
Biochemical and molecular characterization of purified chicken pancreatic phospholipase A 2
Upload
independentCategory
view
2download
0
Life Sciences 76 (2005) 2607–2619
www.elsevier.com/locate/lifescie
A novel anticoagulant purified from fish protein hydrolysate
inhibits factor XIIa and platelet aggregation
Niranjan Rajapakse, Won-Kyo Jung, Eresha Mendis,
Sung-Hoon Moon, Se-Kwon Kim*
Department of Chemistry, Pukyong National University, Daeyon-3-Dong, Nam-Gu, Busan 608-737, Korea
Received 18 October 2004; accepted 10 December 2004
Abstract
A novel fish protein having anticoagulant and antiplatelet properties was enzymatically extracted from the
marine fish, yellowfin sole (Limanda aspera) and purified to homogeneity producing an overall purification fold of
206.6. MALDI-TOF mass spectroscopic and SDS-PAGE analysis identified the purified protein as 12.01 kDa
single-chain monomeric protein. It inhibited the activated coagulation factor XII (FXIIa) by forming an inactive
complex regardless of Zn2+ mediation, and was named, yellowfin sole anticoagulant protein (YAP). In addition,
YAP act to antagonize platelet membrane glycoprotein integrin, to arrest platelet aggregation. However, YAP was
not able to block the adhesion of platelets to collagen, which mediate via major collagen receptors, GPIa/IIa on
platelet membrane. Furthermore, YAP did not possess plasminogen activator-like activity to activate fibrinolysis. In
fact, our findings indicate that YAP binds with FXIIa and platelet membrane integrins to inhibit thrombosis in vitro.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Anticoagulant; Platelet aggregation; Protein hydrolysate; Activated factor XII (FXIIa); Platelet integrin
Introduction
Autoactivation of factor XII (FXII or Hageman factor) into its active form (FXIIa) in contact with
negatively charged surfaces triggers intrinsic pathway of coagulation along with high molecular weight
0024-3205/$ -
doi:10.1016/j.l
* Correspond
E-mail add
see front matter D 2005 Elsevier Inc. All rights reserved.
fs.2004.12.010
ing author. Tel.: +82 51 620 6375; fax: +82 51 628 8147.
ress: [email protected] (S.-K. Kim).

N. Rajapakse et al. / Life Sciences 76 (2005) 2607–26192608
kininogen (HMWK) and prekallikrein (Miller et al., 1980). Some evidences suggest that Zn2+ binding to
FXII induces a conformational change that makes the protein more susceptible for the development of
enzymatic activity during autoactivation (Schousboe, 1993). Therefore, the simple fact that a deficiency
of FXII prolongs activated partial thromboplastin time (APTT) in vitro has been clearly recognized to be
important in coagulation. At the same time, platelets also play an important role in coagulation by
aggregating and adhering to the underline collagen via platelet membrane glycoprotein integrins
(Clemetson and Clemetson, 2001).
The importance of anticoagulants and antiplatelets as treatments for the prevention of ischaemic
events in patients with cardiovascular diseases is becoming increasingly recognized. Other than the
natural anticoagulants found in the coagulation cascade, a number of anticoagulant and antiplatelet
protein compounds have also been identified from diverse natural sources (Urata et al., 2003; Tanaka-
Azevedo et al., 2003). Natural anticoagulants found in above biological systems have been evolved
under genetic control with particular structural sequences to regulate coagulation events. Despite the
traditional means of identifying already existing anticoagulant compounds from these sources, only a
few efforts have been laid to develop novel anticoagulants from natural bioresources. In this regard,
proteolysis can be applied as a potential technique to identify such molecules from sequences of protein
sources. Therefore, in this study we attempted to identify anticoagulant compounds from protein
sequences of fish muscle following enzymatic proteolysis. We report that, the new anticoagulant protein
inhibits coagulation in vitro by forming an inactive complex with FXIIa in the contact system.
Furthermore, we discuss the significance of the anticoagulant protein to act as a platelet aggregation
inhibitor.
Materials and methods
Fish frame and reagents
Yellowfin sole (Limanda aspera) frame was obtained from Daerim Co. (Busan, South Korea). DEAE-
Sephadex A-25, Sephadex G-25, bovine thrombin, fibrinogen, plasmin, p-nitrophenyl phosphate, pepsin,
papain and a-chymotrypsin were purchased from Sigma Chemical Co. (St. Louis, MO). AlcalaseR and
NeutraseR were supplied by Novo Nordisk Co. (Denmark). All PAGE reagents were also purchased
from Sigma Chemical Co. (St. Louis, MO). Coagulation and specific factor assay reagents were products
of Instrumentation Laboratory Co. (Lexington, MA). FXIIa was purchased from ICN Pharmaceuticals
Inc. (Aurora, Ohio, USA). The other reagents were of the highest grade commercially available.
Extraction of fish protein
Yellowfin sole frame containing considerable amount of protein after filleting process was subjected
to hydrolyze separately with seven different proteases, AlcalaseR, NeutraseR, pepsin, papain, a-
chymotrypsin, trypsin and previously extracted tuna pyloric caeca crude enzyme (TPCCE) (Kim et al.,
1997). Hydrolysis was performed for different time intervals and the optimum enzymic conditions were
adjusted as required. After the extraction, hydrolysate was analyzed for the degree of hydrolysis and
expressed as percentage protein content measured by the method of Lowry et al. (1951). Finally, the
hydrolysate was centrifuged at 12000 � g for 10 min at 48C and the supernatant was lyophilized.

N. Rajapakse et al. / Life Sciences 76 (2005) 2607–2619 2609
Blood collection and preparation of plasma
Nine parts of blood was drawn by venipuncture into one part of 3.2% sodium citrate from three
healthy volunteers and pooled. The blood was separately centrifuged at 1600 � g for 10 min and 150 �g for 10 min to obtain platelet poor plasma (PPP) and platelet rich plasma (PRP) respectively. Separated
plasma was utilized for in vitro testing within 24 h.
Plasma-based coagulation assays
All coagulation assays were performed with three individual replicates using an ACLRCoagulation Analyzer (ACLR 7000, Instrumentation Laboratory Co., Lexington, MA) and mean
values were taken. Normal citrated PPP (80 Al) was incubated with the anticoagulant compound (20
Al) for 3 min at 378C. For activated partial thromboplastin time (APTT) assay, 100 Al of APTT
reagent was added to the above mixture and incubated at 378C for 3 min and clotting time was
recorded following the addition of 100 Al of 20 mM CaCl2. In prothrombin time (PT) assay, pre-
warmed 200 Al of PT reagent was added to the incubated mixture of PPP and anticoagulant and
clotting time was recorded. For thrombin time (TT) measurements, 100 Al of anticoagulant was
incubated with human thrombin (100 Al) for 3 min at 378C and clotting time was determined after the
addition of 200 Al of PPP.
Purification of anticoagulant
Lyophilized crude extract was dissolved in 20 mM tris-HCl buffer (pH 7.7) and separated using
a DEAE-Sephadex A-25 ion exchange column (b 35.0 � 280.0 mm). Adsorbed fractions were
collected using a 0–1.5 M NaCl at 215 nm and fractions resulted under a same elution peak were
pooled. Followed by desalting, pooled fractions were lyophilized and tested for APTT assay. An
aqueous solution of potent fraction was loaded onto a Sephadex G-25 gel filtration column (b 20.0 �750.0 mm). Separation was obtained at a flow rate of 0.5 ml/min with distilled water and eluted
fractions (3 ml) were pooled after spectrophotometric measurement. The active fraction selected by
APTT was injected into a capcell pack C18 UG120 (b 20.0 � 250.0 mm) HPLC column (Shiseido
Fine Chemicals, Tokyo, Japan). A linear gradient of acetonitrile (0–50% v/v, in 50 min)
in distilled water was maintained and elution peaks were collected at 215 nm. The peak
responsible for the highest anticoagulant activity was finally purified on a Zorbax SB C18 (b 4.6 �250.0 mm) HPLC column (Agilent Technologies, USA) with a linear gradient of acetonitrile
(0–12.5% v/v, in 75 min). Throughout the purification, all chromatographic procedures were
carried out at 48C.
Determination of purity and molecular weight
The accurate native molecular mass and purity of the anticoagulant were determined by matrix-
assisted laser desorption ionization time-of-flight (MALDI-TOF) mass spectroscopy on a Voyager DE-
PRO mass spectrometer (Applied Biosystems, Boston, MA) at a laser power of 3000 kWcm�2. Reduced
molecular weight was studied by sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-
PAGE) on 12.5% gel following the method of Laemmli (1970).

N. Rajapakse et al. / Life Sciences 76 (2005) 2607–26192610
Identification of amino acid composition and N-terminal sequence
Amino acid composition of a-chymotryptic crude extract was analyzed with an automatic amino acid
analyzer, Biochrom 20 (Biochrom Ltd., Cambridge, UK) after hydrolysis in 6 N HCl at 1108C for 24 h
in a vacuum.
The N-terminal amino acid sequence of the purified anticoagulant was obtained by automated Edman
degradation on a Perkin-Elmer (model 470A, Applied Biosystem Division, Branchburg, NJ) protein
sequencer equipped with an HPLC system.
Determination of specific factor activity
The specific activity of activated coagulation factors was determined by modified clotting assays of
APTT using ILR Test Factor Deficient Plasma assay kits (Instrumentation Laboratory Co., Lexington,
MA). Briefly, various concentrations of anticoagulant treated platelet poor human plasma were tested to
normalize prolong clotting time of specific factor deficient standard human plasma. The percentage
activity of specific coagulation factor was calculated following the manufacturer’s instructions.
PAGE analysis of FXIIa inhibition
Native PAGE was used to identify the inhibition of FXIIa. A mixture of FXIIa and the purified
anticoagulant was incubated at 378C in the presence or absence of Zn2+ (5 nM) for 75 min. After the
reaction, 15 Al aliquots were migrated on a 12.5% separating gel at pH 8.8 using Tris-HCl as the
electrolyte buffer. Calibration molecular weight markers used were aprotinin (MW 6.5 kDa), lysozyme
(MW 14.3 kDa), h-lactoglobulin (MW 18.4 kDa), egg albumin (MW 44.0 kDa) and bovine albumin
(66.0 kDa).
Fibrinolytic assay
Fibrinolytic activity of the purified anticoagulant was estimated by a modified fibrin plate assay of
Dametto et al. (2000) using plasmin as the standard. The fibrinogen solution (4.0 ml of 0.5% human
fibrinogen in 0.1 M barbital buffer, pH 7.7, containing 1.6 mM CaCl2, 0.7 mM MgCl2 and 92.0 mM
NaCl) was mixed with agarose solution (4.0 ml of 2.0% agarose in the same barbital buffer), 0.2 ml of
20 NIH U/ml thrombin in 0.9% saline and 0.1 ml of 1.0 M CaCl2. The mixture was poured into a
Petri dish and allowed to stand for 1 h at room temperature to obtain a fibrin clot layer. Various
concentrations of YAP solutions (0–600 AM) in plasminogen (10 Al) were spotted on the fibrin layer
and incubated at 378C for 10 h. Fibrinolytic activity was determined by measuring two perpendicular
diameters of the lysed zone.
Platelet aggregation inhibition assay
Microplate method of Bendar et al. (1995) was performed to detect platelet aggregation. Briefly, PRP
was adjusted with Ca2+ free Tyrode’s buffer (137.0 mM NaCl, 2.7 mM KCl, 0.5 mM NaH2PO4, 12.0
mM NaHCO3, 1.0 mM MgCl2 and 3.5 g/l bovine serum albumin) to obtain an approximate plate count
of 2 � 105/Al. Above solution (100 Al) was preincubated with 10 Al of differentially concentrated

N. Rajapakse et al. / Life Sciences 76 (2005) 2607–2619 2611
purified anticoagulant (0–600 AM) at 378C for 12 min and 100 Al aliquots were added to individual
wells. Aggregation of platelets was initiated by adding 20 Al of ADP (50 AM) or thrombin (50 nM).
Change in optical density (OD) at 630 nm against PPP was monitored kinetically to evaluate platelet
aggregation by using an Emaxk microplate reader (Molecular Devices Co., Sunnyvale, CA). The rate
change in OD at 630 nm over 5 min was used to measure the platelet aggregation. Diluted PRP in the
absence of purified anticoagulant was kept as the control.
Platelet adhesion inhibition assay
The ability of platelets to adhere with collagen via specific integrin on cell membrane was measured
using a 96-well microtiter plate assay as described by Bellavite et al. (1994). In brief, various
concentrations (0–400 AM) of anticoagulant (10 Al) were preincubated with 150 Al of platelet solutionadjusted with Ca2+ free Tyrode’s buffer (approximately 2 � 105 platelets/Al) at 308C for 10 min. An
aliquot (150 Al) of above mixtures was added to type I collagen plated microtiter plate and incubated at
the same temperature. After 60 min of incubation, non-adherent platelets were completely washed off
with phosphatebuffered saline and adherent platelets were solubilized with 100 Al of buffer containing1.0% Triton X-100 and 5.0 mM p-nitrophenyl phosphate. After incubating for 60 min, the amount of
acid phosphate release (that represents the amount of platelets lysed) was measured using an Emaxkmicroplate reader at 405 nm. The platelet adhesion percentage was calculated using the OD of control,
where PRP was not preincubated with the anticoagulant.
Statistics
Results are presented as mean F SD of the mean (n = 3). Student’s t-test was used to determine the
level of significance.
Results
Extraction and purification of the anticoagulant
In a preliminary study, seven hydrolytic enzymes (as mentioned in the Materials and methods section)
were separately utilized to hydrolyze yellowfin sole frame protein and screened for potent anticoagulant
activity. The anticoagulant activities of hydrolysates were changed with the time course action of
hydrolysis and the highest inhibition of coagulation exhibited after four hours of reaction time (data
not shown). All hydrolysates could significantly prolong APTT but not PT or TT (P b 0.05). Those
results suggested that anticoagulant compounds present in all hydrolysates act to inhibit coagulation
factor(s) in intrinsic pathway of coagulation. Among the enzymes tested, a-chymotrypsin, papin and
trypsin could result comparatively a higher degree of hydrolysis (45–50%). Moreover, the highest
prolongation of APTT (68.4 F 0.5 s) was observed with a-chymotryptic hydrolysate and the same
enzyme was selected for the extraction of anticoagulant protein. The amino acid composition of a-
chymotryptic hydrolysate was rich in acidic amino acids (glutamic acid and aspartic acid), lysine,
leucine and glycine (Table 1). Together they constituted 53.34% of the total amino acid residues. This
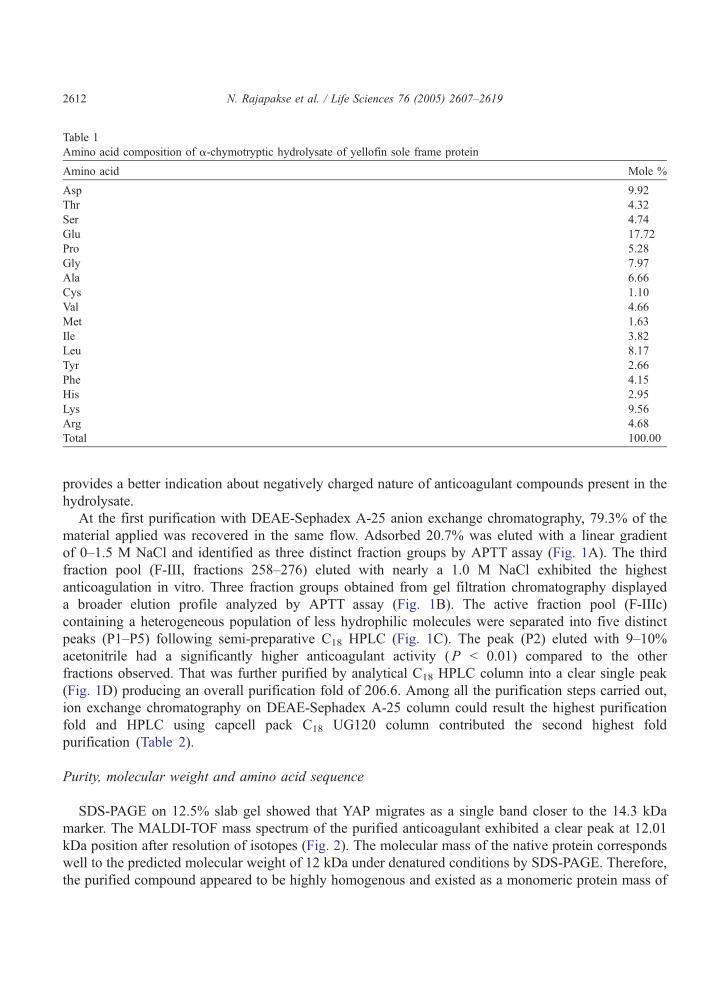
Table 1
Amino acid composition of a-chymotryptic hydrolysate of yellofin sole frame protein
Amino acid Mole %
Asp 9.92
Thr 4.32
Ser 4.74
Glu 17.72
Pro 5.28
Gly 7.97
Ala 6.66
Cys 1.10
Val 4.66
Met 1.63
Ile 3.82
Leu 8.17
Tyr 2.66
Phe 4.15
His 2.95
Lys 9.56
Arg 4.68
Total 100.00
N. Rajapakse et al. / Life Sciences 76 (2005) 2607–26192612
provides a better indication about negatively charged nature of anticoagulant compounds present in the
hydrolysate.
At the first purification with DEAE-Sephadex A-25 anion exchange chromatography, 79.3% of the
material applied was recovered in the same flow. Adsorbed 20.7% was eluted with a linear gradient
of 0–1.5 M NaCl and identified as three distinct fraction groups by APTT assay (Fig. 1A). The third
fraction pool (F-III, fractions 258–276) eluted with nearly a 1.0 M NaCl exhibited the highest
anticoagulation in vitro. Three fraction groups obtained from gel filtration chromatography displayed
a broader elution profile analyzed by APTT assay (Fig. 1B). The active fraction pool (F-IIIc)
containing a heterogeneous population of less hydrophilic molecules were separated into five distinct
peaks (P1–P5) following semi-preparative C18 HPLC (Fig. 1C). The peak (P2) eluted with 9–10%
acetonitrile had a significantly higher anticoagulant activity (P b 0.01) compared to the other
fractions observed. That was further purified by analytical C18 HPLC column into a clear single peak
(Fig. 1D) producing an overall purification fold of 206.6. Among all the purification steps carried out,
ion exchange chromatography on DEAE-Sephadex A-25 column could result the highest purification
fold and HPLC using capcell pack C18 UG120 column contributed the second highest fold
purification (Table 2).
Purity, molecular weight and amino acid sequence
SDS-PAGE on 12.5% slab gel showed that YAP migrates as a single band closer to the 14.3 kDa
marker. The MALDI-TOF mass spectrum of the purified anticoagulant exhibited a clear peak at 12.01
kDa position after resolution of isotopes (Fig. 2). The molecular mass of the native protein corresponds
well to the predicted molecular weight of 12 kDa under denatured conditions by SDS-PAGE. Therefore,
the purified compound appeared to be highly homogenous and existed as a monomeric protein mass of

0.0
1.5
F-IF-II
F-III
Pool
A
F-IIIa
F-IIIb
F-IIIc
Pool
B
D
0
50
150
200
0
50
P1P2
P3
P4P5
C
F-IF-II
F-III
Pool
A
Fraction number 0 50 100 150 200 250 300
0
1
2
3
4
25
30
35
40
45
50
55
60
F-IF-II
F-III
Pool
A
F-IIIa
F-IIIb
F-IIIc
Pool
B
0 20 40
0 20 40 5010 30
60 800.0
0.4
0.8
1.2
1.6
0
50
100
150
200
250
F-IIIa
F-IIIb
F-IIIc
Pool
D
0 15 30 45 60 750.0
0.3
0.6
0.9
1.2
0
5
10
15
20D
0.0
0.5
1.0
1.5
2.0
P1P2
P3
P4P5
C
Retention time (min)
Fraction number
Retention time (min)
100
P1P2
P3
P4P5
C
Abs
orba
nce
at 2
15 n
mA
bsor
banc
e at
215
nm
Ace
toni
trile
(%
)P
rolo
ngat
ion
of A
PT
T (
s)
Abs
orba
nce
at 2
15 n
mA
bsor
banc
e at
215
nm
Ace
toni
trile
(%
)N
aCl (
M)
Pro
long
atio
n of
AP
TT
(s)
Pro
long
atio
n of
AP
TT
(s)
Fig. 1. Purification of a potent coagulation inhibitor from a-chymotryptic hydrolysate of yellowfin sole frame protein. (A) Ion
exchange chromatography on a DEAE-Sephadex A-25 column. Lyophilized crude extract was dissolved in 20 mM tris-HCl
buffer (pH 7.7) and loaded onto the ion exchange column. Adsorbed fractions were eluted with 0–1.5 M NaCl gradient and
collected monitoring absorbance at 280 nm. Eluted fractions were tested for the inhibition of contact pathway coagulation by
APTT assay. (B) Gel filtration chromatography on Sephadex G-25 column. Potent fraction pool (F III) obtained from ion
exchange chromatography was applied onto the gel filtration column and fractions eluted with distilled water were tested for
APTT assay. (C) The lyophilized potent fraction (F IIIc) obtained from gel filtration chromatography was dissolved in distilled
water and injected into a capcell pack C18 UG120 HPLC column. The column was eluted with 0–50% (v/v) acetonitrile gradient
and elution peaks were assayed for APTT. (D) The highest anticoagulation activie peak (P2) obtained from capcell pack C18
UG120 HPLC was finally purified by Zorbax SB C18 HPLC column with 0–12.5% (v/v) acetonitrile gradient.
N. Rajapakse et al. / Life Sciences 76 (2005) 2607–2619 2613
12.01 kDa. We designated this protein as yellowfin sole anticoagulant protein (YAP) and N-terminal
amino acid sequence data determined to be TDGSEDYGILEIDSR.
Inhibition of coagulation and specific factor activity
As shown in Fig. 3, YAP prolonged APTT in a dose-dependant manner. However, APTT was not
affected by low concentrations of YAP. A marked increment in APTT prolongation was observed when
the concentration of YAP was higher than 2 AM. In addition, YAP was tested for PT and TT. Neither PT
nor TT was affected with the treatment of YAP, reflecting the results of crude protein extract (data not
shown). These results indicated that YAP is a specific inhibitor of the intrinsic pathway of coagulation.
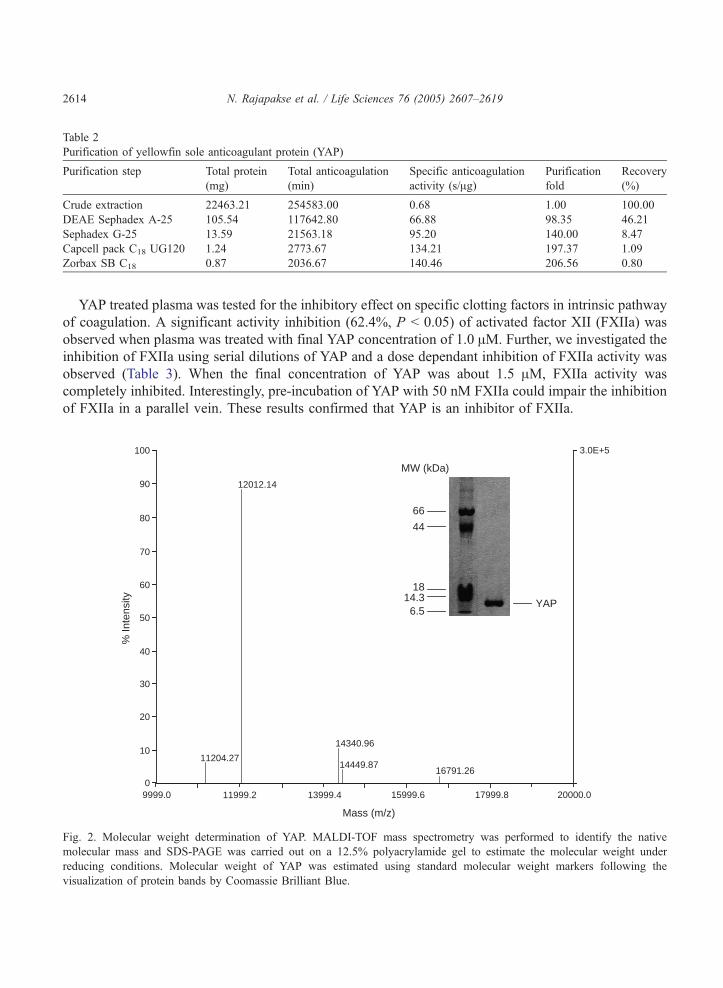
Table 2
Purification of yellowfin sole anticoagulant protein (YAP)
Purification step Total protein
(mg)
Total anticoagulation
(min)
Specific anticoagulation
activity (s/Ag)Purification
fold
Recovery
(%)
Crude extraction 22463.21 254583.00 0.68 1.00 100.00
DEAE Sephadex A-25 105.54 117642.80 66.88 98.35 46.21
Sephadex G-25 13.59 21563.18 95.20 140.00 8.47
Capcell pack C18 UG120 1.24 2773.67 134.21 197.37 1.09
Zorbax SB C18 0.87 2036.67 140.46 206.56 0.80
N. Rajapakse et al. / Life Sciences 76 (2005) 2607–26192614
YAP treated plasma was tested for the inhibitory effect on specific clotting factors in intrinsic pathway
of coagulation. A significant activity inhibition (62.4%, P b 0.05) of activated factor XII (FXIIa) was
observed when plasma was treated with final YAP concentration of 1.0 AM. Further, we investigated the
inhibition of FXIIa using serial dilutions of YAP and a dose dependant inhibition of FXIIa activity was
observed (Table 3). When the final concentration of YAP was about 1.5 AM, FXIIa activity was
completely inhibited. Interestingly, pre-incubation of YAP with 50 nM FXIIa could impair the inhibition
of FXIIa in a parallel vein. These results confirmed that YAP is an inhibitor of FXIIa.
50
YAP
66
44
1814.3
6.5
MW (kDa)
12012.14
16791.2611204.27
14340.96
14449.87
9999.0 11999.2 13999.4 15999.6 17999.8
Mass (m/z)
% In
tens
ity
100
90
80
70
60
40
30
20
10
0
3.0E+5
20000.0
Fig. 2. Molecular weight determination of YAP. MALDI-TOF mass spectrometry was performed to identify the native
molecular mass and SDS-PAGE was carried out on a 12.5% polyacrylamide gel to estimate the molecular weight under
reducing conditions. Molecular weight of YAP was estimated using standard molecular weight markers following the
visualization of protein bands by Coomassie Brilliant Blue.
YAP concentration (µM)
0 42 6 8 10 12
Pro
long
atio
n of
AP
TT
(s)
0
50
100
150
200
250
300
350
Fig. 3. Dose dependant inhibition of contact system induced coagulation by YAP. Various concentrations of YAP were
preincubated with platelet poor plasma and tested for the inhibition of coagulation by APTT assay.
N. Rajapakse et al. / Life Sciences 76 (2005) 2607–2619 2615
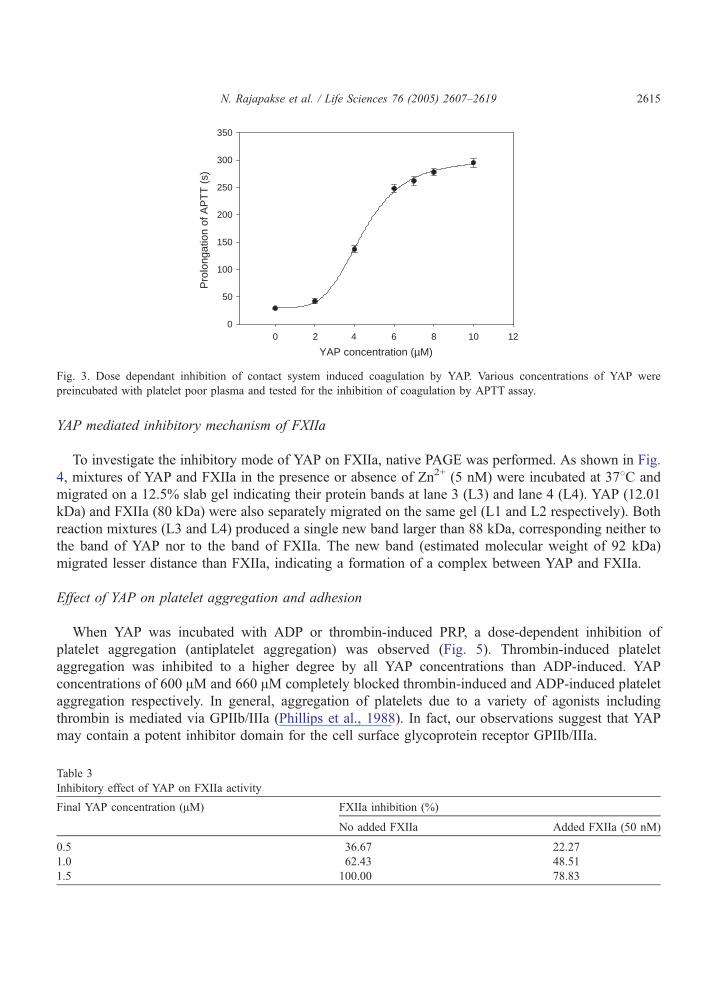
YAP mediated inhibitory mechanism of FXIIa
To investigate the inhibitory mode of YAP on FXIIa, native PAGE was performed. As shown in Fig.
4, mixtures of YAP and FXIIa in the presence or absence of Zn2+ (5 nM) were incubated at 378C and
migrated on a 12.5% slab gel indicating their protein bands at lane 3 (L3) and lane 4 (L4). YAP (12.01
kDa) and FXIIa (80 kDa) were also separately migrated on the same gel (L1 and L2 respectively). Both
reaction mixtures (L3 and L4) produced a single new band larger than 88 kDa, corresponding neither to
the band of YAP nor to the band of FXIIa. The new band (estimated molecular weight of 92 kDa)
migrated lesser distance than FXIIa, indicating a formation of a complex between YAP and FXIIa.
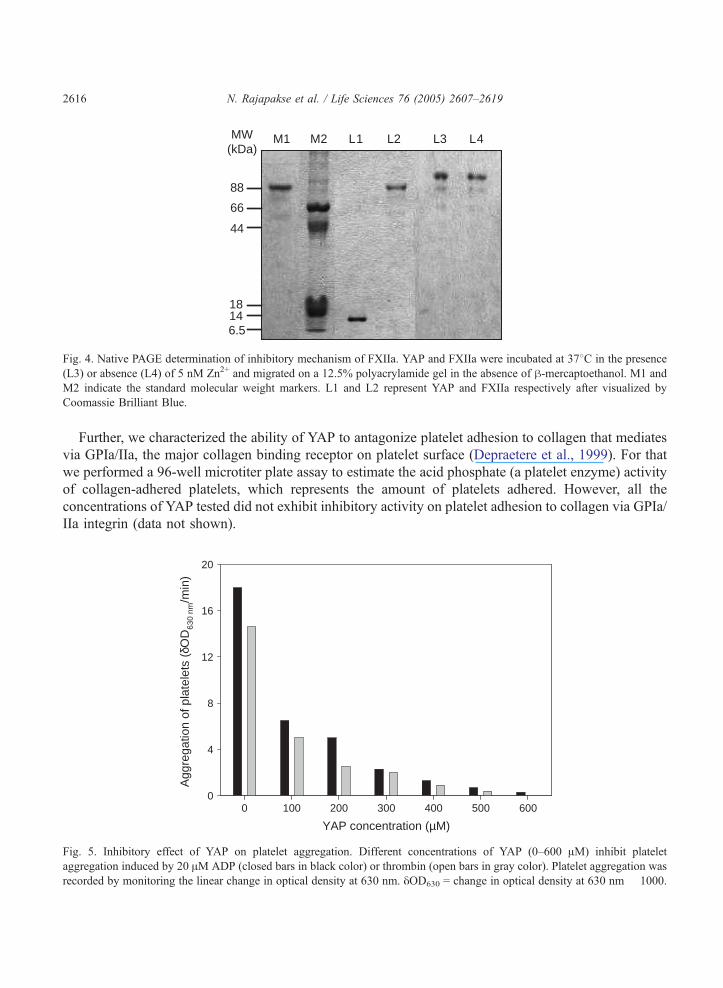
Effect of YAP on platelet aggregation and adhesion
When YAP was incubated with ADP or thrombin-induced PRP, a dose-dependent inhibition of
platelet aggregation (antiplatelet aggregation) was observed (Fig. 5). Thrombin-induced platelet
aggregation was inhibited to a higher degree by all YAP concentrations than ADP-induced. YAP
concentrations of 600 AM and 660 AM completely blocked thrombin-induced and ADP-induced platelet
aggregation respectively. In general, aggregation of platelets due to a variety of agonists including
thrombin is mediated via GPIIb/IIIa (Phillips et al., 1988). In fact, our observations suggest that YAP
may contain a potent inhibitor domain for the cell surface glycoprotein receptor GPIIb/IIIa.
Table 3
Inhibitory effect of YAP on FXIIa activity
Final YAP concentration (AM) FXIIa inhibition (%)
No added FXIIa Added FXIIa (50 nM)
0.5 36.67 22.27
1.0 62.43 48.51
1.5 100.00 78.83
M1 M2 L1 L2 L3 L4
88
66
44
18146.5
MW(kDa)
Fig. 4. Native PAGE determination of inhibitory mechanism of FXIIa. YAP and FXIIa were incubated at 378C in the presence
(L3) or absence (L4) of 5 nM Zn2+ and migrated on a 12.5% polyacrylamide gel in the absence of h-mercaptoethanol. M1 and
M2 indicate the standard molecular weight markers. L1 and L2 represent YAP and FXIIa respectively after visualized by
Coomassie Brilliant Blue.
N. Rajapakse et al. / Life Sciences 76 (2005) 2607–26192616
Further, we characterized the ability of YAP to antagonize platelet adhesion to collagen that mediates
via GPIa/IIa, the major collagen binding receptor on platelet surface (Depraetere et al., 1999). For that
we performed a 96-well microtiter plate assay to estimate the acid phosphate (a platelet enzyme) activity
of collagen-adhered platelets, which represents the amount of platelets adhered. However, all the
concentrations of YAP tested did not exhibit inhibitory activity on platelet adhesion to collagen via GPIa/
IIa integrin (data not shown).
YAP concentration (µM)
0 100 200 300 400 500 600
Agg
rega
tion
of p
late
lets
(δ O
D63
0 nm
/min
)
0
4
8
12
16
20
Fig. 5. Inhibitory effect of YAP on platelet aggregation. Different concentrations of YAP (0–600 AM) inhibit platelet
aggregation induced by 20 AM ADP (closed bars in black color) or thrombin (open bars in gray color). Platelet aggregation was
recorded by monitoring the linear change in optical density at 630 nm. yOD630 = change in optical density at 630 nm � 1000.

N. Rajapakse et al. / Life Sciences 76 (2005) 2607–2619 2617
Discussion
This study was designed aiming to purify a novel anticoagulant from an enzymatic hydrolysate of
marine fish protein source. For this purpose, a-chymotrypsin resulted yellowfin sole protein hydrolysate
was employed consecutive chromatographic separations followed by anticoagulant activity testing.
Finally, a single-chain monomeric protein mass of 12.01 kDa having potent anticoagulant properties was
identified and designated yellowfin sole anticoagulant protein (YAP). As observed in some other reported
anticoagulant proteins (Jung et al., 2002; Hojima et al., 1980), YAP consisted of considerable amount of
negatively charged amino acids in its sequence. The negative charge nature of anticoagulant compounds
expects to facilitate interaction with coagulation factors in the coagulation cascade. Dose-dependant
prolongation of intrinsic pathway of coagulation identified by APTT assay and ineffectiveness to prolong
PT and TT strongly indicated YAP as a specific inhibitor of the intrinsic pathway of coagulation.
We evaluated the inhibitory effect of YAP on specific clotting factors in intrinsic pathway of
coagulation and observed the purified anticoagulant protein is a specific inhibitor of FXIIa.
Autoactivation of factor XII into its active form (FXIIa) in contact with negatively charged surfaces
triggers intrinsic pathway of coagulation and follows sequential activation of number of other
coagulation factors (Miller et al., 1980). Up-to-date, several natural protein sequences have been
identified to inhibit different coagulation factors in intrinsic coagulation cascade (Jung et al., 2002; Urata
et al., 2003). However, only a few reports are available on anticoagulant proteins that inhibit FXIIa. Corn
trypsin inhibitor (12 kDa) with comparatively similar molecular weight acts same as YAP to inhibit
FXIIa (Hojima et al., 1980). Hamadarin, another anticoagulant protein isolated from the saliva of
Anopheles stephensi has shown that it inhibits reciprocal activation of FXII and kallikrein by directly
binding to both FXII and high molecular weight kininogen (Isawa et al., 2002). To investigate the
inhibitory mode of YAP on FXIIa, native PAGE experiments were performed. If YAP cleaves or binds
with FXIIa, the resultant product(s) should migrate on PAGE gel at different rates due to the difference in
molecular weight and relative charge. At 1:1 molar ratio of YAP and FXIIa, a new protein band was
observed and its molecular weight was closer to the estimated combined molecular weight (92 kDa) of
YAP and FXIIa. Therefore, our results confirm that inactivation of FXIIa by YAP is proceeded via
formation of an inactive complex rather than cleavage of FXIIa. It is expected that, Zn2+ induces some
conformational changes in FXII to bind with negatively charged activation surfaces (Schousboe, 1993).
However, incubation of FXIIa with YAP did not reveal the requirement of Zn2+ for the complex
formation between FXIIa and YAP. Further, this suggests that YAP may bind to FXIIa regardless of
metal ion induced conformational change in receptor binding domain. On the other hand, the
physiological concentration of FXIIa in activated normal human plasma is about 375 nM (Tans and
Rosing, 1987) and the complete inhibition of blood coagulation approximately at the same concentration
of YAP, agree with our explanation on the inactive complex formation between FXIIa and YAP.
Activation and aggregation of platelets also play an important role in the regulation of vascular-
platelet hemostasis. These functions are mainly determined by changes of the vascular wall in response
to multiple physiologic agonists such as, adenosine diphosphate (ADP), thrombin, thromboxane A2 and
collagen (Plow and Ginsberg, 1989). Activated platelets interact with each other (aggregation of
platelets) and with some other extracellular components such as collagen and fibrinogen (adhesion of
platelets) mainly via two distinct platelet surface glycoprotein integrins, GPIIb/IIIa (aIIbh3) and GPIa/IIa
(a2h1). Following activation, platelets secrete ADP and other agonists stimulating neighboring platelets
and provoking integrin GPIIb/IIIa-mediated Ca2+-dependent platelet aggregation. Antagonists of platelet

N. Rajapakse et al. / Life Sciences 76 (2005) 2607–26192618
aggregation (platelet inhibitors) work towards blocking the binding of platelet surface integrins with
specific molecular targets during thrombosis (Cohen, 1996). Generally, aggregation of platelets due to a
variety of agonists including thrombin is mediated via GPIIb/IIIa. When we tested YAP for thrombin and
ADP-induced platelet aggregation separately, dose-dependent inhibitions were observed. Therefore, it
can be suggested that YAP may interact with platelet function via GPIIb/IIIa. For clear understanding of
YAP’s binding ability with other main platelet intergrin GPIa/IIa, adhesion of platelets with collagen was
evaluated. However, none of the YAP concentrations could compete with collagen to bind with GPIa/IIa.
Therefore, it can be suggested that YAP inhibit platelet surface integrin GPIIb/IIIa but not GPIa/IIa.
There is a growing interest in the role of FXII and FXIIa as potent initiators of contact activation-
dependent fibrinolysis. This is partly because, FXII shares structural similarity among some other plasma
fibrinolytic proteins namely, plasminogen, tissue-type plasminogen activator (tPA) and urokinase-type
plasminogen activator (uPA) (Tans and Rosing, 1987). Further, DNA sequence data have proven that
FXII exist multiple domains with extensive sequence homology with regions of tPA and fibronectin
(Cool and MacGillivray, 1987). However, the role of FXII as a plasminogen activator in fibrinolysis is
still unclear. But, some evidences strengthen the fact that FXIIa can induce fibrinolysis indirectly, by
activating prekallikrein into kallikrein, which in turn activates uPA and generate tPA in the presence of
bradykinin (Binnema et al., 1991).
Since YAP could bind with FXIIa, it may also have a possibility to bind with plasminogen, an
important zymogen in plasma fibrinolytic system. In this context, we assessed the plasminogen activator-
like property in YAP, utilizing fibrin plate method. But, all of YAP concentrations tested could not lyse
contact fibrin. This indicates that YAP does not involve in activating plasminogen or cleaving fibrin
cross-links by a direct action (data not shown). Therefore, our results speculate that YAP does not act as
a direct fibrinolytic agent by activating plasminogen. On the other hand, it is reasonable to argue YAP as
an indirect inhibitor of fibrinolysis, since it inhibits FXIIa, which suppose to be an initiator of contact
activation-dependant fibrinolysis. Related literature describes that natural fibrinolytic molecule,
hementin isolated from leeches can activate the fibrinolytic system of human plasma and resembles
functional similarity with streptokinase (Swadesh and Budzynski, 1990). However, in this study we did
not assess the ability of YAP to act on other fibrinolytic proteins like urokinase and streptokinase.
In the present work, we identified the new anticoagulant fish protein as an inhibitor of FXIIa and
platelet membrane integrin GPIIb/IIIa. Therefore, we introduce YAP as a multifunctional anticoagulant,
which exerts its action via different mechanisms. To our knowledge, this is the first report on
anticoagulant activity observed in fish protein and it can be suggest that fish protein hydrolysates are
worthwhile targets of investigating potent bioactive molecules.
Acknowledgement
This work was supported by the Brain Korea 21 project.
References
Bellavite, P., Andrioli, G., Guzzo, P., Arigliano, P., Chirumbolo, S., Manzato, F., Santonastaso, C., 1994. A colorimetric method
for the measurement of platelet adhesion in microtiter plates. Analytical Biochemistry 216, 444–450.

N. Rajapakse et al. / Life Sciences 76 (2005) 2607–2619 2619
Bendar, B., Condra, C., Gould, R.J., Connolly, T.M., 1995. Platelet aggregation monitored in a 96-well microplate reader is
useful for evaluation of platelet agonists and antagonists. Thrombosis Research 77, 453–463.
Binnema, D.J., Dooijewaard, G., Turion, P.N.C., 1991. An analysis of the activators of single-chain urokinase-type plasminogen
activator (scu-PA) in the dextran sulphate euglobulin fraction of normal plasma and of plasmas deficient in factor XII and
prekallikrein. Thrombosis and Haemostasis 65, 144–148.
Clemetson, K.J., Clemetson, J.M., 2001. Platelet collagen receptors. Thrombosis and Haemostasis 86, 189–197.
Cohen, M., 1996. Platelet glycoprotein IIb/IIIa receptor inhibitors in coronary artery diseases. Annals of Internal Medicine 124,
843–844.
Cool, D.E., MacGillivray, R.T.A., 1987. Characterization of the human blood coagulation factor XII gene. The Journal of
Biological Chemistry 262, 13662–13673.
Dametto, M., David, A.P., Azzolini, S.S., Campos, I.T.N., Tanaka, A.M., Gomes, A., Andreotti, R., Tanaka, A.S., 2000.
Purification and characterization of a trypsin-like enzyme with fibrinolytic activity present in the abdomen of horn fly.
Haematobia irritans irritans (Diptera: Muscidae). Journal of Protein Chemistry 19, 515–521.
Depraetere, H., Kerekes, A., Deckmyn, H., 1999. The collagen-binding leech products rLAPP and Calin prevent both von
Willebrand factor and a2h1 (GPIa/IIa)-I-domain binding to collagen in a different manner. Thrombosis and Haemostasis 82,
1160–1163.
Hojima, Y., Pierce, J.V., Pisano, J.J., 1980. Hageman factor fragment inhibitor in corn seeds: purification and characterization.
Thrombosis Research 20, 149–162.
Isawa, H., Yuda, M., Orito, Y., Chinzei, Y., 2002. A mosquito salivary protein inhibits activation of the plasma contact system
by binding to factor XII and high molecular weight kininogen. The Journal of Biological Chemistry 277, 27651–27658.
Jung, W.K., Je, J.Y., Kim, H.J., Kim, S.K., 2002. A novel anticoagulant protein from Scapharca broughtonii. Journal of
Biochemistry and Molecular Biology 35, 199–205.
Kim, S.K., Jeon, Y.J., Byeun, H.G., Kim, Y.T., Lee, C.K., 1997. Enzymatic recovery of cod frame proteins with crude protease
from tuna pyloric caeca. Fisheries Science 63, 421–427.
Laemmli, U.K., 1970. Cleavage of structural protein during the assembly of the head of bacteriophage T4. Nature 227,
680–685.
Lowry, O.H., Rosebrough, A.L., Farr, A.L., Randall, R.J., 1951. Protein measurement with Folin-phenol reagent. The Journal of
Biological Chemistry 193, 165–275.
Miller, G., Silverberg, M., Kaplan, A.P., 1980. Autoactivation of human Hageman factor. Biochemical and Biophysical
Research Communications 92, 803–810.
Phillips, D.R., Charo, I.F., Parise, L.V., Fitzgerald, L.A., 1988. The platelet membrane glycoprotein IIb-IIIa complex. Blood 71,
831–843.
Plow, E.F., Ginsberg, M.H., 1989. Cellular adhesion: GPIIb-IIIa as a prototypic adhesion receptor. Progress in Hemostasis and
Thrombosis 9, 117–156.
Schousboe, I., 1993. Contact activation in human plasma is triggered by zinc ion modulation of factor XII (Hageman factor).
Blood Coagulation and Fibrinolysis 4, 671–678.
Swadesh, J.K., Budzynski, I.Y.H.Z., 1990. Purification and characterization of hementin, a fibrinogenolytic protease from the
leech Haementeria ghilianii. Journal of Chromatography 502, 359–369.
Tanaka-Azevedo, A.M., Tanaka, A.S., Sano-Martins, I.S., 2003. A new blood coagulation inhibitor from the snake Bothrops
jararaka plasma: isolation and characterization. Biochemical and Biophysical Research Communications 308, 706–712.
Tans, G., Rosing, J., 1987. Structural and functional characterization of FXII. Seminars in Thrombosis and Hemostasis 13,
1–14.
Urata, J., Shojo, H., Kaneko, Y., 2003. Inhibition mechanisms of hematophagous invertebrate compounds acting on the host
blood coagulation and platelet aggregation pathways. Biochimie 85, 493–500.
Copyright © 2022 FDOKUMEN




















