
A trypanosome-soluble factor induces IP3 formation, intracellular Ca2+ mobilization and...
11
A Trypanosome-soluble Factor Induces IPa Formation, Intracellular Ca 2+ Mobilization and Microfilament Rearrangement in Host Cells Ana Rodriguez, Marc G. Rioult, Ari Ora, and Norma W. Andrews Department of Cell Biology, Yale University School of Medicine, New Haven, Connecticut 06510 Abstract. Lysosomes are recruited to the invasion site during host cell entry by Trypanosoma cruzi, an un- usual process suggestive of the triggering of signal transduction mechanisms. Previous studies showed that trypomastigotes, but not the noninfective epimasti- gotes, contain a proteolytically generated trypomasti- gote factor (PGTF) that induces intracellular free Ca 2+ transients in several mammalian cell types. Using con- focal time-lapse imaging of normal rat kidney (NRK) fibroblasts loaded with the Ca2÷-sensitive dye fluo-3, we show that the initial intracellular free Ca 2÷ concen- tration ([Ca2+]i) transient detected a few seconds after exposure to trypomastigote extracts is a result of Ca 2÷ release from intracellular stores. Removal of Ca 2+ from the extracellular medium or inhibition of Ca 2÷ channels with NiC12 did not affect the response to PGTF, while depletion of intracellular stores with thapsigargin abol- ished it. [Ca2+]i transients induced by PGTF were shown to be coupled to the activity of phospholipase C (PLC), since the specific inhibitor U73122 completely blocked the response, while its inactive analogue U73343 had no effect. In addition, polyphosphoinosi- tide hydrolysis and inositol 1,4,5-trisphosphate (IP3) were detected upon cell stimulation with PGTF, sug- gesting the participation of IP3-sensitive intracellular Ca 2÷ channels. An immediate effect of the signaling induced by PGTF and live trypomastigotes was a rapid and tran- sient reorganization of host cell microfilaments. The re- distribution of F-actin appeared to be a direct conse- quence of increased [Ca2+]i, since thrombin and the Ca 2÷ ionophore ionomycin produced a similar effect, with a time course that corresponded to the kinetics of the elevation in [Ca2+]i . These observations support the hypothesis that PGTF-induced disassembly of the corti- cal actin cytoskeleton may play a role in T. cruzi inva- sion, by facilitating lysosome access to the invasion site. Taken together, our findings suggest that the proteolyt- ically generated trypomastigote factor PGTF is a novel agonist that acts through the PLC/phosphoinositide sig- naling pathway of mammalian cells. I NTRACELLULAR pathogens show remarkable diversity in their mechanisms for invading mammalian cells. Re- cent evidence indicates that host cell invasion is in many cases preceded by the transduction of specific sig- nals across the plasma membrane. Bacterial pathogens such as enteropathogenic Yersinia, Salmonella, and Es- cherichia coli induce responses in their host cells that are believed to facilitate invasion through distinct pathways (Bliska et al., 1993). Although the host cell entry mecha- nism of most intracellular pathogens resembles phago- cytosis, in the sense that it involves F-actin-dependent plasma membrane extensions, exceptions to this rule have been found. One such exception is the intracellular proto- zoan parasite Trypanosoma cruzi, the causative agent of Chagas' disease in man. Address all correspondence to Norma W. Andrews, Dept. of Cell Biology, Yale University School of Medicine, 333 Cedar St., New Haven, CT 06510. Tel.: (203) 785-4313. Fax: (203) 785-7226. Invasion of nonphagocytic vertebrate cells by trypomas- tigotes, the infective stages of T. cruzi, proceeds by a unique mechanism that is independent of pseudopodia formation and actin polymerization (Schenkman et al., 1988; Tardieux et al., 1992). Disruption of the actin cyto- skeleton with cytochalasin D enhances trypanosome entry, indicating that the mechanism of invasion is distinct from phagocytosis (Schenkman et al., 1991; Tardieux et al., 1992). In addition, an unusual process of recruitment and fusion of host lysosomes occurs directly beneath the inva- sion site, and this process is required for parasite internal- ization (Tardieux et al., 1992). These observations suggest that cortical actin rearrangements may be an early step in the T. cruzi invasion mechanism, to allow lysosomes easier access to the plasma membrane. In searching for second messengers involved in the lyso- some recruitment mechanism, it was found that T. cruzi trypomastigotes, but not the noninfective epimastigotes, induce repetitive, pertussis toxin-sensitive, transient eleva- © The Rockefeller University Press, 0021-9525/95/06/1263/11 $2.00 The Journal of Cell Biology, Volume 129, Number 5, June 1995 1263-1273 1263 on January 20, 2015 jcb.rupress.org Downloaded from Published June 1, 1995
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A trypanosome-soluble factor induces IP3 formation, intracellular Ca2+ mobilization and...

A Trypanosome-soluble Factor Induces IPa Formation, Intracellular C a 2+
Mobilization and Microfilament Rearrangement in Host Cells Ana Rodriguez, Marc G. Rioult, Ari Ora, and Norma W. Andrews
Department of Cell Biology, Yale University School of Medicine, New Haven, Connecticut 06510
Abstract. Lysosomes are recruited to the invasion site during host cell entry by Trypanosoma cruzi, an un- usual process suggestive of the triggering of signal transduction mechanisms. Previous studies showed that trypomastigotes, but not the noninfective epimasti- gotes, contain a proteolytically generated trypomasti- gote factor (PGTF) that induces intracellular free Ca 2+ transients in several mammalian cell types. Using con- focal time-lapse imaging of normal rat kidney (NRK) fibroblasts loaded with the Ca2÷-sensitive dye fluo-3, we show that the initial intracellular free Ca 2÷ concen- tration ([Ca2+]i) transient detected a few seconds after exposure to trypomastigote extracts is a result of Ca 2÷ release from intracellular stores. Removal of Ca 2+ from the extracellular medium or inhibition of Ca 2÷ channels with NiC12 did not affect the response to PGTF, while depletion of intracellular stores with thapsigargin abol- ished it. [Ca2+]i transients induced by PGTF were shown to be coupled to the activity of phospholipase C (PLC), since the specific inhibitor U73122 completely blocked the response, while its inactive analogue
U73343 had no effect. In addition, polyphosphoinosi- tide hydrolysis and inositol 1,4,5-trisphosphate (IP3) were detected upon cell stimulation with PGTF, sug- gesting the participation of IP3-sensitive intracellular Ca 2÷ channels.
An immediate effect of the signaling induced by PGTF and live trypomastigotes was a rapid and tran- sient reorganization of host cell microfilaments. The re- distribution of F-actin appeared to be a direct conse- quence of increased [Ca2+]i, since thrombin and the Ca 2÷ ionophore ionomycin produced a similar effect, with a time course that corresponded to the kinetics of the elevation in [Ca2+]i . These observations support the hypothesis that PGTF-induced disassembly of the corti- cal actin cytoskeleton may play a role in T. cruzi inva- sion, by facilitating lysosome access to the invasion site. Taken together, our findings suggest that the proteolyt- ically generated trypomastigote factor PGTF is a novel agonist that acts through the PLC/phosphoinositide sig- naling pathway of mammalian cells.
I NTRACELLULAR pathogens show remarkable diversity in their mechanisms for invading mammalian cells. Re- cent evidence indicates that host cell invasion is in
many cases preceded by the transduction of specific sig- nals across the plasma membrane. Bacterial pathogens such as enteropathogenic Yersinia, Salmonella, and Es- cherichia coli induce responses in their host cells that are believed to facilitate invasion through distinct pathways (Bliska et al., 1993). Although the host cell entry mecha- nism of most intracellular pathogens resembles phago- cytosis, in the sense that it involves F-actin-dependent plasma membrane extensions, exceptions to this rule have been found. One such exception is the intracellular proto- zoan parasite Trypanosoma cruzi, the causative agent of Chagas' disease in man.
Address all correspondence to Norma W. Andrews, Dept. of Cell Biology, Yale University School of Medicine, 333 Cedar St., New Haven, CT 06510. Tel.: (203) 785-4313. Fax: (203) 785-7226.
Invasion of nonphagocytic vertebrate cells by trypomas- tigotes, the infective stages of T. cruzi, proceeds by a unique mechanism that is independent of pseudopodia formation and actin polymerization (Schenkman et al., 1988; Tardieux et al., 1992). Disruption of the actin cyto- skeleton with cytochalasin D enhances trypanosome entry, indicating that the mechanism of invasion is distinct from phagocytosis (Schenkman et al., 1991; Tardieux et al., 1992). In addition, an unusual process of recruitment and fusion of host lysosomes occurs directly beneath the inva- sion site, and this process is required for parasite internal- ization (Tardieux et al., 1992). These observations suggest that cortical actin rearrangements may be an early step in the T. cruzi invasion mechanism, to allow lysosomes easier access to the plasma membrane.
In searching for second messengers involved in the lyso- some recruitment mechanism, it was found that T. cruzi trypomastigotes, but not the noninfective epimastigotes, induce repetitive, pertussis toxin-sensitive, transient eleva-
© The Rockefeller University Press, 0021-9525/95/06/1263/11 $2.00 The Journal of Cell Biology, Volume 129, Number 5, June 1995 1263-1273 1263
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995

tions in the cytosolic free C a 2+ concentration ( [Ca2+]i) 1 of normal rat kidney (NRK) cells (Tardieux et al., 1994). A soluble factor (PGTF) that induces Ca z+ signaling in sev- eral different mammalian cell types was subsequently ex- tracted from trypomastigotes, and found to be dependent on the activity of a leupeptin-sensitive parasite alkaline peptidase (Burleigh and Andrews, 1995). Inhibition of the host cell [Ca2+]i transients, either by interfering with intra- cellular free Ca 2÷ levels or by inhibiting the trypanosome peptidase, decreased parasite entry, thereby suggesting a role for the PGTF-mediated signaling event in invasion (Tardieux et al., 1994; Burleigh and Andrews, 1995).
Signal transduction events mediated by hormones and growth factors are often accompanied by hydrolysis of polyphosphoinositides, transient increases in [Ca2+]i and rapid actin cytoskeletal rearrangements (Chinkers et al., 1979; Kadowaki et al., 1986; Bretscher, 1991; Janmey, 1994). Ca 2÷ signaling has also been shown to be associated with phagosome-lysosome fusion (Jaconi et al., 1990). The T. cruzi-induced Ca 2÷ signal was therefore proposed as playing a role in invasion through reorganization of host cell microfilaments and/or lysosome fusion (Tardieux et al., 1994). In this work we characterize the [Ca2+]i tran- sients induced by the trypomastigote-soluble fraction con- taining the signaling factor PGTF (Burleigh and Andrews, 1995). We show that the initial [Ca2÷]i elevation induced by PGTF in NRK cells is mediated by phospholipase C (PLC), inositol 1,4,5-trisphosphate (IP3) formation, and Ca 2+ release from intraceUular stores. The resulting in- crease in [Ca2+]i induces a transient reorganization of actin microfilaments, in a pattern consistent with a role in facili- tating parasite invasion.
Materials and Methods
Materials
EGTA, leupeptin, thrombin, genistein, staurosporine, thapsigargin, and phalloidin were obtained from Sigma Immunochemical (St. Louis, MO), ionomycin from Calbiochem Novabiochem (La Jolla, CA), and H-7, U73122 and U73343 from BIOMOL Res. Labs., Inc, (Plymouth Meeting, PA). Fluo-3-AM was purchased from Molecular Probes, Inc. (Eugene, OR).
Cells and Parasites
NRK fibroblasts were grown in DMEM containing 10% FBS, at 37°C with 5% CO2. Trypomastigotes from the T. cruzi Y strain were obtained from the supernatant of infected LLC-MK2 cells (Andrews et al., 1987). Epi- mastigotes from the Y strain were cultured in liver infection tryptose me- dium containing 10% FBS at 28°C (Nogueira and Cohn, 1976). Parasites were washed in Hepes-buffered Ringer's solution (Heuser, 1989) and re- suspended in the same solution before exposure to NRK cell monolayers.
Trypanosome Soluble Fraction Trypomastigotes or epimastigotes were washed twice and resuspended in Dulbecco's phosphate-buffered saline containing Ca/+ and Mg 2+ (PBS 2+) at 2 x 108/ml. Parasites were killed by heating at 56°C for 5 min, then fro- zen at -80°C. Trypanosomes were thawed, sonicated on ice with a mi- crotip for two bursts of 15 s at a setting of 2 (Model 250 sonicator; Branson
1. Abbreviations used in this paper. [Ca/+]i , intracellular free Ca 2+ concen- tration; ESF, epimastigote-soluble fraction; IP, inositol monophosphate; IP2, inositol diphosphate; IP3, inositol 1,4,5-trisphosphate; NRK, normal rat kidney; PLC, phospholipase C; PGTF, proteolytically generated trypo- mastigote factor; PKC, protein kinase C; TSF, trypomastigote-soluble fraction.
Ultrasonics Corp., Danbury, CT), and centrifuged at 700 g for 10 rain at 4°C. The postnuclear supernatant was further centrifuged at 100,000 g for 30 min at 4°C and the resulting supernatant fraction was passed through a 1 ml Concanavalin-A-Sepharose column (Pharmacia Chemicals, Uppsala, Sweden). The unbound fraction containing PGTF was collected and stored at -80°C (Burleigh and Andrews, 1995).
Confocal Time-lapse Fluorescence Imaging NRK cells were plated on glass coverslips at 2 × 104 cells/cm 2, cultured for 24 h and loaded with 5 ~M fluo-3-AM (1% DMSO 0.2% pluronic F-127; Molecular Probes) for 45 min at 37°C. Coverslips were mounted in a per- fusion chamber (Warner Instruments, New Haven, CT) and superfused with PBS 2+ or Ca 2÷ free PBS containing 10 mM EGTA at 37°C. Time- lapse images were collected with a confocal scanning laser microscope (EX = 490 nm, EM = 510 nm) (MRC-500; Bio-Rad Labs., Richmond, CA) and recorded on an optical memory disc recorder (TQ-3031F; Pana- sonic, Secaucus, NJ) at 1.8 frames/s (scan time = 0.8 s, download time = 1.0 s) with no averaging. Kinetic plots were obtained from fluorescence values derived by averaging over a 5 × 5 pixel area in a series of images. 10 cells, showing a representative response profile for the whole popula- tion visualized (,'-~30 cells), were analyzed in each experiment.
Polyphosphoinositide Hydrolysis Analysis NRK cells were plated at 1.5 × 104 cells/cm 2 in 12-well dishes and grown for 24 h to subconfluence. Wells were then rinsed twice with 10 mM Hepes-buffered DMEM and incubated in 0.5 ml of this medium contain- ing 2 ixCi/ml [3H] myo-inositol (105 Ci/mmol; Arnersham Corp., Arlington Heights, IL) for 20 h at 37°C. Cells were then treated with 20 mM LiC1 for 1 min before the addition of agonist solution in the presence of 20 mM LiCk After agonist treatment, cells were extracted with 750 I~1 of 20 mM formic acid for 30 min at 4°C. 1-ml packed/volume columns of anion- exchange resin AGIX8, formate form, 100-200 mesh size (Bio Rad Labs., Hercules, CA) were used to bind inositol phosphates. Columns were se- quentially washed with 2 ml of 2 M ammonium formate/0.1 M formic acid, 2 ml of water and 4 ml of 20 mM ammonium hydroxide. Cell extracts were then loaded and the column immediately washed with 3 ml of 40 mM am- monium hydroxide followed by 4 ml of 40 mM ammonium formate. Inosi- tol phosphates were eluted with 4 ml of 2 M ammonium formate/0.1 M formic acid and the radioactivity determined in a scintillation counter (Paris and Pouyssegur, 1986).
HPLC Fractionation of lnositol Phosphates NRK cells were labeled as described above, except that the [3H]myo-ino- sitol concentration was 5 p, Ci/ml and 60-ram dishes were used to grow the cells. Cells were treated with agonist for 15 min as described above and the reaction was terminated by removing the medium and adding boiling distilled water to the cells. Cell lysates were collected, boiled for 5 min, and centrifuged to remove particulate material. The soluble supernatant from two 60-mm dishes was pooled and the volume concentrated to 20 p~l. Samples were injected into a SAX hydropore anion-exchange analytical column (4.6 × 100 mm; Rainin Instrument Co. Inc., Woburn, MA). The flow rate was set at 1 ml/min and 0.8-ml fractions were collected. Inositol was eluted with water for 15 min. Inositol phosphate species were eluted in the order of inositol monophosphate (IP), inositol diphosphate (IP2), IP3 by a 5-min linear gradient of 0-10% 1 M ammonium formate, pH 3.8, followed by 20-min isocratic elution with 10% 1 M ammonium formate. Elution then continued with a 5-rain linear gradient of 10-100% 1 M am- monium formate followed by a 20-min isocratic elution with 1 M ammo- nium formate. The following standards were run in parallel to identify the eluted species: [3H]myo-inositol (105 Ci/mmol; Amersham), [3H]inositol- 1-phosphate, and [3H]inositol 1,4,5-trisphosphate (21 Ci/mmol; Dupont- New England Nuclear, Boston, MA).
F-actin Imaging NRK cells were plated on glass coverslips at 2 × 104 cells/cm 2 and culti- vated for 24 h. Coverslips were washed with PBS ~+ and treated with ago- nist for the indicated periods of time. Fixation was performed with ace- tone at -20°C for 5 min. After fixation, coverslips were washed in TBS and incubated with 1 ixg/ml FITC-labeled phalloidin (Sigma) in TBS con- taining 1% BSA for i h. Preparations were visualized using a fluorescence microscope (Zeiss Axiovert 135; Carl Zeiss, Inc., Thornwood, NY). Line- scan analysis of F-actin was performed on digital images recorded with a
The Journal of Cell Biology, Volume 129, 1995 1264
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995
videocamera (CCD-C72; Dage-MTl, Inc., Michigan City, IN), measuring the fluorescence brightness across a straight line drawn from the cell mar- gin through the nucleus, using MetaMorph software (Universal Imaging Corp., Westchester, PA).
Cell Invasion Assay
Trypomastigotes were washed in Hepes-buffered Ringer's solution (Heuser, 1989) and resuspended in the same solution at 5 × 107/ml. Cov- erslips with NRK cells plated at 2 X 104 cells/cm 2 24 h before, were pre- treated with the indicated drugs for 15 or 30 min, washed and incubated with trypomastigotes for 20 min at 37°C. Three coverslips for each experi- ment were separately washed in cold PBS 2+, fixed in 2% paraformalde- hyde (PFA) for 20 min and incubated for 15 min with 500 mM NH4C1 to quench remaining aldehyde groups of the fixative. The coverslips were then incubated with 10 p~g/ml IgG from a rabbit immunized with T. cruzi trypomastigotes. After washing, rhodamine-conjugated anti-rabbit IgG antibodies (Boehringer Mannheim Corp., Indianapolis, IN) were added for 30 min in TBS containing 1% BSA, followed by washing and staining of host cell and parasite DNA with DAPI (Sigma) diluted to 10 p,g/ml. DAPI-positive parasites with negative or faint immunolabeling were counted as intracellular, whereas parasites strongly labeled with anti-T. cruzi antibodies were considered extracellular and not counted.
Results
Trypomastigote-induced [ Ca2+]i Transients Are 2+ Mediated by Ca Release from IntraceUular Stores
Trypomastigotes induce rapid and repetitive [Ca2+]i tran- sients in host cells (Tardieux et al., 1994). This activity is associated with the soluble fraction of sonicated trypomas- tigotes and is mediated by a novel Ca2+-signaling factor (PGTF) whose generation requires a T. cruzi leupeptin- sensitive alkaline peptidase (Burleigh and Andrews, 1995).
C o n t r o l a
I O0
"~ 0 , ,
g N
0 100 200
Ca 2+ free b
• " d
0 100 200 300
Time (s)
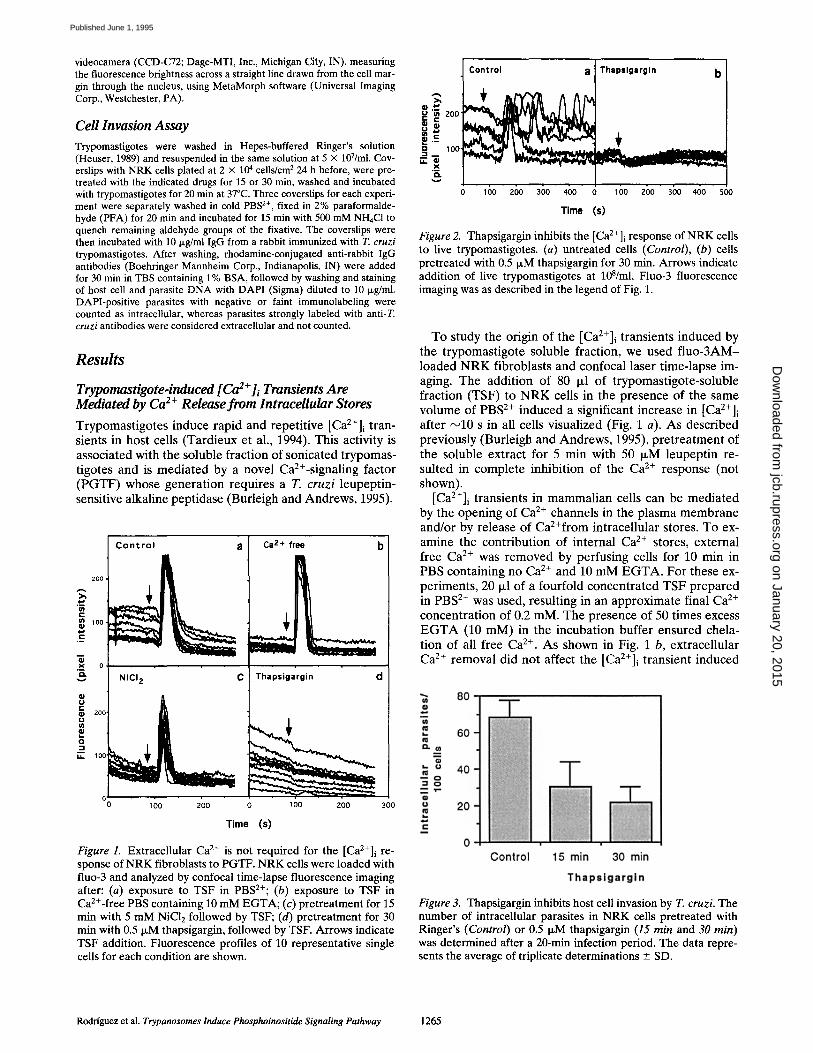
Figure 1. Extracellular Ca 2+ is not required for the [Ca2+]i re- sponse of NRK fibroblasts to PGTF. NRK cells were loaded with fluo-3 and analyzed by confocal time-lapse fluorescence imaging after: (a) exposure to TSF in PBS2+; (b) exposure to TSF in Ca2+-free PBS containing 10 mM EGTA; (c) pretreatment for 15 min with 5 mM NiC12 followed by TSF; (d) pretreatment for 30 min with 0.5 IxM thapsigargin, followed by TSF. Arrows indicate TSF addition. Fluorescence profiles of 10 representative single cells for each condition are shown.
Control a Thapslgargin b
e, '~ 200
=0_ loo-
~.~_
100 zoo 300 400 100 200 300 400 soo
T i m e (s )
Figure 2. Thapsigargin inhibits the [Ca2+]i response of NRK cells to live trypomastigotes. (a) untreated cells (Control), (b) cells pretreated with 0.5 ~M thapsigargin for 30 min. Arrows indicate addition of live trypomastigotes at 108/ml. Fluo-3 fluorescence imaging was as described in the legend of Fig. 1.
To study the origin of the [Ca2+]i transients induced by the trypomastigote soluble fraction, we used f luo-3AM- loaded N R K fibroblasts and confocal laser time-lapse im- aging. The addition of 80 Ixl of trypomastigote-soluble fraction (TSF) to N R K cells in the presence of the same volume of PBS 2÷ induced a significant increase in [Ca2+]i after ~10 s in all cells visualized (Fig. 1 a). As described previously (Burleigh and Andrews, 1995), pretreatment of the soluble extract for 5 min with 50 IxM leupeptin re- suited in complete inhibition of the Ca 2+ response (not shown).
[Ca2+]i transients in mammalian cells can be mediated by the opening of Ca 2÷ channels in the plasma membrane and/or by release of CaZ+from intracellular stores. To ex- amine the contribution of internal Ca 2+ stores, external free Ca e+ was removed by perfusing cells for 10 min in PBS containing no Ca 2÷ and 10 mM E G T A . For these ex- periments, 20 I~1 of a fourfold concentrated TSF prepared in PBS 2÷ was used, resulting in an approximate final Ca 2÷ concentration of 0.2 mM. The presence of 50 times excess E G T A (10 mM) in the incubation buffer ensured chela- tion of all free Ca 2÷. As shown in Fig. 1 b, extracellular Ca 2÷ removal did not affect the [Ca2+]i transient induced
Figure 3. Thapsigargin inhibits host cell invasion by T. cruzi. The number of intracellular parasites in NRK cells pretreated with Ringer's (Control) or 0.5 t~M thapsigargin (15 min and 30 min) was determined after a 20-min infection period. The data repre- sents the average of triplicate determinations _ SD.
Rodriguez et al. Trypanosomes Induce Phosphoinositide Signaling Pathway 1265
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995
b
120
tO0
,~ ~ 80
-6 o 40
N 2o
0 o
c 400.000 "
300.000
200.000
= TSF Thrombin
2 4 6 8 10 12 14 16 Time (min)
Inositol ¢. TSF TSF + leupeptin
E ¢X u
200-~ IP3
IP 1
01 0 2; 40 60
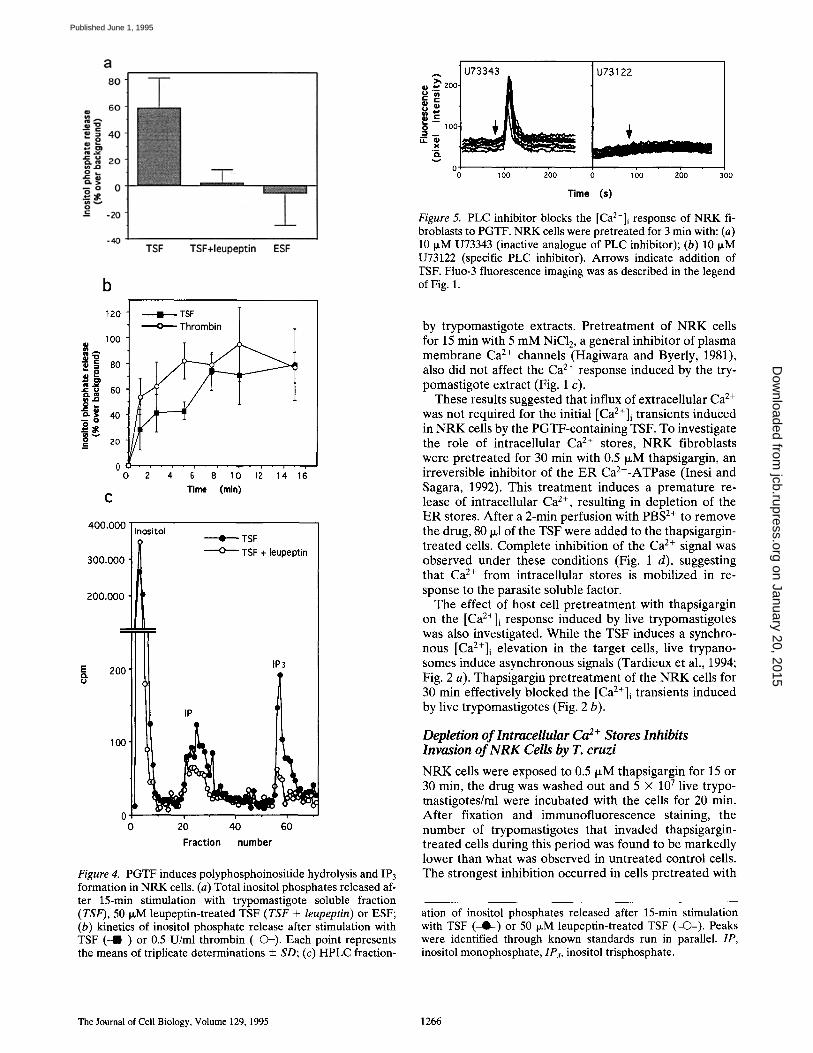
Fraction number Figure 4. PGTF induces polyphosphoinositide hydrolysis and IP3 formation in NRK cells. (a) Total inositol phosphates released af- ter 15-min stimulation with trypomastigote soluble fraction (TSF), 50 I~M leupeptin-treated TSF (TSF + leupeptin) or ESF; (b) kinetics of inositol phosphate release after stimulation with TSF ( -1 - ) or 0.5 U/ml thrombin (-(3-). Each point represents the means of triplicate determinations +-- SD; (c) HPLC fraction-
200. ~'- g ~ 100. m
0
~ ~ U 7 3 3 4 3 !U73122
1 O0 200 300
Time (s) Figure 5. PLC inhibitor blocks the [Ca2+]i response of NRK fi- broblasts to PGTF. NRK cells were pretreated for 3 min with: (a) 10 IxM U73343 (inactive analogue of PLC inhibitor); (b) 10 IxM U73122 (specific PLC inhibitor). Arrows indicate addition of TSF. Fluo-3 fluorescence imaging was as described in the legend of Fig. 1.
by trypomastigote extracts. Pretreatment of NRK cells for 15 min with 5 mM NiC12, a general inhibitor of plasma membrane Ca 2÷ channels (Hagiwara and Byerly, 1981), also did not affect the Ca 2÷ response induced by the try- pomastigote extract (Fig. 1 c).
These results suggested that influx of extracellular Ca 2÷ was not required for the initial [Ca2÷]i transients induced in NRK cells by the PGTF-containing TSF. To investigate the role of intracellular Ca 2÷ stores, N R K fibroblasts were pretreated for 30 min with 0.5 p~M thapsigargin, an irreversible inhibitor of the E R Ca2÷-ATPase (Inesi and Sagara, 1992). This treatment induces a premature re- lease of intracellular Ca 2÷, resulting in depletion of the E R stores. After a 2-min perfusion with PBS 2÷ to remove the drug, 80 ixl of the TSF were added to the thapsigargin- treated cells. Complete inhibition of the Ca 2÷ signal was observed under these conditions (Fig. 1 d), suggesting that Ca 2÷ from intracellular stores is mobilized in re- sponse to the parasite soluble factor.
The effect of host cell pretreatment with thapsigargin on t h e [Ca2+]i response induced by live trypomastigotes was also investigated. While the TSF induces a synchro- nous [Ca2+]i elevation in the target cells, live trypano- somes induce asynchronous signals (Tardieux et al., 1994; Fig. 2 a). Thapsigargin pretreatment of the NRK cells for 30 min effectively blocked the [Ca2+]i t r a n s i e n t s induced by live trypomastigotes (Fig. 2 b).
Depletion of lntraceUular Ca 2+ Stores Inhibits Invasion of N R K Cells by T. cruzi
N R K cells were exposed to 0.5 IxM thapsigargin for 15 or 30 min, the drug was washed out and 5 × 10 7 live trypo- mastigotes/ml were incubated with the cells for 20 min. After fixation and immunofluorescence staining, the number of trypomastigotes that invaded thapsigargin- treated cells during this period was found to be markedly lower than what was observed in untreated control cells. The strongest inhibition occurred in cells pretreated with
ation of inositol phosphates released after 15-min stimulation with TSF (-Q--) or 50 I~M leupeptin-treated TSF (--(3--). Peaks were identified through known standards run in parallel. IP, inositol monophosphate, 1P~, inositol trisphosphate.
The Joumal of Cell Biology, Volume 129, 1995 1266
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995
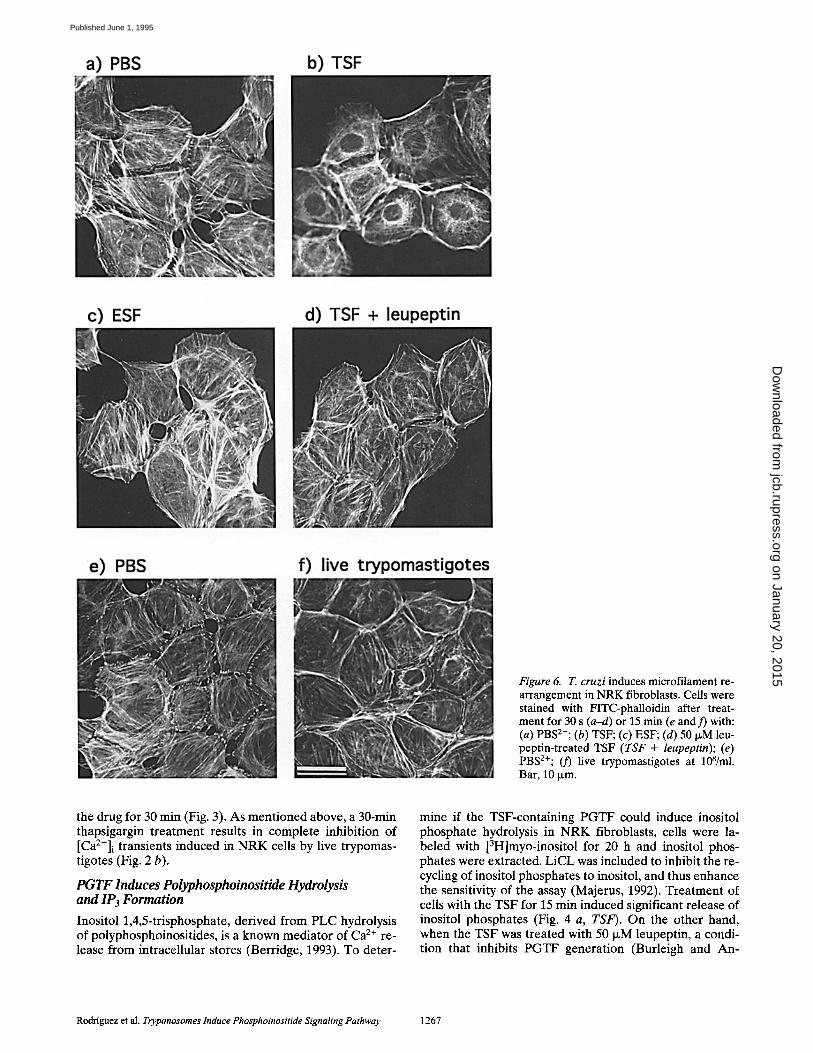
Figure 6. T. cruzi induces microfilament re- arrangement in NRK fibroblasts. Cells were stained with FITC-phalloidin after treat- ment for 30 s (a-d) or 15 min (e and 3') with: (a) PBS2÷; (b) TSF; (c) ESF; (d) 50 I~M leu- peptin-treated TSF (TSF + leupeptin); (e) PBS2+; (3') live trypomastigotes at 108/ml. Bar, 10 ~m.
the drug for 30 min (Fig. 3). As mentioned above, a 30-min thapsigargin treatment results in complete inhibition of [Ca2+]i transients induced in NRK ceils by live trypomas- tigotes (Fig. 2 b).
PGTF Induces Polyphosphoinositide Hydrolysis and IP3 Formation
Inositol 1,4,5-trisphosphate, derived from PLC hydrolysis of polyphosphoinositides, is a known mediator of Ca z+ re- lease from intracellular stores (Berridge, 1993). To deter-
mine if the TSF-containing PGTF could induce inositol phosphate hydrolysis in NRK fibroblasts, cells were la- beled with [3H]myo-inositol for 20 h and inositol phos- phates were extracted. LiCL was included to inhibit the re- cycling of inositol phosphates to inositol, and thus enhance the sensitivity of the assay (Majerus, 1992). Treatment of ceils with the TSF for 15 min induced significant release of inositol phosphates (Fig. 4 a, TSF). On the other hand, when the TSF was treated with 50 p~M leupeptin, a condi- tion that inhibits PGTF generation (Burleigh and An-
Rodriguez et al. Trypanosomes lnduce Phosphoinositide Signaling Pathway 1267
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995
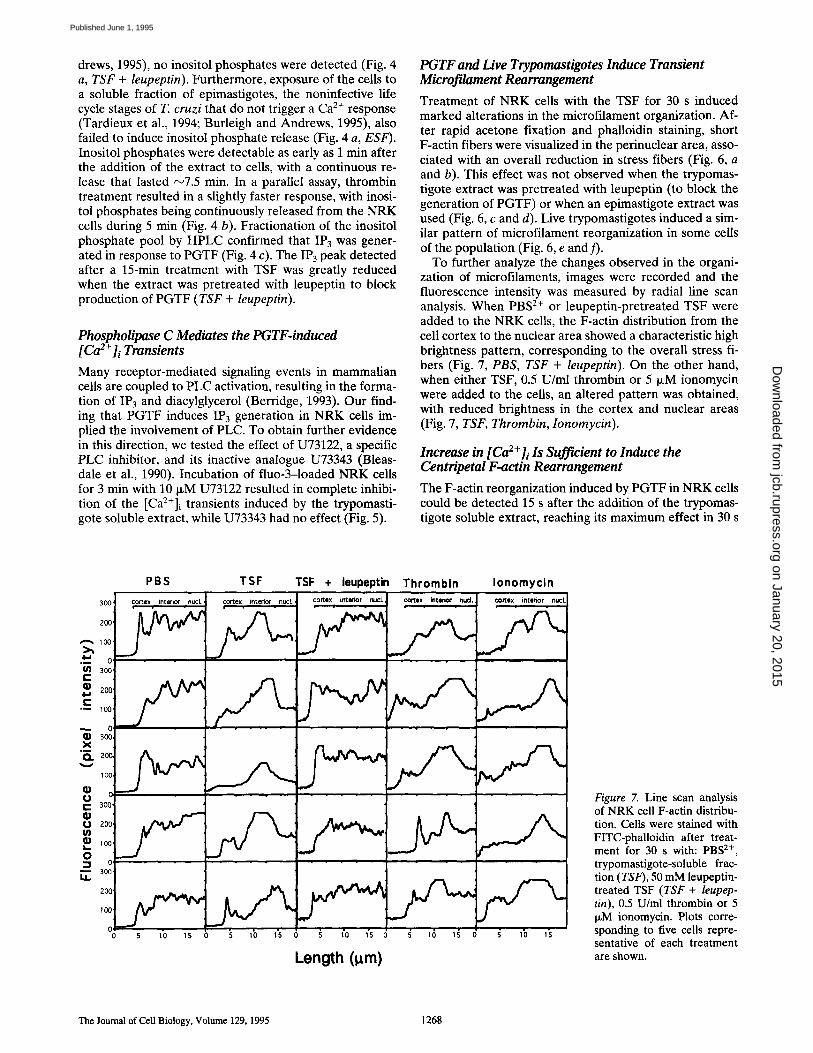
drews, 1995), no inositol phosphates were detected (Fig. 4 a, TSF + leupeptin). Furthermore, exposure of the cells to a soluble fraction of epimastigotes, the noninfective life cycle stages of T. cruzi that do not trigger a Ca 2+ response (Tardieux et al., 1994; Burleigh and Andrews, 1995), also failed to induce inositol phosphate release (Fig. 4 a, ESF). Inositol phosphates were detectable as early as 1 min after the addition of the extract to cells, with a continuous re- lease that lasted ~7.5 min. In a parallel assay, thrombin treatment resulted in a slightly faster response, with inosi- tol phosphates being continuously released from the NRK cells during 5 min (Fig. 4 b). Fractionation of the inositol phosphate pool by HPLC confirmed that IP3 was gener- ated in response to PGTF (Fig. 4 c). The IP3 peak detected after a 15-min treatment with TSF was greatly reduced when the extract was pretreated with leupeptin to block production of PGTF (TSF + leupeptin).
Phospholipase C Mediates the PGTF-induced [ Ca2 + ] i Transients
Many receptor-mediated signaling events in mammalian cells are coupled to PLC activation, resulting in the forma- tion of IP3 and diacylglycerol (Berridge, 1993). Our find- ing that PGTF induces IP3 generation in NRK cells im- plied the involvement of PLC. To obtain further evidence in this direction, we tested the effect of U73122, a specific PLC inhibitor, and its inactive analogue U73343 (Bleas- dale et al., 1990). Incubation of fluo-3-1oaded NRK cells for 3 min with 10 p~M U73122 resulted in complete inhibi- tion of the [ C a 2 + ] i transients induced by the trypomasti- gote soluble extract, while U73343 had no effect (Fig. 5).
PGTF and Live Trypomastigotes Induce Transient Microfilament Rearrangement
Treatment of NRK cells with the TSF for 30 s induced marked alterations in the microfilament organization. Af- ter rapid acetone fixation and phalloidin staining, short F-actin fibers were visualized in the perinuclear area, asso- ciated with an overall reduction in stress fibers (Fig. 6, a and b). This effect was not observed when the trypomas- tigote extract was pretreated with leupeptin (to block the generation of PGTF) or when an epimastigote extract was used (Fig. 6, c and d). Live trypomastigotes induced a sim- ilar pattern of microfilament reorganization in some ceils of the population (Fig. 6, e and f).
To further analyze the changes observed in the organi- zation of microfilaments, images were recorded and the fluorescence intensity was measured by radial line scan analysis. When PBS 2+ or leupeptin-pretreated TSF were added to the NRK cells, the F-actin distribution from the cell cortex to the nuclear area showed a characteristic high brightness pattern, corresponding to the overall stress fi- bers (Fig. 7, PBS, TSF + leupeptin). On the other hand, when either TSF, 0.5 U/ml thrombin or 5 p~M ionomycin were added to the cells, an altered pattern was obtained, with reduced brightness in the cortex and nuclear areas (Fig. 7, TSF, Thrombin, Ionomycin).
Increase in [ Ca2+]i Is Sufficient to Induce the Centripetal F-actin Rearrangement
The F-actin reorganization induced by PGTF in NRK cells could be detected 15 s after the addition of the trypomas- tigote soluble extract, reaching its maximum effect in 30 s
PBS TSF TSF + leupeptin Thrombin Ionomycin 300 cortex interior nucl. cortex interior nucl,
~ ' ~ 100
.~_ o ~ 300
"-- I00
.~ o X
tOO Q;
o
C 3oo
200 f ,oo 0
1~ 3oo i1
200
C 0 5 10 15
cortex interior nucl. c~'tex interior nuct. cortex interior nucl
5 10 1"5 0 s 10 15 0 g 16 15 5 10 1"5
Length (l~m)
Figure 7. Line scan analysis of NRK cell F-actin distribu- tion. Cells were stained with FITC-phalloidin after treat- ment for 30 s with: PBS 2+, trypomastigote-soluble frac- tion (TSF), 50 mM leupeptin- treated TSF (TSF + leupep- tin), 0.5 U/ml thrombin or 5 p~M ionomycin. Plots corre- sponding to five cells repre- sentative of each treatment are shown.
The Journal o f Cell B io logy , Vo lume 129, 1995 1268
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995
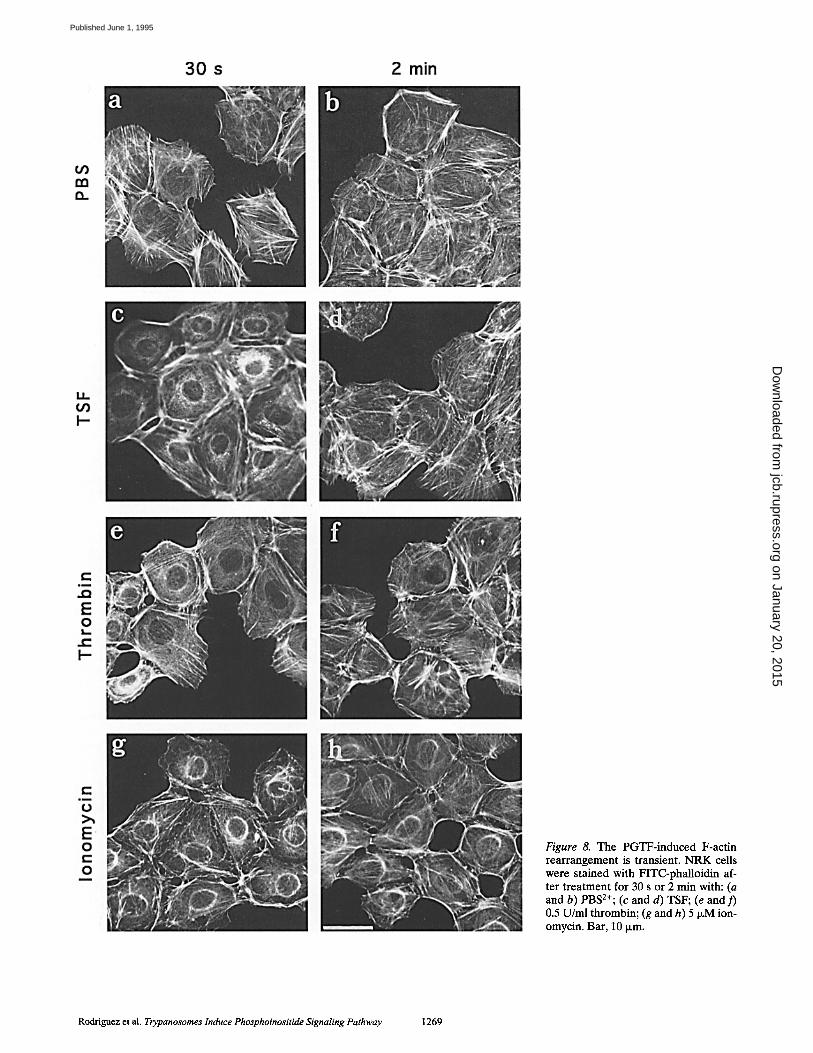
Figure 8. The PGTF-induced F-actin rearrangement is transient. NRK ceils were stained with FITC-phalloidin af- ter treatment for 30 s or 2 min with: (a and b) PBS2+; (c and d) TSF; (e and f) 0.5 U/ml thrombin; (g and h) 5 ~M ion- omycin. Bar, 10 Ixm.
Rodriguez et al. Trypanosomes Induce Phosphoinositide Signaling Pathway 1269
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995
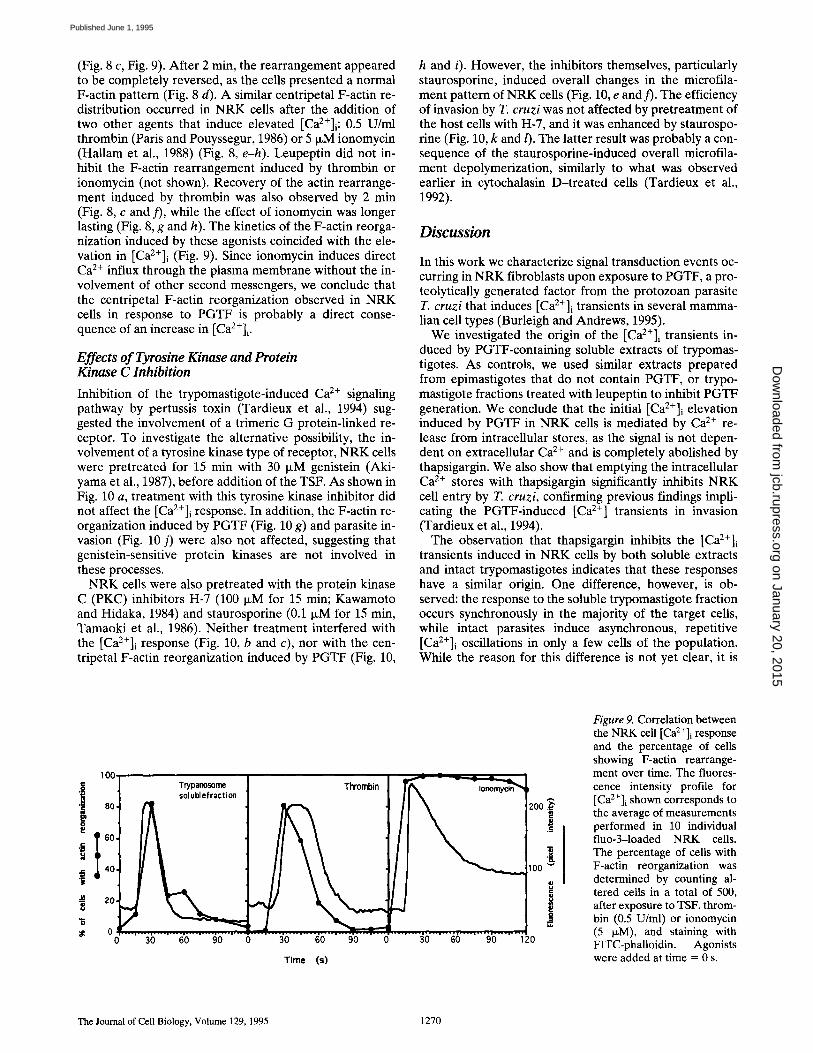
(Fig. 8 c, Fig. 9). After 2 min, the rearrangement appeared to be completely reversed, as the cells presented a normal F-actin pattern (Fig. 8 d). A similar centripetal F-actin re- distribution occurred in NRK cells after the addition of two other agents that induce elevated [Ca2+]i: 0.5 U/ml thrombin (Paris and Pouyssegur, 1986) or 5 IxM ionomycin (Hallam et al., 1988) (Fig. 8, e-h). Leupeptin did not in- hibit the F-actin rearrangement induced by thrombin or ionomycin (not shown). Recovery of the actin rearrange- ment induced by thrombin was also observed by 2 min (Fig. 8, c and )'), while the effect of ionomycin was longer lasting (Fig. 8, g and h). The kinetics of the F-actin reorga- nization induced by these agonists coincided with the ele- vation in [Ca2+]i (Fig. 9). Since ionomycin induces direct Ca 2+ influx through the plasma membrane without the in- volvement of other second messengers, we conclude that the centripetal F-actin reorganization observed in NRK cells in response to PGTF is probably a direct conse- quence of an increase in [Ca2+]i .
Effects of Tyrosine Kinase and Protein Kinase C Inhibition
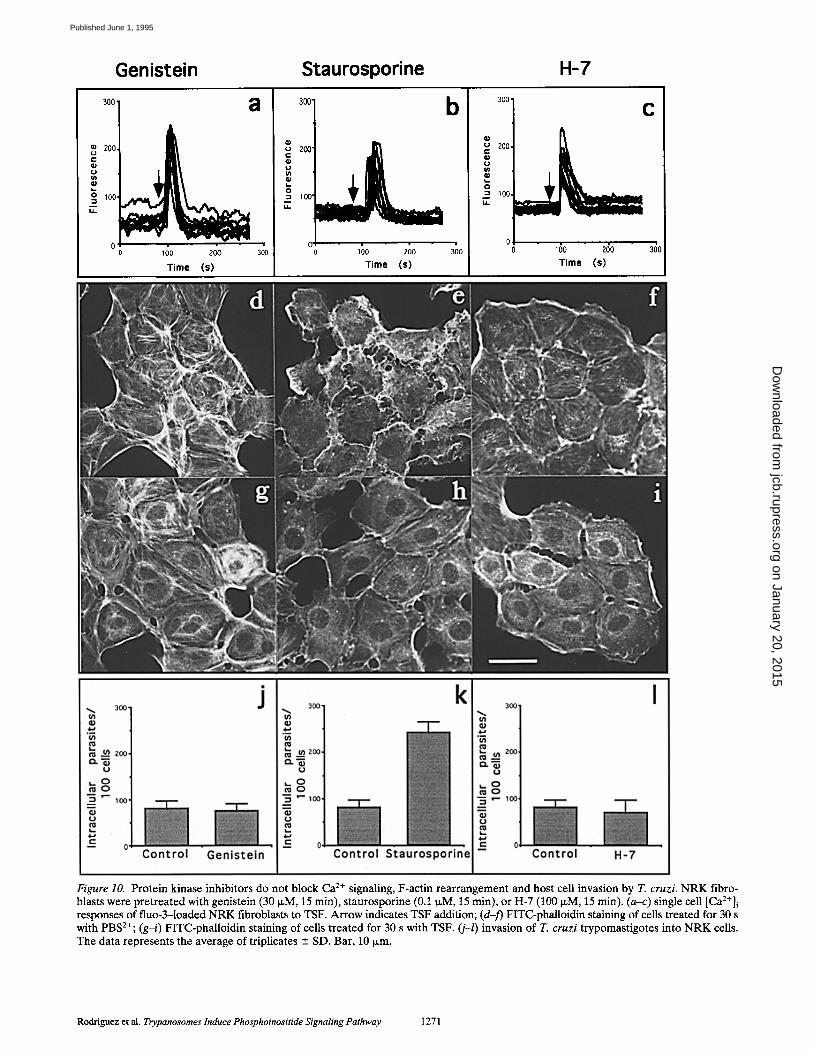
Inhibition of the trypomastigote-induced Ca 2+ signaling pathway by pertussis toxin (Tardieux et al., 1994) sug- gested the involvement of a trimeric G protein-linked re- ceptor. To investigate the alternative possibility, the in- volvement of a tyrosine kinase type of receptor, NRK cells were pretreated for 15 min with 30 }xM genistein (Aki- yama et al., 1987), before addition of the TSF. As shown in Fig. 10 a, treatment with this tyrosine kinase inhibitor did not affect the [Ca2+]i response. In addition, the F-actin re- organization induced by PGTF (Fig. 10 g) and parasite in- vasion (Fig. 10 j) were also not affected, suggesting that genistein-sensitive protein kinases are not involved in these processes.
NRK cells were also pretreated with the protein kinase C (PKC) inhibitors H-7 (100 IxM for 15 min; Kawamoto and Hidaka, 1984) and staurosporine (0.1 IxM for 15 min, Tamaoki et al., 1986). Neither treatment interfered with the [Ca2+]i response (Fig. 10, b and c), nor with the cen- tripetal F-actin reorganization induced by PGTF (Fig. 10,
h and i). However, the inhibitors themselves, particularly staurosporine, induced overall changes in the microfila- ment pattern of NRK cells (Fig. 10, e and f). The efficiency of invasion by T. cruzi was not affected by pretreatment of the host ceils with H-7, and it was enhanced by staurospo- rine (Fig. 10, k and l). The latter result was probably a con- sequence of the staurosporine-induced overall microfila- ment depolymerization, similarly to what was observed earlier in cytochalasin D-treated cells (Tardieux et al., 1992).
Discussion
In this work we characterize signal transduction events oc- curring in NRK fibroblasts upon exposure to PGTF, a pro- teolytically generated factor from the protozoan parasite T. cruzi that induces [Ca2+]i transients in several mamma- lian cell types (Burleigh and Andrews, 1995).
We investigated the origin of the [Ca2+]i transients in- duced by PGTF-containing soluble extracts of trypomas- tigotes. As controls, we used similar extracts prepared from epimastigotes that do not contain PGTF, or trypo- mastigote fractions treated with leupeptin to inhibit PGTF generation. We conclude that the initial [Ca2÷]i elevation induced by PGTF in NRK cells is mediated by Ca 2+ re- lease from intracellular stores, as the signal is not depen- dent on extracellular Ca 2÷ and is completely abolished by thapsigargin. We also show that emptying the intracellular Ca 2÷ stores with thapsigargin significantly inhibits NRK cell entry by T. cruzi, confirming previous findings impli- cating the PGTF-induced [Ca 2+] transients in invasion (Tardieux et al., 1994).
The observation that thapsigargin inhibits the [Ca2+]i transients induced in NRK cells by both soluble extracts and intact trypomastigotes indicates that these responses have a similar origin. One difference, however, is ob- served: the response to the soluble trypomastigote fraction occurs synchronously in the majority of the target cells, while intact parasites induce asynchronous, repetitive [Ca2+]i oscillations in only a few cells of the population. While the reason for this difference is not yet clear, it is
100
i • ~ 80,
~ t 60, ~ 40,
20, 8 0
0 0
Trypanosome Thrornbin ~ sol u bl efracti on / i ~ ~ "
' ,_ . . . , .___, . . . . , . . - . , - . . . , . . . - , - . ._ . . ' . . . , . - - - , .... , - --- , . . . - , --- . , . - ._,__..,____,_.__,__..,....,....,...., .... 30 60 90 0 30 60 90 0 30 60 90 120
Time (s)
b
200 .~ E
lOO
i
Figure 9. Correlation between the NRK cell [Ca2+]i response and the percentage of cells showing F-actin rearrange- ment over time. The fluores- cence intensity profile for [Ca2+]i shown corresponds to the average of measurements performed in 10 individual fluo-3-1oaded NRK cells. The percentage of cells with F-actin reorganization was determined by counting al- tered cells in a total of 500, after exposure to TSF, throm- bin (0.5 U/ml) or ionomycin (5 ixM), and staining with FITC-phalloidin. Agonists were added at time = 0 s.
The Journal of Cell Biology, Volume 129, 1995 1270
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995
Genistein Staurosporine H-7
300' 8
200
1oo
0 o
u ~ 2o0
I 300
001 200
~ 100 ,'7
01 , . . / 0 100 200 300j
Time (s) /
Figure 10. Protein kinase inhibitors do not block Ca 2÷ signaling, F-actin rearrangement and host cell invasion by T. cruzi. NRK fibro- blasts were pretreated with genistein (30 ~,M, 15 min), staurosporine (0.1 N,M, 15 min), or H-7 (100/~M, 15 min). (a-c) single cell [Ca2+]i responses of fluo-3-1oaded NRK fibroblasts to TSF. Arrow indicates TSF addition; (d-f) FITC-phalloidin staining of cells treated for 30 s with PBS2÷; (g-i) FITC-phalloidin staining of cells treated for 30 s with TSF. (j-l) invasion of T. cruzi trypomastigotes into NRK ceils. The data represents the average of triplicates --- SD. Bar, 10 txm.
Rodriguez et al. Trypanosomes Induce Phosphoinositide Signaling Pathway 1271
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995

conceivable that live parasites release small amounts of PGTF intermittently, as they associate with individual host cells. Low concentrations of PGTF could be the factor re- sponsible for the slower Ca 2÷ oscillations observed in cells stimulated by live parasites. The frequency of Ca 2÷ oscilla- tions induced through the inositol phospholipid signaling pathway of mammalian cells often depends on the concen- tration of the extracellular agonist (Woods et al., 1986).
The role of IP3 in releasing Ca 2÷ from intracellular stores through IP3-sensitive channels is well documented (Berridge, 1993). We have detected a specific increase in polyphosphoinositide hydrolysis with the generation of IP3 after addition of PGTF to NRK cells. The kinetics of ino- sitol phosphate release induced by the TSF in the presence of LiC1 was similar to that induced by hormones and growth factors in other cell types (Paris and Pouyssegur, 1986; Rosenbach and Greenlee, 1991). Taken together with the inhibitory effect of a specific PLC inhibitor, our results strongly suggest that IP3 is involved in the intracel- lular Ca 2÷ mobilization induced by PGTF in NRK cells.
Signaling mediated by PLC activation and IP3 formation is usually triggered by the stimulation of specific plasma membrane receptors, often coupled to intracellular media- tors through GTP-binding regulatory proteins (G pro- teins). The earlier finding that pertussis toxin inhibits the T. cruzi-induced [Ca2+]i transients (Tardieux et al., 1994) suggests the involvement of a trimeric G protein contain- ing a PTx-sensitive G~ subunit, perhaps associated with a seven-transmembrane class of receptor. However, we can- not at this point exclude the possibility that PGTF may be a nonspecific agonist similar to the cationic amphiphilic neuropeptides and venon peptides that directly activate G proteins in the absence of a specific receptor (Mousli et al., 1990).
The observed recruitment and fusion of host cell lyso- somes at the invasion site, and the enhancement in para- site entry after disruption of microfilaments with cyto- chalasin D (Tardieux et al., 1992) suggested that actin reorganization might be an early step in the T. cruzi inva- sion process. Ca 2÷ regulation of the actin cytoskeleton through actin-binding proteins is extensively documented (Vanderkhove, 1990; Watts and Howard, 1992; Stossel, 1993; Weeds and Maciver, 1993). There is also evidence that reorganization of the cortical actin cytoskeleton is necessary to allow vesicles to reach exocytotic sites in stimulated secretory cells (Burgouyne and Cheek, 1987; Linstedt and Kelly, 1987; Aunis and Bader, 1988; De Ca- milli et al., 1990). Therefore, we were interested in verify- ing if the [Ca2+]i transients induced by T. cruzi trypomas- tigotes had an effect on the organization of host cell microfilaments. We found that addition of the PGTF-con- taining TSF to NRK cells resulted in a rapid and transient reorganization of the cortical actin cytoskeleton. Live try- pomastigotes induced similar changes, but in an asynchro- nous fashion that allowed detection of F-actin reorganiza- tion in only a few cells of the population at a given time. This observation correlates well with the asynchronous pattern of Ca 2÷ signaling induced in NRK cells by live try- pomastigotes (Tardieux t t al., 1994; Fig. 2).
An increase in [Ca2+]i appears to be sufficient for the rapid centripetal F-actin reorganization induced by T. cruzi in NRK cells. This interpretation is based on two sets
of data. First, a similarly altered microfilament pattern was detected after exposure of the NRK cells to the Ca 2÷ iono- phore ionomycin. Second, there was a good correlation between the kinetics of the Ca 2÷ signal and that of the F-actin rearrangement. Therefore, the [Ca2+]i rise trig- gered by PGTF may be directly responsible for changes in the organization of actin, possibly by activating Ca 2÷- dependent microfilament-severing proteins such as gelso- lin (Watts and Howard, 1992; Weeds and Maciver, 1993). However, we must also consider a direct role for PLC- mediated polyphosphoinositide hydrolysis in disassembly of the cortical cytoskeleton, since it has been proposed that release of phosphoinositide-binding actin regulatory proteins can lead to capping, severing, and sequestration of actin monomers (Goldschmidt-Clemont et al., 1991; Stos- sel, 1993; Janmey 1994).
Centripetal F-actin reorganization such as the one in- duced in NRK cells by T. cruzi has been reported in other cell types, and shown to accompany cellular processes such as locomotion, pseudopod extension, and secretion (Heath, 1983; Norman et al., 1994). In microglial cells, mobilization of Ca 2÷ from intracellular stores by bacterial lipopolysac- charide is associated with reorganization of actin filaments into a diffuse perinuclear pattern (Bader et al., 1994). Such rearrangement appears to involve a reduction in F-actin content in the cortical region and an increase in the cell in- terior (Norman et al., 1994).
It remains to be determined if host cell invasion by T. cruzi involves a local disassembly of the cortical actin cy- toskeleton, and if these changes can be correlated with a localized cytosolic Ca 2÷ wave. An area of elevated [Ca2+]i was recently detected in L6E9 myoblasts around cell-asso- ciated T. cruzi trypomastigotes (Moreno et al., 1994). Ly- sosome recruitment and fusion may be regulated by a lo- calized increase in [Ca2+]i, similarly to what occurs during azurophil granule fusion in neutrophils (Jaconi et al., 1990), neurotransmitter release in synaptic terminals (Smith et al., 1988) and perforin release from cytotoxic T lympho- cytes (Poenie et al., 1987). It is interesting to note that neutrophil azurophil granules and the lytic granules of cytotoxic lymphocytes are considered to have a common biogenesis with lysosomes, based on the presence of hy- drolytic enzymes and of lysosomal membrane glycopro- teins (Jaconi et al., 1990; Griffiths and Isaaz, 1993).
NRK cell pretreatment with genistein, a tyrosine kinase inhibitor, and of H-7 and staurosporine, inhibitors of PKC and other kinases, had no detectable effect on the [Ca2+]i transients or on the centripetal F-actin reorganization in- duced by PGTF. However, staurosporine significantly en- hanced parasite invasion. We believe this effect is related to the overall microfilament disassembly that occurred in NRK cells after exposure to staurosporine. Disruption of the actin cytoskeleton with cytochalasin D causes a similar degree of enhancement in T. cruzi invasion (Tardieux et al., 1992). The disruptive effect of staurosporine on the general organization of the actin cytoskeleton has been of- ten overlooked, and may explain previous observations at- tributed to other protein kinase dependent processes (e.g., Roubey et al., 1991; Zheleznyak and Brown, 1992; Parton et al., 1994). The lack of inhibition by staurosporine and H-7 in our studies also indicate that PKC activity is proba- bly not required for T. cruzi invasion of NRK cells. Fur-
The Journal of Cell Biology, Volume 129, 1995 1272
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995

thermore, the observation that genistein does not affect cell entry by T. cruzi in these cells provides additional evi- dence that the process is distinct from phagocytosis, which has been shown to be inhibited by genistein (Greenberg et al., 1993).
The signal transduction events we detected in NRK fi- broblasts after exposure to the proteolytically-generated trypomastigote factor PGTF are very similar to those fol- lowing cell stimulation by several hormones and growth factors. A few seconds after PGTF stimulation different cell types respond with an elevation in [Ca2+]i that reaches concentrations between 300 and 700 nM (Burleigh and Andrews, 1995). We demonstrate here that there is a rapid, PLC-dependent polyphosphoinositide hydrolysis with production of IP 3, Ca 2+ release from intracellular stores and a dramatic F-actin reorganization in NRK cells. Inhibition of various steps in this signal transduction cas- cade results in block of the T. cruzi invasion process, strongly suggesting a role for PGTF in mediating the para- site-host cell interaction.
We thank A. H. Cornell-Bell for extensive use of the con:focal microscope, H. Tan for photography, M. Cappello for advice with HPLC and B. Bur- leigh for comments on the manuscript.
This work was supported by National Institutes of Health grants RO1AI32056 and RO1AI34867 to N. W. Andrews and A. Rodriguez was supported by a fellowship from CICYT (Spain).
Received for publication 8 February 1995 and in revised form 6 March 1995.
References
Akiyama, T., J. Ishida, S. Nakagawa, H. Ogawara, S. Watanabe, N. Itoh, M. Shibuya, and Y. Fukami. 1987. Genistein, a specific inhibitor of tyrosine-spe- cific protein kinases. J. Biol. Chem. 262:5592-5595.
Andrews, N. W., K. S. Hong, E. S. Robbins, and V. Nussenzweig. 1987. Stage- specific surface antigens expressed during the morphogenesis of vertebrate forms of Trypanosoma cruzi. Exp. Parasitol. 64:474-484.
Aunis, D., and M. F. Bader. 1988. The cytoskeleton as a barrier to exocytosis. Z Exp. Biol. 139:253-266.
Bader, M. F., L. Taupenot, G. Ulrich, D. Aunis, and J. Cieleski-Treska. 1994. Bacterial endotoxin induces [Ca2+]i transients and changes the organization of actin microglia. Glia. 11:336-344.
Berridge, M. J. 1993. Inositol trisphosphate and calcium signaling. Nature (Lond.). 361:315-325.
Bleasdale, J. E., R. Thakur, R. S. Gremban, G. L. Bundy, F. A. Fitzpatrick, R. J. Smith, and S. Bunting. 1990. Selective inhibition of receptor-coupled phos- pholipase C-dependent processes in human platelets and polymorphonu- clear neutrophils. £ Pharmacol. Exp. Ther. 255:756-768.
Bliska, J. B., J. E. Galan, and S. Falkow. 1993. Signal transduction in the mam- malian cell during bacterial attachment and entry. Cell. 73:903-920.
Bretscher, A. 1991. Microfilament structure and function in the cortical cyto- skeleton. Annu. Rev. Cell Biol. 7:903-920.
Burgouyne, R. D., and T. R. Cheek. 1987. Reorganization of peripheral actin filaments as a prelude to exocytosis. Biosci. Rep. 7:281-288.
Burleigh, B., and N. W. Andrews. 1995. A 120 KDa alkaline peptidase from Trypanosoma cruzi is involved in the generation of a novel Ca2÷-signaling factor for mammalian cells. J. BioL Chem. 270:5172-5180.
Chinkers, M., J. A. McKanna, and S. Cohen. 1979. Rapid induction of morpho- logical changes in human carcinoma cells A-431 by epidermal growth factor. J. Cell Biol. 83:260-265.
De Camilll, P., F. Benefenati, F. Valtorta, and P. Greengard. 1990. The syn- apsins. Annu. Rev. Cell. Biol. 111:1069-1079.
Goldschmidt-Clermont, P. J., J. W. Kim, L. M. Machesky, S. G. Rhee, and T. D. Pollard. 1991. Regulation of phospholipase C--/1 by profilin and tyrosine phosphorilation. Science (Wash. DC). 251:1231-1233.
Oreenberg, S., P. Chang, and S. C. Silverstein. 1993. Tyrosine phosphorylation is required for Fc receptor-mediated phagocytosis in mouse macrophages. J. Exp. Med. 177:529-534.
Griffiths, G. M., and S. Isaaz. 1993. Granzymes A and B are targeted to the lytic granules of lymphocytes by the mannose-6- phosphate receptor. A Cell Biol. 120:885496.
Hagiwara, S., and L. Byerly. 1981. Calcium channels. Annu. Rev. Neurosci. 4: 69-125.
Hallam, T. J., R. Jacob, and E. Merrit. 1988. Evidence that agonists stimulate
bivalent-cation influx into human endothelial cells. Biochem. J. 255:179-184. Heath, J. P. 1983. Behaviour and structure of the leading lamella in moving fi-
broblasts. J. Cell Sci. 60:331-354. Heuser, J. E. 1989. Changes in lysosome shape and distribution correlated with
changes in cytoplasmic pH. J. Cell Biol. 108:855--864. Inesi, G., and Y. Sagara. 1992. Thapsigargin, a high affinity and global inhibitor
of intracellular Ca z+ transport ATPases. Arch. Biochem. Biophys. 298:313- 317.
Jaconi, M. E. E., D. P. Lew, J. L. Carpentier, K. E. Magnusson, M. Sjrgren, and O. Stendahl. 1990. Cytosolic free calcium elevation mediates the phago- some-lysosome fusion during phagocytosis in human neutrophils. J. Cell Biol. 110:1555-1564.
Janmey, P. A. 1994. Phosphoinositides and calcium as regulators of cellular ac- tin assembly and disassembly. Annu. Rev. Physiol. 56:169-191.
Kadowaki, T., S. Koyasu, E. Nishida, H, Sakai, F. Takaku, I. Yahara, and M. Kasuga. 1986. Insulin-like growth factors, insulin, and epidermal growth fac- tor cause rapid cytoskeletal reorganization in KB cells. Z Biol. Chem. 261: 16141-16147.
Kawamoto, S., and H. Hidaka. 1984. 1-(5-Isoquinolinesulfonyl)-2-(methylpip- erazine (H-7) is a selective inhibitor of protein kinase C in rabbit platelets. Biochem. Biophys. Res. Commun. 125:258-264.
Linstedt, A. D., and R. B. Kelly. 1987. Overcoming barriers to exocytosis. Trends Neurosci. 10:446--448.
Majerus, P. W. 1992. Inositol phosphate biochemistry. Annu. Rev. Biochem. 61: 225-250.
Mousli, M., J. L. Bueb, C. Bronner, B. Rouot, and Y. Landry. 1990. G protein activation: a mode of action of cationic amphiphilic neuropeptides and venom peptides. Trends Pharmacol. 11:358--362.
Moreno, S. N. J., J. Silva, A. E. Vercesi, and R. Docampo. 1994. Cytosohc-free calcium elevation in Trypanosoma cruzi is required for cell invasion. Z Exp. Med. 180:1535-1540.
Nogueira, N., and Z. A. Cohn. 1976. Trypanosoma cruzi; mechanism of entry and intracellular fate in mammalian ceils. J. Exp. Med. 143:1402-1420.
Norman, J. C., L. S. Price, A. J. Ridley, A. Hall, and A. Koffer. 1994. Actin fila- ment organization in activated mast cells is regulated by heterotrimeric and small GTP-binding proteins. J. Cell Biol. 126:1005-1015.
Paris, S., and J. Pouyssegur. 1986. Pertussis toxin inhibits thrombin-induced ac- tivation of phosphoinositide hydrolysis and Na+/H+ exchange in hamster fibroblasts. EMBO (Eur. Mol. Biol. Org.) J. 5:55-60.
Parton, R. G., B. Joggerst, and K. Simmons. 1994. Regulated internalization of caveolae. Z Cell BioL 127:1199-1215.
Poenie, M., R. Y. Tsien, and A. M. Schmitt-Verhulst. 1987. Sequential activa- tion and lethal hit by [Ca2+]i individual cytolytic T cells and targets. EMBO (Eur. Mol. Biol. Org.) Z 6:2223-2232.
Rosenbach, T., and W. F. Greenlee. 1991. Inositol phosphate formation in the human squamous cell carcinoma line SCC-12 F: studies with bradykinin, the calcium ionophore A23187, and sodium fluoride. J. Invest. Dermatol. 96:116- 122.
Roubey, R. A., G. D. Ross, J. T. Merrill, F. Walton, W. Reed, R. J. Winchester, and J. P. Buyon. 1991. Staurosporine inhibits neutrophil phagocytosis but not iC3b binding mediated by CR3 (CDllb/CD18). J. Immunol. 146:3557- 3562.
Sehenkman, S., N. W. Andrews, V. Nussenzweig, and E. S. Robbins. 1988. Try- panosoma cruzi invade a mammalian epithelial cell in a polarized manner. Cell. 55:157-165.
Schenkman, S., E. S. Robbins, and V. Nussenzweig. 1991. Attachment of Try- panosoma cruzi to mammalian cells requires parasite energy, and invasion can be independent of the target cell eytoskeleton. Infect. Immun. 59:645- 654.
Smith, S. J., and G. J. Agustine. 1988. Calcium ions, active zones and synaptic transmitter release. Trends. Neurosci. 11:458--464.
Stossel, T. P. 1993. On the crawling of animal cells. Science (Wash. DC). 260: 1086-1094.
Tamaoki, T., H. Nomoto, I. Takahashi, Y. Kato, M. Morimoto, and F. Tomita. 1986. Staurosporine, a potent inhibitor of phospholipid/Ca ÷+ dependent protein kinase. Biochem. Biophys. Res. Com. 135:397--402.
Tardieux, I., P. Webster, J. Ravensloot, W. Boron, J. A. Lunn, J. E. Heuser, and N. W. Andrews. 1992. Lysosome recruitment and fusion are early events re- quired for trypanosome invasion of mammalian cells. Cell. 71:1117-1130.
Tardieux, I., M. Nathanson, and N. W. Andrews. 1994. Role in host cell inva- sion of Trypanosoma cruzi-induced eytosolic-free Ca 2+ transients. Z Exp. Med. 179:1017-1022.
Vanderkhove, J. 1990. Actin-binding proteins. Curt. Opin. Cell. Biol. 2:41-50. Watts, R. G., and T. H. Howard. 1992. Evidence for a gelsolin-rich, labile
F-actin pool in human polymorphonuclear leucocytes. Cell. Motil. Cytoskele- ton. 21:25-37.
Weeds, A., and S. Maciver. 1993. F-actin capping proteins. Curt. Opin. Cell Biol. 5:63-69.
Woods, N. M., K. S. Cuthbertson, and P. H. Cobbold. 1986. Repetitive transient rises in cytoplasmic free calcium in hormone-stimulated hepatocytes. Nature ( L o nd.). 319:600--602.
Zheleznyak, A., and E. J. Brown. 1992. Immunoglobulin-mediated phagocyto- sis by human monocytes requires protein kinase C activation. Evidence for protein kinase C transloeation to phagosomes. Z Biol. Chem. 267:12042- 12048.
Rodriguez et al. Trypanosomes lnduce Phosphoinositide Signaling Pathway 1273
on January 20, 2015jcb.rupress.org
Dow
nloaded from
Published June 1, 1995




















