







Bahasa
Halaman
Hukum


www.bba-direct.com
Biochimica et Biophysica Acta 1693 (2004) 73–80
Pro-apoptotic signaling pathway activated by echistatin in GD25 cells
Elena Alimentia, Simona Tafurib, Antonio Scibellia, Danila d’Angeloa, Laura Mannac,Luigi Michele Pavonec, M. Antonietta Belisariob, Norma Staianoa,*
aDipartimento di Strutture, Funzioni e Tecnologie Biologiche, Universita di Napoli Federico II, Naples, ItalybDipartimento di Biochimica e Biotecnologie Mediche, Universita di Napoli Federico II, Naples, Italy
cDipartimento di Scienze Cliniche Veterinarie, Universita di Napoli Federico II, Naples, Italy
Received 30 January 2004; received in revised form 18 May 2004; accepted 19 May 2004
Available online 15 June 2004
Abstract
Disintegrins, low molecular weight RGD-containing polypeptides isolated from snake venoms, have seen use as integrin antagonists in
the field of tumor biology and angiogenesis. In this study, we investigated the molecular mechanism by which the disintegrin echistatin
affects cell adhesion and signaling resulting in an apoptotic response in the GD25 cell system. Wild-type GD25 cells, which lack expression
of the h1 family of integrin, and stable transfectants expressing the A isoform of h1 integrin subunit were used.
Nanomolar concentrations of echistatin detached fibronectin- and vitronectin-adherent GD25 cells from immobilized substratum.
However, prior to inducing detachment of adherent cells, echistatin caused apoptosis as measured by caspase-3 activation. Either cell
detachment or apoptotic response induced by echistatin were more pronounced on fibronectin-adherent GD25 cells than on vitronectin-
adherent ones.
GD25 cell exposure to echistatin caused a reduction of tyrosine phosphorylation levels of pp125FAK, whereas it didn’t affect either Shc
tyrosine phosphorylation levels or Shc–Grb2 functional association. The down-regulation of pp125FAK preceded apoptosis and cell
detachment induced by echistatin. Our results indicate that pp125FAK and not Shc pathway is involved in echistatin-induced apoptotic
response in the GD25 cell system.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Echistatin; GD25 cell; Apoptosis; FAK; Shc
1. Introduction
Disintegrins are a family of low molecular weight
proteins isolated from Crotalidae and Vipiridae snake
venoms [1]. They contain an Arg–Gly–Asp (RGD) motif
which represents a common adhesion recognition site [2].
However, disintegrins with other recognition motifs such as
KGD, VGD or MLD have been identified [3–5]. Disinte-
grins bind with high affinity integrin receptors on platelet
and cell surface, thus inhibiting both platelet aggregation
and cell adhesion [6]. Additional structural features other
than the RGD motif, including the amino acids flanking the
RGD sequence, the pattern of intramolecular S–S bridges,
and the presence in their molecule of other recognition sites
affect disintegrin activity [7]. Thus, structurally distinct
0167-4889/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.bbamcr.2004.05.007
* Corresponding author. Tel.: +39-81-2536108; fax: +39-81-2536097.
E-mail address: [email protected] (N. Staiano).
disintegrins may differently act on cell adhesion and sig-
naling [8,9].
Disintegrins have seen use as integrin antagonists in the
fields of tumor biology and angiogenesis [10,11]. Various
disintegrins have been shown to inhibit angiogenesis either
in vitro and in vivo by inducing apoptosis [12]. Echistatin
was shown to induce an apoptotic response occurring prior
to cell detachment in avh3-transfected human embryonic
kidney-293 epithelial cells [13]. However, the molecular
mechanism of action of disintegrins in causing apoptosis has
not been fully clarified yet.
We previously showed that echistatin inhibits experi-
mental Lewis lung carcinoma-induced metastasis in vivo
[14] and we demonstrated that this disintegrin detaches
fibronectin-adherent melanoma cells by down-regulating
pp125FAK phosphorylation [15]. The echistatin-induced
dephosphorylation of pp125FAK Tyr 118 residue resulted
in a decrease of paxillin tyrosine phosphorylation and in the
disruption of pp125FAK-Src and pp125FAK-paxillin func-

E. Alimenti et al. / Biochimica et Biophysica Acta 1693 (2004) 73–8074
tional complexes [16]. Herein, we also demonstrated that a
reduction of tyrosine phosphorylation of pp125FAK, paxillin
and Shc is involved in apoptosis of COS cells induced by
ochratoxin A, a mycotoxin with nephrotoxic, teratogenic
and carcinogenic activity which seems to be involved in the
pathogenesis of Balkan Endemic Nephropathy and urinary
tract tumors [17].
In this study, we evaluated the ability of echistatin to
affect cell adhesion and signaling in the mouse cell line
GD25. These cells, derived from the embryonic stem cell
clone G201, lack expression of the h1 family of integrin
heterodimers due to the disruption of the h1 subunit gene
[18]. By using wild-type GD25 cells and stable transfectants
of these cells expressing h1A isoform of h1 integrin subunit,
the molecular mechanism by which echistatin induces an
apoptotic response was investigated. In particular, the FAK
and Shc signaling pathways activated by the disintegrin
interaction with integrin receptor(s) were characterized.
2. Materials and methods
2.1. Antibodies
Horseradish peroxidase conjugated goat anti-(mouse
IgG) Ig and horseradish peroxidase conjugated goat anti-
(rabbit IgG) Ig were purchased from Sigma Chemical Co.
(St. Louis, MO, USA); rabbit polyclonal anti-human FAK
IgG from Upstate Biotechnology (Lake Placid, NY, USA);
monoclonal anti-phosphotyrosine (PY20) IgG, monoclonal
mouse anti-Grb2 IgG and rabbit polyclonal anti-Shc Ig from
Transduction Laboratories (Lexington, KY, USA).
2.2. Chemicals
Aprotinin, bovine serum albumin (BSA), Dulbecco
minimum Eagle’s medium (DMEM), human plasma
fibronectin, leupeptin, orthovanadate, pepstatin, N-acetyl-
Asp-Glu-Val-Asp-7-amino-4-methylcoumarin (Ac-DEVD-
AMC), 1,4-piperazinediethane sulfonic acid (PIPES),
ethylenediaminetetraacetic acid (EDTA), 3-[(3-cholamido-
propyl)dimethylammonio]-1-propanesulfonate (CHAPS),
1,1-dichloro-2,2-bis(4-chlorophenyl)ethane (DDT) were
purchased from Sigma Chemical Co.; L-glutamine and tryp-
sin from ICN Biomedicals, Inc. (Aurora, OH, USA); fetal
bovine serum (FBS) fromHyclone Laboratories, Inc. (Logan,
UT, USA); protein A-agarose and protein G-agarose from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
2.3. Cell culture
GD25 cells were kindly provided by Prof. Francesco
Retta (Dipartimento di Genetica, Biologia e Biochimica,
Universita di Torino, Torino, Italy). GD25 cells were
grown on plastic Petri dishes in DMEM supplemented
with 10% FBS and 1% L-glutamine at pH 7.4, and cultured
under standard cell culture conditions (37 jC, 5% CO2). In
the culture medium of GD25 h1A cells hygromycin B was
also added. Cells were harvested for propagation or cell
attachment studies by treatment with 0.25% trypsin/0.02%
EDTA in phosphate-buffered saline (PBS), pH 7.2, and
with 5 mM EDTA in PBS, respectively. Cells were then
washed with DMEM and resuspended in complete DMEM
for propagation or in DMEM with 2% FBS for adhesion
experiments.
2.4. Substrate coating
Tissue culture plastic dishes (35� 10 mm) or 96-multi-
well plastic dishes (Costar, Cambridge, MA, USA) were
coated overnight at 4 jC by incubation with human plasma
fibronectin (10 Ag/ml) diluted in PBS with 1 mM CaCl2 and
1 mM MgCl2, or with vitronectin (5 Ag/ml) diluted in PBS.
After coating, dishes were treated with 1% BSA in PBS for
30 min at 37 jC to block free binding sites on the plastic.
Before plating cells, dishes were rinsed twice with PBS.
2.5. Cell adhesion assay
Freshly suspended cells (100 Al, 75� 104 cells/ml) were
plated onto 96-multiwell plates previously coated with
fibronectin or vitronectin and allowed to adhere for 3 h at
37 jC. Non-adherent cells were removed by gentle washing
with PBS. Adherent cells were exposed to 5 Ag/ml of
echistatin for increasing time intervals (0–6 h) or to
different concentrations of echistatin (0.1–20 Ag/ml) for
3 h and incubated at 37 jC.At the indicated time, detached cells were gently re-
moved by washing with PBS, adherent cells were fixed, and
stained by crystal violet. The colour yields were measured
by a Bio-Rad ELISA reader equipped with a 570 nm filter.
The percentage of cell detachment induced by echistatin
was evaluated as previously described [15].
2.6. Apoptosis assay
Freshly suspended cells (1�106 cells) were plated on
substratum-coated dishes, allowed to adhere at 37 jC for 3 h.
Non-adherent cells were removed by gentle washing with
serum-free medium. Adherent cells were exposed to in-
creasing amounts of echistatin (0.1–10 Ag/ml) for 3 h or to
5 Ag/ml of echistatin for increasing time intervals (0–180
min). At the end of the incubation time, cells were lysed by
ice-cold lysis buffer containing 10 mM Tris–HCl, pH 7.5,
125 mM NaCl, 1% Triton, 10 mM Na4P2O7, 10 mM
Na2HPO4, 22.7 Ag/ml aprotinin (0.1 trypsin inhibitor units/
ml), 10 Ag/ml leupeptin, 0.4 mM Na3VO4. The lysates were
clarified by centrifugation at 12,000� g for 10 min at 4 jC.The amount of proteins in the samples was determined by the
Bio-Rad DC protein assay (Bio-Rad Laboratories). Lysates
containing 150 Ag of proteins were incubated with 50 AM of
Ac-DEVD-AMC in 1 ml of reaction buffer (10 mM PIPES,

E. Alimenti et al. / Biochimica et Biophysica Acta 1693 (2004) 73–80 75
pH 7.4, 2 mM EDTA, 0.1% CHAPS, 5 mM DTT) for 5 h at
37 jC. Fluorescence derived from the release of the 7-amino-
4-methylcoumarin moiety was followed using a Perkin-
Elmer with E excitation at 390 nm and E emission at 460
nm. The data obtained are reported as the variation (D) of the
fluorescence of the treated sample lysate minus the fluores-
cence of the appropriate control sample lysate.
2.7. Protein immunoprecipitation and Western blotting
Freshly suspended cells (1�106 cells) were plated on
substrate-coated dishes and allowed to adhere at 37 jC for
3 h. Non-adherent cells were removed by two gentle wash-
ings with serum-free medium, adherent cells were exposed to
20 Ag/ml of echistatin for 1 or 3 h. At the end of the
incubation time, the proteins were detergent-extracted by
ice-cold lysis buffer, as described in Apoptosis assay. The
lysates were clarified by centrifugation at 12,000� g for 10
min at 4 jC and the amount of proteins in the samples was
determined by the Bio-Rad DC protein assay.
Lysates containing equal amount of proteins were incu-
bated with anti-pp125FAK (1 Ag 100 Ag proteins) or anti-Shc
Ig (0.25 Ag 100 Ag proteins) for 6 h at 4jC. Protein A- or
protein G-agarose was then added to the samples and
incubated overnight at 4jC. Beads were sedimented by
brief centrifugation and immunoprecipitates were washed
extensively with cold lysis buffer. Proteins were resus-
pended in SDS sample buffer, boiled for 5 min in Laemmli
buffer and run on 10% SDS/polyacrilamide gel.
After electrophoresis, the proteins were transferred to
nitrocellulose using a Mini trans-blot apparatus (Bio-Rad
Laboratories) according to manufacturer’s instructions.
Membranes were blocked for 1 h at 42jC with blocking
buffer containing 5% BSA in Tris/NaCl/Pi and incubated
overnight with a 1:2000 dilution of antibody PY20 to probe
for phosphotyrosine-containing proteins. The blots were
washed three times with Tris/NaCl/Pi/Tween (150 mM
NaCl, 20 mM Tris–HCl, pH 7.4, 0.3% Tween 20) and
incubated for 1 h with peroxidase conjugated anti-(mouse
IgG) Ig diluted 1:3000 in Tris/NaCl/Pi, 1% BSA.
To evaluate the amount of Grb2 co-precipitating with
Shc, cellular extracts were immunoprecipitated with anti-
Shc Ig as previously described, and separated by SDS gel
electrophoresis. After electrophoresis, the proteins were
transferred to nitrocellulose using a Mini trans-blot appara-
tus (Bio-Rad Laboratories) according to manufacturer’s
instructions. The membranes were blocked for 1 h with
blocking buffer containing 5% milk in Tris/NaCl/Pi and
incubated overnight with a 1:500 dilution of antibody Grb2.
The blots were washed three times with Tris/NaCl/Pi/Tween
(150 mM NaCl, 20 mM Tris–HCl, pH 7.4, 0.3% Tween 20)
and incubated for 1 h with peroxidase conjugated anti-
(rabbit IgG) Ig diluted 1:3000 in Tris/NaCl/Pi, 2.5% milk.
The proteins were visualized by an ECL chemilumines-
cence kit (Amersham Corp., Little Chalfont, UK). The same
blots were stripped and reprobed using anti-pp125FAK or
anti-Shc Ig to confirm equal loading of pp125FAK or Shc in
the lysates.
3. Results
3.1. Echistatin causes detachment of GD25 cells from
immobilized fibronectin or vitronectin
The ability of echistatin to promote cell detachment from
immobilized substrates was evaluated. Echistatin induced
GD25 cell detachment from immobilized fibronectin, but
this effect was more pronounced on fibronectin-adherent
GD25 h1A cells than on wild-type GD25 cells. Cell
treatment with 20 Ag/ml of echistatin for 3 h induced 87%
of GD25 h1A cell detachment from immobilized fibronec-
tin, while the same dose caused 76% of wild-type GD25 cell
detachment from immobilized fibronectin (Fig. 1, panel A).
Echistatin promoted cell detachment from immobilized
vitronectin, but this effect was more pronounced on wild-
type GD25 cells than on GD25 h1A: 3 h of cell treatment
with 20 Ag/ml echistatin caused 45% and 54% of cell
detachment for GD25 h1A cells and wild-type GD25 cells,
respectively (Fig. 1, panel A).
The time-course of echistatin-induced cell detachment
confirmed the observation that the disintegrin is more active
to promote cell detachment from fibronectin than from
vitronectin and that wild-type cells are easier detached from
vitronectin than GD25 h1A cells, whereas GD25 h1A are
more easily detached from immobilized fibronectin than
wild-type cells (Fig. 1, panel B).
We had previously observed that the attachment to
immobilized fibronectin and vitronectin which bind a5h1
and avh3 integrin receptors on cell surface, respectively, is
efficient for both wild-type GD25 cells and the GD25 h1A
cells. Furthermore, h1A transfected GD25 cells showed a
higher percentage of adhesion to immobilized fibronectin
than wild-type GD25 cells, whereas the percentage of wild-
type GD25 cells adhering to immobilized vitronectin was
higher than that of transfected-ones [19]. This may be due to
the observed overexpression of the avh3 integrin receptor,
specific for vitronectin, on wild-type cells which lack the h1
integrin subunit [20]. Thus, the results obtained indicate that
echistatin is able to promote cell detachment by competing
with both immobilized fibronectin and vitronectin for their
respective integrin receptor. However, echistatin activity
was more pronounced on fibronectin-adherent GD25 cells
than on vitronectin-adherent GD25 cells, thus suggesting
that the disintegrin has higher affinity for fibronectin recep-
tor than for vitronectin receptor.
3.2. Echistatin promotes apoptosis of GD25 cells adhering
to immobilized fibronectin or vitronectin
The ability of echistatin to promote apoptosis in adherent
GD25 cells was evaluated by measuring the activation of
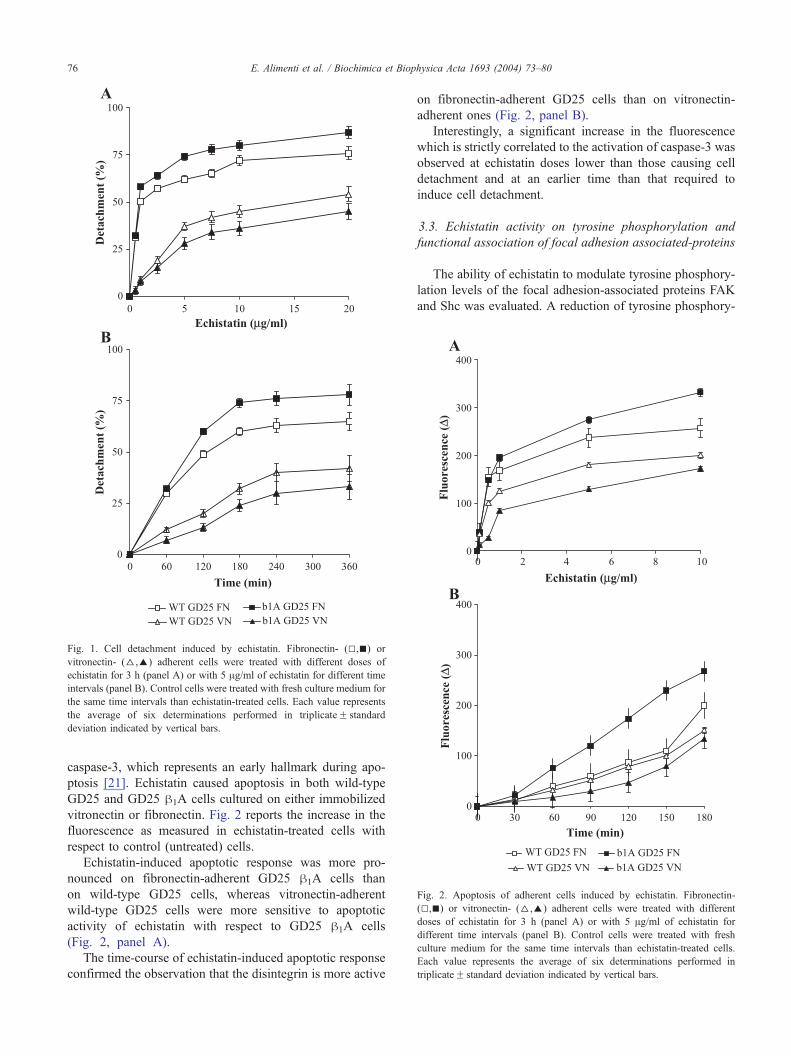
Fig. 2. Apoptosis of adherent cells induced by echistatin. Fibronectin-
(5,n) or vitronectin- (D,E) adherent cells were treated with different
doses of echistatin for 3 h (panel A) or with 5 Ag/ml of echistatin for
different time intervals (panel B). Control cells were treated with fresh
culture medium for the same time intervals than echistatin-treated cells.
Each value represents the average of six determinations performed in
triplicateF standard deviation indicated by vertical bars.
Fig. 1. Cell detachment induced by echistatin. Fibronectin- (5,n) or
vitronectin- (D,E) adherent cells were treated with different doses of
echistatin for 3 h (panel A) or with 5 Ag/ml of echistatin for different time
intervals (panel B). Control cells were treated with fresh culture medium for
the same time intervals than echistatin-treated cells. Each value represents
the average of six determinations performed in triplicateF standard
deviation indicated by vertical bars.
E. Alimenti et al. / Biochimica et Biophysica Acta 1693 (2004) 73–8076
caspase-3, which represents an early hallmark during apo-
ptosis [21]. Echistatin caused apoptosis in both wild-type
GD25 and GD25 h1A cells cultured on either immobilized
vitronectin or fibronectin. Fig. 2 reports the increase in the
fluorescence as measured in echistatin-treated cells with
respect to control (untreated) cells.
Echistatin-induced apoptotic response was more pro-
nounced on fibronectin-adherent GD25 h1A cells than
on wild-type GD25 cells, whereas vitronectin-adherent
wild-type GD25 cells were more sensitive to apoptotic
activity of echistatin with respect to GD25 h1A cells
(Fig. 2, panel A).
The time-course of echistatin-induced apoptotic response
confirmed the observation that the disintegrin is more active
on fibronectin-adherent GD25 cells than on vitronectin-
adherent ones (Fig. 2, panel B).
Interestingly, a significant increase in the fluorescence
which is strictly correlated to the activation of caspase-3 was
observed at echistatin doses lower than those causing cell
detachment and at an earlier time than that required to
induce cell detachment.
3.3. Echistatin activity on tyrosine phosphorylation and
functional association of focal adhesion associated-proteins
The ability of echistatin to modulate tyrosine phosphory-
lation levels of the focal adhesion-associated proteins FAK
and Shc was evaluated. A reduction of tyrosine phosphory-
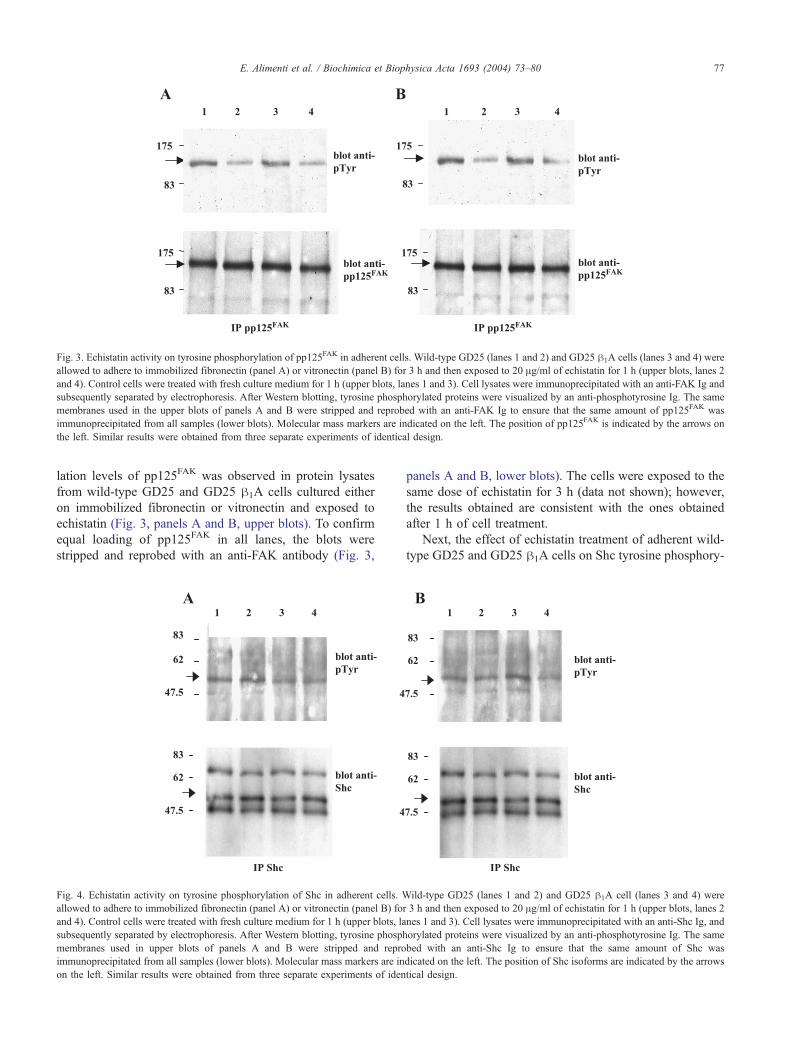
Fig. 3. Echistatin activity on tyrosine phosphorylation of pp125FAK in adherent cells. Wild-type GD25 (lanes 1 and 2) and GD25 h1A cells (lanes 3 and 4) were
allowed to adhere to immobilized fibronectin (panel A) or vitronectin (panel B) for 3 h and then exposed to 20 Ag/ml of echistatin for 1 h (upper blots, lanes 2
and 4). Control cells were treated with fresh culture medium for 1 h (upper blots, lanes 1 and 3). Cell lysates were immunoprecipitated with an anti-FAK Ig and
subsequently separated by electrophoresis. After Western blotting, tyrosine phosphorylated proteins were visualized by an anti-phosphotyrosine Ig. The same
membranes used in the upper blots of panels A and B were stripped and reprobed with an anti-FAK Ig to ensure that the same amount of pp125FAK was
immunoprecipitated from all samples (lower blots). Molecular mass markers are indicated on the left. The position of pp125FAK is indicated by the arrows on
the left. Similar results were obtained from three separate experiments of identical design.
E. Alimenti et al. / Biochimica et Biophysica Acta 1693 (2004) 73–80 77
lation levels of pp125FAK was observed in protein lysates
from wild-type GD25 and GD25 h1A cells cultured either
on immobilized fibronectin or vitronectin and exposed to
echistatin (Fig. 3, panels A and B, upper blots). To confirm
equal loading of pp125FAK in all lanes, the blots were
stripped and reprobed with an anti-FAK antibody (Fig. 3,
Fig. 4. Echistatin activity on tyrosine phosphorylation of Shc in adherent cells. W
allowed to adhere to immobilized fibronectin (panel A) or vitronectin (panel B) for
and 4). Control cells were treated with fresh culture medium for 1 h (upper blots, la
subsequently separated by electrophoresis. After Western blotting, tyrosine phosph
membranes used in upper blots of panels A and B were stripped and repro
immunoprecipitated from all samples (lower blots). Molecular mass markers are in
on the left. Similar results were obtained from three separate experiments of iden
panels A and B, lower blots). The cells were exposed to the
same dose of echistatin for 3 h (data not shown); however,
the results obtained are consistent with the ones obtained
after 1 h of cell treatment.
Next, the effect of echistatin treatment of adherent wild-
type GD25 and GD25 h1A cells on Shc tyrosine phosphory-
ild-type GD25 (lanes 1 and 2) and GD25 h1A cell (lanes 3 and 4) were
3 h and then exposed to 20 Ag/ml of echistatin for 1 h (upper blots, lanes 2
nes 1 and 3). Cell lysates were immunoprecipitated with an anti-Shc Ig, and
orylated proteins were visualized by an anti-phosphotyrosine Ig. The same
bed with an anti-Shc Ig to ensure that the same amount of Shc was
dicated on the left. The position of Shc isoforms are indicated by the arrows
tical design.
E. Alimenti et al. / Biochimica et Biophysica Acta 1693 (2004) 73–8078
lation levels was investigated. Only the p52 isoform of Shc
appears to be phosphorylated in wild-type GD25 and GD25
h1A cells adherent to immobilized substrata. A decrease of
tyrosine phosphorylation of p52 Shc isoform was only
observed in vitronectin-adherent GD25 h1A cells exposed
to echistatin (Fig. 4, panel B, upper blot), whereas the
disintegrin did not modify the tyrosine phosphorylation
levels of Shc protein of both fibronectin- and in vitronec-
tin-adherent wild-type GD25 and GD25 h1A cells (Fig. 4,
panels A and B, upper blots). In order to evaluate whether
the Shc loading was equal in all lanes, the blots were
stripped and reprobed with an anti-Shc antibody (Fig. 4,
panels A and B, lower blots).
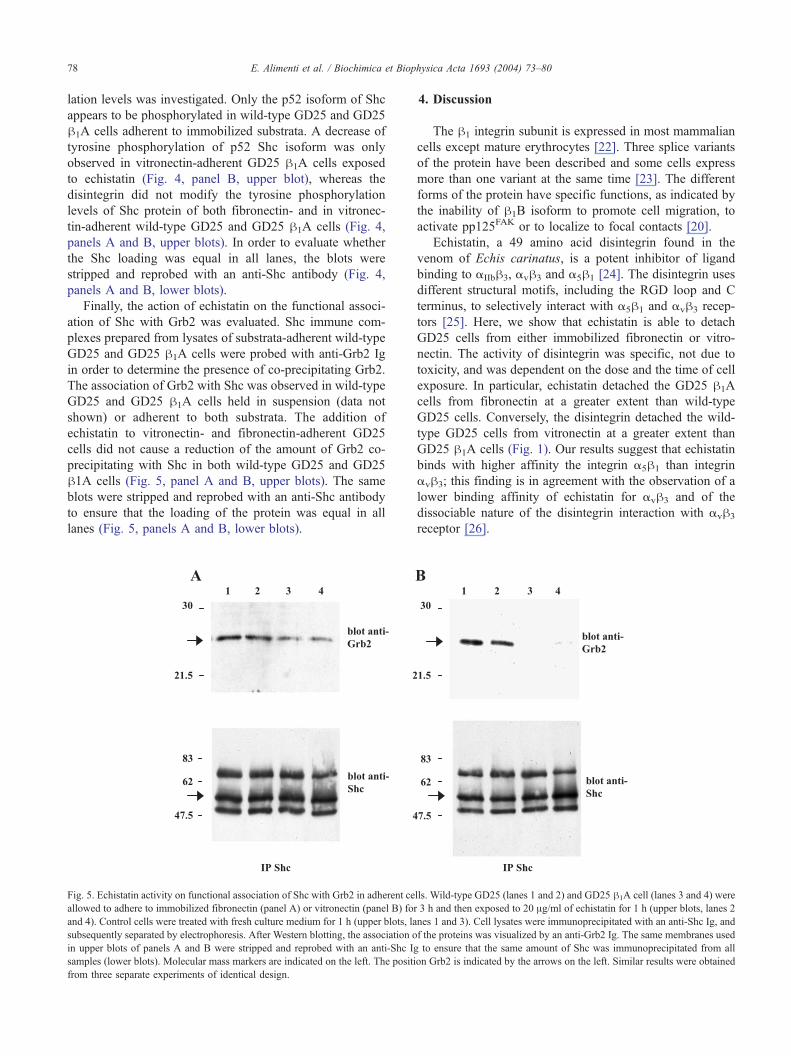
Finally, the action of echistatin on the functional associ-
ation of Shc with Grb2 was evaluated. Shc immune com-
plexes prepared from lysates of substrata-adherent wild-type
GD25 and GD25 h1A cells were probed with anti-Grb2 Ig
in order to determine the presence of co-precipitating Grb2.
The association of Grb2 with Shc was observed in wild-type
GD25 and GD25 h1A cells held in suspension (data not
shown) or adherent to both substrata. The addition of
echistatin to vitronectin- and fibronectin-adherent GD25
cells did not cause a reduction of the amount of Grb2 co-
precipitating with Shc in both wild-type GD25 and GD25
h1A cells (Fig. 5, panel A and B, upper blots). The same
blots were stripped and reprobed with an anti-Shc antibody
to ensure that the loading of the protein was equal in all
lanes (Fig. 5, panels A and B, lower blots).
Fig. 5. Echistatin activity on functional association of Shc with Grb2 in adherent ce
allowed to adhere to immobilized fibronectin (panel A) or vitronectin (panel B) for
and 4). Control cells were treated with fresh culture medium for 1 h (upper blots, la
subsequently separated by electrophoresis. After Western blotting, the association o
in upper blots of panels A and B were stripped and reprobed with an anti-Shc Ig
samples (lower blots). Molecular mass markers are indicated on the left. The positi
from three separate experiments of identical design.
4. Discussion
The h1 integrin subunit is expressed in most mammalian
cells except mature erythrocytes [22]. Three splice variants
of the protein have been described and some cells express
more than one variant at the same time [23]. The different
forms of the protein have specific functions, as indicated by
the inability of h1B isoform to promote cell migration, to
activate pp125FAK or to localize to focal contacts [20].
Echistatin, a 49 amino acid disintegrin found in the
venom of Echis carinatus, is a potent inhibitor of ligand
binding to aIIbh3, avh3 and a5h1 [24]. The disintegrin uses
different structural motifs, including the RGD loop and C
terminus, to selectively interact with a5h1 and avh3 recep-
tors [25]. Here, we show that echistatin is able to detach
GD25 cells from either immobilized fibronectin or vitro-
nectin. The activity of disintegrin was specific, not due to
toxicity, and was dependent on the dose and the time of cell
exposure. In particular, echistatin detached the GD25 h1A
cells from fibronectin at a greater extent than wild-type
GD25 cells. Conversely, the disintegrin detached the wild-
type GD25 cells from vitronectin at a greater extent than
GD25 h1A cells (Fig. 1). Our results suggest that echistatin
binds with higher affinity the integrin a5h1 than integrin
avh3; this finding is in agreement with the observation of a
lower binding affinity of echistatin for avh3 and of the
dissociable nature of the disintegrin interaction with avh3
receptor [26].
lls. Wild-type GD25 (lanes 1 and 2) and GD25 h1A cell (lanes 3 and 4) were
3 h and then exposed to 20 Ag/ml of echistatin for 1 h (upper blots, lanes 2
nes 1 and 3). Cell lysates were immunoprecipitated with an anti-Shc Ig, and
f the proteins was visualized by an anti-Grb2 Ig. The same membranes used
to ensure that the same amount of Shc was immunoprecipitated from all
on Grb2 is indicated by the arrows on the left. Similar results were obtained

E. Alimenti et al. / Biochimica et Biophysica Acta 1693 (2004) 73–80 79
Echistatin was able to promote apoptosis in both types of
GD25 cells cultured on immobilized substrata. The effect of
echistatin was more pronounced on GD25 h1A cells than on
wild-type GD25 cells adhering to fibronectin and more
pronounced on wild-type GD25 cells than on GD25 h1A
cells adhering to vitronectin (Fig. 2). Furthermore, the
apoptotic response induced by echistatin in the GD25 cells
occurred prior to cell detachment. Cell substratum detach-
ment usually induces an apoptotic response (anoikis); how-
ever, echistatin binding to avh3 integrin has been shown to
induce cell death resulting in an apoptotic signal with
different kinetics than the apoptotic signal induced by
matrix detachment [13]. This apoptotic inducing effect of
echistatin may be due to the ability of disintegrin to cause
integrin-mediated death (IMD) [23]. This form of apoptosis
is biologically and biochemically distinct from anoikis,
which is due to cell detachment from substratum. Herein,
IMD is defined as the apoptosis of adherent cells induced by
the recruitment of caspase-8 to the h subunit tails of
unligated integrins. This is a cellular process whereby
adherent cells that find themselves within an inappropriate
extracellular matrix (into the tissues), or those exposed to
integrin antagonists would be actively cued to undergo
apoptosis. Interestingly, while h1 and h3 integrins can
promote this process, h5 does not [27]. Echistatin may
cause IMD on GD25 cells adhering to vitronectin and
fibronectin by binding avh3 and a5h1 integrins.
Because of the complexity of signaling pathways in-
volved in apoptotic response by chemicals and because of
the cell-type specificity of signaling activation that leads to
apoptosis, whether FAK and Shc pathways are involved in
the apoptotic signals induced by echistatin was investigated.
Thus, the ability of echistatin to modulate tyrosine phos-
phorylation levels of the two proteins was evaluated.
Echistatin treatment caused a reduction of tyrosine phos-
phorylation of pp125FAK in protein lysates from GD25 cells
immobilized either on fibronectin or vitronectin (Fig. 3). No
differences were observed in the signaling pathway activat-
ed by echistatin treatment of either wild-type GD25 cells or
transfected-ones. However, integrin signaling independent
of ligand recognition has been described [27].
FAK is a crucial molecule in integrating signals from
integrins and receptor tyrosine kinases in processes such as
cell survival, proliferation and motility [28]. Herein, FAK
phosphorylation levels are closely associated with the sta-
bility of focal adhesions and cell–matrix interactions. Dur-
ing cell adhesion process, turnover of focal adhesions
requires activation of FAK and its functional interactions
with several proteins involved in integrin signaling [29]. For
example, FAK-deficient cells spread more slowly on extra-
cellular matrix proteins exhibiting an increased number of
prominent focal adhesions; these cells migrate poorly in
response to chemotactic signals [30]. Furthermore, cells
lacking paxillin expression, Cas expression or Src family
kinase expression all exhibit defects in focal adhesion
turnover [31]. The disassembly of focal adhesions and the
subsequent GD25 cell detachment from the substratum
induced by echistatin seem to be later events than focal
adhesion protein dephosphorylation. Although disruption of
focal adhesion signaling through FAK clearly precedes cell
detachment after echistatin induced apoptosis, our data also
suggest that caspase-3 activation is not responsible for the
former. Our results suggest that the dephosphorylation of
FAK may contribute to the loss of focal adhesions and cell
detachment.
Using human breast cancer cell lines, Xu et al. [32]
showed that FAK signaling pathway is involved in inhibit-
ing death receptor or death receptor-related apoptosis. The
overexpression in the cells of the carboxyl-terminal domain
of FAK, that is without kinase activity, induced apoptosis
independently of cell adhesion, but required Fas-associating
protein with death domain (FADD), caspase-8 and caspase-
3 [32].
The adapter protein Shc has also been linked to cyto-
skeletal organization of cells under different contexts [33].
Some reports suggest distinct roles for Shc and FAK in focal
adhesions and cell motility [34,35]. However, how Shc
plays a role in the cytoskeletal reorganization in different
cell types is not well understood [36]. GD25 cell exposure
to echistatin does not appear to affect either tyrosine
phosphorylation levels of Shc isoforms (Fig. 4) or the
functional association Shc–Grb2 (Fig. 5).
Our results indicate that FAK pathway, but not Shc
pathway is involved in the pro-apoptotic signals induced
by echistatin in GD25 cell system, during integrin-mediated
death process. However, further studies are needed in order
to identify FAK downstream molecules involved in the
echistatin-induced apoptotic response. These studies aimed
at a better understanding of the molecular mechanism of
action of disintegrins may open the way to therapeutic
targets for human diseases which require a modulation of
integrin function.
Acknowledgements
This work was supported by a grant from Polo delle
Scienze e Tecnologie per la Vita, Universita degli Studi di
Napoli Federico II, Napoli, Italy.
References
[1] R.J. Gould, M.A. Polokoff, P.A. Friedma, T.F. Huang, J.C. Halt, J.J.
Cook, S. Niewiarowski, Disintegrins: a family of integrin inhibitory
proteins from viper venoms, Proc. Soc. Exp. Biol. Med. 195 (1990)
168–171.
[2] M.A. McLane, C. MarcinKiewicz, S. Vijay-Kumar, I. Wierzbieka-
Patynowsky, S. Niewiarowski, Viper venom disintegrins and related
molecules, Proc. Soc. Exp Biol. Med. 219 (1998) 109–119.
[3] C. Brando, C. Marcinkiewicz, B. Goldman, M.A. McLane, S.
Niewiarowski, EC3, a heterodimeric disintegrin from Echis carinatus,
inhibits human and murine alpha4 integrin and attenuates lymphocyte
infiltration of Langherhans islets in pancreas and salivary glands in

E. Alimenti et al. / Biochimica et Biophysica Acta 1693 (2004) 73–8080
nonobase diabetic mice, Biochem. Biophys. Res. Commun. 267
(2000) 413–417.
[4] J.J. Calvete, M. Jurgens, C. Marcinkiewicz, A. Romero, M. Schrader,
S. Niewiarowski, Disulphide-bond pattern and molecular modelling
of the dimeric disintegrin EMF-10, a potent and selective integrin
alpha5beta1 antagonist from Eristocophis macmahoni venom, Bio-
chem J. 345 (2000) 573–581.
[5] Y. Fujii, D. Okuda, Z. Fujimoto, T. Morita, H. Mizuno, Crystallization
and preliminary crystallographic studies of dimeric disintegrins from
the venom of two Agkistrodon snakes, Acta Crystallogr., D Biol.
Crystallogr. 58 (2002) 145–147.
[6] C.P. Blobel, J.M. White, Structure, function and evolutionary rela-
tionship of proteins containing a disintegrin domain, Curr. Opin. Cell
Biol. 4 (1992) 760–765.
[7] E.A. Clark, M. Trikha, F.S. Markland, J.S. Brugge, Structurally dis-
tinct disintegrins contortrostatin and multisquamatin differentially reg-
ulate platelet tyrosine phosphorylation, J. Biol. Chem. 269 (1994)
21940–21943.
[8] A. Scaloni, E. Di Martino, N. Miraglia, A. Pelagalli, R. Della Morte,
N. Staiano, P. Pucci, Amino acid sequence and molecular modelling
of glycoprotein IIb– IIIa and fibronectin receptor iso-antagonists from
Trimeresurus elegans venom, Biochem. J. 319 (1996) 775–782.
[9] T.F. Huang, What have snakes taught us about integrins? Cell. Mol.
Life Sci. 54 (1998) 527–540.
[10] F.S. Markland, Snake venoms and the hemostatic system, Toxicon 36
(1998) 1749–1800.
[11] T.F. Huang, C.H. Yeh, W.B. Wu, Viper venom components affecting
angiogenesis, Haemostasis 31 (2001) 192–206.
[12] C.H. Yeh, H.C. Peng, T.F. Huang, Accutin, a new disintegrin, inhibits
angiogenesis in vitro and in vivo by acting as integrin avh3 antagonist
and inducing apoptosis, Blood 92 (1998) 3268–3276.
[13] D.L. Brassard, E. Maxwell, M. Malkowski, T.L. Nagabhushan, C.C.
Kumar, L. Armstrong, Integrin avh3-mediated activation of apoptosis,
Exp. Cell Res. 251 (1999) 33–45.
[14] D. Spalletti-Cernia, C. Squillacioti, R. Della Morte, P. Laccetti, N.
Staiano, Echistatin inhibits Lewis lung carcinoma cell–matrix adhe-
sion in vitro and experimental metastasis in vivo, Int. J. Oncol. 11
(1997) 757–763.
[15] N. Staiano, C. Garbi, C. Squillacioti, S. Esposito, E. Di Martino, M.A.
Belisario, L. Nitsch, P. Di Natale, Echistatin induces decrease of
pp125FAK phosphorylation, disassembly of actin cytoskeleton and
focal adhesion, and detachment of fibronectin-adherent melanoma
cells, Eur. J. Cell Biol. 73 (1997) 298–305.
[16] R. Della Morte, C. Squillacioti, C. Garbi, P. Derkinderen, M.A.
Belisario, J. Girault, P. Di Natale, L. Nitsch, N. Staiano, Echistatin
inhibits pp125FAK autophosphorylation, paxillin phosphorylation and
pp125FAK-paxillin interaction in fibronectin-adherent melanoma
cells, Eur. J. Biochem. 267 (2000) 5047–5054.
[17] A. Scibelli, S. Tafuri, M.C. Ferrante, E. Alimenti, B. Naso, A.
Lucisano, N. Staiano, R. Della Morte, Ochratoxin A affects COS
cell adhesion and signaling, Toxicol. Appl. Pharmacol. 192 (2003)
222–230.
[18] K. Wennerberg, L. Lohikangas, D. Gullberg, M. Pfaff, S. Johansson,
R. Fassler, h1 Integrin-dependent and -independent polymerization of
fibronectin, J. Cell Biol. 132 (1996) 227–238.
[19] A. Scibelli, S. Tafuri, E. Alimenti, M.A. Belisario, R. Della Morte, N.
Staiano, Biochemical analysis of the molecular mechanism of action
of natural integrin inhibitors, Vet. Res. Commun. 27 (2003) 627–630.
[20] S.F. Retta, F. Balzac, P. Ferrarsi, A.M. Belkin, R. Fassler, M.J.
Humphries, G. De Leo, L. Silengo, G. Tarone, Beta1-integrin cyto-
plasmic subdomains involved in dominant negative function, Mol.
Biol. Cell 9 (1998) 715–731.
[21] T. Patel, G.J. Gores, S.H. Kaufmann, The role of proteases during
apoptosis, FASEB J. 10 (1996) 587–597.
[22] R.O. Hynes, Integrins: bidirectional, allosteric signaling machines,
Cell 110 (2002) 673–687.
[23] S.F. Retta, G. Cassara, M. D’Amato, R. Alessandro, M. Pellegrino, S.
Degani, G. De Leo, L. Silengo, G. Tarone, Cross talk between beta(1)
and alpha(v) integrins: beta(1) affects beta(3) mRNA stability, Mol.
Biol. Cell 12 (2001) 3126–3138.
[24] C. Marcinkiewicz, S. Vijay-Kumar, M.A. McLane, S. Niewiarowski,
Significance of RGD loop and C-terminal domain of echistatin for
recognition of aIIbh3 and avh3 integrins and expression of ligand-
induced binding site, Blood 90 (1997) 1565–1575.
[25] I. Wierzbicka-Patynowski, S. Niewiarowski, C. Marcinkiewicz, J.J.
Calvete, M.M. Marcinkiewicz, M.A. McLane, Structural require-
ments of echistatin for the recognition of avh3 and a5h11 integrins,
J. Biol. Chem. 274 (1999) 37809–37814.
[26] C.C. Kumar, H. Nie, L. Armstrong, R. Zhang, S. Vijay-Kumar, A.
Tsarbopoulos, Chloramine T-induced structural and biochemical
changes in echistatin, FEBS Lett. 429 (1998) 239–248.
[27] D.G. Stupack, X.S. Puente, S. Boutsaboualoy, C.M. Storgard, D.A.
Cheresh, Apoptosis of adherent cells by recruitment of caspase-8 to
unligated integrins, J. Cell Biol. 155 (2001) 459–470.
[28] M.D. Schaller, Biochemical signals and biological responses elicited by
the focal adhesion kinase, Biochim. Biophys. Acta 1540 (2001) 1–21.
[29] D.J. Webb, J.T. Parsons, A.F. Horwitz, Adhesion assembly, disassem-
bly and turnover in migrating cells—over and over and over again,
Nat. Cell Biol. 4 (2002) E97–E100.
[30] D.J. Sieg, C.R. Hauck, D. Ilic, C.K. Klingbeil, E. Schaefer, C.H.
Damsky, D.D. Schlaepfer, FAK integrates growth-factor and integrin
signals to promote cell migration, Nat. Cell Biol. 2 (2000) 249–256.
[31] R.A. Kinghoffer, C. Sachsenmaier, J.A. Cooper, P. Soriano, Src family
kinases are required for integrin but not PDGFR signal transduction,
EMBO J. 18 (1999) 2459–2471.
[32] L.H. Xu, X. Yang, C.A. Bradham, D.A. Brenner, A.S. Baldwin, R.J.
Craven, W.G. Cance, The focal adhesion kinase suppresses transfor-
mation-associated, anchorage-independent apoptosis in human breast
cancer cells. Involvement of death receptor-related signaling path-
ways, J. Biol. Chem. 275 (2000) 30597–30604.
[33] K.S. Ravichandran, Signaling via Shc family adapter proteins, Onco-
gene 20 (2001) 6322–6330.
[34] J. Gu, M. Tamura, R. Pankov, E. Danen, T. Takino, K. Matsumoto, K.
Yamada, Shc and FAK differentially regulate cell motility and direc-
tionality modulated by PTEN, J. Cell Biol. 146 (1999) 389–403.
[35] L. Barberis, K.K. Wary, G. Fiucci, F. Liu, E. Hirsch, M. Brancaccio,
F. Altruda, G. Tarone, F.G. Giancotti, Distinct roles of the adaptor
protein Shc and focal adhesion kinase in integrin signaling to ERK,
J. Biol. Chem. 275 (2000) 36532–36540.
[36] K.M. Lai, T. Pawson, The Shc A phosphotyrosine docking protein
sensitizes cardiovascular signaling in the mouse embryo, Genes Dev.
14 (2000) 1132–1145.
Top Related
Copyright © 2022 FDOKUMEN

