
Process engineering for bioflavour production with metabolically active yeast – a minireview
Upload
independentCategory
view
0download
0
Ž .Brain Research Protocols 2 1998 209–214
Protocol
A rapid method for the isolation of metabolically active mitochondria fromrat neurons and astrocytes in primary culture
Angeles Almeida, Jose M. Medina )´Departamento de Bioquımica y Biologıa Molecular, Facultad de Farmacia, UniÕersidad de Salamanca, Edificio Departamental, AÕda. del Campo Charro,´ ´
37007 Salamanca, Spain
Received 3 November 1997; accepted 10 November 1997
Abstract
Ž .A rapid method about 1.5 h for the isolation of intact functional mitochondria from neurons and astrocytes in primary culture isdescribed. Mitochondria isolated by this method are metabolically active and tightly coupled as shown by respiratory control ratio values,which were about 4 with glutamate–malate as substrate. The activities of marker enzymes revealed the occurrence of a low degree of
Ž . Ž .cytosolic 5% or synaptosomal 5.5% contamination in the mitochondrial fractions. In addition, the activity of citrate synthase wasincreased by 4 fold in both neuronal and astrocytic mitochondria with respect to values found in cell homogenates. These results confirmthat the method affords mitochondrial preparations from cultured brain cells at suitable levels of purity and enrichment for the study oftheir mitochondrial function. Since mitochondrial damage has been associated with the pathogenesis of certain neurodegenerative
Ždiseases, such as Alzheimer’s and Parkinson’s diseases P. Chagnon, C. Betard, Y. Robitaille, A. Cholette, D. Gauvreau, Distribution ofŽ . w xbrain cytochrome oxidase activity in various neurodegenerative disease, Neuroreport 6 1995 711–715 6 ; S.J. Kish, C. Bergeron, A.
Rajput, S. Dozic, F. Mastrogiacomo, L. Chang, J.M. Wilson, L.M. DiStefano, J.N. Nobrega, Brain cytochrome oxidase in Alzheimer’sŽ . w xdisease, J. Neurochem. 59 1992 776–779 10 ; A.H.V. Schapira, J.M. Cooper, D. Dexter, J.B. Clark, P. Jenner, C.D. Marsden,
Ž . w x.Mitochondrial complex I deficiency in Parkinson’s disease, J. Neurochem. 54 1990 823–827 15 , the method described here shed lighton the possible susceptibility of neuronal or astrocytic mitochondria to deleterious effects of these diseases. q 1998 Elsevier Science B.V.
Theme: Endocrine and autonomic regulation
Topic: Respiratory regulation
Keywords: Mitochondria; Isolation; Neurons; Astrocytes; Respiration
1. Type of research
w xØ Isolation of mitochondria from cultured cells 3,9,14 .Ø Mitochondrial respiratory function.Ø Mitochondrial coupling.
2. Time required
About 2 h for preparing astrocyte or neuron culture.Cells were used after 5 days of culture. Cell homogeniza-tion and mitochondrion isolation take approximately 1 hand 30 min. Determinations of mitochondrial respiratory
) Corresponding author. Fax: q34-23-29-45-64.
parameters and marker enzyme activities take about 10min per assay.
3. Materials
Animals. Pregnant albino Wistar rats fed ad libitum on astock laboratory diet were used for the experiments. Toprepare neurons in primary culture, fetuses at 16 d ofgestation were used. Postnatal day-1 newborn rats wereused to prepare astrocytes in primary culture. Approxi-mately 12 neonates or 10 fetuses were used per astrocyteor neuron preparation, respectively.
Ž .Poly-L-lysine coated flasks. Flasks Nunc were incu-Ž .bated at 378C with 10 mgrml wrv poly-L-lysine in
sterile water for 15 min. Poly-L-lysine was removed andflasks were washed once with sterile water.
1385-299Xr98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S1385-299X 97 00044-5

( )A. Almeida, J.M. MedinarBrain Research Protocols 2 1998 209–214210
Special equipment. None required for mitochondrialisolation. A Clark-type oxygen electrode and a thermostati-
Ž .cally controlled 308C and magnetically stirred incubationchamber of about 500 ml capacity is required to measuremitochondrial oxygen consumption in order to check mito-chondrial coupling.
Chemicals and reagents
Ž .Ø Acetyl-CoA SigmaŽ . Ž .Ø Bovine serum albumin BSA Sigma
Ž .Ø Cytosine arabinoside SigmaŽ . Ž .Ø DNase EC 3.1.21.1 type I Sigma
X Ž . Ž .Ø 5,5 -Dithiobis-2-nitrobenzoic acid DTNB SigmaŽ . Ž .Ø Dulbecco’s modified Eagle’s medium DMEM Sigma
Ž . Ž .Ø Earle’s balanced salt solution EBSS SigmaŽ .Ø EDTA Sigma
Ž . Ž .Ø Fetal calf serum FCS Boehringer MannheimŽ .Ø Glutamic acid Sigma
Ž .Ø Mannitol SigmaŽ .Ø Malic acid Sigma
Ž .Ø NADH SigmaŽ .Ø Oxaloacetate SigmaŽ .Ø Poly-L-lysine Sigma
Ž .Ø Potassium chloride MerckŽ .Ø Sucrose Sigma
Ž .Ø Tris SigmaŽ . Ž .Ø Trypsin EC 3.4.21.4 Boehringer Mannheim
4. Detailed procedure
4.1. Cell culture
Primary neuron and astrocyte cultures are prepared inŽ .an incubator at 378C equipped with a CO –air 5:952
w xentry, as previously described 20 :Ø Remove forebrains and mince with a scalpel in EBSS
Žcontaining 20 mgrml DNase and 0.3% BSA solution.A .
Ø Centrifuge at 500=g for 2 min.Ø Incubate the pellet in EBSS containing 0.025% trypsin,
Ž .60 mgrml DNase and 0.3% BSA solution B at 378Cfor 15 min.
Ø Stop trypsinization by adding FCS at a final concentra-tion of 10%.
Ø Centrifuge at 500=g for 5 min.Ø Resuspend the pellet in solution A and pass suspension
8 times through a fire-polished Pasteur pipette.Ø Allow to settle for 4 min and collect supernatant.
Repeat twice.Ø Centrifuge combined fractions at 500=g for 5 min.
ŽØ Resuspend cells in culture medium DMEM supple-Ž . .mented with 10% vrv FCS .
Ø Plate cell suspensions at a density of 2.5 = 105
cellsrcm2 in 75 cm2 flasks coated with poly-L-lysine.
Ø Incubate at 378C in a humidified atmosphere containingŽ .CO –air 5:95 .2
Ø After 3 days of culture, add 10 mM cytosine arabi-noside to neuron culture
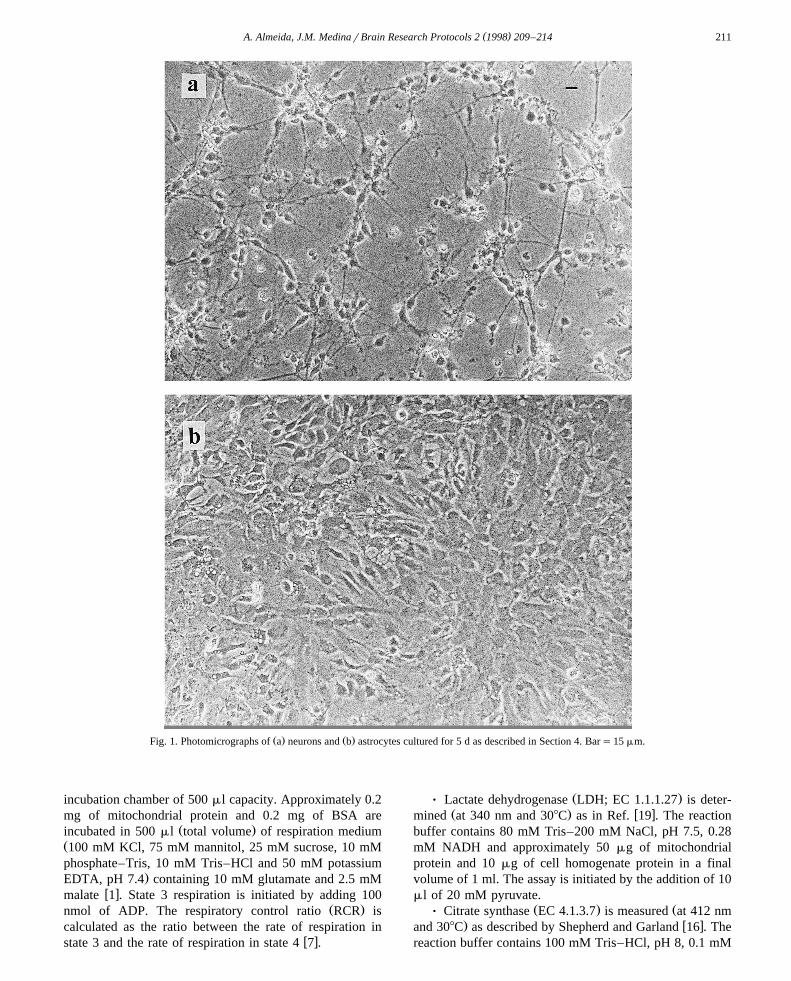
Ž .Cells were used after five days in culture Fig. 1 . AfterŽ5 days, neuronal cultures contained mainly neurons about
.85–90% , as shown by positive immunostaining with spe-w xcific neurofilament antibody 20 . In addition, 5 days of
culture is the time required for maximal type-1 astrocytesŽ .differentiation about 70% and minimal type-2 astrocyte
Ž . Ž .15% and progenitor cells 15% proliferation, as deter-w xmined by flow cytometry 18 . Microglia were always
Žlower than 1% L.I. Sanchez-Abarca and J.M. Medina,´.unpublished results .
( )4.2. Isolation of mitochondria from brain cells see Fig. 2
8 Ž .Ø Harvest about 10 cells neurons or astrocytes with ascraper and wash them twice with 5 ml of isolation
Žmedium 320 mM sucrose, 1 mM potassium EDTA, and.10 mM Tris–HCl pH 7.4 . If microglia are to be avoided,
cell harvesting should be carried out by trypsinization: addŽ .5 ml of isolation medium containing 0.05% wrv trypsin
and incubate at 378C. After 5–7 min, remove trypsinsolution and stop trypsin activity by adding 5 ml ofisolation medium containing 10% FCS. Microglia are re-moved with the trypsin solution.
Ø Centrifuge at 500=g for 5 min at 48C.Ø Resuspend the pellet in 4 ml of isolation medium.Ø Homogenize cells in a tight-fitting glass–teflon ho-
Ž . Ž .mogenizer using 20 for neurons or 30 for astrocytes upŽ Ž .and down strokes at 550 rpm. An aliquot 70 ml of the
.homogenate is frozen to measure enzyme activities.Ø Centrifuge homogenate at 1500=g for 10 min at
48C.Ž .Ø Keep supernatant s-1 on ice and re-homogenize
pellet with a further 3 ml of isolation medium.Ø Centrifuge at 1500=g for 10 min at 48C. Discard
pellet and combine supernatant with s-1.Ø Centrifuge combined supernatants again at 1500=g
for 10 min at 48C.Ø Discard small pellet and centrifuge supernatants at
17 000=g for 11 min at 48C.Ž .Ø Resuspend final pellet mitochondrial fraction in 200
Žml of isolation medium about 4 mg of mitochondrial.proteinrml and keep on ice for enzyme activity determi-
nations and metabolic studies.
4.3. Mitochondrial coupling and marker enzyme actiÕities
Ø The rate of oxygen consumption is measured polaro-graphically using a Clark-type oxygen electrode in a ther-
Ž .mostatically controlled 308C and magnetically stirred
( )A. Almeida, J.M. MedinarBrain Research Protocols 2 1998 209–214 211
Ž . Ž .Fig. 1. Photomicrographs of a neurons and b astrocytes cultured for 5 d as described in Section 4. Bars15 mm.
incubation chamber of 500 ml capacity. Approximately 0.2mg of mitochondrial protein and 0.2 mg of BSA are
Ž .incubated in 500 ml total volume of respiration mediumŽ100 mM KCl, 75 mM mannitol, 25 mM sucrose, 10 mMphosphate–Tris, 10 mM Tris–HCl and 50 mM potassium
.EDTA, pH 7.4 containing 10 mM glutamate and 2.5 mMw xmalate 1 . State 3 respiration is initiated by adding 100
Ž .nmol of ADP. The respiratory control ratio RCR iscalculated as the ratio between the rate of respiration in
w xstate 3 and the rate of respiration in state 4 7 .
Ž .Ø Lactate dehydrogenase LDH; EC 1.1.1.27 is deter-Ž . w xmined at 340 nm and 308C as in Ref. 19 . The reaction
buffer contains 80 mM Tris–200 mM NaCl, pH 7.5, 0.28mM NADH and approximately 50 mg of mitochondrialprotein and 10 mg of cell homogenate protein in a finalvolume of 1 ml. The assay is initiated by the addition of 10ml of 20 mM pyruvate.
Ž . ŽØ Citrate synthase EC 4.1.3.7 is measured at 412 nm. w xand 308C as described by Shepherd and Garland 16 . The
reaction buffer contains 100 mM Tris–HCl, pH 8, 0.1 mM

( )A. Almeida, J.M. MedinarBrain Research Protocols 2 1998 209–214212
Ž .acetyl-CoA, 0.2 mM DTNB, 0.1% vrv Triton X-100 andapproximately 10 mg of mitochondrial protein and 50 mgof cell homogenate protein in a final volume of 1 ml. Theassay is initiated by the addition of 10 ml of 20 mMoxaloacetate.
ŽØ Acetylcholinesterase activity is determined at 412. w xnm and 308C according to the method of Ellman et al. 8 .
The reaction buffer contains 100 mM K HPO –HCl, pH2 4Ž .8, 0.4 mM DTNB, 0.1% vrv Triton X-100 and approxi-
mately 50 mg of mitochondrial protein and 10 mg of cellhomogenate protein in a final volume of 1 ml. The assay isinitiated by the addition of 10 ml of 75 mM acetylthio-choline.
Ø Protein concentrations are determined by the methodw xof Lowry et al. 13 .
5. Results
Table 1 shows the activities of marker enzymes assayedin homogenates and mitochondrial fractions prepared fromneurons and astrocytes in primary culture. The cytosolic
w xmarker enzyme LDH 5 showed about 5% of total ho-mogenate activity in both neuronal and astrocytic mito-chondrial fractions. Acetylcholinesterase, a synaptic marker
w xenzyme 5 , showed about 5.5% of total homogenate activ-ity in the neuronal mitochondrial fraction, activity notbeing detectable in homogenate and mitochondria fromastrocytes. The mitochondrial marker enzyme citrate syn-
w xthase 5 was highly enriched in mitochondrial fractions,activity increasing by 3.7–4-fold in both neuronal andastrocytic mitochondrial fractions as compared to cell ho-mogenates. The yield of mitochondrial protein, relative tocell homogenates, was 6% for astrocytes and 6.5% forneurons.
Mitochondrial fractions showed RCR values of about 4with glutamate–malate as substrate. The ADPrO ratio inmitochondria from both types of cells was very close to 3Ž .Table 1 .
6. Discussion
Cultured mammalian cells are usually quite difficult tobreak open and, together with the often small amounts ofcells available and the low activity of some of the enzymesinvolved, this makes it difficult to isolate pure intact
w xmitochondria from tissue culture cells 14 . Successfulw xmitochondrial preparations isolated from neurons 9,14
w xand glial cells 3,9,14 have been achieved. However, thesemethods afford uncoupled, non-intact mitochondria andare, therefore, not suitable for the study of integratedmitochondrial function. In this work we describe a rapidmethod suitable for the isolation of metabolically-activeintact mitochondria from neurons and astrocytes in pri-mary culture. The measurements of respiratory parameters
Ž .and the activity of marker enzymes Table 1 indicate thatŽ .the method described here affords final pellet Fig. 2 that
is highly enriched in metabolically well preserved mito-chondria. Thus, the activities of the marker enzymes LDHŽ . Ža cytosolic marker enzyme or acetylcholinesterase a
. w xmarker for synaptic or membranous material 11 found inmitochondria from neurons or astrocytes with respect to
Ž .those observed in cell homogenates Table 1 suggest theŽ .occurrence of a low degree of cytosolic 5% or synaptoso-
Ž .mal 5.5% contamination in the mitochondrial fraction. Inaddition, the activity of citrate synthase, a marker of the
w xmitochondrial matrix 5 , is increased by 3.7–4-fold inboth neuronal and astrocytic mitochondrial fractions as
Ž .compared to cell homogenates Table 1 . The low recoveryof LDH and ACHase and the enrichment in citrate syn-thase found in both mitochondrial fractions confirm thepurity of these preparations. The mitochondrial enrichmentin citrate synthase activity was similar to that found in
w xisolated whole brain mitochondria 11 . It is important tonote that the yield of mitochondrial protein, relative to cellhomogenates for neurons and astrocytes was higher thanthose obtained in whole brain for synaptic and nonsynaptic
w xmitochondria 5,11,12,17 . These results suggest that mito-chondrial preparations from cultured brain cells can beobtained at suitable yields, purity and enrichment for study
Table 1Ž . Ž .Characterization of the purity marker enzyme activities and integrity RCR and ADPrO ratio of mitochondrial fractions isolated from astrocytes and
neurons in primary culture
Astrocytes Neurons
Homogenate Mitochondria Homogenate Mitochondria6Ž .LDH nmolrmin per 10 cells 96.1"6.98 5.23"0.58 32.3"2.8 1.80"0.13
6Ž .AChase nmolrmin per 10 cells ND ND 18.6"2.0 0.99"0.09Ž .CS nmolrmin per mg protein 33"1.05 130"12.5 35"1.88 135"3.72
6Ž .Proteins ngr10 cells 250"10.1 15"0.13 130"5.0 9"0.55RCR – 3.46"0.12 – 5.25"0.14
Ž .ADPrO nmol ADPrng atoms O – 2.76"0.06 – 2.94"0.04
Astrocytes and neurons were cultured for 5 d in DMEM supplemented with 10% FCS. Cells were homogenized and mitochondrial fractions were isolatedw xas described in Section 4.2. Results are means " SEM of 4–5 different mitochondrial preparations from different cell cultures. Data are obtained from 2 .
AChase, acetylcholinesterase; CS, citrate synthase; LDH, lactate dehydrogenase; ND, not detectable; RCR, respiratory control ratio.

( )A. Almeida, J.M. MedinarBrain Research Protocols 2 1998 209–214 213
Fig. 2. Flow scheme of the procedure described.
of their mitochondrial function. In this sense, mitochondriaisolated from both astrocytes and neurons had RCR valuesclose to or even above 4 with glutamate–malate as sub-
Ž . w xstrates Table 1 2 . In addition, the ADPrO values foundŽ . w xin astrocytic and neuronal mitochondria Table 1 2
indicate that under our experimental conditions mitochon-w xdria were metabolically active and tightly coupled 4,11,17 .
It should be noted that the RCR values obtained withglutamatermalate as the respiratory substrate in astrocytic
Ž .and neuronal mitochondria Table 1 were comparable tothose found in isolated non-synaptic and synaptic mito-
w xchondria from whole brain 11,17 . This corroborates thesuggestion of Clark and coworkers that the non-synapticmitochondria fraction mainly includes glial mitochondriawhile the synaptic mitochondria fraction mainly resembles
w xneuronal mitochondria 5,11,12 .
6.1. Troubleshooting
Cultured brain cells are quite difficult to break open. Itis important to homogenize cells very slowly at a speed ofabout 550 rpm, not faster, to ensure that the cells arebroken but the mitochondria are left intact. Cells must behomogenized twice in order to increase mitochondrialyield.
Ž .The low microglial contamination less than 1% inastrocyte cultures can be avoided by harvesting cells with
Ž .trypsin treatment see Section 4.2 , although the mitochon-drial yield decreases by about 15% with trypsinization.
6.2. AlternatiÕe and support protocols
Alternative protocols to isolate mitochondria from brainw xcells 3,9,14 afford uncoupled, non-intact mitochondria
and are, therefore, not suitable for the study of integratedmitochondrial function.
Acknowledgements
This work was supported by a grant from the DGICYTŽ .Spain . AA is a recipient of an ‘‘Accion de Incorporacion´ ´de Doctores y Tecnologos’’ del Ministerio de Educacion y´ ´Ciencia, Spain.
References
w x1 A. Almeida, K.L. Allen, T.E. Bates, J.B. Clark, Effect of reperfusionfollowing cerebral ischaemia on the activity of the mitochondrial
Ž .respiratory chain in the gerbil brain, J. Neurochem. 65 19951698–1703.
w x2 A. Almeida, J.M. Medina, Isolation and characterization of tightlycoupled mitochondria from neurons and astrocytes in primary cul-
Ž . Ž .tures, Brain Res. 13580 1997 in press .w x3 N. Auestad, R.A. Korsak, J.W. Morrow, J. Edmond, Fatty acid
oxidation and ketogenesis by astrocytes in primary culture, J. Neu-Ž .rochem. 56 1991 1376–1386.
w x4 J.B. Clark, W.J. Nicklas, The metabolism of rat brain mitochondria.Ž .Preparation and characterization, J. Biol. Chem. 245 1970 4724–
4731.w x Ž .5 J.B. Clark, W.J. Nicklas, Brain mitochondria. In A. Lajtha Eds. ,
Handbook of Neurochemistry, Vol. 7, Plenum Press, New York,1984, pp. 135–159.
w x6 P. Chagnon, C. Betard, Y. Robitaille, A. Cholette, D. Gauvreau,Distribution of brain cytochrome oxidase activity in various neu-
Ž .rodegenerative diseases, Neuroreport 6 1995 711–715.w x7 B. Chance, G.R. Williams, The respiratory chain and oxidative
Ž .phosphorylation, Adv. Enzymol. 17 1956 65–134.w x8 G.L. Ellman, K.D. Courtney, V. Andres, R.M. Featherstone, A new
rapid colorimetric determination of acetylcholinesterase activity,Ž .Biochem. Pharmacol. 7 1961 88–95.
w x9 A. Hamberger, C. Blomstrand, A.L. Lehninger, Comparative studieson mitochondria isolated from neuron-enriched and glial-enriched
Ž .fractions of rabbit and beef brain, J. Cell Biol. 45 1970 221–234.w x10 S.J. Kish, C. Bergeron, A. Rajput, S. Dozic, F. Mastrogiacomo, L.
Chang, J.M. Wilson, L.M. DiStefano, J.N. Nobrega, Brain cy-Ž .tochrome oxidase in Alzheimer’s disease, J. Neurochem. 59 1992
776–779.w x11 J.C.K. Lai, J.B. Clark, Preparation and properties of mitochondria
Ž .derived from synaptosomes, Biochem. J. 154 1976 423–432.w x12 J.C.K. Lai, J.B. Clark, Preparation of synaptic and nonsynaptic
Ž .mitochondria from mammalian brain, Methods Enzymol. 55 197951–60.
w x13 O.H. Lowry, N.J. Rosebrough, A. Lewis-Farr, R.J. Randall, Proteinmeasurement with the Folin phenol reagent, J. Biol. Chem. 193Ž .1951 265–275.
w x14 D. Rickwood, M.T. Wilson, V.M. Darley-Usmar, Isolation andcharacteristics of intact mitochondria. In V.M. Darley-Usmar, D.
Ž .Rickwood, M.T. Wilson Eds. , Mitochondria. A practical approach,IRL Press, Oxford, 1987, pp. 1–16.
w x15 A.H.V. Schapira, J.M. Cooper, D. Dexter, J.B. Clark, P. Jenner,C.D. Marsden, Mitochondrial complex I deficiency in Parkinson’s
Ž .disease, J. Neurochem. 54 1990 823–827.

( )A. Almeida, J.M. MedinarBrain Research Protocols 2 1998 209–214214
w x16 J.A. Shepherd, P.B. Garland, Citrate synthase from rat liver, Meth-Ž .ods Enzymol. 13 1969 11–19.
w x17 N.R. Sims, Rapid isolation of metabolically active mitochondriafrom rat brain and subregions using Percoll density gradient centrifu-
Ž .gation, J. Neurochem. 55 1990 698–707.w x18 A. Tabernero, A. Orfao, J.M. Medina, Astrocyte differentiation in
primary culture followed by flow cytometry, Neurosci. Res. 24Ž .1996 131–138.
w x19 A. Vassault, L-Lactate dehydrogenase. UV method with pyruvateŽ .and NADH. In J. Bergmeyer, M. Graßl Eds. , Methods of Enzy-
matic Analysis, Vol. 3, Verlag Chemie, Weinheim, 1983, pp. 118–126.
w x20 C. Vicario, A. Tabernero, J.M. Medina, Regulation of lactatemetabolism by albumin in rat neurons and astrocytes from primary
Ž .culture, Pediatr. Res. 34 1993 709–715.
Copyright © 2022 FDOKUMEN




















