
1 Rita Armonienė FREEZING TOLERANCE IN WINTER WHEAT
25
ALEKSANDRAS STULGINSKIS UNIVERSITY LITHUANIAN RESEARCH CENTRE FOR AGRICULTURE AND FORESTRY Rita Armonienė FREEZING TOLERANCE IN WINTER WHEAT: GENE IDENTIFICATION AND ANALYSIS Summary of doctoral dissertation Agricultural sciences, agronomy (01A) Akademija, 2014
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of 1 Rita Armonienė FREEZING TOLERANCE IN WINTER WHEAT

1
ALEKSANDRAS STULGINSKIS UNIVERSITY
LITHUANIAN RESEARCH CENTRE FOR AGRICULTURE AND FORESTRY
Rita Armonienė
FREEZING TOLERANCE IN WINTER WHEAT: GENE IDENTIFICATION AND ANALYSIS
Summary of doctoral dissertation
Agricultural sciences, agronomy (01A)
Akademija, 2014

2
The research was carried out at the Lithuanian Research Centre for Agriculture and Forestry, Institute of Agriculture in 2010-2014.
Scientific supervisor: Dr. Gintaras Brazauskas (Lithuanian Research Centre for Agriculture and
Forestry, Agricultural sciences, Agronomy (01A) The dissertation will be defended at the Council of Agronomy
Sciences: Chairman: Prof. habil. dr. Pavelas Duchovskis (Lithuanian Research Centre for
Agriculture and Forestry, Agricultural Sciences, Agronomy (01A) Members: Prof. habil. dr. Izolda Pašakinskienė (Vilnius University, Biomedical
Science, Biology 01B) Dr. Danas Baniulis (Lithuanian Research Centre for Agriculture and
Forestry, Agricultural Sciences, Agronomy (01A) Dr. Algė Leistrumaitė (Lithuanian Research Centre for Agriculture and
Forestry, Agricultural Sciences, Agronomy (01A) Dr. Nils Rostoks (University of Latvia, Biomedical Science, Biology
01B) Defence of the doctoral dissertation will take place at the public meeting
of the Council of Agronomy Science on the 4th of December, 2014 at 13.00 p.m., in the auditorium 211 (4 building) of Aleksandras Stulginskis University.
The summary of the doctoral dissertation was distributed on the 4th of November, 2014.
The doctoral dissertation and summary is available at the Libraries of Lithuanian Research Centre of Agriculture and Forestry and Aleksandras Stulginskis University.

3
ALEKSANDRO STULGINSKIO UNIVERSITETAS
LIETUVOS AGRARINIŲ IR MIŠKŲ MOKSLŲ CENTRAS
Rita Armonienė
ŽIEMINIŲ KVIEČIŲ TOLERANTIŠKUMAS ŽEMOMS TEMPERATŪROMS: GENŲ PAIEŠKA IR ANALIZĖ
Daktaro disertacijos santrauka
Žemės ūkio mokslai, agronomija (01A)
Akademija, 2014

4
Disertacija rengta 2010-2014 metais Lietuvos agrarinių ir miškų mokslų centro filiale Žemdirbystės institute.
Mokslinis vadovas: Dr. Gintaras Brazauskas (Lietuvos agrarinių ir miškų mokslų centras,
Žemės ūkio mokslai, agronomija – 01A)
Disertacija ginama Agronomijos mokslo krypties taryboje:
Pirmininkas: Prof. habil. dr. Pavelas Duchovskis (Lietuvos agrarinių ir miškų mokslų
centras, Žemės ūkio mokslai, agronomija – 01A)
Nariai: Prof. habil. dr. Izolda Pašakinskienė (Vilniaus universitetas,
biomedicinos mokslai, biologija 01B) Dr. Danas Baniulis (Lietuvos agrarinių ir miškų mokslų centras, Žemės
ūkio mokslai, agronomija – 01A) Dr. Algė Leistrumaitė (Lietuvos agrarinių ir miškų mokslų centras,
Žemės ūkio mokslai, agronomija – 01A) Dr. Nils Rostoks (Latvijos universitetas, biomedicinos mokslai, biologija
01B)
Disertacija bus ginama viešame Agronomijos krypties tarybos posėdyje 2014 m. gruodžio mėn. 4 d. 13 val. Aleksandro Stulginskio universiteto IV rūmų posėdžių salėje (211 kab.).
Adresas: Universiteto g. 8A, Akademija, 53361 Kauno r. Disertacijos santrauka išsiųsta 2014 m. lapkričio mėn. 4 d. Disertaciją ir jos santrauką galima peržiūrėti Lietuvos agrarinių ir miškų
mokslų centro ir Aleksandro Stulginskio universiteto bibliotekose.

5
INTRODUCTION
Bread wheat (Triticum aestivum L.) is one of the most widely grown crop in the world, covering more than 200 million hectares. Typically, winter wheat has a higher yield per planted hectare than spring wheat in Lithuania. For this reason winter wheat has a larger cultivation area in Lithuania. Twenty percent of the total cultivated area is occupied by winter wheat, while spring wheat covers only 9 percent. The capability of autumn-seeded varieties to survive winter is often referred to as winter hardiness. Winter hardiness is one of the limiting factors for winter wheat cultivation under Lithuanian conditions. Winter hardiness is a complex trait involving tolerance to freezing, desiccation, anoxia, ice-encasement, resistance to diseases, etc. However, tolerance to low freezing temperatures (freezing tolerance) has been considered as the primary limiting factor in most regions, especially in winters with the fluctuating weather conditions.
Freezing tolerance can be defined as the ability of plants to survive freezing temperatures, prevent damage to the vegetative tissues and minimize other negative effects of freezing temperatures on future yield potential. In winter cereals, frost tolerance is associated with the occurrence of cold acclimation, which is triggered by the induction of the battery of Cor genes after exposure of plants to low but non-freezing temperature for certain periods of time. In particular, Cor genes are transcriptionally activated during cold acclimation, and the accumulated COR proteins lead to the protection of the integrity of cell structures and functionality from freezing damage.
Genetic and molecular mechanisms of freezing tolerance formation in wheat and other plants are widely studied worldwide. Large numbers of genes have been identified, which are induced during cold acclimation and play some role in freezing tolerance of plants. However, the genetic basis of the freezing tolerance induction mechanism, which is determined not only by the interaction among genes, but also by the complex of environmental factors, is still poorly understood. In recent years, in Lithuania freezing tolerance was studied in Fragaria ananassa, Cydonia oblonga, Brasica napus, Miscanthus (Rugienius and Stanys, 2001; Rugienius et al., 2008), Prunus sp. (Zalunskaitė et al., 2008), Triticum aestivum (Ruzgas and Liutkevičius, 2001; Liatukas et al., 2012) and Festuca pratensis (Darginavičienė et al., 2008). The research objective of these studies was to identify the physiological response of plants to low temperatures, however expression of the genetic factors had been studied only in F. ananassa (Lukoševičiūtė et al., 2009; Zaluskaitė et al., 2008) and Prunus sp .(Zalunskaitė et al., 2008) plants.
According to the Ministry of Agriculture, 30% to 50% of the winter wheat crop was killed in two subsequent winters of 2010-2011 and 2009-2010. The global warming will lead to extreme natural conditions during winters and these

6
events will become more frequent and unpredictable (Rapacz et al., 2014), therefore it is necessary to study winter wheat tolerance to freezing temperatures, to identify new genes which would allow a better understanding of the genetic regulation of this process and facilitate the design of DNA markers for molecular selection of freezing tolerant winter wheat genotypes.
Hypothesis
Genes that are differentially expressed during cold acclimation lead to different freezing tolerance of winter wheat.
Research objective To identify new genes of winter wheat induced at low temperatures, to
perform sequence analysis of these genes and to quantify their expression. Experimental tasks:
1. To evaluate freezing tolerance of various winter wheat genotypes under field and controlled conditions. 2. To identify differentially expressed genes at subsequent time points of cold acclimation and to perform analysis of their DNA sequences. 3. To optimize mutagenesis method and to create Targeting Induced Local Lesions in Genomes (TILLING) population of two winter wheat lines. 4. To identify mutations in differentially expressed genes by High Resolution melting (HRM) method, to estimate the impact of mutations on the gene expression during cold acclimation and tolerance to freezing.
Statements to be defended: 1. There are two separate genetic systems of freezing tolerance in winter wheat which determine the duration and the rate of acclimation. 2. cDNA-AFLP technique is suitable for the identification of differentially expressed fragments (DEF) in cold acclimated and non-acclimated wheat. 3. Appropriate mutagen concentrations should be optimized for each winter wheat line to achieve a substantial mutation rate while avoiding serious defects in germination and plant development. 4. Premature stop codon mutation in the Sucrose synthase1 gene significantly reduces it’s expression during cold acclimation and decreases freezing tolerance of the mutant genotype.
Novelty of the research: Results obtained in this study support Fowler and Limin‘s (2007) hypothesis
of two separate genetic systems controlling freezing tolerance in winter wheat which determine the duration and the rate of cold acclimation.
Twenty TDFs were identified as being differentially expressed between cold acclimated and non-acclimated wheat, showing 14 gene homologies to GenBank sequences. Thirteen of these homologies were for the first time shown to be involved in the cold acclimation process. Twelve of these genes had not been available in public databases of wheat sequences.

7
TILLING populations of two winter wheat lines ‘5899-16’ and ‘5450-1’ were created. Two novel alleles of Ss1 gene were identified. Premature stop codon mutation in the Ss1 gene was found to reduce the Ss1 gene expression during cold acclimation and have negative effect on freezing tolerance. Practical application
Identified candidate genes showing differences in expression intensity between cold acclimated and non-acclimated winter wheat crown and leaf tissue are putative targets in marker assisted selection for freezing tolerance in winter wheat. A new TILLING population developed on advanced local breeding material will serve as a valuable source of novel alleles in winter wheat both for practical breeding and applied genetic research purposes. Approval of the experimental results
The main research results were published in 2 scientific articles refereed in the ISI WOS database and as 6 summaries in conference proceedings. One poster presentation and one oral presentation were made at international congresses. Volume and structure of the dissertation
The dissertation is written in Lithuanian. It is composed of an introduction, literature overview, description of materials and methods, experimental results and discussion, conclusions, list of references and list of publications produced by the author in cooperation with co-authors. The dissertation comprises 86 pages, research results are presented in 6 tables and 17 figures, a total of 214 literature references have been cited.
MATERIAL AND METHODS Plant material. Five winter wheat varieties (‘Kovas DS’, ‘Zunda DS’,
‘Kaskada DS’, ‘Vikaras DS’,‘Ada’) and 21 breeding lines (WW-8, WW-13, WW-14, WW-15, WW-18DH, WW-27, WW-29, WW-30, WW- 31, WW-32, WW-35, WW-45, WW-62DH, WW-65DH, WW-66DH, WW-67DH, WW-68DH, WW-70, WW-71, WW-75DH, WW-76DH) developed at the Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, and variety ‘Zentos’ (Lantmännen SW Seed, Germany) as a standard were chosen for evaluation freezing tolerance under field and controlled conditions with cold acclimation and without cold acclimation stage.
Two winter wheat breeding lines ‘5450-1’ and ‘5899-16’ developed at the Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry were used for cDNA-AFLP analysis and generation of TILLING population.
Field experiment. Field experiments were set up in a nine-course crop rotation in Dotnuva during 2010–2012. Winter wheat varieties/lines were sown in 3.6 m2 plots using randomized design with four replications in 2010–2011 and with three replications in 2011–2012 at a seed rate of 4.5 million ha-1. The

8
seeds were not pesticide-treated. The evaluation of winter hardiness in natural conditions was conducted in April of 2011 and 2012 by visual scoring on a 1–9 score basis (Ruzgas, Liutkevičius, 2001). Score 1 means that all plants were killed, while score 9 denotes that all plants have survived.
Laboratory freezing test. The seeds of 27 winter wheat varieties/lines were germinated on the moist filter paper in the Petri dishes. After three days, germinated seeds were sown in plastic pots (225 cm3) filled with universal peat substrate GP0428 (Durpeta, Lithuania). Each genotype was planted in six replications. Plants were grown at 18 °C in a greenhouse with a 16 h photoperiod until the seedlings reached a three-leaf stage. The first experiment included cold acclimation period at 4 °C with an 8 h photoperiod for five weeks in a growth chamber. The second experiment was conducted without cold acclimation period. Freezing test was carried out as described earlier (Skinner, Mackey, 2009) with minor modifications. Freezing tests were carried out in a programmable freezing chamber PE 2412 UY-LX (Angelantoni Industrie, Italy). Plants were kept at 4 °C for one hour, then the temperature in the freezer was reduced from 4 °C to −12 °C over an 8 h period. The temperature was held at the target temperature for 2 h, then raised to 4 °C over 8 h and subsequently maintained at 4 °C for 24 h. After the freezing test, the plants were moved to the greenhouse. Freezing test survival was scored as the proportion of the plants that had re-grown after two weeks.
Fv/Fm measurement. The Fv/Fm was determined in leaves with a hand-held pulse modulated fluorometer OS-30p (Opti-Science Inc., USA) using the Fv/Fm kinetics option. Leaves were cut off after freezing tests and stored in black bags at 4 °C for 24 h. The measurements were taken in 5 replications (leaves) in 5 different spots of the leaf with dark-adaptation clips.
Meteorological conditions. The two field experiments performed during autumn-winter seasons in 2010–2011 and 2011–2012 had very different meteorological conditions. In 2010–2011, the autumn–winter seasons was characterized by a wet second half of autumn and a cold, snowy winter with thaws. In 2011–2012, the autumn–winter seasons hada dry autumn, unusually warm first half of winter and very low temperatures by the end of winter.
Generation of wheat TILLING library. Seeds of ‘5450-1’ and ‘5899-16’ lines were treated with EMS solution with concentrations ranging from 0.5 to 1.0% according to Slade et al. (2005) with minor modifications. Seeds were allowed to germinate in the greenhouse and the number of germinating seeds was scored. The number of survived seedlings was scored two months after treatment. The mutagenized seeds were grown into M1 plants and allowed to self-pollinate. M1 population was advanced to M2. M3 seeds from the individual M2 plants were collected and catalogued.
Plant material for cDNA-AFLP and qRT-PCR. Seeds of winter wheat breeding lines ‘5450-1’ and ‘5899-16’ were sown in plastic pots (125 cm3) filled

9
with universal peat substrate GP0428 (Durpeta, Lithuania). Plants were grown at 20 °C in a plant growth chamber (CLF Plant Climatics, Germany) with a 12 h photoperiod at a light intensity of 600 µmol m-2 s-1 at 70% relative air humidity until the seedlings reached a three-leaf stage. The plants were then cold acclimated by transferring them to a growth chamber set at 5 °C with an 8-h photoperiod at a light intensity of 200 µmol m-2 s-1 and 80% relative air humidity for six weeks. Leaves and crowns were collected in three biological replications at 0, 2, 4 and 6 weeks of cold acclimation, flash frozen in liquid nitrogen and stored at −80 °C until RNA extractions.
For qRT-PCR analysis seeds of wheat line ‘5899-16’ and two mutant lines M631, M692 of M3 generation were sown and samples were collected under the same conditions. All M3 generation seedlings were tested by High Resolution Melting (HRM) analysis for the presence of the mutation.
cDNA-AFLP analysis. The cDNA-AFLP procedure was conducted as described earlier (Jonavičienė et al., 2012) with minor modifications. Selective amplification PCR mix contained 10 μl of 20x diluted pre–amplification product, 0.25 μM of AseI selective primer with two selective nucleotides, 1.5 μM of TaqI selective primer with two selective nucleotides, 0.2 mM dNTP mix, and 0.5 U DreamTaq DNA polymerase (Thermo Scientific, Lithuania) in a 20 μl total reaction volume. Products of selective PCR were fractionated in a 2% agarose gel by electrophoresis at 100 W for 2 h. The fragments were visualized using ethidium bromide and UV light. The reproducibility of transcript-derived fragments (TDFs) was tested by repeating PCR amplifications 2 to 3 times for each primer combination used.
Isolation, cloning and sequence analysis of TDFs. TDFs based on the presence/absence in the samples under comparison were excised from the gel with a surgical blade, avoiding any contaminating fragments, purified with a GeneJETTM Gel Extraction Kit (Thermo Scientific, Lithuania) and eluted with 20 μl of sterile water. Re-amplified DNA fragments were cloned into the pJET1.2/blunt cloning vector using GeneJetTM PCR Cloning Kit (Thermo Scientific, Lithuania) according to the manufacturer’s instructions. Insert sizes were checked by colony PCR and restriction enzyme digestion. Plasmid DNA was isolated with GeneJetTM Plasmid Miniprep Kit (Thermo Scientific, Lithuania) and sequenced at the GATC Biotech (Köln, Germany). At least two clones for each re-amplified TDF were DNA sequenced to ascertain integrity of the excised TDF. Sequence homology analysis of the fragments was carried out against the publicly available non-redundant genes/transcripts (http://www.ncbi.nlm.nih.gov/BLAST) using the BLASTX algorithm.
Genomic DNA isolation. Genomic DNA was extracted from 0.2 g samples of wild type ‘5450-1’ and ‘5899-16’ and M2 generation leaf material according to Lassner et al. (1989) with several modifications. These included additional wash step with chloroform and RNA digestion with ribonuclease A. The DNA

10
was quantified, diluted to 20 ng/μL in deionised water and two-fold pooled according to Botticella et al. (2011).
HRM analysis. Oligonucleotide primers for the target Ss1 gene were designed according to the DNA sequences of extracted TDFs using PrimerBlast (http://www.ncbi.nlm.nih.gov). Primers were selected to specifically amplify 120 bp fragment of Sucrose synthase type 1 gene. The sequences of primers were 5’-TCCTCGGTGTATGTCTCCAG-3’ and 5’-GGTTATTGGAACCGAGCACA-3’ for forward and reverse primer, respectively. HRM analysis was carried out using Melt Doctor HRM Master mix (Life technologies, USA) following manufacturer’s instructions. The PCR was carried out on a 7500 Fast Real-Time PCR System (Life technologies, USA). The reaction conditions were as follows. Enzyme activation was performed at 95 °C for 10 min, followed by two step (denature at 95 °C for 15 s and annealing/extension at 60 °C for 1 min) cycling of 40 cycles. Melt Curve/dissociation analysis was done by denaturation at 95 °C for 10 s, annealing at 60 °C for 1 min, high resolution melting at 95 °C followed by annealing at 60 °C for 15 s. Individuals were genotyped using HRM V.2.0.2 software (Life technologies, USA). Samples identified as putative mutants were selected, re-amplified for each individual in the pool and sequenced at GATC Biotech (Köln, Germany) for the mutation identification.
qRT-PCR and data analysis. Total RNA from leaf and crown tissue was extracted with Rneasy Plant Mini Kit (QIAGEN, Germany) following the manufacturer's protocol. The on-column DNase digestion was performed using RNase-free DNase (QIAGEN, Hilden, Germany). 1 μg aliquot of RNA samples was reverse transcribed with RevertAid™ First Strand cDNA Synthesis Kit (Thermo Scientific, Lithuania). qRT-PCR was performed in triplicate with the primers used in HRM on a 7500 Fast Real-time PCR system (Applied Biosystems, USA) with a SYBR Green PCR master mix kit (Applied Biosystems, USA). The PCR thermal cycling parameters were 50 °C for 2 min, 95 °C for 10 min followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min. The wheat ADP–ribosylation factor gene (Ta2991) was used as an endogenous control using primers developed by Paolacci et al. (2009). Relative expression of the genes of interest was calculated by the 2 –ΔΔCt method (Livak and Schmittgen, 2001) using a control wheat line ‘5899-16’ at 0 week of cold acclimation as calibrator. For both crown and leaf tissue 3 biological replicates x 3 technical replicates were used in qRT-PCR experiments.
The freezing tolerance test for LT50 estimation. The seeds of wild type ‘5899-16’ and M631 were sown in plastic pots (125 cm3) filled with universal peat substrate GP0428 (Durpeta, Lithuania). Plants were grown at 20 °C in a plant growth chamber (CLF Plant Climatics, Germany) with a 12 h photoperiod at a light intensity of 600 µmol m-2 s-1 at 70% relative air humidity until the seedlings reached a three-leaf stage. The plants were then cold acclimated by

11
transferring them to a growth chamber set at 5 °C with an 8 h photoperiod at a light intensity of 200 µmol m-2 s-1 and 80% relative air humidity for four weeks. Freezing tolerance tests were conducted at 0, 2, and 4 weeks of acclimation. Roots and leaves of the wheat plants were trimmed to approx. 6 cm crown and placed in glass tubes containing 0.5 ml of water. Samples were transferred to programmable freezing chamber. Plants were cooled at rate of 1.2 °C h-1 down to −16 °C. Eight crowns were removed at 2 °C intervals from −6 °C to −16 °C for each genotype in each treatment. Sampled crowns were allowed to thaw overnight at 5 °C, planted in peat substrate and transferred to a growth chamber set at 20 °C and 16 h photoperiod. Two weeks later surviving plants were counted and LT50 values were determined.
Data analyses. Statistical analysis was performed using Statistica 7 software (StatSoft Inc., USA). Analysis of variance (ANOVA) was used to determine significance (p < 0.01 and p < 0.05) of the differences between wheat genotypes, EMS concentrations and relative gene expression. Spearman correlation coefficients between mean values of the investigated traits were calculated. The differences between means of wheat genotypes were inspected using Fisher’s protected significant differences post hoc analysis.
The mutation density was calculated as follows (Botticella et al., 2011):
푀푢푡푎푡푖표푛 푑푒푛푠푖푡푦 =(푡표푡푎푙 푠푖푧푒 표푓 푎푚푝푙푖푐표푛)푥 (푡표푡푎푙 푛푢푚푏푒푟 표푓푠푐푟푒푒푛푒푑 푙푖푛푒푠)
(푛푢푚푏푒푟 표푓 푖푑푒푛푡푖푓푖푒푑 푚푢푡푎푡푖표푛푠)
Relative gene expression was calculated as follows (Livak and Schmittgen, 2001):
2 ∆∆ , Ct–number of threshold cycles;
∆C = (C , 푆푠1 − C , 푇푎2291); Ss1 – gene of interest, Ta2291 – housekeeping gene,
∆∆C = ∆C X − C Y, X – ΔCt of cold acclimated and non-acclimated wt‘5899-16’, M631 and
M69l, Y – the ΔCt mean value of biological replications of non-acclimated
wt‘5899-16’. LT50 was estimated according Limin and Fowler, (2002) using non-linear
regression procedures by Statistica10 software (Stat Soft Inc., USA).

12
RESULTS AND DISCUSSION Winter hardiness in the field. The 2010–2011 winter was favourable for
winter hardiness evaluation of wheat genotypes due to the wet autumn, sharp temperature fluctuations with frost and thaw periods and shifting snow cover. Some winter wheat lines were completely killed. Winter hardiness scores ranged from 1.0 (WW-8) to 7.6 (‘Zentos’) (Fig. 1). Winter wheat variety ‘Ada’ also showed high score of winter hardiness (6.6), while most plants of ‘Zunda DS’ were killed. Analysis of variance indicated significant differences in winter hardiness (p < 0.05) between doubled haploid and conventionally-bred wheat genotypes. The majority of doubled haploid wheat lines were completely killed during the 2010–2011 winter. The 2011–2012 winter was not favourable for the evaluation of winter hardiness of wheat genotypes due to beneficial weather conditions for cold acclimation during autumn of 2011 and sufficient snow cover during freezing period in February, 2012. Furthermore, no periods of warming and subsequent deacclimation of plants were observed that year. All wheat genotypes survived the winter of 2011–2012.Winter hardiness scores ranged from 5.8 (WW-31) to 7.8 (‘Kovas DS’) (Fig. 1). No significant differences in winter hardiness (p > 0.05) were found between doubled haploid and conventionally-bred wheat lines.
Wheat genotypes under investigation experienced very different freezing conditions in the field during the two winters. Consequently, no significant correlation for wheat winter hardiness was observed between years (Table 1). Variation coefficients for winter hardiness was 13.3% and 3.5% in 2010–2011and 2011–2012, respectively, suggesting a strong interaction between the hardiness of genotypes investigated and weather conditions experienced.
Freezing tolerance in the laboratory test. Analysis of variance indicated significant differences of Fv/Fm ratio and freezing test survival (p < 0.01) among wheat genotypes. Freezing test survival of non-acclimated plants varied from 20% (WW-31) to 100% (WW-68DH). Plants with high survival ratio also had the highest Fv/Fm ratio. The freezing susceptible wheat plants had an average Fv/Fm ratio value of 0.21 (WW-31), while the freezing tolerant plants had an average Fv/Fm ratio value of 0.77 (WW-67DH) and 0.76 (WW-68DH). Results of our study also show that the ratio of the maximum variable to maximum total fluorescence in dark-adapted state, Fv/Fm, is an efficient method for frost tolerance evaluation in wheat under controlled conditions due to the high correlation between freezing test survival of non-acclimated wheat and Fv/Fm parameter (r = 0.88, p < 0.01). This relationship has also been demonstrated in oat (r = 0.87) (Rizza et al., 2001) and barley (r = 0.91) (Rapacz et al., 2008) under controlled conditions. Rapacz and Wozniczka (2009) have shown a high negative correlation between electrolyte leakage results after freezing test in controlled conditions and Fv/Fm parameter (r = −0.87).

13
Figure 1.Freezing tolerance of wheat genotypes in the field and laboratory experiments: winter hardiness of wheat lines/varieties in 2010–2011 (A) and 2011–2012 (B) field experiments, freezing test survival (C) and Fv/Fm ratio (D) of acclimated wheat lines/varieties, freezing test survival (E) and Fv/Fm ratio (F) of non-acclimated wheat lines/ varieties in laboratory experiments. Notes.WH – winter hardiness, FTS – freezing test survival. Vertical bars represent ± standard deviation (SD) of the mean. The letters above the boxes indicate statistically significant (p < 0.01) differences between winter wheat genotypes.

14
Freezing test of acclimated plants shows no significant differences (p > 0.0) for freezing tolerance between winter wheat genotypes. Freezing test survival varied from 50% (‘Kovas DS’) to 100% (WW-67DH). Average Fv/Fm values varied from 0.55 (‘Kovas DS’) to 0.75 (‘Vikaras DS’). The correlation coefficient between the Fv/Fm parameter and freezing test survival of acclimated plants was 0.41( p < 0.05).
Table 1. Matrix of Spearman correlation coefficients between winter hardiness of two winters in field experiments, freezing test survival and Fv/Fm ratio in laboratory experiments with hardening stage and without hardening stage.
WH 2010/2011
WH 2011/2012
FTS with hardening
Fv/Fm with
hardening
FTS without
hardening
Fv/Fm without
hardening WH 2010/2011 1.00 0.24 0.31 0.24 -0.41* -0.38* WH 2011/2012 1.00 0.15 0.05 0.12 0.14 FTS with hardening 1.00 0.41* 0.10 0.26 Fv/Fm with hardening 1.00 -0.05 -0.00 FTS without hardening 1.00 0.88** Fv/Fm without hardening 1.00
* – p < 0.05, ** – p < 0.01; WH – winter hardiness, FTS – freezing test survival
Low negative, but significant relationship (r = −0.41, p < 0.05) was detected between hardiness scores in 2010–2011 winter and freezing test survival of non-acclimated plants (Table 1). Some of the cultivars with the highest 2010–2011 winter hardiness, ‘Ada’ and WW-32, showed one of the lowest freezing test survival in the freezing test with no acclimation, respectively 25% and 20%. However, ‘Zunda DS’, WW-8DH and some of the other DH wheat lines with the lowest 2010–2011 winter hardiness had one of the highest freezing test survival after freezing with no acclimation (Fig. 1). Our results supports hypothesis of two separate genetic systems of freezing tolerance which are determined by the expression of duration and rate genes (Fowler and Limin, 2007).
cDNA-AFLP analysis of differentially expressed genes. cDNA-AFLP was performed using 48 primer pair combinations, which generated 522 transcript-derived fragments (TDFs) with an average of 11 fragments per primer pair. The size of the TDFs were in the range of 200–2000 bp. Twenty TDFs (3.8%) were identified as being differentially expressed (presence/absence) between cold acclimated and non-acclimated wheat (Table 2). Thirteen TDFs (65%) were up-

15
regulated and 7 TDFs (35%) were down-regulated within 2 weeks after the start of the cold acclimation. About 85% of TDFs exhibited up-regulation in leaf tissue compared to 15% in crown tissue. All of the down-regulated TDFs were detected in leaf tissue profiles. Twenty TDFs were extracted from agarose gels, cloned and sequenced. A total of fourteen TDFs revealed significant (E value < 1e-10) sequence similarities in a BLASTX search against the non-redundant (nr) protein database of GenBank (Table 2).
Table 2. Functional annotation of the 14 TDFs revealing significant (E < 1e-10) sequence similarities in BLASTX search against the non-redundant (nr) protein database of GenBank along with the TDF size and accession numbers in GenBank protein database.
Development of TILLING population. Concentration of the mutagen EMS
had an effect on both germination and development of wheat seedlings. A significant negative relationship (r = −0.59, p < 0.01) between EMS concentration and germination at the 15th day after sowing (when the number of germination reached a plateau) was observed in both genotypes of winter wheat. Analysis of the results of wheat germination average revealed significant differences (p < 0.05) between different EMS concentrations and wheat genotypes. Germination average ranged from 79.9% to 63.3% and from 80.7% to 72.3% in ‘5450-1’ and ‘5899-16’, respectively (Fig. 2). ‘5899-16’ genotype showed significantly (p < 0.01) higher germination rate at the highest (1.0%) EMS concentration treatment compared to ‘5450-1’.
TDF identifier
Plant tissue
Status after 2 weeks of cold acclimation
Homologous protein Species
TDF01 Leaf upregulated Sucrose synthase 1 A. tauschii TDF02 Leaf upregulated Heat-shock protein S. cereale TDF03 Leaf upregulated Serine carboxypeptidase-like A. tauschii TDF04 Leaf upregulated Vacoular-sorting receptor 1 A. tauschii TDF05 Leaf upregulated Zinc finger AN1 domain T. urartu TDF06 Crown upregulated Beta-3-tubulin T. aestivum TDF07 Crown upregulated Phototropin-2-like B. distachyon TDF08 Crown upregulated Digalactosyl diacyl glycerol
synthase 2, chloroplastic T. urartu
TDF09 Leaf downregulated Phosphoglycerate kinase, chloroplastic
A. tauschii
TDF10 Leaf downregulated Signal recognition particle 54 kD protein, chloroplastic
B. distachyon
TDF11 Leaf downregulated Predicted protein H. vulgare TDF12 Leaf downregulated ATP-depentent Clp proteaze
ATP-binding subunit ClpC homolog 1
O. sativa
TDF13 Leaf downregulated Clp-P proteaze subunit T. aestivum TDF14 Leaf downregulated RuBisCO activase A T. urartu
16
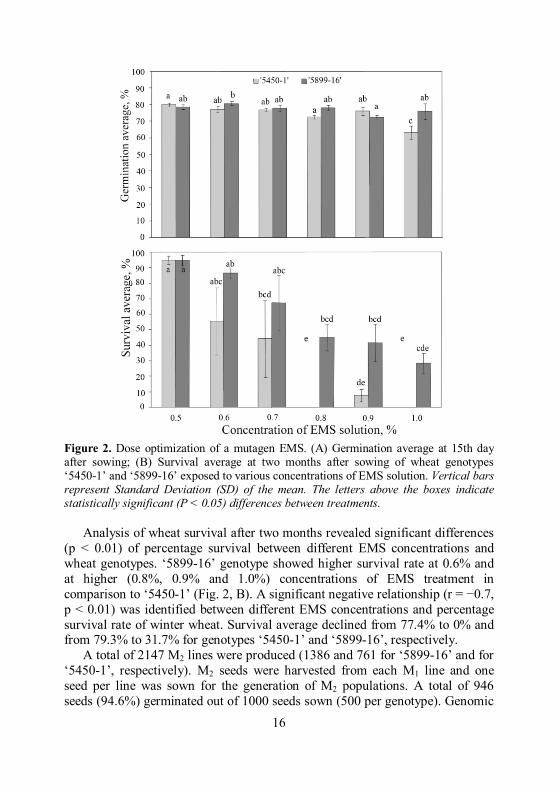
Figure 2. Dose optimization of a mutagen EMS. (A) Germination average at 15th day after sowing; (B) Survival average at two months after sowing of wheat genotypes ‘5450-1’ and ‘5899-16’ exposed to various concentrations of EMS solution. Vertical bars represent Standard Deviation (SD) of the mean. The letters above the boxes indicate statistically significant (P < 0.05) differences between treatments.
Analysis of wheat survival after two months revealed significant differences
(p < 0.01) of percentage survival between different EMS concentrations and wheat genotypes. ‘5899-16’ genotype showed higher survival rate at 0.6% and at higher (0.8%, 0.9% and 1.0%) concentrations of EMS treatment in comparison to ‘5450-1’ (Fig. 2, B). A significant negative relationship (r = −0.7, p < 0.01) was identified between different EMS concentrations and percentage survival rate of winter wheat. Survival average declined from 77.4% to 0% and from 79.3% to 31.7% for genotypes ‘5450-1’ and ‘5899-16’, respectively.
A total of 2147 M2 lines were produced (1386 and 761 for ‘5899-16’ and for ‘5450-1’, respectively). M2 seeds were harvested from each M1 line and one seed per line was sown for the generation of M2 populations. A total of 946 seeds (94.6%) germinated out of 1000 seeds sown (500 per genotype). Genomic

17
DNA was extracted from the leaf material of the four-week-old plants from 486 of the ‘5450-1’ line and from 460 of the ‘5899-16’ line in the M2 population. Seeds were harvested from each of the 756 fertile lines. These seeds make up the M3 TILLING-population.
Mutant discovery by HRM analysis. Exon 8 of differentially expressed Sucrose synthase 1 (Ss1) gene was chosen for mutation screening by HRM analysis in wheat TILLING M2 population. A total of 75.68 kb of DNA was tested and two putative mutants were identified.
Figure 3. HRM analysis of 96 2-fold pooled samples. Aligned curves of fluorescence vs temperature showing wild type (blue) and mutant (red) samples: A) M692; B) M631.
These 2-fold pooled samples of putative mutants had peaks shifted towards a lower temperature (reduced by 0.3 °C) compared to the wild type genotype (Fig. 3). HRM analysis was repeated in separate wells for each sample with predicted mutation. As expected, only single samples of the 2-fold pooled samples showed lower melting temperature. Sequencing results revealed two novel alleles of the Ss1 gene. Both mutations were transitions of the G > A type as

18
expected for the EMS mutagenesis, which acts via alkylation of G residues. Mutant No. 692 was found to have silent mutation (G1119A, K373K) and mutant No. 631 had nonsense (premature stop codon) mutation (G1122A, W374*). Both mutations were in the homozygous state.
An overall mutation density of one mutation per 37.84 Kb of the DNA was observed in our TILLING population, thus appears to be in the same range as previously determined for hexaploid wheat (1 mutation per 40 kb) by HRM (Botticella et al., 2011). This is in agreement with mutation density of 1/40kb and 1/38 kb identified in tetraploid (Slade et al., 2005) and hexaploid (Uauy et al., 2009) wheat, respectively, by traditional TILLING screening based on Cel 1 nuclease digestion.
qRT-PCR analysis of Ss1 gene. qRT-PCR was performed to evaluate the effect of the identified mutations on the expression of the Ss1 gene in crown and leaf tissues during cold acclimation. qRT-PCR analysis showed that premature stop codon mutation has a strong effect on Ss1 gene expression (Fig. 4). Putative knock-out mutant M631 had significantly lower relative expression of Ss1 gene in non-acclimated leaves as well as in crowns and leaves collected at 2, 4 and 6 weeks of cold acclimation compared to the wild type winter wheat line ‘5899-16’ (control) and M692 mutant (silent mutation).
Ss1 gene was strongly repressed during the whole cold-acclimation period in leaf and crown tissues of M631. It was approximately 7-fold and 18-fold lower in leaves and in crowns of M631, respectively, in comparison to the wild type and M692. Hence, premature stop codon in the Ss1 gene probably resulted in a nonsense mediated mRNA decay (NMD) of the aberrant mRNA. NMD is usually induced if the distance between the premature termination codon and the 3‘-end of the transcript exceeds 350 nt (Kalyna et al., 2012). Location of the premature stop codon in our mutant complied with this condition. In wheat, reduced RNA levels have been reported for multiple genes containing premature stop codon mutations including a waxy gene (Saito and Nakamura, 2005), a SbeIIa (Botticella et al., 2011; Slade et al., 2012), and a Vrn1 gene (Chen and Dubcovsky, 2012). Horst et al. (2007) detected premature stop codons in the sucrose synthase LjSus1 and LjSus3 genes of Lotus japonicus. Interestingly, the mutations were in the same exon as in our study. Both mutants (sus1-1 and sus3-1) showed reduced Ss activity. sus3-1 plants had decreased Ss content in the nodules and exhibited defective growth in comparison to the wild type plants (Horst et al., 2007).

19
Figure 4. Relative expression of Ss1 gene during cold acclimation in (A) leaves and (B) crowns of wild type genotype ‘5899-16’ and two mutant genotypes M692 and M631 as revealed by qRT-PCR. Total RNA was extracted from seedling leaves acclimated for 0–6 weeks at 5 °C.ADP–ribosylation factor gene (Ta2991) was used as endogenous control. Vertical bars represent Standard Deviation (SD) of the mean.
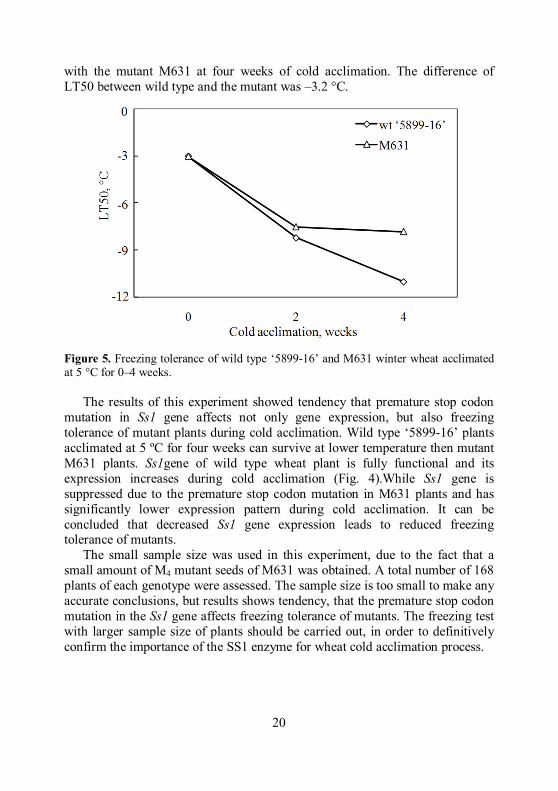
Effect of the premature stop codon mutation in Ss1 gene on the freezing
tolerance. Freezing tolerance test of the wild type and M631winter wheat was performed to determine the effect of premature stop codon mutation in Ss1 gene on the freezing tolerance development during cold acclimation and to confirm the role of Ss1 in wheat cold acclimation process. The two genotypes of winter wheat showed different freezing tolerance at prolonged period of cold acclimation at 5 °C (Fig. 5). There was no difference in LT50 between non-acclimated wild type and mutant plants. Freezing tolerance of both wheat genotypes significantly increased after two weeks of cold acclimation. Meanwhile, the ‘5899-16’ genotype showed higher freezing tolerance compared
20
with the mutant M631 at four weeks of cold acclimation. The difference of LT50 between wild type and the mutant was –3.2 °C.
Figure 5. Freezing tolerance of wild type ‘5899-16’ and M631 winter wheat acclimated at 5 °C for 0–4 weeks.
The results of this experiment showed tendency that premature stop codon mutation in Ss1 gene affects not only gene expression, but also freezing tolerance of mutant plants during cold acclimation. Wild type ‘5899-16’ plants acclimated at 5 ºC for four weeks can survive at lower temperature then mutant M631 plants. Ss1gene of wild type wheat plant is fully functional and its expression increases during cold acclimation (Fig. 4).While Ss1 gene is suppressed due to the premature stop codon mutation in M631 plants and has significantly lower expression pattern during cold acclimation. It can be concluded that decreased Ss1 gene expression leads to reduced freezing tolerance of mutants.
The small sample size was used in this experiment, due to the fact that a small amount of M4 mutant seeds of M631 was obtained. A total number of 168 plants of each genotype were assessed. The sample size is too small to make any accurate conclusions, but results shows tendency, that the premature stop codon mutation in the Ss1 gene affects freezing tolerance of mutants. The freezing test with larger sample size of plants should be carried out, in order to definitively confirm the importance of the SS1 enzyme for wheat cold acclimation process.

21
CONCLUSIONS 1. Low negative, but significant relationship (r = −0.41, p < 0.05) between
2010–2011 winter hardiness in the field and freezing tolerance test survival of non-acclimated plants supports hypothesis of two separate genetic systems controlling freezing tolerance in winter wheat which determine the duration and the rate of cold acclimation. Wheat genotypes with faster de-acclimation (limited duration tolerance) in the field show higher freezing tolerance without acclimation (high rate tolerance) in the freezing test.
2. Twenty TDFs (3.8%) were identified as being differentially expressed between cold acclimated and non-acclimated wheat. Thirteen TDFs (65%) were up-regulated and 7 TDFs (35%) were down-regulated within 2 weeks after the start of cold acclimation.
3. A significant negative relationship (r = −0.7, p < 0.01) was observed between various EMS concentrations and survival rate of winter wheat. Optimal EMS concentrations of 0.7–1.0% and 0.6–0.7% were identified for genotypes '5899-16' and '5450-1', respectively. TILLING population of two winter wheat lines was created, which consists of 2147 M2 and 756 M3 lines.
4. Two novel alleles of Ss1 gene were identified by HRM analysis. Mutant M692 was found to have a silent mutation (G1119A, K373K) and mutant M631 cointained a nonsense (premature stop codon) mutation (G1122A, W374*).
5. qRT-PCR analysis showed that the premature stop codon mutation affects the Ss1 gene expression during cold acclimation. Putative knock-out mutant M631 had significantly lower relative expression of Ss1 gene in non-acclimated leaves as well as in crowns and leaves collected at 2, 4 and 6 weeks of cold acclimation compared to the wild type winter wheat line ‘5899-16’ and mutant M692.
6. Wild type ‘5899-16’ plants acclimated at 4 ºC for four weeks have higher freezing tolerance compared with the M631 plants (LT50 difference of –3.2 °C). This suggests that the Ss1 gene is involved in the process of cold acclimation of winter wheat.
LIST OF PUBLICATIONS
Articles in journals indexed in ISI WOS database: 1. Armonienė R., Liatukas Ž., Brazauskas G. 2013. Evaluation of freezing
tolerance of winter wheat under controlled conditions and it’s correlation with winter hardiness in the field. Žemdirbyste=Agriculture, 100 (4): 417‒424.
2. Armonienė R., Brazauskas G. Non-sense mediated decay of Sucrose synthase 1 mRNA with induced pre-mature chain termination codon during cold acclimation in winter wheat. Turkish Journal of Botany. In press (accepted for publication on the 27nd of August 2014).

22
Abstracts in conference proceedings: 1. Armonienė R., Liatukas Ž., Brazauskas G. Identification and analysis of
winter wheat genes induced at low temperatures. Abstarcts of International Workshop-Seminar: „Plant Biotechnology Advances in Agriculture“, Kaunas distr., Lithuania, 27-28th October 2011, p. 10.
2. Armonienė R., Brazauskas G. The creation of TILLING population in winter wheat and screening for mutations by High Resolution Melting analysis. Abstract book: „5th Baltic congress of genetics”, Kaunas, Lithuania, 19-22th October 2012, p. 22‒23.
3. Armonienė R., Brazauskas G. Žieminių kviečių mutagenezė ir mutacijų aptikimas aukštos skiriamosios gebos DNR lydymosi temperatūros nustatymo metodu. Jaunųjų mokslininkų konferencija: „Jaunieji mokslininkai ‒ žemės ūkio pažangai“, Vilnius, 2012 m. lapkričio 21 d., p. 9‒10.
4. Armonienė R., Brazauskas G. Identification of differentially expressed genes under low temperature in winter wheat. Abstract book: „International Plant Breeding Congress“, Antalya, Turkey, 10-14th November 2013, p. 168.
5. Armonienė R., Brazauskas G. Žemoje teigiamoje temperatūroje diferenciškai ekspresuojamų genų paieška žieminiuose kviečiuose. Antroji jaunųjų mokslininkų konferencija: „Jaunieji mokslininkai ‒ žemės ūkio pažangai. Vilnius, 2013 m. lapkričio 21 d., p. 86
6. Armonienė R., Brazauskas G. Mutation inductionin Sucrose synthase 1 to study cold acclimation in winter wheat. Abstract book: „EUCARPIA Cereals Section - I T M I Joint Conference“.Wernigerode, Germany, June 29 ‒July 4, 2014, p. 221.
ACKNOWELADGEMENTS I would like to express my gratitude to my scientific supervisor dr. Gintaras
Brazauskas for the suggesting the subject of dissertation and valuable advices and guidance during the years of research. Sincere thanks go to dr. Žilvinas Liatukas for wheat varieties and lines provided, and the help to conduct field experiments. Also my sincere thanks go to all the staff of Cereal Breeding Department for help with installation and maintenance of field experiments. I also highly appreciate the help and support of my colleagues and laboratory assistants from the Laboratory of Genetics and Physiology. Most of all I would like to thank my husband Domas for understanding, support and for help of taking care of our children Ūla and Anupras. Thank you everyone for making this dream come true!

23
REZIUMĖ Žieminių kviečių tolerantiškumo žemoms temperatūroms tyrimai buvo
atlikti Lietuvos agrarinių ir miškų mokslų centro filiale Žemdirbystės institute 2010-2014 metais.
Darbo hipotezė. Žieminių kviečių genai, kurie grūdinimosi metu yra ekspresuojami skirtingu intensyvumu, lemia skirtingą kviečių tolerantiškumą žemoms neigiamoms temperatūroms.
Tyrimų tikslas. Identifikuoti naujus žieminių kviečių genus, aktyvuojamus žemose temperatūrose bei atlikti šių genų sekų ir genetinės raiškos analizę.
Tyrimų uždaviniai: 1. Ištirti žieminių kviečių genotipų tolerantiškumą žemoms neigiamoms
temperatūroms natūraliomis ir dirbtinėmis sąlygomis. 2. Nustatyti skirtingose grūdinimosi stadijose diferenciškai ekspresuojamus
genus (kDNR-AFLP analizė) ir atlikti DNR sekų analizę. 3. Optimizuoti mutagenezės metodą ir sukurti dviejų perspektyvių žieminių
kviečių linijų mutagenines populiacijas. 4. Nustatyti skirtingu intensyvumu ekspresuojamų genų mutacijas HRM
metodu, įvertinti nustatytų mutacijų įtaką genų ekspresijai grūdinimosi metu ir žemų neigiamų temperatūrų tolerancijai.
Disertacijos ginamieji teiginiai: 1. Egzistuoja dvi atskiros žieminių kviečių atsparumo šalčiui genetinės
sistemos, kurias lemia užsigrūdinimo trukmės ir užsigrūdinimo spartos genų raiška.
2. kDNR-AFLP metodas tinka diferenciškai ekspresuotų fragmentų (DEF) grūdintuose ir negrūdintuose kviečiuose paieškoms, o šių fragmentų nustatytos sekos gali būti pritaikytos HRM ir TL-PGR analizėms.
3. Kuriant mutageninę kviečių populiaciją, svarbu parinkti tinkamiausias mutageno koncentracijas, kad būtų pasiektas pakankamas mutacijų dažnis ir išvengiama stipraus neigiamo poveikio augalų daigumui ir tolimesniam vystymuisi.
4. Identifikuota priešlaikinio stop kodono mutacija Sacharozės sintazės 1 gene nulemia mutantų M631 nuslopusią genų ekspresiją grūdinimosi metu ir sumažėjusį tolerantiškumą žemoms neigiamoms temperatūroms.
Mokslinio darbo naujumas. Gauti rezultatai palaiko Fowler ir Limin
(2007) hipotezę, kad egzistuoja dvi atskiros žieminio kviečio atsparumo šalčiui genetinės sistemos, kurias lemia užsigrūdinimo trukmės ir užsigrūdinimo spartos genų raiška.
Nustatyta, dvidešimt tarp grūdintų ir negrūdintų žieminių kviečių skirtingu intensyvumu ekspresuojamų sekų. Identifikuota 14 genų homologų, iš kurių 13 buvo pirmą kartą nustatyti kaip genai kandidatai dalyvaujantis grūdinimosi procese. Dvylika iš jų pirmą kartą identifikuoti kviečiuose.

24
Sukurta žieminių kviečių linijų ‘5899-16’ ir ‘5450-1’ mutageninė populiacija. Identifikuoti du nauji Ss1 geno aleliai. Nustatyta, kad priešlaikinio stop kodono mutacija stipriai įtakoja Ss1 geno ekspresiją grūdinimosi metu. Taip pat nustatyta tendencija, kad ši mutacija Ss1 gene, turi įtakos žieminių kviečių atsparumui žemoms neigiamoms temperatūroms.
Praktinis pritaikymas. Rasti genai kandidatai, rodantys skirtingą ekspresijos intensyvumą negrūdintuose ir grūdintuose kviečių lapuose bei krūmijimosi mazguose, yra vertinga pradinė medžiaga tolimesniems tyrimams, siekiant išsiaiškinti šių genų įtaką žieminių kviečių grūdinimosi procese. Sukurtą mutageninę žieminių kviečių populiaciją bus galima naudoti dominančių genų naujų alelių paieškai HRM metodu.
Tyrimų rezultatų aprobavimas. Darbo tema paskelbtos 2 publikacijos žurnaluose, įrašytuose į Mokslinės informacijos instituto (ISI) WOS sąrašą, taip pat 6 santraukos mokslinių konferencijų leidiniuose. Pristatyti 3 stendiniai ir 3 žodiniai pranešimai konferencijose užsienyje ir Lietuvoje.
Disertacijos turinys ir apimtis. Darbas atliktas pagal doktorantūros komisijos patvirtintą metodiką ir LAMMC Žemdirbystės instituto 2011–2016 m. mokslinės veiklos planą. Disertaciją sudaro įvadas, literatūros apžvalga, tyrimo metodų pristatymas, tyrimų rezultatai ir jų aptarimas, išvados, darbe naudotos literatūros sąrašas bei disertantės su bendraautoriais paskelbtų publikacijų sąrašas. Darbas parašytas lietuvių kalba, jo apimtis 86 puslapiai. Darbo rezultatai pateikti 6 lentelėse bei 17 paveikslų. Panaudoti 214 literatūros šaltinių.
IŠVADOS
1. Nustatytas esminis (r = −0,41, p < 0,05) ryšys tarp 2010–2011 m. kviečių žiemkentiškumo lauko eksperimente ir negrūdintų kviečių išgyvenimo po šaldymo testo. Šie rezultatai palaiko hipotezę, kad egzistuoja dvi atskiros žieminių kviečių atsparumo šalčiui genetinės sistemos, kurias lemia užsigrūdinimo trukmės ir užsigrūdinimo spartos genų raiška. Žieminių kviečių genotipai, kurie greičiau praranda žiemkentiškumą (turi ribotą užsigrūdinimo trukmę), yra atsparesni žemoms neigiamoms temperatūroms be grūdinimosi stadijos (turi greitą užsigrūdinimo spartą).
2. kDNR-AFLP analizės metodu buvo identifikuota 20 fragmentų (3,8 %), pasižyminčių skirtinga raiška tarp grūdintų ir negrūdintų kviečių. Nustatyta 13 fragmentų (65 %), kurių ekspresija praėjus 2 savaitėms nuo grūdinimosi pradžios aktyvavosi bei 7 fragmentai (35 %), kurių ekspresija praėjus 2 savaitėms nuo grūdinimosi pradžios nuslopo.
3. Nustatytas patikimas neigiamas ryšys (r = −0,7, p < 0,01) tarp panaudotos EMS koncentracijos ir kviečių procentinio išgyvenamumo. Nustatytos ‘5899-16’ ir ‘5450-1’ genotipams tinkamiausios EMS koncentracijos, atitinkamai 0,7 %–1,0 % ir 0,6 %–0,7 %. Sukurta šių dviejų žieminių

25
kviečių genotipų mutageninė populiacija, kurią sudaro M2 kartos 2147 linijų sėklos ir M3 kartos 756 linijų sėklos.
4. Atlikus Ss1 geno VIII egzono HRM analizę rasti du ‘5899-16’ genotipo mutantai M631 ir M692. Nustatyta, kad mutantas M692 turi tyliąją mutaciją, (G1119A, K373K), tuo tarpu mutantas M631 turi priešlaikinio stop kodono mutaciją (G1122A, W374*).
5. TL-PGR analizė parodė, kad priešlaikinė stop kodono mutacija įtakoja Ss1 geno ekspresiją grūdinimosi metu. Mutantas M631 pasižymėjo patikimai žemesne Ss1 geno santykine ekspresija tiek negrūdintuose lapuose, tiek lapuose ir krūmijimosi mazguose surinktuose po grūdinimo praėjus 2, 4 ir 6 savaitėms, lyginant su laukiniu ‘5899-16’ genotipu ir mutantu M692.
6. Šaldymo testo rezultatai parodė tendenciją, kad grūdinti keturias savaites laukinio tipo kviečiai yra atsparesni žemesnei neigiamai temperatūrai lyginant su M631 kviečiais, turinčiais priešlaikinę stop kodono mutaciją Ss1 gene (LT50 skirtumas –3,2 °C). Tai leidžia daryti prielaidą, kad Ss1 genas dalyvauja žieminių kviečių grūdinimosi procese.
ABOUT THE AUTHOR Rita Armonienė was born in Kaunas, on the 18 th of April, 1985. Graduated
the ‘Varpas’ gymnasium in 2004. In 2008, R. Armonienė graduated Vytautas Magnus University and received a bachelor degree in Biology and in 2010 – master’s degree in Molecular biology and biotechnology. During 2010–2014, was the PhD student at the Lithuanian Research Centre for Agriculture and Forestry. R. Armonienė has been working as junior researcher at the Laboratory of Genetics and Physiology, Institute of Agriculture, LRCAF since 2010.
TRUMPOS ŽINIOS APIE DISERTANTĘ Rita Armonienė gimė 1985 balandžio 18 d. Kaune. 2004m. baigė „Varpo“
gimnaziją. 2004–2010 m. studijavo Vytauto Didžiojo universitete, Gamtos mokslų fakultete. 2008 m. suteiktas biologijos mokslų bakalauro laipsnis, 2010 m. – molekulinės biologijos ir biotechnologijos magistro kvalifikacinis laipsnis. 2010–2014 m. studijavo Lietuvos agrarinių ir miškų mokslų centro doktorantūroje. Nuo 2010 m. dirba LAMMC filiale Žemdirbystės institute, Genetikos ir fiziologijos laboratorijoje.








![iGalerie.cz: [Literatura] Psychologie (Rita L. Atkinson a kol](https://static.fdokumen.com/doc/165x107/63367c2be8daaa60da0fec2d/igaleriecz-literatura-psychologie-rita-l-atkinson-a-kol.jpg)











