
The telomere/telomerase system in autoimmune and systemic immune-mediated diseases
Upload
independentCategory
view
3download
0
A Key Role for Telomerase Reverse Transcriptase Unit inModulating Human Embryonic Stem Cell Proliferation,Cell Cycle Dynamics, and In Vitro Differentiation
CHUNBO YANG,a,b STEFAN PRZYBORSKI,a,c MICHAEL J. COOKE,a,c XIN ZHANG,a,b REBECCA STEWART,a,b
GEORGE ANYFANTIS,a,b STUART P. ATKINSON,a,b GABRIELE SARETZKI,a,d LYLE ARMSTRONG,a,b
MAJLINDA LAKOa,b
aNorth East Institute for Stem Cell Research, bInstitute of Human Genetics, and dCrucible Laboratory, Institute forAgeing and Health, University of Newcastle upon Tyne, International Centre for Life, Newcastle upon Tyne, UnitedKingdom; cSchool of Biological Sciences, University of Durham, Durham, United Kingdom
Key Words. Telomerase reverse transcriptase • Telomerase RNA component • Telomerase • Human embryonic stem cells • Cell cycle •In vitro differentiation
ABSTRACT
Embryonic stem cells (ESC) are a unique cell populationwith the ability to self-renew and differentiate into all threegerm layers. Human ESC express the telomerase reversetranscriptase (TERT) gene and the telomerase RNA (TR)and show telomerase activity, but TERT, TR, and telomeraseare all downregulated during the differentiation process. Toexamine the role of telomerase in human ESC self-renewaland differentiation, we modulated the expression of TERT.Upregulation of TERT and increased telomerase activityenhanced the proliferation and colony-forming ability ofhuman ESC, as well as increasing the S phase of the cellcycle at the expense of a reduced G1 phase. Upregulation ofTERT expression was associated with increases in CYCLIND1 and CDC6 expression, as well as hyperphosphorylationof RB. The differentiated progeny of control ESC showedshortening of telomeric DNA as a result of loss of telomerase
activity. In contrast, the differentiated cells from TERT-overexpressing ESC maintained high telomerase activityand accumulated lower concentrations of peroxides thanwild-type cells, implying greater resistance to oxidativestress. Although the TERT-overexpressing human ESC areable to form teratoma composed of three germ layers invivo, their in vitro differentiation to all primitive and em-bryonic lineages was suppressed. In contrast, downregula-tion of TERT resulted in reduced ESC proliferation, in-creased G1, and reduced S phase. Most importantly,downregulation of TERT caused loss of pluripotency andhuman ESC differentiation to extraembryonic and embry-onic lineages. Our results indicate for the first time animportant role for TERT in the maintenance of human ESCpluripotency, cell cycle regulation, and in vitro differentia-tion capacity. STEM CELLS 2008;26:850–863
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
The telomerase holoenzyme adds repetitive G-rich sequences tochromosome ends to maintain the structure of chromosome endsand protect chromosomes from degradation and end-to-end fu-sion [1–3]. The basic components required for telomerase ac-tivity are the telomerase reverse transcriptase (TERT) unit,which possesses catalytic activity, and the telomerase RNA(TR) component, which contains the template for telomereelongation.
Most tissues express TR, at various levels; however, manyappear to lack telomerase activity, perhaps suggesting thatTERT is the rate-limiting factor for assembly of an activetelomerase complex. In most somatic cells, telomerase activityis silenced or is present at very low levels, whereas cancer-derived cell lines, germline cells, and embryonic stem cellsshow abundant telomerase expression. Beyond its role in telo-mere lengthening, it has been suggested that telomerase prevents
the loss of the G-rich single-stranded overhangs that participate,together with specific telomere-binding proteins, in forming theT-loop structure [4]. Erosion of the telomeric overhang seen insenescent cells is likely to cause collapse of the T-loop structureand uncapping of the telomere end, which eventually signalDNA damage and activate the senescence pathway [5]. Consis-tent with this, a reduction in telomerase activity can causeerosion of the overhangs in normal human cells, whereas ectopicexpression of the TERT gene maintains the overhangs at a lengthsimilar to that found in early passage cells [6].
To date, mounting evidence suggests that telomerase hasadditional extratelomeric roles in mediating cell survival andantiapoptotic functions against various cytotoxic stresses[7–13]. In many cell lines, exogenous telomerase was shown tostimulate cell proliferation, although the mechanistic basis forthese observations is not completely understood. Our data andthose of others have suggested that overexpression of TERT isassociated with alteration in the expression profile of growth-promoting genes and suppression of growth-inhibitory genes
Correspondence: Majlinda Lako, B.Sc., Ph.D., International Centre for Life, Central Parkway, Newcastle upon Tyne NE1 3BZ, UnitedKingdom. Telephone: 00-44-191-241-8688; Fax: 00-44-191-241-8666; e-mail: [email protected] Received August 20, 2007;accepted for publication January 8, 2008; first published online in STEM CELLS EXPRESS January 17, 2008. ©AlphaMed Press 1066-5099/2008/$30.00/0 doi: 10.1634/stemcells.2007-0677
EMBRYONIC STEM CELLS
STEM CELLS 2008;26:850–863 www.StemCells.com
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from

[14–16]. It remains to be elucidated whether TERT can act as atranscription factor or as an effector of gene expression indi-rectly through certain signaling pathways.
Current knowledge of TERT regulation and telomerase ac-tivity derives largely from studies of neoplastic and/or immor-talized cell lines. Although useful for understanding cancermodels, such studies may be less relevant to normal euploidcells, and there is a need to study telomerase regulation innormal development. In this context, studies on embryonic andsomatic stem cells might prove extremely useful, since theyprovide an in vitro model of cell differentiation, where effects oftelomerase modulation can be easily assessed. For example,forced telomerase expression in human mesenchymal stem cellsresults in elongation of telomeres, extended life span, and en-hanced differentiation potential [17]. Similarly, upregulation ofTert expression in the K5-mTert mice promotes epidermal stemcell mobilization and proliferation in the absence of changes intelomere length [18]. Induction of Tert expression in mouse skinepithelium caused the hair follicle cycle to undergo rapid tran-sition from a resting telogen phase to an active anagen phase[19].
In mouse, telomerase activity is important for ESCgrowth: deletions leading to loss of either the telomeraseRNA (mTr) or its reverse transcriptase (mTert) result inprogressive loss of telomeres, genomic instability, aneu-ploidy, telomeric fusions, and eventual reduced growth rate[20 –22]. Data generated by our group have shown that over-expression of Tert in murine ESC enhances self-renewal andimproves resistance to apoptosis, oxidative stress, and in-creased proliferation, suggesting that telomerase functions asa survival enzyme in ESC [14]. Our recent work has indicatedthat human ESC express the TERT and TR genes and showhigh levels of telomerase activity; however, upon differenti-ation, the levels of TERT, TR, and telomerase activity de-crease with the emergence of a maturing population of cells[23]. Downregulation of telomerase activity with differenti-ation has also been reported in embryonic carcinoma cellsand immortalized cells lines that have the ability to beinduced to differentiate into more mature cells, and this hasbeen linked to downregulation in TERT expression ratherthan posttranslational modifications. Importantly, downregu-lation of TERT was found to be an early and direct event ofcellular differentiation and not the cell cycle [24 –27]. Whatis not clear from these studies is whether the downregulationof telomerase activity and TERT is a consequence of differ-entiation or is necessary for the differentiation to proceednormally. Overexpression of TERT appears to block terminaldifferentiation of HaCaT skin keratinocytes, neuronal differ-entiation of NT2 cells, and late development of immortalizedsheep fibroblasts but not bovine adrenocortical cells or hu-man adult bone cells [24 –27]. In view of these results, it is ofgreat value to investigate whether TERT overexpression alsoendows human ESC with these advantageous features.
Cell replacement therapy is a promising treatment for dis-eases such as hematologic malignancies. However, during dif-ferentiation of ESC, the cells are subjected to telomere erosion[23]. In fact, several cases of leukemia with graft failure afterhematopoietic stem cell transplant have been reported to haveunusual telomere shortening [28]. In this study, we overex-pressed and downregulated TERT in human ESC to investigatethe impacts of telomerase modulation on self-renewal, differen-tiation, and viability and to explore the possibility of telomerase-based strategies of large-scale growth of ESC and induceddifferentiation.
MATERIALS AND METHODS
Culture and Differentiation of Human ESCThe human ESC lines H1 (WiCell Research Institute, Madison, WI,http://www.wicell.org) and human embryonic stem (hES)-NCL1[29] were routinely passaged and maintained in human embryonicstem cell (hESC) media on mitotically inactivated mouse embryonicfibroblast feeder layers as described by Stojkovic et al. [29]. Dif-ferentiation was achieved by forming embryoid bodies (EBs) asdescribed previously [29]. One to two passages prior to experi-ments, human ESC were transferred to Matrigel-coated plates (BDBiosciences, San Diego, http://www.bdbiosciences.com) with feed-er-conditioned media, as described previously [29]. Transfection ofTERT and control cDNA was carried out in the H1 cell line (passage29) and hES-NCL1 (passage 24).
Generation of Human TERT-Overexpressing hESCSublinesTen micrograms of the linearized construct pTP6 and pTP6 con-taining full-length human telomerase reverse transcriptase (hTERT)cDNA was transfected into hESC using the Amaxa (Gaithersburg,MD, http://www.amaxa.com) nucleofector kit Cell Line L kit andprogram A-23. Two days after the transfection, stable clones wereselected using puromycin selection (1 �g/ml) for 10 days. Thesurviving human ESC from each cell line were pooled together andexpanded to form the TERT-overexpressing sublines, named H1TERT and hES-NCL1 TERT, or the control sublines, named H1control and hES-NCL1 control. All four sublines were maintainedwith 0.6 �g/ml puromycin to ensure the maintenance of transgene.Every 10–15 passages, quantitative reverse transcription (RT)-poly-merase chain reaction (PCR), Western blot, and flow cytometryanalysis were carried out to confirm TERT overexpression overtime. All four sublines were maintained in culture up to 70 passagespost-transfection.
Flow Cytometry Analysis of hESCFor the flow cytometry analysis, the hESC were collected usingcollagenase IV treatment (1 mg/ml for 5 minutes) followed by briefAccutase incubation (Chemicon, Temecula, CA, http://www.chemicon.com). Human ESC were suspended in staining buffer(phosphate-buffered saline � 5% fetal calf serum) at a concentra-tion of 106 cells per milliliter. Intracellular staining was performedby fixing the cells for 10 minutes in 0.5% formaldehyde followed bygentle permeabilization in ice-cold 90% methanol. The permeabil-ized cells were washed three times in staining buffer before beingstained with the primary antibody (TERT; 1:500; Rockland Immu-nochemicals, Gilbertsville, PA, http://www.rockland-inc.com) for 2hours at room temperature. Three washes in staining buffer werecarried out before staining with the secondary antibody, goat anti-rabbit IgG-fluorescein isothiocyanate (FITC) (final concentration, 6�g/ml; Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com).Cells were washed again three times and resuspended in stainingbuffer before being analyzed with a FACSCalibur instrument (BDBiosciences, San Diego, http://www.bdbiosciences.com) using theCellQuest software (BD Biosciences). For each sample, 10,000events were acquired.
Small Interfering RNAs and TransfectionShort interfering RNAs (siRNAs) were obtained from Invitrogen.The siRNA sequences are shown in supplemental online Table 1.Transfection with a scrambled control siRNA provided by the samecompany was used as a negative control. Transfection of siRNAinto human ESC was carried out using the high-efficiency nucleo-fection kit L from Amaxa and 80 pmol of siRNA (in 2 ml ofmedium) as outlined in the manufacturer’s instructions.
Telomeric Repeat Amplification Protocol AssayTelomeric repeat amplification protocol reactions were carried outusing Telo TAGGG Telomerase PCR ELISA Plus (Roche Diagnos-tics, Indianapolis, http://www.roche-applied-science.com) follow-
851Yang, Przyborski, Cooke et al.
www.StemCells.com
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from

ing the manufacturer’s instructions and as described previously[14].
LightCycler Real-Time PCR AnalysisReal-time PCR analysis was carried out using QuantiTect SYBRGreen PCR Master Mix (Qiagen, Hilden, Germany, http://www1.qiagen.com). GAPDH for each sample was used as the internalcontrol of these real-time analyses. The data were analyzed usingthe LightCycler relative quantification software, version 1.01(Roche Diagnostics). For each gene, the control was normalized to1, and all other values were calculated with respect to this. PCRswere carried out using the primers described in supplemental onlineTable 2.
Western BlottingLysates were electrophoresed on a 10% SDS-polyacrylamide gelelectrophoresis gel and electrophoretically transferred to a poly-vinylidene difluoride membrane (Hybond-P; Amersham Biosciences,Piscataway, NJ, http://www.amersham.com). Membranes wereblocked in Tris-buffered saline with 5% milk and 0.1% Tween. Theblots were probed with primary antibodies overnight and revealedwith horseradish peroxidase-conjugated secondary antibodies. Fullinformation on antibodies used and their concentrations can befound in supplemental online Appendix A.
Alkaline Phosphatase StainingAlkaline phosphatase (AP) staining was carried out using theAlkaline Phosphatase Detection Kit following the manufacturer’sinstructions (Chemicon). The staining method was described previ-ously [29].
Measurement of Cell Proliferation Using the5-Bromo-2�-Deoxyuridine Incorporation MethodHuman ESC proliferation was measured by incorporation of 5-bro-mo-2�-deoxyuridine (BrdU) (Roche Diagnostics) into the genomicDNA during the S phase of the cell cycle. Human ESC wereincubated and processed with a BrdU Flow Kit (BD Biosciences)according to the manufacturer’s protocol. Cells were stained withFITC, anti-BrdU, and 7-aminoactinomycin D. Cells from the samepopulation that were not BrdU-labeled were used as negative con-trol. Flow cytometry analysis was carried out using a FACSCalibur(Becton, Dickinson and Company, San Jose, CA, http://www.bd.com) and CellQuest software.
Cell Cycle AnalysisCell cycle analysis was performed using the CycleTest Plus DNAreagent kit (Becton Dickinson) as described previously [14]. Thedata were analyzed using FlowJo software to generate percentagesof cells in G1, S, and G2/M phases.
Reactive Oxygen Species MeasurementCellular peroxide levels were assessed by staining with 30 �Mdihydrorhodamine 123 (DHR) (Molecular Probes, Eugene, OR,http://probes.invitrogen.com) for 30 minutes at 37° C and analyzedusing FL3 fluorescence as described previously [30].
Telomere LengthThe mean length of telomeric repeats was measured by flow cy-tometry following the hybridization of a fluorescently conjugatedpeptide nucleic acid probe (DakoCytomation, Glostrup, Denmark,http://www.dakocytomation.com). A full description of the protocolis given in supplemental online Appendix A. Relative telomerelength was calculated following the manufacturer’s instructions andexpressed as the percentage of the average telomere fluorescenceper genome. Then, the relative telomere length for the control cells(1,301) was normalized to 100%, and all experimental samples werecalculated with respect to this.
Apoptosis AssayApoptosis assay was carried out using the Annexin V-FITC apop-tosis detection kit (BD Biosciences) as described previously [14].
Karyotype Analysis of hESCChromosome preparations were made using standard cytogenetictechniques and a 16-hour colcemid/BrdU mitotic arrest step. Thekaryotype of ESC was determined by the standard G-banding pro-cedure.
Hematopoietic CFU-GEMM AssayHematopoietic CFU-GEMM assays were performed according tothe manufacturer’s instructions (StemCell Technologies, Vancou-ver, BC, Canada, http://www.stemcell.com) essentially as describedpreviously [14].
Cell Signaling AssaysPanorama antibody microarrays for cell signaling, containing 224different antibodies spotted in duplicate on nitrocellulose-coatedglass, were purchased from Sigma-Aldrich. One milligram of cellextracts prepared from TERT-overexpressing and control sublineswas collected, labeled with Cy3 and Cy5, respectively, and hybrid-ized to the slides according to manufacturer’s instructions. Cy3 andCy5 signals were read on Gene Pix Pro 4.0. More information onanalysis of results is given in supplemental online Appendix A.
Tumor Formation in Severe CombinedImmunodeficient MiceAll procedures involving mice were carried out in accordance withinstitutional guidelines. Approximately 106 hESC were injected intothe testis of adult male severe combined immunodeficient (SCID)mice. After 70–90 days, mice were sacrificed, tissues were dis-sected, and material was analyzed by histology. A full description ofthe method is presented in supplemental online Appendix A.
Statistical AnalysisTwo-tailed pairwise Student’s t test was used to analyze resultsobtained from two samples with one time point. Analysis of vari-ance (two factors with replication) was used to compare multiplesamples (at several time points). The results were considered sig-nificant if p � .05.
RESULTS
Generation and Characterization ofTERT-Overexpressing Human ESC ClonesTwo human ESC lines (H1 and hES-NCL1) were stably trans-fected with the empty construct pTP6 or pTP6-TERT to gener-ate two control sublines (H1 control and hES-NCL1 control)and two TERT-overexpressing sublines (H1 TERT and hES-NCL1 TERT), respectively. Real-time RT-PCR (Fig. 1A) andWestern blotting (Fig. 1B) were carried out to confirm overex-pression of full-length TERT cDNA. To ensure maintenance ofTERT overexpression, all the sublines were maintained in thepresence of 0.6 �g/ml puromycin and analyzed every 10–15passages by real-time RT-PCR and Western blot or flow cytom-etry analysis. This showed maintenance of TERT overexpressionover time in culture (Fig. 1C). Real-time RT-PCR analysisshowed that overexpression of TERT did not result in changes inTR expression (supplemental online Fig. 1). Overexpression ofTERT resulted in the generation of colonies that showed thetypical tightly packed human ESC morphology with large nucleiand visible nucleoli (Fig. 1D). Although H1 TERT and hES-NCL1 TERT ESC sublines overexpressed TERT mRNA atdifferent levels, the telomerase activity levels were similar toeach other (Fig. 1E). Most probably, a 10-fold overexpression of
852 Key Role for TERT in hESC Proliferation and Differentiation
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from
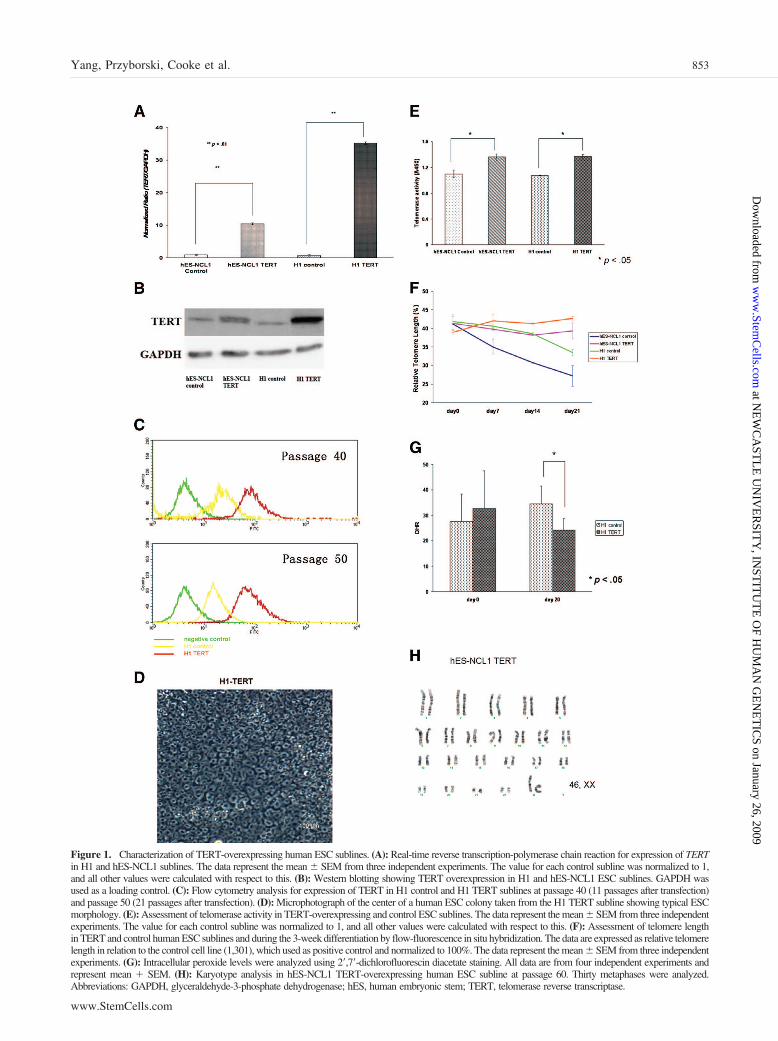
Figure 1. Characterization of TERT-overexpressing human ESC sublines. (A): Real-time reverse transcription-polymerase chain reaction for expression of TERTin H1 and hES-NCL1 sublines. The data represent the mean � SEM from three independent experiments. The value for each control subline was normalized to 1,and all other values were calculated with respect to this. (B): Western blotting showing TERT overexpression in H1 and hES-NCL1 ESC sublines. GAPDH wasused as a loading control. (C): Flow cytometry analysis for expression of TERT in H1 control and H1 TERT sublines at passage 40 (11 passages after transfection)and passage 50 (21 passages after transfection). (D): Microphotograph of the center of a human ESC colony taken from the H1 TERT subline showing typical ESCmorphology. (E): Assessment of telomerase activity in TERT-overexpressing and control ESC sublines. The data represent the mean � SEM from three independentexperiments. The value for each control subline was normalized to 1, and all other values were calculated with respect to this. (F): Assessment of telomere lengthin TERT and control human ESC sublines and during the 3-week differentiation by flow-fluorescence in situ hybridization. The data are expressed as relative telomerelength in relation to the control cell line (1,301), which used as positive control and normalized to 100%. The data represent the mean � SEM from three independentexperiments. (G): Intracellular peroxide levels were analyzed using 2�,7�-dichlorofluorescin diacetate staining. All data are from four independent experiments andrepresent mean � SEM. (H): Karyotype analysis in hES-NCL1 TERT-overexpressing human ESC subline at passage 60. Thirty metaphases were analyzed.Abbreviations: GAPDH, glyceraldehyde-3-phosphate dehydrogenase; hES, human embryonic stem; TERT, telomerase reverse transcriptase.
853Yang, Przyborski, Cooke et al.
www.StemCells.com
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from

TERT, as found in hES-NCL1 TERT subline, is already suffi-cient to saturate the formation of active telomerase complexes.An alternate possibility is that human TR or other componentsof the telomerase holoenzyme became limiting. This couldperhaps explain why the H1 and hES-NCL1 TERT sublinesshowed increases in telomerase activity of only 30% and 26%,respectively, compared with the controls (Fig. 1E). Identicalresults were obtained when different amounts of cellular ex-tracts were used for measurement of telomerase activity (datanot shown), confirming the validity of the results and lack of anyinhibitors which can be detrimental for this type of assay.
In a recent study, we showed that telomerase activity andtelomere length measured by metaphase fluorescence in situhybridization (FISH) decreased during differentiation of humanESC [23]. Using flow-FISH, we were able to confirm a signif-icant decrease in telomere length during the 3-week differenti-ation time course of control sublines (p � .05; Fig. 1F). Mostimportantly, a significant difference was observed between theTERT-overexpressing and control sublines (p � .01; Fig. 1F),since the TERT-overexpressing sublines did not show the telo-mere length decrease observed upon differentiation of the con-trols.
Human ESC have highly efficient antioxidant defensemechanisms, but this protective capacity is progressively lostduring differentiation into embryoid bodies [23]. To investigatewhether continued expression of TERT would interfere with theloss of antioxidant defense during differentiation, we measuredintracellular levels of peroxides by DHR staining. Differentia-tion of control subline into embryoid bodies resulted in 25%increase in cellular DHR fluorescence at day 20, in accordancewith our earlier results [23]. In contrast, peroxide levels de-creased by 25% during differentiation of TERT-overexpressinghuman ESC (Fig. 1G).
To investigate whether continuous overexpression of TERTin human ESC contributes to gross genomic instability, wecarried out G banding on TERT and control sublines at 30 and60 passages after transfection (Fig. 1H). Observation of 20metaphases for each subline showed no deviations from thenormal karyotype.
Overexpression of TERT Results in Enhancement ofHuman ESC Pluripotency and Suppression of the InVitro Differentiation CapacityTo investigate the effects of continuous TERT overexpressionon human ESC pluripotency, we carried out AP staining anal-ysis and observed significantly higher numbers of AP� inTERT-overexpressing sublines compared with the controls (Fig.2A). Human ESC from each subline were also injected into thetestis of SCID mice. One day prior to injection, a small sampleof cells from all sublines was analyzed by real-time RT-PCR(data not shown) and flow cytometry to ensure TERT overex-pression (Fig. 1C). Injection of TERT and control ESC sublinesinto the testis of SCID mice resulted in formation of teratoma.Although similar numbers of ESC were injected from eachsubline, the sizes of tumor were very different, with the TERTteratomas being much larger than the ones resulting from thecontrol sublines (Fig. 2B). In both cases, teratomas were com-posed of cells from all three germ layers, indicating the pluri-potent nature of the cells (Fig. 2C). The identity of cells derivedfrom ectoderm (Fig. 2D) within the tumors was confirmed byimmunohistochemical staining with Nestin, epithelial keratin,neurofilament 200, and glial fibrillary acidic protein. Stainingwith smooth muscle actin was used to confirm presence of cellsderiving from mesoderm, and caudal-related homeobox tran-scription factor antibody (CDX2), shown to play an importantrole in intestinal development, differentiation, and homeostasis
in adults, was used to confirm cells derived from endoderm (Fig.2D). An additional endodermal marker, �-fetoprotein, was usedto identify cells of endodermal origin (Fig. 2D).
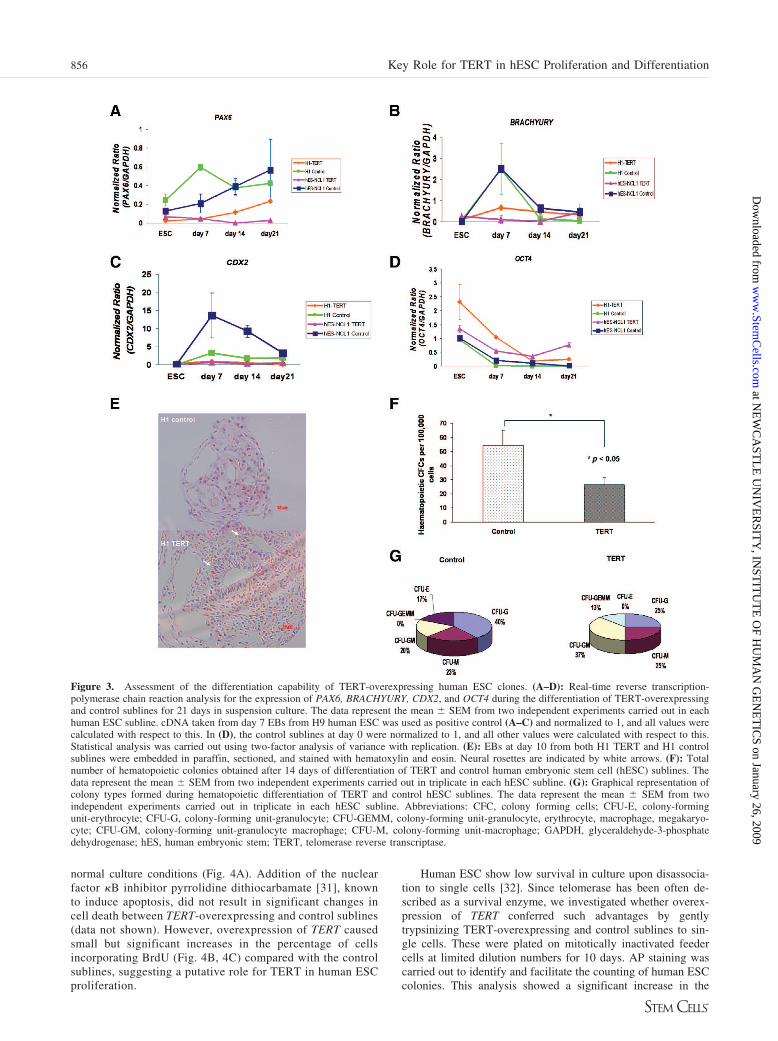
Several recent reports have shown that overexpression ofTERT does suppress terminal differentiation of HaCat skinkeratocytes and NT2 embryonic carcinoma cells [24, 25]. Toinvestigate this more closely in our system, we induced differ-entiation of human ESC using the EB method and quantitated ina series of time points expression of the neuroepithelial marker(PAX6; Fig. 3A), the mesodermal marker (BRACHYURY; Fig.3B), the trophectodermal marker (CDX2; Fig. 3C), primitiveendodermal markers (IHH and GATA6; data not shown), and theprimitive ectoderm marker (FGF5; data not shown). In all cases,we noticed that TERT-overexpressing human ESC sublineswere more resistant to differentiation (p � .05) compared withthe controls, although not completely refractory (Fig. 3A–3C).This was further confirmed by OCT4 real-time RT-PCR analy-sis (Fig. 3D), which showed less drastic reduction of this plu-ripotency marker in TERT sublines compared with controls(p � .05). EBs at day 10 of differentiation from each sublinewere embedded in paraffin, sectioned, and stained with hema-toxylin and eosin. Although this analysis is not quantitative, wewere able to notice neural rosettes at high frequency in EBsmade from TERT-overexpressing sublines. Their presence wasmuch more reduced in the control sublines (Fig. 3E). Sinceneural rosettes are thought to be rich in neural progenitors, it istempting to speculate that overexpression of TERT perhapsleads to higher proliferation and maintenance of progenitors butis inhibitory to their terminal differentiation, although furtherexperimental proof is needed to confirm the validity of thishypothesis.
We also tested the hematopoietic activity of day 14 EBsmade from control and TERT sublines and noticed that this wassignificantly reduced as a consequence of TERT overexpression(Fig. 3F; similar reductions were obtained for days 7 and 21 ofdifferentiation), thus corroborating our data on suppression ofdifferentiation. It is of interest to note that during differentiationof control EBs, the majority of morphologically scored coloniesshowed a differentiated phenotype of granulocytic, macrophage,or erythrocyte lineages (Fig. 3G). Twenty percent of the colo-nies were derived from a bipotential progenitor (granulocyte-macrophage [CFU-GM]), and no mixed colonies (granulocyte-erythrocyte-macrophage-megakaryocyte [CFU-GEMM]) wereobserved. However, during the differentiation of TERT-overex-pressing sublines, we observed the presence of mixed colonies(CFU-GEMM, 12.5%), as well as a higher percentage ofCFU-GM (37.5%), which seems to have occurred at the expenseof a reduction in erythrocytic colonies and granulocytic colo-nies. One possible explanation for this is a higher survival of themore primitive hematopoietic progenitors arising in the cultureor suppression of their differentiation to more mature lineages;however, further work is needed to prove this using the induc-ible TERT overexpression-based system.
Overexpression of TERT Results in Changes inHuman ESC Proliferation, Survival, and Cell CycleWe noticed that TERT-overexpressing sublines grew faster inculture; this could be due to either a decrease in apoptosis, anincrease in human ESC proliferation, or both. To distinguishbetween these possibilities, we performed Annexin V staining todetect early (7-aminoactinomycin D [7-AAD]� Annexin V�)and late apoptotic (7-AAD� Annexin V�) cells (Fig. 4A) andBrdU incorporation to detect DNA synthesizing/proliferatingcells (Fig. 4B, 4C). The results showed that overexpression ofTERT did not affect the rate of apoptosis, since percentages ofapoptotic cells were similar to those of control clones under
854 Key Role for TERT in hESC Proliferation and Differentiation
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from
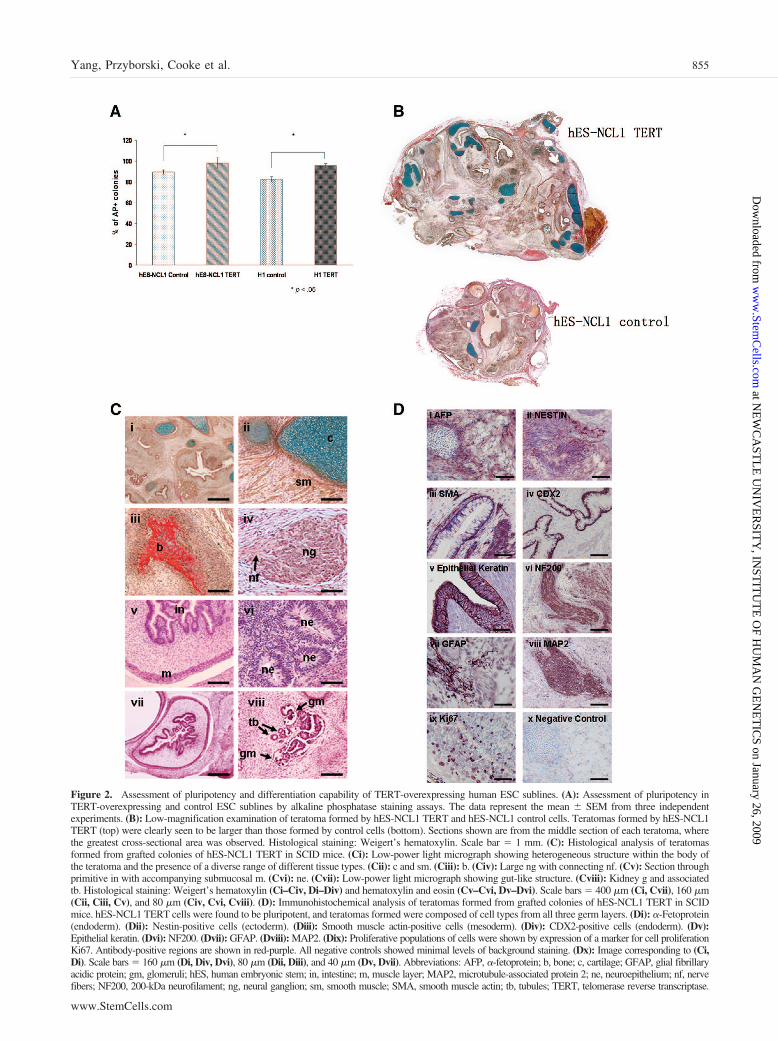
Figure 2. Assessment of pluripotency and differentiation capability of TERT-overexpressing human ESC sublines. (A): Assessment of pluripotency inTERT-overexpressing and control ESC sublines by alkaline phosphatase staining assays. The data represent the mean � SEM from three independentexperiments. (B): Low-magnification examination of teratoma formed by hES-NCL1 TERT and hES-NCL1 control cells. Teratomas formed by hES-NCL1TERT (top) were clearly seen to be larger than those formed by control cells (bottom). Sections shown are from the middle section of each teratoma, wherethe greatest cross-sectional area was observed. Histological staining: Weigert’s hematoxylin. Scale bar � 1 mm. (C): Histological analysis of teratomasformed from grafted colonies of hES-NCL1 TERT in SCID mice. (Ci): Low-power light micrograph showing heterogeneous structure within the body ofthe teratoma and the presence of a diverse range of different tissue types. (Cii): c and sm. (Ciii): b. (Civ): Large ng with connecting nf. (Cv): Section throughprimitive in with accompanying submucosal m. (Cvi): ne. (Cvii): Low-power light micrograph showing gut-like structure. (Cviii): Kidney g and associatedtb. Histological staining: Weigert’s hematoxylin (Ci–Civ, Di–Div) and hematoxylin and eosin (Cv–Cvi, Dv–Dvi). Scale bars � 400 �m (Ci, Cvii), 160 �m(Cii, Ciii, Cv), and 80 �m (Civ, Cvi, Cviii). (D): Immunohistochemical analysis of teratomas formed from grafted colonies of hES-NCL1 TERT in SCIDmice. hES-NCL1 TERT cells were found to be pluripotent, and teratomas formed were composed of cell types from all three germ layers. (Di): �-Fetoprotein(endoderm). (Dii): Nestin-positive cells (ectoderm). (Diii): Smooth muscle actin-positive cells (mesoderm). (Div): CDX2-positive cells (endoderm). (Dv):Epithelial keratin. (Dvi): NF200. (Dvii): GFAP. (Dviii): MAP2. (Dix): Proliferative populations of cells were shown by expression of a marker for cell proliferationKi67. Antibody-positive regions are shown in red-purple. All negative controls showed minimal levels of background staining. (Dx): Image corresponding to (Ci,Di). Scale bars � 160 �m (Di, Div, Dvi), 80 �m (Dii, Diii), and 40 �m (Dv, Dvii). Abbreviations: AFP, �-fetoprotein; b, bone; c, cartilage; GFAP, glial fibrillaryacidic protein; gm, glomeruli; hES, human embryonic stem; in, intestine; m, muscle layer; MAP2, microtubule-associated protein 2; ne, neuroepithelium; nf, nervefibers; NF200, 200-kDa neurofilament; ng, neural ganglion; sm, smooth muscle; SMA, smooth muscle actin; tb, tubules; TERT, telomerase reverse transcriptase.
855Yang, Przyborski, Cooke et al.
www.StemCells.com
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from
normal culture conditions (Fig. 4A). Addition of the nuclearfactor �B inhibitor pyrrolidine dithiocarbamate [31], knownto induce apoptosis, did not result in significant changes incell death between TERT-overexpressing and control sublines(data not shown). However, overexpression of TERT causedsmall but significant increases in the percentage of cellsincorporating BrdU (Fig. 4B, 4C) compared with the controlsublines, suggesting a putative role for TERT in human ESCproliferation.
Human ESC show low survival in culture upon disassocia-tion to single cells [32]. Since telomerase has been often de-scribed as a survival enzyme, we investigated whether overex-pression of TERT conferred such advantages by gentlytrypsinizing TERT-overexpressing and control sublines to sin-gle cells. These were plated on mitotically inactivated feedercells at limited dilution numbers for 10 days. AP staining wascarried out to identify and facilitate the counting of human ESCcolonies. This analysis showed a significant increase in the
Figure 3. Assessment of the differentiation capability of TERT-overexpressing human ESC clones. (A–D): Real-time reverse transcription-polymerase chain reaction analysis for the expression of PAX6, BRACHYURY, CDX2, and OCT4 during the differentiation of TERT-overexpressingand control sublines for 21 days in suspension culture. The data represent the mean � SEM from two independent experiments carried out in eachhuman ESC subline. cDNA taken from day 7 EBs from H9 human ESC was used as positive control (A–C) and normalized to 1, and all values werecalculated with respect to this. In (D), the control sublines at day 0 were normalized to 1, and all other values were calculated with respect to this.Statistical analysis was carried out using two-factor analysis of variance with replication. (E): EBs at day 10 from both H1 TERT and H1 controlsublines were embedded in paraffin, sectioned, and stained with hematoxylin and eosin. Neural rosettes are indicated by white arrows. (F): Totalnumber of hematopoietic colonies obtained after 14 days of differentiation of TERT and control human embryonic stem cell (hESC) sublines. Thedata represent the mean � SEM from two independent experiments carried out in triplicate in each hESC subline. (G): Graphical representation ofcolony types formed during hematopoietic differentiation of TERT and control hESC sublines. The data represent the mean � SEM from twoindependent experiments carried out in triplicate in each hESC subline. Abbreviations: CFC, colony forming cells; CFU-E, colony-formingunit-erythrocyte; CFU-G, colony-forming unit-granulocyte; CFU-GEMM, colony-forming unit-granulocyte, erythrocyte, macrophage, megakaryo-cyte; CFU-GM, colony-forming unit-granulocyte macrophage; CFU-M, colony-forming unit-macrophage; GAPDH, glyceraldehyde-3-phosphatedehydrogenase; hES, human embryonic stem; TERT, telomerase reverse transcriptase.
856 Key Role for TERT in hESC Proliferation and Differentiation
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from
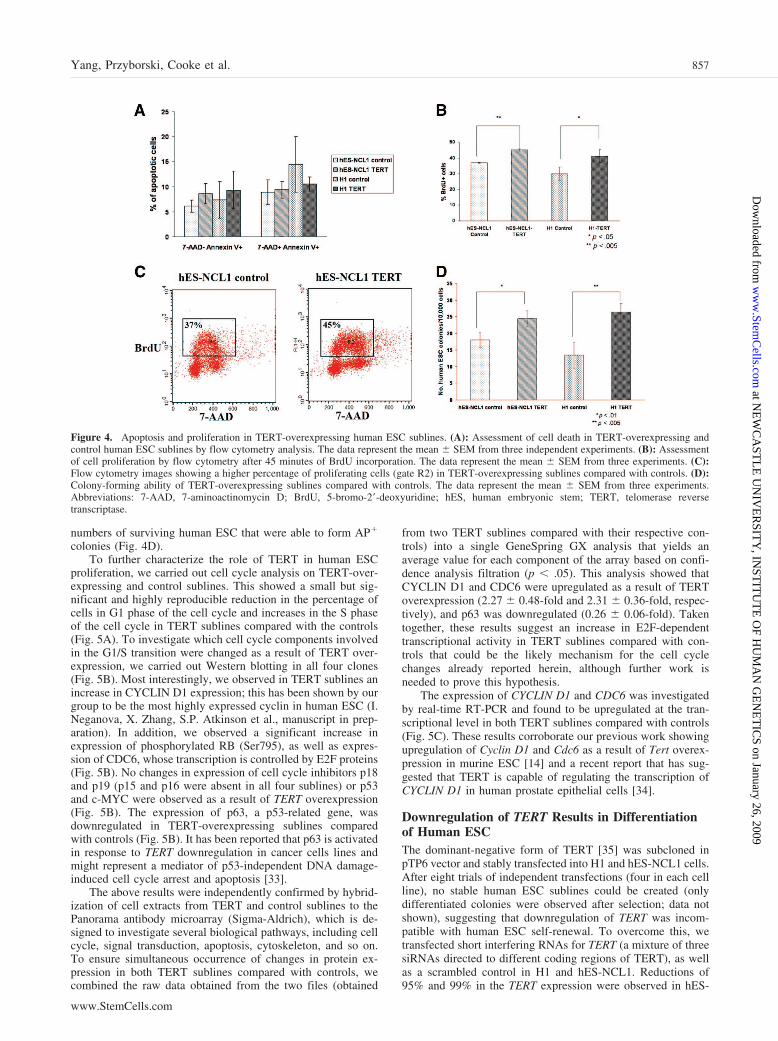
numbers of surviving human ESC that were able to form AP�
colonies (Fig. 4D).To further characterize the role of TERT in human ESC
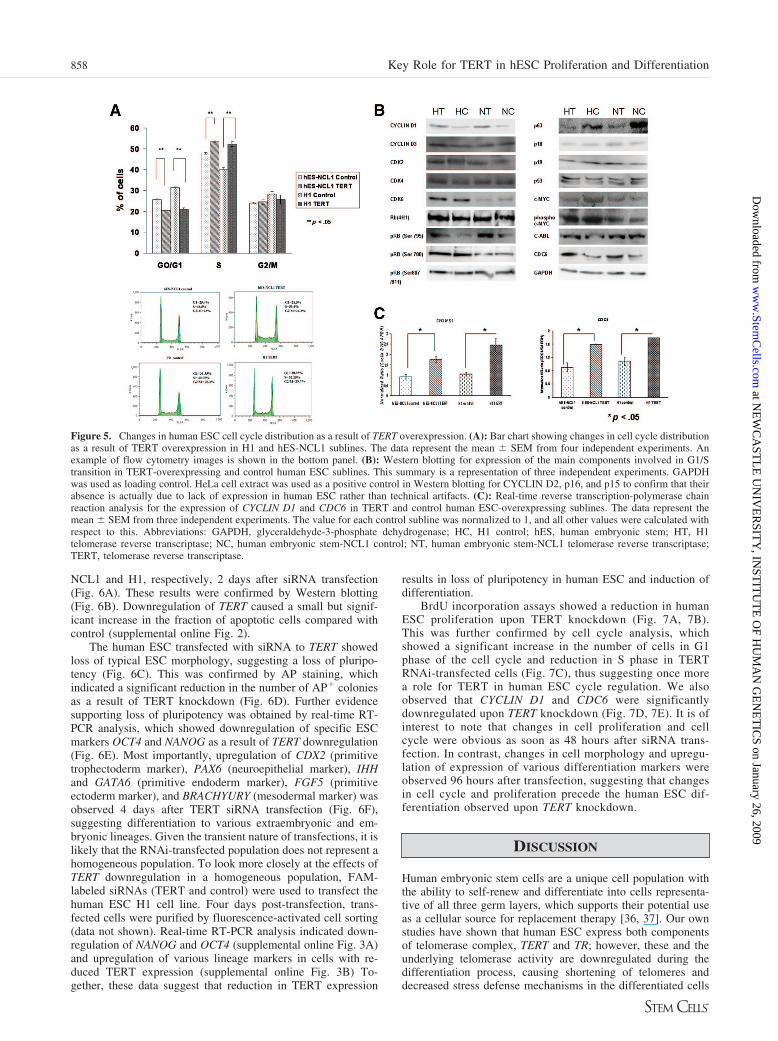
proliferation, we carried out cell cycle analysis on TERT-over-expressing and control sublines. This showed a small but sig-nificant and highly reproducible reduction in the percentage ofcells in G1 phase of the cell cycle and increases in the S phaseof the cell cycle in TERT sublines compared with the controls(Fig. 5A). To investigate which cell cycle components involvedin the G1/S transition were changed as a result of TERT over-expression, we carried out Western blotting in all four clones(Fig. 5B). Most interestingly, we observed in TERT sublines anincrease in CYCLIN D1 expression; this has been shown by ourgroup to be the most highly expressed cyclin in human ESC (I.Neganova, X. Zhang, S.P. Atkinson et al., manuscript in prep-aration). In addition, we observed a significant increase inexpression of phosphorylated RB (Ser795), as well as expres-sion of CDC6, whose transcription is controlled by E2F proteins(Fig. 5B). No changes in expression of cell cycle inhibitors p18and p19 (p15 and p16 were absent in all four sublines) or p53and c-MYC were observed as a result of TERT overexpression(Fig. 5B). The expression of p63, a p53-related gene, wasdownregulated in TERT-overexpressing sublines comparedwith controls (Fig. 5B). It has been reported that p63 is activatedin response to TERT downregulation in cancer cells lines andmight represent a mediator of p53-independent DNA damage-induced cell cycle arrest and apoptosis [33].
The above results were independently confirmed by hybrid-ization of cell extracts from TERT and control sublines to thePanorama antibody microarray (Sigma-Aldrich), which is de-signed to investigate several biological pathways, including cellcycle, signal transduction, apoptosis, cytoskeleton, and so on.To ensure simultaneous occurrence of changes in protein ex-pression in both TERT sublines compared with controls, wecombined the raw data obtained from the two files (obtained
from two TERT sublines compared with their respective con-trols) into a single GeneSpring GX analysis that yields anaverage value for each component of the array based on confi-dence analysis filtration (p � .05). This analysis showed thatCYCLIN D1 and CDC6 were upregulated as a result of TERToverexpression (2.27 � 0.48-fold and 2.31 � 0.36-fold, respec-tively), and p63 was downregulated (0.26 � 0.06-fold). Takentogether, these results suggest an increase in E2F-dependenttranscriptional activity in TERT sublines compared with con-trols that could be the likely mechanism for the cell cyclechanges already reported herein, although further work isneeded to prove this hypothesis.
The expression of CYCLIN D1 and CDC6 was investigatedby real-time RT-PCR and found to be upregulated at the tran-scriptional level in both TERT sublines compared with controls(Fig. 5C). These results corroborate our previous work showingupregulation of Cyclin D1 and Cdc6 as a result of Tert overex-pression in murine ESC [14] and a recent report that has sug-gested that TERT is capable of regulating the transcription ofCYCLIN D1 in human prostate epithelial cells [34].
Downregulation of TERT Results in Differentiationof Human ESCThe dominant-negative form of TERT [35] was subcloned inpTP6 vector and stably transfected into H1 and hES-NCL1 cells.After eight trials of independent transfections (four in each cellline), no stable human ESC sublines could be created (onlydifferentiated colonies were observed after selection; data notshown), suggesting that downregulation of TERT was incom-patible with human ESC self-renewal. To overcome this, wetransfected short interfering RNAs for TERT (a mixture of threesiRNAs directed to different coding regions of TERT), as wellas a scrambled control in H1 and hES-NCL1. Reductions of95% and 99% in the TERT expression were observed in hES-
Figure 4. Apoptosis and proliferation in TERT-overexpressing human ESC sublines. (A): Assessment of cell death in TERT-overexpressing andcontrol human ESC sublines by flow cytometry analysis. The data represent the mean � SEM from three independent experiments. (B): Assessmentof cell proliferation by flow cytometry after 45 minutes of BrdU incorporation. The data represent the mean � SEM from three experiments. (C):Flow cytometry images showing a higher percentage of proliferating cells (gate R2) in TERT-overexpressing sublines compared with controls. (D):Colony-forming ability of TERT-overexpressing sublines compared with controls. The data represent the mean � SEM from three experiments.Abbreviations: 7-AAD, 7-aminoactinomycin D; BrdU, 5-bromo-2�-deoxyuridine; hES, human embryonic stem; TERT, telomerase reversetranscriptase.
857Yang, Przyborski, Cooke et al.
www.StemCells.com
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from
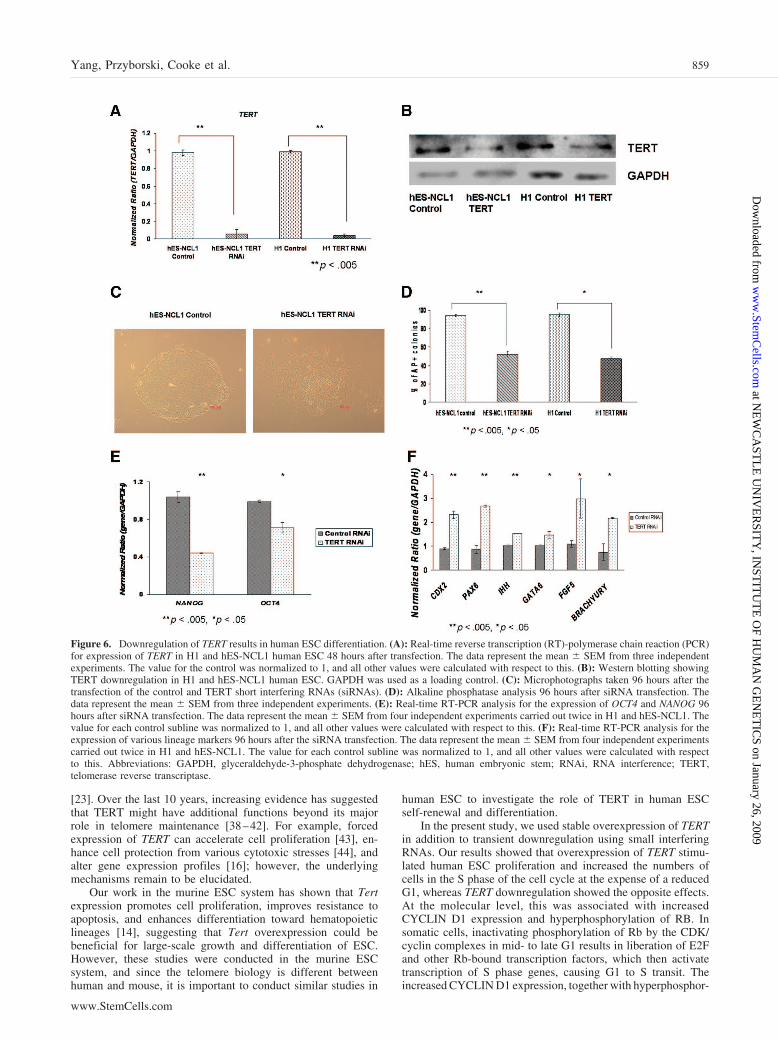
NCL1 and H1, respectively, 2 days after siRNA transfection(Fig. 6A). These results were confirmed by Western blotting(Fig. 6B). Downregulation of TERT caused a small but signif-icant increase in the fraction of apoptotic cells compared withcontrol (supplemental online Fig. 2).
The human ESC transfected with siRNA to TERT showedloss of typical ESC morphology, suggesting a loss of pluripo-tency (Fig. 6C). This was confirmed by AP staining, whichindicated a significant reduction in the number of AP� coloniesas a result of TERT knockdown (Fig. 6D). Further evidencesupporting loss of pluripotency was obtained by real-time RT-PCR analysis, which showed downregulation of specific ESCmarkers OCT4 and NANOG as a result of TERT downregulation(Fig. 6E). Most importantly, upregulation of CDX2 (primitivetrophectoderm marker), PAX6 (neuroepithelial marker), IHHand GATA6 (primitive endoderm marker), FGF5 (primitiveectoderm marker), and BRACHYURY (mesodermal marker) wasobserved 4 days after TERT siRNA transfection (Fig. 6F),suggesting differentiation to various extraembryonic and em-bryonic lineages. Given the transient nature of transfections, it islikely that the RNAi-transfected population does not represent ahomogeneous population. To look more closely at the effects ofTERT downregulation in a homogeneous population, FAM-labeled siRNAs (TERT and control) were used to transfect thehuman ESC H1 cell line. Four days post-transfection, trans-fected cells were purified by fluorescence-activated cell sorting(data not shown). Real-time RT-PCR analysis indicated down-regulation of NANOG and OCT4 (supplemental online Fig. 3A)and upregulation of various lineage markers in cells with re-duced TERT expression (supplemental online Fig. 3B) To-gether, these data suggest that reduction in TERT expression
results in loss of pluripotency in human ESC and induction ofdifferentiation.
BrdU incorporation assays showed a reduction in humanESC proliferation upon TERT knockdown (Fig. 7A, 7B).This was further confirmed by cell cycle analysis, whichshowed a significant increase in the number of cells in G1phase of the cell cycle and reduction in S phase in TERTRNAi-transfected cells (Fig. 7C), thus suggesting once morea role for TERT in human ESC cycle regulation. We alsoobserved that CYCLIN D1 and CDC6 were significantlydownregulated upon TERT knockdown (Fig. 7D, 7E). It is ofinterest to note that changes in cell proliferation and cellcycle were obvious as soon as 48 hours after siRNA trans-fection. In contrast, changes in cell morphology and upregu-lation of expression of various differentiation markers wereobserved 96 hours after transfection, suggesting that changesin cell cycle and proliferation precede the human ESC dif-ferentiation observed upon TERT knockdown.
DISCUSSION
Human embryonic stem cells are a unique cell population withthe ability to self-renew and differentiate into cells representa-tive of all three germ layers, which supports their potential useas a cellular source for replacement therapy [36, 37]. Our ownstudies have shown that human ESC express both componentsof telomerase complex, TERT and TR; however, these and theunderlying telomerase activity are downregulated during thedifferentiation process, causing shortening of telomeres anddecreased stress defense mechanisms in the differentiated cells
Figure 5. Changes in human ESC cell cycle distribution as a result of TERT overexpression. (A): Bar chart showing changes in cell cycle distributionas a result of TERT overexpression in H1 and hES-NCL1 sublines. The data represent the mean � SEM from four independent experiments. Anexample of flow cytometry images is shown in the bottom panel. (B): Western blotting for expression of the main components involved in G1/Stransition in TERT-overexpressing and control human ESC sublines. This summary is a representation of three independent experiments. GAPDHwas used as loading control. HeLa cell extract was used as a positive control in Western blotting for CYCLIN D2, p16, and p15 to confirm that theirabsence is actually due to lack of expression in human ESC rather than technical artifacts. (C): Real-time reverse transcription-polymerase chainreaction analysis for the expression of CYCLIN D1 and CDC6 in TERT and control human ESC-overexpressing sublines. The data represent themean � SEM from three independent experiments. The value for each control subline was normalized to 1, and all other values were calculated withrespect to this. Abbreviations: GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HC, H1 control; hES, human embryonic stem; HT, H1telomerase reverse transcriptase; NC, human embryonic stem-NCL1 control; NT, human embryonic stem-NCL1 telomerase reverse transcriptase;TERT, telomerase reverse transcriptase.
858 Key Role for TERT in hESC Proliferation and Differentiation
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from
[23]. Over the last 10 years, increasing evidence has suggestedthat TERT might have additional functions beyond its majorrole in telomere maintenance [38–42]. For example, forcedexpression of TERT can accelerate cell proliferation [43], en-hance cell protection from various cytotoxic stresses [44], andalter gene expression profiles [16]; however, the underlyingmechanisms remain to be elucidated.
Our work in the murine ESC system has shown that Tertexpression promotes cell proliferation, improves resistance toapoptosis, and enhances differentiation toward hematopoieticlineages [14], suggesting that Tert overexpression could bebeneficial for large-scale growth and differentiation of ESC.However, these studies were conducted in the murine ESCsystem, and since the telomere biology is different betweenhuman and mouse, it is important to conduct similar studies in
human ESC to investigate the role of TERT in human ESCself-renewal and differentiation.
In the present study, we used stable overexpression of TERTin addition to transient downregulation using small interferingRNAs. Our results showed that overexpression of TERT stimu-lated human ESC proliferation and increased the numbers ofcells in the S phase of the cell cycle at the expense of a reducedG1, whereas TERT downregulation showed the opposite effects.At the molecular level, this was associated with increasedCYCLIN D1 expression and hyperphosphorylation of RB. Insomatic cells, inactivating phosphorylation of Rb by the CDK/cyclin complexes in mid- to late G1 results in liberation of E2Fand other Rb-bound transcription factors, which then activatetranscription of S phase genes, causing G1 to S transit. Theincreased CYCLIN D1 expression, together with hyperphosphor-
Figure 6. Downregulation of TERT results in human ESC differentiation. (A): Real-time reverse transcription (RT)-polymerase chain reaction (PCR)for expression of TERT in H1 and hES-NCL1 human ESC 48 hours after transfection. The data represent the mean � SEM from three independentexperiments. The value for the control was normalized to 1, and all other values were calculated with respect to this. (B): Western blotting showingTERT downregulation in H1 and hES-NCL1 human ESC. GAPDH was used as a loading control. (C): Microphotographs taken 96 hours after thetransfection of the control and TERT short interfering RNAs (siRNAs). (D): Alkaline phosphatase analysis 96 hours after siRNA transfection. Thedata represent the mean � SEM from three independent experiments. (E): Real-time RT-PCR analysis for the expression of OCT4 and NANOG 96hours after siRNA transfection. The data represent the mean � SEM from four independent experiments carried out twice in H1 and hES-NCL1. Thevalue for each control subline was normalized to 1, and all other values were calculated with respect to this. (F): Real-time RT-PCR analysis for theexpression of various lineage markers 96 hours after the siRNA transfection. The data represent the mean � SEM from four independent experimentscarried out twice in H1 and hES-NCL1. The value for each control subline was normalized to 1, and all other values were calculated with respectto this. Abbreviations: GAPDH, glyceraldehyde-3-phosphate dehydrogenase; hES, human embryonic stem; RNAi, RNA interference; TERT,telomerase reverse transcriptase.
859Yang, Przyborski, Cooke et al.
www.StemCells.com
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from
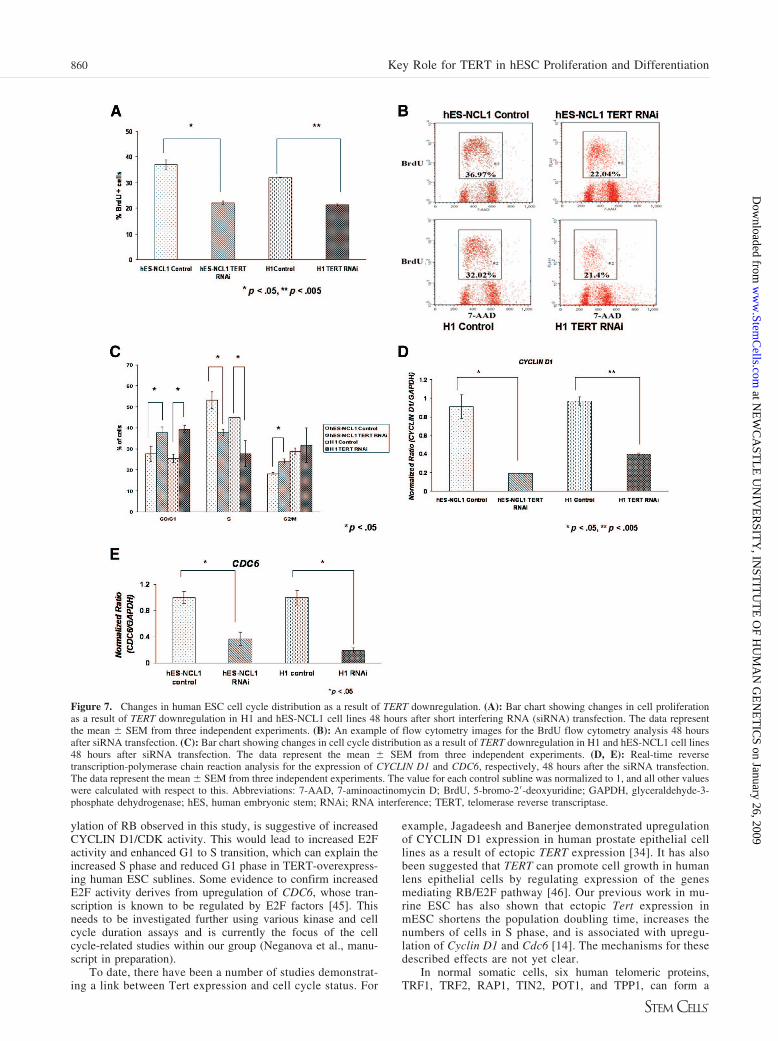
ylation of RB observed in this study, is suggestive of increasedCYCLIN D1/CDK activity. This would lead to increased E2Factivity and enhanced G1 to S transition, which can explain theincreased S phase and reduced G1 phase in TERT-overexpress-ing human ESC sublines. Some evidence to confirm increasedE2F activity derives from upregulation of CDC6, whose tran-scription is known to be regulated by E2F factors [45]. Thisneeds to be investigated further using various kinase and cellcycle duration assays and is currently the focus of the cellcycle-related studies within our group (Neganova et al., manu-script in preparation).
To date, there have been a number of studies demonstrat-ing a link between Tert expression and cell cycle status. For
example, Jagadeesh and Banerjee demonstrated upregulationof CYCLIN D1 expression in human prostate epithelial celllines as a result of ectopic TERT expression [34]. It has alsobeen suggested that TERT can promote cell growth in humanlens epithelial cells by regulating expression of the genesmediating RB/E2F pathway [46]. Our previous work in mu-rine ESC has also shown that ectopic Tert expression inmESC shortens the population doubling time, increases thenumbers of cells in S phase, and is associated with upregu-lation of Cyclin D1 and Cdc6 [14]. The mechanisms for thesedescribed effects are not yet clear.
In normal somatic cells, six human telomeric proteins,TRF1, TRF2, RAP1, TIN2, POT1, and TPP1, can form a
Figure 7. Changes in human ESC cell cycle distribution as a result of TERT downregulation. (A): Bar chart showing changes in cell proliferationas a result of TERT downregulation in H1 and hES-NCL1 cell lines 48 hours after short interfering RNA (siRNA) transfection. The data representthe mean � SEM from three independent experiments. (B): An example of flow cytometry images for the BrdU flow cytometry analysis 48 hoursafter siRNA transfection. (C): Bar chart showing changes in cell cycle distribution as a result of TERT downregulation in H1 and hES-NCL1 cell lines48 hours after siRNA transfection. The data represent the mean � SEM from three independent experiments. (D, E): Real-time reversetranscription-polymerase chain reaction analysis for the expression of CYCLIN D1 and CDC6, respectively, 48 hours after the siRNA transfection.The data represent the mean � SEM from three independent experiments. The value for each control subline was normalized to 1, and all other valueswere calculated with respect to this. Abbreviations: 7-AAD, 7-aminoactinomycin D; BrdU, 5-bromo-2�-deoxyuridine; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; hES, human embryonic stem; RNAi; RNA interference; TERT, telomerase reverse transcriptase.
860 Key Role for TERT in hESC Proliferation and Differentiation
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from

complex called the telosome/shelterin, which determines thestructure of the telomere ends and controls the generation of Tloops, as well as the synthesis of telomeric DNA by telomerase[47]. In normal mammalian cells, which are usually devoid oftelomerase activity, it has been observed that checkpoint pro-teins are on telomeres at each cell cycle and that telomere endsare transiently exposed. However, one of the components of theshelterin complex, TRF2, binds directly to telomere repeat se-quences and inhibits ATM protein kinase, presumably to atten-uate checkpoint signaling at telomeres [48]. The TPP1 compo-nent of shelterin can physically interact with telomerase andstimulate its activity and processivity [49, 50], whereas POT1can inhibit telomerase activity when bound to the telomeric 3�end [51], thus suggesting that a fine balance is likely to existbetween shelterin and telomerase to control the checkpointsignaling and cell cycle progression [52]. Overexpression ofTERT is likely to disturb this balance, thus altering cell cycledynamics. This is an attractive hypothesis; however, it assumesthe presence of shelterin complex and checkpoint signaling inactively dividing cells, such as human ESC, and both of theseissues are unknown and remain to be investigated in this celltype. In addition, the ATM checkpoint signaling that is regu-lated by shelterin is mostly active at the G2/M phase of the cellcycle. Our results do suggest a faster transit of human ESC fromthe G1 to S phase of the cell cycle as a result of TERToverexpression, thus pointing to additional mechanisms bywhich TERT regulates cell cycle progression in these cells.
To investigate whether TERT plays a direct role in thetranscriptional control of CYCLIN D1, we carried out chromatinimmunoprecipitation assays followed by real-time PCR to quan-tify any enrichment of CYCLIN D1 promoter regions bound toTERT. We did not identify any positive enrichment that wouldsuggest that TERT does not bind to the CYCLIN D1 promoter(data not shown). Direct binding of CYCLIN D1 by TERT itselfmay be possible but does not preclude any indirect mechanismsby which TERT and CYCLIN D1 expression might be inter-linked. It has been reported that Cyclin D1 mRNA translation inmurine ESC is mediated via the PI3K/p70 S6 kinase pathway,whereas its degradation is mediated by the PI3K/AKT/GSK3�pathway [53]. It is interesting to note that upon cytokine stim-ulation, human TERT forms physical complexes with AKT,p70S6 kinase, HSP90, and mTOR in human natural killer cells,and TERT activity is post-translationally regulated by the AKT/HSP90/p70SK6 pathway [54]. If this is true in human ESC,ectopic expression of TERT is likely to affect the amount ofavailable AKT in the cell and the phosphorylation of its sub-strate GSK3�, thus changing the degradation rate of CYCLIND1. However, this needs to be further investigated by usinginhibitors of p70SK6, such as rapamycin or PI3K/AKT(LY294002), in conjunction with modulation of TERT expres-sion and quantitation of CYCLIN D1 levels.
It is interesting to note that in both human and murine ESC,Tert is significantly downregulated during the differentiationprocess. Our previous work in murine ESC has suggested thatdownregulation of Tert by overexpression of one of its repres-sors, Zap3, is sufficient to slow down cell growth, reducetelomerase activity and telomere length, and cause changes incell cycle [22]. We have shown in this study that TERT knock-down by RNA interference did result in human ESC differen-tiation along all tested extraembryonic and embryonic lineages,suggesting that reduction of telomerase activity was incompat-ible with human ESC self-renewal. This was further reinforcedby stable overexpression studies, which showed that TERT-overexpressing human ESC sublines were resistant but notcompletely refractory to in vitro differentiation. Furthermore,these human ESC sublines showed lower spontaneous differen-tiation in culture, as well as higher colony-forming ability, when
disassociated into single cells. The underlying mechanism forthe impacts of TERT on the maintenance of human ESC pluri-potency, self-renewal, and differentiation are not clear. Emerg-ing proof suggests that TERT modulation is associated withsignificant changes in the transcriptome [14, 55, 56]. Therefore,one might be able to speculate that TERT may have direct orindirect involvement with the gene expression network thatunderlies the regulation of human ESC pluripotency and self-renewal; however, no transcriptional activity has ever beenlinked with TERT per se. A more attractive hypothesis would bethat upregulation of TERT enhances the maintenance of pluri-potent phenotype by shortening of G1 and enhancement of Sphase of the cell cycle. Clearly, there are examples in theliterature to suggest that lengthening of G1 phase in neuroepi-thelial cells does induce premature differentiation [57]. P19murine embryonic carcinoma cells are particularly vulnerable todifferentiation effects of retinoic acid in G1 phase and refractoryto RA in S phase [58], suggesting that a short G1 phase and longS phase probably protect pluripotent cells from differentiation[59]. Our own work has also shown that activation of cell cycleinhibitors, such as p21 in human ESC, can cause accumulationof cells in G1 phase, resulting in human ESC differentiation (T.Maimets, I. Neganova, L. Armstrong, et al., manuscript submit-ted for publication). On the basis of these findings, we are moreinclined to suggest that TERT has an impact on human ESCpluripotency via the cell cycle changes. Recent data show thattelomere uncapping occurring in progenitor cells of TERT�/�
mice does result in apoptosis in either late S phase or in G2 viaa signaling cascade involving p53, thus suggesting intrinsiclinks between telomerase, telomere replication and capping, andS phase progression [60]. Our work has shown only a smallincrease in apoptosis upon TERT downregulation, suggestingthat human ESC might respond differently to downregulation ofTERT. It is likely that downregulation of TERT results inlengthening of G1 phase, in which human ESC are subjected todifferentiation signals, thus removing them from the replicativeESC pool. Indeed, a similar mechanism has been reported tooperate in murine ESC by p53 in cases of DNA damage [61].Uncapped telomeres resulting from lack of telomerase can alsobe recognized as DNA damage [62]. In this context, it ispossible that a p53-dependent signaling mechanism can result inESC differentiation, as reported by our study. Although nochanges in p53 expression were observed upon upregulation ofTERT, this does not exclude a role for p53-mediated signalingupon TERT downregulation and needs to be investigated fur-ther.
The positive impacts of TERT expression on cell prolifer-ation [14, 63], higher protection to oxidative stress [14, 30], andtelomere length maintenance have led to the suggestion thatoverexpression of TERT can be beneficial for ex vivo expansionof ESC-derived cells and somatic stem cells. This has shownpromising results in mesenchymal and neural progenitor cells,where ectopic TERT has led to extended expansion of these celltypes in culture without impairing their differentiation potential[17, 64]. Our study has shown that overexpression of TERT doesmaintain telomere length and reduces the levels of intracellularperoxides accumulated in the cells during the differentiationprocess. Although TERT overexpression did not cause grosskaryotype abnormalities, in vitro differentiation was impaired.This leads us to suggest that inducible overexpression of TERTusing small molecule activators would have better practicalimpacts. This could be carried out once the differentiated celltype has been produced and needs to be ex vivo expanded or atthe ESC stage if large numbers are required prior to differenti-ation.
In conclusion, our study has shown for the first time a keyrole for TERT in human ESC proliferation, cell cycle regulation,
861Yang, Przyborski, Cooke et al.
www.StemCells.com
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from

maintenance of pluripotent phenotype, and differentiation ca-pacity. Our ongoing studies are focused on investigation of theunderlying mechanisms of TERT actions on human ESC cycleregulation and differentiation. In addition, our investigationsextend to distinguishing between the telomeric and extratelo-meric effects of TERT on human ESC.
ACKNOWLEDGMENTS
We thank Owen Hughes for help with the reading of antibodyarrays, Moira Crosier for help with EB embedding and staining,
Robert Weinberg for the kind gift of the dominant-negativeform of TERT cDNA, Jerome Evans for doing the karyotypicanalysis, and Sun Yung and Dennis Kirk for technical assis-tance. This study was supported by BBSRC grant BBS/B/14779.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Greider CW, Blackburn EH. Identification of a specific telomere termi-nal transferase activity in Tetrahymena extracts. Cell 1985;43:405–413.
2 Morin GB. The human telomere terminal transferase enzyme is a ribo-nucleoprotein that synthesizes TTAGGG repeats. Cell 1989;59:521–529.
3 Lingner J, Cech TR. Telomerase and chromosome end maintenance.Curr Opin Genet Dev 1998;8:226–232.
4 Blackburn EH. Switching and signalling at the telomere. Cell 2001;106:661–673.
5 Stewart SA, Ben-Porath I, Carey VJ et al. Erosion of the telomericsingle-strand overhang at replicative senescence. Nat Genet 2003;33:492–496.
6 Masutomi K, Yu EY, Khurts S et al. Telomerase maintains telomerestructure in normal human cells. Cell 2003;114:241–253.
7 Zhu H, Fu W, Mattson MP. The catalytic subunit of telomerase protectsneurons against amyloid beta-peptide-induced apoptosis. J Neurochem2000;75:117–124.
8 Fu W, Begley JG, Killen MW et al. Anti-apoptotic role of telomerase inpheochromocytoma cells. J Biol Chem 1999;274:7264–7271.
9 Cao Y, Li H, Deb S et al. TERT regulates cell survival independent oftelomerase enzymatic activity. Oncogene 2002;21:3130–3138.
10 Kushner DM, Paranjape JM, Bandyopadhyay B et al. 2–5A antisensedirected against telomerase RNA produces apoptosis in ovarian cancercells. Gynecol Oncol 2000;76:183–192.
11 Fu W, Killen M, Culmsee C et al. The catalytic subunit of telomerase isexpressed in developing brain neurons and serves a cell survival-pro-moting function. J Mol Neurosci 2000;14:3–15.
12 Gorbunova V, Seluanov A, Pereira-Smith OM. Expression of humantelomerase (hTERT) does not prevent stress-induced senescence in nor-mal human fibroblasts but protects the cells from stress-induced apopto-sis and necrosis. J Biol Chem 2002;277:38540–38549.
13 Luiten RM, Pene J, Yssel H et al. Ectopic hTERT expression extends thelife span of human CD4� helper and regulatory T-cell clones andconfers resistance to oxidative stress-induced apoptosis. Blood 2003;101:4512–4519.
14 Armstrong L, Saretzki G, Peters H et al. Overexpression of telomer-ase confers growth advantage, stress resistance, and enhanced differ-entiation of ESCs toward the hematopoietic lineage. STEM CELLS2005;23:516 –529.
15 Lee MK, Hande MP, Sabapathy K. Ectopic mTERT expression in mouseembryonic stem cells does not affect differentiation but confers resis-tance to differentiation- and stress-induced p53-dependent apoptosis.J Cell Sci 2005;118:819–829.
16 Geserick C, Tejera A, Gonzalez-Suarez E et al. Expression of mTertin primary murine cells links the growth-promoting effects of telom-erase to transforming growth factor-beta signalling. Oncogene 2006;25:4310 – 4319.
17 Simonsen JL, Rosada C, Serakinci N et al. Telomerase expressionextends the proliferative life-span and maintains the osteogenic potentialof human bone marrow stromal cells. Nat Biotechnol 2002;20:592–596.
18 Flores I, Cayuela ML, Blasco MA. Effects of telomerase and telomerelength on epidermal stem cell behavior. Science 2005;309:1253–1256.
19 Sarin KY, Cheung P, Gilison D et al. Conditional telomerase induc-tion causes proliferation of hair follicle stem cells. Nature 2005;436:1048 –1052.
20 Niida H, Matsumoto T, Satoh H et al. Severe growth defect in mousecells lacking the telomerase RNA component. Nat Genet 1998;19:203–206.
21 Liu Y, Snow BE, Hande MP et al. The telomerase reverse transcriptaseis limiting and necessary for telomerase function in vivo. Curr Biol2000;10:1459–1462.
22 Armstrong L, Lako M, von Herpe I et al. Reduction of telomeraseactivity during differentiation of embryonal stem cells is mediated by
Zap3 transcriptional repression of murine telomerase reverse transcrip-tase (mTert). Mech Dev 2004;121:1509–1522.
23 Saretzki G, Walter T, Atkinson S et al. Downregulation of multiple stressdefense mechanisms during differentiation of human embryonic stemcells. STEM CELLS 2008;26:455–464.
24 Cerezo A, Stark HJ, Moshir S et al. Constitutive overexpression ofhuman telomerase reverse transcriptase but not c-myc blocks terminaldifferentiation in human HaCaT skin keratinocytes. J Invest Dermatol2003;121:110–119.
25 Richardson RM, Nguyen B, Holt SE et al. Ectopic telomerase expressioninhibits neuronal differentiation of NT2 neural progenitor cells. NeurosciLett 2007;421:168–172.
26 Cui W, Wylie D, Aslam S et al. Telomerase-immortalized sheep fibro-blasts can be reprogrammed by nuclear transfer to undergo early devel-opment. Biol Reprod 2003;69:15–21.
27 Thomas M, Yang L, Hornsby PJ. Formation of functional tissue fromtransplanted adrenocortical cells expressing telomerase reverse transcrip-tase. Nat Biotechnol 2000;18:39–42.
28 Wynn RF, Cross MA, Hatton C et al. Accelerated telomere shortening inyoung recipients of allogeneic bone-marrow transplants. Lancet 1998;351:178–181.
29 Stojkovic M, Lako M, Stojkovic P et al. Derivation of human embryonicstem cells from day-8 blastocysts recovered after three-step in vitroculture. STEM CELLS 2004;22:790–797.
30 Saretzki G, Armstrong L, Leake A et al. Stress defense in murineembryonic stem cells is superior to that of various differentiated murinecells. STEM CELLS 2004;22:962–971.
31 Armstrong L, Hughes O, Yung S et al. The role of PI3K/AKT, MAPK/ERK and NFkappabeta signalling in the maintenance of human embry-onic stem cell pluripotency and viability highlighted by transcriptionalprofiling and functional analysis. Hum Mol Genet 2006;15:1894–1913.
32 Watanabe K, Ueno M, Kamiya D et al. A ROCK inhibitor permitssurvival of dissociated human embryonic stem cells. Nat Biotechnol2007;25:681–686.
33 Shammas MA, Koley H, Batchu RB et al. Telomerase inhibition bysiRNA causes senescence and apoptosis in Barrett’s adenocarcinomacells: Mechanism and therapeutic potential. Mol Cancer 2005;4:24.
34 Jagadeesh S, Banerjee PP. Telomerase reverse transcriptase regulates theexpression of a key cell cycle regulator, cyclin D1. Biochem Biophys ResCommun 2006;347:774–780.
35 Hahn WC, Stewart SA, Brooks MW et al. Inhibition of telomerase limitsthe growth of human cancer cells. Nat Med 1999;5:1164–1170.
36 Carpenter MK, Rosler E, Rao MS. Characterization and differentiation ofhuman embryonic stem cells. Cloning Stem Cells 2003;5:79–88.
37 Stojkovic M, Lako M, Strachan T et al. Derivation, growth andapplications of human embryonic stem cells. Reproduction 2004;128:259 –267.
38 Magnenat L, Tobler H, Muller F. Developmentally regulated telomeraseactivity is correlated with chromosomal healing during chromatin dimi-nution in Ascaris suum. Mol Cell Biol 1999;19:3457–3465.
39 Li H, Cao Y, Berndt MC et al. Molecular interactions between telomer-ase and the tumor suppressor protein p53 in vitro. Oncogene 1999;18:6785–6794.
40 Artandi SE, Alson S, Tietze MK et al. Constitutive telomerase expressionpromotes mammary carcinomas in aging mice. Proc Natl Acad SciU S A 2002;99:8191–8196.
41 Gonzalez-Suarez E, Samper E, Ramirez A et al. Increased epidermaltumors and increased skin wound healing in transgenic mice overex-pressing the catalytic subunit of telomerase, mTERT, in basal keratino-cytes. EMBO J 2001;20:2619–2630.
42 Stewart SA, Hahn WC, O’Connor BF et al. Telomerase contributes totumorigenesis by a telomere length-independent mechanism. Proc NatlAcad Sci U S A 2002;99:12606–12611.
43 Smith LL, Coller HA, Roberts JM. Telomerase modulates expression of
862 Key Role for TERT in hESC Proliferation and Differentiation
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from

growth-controlling genes and enhances cell proliferation. Nat Cell Biol2003;5:474–479.
44 Zhang P, Chan SL, Fu W et al. TERT suppresses apoptosis at a premi-tochondrial step by a mechanism requiring reverse transcriptase activityand 14–3-3 protein-binding ability. FASEB J 2003;17:767–769.
45 de Jager SM, Menges M, Bauer UM et al. Arabidopsis E2F1 binds asequence present in the promoter of S-phase-regulated gene AtCDC6 andis a member of a multigene family with differential activities. Plant MolBiol 2001;47:555–568.
46 Xiang H, Wang J, Mao Y et al. Human telomerase accelerates growth oflens epithelial cells through regulation of the genes mediating RB/E2Fpathway. Oncogene 2002;21:3784–3791.
47 de Lange T. Shelterin: The protein complex that shapes and safeguardshuman telomeres. Genes Dev 2005;19:2100–2110.
48 Karlseder J, Hoke K, Mirzoeva OK et al. The telomeric protein TRF2binds the ATM kinase and can inhibit the ATM-dependent DNA damageresponse. PLoS Biol 2004;2:E240.
49 Wang F, Podell ER, Zaug AJ et al. The POT1-TPP1 telomere complexis a telomerase processivity factor. Nature 2007;445:506–510.
50 Xin H, Liu D, Wan M et al. TPP1 is a homologue of ciliate TEBP-beta andinteracts with POT1 to recruit telomerase. Nature 2007;445:559–562.
51 Lei M, Podell ER, Cech TR. Structure of human POT1 bound totelomeric single-stranded DNA provides a model for chromosome end-protection. Nat Struct Mol Biol 2004;11:1223–1229.
52 Cristofari G, Sikora K, Lingner J. Telomerase unplugged. ACS ChemBiol 2007;2:155–158.
53 Jirmanova L, Afanassieff M, Gobert-Gosse S et al. Differential contri-butions of ERK and PI3-kinase to the regulation of cyclin D1 expressionand to the control of the G1/S transition in mouse embryonic stem cells.Oncogene 2002;21:5515–5528.
54 Kawauchi K, Ihjima K, Yamada O. IL-2 increases human telomerasereverse transcriptase activity transcriptionally and posttranslationallythrough phosphatidylinositol 3�-kinase/Akt, heat shock protein 90, and
mammalian target of rapamycin in transformed NK cells. J Immunol2005;174:5261–5269.
55 Li S, Crothers J, Ha CM et al. Cellular and gene expression responsesinvolved in the rapid growth inhibition of human cancer cells by RNAinterference-mediated depletion of telomerase RNA. J Biol Chem 2005;280:23709–23717.
56 Lai SR, Cunningham AP, Huynh VQ et al. Evidence of extra-telomericeffects of hTERT and its regulation involving a feedback loop. Exp CellRes 2007;313:322–330.
57 Calegari F, Haubensak W, Haffner C et al. Selective lengthening of thecell cycle in the neurogenic subpopulation of neural progenitor cellsduring mouse brain development. J Neurosci 2005;25:6533–6538.
58 Mummery CL, van den Brink CE, de Laat SW. Commitment to differ-entiation induced by retinoic acid in P19 embryonal carcinoma cells iscell cycle dependent. Dev Biol 1987;121:10–19.
59 Lukaszewicz A, Savatier P, Cortay V et al. G1 phase regulation, area-specific cell cycle control, and cytoarchitectonics in the primate cortex.Neuron 2005;47:353–364.
60 Rajaraman S, Choi J, Cheung P et al. Telomere uncapping in progenitorcells with critical telomere shortening is coupled to S-phase progressionin vivo. Proc Natl Acad Sci U S A 2007;104:17747–17752.
61 Lin T, Chao C, Saito S et al. p53 induces differentiation of mouseembryonic stem cells by suppressing Nanog expression. Nat Cell Biol2005;7:165–171.
62 Takai H, Smogorzewska A, de Lange T. DNA damage foci at dysfunc-tional telomeres. Curr Biol 2003;13:1549–1556.
63 Bodnar AG, Ouellette M, Frolkis M et al. Extension of life-span byintroduction of telomerase into normal human cells. Science 1998;279:349–352.
64 Roy NS, Nakano T, Keyoung HM et al. Telomerase immortalization ofneuronally restricted progenitor cells derived from the human fetal spinalcord. Nat Biotechnol 2004;22:297–305.
See www.StemCells.com for supplemental material available online.
863Yang, Przyborski, Cooke et al.
www.StemCells.com
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from

DOI: 10.1634/stemcells.2007-0677 2008;26;850-863; originally published online Jan 17, 2008; Stem Cells
Majlinda Lako George Anyfantis, Stuart P. Atkinson, Gabriele Saretzki, Lyle Armstrong and
Chunbo Yang, Stefan Przyborski, Michael J. Cooke, Xin Zhang, Rebecca Stewart, Differentiation
Embryonic Stem Cell Proliferation, Cell Cycle Dynamics, and In Vitro A Key Role for Telomerase Reverse Transcriptase Unit in Modulating Human
This information is current as of January 26, 2009
& ServicesUpdated Information
http://www.StemCells.com/cgi/content/full/26/4/850including high-resolution figures, can be found at:
Supplementary Material http://www.StemCells.com/cgi/content/full/2007-0677/DC1
Supplementary material can be found at:
at NE
WC
AST
LE
UN
IVE
RSIT
Y, IN
STIT
UT
E O
F HU
MA
N G
EN
ET
ICS on January 26, 2009
ww
w.Stem
Cells.com
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















