
A presynaptically toxic secreted phospholipase A2 is internalized into motoneuron-like cells where...
11
A presynaptically toxic secreted phospholipase A 2 is internalized into motoneuron-like cells where it is rapidly translocated into the cytosol Zala Jenko Pražnikar a , Lidija Kovačič a , Edward G. Rowan b , Rok Romih c , Paola Rusmini d , Angelo Poletti d , Igor Križaj a, ⁎ , Jože Pungerčar a, ⁎ a Department of Molecular and Biomedical Sciences, Jožef Stefan Institute, Ljubljana, Slovenia b Strathclyde Institute of Pharmacy and Biomedical Sciences, University of Strathclyde, Glasgow, Scotland, UK c Institute of Cell Biology, Medical Faculty, University of Ljubljana, Ljubljana, Slovenia d Institute of Endocrinology, University of Milan, Milan, Italy Received 10 October 2007; received in revised form 7 January 2008; accepted 8 January 2008 Available online 26 January 2008 Abstract The molecular mechanism of the presynaptic toxicity of secreted phospholipase A 2 (sPLA 2 ) neurotoxins, including that of ammodytoxin A (AtxA), has not been resolved. Here we report the action of AtxA on mouse motoneuron-like cells, on which it induced characteristic neurotoxic effects on synaptic vesicles and on the reorganization of F-actin. AtxA also released fatty acids from the plasmalemma. Its significantly less neurotoxic V31W mutant showed similar effects on cells but with a much higher rate of hydrolysis than the wild-type, indicating that high enzymatic activity alone is not sufficient for the observed effects. The neurotoxic action was observed by confocal microscopy of a fluorescently labelled AtxA and by electron microscopy of a nanogold-labelled toxin. The Atx-binding proteins were tagged by a photo-cross-linking reagent conjugated to the toxin. AtxA was taken up rapidly by the cells, where it interacted within minutes with calmodulin and 14-3-3 proteins in the cytosol. These data demonstrate, for the first time, the translocation of an sPLA 2 from the extracellular space into the cytosol of a cell. Such an event may thus be important in explaining the action of a range of homologous endogenous sPLA 2 enzymes in mammals whose roles in various cellular processes are not yet completely understood. © 2008 Elsevier B.V. All rights reserved. Keywords: Ammodytoxin; Cytosolic protein; Motoneuron; Neurotoxicity; Secreted phospholipase A 2 ; Synaptic vesicle 1. Introduction Phospholipases A 2 hydrolyze the sn-2 ester bond in gly- cerophospholipids to produce free fatty acids and lysophospho- lipids. Secreted phospholipases A 2 (sPLA 2 s) are widespread in organisms [1], and found in larger quantities in animal venoms. Snake venom sPLA 2 s exhibit a wide variety of pharmacological effects, including neurotoxicity [2]. The venom of the snake Vipera ammodytes ammodytes contains at least two types of sPLA 2 molecules, ammodytoxins (Atxs) and ammodytins (Atns). Ammodytoxins AtxA, B and C are presynaptically toxic sPLA 2 s that act at the neuromuscular junction (NMJ). Ammodytins AtnI 1 and I 2 are non-toxic enzy- mes, and AtnL is an enzymatically inactive sPLA 2 -homologue with myotoxic activity [3]. The main pathological effect of Atxs is to cause complete failure of neuromuscular transmission at vertebrate NMJs, leading to respiratory problems which can lead to the death of the animal [4]. It is known that the enzymatic activity is an absolute requirement for these toxins to induce Available online at www.sciencedirect.com Biochimica et Biophysica Acta 1783 (2008) 1129 – 1139 www.elsevier.com/locate/bbamcr Abbreviations: ACh, acetylcholine; Atn, ammodytin; AtxA, B and C, ammodytoxins A, B and C; CaM, calmodulin; NMJ, neuromuscular junction; PLA 2 , phospholipase A 2 ; POPC, 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho- choline; sPLA 2 , secreted PLA 2 ; sulpho-SBED, sulphosuccinimidyl[2-6-(bioti- namido)-2-(p-azidobenzamido)-hexanoamido]-ethyl-1,3′-dithiopropionate; SV, synaptic vesicle ⁎ Corresponding authors. Department of Molecular and Biomedical Sciences, Jožef Stefan Institute, Jamova cesta 39, SI-1000 Ljubljana, Slovenia. Tel.: +386 1 477 3713; fax: +386 1 477 3984. E-mail addresses: [email protected] (I. Križaj), [email protected] (J. Pungerčar). 0167-4889/$ - see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.bbamcr.2008.01.011
Transcript of A presynaptically toxic secreted phospholipase A2 is internalized into motoneuron-like cells where...

Available online at www.sciencedirect.com
1783 (2008) 1129–1139www.elsevier.com/locate/bbamcr
Biochimica et Biophysica Acta
A presynaptically toxic secreted phospholipase A2 is internalized intomotoneuron-like cells where it is rapidly translocated into the cytosol
Zala Jenko Pražnikar a, Lidija Kovačič a, Edward G. Rowan b, Rok Romih c, Paola Rusmini d,Angelo Poletti d, Igor Križaj a,⁎, Jože Pungerčar a,⁎
a Department of Molecular and Biomedical Sciences, Jožef Stefan Institute, Ljubljana, Sloveniab Strathclyde Institute of Pharmacy and Biomedical Sciences, University of Strathclyde, Glasgow, Scotland, UK
c Institute of Cell Biology, Medical Faculty, University of Ljubljana, Ljubljana, Sloveniad Institute of Endocrinology, University of Milan, Milan, Italy
Received 10 October 2007; received in revised form 7 January 2008; accepted 8 January 2008Available online 26 January 2008
Abstract
The molecular mechanism of the presynaptic toxicity of secreted phospholipase A2 (sPLA2) neurotoxins, including that of ammodytoxin A(AtxA), has not been resolved. Here we report the action of AtxA on mouse motoneuron-like cells, on which it induced characteristic neurotoxiceffects on synaptic vesicles and on the reorganization of F-actin. AtxA also released fatty acids from the plasmalemma. Its significantly lessneurotoxic V31W mutant showed similar effects on cells but with a much higher rate of hydrolysis than the wild-type, indicating that highenzymatic activity alone is not sufficient for the observed effects. The neurotoxic action was observed by confocal microscopy of a fluorescentlylabelled AtxA and by electron microscopy of a nanogold-labelled toxin. The Atx-binding proteins were tagged by a photo-cross-linking reagentconjugated to the toxin. AtxA was taken up rapidly by the cells, where it interacted within minutes with calmodulin and 14-3-3 proteins in thecytosol. These data demonstrate, for the first time, the translocation of an sPLA2 from the extracellular space into the cytosol of a cell. Such anevent may thus be important in explaining the action of a range of homologous endogenous sPLA2 enzymes in mammals whose roles in variouscellular processes are not yet completely understood.© 2008 Elsevier B.V. All rights reserved.
Keywords: Ammodytoxin; Cytosolic protein; Motoneuron; Neurotoxicity; Secreted phospholipase A2; Synaptic vesicle
1. Introduction
Phospholipases A2 hydrolyze the sn-2 ester bond in gly-cerophospholipids to produce free fatty acids and lysophospho-
Abbreviations: ACh, acetylcholine; Atn, ammodytin; AtxA, B and C,ammodytoxins A, B and C; CaM, calmodulin; NMJ, neuromuscular junction;PLA2, phospholipase A2; POPC, 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-choline; sPLA2, secreted PLA2; sulpho-SBED, sulphosuccinimidyl[2-6-(bioti-namido)-2-(p-azidobenzamido)-hexanoamido]-ethyl-1,3′-dithiopropionate; SV,synaptic vesicle⁎ Corresponding authors. Department of Molecular and Biomedical Sciences,
Jožef Stefan Institute, Jamova cesta 39, SI-1000 Ljubljana, Slovenia. Tel.: +3861 477 3713; fax: +386 1 477 3984.
E-mail addresses: [email protected] (I. Križaj), [email protected](J. Pungerčar).
0167-4889/$ - see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.bbamcr.2008.01.011
lipids. Secreted phospholipases A2 (sPLA2s) are widespread inorganisms [1], and found in larger quantities in animal venoms.Snake venom sPLA2s exhibit a wide variety of pharmacologicaleffects, including neurotoxicity [2].
The venom of the snake Vipera ammodytes ammodytescontains at least two types of sPLA2 molecules, ammodytoxins(Atxs) and ammodytins (Atns). Ammodytoxins AtxA, B and Care presynaptically toxic sPLA2s that act at the neuromuscularjunction (NMJ). Ammodytins AtnI1 and I2 are non-toxic enzy-mes, and AtnL is an enzymatically inactive sPLA2-homologuewith myotoxic activity [3]. The main pathological effect of Atxsis to cause complete failure of neuromuscular transmission atvertebrate NMJs, leading to respiratory problems which can leadto the death of the animal [4]. It is known that the enzymaticactivity is an absolute requirement for these toxins to induce

1130 Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
the block of neuromuscular transmission [5], but the exactmolecular mechanism underlying their neurotoxicity has yet tobe fully elucidated. At present, it is suggested that the sPLA2
neurotoxins impair the cycling of synaptic vesicles (SVs) [6]or cause their depletion [7] solely by their enzymatic actionon the outer leaflet of the neuronal plasma membrane or, inaddition to this, by binding to certain intracellular proteinscritical for endo/exocytosis [8]. The “extracellular” theoryassumes that internalization of the sPLA2 neurotoxin, andhence its intracellular action, is not required for presynapticneurotoxicity. The “intracellular” hypothesis, however, is sup-ported by identification in vitro of several high-affinity bindingproteins for AtxA whose localization is intracellular – R25[9,10], calmodulin [11,12], 14-3-3 protein γ and ɛ isoforms [13],and protein disulfide isomerase [14] – as well as by the demon-stration that sPLA2s are relatively stable and enzymaticallyactive in the cytosolic environment [15–17].
The study of the physiological process underlying the re-lease of acetylcholine (ACh) from a motor nerve terminal isextremely difficult, due to its small size and due to the closephysical association with terminal Schwann cells and skeletalmuscle cells. There are several reports on the activity of neu-rotoxic sPLA2s on neuronal cells, e.g. [18–20], but most of thesestudies were performed on embryonal motoneurons isolatedfrom non-mammalian vertebrates or on non-motoneuronalcells. Therefore, in order to further explore the hypothesis thatsPLA2 neurotoxins enter the motor nerve terminal, we have hereemployed an appropriate cellular system. We demonstrated thata mouse neuroblastoma x spinal cord hybrid cell line (NSC34)[21], proven to be a useful model system in a number of studieson dysfunction, degeneration and disease of motoneurons [22–24], is also suitable for studying at least some of the aspects ofthe molecular mechanism of AtxA action. Most importantly, weshowed that the sPLA2 neurotoxin was very rapidly translocatedfrom the extracellular space into the cytosol of these cells, whereit associated with CaM and 14-3-3 proteins.
2. Materials and methods
2.1. Materials
Recombinant AtxA(N79C) and AtnI2 were prepared in Escherichia coli bymethods similar to those described for AtxA andAtxA(V31W) [25,26]. Restrictionendonucleases were obtained from New England BioLabs. T4 polynucleotidekinase was fromMBI Fermentas and T4 DNA ligase from Boehringer Mannheim.Pfu DNA polymerase and RNase One were from Promega, and Taq DNA ligasefromNewEngland BioLabs. Oligonucleotides were fromMWG-Biotech and usedwithout further purification. 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine(POPC) was from Avanti Polar Lipids. Texas Red C2 maleimide and nanogoldmonomaleimide were from Molecular Probes. Dulbecco's modified Eagle'smedium (DMEM) and inactivated foetal bovine serum (FBS) were supplied byInvitrogen.DefinedFBSwas fromHyClone andGibco.Monoclonal anti-CaMwasfrom Upstate Biotechnology (Lake Placid, NY, USA). Polyclonal anti-14-3-3antibodies, goat anti-synaptophysin and goat anti-synaptotagmin antibodies weresupplied by Santa Cruz Biotechnology (Santa Cruz, CA, USA). Mouse monoclo-nal antibodies to clathrin heavy chain were from Abcam (Cambridge, UK).AlexaFluor488-conjugated goat anti-rabbit IgG (green), AlexaFluor488-conjugateddonkey anti-goat IgG (green), AlexaFluor488-conjugated goat anti-mouse IgG(green) and rhodamine–phalloidin were from Molecular Probes. All other che-micals were of at least analytical grade and were from either Sigma or Serva.
2.2. Cell culture conditions
NSC34 cells were maintained in Dulbecco's modified Eagle's medium(DMEM) supplemented with 5% (v/v) defined FBS, 1 mM glutamine andantibiotics (100 units/ml penicillin, and 100 μg/ml streptomycin), and grown at37 °C in a humidified atmosphere of 5% CO2 and 95% air in 25 cm2 flasks; thegrowth medium was changed every 3 days. Cells were replated weekly, bymechanical dissociation, into new culture flasks at a density of 5×105 cells/flask. C2C12 and HEK293 cells were maintained in Dulbecco's modifiedEagle's medium with 20% (v/v) FBS, 1 mM glutamine and antibiotics (asabove) and grown at 37 °C in a humidified atmosphere (5% CO2–95% air) in25 cm2 flasks; the medium was changed every 2–3 days. Cells were replatedweekly, by enzymatic dissociation (trypsin), into new culture flasks at a densityof 1×105 cells/flask.
2.3. Production and purification of recombinant sPLA2s
AtxA(N79C) was prepared by site-directed mutagenesis using PCR, incorporat-ing the mutagenic oligonucleotide 5′-ACC GGG AGT GCG CCATCG TCT G-3′into the amplification product. The outer sense oligonucleotide primer, 5′-TAATACGAC TCA CTA TAG-3′, was complementary to part of the T7 RNA polymerasepromoter site on the plasmid encoding wild-type AtxA, and the outer antisenseoligonucleotide, 5′-GTT TAC TCA TAT ATA CTT TAG-3′, to a region on theplasmid downstream of the stop codon of AtxA cDNA [25]. PCR was performedsimilarly as reported [27] and the amplification product inserted into the expressionplasmid. Recombinant sPLA2s were expressed and refolded in vitro essentially asdescribed [25], and finally purified by FPLC on a Mono S column (Pharmacia) andRP-HPLC using an HP1100 system (Hewlett-Packard).
2.4. Analytical methods
Protein samples were analyzed by SDS-PAGE (Bio-Rad) in the presence of150 mMDTT using 12.5% (w/v) polyacrylamide gels and Coomassie Brilliant BlueR250 staining. Proteins were analyzed by electrospray ionization mass spectrometry(ESI-MS); analysis of the protein was performed using a high-resolution magnetic-sector AutospecQ mass spectrometer (Micromass, UK). N-terminal sequencing wasperformed on an Applied Biosystems Procise 492A protein sequencing system.
2.5. Texas Red C2 maleimide labelling of AtxA(N79C)
Purified AtxA(N79C) was dissolved in 25 mMTris–HCl, pH 8.0, containing0.5M guanidine chloride and 1 mMEDTA, and treated in the dark with a 10-foldmolar excess of Texas Red C2 maleimide for 2 h at room temperature, withconstant stirring. The reaction was stopped by adding a 5-fold molar excess ofglutathione (reduced form) and incubating at room temperature for an additional30 min. After labelling, the components of the conjugation mixture wereseparated using RP-HPLC.
2.6. Nanogold monomaleimide labelling of AtxA(N79C)
HPLC purified AtxA(N79C) was dissolved in 25 mM Tris–HCl, pH 8.0,containing 0.5 M guanidine chloride and 1 mM EDTA, and treated with a 5-foldmolar excess of nanogold monomaleimide, with a 1.4-nm gold cluster, for 1 h atroom temperature without stirring. The reaction was stopped by adding a 5-foldmolar excess of glutathione (reduced form) and incubating at room temperaturefor an additional 30 min, to ensure that no reactive species were present duringthe subsequent purification step. The components of the reaction mixture werefinally separated on a Sephadex G-25 gel filtration column equilibrated with20 mM sodium phosphate, pH 7.4, supplemented with 150 mM NaCl.
2.7. Phospholipase activity
NSC34, C2C12 and HEK293 cells were grown to 70–90% confluence, theculture medium was removed and the cells dislodged. An equal volume of thecomplete medium was added, and the cells were pelleted and washed twice withHanks' balanced salt solution with calcium and magnesium. Assays were performed

1131Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
with approx. 5×105 cells in Hanks' balanced salt solution at 37 °C. Unilamellarphospholipid vesicles, with a diameter of 0.1 μm and containing POPC, wereprepared by extrusion. Fatty acid release from intact cells and POPC vesicles byexogenously added 0.5–2000 ng sPLA2s was determined using the fluorescencedisplacement assay described previously [26]. The initial rate of hydrolysis ofphospholipids by sPLA2s was measured by monitoring the displacement of afluorescent fatty acid analogue from fatty acid-binding protein (FABP), similarly asreported [28]. Assays were performed in Hanks' balanced salt solution with1.26 mM Ca2+ and 0.9 mM Mg2+ (Invitrogen, Carlsbad, CA, USA) containing30 μM phospholipid or cells, 1 μM 11-dansylundecanoic acid and 10 μg re-combinant FABP. Solutions, with a final volume of 1.3 ml, were assayed in plasticfluorometric cuvettes at 37 °Cwith magnetic stirring, using a Perkin-Elmer LS50Bfluorometer. Excitation was at 350 nm and emission at 500 nm, with 10-nm slitwidths. Reactions were started by adding 0.5–2000 ng sPLA2, typically in 1 μl. Inthis range, a dose–response of fatty acid release due to sPLA2 activity wasobserved. All dilutions were prepared in buffer containing 1 mg/ml fatty acid-freebovine serumalbumin (BSA; Sigma) to prevent loss of enzymedue to adsorption tothe walls of the tube. Assays were calibrated by adding a known amount of oleicacid (Sigma) dissolved in methanol and monitoring the decrease in fluorescence.We chose amounts which resulted in a slope of approximately 45 degrees (forPOPC: AtxA, 10 ng; AtxA(V31W), 1 ng; AtnI2, 10 ng; for NSC34 cells: AtxA,2 ng; AtxA(V31W), 0.3 ng; AtnI2, 10 ng; for C2C12 cells: AtxA, 4 ng; AtxA(V31W), 0.5 ng; AtnI2, 10 ng; and for HEK293 cells: AtxA, 8 ng; AtxA(V31W),1 ng; AtnI2, 20 ng). Cells were exposed to the toxins for 2 min, but the initial rate ofhydrolysis of phospholipids was determined over the first 5–10 s. Cell counts werefirst determined using a hemocytometer in the presence of Trypan blue to measureviability, and the fatty acid release was then measured. Cell counts were alsodetermined after experiments, but the cell viability observed before and afterexperiment was unaltered.
2.8. Toxicity
Lethal potency was determined by intraperitoneal injection of a toxin intoBALB/c albino mice. Five dose levels and five mice per dose were used for thetoxin. The samples of recombinant toxin were diluted to a final volume of 0.5 mlin 0.9% (w/v) NaCl just prior to application and the LD50 determined after 24 hof observation. All experimental procedures on mice were carried out inaccordance with the EC Council Directive regarding animal experimentation.
2.9. F-actin staining
The NSC34 cells were plated on 4-well chamber tissue culture plates andexposed to 100 nM AtxA for 0.5, 1 and 2 h. Cells were washed twice withphosphate buffered saline (PBS) and then fixed with 3.7% paraformaldehyde(PFA; 100 μl) for 10 min, after which they were washed with PBS (3×, 15 min).Rhodamine–phalloidin (1% w/v final; ∼66 nM in PBS) was then added to eachsample, and subsequently incubated in the dark for 60 min. Cells were washed inPBS, and F-actin was visualized using epifluorescence or confocal microscopy.In some experiments the cells were incubated with PBS containing 2 mM SrCl2instead of calcium (2 mM) to test the PLA2-dependent action of AtxA.
2.10. Immunocytostaining
The NSC34 cells were plated onto a sterile cover glass or 4-well chamberslides and incubated at 37 °C with 5% CO2. The cells were treated with 100 or500 nM AtxA in DMEM for 5, 10, 15, 30 and 60 min in a 37 °C, 5% CO2
humidified incubator, supplemented with 2 mMCaCl2. Cells were washed twicewith cold PBS, and then fixed at room temperature with 3.7% PFA in PBSfor 10 min. After fixing, the cells were washed three times with PBS andpermeabilized with PBS containing 0.1% Triton X-100 and 2% BSA, for 5 minat room temperature, washed three times with PBS and incubated overnight at4 °C with the primary antibodies (rabbit anti-AtxA, goat anti-synaptophysin ormouse anti-clathrin) diluted in blocking buffer. Three further washes for 10 mineach followed, and the secondary antibodies (anti-rabbit or anti-goat or anti-mouse), diluted in blocking buffer, were then added. After 1 h incubation atroom temperature, the slides were washed three times in PBS, for 10 min each,and mounted using Mowiol (Calbiochem). Slides were examined on an inverted
epifluorescent microscope (Olympus IX71 equipped with a digital CCD cameraDP50), using a reflected light fluorescence illuminator, or by confocal microscopy.
2.11. Confocal microscopy imaging of AtxA internalization
Imaging was performed by a Zeiss LSM510 laser scanning confocal mic-roscope with the detector gain adjusted to eliminate background autofluores-cence. The signal from Texas Red conjugated to AtxC(N79C) was observeddirectly upon excitation with a 568-nm argon/krypton laser and a 650-nm longpass filter. A 63× (NA 1.4) or 40× (NA 1.2) oil immersion objective was used tovisualize the cells in all experiments. Images were analyzed using the Zeissbiophysical software package.
2.12. Labelling of Atx-binding proteins in NSC34 cells
The NSC34 or (negative) control HEK293 cells were plated on a sterile 6-wellplate and incubated at 37 °C with 5% CO2. The intact cells or, alternatively, theirlysates (positive control) were treated with 100 nM AtxC conjugated to sul-phosuccinimidyl[2-6-(biotinamido)-2-(p-azidobenzamido)-hexanoamido]-ethyl-1,3′-dithiopropionate (sulpho-SBED; Pierce Biotechnology, Rockford, IL, USA),prepared as described [29], in DMEM for 1, 2, 5, 15, 30 and 60 min in a 37 °C, 5%CO2 humidified incubator. Subsequently, cells were cooled on ice and exposed for10 min to five 15 W 312-nm UV lamps at a distance of 5 cm. Labelled cells werewashed twice with PBS, and extracts prepared. Sulpho-SBED–AtxC-labelledsampleswere separated on amonomeric avidin–Sepharose column. After applyingthe sample, the columnwas thoroughly washed with PBS and eluted with 100 mMglycine–HCl, pH 2.0. Protein-containing fractions were collected and analyzed asreported [29]. The position ofCaMand 14-3-3 proteins on the blotswas determinedimmunologically using mouse monoclonal anti-CaM antibodies at 1 μg/ml andrabbit polyclonal anti-14-3-3 antibodies at 1 μg/ml, respectively, and a BMchemiluminescence Western blotting detection system as specified by themanufacturer. Kodak BioMax Light Film (Sigma-Aldrich) was used for detection.
2.13. Transmission electron microscopy
NSC34 cells were plated in a sterile 6-well plate and incubated at 37 °C with5% CO2. They were treated with 100 nM AtxA, AtxA(N79C)-nanogold orunconjugated nanogold monomaleimide for 1 h. After incubation, for con-ventional electron microscopy, culture medium was replaced by a fixativecontaining 4% PFA and 2% glutaraldehyde in cacodylate buffer, pH 7.3. Cellswere fixed for 3 h at 4 °C, washed and postfixed in 1%OsO4 for 1 h. Finally, cellswere dehydrated with graded ethanol and embedded in Epon 812 (ElectronMicroscopy Sciences, Fort Washington, PA, USA). Ultrathin sections werecontrasted with uranyl acetate and lead citrate and examined with a PhilipsCM100 transmission microscope. For toxin-nanogold localization, cells wereimmediately fixed in 1% glutaraldehyde in PBS on ice for 30 min. Fixative wasthen washed away with PBS, then with deionized water (18 MΩ) and with0.02 M citrate buffer, pH 7.0, to reduce background. HQ Silver (Nanoprobes,Yaphank, NY, USA) was applied for 4 min in the dark at room temperature.Samples were thoroughly rinsed with deionized water and postfixed in 0.1%OsO4. Dehydration, embedding and preparation of ultrathin sections wereperformed as for conventional electron microscopy.
3. Results
3.1. Properties of recombinant AtnI2, AtxA(N79C) and itscovalent conjugates
Recombinant sPLA2s were produced in E. coli as inactiveinclusion bodies and refolded in vitro. Their homogeneity wasdemonstrated by SDS-PAGE, analytical RP-HPLC, ESI-MSand N-terminal protein sequencing (data not shown). In ad-dition, no post-translational modifications (also absent in na-tural proteins [3,4]) were observed. The basic biochemical
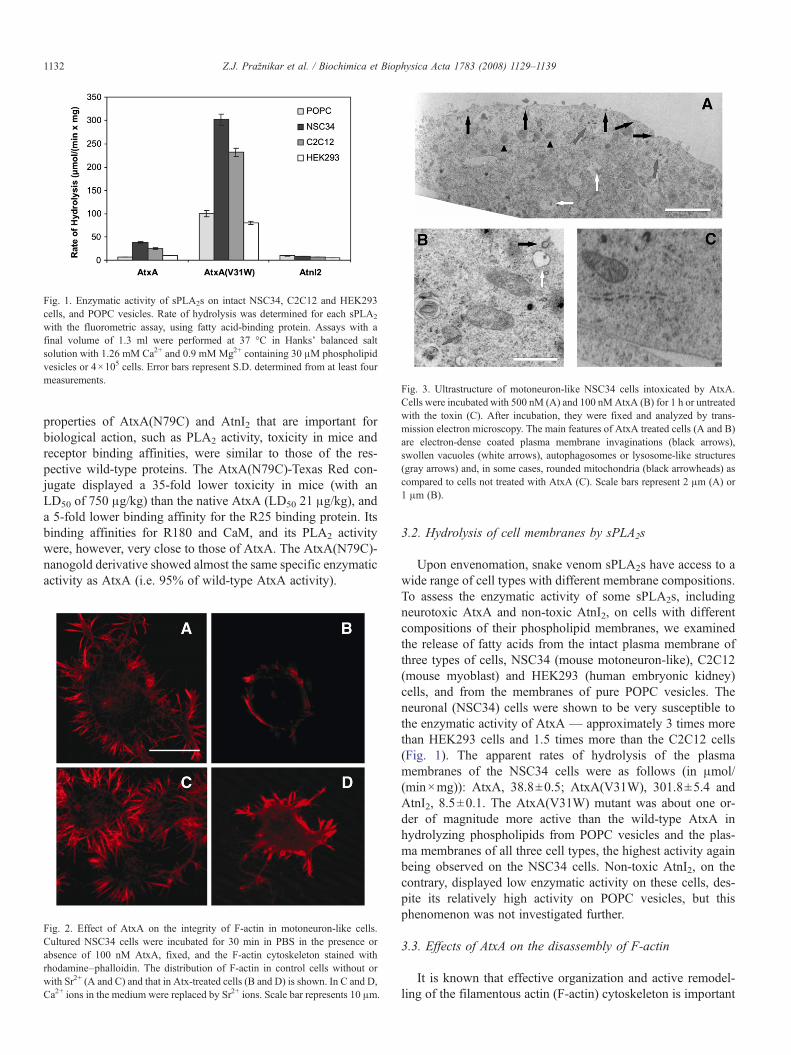
Fig. 1. Enzymatic activity of sPLA2s on intact NSC34, C2C12 and HEK293cells, and POPC vesicles. Rate of hydrolysis was determined for each sPLA2
with the fluorometric assay, using fatty acid-binding protein. Assays with afinal volume of 1.3 ml were performed at 37 °C in Hanks' balanced saltsolution with 1.26 mM Ca2+ and 0.9 mMMg2+ containing 30 μM phospholipidvesicles or 4×105 cells. Error bars represent S.D. determined from at least fourmeasurements.
Fig. 3. Ultrastructure of motoneuron-like NSC34 cells intoxicated by AtxA.Cells were incubated with 500 nM (A) and 100 nMAtxA (B) for 1 h or untreatedwith the toxin (C). After incubation, they were fixed and analyzed by trans-mission electron microscopy. The main features of AtxA treated cells (A and B)are electron-dense coated plasma membrane invaginations (black arrows),swollen vacuoles (white arrows), autophagosomes or lysosome-like structures(gray arrows) and, in some cases, rounded mitochondria (black arrowheads) ascompared to cells not treated with AtxA (C). Scale bars represent 2 μm (A) or1 μm (B).
1132 Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
properties of AtxA(N79C) and AtnI2 that are important forbiological action, such as PLA2 activity, toxicity in mice andreceptor binding affinities, were similar to those of the res-pective wild-type proteins. The AtxA(N79C)-Texas Red con-jugate displayed a 35-fold lower toxicity in mice (with anLD50 of 750 μg/kg) than the native AtxA (LD50 21 μg/kg), anda 5-fold lower binding affinity for the R25 binding protein. Itsbinding affinities for R180 and CaM, and its PLA2 activitywere, however, very close to those of AtxA. The AtxA(N79C)-nanogold derivative showed almost the same specific enzymaticactivity as AtxA (i.e. 95% of wild-type AtxA activity).
Fig. 2. Effect of AtxA on the integrity of F-actin in motoneuron-like cells.Cultured NSC34 cells were incubated for 30 min in PBS in the presence orabsence of 100 nM AtxA, fixed, and the F-actin cytoskeleton stained withrhodamine–phalloidin. The distribution of F-actin in control cells without orwith Sr2+ (A and C) and that in Atx-treated cells (B and D) is shown. In C and D,Ca2+ ions in the medium were replaced by Sr2+ ions. Scale bar represents 10 μm.
3.2. Hydrolysis of cell membranes by sPLA2s
Upon envenomation, snake venom sPLA2s have access to awide range of cell types with different membrane compositions.To assess the enzymatic activity of some sPLA2s, includingneurotoxic AtxA and non-toxic AtnI2, on cells with differentcompositions of their phospholipid membranes, we examinedthe release of fatty acids from the intact plasma membrane ofthree types of cells, NSC34 (mouse motoneuron-like), C2C12(mouse myoblast) and HEK293 (human embryonic kidney)cells, and from the membranes of pure POPC vesicles. Theneuronal (NSC34) cells were shown to be very susceptible tothe enzymatic activity of AtxA — approximately 3 times morethan HEK293 cells and 1.5 times more than the C2C12 cells(Fig. 1). The apparent rates of hydrolysis of the plasmamembranes of the NSC34 cells were as follows (in μmol/(min×mg)): AtxA, 38.8±0.5; AtxA(V31W), 301.8±5.4 andAtnI2, 8.5±0.1. The AtxA(V31W) mutant was about one or-der of magnitude more active than the wild-type AtxA inhydrolyzing phospholipids from POPC vesicles and the plas-ma membranes of all three cell types, the highest activity againbeing observed on the NSC34 cells. Non-toxic AtnI2, on thecontrary, displayed low enzymatic activity on these cells, des-pite its relatively high activity on POPC vesicles, but thisphenomenon was not investigated further.
3.3. Effects of AtxA on the disassembly of F-actin
It is known that effective organization and active remodel-ling of the filamentous actin (F-actin) cytoskeleton is important

Fig. 4. Cellular distribution of AtxA(N79C)-Texas Red in NSC34 cells at different times. Cells in culture were exposed to 100 nM AtxA(N79C)-Texas Red inDMEM buffer (i.e. low potassium non-depolarizing medium) with 2 mM Ca2+ ions for 2 min (A), 15 min (B,D,E) and 60 min (C). They were then fixed and analyzedby confocal microscopy. After fixing the cells, labelled AtxA (red; D) was stained with rabbit anti-AtxA antibodies (green; E). Scale bars represent 25 μm (A,C)or 10 μm (D).
1133Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
for a number of cellular functions, including intracellular mem-brane fusion, exocytosis of neurotransmitter, and endocytosis [30].Since the disassembly of this structure by these toxins has beendemonstrated in ratmotoneuron and in severalmodel cell lines usedto study the action of neurotoxic sPLA2 [31–33], it was importantto know whether AtxA induces the same effect in our modelNSC34 cells. Following 30 min incubation with AtxA, the cha-racteristic rhodamine–phalloidin staining of F-actin (Fig. 2A) ofthe NSC34 cells was largely lost (Fig. 2B), indicating fragmenta-tion of the F-actin cytoskeleton. The neurotoxin-induced disas-sembly of F-actin was prevented by substituting Ca2+ with Sr2+
ions in the incubation medium (Fig. 2D), suggesting that this
Fig. 5. Internalization of AtxA(N79C)-Texas Red into motoneuron-like cells in the pr(N79C)-Texas Red in DMEM buffer with 2 mM Ca2+ ions for 30 min (A), or withoumicroscopy. Scale bar represents 10 μm (B).
action depends on the phospholipase activity of AtxA, which isinhibited by Sr2+ ions. In addition, as shown below in section 3.5,internalization ofAtxA occurred very slowly in the absence of Ca2+
ions. No effects on F-actin organization were observed whencells were incubated with extracellular Sr2+ instead of Ca2+, with-out adding AtxA (Fig. 2C). In the control C2C12 cells, AtxAinduced no changes in F-actin staining pattern (data not shown).
3.4. Ultrastructural changes in NSC34 cells induced by AtxA
Electron microscope examination of NSC34 cells exposed to100 nM or 500 nM AtxA for 1 h revealed clear features of
esence and absence of Ca2+ ions. Cells in culture were exposed to 100 nM AtxAt Ca2+ ions for 30 min (B). They were then fixed and analyzed by fluorescence
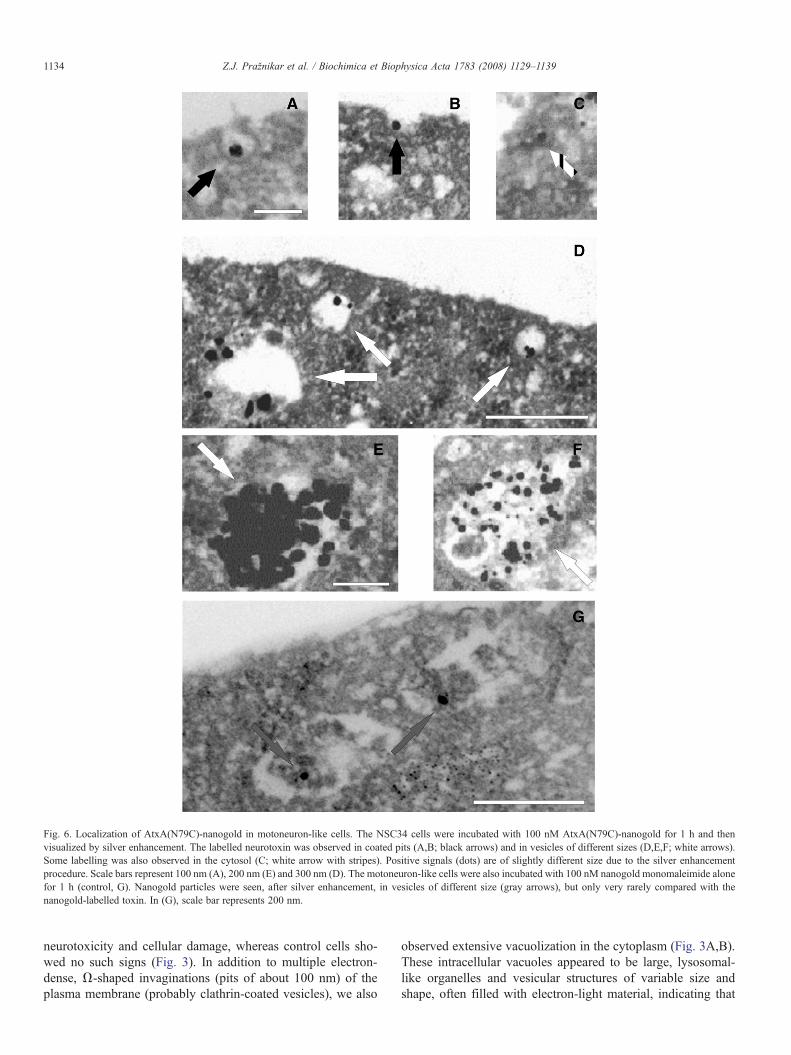
Fig. 6. Localization of AtxA(N79C)-nanogold in motoneuron-like cells. The NSC34 cells were incubated with 100 nM AtxA(N79C)-nanogold for 1 h and thenvisualized by silver enhancement. The labelled neurotoxin was observed in coated pits (A,B; black arrows) and in vesicles of different sizes (D,E,F; white arrows).Some labelling was also observed in the cytosol (C; white arrow with stripes). Positive signals (dots) are of slightly different size due to the silver enhancementprocedure. Scale bars represent 100 nm (A), 200 nm (E) and 300 nm (D). The motoneuron-like cells were also incubated with 100 nM nanogold monomaleimide alonefor 1 h (control, G). Nanogold particles were seen, after silver enhancement, in vesicles of different size (gray arrows), but only very rarely compared with thenanogold-labelled toxin. In (G), scale bar represents 200 nm.
1134 Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
neurotoxicity and cellular damage, whereas control cells sho-wed no such signs (Fig. 3). In addition to multiple electron-dense, Ω-shaped invaginations (pits of about 100 nm) of theplasma membrane (probably clathrin-coated vesicles), we also
observed extensive vacuolization in the cytoplasm (Fig. 3A,B).These intracellular vacuoles appeared to be large, lysosomal-like organelles and vesicular structures of variable size andshape, often filled with electron-light material, indicating that

Fig. 7. Co-localization of AtxA(N79C)-Texas Red and clathrin in NSC34 cells. After fixing the cells, clathrin (green; A–C) was stained by appropriate antibodies, andcells analyzed by epifluorescence microscopy. Prior to fixing, cells were exposed to 100 nM AtxA(N79C)-Texas Red conjugate in DMEM buffer with 2 mM calciumfor 2 min (red; D), 5 min (red; E), and 30 min (red; F). Scale bar represents 20 μm. In (G–I), yellow areas indicate the regions where the toxin and clathrin are co-localized.
1135Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
they could be autophagosomes and/or derived from endocytosisof the plasma membrane. More cellular damage was observed incells incubated with AtxA for longer periods (i.e. 3 h). Forexample, more mitochondria appeared rounded and their in-ternal structure became less defined. Notably, the abnormalitiesdetected in the AtxA-intoxicated NSC34 cells were similarto those reported in nerve terminals at the mammalian NMJexposed to sPLA2 neurotoxins [7].
3.5. AtxA enters rapidly into motoneuron-like cells
To study whether the sPLA2 neurotoxin AtxA is internalizedinto NSC34 cells, we prepared a recombinant AtxA chemicallylabelled with a fluorescent probe suitable for real-time mo-nitoring. Texas Red fluorophore was attached to a singlecysteine residue introduced into the molecule by replacingAsn79 with Cys by site-directed mutagenesis. After addingAtxA(N79C)-Texas Red conjugate (100 nM) to the culturemedium, its uptake by the NSC34 cells was visualized atdifferent times. The conjugate was observed to have enteredthe cells; after 2–15 min of exposure to AtxA(N79C)-TexasRed, in non-depolarizing medium, cells displayed a granularstaining pattern (Fig. 4A,B). After 30–60 min, larger AtxA(N79C)-Texas Red-containing granules or vesicles were for-med (Fig. 4C). Texas Red fluorescence was not detected in theregion of the cell nuclei. The spotty cytoplasmic distribution ofthe red signal observed by confocal fluorescence microscopyon live cells was confirmed on fixed cells by using eitherunlabelled (data not shown) or fluorescently labelled AtxA(Fig. 4E), both stained by anti-AtxA antibodies. In all cases the
cell uptake of AtxAwas found to be dependent on extracellularCa2+. In the absence of Ca2+ ions in the extracellular medium,AtxA internalization occurred only very slowly (Fig. 5B).
In order to identify intracellular structures involved in theinternalization of the toxin we examined the intracellular loca-tion of nanogold-conjugated AtxA(N79C) in the NSC34 cellswith transmission electron microscopy. AtxA(N79C)-nanogoldwas closely associated with electron-dense (probably clathrin)coated pits at the surface of the plasma membrane (Fig. 6A,B).In addition, nanogold labelling was found mainly in vesicu-lar structures ranging from 100 to 600 nm in diameter (Fig. 6,D–F). A small portion of the gold labelling was also observed inthe cytosol (Fig. 6C) and, probably, in damaged mitochondria.The NSC34 cells were also examined 1 h after being treatedwith nanogold monomaleimide only (control, Fig. 6G). A fewgold particles were observed in certain vesicles, but much morerarely than in the case of the nanogold-conjugated toxin. Theultrastructural analysis of the subcellular localization of theneurotoxin thus confirmed the spotty distribution of AtxAobserved by confocal fluorescence microscopy. The dynamicsand nature of the toxin internalization were observed by addingthe AtxA(N79C)-Texas Red conjugate to NSC34 cells, withsubsequent immunostaining of endogenous clathrin. Just twominutes after the treatment, the toxin was present mainly be-neath the plasma membrane and already co-localized withclathrin (Fig. 7A,D,G). At 15 min, AtxA and clathrin co-lo-calized predominantly in vesicles in the cytosol (Fig. 7B,E,H).By 30 min, the internalization process was already finished(Fig. 7C,F,I). In the light of this, the results presented above arebest explained by the observed internalization of AtxA into
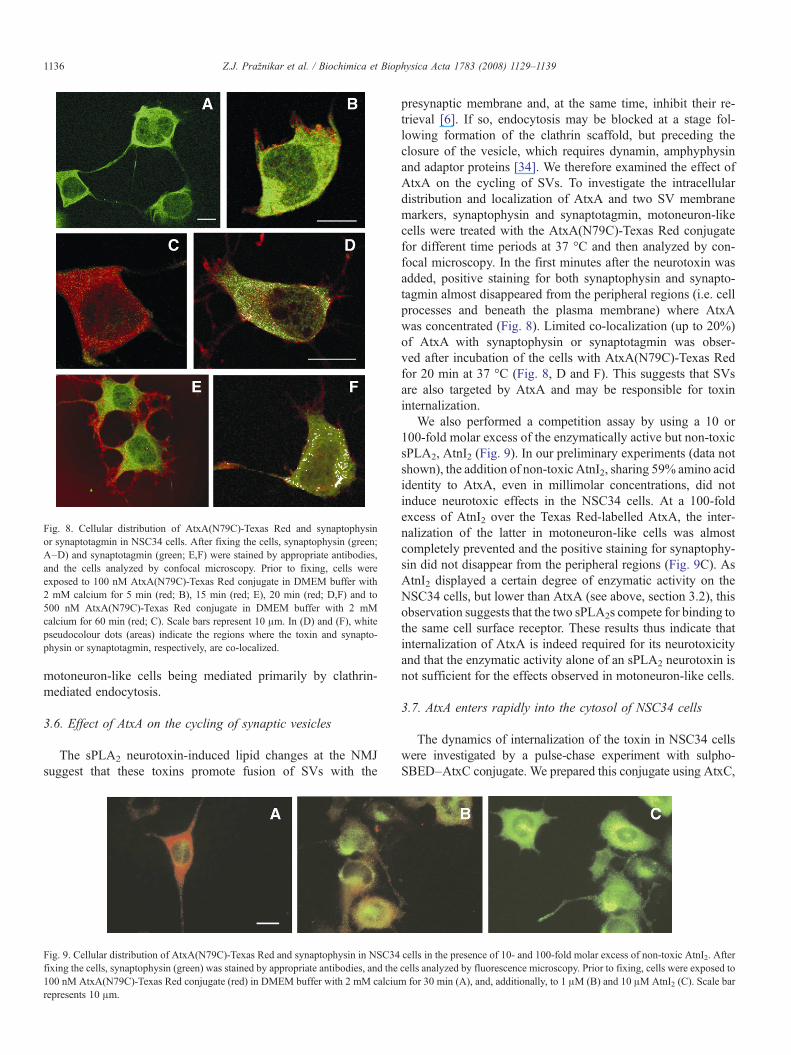
Fig. 8. Cellular distribution of AtxA(N79C)-Texas Red and synaptophysinor synaptotagmin in NSC34 cells. After fixing the cells, synaptophysin (green;A–D) and synaptotagmin (green; E,F) were stained by appropriate antibodies,and the cells analyzed by confocal microscopy. Prior to fixing, cells wereexposed to 100 nM AtxA(N79C)-Texas Red conjugate in DMEM buffer with2 mM calcium for 5 min (red; B), 15 min (red; E), 20 min (red; D,F) and to500 nM AtxA(N79C)-Texas Red conjugate in DMEM buffer with 2 mMcalcium for 60 min (red; C). Scale bars represent 10 μm. In (D) and (F), whitepseudocolour dots (areas) indicate the regions where the toxin and synapto-physin or synaptotagmin, respectively, are co-localized.
1136 Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
motoneuron-like cells being mediated primarily by clathrin-mediated endocytosis.
3.6. Effect of AtxA on the cycling of synaptic vesicles
The sPLA2 neurotoxin-induced lipid changes at the NMJsuggest that these toxins promote fusion of SVs with the
Fig. 9. Cellular distribution of AtxA(N79C)-Texas Red and synaptophysin in NSC34fixing the cells, synaptophysin (green) was stained by appropriate antibodies, and the100 nM AtxA(N79C)-Texas Red conjugate (red) in DMEM buffer with 2 mM calciurepresents 10 μm.
presynaptic membrane and, at the same time, inhibit their re-trieval [6]. If so, endocytosis may be blocked at a stage fol-lowing formation of the clathrin scaffold, but preceding theclosure of the vesicle, which requires dynamin, amphyphysinand adaptor proteins [34]. We therefore examined the effect ofAtxA on the cycling of SVs. To investigate the intracellulardistribution and localization of AtxA and two SV membranemarkers, synaptophysin and synaptotagmin, motoneuron-likecells were treated with the AtxA(N79C)-Texas Red conjugatefor different time periods at 37 °C and then analyzed by con-focal microscopy. In the first minutes after the neurotoxin wasadded, positive staining for both synaptophysin and synapto-tagmin almost disappeared from the peripheral regions (i.e. cellprocesses and beneath the plasma membrane) where AtxAwas concentrated (Fig. 8). Limited co-localization (up to 20%)of AtxA with synaptophysin or synaptotagmin was obser-ved after incubation of the cells with AtxA(N79C)-Texas Redfor 20 min at 37 °C (Fig. 8, D and F). This suggests that SVsare also targeted by AtxA and may be responsible for toxininternalization.
We also performed a competition assay by using a 10 or100-fold molar excess of the enzymatically active but non-toxicsPLA2, AtnI2 (Fig. 9). In our preliminary experiments (data notshown), the addition of non-toxic AtnI2, sharing 59% amino acididentity to AtxA, even in millimolar concentrations, did notinduce neurotoxic effects in the NSC34 cells. At a 100-foldexcess of AtnI2 over the Texas Red-labelled AtxA, the inter-nalization of the latter in motoneuron-like cells was almostcompletely prevented and the positive staining for synaptophy-sin did not disappear from the peripheral regions (Fig. 9C). AsAtnI2 displayed a certain degree of enzymatic activity on theNSC34 cells, but lower than AtxA (see above, section 3.2), thisobservation suggests that the two sPLA2s compete for binding tothe same cell surface receptor. These results thus indicate thatinternalization of AtxA is indeed required for its neurotoxicityand that the enzymatic activity alone of an sPLA2 neurotoxin isnot sufficient for the effects observed in motoneuron-like cells.
3.7. AtxA enters rapidly into the cytosol of NSC34 cells
The dynamics of internalization of the toxin in NSC34 cellswere investigated by a pulse-chase experiment with sulpho-SBED–AtxC conjugate. We prepared this conjugate using AtxC,
cells in the presence of 10- and 100-fold molar excess of non-toxic AtnI2. Aftercells analyzed by fluorescence microscopy. Prior to fixing, cells were exposed tom for 30 min (A), and, additionally, to 1 μM (B) and 10 μMAtnI2 (C). Scale bar

Fig. 10. Labelling and identification of the Atx-binding proteins in NSC34 cells with sulpho-SBED–AtxC. Cells were incubated with 100 nM sulpho-SBED–AtxC inthe dark and then exposed to UV light. Cell extracts were then prepared, the proteins separated on an avidin–Sepharose affinity column, and dissolved in reductiveSDS-PAGE sample buffer. After electrophoresis and Western blotting to a PVDF membrane, the biotin-containing bands were visualized by streptavidin–HRP-ECLdetection system (A,C) or using antibodies against 14-3-3 proteins (B) and CaM (D).
1137Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
a natural double mutant of AtxA, which is highly similar to AtxAin activity but shows much lower affinity for various surfacesincluding plasticware and glassware. On irradiation with UV, thephotoreactive AtxC conjugate is covalently attached to proteinsthat are in its close proximity, shown by transfer of the biotin labelto these molecules. Following the exposure of NSC34 cells tosulpho-SBED–AtxC, the photo-reaction was triggered in incuba-tion mixture aliquots at different points in time. Both cytosolicAtxC binding proteins, CaM (Fig. 10A,B) and 14-3-3 proteins(Fig. 10C,D), were biotinylated. The internalization of the probewas rapid, as labelling of CaM and 14-3-3 proteins was observed,respectively, after 2 and 5 min exposure of motoneuron-like cellsto the photoreactive conjugate. No interaction between the sPLA2
neurotoxin and the two cytosolic Atx-binding proteins wasobserved within the first minute of the treatment. In the case ofnon-neuronal HEK293 cells, labelling of CaM was only detectedif the cells were lysed before the sulpho-SBED–AtxC was addedbut not if the intact cells were exposed to the photo-probe. Thisindicates that internalization of Atx into the cytosol is a processspecific to neuronal cells.
4. Discussion
Despite a number of studies on the presynaptic toxicity ofsnake venom sPLA2s, the precise mode of their action at themolecular level is not fully understood. In the present in-vestigation, we approached this problem by investigating theeffect of Atxs on NSC34 cells, which show several phenotypicproperties of motoneurons including the ability to synthesize,
store and release ACh, and even to induce ACh receptor clusterson cocultured myotubes [21]. We demonstrated that this cell linemay also be used as a model of mouse motoneurons, whoseterminals are the natural target for snake sPLA2 neurotoxins.The application of nanomolar concentrations of AtxA to themotoneuron-like cells induced typical neurotoxic lesions closelyresembling those observed at the rat NMJ intoxicated by simi-lar sPLA2 neurotoxins, β-bungarotoxin, notexin and taipoxin[7,32,35].
AtxA was effective in degrading the plasma membranephospholipids of various cell types, with the motoneuron-likeNSC34 cells being the most susceptible of the three tested. Thesingle-site mutant of AtxA (V31W) was about an order ofmagnitude more effective than the wild-type AtxA in releasingfatty acids from the plasma membrane of intact cells and PCvesicles, but it showed quantitatively similar effects on NSC34cells on the disorganization of F-actin cytoskeleton and on SVs.Moreover, its toxicity in mice was even lower, by approximately6-fold (LD50 135 vs. 21 μg/kg), than that of the wild-type AtxA[26]. On the other hand, the enzymatically active but non-toxicAtnI2 was not able to induce any toxic effects on motoneuron-like cells. Based on our results, the (high) enzymatic activityof an sPLA2 neurotoxin is not alone sufficient to induce neu-rotoxicity. The recent finding that the N-terminal region ofanother presynaptic sPLA2 neurotoxin, OS2, is more importantfor central neurotoxicity than the enzymatic activity [36] isconsistent with our observation.
Following the presumed specific binding to a cell membranereceptor, AtxA is rapidly taken up by the motoneuron-like cells

1138 Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
and also found, at least to a certain extent, to be translocated intothe cytosol. These results are the first direct demonstration thatan sPLA2 is able to translocate from the extracellular space intothis cellular compartment. This confirms our suggestion thatAtxA arrives in the nucleus of the hippocampal neuron throughits cytosol [16]. β-bungarotoxin uptake into the soma of hippo-campal neurons [37] and fast taipoxin uptake into chromaffincells [31] have also been reported but their cytosolic localiza-tion was not established. Fluorescence microscopy showedgranular distribution of labelled AtxA within the motoneuron-like cells, suggesting vesicular internalization. The co-loca-lization of AtxA with clathrin observed in the process ofinternalization indicates that the endocytosis is largely clathrin-mediated. After a longer period of intoxication, formation oflarger AtxA-containing vesicles was observed. These resultswere confirmed by ultrastructural analysis using nanogold-labelled AtxA. The toxin derivative was seen in coated pits atthe plasma membrane, in endosomes, and in vesicles of dif-ferent sizes in the cytoplasm. The distribution of nanogold-labelled AtxA in the NSC34 cells was very similar to that of thegold-labelled tetanus toxin internalized by motoneuronal cellsfrom the rat spinal cord [38]. Internalization of AtxA wassignificantly enhanced by the presence of millimolar extra-cellular Ca2+, suggesting that the phospholipase activity ofAtxA, or some other Ca2+-dependent process, is important fortranslocation to occur. Limited co-localization of AtxA withtwo SV marker proteins, synaptophysin and synaptotagmin,indicates that a certain amount of the toxin may enter the cellsby recycling SVs at an early stage of intoxication, before thecomplete arrest of SV cycling. Although intracellular traffickingof the toxin is not yet understood, specific affinity photolabel-ling by Atx of two cytosolic proteins, CaM and 14-3-3 proteins,in the intact motoneuron-like cells showed that AtxA wastranslocated into the cytosol within a few minutes of into-xication. Indeed, a small amount of positive nanogold labellingwas localized in the cytosol. Furthermore, the toxin may alsobe targeted to certain organelles, such as mitochondria, butthis remains to be confirmed. The mitochondrial and nuclearlocalization of rat group IIA sPLA2 [39] and nuclear loca-lization of group V sPLA2 [40] may involve the translocationof these endogenous sPLA2s from the cytosol, as is indicated tobe the case of AtxA in our study. In general, the translocation of(endogenous mammalian) sPLA2s from vesicular structures,including certain organelles, to the cytosol and vice versa maythus be important for their function.
Our results do not fully support the hypothesis that sPLA2
neurotoxin-induced hydrolysis of the plasma membrane is suf-ficient for presynaptic neurotoxicity [6,41]. It appears ratherthat, at least in the case of AtxA, internalization of the sPLA2
neurotoxin is essential for the complete and specific neuro-toxic effect, as was confirmed by the competition assay. Weassume that AtxA is rapidly internalized into the motoneuron-like cells by receptor-dependent clathrin-mediated endocyto-sis and/or recycling SVs, and translocated from endosomes and/or SVs, which may be sponsored by its catalytic activity,into the cytosol and from there to certain organelles such asmitochondria. Intracellular trafficking of the toxin, its binding
to important cellular targets, such as CaM and 14-3-3 proteins,probably together with its still considerable enzymatic activityin the cytosol [16], finally lead to the irreversible blockade ofSV recycling, followed by further degeneration of the nerve cell.Very recently, the fusion of at least a fraction of the vesiclesendocytosed during synaptic activity with early endosomes inintact nerve terminals has been strongly indicated [42], pro-viding a further possible element in the toxin trafficking.Important and distinctive roles of CaM, 14-3-3 proteins and theF-actin cytoskeleton in both endo- and exocytosis, includingthat in neuronal cells, are well established [43–46].
In summary, our data provide the first experimental evidencethat an sPLA2 neurotoxin is internalized into motoneuron-likecells, where it is translocated into the cytosol and interacts withCaM and 14-3-3 proteins, and to certain organelles. Althoughthe real target and action of presynaptic sPLA2 toxins on motornerve terminals may be somewhat different from those observedin motoneuron-like cells, our results provide a firm basis for anexplanation of the molecular mechanism of presynaptic sPLA2
neurotoxicity in terms of the intracellular action of these toxins.Furthermore, the cytosolic translocation of endocytosed sPLA2,demonstrated here for the first time, opens up a new way to theunderstanding of many (patho)physiological effects of an arrayof endogenous sPLA2s on different mammalian cells.
Acknowledgements
We sincerely thank Prof. Neil R. Cashman, University ofToronto, Canada, for giving us permission to work on theNSC34 cells. We also thank Anita Štrekelj for constructing theAtxA(N79C) mutant, Uroš Logonder for providing anti-AtxAantibodies, Dr. Marko Kreft for helping us with confocalmicroscopy and Dr. Roger H. Pain for the critical reading of themanuscript. This work was supported by grant P1-0207-0106from the Slovenian Ministry of Higher Education, Science andTechnology, by NATOCollaborative Linkage Grant No. 980899and by an EMBO short-term fellowship (to Jože Pungerčar).
References
[1] I. Kudo, M. Murakami, Phospholipase A2 enzymes, Prostaglandins OtherLipid Mediat. 68–69 (2002) 3–58.
[2] R.M. Kini, Excitement ahead: structure, function and mechanism of snakevenom phospholipase A2 enzymes, Toxicon 42 (2003) 827–840.
[3] L.G. Thouin, A. Ritonja, F. Gubenšek, F.E. Russell, Neuromuscular andlethal effects of phospholipase A from Vipera ammodytes venom, Toxicon20 (1982) 1051–1058.
[4] C.Y. Lee, M.C. Tsai, Y.M. Chen, A. Ritonja, F. Gubenšek, Mode ofneuromuscular blocking action of toxic phospholipases A2 from Viperaammodytes venom, Arch. Int. Pharmacodyn. Ther. 268 (1984) 313–324.
[5] C.C. Chang, M.J. Su, Presynaptic toxicity of the histidine-modified,phospholipase A2-inactive, β-bungarotoxin, crotoxin and notexin, Toxicon20 (1982) 895–905.
[6] M. Rigoni, P. Caccin, S. Gschmeissner, G. Koster, A.D. Postle, O.Rossetto, G. Schiavo, C. Montecucco, Equivalent effects of snake PLA2
neurotoxins and lysophospholipid–fatty acid mixtures, Science 310 (2005)1678–1680.
[7] S. Prasarnpun, J. Walsh, J.B. Harris, β-bungarotoxin-induced depletion ofSVs at the mammalian NM junction, Neuropharmacology 47 (2004)304–314.

1139Z.J. Pražnikar et al. / Biochimica et Biophysica Acta 1783 (2008) 1129–1139
[8] J. Pungerčar, I. Križaj, Understanding the molecular mechanism under-lying the presynaptic toxicity of secreted phospholipases A2, Toxicon 50(2007) 871–892.
[9] N. Vučemilo, A. Čopič, F. Gubenšek, I. Križaj, Identification of a newhigh-affinity binding protein for neurotoxic phospholipases A2, Biochem.Biophys. Res. Commun. 251 (1998) 209–212.
[10] J. Šribar, A. Čopič, M. Poljšak-Prijatelj, J. Kuret, U. Logonder, F.Gubenšek, I. Križaj, R25 is an intracellular membrane receptor for a snakevenom secretory phospholipase A2, FEBS Lett. 553 (2003) 309–314.
[11] J. Šribar, A. Čopič, A. Pariš, N.E. Sherman, F. Gubenšek, J.W. Fox, I.Križaj, A high affinity acceptor for phospholipase A2 with neurotoxicactivity is a calmodulin, J. Biol. Chem. 276 (2001) 12493–12496.
[12] P. Prijatelj, J. Šribar, G. Ivanovski, I. Križaj, F. Gubenšek, J. Pungerčar,Identification of a novel binding site for calmodulin in ammodytoxin A, aneurotoxic group IIA phospholipase A2, Eur. J. Biochem. 270 (2003)3018–3025.
[13] J. Šribar, N.E. Sherman, P. Prijatelj, G. Faure, F. Gubenšek, J.W. Fox, A.Aitken, J. Pungerčar, I. Križaj, The neurotoxic phospholipase A2 associates,through a non-phosphorylated binding motif, with 14-3-3 protein γ and εisoforms, Biochem. Biophys. Res. Commun. 302 (2003) 691–696.
[14] J. Šribar, G. Anderluh, J.W. Fox, I. Križaj, Protein disulphide isomerasebinds ammodytoxin strongly: possible implications for toxin trafficking,Biochem. Biophys. Res. Commun. 329 (2005) 733–737.
[15] A.G. Singer, F. Ghomashchi, C. Le Calvez, J. Bollinger, S. Bezzine, M.Rouault, M. Sadilek, M. Lazdunski, G. Lambeau, M.H. Gelb, Interfacialkinetic and binding properties of the complete set of human and mousegroups I, II, V, X, and XII secreted phospholipases A2, J. Biol. Chem. 277(2002) 48535–48549.
[16] U. Petrovič, J. Šribar, A. Pariš, M. Rupnik, M. Kržan, N. Vardjan, F.Gubenšek, R. Zorec, I. Križaj, Ammodytoxin, a neurotoxic secretedphospholipase A2, can act in the cytosol of the nerve cell, Biochem.Biophys. Res. Commun. 324 (2004) 981–985.
[17] U. Petrovič, J. Šribar, M. Matis, G. Anderluh, J. Peter-Katalinić, I. Križaj, F.Gubenšek,Ammodytoxin, a secretory phospholipaseA2, inhibitsG2 cell cyclearrest in yeast Saccharomyces cerevisiae, Biochem. J. 391 (2005) 383–388.
[18] N. Hirokawa, Characterization of various nervous tissues of the chickembryos through responses to chronic application and immunocytochem-istry of β-bungarotoxin, J. Comp. Neurol. 180 (1978) 449–466.
[19] R. Pittman, R.W. Oppenheim, I.W. Chu-Wang, β-bungarotoxin inducedneuronal degeneration in the chick embryo spinal cord, Brain Res. 153(1978) 199–204.
[20] H. Rehm, T. Schäfer, H. Betz, β-bungarotoxin-induced cell-death ofneurons in chick retina, Brain Res. 250 (1982) 309–319.
[21] N.R. Cashman, H.D. Durham, J.K. Blusztajn, K. Oda, T. Tabira, I.T. Shaw,S. Dahrouge, J.P. Antel, Neuroblastoma x spinal cord (NSC) hybrid celllines resemble developing motor neurons, Dev. Dyn. 194 (1992) 209–221.
[22] F. Piccioni, P. Pinton, S. Simeoni, P. Pozzi, U. Fascio, G. Vismara, L.Martini, R. Rizzuto, A. Poletti, Androgen receptor with elongatedpolyglutamine tract forms aggregates that alter axonal trafficking andmitochondrial distribution in motoneuronal processes, FASEB J. 16 (2002)1418–1420.
[23] F.M. Menzies, M.R. Cookson, R.W. Taylor, D.M. Turnbull, Z.M.Chrzanowska-Lightowlers, L. Dong, D.A. Figlewicz, P.J. Shaw, Mito-chondrial dysfunction in a cell culture model of familial amyotrophiclateral sclerosis, Brain 125 (2002) 1522–1533.
[24] W. Wen, T. Sanelli, W. Ge, W. Strong, M.J. Strong, Activated microglialsupernatant induced motor neuron cytotoxicity is associated withupregulation of the TNFR1 receptor, Neurosci. Res. 55 (2006) 87–95.
[25] J. Pungerčar, I. Križaj, N.S. Liang, F. Gubenšek, An aromatic, but not abasic, residue is involved in the toxicity of group-II phospholipase A2
neurotoxins, Biochem. J. 341 (1999) 139–145.[26] T. Petan, I. Križaj, M.H. Gelb, J. Pungerčar, Ammodytoxins, potent
presynaptic neurotoxins, are also highly efficient phospholipase A2
enzymes, Biochemistry 44 (2005) 12535–12545.
[27] T. Petan, I. Križaj, F. Gubenšek, J. Pungerčar, Phenylalanine-24 in theN-terminal region of ammodytoxins is important for both enzymicactivity and presynaptic toxicity, Biochem. J. 363 (2002) 353–358.
[28] S. Bezzine, R.S. Koduri, E. Valentin, M. Murakami, I. Kudo, F.Ghomashchi, M. Sadilek, G. Lambeau, M.H. Gelb, Exogenously addedhuman group X secreted phospholipase A2 but not the group IB, IIA, andV enzymes efficiently release arachidonic acid from adherent mammaliancells, J. Biol. Chem. 275 (2000) 3179–3191.
[29] L. Kovačič, J. Šribar, I. Križaj, A new photoprobe for studying biologicalactivities of secreted phospholipases A2, Bioorg. Chem. 35 (2007)295–305.
[30] G. Eitzen, Actin remodeling to facilitate membrane fusion, Biochim.Biophys. Acta 1641 (2003) 175–181.
[31] P. Neco, O. Rossetto, A. Gil, C. Montecucco, L.M. Gutierrez, Taipoxininduces F-actin fragmentation and enhances release of catecholamines inbovine chromaffin cells, J. Neurochem. 85 (2003) 329–337.
[32] J.B. Harris, B.D. Grubb, C.A. Maltin, R. Dixon, The neurotoxicity of thevenom phospholipases A2, notexin and taipoxin, Exp. Neurol. 161 (2000)517–526.
[33] M. Rigoni, G. Schiavo, A.E. Weston, P. Caccin, F. Allegrini, M. Pennuto,F. Valtorta, C. Montecucco, O. Rossetto, Snake presynaptic neurotoxinswith phospholipase A2 activity induce punctate swellings of neurites andexocytosis of SVs, J. Cell Biol. 117 (2004) 3561–3570.
[34] O. Cremona, P. De Camilli, Synaptic vesicle endocytosis, Curr. Opin.Neurol. 7 (1997) 323–330.
[35] R.W. Dixon, J.B. Harris, Nerve terminal damage by β-bungarotoxin: itsclinical significance, Am. J. Pathol. 154 (1999) 447–455.
[36] M. Rouault, L.D. Rash, P. Escoubas, E. Boilard, J. Bolinger, B. Lomonte,T. Maurin, C. Guillaume, S. Canaan, C. Deregnaucourt, J. Schrevel, A.Doglio, J.M. Gutierrez, M. Lazdunski, M.H. Gelb, G. Lambeau,Neurotoxicity and other pharmacological activities of the snake venomphospholipase A2 OS2: the N-terminal region is more important thanenzymatic activity, Biochemistry 45 (2006) 5800–5816.
[37] M. Herkert, O. Shakhman, E. Schweins, C.M. Becker, β-bungarotoxin is apotent inducer of apoptosis in cultured rat neurons by receptor-mediatedinternalization, Eur. J. Neurosci. 14 (2001) 821–828.
[38] S. Roux, C. Colasante, C.S. Cloment, J. Barbier, T. Curie, E. Girard, J.Molgo, P. Brulet, Internalization of a GFP-tetanus toxin C-terminalfragment fusion protein at mature mouse neuromuscular junctions, Mol.Cell. Neurosci. 30 (2005) 572–582.
[39] L. Macchioni, L. Corazzi, V. Nardicchi, R. Mannucci, C. Arcuri, S.Porcellati, T. Sposini, R. Donato, G. Goracci, Rat brain cortexmitochondria release group II secretory phospholipase A2 under reducedmembrane potential, J. Biol. Chem. 279 (2004) 37860–37869.
[40] V. Nardicchi, L. Macchioni, M. Ferrini, G. Gorraci, The presence of asecretory phospholipase A2 in the nuclei of neuronal and glial cells of ratbrain cortex, Biochim. Biophys. Acta 1771 (2007) 1345–1352.
[41] O. Rossetto, L. Morbiato, P. Caccin, M. Rigoni, C. Montecucco,Presynaptic enzymatic neurotoxins, J. Neurochem. 97 (2006) 1534–1545.
[42] S.O. Rizzoli, I. Bethani, D. Zwilling, D. Wenzel, T.J. Siddiqui, D.Brandhorst, R. Jahn, Evidence for early endosome-like fusion of recentlyendocytosed synaptic vesicles, Traffic 7 (2006) 1163–1176.
[43] S. Quetglas, C. Iborra, N. Sasakawa, L. De Haro, K. Kumakura, K. Sato,C. Leveque, M. Seagar, Calmodulin and lipid binding to synaptobrevinregulates calcium-dependent exocytosis, EMBO J. 21 (2002)3970–3979.
[44] D. Roth, R.D. Burgoyne, Stimulation of catecholamine secretion fromadrenal chromaffin cells by 14-3-3 proteins is due to reorganisation of thecortical actin network, FEBS Lett. 37 (1995) 77–81.
[45] S. Sankaranarayanan, P.P. Atluri, T.A. Ryan, Actin has a molecularscaffolding, not propulsive, role in presynaptic function, Nat. Neurosci. 6(2003) 127–135.
[46] C. Dillon, Y. Goda, The actin cytoskeleton: integrating form and functionat the synapse, Annu. Rev. Neurosci. 28 (2005) 25–55.




















