
Size and topology of exogenous DNA as determinant ... - Nature
9
Gene Therapy (2002) 9, 1500–1507 2002 Nature Publishing Group All rights reserved 0969-7128/02 $25.00 www.nature.com/gt RESEARCH ARTICLE Size and topology of exogenous DNA as determinant factors of transgene transcription in mammalian cells H Kamiya, J Yamazaki and H Harashima Graduate School of Pharmaceutical Sciences, Hokkaido University, Sapporo, Japan We prepared a series of linearized DNA duplexes of various lengths in order to examine the effects of topology and the size of exogenous, plasmid-derived DNAs on transgene expression. These linearized DNA duplexes were capped at each end with a highly stable loop (5’-GCGAAGC-3’) to pro- duce a dumbbell-shaped construct that is refractory to exon- uclease digestion in comparison to the analogous uncapped DNA duplexes. Intranuclear microinjection of the DNA dumb- bells into simian COS-7 cells allowed the expression of the green fluorescent protein (GFP) gene on the linearized mol- ecules, which was expressed five- to 10-fold more than that on the circular DNA of the same size. In addition, the expression by the dumbbell DNA was higher than that by the circular plasmid for at least 14 days. Interestingly, the size of the dumbbell DNA affected the transgene expression upon their microinjection into cell nuclei. The GFP Keywords: topology; size; linearization; transcription Introduction Nonviral vectors are highly attractive in gene therapy due to their excellent safety profile, despite their low transgene expression efficiency in comparison with viral vectors. 1 It is thought that the lower expression of non- viral vectors is because of their insufficient and non-uni- form delivery into the nucleus and the low transcription efficiency of the transgene in the nucleus. Much effort has been made to improve the delivery of exogenous DNA into the nucleus. 2–6 In addition, strong promoters and various poly(A) signal sequences have been used to increase transgene expression. However, less attention has been paid to the structure of plasmid-based DNA to enhance the transcription of a transgene. We hypothesized that the topology and the size of the DNA may be important determinants for transgene expression in nonviral vector systems. Plasmid-derived DNAs isolated from Escherichia coli have been used for nonviral delivery systems. They are circular duplexes, and this may be one of the reasons for their low expression efficiency, because the proteins involved in transcription may be loaded on to circular DNA less efficiently than on to linear DNA. In addition, the plas- Correspondence: H Kamiya, Graduate School of Pharmaceutical Sciences, Hokkaido University, Kita-12, Nishi-6, Kita-ku, Sapporo 060-0812, Japan Received 13 November 2001; accepted 6 June 2002 expression efficiency increased with decreasing DNA size below a DNA size of 5.7 kb. The effects of topology and size on the expression of DNAs transfected with cationic lipids are similar to those of DNAs microinjected into cell nuclei. In contrast, microinjection into the cytosol showed the inverse size dependency over a range of 2.3 to 9.4 kb. Thus, transcription of a transgene in the nucleus, but not endo- cytosis or nuclear entry, was influenced by the exogenous DNA structure, and this was the primary determinant of transgene expression upon transfection under our con- ditions. These results indicate that small, linearized DNA duplexes that are end-capped with a highly stable loop (dumbbell-shaped DNA) would be very useful for nonviral gene therapy. Gene Therapy (2002) 9, 1500–1507. doi:10.1038/sj.gt.3301831 mid-derived DNAs contain regions, such as the origin of replication and the antibiotic resistance gene, which are unnecessary for gene expression in mammalian cells and could be deleted. The expression of transgenes on short DNA molecules lacking the extra portions may be efficient. To our knowledge, the influences of DNA struc- ture on transcription in the nucleus have never been examined, although others have tested the effects of DNA structure by transfection. 6–9 To address our hypothesis systematically, we investigated transgene expression by a combination of intranuclear and cytoplasmic micro- injections, and by transfection. The DNA fragment, 5’-GCGAAGC-3’, forms a highly thermostable loop. 10,11 This loop was attached to the ends of linearized DNA by enzymatic ligation to make the ter- minus rigid and to prevent digestion by exonuclease(s). To examine our hypothesis, linearized DNA dumbbells of various lengths were prepared, and their GFP expression was tested by intranuclear and cytosolic microinjections, and by cationic lipid-mediated transfec- tion. Our results suggest that (1) a transgene on a lin- earized DNA dumbbell is expressed five- to 10-fold more than that on circular DNA; and (2) smaller linearized DNA dumbbells showed increased transgene expression. In addition, (3) the order of the expression efficiency of DNA microinjected into the nucleus is very similar to that of transfected DNA, indicating that the structure and the size of DNA molecules affect the transcription, rather than endocytosis or nuclear entry.
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Size and topology of exogenous DNA as determinant ... - Nature

Gene Therapy (2002) 9, 1500–1507 2002 Nature Publishing Group All rights reserved 0969-7128/02 $25.00
www.nature.com/gt
RESEARCH ARTICLE
Size and topology of exogenous DNA as determinantfactors of transgene transcription in mammalian cells
H Kamiya, J Yamazaki and H HarashimaGraduate School of Pharmaceutical Sciences, Hokkaido University, Sapporo, Japan
We prepared a series of linearized DNA duplexes of variouslengths in order to examine the effects of topology and thesize of exogenous, plasmid-derived DNAs on transgeneexpression. These linearized DNA duplexes were capped ateach end with a highly stable loop (5’-GCGAAGC-3’) to pro-duce a dumbbell-shaped construct that is refractory to exon-uclease digestion in comparison to the analogous uncappedDNA duplexes. Intranuclear microinjection of the DNA dumb-bells into simian COS-7 cells allowed the expression of thegreen fluorescent protein (GFP) gene on the linearized mol-ecules, which was expressed five- to 10-fold more than thaton the circular DNA of the same size. In addition, theexpression by the dumbbell DNA was higher than that bythe circular plasmid for at least 14 days. Interestingly, thesize of the dumbbell DNA affected the transgene expressionupon their microinjection into cell nuclei. The GFP
Keywords: topology; size; linearization; transcription
IntroductionNonviral vectors are highly attractive in gene therapydue to their excellent safety profile, despite their lowtransgene expression efficiency in comparison with viralvectors.1 It is thought that the lower expression of non-viral vectors is because of their insufficient and non-uni-form delivery into the nucleus and the low transcriptionefficiency of the transgene in the nucleus. Much effort hasbeen made to improve the delivery of exogenous DNAinto the nucleus.2–6 In addition, strong promoters andvarious poly(A) signal sequences have been used toincrease transgene expression. However, less attentionhas been paid to the structure of plasmid-based DNA toenhance the transcription of a transgene.
We hypothesized that the topology and the size of theDNA may be important determinants for transgeneexpression in nonviral vector systems. Plasmid-derivedDNAs isolated from Escherichia coli have been used fornonviral delivery systems. They are circular duplexes,and this may be one of the reasons for their lowexpression efficiency, because the proteins involved intranscription may be loaded on to circular DNA lessefficiently than on to linear DNA. In addition, the plas-
Correspondence: H Kamiya, Graduate School of Pharmaceutical Sciences,Hokkaido University, Kita-12, Nishi-6, Kita-ku, Sapporo 060-0812, JapanReceived 13 November 2001; accepted 6 June 2002
expression efficiency increased with decreasing DNA sizebelow a DNA size of 5.7 kb. The effects of topology and sizeon the expression of DNAs transfected with cationic lipidsare similar to those of DNAs microinjected into cell nuclei.In contrast, microinjection into the cytosol showed theinverse size dependency over a range of 2.3 to 9.4 kb. Thus,transcription of a transgene in the nucleus, but not endo-cytosis or nuclear entry, was influenced by the exogenousDNA structure, and this was the primary determinant oftransgene expression upon transfection under our con-ditions. These results indicate that small, linearized DNAduplexes that are end-capped with a highly stable loop(dumbbell-shaped DNA) would be very useful for nonviralgene therapy.Gene Therapy (2002) 9, 1500–1507. doi:10.1038/sj.gt.3301831
mid-derived DNAs contain regions, such as the origin ofreplication and the antibiotic resistance gene, which areunnecessary for gene expression in mammalian cells andcould be deleted. The expression of transgenes on shortDNA molecules lacking the extra portions may beefficient. To our knowledge, the influences of DNA struc-ture on transcription in the nucleus have never beenexamined, although others have tested the effects of DNAstructure by transfection.6–9 To address our hypothesissystematically, we investigated transgene expression bya combination of intranuclear and cytoplasmic micro-injections, and by transfection.
The DNA fragment, 5’-GCGAAGC-3’, forms a highlythermostable loop.10,11 This loop was attached to the endsof linearized DNA by enzymatic ligation to make the ter-minus rigid and to prevent digestion by exonuclease(s).To examine our hypothesis, linearized DNA dumbbellsof various lengths were prepared, and their GFPexpression was tested by intranuclear and cytosolicmicroinjections, and by cationic lipid-mediated transfec-tion. Our results suggest that (1) a transgene on a lin-earized DNA dumbbell is expressed five- to 10-fold morethan that on circular DNA; and (2) smaller linearizedDNA dumbbells showed increased transgene expression.In addition, (3) the order of the expression efficiency ofDNA microinjected into the nucleus is very similar to thatof transfected DNA, indicating that the structure and thesize of DNA molecules affect the transcription, ratherthan endocytosis or nuclear entry.
v.kerguelen
Text Box
RETRACTED

Effects of DNA structure on transcriptionH Kamiya et al
1501Results
Experimental schemeWe constructed plasmids, linearized DNA, and dumb-bells of different lengths in order to compare theexpression of the green fluorescent protein (GFP) trans-gene as a function of DNA size. We inserted an‘expression cassette’ containing the cytomegalovirus(CMV) promoter, gene, and poly(A) signal of pQBI 25 tothe backbone of another plasmid, to eliminate effects ofthe neo expressing component on pQBI 25. This new plas-mid, pQBI 25-63, contained the ampicillin resistance geneand the origin of replication, which function in E. coli,and the GFP expression cassette (Figure 1a). The sizes ofplasmid DNAs were increased and decreased by dupli-cation and elimination, respectively, of the backboneparts of pQBI 25-63 (see Materials and methods).
We performed restriction digests on the various plas-mids in order to create linearized DNAs, and cappedtheir termini with a loop-forming oligonucleotide(s) byligase treatment to form a series of DNA dumbbells. Thecapping sequence, 5’-GCGAAGC-3’, is extraordinarilythermostable,10,11 and was incorporated into the loop-for-ming oligonucleotides used in this study (Figure 1b). TheDNA constructs were purified by low melting point aga-rose gel electrophoresis, and subsequent extraction fromthe gel by heating and organic solvent treatments.
It is known that light irradiation in the presence of ethi-dium bromide (EtBr), organic solvent extractions, andUV-irradiation damage DNA,12–16 and that the formationof DNA lesions results in transcriptional inhibition anderrors.17–20 Thus, we purified closed circular moleculesand linearized DNAs in the same gel to compare theeffect of the topology precisely. In addition, we used UV-shadowing instead of EtBr staining for DNA detection inorder to minimize DNA damage during manipulation ofthe plasmid-derived constructs.
Figure 1 Structures of DNAs containing the GFP gene used in thisstudy. (a) Structure of the parental DNA, pQBI 25-63. (b) Oligonucleot-ides forming a highly stable loop used for capping of linearized DNA.These oligonucleotides contain ends that are compatible with the restric-tion enzyme-cleaved ends indicated on the left. (c) Schematic presentationof the DNA dumbbells containing the expression cassette. CMV, CMVpromoter; AmpR, E. coli ampicillin resistance gene; ori, E. coli repli-cation origin.
Gene Therapy
To confirm the end-capping by the hairpin-formingoligonucleotide(s), linearized, but uncapped, and dumb-bell DNA molecules were treated with BAL 31 nuclease,which possesses 5’-to-3’ and 3’-to-5’ exonuclease activi-ties, and were analyzed by 1% agarose gel electro-phoresis. The capped DNA was more resistant to BAL 31nuclease than the uncapped DNA (data not shown).Thus, the expected DNA molecules were obtained byour procedures.
Effect of topology on GFP gene transcriptionFirst, we examined whether the topology of plasmid-derived DNAs affects the transgene expression(transcription). Circular DNA molecules and DNAdumbbells were microinjected into the nuclei of simianCOS-7 cells, together with tetramethylrhodamine-labeleddextran. The presence of the labeled dextran in thenucleus in living cells is a marker of successful microin-jection. GFP expression was monitored by fluorescenceand confocal microscopy, and the ratio of cells expressingGFP to those containing the labeled dextran in theirnuclei was calculated.
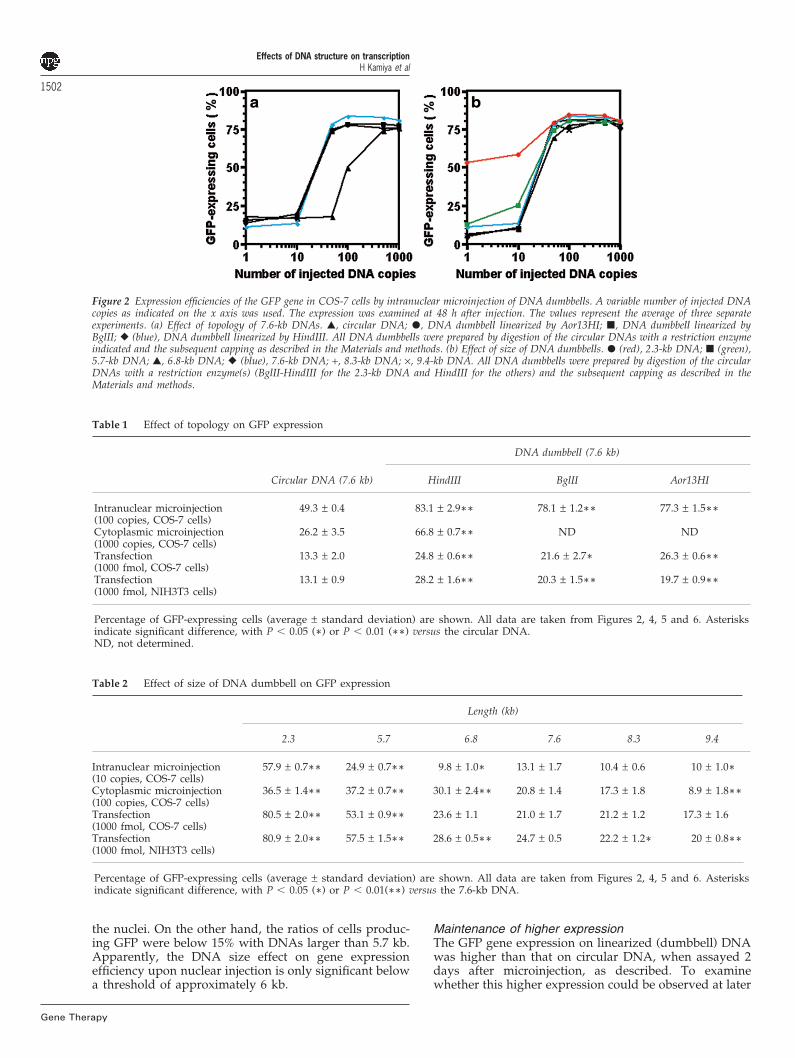
The parental plasmid, pQBI 25-63 (7.6 kb), was used toprepare circular and linearized molecules. The pQBI 25-63 plasmid was treated with BglII, Aor13HI, and HindIIIin order to examine the influence of the position of the‘expression cassette’ (Figure 1c). When microinjected intocell nuclei, the linearized DNA dumbbells showed anenhanced expression ratio as compared with that of thecircular duplex (Figure 2a, Table 1). When 50 copies wereinjected, approximately 75% of the cells expressed GFP,in the case of the linearized DNA dumbbells. On theother hand, the expression was only 18% for the circularDNA. When a 10-fold greater amount (500 copies) of thecircular DNA was injected, 74% GFP expression wasobtained. The copy numbers for 50% expression, calcu-lated from the dose-expression curve, were 20 and 100copies for the dumbbell and circular duplexes, respect-ively. Thus, the linearization and capping facilitatedtransgene expression by about five- to 10-fold. When theexpression cassette was located near the ends (HindIII-and BglII-digested duplexes) or near the center (Aor13HI-treated DNA) of the DNA dumbbells, GFP expressionwas very similar (Figures 1c and 2a). Thus, no obviouseffect was observed with the different locations of theexpression cassette.
Effect of size on GFP gene expressionThe enhanced gene expression caused by plasmid lin-earization and dumbbell formation prompted us to inves-tigate the effect of the length of the linearized DNA onGFP expression. Six different-sized molecules were exam-ined: 5.7 to 9.4 kb DNAs were prepared by HindIII diges-tions of the corresponding plasmids, and a 2.3-kb duplexwas obtained by HindIII-BglII double digestion of pQBI25-63 (Figure 1c). Even the smallest DNA duplex containsthe minimal sequence required for GFP expression. Sur-prisingly, GFP expression was highly correlated with theDNA dumbbell length, as shown in Figure 2b and Table2. The best expression was obtained with the smallestDNA (2.3 kb), followed by the 5.7-kb DNA. The DNAdumbbells with 6.8–9.3 kb lengths showed similarexpression patterns (Figure 2b). For example, 58% and25% of cells expressed GFP when 10 copies of the 2.3-kband 5.7-kb DNAs, respectively, were microinjected into
Effects of DNA structure on transcriptionH Kamiya et al
1502
Gene Therapy
Figure 2 Expression efficiencies of the GFP gene in COS-7 cells by intranuclear microinjection of DNA dumbbells. A variable number of injected DNAcopies as indicated on the x axis was used. The expression was examined at 48 h after injection. The values represent the average of three separateexperiments. (a) Effect of topology of 7.6-kb DNAs. �, circular DNA; �, DNA dumbbell linearized by Aor13HI; �, DNA dumbbell linearized byBglII; � (blue), DNA dumbbell linearized by HindIII. All DNA dumbbells were prepared by digestion of the circular DNAs with a restriction enzymeindicated and the subsequent capping as described in the Materials and methods. (b) Effect of size of DNA dumbbells. � (red), 2.3-kb DNA; � (green),5.7-kb DNA; �, 6.8-kb DNA; � (blue), 7.6-kb DNA; +, 8.3-kb DNA; ×, 9.4-kb DNA. All DNA dumbbells were prepared by digestion of the circularDNAs with a restriction enzyme(s) (BglII-HindIII for the 2.3-kb DNA and HindIII for the others) and the subsequent capping as described in theMaterials and methods.
Table 1 Effect of topology on GFP expression
DNA dumbbell (7.6 kb)
Circular DNA (7.6 kb) HindIII BglII Aor13HI
Intranuclear microinjection 49.3 ± 0.4 83.1 ± 2.9∗∗ 78.1 ± 1.2∗∗ 77.3 ± 1.5∗∗(100 copies, COS-7 cells)Cytoplasmic microinjection 26.2 ± 3.5 66.8 ± 0.7∗∗ ND ND(1000 copies, COS-7 cells)Transfection 13.3 ± 2.0 24.8 ± 0.6∗∗ 21.6 ± 2.7∗ 26.3 ± 0.6∗∗(1000 fmol, COS-7 cells)Transfection 13.1 ± 0.9 28.2 ± 1.6∗∗ 20.3 ± 1.5∗∗ 19.7 ± 0.9∗∗(1000 fmol, NIH3T3 cells)
Percentage of GFP-expressing cells (average ± standard deviation) are shown. All data are taken from Figures 2, 4, 5 and 6. Asterisksindicate significant difference, with P � 0.05 (∗) or P � 0.01 (∗∗) versus the circular DNA.ND, not determined.
Table 2 Effect of size of DNA dumbbell on GFP expression
Length (kb)
2.3 5.7 6.8 7.6 8.3 9.4
Intranuclear microinjection 57.9 ± 0.7∗∗ 24.9 ± 0.7∗∗ 9.8 ± 1.0∗ 13.1 ± 1.7 10.4 ± 0.6 10 ± 1.0∗(10 copies, COS-7 cells)Cytoplasmic microinjection 36.5 ± 1.4∗∗ 37.2 ± 0.7∗∗ 30.1 ± 2.4∗∗ 20.8 ± 1.4 17.3 ± 1.8 8.9 ± 1.8∗∗(100 copies, COS-7 cells)Transfection 80.5 ± 2.0∗∗ 53.1 ± 0.9∗∗ 23.6 ± 1.1 21.0 ± 1.7 21.2 ± 1.2 17.3 ± 1.6(1000 fmol, COS-7 cells)Transfection 80.9 ± 2.0∗∗ 57.5 ± 1.5∗∗ 28.6 ± 0.5∗∗ 24.7 ± 0.5 22.2 ± 1.2∗ 20 ± 0.8∗∗(1000 fmol, NIH3T3 cells)
Percentage of GFP-expressing cells (average ± standard deviation) are shown. All data are taken from Figures 2, 4, 5 and 6. Asterisksindicate significant difference, with P � 0.05 (∗) or P � 0.01(∗∗) versus the 7.6-kb DNA.
the nuclei. On the other hand, the ratios of cells produc-ing GFP were below 15% with DNAs larger than 5.7 kb.Apparently, the DNA size effect on gene expressionefficiency upon nuclear injection is only significant belowa threshold of approximately 6 kb.
Maintenance of higher expressionThe GFP gene expression on linearized (dumbbell) DNAwas higher than that on circular DNA, when assayed 2days after microinjection, as described. To examinewhether this higher expression could be observed at later
Effects of DNA structure on transcriptionH Kamiya et al
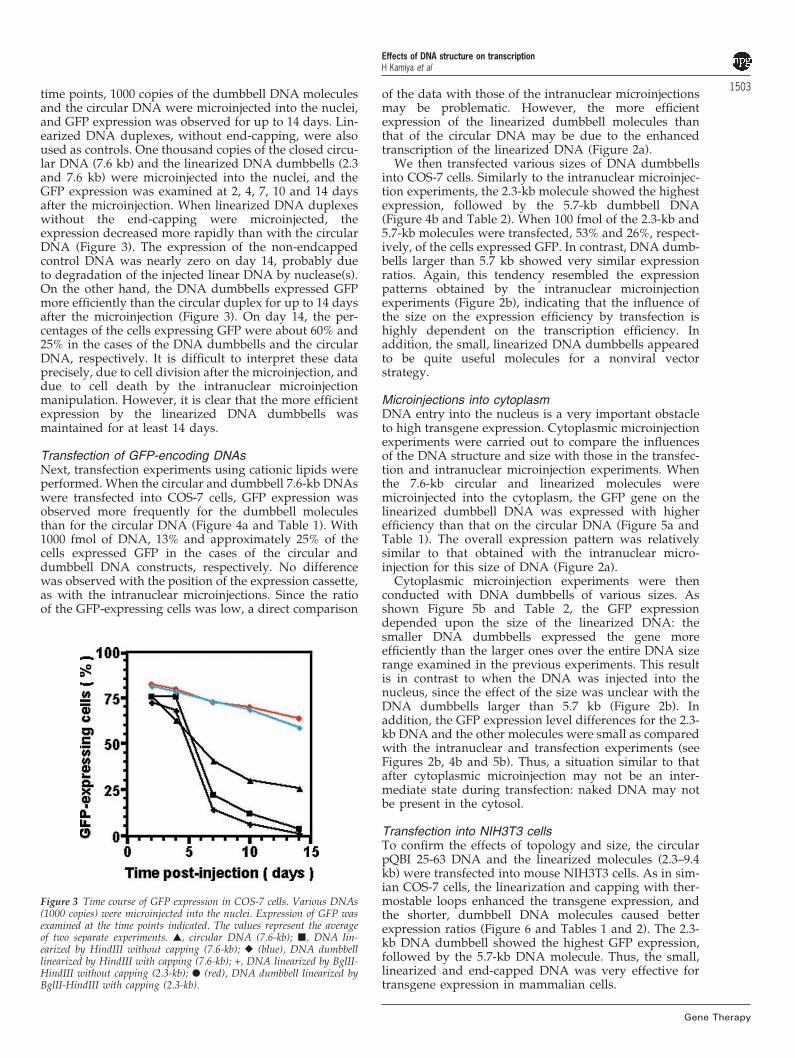
1503time points, 1000 copies of the dumbbell DNA moleculesand the circular DNA were microinjected into the nuclei,and GFP expression was observed for up to 14 days. Lin-earized DNA duplexes, without end-capping, were alsoused as controls. One thousand copies of the closed circu-lar DNA (7.6 kb) and the linearized DNA dumbbells (2.3and 7.6 kb) were microinjected into the nuclei, and theGFP expression was examined at 2, 4, 7, 10 and 14 daysafter the microinjection. When linearized DNA duplexeswithout the end-capping were microinjected, theexpression decreased more rapidly than with the circularDNA (Figure 3). The expression of the non-endcappedcontrol DNA was nearly zero on day 14, probably dueto degradation of the injected linear DNA by nuclease(s).On the other hand, the DNA dumbbells expressed GFPmore efficiently than the circular duplex for up to 14 daysafter the microinjection (Figure 3). On day 14, the per-centages of the cells expressing GFP were about 60% and25% in the cases of the DNA dumbbells and the circularDNA, respectively. It is difficult to interpret these dataprecisely, due to cell division after the microinjection, anddue to cell death by the intranuclear microinjectionmanipulation. However, it is clear that the more efficientexpression by the linearized DNA dumbbells wasmaintained for at least 14 days.
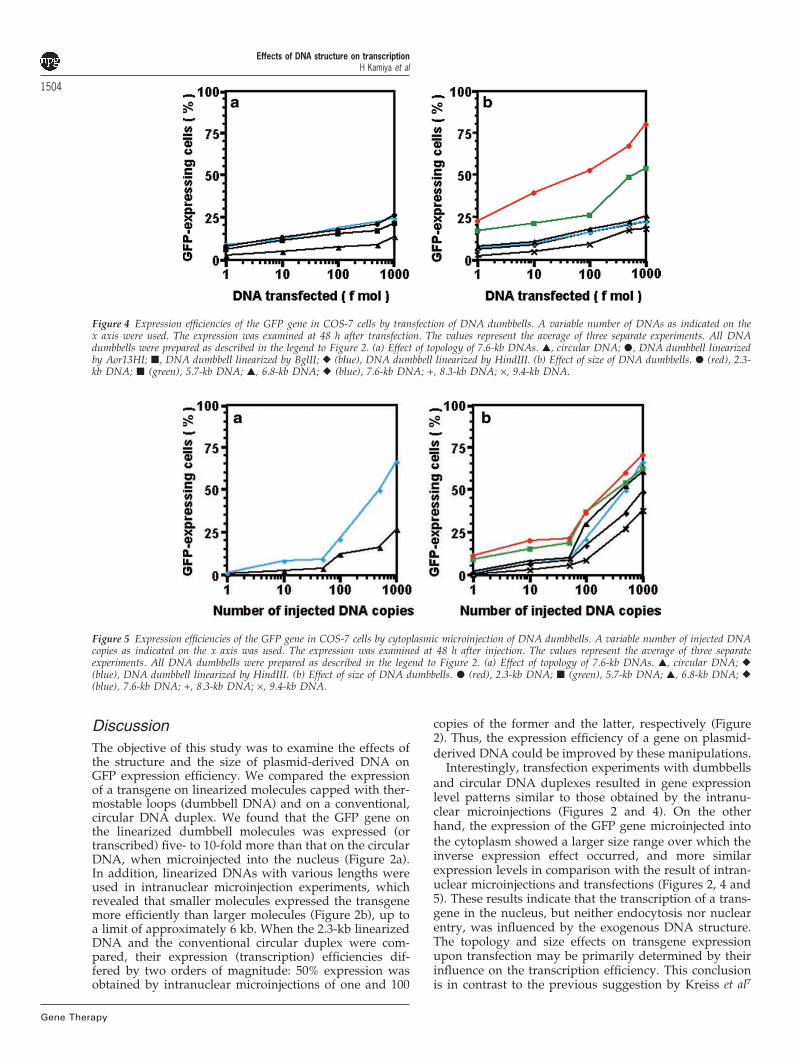
Transfection of GFP-encoding DNAsNext, transfection experiments using cationic lipids wereperformed. When the circular and dumbbell 7.6-kb DNAswere transfected into COS-7 cells, GFP expression wasobserved more frequently for the dumbbell moleculesthan for the circular DNA (Figure 4a and Table 1). With1000 fmol of DNA, 13% and approximately 25% of thecells expressed GFP in the cases of the circular anddumbbell DNA constructs, respectively. No differencewas observed with the position of the expression cassette,as with the intranuclear microinjections. Since the ratioof the GFP-expressing cells was low, a direct comparison
Figure 3 Time course of GFP expression in COS-7 cells. Various DNAs(1000 copies) were microinjected into the nuclei. Expression of GFP wasexamined at the time points indicated. The values represent the averageof two separate experiments. �, circular DNA (7.6-kb); �, DNA lin-earized by HindIII without capping (7.6-kb); � (blue), DNA dumbbelllinearized by HindIII with capping (7.6-kb); +, DNA linearized by BglII-HindIII without capping (2.3-kb); � (red), DNA dumbbell linearized byBglII-HindIII with capping (2.3-kb).
Gene Therapy
of the data with those of the intranuclear microinjectionsmay be problematic. However, the more efficientexpression of the linearized dumbbell molecules thanthat of the circular DNA may be due to the enhancedtranscription of the linearized DNA (Figure 2a).
We then transfected various sizes of DNA dumbbellsinto COS-7 cells. Similarly to the intranuclear microinjec-tion experiments, the 2.3-kb molecule showed the highestexpression, followed by the 5.7-kb dumbbell DNA(Figure 4b and Table 2). When 100 fmol of the 2.3-kb and5.7-kb molecules were transfected, 53% and 26%, respect-ively, of the cells expressed GFP. In contrast, DNA dumb-bells larger than 5.7 kb showed very similar expressionratios. Again, this tendency resembled the expressionpatterns obtained by the intranuclear microinjectionexperiments (Figure 2b), indicating that the influence ofthe size on the expression efficiency by transfection ishighly dependent on the transcription efficiency. Inaddition, the small, linearized DNA dumbbells appearedto be quite useful molecules for a nonviral vectorstrategy.
Microinjections into cytoplasmDNA entry into the nucleus is a very important obstacleto high transgene expression. Cytoplasmic microinjectionexperiments were carried out to compare the influencesof the DNA structure and size with those in the transfec-tion and intranuclear microinjection experiments. Whenthe 7.6-kb circular and linearized molecules weremicroinjected into the cytoplasm, the GFP gene on thelinearized dumbbell DNA was expressed with higherefficiency than that on the circular DNA (Figure 5a andTable 1). The overall expression pattern was relativelysimilar to that obtained with the intranuclear micro-injection for this size of DNA (Figure 2a).
Cytoplasmic microinjection experiments were thenconducted with DNA dumbbells of various sizes. Asshown Figure 5b and Table 2, the GFP expressiondepended upon the size of the linearized DNA: thesmaller DNA dumbbells expressed the gene moreefficiently than the larger ones over the entire DNA sizerange examined in the previous experiments. This resultis in contrast to when the DNA was injected into thenucleus, since the effect of the size was unclear with theDNA dumbbells larger than 5.7 kb (Figure 2b). Inaddition, the GFP expression level differences for the 2.3-kb DNA and the other molecules were small as comparedwith the intranuclear and transfection experiments (seeFigures 2b, 4b and 5b). Thus, a situation similar to thatafter cytoplasmic microinjection may not be an inter-mediate state during transfection: naked DNA may notbe present in the cytosol.
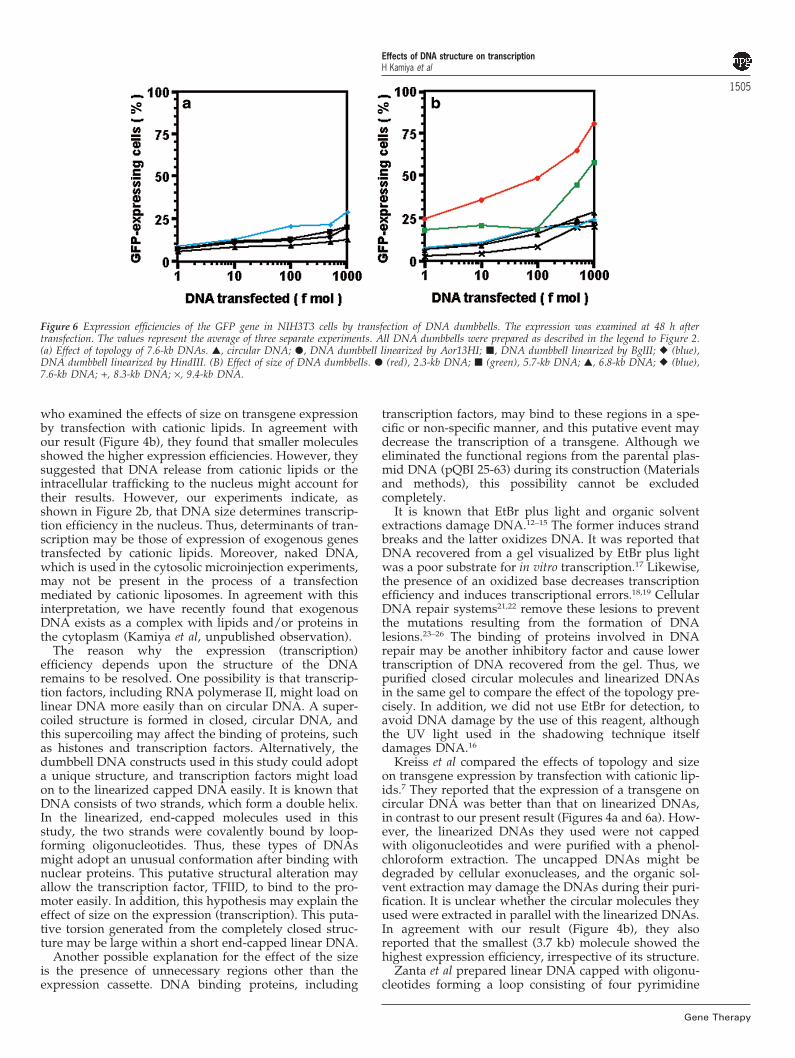
Transfection into NIH3T3 cellsTo confirm the effects of topology and size, the circularpQBI 25-63 DNA and the linearized molecules (2.3–9.4kb) were transfected into mouse NIH3T3 cells. As in sim-ian COS-7 cells, the linearization and capping with ther-mostable loops enhanced the transgene expression, andthe shorter, dumbbell DNA molecules caused betterexpression ratios (Figure 6 and Tables 1 and 2). The 2.3-kb DNA dumbbell showed the highest GFP expression,followed by the 5.7-kb DNA molecule. Thus, the small,linearized and end-capped DNA was very effective fortransgene expression in mammalian cells.
Effects of DNA structure on transcriptionH Kamiya et al
1504
Gene Therapy
Figure 4 Expression efficiencies of the GFP gene in COS-7 cells by transfection of DNA dumbbells. A variable number of DNAs as indicated on thex axis were used. The expression was examined at 48 h after transfection. The values represent the average of three separate experiments. All DNAdumbbells were prepared as described in the legend to Figure 2. (a) Effect of topology of 7.6-kb DNAs. �, circular DNA; �, DNA dumbbell linearizedby Aor13HI; �, DNA dumbbell linearized by BglII; � (blue), DNA dumbbell linearized by HindIII. (b) Effect of size of DNA dumbbells. � (red), 2.3-kb DNA; � (green), 5.7-kb DNA; �, 6.8-kb DNA; � (blue), 7.6-kb DNA; +, 8.3-kb DNA; ×, 9.4-kb DNA.
Figure 5 Expression efficiencies of the GFP gene in COS-7 cells by cytoplasmic microinjection of DNA dumbbells. A variable number of injected DNAcopies as indicated on the x axis was used. The expression was examined at 48 h after injection. The values represent the average of three separateexperiments. All DNA dumbbells were prepared as described in the legend to Figure 2. (a) Effect of topology of 7.6-kb DNAs. �, circular DNA; �(blue), DNA dumbbell linearized by HindIII. (b) Effect of size of DNA dumbbells. � (red), 2.3-kb DNA; � (green), 5.7-kb DNA; �, 6.8-kb DNA; �(blue), 7.6-kb DNA; +, 8.3-kb DNA; ×, 9.4-kb DNA.
DiscussionThe objective of this study was to examine the effects ofthe structure and the size of plasmid-derived DNA onGFP expression efficiency. We compared the expressionof a transgene on linearized molecules capped with ther-mostable loops (dumbbell DNA) and on a conventional,circular DNA duplex. We found that the GFP gene onthe linearized dumbbell molecules was expressed (ortranscribed) five- to 10-fold more than that on the circularDNA, when microinjected into the nucleus (Figure 2a).In addition, linearized DNAs with various lengths wereused in intranuclear microinjection experiments, whichrevealed that smaller molecules expressed the transgenemore efficiently than larger molecules (Figure 2b), up toa limit of approximately 6 kb. When the 2.3-kb linearizedDNA and the conventional circular duplex were com-pared, their expression (transcription) efficiencies dif-fered by two orders of magnitude: 50% expression wasobtained by intranuclear microinjections of one and 100
copies of the former and the latter, respectively (Figure2). Thus, the expression efficiency of a gene on plasmid-derived DNA could be improved by these manipulations.
Interestingly, transfection experiments with dumbbellsand circular DNA duplexes resulted in gene expressionlevel patterns similar to those obtained by the intranu-clear microinjections (Figures 2 and 4). On the otherhand, the expression of the GFP gene microinjected intothe cytoplasm showed a larger size range over which theinverse expression effect occurred, and more similarexpression levels in comparison with the result of intran-uclear microinjections and transfections (Figures 2, 4 and5). These results indicate that the transcription of a trans-gene in the nucleus, but neither endocytosis nor nuclearentry, was influenced by the exogenous DNA structure.The topology and size effects on transgene expressionupon transfection may be primarily determined by theirinfluence on the transcription efficiency. This conclusionis in contrast to the previous suggestion by Kreiss et al7
Effects of DNA structure on transcriptionH Kamiya et al
1505
Figure 6 Expression efficiencies of the GFP gene in NIH3T3 cells by transfection of DNA dumbbells. The expression was examined at 48 h aftertransfection. The values represent the average of three separate experiments. All DNA dumbbells were prepared as described in the legend to Figure 2.(a) Effect of topology of 7.6-kb DNAs. �, circular DNA; �, DNA dumbbell linearized by Aor13HI; �, DNA dumbbell linearized by BglII; � (blue),DNA dumbbell linearized by HindIII. (B) Effect of size of DNA dumbbells. � (red), 2.3-kb DNA; � (green), 5.7-kb DNA; �, 6.8-kb DNA; � (blue),7.6-kb DNA; +, 8.3-kb DNA; ×, 9.4-kb DNA.
who examined the effects of size on transgene expressionby transfection with cationic lipids. In agreement withour result (Figure 4b), they found that smaller moleculesshowed the higher expression efficiencies. However, theysuggested that DNA release from cationic lipids or theintracellular trafficking to the nucleus might account fortheir results. However, our experiments indicate, asshown in Figure 2b, that DNA size determines transcrip-tion efficiency in the nucleus. Thus, determinants of tran-scription may be those of expression of exogenous genestransfected by cationic lipids. Moreover, naked DNA,which is used in the cytosolic microinjection experiments,may not be present in the process of a transfectionmediated by cationic liposomes. In agreement with thisinterpretation, we have recently found that exogenousDNA exists as a complex with lipids and/or proteins inthe cytoplasm (Kamiya et al, unpublished observation).
The reason why the expression (transcription)efficiency depends upon the structure of the DNAremains to be resolved. One possibility is that transcrip-tion factors, including RNA polymerase II, might load onlinear DNA more easily than on circular DNA. A super-coiled structure is formed in closed, circular DNA, andthis supercoiling may affect the binding of proteins, suchas histones and transcription factors. Alternatively, thedumbbell DNA constructs used in this study could adopta unique structure, and transcription factors might loadon to the linearized capped DNA easily. It is known thatDNA consists of two strands, which form a double helix.In the linearized, end-capped molecules used in thisstudy, the two strands were covalently bound by loop-forming oligonucleotides. Thus, these types of DNAsmight adopt an unusual conformation after binding withnuclear proteins. This putative structural alteration mayallow the transcription factor, TFIID, to bind to the pro-moter easily. In addition, this hypothesis may explain theeffect of size on the expression (transcription). This puta-tive torsion generated from the completely closed struc-ture may be large within a short end-capped linear DNA.
Another possible explanation for the effect of the sizeis the presence of unnecessary regions other than theexpression cassette. DNA binding proteins, including
Gene Therapy
transcription factors, may bind to these regions in a spe-cific or non-specific manner, and this putative event maydecrease the transcription of a transgene. Although weeliminated the functional regions from the parental plas-mid DNA (pQBI 25-63) during its construction (Materialsand methods), this possibility cannot be excludedcompletely.
It is known that EtBr plus light and organic solventextractions damage DNA.12–15 The former induces strandbreaks and the latter oxidizes DNA. It was reported thatDNA recovered from a gel visualized by EtBr plus lightwas a poor substrate for in vitro transcription.17 Likewise,the presence of an oxidized base decreases transcriptionefficiency and induces transcriptional errors.18,19 CellularDNA repair systems21,22 remove these lesions to preventthe mutations resulting from the formation of DNAlesions.23–26 The binding of proteins involved in DNArepair may be another inhibitory factor and cause lowertranscription of DNA recovered from the gel. Thus, wepurified closed circular molecules and linearized DNAsin the same gel to compare the effect of the topology pre-cisely. In addition, we did not use EtBr for detection, toavoid DNA damage by the use of this reagent, althoughthe UV light used in the shadowing technique itselfdamages DNA.16
Kreiss et al compared the effects of topology and sizeon transgene expression by transfection with cationic lip-ids.7 They reported that the expression of a transgene oncircular DNA was better than that on linearized DNAs,in contrast to our present result (Figures 4a and 6a). How-ever, the linearized DNAs they used were not cappedwith oligonucleotides and were purified with a phenol-chloroform extraction. The uncapped DNAs might bedegraded by cellular exonucleases, and the organic sol-vent extraction may damage the DNAs during their puri-fication. It is unclear whether the circular molecules theyused were extracted in parallel with the linearized DNAs.In agreement with our result (Figure 4b), they alsoreported that the smallest (3.7 kb) molecule showed thehighest expression efficiency, irrespective of its structure.
Zanta et al prepared linear DNA capped with oligonu-cleotides forming a loop consisting of four pyrimidine

Effects of DNA structure on transcriptionH Kamiya et al
1506
Gene Therapy
bases.6 This DNA was similar to the shortest DNA (2.3kb) used in our study in that it contained only the essen-tial region for transgene expression in addition to theircap structures. They observed that nearly equal luciferaseexpression was obtained by the transfection of linearizedDNA with end-capping, depending upon the cell strain.However, we found that the shortest DNA was much bet-ter than the closed circular molecule (Figures 4 and 6).This discrepancy may be due to the purification method.They separated the linearized DNA by ultracentrifug-ation. This ultracentrifugation is usually carried out inthe presence of EtBr, and the linearized DNA that theyused may have been damaged by this reagent, althougha detailed experimental procedure was not provided. Itis also unknown whether the circular molecule they usedwas re-purified in parallel with the linearized DNA.Thus, direct comparisons of the results of previouswork6,7 and our results are problematic, since the otherinvestigators may not have purified the circular DNAsand linearized DNAs at the same time.
Very recently, Schakowski et al reported geneexpression from linearized and capped molecules, similarto those used in this work.8 They compared the transgeneexpression of the linearized minimal-size DNAs and thatof the circular DNAs using polyethylenimine and lipofec-tamine, and observed that the linearized DNAs improvedthe amount of the transgene product. However, the per-centage of cells expressing the gene was similar in thecases of circular and linearized DNAs. We also used theratio of cells expressing GFP, and found that the shortestlinearized molecule (2.3 kb) was a much better vector(Figures 4 and 6). The reason for these discrepanciesbetween our present study and that of Schakowski et alis not clear. One possibility could be the differences inthe loop structure. They used a loop consisting of fourthymine bases. In contrast, we selected a highly thermost-able loop to make the terminus rigid, as well as to pre-vent digestion by exonuclease(s). The difference in theterminal structure may affect the stability in cells and/orthe protein-binding. Alternatively, some inhibitory fac-tors may have been present after the chromatographystep which they used for the purification of linearizedvectors.
It should be emphasized that our results were obtainedby microinjections in addition to transfection. Thus, wecould reach the important conclusion that the effects ofthe topology and the size on transgene expression bytransfection were highly dependent on the transcriptionefficiency. Of the DNAs studied, the linearized DNAdumbbell molecule with the minimum size was mosteffective for transgene expression, because it had thehighest transcription. It is well understood that improve-ments of endocytosis and nuclear entry by variousmethods are necessary to enhance transgene expressionefficiency.2–5 The results obtained in this study, togetherwith improved delivery systems, will help to producenonviral vector systems with clinical applications.
Materials and methods
GeneralNIH3T3 cells were from the American Type Culture Col-lection (Manassas, VA, USA). COS-7 cells were from theRIKEN Cell Bank (Tsukuba, Japan). The pQBI 25 and
pQBI 63 plasmids were from Takara (Kusatsu, Japan).Tetramethylrhodamine-labeled dextran (MW 70 000) wasfrom Molecular Probes (Eugene, OR, USA).
OligonucleotidesOligonucleotides for end-capping were designed toincorporate the stable loop-forming sequence, 5’-GCGAAGC-3’, in the middle and to make a 5’-protrudingend compatible with the HindIII, BglII, or Aor13HI restric-tion enzyme: 5’-AGCTTATGGCGAAGCCATA-3’, 5’-GATCTATGGCGAAGCCATA-3’, or 5’-CCGGAATGGCGAAGCCATT-3’, where italicized and underlinedsequences represent sticky end and loop-forming sites,respectively. The oligonucleotides listed above wereobtained from Hokkaido System Science (Sapporo, Japan)in the purified form. These oligonucleotides were 5’-phosphorylated by T4 polynucleotide kinase and ATP,and were annealed before ligation to linearized DNAs.
Construction of plasmids with various lengthsThe DraIII site of the pQBI 25 plasmid was converted toa HindIII site by oligonucleotide linker-mediatedmutagenesis, and an ‘expression cassette’ containing theCMV promoter, the GFP gene, and the bovine growthhormone poly(A) sequence was excised from this modi-fied plasmid by HindIII and BglII digestions. This cassettewas purified by low melting point agarose gel electro-phoresis and was inserted into the pQBI 63 plasmiddigested by the same restriction enzymes. This new plas-mid, designated as pQBI 25-63 (7.6 kb, Figure 1a), wasused as the parental DNA for the construction of otherplasmids. Plasmids with 5.7 and 6.8 kb lengths wereobtained by self-ligation of large blunt-ended BglII–NruIand BglII–MluI fragments of pQBI 25-63, respectively.The insertion of small, blunt-ended BglII–NruI and BglII–MluI fragments into the BglII site of pQBI 25-63 yieldedplasmids with 9.4 and 8.3 kb lengths, respectively. Theseplasmid DNAs were purified with a Qiagen (Valencia,CA, USA) EndoFree Mega kit.
Preparation of linearized DNAsThe pQBI 25-63 plasmid was digested with either BglII,Aor13HI, or HindIII according to the conditions rec-ommended by the suppliers. The digested DNAs (4.3pmol) were mixed and joined with 430 pmol of the corre-sponding phosphorylated oligonucleotides with a Lig-ation High Kit (Toyobo, Osaka, Japan). After ethanol pre-cipitation, the ligated DNA was loaded on to a 1% lowmelting point agarose gel and the electrophoresis wasdone in TAE buffer.27 The gel was placed on to a thin-layer chromatography plate containing fluorescentmaterials. The linearized DNA was recovered from thegel by visualization by UV irradiation from the upperside. The DNA was purified as described in the litera-ture.27 The other linearized molecules were prepared bya similar procedure.
MicroinjectionCells were microinjected at day 1 after plating. In thisprocedure, we used a semi-automatic injection system(Eppendorf transjector 5246, Hamburg, Germany)attached to the Eppendorf micromanipulator 5171.Intranuclear and cytosolic microinjections were perfor-med with the Z (depth) limit option using a 0.2-s injectiontime and a 30–200 hectopascal-injection pressure. The

Effects of DNA structure on transcriptionH Kamiya et al
1507DNA was diluted with an injection buffer solution (0.5%tetramethylrhodamine-labeled dextran in phosphatebuffered saline). At 48 h (unless indicated) after injection,the cells were washed with PBS twice and then werefixed with 25% glutaraldehyde at room temperature for30 min.
TransfectionDNA transfection was carried out with the LipofectaminePlus Reagent (Invitrogen, Carlsbad, CA, USA) accordingto the supplier’s instructions. COS-7 or NIH3T3 cells (3.5–5 × 105 cells) were incubated overnight in DMEM with10% fetal calf serum under a 5% CO2/95% air atmosphereat 37°C. The DNA was mixed with the lipids and wastransfected into the cells. After 3 h, the lipid–DNA com-plex was removed and the cells were incubated inDMEM with 10% fetal calf serum. At 48 h after the trans-fection initiation, the cells were washed with PBS twiceand then were fixed with 25% glutaraldehyde at roomtemperature for 30 min.
Exonuclease treatmentLinearized DNA molecules with and without capping(100 ng) were treated with 0.15 U of BAL 31 nuclease(Takara) at 30°C for 10 min. After the addition of threevolumes of 24 mM EDTA, the treated DNA was precipi-tated with ethanol and was analyzed by 1% agarose gelelectrophoresis.
AcknowledgementsWe thank Dr Hirotake Kitaura of Hokkaido University,for his technical advice. We are grateful to Dr Paul THenderson of MIT for reading the manuscript. This workwas supported in part by Grants-in-Aid for ScientificResearch on Priority Areas from the Ministry of Edu-cation, Culture, Sports, Science, and Technology of Japan.
References1 Rolland AP. From genes to gene medicines: recent advances in
nonviral gene delivery. Crit Rev Ther Drug Carrier Syst 1998; 15:143–198.
2 Gao X, Huang L. Potentiation of cationic liposome-mediatedgene delivery by polycations. Biochemistry 1996; 35: 1027–1036.
3 Simoes S et al. Gene delivery by negatively charged ternary com-plexes of DNA, cationic lipsomes and transferrin or fusigenicpeptides. Gene Therapy 1998; 5: 955–964.
4 Liang E, Hughes J. Characterization of a pH-sensitive surfactant,dodecyl-2-(1’-imidazolyl) propionate (DIP), and preliminarystudies in liposome mediated gene transfer. Biochim Biophys Acta1998; 1369: 39–50.
5 Schoen P et al. Gene transfer mediated by fusion protein hemag-glutinin reconstituted in cationic lipid vesicles. Gene Therapy1999; 6: 823–832.
6 Zanta MA, Belguise-Valladier P. Behr J-P. Gene delivery: a sin-gle nuclear localization signal peptide is sufficient to carry DNAto the cell nucleus. Proc Natl Acad Sci USA 1999; 96: 91–96.
Gene Therapy
7 Kreiss P et al. Plasmid DNA size does not affect the physico-chemical properties of lipoplexes but modulates gene transferefficiency. Nucleic Acids Res 1999; 27: 3792–3798.
8 Schakowski F et al. A novel minimal-size vector (MIDGE)improves transgene expression in colon carcinoma cells andavoids transfection of undesired DNA. Mol Ther 2001; 5: 793–800.
9 Darquet A et al. Minicircle: an improved DNA molecule for invitro and in vivo gene transfer. Gene Therapy 1999; 6: 209–218.
10 Hirao I et al. Stability difference between DNA and RNA mini-hairpins containing two G-C pairs. Nucleic Acids Symp Ser 1992;27: 127–128.
11 Yoshizawa S et al. GNA trinucleotide loop sequences producingextraordinarily stable DNA minihairpins. Biochemistry 1997; 36:4761–4767.
12 Deniss IS, Morgan AR. Studies on the mechanism of DNA cleav-age by ethidium. Nucleic Acids Res 1976; 3: 315–323.
13 Krishnamurthy G, Polte T, Rooney T. Hogan ME A photochem-ical method to map ethidium bromide binding sites on DNA:application to a bent DNA fragment. Biochemistry 1990; 29:981–988.
14 Claycamp HG. Phenol sensitization of DNA to subsequent oxi-dative damage in 8-hydroxyguanine assays. Carcinogenesis 1992;13: 1289–1292.
15 Helbock HJ et al. DNA oxidation matters: the HPLC-electro-chemical detection assay of 8-oxo-deoxyguanosine and 8-oxo-guanine. Proc Natl Acad Sci USA 1998; 95: 288–293.
16 Douki T, Cadet J. Individual determination of the yield of themain UV-induced dimeric pyrimidine photoproducts in DNAsuggests a high mutagenicity of CC photolesions. Biochemistry2001; 40: 2495–2501.
17 Grundemann D, Schomig E. Protection of DNA during prepara-tive agarose gel electrophoresis against damage induced byultraviolet light. Biotechniques 1996; 21: 898–903.
18 Liu J, Doetsch PW. Escherichia coli RNA and DNA polymerasebypass of dihydrouracil: mutagenic potential via transcriptionand replication. Nucleic Acids Res 1998; 26: 1707–1712.
19 Viswanathan A, Doetsch PW. Effects of nonbulky DNA basedamages on Escherichia coli RNA polymerase-mediated elong-ation and promoter clearance. J Biol Chem 1998; 273: 21276–21281.
20 Selby CP, Drapkin R, Reinberg D, Sancar A. DNA excisionrepair. Nucleic Acids Res 1997; 25: 787–793.
21 Sancar A. DNA excision repair. Annu Rev Biochem 1996; 65:43–81.
22 Wallace SS. Enzymatic processing of radiation-induced free rad-ical damage in DNA. Radiat Res 1998; 150: S60–S79.
23 Kamiya H et al. c-Ha-ras containing-hydroxyguanine at codon12 induces point mutations at the modified and adjacent pos-itions. Cancer Res 1992; 52: 3483–3485.
24 Kamiya H et al. 8-Hydroxyguanine (7,8-dihydro-8-oxoguanine)in hot spots of the c-Ha-ras gene. Effects of sequence contextson mutation spectra. Carcinogenesis 1995; 16: 883–889.
25 Kamiya H et al. 8-Hydroxyadenine (7, 8-dihydro-8-oxoadenine)induces misincorporation in in vitro DNA synthesis andmutations in NIH3T3 cells. Nucleic Acids Res 1995; 23: 2893–2899.
26 Kamiya H, Kasai H. Mutations induced by 2-hydroxyadenineon a shuttle vector during leading and lagging strand synthesesin mammalian cells. Biochemistry 1997; 36: 11125–11130.
27 Sambrook J, Fritsch EF, Maniatis T. (1989) Molecular Cloning: ALaboratory Manual, 2nd edn. Cold Spring Harbor LaboratoryPress: Cold Spring Harbor, pp 6.30–6.31.

RETRACTION
Size and topology of exogenous DNA as determinantfactors of transgene transcription in mammalian cells
H Kamiya, J Yamazaki and H HarashimaGraduate School of Pharmaceutical Sciences, Hokkaido University, Sapporo, Japan
Gene Therapy (2006) 13, 932. doi:10.1038/sj.gt.3302795
Reference to: Gene Therapy (2002) 9, 1500–1507.doi:10.1038/sj.gt.3301831
In November 2002, Gene Therapy published an articleby Professor Hiroyuki Kamiya and colleagues, whichdescribed an experiment that sought to constructtopology altered DNA from which a transgene productcould be efficiently produced.
In December 2005, the authors informed us that theyhad been unable to reproduce the results of this studyand would like to have the paper retracted.
Having considered the authors explanation, which ispublished below, we have decided to withdraw this paper.
Statement by corresponding author Hiroyuki Kamiya:
Recently, we added a functional DNA sequence toplasmid DNA and examined its effects on transgeneexpression. At that time, we also linearized the newplasmid DNA, as reported in the Gene Therapy paper. Ascompared with the circular plasmid DNA, the linearizedDNA (DNA dumbbell) did not show the expected,enhanced expression. We checked expression level of theoriginal (without the functional sequence mentionedabove) DNA dumbbell and found that the DNAtopology had no dramatic effect. We constructed theseDNA dumbbells by ligation of oligoDNAs and plasmidDNA digested with restriction enzyme(s). To exclude the
possibility that the efficiency of the intermolecularligation had an influence on the expression, we con-structed another plasmid DNA that enables intramole-cular ligation. However, the DNA dumbbell preparedfrom the new plasmid DNA did not increase theexpression. Thus, we have to conclude that the DNAdumbbell topology did not enhance transgene expression.
The purpose of our research was to construct topologyaltered DNA from which a transgene product can beefficiently produced. Although somewhat similar effectswere observed by us1 and others,2 the effects of thetopology seem to be highly dependent on the experi-mental conditions.
We regret that most of the results reported in our GeneTherapy paper are not reproducible and we would like toapologize for this retraction.
References
1 Tanimoto M, Kamiya H, Minakawa N, Matsuda A, Harashima H.No enhancement of nuclear entry by direct conjugation of anuclear localization signal peptide to linearized DNA. BioconjChem 2003; 14: 1197–1202.
2 van der Aa MA, Koning GA, d’Oliveira C, Oosting RS, WilschutKJ, Hennink WE et al. An NLS peptide covalently linked to linearDNA does not enhance transfection efficiency of cationic polymerbased gene delivery systems. J Gene Med 2005; 7: 208–217.
Gene Therapy (2006) 13, 932& 2006 Nature Publishing Group All rights reserved 0969-7128/06 $30.00
www.nature.com/gt




















