
A phylogenetic analysis of the genus Leuconostoc based on reverse transcriptase sequencing of 16 S...
11
FEMS Microbiology Letters 70 (1990) 73-84 73 Publishedby Elsevier FEMSLE 04046 A phylogenetic analysis of the genus Leuconostoc based on reverse transcriptase sequencing of 16 S rRNA A.J. Martinez-Murcia and M.D. Collins Department of Microbiology,AFRC Institute of Food Research, Reading Laboratory, Shinfiel~ Reading, U.K. Received24 January 1990 Accepted 12 March 1990 Key words: Leuconostoc; Lactobacillus; Taxonomy; Phylogeny; rRNA 1. SUMMARY The phylogenetic interrelationships of members of the genus Leuconostoc and some heterofermen- tative lactobaciUi, which phenotypically resemble leuconostocs, were investigated by comparative analysis of their 16 S rRNA sequences. The six species, Leuconostoc mesenteroides, Leu. carno- sum, Leu. citreum, Leu. gelidum, Leu. lactis and Leu. pseudomesenteroides exhibited a high degree of sequence similarity with each other and formed a phylogenetically coherent group, quite separate from all other lactic acid bacteria investigated. The species Leg paramesenteroides was found to be phylogenetically distinct from the Leu. mesen- teroides group of species and formed a natural grouping with the heterofermentative lactobacilli, Lb. confusus, Lb. kandleri, Lb. minor and Lb. viridescens. The rRNA sequence of the acidophilic species, Leg oenos, displayed exceptionally low levels of homology with all of the other taxa examined. The 16 S sequence of Leu. oenos showed Correspondence to: Dr M.D. Collins,AFRC Institute of Food Research, ReadingLaboratory, Shinfield,Reading,RG2 9AT, U.K. major nucleotide differences in relatively highly conserved positions of the molecule indicating this species is phylogenetically distinct and warrants a separate genus. 2. INTRODUCTION The genus Leuconostoc comprises a collection of catalase-negative, gas-forming, non-motile spherical (but often lenticular) shaped bacteria [1]. The intra- and inter-generic relationships of Leuconostoc species have always been equivocal. Leuconostocs can be readily separated from streptococci by their ability to produce gas from glucose. The relationship of leuconostocs to cer- tain gas-producing heterofementative lactobacilli (e.g. Lactobacillus confuses, Lb. viridescens), has, however, long been controversial. The separation of leuconostocs from lactobacilli is based prim- arily on differences in morphology (leuconostocs are more coccoid than rodlike, and lactobacilli are more rodlike than coccoid). It is now generally recognized, however, that morphology is a poor indicator of relatedness and that the morphology of leuconostocs and many heterofermentative lactobacilli overlap [2]. 0378-1097/90/$03.50 © 1990Federationof EuropeanMicrobiological Societies
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of A phylogenetic analysis of the genus Leuconostoc based on reverse transcriptase sequencing of 16 S...

FEMS Microbiology Letters 70 (1990) 73-84 73 Published by Elsevier
FEMSLE 04046
A phylogenetic analysis of the genus Leuconostoc based on reverse transcriptase sequencing of 16 S rRNA
A.J. Mar t inez-Murc ia and M.D. Coll ins
Department of Microbiology, AFRC Institute of Food Research, Reading Laboratory, Shinfiel~ Reading, U.K.
Received 24 January 1990 Accepted 12 March 1990
Key words: Leuconostoc; Lactobacillus; Taxonomy; Phylogeny; rRNA
1. SUMMARY
The phylogenetic interrelationships of members of the genus Leuconostoc and some heterofermen- tative lactobaciUi, which phenotypically resemble leuconostocs, were investigated by comparative analysis of their 16 S rRNA sequences. The six species, Leuconostoc mesenteroides, Leu. carno- sum, Leu. citreum, Leu. gelidum, Leu. lactis and Leu. pseudomesenteroides exhibited a high degree of sequence similarity with each other and formed a phylogenetically coherent group, quite separate from all other lactic acid bacteria investigated. The species Leg paramesenteroides was found to be phylogenetically distinct from the Leu. mesen- teroides group of species and formed a natural grouping with the heterofermentative lactobacilli, Lb. confusus, Lb. kandleri, Lb. minor and Lb. viridescens. The rRNA sequence of the acidophilic species, Leg oenos, displayed exceptionally low levels of homology with all of the other taxa examined. The 16 S sequence of Leu. oenos showed
Correspondence to: Dr M.D. Collins, AFRC Institute of Food Research, Reading Laboratory, Shinfield, Reading, RG2 9AT, U.K.
major nucleotide differences in relatively highly conserved positions of the molecule indicating this species is phylogenetically distinct and warrants a separate genus.
2. INTRODUCTION
The genus Leuconostoc comprises a collection of catalase-negative, gas-forming, non-motile spherical (but often lenticular) shaped bacteria [1]. The intra- and inter-generic relationships of Leuconostoc species have always been equivocal. Leuconostocs can be readily separated from streptococci by their ability to produce gas from glucose. The relationship of leuconostocs to cer- tain gas-producing heterofementative lactobacilli (e.g. Lactobacillus confuses, Lb. viridescens), has, however, long been controversial. The separation of leuconostocs from lactobacilli is based prim- arily on differences in morphology (leuconostocs are more coccoid than rodlike, and lactobacilli are more rodlike than coccoid). It is now generally recognized, however, that morphology is a poor indicator of relatedness and that the morphology of leuconostocs and many heterofermentative lactobacilli overlap [2].
0378-1097/90/$03.50 © 1990 Federation of European Microbiological Societies
DNA-rRNA hybridization studies by Garvie [3] have indicated that the non-acidophilic species Leu. mesenteroides (and subspecies), Le~ lactis and Le~ paramesenteroides form a natural group distinct from Lb. confusus and Lb. viridescens. These studies also demonstrated Le~ oenos was unrelated to other leuconostocs and hetero- fermentative lactobacilli [3]. More recent DNA- rRNA hybridization studies by Schillinger et al. [4] have confirmed the distinctiveness of Le~ oenos. It was also shown that Leu. parame- senteroides was more closely related to Lb. con- fusus and Lb. viridescens rather than to other non-acidophilic leuconostocs [4]. Full sequencing of I6 S rRNA using reverse transcriptase is now being used increasingly as an alternative to DNA- rRNA hybridizations and oligonucleotide cata- loguing for elucidating the natural interrelation- ships of microorganisms [5,6]. Complete sequence determination produces more precise data than hybridization and cataloguing procedures and provides an unequivocal means for t~et~;mL-fing phylogenetic interrelationships [7]. Therefore, in the present study the 16 S rRNA primary struc- tures of members of the genus Leuconoswc (in. chiding the newly described species Le~ carno. sum, Le~ citreum, Le~ gelidum and Le~ pseudo-
mesenteroides) and phenotypically similar hetero- fermentative lactobacilli have been determined in an attempt to clarify their inter- and intra-generic relationships.
3. MATERIALS A N D M E T H O D S
3.1. Cultures and cultivation Details of the test strains examined are listed in
Tables 1 and 2. Leuconostoc oenos was grown in Acid Tomato Broth (glucose, 10 g; peptone, 10 g; yeast extract, 5 g; MgSO4" 7H20, 0.2 g; MnSO 4 • 4H20, 0.05 g; tomato juice, 100 ml; H20 , 1 litre; pH 4.8) at 22°C. LactobaciUi and the lenconos- toes were cultivated in MRS broth (Oxoid) at 25 ° C. Cultures were harvested in late exponential phase by centrifugation and washed in deiouized water.
3.2. Extraction and sequence determination of RNA Total cellular rRNA was extracted from ca. 2 g
of wet cells [8]. Nucleotide sequences were de- termined by the didenxynuclentide method [9] using AMV reverse transcriptase. The sequences of oli8onucleotide primers and their 16 S rRNA
Table 1
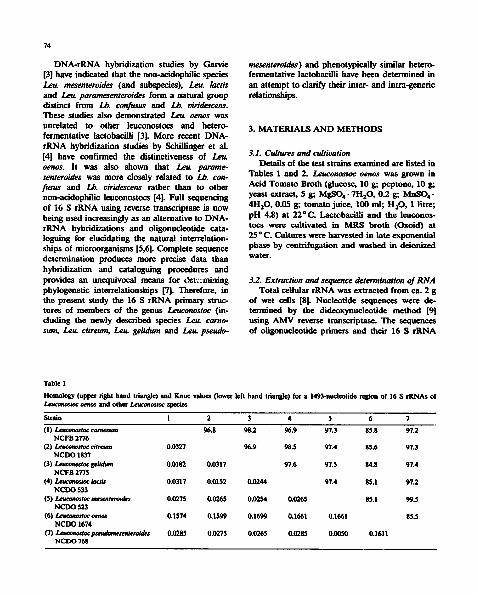
Homology (upper right hand triangle ) and Knuc values (lower left hand triani0e) for a 1493-nucleotide region of 16 S rRNAs of Leuconoszoc oenos and other Leuconoszac species
Strain 1 2 3 4 5 6 7
(1) Leuconoswc camosum 96,8 98.2 96.9 97.3 85.8 97.2 NCFB 2776
(2) Leucom~loc citreum 0.0327 96.9 98.5 97.4 85.6 97.3 NCDO 1837
(3) I,x.uconostoc gelidum 0,0182 0.0317 97.6 97.5 84.8 97.4 NCFB 2775
(4) Leucom~stoc/act/s 0,0317 0.0152 0.0244 97.4 85.1 97.2 NCDO 533
( 5 ) / . . , ~ Z o c mesemero~des 0.0275 0,0265 0.0254 0.0265 85.1 99.5 NCDO 523
(6)/,eut'onosWc otmos 0,1574 0,1599 0.1699 0.1661 0.1661 85.5 NCDO 1674
(7) lx'uconostocpset~m~semeroides 0.0285 0.0275 0.0265 0.0285 0.0050 0.1611 NCDO 768

target sites were as described by Lane et al. [6] and Embley et al. [8] except for a D N A primer of sequence 5" TCAGTCTCTCAACTCGGCTA,
75
which was employed for the ca. 300 region (com- plimentary to positions 287 to 306; Escherichia coil).
Leu. c~. Leu. cx. N~:~:
N~:N: Leu. ce. Leu.c~. N~:~:
N~:N: Leu.c~. LeU.C~. Leu. e .
~ : ~ : Leu . ce . Leu. cx. ~ :~:
u . l ~ .
Leu. cO. Leu.c].. l~u . qe. Leu . l a . I,eu.me. heu.oe. N~:N:
u. c~. u .c~ . u.ge. u.~a. u.me. o.o~.
~:~: Leu.ce. Leu. cx. Leu. ae . Leu. l a .
LeUoCe. Leu.l~a. Leu.~s. L~u. c~. Imu.c~. ~u.qe.
u . I e . u .me . u . oe .
Leu. ce. LeU. cx . N~:~:
N~:N: Leu.co. Leu.cx. Leu. . L e u . ~ .
l~u . .
7 GAC~UCCUGGCUCAGGAUNAACGCUGGCGGCGUC-CCUh~U '~CNUGCNAGUCGAACCACA~~C~ GAGAGUUUGaUCCUGGCUCc~ . . . -CAUNAACGCUGGCGGCGUGCCUaAUaCaUGCaAGUCGAA~~CuuUc~
~UCCIF-,GCIF..AC-C~UI~ACGCUGGCC-GCGUC.~CURAU~CAUGCAAGUCGAAC . . UA UCU GAG~GUUuC~~GGAUNAACGCUNGCGGCGUgCClmAUACaUGCRAGUCGAACCACAGCGRAA uUCRAGU
C ~ G O G G A U N A C C U G C C U C A A ~ U aACa UUUGGAAACa GAIR~CONAUACCGAAIR~@~%ACI/UaGUAU GGGUGAGUuACNCGUGGAUNA~~CAUUIFoGaAAC~GA~. ,CUGAU&CCGaAUGAAACUU~U ~GUGGAUAACCUaCCUCI~GGNNGC-GGAUaACaUUUGGRAACaGAUGCUaAUACCGAAU~AACUUC~U
c~ GGGUGAGUNACAC U~AC~aAGGCU G G G N U a A C A ~ U CCU~t~AA AU
GU~,AGA~AC U N A ~ C N G ~ U G C U A A U GI~ACNACNAAUCA GA~JGGCGRAC~GUAACACGUGGACAACCUGCCUCA~GGCUGGGGAUAACAUUU~G~UGCUAAUAC~~~
CC.~UGAUAC~ARRAGGCGCUACGGCGUCACCU NGAG A U G G A U C C ~ U U A G U U A ~ . ~ / R A A G G C C U A C ~ U G A U C C ~ A U G A U A ~ A R ~ A ~ A C G G C G U C A C C U AGAG AUGGAUCCGCGGUGCAUUAGUU&~I/A~AGGCUoAC~I/C~AU CC-CAUGAUACAACK~UUAAAAGGCC~JACGGCGUCACCU AGAG AUGGAUCCGCGGUGCAUUAGUUA~AAAGGCCRAcCA~C~CA~UGAU C C . ~ A U G A U A C R A A ~ U N C ~ G U C A C C U AGAG AUGGGUCCGC~UUAGUI/AOtn~-~GUGC~UNAAGGCCUACCRAGACRAUGAU CGCAUGACA GGCGUNACCU AGAG AUGGAUCCGCGGUC-CAUUAGI~Ja~AAAGGCCUAC~UGAU CAC&UGUGAUCUNUUUC.~AAGGUCCI/NUUGGAUCG CU AGAGGAUGGUCUUGCGGCGUAUUAGCUNGUUGGUAGGGUAGAAGCCUAC~I&UGAU " GAUGGt~JCUGCUAUCACUAAGAG AUGGACCCGC~UUAGCUaOUUGGURAGGUAACGGCUNACCAA~UGAU CGCAUGACACAAAGU0 GUCACCU AGAG AUGGAUCCGCGOUGCAUUAGWA~DJC~JC~..~-UaAAC-GCCUACCA~G~CAAUGAU
GA CA CUG~ C-C~CUNAACUC UAC GGC UGGGCGaAA
U& UC CA GA CCNaACC~CUACGGGA GAAA
G C A U & G C C C ~ G U 1 / G A G A C . ~ C U N A U C G G C C A C . A ~ ~ C A C G G C C N N A A C U C C U A C ~ ~ & ~ ~ ~ G C G U A C ~ C ~ G A G A ~ C G G C C A ~ C U G A G A C u C U G C C C m U ~ C U C C U A C G G G A G G C U G C ~ U b ' U U C C C - C U A ~ GCAUAGCCGAGUUGAGAGACUNAUCGGCCACt~UGGGACUNAGACACGGCCCNUNCUCCUAC~GC.~~UCU~~ G C A U A G C C G A G U U G A G A G A C U N A U C G G C C A C a ~ C U G A C ~ C a C G G C C U N A A C U C C U A C G G G A G G C ~ a ~ ~ C ~
~CUN AUGOAGCNACGCCGCGUGUGUGAUGRAGGCUUUCGGGUCGUNAAGCNCUGUUGUA~~UA~C~ CUbA CUHAUGGAGCNACGCCGCGU ~ . U G U G A U G A A G G C U t n J C G G G U C G U a A A G C N C U G U U G U A U ~ ~ U A ~ ~
GU UGuc, auc~a A u~aa acu~uu~u&Uc, C~aauaaauu~ua uA uu~u~u u~c,a
b"OQ~CC, C~GC¢;A CC~C C~ GI~"t ~0C&UCAAC~3C UUUCGC~GUC GUA.:bq~ ~ ~ A ~ ~ ~ a ~ ~ ~ /~A GUG~GUGAUGAAG UUUC ~ NCUGUUAUAAGAGAAGRAC CACUGAGAG 7,JCAGUGU GUGA
4 ~ G U ~ C C a U a C C A G A ~ A C - C ~ C G G C U m ~ U A C G U G C C A G C A G C C G C 0 0 " U N A U A C O U A U ~ C C ~ C ~ a ~ ~ CGGUACCAUACCAC'RAAG~.~A~GGCU~AAU&~GUGCCAGCAGCC~CGGU~AUACGUAUGUGCCN~GC~UUAUC~GGAUuuA~~~
, ~ U C ~ C U C C ~ r ~ , A ~ * U ~ C U C ~ U U ~ C U ~ C ~ a ~ C * ~ u ~ ~ U U GOUUGAUUAA~~GCCCGG~GCIF, AACUCCGGNAUGGCAUUGGAAACUGGUU~A AGAC-GU . . . . u
uu UCAA UU U UcAJ~ AGACGGUU~AUUaAGUCUoA~C.~(;CGGAGCUCAACUCCGGAAUGGCa UUGGAAACUGGUUAA _ ~ ( ~ ; U ~ _ G U A C ~ d ~ I } ~ ~ U
GU UAU~ CCAGUGGCGAA U~
CA~tR~C.~&CCC01~UAGOCCaCACCG0.~J~CGa~AC, G U ~ & C ~ C ~ C ~ t F d 0 C ~ C ~ & ~ C ~ CUI~.CGr..~00
c c u r . ~ . ~ ^ c c . . ~ c c c o . . ~ c ~ u ~ c u c ~ c . o ~ u u c ~ c c c ~ c ~ c . c c ~ m ~ c ~ c . c ^ ~ , ~ ~ ~ C~3%]~T, GAGOACC.~C CC, CA.~G~IK~AaC ~ C.-GA~OUC~ C ~ C C G C A CAAGC C~ UGC~GCAU G ~ ~ C ~ ~ C ~
Fig. I. P r i m l y s t a t u r e of the 16 S rRNAs from Leu. c a r m m ~ (Leu. ca.) NCFB2776; Le~ citreum (Leu. ci.) NCDO1837; g e / ~ ( L ~ . I~.) NCFB2"F/5, L e ~ / a c t u (Leu. la.) NCDO533; Le~ nl~enteroides (Leu. me,) NCDO523; Le~ pseudomesenteroides (Leu. ps.) NCDO768; /.,eu. para~esentero~des (Leu. p~ ) NCDOS03 and Lea oenos (Lea. oe.) NCDOI674. The first and last nucleofide in the sequences of ~ mesenteroides are anak~ous to positions 7 and 1483 of the E. coil 16 S rRNA sequence [21]. N,
undetermined nucleodd¢. Spaces in the sequences correspoad to a l i ~ m c m gaps.

983 Leu. C~. ACCAGGUC UUGACNUCCUUUGAAGCUNUUAG~AAUNGAAGUGUUCUC~CGGAGACA~AGUGACAGGUGGNGCAUNGU~C~~G Leu. c~. A~CAGGUCUUGACAUCCUUUGAAGCUuNUAGAGAUAGA~GUQUUCUCUUCGGAGAC~AAGUGACAGGUGGuGCAUGGUCGUC~~G Leu. e. ACCA U UU NU UUU UUNNA Ua UGUU
ACCAGGUCUUGACAUCCtRJUGAAGCUNUUAGAGAUAGAAGUG~RJCU a UNdUe LeU ....... oe. A C C A G G U C U U G A C A U A C C A A U G A U C G C U N N N G U A A U G R A A ~ ~ - A ~ a ~ ~ u ~ ~ ~ Leu. pa, ACCAGGUCUUGACAUCCCUNGCUNAUCCUAGAAAUAGGACGGUU CUUC~CAA GUGAG Leu. ps. ACCAGGUCuUGACAUCCUUUG~AGCUUUUAGAGAUAGAAGUGUUCUC(R~CGGAGACA~AGUC4%CAGGUGGNGC~UNGUC~~~G
AUGUUGGGUUNAGUCCC aA C~% aACCCUUaUUGUUaGUU CCAGCa~U'NaGA aGCGAGACUGC ~euU: ~i ~ : AUGUUGGGUURAG U C C C ~ G A ~ C C C b ~ / A U U G ~ J A ~ C A G C A U U C A G ~ ~ G C G A G A C U G C ~ ~ ~u. ~e : AUGUUGGGUUNAGUCC~GCaACGAGCGCaA~CCUNNDUGUUAGUUGCC~GCaUUNaGA~AGUGAGACUGCCGGUGACNA~~
AUGUUGGGUUAAGUCCCGCAAC GCAACCC(/N/&UUGUUAGUUC-CCA A GACUGCC CAAACCGGAGGAAGG
Leu. oe. AUGUUGGGUUAAGUCCCGCa ACGAC-CGCAACCCUNGUUAUUAGUUGC UUUA AAUGAGACU C URAACCGGA GG Leu. pa. AUGUUGGG UUAAGUCCCGCAACGAGCGCNACCCUUN%R3NDUa G~UGC~UUCA~aGUGA~UU~C~GA~%/~CGGA~ Leu. ps. AUGUUGGGUUAAGUCCCGCAACGAGCGCa&CCCUUAUUGUUAGUUGCCAGCAUUCaGAUGGGCACUCUaGCGAGA~C~C~
Leu. c~. CGGGGACGACGUCAGAUCAUCAUGCCCCUUAUGACC UGGGCUACACACGUGCUACaAUGGCGUAUACaACGA~CAACCUGCGAAGG~RAUC Leu.cl. C~ACGACGUCAGAUCAUCAUGCCCC 'JUAUGACCUNGGCUACACACGUGCUACAAUGGCGUAUACAACGAGUUGCCAACC"JGC~M~ Leu.qe. CGGGGACGACGUCAGAUCAUC/~UGCCCC~ ~UAUGACCUGGGCUACACACGUGCUACaAUGGCGU&UACaACGAGUUGCCAACCUGUGAA~M~ Leu.la. CGGGGACGACGUCAGAUCAUCAUGCCCC~ YUAUGACCUNGGCUACACACGUGCUACRAUGCCGUAUACAACGAGDUC~CCAACCCGCGA~~ Leu .me. CGGGGACGACGUCAGAUcAUCAUGCCCCUuAUGACcUNGGCUACACACGUGCUACAAUGGCGUAUACAAcG~GUUGCC~ACCCGCGA~aA~ Leu. oe. UGGGGACGACGUCa GAUCa UC a UGCCCCUUAUGACCUNGGCNACACACGUGCUACAAUGGC, AAGUACAACGAGUCGCAAACCGGCGACGGURAGCUAAUC
U~AUGACGUCAAAUCAU CCUUAU CCUNGGCUACACACGUGCUACAAU UAU Cad& CGC AAUC Leu.pa. CGGGGACGACGU~AGAUCAU~CCUUAU~CCUGc.C.CUACACACGUC~UACAA~C.~G~G~A~GA~C~.~%~ Leu.ps.
Leu. c¢. UCUUAAAGUACGUCUCAG~UcGGAcUGCAGUcUGCAACUCGACUNCACGAAGUCGGAAUCGCUAGUAAUCGCGGAUCAGCAcGCCGC~u~ Leu. c1. U~UUAAAGUA~GUCUCAGUUCGGACUGCAGUCUGCAACUNGACUGCAGC~AGUCGG~AUCGCUAGUAAUCGC~~C~~ Leu. e. UCDUAAAGUACGUCUCAGUU UC-CAGUCUGCAACUCGACUGCACGAAGUCGGAAUCGCUA UCGC UCAGCACGCCGCGGUGAAUAC I~U. ~a. UCUUAAAGUACGUCUCA~UU~UGCAGUCUGCAACUCGACUGCACGAAGUCGGAAUC~UA~UCGC~UCAC~CAC~C~UAC~ Leu.me. UCU uNAAGUACGUCUCAGUU~GGAUUGUAGUCUGCMCUcGACUNCAUGAAGUCGGRAUCGCU&GUAAUCGCGGAUCA~C~C~UAC~ Leu . oe . UCDUAAAACUNCUcUcAGUUCGGACUGGAGUcUGCa&CUCGACUCCACGAAGuCGGAAUCGCUAGUA~UcGCGAAUcaGCaUGUCC-CGGUGAAUaCG~JC Leu. pa. UCUUAAAGUAUGUC UCAGUUCGGAUUGUAGGCUGCAAC UCGCCUGCAUCAAGUCGGAAUCGCUAGUAAUCGCGGAUC&~C~UAC~ Leu.ps. U ~t~AAAGUACGUCUCAGUUCGGAUUGUAGUCUGCAACUCGACUaCAUGAAGUCGC~.AUCGCUAGUAAUC~&~C~C~UAc~
Leu. c~. CCGGGUCUUGUACaCACCGCCCGUCACACNAUGGAAGWJUGUAAUGCCCAAAGCCGGUGGCCUAACC GUUAGGAGGGAACCGUCUUAGGCAGGACAGAU Leu.cl. C CGGG UC UUGUAC aCACCGCCCGUCACACCAUGGGAGUUUGUMUGCCCAAAGCCGGUGGCCUAAC b~'NGGGAGGGAGC C ~ 3 M ~ C A G A U Leu. e. CCGGGUCUUGUACACACCGCCCGUCACACCAUGAGAGUUUGUMUGCCCAAAGCCGGUGGCCUAACC NNNAGGAGGGRACCGUCUUAGGCAGGACAGAU
C CC4~GUC UUGUACACACCGCCCGUCACACCAUGGGAGUUUGUAAUGCCCAAAGCCGGUGGC uuNU GCCGUCUNA U L~U. ~a. CCGGG UC UUG UACA CACCGCCCGUCACACCAUGGGAGUUUGUAAUGCCCAAAGCCGGU~GCCC~CUL~UA~GGAN~C~~U Leu .me. Leu.oe. CCGGGUNUNGUACaCACCGCCNGUCAAAUCNUGGGAGUCGGAAGUACCCAAAGUCGCUUGGCt~AC ~ AGAGGCCGGUGCCUAAGGUAARAUCC4&U
CCGGGUC UUGUACACAC CGCCCGUCACACCAUGAGAGUUUGUAACACCCAAA GC UGGGG CUUUU GGA~CAGCC GGUGGGACAGA CGGU UAAC A CA CGUCUAA U cc~ ..... ~c~c ....... o ..... c ~ . ~ = ~ . ~ . G = C ~ c ~ c c u . . ^ ~ C ~ ^ G G ~ . Leu.ps.
Fig. ] continued.
The sequences generated were aligned with those of some reference lactic acid bacteria ([10- 13]: Collins and associates, unpublished) and ho- mology values determined using the Beckman Mi- crogenie program [14]. Calculation of evolutionary distance (Knuc) values [15] and generation of a phylogenetic tree [16] by the distance matrix method contained in a program written by Felsen- stein [17] for the IBM PC (Philip version 3.1) followed described methods.
4. RESULTS AND DISCUSSION
The 16 S rRNA sequences of eight species of Leuconostoc are shown in Fig. 1. The reverse tran- scriptase (RT) sequences comprised between 1479 and 1495 nucleotides representing ca. %% of the total primary sequences. The ]euconostoc se- quences were aligned and percentage homologies calculated for a continuous stretch of 1493 bases (ranging from positions 7 and 1483 on the E. coli numbering 3ystem) (Table 1). Homologies with four phenotypicaliy similar heterofermentative
lactobacilli and twenty reference strains repre- senting nine genera ([10-13,18] and Collins and associates, unpublished) were also determined (Table 2). The latter values were based on a smaller number of 1340 nucleotides (ranging from posi- tions 107 to 1433 on the E. coil numbering sys- tem) due to incompleteness of sequence data for some of the reference species. Unrooted phyio- genetic trees constructed from calculated evolu- tionary distance values from the two data sets are shown in Figs. 2 and 3. It is evident from the branching pattern of the trees (Fig. 3) that the Leuconostoc species fall into three distinct groups/lines. The six species Leu, mesenteroides, Leu. carnosum, Leu. citreum, Leu. gelidum, Leu. lactis and Leu. pseudomesenteroides exhibited very high sequence homologies with each other (ca. 97 to 99%) and form a phylogenetically natural group. Within the Leu. mesenteroides cluster (designated Leuconostoc sensu stricto ) considerable intragener- ic structure was evident. Leu. mesenteroides and Leu. pseudomesenteroides were found to be very closely related (5 nucleotide differences in 1483 bases). Similarly Leu. lactis and Leu. citreum
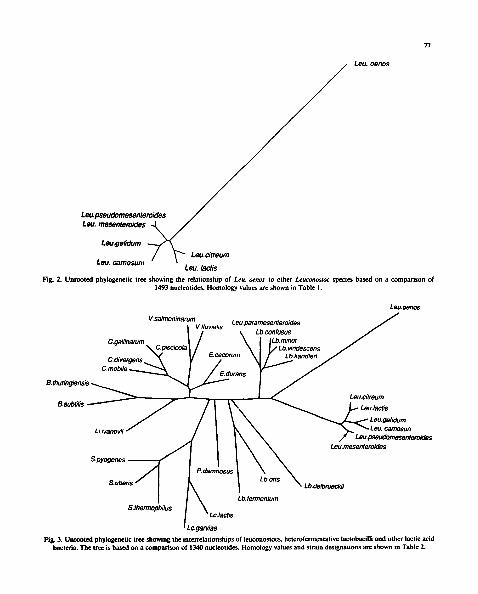
"/7
Lou. oenos
Leu.pseudomesenteroides Leu. rnesenteroides ~ /
Leu.gelidum ~ / ~ ~" Leu.citreum
Leu. carnosum " I Leu. lactis
Fig. 2. Unrooted phylogenetic tree showing the relationship of Leu. oenos to other Leuconostoc species based on a comparison of 1493 nucleotides. Homology values are shown in Table 1.
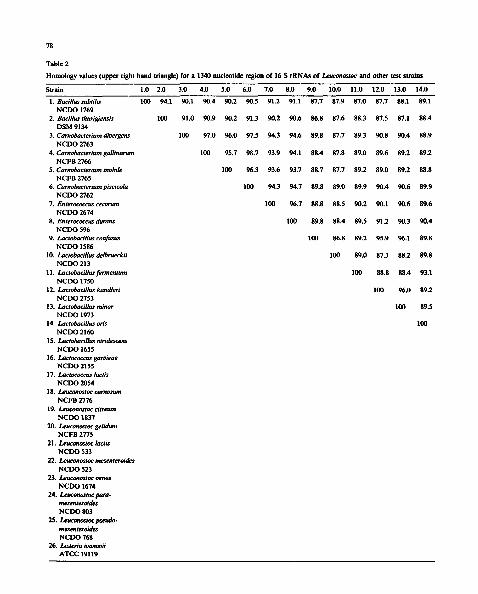
Leu.oenos
Vsalmoninarum . . . . . . Leu.paramesenteroides I v.nuwalls Lbconfusus
C.gallinamm - . . I / ~ I ' | Lb'min°r C.piscicola| i \ I I f Lb.viridescens
C.divergens " % ( I / E.cecorum ~ I [ Lb.kandleri /
/ - I I \ \ J . Leu.lactis • " ~ *" ~ Leu.gelidum
Li ivanovii ~ / I ~ ~ ~ Leu. camosun
/ I | I \ \ ~ou~e.e.,o,o~. ,.,,o~.e~ ,¢" I I 1 \ \
/ I I P.d . . . . . . . | \ N
S.thermophilus I \ Lc.lactis
! Lc.garviae
Fig. 3. Unroofed phylogenetic tree showing the interrelationships of leuconostocs, heterofermentative lactobacilli and other lactic acid bacteria. The tree is based on a comparison of 1340 nucleotides. Homology values and strain designations are shown in Table 2.
78
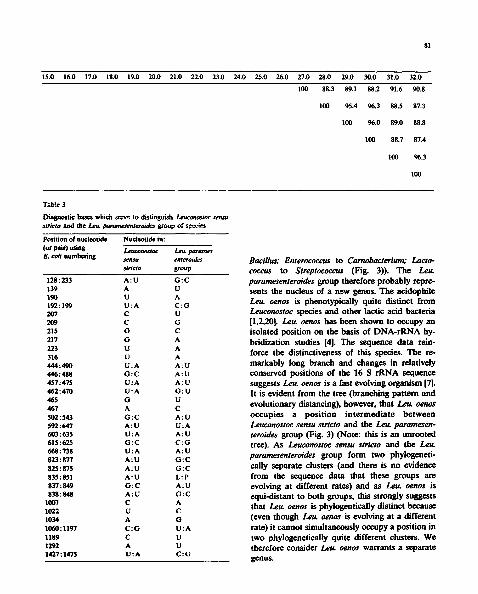
Table 2
Homology values (upper right hand triangle) for a 1340 nucleotide region of 16 S rRNAs of Leuconostoc and other test strains
Strain 1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0 10.0 11.0 12.0 13.0 14.0
1. Bacillus subtilis NCDO 1769
2. Bacillus thurigiensis DSM 9134
3. Carnobacterium divergens NCDO 2763
4. Carnobacterium gallinarum NCFB 2766
5. Carnobacterium mobile NCFB 2765
6. Carnobacterium piscicola NCDO 2762
7. Enterococcus cecorum NCDO 2674
8. Enterococcus durans NCDO 596
9. Lactobacillus confusus NCDO 1586
10. Lactobacillus delbrueckii NCDO 213
11. Lactobacillus fermentum NCDO 1750
12. Lactobacillus kandleri NCDO 2753
13. Lactobacillus minor NCDO 1973
14. Lactobacillus otis NCDO 2160
15. Lactobacillus viridescens NCDO 1655
16. Lactococcus garvieae NCDO 2155
17. Lactococcus lactis NCDO 2054
18. Leuconostoc carnosum NCFB 2776
19. Leuconostoc citreum NCDO 1837
20. Leuconostoc gelidum NCFB 2775
21. Leuconostoc lactis NCDO 533
22. Leuconostoc mesenteroides NCDO 523
23. Leuconostoc oenos NCDO 1674
24. Leuconost~ para. mesenteroides NCDO 803
25. Leuconostoc pseudo. meremeroides NCDO 768
26. 12ateria i ~ i i ATCC 19119
100 94.1 90.1 90.4 90.2 90.5 91.2 91.1 87.7 87.9 87.0 87.7 88.1 89.1
100 91.0 90.9 90.2 91.3 90.2 90.6 86.8 87.6 88.3 87.5 87.1 88.4
100 97.0 96.0 97.5 94.3 94.6 89.8 87.7 89.3 90.8 90.4 88.9
100 95.7 98.7 93.9 94.1 88.4 87.8 89.0 89.6 89.2 89.2
100 96.3 93.6 93.7 88.7 87.7 89.2 89.0 89.2 88.8
100 94.3 94.7 89.8 89.0 89.9 90.4 90.6 89.9
100 96.7 88.8 88.5 90.2 90.1 90.6 89.6
100 89.8 88.4 89.5 91.2 90.3 90.4
100 86.8 89.2 95.9 96.1 89.8
100 89.0 87.3 88.2 89.8
100 88.8 88.4 93.1
100 96.0 89.2
100 89.5
100

15.0 16.0 17.0 18.0 19.0 20.0 21.0 22.0 23.0 24.0 25.0 26.0 27.0 28.0 29.0 30.0 31.0 32.0
87.7 85.2 86.2 83.9 84.8 85.3 84.7 85.6 81.9 87.5 85.4 92.4 89.3 86.7 85.9 86.3 89.7 89.9
87.6 85.8 85.9 85.0 85.6 84.8 85.1 86.2 82.7 86.8 85.8 90.8 89.7 87.3 85.3 87.2 91.0 91.3
90.6 87.4 88.5 85.8 85.9 86.4 85.9 86.5 83.4 89.2 86.6 93.3 91.6 89.0 88.9 89.2 93.4 94.1
89.0 87.7 88.3 85.3 84.7 85.8 85.4 86.2 82.7 88.6 86.2 92.6 91.3 88.6 88.0 88.5 92.3 93.2
89.4 87.6 88.1 84.9 84.7 86.0 84.8 86.5 83.7 88.6 85.9 92.4 91.2 88.0 87.5 87.5 93.4 93.7
89.9 88.4 88.1 85.1 85.0 86.3 85.2 86.2 84.8 90.0 85.9 92.6 91.8 89.3 88.9 89.3 93.4 93.8
90.0 90.2 89.9 86.8 87.2 87.9 88.0 87.4 84.4 89.4 88.6 92.8 91.5 90.0 89.8 90.8 94.5 93.4
90.6 89.5 89.5 86.7 87.5 88.0 87.4 88.2 83.9 90.2 88.3 93.5 92.1 90.2 90.7 90.8 94.6 94.5
96.2 86.9 86.6 90.7 90.8 91.5 90.8 91.5 85.5 96.0 91.5 88.0 "~8.7 88.0 87.4 86.7 87.4 89.0
88.1 86.8 85.7 86.2 87.4 87.7 88.0 88.5 84.8 86.5 87.7 87.2 89.5 85.9 84.4 84.2 88.0 87.4
88.5 87.8 85.8 86.7 87.1 87.6 87.1 88.4 85.5 89.0 88.2 88.7 92.0 86.9 86.0 87.0 89.9 87.0
97.0 86.5 86.9 90.4 90.2 90.6 90.4 90.9 85.9 94.6 90.8 88.6 89.4 88.1 86.6 87.8 89.3 89.3
98.4 86.6 87.1 89.3 90.1 90.2 89.8 90.8 86.3 95.2 90.4 88.7 89.5 87.7 86.3 87.0 88.4 89.4
89.4 87.2 85.1 87.2 88.9 88.2 89.0 89.1 86.3 89.5 88.4 88.1 91.9 86.8 84.9 86.2 90.2 87.5
100 87.1 86.6 89.8 90.8 89.8 90.3 90.7 86.8 95.6 90.6 88.6 89.8 87.7 87.1 88.1 89.2 89.4
100 95.0 84.8 84.3 85.2 83.9 84.5 83.4 86.7 8 4 . 1 87.4 87.9 91.0 89.9 90.0 87.3 87.8
100 83.9 84.5 85.0 84.4 84.7 8 4 . 1 87.7 84.4 87.8 89.6 92.5 91.4 91.7 87.5 88.4
100 97.5 98.3 97.2 97.5 86.7 89.3 97.2 85.3 87.4 84.9 85.2 85.6 85.4 85.2
100 97.1 98.9 97.8 86.8 90.5 97.6 85.9 87.7 85.2 85.6 84.2 85.2 83.9
100 97.5 97.6 85.9 90.4 97.4 85.3 88.0 85.9 86.0 86.0 85.9 84.7
100 97.8 86.5 90.9 97.3 85.9 87.7 85.9 85.5 84.6 86.3 84.4
100 86.0 90.1 99.6 87.0 87.4 86.8 85.5 86.0 87.3 86.2
100 86.2 86.8 84.4 85.7 84.5 84.0 84.2 82.5 82.7
100 90.3 87.6 89.5 87.1 86.8 87.3 88.2 87.8
100 87.2 87.3 86.8 85.9 85.6 87.1 85.9
100 89.9 87.5 88.1 88.7 91.8 91.4

80
Table 2 (continued)
Strain 1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0 10.0 11.0 12.0 13.0 14.0
27. Pediococcus damnosus NCDO 1832
28. Streptococcus pyogenos NCDO 2381
29. Streptococcus thermophilus NCDO 573
30. Streptococcus uberis NCDO 643
31. Vagococcus fluvialis NCDO 2497
32. Vagococcus salmoninarum NCFB 2777
formed a subgroup, as did the two recently de- scribed species, Let~ carnosum and Le~ gelidum [19] from meat.
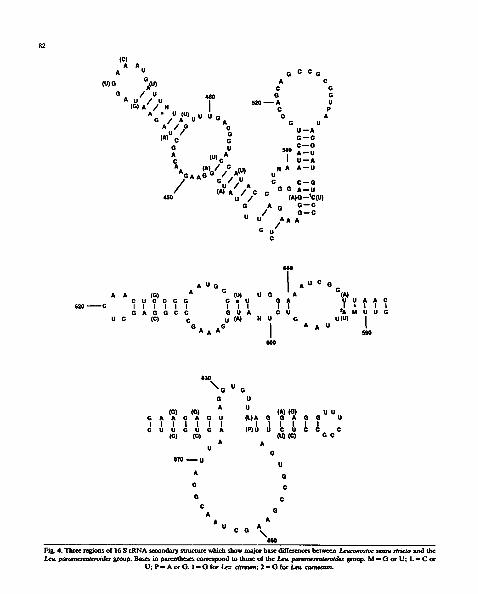
Le~ paramesenteroides was found to be phylo- genetically quite separate from Leuconostoc sensu stricto but showed high sequence relatedness with the four heterofermentative lactobacilhis species Lb. confusus, Lb. kandleri, Lb. minor and Lb. viridescens (Table 2). Within the Leu. paramesen- teroides cluster specific relationships between Lb. confusus and Leu. paramesenteroides and Lb. minor and Lb. viridescens were apparent (Fig. 3). The Le~ paramesenteroides cluster exhibited rela- tively high intragroup sequence relatedness (ca. 95 to 96%). Significantly lower sequence homologies were shown with members of Leuconostoc sensu stricto (ca. 89 to 91~) (Table 2). An analysis of the sequence al ignments demonstra ted that Leuconostoc sensu stricto and the Le~ paramesen- teroides group could be readily distinguished from each other. Diagnostic bases, which serv© to dis- tinguish these two groups from each other, are shown in Table 3. The positions of the most important differences maintaining secondary structure base pairing are shown in Fig. 4.
The acidophilic species Le~ oenos was found to be phylogenetically distinct from Leuconostoc sensu stricto and the Le~ paramesenteroides clus- ter exhibiting low degrees of sequence homology (ca. 86%) with both groups. Similarly low degrees of sequence relatedness were also found with all of
the other test strains examined (Table 2). Leu. oenos possessed an extraordinarily long stalk, the branch length (evolutionary distance) of which was approximately equidistant to the Leuconostoc sensu stricto and Le~ paramesenteroides clusters (Fig. 3). An inspection of the RT data revealed that sequence of Leu. oenos was most unusual and differed from those of Leuconostoc sensu stricto, Le~ paramesenteroides group and all of the other taxa examined in a large number (ca. 30) of rela- tively conserved positions.
The results of the pzesent study clearly demon- strate that the genus Leuconostoc, as presently defined [1], is phylogenetically heterogeneous. The RT sequence data show that the genus Leuconos- toc should be restricted to Le~ mesenteroides (type species) and closely related species (viz: Le~ carnosum, Leu. citreum, Leu. gelidum, Le~ lactis, Le~ pseudomesenteroides ). The data further dem- onstrate Le~ paramesenteroides is phylogeneti- tally mole closely related to the heterofermenta- five lactobacilfi, Lb. confusus, Lb. kandleri, Lb. minor and Lb. viridescens than Leuconostoc sensu stricto. These findings are in accordance with the recent DNA-rRNA hybridization studies of Schil- linger et al. [4]. In terms of sequence relatedness (homologies and signatures) and the branching pattern of the tree l , euconostoc sensu stricto and the Left paramesenteroides group are as distinct from each other as are several other phylogeneti- cally separate taxa (for example Staphylococcus to
15.0 16.0 17.0 18.0 19.0 20.0 21.0 22.0 23.0 24.0 25.0 26.0 27.0 28.0 29.0 30.0 31.0 32.0
I00 88.3 89.1 88.2 91.6 90.8
100 95.4 96.3 88.5 87.3
100 96.0 89.0 88.8
100 88.7 87.4
100 96.3
100
Table 3
Diagnostic bases which s~. e to cfistingtnsh Leuconostoc sensu $tricto and the Lelt paranws~'tteroides group of ~'~'ci._'es
Position of nudeotide (or pair) using E. co//numbc~n 8
Nucleotide in:
Lt'uconoszoc Le~ parames enteroides $¢nxu
stricto Stoup
128:233 A:U G:C 139 A U 190 U A 192:199 U:A C:G 2O7 C U 2O9 C G 215 G C 217 G A 223 U A 316 U A 444:490 U :A A :U 446:488 G:C A:U 457:475 U : A A : U 462:470 U : A G :U 465 G U 46"/ A C 502:543 G:C A:U $92:647 A:U U:A 603:635 U:A A:U 615:625 G:C C:G 668:738 U:A A:U 823:877 A:U G:C 825:875 A:U G:C 835:851 A:U L:P 837:849 G:C A:U 838:848 A:U G:C
1007 C A 1022 U C 1034 A G 1060:1197 C:G U:A 1189 C U 1292 A U 1427:1475 U:A C:G
Bacillus; Enterococcus to Carnobacterimn; Lacto- coccus to Streptococcus (Fig. 3)). The Leu. paramesenteroides group therefore probably repre- sents the nucleus of a new genus. The acidophile Le~ oenos is phenotypically quite distinct from Leuconostoc species and other lactic acid bacteria [1,2,20]. Le~ oenos has been shown to occupy an isolated position on the basis of DNA-rRNA hy- bridization studies [4]. The sequence data rein- force the distinctiveness of this species. The re- markably long branch and changes in relatively conserved positions of the 16 S rRNA sequence suggests Le~ oenos is a fast evolving organism [7]. It is evident from the tree (branching pattern and evolutionary distancing), however, tha t / . zu , oenos
occupies a posi t ion in te rmedia te be tween Leuconostoc sensu stricto and the Leu. paramesen- teroides group (Fig. 3) (Note: this is an unrooted tree). As Leuconostoc sensu stricto and the L e ~
paramesenteroides group form two phylogeneti- cally separate clusters (and there is no evidence from the sequence data that these groups are evolving at different rates) and as Le~ oenos is equi-distant to both groups, this strongly suggests that Le~ oenos is phylogentically distinct because (even though Le~ oenos is evolving at a different rate) it cannot simultaneously occupy a position in two phylogenetically quite different clusters. We therefore consider Leu. oenos warrants a separate genus.
'~ A A U G C C G
(U) G GA(U ) C A C G
G U / U 480 G G u
Aoi,°,*:'°° Oo o" (A)U/C / O U - - A
G G- -C C--G
G U 510 A- -U A (u)cA i u - - A
N A A- -U CA" (')~/ / "~ g c--o A A G G O / / u
(A) C G G A - - U A
4SO A U I / C (A)G--~C(U)
U U A A
G u / c
620 - - C
64O
A U C G
A A U G
A A (cG) c G G A G U G
G A G G C C G U A C U U C (¢) C U (A) N U G
G A AG i A A U A
G(~
U(~ [
,5@O
830
~ G U G G U A U
(c) 1¢1 (u) 1¢1 G C A A u
G 870 ~ U
U A G O O
G C
c A G
A A U A
¢ G \
m
Fig. 4. Three regions of 16 S rRNA secondary structure which show major base differences between l, euconostoc sensu stricto and the paramesenteroides group. Bases in parenthes~ ~ n d to those of the L, eu. paramesenteroides group. M = (3 or U; L = C or
U; P = A o r G. 1 = ( } for L~.,. citresm,; 2 - G for Le~ ¢~Tm~m.

R E F E R E N C E S
[I] Oarvie, E.l. (1986) In Bargey's Manual of Systematic Bacteriology, Vol. 2 (Sneath, P.H.A., Mair, N.S., Sharpe, M.E. and Holt, J.O., eds.), pp. 1071-1075, Williams and Wilkins, Baltimore.
[2] Oarvie, E.I. (1984) Methods. Microbiol. 16,147-178. [3] Oande~ E,I. (1981) J. Oan. Microbiol. 127, 209-212. [4] Schillinger, U., Holzapfel, W. and Kandler, O. (1989)
Syst. AppL Microbiol. 12, 48-55. [5] Qu, L.H., Michot, B. and BacheUerie, J.P. (1983) Nucleic
Aeids Res. 11, 5903-5920. [6] lane, D.J., Pace, B., Olsen, G.J., Staid, D.A., Sogin, M.L.
and Pace, N.IL (1985) Proc. Natl. Acad. Sci., U.S.A. 82, 6955-6969.
[7] Woese~ C.IL (1987) Mier6biol. Rev. 51, 221-271. [8] Embley, T.M., Smida, J. and Stackebrandt, E. (1988) J.
Gen. Microbiol. 134, 961-966. [9] Sanger, F., Nicklen, S. and Coulson, A.R. (1977) Proc.
Natl. Acad" Sci. U.S.A. 74, 5463-5467. [10] Williams, A.M., Farrow, J.A.E. and Collins, M.D. (1989)
Left. Appl. Microbiol. 8,185-189. [11] Collins, M.D., Ash, C., Farrow, J.A.E., Wallbanks, S. and
Williams, A.M. (1989) J. Appl. Bacteriol. 67, 453-460.
83
[12] Walibmdc.s, S., Martinez-Mureia, A.J., Fryer, J.L., Phillips, B.A. and Collins, M.D. (1989) Int. J. Syst. Bacteriol. In Press.
[13] Williams, A.M. and Collins, M.D. (1989) J. Appl. Bacteriol. In Press.
[14] Queen, C. and Kom, LJ. (1984) Nucleic Acids Res. 12, 581-599.
[15] Hori, H. and Osawa, S. (1979) Proc. Natl. Acad. Sci., U.S.A. 76, 381-385.
[16] Fitch, W.M. and Margoliash, E. (1967) Science 155, 279- 284.
[17] Felsenstein, J. (1982) Q. Rev. Biol. 57, 379-404. [18] Green, CJ., Stewart, G.C., Hollis, M.A., Void, B.S. and
Bott, K. (1985) Gene 37, 261-266. [19] Shaw, B.G. and Harding, C.D. (1989) InL J. Syst. Bacteriol.
39, 217-223. [20] Priest, F.G. and Barbour, E.A. (1985) In Computer-as-
sisted Bacterial Systematics. Special publication of the Society for General Microbiology (Goodfeilow, M., Jones, D. and Priest, F.D. eds.), pp 138-163, Academic Press, London.
[21] Brosius, J., Palmer, J.L., Kennedy, J.P. and Noller, H.F. (1978) Proc. Natl. Acad. Sci. U.S.A. 75, 4801-4805.














![5 H-pyrrolo[1,2- b][1,2,5]benzothiadiazepines (PBTDs): A novel class of non-nucleoside reverse transcriptase inhibitors](https://static.fdokumen.com/doc/165x107/63144331fc260b71020f7b4c/5-h-pyrrolo12-b125benzothiadiazepines-pbtds-a-novel-class-of-non-nucleoside.jpg)

![Synthesis, biological evaluation, and binding mode of novel 1-[2-(diarylmethoxy) ethyl]-2-methyl-5-nitroimidazoles targeted at the HIV-1 reverse transcriptase](https://static.fdokumen.com/doc/165x107/6335adc702a8c1a4ec01e00d/synthesis-biological-evaluation-and-binding-mode-of-novel-1-2-diarylmethoxy.jpg)



