Triplet State Phosphorescence in Tris(8-hydroxyquinoline) Aluminum Light Emitting Diode Materials

Report
A Conserved Proline Triplet i
n Val-tRNA Synthetase andthe Origin of Elongation Factor PGraphical Abstract
Highlights
The only conserved polyproline stretch is present in ValS
The proline triplet in ValS is critical for tRNAVal charging and edit-
ing activities
Mutations within the proline triplet of ValS reduce growth and
viability of E. coli
The invariant proline triplet in ValS may explain the coevolution
of EF-P
Starosta et al., 2014, Cell Reports 9, 1–8October 23, 2014 ª2014 The Authorshttp://dx.doi.org/10.1016/j.celrep.2014.09.008
Authors
Agata L. Starosta, Jurgen Lassak, ..., Kirs-
ten Jung, Daniel N. Wilson
In Brief
Elongation factor P is required for ribo-
somes to synthesize polyproline-contain-
ing proteins. Here, Starosta et al. reveal
that only one polyproline-containing pro-
tein is invariant throughout the three do-
mains of life, namely the essential valine
tRNA synthetase ValS, suggesting that
the invariant proline triplet in ValS may
explain the coevolution of EF-P.

Cell Reports
Report
A Conserved Proline Triplet in Val-tRNASynthetase and the Origin of Elongation Factor PAgata L. Starosta,1 Jurgen Lassak,2 Lauri Peil,3,4 Gemma C. Atkinson,4,5 Christopher J. Woolstenhulme,6 Kai Virumae,7
Allen Buskirk,6 Tanel Tenson,4 Jaanus Remme,7 Kirsten Jung,2,8 and Daniel N. Wilson1,8,*1Gene Center and Department for Biochemistry, University of Munich, Feodor-Lynen-Straße 25, 81377 Munich, Germany2Department of Biology I, Microbiology, Ludwig-Maximilians-Universitat Munchen, 82152 Martinsried, Germany3Wellcome Trust Centre for Cell Biology, University of Edinburgh, Edinburgh EH8 9YL, UK4Institute of Technology, University of Tartu, Tartu 50090, Estonia5Department of Molecular Biology and Laboratory for Molecular InfectionMedicine Sweden (MIMS), Umea University, Umea 901 87, Sweden6Department of Chemistry and Biochemistry, Brigham Young University, Provo, UT 84602, USA7Institute of Molecular and Cell Biology, University of Tartu, Tartu 50090, Estonia8Center for Integrated Protein Science Munich (CiPSM) at the University of Munich, 80539 Munich, Germany
*Correspondence: [email protected]
http://dx.doi.org/10.1016/j.celrep.2014.09.008
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
SUMMARY
Bacterial ribosomes stall on polyproline stretchesand require the elongation factor P (EF-P) to relievethe arrest. Yet it remains unclear why evolution hasfavored the development of EF-P rather than select-ing against the occurrence of polyproline stretchesin proteins. We have discovered that only a singlepolyproline stretch is invariant across all domains oflife, namely a proline triplet in ValS, the tRNA synthe-tase, that charges tRNAVal with valine. Here, we showthat expression of ValS in vivo and in vitro requiresEF-P and demonstrate that the proline triplet locatedin the active site of ValS is important for efficientcharging of tRNAVal with valine and preventing forma-tion of mischarged Thr-tRNAVal as well as efficientgrowth of E. coli in vivo. We suggest that the criticalrole of the proline triplet for ValS activity may explainwhy bacterial cells coevolved the EF-P rescuesystem.
INTRODUCTION
Polymerization of amino acids by ribosomes to form polypeptide
chains is a fundamental process in all cells. Ribosomes can poly-
merize most polypeptide chains without difficulty, but distinct
amino acid combinations pose serious problems. For example,
three or more consecutive proline residues induce translational
arrest by preventing peptide-bond formation (Doerfel et al.,
2013; Ude et al., 2013; Woolstenhulme et al., 2013). Ribosome
stalling results from the slow rate of peptide-bond formation be-
tween the peptidyl-Pro-Pro-tRNA located in the P site and the
Pro-tRNA in the A site (Doerfel et al., 2013). Ribosome stalling
in the absence of elongation factor P (EF-P) has also been
observed at diprolyl motifs (XPPY), with the efficiency of stalling
dependent on the nature of the amino acid located before (X) and
after (Y) the diprolyl motif (PP) (Hersch et al., 2013; Peil et al.,
2013; Woolstenhulme et al., 2013). The translational arrest at
PPP and XPPY motifs was in all cases relieved by the presence
of the translation elongation factor EF-P (Peil et al., 2013), which
binds to the stalled ribosomes and stimulates peptide-bond
formation (Doerfel et al., 2013; Ude et al., 2013). A conserved
lysine residue of Escherichia coli EF-P is subject to posttransla-
tional modification by YjeA, YjeK, and YfcM (EpmA, EpmB, and
EpmC) (Navarre et al., 2010; Peil et al., 2012; Yanagisawa
et al., 2010), and this lysinylation modification is required for
the rescue activity of EF-P (Doerfel et al., 2013; Peil et al.,
2013; Ude et al., 2013). The equivalent lysine of the ortholog of
EF-P in archaea and eukaryotes, IF-5A (a/eIF-5A), is also post-
translationally modified, but via hypusinylation rather than lysiny-
lation (Park et al., 2010). In yeast, the posttranslational modi-
fication of eIF-5A is also critical for rescue of ribosomes stalled
on polyproline stretches (Gutierrez et al., 2013). Moreover, the
associated archaeal and eukaryotic enzymes, deoxyhypusine
synthase (DHS) and deoxyhypusine hydroxylase (DOHH), are
evolutionarily unrelated to their bacterial counterparts (Park
et al., 2010). Thus, nature has evolved not only specialized trans-
lation factors to overcome stalling at polyproline stretches but
also independent sets of modification enzymes to activate these
factors. This in itself implies that the benefits of retaining polypro-
line stretches significantly outweigh the cost of implementing
and maintaining the EF-P and a/eIF5A rescue systems.
RESULTS
Conservation of Polyproline StretchesIn order to understand which proteins have in fact retained
a polyproline stretch (three or more consecutive prolines)
throughout evolution, we initially examined the number and
conservation of polyproline-containing proteins across 1,273
completely sequenced bacterial genomes. As seen in Figure 1A,
the number of polyproline-containing proteins varies across the
different bacterial phyla and is generally higher in bacteria with
larger genomes, for example, the delta-proteobacterium Soran-
gium cellulosum, which has the largest sequenced genome to
date (9,367 protein-coding genes [CDS]) (Schneiker et al.,
Cell Reports 9, 1–8, October 23, 2014 ª2014 The Authors 1
Please cite this article in press as: Starosta et al., A Conserved Proline Triplet in Val-tRNA Synthetase and the Origin of Elongation Factor P, Cell Re-ports (2014), http://dx.doi.org/10.1016/j.celrep.2014.09.008

2007), has the most polyproline-containing proteins (i.e., 2,406
occurrences in 1,779 of the 9,367 proteins, or 19%). By compar-
ison, a typical K12 E. coli strain has�100 polyproline-containing
proteins from a total of �4,100 CDS (2%). Curiously, we identi-
fied 19 bacterial genomes ranging in size from 182 to 2,013
CDS encoding only a single polyproline-containing protein
(0.05%–0.5%) (Figures 1A and 1B). These include both free-
living bacteria of theMycoplasma genus aswell asmany obligate
endosymbiotic bacteria, such as Carsonella ruddii, which
cohabit with sap-feeding insects, such as aphids, psyllids, and
cicadas (Figure 1B). In all cases, the single polyproline-contain-
ing CDS was identified as ValS (E. coli nomenclature), the Val-
tRNA synthetase that aminoacylates tRNAVal with the amino
acid valine. The valS gene, encoding ValS, is an essential gene
(Gerdes et al., 2003) and is correspondingly present in all do-
mains of life. The presence and location of the proline triplet in
ValS is invariant not only in bacteria but also in the 205 archaeal
and 98 eukaryotic genomes that we analyzed (Figure 1C). In fact,
the proline triplet has a higher conservation than the ‘‘HIGH’’
motif that defines ValS as a class I tRNA synthetase (Arnez and
Moras, 1997). No conserved polyproline stretches longer than
a triplet were identified. The protein with the next-highest con-
servation of a polyproline stretch was the translational GTPase
LepA, which contains a proline triplet in �65% of the 1,273
sequenced genomes, and, unlike valS, the lepA gene is not
essential for viability (Baba et al., 2006). Given that XPPY motifs
other than PPP also cause stalling (albeit at lower levels than
PPP), which is relieved by EF-P (Peil et al., 2013), we also
analyzed the bacterial genomes for conservation of XPPYmotifs.
We detected only one diprolyl motif as being correspondingly
conserved in all domains of life, namely, the GPP motif found
within IleS, the Ile-tRNA synthetase that aminoacylates tRNAIle
with the amino acid isoleucine. The presence and location of
the GPP motif in IleS is homologous to the PPP motif in ValS,
also being located adjacent to the HIGH motif (Figure 1C). We
have previously demonstrated that ribosomes stall only weakly
at GPP motifs in the absence of EF-P, whereas much stronger
stalling is observed at PPP motifs (Peil et al., 2013).
ValS Expression Is Dependent on Modified EF-PTo show that active EF-P is in fact required for ValS expression,
we analyzed our previous stable isotope labeling and mass
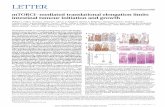
Figure 1. A Conserved Proline Triplet in ValS Confers EF-P Dependence for Expression
(A) Scatterplot illustrating the total number of polyproline-containing proteins relative to genome size (in terms of open reading frame [ORF] number) of species
within different bacterial phyla.
(B) Zoom of (A) highlighting bacteria with the smallest genomes.
(C) Sequence alignment illustrating the conservation of PPP and GPP in Val- and Ile-tRNA synthetases, respectively, across the three domains of life, compared
with the highly conserved HIGH motif.
(D) Heatmap representation of changes in expression levels in vivo (blue, downregulated; red, upregulated; white, unchanged; gray, not identified or quantified) of
selected proteins when comparing Defp, DyjeA, DyjeK, and DyfcM strains against a wild-type E. coli internal standard. Data from two biological replicates were
taken from Peil et al. (2013).
(E) Schematic illustration of the gene arrangement of the ilvL-BN and ilvL-GMEDA operons, with the corresponding sequence of the leader peptides IvbL and IlvL.
Valine residues essential for attenuation are shown in red.
(F) Autoradiograph of SDS-PAGE monitoring the in vitro translation at 3 min and 7 min of ValS, in the absence (�EF-P) and presence of EF-P (+EF-P), with full-
length (FL) ValS and peptidyl-tRNA (25 kDa) as indicated.
(G) Toeprinting of ValS in the absence (�) and presence (+) of active EF-P. G and C sequence lanes are included.
2 Cell Reports 9, 1–8, October 23, 2014 ª2014 The Authors
Please cite this article in press as: Starosta et al., A Conserved Proline Triplet in Val-tRNA Synthetase and the Origin of Elongation Factor P, Cell Re-ports (2014), http://dx.doi.org/10.1016/j.celrep.2014.09.008

spectrometry data that monitored the changes in expression
levels of proteins in E. coli strains lacking the efp, yjeK, yjeA,
and yfcM genes relative to the parental wild-type E. coli strain
(Peil et al., 2013). As predicted, ValS was heavily downregulated
when EF-P was absent (Defp) or EF-P was inactive because of
lack of lysinylation (DyjeK/DyjeA) (Figure 1D). The lack of YfcM
(DyfcM), which hydroxylates the lysinylated EF-P (Peil et al.,
2012), did not lead to downregulation of ValS, consistent
with previous reports that lysinylated, but not hydroxylated,
EF-P still retains the ability to stimulate peptide-bond formation
and thus relieve stalling at polyproline stretches (Doerfel et al.,
2013; Peil et al., 2013; Ude et al., 2013). The downregulation
of ValS in the absence of active EF-P is specific, since down-
regulation of other tRNA synthetases, such as LeuS or IleS, is
not observed (Figure 1D). The lack of dependence of IleS
expression on EF-P is consistent with our previous findings
that the GPP motif only induces weak translational stalling
compared to PPP (2-fold versus 20-fold, respectively) (Peil
et al., 2013).
Notably, we also observed upregulation of the proteins
encoded in the ilvBN and ilvGEDA operons involved in the
biogenesis of isoleucine, leucine, and valine. This can be ex-
plained by transcriptional attenuation of the ilvBN and ilvGEDA
operons (Yanofsky, 1987). When amino acids such as valine
are limiting, the reduction in Val-tRNAVal causes ribosomes to
stall during translation of the upstream valine-rich leader pep-
tides IvbL and IlvL. This in turn prevents formation of a down-
stream transcription termination structure, leading to induction
of expression of the downstream genes in the operon (Yanof-
sky, 1987) (Figure 1E). Similarly, a reduction in charged Val-
tRNAVal due to the downregulation of ValS would therefore
explain the upregulation of ilvBN and ilvGEDA operons in the
absence of active EF-P (Figure 1D). In contrast, the absence
of active EF-P had little effect on the expression of the ilvI
and ilvH, which are not attenuated, indicating that the upregu-
lation of the ilvBN and ilvGEDA operons is specific. Although
the efp gene is not essential in E. coli (Baba et al., 2006; Peil
et al., 2012, 2013), the loss of EF-P leads to a reduction in
growth rate (Yanagisawa et al., 2010), presumably due to the
reduction in levels of important cellular polyproline-containing
proteins. Since we have demonstrated that the lack of EF-P
leads to a strong downregulation of ValS (Figure 1D), we tested
and could show that expression of ValS could partially rescue
the growth defect due to lack of the efp gene in E. coli
(Figure S1).
We also demonstrated that ValS expression is dependent on
EF-P in vitro (Figure 1F): In the absence of EF-P, translation of
ValS in vitro leads to accumulation of a �25 kDa band, which
correlates with that of the ValS nascent polypeptide chain trans-
lated up to the proline triplet (�5 kDa), but still remaining
attached to a tRNA (�20 kDa tRNA). As expected, the band
for the stalled peptidyl-tRNA is absent when EF-P is included
in the in vitro translation reaction (Figure 1F). Toeprinting
confirmed that when EF-P is absent, the ribosome stalls at the
proline triplet of ValS, specifically with the second proline codon
of the proline triplet located in the P site (Figure 1G). Collectively,
these findings demonstrate that expression of ValS is strictly
dependent on the presence of active EF-P in vivo and in vitro.
The Proline Triplet Is Located at the Active Site of ValSIf the EF-P rescue system really evolved to allow the proline
triplet in ValS to be retained, this would imply that the proline
triplet in ValS is critically important for function, as indicated by
its universal conservation. Crystal structures of T. thermophilus
ValS reveal that the proline triplet (P41–P43, E. coli numbering)
resides in the active site of ValS, namely where valine is activated
by ATP to form valyl-AMP and then transferred to tRNAVal to form
Val-tRNAVal (Figure 2A) (Fukai et al., 2000, 2003). The side chain
of Pro41 of ValS contacts the g1-CH3 group of valyl-AMP,
whereas Pro42 and Pro43 appear to also contribute to the stable
binding of valyl-AMP by positioning the a-CO group of Pro42 to
hydrogen bond with the a-NH3 group of valyl-AMP (Figure 2A)
(Fukai et al., 2000, 2003). In IleS, the presence of Gly45, rather
than Pro, allows accommodation of isoleucyl-AMP (Figure 2B)
(Nureki et al., 1998; Silvian et al., 1999), consistent with the
high conservation of the GPP motif in Ile-tRNA synthetases (Fig-
ure 1C). In contrast, accommodation of isoleucyl-AMP in ValS is
less favorable due to the close distance between Pro41 and the
g1-CH3 group of isoleucyl-AMP (Figure 2C), which led to the pro-
posed role of Pro41 in ValS in the discrimination between valine
and isoleucine (Fukai et al., 2000). In a simplified model, one
might predict that mutation of P41G in ValS should increase
the level of misincorporation of isoleucine (Figure 2D); however,
this has not been tested.
Critical Importance of the Proline Triplet for ValSCharging ActivityTo experimentally validate the role of Pro41 aswell as the general
functional importance of the proline triplet for ValS activity, we
generated the three single ValS Pro-to-Gly mutants (ValS-GPP,
-PGP and -PPG) as well as a triple PPP-to-GGG mutant (ValS-
GGG). The ability of the ValS mutants to charge tRNAVal with
[14C]valine was assessed and compared to wild-type ValS (Fig-
ure 2E). Under our reaction conditions at 37�C,maximal charging
of tRNAVal with [14C]valine by wild-type ValS occurred within the
first minute (Figure 2E). In contrast, all ValS mutants were less
efficient than wild-type; the ValS-PPG and -GGG mutants were
completely devoid of activity, whereas the ValS-PGP and -GPP
mutants retained some activity but at lower levels than the
wild-type ValS (Figure 2E).
Charging of tRNAVal by ValS occurs in two steps. First, valine
is activated with ATP to form Val-AMP, leading to the release of
pyrophosphate (Figure 2F). Second, the valine is then trans-
ferred to the tRNAVal to form Val-tRNAVal, with a concomitant
release of AMP (Figure 2G). Initially, we employed thin-layer
chromatography (TLC) and g[32P]-ATP to monitor the release
of pyrophosphate (g[32P]-PPi) by wild-type ValS, or the mutant
ValS-GPP, in the presence of valine, and in the presence or
absence of tRNAVal (Figure 2F). Pyrophosphate (PPi) release is
monitored indirectly by treatment with pyrophosphatase that
converts the PPi to inorganic monophosphate (Pi), which is
then visualized directly by TLC. As seen in Figure 2F, Pi for-
mation is observed only for wild-type ValS in the presence of
the pyrophosphatase and, as expected, is stimulated by the
presence of tRNAVal. The migration position of Pi on the TLC
was determined using a control reaction of g[32P]-ATP treated
with Apyrase, an ATP diphosphatase that hydrolyzes ATP
Cell Reports 9, 1–8, October 23, 2014 ª2014 The Authors 3
Please cite this article in press as: Starosta et al., A Conserved Proline Triplet in Val-tRNA Synthetase and the Origin of Elongation Factor P, Cell Re-ports (2014), http://dx.doi.org/10.1016/j.celrep.2014.09.008

sequentially to yield AMP and Pi. Surprisingly, the ValS-GPP
mutant was observed to produce large quantities of g[32P]-con-
taining material that migrated at a position similar to but distinct
from Pi (termed Pi*). Moreover, formation of Pi* by the ValS-GPP
mutant was independent of pyrophosphatase treatment or the
presence of tRNAVal (Figure 2F). Pi* was also produced by the
ValS-PGP and ValS-PPG mutants in a pyrophosphatase- and
tRNAVal-independent fashion, whereas the ValS-GGG mutant
was virtually inactive (Figure S2A). Based on the retention factor
(Rf) of Pi* (Rf 0.69), we can conclude that Pi* is not AMP (Rf 0.83),
ADP (Rf 0.17), ATP (Rf 0), or monophosphate Pi (Rf 0.56).
Furthermore, Pi* cannot be PPi, since PPi is not observable on
our TLCs and addition of pyrophosphatase does not influence
the migration of Pi* (Figure 2F). PPi generated by tRNA synthe-
tases has been reported to attack ATP and ADP to generate
diadenosine polyphosphates, such as Ap3A and Ap4A via
phosphorolysis (Plateau and Blanquet, 1976); however, we
can exclude that Pi* is Ap3A or Ap4A, since these compounds
are resistant to phosphatase treatment whereas Pi* is not (Fig-
ure S2B). Further experiments will be required to determine
the exact nature of Pi*.
Since the ValS mutants produce Pi*, rather than PPi, we
reasoned that the ValS mutants may also catalyze a reaction
other than canonical ATP to AMP, as catalyzed by a ValS-
tRNA synthetase. To address this, charging assays were per-
formed as before but using a[32P]-ATP instead of g[32P]-ATP.
Each reaction contained the amino acid valine, together with
either wild-type ValS or one of the ValS mutants, and was per-
formed in the absence or presence of tRNAVal (Figure 2G). As ex-
pected, wild-type ValS produced AMP only in the presence of
tRNAVal. AMP formation was also observed for the ValS-GPP,
PGP, and, to a lesser extent, the PPG mutant, consistent with
some degree of charging activity observed in Figure 2E, whereas
ValS-GGG was virtually inactive. Strikingly, unlike wild-type
ValS, the ValS-GPP, -PGP, and -PPG mutants converted large
amounts of a[32P]-ATP to a[32P]-ADP (Figure 2G). The migration
position on the TLC of a[32P]-ADP (and a[32P]-AMP) was deter-
mined using a control reaction where a[32P]-ATP was treated
with increasing concentrations of Apyrase that hydrolyzes
sequentially ATP to ADP and then AMP and Pi. Furthermore,
the generation of a[32P]-ADP by the ValS-GPP, -PGP and
-PPG mutants was independent of the presence of tRNAVal
Figure 2. PPP at the Active Site of ValS Is Required for Efficient tRNA Charging
(A) Location of PPP in the active site of ValS relative to Val-AMP (Protein Data Bank [PDB] ID 1GAX) (Fukai et al., 2000).
(B) Active site of IleS with Ile-AMP (1JZQ) (Nakama et al., 2001).
(C) ValS from (A) but with superimposed position of Ile-AMP from (B).
(D) Experiments performed as in (C) but with in silico Pro41Gly mutation.
(E) Charging efficiency of tRNAVal with valine by wild-type (WT) ValS and ValS mutants as a function of time (min).
(F and G) Autoradiograph of TLC separation of (F) g[32P]-Pi from g[32P]-ATP, and (G) a[32P]-AMP and a[32P]-ADP from a[32P]-ATP, when wild-type ValS (WT), or
ValS mutant(s) were incubated with valine, in the absence (�) and presence (+) of pyrophosphatase (PPi-ase) and/or deacylated tRNAVal. ATP at the origin in-
dicates where the sampleswere loaded onto the TLC plate. In (F), themigration position of Pi is determined by treatment of g[32P]-ATPwith Apyrase (A), and in (G),
the migration of AMP and ADP was determined by treatment of a[32P]-ATP with increasing concentrations of Apyrase.
4 Cell Reports 9, 1–8, October 23, 2014 ª2014 The Authors
Please cite this article in press as: Starosta et al., A Conserved Proline Triplet in Val-tRNA Synthetase and the Origin of Elongation Factor P, Cell Re-ports (2014), http://dx.doi.org/10.1016/j.celrep.2014.09.008

and also occurredwhen no amino acid was present (Figure S2C).
Moreover, the a[32P]-ADP generated by the ValS-GPP, -PGP,
and -PPG mutants could be converted to a[32P]-AMP by
Apyrase (Figure S2D). In summary, these assays suggest that
mutations within the conserved proline triplet of ValS not only
reduce the efficiency of tRNA charging but also cause the ValS
mutants to nonproductively hydrolyze ATP to ADP.
The Proline Triplet Is Important for the EditingActivity of ValSTo test the hypothesis that mutation of Pro41 in ValS distin-
guishes between valine and isoleucine (Figure 2D), we performed
mischarging assays where [14C]isoleucine replaced [14C]valine.
No mischarging of tRNAVal with [14C]isoleucine by wild-type
ValS, or any of the ValSmutants, was observed (data not shown).
Furthermore, introduction of T222P mutation that disables the
ValS editing function did not promote Ile-tRNAVal formation
(data not shown), consistent with Ile not being a substrate for
editing by ValS (Tardif et al., 2001). Similarly, isoleucine did not
stimulate AMP formation by ValS-GPP (Figure S2E), collectively
suggesting that Pro41 is not the only residue in ValS involved in
discrimination of valine from isoleucine. On the other hand, thre-
onine, which is isosteric with valine, is readily activated by ValS
and transferred to tRNAVal, but the mischarged Thr-tRNAVal
that forms is rapidly deacylated by the editing domain of ValS
(Fersht and Kaethner, 1976) (Figures 3A and 3B). Indeed, in mis-
charging assays where [14C]threonine replaced [14C]valine, we
observed a low rate of Thr-tRNAVal formation by wild-type
ValS, which was enhanced when the editing domain was
disabled by the T222P mutation (Doring et al., 2001) (Figures
3B and 3C). Formation of Thr-tRNAVal was also enhanced by
the ValS-GPP and -PGP mutations. The introduction of the
T222P mutation in ValS-GPP did not further enhance mischarg-
ing, indicating that the GPP mutation may disable the editing
function of ValS (Figure 3C). Surprisingly, mischarging was sup-
pressed when the T222P was introduced into the ValS-PGP
mutant (Figure 3C), suggesting an intricate communication exists
between the activation and editing domains of ValS. This inter-
play was also observed in the TLC analyses measuring the mis-
activation of threonine, namely, in that the nonproductive ADP
formation by the ValS-GPP and -PGP mutants was suppressed
in the presence of the additional T222P mutation (Figure 3D).
Collectively, the in vitro assays indicate that the conserved pro-
line triplet of ValS is important not only for efficient tRNA charging
but also for communication with the editing domain to ensure
efficient deacylation of mischarged Thr-tRNAVal.
The Proline Triplet of ValS Is Important forViability of E. coliGiven the defects of the ValS mutants in vitro, we assessed their
functionality in vivo using a genetic complementation system
(Figure 4A). The DvalS E. coli strain was complemented with a
plasmid bearing ValS containing an N-terminal 6xHistidine
(6xHis) tag and small ubiquitin-like modifier (SUMO) tag (Hay,
2005) followed by a WFCWS linker. The resulting JL001 strain
(Table S1) was viable but exhibited a slightly reduced growth
rate, presumably because of the lower expression level of the
plasmid-encoded ValS (Figure 4B). Next a plasmid expressing
Ulp1 (pUlp1) was introduced into JL001. Ulp1 encodes a
SUMO-specific protease that cleaves the SUMO tag from ValS
exposing the N-terminal WFCWS sequence motif that, in accor-
dance with the N-end degradation rule, leads to recognition and
degradation of the ValS protein by the ClpAP protease (Dougan
et al., 2010; Wang et al., 2007) (Figure 4A). Under the conditions
used, western blotting against the 6xHis tag indicated a rapid
loss of ValS protein in the cell within minutes of induction of
Ulp1 expression (Figure S3). Consistently, induction of Ulp1
expression also led to strong reduction in growth rate of the
JL001 strain (Figure 4B). In the next step, we repeated the growth
experiments with the JL001 strain bearing pUlp1 (or a control
plasmid) together with an additional plasmid bearing an IPTG-in-
ducible copy of either the wild-type ValS (pValS-PPP; Figure 4C)
Figure 3. PPP at the Active Site of ValS Is Required to Prevent Thr-tRNAVal Formation
(A) Scheme for misactivation and mischarging of tRNAVal with threonine and subsequent deacylation by ValS.
(B) Overview of ValS bound with tRNA (yellow) and Thr-AMP (orange) (PDB 1GAX) (Fukai et al., 2000). PPP and T222 are indicated by red spheres. Inset shows the
active site of ValS with PPP relative to Thr-AMP.
(C) Mischarging of tRNAVal with threonine by wild-type (WT) ValS and ValS mutants as a function of time (min).
(D) Autoradiograph of TLC separation of a[32P]-AMP and a[32P]-ADP from a[32P]-ATP, when wild-type ValS (WT) or ValSmutant(s) were incubated with threonine,
in the absence (�) and presence (+) of deacylated tRNAVal. ATP at the origin indicates where the sampleswere loaded onto the TLCplate and apyrase treatment of
ATP provides markers for ADP and AMP positions.
Cell Reports 9, 1–8, October 23, 2014 ª2014 The Authors 5
Please cite this article in press as: Starosta et al., A Conserved Proline Triplet in Val-tRNA Synthetase and the Origin of Elongation Factor P, Cell Re-ports (2014), http://dx.doi.org/10.1016/j.celrep.2014.09.008

or one of the ValS mutants (ValS-GPP, -PGP, -PPG, or -GGG;
Figures 4D–4G). As expected, the presence of an additional
copy of the wild-type ValS (pValS-PPP) could rescue the growth
of JL001 following the degradation of the SUMO-ValS that oc-
curs upon induction of Ulp1 expression (Figure 4C). Similarly,
pValS-GPP was also able to rescue growth upon Ulp1 expres-
sion (Figure 4D), albeit not as efficiently as pValS-PPP. By
contrast, the pValS-PGP, -PPG, and -GGG displayed little or
no ability to rescue growth in the absence of wild-type ValS (Fig-
ures 4E–4G). Unfortunately, attempts to transform plasmid li-
braries of pValS-XXX mutants to select for viable mutations of
the proline triplet failed; however, given the ability of the ValS-
GPP mutant to rescue, we therefore generated all possible
pValS-XPP mutants and monitored their ability to rescue growth
in the absence of wild-type ValS (Figure 4H). Of the 20 pValS-
XPP mutants tested, six mutants (PPP, GPP, APP, SPP, CPP,
and TPP) rescued growth in the absence of wild-type ValS,
with only APP and SPP being able to sustain growth at levels
comparable to PPP and GPP (Figure 4H).
DISCUSSION
We identified only a single polyproline stretch that is invariant
across all bacterial genomes, namely a proline triplet present in
ValS, the Val-tRNA synthetase (Figures 1B and 1C), and demon-
strated that efficient expression of ValS in vivo and in vitro re-
quires the presence of EF-P. Since the PPP triplet in ValS is
invariant in bacteria, archaea, and eukaryotes (Figure 1C), this
leads us to propose that EF-P and a/eIF-5A may have initially
(co)evolved to facilitate primarily the expression of ValS. We
cannot, however, exclude that other polyproline-containing
proteins contributed to the evolution of EF-P or that EF-P and
a/eIF-5A evolved to facilitate expression of another protein that
contained a polyproline-stretch at the time but that was subse-
quently lost from some or all of the currently available bacterial
genomes.
Regardless of whether EF-P and a/eIF-5A evolved to facilitate
expression of ValS or not, our study nevertheless identified a
highly conserved PPPmotif located in the active site of ValS (Fig-
ure 2A) that is critical for the tRNA charging activity (Figure 2E)
and productive ATPase activity (Figures 2F and 2G) of ValS.
Although comparisons of ValS and IleS led to the suggestion
that Pro41 of ValS is important for the discrimination of valine
from isoleucine (Fukai et al., 2000), we find no evidence to sup-
port this. Instead, we find that mutations within the PPP motif
of ValS enhance formation of mischarged Thr-tRNAVal (Fig-
ure 3C). Moreover, we could also demonstrate that the T222P
mutation within the editing site of ValS could suppress the
nonproductive formation of ADP by the ValS mutants (Figure 4D)
and in the case of ValS-PGP was even able to rescue the editing
Figure 4. The Proline Triplet of ValS Is Required for Viability of E. coli
(A) Scheme for the ValS complementation system.
(B–G) Growth curves (OD600nm) of E. coli strains (B) BW25113, JL001, JL001 + pUlp1, or (C–G) JL001 bearing additional plasmids expressing pUlp1, and/or (C)
wild-type ValS (pValS-PPP) or ValS mutants (D) pValS-GPP, (E) pValS-PGP, (F) pValS-PPG, or (G) pValS-GGG.
(H) Comparison of growth (after 90 min) of JL001 bearing plasmids expressing pUlp1 and pValS/XPPmutants normalized by subtraction of background growth in
the presence of pUlp1 alone. The error bars represent the SD from the mean of three biological replicates.
6 Cell Reports 9, 1–8, October 23, 2014 ª2014 The Authors
Please cite this article in press as: Starosta et al., A Conserved Proline Triplet in Val-tRNA Synthetase and the Origin of Elongation Factor P, Cell Re-ports (2014), http://dx.doi.org/10.1016/j.celrep.2014.09.008

activity (Figure 4C). Collectively, these findings suggest an inti-
mate communication between the aminoacylation and editing
sites on ValS that warrants further study.
Consistent with our in vitro assays, the ValS-GPP mutant was
able to rescue growth and viability of an E. coli strain lacking
wild-type valS gene, but not as efficiently as ValS-PPP, whereas
ValS-PGP, -PPG, and -GGG displayed little or no rescue pheno-
type. Testing of all possible ValS-XPP mutants demonstrated
that it is possible to select for alternative motifs, such as APP
or SPP, that support viability of E. coli under optimal conditions;
however, it remains to be tested whether such mutants are also
impaired in tRNA charging activities as observed for the ValS-
GPP mutant.
The critical importance of the proline triplet of ValS for tRNA
charging and viability of E. coli would provide an explanation
for why nature needed to coevolve EF-P and a/eIF-5A to facili-
tate expression during translation of the ValS.
EXPERIMENTAL PROCEDURES
Oligonucleotides, Plasmids, and Bacterial Strains
Primers, plasmids, and strains used in this study are listed in Table S1.
Bioinformatic Analysis
To search for ValRS protein sequences and determine the conservation of the
PPP motif, 1,273 bacterial, 205 archaeal, and 98 eukaryotic genomes were
searched with a hidden Markov model (HMM) profile (Eddy, 1998) with an E
value cutoff of e-70 as described previously. The HMM was generated from
a MAFFT (Katoh et al., 2005) alignment of ValRS homologs identified in an
initial BlastP search (Altschul et al., 1990). As the HMM also hits the close rela-
tive Ile-tRNA synthetase, phylogenetic analysis using FastTree (Price et al.,
2010) was carried out to extract sequences from the ValRS clade of orthologs.
The resulting sequences were aligned, and the PPP motif region was in-
spected for conservation.
tRNAVal Charging Assays
Charging reactions and assays tomonitor aminoadenylate formationwere per-
formed as described previously (Splan et al., 2008).
Growth Conditions
Lysogeny broth (LB)was used as complexmedium (Bertani, 1951) andmodified
with NaCl. When indicated, LBwas supplemented with 0.4% (w/v) glucose as a
carbon source to suppress Ulp1 expression before induction with L-arabinose
0.2% (w/v). A total of 1 mM isopropyl-b-D-1-thiogalactopyranoside (IPTG)
(Sigma-Aldrich) was used to induce expression of pQE70-ValS (pValS-PPP)
andGPP, PGP, PPG, andGGG variants. Antibioticswere used when necessary
with the following concentrations: 100 mg/ml ampicillin sodium salt, 50 mg/ml
kanamycin sulfate, 34 mg/ml chloramphenicol, 20 mg/ml gentamycin sulfate.
ValS Silencing
E. coli BW2113 was used to delete valS via pRED/ET recombination technol-
ogy. Due to the fact that valS is an essential gene, gene deletionwas performed
in a strain containing plasmid pBBR1MCS-5-PT7-SUMO-WFCWS-valS en-
coding SUMO-ValS (ValS*). The resultant strain JL001 was cotransformed
with pBAD33 or pBAD33-ulp1, respectively, and pQE70 encoding either
wild-type ValS or ValS-GPP, -PGP, -PPG, or -GGGmutants. Cells were plated
on LB agar containing chloramphenicol and ampicillin for transformant selec-
tion aswell as IPTG to induce pQE70-ValS expression and glucose to suppress
pBAD activity by catabolite repression. Transformants were precultured over-
night in LB + glucose + chloramphenicol (pBAD33) + gentamycin sulfate
(pBBR1MCS-5) + ampicillin sodium salt (pQE70) + kanamycin sulfate (DvalS::
kan). A second preculture was started by diluting the overnight culture 1:100.
Cells were grown to an optical density of 600 nm (OD600nm) = 0.3–0.7. Subse-
quently, cells were washed twice in LB + 0.2% (w/v) arabinose and adjusted to
an OD600nm = 0.1. The main culture was started by a further dilution step to
OD600nm = 0.01 in a microtiter plate in a final volume of 200 ml LB + 0.2% arab-
inose + chloramphenicol. Growth was recorded in a Tecan reader (Infinite
M200 Pro) by measuring OD600nm every 10 min for 8 hr. In parallel, Ulp1 pro-
duction and ValS degradation was monitored by western blot analysis. Cells
of the JL001 strain harboring pBAD33-ulp1 and pBBR1MCS-5-PT7-SUMO-
WFCWS-valS were harvested at different times after arabinose induction. As
a control, JL001 cells harboring pBAD33 and pBBR1MCS-5-PT7-SUMO-
WFCWS-valS were analyzed.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures,
three figures, and one table and can be found with this article online at
http://dx.doi.org/10.1016/j.celrep.2014.09.008.
ACKNOWLEDGMENTS
We would like to thank Ingrid Weitl for technical assistance and Michael Graf
for help with purification of the ValS mutants. This research was supported
by grants from the Deutsche Forschungsgemeinschaft (FOR1805 and
WI3285/4-1 to D.N.W., and Exc114/2 to K.J.), the Estonian Science Founda-
tion (ETF9020 to G.C.A), and IUT 14021 (JR). L.P. and G.A. received support
from the European Social Fund program Mobilitas grant MJD144 and
MJD99, respectively. A.L.S. is funded by an AXA Research Fund postdoctoral
fellowship, and L.P. is supported by the Marie Curie FP7-PEOPLE-2011-IEF
postdoctoral fellowship.
Received: April 25, 2014
Revised: August 5, 2014
Accepted: September 5, 2014
Published: October 9, 2014
REFERENCES
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990). Basic
local alignment search tool. J. Mol. Biol. 215, 403–410.
Arnez, J.G., and Moras, D. (1997). Structural and functional considerations of
the aminoacylation reaction. Trends Biochem. Sci. 22, 211–216.
Baba, T., Ara, T., Hasegawa, M., Takai, Y., Okumura, Y., Baba, M., Datsenko,
K.A., Tomita, M., Wanner, B.L., and Mori, H. (2006). Construction of Escheri-
chia coli K-12 in-frame, single-gene knockout mutants: the Keio collection.
Mol. Syst. Biol. 2, 0008.
Bertani, G. (1951). Studies on lysogenesis. I. The mode of phage liberation by
lysogenic Escherichia coli. J. Bacteriol. 62, 293–300.
Doerfel, L.K., Wohlgemuth, I., Kothe, C., Peske, F., Urlaub, H., and Rodnina,
M.V. (2013). EF-P is essential for rapid synthesis of proteins containing
consecutive proline residues. Science 339, 85–88.
Doring, V., Mootz, H.D., Nangle, L.A., Hendrickson, T.L., de Crecy-Lagard, V.,
Schimmel, P., and Marliere, P. (2001). Enlarging the amino acid set of Escher-
ichia coli by infiltration of the valine coding pathway. Science 292, 501–504.
Dougan, D.A., Truscott, K.N., and Zeth, K. (2010). The bacterial N-end rule
pathway: expect the unexpected. Mol. Microbiol. 76, 545–558.
Eddy, S.R. (1998). Profile hidden Markov models. Bioinformatics 14, 755–763.
Fersht, A.R., and Kaethner, M.M. (1976). Enzyme hyperspecificity. Rejection of
threonine by the valyl-tRNA synthetase by misacylation and hydrolytic editing.
Biochemistry 15, 3342–3346.
Fukai, S., Nureki, O., Sekine, S., Shimada, A., Tao, J., Vassylyev, D.G., and Yo-
koyama, S. (2000). Structural basis for double-sieve discrimination of L-valine
from L-isoleucine and L-threonine by the complex of tRNA(Val) and valyl-tRNA
synthetase. Cell 103, 793–803.
Fukai, S., Nureki, O., Sekine, S., Shimada, A., Vassylyev, D.G., and Yokoyama,
S. (2003). Mechanism of molecular interactions for tRNA(Val) recognition by
valyl-tRNA synthetase. RNA 9, 100–111.
Cell Reports 9, 1–8, October 23, 2014 ª2014 The Authors 7
Please cite this article in press as: Starosta et al., A Conserved Proline Triplet in Val-tRNA Synthetase and the Origin of Elongation Factor P, Cell Re-ports (2014), http://dx.doi.org/10.1016/j.celrep.2014.09.008

Gerdes, S.Y., Scholle, M.D., Campbell, J.W., Balazsi, G., Ravasz, E., Daugh-
erty, M.D., Somera, A.L., Kyrpides, N.C., Anderson, I., Gelfand, M.S., et al.
(2003). Experimental determination and system level analysis of essential
genes in Escherichia coli MG1655. J. Bacteriol. 185, 5673–5684.
Gutierrez, E., Shin, B.S., Woolstenhulme, C.J., Kim, J.R., Saini, P., Buskirk,
A.R., and Dever, T.E. (2013). eIF5A promotes translation of polyproline motifs.
Mol. Cell 51, 35–45.
Hay, R.T. (2005). SUMO: a history of modification. Mol. Cell 18, 1–12.
Hersch, S.J.,Wang, M., Zou, S.B., Moon, K.M., Foster, L.J., Ibba,M., and Nav-
arre, W.W. (2013). Divergent protein motifs direct elongation factor P-medi-
ated translational regulation in Salmonella enterica and Escherichia coli.
MBio 4, e00180–e13.
Katoh, K., Kuma, K., Toh, H., and Miyata, T. (2005). MAFFT version 5: im-
provement in accuracy of multiple sequence alignment. Nucleic Acids Res.
33, 511–518.
Nakama, T., Nureki, O., and Yokoyama, S. (2001). Structural basis for the
recognition of isoleucyl-adenylate and an antibiotic, mupirocin, by isoleucyl-
tRNA synthetase. J. Biol. Chem. 276, 47387–47393.
Navarre, W.W., Zou, S.B., Roy, H., Xie, J.L., Savchenko, A., Singer, A., Edvo-
kimova, E., Prost, L.R., Kumar, R., Ibba, M., and Fang, F.C. (2010). PoxA, yjeK,
and elongation factor P coordinately modulate virulence and drug resistance in
Salmonella enterica. Mol. Cell 39, 209–221.
Nureki, O., Vassylyev, D.G., Tateno, M., Shimada, A., Nakama, T., Fukai, S.,
Konno, M., Hendrickson, T.L., Schimmel, P., and Yokoyama, S. (1998).
Enzyme structure with two catalytic sites for double-sieve selection of sub-
strate. Science 280, 578–582.
Park, M.H., Nishimura, K., Zanelli, C.F., and Valentini, S.R. (2010). Functional
significance of eIF5A and its hypusinemodification in eukaryotes. Amino Acids
38, 491–500.
Peil, L., Starosta, A.L., Virumae, K., Atkinson, G.C., Tenson, T., Remme, J., and
Wilson, D.N. (2012). Lys34 of translation elongation factor EF-P is hydroxylated
by YfcM. Nat. Chem. Biol. 8, 695–697.
Peil, L., Starosta, A.L., Lassak, J., Atkinson, G.C., Virumae, K., Spitzer, M.,
Tenson, T., Jung, K., Remme, J., and Wilson, D.N. (2013). Distinct XPPX
sequence motifs induce ribosome stalling, which is rescued by the translation
elongation factor EF-P. Proc. Natl. Acad. Sci. USA 110, 15265–15270.
Plateau, P., and Blanquet, S. (1976). Synthesis of NpnN’ (n = 3 or 4) in vitro and
in vivo. In Ap4A and Other Dinucleoside Polyphosphates, A.G. McLennan, ed.
(Boca Raton: CRC Press), pp. 81–107.
Price, M.N., Dehal, P.S., and Arkin, A.P. (2010). FastTree 2—approximately
maximum-likelihood trees for large alignments. PLoS ONE 5, e9490.
Schneiker, S., Perlova, O., Kaiser, O., Gerth, K., Alici, A., Altmeyer, M.O., Bar-
tels, D., Bekel, T., Beyer, S., Bode, E., et al. (2007). Complete genome
sequence of the myxobacterium Sorangium cellulosum. Nat. Biotechnol. 25,
1281–1289.
Silvian, L.F., Wang, J., and Steitz, T.A. (1999). Insights into editing from an
ile-tRNA synthetase structure with tRNAile and mupirocin. Science 285,
1074–1077.
Splan, K.E., Musier-Forsyth, K., Boniecki, M.T., and Martinis, S.A. (2008).
In vitro assays for the determination of aminoacyl-tRNA synthetase editing ac-
tivity. Methods 44, 119–128.
Tardif, K.D., Liu, M., Vitseva, O., Hou, Y.M., and Horowitz, J. (2001). Misacyla-
tion and editing by Escherichia coli valyl-tRNA synthetase: evidence for two
tRNA binding sites. Biochemistry 40, 8118–8125.
Ude, S., Lassak, J., Starosta, A.L., Kraxenberger, T., Wilson, D.N., and Jung,
K. (2013). Translation elongation factor EF-P alleviates ribosome stalling at pol-
yproline stretches. Science 339, 82–85.
Wang, K.H., Sauer, R.T., and Baker, T.A. (2007). ClpS modulates but is not
essential for bacterial N-end rule degradation. Genes Dev. 21, 403–408.
Woolstenhulme, C.J., Parajuli, S., Healey, D.W., Valverde, D.P., Petersen,
E.N., Starosta, A.L., Guydosh, N.R., Johnson, W.E., Wilson, D.N., and Buskirk,
A.R. (2013). Nascent peptides that block protein synthesis in bacteria. Proc.
Natl. Acad. Sci. USA 110, E878–E887.
Yanagisawa, T., Sumida, T., Ishii, R., Takemoto, C., and Yokoyama, S. (2010).
A paralog of lysyl-tRNA synthetase aminoacylates a conserved lysine residue
in translation elongation factor P. Nat. Struct. Mol. Biol. 17, 1136–1143.
Yanofsky, C. (1987). Operon specific control by transcription attenuation.
Trends Genet. 3, 356–360.
8 Cell Reports 9, 1–8, October 23, 2014 ª2014 The Authors
Please cite this article in press as: Starosta et al., A Conserved Proline Triplet in Val-tRNA Synthetase and the Origin of Elongation Factor P, Cell Re-ports (2014), http://dx.doi.org/10.1016/j.celrep.2014.09.008

Cell Reports, Volume 9
Supplemental Information
A Conserved Proline Triplet in Val-tRNA
Synthetase and the Origin of Elongation Factor P
Agata L. Starosta, Jürgen Lassak, Lauri Peil, Gemma C. Atkinson, Christopher J.
Woolstenhulme, Kai Virumäe, Allen Buskirk, Tanel Tenson, Jaanus Remme,
Kirsten Jung, and Daniel N. Wilson

Supplemental Figures
Figure S1, Related to Figure 1, Expression of ValS can improve growth of E. coli strains lacking EF-P. Growth curves (OD600nm) of E. coli strain BW25113 without plasmid (wt) or containing plasmids pValS-PPP, pValS-GPP or pValS-GGG, compared with E. coli strain BW25113 lacking efp (JW4106,(Baba et al., 2006)) without plasmid (Δefp) or containing plasmids pValS-PPP, pValS-GPP or pValS-GGG. As expected, overexpression of the inactive ValS-GGG mutant did improve the growth of the Δefp strain, but rather impaired growth further. In contrast, overexpression of the wildtype ValS and the ValS-GPP slightly improved the growth of the Δefp strain, but not to the levels of the wildtype strain. The more efficient rescue by ValS-GPP compared to wildtype ValS may result because ValS contains PPP and therefore requires EF-P for expression, whereas the ValS mutant, although less active than wildtype ValS, contains GPP conferring less dependence on EF-P for expression (Peil et al., 2013). The lack of complete rescue by ValS overexpression is expected given that many E. coli proteins, both essential and non-essential, are dependent on EF-P for expression and their loss leads to growth defects in E. coli (Baba et al., 2006).

Figure S2, Related to Figure 2F-G, Monitoring aminoadenylate formation by ValS and mutants. (A-E) Autoradiograph of thin-layer chromatographic separation of (A-B) γ[32P]-Pi from γ[32P]-ATP, and (C-D) α[32P]-AMP and α[32P]-ADP from α[32P]-ATP, when wildtype ValS (wt), or ValS mutants GPP, PGP, PPG or GGG were incubated with (A,B,D) valine or (C) no amino acid or (E) isoleucine, in the absence (-) and presence (+) of deacylated tRNAVal. The origin where the samples were loaded onto the TLC plate correlates with the position of ATP. In (B), reactions were treated with pyrophosphatase (PPi-ase) or alkaline phosphatase (AP) as indicated. In (D), the migration of AMP and ADP was determined by treatment of α[32P]-ATP with increasing concentrations of Apyrase.

Figure S3, Related to Figure 4B-H, Detection of ValS and Ulp1 by Western blotting. In parallel Ulp1 production and ValS degradation was monitored by Western Blot analysis against the 6x His tags. Cells of the JL001 strain harboring pBAD33-ulp1 and pBBR1MCS-5-PT7-SUMO-WFCWS-valS were harvested 0min, 1min, 5min, 10min, 20min, 30min, 45min, 60min and 120min after arabinose induction. As control JL001 cells harboring pBAD33 and pBBR1MCS-5-PT7-SUMO-WFCWS-valS were analyzed.

Supplemental Table Table S1 Strains, plasmids and oligonucleotides Strains and plasmids Relevant genotype or description Source or reference Escherichia coli strains DH5αλpir recA1 gyrA (lacIZYA-argF) (80d lac [lacZ] M15) pir RK6 (Macinga et al., 1995)
BW25113 Δ(araD-araB)567, ΔlacZ4787(::rrnB-3), lambda-, rph-1, Δ(rhaD-rhaB)568, hsdR514 (Datsenko and Wanner,
2000)
JL001 BW25113 valS::kan pBBR1MCS-5-PT7-SUMO-WFCWS-valS; Kanr Gmr This study
Plasmids pRedET Red/ET expression plasmid Tetr Gene Bridges
pFRT-PGK-gb2-neo-FRT PCR-template (plasmid DNA) for generating a FRT-flanked PGK-gb2-neo cassette; Kanr
Gene Bridges
pTB145 lacIq PT7::h-ulp1(403-621) pBR/colE1; Ampr (Bendezu et al., 2009) pTB146 lacIq PT7::h-sumo pBR/colE1; Ampr (Bendezu et al., 2009) pBAD33 ori-p15a araC PBAD; Cmr (Guzman et al., 1995) pUlp1 (pBAD33-ulp1) ulp1 in pBAD33; Cmr This study pBBR1MCS-5 oriT mobRK2 pBBR origin of replication lacZα; Gmr (Kovach et al., 1995) pBBR1MCS-5-PT7-SUMO-WFCWS-valS
SUMO-WFCWS-valS in pBBR1MCS-5 This study
pET21b-ValS (wt + mutants) valS cloned into pET21b (Novagen/Merck) expression plasmid
This study
pValS (pQE70-ValS) (wt + mutants) valS cloned into pQE70 (Qiagen) expression plasmid This study
Oligonucleotides
Construction of valS::kan inframe deletion
DvalS-us
GCT ATA AAG CCT ACC GCG TGG CTG GTT TCA ACC TGA ATA CGG CAA CCT GGA AAT AAA ATT AAC CCT CAC TAA AGG GCG
This study
DvalS-ds
GAG ATA AAA AAG GCC GGA GCA TGC TCC GGC CTT CGT TTT CAT CAC TGT GTT TTG ATA ATA CGA CTC ACT ATA GGG CTC
This study
valS-chk-Fw CGC AAA AGC GTA GCA GCA GC This study
valS-chk-Rev TGG GCG ATT TGG ATG ACC TTC C This study
Construction of pBBR1MCS-5-PT7-SUMO-WFCWS-valS
T7 Promoter Primer TAA TAC GAC TCA CTA TAG GG
SUMO-WFCWS-ValS-OL-Rev
GCT CCA GCA AAA CCA ACC ACC AAT CTG TTC TCT GTG AGC
This study
SUMO-WFCWS-ValS-OL-Fw
TGG TTT TGC TGG AGC GAA AAG ACA TAT AAC CCA CAA GAT ATC GA
This study
XmaI-ValS-Rev GCA GTC ACC CGG GTT ACA GCG CGG CGA TAA CAG C This study
ValS-636-Seq-Fw GGT AAA GAT TAT CTG GTG GTC GCG This study
ValS-1378-Seq-Fw TCG GTG CTG ATG TTG TCC TGC This study
ValS-2091-Seq-Fw CGA TAT CGC CGC AGG CAT TC This study
Construction of pBAD33-ulp1
SacI-His6-ulp1-Fw GCT GCG GAG CTC AAT AAT TTT GTT TAA CTT TAA GAA GGA GAT ATA CC
This study
XmaI-ulp1-Rev GCG TCC CGG GTT ATT TTA AAG CGT CGG TTA AAA TCA AAT GG
This study
pBAD-Fw GGC GTC CAC ACT TTG CTA TGC (Lassak et al., 2010)
pBAD-Rev CAG TTC CCT ACT CTC GCA TG (Lassak et al., 2010)
Construction of ET21b-valS and pQE70-valS: wt and mutants
FOR valS pQE70 GAA TTC ATT AAA GAG GAG AAA TTA AGC ATG CTT GAA AAG ACA TAT AAC CCA CAA G
This study
REV valS pQE70 CAG GAG TCC AAG CTC AGC TAA TTA AGC TTT TAC AGC GCG GCG ATA ACA GCC TG
This study
FOR pET21b-valS GTT TAA CTT TAA GAA GGA GAT ATA CAT ATG GAA AAG ACA TAT AAC CCA CAA G
This study
REV pET21b-valS GGC CGC AAG CTT GTC GAC GGA GCT CCC CAG CGC GGC GAT AAC AG CCT G
This study
FOR GGG valS GAA AGT TTC TGC ATC ATG ATC GGT GGT GGT AAC GTC This study

ACC GGC AGT TTG CA
REV GGG valS TGC AAA CTG CCG GTG ACG TTA CCA CCA CCG ATC ATG ATG CAG AAA CTT TC
This study
FOR GPP valS GAA AGT TTC TGC ATC ATG ATC GGT CCG CCG AAC GTC ACC GGC AGT TTG CA
This study
REV GPP valS TGC AAA CTG CCG GTG ACG TTC GGC GGA CCG ATC ATG ATG CAG AAA CTT TC
This study
FOR PGP valS GAA AGT TTC TGC ATC ATG ATC CCG GGT CCG AAC GTC ACC GGC AGT TTG CA
This study
REV PGP valS TGC AAA CTG CCG GTG ACG TTC GGA CCC GGG ATC ATG ATG CAG AAA CTT TC
This study
FOR PPG valS GAA AGT TTC TGC ATC ATG ATC CCG CCG GGT AAC GTC ACC GGC AGT TTG CA
This study
REV PPG valS TGC AAA CTG CCG GTG ACG TTA CCC GGC GGG ATC ATG ATG CAG AAA CTT TC
This study
FOR valS T222P GATTATCTGGTGGTCGCGACTCCCCGTCCAGAAACCCTGCTGGG
This study
REV valS T222P CCCAGCAGGGTTTCTGGACGGGGAGTCGCGACCACCAGATAATC
This study
pQE70-Seq-Fw CGA GGC CCT TTC GTC TTC ACC This study
pQE70-Seq-Rev AGG TCA TTA CTG GAT CTA TCA ACA GG This study
ValS-APP-OL-Fw TGC ATC ATG ATC GCG CCG CCG AAC GTC This study
ValS-APP-OL-Rev GAC GTT CGG CGG CGC GAT CAT GAT GCA This study
ValS-CPP-OL-Fw TGC ATC ATG ATC TGC CCG CCG AAC GTC This study
ValS-CPP-OL-Rev GAC GTT CGG CGG GCA GAT CAT GAT GCA This study
ValS-DPP-OL-Fw TGC ATC ATG ATC GAT CCG CCG AAC GTC This study
ValS-DPP-OL-Rev GAC GTT CGG CGG ATC GAT CAT GAT GCA This study
ValS-EPP-OL-Fw TGC ATC ATG ATC GAA CCG CCG AAC GTC This study
ValS-EPP-OL-Rev GAC GTT CGG CGG TTC GAT CAT GAT GCA This study
ValS-FPP-OL-Fw TGC ATC ATG ATC TTT CCG CCG AAC GTC This study
ValS-FPP-OL-Rev GAC GTT CGG CGG AAA GAT CAT GAT GCA This study
ValS-HPP-OL-Fw TGC ATC ATG ATC CAT CCG CCG AAC GTC This study
ValS-HPP-OL-Rev GAC GTT CGG CGG ATG GAT CAT GAT GCA This study
ValS-IPP-OL-Fw TGC ATC ATG ATC ATT CCG CCG AAC GTC This study
ValS-IPP-OL-Rev GAC GTT CGG CGG AAT GAT CAT GAT GCA This study
ValS-KPP-OL-Fw TGC ATC ATG ATC AAA CCG CCG AAC GTC This study
ValS-KPP-OL-Rev GAC GTT CGG CGG TTT GAT CAT GAT GCA This study
ValS-LPP-OL-Fw TGC ATC ATG ATC CTG CCG CCG AAC GTC This study
ValS-LPP-OL-Rev GAC GTT CGG CGG CAG GAT CAT GAT GCA This study
ValS-MPP-OL-Fw TGC ATC ATG ATC ATG CCG CCG AAC GTC This study
ValS-MPP-OL-Rev GAC GTT CGG CGG CAT GAT CAT GAT GCA This study
ValS-NPP-OL-Fw TGC ATC ATG ATC AAC CCG CCG AAC GTC This study
ValS-NPP-OL-Rev GAC GTT CGG CGG GTT GAT CAT GAT GCA This study
ValS-QPP-OL-Fw TGC ATC ATG ATC CAG CCG CCG AAC GTC This study
ValS-QPP-OL-Rev GAC GTT CGG CGG CTG GAT CAT GAT GCA This study
ValS-RPP-OL-Fw TGC ATC ATG ATC CGC CCG CCG AAC GTC This study
ValS-RPP-OL-Rev GAC GTT CGG CGG GCG GAT CAT GAT GCA This study
ValS-SPP-OL-Fw TGC ATC ATG ATC AGC CCG CCG AAC GTC This study
ValS-SPP-OL-Rev GAC GTT CGG CGG GCT GAT CAT GAT GCA This study
ValS-TPP-OL-Fw TGC ATC ATG ATC ACC CCG CCG AAC GTC This study
ValS-TPP-OL-Rev GAC GTT CGG CGG GGT GAT CAT GAT GCA This study

ValS-VPP-OL-Fw TGC ATC ATG ATC GTG CCG CCG AAC GTC This study
ValS-VPP-OL-Rev GAC GTT CGG CGG CAC GAT CAT GAT GCA This study
ValS-WPP-OL-Fw TGC ATC ATG ATC TGG CCG CCG AAC GTC This study
ValS-WPP-OL-Rev GAC GTT CGG CGG CCA GAT CAT GAT GCA This study
ValS-YPP-OL-Fw TGC ATC ATG ATC TAT CCG CCG AAC GTC This study
ValS-YPP-OL-Rev GAC GTT CGG CGG ATA GAT CAT GAT GCA This study

Supplemental Experimental Procedures
Oligonucleotides, plasmids and bacterial strains
Primers, plasmids and strains used in this study are listed in Table S1. Strain ΔvalS
was constructed from BW25113 harboring the pBBR1MCS-5-PT7-SUMO-WFCWS-
valS by using the pRED®/ET® recombination technology in accordance to the
technical protocol of the Quick & Easy E. coli Gene Deletion Kit of Gene Bridges
(www.genebridges.com). Construction of plasmids encoding ulp1 and PT7-SUMO-
WFCWS-ValS, respectively, are described in Table S1.
Molecular biology methods
Enzymes and kits were used according to the manufacturer´s directions. Genomic
DNA was obtained by using the DNeasy blood and tissue kit (Qiagen) and plasmid
DNA was isolated using a Hi-Yield plasmid Mini Kit (Suedlabor). DNA fragments
were purified from agarose gels by employing a High-Yield PCR cleanup and gel
extraction kit (Suedlabor). Restriction endonucleases were purchased from New
England Biolabs (NEB). Sequence amplifications by PCR were performed utilizing
the Phusion high-fidelity DNA polymerase (Finnzymes) or the Taq DNA polymerase
(NEB), respectively. Standard methods were performed according to Sambrook
(Sambrook et al., 1989).
Purification of ValS
The valS gene was cloned into pET21b using NdeI and SacI restriction sites and then
valS mutants were generated using site directed mutagenesis strategy. C-term-His-
tagged-ValS proteins were expressed in BL21(pLysS) and purified using Protino Ni-
NTA agarose beads (Macherey-Nagel) and HiLoad 16/60 Superdex75 preparative gel
filtration column (GE Healthcare) using 50 mM Hepes pH 7.8, 50 mM KCl, 100 mM
NaCl, 10 mM MgCl2, 2% glycerol, 5 mM β-mercaptoethanol
tRNAVal charging assay
Charging reactions were performed as described (Splan et al., 2008). Briefly, each
reaction (50 µL) contained 500 pmol [14C]-Valine (Perkin Elmer) or [14C]-Threonine
(Hartman Analytics), 100 µmol ATP, 7.5 pmol ValS in 100 mM Hepes pH 7.4,

20 mM KCl, 15 mM MgCl2, 10 mM DTT and 50 pmol tRNAVal (Sigma Aldrich).
Each reaction was incubated at 37C for 1-30 min and then quenched with 5 µL of
3 M sodium acetate pH 5.0. Excess of free amino acid was removed using Amicon
Ultra 3.5K (Merck-Millipore) (3x wash with cold buffer) and then the radioactivity of
tRNA samples was measured using scintillation counting.
Monitoring of PPi and AMP formation
Assays to monitor aminoadenylate formation were performed as previously described
(Splan et al., 2008). Briefly, reactions contained 100 µmol amino acid, 600 pmol
ATP, 2 µCi α[32P]-ATP or [32P]-ATP (Hartmann Analytic), 15 pmol ValS in
100 mM Hepes pH 7.4, 20 mM KCl, 15 mM MgCl2, 10 mM DTT and 400 pmol
tRNAVal (where indicated) in 50 µL. Reactions were incubated at 37C for 5 min and
then quenched with 5 µL of 3 M sodium acetate pH 5.0. Where indicated, 0.1 U of
Inorganic Pyrophosphatase from E. coli (NEB) or 5 mU of Apyrase (NEB)
(supplemented with Apyrase buffer) was added. ATP/ADP/AMP standards were
prepared using dilutions of Apyrase (0/0.05/0.5/5 mU). Thin-layer chromatography
was performed using 20 x 10 cm TLC PEI Cellulose F plates (Merck-Millipore),
which were pre-equilibrated in water. Samples (0.5 µL) were spotted 1 cm from the
bottom edge of the plate and the nucleotides separated by running in tanks containing
0.15 M formic acid-0.15 M LiCl. Plates were then dried and exposed to Amersham
Hyperfilm MP (GE Healthcare) for >1h.
Western blotting analysis
To monitor SUMO-ValS degradation and Ulp1 production, exponentially grown cells
were harvested at certain times after arabinose induction. Cells were resuspended in
water yielding an OD600nm of 10. 10 µl of the cell suspension was mixed with an
equivalent volume of Laemmli buffer and boiled for 5min before separation by
sodium dodecyl sulfate (SDS) polyacrylamide gel electrophoresis (PAGE) using
12.5% gels (Laemmli, 1970). SUMO-ValS and Ulp1 were detected by Western
immunoblotting according to standard protocols (Sambrook et al., 1989) using mouse
monoclonal anti 6xHis tag antibody coupled to alkaline phosphatase (Abcam).

ValS-XPP library
pQE70-ValS-XPP library was constructed as essentially described in van den Ent &
Loewe (van den Ent and Lowe, 2006) after generation of a mutant fragment of valS
by overlap extension PCR (Lassak et al., 2010). valS::kan PT7-SUMO-valS was
transformed with pBAD33 or pBAD33-ulp1 and a pQE70-ValS-XPP variant.
Overnight (ON) cultures of transformants were grown to exponential phase in LB
under repressing conditions (+0.2% Glucose). Subsequently cells were washed and
diluted to OD 600nm= 0.05 into fresh medium lacking Glucose but containing 0.2%
arabinose. Growth was recorded by measuring OD600nm after 1.5h incubation at
37°C.
Complementation analysis
JW4106 (Δefp) was transformed with pQE70-vectors containing either a copy of efp
or ValS-PPP, ValS-GPP or ValS-GGG. ON cultures of transformants were grown to
exponential phase in LB under repressing conditions (+0.2% Glucose). Subsequently,
cells were washed and diluted into fresh medium lacking Glucose but containing
1mM IPTG. Doubling times were calculated from exponentially grown cells.
Supplemental References
Baba, T., Ara, T., Hasegawa, M., Takai, Y., Okumura, Y., Baba, M., Datsenko, K.A.,
Tomita, M., Wanner, B.L., and Mori, H. (2006). Construction of Escherichia coli
K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol
2, 2006 0008.
Bendezu, F.O., Hale, C.A., Bernhardt, T.G., and de Boer, P.A. (2009). RodZ (YfgA)
is required for proper assembly of the MreB actin cytoskeleton and cell shape in
E. coli. The EMBO journal 28, 193-204.
Datsenko, K.A., and Wanner, B.L. (2000). One-step inactivation of chromosomal
genes in Escherichia coli K-12 using PCR products. Proceedings of the National
Academy of Sciences of the United States of America 97, 6640-6645.
Guzman, L.M., Belin, D., Carson, M.J., and Beckwith, J. (1995). Tight regulation,
modulation, and high-level expression by vectors containing the arabinose PBAD
promoter. Journal of bacteriology 177, 4121-4130.

Kovach, M.E., Elzer, P.H., Hill, D.S., Robertson, G.T., Farris, M.A., Roop, R.M.,
2nd, and Peterson, K.M. (1995). Four new derivatives of the broad-host-range
cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes.
Gene 166, 175-176.
Laemmli, U.K. (1970). Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature 227, 680-685.
Lassak, J., Henche, A.L., Binnenkade, L., and Thormann, K.M. (2010). ArcS, the
cognate sensor kinase in an atypical Arc system of Shewanella oneidensis MR-1.
Applied and environmental microbiology 76, 3263-3274.
Macinga, D.R., Parojcic, M.M., and Rather, P.N. (1995). Identification and analysis
of aarP, a transcriptional activator of the 2'-N-acetyltransferase in Providencia
stuartii. Journal of bacteriology 177, 3407-3413.
Peil, L., Starosta, A.L., Lassak, J., Atkinson, G.C., Virumae, K., Spitzer, M., Tenson,
T., Jung, K., Remme, J., and Wilson, D.N. (2013). Distinct XPPX sequence
motifs induce ribosome stalling, which is rescued by the translation elongation
factor EF-P. Proc Natl Acad Sci U S A 110, 15265-15270.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular Cloning, a laboratory
manual: 2nd edition (Cold Spring Harbour, NY: Cold Spring Harbour Laboratory
Press).
Splan, K.E., Musier-Forsyth, K., Boniecki, M.T., and Martinis, S.A. (2008). In vitro
assays for the determination of aminoacyl-tRNA synthetase editing activity.
Methods 44, 119-128.
van den Ent, F., and Lowe, J. (2006). RF cloning: a restriction-free method for
inserting target genes into plasmids. Journal of Biochemical and Biophysical
Methods 67, 67-74.
Copyright © 2022 FDOKUMEN