
A metaproteomic assessment of winter and summer bacterioplankton from Antarctic Peninsula coastal...
18
ORIGINAL ARTICLE A metaproteomic assessment of winter and summer bacterioplankton from Antarctic Peninsula coastal surface waters Timothy J Williams 1 , Emilie Long 1,2 , Flavia Evans 1 , Mathew Z DeMaere 1 , Federico M Lauro 1 , Mark J Raftery 3 , Hugh Ducklow 4 , Joseph J Grzymski 5 , Alison E Murray 5 and Ricardo Cavicchioli 1 1 School of Biotechnology and Biomolecular Sciences, The University of New South Wales, Sydney, New South Wales, Australia; 2 De´partement de formation ‘Biologie Cellulaire et Mole´culaire’, UFR 927, Universite´ Pierre et Marie Curie (UPMC) Paris VI, Paris, France; 3 Bioanalytical Mass Spectrometry Facility, The University of New South Wales, Sydney, New South Wales, Australia; 4 The Ecosystems Center, Marine Biological Laboratory, Woods Hole, MA, USA and 5 Desert Research Institute, Division of Earth and Ecosystem Sciences, Reno, NV, USA A metaproteomic survey of surface coastal waters near Palmer Station on the Antarctic Peninsula, West Antarctica, was performed, revealing marked differences in the functional capacity of summer and winter communities of bacterioplankton. Proteins from Flavobacteria were more abundant in the summer metaproteome, whereas winter was characterized by proteins from ammonia-oxidizing Marine Group I Crenarchaeota. Proteins prevalent in both seasons were from SAR11 and Rhodobacterales clades of Alphaproteobacteria, as well as many lineages of Gammaproteobacteria. The metaproteome data were used to elucidate the main metabolic and energy generation pathways and transport processes occurring at the microbial level in each season. In summer, autotrophic carbon assimilation appears to be driven by oxygenic photoautotrophy, consistent with high light availability and intensity. In contrast, during the dark polar winter, the metaproteome supported the occurrence of chemolithoautotrophy via the 3-hydroxypropionate/4-hydroxybutyrate cycle and the reverse tricarboxylic acid cycle of ammonia-oxidizing archaea and nitrite-oxidizing bacteria, respectively. Proteins involved in nitrification were also detected in the metaproteome. Taurine appears to be an important source of carbon and nitrogen for heterotrophs (especially SAR11), with transporters and enzymes for taurine uptake and degradation abundant in the metaproteome. Divergent heterotrophic strategies for Alphaproteobacteria and Flavobacteria were indicated by the metaproteome data, with Alphaproteobacteria capturing (by high-affinity transport) and processing labile solutes, and Flavobacteria expressing outer membrane receptors for particle adhesion to facilitate the exploitation of non-labile substrates. TonB-dependent receptors from Gammaproteo- bacteria and Flavobacteria (particularly in summer) were abundant, indicating that scavenging of substrates was likely an important strategy for these clades of Southern Ocean bacteria. This study provides the first insight into differences in functional processes occurring between summer and winter microbial communities in coastal Antarctic waters, and particularly highlights the important role that ‘dark’ carbon fixation has in winter. The ISME Journal (2012) 6, 1883–1900; doi:10.1038/ismej.2012.28; published online 26 April 2012 Subject Category: integrated genomics and post-genomics approaches in microbial ecology Keywords: marine microorganisms; metaproteomics; antarctic microbiology; southern ocean microbiology Introduction The Southern Ocean encircles the Antarctic con- tinent, and influences the climate of the entire Earth. It is a major sink for global carbon dioxide emissions, accounting for around 30% of global ocean uptake of CO 2 (AASSP, 2011). Global thermo- haline circulation is driven by the formation of deep water masses in the Southern Ocean that, in turn, drive global nutrient cycling throughout the oceans. As the southernmost part of the global ocean Correspondence: R Cavicchioli, School of Biotechnology and Biomolecular Sciences, The University of New South Wales, BABS, Sydney, New South Wales 2052, Australia. E-mail: [email protected] or AE Murray, Desert Research Institute, Division of Earth and Ecosystem Sciences, 2215 Raggio Parkway, Reno, NV 89512, USA. E-mail: [email protected] Received 23 October 2011; revised 23 February 2012; accepted 24 February 2012; published online 26 April 2012 The ISME Journal (2012) 6, 1883–1900 & 2012 International Society for Microbial Ecology All rights reserved 1751-7362/12 www.nature.com/ismej
Transcript of A metaproteomic assessment of winter and summer bacterioplankton from Antarctic Peninsula coastal...

ORIGINAL ARTICLE
A metaproteomic assessment of winter and summerbacterioplankton from Antarctic Peninsula coastalsurface waters
Timothy J Williams1, Emilie Long1,2, Flavia Evans1, Mathew Z DeMaere1,Federico M Lauro1, Mark J Raftery3, Hugh Ducklow4, Joseph J Grzymski5, Alison E Murray5
and Ricardo Cavicchioli1
1School of Biotechnology and Biomolecular Sciences, The University of New South Wales, Sydney,New South Wales, Australia; 2Departement de formation ‘Biologie Cellulaire et Moleculaire’, UFR 927,Universite Pierre et Marie Curie (UPMC) Paris VI, Paris, France; 3Bioanalytical Mass Spectrometry Facility,The University of New South Wales, Sydney, New South Wales, Australia; 4The Ecosystems Center, MarineBiological Laboratory, Woods Hole, MA, USA and 5Desert Research Institute, Division of Earth and EcosystemSciences, Reno, NV, USA
A metaproteomic survey of surface coastal waters near Palmer Station on the Antarctic Peninsula,West Antarctica, was performed, revealing marked differences in the functional capacity of summerand winter communities of bacterioplankton. Proteins from Flavobacteria were more abundant in thesummer metaproteome, whereas winter was characterized by proteins from ammonia-oxidizingMarine Group I Crenarchaeota. Proteins prevalent in both seasons were from SAR11 andRhodobacterales clades of Alphaproteobacteria, as well as many lineages of Gammaproteobacteria.The metaproteome data were used to elucidate the main metabolic and energy generation pathwaysand transport processes occurring at the microbial level in each season. In summer, autotrophiccarbon assimilation appears to be driven by oxygenic photoautotrophy, consistent with high lightavailability and intensity. In contrast, during the dark polar winter, the metaproteome supported theoccurrence of chemolithoautotrophy via the 3-hydroxypropionate/4-hydroxybutyrate cycle and thereverse tricarboxylic acid cycle of ammonia-oxidizing archaea and nitrite-oxidizing bacteria,respectively. Proteins involved in nitrification were also detected in the metaproteome. Taurineappears to be an important source of carbon and nitrogen for heterotrophs (especially SAR11), withtransporters and enzymes for taurine uptake and degradation abundant in the metaproteome.Divergent heterotrophic strategies for Alphaproteobacteria and Flavobacteria were indicated by themetaproteome data, with Alphaproteobacteria capturing (by high-affinity transport) and processinglabile solutes, and Flavobacteria expressing outer membrane receptors for particle adhesion tofacilitate the exploitation of non-labile substrates. TonB-dependent receptors from Gammaproteo-bacteria and Flavobacteria (particularly in summer) were abundant, indicating that scavenging ofsubstrates was likely an important strategy for these clades of Southern Ocean bacteria. This studyprovides the first insight into differences in functional processes occurring between summer andwinter microbial communities in coastal Antarctic waters, and particularly highlights the importantrole that ‘dark’ carbon fixation has in winter.The ISME Journal (2012) 6, 1883–1900; doi:10.1038/ismej.2012.28; published online 26 April 2012Subject Category: integrated genomics and post-genomics approaches in microbial ecologyKeywords: marine microorganisms; metaproteomics; antarctic microbiology; southern oceanmicrobiology
Introduction
The Southern Ocean encircles the Antarctic con-tinent, and influences the climate of the entireEarth. It is a major sink for global carbon dioxideemissions, accounting for around 30% of globalocean uptake of CO2 (AASSP, 2011). Global thermo-haline circulation is driven by the formation of deepwater masses in the Southern Ocean that, in turn,drive global nutrient cycling throughout the oceans.As the southernmost part of the global ocean
Correspondence: R Cavicchioli, School of Biotechnology andBiomolecular Sciences, The University of New South Wales,BABS, Sydney, New South Wales 2052, Australia.E-mail: [email protected] orAE Murray, Desert Research Institute, Division of Earth andEcosystem Sciences, 2215 Raggio Parkway, Reno, NV 89512, USA.E-mail: [email protected] 23 October 2011; revised 23 February 2012; accepted 24February 2012; published online 26 April 2012
The ISME Journal (2012) 6, 1883–1900& 2012 International Society for Microbial Ecology All rights reserved 1751-7362/12
www.nature.com/ismej

network, this polar region undergoes extreme sea-sonal variations in sea-ice cover and illumination,which greatly influences the primary productivity ofthe indigenous marine microorganisms. During theaustral summer, Antarctic surface water is exposedto continuous high solar irradiation causing sea-iceand glacier-ice melt, and the waters are highlyproductive. In contrast, the prolonged cold anddarkness during the austral winter permits sea-iceexpansion and minimal or no photosynthetic pri-mary production.
Although microorganisms that depend uponoxygenic photosynthesis decline during the winterseason, some groups of microorganisms increase inabundance (Murray and Grzymski, 2007). Shifts inmicrobial community composition between summerand winter have been documented (Murray et al.,1998; Massana et al., 2001; Murray and Grzymski,2007; Piquet et al., 2011; Ghiglione and Murray,2012; Grzymski et al., 2012). However, little isknown about global gene expression of SouthernOcean microbial communities as relevant studies(for example, metaproteomics, metatranscriptomicsand stable isotope metagenomics) have not beenperformed.
By determining which proteins have been synthe-sized by microorganisms present at the time ofsampling, metaproteomics enables the reconstruc-tion of microbial processes and metabolic pathwaysthat are central to the functioning of the ecosystem.Marine metaproteomic investigations have beenperformed on the ocean gyre of the Sargasso Sea(Sowell et al., 2009), ocean gyre and coastal sites inthe South Atlantic (Morris et al., 2010), and a coastalregion near Oregon (Sowell et al., 2011). In Antarcticenvironments, metaproteomics has been applied totwo marine-derived lakes (Ng et al., 2010; Lauroet al., 2011; Yau et al., 2011). These studies havebeen facilitated by an expanding database ofgenomic and metagenomic data that provides aframework for the identification of proteins by massspectrometry (Thomas et al., 2007).
In the current study, we used metaproteomics toelucidate the predominant metabolic processes per-formed by bacterioplankton present in surface coastalwaters of the West Antarctic Peninsula (WAP). Thissite, in the coastal waters of Anvers Island, is in thestudy region of the Palmer Long Term EcologicalResearch (LTER) program, where a 420-year researchprogram has focused on the Antarctic pelagic marineecosystem. This region is experiencing one of themost rapid warming events on Earth (Turner et al.,2005). Using a customized metagenomic databasethat included metagenome data for summer andwinter communities from the WAP (Grzymski et al.,2012), metagenome sequence from six librariessequenced from the Southern Ocean off East Antarc-tica and genome sequences of numerous marinemicroorganisms, we identified proteins from AntarcticPeninsula summer and winter metaproteomesamples collected during the International Polar Year.
By examining expressed proteins in the samples, wewere able to infer the major trophic pathways andprocesses and consider the significance of thedifferences between summer and winterbacterioplankton.
Materials and methods
Sample collectionSeawater samples were collected from coastal sur-face waters off of Anvers Island, Western AntarcticPeninsula. Samples for metaproteomic analysis(200 l each) were collected near LTER Station Bon six occasions in the austral summer of 2008(23 February, 26 February, 28 February, 1 March and4 March), and on three dates in austral winter of2008 (28 July, 12 August and 19 August) at LTERStation B on the first two samples and at theseawater intake for the third sample. The depth ofthe water column at Station B is B70 m and B6 m atthe seawater intake. Physiochemical data collectedcoincident with metaproteome samples were aver-aged across sample dates in 2008 to provide anenvironmental context for the conditions betweensummer and winter. Seawater physical, chemicaland biological properties were determined usingstandard LTER methods (http://pal.lternet.edu/pub-lications/documents/protocols/). 3H-Leucine incor-poration rates were performed as describedpreviously (Kirchman et al., 1985). Dissolvedorganic carbon (DOC) was measured with a Shi-madzu TOC-V high temperature combustion totalcarbon analyzer as described previously (Sharpet al., 1993; Farmer and Hansell, 2007). Seawaterwas processed by prefiltering through sequential5.0- and 2.5-mm filters (Polygard, Millipore,Billerica, MA, USA) and cells concentrated using atangential flow filtration system and the cellsharvested on 25-mm 0.2-mm Supor filters(Millipore). Samples were immersed in a proteaseinhibitor solution (Ng et al., 2010), which included2.5 mM EGTA, 2.5 mM EDTA, 0.1 mM Tris-EDTA (pH8), 1 mM PMSF (freshly prepared) and 50 mm pro-tease inhibitor cocktail VI (Calbiochem, San Diego,CA, USA), and the sample tubes placed into liquidN2 and stored at � 80 1C until extraction.
MetaproteomicsProtein extraction from filter membranes followedthe method of Ng et al. (2010), using 20-ml lysisbuffer containing 10 mM Tris-EDTA (pH 8.0; Univar,Sydney, Australia), 20 ml of protease inhibitor cock-tail VI (Calbiochem), 0.1% sodium dodecyl sulfate(Univar) and 1 mM dithiothreitol (Sigma-Aldrich,Sydney, Australia). The only departure from themethod was that five sonications were carried outat the following settings: 40 s intervals, 0.5 s pulseon/0.5 s pulse off, 20% amplitude and that 3-kDarather than 5-kDa Amicon filter units were used.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1884
The ISME Journal

One-dimensional SDS–PAGE and in-gel trypsindigestion also followed Ng et al. (2010), except gelswere stained with Coomassie blue. Digested pep-tides were separated by nano-LC using an Ultimate3000 HPLC and autosampler system (Dionex,Amsterdam, Netherlands). Samples (2.5 ml) wereconcentrated and de-salted onto a micro C18precolumn (500 mm� 2 mm, Michrom Bioresources,Auburn, CA, USA) with H2O:CH3CN (98:2, 0.1%TFA) at 15 ml min� 1. After a 4-min wash the pre-column was switched (Valco 10 port valve, Dionex)into line with a fritless nano column (75 m�B10 cm) containing C18 media (5 m, 200 A Magic,Michrom Bioresources). Peptides were eluted usinga linear gradient of H2O:CH3CN (98:2, 0.1% formicacid) to H2O:CH3CN (64:36, 0.1% formic acid) at250 nl min� 1 over 30 min. High voltage (2000 V) wasapplied to low volume tee (Upchurch Scientific,Oak Harbor, WA, USA) and the column tip positio-nedB0.5 cm from the heated capillary (T¼ 280 1C)of an Orbitrap Velos (Thermo Electron, Bremen,Germany) mass spectrometer. Positive ions weregenerated by electrospray and the Orbitrap operatedin data-dependent acquisition mode. A survey scanm/z 350–1750 was acquired in the Orbitrap(Resolution¼ 30 000 at m/z 400, with an accumula-tion target value of 1 000 000 ions) with lockmassenabled. Up to the 10 most abundant ions (45000counts) with charge states 4þ 2 were sequentiallyisolated and fragmented within the linear ion trapusing collisionally induced dissociation with anactivation q¼ 0.25 and activation time of 30 ms at atarget value of 30 000 ions. m/z ratios selected forMS/MS were dynamically excluded for 30 s. Peaklists were generated using Mascot Daemon/extract_msn (Matrix Science, Thermo, London,UK) using the default parameters, and submittedto Mascot 2.1 (Matrix Science). An Antarcticmetaproteome sequence database, AntComb, wasconstructed from: fosmid libraries (IMG Acc:2008193000, 2008193001, 2012990003, 2040502005and 2040502004), individual marine microbialgenomes (RefSeq Project ID: 202, 58903, 54247,58401, 59427, 57855, 54575, 54169, 51877, 54583,54265, 54403, 54255, 54577, 54163, 54227, 54185,58403, 54207, 54205, 54623, 54467, 58597, 54183,57863, 52598, 54259, 58183, 13044 and 51609) andAntarctic (Southern Ocean off East Antarctica)metagenome samples (SRA Acc: SRX024734,SRX024735, SRX024799, SRX025108, SRX024736and SRX024800). Gene predictions for fosmiddata were taken from IMG. Fosmid end, or fullysequenced fosmids, were annotated through theIMG, which incorporates a full annotation systemincluding BlastP to the RefSeq database (Markowitzet al., 2012). Predictions for Celera WGS assembledmetagenome data were produced in-house withSHAP (DeMaere et al., 2011). The combined set oftranslated gene predictions were reduced to a non-redundant set with cdhit-est using a global sequenceidentity cutoff of 100%. MS/MS data analysis and
validation of protein identifications were performedas previously described (Ng et al., 2010) except thatthe databases used were NR and AntComb. Mascotsearches with a false discovery rate 45% wererejected. All other Mascot results were combinedand validated using a MudPit analysis in Scaffold3.0 (Proteome Software Inc., Portland, OR, USA)using the following parameters: minimal probabilityof peptide identification, 95%; minimal probabilityof protein identification, 99%. Protein matches wereonly accepted if they were identified by a minimumof two unique peptides. All proteins were manuallyannotated with the aid of BLASTP (Allen et al.,2009), and the protein hit that showed the highestsequence identity was recorded, including theorganism name (Supplementary Table S1). Todetermine if individual phylogenetic groups andinferred metabolic functions exhibited seasonalvariation, spectral counts were analyzed statisticallyby using the resampling method implemented in thescript ALL_scrambler.pl (Allen et al., 2009). Briefly,a subset (n¼ 20 000) of the spectra from summer andwinter was selected at random, with replacement,classified functionally or phylogenetically, andcompared. This process was repeated 10 000 timesand the median difference between groups wasrecorded. The statistically significant differenceswere then assessed at a 99% confidence level asdescribed previously (Rodriguez-Brito et al., 2006).
Results and discussion
Physicochemical data for the summer and wintersamplesChlorophyll a values were 1.2–3.8 mg l�1 for summerand 0.037–0.171 mg l�1 for winter. DOC values forsummer and winter samples were 48.8 and41.8 mmol l�1, respectively. Bacterial production insummer was much higher as indicated by leucineincorporation rates: average 12.7±1.6 vs 2±0.4 pmolleucine h�1. Average water temperature was 1.4 1C and� 0.92 1C in summer and winter, respectively.Nutrient levels averaged 1.53±0.04mM PO4, 55.22±6.15mM Si(OH)4 and 30.45 0.58mM NO2
� þNO3� for
the last three summer samples (29 Febuary–6 March)and 2.03±0.04mM PO4, 51.75±7.40mM Si(OH)4 and30.56±0.16mM NO2
� þNO3� over the three winter
sample dates.
Phylogenetic diversity represented in the AntarcticmetaproteomeA total of 1061 proteins were detected (310 uniqueto the austral summer, 349 unique to the australwinter and 402 that were detected in both data sets);the full list of proteins is in SupplementaryTable S1. Of the 1061 identified proteins, 755proteins were from the AntComb database, 323 werefrom NR and 17 were common to both. This suggeststhat protein sequence data were far better repre-sented in fosmids and metagenomic data than in
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1885
The ISME Journal

microbial genomes contained in NR. Of the 1061proteins, these included 831 proteins with highestsequence identity to bacterial proteins, 189 archaealproteins consisting mainly of Marine Group ICrenarchaeota (MGI; 188), 32 eukaryotic proteinsand seven phage proteins. Within the bacterialsubset, the majority had the best match (highestsequence identity) to proteins from members of theAlphaproteobacteria (404), followed by Gammapro-teobacteria (322), Bacteroidetes (56), Betaproteobac-teria (18), Nitrospirae (4) and Cyanobacteria (4).Matches to the Bacteroidetes clade (¼ Cytophaga-Flavobacterium-Bacteroides or CFB group) weremainly to Flavobacteria (54 proteins). Our data areconsistent with the phylogenetic diversity obtainedby previous gene-based analyses of coastal SouthernOcean waters, which identified the relative abun-dance of MGI, Alphaproteobacteria, Gammaproteo-bacteria and Flavobacteria (Murray and Grzymski,2007; Ghiglione and Murray, 2012; Grzymski et al.,2012). Cyanobacteria have been reported to be anegligible component of the marine picoplankton inAntarctic waters (Ghiglione and Murray, 2012).Nevertheless, we detected proteins with bestmatches to cyanobacterial viruses (cyanophages),including capsid protein Gp23 from Myoviruses(Supplementary Table S1). These cyanophage canexceed 200 nm in length (Sullivan et al., 2005)allowing them to be preferentially captured on thefilters. Consistent with this, low numbers of geneswith matches to Prochlorococcus spp., Synechococ-cus spp. and their phages have been detected fromlarge-scale shotgun metagenomics analyses of pela-gic Southern Ocean waters from East Antarctica(Wilkins D et al., unpublished data), but not in theAntarctic Peninsula metagenomes reported byGrzymski et al. (2012).
Within the Alphaproteobacteria, nearly half (199)of the matches had highest identity to proteins fromthe SAR11 clade (including Candidatus Pelagibacterubique); other proteins had the best match tomembers of Rhodobacterales (129), Sphingomona-dales (37), Rhizobiales (21) and uncultivatedSAR116 cluster (9). Within Rhodobacterales, pro-teins affiliated with the Roseobacter clade predomi-nated (105). The majority of Sphingomonadalesproteins had 100% sequence identity to the marineoligotroph Sphingopyxis alaskensis. Within theGammaproteobacteria, most of the proteins had thebest match to members of four groups: Alteromona-dales (83), gammaproteobacterial sulfur oxidizerEOSA-1 (GSO–EOSA-1) complex (Walsh et al.,2009; Swan et al., 2011) (72), Oceanospirillales(63) and the Oligotrophic Marine Gammaproteobac-teria (OMG) group (Cho and Giovannoni, 2004) (39).Alteromonadales were represented by proteins with100% sequence identity to Pseudoalteromonashaloplanktis, Moritella sp. PE36 and bacteriumTW-7. Matches to the GSO–EOSA-I complex areindicative of the presence of the uncultivated sistergroups SUP05 and/or ARCTIC96BD-19 (see section
GSO–EOSA-1 bacteria ARCTIC96BD-19/SUP05below). Within Oceanospirillales, the most commonmatches were to proteins with the highest identity toNeptuniibacter caesariensis or Marinomonas spp.Proteins from the Bacteroidetes had the bestmatch to proteins from Flavobacteria, includingPolaribacter spp., Zunongwangia profunda,Psychroflexus torquis, and the uncultivated marinebacterium MS024-2A.
All the archaeal proteins matched best to mem-bers of MGI (detected exclusively in winter), exceptfor two that matched to the Antarctic methanogenMethanococcoides burtonii. MGI proteins had thebest match to the ammonia-oxidizing archaea (AOA)Nitrosopumilus maritimus or Cenarchaeum symbio-sum (Hallam et al., 2006a, b; Walker et al., 2010).The dominance of proteins for MGI is consistentwith the minimal phylogenetic diversity previouslyobserved for the Archaea in surface waters ofthe Southern Ocean compared with the diversity ofthe Bacteria (Massana et al., 2001; Murray andGrzymski, 2007; Grzymski et al., 2012).
Matches to eukaryotic phytoplankton were moreabundant in summer with 32 and 8 total matches insummer vs winter, respectively. Spectral countswere also significantly higher for eukaryoticproteins in summer. These proteins includedphotosystem proteins and ribulose bisphosphatecarboxylase/oxygenase (RuBisCO) from algae, con-sistent with algae being the major drivers of CO2
fixation in summer (Murray and Grzymski, 2007).Phytoplankton is undoubtedly the dominant agentof light-mediated carbon fixation in summer forwaters off the Antarctic Peninsula. Nanoplanktonand larger cells would be expected to be capturedby the 1.6- or 2.5-mm prefilters that were used forsampling (see Materials and methods). The eukar-yotic proteins present on the 0.2-mm filters are likelyto derive from organismal turnover, or from suffi-ciently small unicellular algae such as Ostreococcustauri. Although bacterial RuBisCO was not detectedin the metaproteome, a chemolithoautotroph-relatedform I RuBisCO was present in several fosmidssequenced in the WAP metagenome (Grzymski et al.,2012), and the winter metaproteome did contain abacterial CbbQ protein (Supplementary Table S1),which may have a role in post-translation activationof RuBisCO (Hayashi et al., 1997).
Phylogenetic differences between summer and winterThe protein assignments illustrated that specifictaxa of bacteria and archaea had different seasonalabundances (Figure 1). Spectra assigned to MGI,SAR11 and GSO–EOSA-1 were significantly moreabundant in the winter metaproteome, whereasspectra assigned to Bacteroidetes, Sphingomonadales,Oceanospirillales, Alteromonadales and OMG weresignificantly more abundant in the summer meta-proteome. By contrast, the contribution of SAR11proteins to inferred phylogenetic diversity was
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1886
The ISME Journal
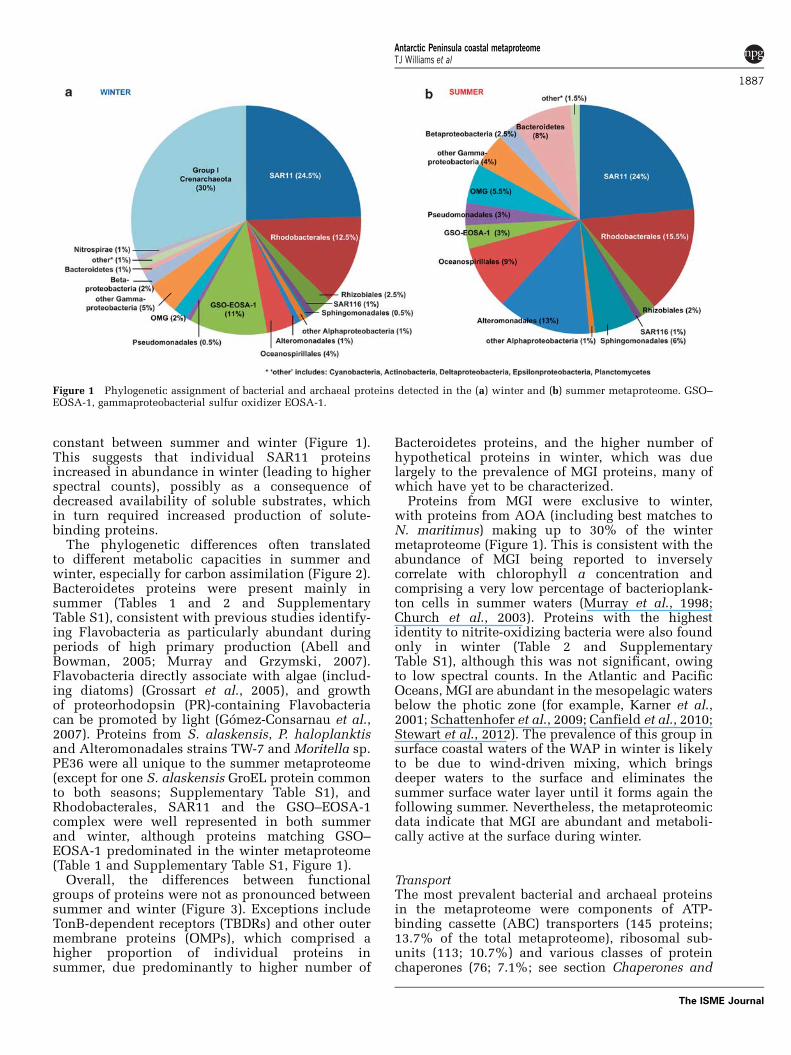
constant between summer and winter (Figure 1).This suggests that individual SAR11 proteinsincreased in abundance in winter (leading to higherspectral counts), possibly as a consequence ofdecreased availability of soluble substrates, whichin turn required increased production of solute-binding proteins.
The phylogenetic differences often translatedto different metabolic capacities in summer andwinter, especially for carbon assimilation (Figure 2).Bacteroidetes proteins were present mainly insummer (Tables 1 and 2 and SupplementaryTable S1), consistent with previous studies identify-ing Flavobacteria as particularly abundant duringperiods of high primary production (Abell andBowman, 2005; Murray and Grzymski, 2007).Flavobacteria directly associate with algae (includ-ing diatoms) (Grossart et al., 2005), and growthof proteorhodopsin (PR)-containing Flavobacteriacan be promoted by light (Gomez-Consarnau et al.,2007). Proteins from S. alaskensis, P. haloplanktisand Alteromonadales strains TW-7 and Moritella sp.PE36 were all unique to the summer metaproteome(except for one S. alaskensis GroEL protein commonto both seasons; Supplementary Table S1), andRhodobacterales, SAR11 and the GSO–EOSA-1complex were well represented in both summerand winter, although proteins matching GSO–EOSA-1 predominated in the winter metaproteome(Table 1 and Supplementary Table S1, Figure 1).
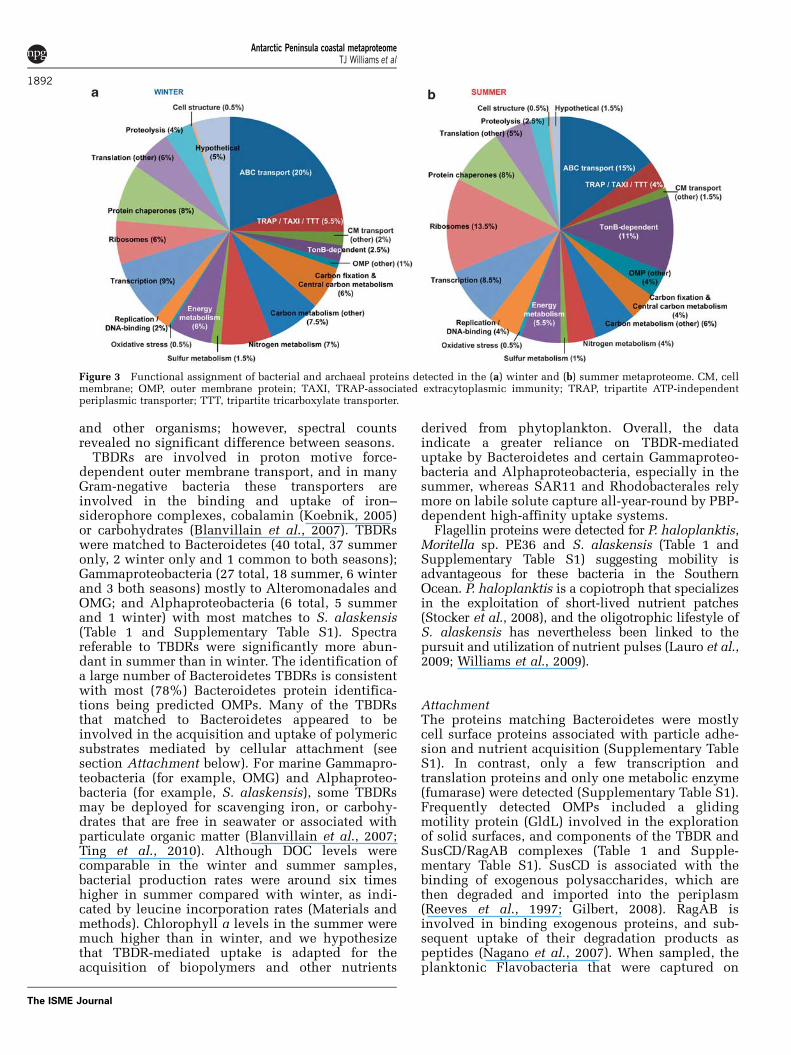
Overall, the differences between functionalgroups of proteins were not as pronounced betweensummer and winter (Figure 3). Exceptions includeTonB-dependent receptors (TBDRs) and other outermembrane proteins (OMPs), which comprised ahigher proportion of individual proteins insummer, due predominantly to higher number of
Bacteroidetes proteins, and the higher number ofhypothetical proteins in winter, which was duelargely to the prevalence of MGI proteins, many ofwhich have yet to be characterized.
Proteins from MGI were exclusive to winter,with proteins from AOA (including best matches toN. maritimus) making up to 30% of the wintermetaproteome (Figure 1). This is consistent with theabundance of MGI being reported to inverselycorrelate with chlorophyll a concentration andcomprising a very low percentage of bacterioplank-ton cells in summer waters (Murray et al., 1998;Church et al., 2003). Proteins with the highestidentity to nitrite-oxidizing bacteria were also foundonly in winter (Table 2 and SupplementaryTable S1), although this was not significant, owingto low spectral counts. In the Atlantic and PacificOceans, MGI are abundant in the mesopelagic watersbelow the photic zone (for example, Karner et al.,2001; Schattenhofer et al., 2009; Canfield et al., 2010;Stewart et al., 2012). The prevalence of this group insurface coastal waters of the WAP in winter is likelyto be due to wind-driven mixing, which bringsdeeper waters to the surface and eliminates thesummer surface water layer until it forms again thefollowing summer. Nevertheless, the metaproteomicdata indicate that MGI are abundant and metaboli-cally active at the surface during winter.
TransportThe most prevalent bacterial and archaeal proteinsin the metaproteome were components of ATP-binding cassette (ABC) transporters (145 proteins;13.7% of the total metaproteome), ribosomal sub-units (113; 10.7%) and various classes of proteinchaperones (76; 7.1%; see section Chaperones and
Figure 1 Phylogenetic assignment of bacterial and archaeal proteins detected in the (a) winter and (b) summer metaproteome. GSO–EOSA-1, gammaproteobacterial sulfur oxidizer EOSA-1.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1887
The ISME Journal

quality control below) (Table 1 and SupplementaryTable S1). Proteins involved in high-affinity transportwere highly represented, similar to findings frommetaproteomic studies of the Sargasso Sea (Sowellet al., 2009) and Oregon coast (Sowell et al., 2011).
The majority of ABC transport proteins wereperiplasmic-binding proteins (PBPs), which weremuch more abundant (138 of the 145 total) than themembrane-associated ATPase and permease compo-nents (Supplementary Table S1). Other high-affinitytransporters include tripartite ATP-independentperiplasmic (TRAP) transporters (32 proteins,including TAXI) and tripartite tricarboxylate trans-porters (TTT, 5 proteins), and both types wererepresented in the metaproteome solely by PBPs(Supplementary Table S1).
The relatively high abundance of PBPs relative toATPase and/or permease components can be attrib-uted to high expression levels to enhance thefrequency of solute capture. Achieving conversionof the imported substrates also requires a relativelyminimal number of cytoplasmic enzymes relative toPBPs. Membrane proteins are also more difficult toextract and solubilize from biomass thereby poten-tially reducing their frequency of detection relative to
inherently soluble proteins such as PBPs (Burg et al.,2010; Morris et al., 2010; Williams et al., 2010).However, despite not using methods to specificallyenrich for membrane proteins, we detected severalclasses of OMPs including 73 TBDRs and cellmembrane proteins such as PR (3), ammoniumtransporters (3) and sodium/solute symporters (4;Table 1 and Supplementary Table S1).
ABC, TRAP and TTT transporter proteins wereidentified in summer and winter samples, and themajority (85%) matched to marine Alphaproteobac-teria (Table 1 and Supplementary Table S1). A largenumber of PBPs of ABC transporters matched tomembers of SAR11 (54) or Roseobacter (43) clades.Based on sequence identity, these were inferred to bespecific for polyamines, amino acids, carbohydrates,taurine/sulfonates, oligopeptides, glycerol-3-phos-phate, phosphonate or iron. Based on spectral counts,ABC transporter PBPs (for general amino acids,branched-chain amino acids, glycine betaine andsugars) and TRAP transporter PBPs were significantlymore abundant in winter than in summer.
The large number of ABC transporter PBPs foramino acids and polyamines (putrescine/spermi-dine) that appear to derive from members of the
Figure 2 Depiction of the metabolic characteristics of winter and summer communities inferred from the metaproteome. Individualgroups were inferred to be present based on best protein matches. Eukaryotic groups are indicated by squares, bacterial clades by ellipsesand the sole shown archaeal clade by a diamond. Groups are shaded based on their seasonal distribution: predominantly found in winter(blue) vs summer (red), or abundant in both seasons (green). The microbial processes that appear to be the most important in the cyclingof carbon, nitrogen and sulfur, and for energy metabolism are shown. For convenience, dissolved inorganic carbon is shown as CO2.Pathways or processes that are particularly important for energy generation are signified using a yellow lightning bolt. ABC, ATP-bindingcassette; DMSP, dimethylsulfoniopropionate; OMP, outer membrane proteins; PR, proteorhodopsin; TBDR, TonB-dependent receptor;TRAP, tripartite ATP-independent periplasmic transporter; TTT, tripartite tricarboxylate transporter.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1888
The ISME Journal
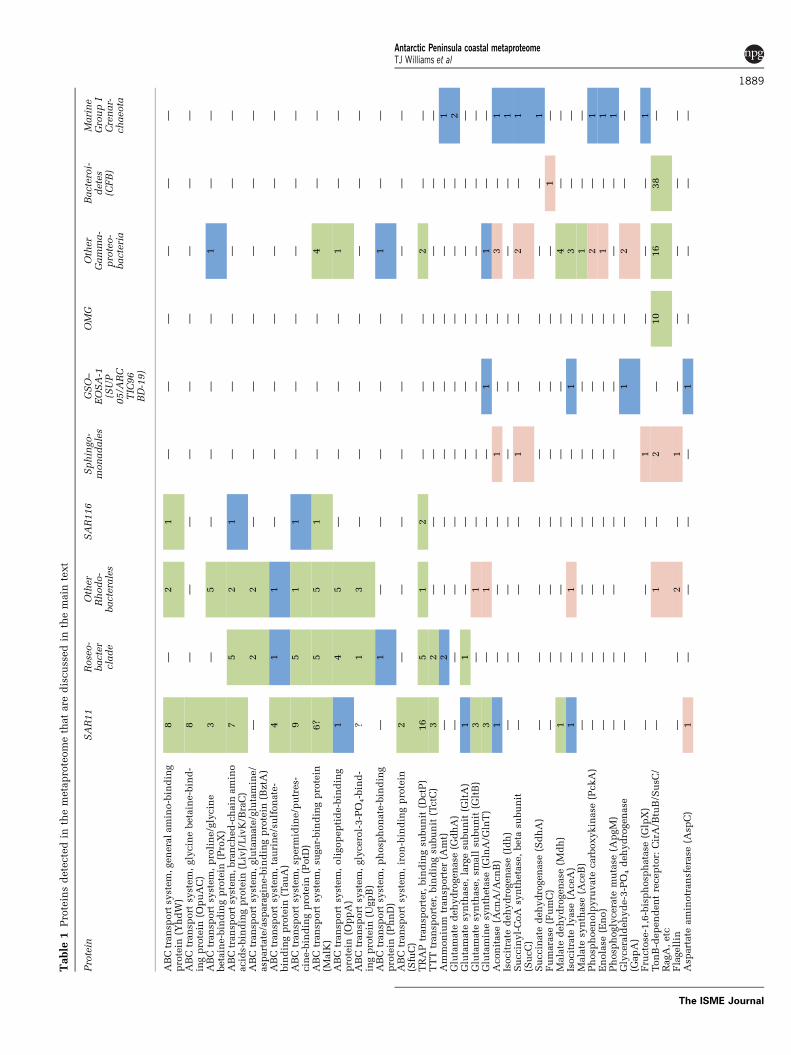
Table
1P
rote
ins
dete
cted
inth
em
eta
pro
teom
eth
at
are
dis
cu
ssed
inth
em
ain
text
Pro
tein
SA
R11
Rose
o-
bacte
rcla
de
Oth
er
Rh
od
o-
bacte
rale
s
SA
R116
Sp
hin
go-
mon
ad
ale
sG
SO
–E
OS
A-1
(SU
P05/A
RC
TIC
96
BD
-19)
OM
GO
ther
Gam
ma-
pro
teo-
bacte
ria
Bacte
roi-
dete
s(C
FB
)
Mari
ne
Gro
up
IC
ren
ar-
ch
aeota
AB
Ctr
an
sport
syst
em
,gen
era
lam
ino-b
ind
ing
pro
tein
(Yh
dW
)8
—2
1—
——
——
—
AB
Ctr
an
sport
syst
em
,gly
cin
ebeta
ine-b
ind
-in
gp
rote
in(O
pu
AC
)8
——
——
——
——
—
AB
Ctr
an
sport
syst
em
,p
roli
ne/g
lycin
ebeta
ine-b
ind
ing
pro
tein
(Pro
X)
3—
5—
——
—1
——
AB
Ctr
an
sport
syst
em
,bra
nch
ed
-ch
ain
am
ino
acid
s-bin
din
gp
rote
in(L
ivJ/
Liv
K/B
raC
)7
52
1—
——
——
—
AB
Ctr
an
sport
syst
em
,glu
tam
ate
/glu
tam
ine/
asp
art
ate
/asp
ara
gin
e-b
ind
ing
pro
tein
(BztA
)—
22
——
——
——
—
AB
Ctr
an
sport
syst
em
,ta
uri
ne/s
ulf
on
ate
-bin
din
gp
rote
in(T
au
A)
41
1—
——
——
——
AB
Ctr
an
sport
syst
em
,sp
erm
idin
e/p
utr
es-
cin
e-b
ind
ing
pro
tein
(PotD
)9
51
1—
——
——
—
AB
Ctr
an
sport
syst
em
,su
gar-
bin
din
gp
rote
in(M
alK
)6?
55
1—
——
4—
—
AB
Ctr
an
sport
syst
em
,oli
gop
ep
tid
e-b
ind
ing
pro
tein
(Op
pA
)1
45
——
——
1—
—
AB
Ctr
an
sport
syst
em
,gly
cero
l-3-P
O4-b
ind
-in
gp
rote
in(U
gp
B)
?1
3—
——
——
——
AB
Ctr
an
sport
syst
em
,p
hosp
hon
ate
-bin
din
gp
rote
in(P
hn
D)
—1
——
——
—1
——
AB
Ctr
an
sport
syst
em
,ir
on
-bin
din
gp
rote
in(S
fuC
)2
——
——
——
——
—
TR
AP
tran
sport
er,
bin
din
gsu
bu
nit
(DctP
)16
51
2—
——
2—
—T
TT
tran
sport
er,
bin
din
gsu
bu
nit
(TctC
)3
2—
——
——
——
—A
mm
on
ium
tran
sport
er
(Am
t)—
2—
——
——
——
1G
luta
mate
deh
yd
rogen
ase
(Gd
hA
)—
——
——
——
——
2G
luta
mate
syn
thase
,la
rge
subu
nit
(Glt
A)
11
——
——
——
——
Glu
tam
ate
syn
thase
,sm
all
subu
nit
(Glt
B)
3—
1—
——
——
——
Glu
tam
ine
syn
theta
se(G
lnA
/Gln
T)
3—
1—
—1
—1
——
Acon
itase
(Acn
A/A
cn
B)
1—
——
1—
—3
—1
Isocit
rate
deh
yd
rogen
ase
(Id
h)
——
——
——
——
—1
Su
ccin
yl-
CoA
syn
theta
se,
beta
subu
nit
(Su
cC
)—
——
—1
——
2—
1
Su
ccin
ate
deh
yd
rogen
ase
(Sd
hA
)—
——
——
——
——
1F
um
ara
se(F
um
C)
——
——
——
——
1—
Mala
ted
eh
yd
rogen
ase
(Md
h)
1—
——
——
—4
——
Isocit
rate
lyase
(AceA
)1
—1
——
1—
3—
—M
ala
tesy
nth
ase
(AceB
)—
——
——
——
1—
—P
hosp
hoen
olp
yru
vate
carb
oxykin
ase
(PckA
)—
——
——
——
2—
1E
nola
se(E
no)
——
——
——
—1
—1
Ph
osp
hogly
cera
tem
uta
se(A
pgM
)—
——
——
——
——
1G
lycera
ldeh
yd
e-3
-PO
4d
eh
yd
rogen
ase
(Gap
A)
——
—1
—2
——
Fru
cto
se-1
,6-b
isp
hosp
hata
se(G
lpX
)—
——
—1
——
—1
Ton
B-d
ep
en
den
tre
cep
tor:
Cir
A/B
tuB
/Su
sC/
RagA
,etc
—1
—2
—10
16
38
—
Fla
gell
in—
—2
—1
——
——
—A
spart
ate
am
inotr
an
sfera
se(A
spC
)1
——
——
1—
——
—
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1889
The ISME Journal
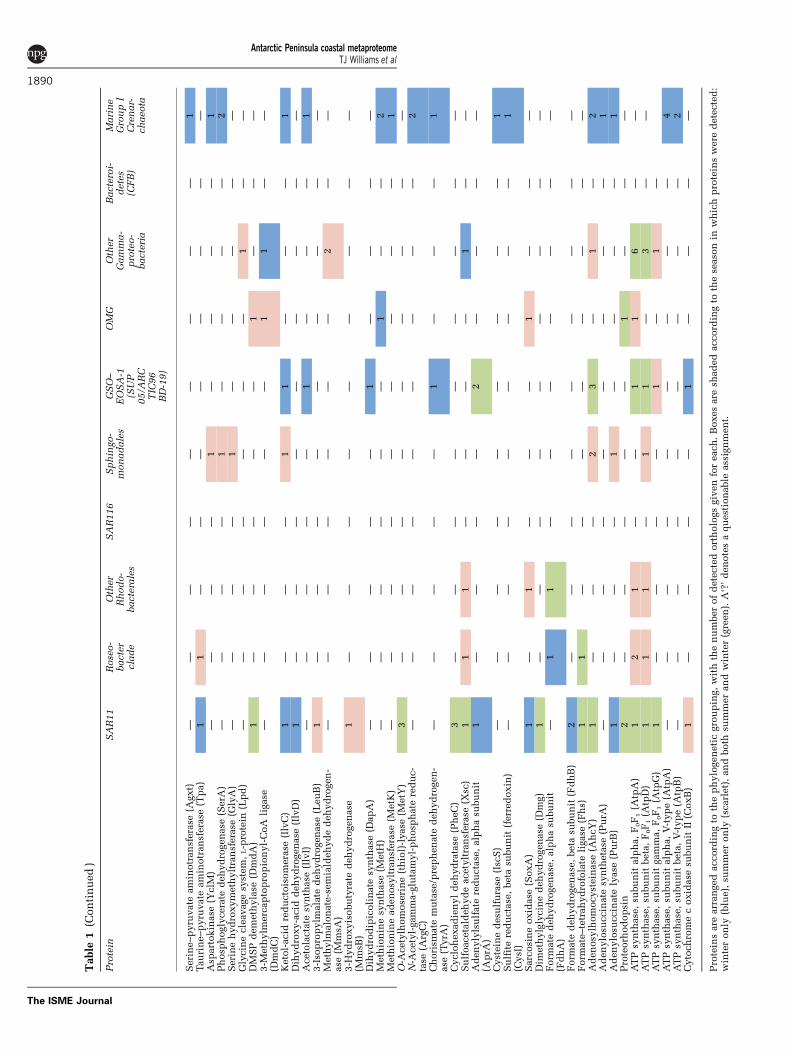
Table
1(C
on
tin
ued
)
Pro
tein
SA
R11
Rose
o-
bacte
rcla
de
Oth
er
Rh
od
o-
bacte
rale
s
SA
R116
Sp
hin
go-
mon
ad
ale
sG
SO
–E
OS
A-1
(SU
P05/A
RC
TIC
96
BD
-19)
OM
GO
ther
Gam
ma-
pro
teo-
bacte
ria
Bacte
roi-
dete
s(C
FB
)
Mari
ne
Gro
up
IC
ren
ar-
ch
aeota
Seri
ne–p
yru
vate
am
inotr
an
sfera
se(A
gxt)
——
——
——
——
—1
Tau
rin
e–p
yru
vate
am
inotr
an
sfera
se(T
pa)
11
——
——
——
——
Asp
art
okin
ase
(YclM
)—
——
—1
——
——
1P
hosp
hogly
cera
ted
eh
yd
rogen
ase
(SerA
)—
——
—1
——
——
2S
eri
ne
hyd
roxym
eth
ylt
ran
sfera
se(G
lyA
)—
——
—1
——
——
—G
lycin
ecle
avage
syst
em
,L-p
rote
in(L
pd
)—
——
——
——
1—
—D
MS
Pd
em
eth
yla
se(D
md
A)
1—
——
——
1—
——
3-M
eth
ylm
erc
ap
top
rop
ion
yl-
CoA
ligase
(Dm
dC
)—
——
——
—1
1—
—
Keto
l-acid
red
ucto
isom
era
se(I
lvC
)1
——
—1
1—
——
1D
ihyd
roxy-a
cid
deh
yd
rogen
ase
(Ilv
D)
1—
——
——
——
——
Aceto
lacta
tesy
nth
ase
(Ilv
I)—
——
——
1—
——
13-I
sop
rop
ylm
ala
ted
eh
yd
rogen
ase
(Leu
B)
1—
——
——
——
——
Meth
ylm
alo
nate
-sem
iald
eh
yd
ed
eh
yd
rogen
-ase
(Mm
sA)
——
——
——
—2
——
3-H
yd
roxyis
obu
tyra
ted
eh
yd
rogen
ase
(Mm
sB)
1—
——
——
——
——
Dih
yd
rod
ipic
oli
nate
syn
thase
(Dap
A)
——
——
—1
——
——
Meth
ion
ine
syn
thase
(MetH
)—
——
——
—1
——
2M
eth
ion
ine
ad
en
osy
ltra
nsf
era
se(M
etK
)—
——
——
——
——
1O
-Acety
lhom
ose
rin
e(t
hio
l)-l
yase
(MetY
)3
——
——
——
——
—N
-Acety
l-gam
ma-g
luta
myl-
ph
osp
hate
red
uc-
tase
(Arg
C)
——
——
——
——
—2
Ch
ori
smate
mu
tase
/pre
ph
en
ate
deh
yd
rogen
-ase
(TyrA
)—
——
——
1—
——
1
Cyclo
hexad
ien
yl
deh
yd
rata
se(P
heC
)3
——
——
——
——
—S
ulf
oaceta
ldeh
yd
eacety
ltra
nsf
era
se(X
sc)
11
1—
——
—1
——
Ad
en
yly
lsu
lfate
red
ucta
se,
alp
ha
subu
nit
(Ap
rA)
1—
——
—2
——
——
Cyst
ein
ed
esu
lfu
rase
(Isc
S)
——
——
——
——
—1
Su
lfit
ere
du
cta
se,
beta
subu
nit
(ferr
ed
oxin
)(C
ysI
)—
——
——
——
——
1
Sarc
osi
ne
oxid
ase
(SoxA
)1
—1
——
—1
——
—D
imeth
ylg
lycin
ed
eh
yd
rogen
ase
(Dm
g)
1—
——
——
——
——
Form
ate
deh
yd
rogen
ase
,alp
ha
subu
nit
(Fd
hA
)—
11
——
——
——
—
Form
ate
deh
yd
rogen
ase
,beta
subu
nit
(Fd
hB
)2
——
——
——
——
—F
orm
ate
–te
trah
yd
rofo
late
ligase
(Fh
s)1
1—
——
——
——
—A
den
osy
lhom
ocyst
ein
ase
(Ah
cY
)1
——
—2
3—
1—
2A
den
ylo
succin
ate
syn
theta
se(P
urA
)—
——
——
——
——
1A
den
ylo
succin
ate
lyase
(Pu
rB)
1—
——
1—
——
—1
Pro
teorh
od
op
sin
2—
——
——
1—
——
AT
Psy
nth
ase
,su
bu
nit
alp
ha,
F0F
1(A
tpA
)1
21
——
11
6—
—A
TP
syn
thase
,su
bu
nit
beta
,F
0F
1(A
tpD
)1
11
—1
1—
3—
—A
TP
syn
thase
,su
bu
nit
gam
ma,
F0F
1(A
tpG
)1
——
——
1—
1—
AT
Psy
nth
ase
,su
bu
nit
alp
ha,
V-t
yp
e(A
tpA
)—
——
——
——
——
4A
TP
syn
thase
,su
bu
nit
beta
,V
-typ
e(A
tpB
)—
——
——
——
——
2C
yto
ch
rom
ec
oxid
ase
subu
nit
II(C
oxB
)1
——
——
1—
——
—
Pro
tein
sare
arr
an
ged
accord
ing
toth
ep
hylo
gen
eti
cgro
up
ing,
wit
hth
en
um
ber
of
dete
cte
dort
holo
gs
giv
en
for
each
.B
oxes
are
shad
ed
accord
ing
toth
ese
aso
nin
wh
ich
pro
tein
sw
ere
dete
cte
d:
win
ter
on
ly(b
lue),
sum
mer
on
ly(s
carl
et)
,an
dboth
sum
mer
an
dw
inte
r(g
reen
).A
‘?’
den
ote
sa
qu
est
ion
able
ass
ign
men
t.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1890
The ISME Journal

Roseobacter clade (Supplementary Table S1) areconsistent with genomic studies implicating marineRoseobacter spp. as using these organic solutes assources of carbon and nitrogen (Moran et al., 2007).
The Roseobacter clade is a dominant constituent ofbacterioplankton, especially in coastal waters,demonstrating metabolic versatility and a capacityto associate with phytoplankton blooms as a meansof deriving metabolites from exudates (Buchan et al.,2005; Moran et al., 2007). Nevertheless, unlikeheterotrophic Bacteroidetes and Gammaproteobac-teria groups, based on spectral counts, Rhodobacter-ales was not significantly more abundant in summerthan in winter. ABC transporter proteins were alsoidentified in the winter metaproteome for phospho-nate transport, matching to members of the Rhodo-bacterales and Gammaproteobacteria (Table 1 andSupplementary Table S1).
The ABC transporter PBPs that best matched toSAR11 included those specific for polyamines(spermidine/putrescine), general amino acids,branched-chain amino acids, glycine betaine, taur-ine/sulfonates, iron and possibly sugars (Table 1 andSupplementary Table S1). The expression of puta-tive sugar transporters accords with the ability ofcoastal SAR11 isolates to utilize energy-rich carbo-hydrates (Schwalbach et al., 2010). In other meta-proteome studies of SAR11, summer populations offthe Oregon coast were similarly dominated bytransport proteins for carbon- and nitrogen-contain-ing compounds, rather than phosphate (Sowellet al., 2011). In contrast, a study of the ocean gyreof the Sargasso Sea identified abundant SAR11phosphate and phosphonate ABC transporters(Sowell et al., 2009).
Unlike ABC transporters, TRAP and TTT trans-porters utilize a proton motive force rather than ATPhydrolysis to drive solute uptake (Forward et al.,1997; Winnen et al., 2003). The expression of theseindicates that carboxylic acids are likely to betransported and metabolized for growth, predomi-nantly by the Roseobacter and SAR11 clades (Moranand Zepp, 1997; Moran et al., 2007).
Many of the SAR11 proteins in the metaproteomethat are implicated in carbon and nitrogen substrateuptake are the same as those detected for SAR11 inthe Sargasso Sea: ABC transporters for polyamines,amino acids, and taurine and TRAP dicarboxylatetransporters (Sowell et al., 2009). However, theSargasso Sea SAR11 metaproteome was notable forthe absence of iron transport proteins, and this wasattributed to high levels of iron in Sargasso Seasummer surface waters due to eolian input from theSaharan desert (Sowell et al., 2009). The expressionof sfuC (iron-binding PBP ABC transport protein),cspL (RNA-binding protein implicated in control-ling SfuC translation in response to iron scarcity)and groES was elevated during iron-limited growthof SAR11, and SfuC has been suggested to be auseful ecological marker for iron limitation (Smithet al., 2010). In the current metaproteome, all threeproteins (SfuC, CspL and GroES) from SAR11 weredetected in the summer (Supplementary Table S1),when iron limitation may be more acute due to moreintense competition for iron with phytoplankton
Table 2 Proteins inferred to belong to carbon fixation andnitrification pathways identified in the winter metaproteome
Carbon fixation and nitrification
Archaea N. maritimushomolog
Ammonia oxidationAmmonium transporter Nmar_1698 (100%)Ammonia monooxygenase, subunit B Nmar_1503 (100%)Ammonia monooxygenase, subunit B Nmar_1503 (84%)
3-Hydroxypropionate/4-hydroxybutyrate cycleAcetyl-CoA/propionyl-CoA carboxylase(ATP-binding subunit)
Nmar_0273 (86%)
Acetyl-CoA/propionyl-CoA carboxylase(ATP-binding subunit)
Nmar_0273 (74%)
Acetyl-CoA/propionyl-CoA carboxylase(carboxyltransferase subunit)
Nmar_0272 (84%)
Malonic semialdehyde reductase Nmar_1043 (77%)Malonic semialdehyde reductase Nmar_1043 (77%)3-Hydroxypropionyl-CoA synthetize Nmar_1309 (84%)Acryloyl-CoA reductase Nmar_1622 (81%)Acryloyl-CoA reductase Nmar_1622 (80%)Methylmalonyl-CoAmutase, large subunit
Nmar_0954 (87%)
Succinic semialdehyde reductase Nmar_1110 (90%)4-Hydroxybutyryl-CoA synthetase(AMP-forming)
Nmar_0206 (90%)
4-Hydroxybutyryl-CoA dehydratase Nmar_0207 (97%)4-Hydroxybutyryl-CoA dehydratase Nmar_0207 (91%)Crotonyl-CoA hydratase Nmar_1308 (100%)3-Hydroxybutyryl-CoA dehydrogenase(NADþ )
Nmar_1028 (87%)
Acetoacetyl-CoA beta-ketothiolase Nmar_1631 (88%)
Energy conservation (electron transport chain)Multicopper oxidase (NirK-like) Nmar_1667 (87%)Blue (Type 1) copper domain protein Nmar_0185 (72%)Plastocyanin-family protein Nmar_0343 (63%)
Bacteria Closest homolog
Ammonia oxidationAmmonia monooxygenase,subunit B
AmoB, Nitrosospira sp. NpAV(Betaproteobacteria) (100%)
Reverse tricarboxylic acid cyclePyruvate:ferredoxin oxi-doreductase, gammasubunit
PorG, Candidatus Leptospirillumrubarum (Nitrospirae) (100%)
Nitrite oxidationNitrite oxidoreductase,alpha subunit
NxrA, Candidatus Nitrospiradefluvii (Nitrospirae) (69%)
Nitrite oxidoreductase,alpha subunit
NarG, Candidatus Kuenenia stutt-gartiensis (Planctomycetes) (78%)
The closest match of each protein is shown for the ammonia-oxidizingarchaeon N. maritimus, ammonia-oxidizing Betaproteobacteria, andnitrite-oxidizing Nitrospirae and Planctomycetes. For some proteins,multiple orthologs were identified in the metaproteome. Percentagesin parentheses represent percentage amino-acid identity.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1891
The ISME Journal
and other organisms; however, spectral countsrevealed no significant difference between seasons.
TBDRs are involved in proton motive force-dependent outer membrane transport, and in manyGram-negative bacteria these transporters areinvolved in the binding and uptake of iron–siderophore complexes, cobalamin (Koebnik, 2005)or carbohydrates (Blanvillain et al., 2007). TBDRswere matched to Bacteroidetes (40 total, 37 summeronly, 2 winter only and 1 common to both seasons);Gammaproteobacteria (27 total, 18 summer, 6 winterand 3 both seasons) mostly to Alteromonadales andOMG; and Alphaproteobacteria (6 total, 5 summerand 1 winter) with most matches to S. alaskensis(Table 1 and Supplementary Table S1). Spectrareferable to TBDRs were significantly more abun-dant in summer than in winter. The identification ofa large number of Bacteroidetes TBDRs is consistentwith most (78%) Bacteroidetes protein identifica-tions being predicted OMPs. Many of the TBDRsthat matched to Bacteroidetes appeared to beinvolved in the acquisition and uptake of polymericsubstrates mediated by cellular attachment (seesection Attachment below). For marine Gammapro-teobacteria (for example, OMG) and Alphaproteo-bacteria (for example, S. alaskensis), some TBDRsmay be deployed for scavenging iron, or carbohy-drates that are free in seawater or associated withparticulate organic matter (Blanvillain et al., 2007;Ting et al., 2010). Although DOC levels werecomparable in the winter and summer samples,bacterial production rates were around six timeshigher in summer compared with winter, as indi-cated by leucine incorporation rates (Materials andmethods). Chlorophyll a levels in the summer weremuch higher than in winter, and we hypothesizethat TBDR-mediated uptake is adapted for theacquisition of biopolymers and other nutrients
derived from phytoplankton. Overall, the dataindicate a greater reliance on TBDR-mediateduptake by Bacteroidetes and certain Gammaproteo-bacteria and Alphaproteobacteria, especially in thesummer, whereas SAR11 and Rhodobacterales relymore on labile solute capture all-year-round by PBP-dependent high-affinity uptake systems.
Flagellin proteins were detected for P. haloplanktis,Moritella sp. PE36 and S. alaskensis (Table 1 andSupplementary Table S1) suggesting mobility isadvantageous for these bacteria in the SouthernOcean. P. haloplanktis is a copiotroph that specializesin the exploitation of short-lived nutrient patches(Stocker et al., 2008), and the oligotrophic lifestyle ofS. alaskensis has nevertheless been linked to thepursuit and utilization of nutrient pulses (Lauro et al.,2009; Williams et al., 2009).
AttachmentThe proteins matching Bacteroidetes were mostlycell surface proteins associated with particle adhe-sion and nutrient acquisition (Supplementary TableS1). In contrast, only a few transcription andtranslation proteins and only one metabolic enzyme(fumarase) were detected (Supplementary Table S1).Frequently detected OMPs included a glidingmotility protein (GldL) involved in the explorationof solid surfaces, and components of the TBDR andSusCD/RagAB complexes (Table 1 and Supple-mentary Table S1). SusCD is associated with thebinding of exogenous polysaccharides, which arethen degraded and imported into the periplasm(Reeves et al., 1997; Gilbert, 2008). RagAB isinvolved in binding exogenous proteins, and sub-sequent uptake of their degradation products aspeptides (Nagano et al., 2007). When sampled, theplanktonic Flavobacteria that were captured on
Figure 3 Functional assignment of bacterial and archaeal proteins detected in the (a) winter and (b) summer metaproteome. CM, cellmembrane; OMP, outer membrane protein; TAXI, TRAP-associated extracytoplasmic immunity; TRAP, tripartite ATP-independentperiplasmic transporter; TTT, tripartite tricarboxylate transporter.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1892
The ISME Journal

filters may have had minimal metabolic activity, butwere poised to detect and exploit particulate matter.The metaproteomic data corroborate the view thatmarine Flavobacteria are specialized for attachmentand growth on detrital particles (especially asso-ciated with the decay phase of phytoplanktonblooms), and rely mostly on biopolymers for carbonand energy, in preference to the labile solutesfavored by heterotrophic Proteobacteria (Cottrelland Kirchman, 2000; Abell and Bowman, 2005;Gonzalez et al., 2008).
Ammonia oxidation and carbon fixationTwo chemolithoautotrophic pathways are indicatedin the Antarctic winter metaproteome: the 3-hydro-xypropionate/4-hydroxybutyrate (3HP/4HB) cycleand the reverse tricarboxylic acid cycle (Table 2).The former is represented by proteins assigned tothe 3HP/4HB cycle, a uniquely archaeal pathwayfor carbon fixation present in AOA such asN. maritimus. The full pathway has yet to becharacterized, but homologs of proteins referred tothis pathway in MGI and other archaeal speciesthat use the 3HP/4HB cycle (Berg et al., 2007;Kockelkorn and Fuchs, 2009; Walker et al., 2010;Ramos-Vera et al., 2011) were found exclusivelyin the winter metaproteome (Table 2 andSupplementary Table S2). These data are consistentwith the high abundance of MGI in coastal surfaceAntarctic waters, especially in winter (Murray et al.,1998; Church et al., 2003; Murray and Grzymski,2007; Grzymski et al., 2012).
For AOA, ammonia oxidation is intimately linkedto the 3HP/4HB cycle because it provides the energynecessary for carbon fixation. We identified proteinsfrom Crenarchaeota for ammonia transport andammonia oxidation; a previous metaproteomicstudy detected ammonia monooxygenase subunitsfrom N. maritimus at coastal upwelling sites in theSouth Atlantic, providing evidence for archaealnitrification at the surface in summer (Morriset al., 2010). Our study also detected componentsof the proposed respiratory pathway for AOA,including NADH dehydrogenase, multicopperoxidase, and other copper-containing proteins andferredoxin involved in electron transfer reactionsrequired for energy metabolism (Table 2 andSupplementary Table S2) (Konneke et al., 2005;Walker et al., 2010). Our data support the hypothesisthat MGI are important in nitrification and carbonfixation in Antarctic coastal surface waters duringthe winter months.
The presence of an ammonia monooxygenasesubunit from a nitrifying member of the Betaproteo-bacteria in the winter metaproteome indicates thatammonia-oxidizing bacteria (AOB) are also presentin winter surface waters. Notably, two ammonia-oxidizing Betaproteobacteria-associated RuBisCOgenes were detected in the winter metagenome(Grzymski et al., 2012). Ammonia oxidation by
AOA and AOB leads to nitrite, which is subse-quently oxidized to nitrate to complete the processof nitrification. Two nitrite oxidoreductase alphasubunits were detected in the winter metaproteome,which showed the highest identities to proteinsfrom Nitrospirae (Lucker et al., 2010) and anammoxPlanctomycetes (Strous et al., 2006) (Table 2 andSupplementary Table S1). The presence of pyruva-te:ferredoxin oxidoreductase, also from the Nitros-pirae clade (Table 2 and Supplementary Table S1),provides the sole evidence for operation of thereverse tricarboxylic acid cycle for carbon fixation inour metaproteome data. This chemoautotrophicpathway has been documented for the nitrite-oxidizing bacteria Ca. Nitrospira defluvii andCa. Leptospirillum rubarum (both Nitrospirae) withthe potential for mixotrophic growth inferred for thelatter (Goltsman et al., 2009; Lucker et al., 2010).
Other detected MGI proteins include thosebelonging to an oxidative Krebs cycle, and gluco-neogenesis (including fructose-1,6-bisphosphatase),with phosphoenolpyruvate carboxykinase used togenerate phosphoenolpyruvate from oxaloacetate(Table 1 and Supplementary Table S1) (Dansonet al., 2007). MGI biosynthetic pathways alsoidentified in the metaproteome include enzymesof individual amino-acid biosynthesis pathways,and the biosynthesis of nucleotides and variouscofactors/coenzymes, including cobalamin (Supple-mentary Table S2).
A potential for mixotrophic growth has beensuggested for N. maritimus, based on the genomicpresence of genes that encode transporters fororganic compounds (amino acids, oligopeptidesand taurine; Walker et al., 2010). Mixotrophicgrowth has also been inferred for the soil AOANitrosphaera viennensis, based on stronger growthwhen provided with pyruvate, albeit carbon assim-ilation is driven predominantly by CO2 (as bicarbo-nate) fixation (Tourna et al., 2011). No unambiguousABC transporters from Crenarchaeota were detectedin the winter metaproteome (SupplementaryTable S2). It is possible that the proteases andamino-acid degradation enzymes that were detectedwere associated with protein turnover by theproteasome. An archaeal cysteine desulfurasewas detected in the metaproteome, indicatingthat cysteine can serve as a source of sulfur forbiosynthetic purposes, such as iron–sulfur clusterproteins, which were especially abundant in themetaproteome and attributable to MGI (Table 1 andSupplementary Table S1).
Ammonia assimilationThe identification of proteins associated with theuptake and incorporation of ammonia (ammoniumtransporter, glutamine synthetase (GS), glutamatesynthase (GOGAT) and glutamate dehydrogenase(GDH)) indicate that the Antarctic coastal bacteriaand archaea that were sampled were metabolically
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1893
The ISME Journal

poised to assimilate inorganic reduced nitrogen(Table 1 and Supplementary Table S1). However,the metaproteome data also suggest that thesebacteria and archaea adopt divergent pathways forammonia assimilation, consistent with proposedniche partitioning in marine habitats (Martens-Habbena et al., 2009). The bacteria (includingSAR11 and GSO–EOSA-1) appear to employ theGS–GOGAT cycle, whereas AOA utilize GDH(Table 1 and Supplementary Table S1). An extre-mely high specific affinity for dissolved ammoniahas been demonstrated for N. maritimus strainSCM1, providing this AOA with a competitiveadvantage under oligotrophic conditions (Martens-Habbena et al., 2009). One suggestion is that theextremely high capacity for ammonia uptake byAOA is due to overexpression of ammoniumtransporters (Stewart et al., 2012). If so, then theresulting intracellular accumulation could allow forammonia assimilation via the lower-affinity enzymeGDH rather than the higher-affinity GS.
Taurine degradationTaurine (2-aminoethanesulfonic acid) is an organo-sulfonate found in the tissues of marine inverte-brates and certain algae, and is a potential source ofcarbon, nitrogen and sulfur. Further, growth ofSAR11 cultures on taurine has previously beenexperimentally demonstrated (Schwalbach et al.,2010). Proteins that could be assigned to a functionin the taurine degradation pathway were identifiedin the metaproteome, including taurine–pyruvateaminotransferase and sulfoacetaldehyde acetyltrans-ferase (Table 1 and Supplementary Table S1;Figure 4). Complete taurine degradation yieldsacetyl-CoA for carbon assimilation, and ammoniathat can be assimilated as a nitrogen source via theGS–GOGAT cycle. The use of acetate as a carbonsource for cellular biomass production is consistentwith glyoxylate cycle enzymes also being detected(isocitrate lyase and malate synthase) for a range ofheterotrophs, including SAR11 (Table 1 andSupplementary Table S1). Acetate is also a productof dimethylsulfoniopropionate (DMSP) degradation(see section Methionine and DMSP degradationbelow). The detection of taurine degradationenzymes from a range of bacteria, combined withthe presence of ABC-type taurine/sulfonate trans-porters, suggests that this organosulfonate is afavored substrate in coastal Antarctic Peninsulawaters. SAR11 taurine transporters were alsodetected in the Sargasso Sea (Sowell et al., 2009).
Taurine desulfonation by sulfoacetaldehyde acet-yltransferase releases sulfite anion (Figure 4), whichis highly reactive and can cause damage to DNA andproteins (Kappler, 2011). SAR11 is unable toreductively assimilate oxidized sulfur, possiblybecause of the bioenergetic costs required for theimport and subsequent reduction of sulfate (Trippet al., 2008). The ability to oxidize sulfite to sulfate
is suggested by the presence of adenylylsulfate(APS) reductase (AprAB) and a putative membraneanchor protein (AprM) in SAR11 genomes; theSAR11 APS reductase alpha subunit (AprA) wasdetected in the metaproteome (Table 1 and Supple-mentary Table S1). This suggests that sulfite isoxidatively bound to AMP to generate the activatedintermediate APS, as a mechanism for the intracellulardetoxification of sulfite (Meyer and Kuever, 2007).
There is no evidence in SAR11 genomes of ATPsulfurylase or APS adenylyltransferase (APAT) forsubstrate-level phosphorylation to ATP or ADP.Nevertheless, immediately adjacent to the aprABMgene cluster is a gene that encodes a HIT familyhydrolase, homologous to adenylylsulfatase. Weposit that SAR11 is capable of enzymatic hydrolysisof APS to AMP and sulfate, or, owing to the inherentinstability of the phosphosulfur anhydride bond,hydrolysis of APS is spontaneous (Figure 4). Accor-ding to our hypothesis, AMP would be regeneratedand sulfate subsequently released from the cell.Efficient detoxification of sulfite requires availableintracellular AMP. Therefore, as an oligotroph thismechanism could be advantageous to SAR11 asAMP would be recycled for subsequent sulfitedetoxification. If taurine is metabolized in this wayby SAR11 and other heterotrophic bacteria, thebreakdown of taurine to produce inorganic sulfurcompounds could represent an important route bywhich these sulfur species (sulfate, sulfite andthiosulfate) are produced at the ocean surface.
Glycine betaine degradation and one-carbon (C1)metabolismGlycine betaine is a compatible solute and apotentially important cryoprotectant in the Antarcticenvironment. The large number of glycine betainetransport proteins identified from both seasons(Table 1 and Supplementary Table S1) suggests thatbacteria are importing this osmolyte. Glycine betainecan also be catabolized to glycine; this may be ofparticular importance for SAR11, which has beenfound to be functionally auxotrophic for glycine(Tripp et al., 2009). Two of the three proteinsinvolved in the degradation pathway of glycinebetaine to glycine (Sun et al., 2011) were detectedin the metaproteome (dimethylglycine dehydrogen-ase and sarcosine oxidase), with best matches toSAR11 (Table 1 and Supplementary Table S1;Figure 5). Both of these oxidation steps generateformaldehyde as a byproduct, which would sponta-neously degrade to formate. Formate could then beoxidized to CO2 by formate dehydrogenase and serveas a source of reductant, with the CO2 released fromthe cell (Sowell et al., 2008). Alternatively, theformate could be activated by formyl-tetrahydro-folate ligase and directed to C1 metabolism forbiosynthesis (Figure 5). Proteins pertaining to bothpossible fates were detected in the metaproteome(Supplementary Table S1; Figure 5).
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1894
The ISME Journal

Other proteins related to C1 metabolism includetwo methanol dehydrogenase large subunit proteinsfrom summer and winter that belong to the OM43clade of Betaproteobacteria, with best matches to theobligately methylotrophic strain HTCC2181(Giovannoni et al., 2008). Consistent with the viewthat methylotrophic members of the OM43 clade areimportant components of coastal marine ecosystems(Giovannoni et al., 2008; Sowell et al., 2011), ourdata imply that C1 compounds such as methanoland formaldehyde are able to be metabolized.Possible sources of methanol include depositionfrom the atmosphere, a byproduct of marine phyto-plankton growth (Heikes et al., 2002) or photoche-mical degradation of surface DOC (Dixon et al.,2011).
Methionine and DMSP degradationThe ABC transporter for glycine betaine (OpuA) hasalso been shown to import DMSP, an algal osmolyte
and reduced sulfur source (Holtmann and Bremer,2004). The general amino-acid transporter (Yhd) hasbeen proposed to import methionine in SAR11(Sowell et al., 2009). In the absence of genes forthe reductive assimilation of sulfur, SAR11 requiresexogenous sources of reduced sulfur, such asmethionine or DMSP (Tripp et al., 2008). WhileSAR11 proteins corresponding to the postulatedmethionine degradation pathway (Tripp et al., 2008)were not detected, two proteins in the pathway(adenosylhomocysteinase and O-acetyl-homoserine[thiol]-lyase) that matched other bacterioplanktonwere detected (Table 1 and Supplementary TableS1). The former matched to a range of bacteria, andMGI, suggesting that the biosynthesis of cysteinefrom methionine is a relatively common pathway inthe Antarctic community that was sampled.
A novel pathway for the assimilation of DMSPwas recently demonstrated for the Roseobacter cladebacterium Ruegeria pomeroyi (Reisch et al., 2011).Following initial demethylation of DMSP, the
Figure 4 Putative pathway for the degradation of taurine in SAR11 and associated detoxification of sulfite. Genes for all proteins werefound in SAR11 genomes; bolded and underlined proteins were detected in the metaproteome; and spectral counts per protein are shownfor winter (first number after the protein) and summer (second number after the protein). Ald, alanine dehydrogenase; AprAB,adenylylsulfate (APS) reductase; GlnT, glutamine synthetase; GltAB, glutamate synthase; Pta, phosphotransacetylase; TauABC, taurine/sulfonate ABC transporter; Tpa, taurine-pyruvate aminotransferase; Xsc, sulfoacetaldehyde acetyltransferase.
Figure 5 Putative pathway for the degradation of glycine betaine in SAR11. Genes for all proteins were found in SAR11 genomes;bolded and underlined proteins were detected in the metaproteome; and spectral counts per protein are shown for winter (first numberafter the protein) and summer (second number after the protein). BhmT, betaine-homocysteine methyltransferase; Dmg, dimethylglycinedehydrogenase; FhdAB, formate dehydrogenase; Fhs, formate-tetrahydrofolate ligase; OpuACAB, glycine betaine ABC transporter;ProXYZ, proline/glycine betaine ABC transporter; SoxA, sarcosine oxidase (monomeric); THF, tetrahydrofolate.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1895
The ISME Journal

intermediate methylmercaptopropionate (MMPA) isfurther catabolized to methanethiol, which can bereadily assimilated as a reduced sulfur source(Reisch et al., 2011). SAR11 was also shown topossess enzyme activities for the first two steps ofMMPA catabolization (Reisch et al., 2011). The firststep is mediated by DMSP demethylase (DmdA), ahomolog of the glycine cleavage T-protein (GcvT).GcvT homologs were detected in the metaproteome,and we infer two (one from SAR11 and one fromOMG) to be DmdA (Table 1 and SupplementaryTable S1). SAR11 proteins for the subsequent MMPAcatabolization steps were not detected in themetaproteome, although two proteins that corre-spond to the enzyme that catalyzes the second stepof MMPA catabolization, methylmercaptopropionyl-CoA ligase (DmdC), were detected for Gammapro-teobacteria (including OMG) (Table 1 andSupplementary Table S1). A SAR11 acetaldehydedehydrogenase was detected for the oxidation ofacetaldehyde (another product of MMPA cataboliza-tion) to acetate.
GSO–EOSA-1 bacteria (ARCTIC96BD-19/SUP05)Many proteins in the summer and winter metapro-teome had the highest identity to proteins frommembers of the SUP05 cluster of Gammaproteo-bacteria, including the clam symbiont Ca. Ruthiamagnifica (Newton et al., 2007), and the free-livingSUP05 Walsh et al., 2009). The SUP05 and ARC-TIC96BD-19 clusters together make up the GSO–EOSA-1 complex, and pelagic members of thisgroup are inferred to be major contributors to globalchemolithoautotrophic carbon fixation via sulfuroxidation (Walsh et al., 2009; Swan et al., 2011). TheSUP05 cluster has been reported in high abundancein some oxygen minimum zones (Walsh et al., 2009),and the closely related ARCTIC98BD-19 cluster havebeen recovered from diverse oceanic habitats andwere recently reported to be widespread in themesopelagic Atlantic and Pacific oceans (Swanet al., 2011). Overall, our metaproteomic datasuggest that GSO–EOSA-1 bacteria are metabolicallyactive in the aerobic surface waters of the Antarcticcoast, with a higher representation in winter. Aswell as components of the transcription and transla-tion machinery, these included proteins pertainingto ammonia assimilation, energy generation (sub-units of cytochrome c oxidase (Cox) and ATPsynthase) and biosynthesis (for example, aminoacids and nucleotides) (Table 1 and SupplementaryTable S1).
SUP05 has the potential for chemolithoautotrophyusing reduced sulfur compounds coupled to oxygenor nitrate reduction and carbon fixation using formII RuBisCO (Walsh et al., 2009), while the genomesequence of a member of the ARCTIC96BD-19cluster indicates the capacity for carbon fixationusing form I RuBisCO and sulfur oxidation (AprA),and the potential for mixotropic growth (Swan et al.,
2011). Sequences from the winter metagenomiclibrary matched genes in Ca. R. magnifica requiredfor APS reductase, sulfur oxidation (Sox) andreverse dissimilatory sulfate reduction pathway(Grzymski et al., 2012), with the latter possibly usedfor the oxidation of stored sulfur (S0). The wintermetagenome also contained closely relatedsequences to ARCTIC96BD-19 cluster related formI RuBisCO (Swan et al., 2011). The detection ofCoxII in the winter metaproteome indicates anaerobic metabolism by GSO–EOSA-1 bacteria,whereas the detection of AprA (both seasons;Supplementary Table S1) could indicate sulfuroxidation at the surface. Studies examining themetabolic activity of GSO–EOSA-1 bacteria inaerobic waters are needed in order to determinethe ecological role that these bacterioplankton havein Southern Ocean waters.
PRPRs are photoactive cell membrane proteins inbacteria (Beja et al., 2000, 2001; DeLong and Beja,2010). Three PRs were detected in our metapro-teome (both seasons), two of which had bestmatches to SAR11 and the third to the OMG group(Table 1 and Supplementary Table S1). Two of thePRs are inferred to be green-light-absorbing (oneSAR11 and one OMG) and the other blue-light-absorbing (SAR11) (Man et al., 2003) (Table 1 andSupplementary Table S1). Laboratory studies ofCa. P. ubique strain HTCC1062 (which has the‘green’ PR) indicate that PR serves as a light-dependent proton pump, and is important underilluminated, carbon-limiting conditions when PR-mediated ATP generation can substitute for respira-tion (Giovannoni et al., 2005; Steindler et al., 2011).Although ‘green’ PR is optimal for capturing light atthe surface and ‘blue’ for light deep in the photiczone, both variants are found in shallow water(Fuhrman et al., 2008). Constitutive expression ofPR for light harvesting in Ca. P. ubique may facilitatethe ability to immediately respond to cellular energydeficits caused by carbon starvation (Steindler et al.,2011).
Chaperones and quality controlProtein chaperones were highly abundant in themetaproteome, including DnaK, HtpG, peptidyl–prolyl cis/trans isomerases, trigger factor, andbacterial GroEL/GroES and the equivalent archaealchaperonin Cpn60. Proteins of the FtsH–HflKCmembrane complex involved in proteolytic turnoverof membrane and soluble proteins were alsodetected. Chaperones and proteases were abundantin both seasons, and the high abundance of theseproteins has been linked to microbial survival insurface ocean water as a response to exposure toenvironmental stresses (Sowell et al., 2009, 2011), aswell as oxidation of endogenous biomass via
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1896
The ISME Journal

proteolysis under carbon-limiting conditions(Steindler et al., 2011). Additionally, proteasome,exosome and degradosome (particularly polyribo-nucleotide nucleotidyltransferase) componentswere detected. Consistent with the detection ofMGI proteins, the archaeal proteasome and exosomeproteins were found exclusively in winter. Proteo-mic studies of the Antarctic archaeon M. burtoniihave linked the abundance of proteasome andexosome proteins to cold adaptation (Williamset al., 2010). One possibility is that the inhibitoryeffects of low temperature on translation, includingcorrect folding of the nascent polypeptide, leadsto an increase in the number of irretrievablymisfolded proteins. Cold temperatures also leadto an increase in adventitious secondary structuresformed by RNA transcripts, which can be resolvedby DEAD box RNA helicases, the latter were foundin the metaproteome (two in winter and onein summer) (Supplementary Table S1). Increasedexosome and degradosome levels are consistentwith the need to degrade irreparably damaged ortangled RNA.
Conclusions
Our metaproteomic analysis provided insight intofunctionally important transport, metabolism andenergy generation processes in surface water com-munities in the coastal WAP. Bacterial ABC, TRAPand TBDR transporters have a critical rolein nutrient scavenging. We have obtained evidencethat glycine betaine is being metabolized to glycinein Antarctic coastal waters to redress conditionalglycine auxotrophy in SAR11 (Tripp et al., 2009).We also posit that the purpose of APS reductase inlieu of sulfate reduction by SAR11 (Tripp et al.,2008) is to detoxify sulfite that is generatedendogenously (for example, by taurine desulfona-tion) without providing any benefit in energyconservation. Thus, the prevalence of aprA infunctional gene studies of environmental samplesmay be partly explained by the utility of a reverse-acting APS reductase in organic sulfur compounddegradation, including by SAR11 (Meyer andKuever, 2007).
MGI have been proposed to have an importantrole in oceanic nitrification, based on the detectionof these organisms in high abundance in coastalAntarctic surface waters (DeLong et al., 1994;Murray and Grzymski, 2007) and studies from theNorth Sea, which demonstrated that MGI abundancecorrelates with ammonia oxidation to nitrite(Wuchter et al., 2006). Components of the 3HP/4HB cycle and the oxidation of ammonia to nitritewere detected in the winter metaproteome indicat-ing that this pathway is active and the MGIare causing nitrification in coastal WAP waters.Ammonia-oxidizing and nitrite-oxidizing bacteriawere also represented in the data, indicating that
nitrification in these waters involved multiplespecies. As this nitrification process fuels thefixation of inorganic carbon, the metaproteomecorroborates metagenomic and SSU ribosomalRNA interpretations that the chemolithoautotrophicarchaea and bacteria constitute a major CO2 sinkduring the cold, dark winter months (Grzymskiet al., 2012). Overall, these data emphasize therole that specific types of Southern Ocean micro-organisms have, year round, in primary productionand remineralization in a coastal region of WestAntarctica.
Acknowledgements
We extend our thanks to our field team M Erickson,K Myers, V Peng and J-F Ghiglione for their expertassistance sampling in Antarctica and to the PalmerStation personnel in the US Antarctic Program forsupport, especially during the winter field season. Thework was supported by the NSF grants, ANT 0632389 toAEM and JJG, and ANT 0632278 and ANT 0217282 to HD.The work of the Australian contingent was supported bythe Australian Research Council. Mass spectrometricresults were obtained at the Bioanalytical Mass Spectro-metry Facility within the Analytical Centre of theUniversity of New South Wales. This work was under-taken using infrastructure provided by NSW Governmentco-investment in the National Collaborative ResearchInfrastructure Scheme. Subsidized access to this facilityis gratefully acknowledged.
References
Abell GCJ, Bowman JP. (2005). Ecological and biogeo-graphic relationships of class Flavobacteria in theSouthern Ocean. FEMS Microbiol Ecol 51: 265–277.
Allen M, Lauro FM, Williams TJ, Burg D, Siddiqui KS, DeFrancisci D et al. (2009). The genome sequence of thepsychrophilic archaeon, Methanococcoides burtonii:the role of genome evolution in cold-adaptationISME J 3: 1012–1035.
Australian Commonwealth Government Australian Ant-arctic Science Strategic Plan 2011–12 to 2020–21(2011). Australian Commonwealth Government: ISBN978 1 876934 16 6.
Beja O, Aravind L, Koonin EV, Suzuki MT, Hadd A,Nguyen LP et al. (2000). Bacterial rhodopsin: evidencefor a new type of phototrophy in the sea. Science 15:1902–1906.
Beja O, Spudich EN, Spudich JL, Leclerc M, DeLong EF.(2001). Proteorhodopsin phototrophy in the ocean.Nature 411: 786–789.
Berg IA, Kockelkorn D, Buckel W, Fuchs G. (2007). A3-hydroxypropionate/4-hydroxybutyrate autotrophiccarbon dioxide assimilation pathway in Archaea.Science 318: 1782–1786.
Blanvillain S, Meyer D, Boulanger A, Lautier M, Guynet C,Denance N et al. (2007). Plant carbohydrate scaven-ging through TonB-dependent receptors: a featureshared by phytopathogenic and aquatic bacteriaPLoS One 2: 224.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1897
The ISME Journal

Buchan A, Gonzalez JM, Moran MA. (2005). Overview ofthe marine Roseobacter lineage. Appl Environ Micro-biol 71: 5665–5677.
Burg DW, Lauro FM, Williams TJ, Raftery MJ, Guilhaus M,Cavicchioli R. (2010). Analyzing the hydrophobicproteome of the antarctic archaeon Methanococcoidesburtonii using differential solubility fractionationJ Proteome Res 9: 664–676.
Canfield DE, Stewart FJ, Thamdrup B, De Brabandere L,Dalsgaard T, Delong EF et al. (2010). A cryptic sulfurcycle in oxygen-minimum-zone waters off the Chileancoast. Science 330: 1375–1378.
Cho JC, Giovannoni SJ. (2004). Cultivation and growthcharacteristics of a diverse group of oligotro-phic marine Gammaproteobacteria. Appl EnvironMicrobiol 70: 432–440.
Church MJ, DeLong EF, Ducklow HW, Karner MB, PrestonCM, Karl DM. (2003). Abundance and distribution ofplanktonic Archaea and Bacteria in the waters westof the Antarctic Peninsula. Limnol Oceanogr 48:1893–1902.
Cottrell MT, Kirchman DL. (2000). Natural assemblages ofmarine Proteobacteria and members of the Cytophaga-Flavobacter cluster consuming low- and high-mole-cular weight dissolved organic matter. Appl EnvironMicrobiol 66: 1692–1697.
Danson MJ, Lamble HJ, Hough DW. (2007). Centralmetabolism. In: Caviccholi R (ed): Archaea: Molecularand Cellular Biology. ASM Press: Washington DC,pp 260–287.
DeLong EF, Beja O. (2010). The light-driven proton pumpproteorhodopsin enhances bacterial survival duringtough times. PLoS Biol 8: e1000359.
DeLong EF, Wu KY, Prezelin BB, Jovine RVM. (1994).High abundance of Archaea in Antarctic marinepicoplankton. Nature 371: 695–697.
DeMaere MZ, Lauro FM, Thomas T, Yau S, Cavicchioli R.(2011). Simple high-throughput annotation pipeline(SHAP). Bioinformatics 27: 2431–2432.
Dixon JL, Beale R, Nightingale PD. (2011). Rapid biologicaloxidation of methanol in the tropical Atlantic:significance as a microbial carbon source. BiogeosciDiscuss 8: 3899–3921.
Farmer CT, Hansell DH. (2007). Determination of dis-solved organic carbon and total dissolved nitrogen insea water. In: Dickson AG, Sabina CL, Christian JR(eds): Guide to Best Practices for Ocean CO2 Measure-ments. PICES Special Publication: Sydney, BritishColumbia, Canada, p 3191.
Forward JA, Behrendt MC, Wyborn NR, Cross R, Kelly DJ.(1997). TRAP transporters: a new family of periplas-mic solute transport systems encoded by the dctPQMgenes of Rhodobacter capsulatus and by homologs indiverse gram-negative bacteria. J Bacteriol 179:5482–5493.
Fuhrman JA, Schwalbach MS, Stingl U. (2008). Proteo-rhodopsins: an array of physiological roles? Nat RevMicrobiol 6: 488–494.
Ghiglione JF, Murray AE. (2012). Pronounced summer towinter differences and higher wintertime richness incoastal sub-Antarctic and Antarctic marine bacterio-plankton. Environ Microbiol 14: 617–629.
Gilbert HJ. (2008). Sus out sugars in. Structure 16:987–989.
Giovannoni SJ, Bibbs L, Cho JC, Stapels MD, Desiderio R,Vergin KL et al. (2005). Proteorhodopsin in theubiquitous marine bacterium SAR11. Nature 438: 82–85.
Giovannoni SJ, Hayakawa DH, Tripp HJ, Stingl U, GivanSA, Cho JC et al. (2008). The small genome of anabundant coastal ocean methylotroph. Environ Micro-biol 10: 1771–1782.
Goltsman DS, Denef VJ, Singer SW, VerBerkmoes NC,Lefsrud M et al. (2009). Community genomic andproteomic analyses of chemoautotrophic iron-oxidiz-ing ‘‘Leptospirillum rubarum’’ (Group II) and ‘‘Leptos-pirillum ferrodiazotrophum’’ (Group III) bacteria inacid mine drainage biofilms. Appl Environ Microbiol75: 4599–4615.
Gomez-Consarnau L, Gonzalez JM, Coll-Llado M, GourdonP, Pascher T, Neutze R et al. (2007). Light stimulatesgrowth of proteorhodopsin-containing marine Flavo-bacteria. Nature 445: 210–213.
Gonzalez JM, Fernandez-Gomez B, Fernandez-Guerra A,Gomez-Consarnau L, Sanchez O, Coll-Llado M et al.(2008). Genome analysis of the proteorhodopsin-containing marine bacterium Polaribacter sp.MED152 (Flavobacteria). Proc Natl Acad Sci USA105: 8724–8729.
Grossart H-P, Levold F, Allgaier M, Simon M, Brinkhoff T.(2005). Marine diatom species harbour distinct bacter-ial communities. Environ Microbiol 7: 860–873.
Grzymski JJ, Riesenfeld CS, Williams TJ, Dussaq AM,Ducklow H, Erickson M et al. (2012). A metagenomicassessment of winter and summer bacterioplanktonfrom Antarctica Peninsula coastal surface waters. ISME J;e-pub ahead of print; doi:10.1038/ismej.2012.31.
Hallam SJ, Konstantinidis KT, Putnam N, Schleper C,Watanabe Y, Sugahara J et al. (2006a). Genomicanalysis of the uncultivated marine crenarchaeoteCenarchaeum symbiosum. Proc Natl Acad Sci USA103: 18296–18301.
Hallam SJ, Mincer TJ, Schleper C, Preston CM, Roberts K,Richardson PM et al. (2006b). Pathways of carbonassimilation and ammonia oxidation suggested byenvironmental genomic analyses of marine Crenarch-aeota. PLoS Biol 4: e95.
Hayashi NR, Arai H, Kodama T, Igarashi Y. (1997). Thenovel genes, cbbQ and cbbO, located downstreamfrom the RubisCO genes of Pseudomonas hydroge-nothermophila, affect the conformational states andactivity of RubisCO. Biochem Biophys Res Commun241: 565–569.
Heikes BG, Chang W, Pilson MEQ, Swift E, Singh HB,Guenther A et al. (2002). Atmospheric methanolbudget and ocean implication. Global BiogeochemCycles 16: 1133.
Holtmann G, Bremer E. (2004). Thermoprotection ofBacillus subtilis by exogenously provided glycinebetaine and structurally related compatible solutes:involvement of Opu transporters. J Bacteriol 186:1683–1693.
Kappler U. (2011). Bacterial sulfite-oxidizing enzymes.Biochim Biophys Acta 1807: 1–10.
Karner M, DeLong EF, Karl DM. (2001). Archaeal dom-inance in the mesopelagic zone of the Pacific Ocean.Nature 409: 507–509.
Kirchman D, K’Nees E, Hodson R. (1985). Leucineincorporation and its potential as a measure of proteinsynthesis by bacteria in natural aquatic systems. ApplEnviron Microbiol 49: 599–607.
Kockelkorn D, Fuchs G. (2009). Malonic semialdehydereductase, succinic semialdehyde reductase, and succi-nyl-coenzyme A reductase from Metallosphaera sedula:enzymes of the autotrophic 3-hydroxypropionate/
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1898
The ISME Journal

4-hydroxybutyrate cycle in Sulfolobales. J Bacteriol 191:6352–6362.
Koebnik R. (2005). TonB-dependent trans-envelope signal-ling: the exception or the rule? Trends Microbiol 13:343–347.
Konneke M, Bernhard AE, de la Torre JR, Walker CB,Waterbury JB, Stahl DA. (2005). Isolation of anautotrophic ammonia-oxidizing marine archaeonNature 437: 543–546.
Lauro FM, DeMaere MZ, Yau S, Brown M, Ng C, Wilkins Det al. (2011). An integrative study of a meromictic lakeecosystem in Antarctica. ISME J 5: 879–895.
Lauro FM, McDougald D, Thomas T, Williams TJ, Egan S,Rice S et al. (2009). The genomic basis of trophicstrategy in marine bacteria. Proc Natl Acad Sci USA106: 15527–15533.
Lucker S, Wagner M, Maixner F, Pelletier E, Koch H,Vacherie B et al. (2010). A Nitrospira metagenomeilluminates the physiology and evolution of globallyimportant nitrite-oxidizing bacteria. Proc Natl AcadSci USA 107: 13479–13484.
Man D, Wang W, Sabehi G, Aravind L, Post AF, Massana Ret al. (2003). Diversification and spectral tuning inmarine proteorhodopsins. EMBO J 22: 1725–1731.
Markowitz VM, Chen IM, Chu K, Szeto E, Palaniappan K,Grechkin Y et al. (2012). IMG/M: the integratedmetagenome data management and comparative ana-lysis system. Nucleic Acids Res 40: (Database issue)D123–D129.
Martens-Habbena W, Berube PM, Urakawa H, de la TorreJR, Stahl DA. (2009). Ammonia oxidation kineticsdetermine niche separation of nitrifying Archaea andBacteria. Nature 461: 976–979.
Massana R, Pedros-Alio C, Casamayor EO, Gasol JM.(2001). Changes in marine bacterioplankton phyloge-netic composition during incubations designed tomeasure biogeochemically significant parameters.Limnol Oceanogr 46: 1181–1188.
Meyer B, Kuever J. (2007). Molecular analysis of thediversity of sulfate-reducing and sulfur-oxidizingprokaryotes in the environment, using aprA as func-tional marker gene. Appl Environ Microbiol 73: 7664–7679.
Moran MA, Belas R, Schell MA, Gonzalez JM, Sun F, SunS et al. (2007). Ecological genomics of marineRoseobacters. Appl Environ Microbiol 73: 4559–4569.
Moran MA, Zepp RG. (1997). Role of photoreactions in theformation of biologically labile compounds fromdissolved organic matter. Limnol Oceanogr 42:1307–1316.
Morris RM, Nunn BL, Frazar C, Goodlett DR, Ting YS,Rocap G. (2010). Comparative metaproteomics revealsocean-scale shifts in microbial nutrient utilization andenergy transduction. ISME J 4: 673–685.
Murray AE, Grzymski JJ. (2007). Diversity and genomics ofAntarctic marine micro-organisms. Philos Trans R SocLond B Biol Sci 362: 2259–2271.
Murray AE, Preston CM, Massana R, Taylor LT, Blakis A,Wu K et al. (1998). Seasonal and spatial variability ofbacterial and archaeal assemblages in the coastalwaters off Anvers Island, Antarctica. Appl EnvironMicrobiol 64: 2585–2595.
Nagano K, Murakami Y, Nishikawa K, Sakakibara J,Shimozato K, Yoshimura F. (2007). Characterizationof RagA and RagB in Porphyromonas gingivalis: studyusing gene deletion mutants. J Med Microbiol 56:1536–1548.
Newton IL, Woyke T, Auchtung TA, Dilly GF, Dutton RJ,Fisher MC et al. (2007). The Calyptogena magnificachemoautotrophic symbiont genome. Science 315:998–1000.
Ng C, DeMaere MZ, Williams TJ, Lauro FM, Raftery M,Gibson JA et al. (2010). Metaproteogenomic analysis ofa dominant green sulfur bacterium from Ace Lake,Antarctica. ISME J 4: 1002–1019.
Piquet AM, Bolhuis H, Meredith MP, Buma AG. (2011).Shifts in coastal Antarctic marine microbial commu-nities during and after melt water-related surfacestratification. FEMS Microbiol Ecol 76: 413–427.
Ramos-Vera WH, Weiss M, Strittmatter E, Kockelkorn D,Fuchs G. (2011). Identification of missing genes andenzymes for autotrophic carbon fixation in crenarch-aeota. J Bacteriol 193: 1201–1211.
Reeves AR, Wang G-R, Salyers AA. (1997). Characteriza-tion of four outer membrane proteins that play a rolein utilization of starch by Bacteroides thetaiotaomi-cron. J Bacteriol 179: 643–649.
Reisch CR, Stoudemayer MJ, Varaljay VA, Amster IJ,Moran MA, Whitman WB. (2011). Novel pathway forassimilation of dimethylsulphoniopropionate wide-spread in marine bacteria. Nature 473: 208–211.
Rodriguez-Brito B, Rohwer F, Edwards RA. (2006). Anapplication of statistics to comparative metagenomics.BMC Bioinformatics 7: 162.
Sharp JH, Benner R, Carlson CA, Dow R, Fitzwater SE.(1993). Re-evaluation of high temperature combustionand chemical oxidation measurements of dissolvedorganic carbon in seawater. Limnol Oceanogr 38:1774–1782.
Schattenhofer M, Fuchs BM, Amann R, Zubkov MV,Tarran GA, Pernthaler J. (2009). Latitudinal distribu-tion of prokaryotic picoplankton populations in theAtlantic Ocean. Environ Microbiol 11: 2078–2093.
Schwalbach MS, Tripp HJ, Steindler L, Smith DP,Giovannoni SJ. (2010). The presence of the glycolysisoperon in SAR11 genomes is positively correlatedwith ocean productivity. Environ Microbiol 12:490–500.
Smith DP, Kitner JB, Norbeck AD, Clauss TR, Lipton MS,Schwalbach MS et al. (2010). Transcriptional andtranslational regulatory responses to iron limitation inthe globally distributed marine bacterium CandidatusPelagibacter ubique. PLoS One 5: e10487.
Sowell SM, Abraham PE, Shah M, Verberkmoes NC, SmithDP, Barofsky DF et al. (2011). Environmental proteo-mics of microbial plankton in a highly productivecoastal upwelling system. ISME J 5: 856–865.
Sowell SM, Norbeck AD, Lipton MS, Nicora CD, CallisterSJ, Smith RD et al. (2008). Proteomic analysis ofstationary phase in the marine bacterium ‘‘CandidatusPelagibacter ubique’’. Appl Environ Microbiol 74:4091–4100.
Sowell SM, Wilhelm LJ, Norbeck AD, Lipton MS, NicoraCD, Barofsky DF et al. (2009). Transport functionsdominate the SAR11 metaproteome at low-nutrientextremes in the Sargasso Sea. ISME J 3: 93–105.
Steindler L, Schwalbach MS, Smith DP, Chan F, Giovan-noni SJ. (2011). Energy starved Candidatus Pelagibac-ter ubique substitutes light-mediated ATP productionfor endogenous carbon respiration. PLoS One 6:e19725.
Stewart FJ, Ulloa O, Delong EF. (2012). Microbialmetatranscriptomics in a permanent marine oxygenminimum zone. Environ Microbiol 14: 23–40.
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1899
The ISME Journal

Stocker R, Seymour JR, Samadani A, Hunt DE, Polz MF.(2008). Rapid chemotactic response enables marinebacteria to exploit ephemeral microscale nutrientpatches. Proc Natl Acad Sci USA 105: 4209–4214.
Strous M, Pelletier E, Mangenot S, Rattei T, Lehner A,Taylor MW et al. (2006). Deciphering the evolutionand metabolism of an anammox bacterium from acommunity genome. Nature 440: 790–794.
Sullivan MB, Coleman ML, Weigele P, Rohwer F,Chisholm SW. (2005). Three Prochlorococcus cya-nophage genomes: signature features and ecologicalinterpretations. PLoS Biol 3: e144.
Sun J, Steindler L, Thrash JC, Halsey KH, Smith DP, CarterAE et al. (2011). One carbon metabolism in SAR11pelagic marine bacteria. PLoS One 6: e23973.
Swan BK, Martinez-Garcia M, Preston CM, Sczyrba A,Woyke T, Lamy D et al. (2011). Potential for chemo-lithoautotrophy among ubiquitous bacteria lineages inthe dark ocean. Science 333: 1296–1300.
Thomas T, Egan S, Burg D, Ng C, Ting L, Cavicchioli R.(2007). The integration of genomics and proteomicsinto marine, microbial ecology. Theme series on‘‘Genomics, proteomics and metabolomics in marineecology’’. Mar Ecol Prog Ser 332: 291–299.
Ting L, Williams TJ, Cowley MJ, Lauro FM, Guilhaus M,Raftery MJ et al. (2010). Cold adaptation in themarine bacterium, Sphingopyxis alaskensis, assessedusing quantitative proteomics. Environ Microbiol 12:2658–2676.
Tourna M, Stieglmeier M, Spang A, Konneke M, Schintl-meister A, Urich T et al. (2011). Nitrososphaeraviennensis, an ammonia oxidizing archaeon from soil.Proc Natl Acad Sci USA 108: 8420–8425.
Tripp HJ, Kitner JB, Schwalbach MS, Dacey JW, WilhelmLJ, Giovannoni SJ. (2008). SAR11 marine bacteriarequire exogenous reduced sulphur for growth. Nature452: 741–744.
Tripp HJ, Schwalbach MS, Meyer MM, Kitner JB, BreakerRR, Giovannoni SJ. (2009). Unique glycine-activatedriboswitch linked to glycine-serine auxotrophy inSAR11. Environ Microbiol 11: 230–238.
Turner J, Colwell SR, Marshall GJ, Lachlan-Cope TA,Carleton AM, Jones PD et al. (2005). Antarctic climatechange during the last 50 years. Int J Climatol 25:279–294.
Walker CB, de la Torre JR, Klotz MG, Urakawa H, Pinel N,Arp DJ et al. (2010). Nitrosopumilus maritimusgenome reveals unique mechanisms for nitrificationand autotrophy in globally distributed marine cre-narchaea. Proc Natl Acad Sci USA 107: 8818–8823.
Walsh DA, Zaikova E, Howes CG, Song YC, Wright JJ,Tringe SG et al. (2009). Metagenome of a versatilechemolithoautotroph from expanding oceanic deadzones. Science 326: 578–582.
Williams TJ, Burg DW, Raftery MJ, Poljak A, Guilhaus M,Pilak O et al. (2010). Global proteomic analysis of theinsoluble, soluble, and supernatant fractions of thepsychrophilic archaeon Methanococcoides burtoniiPart I: the effect of growth temperature. J ProteomeRes 9: 640–652.
Williams TJ, Ertan H, Ting L, Cavicchioli R. (2009). Carbonand nitrogen substrate utilization in the marinebacterium Sphingopyxis alaskensis strain RB22. ISMEJ 3: 1036–1052.
Winnen B, Hvorup RN, Saier MH Jr. (2003). The tripartitetricarboxylate transporter (TTT) family. Res Microbiol154: 457–465.
Wuchter C, Abbas B, Coolen MJ, Herfort L, van BleijswijkJ, Timmers P et al. (2006). Archaeal nitrification in theocean. Proc Natl Acad Sci USA 103: 12317–12322.
Yau S, Lauro FM, DeMaere MZ, Brown MV, Thomas T,Raftery MJ et al. (2011). Virophage control of Antarcticalgal host–virus dynamics. Proc Natl Acad Sci USA108: 6163–6168.
Supplementary Information accompanies the paper on The ISME Journal website (http://www.nature.com/ismej)
Antarctic Peninsula coastal metaproteomeTJ Williams et al
1900
The ISME Journal




















