
A Novel Bioluminescent Tumor Model of Human Renal Cancer Cell Lines: An In Vitro and In Vivo...
13
of August 4, 2014. This information is current as Function Negatively Regulates NKG2D-Mediated A Truncated Human NKG2D Splice Isoform Taku Kambayashi H. Bachmann, James R. Carlyle, Cynthia L. Baldwin and Mobin A. Karimi, Oscar A. Aguilar, Baixiang Zou, Michael ol.1400920 http://www.jimmunol.org/content/early/2014/08/01/jimmun published online 4 August 2014 J Immunol Material Supplementary DCSupplemental.html http://www.jimmunol.org/content/suppl/2014/08/01/content.1400920. Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2014 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology at Univ of Pennsylvania Library on August 4, 2014 http://www.jimmunol.org/ Downloaded from at Univ of Pennsylvania Library on August 4, 2014 http://www.jimmunol.org/ Downloaded from at Univ of Pennsylvania Library on August 4, 2014 http://www.jimmunol.org/ Downloaded from at Univ of Pennsylvania Library on August 4, 2014 http://www.jimmunol.org/ Downloaded from at Univ of Pennsylvania Library on August 4, 2014 http://www.jimmunol.org/ Downloaded from at Univ of Pennsylvania Library on August 4, 2014 http://www.jimmunol.org/ Downloaded from
Transcript of A Novel Bioluminescent Tumor Model of Human Renal Cancer Cell Lines: An In Vitro and In Vivo...

of August 4, 2014.This information is current as
FunctionNegatively Regulates NKG2D-Mediated A Truncated Human NKG2D Splice Isoform
Taku KambayashiH. Bachmann, James R. Carlyle, Cynthia L. Baldwin and Mobin A. Karimi, Oscar A. Aguilar, Baixiang Zou, Michael
ol.1400920http://www.jimmunol.org/content/early/2014/08/01/jimmun
published online 4 August 2014J Immunol
MaterialSupplementary
DCSupplemental.htmlhttp://www.jimmunol.org/content/suppl/2014/08/01/content.1400920.
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2014 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
A Truncated Human NKG2D Splice Isoform NegativelyRegulates NKG2D-Mediated Function
Mobin A. Karimi,*,† Oscar A. Aguilar,‡ Baixiang Zou,* Michael H. Bachmann,x
James R. Carlyle,‡ Cynthia L. Baldwin,*,1 and Taku Kambayashi†,1
NKG2D is a stimulatory receptor expressed by NK cells and a subset of T cells. NKG2D is crucial in diverse aspects of innate and
adaptive immune functions. In this study, we characterize a novel splice variant of humanNKG2D that encodes a truncated receptor
lacking the ligand-binding ectodomain. This truncated NKG2D (NKG2DTR) isoform was detected in primary human NK and
CD8+ T cells. Overexpression of NKG2DTR severely attenuated cell killing and IFN-g release mediated by full-length NKG2D
(NKG2DFL). In contrast, specific knockdown of endogenously expressed NKG2DTR enhanced NKG2D-mediated cytotoxicity,
suggesting that NKG2DTR is a negative regulator of NKG2DFL. Biochemical studies demonstrated that NKG2DTR was bound
to DNAX-activated protein of 10 kDa (DAP10) and interfered with the interaction of DAP10 with NKG2DFL. In addition,
NKG2DTR associated with NKG2DFL, which led to forced intracellular retention, resulting in decreased surface NKG2D
expression. Taken together, these data suggest that competitive interference of NKG2D/DAP10 complexes by NKG2DTR constitutes
a novel mechanism for regulation of NKG2D-mediated function in human CD8+ T cells and NK cells. The Journal of Immunology,
2014, 193: 000–000.
Natural killer group 2, member D (NKG2D) is a homo-dimeric, stimulatory type II transmembrane receptorexpressed by most NK cells, CD8+ T cells, gd T cells,
and NKT cells in humans (1, 2). NKG2D transmits activatingsignals through its interaction with the signal-transducing adaptormolecule, DNAX-activated protein of 10 kDa (DAP10), which cantrigger cell-mediated cytotoxicity and cytokine production. NKG2Drecognizes stress-inducible ligands structurally related to MHC class Imolecules including MHC class I chain–related gene (MIC)A, MICB,and UL16-binding protein 1,2,3,4 in humans and retinoic acid early-inducible (Rae)-1, H60, and murine UL16-binding protein–liketranscript 1 in mice. These ligands are minimally expressed by nor-mal tissues but can be upregulated by cellular stress and DNA damageresponse pathways, such as those initiated during viral infection, tu-morigenesis, and exposure to DNA alkylating chemotherapeuticagents (3–5).
NKG2D plays a crucial role in many aspects of immune functionand immune surveillance. For example, NKG2D blockade or de-
ficiency has been shown to enhance susceptibility toward inducible
and spontaneously growing tumors (6–9). NKG2D also contributes
to viral immunity. In CMV infection studies, deletion of virally
encoded proteins that prevent expression of NKG2D ligands
moderates virulence during early infection, an effect that can be
reversed by NKG2D blockade (10, 11). In addition to NK cells,
NKG2D also plays an important role in CD8+ T cell cytotoxicity.
Human CD8+ T cells activated ex vivo by TCR cross-linking in
the presence of IL-2 and IFN-g upregulate cytotoxic effector
molecules and potently lyse a broad range of malignant cell lines
and primary tumor samples (12–14). The cytotoxic effect of these
human CD8+ T cells is NKG2D dependent, because NKG2D
knockdown attenuates their killing activity (15).NKG2D gene expression is subject to posttranscriptional reg-
ulation by splicing variation (16). In mice, NKG2D isoformsgenerated by alternative mRNA splicing include long (NKG2D-L)and short (NKG2D-S) variants, which allows NKG2D-S to pairwith DAP12 in addition to DAP10 (17, 18). The human NKG2Dgene is expressed from at least three distinct alleles, and severalisoforms at the mRNA level have been described, including analternatively spliced variant that introduces a nonsense mutationresulting in a protein that lacks the entire extracellular ligandbinding domain (19). To date, the significance of this splicingvariation is not well understood, and the functional aspects of thehuman splice variants have not been analyzed.In this study, we characterized a novel splice variant of human
NKG2D in CD8+ T cells and NK cells. The splice variant encodes
a truncated form of NKG2D (NKG2DTR), which acts as a domi-
nant-negative regulator of NKG2D function by downregulating
full-length NKG2D (NKG2DFL) surface expression through in-
tracellular retention. Thus, NKG2DTR acts as a negative regulator
of NKG2D-mediated function. Our data provide novel insights
into the regulatory mechanisms of cytolysis as a potential op-
portunity to strengthen the broad antitumor potential of these cells.
*Immunology Research Group, Department of Veterinary and Animal Sciences,University of Massachusetts, Amherst, MA01003; †Department of Pathology andLaboratory Medicine, Perelman School of Medicine, University of Pennsylvania,Philadelphia, PA 19104; ‡Department of Immunology, University of Toronto, TorontoM4N 3M5, Canada; and xDepartment of Pediatrics, Stanford University School ofMedicine, Stanford, CA 94305
1C.L.B. and T.K. contributed equally to the manuscript.
Received for publication April 9, 2014. Accepted for publication July 9, 2014.
This work was supported by the Translational Center of Excellence in HematologicalMalignancies of the Abramson Cancer Center, University of Pennsylvania, and Na-tional Institutes of Health Grants R01HL107589 and R01HL111501. M.A.K. wassupported by National Institutes of Health Grant T32-HL-007775-20.
Address correspondence and reprint requests to Dr. Mobin A. Karimi and Dr. TakuKambayashi, Department of Pathology and Laboratory Medicine, 288 John Mor-gan Building, 3620 Hamilton Walk, Philadelphia, PA 19104. E-mail addresses:[email protected] (M.A.K.) and [email protected] (T.K.)
The online version of this article contains supplemental material.
Abbreviations used in this article: DAP10, DNAX-activated protein of 10 kDa; HA,hemagglutinin; MICA, MHC class I chain–related gene A; NKG2DFL, full-lengthNKG2D; NKG2DTR, truncated NKG2D variant; shRNA, short hairpin RNA; sMICA,soluble MICA.
Copyright� 2014 by The American Association of Immunologists, Inc. 0022-1767/14/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1400920
Published August 4, 2014, doi:10.4049/jimmunol.1400920 at U
niv of Pennsylvania Library on A
ugust 4, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

Materials and MethodsReagents, cell lines, and flow cytometry
Monoclonal Abs were purchased from eBioscience (San Diego, CA): anti-human OKT3, anti-CD28 clone CD28-6; BD Biosciences (San Jose, CA):anti-CD16 FITC clone 3G8 BD, anti CD56 PE clone B159, anti-CD314/NKG2D PE or allophycocyanin, BD clone 1D11, isotype controls (IgG)as either FITC or PE conjugates; R&D Systems (Minneapolis, MN): anti-human NKG2D clone 1 49810; Sigma-Aldrich (St. Louis, MO): anti–g-tubulin clone 4D11; Fisher Scientific (Pittsburgh, PA) anti-hemagglutinin(HA) HRP clone HA7, anti-Flag clone M2 HRP, anti-Myc HRP A5598;Santa Cruz Biotechnology (Santa Cruz, CA): anti-human NKG2D cloneN-20, NKG2D peptide N-20, DAP10 peptide; Abcam (Cambridge, MA):anti-human NKG2D 5C6. All cell culture reagents and chemicals werepurchased from Invitrogen (Grand Island, NY) and Sigma-Aldrich, re-spectively, unless otherwise specified. COS-7 cells and HEK293T cellswere cultured in complete DMEM (10% FCS; Invitrogen), 2 mM L-glu-tamine, 100 U/ml penicillin, and 100 mg/ml streptomycin). The P815murine mastocytoma cell line was purchased from the American TypeCulture Collection (Manassas, VA) and cultured as recommended. Flowcytometry was performed on Ab-labeled or transduced cells by a BD LSR-II instrument (BD Biosciences). Transduced cells were cell sorted witha FACSAria (BD Biosciences).
Generation of activated CD8+ T cells and NK cells
Human PBLs were obtained from buffy coats of healthy donors by Ficoll–Hypaque density centrifugation. CD8+ T cells were enriched with CD8-coated MicroBeads, followed by cell sorting by FACS. CD8+ T cells wereactivated either with plate-bound anti-CD3 plus anti-CD28 (10 ng/ml) orwith 1000 U/ml human IFN-g (Genentech, South San Francisco), followedon day 1 by stimulation with anti-CD3 (25 ng/ml; OrthoBioTech, (Hor-sham, PA) and recombinant human IL-2 (300 U/ml; Chiron, (Emeryville,CA) as described previously (20). Every 3–5 d thereafter, new completeRPMI 1640 medium was added and supplemented with IL-2 (300 U/ml) tomaintain a cell density of 1.5–23 106/ml for a total of 14 d. NK cells werepositively selected by incubating PBMCs with CD16-coated MicroBeadsusing MACS (Miltenyi Biotec, Auburn, CA). NK cells were further pu-rified by cell sorting for the CD56+ population as described previously(15). Consent was obtained from all donors of PBL buffy coats underprotocols approved by the Institutional Review Board at the University ofMassachusetts (IRB2011-1051).
PCR and sequence analysis of NKG2D cDNA
RNA was isolated from purified CIK cells using the RNeasy Mini Kit(Qiagen) and reverse transcribed with the iScript cDNA Synthesis Kit (Bio-Rad). Primers used for PCR amplification of NKG2D cDNAs are 59-CTGGGAGATGAGTGAATTTCATA-39 (forward) and 59-GACTTCAC-CAGTTTAAGTAAATC-39 (reverse) PCR amplification was performed asdescribed previously (16). PCR products were separated by agarose gelelectrophoresis and subsequently cloned into TA vectors (Invitrogen).Cloned inserts were sequenced with an automated DNA sequence analyzer.DNA sequences of human NKG2D splice variants were analyzed usingVector NTI software (Invitrogen). Primers used for NKG2DTR are 59-ACGATGGCAAAAGCAAAGATGTCC-39 (forward) and 59-ACGGATT-CCCATGGCTACAGCG-39 (reverse).
Specific knockdown of the NKG2DTR alternative splice variantby short hairpin RNA
The oligonucleotide sequence 59-TCCAGAAATTTGTAAACTA-39 (136–155 bp of intron 4) was designed to target a unique position within intron 4specifically retained in NKG2DTR mRNA but has no sequence homologyto NKG2DFL mRNA as determined by basic local alignment search toolsequence analysis. This sequence was incorporated within a short hairpinstructure, using the stem loop sequence 59-TCAAGAGA-39, and cloned inthe plasmid pSICO-GFP, as described previously (21). HEK293T cellswere transfected using the calcium phosphate method with 20 mg pSICOlentivirus, 10 mg vesicular stomatitis virus G plasmid, and 15 mgCMVDR8.74, according to standard protocols. After 16 h, the medium waschanged, and recombinant lentivirus vectors were harvested 24–48 h later.Lentiviral infection of activated CD8+ T cell or NK cell was performedthree times at 24-h intervals. For each infection, cells were plated in 48-well plates at 1 3 105 cells/well and infected in the presence of protamineand hexadimethrin. Spin infection was performed at 1200 rpm for 90 minat 37˚C. Four days after the first infection, transduced cells were isolatedby FACS sorting of GFP+ cells to .97% purity.
Generation of soluble MHC class I chain–related gene A
The cDNA encoding the human NKG2D ligand MICAwas obtained by RT-PCR on RNA from normal human B cells and subcloned into thepFUSE-hFC1 plasmid (Invivogen) to create an MHC class I chain–related gene A (MICA)–Fc-fusion protein between MICA and the Fcregion (CH2 and CH3 domains) of the human IgG1 H chain. HEK293Tcells were transduced with the pFUSE-hFC1-MICA plasmid using Lipo-fectamine 2000. Supernatants containing soluble MICA (sMICA)-Fcfusion protein from transfected HEK293T cells were collected and sMICA-Fc was concentrated using Millipore ultra centrifuge filter (10k) and wasfurther purified by an Fc-binding column (Hiprop; GE Healthcare lifeScience). The purified sMICA was used to stimulate CD8+ T cells.
IFN-g production by activated CD8+ T cells
CD8+ T cells were retrovirally transduced with pQCN-NKG2DFL-Flag-IRES-GFP or pQCN-NKG2DTR-myc-IRES-GFP. Transduced cells were sortedfor GFP, and 5 3 106 sorted cells were cultured as described earlier.Cells were washed and stimulated with plate-bound anti-NKG2D(5 mg/ml), sMICA (10 mg/ml), or anti-CD3 (10 mg/ml). Twelve hourslater, cell-free supernatants were collected, and IFN-g content wasmeasured by ELISA (eBioscience), according to the manufacturer’sinstructions.
Expression vector, retrovirus, and lentivirus construction andtransduction
Flag-tagged NKG2DFL and Myc-tagged NKG2DTR were cloned intopcDNATOPO (Invitrogen, Carlsbad, CA) and subcloned into the retroviralvector pQCNXIX (BD Clontech, Mountain View, CA) generating pQCN-NKG2DFL-Flag-IRES-GFP and pQCN-NKG2DTR-myc-IRES-GFP. In addi-tion, Myc-tagged NKG2DTR and HA-tagged DAP10 were cloned into pC-GFP plasmid (BD Clontech) to create in-frame C-terminal GFP fusionproteins and subcloned into the retroviral vectors pQCNXIX, pQCN-NKG2DTR-myc-IRES-GFP, or pQCN-NKG2DFL-Flag-IRES-GFP generatingthe constructs pQCN-DAP10HA = GFP, pQCN-NKG2DFL-Flag-IRES-NKG2DTR-Myc = GFP, NKG2DFL-Flag-IRES-DAP10HA = GFP, or NKG2DTR-Myc-IRES-DAP10HA.GFP. DAP10HA also was cloned into the pLVX-Dsred-Monomer-N-1 (BD Clontech) to create an in-frame N-terminal RFP fusionprotein. DAP10HA = RFP was subcloned into the retroviral vector pQCNXIXgenerating pQCN-DAP10HA = RFP. To create retroviral vector particles,Phoenix A cells were transfected with these retroviral vector plasmids usingLipofectamine 2000 (Invitrogen). Forty-eight to 72 h posttransfection,supernatants were collected and, along with 5 mg/ml protamine sulfate, addedto CD8+ T cells. Cells were centrifuged for 90 min at 200 3 g, and resus-pended in RPMI 1640 medium. The transduction efficiency ranged from 20to 40%. Transduced cells were sorted by flow cytometry for expression ofGFP, RFP, Flag-tag, myc-tag, or NKG2D C terminus using a specific Ab. Inexperiments where all three proteins were transduced (NKG2DFL, NKG2DTR,and DAP10), the cells were transduced with two separate retroviruses.In experiments involving transient transfections, HEK293T and COS-7cells were transfected with the constructs using Lipofectamine 2000.pcDNA3.1 vectors encoding NKG2DFL-Flag, NKG2DTR-Myc, and DAP10HA
were used in transient transfection experiments using COS-7 and HEK293Tcells.
51Cr release cytotoxicity assay
P815 cells were labeled with 51Cr (DuPont-NEN, Boston, MA), washed,resuspended in complete RPMI 1640 medium, and plated in 96-well platesat a concentration of 1 3 104 cells/ml in triplicates. Anti-NKG2D Ab(5 mg/ml) was added and incubated for 30 min before washing and plating.Effector cells were added at specified ratios and incubated at 37˚C for 4 h.At the end of each assay, supernatants were collected and counted usinga gamma counter (Cobra/AII). Cytotoxicity was calculated by the fol-lowing equation: percent-specific lysis = 100 3 ([test release] 2 [spon-taneous release])/([maximal release] 2 [spontaneous release]).
Western blotting and immunoprecipitation
Cells were lysed in freshly prepared lysis buffer (10 mM Tris [pH 8],150 mM NaCl, 1 mM EDTA, 1% Nonidet-P40, 0.5% deoxycholate, 0.1%SDS, complete Protease Inhibitor Cocktail [Roche, Palo Alto, CA], and 500mM PMSF) and centrifuged for 10 min at 4˚C. Aliquots containing 70 mgprotein were separated on an 18% denaturing polyacrylamide gel andtransferred to polyvinylfluoride membranes for immunoblot analysis usingAbs specific to Flag, myc, HA, or different regions of NKG2D. Inexperiments involving peptide blockade, anti-NKG2D Ab (1 mg/ml) wasmixed with either specific (anti-NKG2D peptide N-20:P; Santa Cruz
2 AN NKG2D SPLICE VARIANT REGULATES NKG2D FUNCTION
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

Biotechnology) or nonspecific peptide (Abcam aggrecan peptide;Abcam) at 2 mg/ml in 2 ml blocking buffer. After agitation overnight at4˚C, Western blot analysis was performed on the cell lysates. For immu-noprecipitation, Flag-tagged, myc-tagged, and HA-tagged proteins inlysates of transduced cells were recovered by incubation with 10 mganti-Flag, anti-myc, or anti-HA Ab for 2 h at 4˚C, followed by bindingto protein G–agarose beads (Pierce). After washing, immunoprecipi-tated lysates were separated and resolved by Western blot analysis asdescribed above.
Immunofluorescence-based cell imaging
COS-7 cells (CRL-1651; American Type Culture Collection) were trans-fected using Extreme Gene (Roche) following the manufacturer’sinstructions. Transfected cells were plated on glass coverslips coated with10 mg/ml bovine fibronectin (Sigma-Aldrich). Cells were fixed in PBSwith 3.7% formaldehyde, permeabilized in PBS containing 0.5% TritonX-100, blocked with PBS containing 1% BSA and 10% goat serum, fol-lowed by incubation with anti-Flag Ab. Cells were washed and stainedwith goat anti-mouse Dilight-488 secondary Ab. Nuclei were stained usingDAPI. Cells were mounted in Vectashield (Vector) and imaged usinga Zeiss 200M inverted microscope with the Axiovision software.
Statistical analysis
Statistical significant differences in the observed mean values of experi-mental groups were determined by ANOVA using Excel (Microsoft, Seattle,WA) or Prism computer software (GraphPad, San Diego, CA).
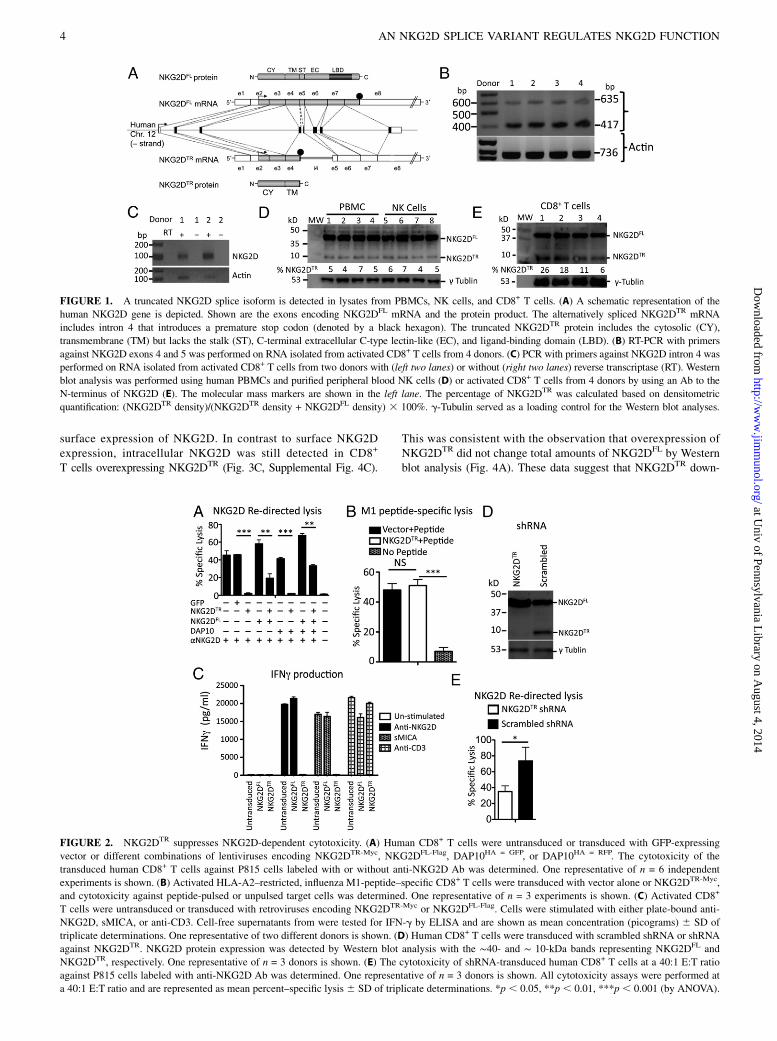
ResultsIdentification of an NKG2D splice isoform in human CD8+
T cells and NK cells
Human NKG2D is encoded by a single gene consisting of 10 exonslocated in the NK gene complex (22). Exons 2–4 encode the
intracellular/transmembrane domain, followed by exons 5–8 that
encode the ligand-binding ectodomain (Fig. 1A) (16). Analysis of
NKG2D gene transcripts expressed from cells in human peripheral
blood identified an additional unexpected NKG2D mRNA transcript
by RT-PCR using primers specific to regions in exons 4 and 5
(Fig. 1B). DNA sequence analysis demonstrated that the unexpected
PCR product resulted from an alternatively spliced NKG2D tran-
script that included intron 4 retained between exons 4 and 5.
This intron encoded a translational stop codon, potentially creating
a truncated form of NKG2D (NKG2DTR) without the extracellular
ligand-binding domain (exons 5–8; Fig. 1A). PCR using primers
specific to regions in intron 4 showed a band in mRNA from CD8+
T cells only in the presence of reverse transcriptase, confirming the
presence of NKG2D mRNA with retained intron 4 (Fig. 1C).To determine whether a truncated protein corresponding to
this transcript was detectable in these cells, we probed forNKG2D protein in PBMCs, NK cells, and activated CD8+
T cells. Using an NKG2D N terminus–specific Ab, Western blotanalysis revealed two bands with molecular masses of ∼40 and∼10 kDa, corresponding to the expected sizes of NKG2DFL andNKG2DTR, respectively in PBMCs, CD8+ T cells, and NK cells(Fig. 1D, 1E). Both bands corresponded to specific recognitionby the Ab, because the addition of an NKG2D derived but nota nonspecific peptide specifically blocked detection of bothbands (Supplemental Fig. 1). Next, we investigated whether therelative abundance of the truncated NKG2D isoform is influ-enced by cell activation. CD8+ T cells from three differentdonors were stimulated using anti-CD3 in the presence of IFN-gand IL-2. Cell lysates from CD8+ T cells activated for 0, 24, 48,or 72 h were analyzed for NKG2DFL and NKG2DTR proteinexpression. Western blot analysis showed that the relative per-centage of NKG2DTR to NKG2DFL did not change after acti-vation (Supplemental Fig 2).
NKG2DTR suppresses NKG2D-dependent function
Given that NKG2DTR lacks an extracellular domain, we predictedthat expression of NKG2DTR would impact the function ofNKG2DFL. To test the role of NKG2DTR in NKG2D-dependentcytotoxicity, we used a gain and loss of function approach in CD8+
T cells. Overexpression of NKG2DTR significantly decreasedNKG2D-dependent cytotoxicity (redirected lysis by anti-NKG2DAb) by activated human CD8+ T cells. Decreased cytotoxicity byNKG2DTR was observed even when NKG2DFL and DAP10 wereoverexpressed in these cells (Fig. 2A, Supplemental Fig. 3B).Similar results were obtained with cytotoxicity against K562 cells,which endogenously express the NKG2D ligands MICA and MICB(Supplemental Fig. 3B). Importantly, the transduction of activatedCD8+ T cells with NKG2DTR was not associated with global ad-verse effects on cytotoxic function, because TCR-dependent lysis ofHLA-A2–restricted influenza M1 peptide–specific activated CD8+
T cells against Ag-pulsed target cells was not affected by trans-duction with NKG2DTR (Fig. 2B).Because NKG2D induces cytokine production from activated
CD8+ T cells, we next examined whether NKG2DTR impactsNKG2D-mediated IFN-g release by CD8+ T cells. CD8+ T cellswere retrovirally transduced with either NKG2DTR or NKG2DFL
and stimulated with plate-immobilized anti-NKG2D, sMICA, oranti-CD3 for 12 h. Compared with CD8+ T cells untransduced ortransduced with NKG2DFL, CD8+ T cells overexpressing NKG2DTR
produced little IFN-g in response to NKG2D stimulation (Fig. 2C).In contrast, anti-CD3 stimulation induced similar IFN-g levels in allCD8+ T cells, suggesting that the inhibitory effect of NKG2DTR isspecific to NKG2D stimulation.To determine whether the endogenous NKG2DTR splice variant
physiologically regulates endogenous NKG2DFL function, wedesigned short hairpin RNA (shRNA) sequences to specificallyknockdown NKG2DTR expression while leaving NKG2DFL ex-pression intact. Because the NKG2DTR splice variant sequenceretains intron 4, we targeted a unique position within this regionthat shares no homology with the NKG2DFL sequence (intron 4,136–155 bp). Control and experimental shRNA oligonucleotideswere delivered by lentiviral vectors into activated human CD8+
T cells. Immunoblotting analyses confirmed specific knockdownof the endogenous NKG2DTR protein because there was a 78–93%reduction of the NKG2DTR to NKG2DFL ratio in cells transducedwith experimental shRNA compared with scrambled shRNA(Fig. 2D). Specific knockdown of endogenous NKG2DTR in ac-tivated CD8+ T cells resulted in significantly enhanced cytotox-icity mediated by NKG2DFL (Fig. 2E). Thus, NKG2DTR acts asa physiologic dominant-negative regulator of NKG2DFL-depen-dent cytotoxicity.
NKG2DTR downregulates surface expression of NKG2DFL
To determine how NKG2DTR negatively regulates NKG2DFL, weexamined the cell surface expression of NKG2D on activatedCD8+ T cells after transduction with different combinations ofNKG2DTR, NKG2DFL, and/or DAP10. Activated CD8+ T cellsoverexpressing NKG2DTR, with or without DAP10 overexpression,showed a significant reduction in cell surface NKG2D expres-sion (Supplemental Fig. 4A). Importantly, the overexpression ofNKG2DFL and/or DAP10 was unable to overcome the negativeeffect of NKG2DTR on NKG2D cell surface expression (Fig.3A, Supplemental Fig. 4A). Next, NKG2DTR was specificallyknocked down in CD8+ T cells and cell surface NKG2D wasassessed. Silencing of NKG2DTR significantly increased sur-face expression of NKG2DFL (Fig. 3B, Supplemental Fig. 4B).These data suggest that NKG2DTR negatively regulates cell
The Journal of Immunology 3
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
surface expression of NKG2D. In contrast to surface NKG2Dexpression, intracellular NKG2D was still detected in CD8+
T cells overexpressing NKG2DTR (Fig. 3C, Supplemental Fig. 4C).
This was consistent with the observation that overexpression ofNKG2DTR did not change total amounts of NKG2DFL by Westernblot analysis (Fig. 4A). These data suggest that NKG2DTR down-
FIGURE 1. A truncated NKG2D splice isoform is detected in lysates from PBMCs, NK cells, and CD8+ T cells. (A) A schematic representation of the
human NKG2D gene is depicted. Shown are the exons encoding NKG2DFL mRNA and the protein product. The alternatively spliced NKG2DTR mRNA
includes intron 4 that introduces a premature stop codon (denoted by a black hexagon). The truncated NKG2DTR protein includes the cytosolic (CY),
transmembrane (TM) but lacks the stalk (ST), C-terminal extracellular C-type lectin-like (EC), and ligand-binding domain (LBD). (B) RT-PCR with primers
against NKG2D exons 4 and 5 was performed on RNA isolated from activated CD8+ T cells from 4 donors. (C) PCR with primers against NKG2D intron 4 was
performed on RNA isolated from activated CD8+ T cells from two donors with (left two lanes) or without (right two lanes) reverse transcriptase (RT). Western
blot analysis was performed using human PBMCs and purified peripheral blood NK cells (D) or activated CD8+ T cells from 4 donors by using an Ab to the
N-terminus of NKG2D (E). The molecular mass markers are shown in the left lane. The percentage of NKG2DTR was calculated based on densitometric
quantification: (NKG2DTR density)/(NKG2DTR density + NKG2DFL density) 3 100%. g-Tubulin served as a loading control for the Western blot analyses.
FIGURE 2. NKG2DTR suppresses NKG2D-dependent cytotoxicity. (A) Human CD8+ T cells were untransduced or transduced with GFP-expressing
vector or different combinations of lentiviruses encoding NKG2DTR-Myc, NKG2DFL-Flag, DAP10HA = GFP, or DAP10HA = RFP. The cytotoxicity of the
transduced human CD8+ T cells against P815 cells labeled with or without anti-NKG2D Ab was determined. One representative of n = 6 independent
experiments is shown. (B) Activated HLA-A2–restricted, influenza M1-peptide–specific CD8+ T cells were transduced with vector alone or NKG2DTR-Myc,
and cytotoxicity against peptide-pulsed or unpulsed target cells was determined. One representative of n = 3 experiments is shown. (C) Activated CD8+
T cells were untransduced or transduced with retroviruses encoding NKG2DTR-Myc or NKG2DFL-Flag. Cells were stimulated with either plate-bound anti-
NKG2D, sMICA, or anti-CD3. Cell-free supernatants from were tested for IFN-g by ELISA and are shown as mean concentration (picograms) 6 SD of
triplicate determinations. One representative of two different donors is shown. (D) Human CD8+ T cells were transduced with scrambled shRNA or shRNA
against NKG2DTR. NKG2D protein expression was detected by Western blot analysis with the ∼40- and ∼ 10-kDa bands representing NKG2DFL and
NKG2DTR, respectively. One representative of n = 3 donors is shown. (E) The cytotoxicity of shRNA-transduced human CD8+ T cells at a 40:1 E:T ratio
against P815 cells labeled with anti-NKG2D Ab was determined. One representative of n = 3 donors is shown. All cytotoxicity assays were performed at
a 40:1 E:T ratio and are represented as mean percent–specific lysis 6 SD of triplicate determinations. *p , 0.05, **p , 0.01, ***p , 0.001 (by ANOVA).
4 AN NKG2D SPLICE VARIANT REGULATES NKG2D FUNCTION
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

regulates surface NKG2D expression by retaining NKG2DFL intra-cellularly.
NKG2DTR pairs with DAP10 and NKG2DFL, interferes withthe association of DAP10 with NKG2DFL, and retainsNKG2DFL in the intracellular compartment
We reasoned that NKG2DTR could attenuate cell surface expres-sion of NKG2D by competing for binding to DAP10 and/or bypairing with NKG2DFL. To test these possibilities, NKG2DTR wasoverexpressed, and the ability of DAP10 to coimmunoprecipitatewith NKG2DFL was assessed. Compared with vector-transduced
CD8+ T cells, NKG2DTR overexpression markedly decreasedbinding of NKG2DFL with DAP10 (Fig. 4A). Conversely, knock-down of NKG2DTR resulted in enhanced association of NKG2DFL
with DAP10 (Fig. 4B).Next, to test whether NKG2DTR associates with DAP10 and/or
form heterodimers with NKG2DFL, the three constructs weretransiently transfected into HEK293T cells. NKG2DFL coimmu-noprecipitated with NKG2DTR, indicating that NKG2DFL andNKG2DTR can form heterodimers (Fig. 5A, 5B). DAP10 also wascoimmunoprecipitated with both NKG2DFL and NKG2DTR (Fig. 5B,5C), indicating that both isoforms are able to associate withDAP10. The expression of NKG2DTR did not affect the associa-tion of DAP10 with NKG2DFL in these experiments, suggestingthat overexpression of DAP10 overcomes the inhibition of theNKG2DFL/DAP10 interaction by NKG2DTR seen in CD8+ T cells(Fig. 4A). Alternatively, these differences may arise because thelatter immunoprecipitation studies (Fig. 5) were performed withHEK293T cells instead of activated CD8+ T cells. However,DAP10 overexpression was insufficient to reverse NKG2DTR-mediated inhibition of NKG2D surface expression (Fig. 3A) orrestore NKG2D-mediated cytotoxic activity of CD8+ T cells(Fig. 2A). Thus, these data suggest that NKG2DTR interferencewith the DAP10/NKG2DFL interaction is only partially respon-sible for the reduced NKG2DFL surface expression by NKG2DTR
cannot fully explain why cell surface expression of NKG2D isdecreased by NKG2DTR.To determine the intracellular localization of the full-length
and truncated NKG2D molecules, we overexpressed NKG2DTR
and/or NKG2DFL with DAP10 and membrane-targeted mCherryin COS-7 cells. Similar to the expression pattern seen in CD8+
T cells, only NKG2DFL but not NKG2DTR was found at the cellsurface of transduced COS-7 cells (Fig. 6A, Supplemental Fig.4D). Using anti-Flag Abs to detect NKG2DFL/FLAG, we foundthat the distribution of NKG2DFL was consistent with that ofa typical transmembrane protein; the majority of NKG2DFL wasclustered at the cell membrane with some perinuclear fluores-cence (Golgi) observed (Fig. 6B). In contrast, in cells expressingboth NKG2DFL and NKG2DTR, the staining was restricted toperinuclear localization, and almost no surface membrane
FIGURE 3. NKG2DTR downregulates surface expression of NKG2DFL. (A) Human CD8+ T cells were untransduced, transduced with vector (GFP), or
with different combinations of retroviruses encoding NKG2DFL-Flag, NKG2DTR-myc = GFP, NKG2DTR-myc, and DAP10HA = GFP and analyzed for NKG2DFL
surface expression. NKG2DTR-myc = GFP was used in double transduction of NKG2DTR and NKG2DFL. Filled histograms and solid lines represent cells
stained with IgG control Ab and anti-NKG2D, respectively. One representative of n = 6 independent experiments is shown. (B) Human CD8+ T cell cells
were transduced with vectors expressing either scrambled shRNA (solid line) or NKG2DTR shRNA (dotted line) and analyzed for NKG2DFL surface
expression. The filled histograms represent cells stained with IgG control Ab. One representative of n = 3 donors is shown. (C) Human CD8+ T cells were
transduced with GFP alone or with NKG2DTR-Myc. Surface and intracellular NKG2D was detected by flow cytometry. Filled histograms and solid lines
represent cells stained with IgG control Ab and anti-NKG2D, respectively. One representative of n = 3 experiments is shown.
FIGURE 4. NKG2DTR does not change total NKG2DFL expression but
blocks the interaction of NKG2DFL with DAP10. (A) Human CD8+ T cells
were transduced with either vector (GFP alone) or NKG2DTR. Total cell
lysates were blotted for NKG2D, g-tubulin (loading control), or DAP10.
Cell lysates were also immunoprecipitated (IP) with anti-NKG2DFL Ab
and immunoblotted (IB) with Ab against NKG2DFL or anti-DAP10. One
representative of n = 3 independent experiments is shown. (B) Total cell
lysates from CD8+ T cells untransduced (left lane), transduced with
scrambled shRNA (middle lane), and NKG2DTR shRNAvector (right lane)
were blotted for NKG2D, g-tubulin (loading control), or DAP10. Cell
lysates also were IP with anti-NKG2DFL Ab and IB with Ab against
NKG2DFL or anti-DAP10. One representative of n = 3 donors is shown.
The Journal of Immunology 5
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
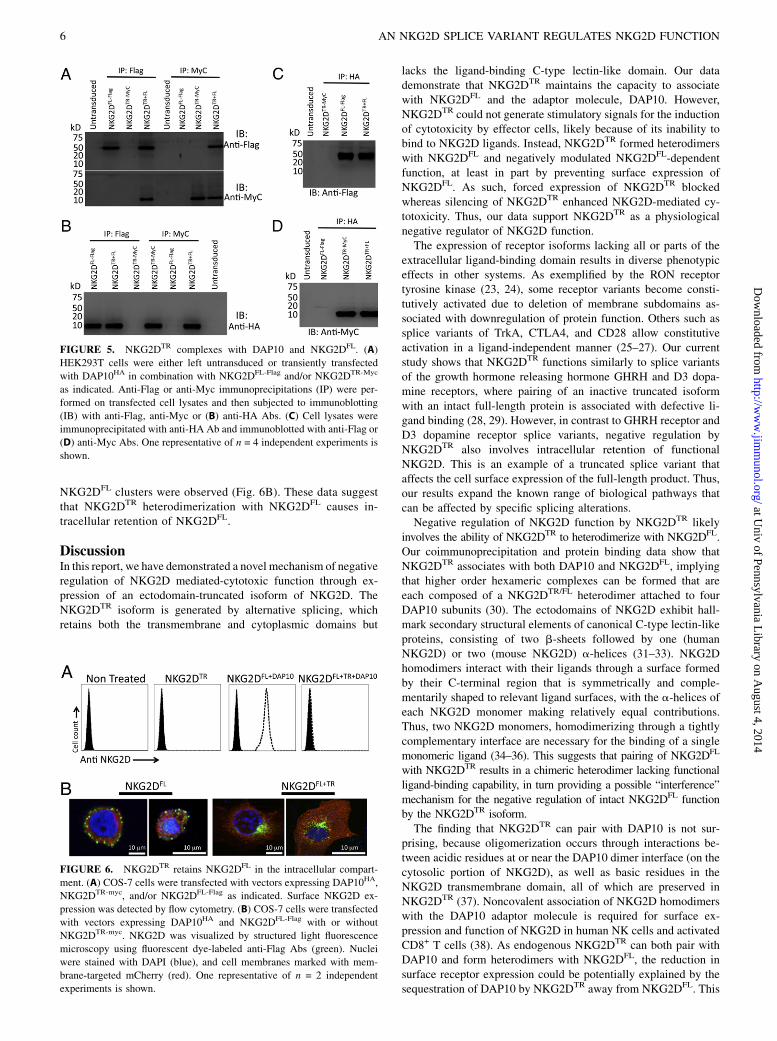
NKG2DFL clusters were observed (Fig. 6B). These data suggestthat NKG2DTR heterodimerization with NKG2DFL causes in-tracellular retention of NKG2DFL.
DiscussionIn this report, we have demonstrated a novel mechanism of negativeregulation of NKG2D mediated-cytotoxic function through ex-pression of an ectodomain-truncated isoform of NKG2D. TheNKG2DTR isoform is generated by alternative splicing, whichretains both the transmembrane and cytoplasmic domains but
lacks the ligand-binding C-type lectin-like domain. Our datademonstrate that NKG2DTR maintains the capacity to associatewith NKG2DFL and the adaptor molecule, DAP10. However,NKG2DTR could not generate stimulatory signals for the inductionof cytotoxicity by effector cells, likely because of its inability tobind to NKG2D ligands. Instead, NKG2DTR formed heterodimerswith NKG2DFL and negatively modulated NKG2DFL-dependentfunction, at least in part by preventing surface expression ofNKG2DFL. As such, forced expression of NKG2DTR blockedwhereas silencing of NKG2DTR enhanced NKG2D-mediated cy-totoxicity. Thus, our data support NKG2DTR as a physiologicalnegative regulator of NKG2D function.The expression of receptor isoforms lacking all or parts of the
extracellular ligand-binding domain results in diverse phenotypiceffects in other systems. As exemplified by the RON receptortyrosine kinase (23, 24), some receptor variants become consti-tutively activated due to deletion of membrane subdomains as-sociated with downregulation of protein function. Others such assplice variants of TrkA, CTLA4, and CD28 allow constitutiveactivation in a ligand-independent manner (25–27). Our currentstudy shows that NKG2DTR functions similarly to splice variantsof the growth hormone releasing hormone GHRH and D3 dopa-mine receptors, where pairing of an inactive truncated isoformwith an intact full-length protein is associated with defective li-gand binding (28, 29). However, in contrast to GHRH receptor andD3 dopamine receptor splice variants, negative regulation byNKG2DTR also involves intracellular retention of functionalNKG2D. This is an example of a truncated splice variant thataffects the cell surface expression of the full-length product. Thus,our results expand the known range of biological pathways thatcan be affected by specific splicing alterations.Negative regulation of NKG2D function by NKG2DTR likely
involves the ability of NKG2DTR to heterodimerize with NKG2DFL.Our coimmunoprecipitation and protein binding data show thatNKG2DTR associates with both DAP10 and NKG2DFL, implyingthat higher order hexameric complexes can be formed that areeach composed of a NKG2DTR/FL heterodimer attached to fourDAP10 subunits (30). The ectodomains of NKG2D exhibit hall-mark secondary structural elements of canonical C-type lectin-likeproteins, consisting of two b-sheets followed by one (humanNKG2D) or two (mouse NKG2D) a-helices (31–33). NKG2Dhomodimers interact with their ligands through a surface formedby their C-terminal region that is symmetrically and comple-mentarily shaped to relevant ligand surfaces, with the a-helices ofeach NKG2D monomer making relatively equal contributions.Thus, two NKG2D monomers, homodimerizing through a tightlycomplementary interface are necessary for the binding of a singlemonomeric ligand (34–36). This suggests that pairing of NKG2DFL
with NKG2DTR results in a chimeric heterodimer lacking functionalligand-binding capability, in turn providing a possible “interference”mechanism for the negative regulation of intact NKG2DFL functionby the NKG2DTR isoform.The finding that NKG2DTR can pair with DAP10 is not sur-
prising, because oligomerization occurs through interactions be-tween acidic residues at or near the DAP10 dimer interface (on thecytosolic portion of NKG2D), as well as basic residues in theNKG2D transmembrane domain, all of which are preserved inNKG2DTR (37). Noncovalent association of NKG2D homodimerswith the DAP10 adaptor molecule is required for surface ex-pression and function of NKG2D in human NK cells and activatedCD8+ T cells (38). As endogenous NKG2DTR can both pair withDAP10 and form heterodimers with NKG2DFL, the reduction insurface receptor expression could be potentially explained by thesequestration of DAP10 by NKG2DTR away from NKG2DFL. This
FIGURE 5. NKG2DTR complexes with DAP10 and NKG2DFL. (A)
HEK293T cells were either left untransduced or transiently transfected
with DAP10HA in combination with NKG2DFL-Flag and/or NKG2DTR-Myc
as indicated. Anti-Flag or anti-Myc immunoprecipitations (IP) were per-
formed on transfected cell lysates and then subjected to immunoblotting
(IB) with anti-Flag, anti-Myc or (B) anti-HA Abs. (C) Cell lysates were
immunoprecipitated with anti-HA Ab and immunoblotted with anti-Flag or
(D) anti-Myc Abs. One representative of n = 4 independent experiments is
shown.
FIGURE 6. NKG2DTR retains NKG2DFL in the intracellular compart-
ment. (A) COS-7 cells were transfected with vectors expressing DAP10HA,
NKG2DTR-myc, and/or NKG2DFL-Flag as indicated. Surface NKG2D ex-
pression was detected by flow cytometry. (B) COS-7 cells were transfected
with vectors expressing DAP10HA and NKG2DFL-Flag with or without
NKG2DTR-myc. NKG2D was visualized by structured light fluorescence
microscopy using fluorescent dye-labeled anti-Flag Abs (green). Nuclei
were stained with DAPI (blue), and cell membranes marked with mem-
brane-targeted mCherry (red). One representative of n = 2 independent
experiments is shown.
6 AN NKG2D SPLICE VARIANT REGULATES NKG2D FUNCTION
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

is supported by our data showing that knockdown of NKG2DTR
enhances while the overexpression of NKG2DTR inhibits the in-teraction of NKG2DFL with DAP10. However, the inability ofDAP10 overexpression to restore NKG2D surface expression andfunction argues against this notion. Although the sequestration ofDAP10 by NKG2DTR could still play a role in the inhibitoryfunction of NKG2DTR, features of NKG2DTR other than DAP10binding are likely responsible for intracellular NKG2D retention.Alternatively, the association of NKG2DTR with NKG2DFL maycause misfolding of this heterodimeric protein, which abrogatescell surface expression of NKG2DFL. Although we do not knowthe exact cellular compartment where these complexes reside, theabsence of cell surface expression of NKG2DFL when NKG2DTR
is overexpressed suggest that NKG2DTR does not make it to thecell surface. Further studies will be required to thoroughly in-vestigate the trafficking of these complexes.The demonstrated negative effect of NKG2DTR on NKG2DFL
function and overall NKG2D-dependent cytotoxicity, both atphysiologically relevant endogenous levels and those enforced byoverexpression, suggests potential roles for NKG2DTR under bothphysiological and pathological conditions. The mechanisms un-derlying the regulation of NKG2DTR expression among individualsis unknown. However, the manipulation of NKG2DTR splice variantexpression may have clinical implications. Both activated CD8+
T cells and NK cells have considerable therapeutic potential, par-ticularly in immune-based approaches to tumor therapy (39). Thus,it is possible that expression of NKG2DTR in donor lymphocyteswould negatively impact graft-versus-tumor effects in bone marrowtransplantation. Selection of donors with relatively less expressionof NKG2DTR might be beneficial for an augmented graft-versus-tumor effect. Alternatively, the expression of NKG2DTR could besuppressed by RNA interference before adoptive transfer intorecipients to enhance the cytotoxic potential of CD8+ T cells andNK cells. Although it is unknown whether chimeric AgR-expressingT cells require NKG2D for maximal effector function, silencing ofNKG2DTR might also be an attractive option to enhance the activityof genetically engineered tumor-targeting T cells.In summary, we have identified a novel and important regulatory
mechanism that regulates NKG2D-mediated cytotoxic function inactivated CD8+ T cells. Because NKG2DTR affects the cell surfaceexpression of NKG2D, the full delineation of the factors affectingthe interaction between NKG2DTR and NKG2DFL, along with anelucidation of internal and external regulators of NKG2D splicing,may open doors to specific modulation of NKG2D function. Thiswill enhance our understanding of the regulation of immuneresponses, which may lead to better treatment options for severaldiseases, including cancer and autoimmune disorders.
AcknowledgmentsWe thank Drs. Barbara Osborne, Samuel Black, Mark Robert Nicolls, Gary
Koretzky, Avinash Bhandoola, Edward Behrens, and Daniel Chapman for
careful reading of this manuscript. We also thank Ted Hudgens for his help
with statistical analysis and Theresa Leichner for technical support.
DisclosuresThe authors have no financial conflicts of interest.
References1. Lanier, L. L. 2005. NK cell recognition. Annu. Rev. Immunol. 23: 225–274.2. Raulet, D. H. 2003. Roles of the NKG2D immunoreceptor and its ligands. Nat.
Rev. Immunol. 3: 781–790.3. Bauer, S., V. Groh, J. Wu, A. Steinle, J. H. Phillips, L. L. Lanier, and T. Spies.
1999. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 285: 727–729.
4. Cerwenka, A., A. B. Bakker, T. McClanahan, J. Wagner, J. Wu, J. H. Phillips,and L. L. Lanier. 2000. Retinoic acid early inducible genes define a ligand familyfor the activating NKG2D receptor in mice. Immunity 12: 721–727.
5. Gasser, S., S. Orsulic, E. J. Brown, and D. H. Raulet. 2005. The DNA damagepathway regulates innate immune system ligands of the NKG2D receptor. Nature436: 1186–1190.
6. Smyth, M. J., J. Swann, E. Cretney, N. Zerafa, W. M. Yokoyama, andY. Hayakawa. 2005. NKG2D function protects the host from tumor initiation. J.Exp. Med. 202: 583–588.
7. Takaki, R., Y. Hayakawa, A. Nelson, P. V. Sivakumar, S. Hughes, M. J. Smyth,and L. L. Lanier. 2005. IL-21 enhances tumor rejection through a NKG2D-dependent mechanism. J. Immunol. 175: 2167–2173.
8. Diefenbach, A., E. R. Jensen, A. M. Jamieson, and D. H. Raulet. 2001. Rae1 andH60 ligands of the NKG2D receptor stimulate tumour immunity. Nature 413:165–171.
9. Guerra, N., Y. X. Tan, N. T. Joncker, A. Choy, F. Gallardo, N. Xiong,S. Knoblaugh, D. Cado, N. M. Greenberg, and D. H. Raulet. 2008. NKG2D-deficient mice are defective in tumor surveillance in models of spontaneousmalignancy. Immunity 28: 571–580.
10. Lodoen, M., K. Ogasawara, J. A. Hamerman, H. Arase, J. P. Houchins,E. S. Mocarski, and L. L. Lanier. 2003. NKG2D-mediated natural killer cellprotection against cytomegalovirus is impaired by viral gp40 modulation ofretinoic acid early inducible 1 gene molecules. J. Exp. Med. 197: 1245–1253.
11. Krmpotic, A., D. H. Busch, I. Bubic, F. Gebhardt, H. Hengel, M. Hasan,A. A. Scalzo, U. H. Koszinowski, and S. Jonjic. 2002. MCMV glycoprotein gp40confers virus resistance to CD8+ T cells and NK cells in vivo. Nat. Immunol. 3:529–535.
12. Schmidt-Wolf, I. G., R. S. Negrin, H. P. Kiem, K. G. Blume, and I. L. Weissman.1991. Use of a SCID mouse/human lymphoma model to evaluate cytokine-induced killer cells with potent antitumor cell activity. J. Exp. Med. 174: 139–149.
13. Hoyle, C., C. D. Bangs, P. Chang, O. Kamel, B. Mehta, and R. S. Negrin. 1998.Expansion of Philadelphia chromosome-negative CD3+CD56+ cytotoxic cellsfrom chronic myeloid leukemia patients: in vitro and in vivo efficacy in severecombined immunodeficiency disease mice. Blood 92: 3318–3327.
14. Alvarnas, J. C., Y. C. Linn, E. G. Hope, and R. S. Negrin. 2001. Expansion ofcytotoxic CD3+CD56+ cells from peripheral blood progenitor cells of patientsundergoing autologous hematopoietic cell transplantation. Biol. Blood MarrowTransplant. 7: 216–222.
15. Karimi, M., T. M. Cao, J. A. Baker, M. R. Verneris, L. Soares, and R. S. Negrin.2005. Silencing human NKG2D, DAP10, and DAP12 reduces cytotoxicity ofactivated CD8+ T cells and NK cells. J. Immunol. 175: 7819–7828.
16. Uhrberg, M., N. M. Valiante, B. P. Shum, H. G. Shilling, K. Lienert-Weidenbach,B. Corliss, D. Tyan, L. L. Lanier, and P. Parham. 1997. Human diversity in killercell inhibitory receptor genes. Immunity 7: 753–763.
17. Diefenbach, A., E. Tomasello, M. Lucas, A. M. Jamieson, J. K. Hsia, E. Vivier, andD. H. Raulet. 2002. Selective associations with signaling proteins determinestimulatory versus costimulatory activity of NKG2D. Nat. Immunol. 3: 1142–1149.
18. Gilfillan, S., E. L. Ho, M. Cella, W. M. Yokoyama, and M. Colonna. 2002.NKG2D recruits two distinct adapters to trigger NK cell activation and co-stimulation. Nat. Immunol. 3: 1150–1155.
19. Shum, B. P., L. R. Flodin, D. G. Muir, R. Rajalingam, S. I. Khakoo, S. Cleland,L. A. Guethlein, M. Uhrberg, and P. Parham. 2002. Conservation and variation inhuman and common chimpanzee CD94 and NKG2 genes. J. Immunol. 168: 240–252.
20. Verneris, M. R., M. Karami, J. Baker, A. Jayaswal, and R. S. Negrin. 2004. Roleof NKG2D signaling in the cytotoxicity of activated and expanded CD8+ T cells.Blood 103: 3065–3072.
21. Reynolds, A., D. Leake, Q. Boese, S. Scaringe, W. S. Marshall, andA. Khvorova. 2004. Rational siRNA design for RNA interference. Nat. Bio-technol. 22: 326–330.
22. Glienke, J., Y. Sobanov, C. Brostjan, C. Steffens, C. Nguyen, H. Lehrach,E. Hofer, and F. Francis. 1998. The genomic organization of NKG2C, E, F, andD receptor genes in the human natural killer gene complex. Immunogenetics 48:163–173.
23. Wei, X., L. Hao, S. Ni, Q. Liu, J. Xu, and P. H. Correll. 2005. Altered exon usagein the juxtamembrane domain of mouse and human RON regulates receptoractivity and signaling specificity. J. Biol. Chem. 280: 40241–40251.
24. Wang, M. H., H. L. Fung, and Y. Q. Chen. 2000. Regulation of the RON receptortyrosine kinase expression in macrophages: blocking the RON gene transcriptionby endotoxin-induced nitric oxide. J. Immunol. 164: 3815–3821.
25. Tacconelli, A., A. R. Farina, L. Cappabianca, G. Desantis, A. Tessitore,A. Vetuschi, R. Sferra, N. Rucci, B. Argenti, I. Screpanti, et al. 2004. TrkAalternative splicing: a regulated tumor-promoting switch in human neuroblas-toma. Cancer Cell 6: 347–360.
26. Vijayakrishnan, L., J. M. Slavik, Z. Illes, R. J. Greenwald, D. Rainbow, B. Greve,L. B. Peterson, D. A. Hafler, G. J. Freeman, A. H. Sharpe, et al. 2004. An au-toimmune disease-associated CTLA-4 splice variant lacking the B7 bindingdomain signals negatively in T cells. Immunity 20: 563–575.
27. Hanawa, H., Y. Ma, S. A. Mikolajczak, M. L. Charles, T. Yoshida, R. Yoshida,C. A. Strathdee, D. W. Litchfield, and A. Ochi. 2002. A novel costimulatory sig-naling in human T lymphocytes by a splice variant of CD28. Blood 99: 2138–2145.
28. Elmhurst, J. L., Z. Xie, B. F. O’Dowd, and S. R. George. 2000. The splicevariant D3nf reduces ligand binding to the D3 dopamine receptor: evidence forheterooligomerization. Brain Res. Mol. Brain Res. 80: 63–74.
29. McElvaine, A. T., and K. E. Mayo. 2006. A dominant-negative human growthhormone-releasing hormone (GHRH) receptor splice variant inhibits GHRHbinding. Endocrinology 147: 1884–1894.
The Journal of Immunology 7
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

30. Upshaw, J. L., L. N. Arneson, R. A. Schoon, C. J. Dick, D. D. Billadeau, andP. J. Leibson. 2006. NKG2D-mediated signaling requires a DAP10-bound Grb2-Vav1intermediate and phosphatidylinositol-3-kinase in human natural killer cells. Nat.Immunol. 7: 524–532.
31. Wolan, D. W., L. Teyton, M. G. Rudolph, B. Villmow, S. Bauer, D. H. Busch,and I. A. Wilson. 2001. Crystal structure of the murine NK cell-activating re-ceptor NKG2D at 1.95 A. Nat. Immunol. 2: 248–254.
32. Li, P., D. L. Morris, B. E. Willcox, A. Steinle, T. Spies, and R. K. Strong. 2001.Complex structure of the activating immunoreceptor NKG2D and its MHC classI-like ligand MICA. Nat. Immunol. 2: 443–451.
33. Li, P., G. McDermott, and R. K. Strong. 2002. Crystal structures of RAE-1beta andits complex with the activating immunoreceptor NKG2D. Immunity 16: 77–86.
34. O’Callaghan, C. A., A. Cerwenka, B. E. Willcox, L. L. Lanier, andP. J. Bjorkman. 2001. Molecular competition for NKG2D: H60 and RAE1compete unequally for NKG2D with dominance of H60. Immunity 15: 201–211.
35. Steinle, A., P. Li, D. L. Morris, V. Groh, L. L. Lanier, R. K. Strong, and T. Spies.2001. Interactions of human NKG2D with its ligands MICA, MICB, and homologs ofthe mouse RAE-1 protein family. Immunogenetics 53: 279–287.
36. McFarland, B. J., T. Kortemme, S. F. Yu, D. Baker, and R. K. Strong. 2003.Symmetry recognizing asymmetry: analysis of the interactions between theC-type lectin-like immunoreceptor NKG2D and MHC class I-like ligands.Structure 11: 411–422.
37. Garrity, D., M. E. Call, J. Feng, and K. W. Wucherpfennig. 2005. The activatingNKG2D receptor assembles in the membrane with two signaling dimers intoa hexameric structure. Proc. Natl. Acad. Sci. USA 102: 7641–7646.
38. Billadeau, D. D., J. L. Upshaw, R. A. Schoon, C. J. Dick, and P. J. Leibson. 2003.NKG2D-DAP10 triggers human NK cell-mediated killing via a Syk-independentregulatory pathway. Nat. Immunol. 4: 557–564.
39. Leen, A. M., C. M. Rooney, and A. E. Foster. 2007. Improving T cell therapy forcancer. Annu. Rev. Immunol. 25: 243–265.
8 AN NKG2D SPLICE VARIANT REGULATES NKG2D FUNCTION
at Univ of Pennsylvania L
ibrary on August 4, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from
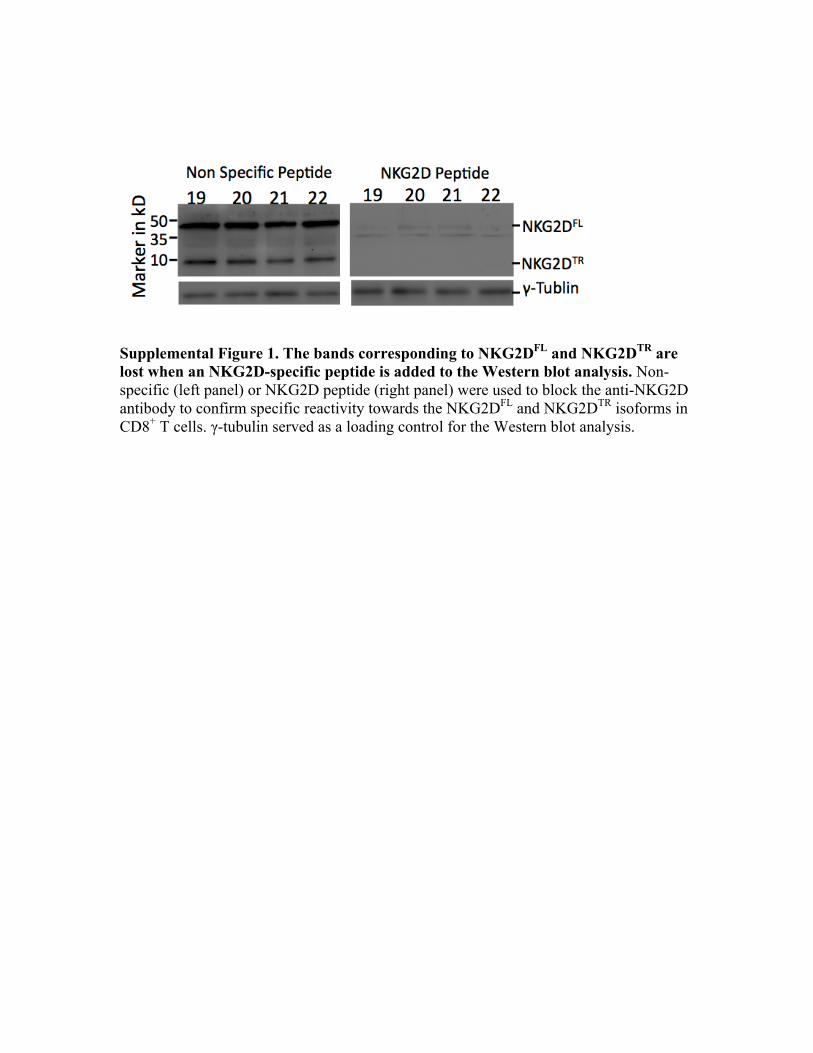
Supplemental Figure 1. The bands corresponding to NKG2DFL and NKG2DTR are lost when an NKG2D-specific peptide is added to the Western blot analysis. Non-specific (left panel) or NKG2D peptide (right panel) were used to block the anti-NKG2D antibody to confirm specific reactivity towards the NKG2DFL and NKG2DTR isoforms in CD8+ T cells. γ-tubulin served as a loading control for the Western blot analysis.
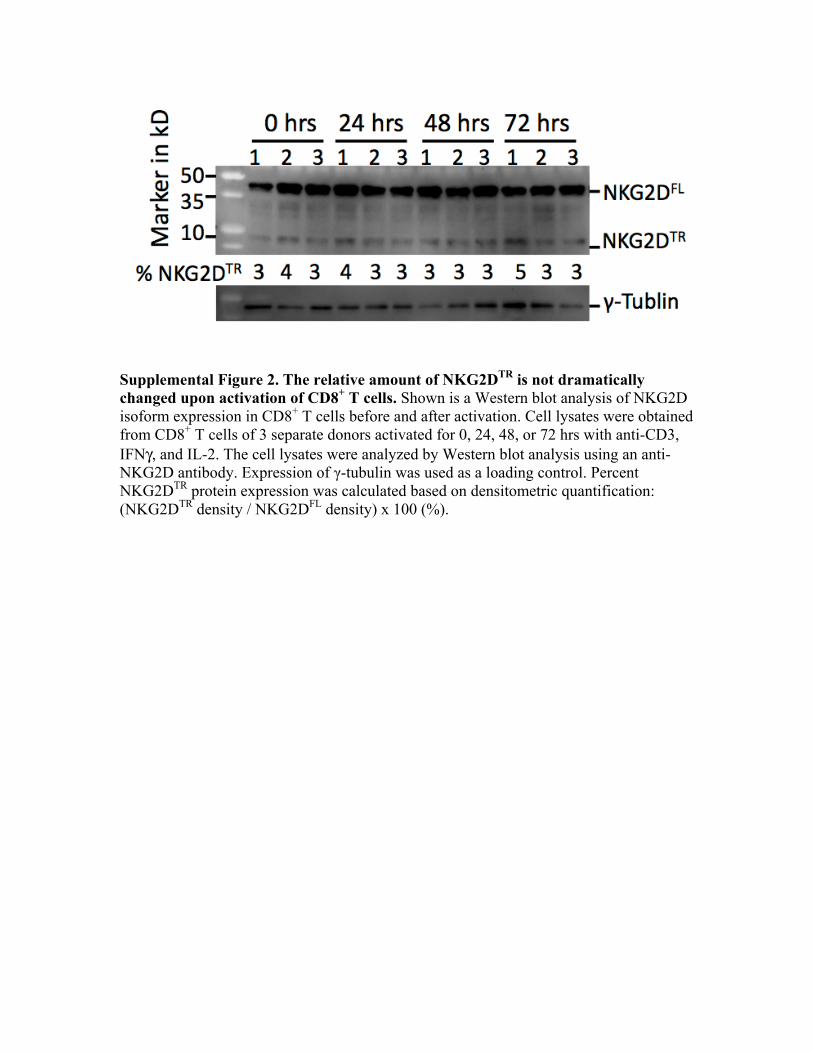
Supplemental Figure 2. The relative amount of NKG2DTR is not dramatically changed upon activation of CD8+ T cells. Shown is a Western blot analysis of NKG2D isoform expression in CD8+ T cells before and after activation. Cell lysates were obtained from CD8+ T cells of 3 separate donors activated for 0, 24, 48, or 72 hrs with anti-CD3, IFNγ, and IL-2. The cell lysates were analyzed by Western blot analysis using an anti-NKG2D antibody. Expression of γ-tubulin was used as a loading control. Percent NKG2DTR protein expression was calculated based on densitometric quantification: (NKG2DTR density / NKG2DFL density) x 100 (%).

Supplemental Figure 3. NKG2D-mediated re-directed and K562 lysis is inhibited by NKG2DTR. Human CD8+ T cells were untransduced or transduced with GFP-expressing vector or different combinations of lentiviruses encoding NKG2DTR-Myc, NKG2DFL-Flag, DAP10HA=GFP, or DAP10HA=RFP. (A) The cytotoxicity of the transduced human CD8+ T cells against P815 cells labeled with or without anti-NKG2D antibody was determined at a 10:1 E:T ratio. (B) The cytotoxicity of the transduced human CD8+ T cells against K562 cells was determined at the 40:1 and 10:1 E:T ratios. One representative of at leats n = 3 independent experiments is shown. *** indicates a statistically significant difference of p<0.001 between the groups using ANOVA with Tukey’s post-test.
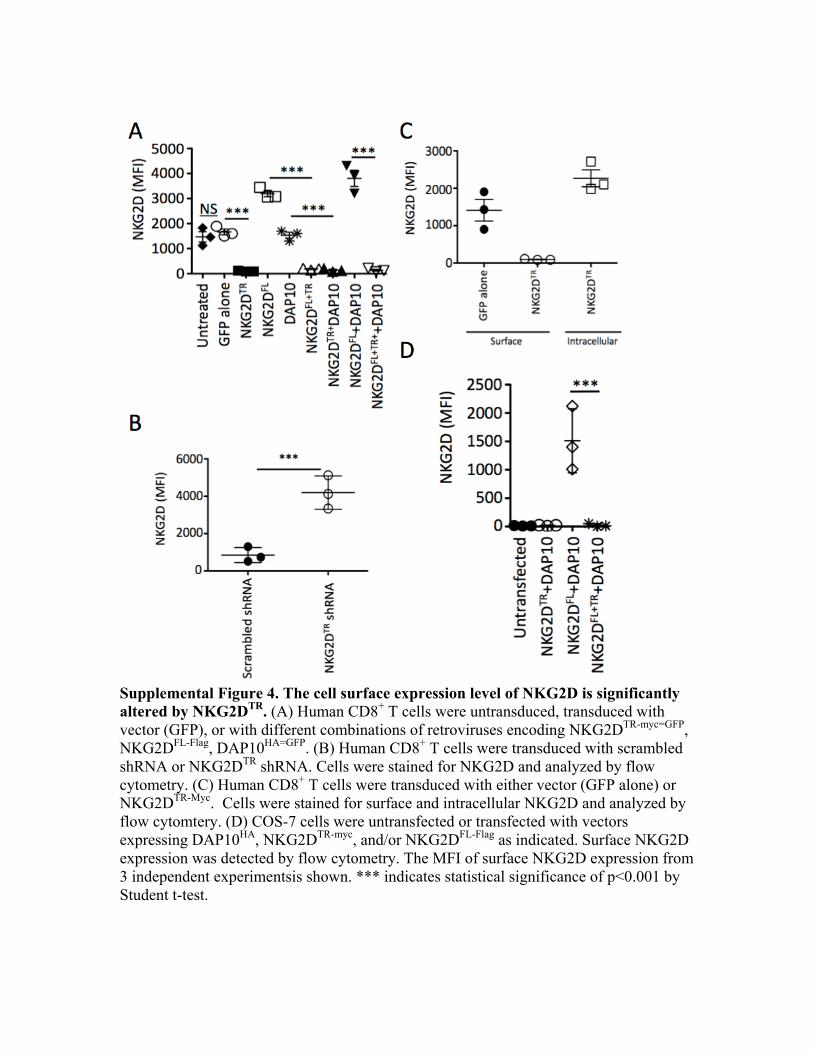
Supplemental Figure 4. The cell surface expression level of NKG2D is significantly altered by NKG2DTR. (A) Human CD8+ T cells were untransduced, transduced with vector (GFP), or with different combinations of retroviruses encoding NKG2DTR-myc=GFP, NKG2DFL-Flag, DAP10HA=GFP. (B) Human CD8+ T cells were transduced with scrambled shRNA or NKG2DTR shRNA. Cells were stained for NKG2D and analyzed by flow cytometry. (C) Human CD8+ T cells were transduced with either vector (GFP alone) or NKG2DTR-Myc. Cells were stained for surface and intracellular NKG2D and analyzed by flow cytomtery. (D) COS-7 cells were untransfected or transfected with vectors expressing DAP10HA, NKG2DTR-myc, and/or NKG2DFL-Flag as indicated. Surface NKG2D expression was detected by flow cytometry. The MFI of surface NKG2D expression from 3 independent experimentsis shown. *** indicates statistical significance of p<0.001 by Student t-test.




















