, H., Yasukawa,
M., Saito, S. & Suzuki, S. (1957) Jap. J . Exp. Med. 27,
20. Katsuta, H., Takaoka, T., Oishi, Y., Baba, T. & Chang, K. C.
21. Matsuhashi, M., Matsuhashi, S. & Lynen, F. (1964) Biochem.
22. Nisselbaum, J. S. & Bodansky, 0. (1959) J . Biol. Chem. 234,
23. Spector, A. A. & Hoak, J. C . (1969) Anal. Biochem. 32, 297-
24. Folch, J., Lees, M. & Sloane-Stanley, G. H. (1957) J . Bid .
25. Bray, G. A. (1960) Anal. Biochem. I, 279-285. 26. Yamashita, S., Nakaya. N., Miki, Y. & Numa, S. (1975) Proc.
Nut1 Acad. Sci. U.S.A. 72, 600 - 603. 27. Hashimoto, T. & Numa, S. (1971) Eur. J . Biuchem. 18, 319-
331. 28. Ouchterlony. 0 . (1949) Actu Path. Microhiol. 26, 507-515. 29. Scheidegger, J. J . (1955) lnt. Arch. Allergy Appl. Immunol. 7,
30. Weber, K. & Osborn, M. (1969) J . Bid . Chem. 244,4406-4412. 31. Patterson, M. S. & Greene, R. C . (1965) Anal. Chern. 37, 854.
(1951) J. Bid . Chem. 193, 265-275.
443 - 458.
(1954) Jap. J . Exp. Med. 24, 125-139.
2,340,263 - 289.
3276- 3280.
302.
Chem. 226,497 - 509.
103-110.
Eur. J . Biochem. 54 (1975)

K. Kitajima, S. Tashiro, and S. Numa 383
32. Eckey, E. W. (1954) Vegetable Fats and Oils, pp. 283-287,
33. Schimke, R. T. & Doyle, D. (1970) Annu. Rev. Biochem. 39,
34. Koch, B. L. (1962) J. Theor. Biol. 3, 283-303. 35. Bortz, W., Abraham, S. & Chaikoff, I. L. (1963) J. Biol. Chem.
36. Allmann, D. W. & Gibson, D. M. (1965) J. Lipid Res. 6,
37. Muto, Y. & Gibson, D. M. (1970) Biochem. Biophys. Res.
38. Donaldson, W. E. & Liou, G. I. (1972) Fed. Proc. 31, 698.
Reinhold Publishing Corporation, New York.
929 - 976.
238, 1266- 1272.
51-62.
Commun. 24,750-757.
39. Gozukara, E. M., Frolich, M. & Holten, 0. (1972) Biochim.
40. Musch, K., Ojakian, M. A. &Williams, M. A. (1974) Biochim.
41. Jacobs, R. A,, Sly, W. S. & Majerus, P. W. (1973) J. Biol.
42. Alberts, A. W., Ferguson, K., Hennessy, S. & Vagelos, P. R.
43. Goodridge, A. G., Garay, A. & Silpananta, P. (1974) J. Biol.
44. Kamiryo, T. & Numa, S. (1973) FEBS Lett. 38, 29-32. 45. Kagawa, Y., Takaoka, T. & Katsuta, H. (1969) J. Biochem.
Biophys. Acta, 286, 155- 163.
Biophys. Acta, 337, 343 - 348.
Chem. 248,1268- 1276.
(1974) J. Biol. Chem. 249, 5241 - 5249.
Chem. 249,1469- 1475.
(Tokyo) 65,799-808.
K. Kitajima, S. Tashiro, and S. Numa, Department of Medical Chemistry, Kyoto University Faculty of Medicine, Yoshida, Sakyo-ku, Kyoto, Japan 606
Eur. J. Biochem. 54 (1975)




















