
Solid state and thermal behavior of 17-estradiol in ammonioethyl methacrylate ester copolymer
Upload
independentCategory
view
0download
0
BIOLOGY OF REPRODUCTION 78, 101–114 (2008)Published online before print 10 October 2007.DOI 10.1095/biolreprod.107.063909
17Beta-Estradiol Induces the Translocation of the Estrogen Receptors ESR1 and ESR2to the Cell Membrane, MAPK3/1 Phosphorylation and Proliferation of CulturedImmature Rat Sertoli Cells1
Thaıs F.G. Lucas,3 Erica R. Siu,3 Carlos A. Esteves,3 Hugo P. Monteiro,4 Cleida A. Oliveira,5
Catarina S. Porto,3 and Maria Fatima M. Lazari2,3
Section of Experimental Endocrinology,3 Department of Pharmacology, and Department of Biochemistry,4
Universidade Federal de Sao Paulo, Escola Paulista de Medicina, Sao Paulo 04044-020, BrazilDepartment of Morphology,5 Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais 31270-901, Brazil
ABSTRACT
The aim of the present study was to determine themechanisms involved in estrogen actions in cultured rat Sertolicells. RT-PCR detected transcripts for the estrogen receptorsESR1 and ESR2 in cultured immature Sertoli cells and in thetestis of 15-, 28-, and 120-day-old rats. The expression of ESR1and ESR2 was confirmed in Sertoli cells by immunofluorescenceand Western blot. Immunohistochemistry with cryosections oftestes from immature and adult rats revealed that ESR1 is presentin Sertoli, Leydig, and some peritubular myoid cells, and ESR2 ispresent in multiple cell types, including germ cells. Treatment ofSertoli cells with 17beta-estradiol (E
2) induced a translocation of
ESR1 and ESR2 to the plasma membrane and a concomitantphosphorylation of MAPK3/1. Both effects reached a maximumafter 10 min and were blocked by PP2, an inhibitor of the SRCfamily of protein tyrosine kinases, and by the antiestrogen ICI182,780 (ICI). MAPK3/1 phosphorylation was also decreased inthe presence of AG 1478, an inhibitor of the epidermal growthfactor receptor (EGFR) kinase, and in the presence of MAP2K1/2inhibitor UO126. Treatment with E
2for 24 h increased the
incorporation of [methyl-3H]thymidine, which was blocked byICI. These results indicate that E
2activates an SRC-mediated
translocation of estrogen receptors to the plasma membrane,which results in the activation of EGFR and the mitogen-activated protein kinase signaling pathway. In addition, activa-tion of ESR1 and/or ESR2 by E
2is involved in proliferation of
immature Sertoli cells. The estrogen actions in Sertoli cells mightbe a key step mediating cellular events important for spermato-genesis and fertility.
estradiol, estradiol receptor, kinases, mechanisms of hormoneaction, Sertoli cells
INTRODUCTION
Estrogen plays an important role in the development,differentiation, and growth of the male reproductive system[1–6]. Estrogens are synthesized from androgens by thearomatase complex, which contains the cytochrome P450enzyme encoded by the CYP19 gene [6, 7]. Aromataseexpression is detected in Leydig and Sertoli cells, spermato-gonia, spermatocytes, elongate spermatids, and spermatozoa inadult mice and rats [3, 7] and in Sertoli cells isolated fromimmature rat testis [8].
In order to mediate its biological effects, estrogen interactswith specific receptors ESR1 and ESR2 (also known as ERaand ERb, respectively), which are widely distributed in themale reproductive tract [4, 9–11]. In the testis of adult rats andmice, ESR1 immunostaining is detected in Leydig andperitubular myoid cells [3, 4, 9, 11–13]. Several studiesreported the absence of ESR1 in germ cells, but Pelletier et al.[14] detected ESR1 in spermatocytes and spermatids. Althoughmost of the studies did not find a positive ESR1 immuno-staining in Sertoli cells, a faint and diffuse staining, interpretedas an artifact, was detected in seminiferous cords fromimmature rats [9] and baboon testis [15], and a positiveimmunostaining was detected in Sertoli cells of immature andmature boars [16, 17]. ESR2 expression is detected in multiplecell types, including the Sertoli cells, and in some but not allgerm cells [12, 13]. Recently, ESR1 mRNA and proteinexpression has been shown in the SK11 cell line derived fromSertoli cells of 10-day-old H2Kb-tsA58 transgenic mice [18].
The mechanisms by which estrogens influence male fertilityremain uncertain. Estrogen effects on reproductive functionhave been studied using mice with targeted disruptionof estrogen receptors (Esr1�/�, Esr2�/�, Esr1�/�/Esr2�/�),aromatase enzyme (Cyp19�/�), and animals treated with theantiestrogen ICI 182,780 (ICI) [3, 5, 11, 19, 20]. The studieswith knockout animals have shown that the spermato-genesis, steroidogenesis, and fertility of Esr1�/�, Esr1�/�/Esr2�/�, and Cyp19�/� animals are affected, but none of theseparameters is affected in Esr2�/� animals. In fact, infertility inEsr1�/� seems to be mainly due to disruption of fluidreabsorption in efferent ductules, and the backpressure of theaccumulating luminal fluids leads to a progressive degenerationof the testicular tissue and to dilation of seminiferous tubules[3, 21]. There is evidence that estrogen also affects Sertoli cellproliferation and may suppress differentiation [19], but nochanges were observed in the number of Sertoli cells andspermatogonia in Esr1�/� mice [20]. On the other hand, ESR2inactivation increases the number of spermatogonias by morethan 50% in neonatal mice [22]. It is surprising that, in spite ofthe evidence that ESR2 regulates mitosis of spermatogonia,
1Supported by Fundacao de Amparo a Pesquisa do Estado de Sao Paulo(FAPESP, grant 2004/01152-0, to C.S.P.). Research fellowship (C.S.P.)was supported by Conselho Nacional de Desenvolvimento Cientifico eTecnologico, CNPq. Doctoral fellowships supported by CNPq (T.F.G.L.)and FAPESP (E.R.S.). Master fellowship supported by Coordenacao deAperfeicoamento de Pessoal de Nıvel Superior, CAPES (C.A.E.).2Correspondence: Maria Fatima M. Lazari, Section of ExperimentalEndocrinology, Department of Pharmacology, Universidade Federal deSao Paulo, Escola Paulista de Medicina, Rua Tres de maio 100, INFAR,Vila Clementino, Sao Paulo, SP 04044-020, Brazil.FAX: 5511 5576 4448; e-mail: [email protected]
Received: 2 July 2007.First decision: 28 July 2007.Accepted: 8 October 2007.� 2008 by the Society for the Study of Reproduction, Inc.ISSN: 0006-3363. http://www.biolreprod.org
101

disturbances of sperm production were not evident in Esr2�/�
mice [23]. Paradoxically, spermatogenic arrest occurs inEsr1�/� mice [21], suggesting that testicular cells regulateSertoli cell support of germ cell development throughunidentified ESR1-mediated mechanisms. This line of evi-dence is supported by experiments in which germ cells fromdonor males homozygous for the mutation Esr1�/� weretransplanted to testes of wild-type Esr1þ/þ recipient micedepleted of germ cells. When mated to wild-type females, therecipients sired offspring heterozygous for the mutationEsr1þ/�, but retained the coat-color marker of Esr1�/� donormice. This finding confirmed that somatic cells in the testis, butnot germ cells, require ESR1 in order to support the process ofspermatogenesis [24, 25].
In addition to the classic genomic mechanism of estrogenaction, mediated by ESR1 and ESR2, there is now convincingevidence that the steroid also exerts rapid, nongenomic actionsinitiated at the cell surface. The rapid action of estrogen can bedivided into two major categories: classical receptor-mediatedresponses, which involve translocation of estrogen receptorsfrom nuclei to the plasma membrane [26, 27], and nonclassicalresponses, mediated through proteins other than estrogenreceptors, such as the G-protein coupled receptor GPR30[28–32]. Estrogen nongenomic signaling involves a series ofcell type-dependent events that includes mobilization of secondmessengers, interaction with membrane receptors such asinsulin-like growth factor 1 receptor (IGF1R) and epidermalgrowth factor receptor (EGFR), and stimulation of effectormolecules, such as the SRC family of tyrosine kinases and thephosphatidylinositol 3-Kinase, the serine/threonine proteinkinase (AKT), and the mitogen-activated protein kinases(MAPKs) [26, 27, 33–35].
Because the physiological role of estrogen in Sertoli cells isnot completely understood and there is some controversy inthe literature about ESR1 expression in these cells, the aim ofthe present study was to investigate the expression of ESR1and ESR2 and the effects of 17b-estradiol (E
2) on the
subcellular distribution, signaling, and function of thesereceptors in rat Sertoli cells.
MATERIALS AND METHODS
Cell Culture
Primary cultures of Sertoli cells were obtained from 15-day-old male Wistarrats housed in the Animal Facility at Instituto de Farmacologia e BiologiaMolecular (INFAR), Universidade Federal de Sao Paulo-Escola Paulista deMedicina (UNIFESP-EPM), and maintained on a 12L:12D lighting schedule at238C with food and water ad libitum. The experimental procedures wereconducted according to guidelines for the care and use of laboratory animals asapproved by the Research Ethical Committee from UNIFESP-EPM. The testeswere removed and decapsulated, and Sertoli cells were prepared as previouslydescribed [36–38]. Cells were plated at a density of approximately 4 3 106 cellsper milliliter in phenol-red free Ham F12/Dulbecco modified Eagle medium(F12/DMEM 1:1; Gibco; Invitrogen, Grand Island, NY) containing 0.02 g/Lgentamicin (pH 7.2–7.4; Sigma Chemical Co., St. Louis, MO) andsupplemented with 10 lg/ml insulin, 10 lg/ml transferrin, 10 ng/ml sodiumselenite, and 10 ng/ml epidermal growth factor (EGF; Sigma). The cells weregrown in a humidified atmosphere of 5% CO
2:95% air at 358C and, after 48 h,
treated with 20 mM Tris-HCL (pH 7.4) to lyse residual germ cells [39], andallowed to grow for another 24 h. Culture medium was replaced by another onewithout supplements 20 h before the experiments with Sertoli cells. At thisstage, the cells were 90%–95% confluent, and the amount of viable cells ineach culture, as determined by trypan blue exclusion, was more than 90%. Forproliferation assays, Sertoli cells were prepared as described above and platedat a low density, and at the day of experiments the confluence was 50%–60%.
The presence of other cell types in the primary culture of Sertoli cells wasevaluated by several criteria. Cells were grown on coverslips coated withgelatin (0.1%) for both morphological and immunocytochemistry analyses.Under a phase-contrast microscope, Sertoli cells were the predominant cell type
after hypotonic shock (data not shown). Hematoxylin-eosin staining showedthat, under the described culture conditions, the cells presented thecharacteristic rounded nuclei and irregular polygonal shape with well-formedcytoplasmic extensions (Fig. 1A), as previously reported for cultured Sertolicells [40]. Further analyses included the immunocytochemical detection of twomajor proteins secreted by Sertoli cells: transferrin and sulfated glycoprotein 2,also known as clusterin (CLU) [41, 42]. Transferrin and CLU were detected byusing a rabbit polyclonal antibody against human transferrin (1:50 dilution;DAKO, Carpinteria, CA) and a mouse monoclonal antibody against rat CLU(1:50 dilution, clone 6E9) [43], and secondary antibodies Alexa Fluor488-labeled anti-rabbit and Alexa Fluor 594-labeled anti-mouse, respectively(1:300 dilution; Molecular Probes, Eugene, OR; Fig. 1, B and C). All theseanalyses confirmed the absolute predominance of Sertoli cells in the primaryculture.
RT-PCR Assays
Total RNA was extracted with the TRIzol reagent (Invitrogen, Carlsbad,CA) from testes of 15-, 28-, and 120-day-old rats and from Sertoli cells culturedfor 5 days, according to the standard protocol [44], and checked for ribosomalRNA integrity by agarose gel electrophoresis. A 2:1 ratio of sharp and clearethidium-bromide stained 28S:18S ribosomal RNA bands was observed for allsamples. Total RNA (5 lg) was used to synthesize the first strand cDNA at428C with the ThermoScritp RT-PCR kit (Invitrogen) and the random hexamerssupplied with the kit. Reactions in the absence of reverse transcriptase werealso included for each RNA tested in order to check for genomiccontamination. The resulting cDNA was used in PCRs with primers(Invitrogen) designed to amplify specific regions of the different estrogenreceptors (Table 1). Esr2_v2 (ERb2) refers to the splice variant 2 of Esr2 foundonly in rodents, which differs from the full-length wild-type variant 1, Esr2_v1(ERb1), because it contains an insertion of 54 bp in the ligand-binding domain[45]. This variant has also been called bins [46] and differs from the ERbcx/b2splice variant found in humans [47], which presents a truncation at thecarboxyterminal region that leads to the loss of 61 amino acids. Theamplification of the housekeeping gene glyceraldehyde-3-phosphate dehydro-genase (Gapdh) was used as an internal control using the following primers:forward 50-CGGGAAGCTTGTGATCAATGG-30 and reverse 50-GGCAGT-GATGCCATGGACTG-30 [48]. The PCR mixture consisted of 2 ll of cDNA,1.5 mM MgCl
2, 0.2 mM dNTP, 0.5 lM of each primer, and 2.5 U of Taq DNA
polymerase (Invitrogen) in the appropriate PCR buffer. Samples were amplifiedin an Applied Biosystem 9600 Fast Thermal Cycler (Applied Biosystems,Foster City, CA). The thermal cycles for amplification of the Esr1 transcriptwere: a denaturation cycle at 948C for 5 min, a touch-down protocol of threecycles at annealing temperature 758C for 1 min, three cycles at annealingtemperature 708C for 1 min, 35 cycles at annealing temperature 658C for 1 min,and a final extension of 728C for 7 min. For amplification of Esr2_v1 andEsr2_v2 transcripts, the following thermal cycles were used: a denaturationcycle at 948C for 5 min, a touch down protocol of three cycles at annealingtemperature 758C for 1 min, three cycles at annealing temperature 708C for 1min, three cycles at annealing temperature 658C for 1 min, 35 cycles at annealingtemperature 608C for 1 min, and a final extension of 728C for 7 min. Foramplification of the Gapdh transcript, a denaturation cycle at 948C for 5 min, 35cycles at 948C for 5 min, 628C for 1 min, 728C for 2 min, and a final extensioncycle of 728C for 7 min were used. The PCR products were resolved ontoagarose gels (1.5%) containing ethidium bromide (0.5 lg/ml) and visualizedunder UV transillumination. After extraction from the gel (gel extration kit;QIAgen, Hilden, Germany), the identity of each product was confirmed bydirect nucleotide sequencing in an ABI PRISM 377 automated sequencer(Applied Biosystems) using the DYEnamic ET Terminator Sequencing kit(Amersham Biosciences, Piscataway, NJ).
Immunohistochemistry
Testes from 15- and 120-day-old rats were removed, embedded in JungTissue Freezing Medium (Leica Instruments, Nussloch, Germany), frozen indry, ice-cold acetone, and maintained at�758C until use. Frozen sections (8-lmthickness) of testis were cut at�208C in a cryostat (Microm HM550; MicromInternational GmbH, Walldorf, Germany) and mounted on silanized slides(DAKO). Sections were air dried, fixed in 4% (v/v) paraformaldehyde(Electron Microscopy Sciences, Hatfield, PA) diluted in PBS (137 mM NaCl,2.68 mM KCl, 6.03 mM Na
2HPO
4, and 1.47 mM KH
2PO
4; pH 7.4; Sigma)
[10] for 20 min at room temperature, and then incubated with 0.1 M glycine(Sigma). In order to block the endogenous peroxidase activity, slides weretreated with 3% (v/v) H
2O
2and washed with PBS. Nonspecific antibody
binding was minimized by incubation with blocking solution (PBS containing0.01% saponin and 3% BSA, Sigma) for 1 h at room temperature, and theendogenous biotin was blocked by incubation with Avidin/Biotin Blocking Kit
102 LUCAS ET AL.

(Vector Laboratories, Inc., Burlingame, CA). Slides were incubated with one ofthe following primary antibodies (Santa Cruz Biotechnology, San Diego, CA):rabbit polyclonal antibody raised against the aminoterminal region of ESR1 ofhuman origin (H184), rabbit polyclonal antibody raised against the carboxy-terminal region of ESR1 of mouse origin (MC20), and goat polyclonal antibodyraised against the aminoterminal region of ESR2 of mouse origin (Y19), at 1:50dilution in blocking solution overnight at 48C. Afterwards, slides were washedwith PBS and blocking solution, and incubated with the appropriatebiotinylated secondary antibody (Santa Cruz Biotechnology): donkey anti-rabbit IgG (for ESR1) or rabbit anti-goat IgG (for ESR2) at 1:300 dilution inblocking solution for 1 h at room temperature. The immunoreaction wasvisualized using avidin-biotin complex (Vector Laboratories) for 90 min atroom temperature, followed by several washes with PBS and incubation with0.05% (w/v) 3,30diaminobenzidine containing 0.01% (v/v) H
2O
2in PBS. Slides
were mounted with Permount (Fisher Scientific, Fair Lawn, NJ). Negativecontrol slides were performed either with the primary antibody preadsorbedwith its respective blocking peptide or in the absence of primary antibody. The
sections were visualized with a Nikon E800 microscope (Nikon, Melville, NY).Images were processed using a CoolSNAP-Pro CCD digital camera and theImage-Pro Express Software Program (Media Cybernetics, Silver Spring, MD).
Immunofluorescence
Sertoli cells were grown as described above on coverslips coated withgelatin (0.1%) and placed into six-well plates. Cells were incubated in theabsence (control, C) and presence of E
2(0.1 nM; Sigma) for 5, 10, 15, and 30
min. In another series of experiments, the cells were untreated or pretreatedwith ICI (1 nM; Faslodex; AstraZenica, Sao Paulo, Brazil) or the selectiveinhibitor of the SRC family of protein tyrosine kinases PP2 (4-amino-5-(4-chlorophenyl)-7-(t-butyl) pyrazolo[3,4-d]pyrimidine; 5 nM; Calbiochem,Darmstadt, Germany) for 30 min at 358C. Afterwards, the cells were stimulatedwith E
2(0.1 nM, 10 min). Concentrations of E
2and ICI were chosen from
preliminary experiments, in agreement with previous reports [49, 50]. Medium
TABLE 1. Oligonucleotides used for RT-PCR.
Transcript (accession no.) Pair of primers Orientation Sequence (50–30) Nucleotide position
Esr1 (Y00102) 1 Forward TCCAAGGGAACGAGCTGGAG 262 to 281Reverse TACGAAAAGACCGCCGAGGA 1007 to 988
Esr2_v1 and Esr2_v2 (U57439) 2a Forward AACCGCCATGAGTATTCAGC 396 to 415Reverse GGCCATCACTGAGACTGTAGGTTC 1879 to 1856
Esr2_v2 (AF042059) 3 Forward AACCGCCATGAGTATTCAGC �21 to �3Reverse CTGCGCAACGTGCCAGTGAGG 990 to 970
a Primers are able to amplify both Esr2 variants; the expected size of the amplicons should be 1484 bp for variant 1 and 1538 bp for variant 2.
FIG. 1. Characterization of Sertoli cellculture from 15-day-old rats. Cells werecultured for 5 days, fixed in paraformalde-hyde, and stained with hematoxylin-eosin(A). Morphological studies showed Sertolicells with rounded nuclei (arrows) andirregular polygonal shapes with well-formed cytoplasmic extensions (arrow-heads). Specific immunostaining for trans-ferrin using a rabbit polyclonal antibodyagainst human transferrin (B), and CLUusing a mouse monoclonal antibody againstrat CLU (C) was observed in primary cultureof Sertoli cells. Scale bars as indicated. Thedata shown are representative of at leastthree independent experiments.
ESTROGEN AND MAPK3/1 ACTIVATION IN SERTOLI CELL 103

was removed, the cells were washed with PBS, fixed in 2% paraformaldehydefor 20 min at room temperature, and washed with PBS containing 0.1 Mglycine. Cells were then permeabilized with 0.01% saponin and blocked withPBS containing 1% BSA for 10 min at room temperature. After blocking, cellswere incubated with one of the following primary antibodies: rabbit anti-ESR1antibodies (H184 and MC20), mouse monoclonal antibody raised against thefull-length human ESR1 (NCL-ER-6F11; Novocastra Laboratories, Newcastle,UK) and goat anti-ESR2 antibody (Y19), at 1:50 dilution in PBS containing0.01% saponin and 1% BSA, for 1 h at room temperature. Afterwards, cellswere washed with PBS containing 0.01% saponin and 1% BSA and incubatedwith the appropriate Alexa Fluor 594-labelled secondary antibody (MolecularProbes): anti-rabbit, anti-mouse, or anti-goat antibodies (1:300) for 30 min atroom temperature. Coverslips were washed and mounted on slides withFluoromount G (Electron Microscopy Sciences). Negative control slides wereperformed either with the primary antibody preadsorbed with its respectiveblocking peptide (Santa Cruz Biotechnology) or in the absence of primaryantibody. The slides were also incubated with DAPI (40,60-diamidino-2-phenylindole; Sigma) for cell nuclei visualization. Immunolocalization of thereceptors was visualized under a Nikon E800 fluorescence microscopeequipped with a short arc mercury lamp, respective exciter filters of 330–380nm and 528–553 nm, and respective barrier filters of 435–485 nm and 600–660nm (Nikon). Images were processed with a CoolSNAP-Pro CCD digital cameraand the Image-Pro Express Software Program.
Confocal Laser Scanning Microscopy
Cells grown on coverslips coated with gelatin (0.1%) were incubated in theabsence (control, C) and presence of E
2(0.1 nM; Sigma) for 10 min, and
immunofluorescence assays were performed as described above, with anadditional staining step using either a green nuclear marker (Yo-Pro1;Molecular Probes) or FITC-conjugated lectin from Triticum vulgaris (Sigma),which is also known as wheat germ agglutinin (WGA) and selectively binds toN-acetylglucosamine and sialic acid residues, common to many membraneglycoproteins. Images were obtained using an inverted LSM 510 confocal laserscanning microscope (Carl Zeiss Inc., Oberkochen, Germany). An Ar-ion 488-nm and a He-Ne 543-nm laser were used for excitation of green and redfluorescences, respectively. Serial z-axis sections were collected at a 0.4-lmthickness with a 63X PlanAploChromat objective (Carl Zeiss Inc.). Analysisand reconstruction of confocal microscopy images were performed using thefree software Image J 1.37 [51]. Images are presented as projections of opticalimages stacks.
Western Blot for Detection of Estrogen Receptors
Primary Sertoli cell cultures were washed with ice-cold PBS, lysed in ice-cold lysis buffer (50 mM Tris, 150 mM NaCl, 0.05% sodium deoxycholate, 1%NP-40, 1 mM EDTA, 0.1% SDS, 1 lg/ml aprotinin, 1 lg/ml leupeptin, 1 lg/mlpepstatin A, 1 mM PMSF; pH 8.0), and centrifuged at 11 200 3 g for 30 min at48C. Proteins from supernatant (50 and 100 lg per lane) were incubated withsample buffer containing dithiothreitol and b-mercaptoethanol and subjected to7.5% SDS-PAGE. Proteins were electrotransferred onto PVDF membranes(0.45-lm pore size; ImmobilonP; Millipore, Bedford, MA) overnight, 20 V at48C. Membranes were blocked in Tris-buffered saline (TBS; 10 mM Tris, 150mM NaCl; pH 8.0) containing 0.2% Tween 20 and 5% nonfat dry milk for 2 hat room temperature. ESR1 and ESR2 were detected by probing membraneswith rabbit anti-ESR1 antibodies and goat anti-ESR2 antibody diluted 1:200 inblocking solution, for 2 h at room temperature. Membranes were washed inTBS containing 0.2% Tween 20 (TBS-T), and the antigens bound to primaryantibodies were detected with the appropriate horseradish peroxidase (HRP)-conjugated anti-IgG secondary antibodies (donkey anti-rabbit [1:5000] andrabbit anti-goat [1:4000]; Jackson Immunoresearch Lab, West Grove, PA) for1 h at room temperature. Thereafter, the membranes were washed with TBS-T,and immunoreactive bands were visualized onto preflashed Biomax XAR film(Eastman Kodak Co., Rochester, NY) by enhanced chemiluminescence reagent(Luminol; PerkinElmer, Boston, MA). In negative controls, primary antibodieswere preadsorbed with the respective peptide immunogen and processed asdescribed above. Apparent molecular masses of protein bands were determinedfrom molecular mass standards (prestained protein marker, broad range 6–175kDa; New England Biolabs, Boston, MA).
Western Blot for Detection of MAPK3/1 and Phospho-MAPK3/1
Primary Sertoli cell cultures were incubated in the absence (control, C) andpresence of E
2(0.1 nM; Sigma) for 5–30 min at 358C. In another series of
experiments, the cells were untreated or pretreated with ICI (1 nM), PP2 (5 nM;
Calbiochemical, San Diego, CA), the MAP2K1/2 inhibitor U0126 (100 lM;Cell Signaling Technology, Beverly, MA) for 30 min, or the EGFR kinaseinhibitor AG 1478 (4-(3-chloroanilino)-6,7-dimethoxyquinazoline; 50 lM;Calbiochem) for 15 min at 358C. Afterward, the cells were stimulated with E
2
(0.1 nM, 10 min). EGF (10 ng/ml, 10 min, 358C) was used as control. Mediumwas removed, the cells were washed with ice-cold PBS and lysed in ice-coldlysis buffer (20 mM HEPES [pH 7.5], 150 mM NaCl, 10% glycerol, 1% TritonX-100, 1.5 mM MgCl
2, 1 mM EGTA, 1 lg/ml aprotinin, 1 lg/ml leupeptin, 1
mM PMSF, 2 mM Na3VO
4, 50 mM NaF, and 10 mM Na
4P
20
7), as previously
described [52]. Total cellular proteins (60 lg per lane) were incubated withsample buffer containing b-mercaptoethanol and subjected to 10% SDS/PAGE.Proteins were electrotransferred onto PVDF membranes overnight, 20 V at 48C.Membranes were blocked in TBS-t containing 5% nonfat dry milk (pH 7.6) for2 h at room temperature. After washes in TBS-T, membranes were incubatedwith a rabbit polyclonal antibody raised against a synthetic peptide derivedfrom the sequence of rat MAPK3/1 (p44/p42 MAP kinase, Erk1/Erk2, #9102;Cell Signaling Technology) or anti-phospho-MAPK3/1 antibody (Thr202/Tyr204, #9101; Cell Signaling Technology) diluted in blocking solution(1:1000 and 1:2000, respectively) overnight at 48C. Proteins were visualizedafter incubation with donkey anti-rabbit HRP-conjugated secondary antibody(Amersham Biosciences) diluted in TBS-T (1:3000) for 1 h at roomtemperature by enhanced chemiluminescence reagent (ECL; AmershamBiosciences). Apparent molecular masses of these proteins were determinedfrom molecular mass standards (New England Biolabs).
Band intensities of total MAPK3/1 and phosphorylated MAPK3/1 fromindividual experiments were quantified by densitometric analysis of linear-range autoradiograms by using the Epson Expression 1680 scanner (EpsonAmerica Inc., Long Beach, CA) and Quick Scan 2000 WIN software (HelenaLaboratories, Beaumont, TX). Results were normalized based on total MAPK3/1 expression in each sample and plotted (mean 6 SEM) in relation to control(C¼ 1).
[Methyl-3H]thymidine Incorporation Assays
Incorporation of [methyl-3H]thymidine into cell DNA was estimated asdescribed by Guizzetti et al. [53]. Previous studies in our laboratory indicatedthat [methyl-3H]thymidine (0.074 MBq/ml, specific activity 2923 GBq/ml;Amersham Biosciences) incorporation in cultured Sertoli cells was time-dependent and linear from 2 to 10 h of incubation. All studies were performedusing 6 h of [methyl-3H]thymidine incubation [38].
Primary Sertoli cell cultures were initially incubated with 0.074 MBq/ml[methyl-3H]thymidine for 6 h at 358C. Incubation was continued in the absence(basal incorporation) and presence of E
2(0.1 nM; Sigma) for 24 h at 358C. In
another series of experiments, the cells were treated with ICI (1 nM) for 30 minat 358C before addition of E
2(0.1 nM) for 24 h. The reaction was stopped by
cooling the cells at 08C. The medium was aspirated and the cells rinsed twicewith ice-cold PBS and 5% trichloroacetic acid (Sigma). The cells were thensolubilized with 0.5 N NaOH, collected with cotton-swabs, and transferred to5 ml OptiPhase HiSafe-3 scintillation liquid (PerkinElmer Life ScienceProducts). Bound radioactivity was determined in a scintillation b counter(LS 6000 IC; Beckman Coulter Inc., Palo Alto, CA). Results were expressed aspercentage of [methyl-3H]thymidine incorporation above basal level.
Protein Assays
Protein concentration was determined by Bio-Rad protein assay, using BSAas standard (Bio-Rad Laboratories, Hercules, CA).
Statistical Analysis
Data were expressed as mean 6 SEM. Statistical analysis was carried outby ANOVA followed by the Newman-Keuls test for multiple comparisons orby Student t-test to compare the differences between two data. P , 0.05 wasconsidered significant.
RESULTS
Expression of ESR1 and ESR2 in Rat Testis and CulturedSertoli Cells
RT-PCR experiments detected the presence of Esr1 andEsr2_v2-related transcripts in cultured Sertoli cells from 15-day-old rats and in testes from 15-, 28-, and 120-day-old rats(Fig. 2). A transcript related to Esr2_v1 was also detected incultured Sertoli cells and in testis from 15-day-old animals
104 LUCAS ET AL.
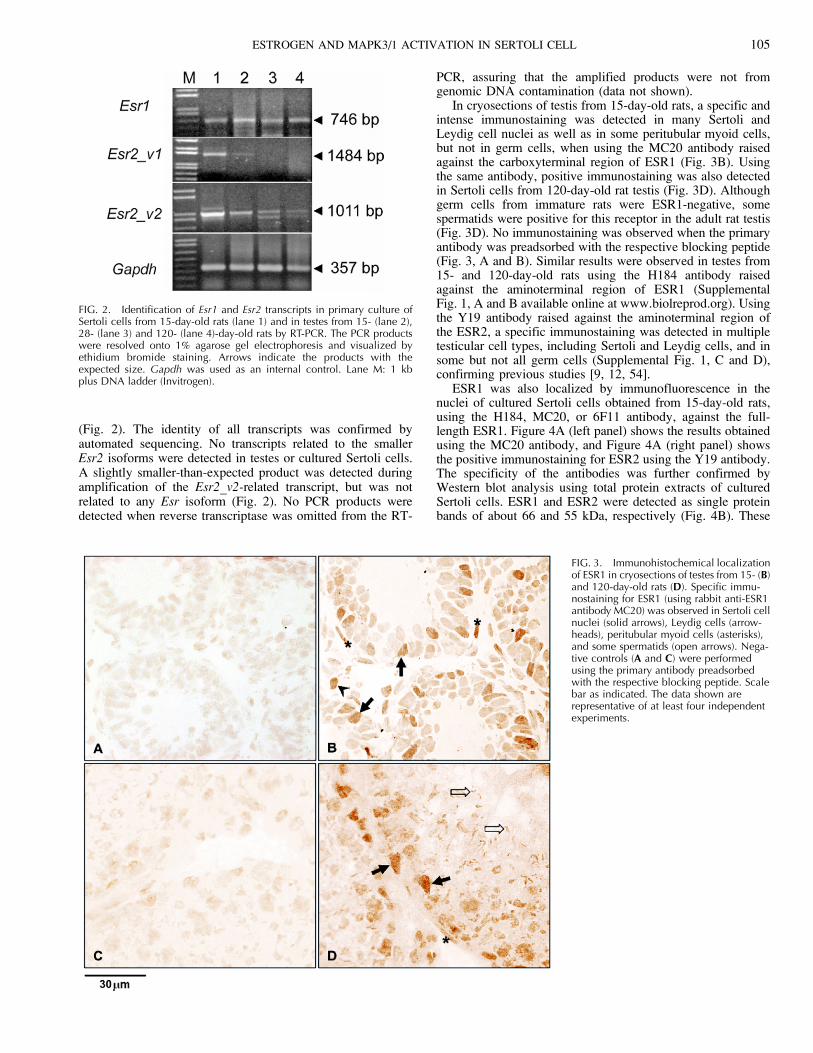
(Fig. 2). The identity of all transcripts was confirmed byautomated sequencing. No transcripts related to the smallerEsr2 isoforms were detected in testes or cultured Sertoli cells.A slightly smaller-than-expected product was detected duringamplification of the Esr2_v2-related transcript, but was notrelated to any Esr isoform (Fig. 2). No PCR products weredetected when reverse transcriptase was omitted from the RT-
PCR, assuring that the amplified products were not fromgenomic DNA contamination (data not shown).
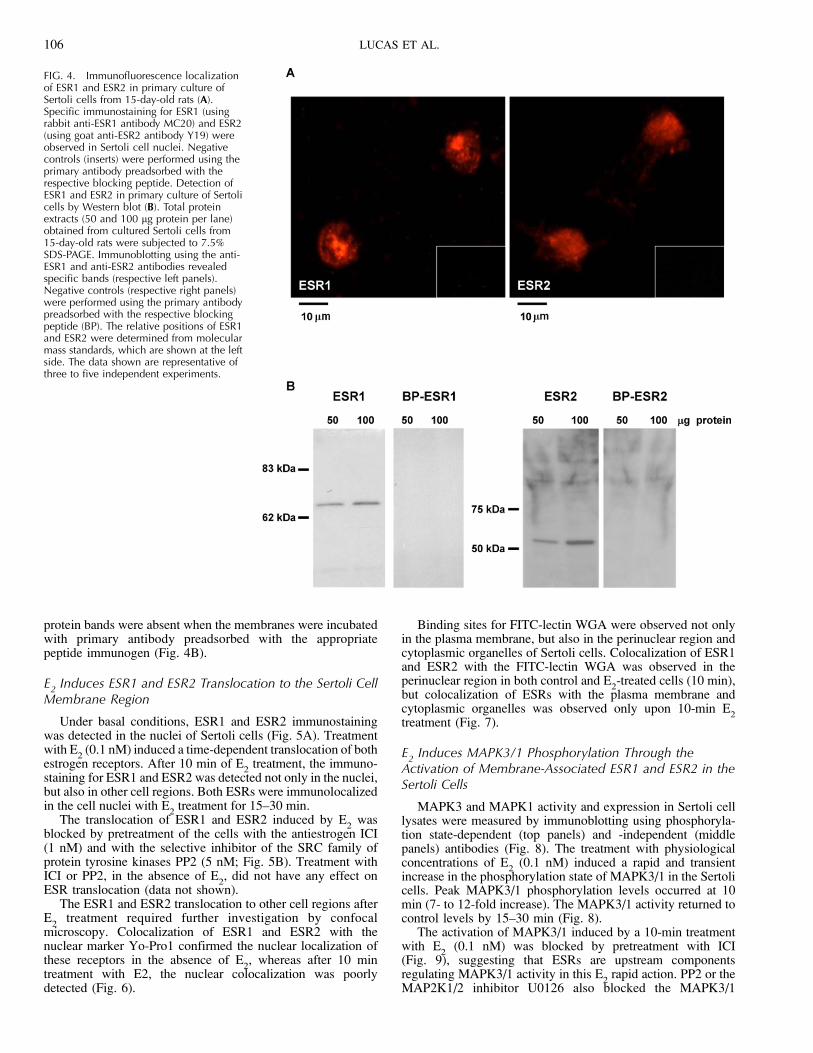
In cryosections of testis from 15-day-old rats, a specific andintense immunostaining was detected in many Sertoli andLeydig cell nuclei as well as in some peritubular myoid cells,but not in germ cells, when using the MC20 antibody raisedagainst the carboxyterminal region of ESR1 (Fig. 3B). Usingthe same antibody, positive immunostaining was also detectedin Sertoli cells from 120-day-old rat testis (Fig. 3D). Althoughgerm cells from immature rats were ESR1-negative, somespermatids were positive for this receptor in the adult rat testis(Fig. 3D). No immunostaining was observed when the primaryantibody was preadsorbed with the respective blocking peptide(Fig. 3, A and B). Similar results were observed in testes from15- and 120-day-old rats using the H184 antibody raisedagainst the aminoterminal region of ESR1 (SupplementalFig. 1, A and B available online at www.biolreprod.org). Usingthe Y19 antibody raised against the aminoterminal region ofthe ESR2, a specific immunostaining was detected in multipletesticular cell types, including Sertoli and Leydig cells, and insome but not all germ cells (Supplemental Fig. 1, C and D),confirming previous studies [9, 12, 54].
ESR1 was also localized by immunofluorescence in thenuclei of cultured Sertoli cells obtained from 15-day-old rats,using the H184, MC20, or 6F11 antibody, against the full-length ESR1. Figure 4A (left panel) shows the results obtainedusing the MC20 antibody, and Figure 4A (right panel) showsthe positive immunostaining for ESR2 using the Y19 antibody.The specificity of the antibodies was further confirmed byWestern blot analysis using total protein extracts of culturedSertoli cells. ESR1 and ESR2 were detected as single proteinbands of about 66 and 55 kDa, respectively (Fig. 4B). These
FIG. 2. Identification of Esr1 and Esr2 transcripts in primary culture ofSertoli cells from 15-day-old rats (lane 1) and in testes from 15- (lane 2),28- (lane 3) and 120- (lane 4)-day-old rats by RT-PCR. The PCR productswere resolved onto 1% agarose gel electrophoresis and visualized byethidium bromide staining. Arrows indicate the products with theexpected size. Gapdh was used as an internal control. Lane M: 1 kbplus DNA ladder (Invitrogen).
FIG. 3. Immunohistochemical localizationof ESR1 in cryosections of testes from 15- (B)and 120-day-old rats (D). Specific immu-nostaining for ESR1 (using rabbit anti-ESR1antibody MC20) was observed in Sertoli cellnuclei (solid arrows), Leydig cells (arrow-heads), peritubular myoid cells (asterisks),and some spermatids (open arrows). Nega-tive controls (A and C) were performedusing the primary antibody preadsorbedwith the respective blocking peptide. Scalebar as indicated. The data shown arerepresentative of at least four independentexperiments.
ESTROGEN AND MAPK3/1 ACTIVATION IN SERTOLI CELL 105
protein bands were absent when the membranes were incubatedwith primary antibody preadsorbed with the appropriatepeptide immunogen (Fig. 4B).
E2
Induces ESR1 and ESR2 Translocation to the Sertoli CellMembrane Region
Under basal conditions, ESR1 and ESR2 immunostainingwas detected in the nuclei of Sertoli cells (Fig. 5A). Treatmentwith E
2(0.1 nM) induced a time-dependent translocation of both
estrogen receptors. After 10 min of E2
treatment, the immuno-staining for ESR1 and ESR2 was detected not only in the nuclei,but also in other cell regions. Both ESRs were immunolocalizedin the cell nuclei with E
2treatment for 15–30 min.
The translocation of ESR1 and ESR2 induced by E2
wasblocked by pretreatment of the cells with the antiestrogen ICI(1 nM) and with the selective inhibitor of the SRC family ofprotein tyrosine kinases PP2 (5 nM; Fig. 5B). Treatment withICI or PP2, in the absence of E
2, did not have any effect on
ESR translocation (data not shown).The ESR1 and ESR2 translocation to other cell regions after
E2
treatment required further investigation by confocalmicroscopy. Colocalization of ESR1 and ESR2 with thenuclear marker Yo-Pro1 confirmed the nuclear localization ofthese receptors in the absence of E
2, whereas after 10 min
treatment with E2, the nuclear colocalization was poorlydetected (Fig. 6).
Binding sites for FITC-lectin WGA were observed not onlyin the plasma membrane, but also in the perinuclear region andcytoplasmic organelles of Sertoli cells. Colocalization of ESR1and ESR2 with the FITC-lectin WGA was observed in theperinuclear region in both control and E
2-treated cells (10 min),
but colocalization of ESRs with the plasma membrane andcytoplasmic organelles was observed only upon 10-min E
2treatment (Fig. 7).
E2
Induces MAPK3/1 Phosphorylation Through theActivation of Membrane-Associated ESR1 and ESR2 in theSertoli Cells
MAPK3 and MAPK1 activity and expression in Sertoli celllysates were measured by immunoblotting using phosphoryla-tion state-dependent (top panels) and -independent (middlepanels) antibodies (Fig. 8). The treatment with physiologicalconcentrations of E
2(0.1 nM) induced a rapid and transient
increase in the phosphorylation state of MAPK3/1 in the Sertolicells. Peak MAPK3/1 phosphorylation levels occurred at 10min (7- to 12-fold increase). The MAPK3/1 activity returned tocontrol levels by 15–30 min (Fig. 8).
The activation of MAPK3/1 induced by a 10-min treatmentwith E
2(0.1 nM) was blocked by pretreatment with ICI
(Fig. 9), suggesting that ESRs are upstream componentsregulating MAPK3/1 activity in this E
2rapid action. PP2 or the
MAP2K1/2 inhibitor U0126 also blocked the MAPK3/1
FIG. 4. Immunofluorescence localizationof ESR1 and ESR2 in primary culture ofSertoli cells from 15-day-old rats (A).Specific immunostaining for ESR1 (usingrabbit anti-ESR1 antibody MC20) and ESR2(using goat anti-ESR2 antibody Y19) wereobserved in Sertoli cell nuclei. Negativecontrols (inserts) were performed using theprimary antibody preadsorbed with therespective blocking peptide. Detection ofESR1 and ESR2 in primary culture of Sertolicells by Western blot (B). Total proteinextracts (50 and 100 lg protein per lane)obtained from cultured Sertoli cells from15-day-old rats were subjected to 7.5%SDS-PAGE. Immunoblotting using the anti-ESR1 and anti-ESR2 antibodies revealedspecific bands (respective left panels).Negative controls (respective right panels)were performed using the primary antibodypreadsorbed with the respective blockingpeptide (BP). The relative positions of ESR1and ESR2 were determined from molecularmass standards, which are shown at the leftside. The data shown are representative ofthree to five independent experiments.
106 LUCAS ET AL.

activation induced by E2
(0.1 nM) for 10 min (Fig. 9). Thetreatment with ICI (1 nM), PP2, or U0126 for 30 min, in theabsence of E
2, did not have any effect on MAPK3/1
phosphorylation (data not shown). To further confirm theinvolvement of EGFR, the EGFR kinase inhibitor AG 1478was tested and markedly decreased E
2-induced MAPK3/1
phosphorylation. EGF alone was used as a positive control andstrongly activated MAPK3/1, and this effect was blocked byAG 1478 (Fig. 10). No differences were observed in totalMAPK3/1 protein expression under any of these conditions(middle panels of Figures 8–10).
E2
Stimulates Sertoli Cell Proliferation
E2
caused an enhancement of 41.3% 6 1.3% of[methyl-3H]thymidine incorporation after 24 h incubation
(Fig. 11). In addition, the pretreatment of Sertoli cells withICI markedly reduced the [methyl-3H]thymidine incorporationinto DNA induced by E
2(Fig. 11). In the absence of E
2, the
antiestrogen did not have any effect on [methyl-3H]thymidineincorporation (Fig. 11).
DISCUSSION
The present study indicates that estrogen promotes Sertolicell proliferation and the activation of a signaling cascade thatinitiates at the plasma membrane and involves SRC, EGFR,and MAPK3/1 activation. Furthermore, it was identified thatboth ESR1 and ESR2 are present in Sertoli cells and thetreatment with E
2induces the translocation of both to the
plasma membrane region.
FIG. 5. E2-induced translocation of ESR1
and ESR2 to the Sertoli cell membraneregion. ESR1 and ESR2 immunostaining wasdetected in the Sertoli cell nuclei underbasal conditions (C, control, vehicle-treatedcells). After incubation with E
2(0.1 nM) for
10 min, ESR1 and ESR2 were detected in ornear the plasma membrane and in thecytosol of Sertoli cells. Both ESRs weredetected in the Sertoli cell nuclei after 15 to30 min of E
2incubation (A). E
2-induced
translocation of ESR1 and ESR2 to theplasma membrane region was blocked bypretreatment of the cells with the antiestro-gen ICI (1 nM; 30 min) and the selectiveinhibitor of the SRC family of proteintyrosine kinases PP2 (5 nM, 30 min; B). Thedata shown are representative of three tofive independent experiments.
ESTROGEN AND MAPK3/1 ACTIVATION IN SERTOLI CELL 107
ESR1 and ESR2 are encoded by two different genes locatedon different chromosomes. Multiple RNA splicing variants forESR1 [55] and alternative splicing variants [56–58] andpolymorphism [59] for ESR2 have been reported for human.Such a molecular diversity due to alternative splicingmechanisms of the correpondent genes is also observed inrodents [45, 60, 61]. Esr1 splicing variants were not detected incultured Sertoli cells or in testis, and only a single band forESR1 was detected in cultured Sertoli cells by the Western blotanalysis. On the other hand, two splicing variants were detectedfor Esr2 both in cultured Sertoli cells and in testis: the Esr2_v1isoform, with high similarity to the human ESR2 [62, 63], andthe Esr2_v2, which contains an in-frame insert of 54
nucleotides that has only been found in tissues from rodents[45]. Esr2_v2 encodes a protein that differs from the isoform 1of ESR2 only by an insertion of 18 aminoacids in the ligand-binding domain. In fact, in Western blot analysis only oneprotein band was distinguished when an antibody thatrecognizes the aminoterminal region of the receptor commonto both ESR2 isoforms was used. Petersen et al. [45] reportedan identical migration pattern for the recombinant ESR2isoforms 1 and 2 expressed in HEK293T cells. Interestingly, inthe present study an Esr2_v1-related transcript was detectedonly in cultured Sertoli cells and in testes from 15-day-oldanimals but not from older animals, whereas an Esr2_v2-related transcript was detected in the testes of all studied ages.
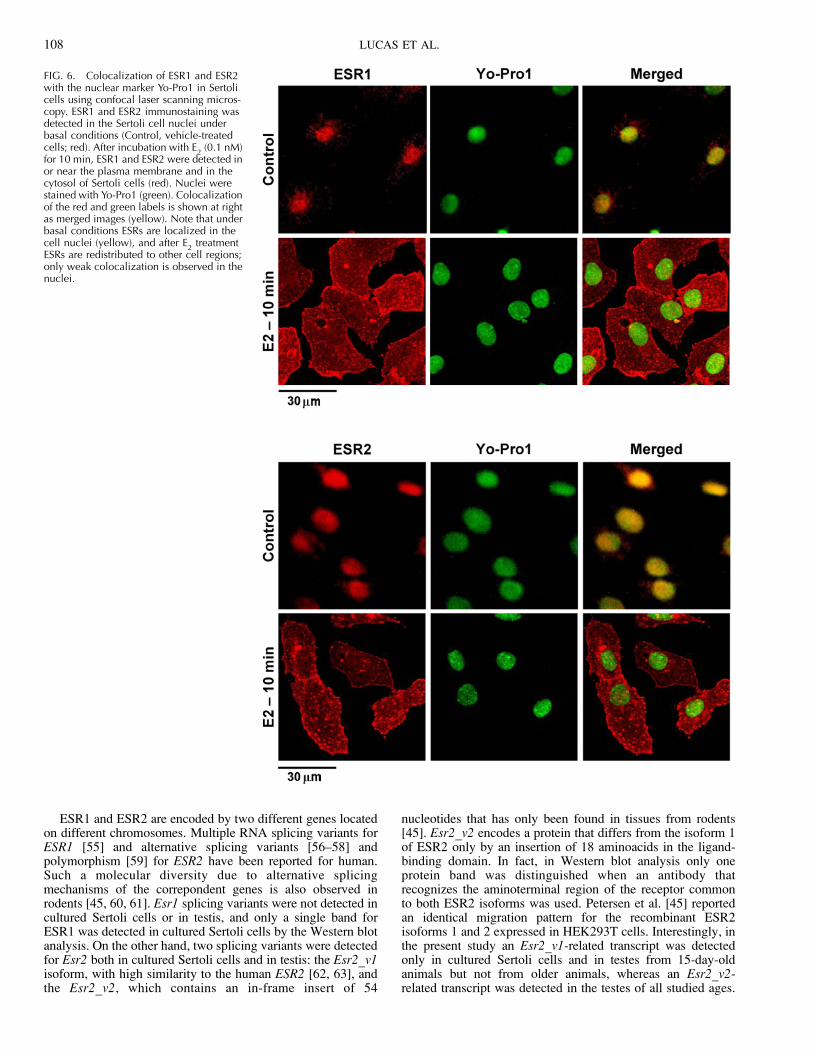
FIG. 6. Colocalization of ESR1 and ESR2with the nuclear marker Yo-Pro1 in Sertolicells using confocal laser scanning micros-copy. ESR1 and ESR2 immunostaining wasdetected in the Sertoli cell nuclei underbasal conditions (Control, vehicle-treatedcells; red). After incubation with E
2(0.1 nM)
for 10 min, ESR1 and ESR2 were detected inor near the plasma membrane and in thecytosol of Sertoli cells (red). Nuclei werestained with Yo-Pro1 (green). Colocalizationof the red and green labels is shown at rightas merged images (yellow). Note that underbasal conditions ESRs are localized in thecell nuclei (yellow), and after E
2treatment
ESRs are redistributed to other cell regions;only weak colocalization is observed in thenuclei.
108 LUCAS ET AL.

The physiological implication of the existence of several ESR2isoforms has been investigated. The human ESR2 isoform 1apparently is the only fully functional one. However, the otherisoforms can heterodimerize with ESR2 isoform 1 under thestimulation of estrogens and enhance the transactivation [64].Furthermore, the human ESR2 isoform 2 has been proposed toact as a dominant negative receptor, preventing estrogen actionin gonocytes [58]. The differential distribution of ESR2variants in human testicular cells may indicate that thesevariants have specific functions in spermatogenesis [57]. Onthe other hand, the rodent ESR2 isoform 2 seems to befunctional, although much higher concentrations of estrogenare required for its activity [45].
The expression of ESR1 and ESR2 in the fetal testis occursvery early in development, and their distribution in the differenttesticular cells has been extensively studied in mammals [5, 11,65]. Immunohistochemical data have shown that ESR1 ispresent in the undifferentiated gonad and in the fetal Leydigcells until birth in rodents [19]. The most consistent data acrossspecies has been the presence of ESR1 in the Leydig cells.There are conflicting reports of ESR1 in Sertoli cells, germcells, and spermatozoa [4, 9, 13, 16, 17, 66–68]. Our resultsdemonstrated the presence of ESR1 in Sertoli cells fromimmature and adult rats by using antibodies raised against thecarboxyterminal (presented data) or aminoterminal region ofESR1 (Supplemental Fig. 1). It was also confirmed that germ
FIG. 7. Colocalization of ESR1 and ESR2with FITC-lectin WGA in Sertoli cells usingconfocal laser scanning microscopy. ESR1and ESR2 immunostaining was detected inthe Sertoli cell nuclei under basal condi-tions (Control, vehicle-treated cells; red).After incubation with E
2(0.1 nM) for 10
min, ESR1 and ESR2 were detected in ornear the plasma membrane and in thecytosol of Sertoli cells (red). Plasma mem-brane, cytoplasmic organelles, and theperinuclear region were stained with FITC-lectin (green), which selectively binds to N-acetylglucosamine and sialic acid residues.Colocalization of the red and green labels isshown at right as merged images (yellow).Both control and E
2-treated cells (10 min)
showed colocalization of ESRs in theperinuclear region (yellow, arrows). After E
2treatment, ESRs were redistributed to othercell regions, including the plasma mem-brane (yellow, arrowheads).
ESTROGEN AND MAPK3/1 ACTIVATION IN SERTOLI CELL 109

cells from immature rats are ESR1-negative, though somespermatids were positive for this receptor in the adult animal.Indeed, the presence of ESR1 in rat and human spermatozoahas been reported recently [3, 67, 68]. ESR1 was also detectedby immunofluorescence in the nuclei of cultured Sertoli cellsobtained from 15-day-old rats. Colocalization of ESR1 andCLU further confirmed that ESR1 was in fact expressed bySertoli cells (data not shown). One possibility to explain theapparent discrepancies observed in the literature regarding tothe presence of ESR1 in Sertoli cells could be related to themethodologies used. We examined various fixing protocolsand antibody sources to obtain a suitable immunohistochemicalprocedure for ESR1 detection in rat testes. In fact, using testesfrom adult rats perfused with 10% neutral buffer formalinembedded in paraffin and the antibodies described above, wewere not able to detect any immunoreaction for ESR1 in Sertolicells, but Leydig cells and peritubular myoid cells wereimmunostained (data not shown).
There is more agreement in the literature concerning theimmunolocalization of the ESR2 protein. We detected ESR2-specific immunostaining in multiple cell types, includingSertoli and Leydig cells, and in some but not all germ cells,which confirms previous studies [9, 12, 54]. It is worthmentioning that ESR2 is coexpressed with ESR1 in tumoralseminoma cells, where it may counteract the tumor cellproliferation mediated by ESR1 [69].
A series of E2-induced, membrane-initiated steroid signaling
has been described in benign and malignant cells of variousorigins. These effects occur from seconds to minutes after
administration of E2
and involve rapid activation of manysignaling molecules [20, 26, 27, 34, 35]. In the presentstudy, treatment with E
2induced a translocation of ESR1 and
ESR2 from the nuclei to the plasma membrane of Sertoli cells.In addition to posttranslation modifications of ESR1 bypalmitoylation [70], other mechanisms have been proposed toexplain the translocation of ESR1 to the cell membrane inestrogen-stimulated cells [26, 27]. For example, in humanmammary cancer MCF-7 cells, the translocation of the ESR1 tothe cell membrane involves SHC1 as a transporter. In responseto E
2, SHC1 is phosphorylated and binds to ESR1. At the same
time, the SHC1 binding sites on the IGF1R are phosphorylated,allowing SHC1 to bind to the IGF1R. ESR1 can then betethered to the membrane through SHC1 interaction withIGF1R [50, 71, 26]. SHC1 is a substrate of activated SRC and,once phosphorylated, interacts with other proteins [26],suggesting a possible role of SRC as a signal integrator inestrogen-stimulated MCF-7 cells, since estrogen stimulatesMAPK activation through the interaction of SRC with ESR1[72–74]. There is evidence that ESR2 could also localize in
FIG. 8. Effects of E2
on MAPK3/1 phosphorylation in primary culture ofSertoli cells from 15-day-old rats. Cells were incubated in the absence (C,control, vehicle-treated cells) and presence of E
2(0.1 nM) for 5 to 30 min
and lysed in ice-cold lysis buffer. Total cellular proteins (60 lg per lane)were resolved in 10% SDS/PAGE, transferred to PVDF membrane, andprobed with antibody specific for phosphorylated MAPK3/1 (top panels) orwith antibody that recognizes total (phosphorylation state-independent)MAPK3/1 proteins (middle panels). The relative positions of phosphory-lated MAPK3/1 and total MAPK3/1 proteins are shown at the right. Thedata shown are representative of three to four independent experiments.Bottom panel: The bars represent the densitometric analysis of theWestern blot. Solid bars ¼ MAPK3; Open bars ¼ MAPK1. Results werenormalized to total MAPK3/1 expression in each sample and plotted(mean 6 SEM) in relation to control (C ¼ 1). Different letters indicatestatistical significance (P , 0.05, Newman-Keuls test).
FIG. 9. Participation of estrogen receptors, SRC, and MAP2K1/2 onE
2-induced MAPK3/1 phosphorylation in primary culture of Sertoli cells
from 15-day-old rats. Cells were untreated or pretreated with ICI (1 nM),PP2 (5 nM), or MAP2K1/2 inhibitor (U0126, 100 lM) for 30 min.Afterwards, the cells were stimulated with E
2(0.1 nM, 10 min) and lysed
in ice-cold lysis buffer. Total cellular proteins (60 lg per lane) wereresolved in 10% SDS-PAGE, transferred to PVDF membrane, and probedwith antibody specific for phosphorylated MAPK3/1 (top panels) or withantibody that recognizes total (phosphorylation state-independent)MAPK3/1 proteins (middle panels). The relative positions of phosphory-lated MAPK3/1 and total MAPK3/1 proteins are shown at the right. Thedata shown are representative of three to four independent experiments.Bottom panel: The bars represent the densitometric analysis of theWestern blot. Solid bars ¼ MAPK3; Open bars ¼ MAPK1. Results werenormalized to total MAPK3/1 expression in each sample and plotted(mean 6 SEM) in relation to control (C ¼ 1). Different letters indicatestatistical significance (P , 0.05, Newman-Keuls test).
110 LUCAS ET AL.

caveolae and participate in nongenomic signaling through theinteraction of SRC with ESR2 [75–76]. As presently found, theE
2-induced translocation of ESR1 and ESR2 to the plasma
membrane region of Sertoli cells was blocked by pretreatmentof the cells with a selective inhibitor of the SRC family ofprotein tyrosine kinases, suggesting that SRC plays a crucialrole in the translocation of ESR1 and ESR2 from the nucleus tothe cell membrane. Fan et al. [77] have recently shown asimilar mechanism of SRC- and EGFR-mediated translocationof ESR1 out of the nucleus induced by tamoxifen in MCF-7cells resistant to tamoxifen, and they suggest that thismechanism could contribute to acquired tamoxifen resistance.
Membrane-associated ESR1 transduces estrogen rapidsignals, leading to the activation of many signaling molecules,such as MAPK, AKT, KRAS, RAF1, and protein kinase C,alteration of maxi-K
þchannels, increase in intracellular Ca
þþ
levels, release of nitric oxide, and stimulation of prolactinsecretion [26, 27, 78]. In addition to the activation of thesesignaling molecules, membrane receptors for growth factors,such as IGF1R and EGFR, can also be activated by estrogenrapid action, leading to activation of downstream MAPK andAKT signaling pathways [26, 27, 79, 80]. The translocation ofESR1 and ESR2 to the Sertoli cell membrane occurred after E
2
treatment in the same time frame as MAPK3/1 activation, andthese effects were also blocked by a selective inhibitor of theSRC. Furthermore, the antiestrogen ICI blocked E
2-induced
ESR1 and ESR2 translocation to the membrane and MAPK3/1phosphorylation, indicating the direct involvement of ESRs inthese events. These estrogen actions and induced pathwayshave also been demonstrated in many other cell types,including MCF-7 breast cancer cells [72], osteoblasts [81],rat brain [82], human colon carcinoma-derived Caco-2 cells[83], and LNCaP prostate cancer cells [84]. The evidence of arelationship between the EGFR and estrogen has beenpreviously provided by data showing that neutralizingantibodies against EGF inhibited estrogen-mediated prolifera-tion in the mice uterus [85]. In Sertoli cells, an EGFR kinaseinhibitor decreased the E
2-induced MAPK3/1 phosphorylation,
confirming the involvement of EGFR in this activity. Whethera linear pathway involving a sequential activation of IGF1R,metalloproteinases, and EGFR by E
2is also present in Sertoli
cells, such as recently demonstrated in MCF-7 breast cancercells [80], still remains to be determined. Furthermore, in awide variety of cells, besides the localization in nucleus andplasma membranes, ESR1 and ESR2 have also been localizedin the mitochondria [86]. The relative subcellular localizationof the receptors is altered in cancer cells, and ESR2 has beenrecently shown to shift from the mitochondria to the nucleusduring neoplastic transformation of epithelial breast cells [87].It would be interesting to determine whether, in Sertoli cells,ESRs may also localize in mitochondria and the influence ofestrogen on their subcellular distribution.
In order to provide further evidence of the importance ofestrogen on Sertoli cell biology, we examined the effects of E
2on Sertoli cell proliferation. After 24 h of E
2incubation, an
increase of 41% of [methyl-3H]thymidine incorporation wasobserved. Moreover, ICI blocked this effect, suggesting thatthe E
2plays a role in the Sertoli cell proliferation.
In the developing testis, estrogen plays a significant role inestablishing Sertoli cell function [19] and potentially even inestablishing Sertoli cell-germ cell adhesion [88]. Immature,
FIG. 10. Involvement of EGFR on E2-induced MAPK3/1 phosphorylation
in primary culture of Sertoli cells from 15-day-old rats. Cells wereuntreated or pretreated with the EGFR kinase inhibitor AG 1478 (50 lM)for 15 min. Afterwards, the cells were stimulated with E
2(0.1 nM, 10 min)
and lysed in ice-cold lysis buffer. EGF (10 ng/ml) was used as a positivecontrol. Total cellular proteins (60 lg per lane) were resolved in 10% SDS-PAGE, transferred to PVDF membrane, and probed with antibody specificfor phosphorylated MAPK3/1 (top panels) or with antibody that recognizestotal (phosphorylation state-independent) MAPK3/1 proteins (middlepanels). The relative positions of phosphorylated MAPK3/1 and totalMAPK3/1 proteins are shown at the right. The data shown arerepresentative of three to four independent experiments. Bottom panel:The bars represent the densitometric analysis of the Western blot. Solidbars ¼ MAPK3; Open bars ¼ MAPK1. Results were normalized to totalMAPK3/1 expression in each sample and plotted (mean 6 SEM) inrelation to control (C¼ 1). Different letters indicate statistical significance(P , 0.05, Newman-Keuls test).
FIG. 11. Effects of E2
on Sertoli cell proliferation. Primary culturesof Sertoli cells from 15-day-old rats were initially incubated with0.074 MBq/ml [methyl-3H]thymidine for 6 h at 358C. Incubation wascontinued in the absence (basal incorporation) and presence of E
2(0.1
nM) for 24 h. Cells were treated with ICI (1 nM) for 30 min before additionof E
2(0.1 nM) for 24 h, or only treated with ICI (1 nM) for 30 min. The
reaction was stopped, the cells rinsed with ice-cold PBS and trichloro-acetic acid, and then solubilized with NaOH. Bound radioactivity wasdetermined and the results were expressed as percentage of incorporationabove basal level (mean 6 SEM of three to five independent experimentsperformed in triplicate). Basal incorporation of [methyl-3H]thymidine was1440.7 6 164.4 disintegrations per minute per well. *[methyl-3H]thymi-dine incorporation activation significantly greater than basal level (P ,0.05, Student t-test).
ESTROGEN AND MAPK3/1 ACTIVATION IN SERTOLI CELL 111

undifferentiated Sertoli cells may generate estrogen themselvesto act in an autocrine manner [3]. Current evidence suggests thatthe effect of E
2on Sertoli cells depends on the concentration
of the hormone. Administration of high doses (10 lg per animalduring 10 days) of diethylstilbestrol to neonatal rats suppressedFSH plasma levels and induced changes that were consistentwith a permanent impairment of functional maturation of Sertolicells [89], whereas lower doses (0.1 lg per animal) decreasedSertoli cell numbers without interfering with FSH levels [90].On the other hand, in immature bank voles, administration oflow doses of E
2(0.1 lg/g body weight) accelerated the onset
of spermatogenesis, and high doses (10 lg/g body weight)caused a disruption of testicular structure and tubular atrophy[91]. The present in vitro study shows that, in cultureconditions, physiological concentrations of E
2(0.1 nM) can
induce the proliferation of Sertoli cells from 15-day-old rats,and corroborates previous findings that low concentrations ofE
2may have positive effects on testicular function. It is very
likely that estrogens not only suppress Sertoli cell differentia-tion, but also act as a mitogen in this cell proliferation during theneonatal period.
It is interesting that in the Esr1�/� testis there is significantlyless seminiferous tubular secretion than in the wild-type testis[1]. The same effect was suggested for the Cyp19�/� testis [92].Mice subjected to a long-term treatment with ICI showedhypospermatogenesis and abnormal germ cell development,but no seminiferous tubular dilation or increase in testis weightwas observed [93]. Taken together, these data suggest thatestrogen-ESRs play a role in Sertoli cell function. Thus, thesignaling mechanism elicited by estrogen involving transloca-tion of ESRs to the plasma membrane and activation of MAPKsignaling may have important implications for Sertoli cellfunction and spermatogenesis. A recent study described asimilar signaling pathway elicited in Sertoli cells by testoster-one acting on plasma membrane-associated androgen receptors[94].
A sequential event induced by estrogen and involvingactivation of G proteins, matrix metalloproteinases 2 and 9,release of heparin-binding EGF, and finally EGFR activationon the cell membrane has also been reported to lead toactivation of MAPK and AKT [28–31]. The present data showthat the E
2effect on MAPK3/1 activation involves the classical
ESRs because it can be blocked by the antagonist ICI.Similarly, the E
2-induced proliferation of Sertoli cells was
blocked by ICI, demonstrating the involvement of ESRs inthis process. It is not clear yet whether the proliferative effectof E
2on cultured Sertoli cells involves a genomic and/or a non-
genomic action of ESR1 and ESR2.Our studies do not exclude the possibility that the G protein-
coupled receptor GPR30 may contribute to E2-induced
MAPK3/1 activation and Sertoli cell proliferation, and therelative contribution of each receptor to control E
2action in
Sertoli cells still remains to be determined. In ovarian cancercells, it has been recently demonstrated that both GPR30 andESR1 must be present to observe E
2-mediated activation of
EGFR signaling, MAPK3/1 activation, and cell proliferation[32]. On the other hand, in MCF-7 breast cancer cells, E
2stimulated cell proliferation via sequential activation ofIGF1R and EGFR, and knockdown or blockade of ESR1, butnot of GPR30, blocked E
2-induced MAPK activation [80]. The
apparent discrepancies found in the literature seem to resultfrom differences in cellular context. Further studies areunderway to determine the role of GPR30 in rat Sertoli cells.
In conclusion, these results indicate that the effect of E2
onSertoli cell function involves the following steps: i) transloca-tion of ESR1 and ESR2 to the plasma membrane mediated by
the non-receptor tyrosine kinase SRC and ii) activation ofEGFR and MAPK signaling pathway. In addition, E
2induces
Sertoli cell proliferation. These actions of E2
might be one ofthe key steps that mediate the estrogen-dependent activation ofcellular events in Sertoli cells. Additional studies are nowrequired to further elucidate the role of each ESR in Sertoli cellbiology and, consequently, in spermatogenesis and male(in)fertility.
ACKNOWLEDGMENTS
We thank Espedita M.J. Silva Santos, Maria Damiana Silva, andSabrina A. Souza e Silva for technical assistance.
REFERENCES
1. Hess RA, Bunick D, Bahr J, Taylor JA, Korach KS, Lubahn DB. A rolefor oestrogens in the male reproductive system. Nature 1997; 390:509–512.
2. Sharpe RM. Hormones and testis development and the possible adverseeffects of environmental chemicals. Toxicol Lett 2001; 120:221–232.
3. Carreau S, Delalande C, Silandre D, Bourguiba S, Lambard S. Aromataseand estrogen receptors in male reproduction. Mol Cell Endocrinol 2006;246:65–68.
4. Hess RA, Carnes K. The role of estrogen in testis and the malereproductive tract: a review and species comparison. Anim Reprod 2004;1:5–30.
5. Akingbemi BT. Estrogen regulation of testicular function. Reprod BiolEndocrinol 2005; 3:2–13.
6. Simpson ER, Mahendroo MS, Means GD, Kilgore MW, HinshelwoodMM, Graham-Lorence S, Amarneh B, Ito Y, Fisher CR, Michael MD.Aromatase cytochrome P450, the enzyme responsible for estrogenbiosynthesis. Endocr Rev 1994; 15:342–355.
7. Carreau S. The testicular aromatase: from gene to physiological role.Reprod Biol 2002; 2:5–12.
8. Dorrington JH, Bendell JJ, Khan SA. Interactions between FSH, estradiol-17 beta and transforming growth factor-beta regulate growth anddifferentiation in the rat gonad. J Steroid Biochem Mol Biol 1993; 44:441–447.
9. Fisher JS, Millar MR, Majdic G, Saunders PT, Fraser HM, Sharpe RM.Immunolocalisation of oestrogen receptor-alpha within the testis andexcurrent ducts of the rat and marmoset monkey from perinatal life toadulthood. J Endocrinol 1997; 153:485–495.
10. Yamashita S. Localization of estrogen and androgen receptors in malereproductive tissues of mice and rats. Anat Rec Discov Mol Cell Evol Biol2004; 279:768–778.
11. Sierens JE, Sneddon SF, Collins F, Millar MR, Saunders PT. Estrogens intestis biology. Ann N Y Acad Sci 2005; 1061:65–76.
12. Saunders PT, Fisher JS, Sharpe RM, Millar MR. Expression of oestrogenreceptor beta (ER beta) occurs in multiple cell types, including some germcells, in the rat testis. J Endocrinol 1998; 156:R13–R17.
13. Zhou Q, Nie R, Prins GS, Saunders PT, Katzenellenbogen BS, Hess RA.Localization of androgen and estrogen receptors in adult male mousereproductive tract. J Androl 2002; 23:870–881.
14. Pelletier G, Labrie C, Labrie F. Localization of oestrogen receptor alpha,oestrogen receptor beta and androgen receptors in the rat reproductiveorgans. J Endocrinol 2000; 165:359–370.
15. Albrecht ED, Billiar RB, Aberdeen GW, Babischkin JS, Pepe GJ.Expression of estrogen receptors alpha and beta in the fetal baboon testisand epididymis. Biol Reprod 2003; 70:1106–1113.
16. Ramesh R, Pearl CA, At-Taras E, Roser JF, Berger T. Ontogeny ofandrogen and estrogen receptor expression in porcine testis: effect ofreducing testicular estrogen synthesis. Anim Reprod Sci; 15 November2006; PMID: 17157457.
17. Mutembei HM, Pesch S, Schuler G, Hoffmann B. Expression of oestrogenreceptors alpha and beta and of aromatase in the testis of immature andmature boars. Reprod Domest Anim 2005; 40:228–236.
18. Sneddon SF, Walther N, Saunders PT. Expression of androgen andestrogen receptors in sertoli cells: studies using the mouse SK11 cell line.Endocrinology 2005; 146:5304–5312.
19. O’Donnell L, Robertson KM, Jones ME, Simpson ER. Estrogen andspermatogenesis. Endocr Rev 2001; 21:497–511.
20. Hewitt SC, Harrell JC, Korach KS. Lessons in estrogen biology fromknockout and transgenic animals. Annu Rev Physiol 2005; 67:285–308.
21. Eddy EM, Washburn TF, Bunch DO, Goulding EH, Gladen BC, Lubahn
112 LUCAS ET AL.

DB, Korach KS. Targeted disruption of the estrogen receptor gene in malemice causes alteration of spermatogenesis and infertility. Endocrinology1996; 137:4796–4805.
22. Delbes G, Levacher C, Pairault C, Racine C, Duquene C, Krust A, HabertR. Estrogen receptor (beta)-mediated inhibitor of male germ cell linedevelopment in mice by endogenous estrogens during perinatal life.Endocrinology 2004; 145:3395–3403.
23. Ogawa S, Chan J, Chester AE, Gustafsson JA, Korach KS, Pfaff DW.Survival of reproductive behaviors in estrogen receptor beta gene-deficient(betaERKO) male and female mice. Proc Natl Acad Sci U S A 1999; 96:12887–12892.
24. Mahato D, Goulding EH, Korach KS, Eddy EM. Spermatogenic cells donot require estrogen receptor-alpha for development or function.Endocrinology 2000; 14:1273–1276.
25. Mahato D, Goulding EH, Korach KS, Eddy EM. Estrogen receptor-alphais required by the supporting somatic cells for spermatogenesis. Mol CellEndocrinol 2001; 178:57–63.
26. Song RXD, Santen RJ. Membrane initiated estrogen signaling in breastcancer. Biol Reprod 2006; 75:9–16.
27. Manavathi B, Kumar R. Steering estrogen signals from the plasmamembrane to the nucleus: two sides of the coin. J Cell Physiol 2006; 207:594–604.
28. Filardo EJ. Epidermal growth factor receptor (EGFR) transactivation byestrogen via the G-protein-coupled receptor, GPR30: a novel signalingpathway with potential significance for breast cancer. J Steroid BiochemMol Biol 2002; 80:231–238.
29. Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER. Atransmembrane intracellular estrogen receptor mediates rapid cellsignaling. Science 2005; 307:1625–1630.
30. Filardo EJ, Thomas P. GPR30: a seven-transmembrane-spanning estrogenreceptor that triggers EGF release. Trends Endocrinol Metab 2005; 16:362–367.
31. Prossnitz ER, Arterburn JB, Sklar LA. GPR30: a G protein-coupledreceptor for estrogen. Mol Cell Endocrinol 2007; 265/266:138–142.
32. Albanito L, Madeo A, Lappano R, Vivacqua A, Rago V, Carpino A,Oprea TI, Prossnitz ER, Musti AM, Ando S, Maggiolini M. G protein-coupled receptor 30 (GPR30) mediates expression changes and growthresponse to 17b-estradiol and selective GPR30 ligand G-1 in ovariancancer cells. Cancer Res 2007; 67:1859–1866.
33. Hewitt SC, Deroo BJ, Korach KS. Signal transduction. A new mediatorfor an old hormone? Science 2005; 307:1572–1573.
34. Bjornstrom L, Sjoberg M. Mechanisms of estrogen receptor signaling:convergence of genomic and nongenomic actions on target genes. MolEndocrinol 2005; 19:833–842.
35. Song RXD, Zhang Z, Santen RJ. Estrogen rapid action via proteincomplex formation involving ERa and Src. Trends Endocrinol Metab2005; 16:347–353.
36. Skinner MK, Fritz IB. Testicular peritubular cells secrete a protein underandrogen control that modulates Sertoli cells functions. Proc Natl Acad SciU S A 1985; 82:114–118.
37. Grima J, Zhu LJ, Zong SD, Cattedrall JF, Bardin CW, Cheng CY. Rattestis is a newly identified component of the junctional complexes invarious tissues whose mRNA is predominantly expressed in the testis andovary. Biol Reprod 1995; 52:340–355.
38. Lucas TFG, Avellar MC, Porto CS. Effects of carbachol on rat Sertoli cellproliferation and muscarinic acetylcholine receptors regulation: an in vitrostudy. Life Sci 2004; 75:1761–1773.
39. Galdieri M, Ziparo E, Palombi F, Russo MA, Stefanini M. Pure Sertolicells culture: a new model for the study of somatic-germ cell interactions. JAndrol 1981; 5:249–259.
40. Tung PS, Dorrington JH, Fritz IB. Structural changes inducted by follicle-stimulating hormone or dibutyryl cyclic AMP on presumptive Sertoli cellsin culture. Proc Natl Acad Sci U S A 1975; 72:1838–1842.
41. Skinner MK, Griswold MD. Sertoli cells synthesize and secretetransferrin-like protein. J Biol Chem 1980; 255:9523–9525.
42. Griswold MD, Roberts K, Bishop P. Purification and characterization of asulfated glycoprotein secreted by Sertoli cells. Biochemistry 1986; 25:7265–7270.
43. Lakins J, Bennett SA, Chen JH, Arnold JM, Morrissey C, Wong P,O’Sullivan J, Tenniswood M. Clusterin biogenesis is altered duringapoptosis in the regressing rat ventral prostate. J Biol Chem 1998; 273:27887–27895.
44. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acidguanidiniumthiocyanate-phenol-chloroform extraction. Anal Biochem1987; 162:156–159.
45. Petersen DN, Tkalcevic GT, Koza-Taylor PH, Turi TG, Brown TA.Identification of estrogen receptor b2, a functional variant of estrogen
receptor b expressed in normal rat tissue. Endocrinology 1998; 139:1082–1092.
46. Saji S, Sakaguchi H, Andersson S, Warner M, Gustafsson J. Quantitativeanalysis of estrogen receptor proteins in rat mammary gland. Endocrinol-ogy 2001; 142:3177–3186.
47. Moore JT, McKee DD, Slentz-Kesler K, Moore LB, Jones SA, Horne EL,Su JL, Kliewer SA, Lehmann JM, Willson TM. Cloning and character-ization of human estrogen receptor beta isoforms. Biochem Biophys ResCommun 1998; 247:75–78.
48. Scofield MA, Liu F, Abel PW, Jeffries WB. Quantification of steady stateexpression of mRNA for alpha-1 adrenergic receptor subtypes usingreverse transcription and a competitive polymerase chain reaction. JPharmacol Exp Ther 1995; 275:1035–1042.
49. Wong JK, Le HH, Zsarnovszky A, Belcher SM. Estrogens and ICI182,780 (faslodex) modulate mitosis and cell death in immature cerebellarneurons via rapid activation of p44/p42 mitogen-activated protein kinase. JNeurosci 2003; 23:4984–4995.
50. Song RX, Barnes CJ, Zhang Z, Bao Y, Kumar R, Santen RJ. The role ofShc and insulin-like growth factor 1 receptor in mediating the translocationof estrogen receptor a to the plasma membrane. Proc Natl Acad Sci U S A2004; 101:2076–2081.
51. Rasband WS. ImageJ Software. U.S. National Institutes of Health,Bethesda, Maryland. 2007. World Wide Web (URL: http://rsb.info.nih.gov/ij/). (13 October 2006).
52. Monteiro HP, Oliveira JR, Curcio MF, Moraes MS, Arai RJ. Tyrosinephosphorylation in nitric oxide-mediated signaling events. MethodsEnzymol 2005; 396:350–358.
53. Guizzetti M, Costa P, Peters J, Costa LG. Acetylcholine as a mitogen:muscarinic receptor-mediated proliferation of rat astrocytes and humanastrocytoma cells. Eur J Pharmacol 1996; 297:265–273.
54. van Pelt AM, de Rooij DG, van der Burg B, van der Saag PT, GustafssonJA, Kuiper GG. Ontogeny of estrogen receptor-beta expression in rattestis. Endocrinology 1999; 140:478–483.
55. Flouriot G, Griffin C, Kenealy M, Sonntag-Buck V, Gannon F.Differentially expressed messenger RNA isoforms of the human estrogenreceptor-alpha gene are generated by alternative splicing and promoterusage. Mol Endocrinol 1998; 12:1939–1954.
56. Zhou W, Liu Z, Wu J, Liu J, Hyder SM, Antoniou E, Lubahn DB.Identification and characterization of two novel splicing isoforms ofhuman estrogen-related receptor beta. J Clin Endocrinol Metab 2006; 91:569–579.
57. Aschim EL, Saether T, Wiger R, Grotmol T, Haugen TB. Differentialdistribution of splice variants of estrogen receptor beta in human testicularcells suggests specific functions in spermatogenesis. J Steroid BiochemMol Biol 2004; 92:97–106.
58. Gaskell TL, Robinson LLL, Groome NP, Anderson RA, Saunders PTK.Differential expression of two estrogen receptor b isoforms in the humanfetal testis during the second trimester of pregnancy. J Clin EndocrinolMetabol 2003; 88:424–432.
59. Aschim EL, Giwereman A, Stahl O, Eberhard J, Cwikiel M, NordenskjoldA, Haugen TB, Grotmol T, Giwereman YL. The RsaI polymorphism inthe estrogen receptor-b gene is associated with male infertility. J ClinEndocrinol Metab 2005; 90:5313–5343.
60. Kuiper GG, Gustafsson JA. The novel estrogen receptor-beta subtype:potential role in the cell- and promoter-specific actions of estrogens andanti-estrogens. FEBS Lett 1997; 410:87–90.
61. Varayoud J, Ramos JG, Monje L, Bosquiazzo V, Munoz-de-Toro M,Luque EH. The estrogen receptor alpha sigma3 mRNA splicing variant isdifferentially regulated by estrogen and progesterone in the rat uterus. JEndocrinol 2005; 186:51–60.
62. Saunders PTK, Miliar MR, Macpherson S, Irvine DS, Groome NP, EvansLR, Sharpe RM, Scobie GA. ERb1 and the ERb2 splice variant (ERbcx/b2) are expressed in distinct cell populations in the adult human testis. JClin Endocrinol Metab 2002; 87:2706–2715.
63. Saunders PT. Does estrogen receptor beta play a significant role in humanreproduction? Trends Endocrinol Metab 2005; 16:222–227.
64. Leung YK, Mak P, Hassan S, Shuk-Mei H. Estrogen receptor (ER)-bisoforms: a key to understanding ER-b signaling. Proc Natl Acad Sci U SA 2006; 103:13162–13167.
65. Delbes G, Levacher C, Habert R. Estrogen effects on fetal and neonataltesticular development. Reproduction 2006; 132:527–538.
66. Nie R, Zhou Q, Jassim E, Saunders PT, Hess RA. Differential expressionof estrogen receptors alpha and beta in the reproductive tracts of adult maledogs and cats. Biol Reprod 2002; 66:1161–1168.
67. Aquila S, Sisci D, Gentile M, Middea E, Catalano S, Carpino A, Rago V,Ando S. Estrogen receptor (ER)alpha and ER beta are both expressed inhuman ejaculated spermatozoa: evidence of their direct interaction with
ESTROGEN AND MAPK3/1 ACTIVATION IN SERTOLI CELL 113

phosphatidylinositol-3-OHkinase/Akt pathway. J Clin Endocrinol Metab2004; 89:1443–1451.
68. Lambard S, Galeraud-Denis I, Saunders PT, Carreau S. Human immaturegerm cells and ejaculated spermatozoa contain aromatase and oestrogenreceptors. J Mol Endocrinol 2004; 32:279–289.
69. Roger C, Lambard S, Bouskine A, Mograbi B, Chevallier D, Nebout M,Pointis G, Carreau S, Fenichel P. Estrogen-induced growth inhibition ofhuman seminoma cells expressing estrogen receptor b and aromatase. JMol Endocrinol 2005; 35:191–199.
70. Acconcia F, Ascenzi P, Bocedi A, Spisni E, Tomasi V, Trentalance A,Visca P, Marino M. Palmitoylation-dependent estrogen receptor alphamembrane localization: regulation by 17beta-estradiol. Mol Biol Cell2005; 16:231–237.
71. Song RX, McPherson RA, Adam L, Bao Y, Shupnik M, Kumar R, SantenRJ. Linkage of rapid estrogen action to MAPK activation by ERalpha-Shcassociation and Shc pathway activation. Mol Endocrinol 2002; 16:116–127.
72. Migliaccio A, Di Domenico M, Castoria G, de Falco A, Bontempo P, NolaE, Auricchio F. Tyrosine kinase/p21ras/MAP-kinase pathway activation byestradiol-receptor complex in MCF-7 cells. EMBO J 1996; 15:1292–1300.
73. Wong CW, McNally C, Nickbarg E, Komm BS, Cheskis BJ. Estrogenreceptor-interacting protein that modulates its nongenomic activity-crosstalk with Src/Erk phosphorylation cascade. Proc Natl Acad Sci U SA 2002; 99:14783–14788.
74. Vadlamudi RK, Manavathi B, Balasenthil S, Nair SS, Yang Z, Sahin AA,Kumar R. Functional implications of altered subcellular localization ofPELP1 in breast cancer cells. Cancer Res 2005; 65:7724–7732.
75. Chambliss KL, Yuhanna IS, Anderson RGW, Mendelsohn ME, ShaulPW. ERb has nongenomic action in caveolae. Mol Endocrinol 2002; 16:938–946.
76. Pietras RJ, Marquez DC, Chen HW, Tsai E, Weinberg O, Fishbein M.Estrogen and growth factor receptor interaction in human breast and non-small cell lung cancer cells. Steroids 2005; 70:372–381.
77. Fan P, Wang J, Santen RJ, Yue W. Long-term treatment with tamoxifenfacilitates translocation of estrogen receptor a out of the nucleus andenhances its interaction with EGFR in MCF-7 breast cancer cells. CancerRes 2007; 67:1352–1360.
78. Levin ER. Integration of the extranuclear and nuclear actions of estrogen.Mol Endocrinol 2005; 19:1951–1959.
79. Migliaccio A, Castoria G, Di Domenico M, Ciociola A, Lombardi M, DeFalco A, Nanayakkara M, Bottero D, De Stasio R, Varricchio L, AuricchioF. Crosstalk between EGFR and extranuclear steroid receptors. Ann N YAcad Sci 2006; 1089:194–200.
80. Song RXD, Zhang Z, Chen Y, Bao Y, Santen RJ. Estrogen signaling via alinear pathway involving insulin-like growth factor I receptor, matrixmetalloproteinases, and epidermal growth factor receptor to activatemitogen-activated protein kinase in MCF-7 breast cancer cells. Endocri-nology 2007; 148:4091–4101.
81. Endoh H, Sasaki H, Maruyama K, Takeyama K, Waga I, Shimizu T, KatoS, Kawashima H. Rapid activation of MAP kinase by estrogen in the bonecell line. Biochem Biophys Res Commun 1997; 235:99–102.
82. Bi R, Broutman G, Foy MR, Thompson RF, Baudry M. The tyrosine
kinase and mitogen-activated protein kinase pathways mediate multipleeffects of estrogen in hippocampus. Proc Natl Acad Sci U S A 2000; 97:3602–3607.
83. Di Domenico M, Castoria G, Bilancio A, Migliaccio A, Auricchio F.Estradiol activation of human colon carcinoma-derived Caco-2 cellgrowth. Cancer Res 1996; 56:4516–4521.
84. Migliaccio A, Di Domenico M, Castoria G, Nanayakkara M, Lombardi M,de Falco A, Bilancio A, Varricchio L, Ciociola A, Auricchio F. Steroidreceptor regulation of epidermal growth factor signaling through Src inbreast and prostate cancer cells: steroid antagonist action. Cancer Res2005; 65:10585–10593.
85. Nelson KG, Takahashi T, Bossert NL, Walmer DK, McLachlan JA.Epidermal growth factor replaces estrogen in the stimulation of femalegenital-tract growth and differentiation. Proc Natl Acad Sci U S A 1991;88:21–25.
86. Chen JQ, Yager JD, Russo J. Regulation of mitochondrial respiratorychain structure and function by estrogens/estrogen receptors and potentialphysiological/pathophysiological implications. Biochim Biophys Acta2005; 1746:1–17.
87. Chen JQ, Russo PA, Cooke C, Russo IH, Russo J. ERb shifts frommitochondria to nucleus during estrogen-induced neoplastic transforma-tion of human breast epithelial cells and is involved in estrogen-inducedsynthesis of mitochondrial respiratory chain proteins. Biochim BiophysActa; 29 May 2007; PMID:17604135.
88. MacCalman CD, Getsios S, Farookhi R, Blaschuk OW. Estrogenspotentiate the stimulatory effects of follicle-stimulating hormone on N-cadherin messenger ribonucleic acid levels in cultured mouse Sertoli cells.Endocrinology 1997; 138:41–48.
89. Sharpe RM, Atanassova N, McKinnell C, Parte P, Turner KJ, Fisher JS,Kerr JB, Groome NP, Macpherson S, Millar MR, Saunders PT.Abnormalities in functional development of the Sertoli cells in rats treatedneonatally with diethylstilbestrol: a possible role for estrogens in Sertolicell development. Biol Reprod 1998; 59:1084–1094.
90. Atanassova NN, Walker M, McKinnell C, Fisher JS, Sharpe RM.Evidence that androgens and oestrogens, as well as follicle-stimulatinghormone, can alter Sertoli cell number in the neonatal rat. J Endocrinol2005; 184:107–117.
91. Gancarczyk M, Paziewska-Hejmej A, Carreau S, Tabarowski Z, BilinskaB. Dose- and photoperiod-dependent effects of 17beta-estradiol and theanti-estrogen ICI 182,780 on testicular structure, acceleration ofspermatogenesis, and aromatase immunoexpression in immature bankvoles. Acta Histochem 2004; 106:269–278.
92. Robertson KM, O’Donnell L, Simpson ER, Jones ME. The phenotype ofthe aromatase knockout mouse reveals dietary phytoestrogens impactsignificantly on testis function. Endocrinology 2002; 143:2913–2921.
93. Cho HW, Nie R, Carnes K, Zhou Q, Sharief NA, Hess RA. Theantiestrogen ICI 182,780 induces early effects on the adult male mousereproductive tract and long-term decreased fertility without testicularatrophy. Reprod Biol Endocrinol 2003; 1:1–17.
94. Cheng J, Watkins SC, Walker WH. Testosterone activates mitogen-activated protein kinase via Src kinase and the epidermal growth factorreceptor in sertoli cells. Endocrinology 2007; 148:2066–2074.
114 LUCAS ET AL.
Copyright © 2022 FDOKUMEN




















