
Spatial attention and conscious perception: the role of endogenous and exogenous orienting
Upload
independentCategory
view
1download
0
CANCER STEM CELLS
Proliferation of Human Glioblastoma Stem Cells Occurs
Independently of Exogenous Mitogens
JOHN J. P. KELLY,a,b
OWEN STECHISHIN,a,b
ANDREW CHOJNACKI,a,b
XUEQING LUN,cBEICHEN SUN,
c
DONNA L. SENGER,c,d PETER FORSYTH,a,c,d,e ROLAND N. AUER,a,e,f JEFF F. DUNN,a,g
J. GREGORY CAIRNCROSS,a,d,e IAN F. PARNEY,a,d,e SAMUEL WEISSa,b
aHotchkiss Brain Institute, bDepartment of Cell Biology and Anatomy, cClark Smith Integrative Brain Tumor
Research Center and Southern Alberta Cancer Research Institute, dDepartment of Oncology, eDepartment of
Clinical, Neurosciences, fDepartment of Pathology and Laboratory Medicine, and gDepartment of Diagnostic
Imaging, University of Calgary, Calgary, Alberta, Canada
Key Words. Neural stem cell • Brain tumor stem cell • Glioma • Glioblastoma • Mitogen
ABSTRACT
Primary glial tumors of the central nervous system, mostcommonly glioblastoma multiforme (GBM), are aggressivelesions with a dismal prognosis. Despite identification and
isolation of human brain tumor stem cells (BTSCs), charac-teristics that distinguish BTSCs from neural stem cells
remain to be elucidated. We cultured cells isolated fromgliomas, using the neurosphere culture system, to under-stand their growth requirements. Both CD1331 and
CD1332
adult GBM BTSCs proliferated in the absence ofexogenous mitogenic stimulation and gave rise to multipo-
tent GBM spheres that were capable of self-renewal. Epi-dermal growth factor (EGF) and fibroblast growth factor-2enhanced GBM BTSC survival, proliferation, and subse-
quent sphere size. Blockade of EGF receptor (EGFR) signal-ing reduced exogenous mitogen-independent GBM spheregrowth. Implantation of as few as 10 exogenous mitogen-in-
dependent GBM BTSCs led to the formation of highly inva-sive intracranial tumors, which closely resembled human
GBMs, in immunocompromised mice. These results demon-strate that exogenous mitogen independence, mediated inpart through EGFR signaling, is one characteristic that dis-
tinguishes CD1331
and CD1332
GBM BTSCs from neuralstem cells. This novel experimental system will permit the
elucidation of additional constitutively activated mechanismsthat promote GBM BTSC survival, self-renewal, and prolif-eration. STEM CELLS 2009;27:1722–1733
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Glioblastoma multiforme (GBM), the most common primarybrain tumor, is highly aggressive with a median survival of 1year. Historically, the astrocyte was believed to undergo trans-formation, giving rise to GBM [1]. However, identification ofneural stem cells (NSCs) [2] and immature progenitors [3, 4]within the adult mammalian central nervous system (CNS)has brought this dogma into question.
The biology of NSCs can be studied using the neuro-sphere culture system in which one NSC gives rise to oneneurosphere in vitro [5]. Human NSCs require the addition ofexogenous epidermal growth factor (EGF) and fibroblastgrowth factor-2 (FGF2) to culture medium to proliferate and
form neurospheres [6–9]. An approach using the neurosphereculture system also contributed to the discovery of brain tu-mor stem cells (BTSCs) [10]. BTSCs were initially identifiedas CD133þ-, EGF-, and FGF2-responsive cells that form mul-tipotent brain tumor spheres in cell culture [10]. Recently,CD133� GBM BTSCs have also been identified [11]. Impor-tantly, all BTSCs initiate tumors in immunocompromisedmice that phenocopy their original tumor [12].
One hallmark of cancer cells in general is their ability todivide in the absence of exogenous mitogenic stimulation,leading to unregulated proliferation [13]. This characteristichas yet to be ascribed to BTSCs. We hypothesized thatBTSCs, like other cancer cells, would grow in serum-free cul-ture medium (SFM) without exogenous mitogenic stimulation.Thus, we set out to test whether BTSCs could proliferate in
Author contributions: J.J.P.K.: Conception and design, collection and/or assembly of data, data analysis and interpretation, manuscriptwriting; O.S.: Conception and design, collection and/or assembly of data, data analysis and interpretation; A.C.: Conception and design,data analysis and interpretation, manuscript writing; X.L., J.F.D.: Collection and/or assembly of data, data analysis and interpretation;B.S.: Collection and/or assembly of data; D.L.S., R.N.A.: Conception and design, data analysis and interpretation; P.F.: Conception anddesign, financial support, data analysis and interpretation; J.G.C.: Conception and design, financial support, administrative support, dataanalysis and interpretation; I.F.P.: Conception and design, provision of study material or patients, financial support, data analysis andinterpretation; S.W.: Conception and design, financial support, administrative support, data analysis and interpretation, manuscriptwriting, final approval of manuscript.
Correspondence: Samuel Weiss, Ph.D., Hotchkiss Brain Institute, Faculty of Medicine, University of Calgary, Room 2263, 3330Hospital Drive N.W., Calgary, Alberta, Canada T2N 4N1. Telephone: 403-220-3994; Fax: 403-210-9382; e-mail: [email protected] Received December 16, 2008; accepted for publication April 14, 2009; first published online in STEM CELLS EXPRESS April 23,2009. VC AlphaMed Press 1066-5099/2009/$30.00/0 doi: 10.1002/stem.98
STEM CELLS 2009;27:1722–1733 www.StemCells.com

SFM in the absence of exogenous EGF þ FGF2, while main-taining their capacity for self-renewal and tumor initiation invivo.
We demonstrate that both CD133þ and CD133� GBMBTSCs, cultured in SFM without exogenous EGF þ FGF2,proliferate and generate multipotent GBM spheres. These kar-yotypically abnormal BTSCs self-renew, differentiate aber-rantly along multiple lineage pathways, and initiate highlyinvasive intracranial tumors in immunocompromised mice.Addition of EGF þ FGF2 to GBM BTSC cultures enhancedcell survival and proliferation and altered their phenotypicpotential in vitro but not in vivo. Blockade of EGF receptor(EGFR) signaling reduced exogenous mitogen-independentGBM sphere growth. These results identify a fundamental dif-ference between the growth requirements of GBM BTSCs andnormal NSCs [6–9] and have led to the development of anovel experimental system for the investigation of GBM.
MATERIALS AND METHODS
GBM BTSC Sphere and Human Temporal LobeCell Culture
Fresh tumor samples and normal human temporal lobe sampleswere obtained from adult patients (age >16) during their opera-tive procedure, after informed consent was obtained. Tumorswere graded at the Foothills Medical Centre, University of Cal-gary/Calgary Health Region, by a neuropathologist according toWorld Health Organization guidelines [1]. For cell culture, tumortissue or temporal lobe tissue was washed in sterile 1� phos-phate-buffered saline (PBS) containing penicillin and streptomy-cin. Specimens were finely minced and placed in SFM containingDulbecco’s modified Eagle’s medium/Ham’s F12 (1:1) with5 mM HEPES buffer, 0.6% glucose, 3 mM sodium bicarbonate,2 mM glutamine, 25 lg/ml insulin, 100 lg/ml transferrin, 20 nMprogesterone, 10 lM putrescine, and 30 nM selenite. A single-cell suspension was obtained by performing a series of mechani-cal dissociations, followed by filtration through a 40-lm filter.Red blood cells were removed by hypotonic lysis. Finally, cellswere resuspended in SFM, counted using trypan blue to excludedead cells, and plated at a density of 20,000 viable cells per ml.For brain tumor and temporal lobe cell culture two conditionswere used, SFM alone and SFM supplemented with EGF (20 ng/ml; Peprotech, Rocky Hill, NJ, http://www.peprotech.com),FGF2 (20 ng/ml; R&D Systems Inc., Minneapolis, http://www.rndsystems.com), and heparin sulfate (2 lg/ml; R&D Systems).Brain tumor and temporal lobe cell cultures were fed weekly byremoving half of the existing medium and replacing an equal vol-ume of SFM or SFM with EGF þ FGF2. Brain tumor spheres(none from temporal lobe appeared) were evident as early as 1week after plating. Spheres were grown for at least 3 weeks oruntil they reached a size adequate (�100-200 lm) for plating,differentiation, and passaging.
GBM BTSC Sphere Differentiation
Tumor spheres were plated on poly(L-ornithine)-coated glass cov-erslips, were allowed to differentiate for 7 days in SFM with 1%fetal bovine serum (Invitrogen, Carlsbad, CA, http://www.invitro-gen.com), and then were fixed using 4% paraformaldehyde.
Immunocytochemical Staining of GBMSpheres and Cells
Triple immunostaining of differentiated GBM spheres was per-formed by incubating coverslips for 2 hours at 37�C in 1� PBScontaining 10% normal goat serum (NGS) and mouse IgM anti-O4 (1:10; Chemicon, Temecula, CA, http://www.chemicon.com).Coverslips were washed with 1� PBS after and between each
subsequent step. Next, a biotin-goat anti-mouse IgM (1:200; Jack-son Immunoresearch Laboratories, West Grove, PA, http://www.jacksonimmuno.com) was applied for 1 hour at 37�C fol-lowed by streptavidin-cyanine 3 (SA-Cy3) (1:2,000). Coverslipswere then incubated for 2 hours at 37�C in 0.3% Triton X-100 inPBS (PBS-T) containing 10% NGS, rabbit anti-glial fibrillaryacidic protein (GFAP) (1:400; BTI Inc., Stoughton, MA, http://www.btiinc.com), mouse anti-microtubule-associated protein-2 (MAP2) (1:1,000; Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com), and/or mouse-anti-b-III-tubulin (1:1,000;Sigma-Aldrich). Finally, goat anti-mouse IgG fluorescein isothio-cyanate (FITC) (1:100; Jackson Immunoresearch), goat anti-rabbitrhodamine (1:200; Jackson Immunoresearch), and Hoechst 33258(1:100; Sigma-Aldrich) were applied together for 1 hour at 37�C.Immunolabeling of GBM spheres for nestin was performed byincubating coverslips for 2 hours at 37�C in PBS-T containing10% NGS and mouse IgG anti-nestin (1:100; Chemicon). Cover-slips were then incubated for 1 hour at 37�C with biotin-goatanti-mouse IgG (1:200; Jackson Immunoresearch), followed bySA-Cy3 (1:2,000) with Hoechst 33258. Immunolabeling forEGFR was performed using a sheep anti-EGFR antibody (1:100;Upstate, Charlottesville, VA, http://www.upstate.com) followedby a biotin donkey anti-sheep antibody (1:200; Jackson Immunor-esearch) and then streptavidin-FITC (1:1,000) together withHoechst 33258.
GBM Sphere Self-Renewal Assay
Self-renewal capacity was examined by mechanically dissociatingprimary tumor spheres into a single-cell suspension using aP1000 followed by a P200 Pipetman tip (Gilson, Inc., Middleton,WI, http://www.pipetman.com). Cell suspensions were washedtwice in SFM, inspected, and, if necessary, filtered through a 40-lm mesh filter to obtain a single-cell suspension. Cell countswere performed, and the cells were replated at a density �20,000cells per ml in either 24-well plates (uncoated; Nunc, Rochester,NY, http://www.nuncbrand.com) or 25-cm2 flasks (uncoated;Nunc) in their original growth condition. Cultures were fed andobserved weekly for secondary sphere formation. Secondaryspheres were handled in the same manner as primary GBMspheres for differentiation and passaging.
GBM Sphere Growth Assays
Dissociated GBM spheres were seeded at 500 cells per well in96-well uncoated plates (Nunc) for both antibody and inhibitorblockade experiments. Cultures were performed with four inde-pendent GBM BTSC lines, in triplicate for each line with at leastthree wells per condition for each replicate. For antibody block-ade experiments, either an EGFR function blocking monoclo-nal antibody (4 lg/ml; Calbiochem, San Diego, http://www.emdbiosciences.com) or a mouse IgG2a isotype control (R&DSystems), were added to each well. For EGFR kinase inhibitionexperiments, the following small molecule tyrphostin receptor ty-rosine kinases inhibitors (Calbiochem) were used: AG9 (5 lM),AG1295 (5 lM), AG1478 (5 lM), and PD168393 (650 nM). Di-methyl sulfoxide was used in an identical volume as the control.
To evaluate protein expression of EGFR and phospho-EGFRwe cultured exogenous mitogen-independent GBM BTSCs inSFM under the antibody and small molecule inhibitor conditionsdescribed above for 1 hour. Cellular protein was then harvestedby lysing cells in RIPA buffer with complete mini protease inhib-itor cocktail (Roche Diagnostics, Basel, Switzerland, http://www.roche-applied-science.com) and Halt phosphatase cocktail (ThermoFisher Scientific, Waltham, MA, http://www.thermofisher.com)added; protein content was measured with the Bio-Rad ProteinAssay (Bio-Rad, Hercules, CA, http://www.bio-rad.com). Then 15lg of protein was loaded per lane on 6% acrylamide gel andtransblotted to nitrocellulose membranes (Bio-Rad). Blots wereblocked with 5% nonfat milk (Bio-Rad) in Tris-bufferedsaline-Tween 20 (TBS-T). Primary phospho (p)-EGFR and EGFRantibodies (Cell Signaling Technology, Beverly, MA, http://
Kelly, Stechishin, Chojnacki et al. 1723
www.StemCells.com

www.cellsignal.com) were used at 1:1,500 in 3% milk-TBS-T.Secondary donkey anti-mouse IgG horseradish peroxidase (HRP)(for p-EGFR) and goat anti-rabbit IgG HRP (Chemicon) were usedat 1:7,500 in 1% milk-TBS-T. Bands were visualized with an ECLPlus Western Blotting Detection System and Hyperfilm (Amer-sham Biosciences, Piscataway, NJ, http://www.gelifesciences.com).Blots were first probed for p-EGFR and then stripped with RestoreWestern Blot Stripping Buffer (Thermo Fisher Scientific),reblocked in 5% milk, and reprobed for EGFR.
Karyotyping
Cells were treated with medium containing Colcemid (10 mg/ml)for 2 hours and were resuspended in a hypotonic solution con-taining 1% sodium citrate for 30 minutes. Cells were then washedin a fixative solution containing methanol and acetic acid in a 3:1ratio. Cells were spread onto slides and G-banding staining wasperformed. Metaphases were analyzed and photographed.
Chromagen In Situ Hybridization andp53 Sequencing
EGFR Chromagen in situ hybridization (CISH) was performed onboth formalin-fixed paraffin-embedded tissue samples and corre-sponding ethanol-fixed fresh cellular samples using the SPoT-Light EGFR CISH probe, according to standard protocols (http://www.invitrogen.com). For p53 sequencing RNA was extractedfrom fresh GBM tissue and sphere cultures using RNeasy (Qia-gen, Hilden, Germany, http://www1.qiagen.com) according to themanufacturer’s instructions, and 500 ng of RNA was reverse tran-scribed with the SuperScript III First-Strand Synthesis System(Invitrogen) using poly(T) primers. Then 2 ml of cDNA was usedin a 50-ll reverse transcriptase-polymerase chain reaction (RT-PCR) reaction (Invitrogen) to amplify a 1,267-base pair fragmentcontaining the entire p53 open reading frame as described previ-ously [14]. The RT-PCR product was purified by agarose gelelectrophoresis and isolated with the QIAquick Gel ExtractionKit (Qiagen). Automated DNA sequencing was performed at theUniversity of Calgary Core DNA Services facility.
GBM Sphere Cell Implantation intoImmunocompromised Mice
Low passage number GBM spheres (implantation series 1:BT012, BT025, and BT030 between passages 5 and 7; implanta-tion series 2: BT012 at passage >10) were mechanically dissoci-ated to single-cell suspensions. U87 glioma (U87MG) cells weretrypsinized to obtain a single-cell suspension and used as controlsfor implantation experiments. All cells were washed twice inSFM, and a cell count was performed using trypan blue exclusionto identify viable cells. Either 10, 100, 1,000, 10,000 or 100,000viable cells were resuspended in 3 ll of SFM for stereotactic im-plantation into the right striatum of 6- to 8-week-old CB-17NOD-SCID mice (The Jackson Laboratory, Bar Harbor, ME,http://www.jax.org). Coordinates for stereotactic implantationwere as follows: anteroposterior �1.0, mediolateral 2.0, and dor-soventral 3.0. Mice were allowed to survive up to 12 weeks andthen were killed. Brains were removed and observed for grossmorphological evidence of tumor formation. Each brain was cutin the coronal plane to obtain two segments, each containing partof the tumor. One half was immediately cryopreserved, and theother fixed in formalin before immunohistochemical analysis.
Immunohistochemistry
For histological examination of tumors that arose in SCID miceafter GBM BTSC implantation, H&E and toluidine blue stainingwas performed according to standard protocols. An indirect im-munoperoxidase method was used for immunohistochemical anal-ysis of frozen and paraffin-embedded sections. Immunostainingagainst human nuclear antigen was performed on formalin-fixedfrozen sections. Sections were incubated with a mouse-anti-human nuclear antigen antibody (1:50; Chemicon) for 16 h at
4�C followed by a biotinylated horse anti-mouse antibody (VectorLaboratories, Burlingame, CA, http://www.vectorlabs.com). Avi-din-biotin-peroxidase complexes were formed using an ABC kit(Vector Laboratories). Diaminobenzidine (DAB) was converted toa brown reaction product by peroxidase; toluidine blue was usedas a nuclear counterstain. The following primary antibodies wereused for immunohistochemical analysis of formalin-fixed paraf-fin-embedded sections: mouse anti-GFAP (1:300; R&D Systems),mouse-anti-nestin (1:200; Chemicon), mouse-anti-MAP2(1:1,500; BD Pharmingen, San Diego, http://www.bdbiosciences.com/index_us.shtml), and rabbit anti-myelin basic protein (MBP)(1:2,000; DAKO, Glostrup, Denmark, http://www.dako.com).Sections were incubated with each specific primary antibodylisted above for 16 h at 4�C followed by a biotinylated horseanti-mouse/goat anti-rabbit antibody (Vector Laboratories). Avi-din-biotin-peroxidase complexes were formed using an ABC kit.Peroxidase converted DAB to a brown reaction product, andhematoxylin was used as a blue nuclear counterstain.
Cell Cycle Analysis by Flow Cytometry
Dissociated GBM sphere cells were cultured at 20,000 cells perml in SFM for 7 days and then were changed to either SFM orSFM with EGF þ FGF2 for an additional 7 days (n ¼ 3 differentBTSC cultures and 3 individual samples per growth condition).The resulting GBM spheres were then dissociated to single-cellsuspensions, washed with PBS, resuspended in 500 ll of PBS,and fixed by adding 500 ll of absolute ethanol. Cells were incu-bated at 4�C overnight before staining. Cells were then pelleted,washed, and resuspended in propidium iodide (PI) solution con-taining 200 lg/ml PI and 0.4 mg/ml RNase. Cells were incubatedfor 1 hour at 4�C and scanned using a BD FACSCalibur flow cy-tometer and analyzed using ModFit LT software (Becton Dickin-son and Company, Franklin Lakes, NJ, http://www.bd.com).
Flow Cytometry and Florescence Activated CellSorting for CD133
GBM cells were washed and resuspended in PBS þ 0.5% bovineserum albumin (PBS-BSA) then incubated with either a monoclo-nal CD133/1-phycoerytherin (CD133-PE)-conjugated antibody ora phycoerythrin-conjugated mouse IgG1 isotype control antibody(both 1:10, according to the manufacturer’s instructions; MiltenyiBiotec, Bergisch Gladbach, Germany, http://www.miltenyibiotec.com) for 60 minutes at 4�C. Cells were then washed with 2 ml ofPBS-BSA, centrifuged at 800 rpm for 5 minutes, resuspended in0.5 ml of PBS-BSA, and analyzed by flow cytometry. Viablecells were identified using 7-amino actinomycin D. Cell sorting(BD FACSVantage SE; Becton Dickinson and Company) wasperformed using a staining method identical to that for CD133-PE described above. Sorted cells were analyzed for purity byflow cytometry with a FACSCalibur flow cytometer (BectonDickinson and Company) and FlowJo software (Tree Star, Inc.,Ashland, OR, http://www.treestar.com).
Limiting Dilution Assay
GBM single-cell suspensions, obtained after dissociation of newlyobtained GBM tissue or after dissociation of sphere cultures ofestablished GBM BTSC lines, underwent fluorescence-activatedcell sorting (FACS) purification into CD133þ- and CD133�-enriched fractions. Purity of the positive and negative fractionswas confirmed by flow cytometry. Both cell fractions were thenplated in uncoated 96-well plates at three dilutions that included1,000 (n ¼ 4-5 wells), 100 (n ¼ 8-10 wells), and 10 (n ¼ 20wells) cells per well in 100 ll of SFM. Plates were fed weeklyuntil spheres were an adequate size (>100 lm diameter) forcounting.
Animal Magnetic Resonance Imaging
Multiecho T2 (repetition time ¼ 5 s, 16 echos, 10 millisecondseach) sequences were performed at the University of Calgary Ex-perimental Imaging Centre using a 9.4-T Bruker horizontal-bore
1724 Exogenous Mitogen-Independent BTSCs

magnetic resonance (MR) system. Images were reviewed usingImageJ software.
Statistics
t tests were performed using Microsoft Excel data analysis for com-parison of two conditions. Results are displayed as the mean �SEM. One-way analysis of variance was performed using GraphPadPrismTM for statistical comparisons among three or more groupsand were followed by Tukey’s test for post hoc comparisonbetween each group. Results are displayed as mean � SEM.
RESULTS
GBM Cells Grow as Spheres Without ExogenousMitogenic Stimulation
To date, studies examining BTSCs have used EGF þ FGF2to stimulate brain tumor sphere formation [10, 15]. One hall-mark of a cancer cell is the ability to proliferate independ-ently of exogenous mitogenic stimulation [13]. We reasonedthat BTSCs, isolated from adult human gliomas, should prolif-erate in SFM in the absence of exogenous EGF þ FGF2. In afirst series of adult human brain cultures, we prepared single-cell suspensions from 11 fresh GBM specimens and four freshtemporal lobe specimens and cultured each using the neuro-sphere culture system. Eight of 11 GBMs generated spheresin SFM alone and in SFM with EGF þ FGF2 (supporting in-formation Table S1). None of the temporal lobe specimens,dissociated and cultured in an identical fashion, generatedneurospheres in either SFM alone or SFM with EGF þ FGF2(supporting information Table S2). Cultures generated fromGBMs, after 21 days in SFM alone, demonstrate the typicalgrowth patterns observed previously for BTSCs [10, 15]: anadherent monolayer of differentiated cells (Fig. 1A) andphase-bright, floating GBM spheres that presumably arisefrom GBM BTSCs (Fig. 1B). Insulin-like growth factor-1(IGF-1) is required for EGF and/or FGF2 to induce prolifera-tion and expansion of NSCs [16]. To eliminate any confound-ing effects of IGF-1 signaling and to ensure that GBM BTSCswere truly exogenous mitogen-independent cells, we per-formed primary cultures using SFM without insulin in parallelwith our regular cultures [16]. GBM BTSCs generated spheresin the absence of insulin (n ¼ 3, data not shown). To confirmthat cells isolated from GBMs were not normal NSCs contam-inating the tumor cell population, we examined the karyotypesof GBM BTSCs. Cells isolated from GBM spheres displayedabnormal karyotypes and thus are probably not normal NSCs(supporting information Fig. S1). Furthermore, we evaluatedgenetic characteristics relevant to GBM, including EGFR andp53, in two examples of GBM BTSCs compared with theoriginal tumors. Evaluation of EGFR copy number usingCISH demonstrated that GBM BTSCs in line BT012 main-tained low-level copy amplification virtually identical to thatobserved in the parent tumor (supporting information Fig.S2). Similarly, evaluation of the p53 mutation status by DNAsequencing revealed identical mutations in both the originaltumor and the corresponding GBM BTSCs in line BT073(supporting information Fig. S2). These data suggest thatGBMs contain BTSCs that have undergone genetic changesresulting in their transformation and acquisition of the abilityto proliferate and form spheres without exogenous mitogens.
EGF Together with FGF2 Increases GBM SphereFormation and Size
EGF and FGF2 are mitogens for adult mammalian NSCs. Todetermine how EGF together with FGF2 affects GBM BTSCs,
we compared the growth of spheres from primary GBM cellsthat had been cultured in either SFM alone or SFM with EGFþ FGF2. GBM spheres generated in SFM alone or in SFMwith EGF þ FGF2 were morphologically identical (supportinginformation Fig. S3). However, the addition of EGF þ FGF2to primary cultures significantly increased the number ofGBM spheres by 50% over the number of spheres generatedin SFM alone (Table 1). Furthermore, the average diameter ofprimary GBM spheres grown in SFM significantly increased2.4-fold by the addition of EGF þ FGF2 (Table 1), and thisled to a corresponding increase in the number of cellsobtained after dissociation of these spheres (data not shown).
To understand how exogenous EGF þ FGF2 increasesGBM sphere number and size, we used flow cytometry, basedon propidium iodide incorporation, to compare apoptosis andthe cell cycle of exogenous mitogen-independent GBMBTSCs with those of cells stimulated with EGF þ FGF2. Ex-ogenous mitogen-independent GBM spheres were dissociatedto single-cell suspensions and then were cultured for 1 weekin the absence or presence of EGF þ FGF2. The addition ofEGFþFGF2 modestly altered the cell cycle of exogenousmitogen-independent GBM BTSCs by significantly increasingthe number of cells in S phase by 53% (Table 1). Most nota-bly, the number of cells undergoing apoptosis was signifi-cantly reduced by 55% with the addition of EGF þ FGF2(Table 1). Thus, EGF þ FGF2-induced increases in bothGBM sphere cell survival and entry into S phase probablyaccount for the increased number and size of GBM spheres.
Exogenous Mitogen-Independent GBM SpheresDemonstrate Multilineage DifferentiationThat Is Altered by EGF 1 FGF2
A fundamental characteristic of NSCs is multipotency, theability to generate neurons, astrocytes, and oligodendrocytes[2]. We sought to determine whether exogenous mitogen-in-dependent BTSCs isolated from GBMs were multipotent.GBM spheres derived from each specimen were differentiatedto assess their phenotypic potential. GBM spheres generatedin SFM alone were capable of differentiating into cellsexpressing markers of astrocytes (GFAP), oligodendrocytes(O4), and neurons (MAP2) (Fig. 1C–1F) in six of eight cul-tures (Fig. 1K). The GBM spheres from BT020 and BT025were also multipotent but did not differentiate into neurons(Fig. 1K). Differentiated cells from exogenous mitogen-inde-pendent primary GBM spheres that simultaneously expressedmore than one mature CNS cell lineage marker were alsoobserved (supporting information Fig. S4A–4D, 4E–4H, 4I–4L). In seven of eight cultures, GBM spheres generated inSFM with EGF þ FGF2 produced spheres that were capableof differentiating into cells that expressed astrocytic, oligo-dendroglial, and neuronal markers (Fig. 1G–1J, 1L). More-over, addition of EGF þ FGF2 during the primary culture pe-riod altered the phenotypic potential of GBM spheres, incomparison with spheres generated in SFM alone, in seven ofeight cultures (Fig. 1K–1L). However, the impact of EGF þFGF2 on phenotypic potential was different in each primaryculture. These findings further demonstrate that GBM spheresisolated in our experiments were derived from BTSCs withaltered differentiation programs, rather than from normalNSCs that had migrated to and infiltrated the tumor.
Exogenous Mitogen-Independent GBMBTSCs Self-Renew
A cardinal property of adult human NSCs is their capacity formitogen-dependent self-renewal [8]. Previous studies haveshown that BTSCs also self-renew in the presence of EGF þ
Kelly, Stechishin, Chojnacki et al. 1725
www.StemCells.com

Figure 1. GBM brain tumor stem cells (BTSCs) proliferate into multipotent spheres and self-renew without exogenous mitogenic stimulation.(A, B): Phase-contrast photomicrographs demonstrate the typical appearance of GBM cells grown using the neurosphere culture system in SFMalone (without EGF þ FGF2); a monolayer of differentiated cells (A) and a floating GBM sphere (B). (C–J): Primary GBM spheres generated inSFM without exogenous EGF þ FGF2 (C–F) or with EGF þ FGF2 (G–J) differentiate into cells that immunostain for characteristic astrocytic(GFAP; C, I), oligodendroglial (O4; D, H), and neuronal (MAP2; E, I) markers confirming their multipotency. (F, J): Merged images. Nucleiwere counterstained with Hoechst 33258. (K–L): Each of the eight GBMs that generated primary spheres in either SFM alone (K) or SFM withEGF þ FGF2 (L) demonstrated individual variation in the sphere phenotypes observed between those generated in the absence (K) or presenceof EGF þ FGF2 (L). (M): Seven of eight primary GBM sphere cultures self-renewed when generated in SFM without or with EGF þ FGF2.(N): Secondary exogenous mitogen independent GBM spheres remained multipotent and continued to generate astrocytes, oligodendrocytes andneurons. Sphere phenotypes are indicated by the following: A, astrocytes only; A/O, astrocytes and oligodendrocytes; A/N, astrocytes and neu-rons; A/O/N, astrocytes, oligodendrocytes, and neurons (n � 10 spheres per sample). Scale bars ¼ 10 lm (A), 50 lm (B), 50 lm (C–J, O).Abbreviations: EGF, epidermal growth factor; FGF2, fibroblast growth factor-2; GBM, glioblastoma multiforme; GFAP, glial fibrillary acidic pro-tein; MAP2, microtubule-associated protein-2; SFM, serum-free culture medium.
1726 Exogenous Mitogen-Independent BTSCs

FGF2 [10]. To test BTSC self-renewal in the absence of ex-ogenous mitogens, primary GBM spheres, grown in SFMalone were dissociated to single cells and replated in SFM.Seven of eight GBM cultures that generated primary spheresin SFM alone generated secondary spheres; only BT028 didnot generate secondary spheres (Fig. 1M). Immunocytochemi-cal analysis was performed to assess the phenotypes of sec-ondary GBM spheres. Exogenous mitogen-independentBTSCs retained their capacity for multilineage differentiationwhen passaged (Fig. 1N). Thus, exogenous mitogen-independ-ent GBM BTSCs are capable of self-renewal with retention ofmultilineage potential.
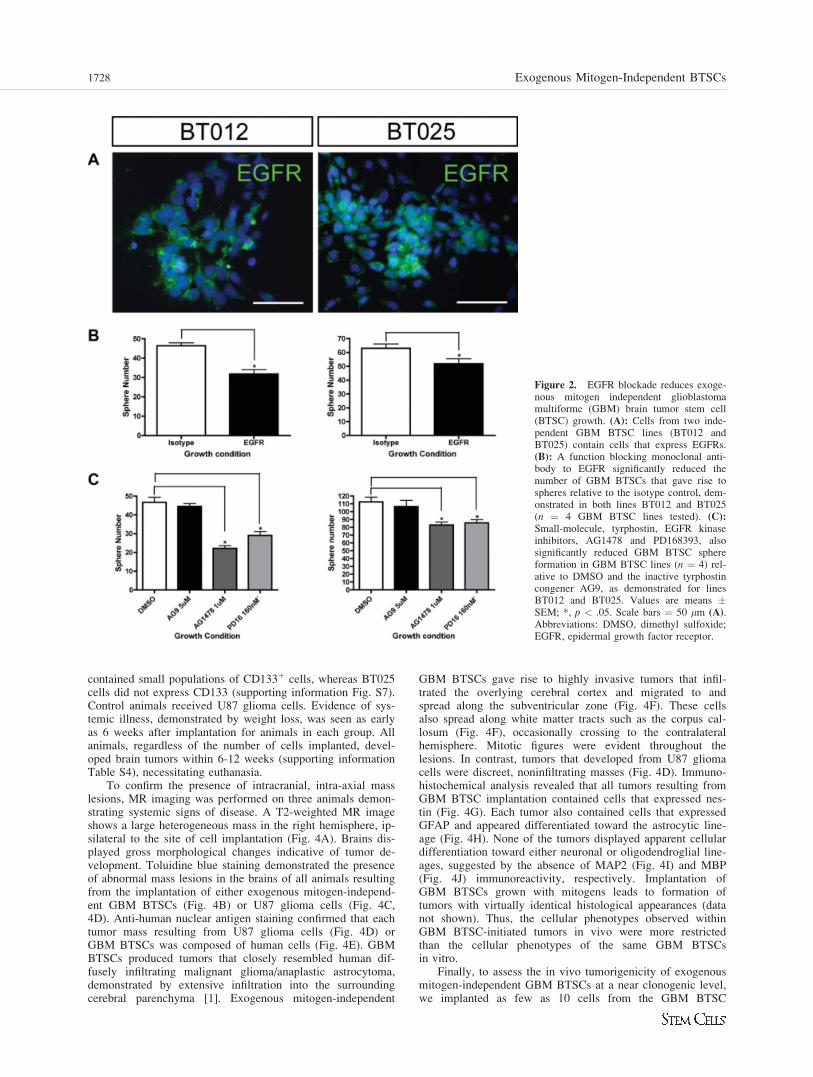
Exogenous Mitogen-Independent GBM BTSCProliferation Is Mediated, in Part, byEGFR Signaling
EGF signaling through the EGFR is fundamental for the pro-liferation of not only NSCs [2, 5, 17] but also astrocytes [18].EGFR mutation or amplification occurs in 50-60% of high-grade gliomas, indicating the importance of EGF signaling tothe pathobiology of these cancers [19, 20]. To gain insightinto the mechanisms underlying exogenous mitogen-independ-ent GBM BTSC proliferation we first examined the expres-sion of EGFR in GBM BTSC cultures. Cells from GBMBTSC lines (n ¼ 4) expressed EGFR by both RT-PCR (notshown) and immunocytochemical staining (Fig. 2A), withoutevidence for expression of the constitutively active mutantEGF receptor, EGFRVIII, known to occur in GBM (notshown). We then asked whether blocking signaling throughEGFR would eliminate exogenous mitogen-independent GBMBTSC growth. Exogenous mitogen-independent GBM BTSCs(n ¼ 4 lines) were dissociated to single-cell suspensions andcultured in the presence of an EGFR function-blocking mono-clonal antibody. Monoclonal antibody blockade of EGFR sig-nificantly decreased the number of spheres formed by GBMBTSCs (Fig. 2B). Furthermore, inhibition of EGFR using tar-geted, small molecule receptor tyrosine kinase inhibitors alsoled to a significant reduction in the number of spheres formedby GBM BTSCs (Fig. 2C). Each BTSC line examined exhib-ited different degrees of growth reduction by EGFR inhibition(Fig. 2B, 2C), and further growth reduction was not achievedby increasing the concentration of either the blocking anti-body or the small molecule inhibitors (not shown). Specificityof both the anti-EGFR monoclonal antibody and the smallmolecule inhibitors compared with that of control treatmentswas confirmed by their ability to selectively reduce phospho-
rylation of the EGFR (supporting information Fig. S5). Thus,exogenous mitogen-independent GBM BTSC proliferation ismediated, in part, through endogenous activation of EGFRand its signaling pathways. Because abrogation of EGFR sig-naling reduces, but does not eliminate exogenous mitogen-in-dependent GBM BTSC proliferation, other mechanisms mustcontribute to constitutive GBM BTSC growth.
Both CD1331 and CD1332 GBM Cells ProliferateWithout Exogenous Mitogenic Stimulation
The NSC marker CD133 has been used to identify and pro-spectively isolate BTSCs from primary brain tumors [10, 12].It has recently been demonstrated, however, that some GBMBTSCs do not express CD133 [11]. We set out to prospec-tively determine whether CD133 expression predicts exoge-nous mitogen-independent GBM cell growth. We used threeGBM BTSC cultures (BT012, BT048, and BT053) (support-ing information Table S1), each of which was established inSFM and has maintained a population of CD133þ cells overmultiple passages. We prepared single-cell suspensions fromthe BTSC lines and then analyzed each for CD133 expressionby flow cytometry. We performed FACS on these cells, basedon CD133 expression, and plated both the CD133�- andCD133þ-enriched fractions in a limiting dilution assay inSFM. For one representative BTSC line, BT048 (Fig. 3A,supporting information Fig. S6A), both highly enrichedCD133þ (supporting information Fig. S6B) and CD133� (sup-porting information Fig. S6C) cell populations proliferated inSFM (Fig. 3Aa) and gave rise to multipotent spheres (Fig.3Aa0), as did each of the other two BTSC lines (not shown).Cells from both CD133þ and CD133� spheres from all linesexpressed the NSC marker nestin (Fig. 3Aa00).
To confirm this finding and to examine the impact ofCD133 expression on exogenous mitogen-independent BTSCgrowth in primary culture we obtained fresh tissue from sevenadditional GBMs (supporting information Tables S1 and S3).Single-cell suspensions were prepared, CD133 expression wasdetermined by flow cytometry, and FACS was performed.Both CD133� and, when present, CD133þ fractions wereplated in a limiting dilution assay in SFM. Four of sevenGBMs contained CD133þ cells (supporting information TableS3; Fig. 3B) in the primary tumor that proliferated in the ab-sence of exogenous EGF þ FGF2 (Fig. 3Bb) and gave rise tomultipotent spheres (Fig. 3Bb0). Cells from these spheres alsoexpressed nestin (Fig. 3Bb00). Six of seven GBMs containedCD133� cells (Fig. 3C; supporting information Table S3) thatalso gave rise to brain tumor spheres in the absence of exoge-nous EGF þ FGF2 (Fig. 3Cc). Spheres that arose fromCD133� cells were also multipotent (Fig. 3Cc0) and expressednestin (Fig. 3Cc00). Only one GBM, BT075, containedCD133� cells that did not survive in culture under any condi-tion. Thus, exogenous mitogen-independent GBM BTSCgrowth occurs irrespective of CD133 expression.
Exogenous Mitogen-Independent GBM BTSCs FormIntracranial Tumors in Immunocompromised Mice
Our in vitro cell culture results suggested that exogenousmitogen-independent sphere-forming cells isolated from bothCD133þ and CD133� adult GBMs were BTSCs. In vivo tu-mor initiation is a more rigorous test of cancer stem cell iden-tity [21]. We sought to confirm the in vivo tumorigenicity ofcells isolated from GBM spheres generated without exogenousmitogens. Exogenous mitogen-independent GBM sphere cellswere implanted into the brains of NOD-SCID mice (n ¼ 3BTSC lines, BT012, BT025, and BT030, between passages 5and 7) (supporting information Table S4). BT012 and BT030
Table 1. Actions of EGF 1 FGF2 on GBM brain tumor stemcells
SFM EGF þ FGF2
No. primary GBM spheres (per200,000 cells)
198 � 28 297 � 43a
Primary GBM sphere size (lm) 97.5 � 3.3 233.8 � 25.1a
Cell cycle analysis (%)G1 72.7 � 5.0 67.0 � 4.0S 10.2 � 2.8 15.6 � 3.6a
G2 17.1 � 7.5 17.4 � 7.5Apoptosis(%) 13.6 � 2.8 6.1 � 3.2a
For numbers and size of primary spheres, n ¼ 4; for cell cycleanalysis and apoptosis, n ¼ 3. Data are means � SEM.ap < .05.Abbreviations: EGF, epidermal growth factor; FGF2, fibroblastgrowth factor-2; GBM, glioblastoma multiforme; SFM, serum-free medium.
Kelly, Stechishin, Chojnacki et al. 1727
www.StemCells.com
contained small populations of CD133þ cells, whereas BT025cells did not express CD133 (supporting information Fig. S7).Control animals received U87 glioma cells. Evidence of sys-temic illness, demonstrated by weight loss, was seen as earlyas 6 weeks after implantation for animals in each group. Allanimals, regardless of the number of cells implanted, devel-oped brain tumors within 6-12 weeks (supporting informationTable S4), necessitating euthanasia.
To confirm the presence of intracranial, intra-axial masslesions, MR imaging was performed on three animals demon-strating systemic signs of disease. A T2-weighted MR imageshows a large heterogeneous mass in the right hemisphere, ip-silateral to the site of cell implantation (Fig. 4A). Brains dis-played gross morphological changes indicative of tumor de-velopment. Toluidine blue staining demonstrated the presenceof abnormal mass lesions in the brains of all animals resultingfrom the implantation of either exogenous mitogen-independ-ent GBM BTSCs (Fig. 4B) or U87 glioma cells (Fig. 4C,4D). Anti-human nuclear antigen staining confirmed that eachtumor mass resulting from U87 glioma cells (Fig. 4D) orGBM BTSCs was composed of human cells (Fig. 4E). GBMBTSCs produced tumors that closely resembled human dif-fusely infiltrating malignant glioma/anaplastic astrocytoma,demonstrated by extensive infiltration into the surroundingcerebral parenchyma [1]. Exogenous mitogen-independent
GBM BTSCs gave rise to highly invasive tumors that infil-trated the overlying cerebral cortex and migrated to andspread along the subventricular zone (Fig. 4F). These cellsalso spread along white matter tracts such as the corpus cal-losum (Fig. 4F), occasionally crossing to the contralateralhemisphere. Mitotic figures were evident throughout thelesions. In contrast, tumors that developed from U87 gliomacells were discreet, noninfiltrating masses (Fig. 4D). Immuno-histochemical analysis revealed that all tumors resulting fromGBM BTSC implantation contained cells that expressed nes-tin (Fig. 4G). Each tumor also contained cells that expressedGFAP and appeared differentiated toward the astrocytic line-age (Fig. 4H). None of the tumors displayed apparent cellulardifferentiation toward either neuronal or oligodendroglial line-ages, suggested by the absence of MAP2 (Fig. 4I) and MBP(Fig. 4J) immunoreactivity, respectively. Implantation ofGBM BTSCs grown with mitogens leads to formation oftumors with virtually identical histological appearances (datanot shown). Thus, the cellular phenotypes observed withinGBM BTSC-initiated tumors in vivo were more restrictedthan the cellular phenotypes of the same GBM BTSCsin vitro.
Finally, to assess the in vivo tumorigenicity of exogenousmitogen-independent GBM BTSCs at a near clonogenic level,we implanted as few as 10 cells from the GBM BTSC
Figure 2. EGFR blockade reduces exoge-nous mitogen independent glioblastomamultiforme (GBM) brain tumor stem cell(BTSC) growth. (A): Cells from two inde-pendent GBM BTSC lines (BT012 andBT025) contain cells that express EGFRs.(B): A function blocking monoclonal anti-body to EGFR significantly reduced thenumber of GBM BTSCs that gave rise tospheres relative to the isotype control, dem-onstrated in both lines BT012 and BT025(n ¼ 4 GBM BTSC lines tested). (C):
Small-molecule, tyrphostin, EGFR kinaseinhibitors, AG1478 and PD168393, alsosignificantly reduced GBM BTSC sphereformation in GBM BTSC lines (n ¼ 4) rel-ative to DMSO and the inactive tyrphostincongener AG9, as demonstrated for linesBT012 and BT025. Values are means �SEM; *, p < .05. Scale bars ¼ 50 lm (A).Abbreviations: DMSO, dimethyl sulfoxide;EGFR, epidermal growth factor receptor.
1728 Exogenous Mitogen-Independent BTSCs

culture, BT012 (supporting information Table S5), which con-tains a small population of CD133þ cells (supporting informa-tion Fig. S7), into immunocompromised mice. Animals weresacrificed 12 weeks after implantation. Each animal, including
all six animals implanted with 10 cells, demonstrated invasivetumor growth (Fig. 5A–5D) over this time period. We reiso-lated cells from the tumors of two animals that received 1,000cell implants and recultured these in SFM to demonstrate their
Figure 3. CD133þ and CD133� glioblastoma multiforme (GBM) brain tumor stem cells (BTSCs) proliferate without exogenous epidermalgrowth factor (EGF) þ fibroblast growth factor 2 (FGF2) and give rise to multipotent GBM spheres. (A, B, C): Flow cytometric histograms dem-onstrate the presence of both CD133þ and CD133� cells in an established GBM BTSC culture (A) and in two primary GBM samples (B, C).(Aa, Bb, Cc): Highly fluorescence-activated cell sorter-purified CD133þ (Aa, Bb) and CD133� (Aa, Cc) primary GBM cells gave rise to spheresin limiting dilution assays in serum-free medium without EGF þ FGF2. Values are means � SEM; *, p < .05. (Aa0, Bb0, Cc0): All primaryGBM spheres demonstrated multipotency by the presence of astrocytes, oligodendrocytes, and neurons, regardless of whether spheres arose fromCD133þ cells (Bb0) or CD133� cells. (Aa0, Cc0). (Aa00, Bb00, Cc00): GBM spheres that arose from both CD133þ (Bb00) and CD133� (Aa00, Cc00)cells expressed nestin. Scale bars ¼ 50 lm (a0), 50 lm (b0, c0), 50 lm (a00–c00). Abbreviations: B-Tub, b-tubulin; GFAP, glial fibrillary acidic pro-tein; Hoechst, Hoechst 33258; Max, maximum; PE, phycoerythrin.
Kelly, Stechishin, Chojnacki et al. 1729
www.StemCells.com

ability to proliferate as spheres (Fig. 5E) and retain multipo-tency in vitro (Fig. 5F–5H). Thus, GBM spheres, generated inthe absence of EGF þ FGF2, contain exogenous mitogen-in-dependent GBM BTSCs that initiate highly invasive tumorsin vivo.
DISCUSSION
We set out to characterize the growth requirements of GBMBTSCs using the neurosphere culture system. We found thatboth CD133þ and CD133� GBM BTSCs proliferated in theabsence of exogenous mitogens, giving rise to multipotentGBM spheres. The addition of EGF þ FGF2 to GBM spherecultures increased the survival and proliferation of GBM
BTSCs and altered their phenotypic potential in vitro. Exoge-nous mitogen-independent GBM sphere cells behaved likeBTSCs when transplanted into immunocompromised mice, byinitiating highly invasive tumors that resembled humanGBMs.
Exogenous mitogen-independent GBM BTSCs possess thein vitro characteristics previously ascribed to BTSCs, includ-ing the ability to proliferate, self-renew, and differentiatedown multiple neural cell lineages [10]. This finding demon-strates a fundamental difference between the growth require-ments of NSCs, which require exogenous mitogens to prolif-erate [6–9], and GBM BTSCs. Three possibilities couldexplain why GBM BTSCs proliferate without exogenous mi-togenic stimulation: (a) autocrine/paracrine stimulation of re-ceptor tyrosine kinase signaling pathways occurs, (b) cellcycle pathways are constitutively activated, or (c) apoptoticpathways are inactivated [22]. Each of these mechanisms andtheir corresponding genetic mutations have been demonstratedin GBM [23]. Specific examples that might affect GBMBTSCs include EGF receptor mutations [24, 25], autocrine/paracrine platelet-derived growth factor signaling [26], p53mutation [27], and phosphatase and tensin homolog inactiva-tion [25]. Our findings suggest that constitutive activation andsignaling through the wild-type EGFR, probably mediated byautocrine/paracrine mechanisms, plays a role in exogenousmitogen-independent GBM BTSC growth. However, abroga-tion of EGFR signaling did not completely abolish thisgrowth. We are currently investigating additional, putativemechanisms underlying exogenous mitogen-independentGBM BTSC proliferation and sphere formation.
In our study, we found that cells isolated from only 85%(17 of 20) of GBMs survived, proliferated, and generatedspheres in serum-free conditions regardless of whether exoge-nous mitogens were absent or present and irrespective ofCD133 status. The reasons that some GBMs do not formspheres in the neurosphere culture system remain uncertain.This finding probably reflects the inter- and intratumoral het-erogeneity between individual GBMs and the correspondingnumber of BTSCs present in each sample, because samples
Figure 4. Cells from exogenous mitogen-independent glioblastomamultiforme (GBM) spheres form intracranial tumors in immunocom-promised mice. (A): A T2-weighted magnetic resonance image dem-onstrates a large heterogeneous lesion in the right hemisphere of aNOD-SCID mouse after implantation of exogenous mitogen-inde-pendent GBM Brain tumor stem cells (BTSCs). (B, C): Toluidineblue staining demonstrates the presence of large tumors resultingfrom implantation of exogenous mitogen-independent GBM spherecells (B) and U87MG cells (C). (D, F): Antihuman nuclear antigenstaining confirmed the presence of human cells within and composingthe bulk of each tumor that developed in NOD-SCID mice after im-plantation of either U87MG cells (D) or GBM BTSCs (E). ControlU87MG cells formed intracranial tumors with distinct borders and didnot invade surrounding brain parenchyma (D). (F): Brain tumors thatformed in NOD-SCID mice after implantation of exogenous mitogen-independent GBM BTSCs infiltrated overlying cortex (thick arrow),migrated along the corpus callosum (medium arrow), and spread tothe subventricular zone (bordering the lateral ventricles) (thin arrow).(G–J): Brain tumors that formed in NOD-SCID mice after implanta-tion of exogenous mitogen-independent GBM sphere cells containcells that express nestin (G) and GFAP (H). Expression of markersindicating either neuronal (MAP2) (I) or oligodendroglial (MBP) (J)differentiation was not observed in any of tumors that grew in thebrains of NOD-SCID mice after GBM BTSC implantation. Abbrevia-tions: CC, corpus callosum; Cx, cortex; GFAP, glial fibrillary acidicprotein; LV, lateral ventricles; MAP2, microtubule-associated protein-2; MBP, myelin basic protein.
1730 Exogenous Mitogen-Independent BTSCs

differ in location and tissue volume. Finally, some tumorsmay not grow in vitro simply because the cells cannot with-stand the process required to generate a single-cell suspensionfor culturing. The in vitro growth characteristics of individualGBMs may reflect their underlying genetic, molecular, and bi-ological properties. This possibility will need to be investi-
gated by examining many samples to establish whethergrowth characteristics in vitro reflect molecular profiles, natu-ral history and prognosis for individual patients.
Most previous studies investigating BTSCs have used theneurosphere culture system, adding EGF þ FGF2 to stimulatebrain tumor sphere formation and prevent BTSC
Figure 5. As few as 10 exogenous mitogen-independent glioblastoma multiforme (GBM) brain tumor stem cells (BTSCs) initiate invasivetumors and can be reisolated in serum-free medium (SFM). (A–D): Representative coronal H&E sections from two animals, m1 (A, B) and m2(C, D), demonstrate that as few as 10 exogenous mitogen-independent GBM cells initiate highly invasive tumors in immunocompromised mice.Exogenous mitogen-independent GBM BTSCs can be reisolated after tumor initiation in vivo and maintain multipotency in vitro. (E): TypicalGBM spheres arose after reculture, in SFM, of biopsy samples from the brains of immunocompromised mice implanted with exogenous mitogen-independent GBM BTSCs 12 weeks earlier. (F–H): Recultured exogenous mitogen-independent GBM spheres retained multipotency in vitro andgave rise to neurons (arrowhead, G), oligodendrocytes (arrow, G), and astrocytes (arrow, H) after differentiation. Scale bars ¼ 100 lm (E), 100lm (F). Abbreviations: B-III-Tub, b-III-tubulin; GFAP, glial fibrillary acidic protein; Hoechst, Hoechst 33258; LV, lateral ventricle; MAP2,microtubule-associated protein-2
Kelly, Stechishin, Chojnacki et al. 1731
www.StemCells.com

differentiation [10, 15, 28]. Because exogenous mitogens arenot always necessary for GBM BTSC survival and prolifera-tion in vitro, the question of how mitogens affect GBM cells,cultured using the neurosphere culture system, must beaddressed. In our study, addition of EGF þ FGF2 to GBMBTSC cultures enhanced the survival and proliferation ofthese BTSCs and altered their phenotypic potentials in vitro.Mitogens such as EGF or FGF2 may alter the fundamentalproperties of a true cancer stem cell by influencing cellulardifferentiation and phenotypic potential [29]. Increasing cellu-lar proliferation in BTSC cultures may inadvertently lead toamplification and characterization of a subgroup of cells thatare responsive to exogenous mitogens but are not true BTSCs.In addition, because GBM cells possess inherent genetic insta-bility, increasing their proliferation may increase the rate ofspontaneous mutations, leading to irreversible changes overtime. Whether these mitogens lead to isolation of a differentcell population or irreversibly change the genetic makeup ofBTSCs over time remains to be determined. Providing BTSCswith the minimal essential nutrients for survival in vitro, with-out additional mitogenic or differentiating agents is probably animportant factor for maintenance of the original stem cell-likeproperties of BTSCs [28]. Establishing an in vitro system forisolating and maintaining GBM BTSCs that closely resemblethe original tumor cells is crucial to further investigate thesecells and develop novel and relevant therapeutics for GBM.
Exogenous mitogen-independent GBM BTSCs demon-strate the defining characteristic of BTSCs in vivo: the abilityto initiate tumors in immunocompromised mice that recapitu-late the parental tumor [21]. Previous reports have shown thattumors initiated by cells from GBM spheres grown in EGF þFGF2 are much more similar to the human disease than aretumors produced by glioma cell lines [15, 28]. We alsoobserved a significant difference between the invasiveness ofexogenous mitogen-independent GBM BTSCs compared withthat of U87MG cells, the latter of which neither invaded normigrated throughout the brain parenchyma. GBM BTSCs gen-erated with and without EGF þ FGF2 were capable of intra-cranial tumor formation in NOD-SCID mice, and thesetumors were histologically and immunohistochemically indis-tinguishable from one another. We demonstrate for the firsttime that tumor initiation occurs even when only 10 exoge-nous mitogen-independent GBM cells are implanted. Thus,exogenous mitogen-independent GBM BTSC tumor formation
represents a further simplification of the in vivo BTSC animalmodel system for studying human GBM.
CONCLUSION
Both CD133þ and CD133� GBM BTSCs can be distin-guished from NSCs by their unique ability to survive and pro-liferate independently of exogenous mitogenic stimulation.Exogenous mitogen-independent GBM BTSC growth is medi-ated in part by EGFR signaling. The novel experimental sys-tem described here, comprising complementary in vitro and invivo systems, can be used to further investigate GBM and themechanisms that drive GBM BTSC growth. This experimentalsystem will be useful for identifying biological pathways thathave distinct effects on BTSCs and NSCs, enabling the identi-fication of therapeutic targets that could eradicate BTSCswithout affecting NSCs.
ACKNOWLEDGMENTS
This work was supported by a Canadian Institutes of HealthResearch operating grant to S.W. Additional support was pro-vide by a Program Project grant from the National Cancer Insti-tute of Canada with funds from the Terry Fox Foundation to P.F.and D.L.S., the Alberta Cancer Foundation Chair in Brain TumorResearch held by J.G.C., a Natural Sciences and EngineeringResearch Council of Canada studentship award to O.S., andAlberta Heritage Foundation for Medical Research Clinical Fel-lowship, Clinical Investigator and Scientist Awards, respec-tively, to J.J.P.K, I.F.P., and S.W. We thank L. Kennedy andL. Robertson at The University of Calgary Flow Cytometry CoreFacility; Dr. D. Demetrick and S. Hyunh at the University ofCalgary Molecular Pathology Laboratory.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Louis DN, Ohgaki H, Weistler OD et al. WHO Classification ofTumors of the Central Nervous System. 4th ed. Lyon, France: IARC,2007.
2 Reynolds BA, Weiss S. Generation of neurons and astrocytes fromisolated cells of the adult mammalian central nervous system. Science1992;255:1707–1710.
3 Curtis MA, Kam M, Nannmark U et al. Human neuroblasts migrate tothe olfactory bulb via a lateral ventricular extension. Science 2007;315:1243–1249.
4 Doetsch F, Petreanu L, Caille I et al. EGF converts transit-amplifyingneurogenic precursors in the adult brain into multipotent stem cells.Neuron 2002;36:1021–1034.
5 Reynolds BA, Weiss S. Clonal and population analyses demonstratethat an EGF-responsive mammalian embryonic CNS precursor is astem cell. Dev Biol 1996;175:1–13.
6 Chojnacki A, Kelly JJ, Hader W et al. Distinctions between fetal andadult human platelet-derived growth factor-responsive neural precur-sors. Ann Neurol 2008;64:127–142.
7 Nunes MC, Roy NS, Keyoung HM et al. Identification and isolationof multipotential neural progenitor cells from the subcortical whitematter of the adult human brain. Nat Med 2003;9:439–447.
8 Sanai N, Tramontin AD, Quinones-Hinojosa A et al. Unique astrocyteribbon in adult human brain contains neural stem cells but lacks chainmigration. Nature 2004;427:740–744.
9 Villa A, Snyder EY, Vescovi A et al. Establishment and properties ofa growth factor-dependent, perpetual neural stem cell line from thehuman CNS. Exp Neurol 2000;161:67–84.
10 Singh SK, Clarke ID, Terasaki M et al. Identification of a cancer stemcell in human brain tumors. Cancer Res 2003;63:5821–5828.
11 Beier D, Hau P, Proescholdt M et al. CD133þ and CD133� glioblas-toma-derived cancer stem cells show differential growth characteristicsand molecular profiles. Cancer Res 2007;67:4010–4015.
12 Singh SK, Hawkins C, Clarke ID et al. Identification of human braintumour initiating cells. Nature 2004;432:396–401.
13 Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000;100:57–70.
14 Kato H, Kato S, Kumabe T et al. Functional evaluation of p53and PTEN gene mutations in gliomas. Clin Cancer Res 2000;6:3937–3943.
15 Galli R, Binda E, Orfanelli U et al. Isolation and characterization oftumorigenic, stem-like neural precursors from human glioblastoma.Cancer Res 2004;64:7011–7021.
16 Arsenijevic Y, Weiss S, Schneider B et al. Insulin-like growth factor-Iis necessary for neural stem cell proliferation and demonstrates dis-tinct actions of epidermal growth factor and fibroblast growth factor-2. J Neurosci 2001;21:7194–7202.
1732 Exogenous Mitogen-Independent BTSCs

17 Reynolds BA, Tetzlaff W, Weiss S. A multipotent EGF-responsivestriatal embryonic progenitor cell produces neurons and astrocytes.J Neurosci 1992;12:4565–4574.
18 Kornblum HI, Hussain R, Wiesen J et al. Abnormal astrocyte develop-ment and neuronal death in mice lacking the epidermal growth factorreceptor. J Neurosci Res 1998;53:697–717.
19 Maher EA, Furnari FB, Bachoo RM et al. Malignant glioma:genetics and biology of a grave matter. Genes Dev 2001;15:1311–1333.
20 Heimberger AB, Hlatky R, Suki D et al. Prognostic effect of epider-mal growth factor receptor and EGFRvIII in glioblastoma multiformepatients. Clin Cancer Res 2005;11:1462–1466.
21 Vescovi AL, Galli R, Reynolds BA. Brain tumour stem cells. Nat RevCancer 2006;6:425–436.
22 Vogelstein B, Kinzler KW. Cancer genes and the pathways they con-trol. Nat Med 2004;10:789–799.
23 Rasheed BK, Wiltshire RN, Bigner SH et al. Molecular pathogenesisof malignant gliomas. Curr Opin Oncol 1999;11:162–167.
24 Lee JC, Vivanco I, Beroukhim R et al. epidermal growth factor recep-tor activation in glioblastoma through novel missense mutations in theextracellular domain. PLoS Med 2006;3:e485.
25 Mellinghoff IK, Wang MY, Vivanco I et al. Molecular determinantsof the response of glioblastomas to EGFR kinase inhibitors. N Engl JMed 2005;353:2012–2024.
26 Guha A, Dashner K, Black PM et al. Expression of PDGF and PDGFreceptors in human astrocytoma operation specimens supports the ex-istence of an autocrine loop. Int J Cancer 1995;60:168–173.
27 Sidransky D, Mikkelsen T, Schwechheimer K et al. Clonal expansionof p53 mutant cells is associated with brain tumour progression. Na-ture 1992;355:846–847.
28 Lee J, Kotliarova S, Kotliarov Y et al. Tumor stem cells derived fromglioblastomas cultured in bFGF and EGF more closely mirror the phe-notype and genotype of primary tumors than do serum-cultured celllines. Cancer Cell 2006;9:391–403.
29 Linskey ME. Glial ontogeny and glial neoplasia: the search for clo-sure. J Neurooncol 1997;34:5–22.
See www.StemCells.com for supporting information available online.
Kelly, Stechishin, Chojnacki et al. 1733
Copyright © 2022 FDOKUMEN




















