
Synthesis of nano‑fibers containing nano‑curcumin in zein ...

DOI 10.1105/tpc.010431; originally published online March 8, 2002;Plant Cell
LarkinsCheol Soo Kim, Young-min Woo, Amy M. Clore, Ronald J. Burnett, Newton P. Carneiro and Brian A.
Endoplasmic Reticulum Membranes, Influence Protein Body Formation in Maize EndospermZein Protein Interactions, Rather Than the Asymmetric Distribution of Zein mRNAs on
This information is current as of October 30, 2013
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists

This article is published in
The Plant Cell
Online,
The Plant Cell
Preview Section, which publishes manuscripts accepted for publicationafter they have been edited and the authors have corrected proofs, but before the final, complete issue is published online. Early posting
of articles reduces normal time to publication by several weeks.
The Plant Cell
Preview, www.aspb.org © 2001 American Society of Plant Biologists 1 of 18
Zein Protein Interactions, Rather Than the Asymmetric Distribution of Zein mRNAs on Endoplasmic Reticulum Membranes, Influence Protein Body Formation in Maize Endosperm
Cheol Soo Kim, Young-min Woo, Amy M. Clore, Ronald J. Burnett, Newton P. Carneiro, and Brian A. Larkins
1
Department of Plant Sciences, University of Arizona, Tucson, Arizona 85721
Prolamin-containing protein bodies in maize endosperm are composed of four different polypeptides, the
�
-,
�
-,
�
-, and
�
-zeins. The spatial organization of zeins within the protein body, as well as interactions between them, suggests that thelocalized synthesis of
�
-zeins could initiate and target protein body formation at specific regions of the rough endoplas-mic reticulum. To investigate this possibility, we analyzed the distribution of mRNAs encoding the 22-kD
�
-zein and the27-kD
�
-zein proteins on cisternal and protein body rough endoplasmic reticulum membranes. In situ hybridization re-vealed similar frequencies of the mRNAs in both regions of the endoplasmic reticulum, indicating that the transcripts aredistributed more or less randomly. This finding implies that zein protein interactions determine protein body assembly.To address this question, we expressed cDNAs encoding
�
-,
�
-,
�
-, and
�
-zeins in the yeast two-hybrid system. Wefound strong interactions among the 50-, 27-, and 16-kD
�
-zeins and the 15-kD
�
-zein, consistent with their colocaliza-tion in developing protein bodies. Interactions between the 19- and 22-kD
�
-zeins were relatively weak, although each ofthem interacted strongly with the 10-kD
�
-zein. Strong interactions were detected between the
�
- and
�
-zeins and the16-kD
�
-zein and the 15-kD
�
-zein; however, the 50- and 27-kD
�
-zeins did not interact with the
�
- and
�
-zein proteins.We identified domains within the 22-kD
�
-zein that bound preferentially the
�
- and
�
-zeins and the
�
- and
�
-zeins. Affin-ities between zeins generally were consistent with results from immunolocalization experiments, suggesting an impor-tant role for the 16-kD
�
-zein and the 15-kD
�
-zein in the binding and assembly of
�
-zeins within the protein body.
INTRODUCTION
The mechanisms by which cells organize proteins in their cy-toplasm and organelles have been studied actively in recentyears. It is known that unique peptides (signal/transit/target-ing) within proteins direct their movement either cotransla-tionally or post-translationally into the secretory system(endoplasmic reticulum, Golgi, vacuoles), double membraneorganelles (plastids, mitochondria), and the nucleus (Bar-Peled et al., 1996; Keegstra and Cline, 1999; Vitale andDenecke, 1999). Another emerging paradigm is the roleplayed by the cytoskeleton in mediating mRNA targeting tospecific regions of the cell, where translation brings aboutprotein localization. This is an efficient mechanism that allowsboth soluble and secreted proteins to be made proximal to
where they are needed, and it also provides a mechanism toseparate proteins that otherwise might interact inappropri-ately (Rings et al., 1994). Evidence of mRNA sorting was ob-tained first from studies of embryogenesis in
Xenopus
and
Drosophila
, in which the asymmetric distribution of mRNAs inthe oocyte and early embryo, respectively, was shown to beessential for the development of polarity (St. Johnston, 1995).
The mechanisms of mRNA sorting have not been investi-gated widely in plants, although Okita and co-workers de-scribed evidence for this phenomenon in developing riceendosperm (Li et al., 1993a). Rice seed contain two types ofstorage proteins: prolamins (oryzins), which form accretionsdirectly within the lumen of the rough endoplasmic reticulum(ER), and glutelins, which are synthesized on rough ERmembranes but are transported to the vacuole, where theyform protein bodies. Li et al. (1993a) reported that althoughboth mRNAs are found in membrane-bound polysomes, oryzinmRNAs are localized preferentially to the ER surroundingprolamin-containing protein bodies and glutelin mRNA is asso-ciated predominantly with polysomes on the cisternal ER. Themechanism(s) responsible for the asymmetric distribution of
1
To whom correspondence should be addressed. E-mail [email protected]; fax 520-621-3692.Article, publication date, and citation information can be found atwww.plantcell.org/cgi/doi/10.1105/tpc.010431.

2 of 18 The Plant Cell
these two types of mRNAs is unknown, but recent studiesindicate a role for the mRNA 3
�
noncoding sequences (Choiet al., 2000).
Studies of storage protein synthesis in maize endospermsuggest that the cytoskeleton also plays a role in targetingmaize prolamin (zein) mRNAs to the ER. Zein protein bodiesform in the lumen of the ER throughout these large (100 to200
�
m diameter), nearly isodiametric cells (Lopes and Lar-kins, 1993). Consequently, there must be mechanisms thattransport zein mRNAs from the nucleus to distant sites onthe ER, organize zeins within the protein body, and limit pro-tein body enlargement. Abe et al. (1991) and Stankovic et al.(1993) discovered associations between actin filaments,polysomes, and protein bodies after the homogenization ofdeveloping maize endosperm in a cytoskeleton-stabilizingbuffer. Other studies examining cytoskeleton organization indeveloping endosperm showed that actin occurs as fine fila-ments in the cortex and cytoplasm as well as in clusters be-tween starch grains (Clore et al., 1996). Not only does thedistribution of actin closely match the location of proteinbodies, but actin also surrounds protein bodies. Some mi-crotubules in these cells are found in a multidirectional arraybetween the starch grains in close apposition to proteinbodies. It also was found that eEF1a is localized aroundprotein bodies, where it is merged into a complex with actin.However, the function and mechanism of zein mRNA cyto-skeleton associations remain to be elucidated.
The mechanisms by which zeins assemble into proteinbodies are poorly understood. Zein protein bodies are com-posed of three structurally distinct types of proteins:
�
-zein,
�
-zein (which includes
�
-zein), and
�
-zein (Larkins et al.,1989).
�
-Zeins, which typically are the most abundant, con-tain
�
40 N-terminal amino acids that precede a series ofnine or 10 repeated peptides of 20 amino acids. These re-peats are predicted to be
�
-helical and wind the protein intoa rod-shaped molecule (Argos et al., 1982; Garratt et al.,1993).
�
-Zein, which is related to the
�
-zeins (Woo et al.,2001), contains no repetitive peptides and appears to con-sist mostly of
�
-sheet and turn conformation (Pedersen etal., 1986). Each of the
�
-zeins has a unique N-terminal se-quence (Prat et al., 1985, 1987; Woo et al., 2001). In the50-kD
�
-zein, this region is 136 amino acids in length and isvery His rich (18%). The 27-kD
�
-zein protein has a series ofeight tandem hexapeptide repeats (PPPVHL) that occur 11amino acids after the N terminus. This Pro-rich sequenceappears to function as an ER-retention mechanism (Geli etal., 1994). The first eight amino acids of the 16-kD
�
-zeinprotein are identical to those of the 27-kD
�
-zein, but it hasthree degenerate versions of the Pro-rich repeat (PPPF/HH/YM/L). Approximately the last 140 amino acids of the
�
- and
�
-zeins are 85% identical. The
�
-zeins, which are the mosthydrophobic proteins of the group, contain no repetitive pep-tides and are exceptionally rich in Met (23%) and Cys (4%)(Kirihara et al., 1988; Chui and Falco, 1995).
The pattern of temporal and spatial accumulation of zeinsin protein bodies implies that specific interactions between
them are important for their association (Lending and Larkins,1989; Esen and Stetler, 1992). The smallest protein bodiesconsist of aggregates of
�
- and
�
-zeins. The
�
- and
�
-zeinssubsequently penetrate this network, filling the interior ofthe protein body and expanding it until it reaches a diameterof 1 to 2
�
m. Immunogold labeling showed that the
�
- and
�
-zeins are concentrated toward the periphery of the proteinbody, although they also are detected in the interior. Thissequential pattern of zein protein accumulation is consistentwith the temporal and spatial distribution of their mRNAs inendosperm cells (Woo et al., 2001).
Experiments with transgenic plants also support the hy-pothesis that specific interactions between zeins influenceprotein body assembly. Studies in which genes encodingone or more types of zein proteins were expressed in devel-oping endosperm (Coleman et al., 1996) or other tissues oftransgenic tobacco plants (Bagga et al., 1997) provided evi-dence that
�
- and
�
-zeins interact with
�
- and
�
-zeins, pro-moting their retention in the ER and their incorporation intoprotein bodies. When synthesized individually,
�
- and
�
-zeinsformed protein accretions that were retained within the ERand that appeared to be reasonably stable over prolongedperiods of time. However, when
�
- and
�
-zeins were synthe-sized alone, they were not retained in the ER and appearedto become degraded. The PPPVHL repeats at the N termi-nus of the 27-kD
�
-zein protein could provide the mecha-nism for its retention in the ER (Geli et al., 1994). It wassuggested that this sequence can form an amphipathic helixthat interacts with the surface of the ER (Rabanal et al.,1993). Perhaps this Pro-rich sequence nucleates proteinbody formation, leading to interactions between
�
-zeins viatheir conserved C-terminal regions. These proteins thencould bind and retain the
�
- and
�
-zeins, leading to their ac-cumulation in the protein body.
Because zein protein bodies enlarge to form uniformspherical protein accretions in what appear to be localizedregions of the ER, a hypothesis that explains the previousobservations describing protein body formation in maize en-dosperm involves targeting of these mRNAs, or at least the
�
-zein mRNAs, to distinct regions of the rough ER. To inves-tigate the importance of mRNA targeting in zein proteinbody formation, we characterized the subcellular distribu-tion of zein mRNAs in developing endosperm. On the basisof in situ hybridization, we did not find differences in the oc-currence of the 27-kD
�
-zein and the 22-kD
�
-zein mRNAson cisternal and protein body ER. Thus, although zeinmRNA trafficking to the ER is possible, the mRNAs do notseem to be targeted asymmetrically to domains of the cis-ternal and protein body ER.
To examine the interactions between zein proteins andtheir role in protein body assembly, cDNAs encoding thevarious types of zeins were expressed in the yeast two-hybrid system. These experiments revealed strong affinitiesbetween
�
- and
�
-zeins and relatively weak interactions be-tween
�
-zeins. Surprisingly, there were stronger interactionsbetween the 15-kD
�
-zein and the 16-kD
�
-zein and the 19-
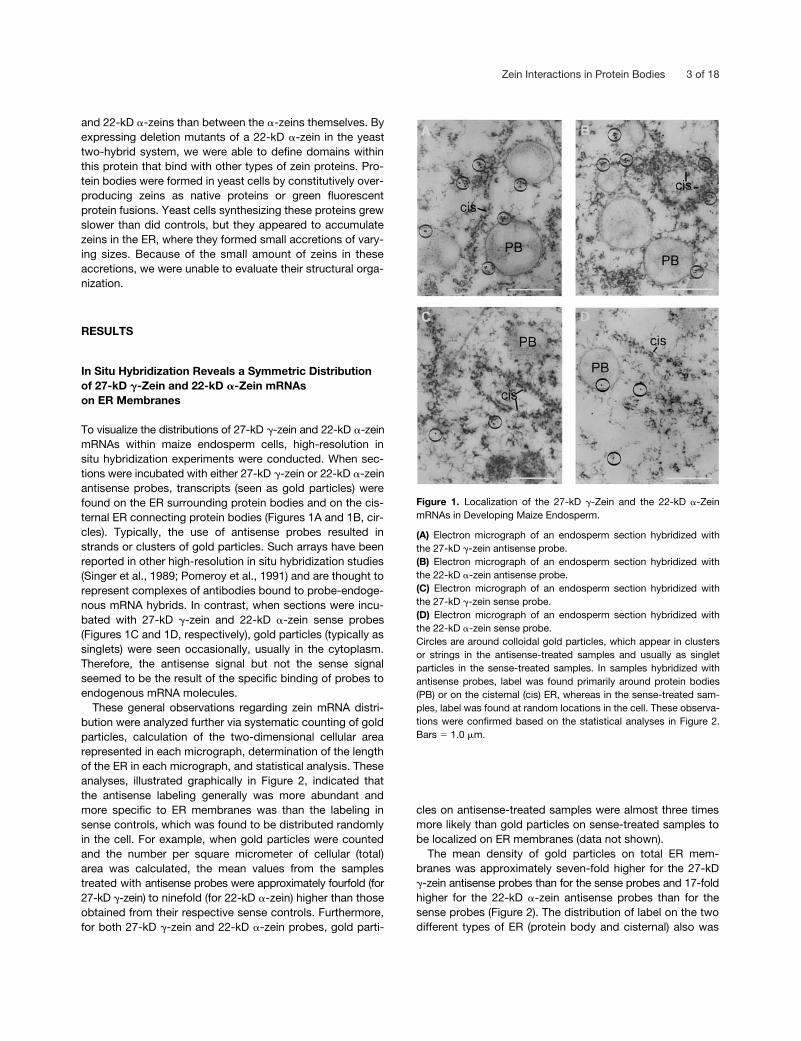
Zein Interactions in Protein Bodies 3 of 18
and 22-kD
�
-zeins than between the
�
-zeins themselves. Byexpressing deletion mutants of a 22-kD
�
-zein in the yeasttwo-hybrid system, we were able to define domains withinthis protein that bind with other types of zein proteins. Pro-tein bodies were formed in yeast cells by constitutively over-producing zeins as native proteins or green fluorescentprotein fusions. Yeast cells synthesizing these proteins grewslower than did controls, but they appeared to accumulatezeins in the ER, where they formed small accretions of vary-ing sizes. Because of the small amount of zeins in theseaccretions, we were unable to evaluate their structural orga-nization.
RESULTS
In Situ Hybridization Reveals a Symmetric Distribution of 27-kD
�
-Zein and 22-kD
�
-Zein mRNAs on ER Membranes
To visualize the distributions of 27-kD
�
-zein and 22-kD
�
-zeinmRNAs within maize endosperm cells, high-resolution insitu hybridization experiments were conducted. When sec-tions were incubated with either 27-kD
�
-zein or 22-kD
�
-zeinantisense probes, transcripts (seen as gold particles) werefound on the ER surrounding protein bodies and on the cis-ternal ER connecting protein bodies (Figures 1A and 1B, cir-cles). Typically, the use of antisense probes resulted instrands or clusters of gold particles. Such arrays have beenreported in other high-resolution in situ hybridization studies(Singer et al., 1989; Pomeroy et al., 1991) and are thought torepresent complexes of antibodies bound to probe-endoge-nous mRNA hybrids. In contrast, when sections were incu-bated with 27-kD
�
-zein and 22-kD
�
-zein sense probes(Figures 1C and 1D, respectively), gold particles (typically assinglets) were seen occasionally, usually in the cytoplasm.Therefore, the antisense signal but not the sense signalseemed to be the result of the specific binding of probes toendogenous mRNA molecules.
These general observations regarding zein mRNA distri-bution were analyzed further via systematic counting of goldparticles, calculation of the two-dimensional cellular arearepresented in each micrograph, determination of the lengthof the ER in each micrograph, and statistical analysis. Theseanalyses, illustrated graphically in Figure 2, indicated thatthe antisense labeling generally was more abundant andmore specific to ER membranes was than the labeling insense controls, which was found to be distributed randomlyin the cell. For example, when gold particles were countedand the number per square micrometer of cellular (total)area was calculated, the mean values from the samplestreated with antisense probes were approximately fourfold (for27-kD
�
-zein) to ninefold (for 22-kD
�
-zein) higher than thoseobtained from their respective sense controls. Furthermore,for both 27-kD
�
-zein and 22-kD
�
-zein probes, gold parti-
cles on antisense-treated samples were almost three timesmore likely than gold particles on sense-treated samples tobe localized on ER membranes (data not shown).
The mean density of gold particles on total ER mem-branes was approximately seven-fold higher for the 27-kD
�
-zein antisense probes than for the sense probes and 17-foldhigher for the 22-kD
�
-zein antisense probes than for thesense probes (Figure 2). The distribution of label on the twodifferent types of ER (protein body and cisternal) also was
Figure 1. Localization of the 27-kD �-Zein and the 22-kD �-ZeinmRNAs in Developing Maize Endosperm.
(A) Electron micrograph of an endosperm section hybridized withthe 27-kD �-zein antisense probe.(B) Electron micrograph of an endosperm section hybridized withthe 22-kD �-zein antisense probe.(C) Electron micrograph of an endosperm section hybridized withthe 27-kD �-zein sense probe.(D) Electron micrograph of an endosperm section hybridized withthe 22-kD �-zein sense probe.Circles are around colloidal gold particles, which appear in clustersor strings in the antisense-treated samples and usually as singletparticles in the sense-treated samples. In samples hybridized withantisense probes, label was found primarily around protein bodies(PB) or on the cisternal (cis) ER, whereas in the sense-treated sam-ples, label was found at random locations in the cell. These observa-tions were confirmed based on the statistical analyses in Figure 2.Bars � 1.0 �m.

4 of 18 The Plant Cell
analyzed (Figure 2). The density (i.e., number per microme-ter) of gold particles was 0.138
0.014 on the protein bodyER of samples treated with 27-kD
�
-zein antisense probesand 0.194
0.114 for samples treated with 22-kD
�
-zeinantisense probes. The density of gold particles was 0.110
0.059 on the cisternal ER of samples treated with 27-kD
�
-zeinantisense probes and 0.210
0.116 for samples treatedwith 22-kD
�-zein antisense probes. Therefore, taking intoaccount standard error values, the extent of labeling on theprotein body ER was similar to that on the cisternal ER forboth 27-kD �-zein and 22-kD �-zein antisense probes. Mostimportantly, there was no asymmetry in the distribution of27-kD �-zein versus 22-kD �-zein mRNAs, because bothwere found in approximately equivalent amounts on proteinbody as well as cisternal ER.
Analysis of Zein Protein Interactions with the Yeast Two-Hybrid System
Interactions between the different types of zein proteins ap-pear to mediate their association into protein bodies (Colemanet al., 1996; Bagga et al., 1997). To investigate the nature ofthese interactions and the roles of specific zeins in proteinbody formation, we expressed zein coding sequences, afterremoval of their signal peptides, in yeast two-hybrid vectors(Bai and Elledge, 1996). The different types of zein geneswere expressed pairwise in both activation domain (pACT2)and DNA binding domain (pAS2) plasmids, and the strengthof the interaction was evaluated by the intensity of the X-Galcolor reaction (Figure 3A) and, more sensitively, by growth inmedium containing 3-aminotriazole (Figure 3B). The 50-kD�-zein interacted strongly with itself and the 27- and 16-kD�-zeins in both the activation and DNA binding domain plas-mids. It showed very weak or no interaction with the 22- and
19-kD �-zeins and the 15-kD �-zein, although there wasweak interaction with the 10-kD �-zein when it was in theactivation domain vector. The 27-kD �-zein interacted morestrongly with the 50- and 16-kD �-zeins than it did with it-self; these interactions were strongest when it was in the ac-tivation domain plasmid. The 27-kD �-zein had a very weakinteraction with the 19-kD �-zein, and this was true as wellof the reciprocal relationship.
Unlike the 27-kD �-zein, the 16-kD �-zein interactedstrongly with itself and the 15-kD �-zein. It had a stronginteraction with the 22-kD �-zein and a strong or weak inter-action with the 19-kD �-zein and the 10-kD �-zein, depend-ing on whether it was expressed in the activation or the DNAbinding domain vector. The 22-kD �-zein was expressed inyeast two-hybrid vectors as the mature protein as well as aC-terminal green fluorescent protein (GFP) fusion. The ma-ture protein interacted weakly with itself, but the strength ofthe interaction was increased with the 22-kD �-zein::GFPfusion in the activation domain vector. The 22-kD �-zein hada strong interaction with the 16-kD �-zein in both yeast two-hybrid vectors and when expressed with a C-terminal GFPfusion. It had a strong or weak interaction with the 15-kD�-zein, depending on the expression vector; the strength ofthis interaction was increased by the C-terminal GFP fusion.The 22-kD �-zein had a strong interaction with the 10-kD�-zein in both vectors, with or without the GFP fusion. Incontrast to the 22-kD �-zein, the 19-kD �-zein interactedstrongly with itself but weakly with the 15-kD �-zein and the10-kD �-zein. The 15-kD �-zein interacted strongly with it-self, the 16-kD �-zein, and the 10-kD �-zein. Besides thepreviously described interactions of the 10-kD �-zein, it alsointeracted strongly with itself.
During protein body assembly, �-zeins appear to pene-trate a network of �-zeins and then associate, along with thesmall amount of �-zeins, and fill the interior of the proteinbody (Lending and Larkins, 1989; Esen and Stetler, 1992).Because little is known about the interactions between �- and�-zeins, we used the yeast two-hybrid system to examinepeptide domains within a 22-kD �-zein that interact withother types of zein proteins. For these experiments, a con-struct encoding a 22-kD �-zein with a C-terminal GFP fusionwas used so that the proteins could be visualized subse-quently in transformed yeast cells. We expressed the 22-kD�-zein::GFP protein in the pACT2 plasmid, and the otherzein proteins were expressed in the pAS2 vector. As shownin Figure 4 (construct a), the 22-kD �-zein::GFP fusion pro-tein interacted strongly with the 22-kD �-zein, 15-kD �-zein,10-kD �-zein, and 16-kD �-zein proteins. These results weresimilar to those obtained with the mature zein proteins (Fig-ure 3) and demonstrated that the GFP fusion did not interferewith the interactions between zeins leading to an activeGAL4 transcription factor. Furthermore, no �-galactosidaseactivity was detected in yeast cells with a pACT2 constructexpressing GFP alone and a pAS2 construct expressing zeinproteins (Figures 4A and 4B, construct i).
The �-zein proteins contain a distinct N-terminal leader of
Figure 2. Distribution of Colloidal Gold Particles per Micrometer ofProtein Body versus Cisternal ER on Endosperm Sections Treatedwith 22-kD �-Zein and 27-kD �-Zein Antisense and Sense Probes(as Illustrated in Figure 1).
The method for counting gold particles is described in Methods; er-ror bars indicate standard error. cis, cisternal; PB, protein body.

Zein Interactions in Protein Bodies 5 of 18
�40 amino acids followed by a series of eight or nine 20–amino acid repeated peptides and a C terminus of �35amino acids (Argos et al., 1982). A series of deletion mu-tants was constructed in which the N-terminal leader, a vari-able number of the repeated peptides, or the C-terminalregion was deleted. These sequences were cloned into thepACT2 vector as GFP fusions, and the interactions with zeinproteins in the pAS2 plasmid were tested. When all of therepeated peptides were deleted, leaving only the N- andC-terminal ends (Figures 4A and 4B, construct j), there wasno evidence of �-galactosidase activity in yeast cells, sug-
gesting that the repeated peptide domain is important forthe interaction between the 22-kD �-zein and the othertypes of zein proteins. The sequential deletion of the nine re-peated peptides (Figures 4A and 4B, constructs a to h) in-creasingly weakened the yeast two-hybrid interactions.There were strong interactions with constructs missing theN terminus or the N terminus plus the first two repeatedpeptides (Figures 4A and 4B, constructs b and c). Deletionof the N terminus and the first four repeated peptidesblocked the yeast two-hybrid interactions (Figures 4A and4B, construct d), but a mutant missing six peptide repeatshad strong interactions (Figures 4A and 4B, construct e),and a mutant missing seven repeats had weak interactions(Figures 4A and 4B, construct f). Mutants missing eight ornine repeated peptides (C-terminal peptide only) alsoshowed no interaction in the yeast two-hybrid assay (Fig-ures 4A and 4B, constructs g and h). These results sug-gested that one or more of the repeated peptides, perhapstoward the C terminus, is important for the interactions be-tween the 22-kD �-zein and the other types of zein proteins.
To further define the regions within the 22-kD �-zein thatbind with the other zein proteins, an additional series of de-letion mutants was constructed with a variable number ofrepeated peptides with or without the N- and C-terminalleader sequences. Construct k in Figure 4 contains theN-terminal leader, the last three peptide repeats, and the Cterminus. This protein interacted strongly with the 16-kD�-zein and the 15-kD �-zein but interacted weakly with the22-kD �-zein and the 10-kD �-zein. If the N- and C-terminalpeptides were removed from construct k (Figures 4A and4B, construct l), there was no interaction with any of the zeinproteins, again suggesting the importance of the N- andC-terminal peptide sequences. Because the last two re-peated peptides plus the C-terminal sequence (Figures 4Aand 4B, construct f) promoted a weak interaction with otherzein proteins, an additional construct was made in which thelast 18 amino acids of the C terminus were deleted (Figure 4,construct m). Like construct f, construct m also had a weakinteraction with these proteins, suggesting that the first half ofthe C terminus is functionally important for zein interactions.
To examine the interactions of the N-terminal sequence ofthe 22-kD �-zein, the last three peptide repeats plus the Cterminus were eliminated (Figure 4, construct n), becausethis region appeared to be sufficient to promote a strongyeast two-hybrid interaction with multiple types of zein pro-teins. Surprisingly, this construct gave a strong �-galactosi-dase reaction with each of the other zeins. Therefore, wemade constructs consisting of only the N terminus plus twopeptide repeats or just the first two peptide repeats alone(Figures 4A and 4B, constructs o and p). With construct o,there was a strong interaction with the 22-kD �-zein and the10-kD �-zein and a weak interaction with 16-kD �-zein andthe 15-kD �-zein. However, no interaction was apparentwhen only the first two repeated peptides were expressed,indicating that the N terminus must play an important role inthese interactions.
Figure 3. Assay of Zein Interactions with the Yeast Two-HybridSystem.
After removal of the signal peptide sequence, zein coding regionswere subcloned into the pAS2 and pACT2 plasmids of the yeasttwo-hybrid system and transformed reciprocally into yeast cells.(A) Yeast cells inoculated on filters and grown with X-Gal overnight.(B) Yeast grown on plates containing 3-aminotriazole for 6 days. Strong(s) indicates that colonies expressing the two indicated zeins had a pos-itive reaction in the X-Gal filter lift assay and scored positively for growthon medium containing 3-aminotriazole compared with negative con-trols. Weak (w) indicates that colonies expressing the indicated zeinsshowed a slight color reaction in the X-Gal assay but grew slowly onselection medium containing 3-aminotriazole compared with controls.() indicates that no X-Gal reaction was detected.

6 of 18 The Plant Cell
On the basis of the analyses of the 22-kD �-zein mutants, itappeared that amino acid sequences within the N- and C-ter-minal regions, as well as a small number of repeated pep-tides, promote interactions with the 22-kD �-zein, 15-kD�-zein, 16-kD �-zein, and 10-kD �-zein proteins. Becauseconstructs containing the last two repeated peptides, all orpart of the C terminus (Figures 4A and 4B, constructs f andm) and the N terminus, and the first two repeated peptidesgave strong or at least weak interactions with this subset ofzein proteins, two additional constructs were tested. The firstcontained the N terminus, the first two peptide repeats, the lasttwo peptide repeats, and the first 17 amino acids of the C ter-minus (Figures 4A and 4B, construct q). This protein inter-acted strongly with these zein proteins, consistent with theother experiments showing the importance of the N- and C-ter-minal sequences plus a small number of peptide repeatsfor promoting interactions with other zein proteins. With aview to using this truncated version of the 22-kD �-zein as avehicle to produce fusion proteins in transgenic plants, we in-serted GFP between the first two and last two repeated pep-
tides of construct q to evaluate how the hybrid protein(Figures 4A and 4B, construct r) would interact with theseproteins. As was true of construct q, this protein interactedvery strongly with this group of zein proteins based on growthin 3-aminotriazole and the �-galactosidase filter assay.
Because the �-galactosidase filter assay does not providea quantitative measure of the two-hybrid interaction be-tween zein proteins, a second set of experiments was per-formed in which �-galactosidase activity was assayed inlysates of yeast cells expressing combinations of zein pro-teins. Figure 5A shows the �-galactosidase activity of yeastcells expressing the full-length 22-kD �-zein in the pACT2plasmid and the other types of zeins in the pAS2 plasmid.For this assay, a 300-�L aliquot of an overnight culture wasinoculated in fresh medium, and the yeast cells were har-vested 10 hr later when the cultures reached an OD600 of 0.8OD. After cell lysis, �-galactosidase was assayed spectro-photometrically using o-nitrophenyl �-D-galactopyranoside(ONPG) as a substrate. Fundamentally, the results of thisanalysis were consistent with those obtained from the filterassay, although the measurement of �-galactosidase activitywas much more sensitive and quantitative. The 22-kD �-zeininteracted most strongly with the 16-kD �-zein and the 15-kD�-zein. It had a stronger interaction with the 10-kD �-zeinthan it did with itself or the 19-kD �-zein. Interactions withthe 50- and 27-kD �-zeins were barely detectable with thisassay.
We next assayed the strength of the two-hybrid interactionsusing the 22-kD �-zein deletion mutants (Figure 4A, con-structs k, o, and q) and construct r, which is equivalent to con-struct q but with GFP inserted into the center of the repeatedpeptide domain. These assays were performed similar to thatfor the full-length 22-kD �-zein::GFP construct, but a cultureperiod of 6 hr was sufficient to reach an OD of 0.8. The resultsobtained with constructs k (Figure 5B), o (Figure 5C), and q(Figure 5D) were similar to those of the filter assay (Figure 4).Constructs k and q had the strongest interactions with the16-kD �-zein and the 15-kD �-zein; in fact, the relative amountof �-galactosidase activity was similar in the correspondingyeast strains. The addition of the two peptide repeats at the Nterminus strengthened the interaction with the 22-kD �-zeinand the 10-kD �-zein (cf. Figures 5B and 5D). The �-galactosi-dase activity from construct o, which is missing the last sevenrepeated peptides and the C terminus, was the reciprocal ofthat from constructs k and q: interactions with the 16-kD �-zeinand the 15-kD �-zein were weaker than those with the 22-kD�-zein and the 10-kD �-zein. The activities of �-galactosidaseassays with construct r were very similar to those with con-struct q; however, construct r had a slightly stronger interac-tion with the 16-kD �-zein.
Synthesis and Processing of Zein Proteins in Yeast Cells
We tested yeast cells as a heterologous system to synthe-size zeins and form protein bodies. The coding sequences
Figure 4. Assay of Yeast Two-Hybrid Interactions between 22-kD�-Zein Deletion Mutants and Other Types of Zein Proteins.
(A) Construction of the 22-kD �-zein deletion mutants in the pACT2plasmid. White boxes correspond to the N-terminal leader followingthe signal peptide; dotted, hatched, and cross-hatched boxes (1 to9) correspond to the 20–amino acid repeated peptides; the C-termi-nal 17 amino acids (17 aa) are indicated by solid gray boxes; and theC-terminal GFP fusion is shown by black boxes.(B) The nature of the yeast two-hybrid interaction between 22-kD�-zein deletion mutants and zein proteins. Strong (s) indicates thatcolonies expressing the two indicated zeins showed a positive reac-tion in the X-Gal filter lift assay and scored positively for growth onmedium containing 3-aminotriazole compared with negative controls.Weak (w) indicates that colonies expressing the indicated zeinsshowed a slight color reaction in the X-Gal assay but grew slowly onselective medium containing 3-aminotriazole compared with con-trols. () indicates that no X-Gal reaction was detected.

Zein Interactions in Protein Bodies 7 of 18
of genes encoding the 22-kD �-zein, 15-kD �-zein, and 27-kD�-zein were subcloned into the yeast expression vectorspGPD414 and pGPD426 between the glyceraldehyde-3-phos-phate dehydrogenase constitutive promoter and the cyto-chrome c oxidase terminator (Mumberg et al., 1995). Yeastcells synthesizing �-, �-, �-, and �-zeins, or GFP, werefound to grow more slowly than empty vector controls. Typ-ically, freshly inoculated cultures of the empty vector controlstrain reached an A600 of 1.0 OD (midlog phase) in �12 to 15
hr, whereas cells expressing any one of the heterologouszein proteins reached this OD in �25 to 30 hr. Strains syn-thesizing GFP or the 22-kD �-zein::GFP, 15-kD �-zein::GFP,and 16-kD �-zein::GFP fusions (see below) grew more rap-idly than those producing native zein proteins; those ex-pressing the 15-kD �-zein and the 27- and 16-kD �-zeinsgrew slowest. We observed different phenotypes for cul-tures producing individual and combinations of zein pro-teins. Cells synthesizing 22-kD �-zein::GFP with the 15-kD�-zein or 22-kD �-zein::GFP with the 16-kD �-zein tended toclump in solution and formed more friable colonies onplates.
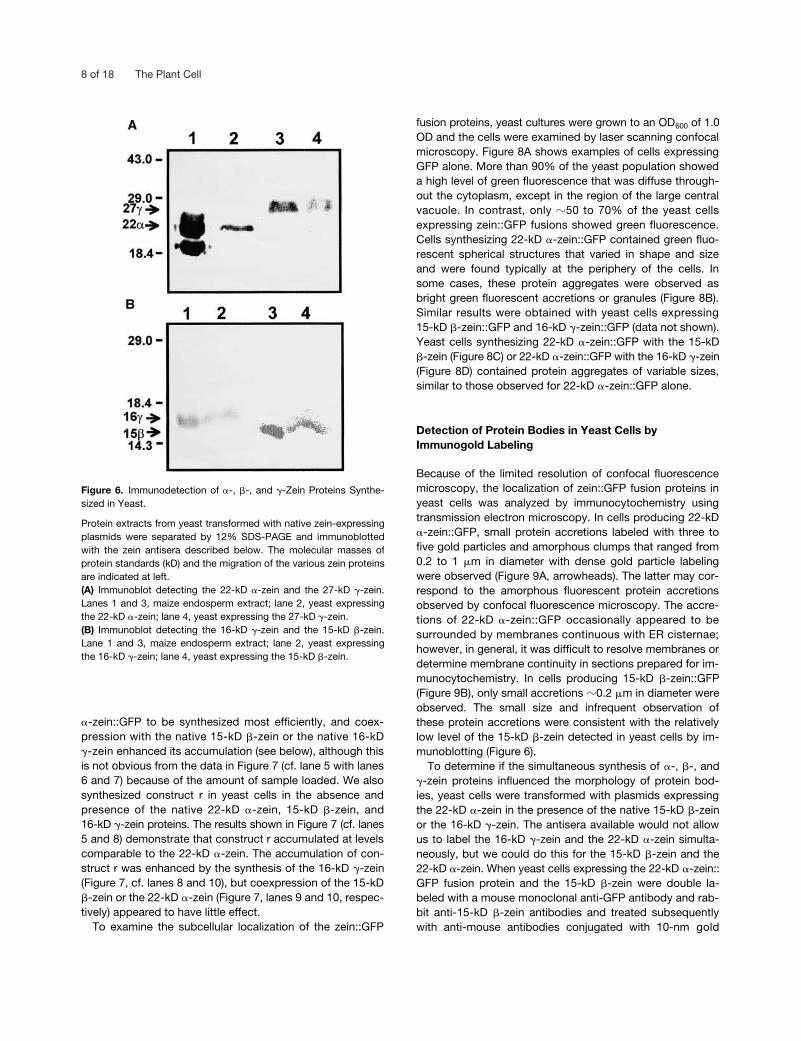
Figure 6 shows immunoblots made after SDS-PAGE sep-aration of the 22-kD �-zein, 15-kD �-zein, and 27-kD �-zeinsynthesized in yeast cell cultures. The 22-kD �-zein and the27-kD �-zein were produced in sufficient amounts that theywere detected in 5 �L of cell lysate (Figure 6A, lanes 2 and4), but the 15-kD �-zein and the 16-kD �-zein accumulatedin smaller amounts (Figure 6B, lanes 2 and 4), and it wasnecessary to load four times more lysate to detect themeasily. Although zeins from yeast extracts tended to formsmeared bands during SDS-PAGE (Figures 6A and 6B, cf.lanes 1 and 2 with lanes 3 and 4), especially with overloadedsamples, their apparent molecular masses were similar tothose of native zein proteins. In underloaded gels, the pro-teins showed identical mobility with native zein proteins(data not shown). Thus, these proteins appeared to undergonormal signal peptide cleavage in yeast cells, a conclusionsupported by the subcellular localization of the proteinswithin membrane vesicles, presumably the ER (see below).
Zein::GFP Fusion Proteins Form Fluorescent Accretions in Yeast Cells
To determine the subcellular localization of zein proteins inyeast and examine the interactions between them that influ-ence protein body formation, we constructed C-terminalGFP fusion proteins with cDNAs encoding the 22-kD �-zein,16-kD �-zein, and 15-kD �-zein and expressed them usingthe pGPD414 and pGPD426 vectors. Cells were grown onselection medium lacking tryptophan or uracil to an OD600 of1.0 and lysed, and the proteins were separated by SDS-PAGE. Figure 7 shows the immunodetection of GFP andzein::GFP fusion proteins, produced individually or in com-bination with native zeins, in yeast cells. A single polypep-tide band of the expected molecular mass was detected foreach construct. Thus, each protein appeared to be pro-cessed appropriately and accumulated stably; that is, theextracts showed no evidence of proteolysis or impaired pro-cessing of signal peptides.
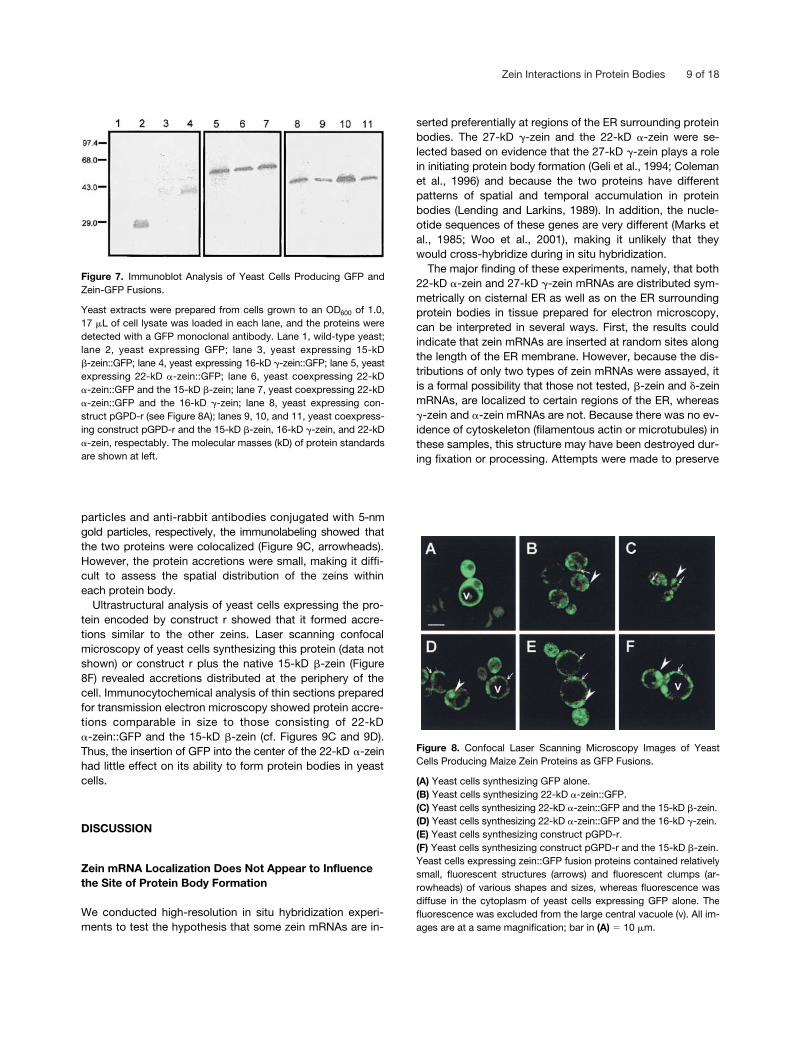
On the basis of the relative staining intensity in the proteingel blot, a smaller amount of the 15-kD �-zein::GFP fusionprotein accumulated in yeast cells compared with the 16-kD�-zein::GFP and 22-kD �-zein::GFP fusion proteins (Figure 7,cf. lane 3 with lanes 4 and 5). Generally, we found the 22-kD
Figure 5. Assay of �-Galactosidase Activity Resulting from YeastTwo-Hybrid Interactions between Selected 22-kD �-Zein DeletionMutants and Other Zein Proteins.
Activation of the LacZ reporter gene, measured as �-galactosidaseactivity with ONPG substrate, was assayed to measure the strengthof the yeast two-hybrid interaction. The values shown are averages ofthree experiments. A diagram illustrating the structure of the 22-kD�-zein deletion mutant (see Figure 4A) in the pACT2 plasmid is pro-vided for each assay.(A) Interaction of the complete 22-kD �-zein (construct pACT2-a) withother zeins.(B) Yeast coexpressing construct pACT2-k and the 22-kD �-zein,16-kD �-zein, 15-kD �-zein, and 10-kD �-zein.(C) Yeast coexpressing construct pACT2-o and the 22-kD �-zein,16-kD �-zein, 15-kD �-zein, and 10-kD �-zein.(D) Yeast coexpressing construct pACT2-q and the 22-kD �-zein,16-kD �-zein, 15-kD �-zein, and 10-kD �-zein.(E) Yeast coexpressing construct pACT2-r and the 22-kD �-zein,16-kD �-zein, 15-kD �-zein, and 10-kD �-zein.Error bars indicate SD.
8 of 18 The Plant Cell
�-zein::GFP to be synthesized most efficiently, and coex-pression with the native 15-kD �-zein or the native 16-kD�-zein enhanced its accumulation (see below), although thisis not obvious from the data in Figure 7 (cf. lane 5 with lanes6 and 7) because of the amount of sample loaded. We alsosynthesized construct r in yeast cells in the absence andpresence of the native 22-kD �-zein, 15-kD �-zein, and16-kD �-zein proteins. The results shown in Figure 7 (cf. lanes5 and 8) demonstrate that construct r accumulated at levelscomparable to the 22-kD �-zein. The accumulation of con-struct r was enhanced by the synthesis of the 16-kD �-zein(Figure 7, cf. lanes 8 and 10), but coexpression of the 15-kD�-zein or the 22-kD �-zein (Figure 7, lanes 9 and 10, respec-tively) appeared to have little effect.
To examine the subcellular localization of the zein::GFP
fusion proteins, yeast cultures were grown to an OD600 of 1.0OD and the cells were examined by laser scanning confocalmicroscopy. Figure 8A shows examples of cells expressingGFP alone. More than 90% of the yeast population showeda high level of green fluorescence that was diffuse through-out the cytoplasm, except in the region of the large centralvacuole. In contrast, only �50 to 70% of the yeast cellsexpressing zein::GFP fusions showed green fluorescence.Cells synthesizing 22-kD �-zein::GFP contained green fluo-rescent spherical structures that varied in shape and sizeand were found typically at the periphery of the cells. Insome cases, these protein aggregates were observed asbright green fluorescent accretions or granules (Figure 8B).Similar results were obtained with yeast cells expressing15-kD �-zein::GFP and 16-kD �-zein::GFP (data not shown).Yeast cells synthesizing 22-kD �-zein::GFP with the 15-kD�-zein (Figure 8C) or 22-kD �-zein::GFP with the 16-kD �-zein(Figure 8D) contained protein aggregates of variable sizes,similar to those observed for 22-kD �-zein::GFP alone.
Detection of Protein Bodies in Yeast Cells by Immunogold Labeling
Because of the limited resolution of confocal fluorescencemicroscopy, the localization of zein::GFP fusion proteins inyeast cells was analyzed by immunocytochemistry usingtransmission electron microscopy. In cells producing 22-kD�-zein::GFP, small protein accretions labeled with three tofive gold particles and amorphous clumps that ranged from0.2 to 1 �m in diameter with dense gold particle labelingwere observed (Figure 9A, arrowheads). The latter may cor-respond to the amorphous fluorescent protein accretionsobserved by confocal fluorescence microscopy. The accre-tions of 22-kD �-zein::GFP occasionally appeared to besurrounded by membranes continuous with ER cisternae;however, in general, it was difficult to resolve membranes ordetermine membrane continuity in sections prepared for im-munocytochemistry. In cells producing 15-kD �-zein::GFP(Figure 9B), only small accretions �0.2 �m in diameter wereobserved. The small size and infrequent observation ofthese protein accretions were consistent with the relativelylow level of the 15-kD �-zein detected in yeast cells by im-munoblotting (Figure 6).
To determine if the simultaneous synthesis of �-, �-, and�-zein proteins influenced the morphology of protein bod-ies, yeast cells were transformed with plasmids expressingthe 22-kD �-zein in the presence of the native 15-kD �-zeinor the 16-kD �-zein. The antisera available would not allowus to label the 16-kD �-zein and the 22-kD �-zein simulta-neously, but we could do this for the 15-kD �-zein and the22-kD �-zein. When yeast cells expressing the 22-kD �-zein::GFP fusion protein and the 15-kD �-zein were double la-beled with a mouse monoclonal anti-GFP antibody and rab-bit anti-15-kD �-zein antibodies and treated subsequentlywith anti-mouse antibodies conjugated with 10-nm gold
Figure 6. Immunodetection of �-, �-, and �-Zein Proteins Synthe-sized in Yeast.
Protein extracts from yeast transformed with native zein-expressingplasmids were separated by 12% SDS-PAGE and immunoblottedwith the zein antisera described below. The molecular masses ofprotein standards (kD) and the migration of the various zein proteinsare indicated at left.(A) Immunoblot detecting the 22-kD �-zein and the 27-kD �-zein.Lanes 1 and 3, maize endosperm extract; lane 2, yeast expressingthe 22-kD �-zein; lane 4, yeast expressing the 27-kD �-zein.(B) Immunoblot detecting the 16-kD �-zein and the 15-kD �-zein.Lane 1 and 3, maize endosperm extract; lane 2, yeast expressingthe 16-kD �-zein; lane 4, yeast expressing the 15-kD �-zein.
Zein Interactions in Protein Bodies 9 of 18
particles and anti-rabbit antibodies conjugated with 5-nmgold particles, respectively, the immunolabeling showed thatthe two proteins were colocalized (Figure 9C, arrowheads).However, the protein accretions were small, making it diffi-cult to assess the spatial distribution of the zeins withineach protein body.
Ultrastructural analysis of yeast cells expressing the pro-tein encoded by construct r showed that it formed accre-tions similar to the other zeins. Laser scanning confocalmicroscopy of yeast cells synthesizing this protein (data notshown) or construct r plus the native 15-kD �-zein (Figure8F) revealed accretions distributed at the periphery of thecell. Immunocytochemical analysis of thin sections preparedfor transmission electron microscopy showed protein accre-tions comparable in size to those consisting of 22-kD�-zein::GFP and the 15-kD �-zein (cf. Figures 9C and 9D).Thus, the insertion of GFP into the center of the 22-kD �-zeinhad little effect on its ability to form protein bodies in yeastcells.
DISCUSSION
Zein mRNA Localization Does Not Appear to Influence the Site of Protein Body Formation
We conducted high-resolution in situ hybridization experi-ments to test the hypothesis that some zein mRNAs are in-
serted preferentially at regions of the ER surrounding proteinbodies. The 27-kD �-zein and the 22-kD �-zein were se-lected based on evidence that the 27-kD �-zein plays a rolein initiating protein body formation (Geli et al., 1994; Colemanet al., 1996) and because the two proteins have differentpatterns of spatial and temporal accumulation in proteinbodies (Lending and Larkins, 1989). In addition, the nucle-otide sequences of these genes are very different (Marks etal., 1985; Woo et al., 2001), making it unlikely that theywould cross-hybridize during in situ hybridization.
The major finding of these experiments, namely, that both22-kD �-zein and 27-kD �-zein mRNAs are distributed sym-metrically on cisternal ER as well as on the ER surroundingprotein bodies in tissue prepared for electron microscopy,can be interpreted in several ways. First, the results couldindicate that zein mRNAs are inserted at random sites alongthe length of the ER membrane. However, because the dis-tributions of only two types of zein mRNAs were assayed, itis a formal possibility that those not tested, �-zein and �-zeinmRNAs, are localized to certain regions of the ER, whereas�-zein and �-zein mRNAs are not. Because there was no ev-idence of cytoskeleton (filamentous actin or microtubules) inthese samples, this structure may have been destroyed dur-ing fixation or processing. Attempts were made to preserve
Figure 7. Immunoblot Analysis of Yeast Cells Producing GFP andZein-GFP Fusions.
Yeast extracts were prepared from cells grown to an OD600 of 1.0,17 �L of cell lysate was loaded in each lane, and the proteins weredetected with a GFP monoclonal antibody. Lane 1, wild-type yeast;lane 2, yeast expressing GFP; lane 3, yeast expressing 15-kD�-zein::GFP; lane 4, yeast expressing 16-kD �-zein::GFP; lane 5, yeastexpressing 22-kD �-zein::GFP; lane 6, yeast coexpressing 22-kD�-zein::GFP and the 15-kD �-zein; lane 7, yeast coexpressing 22-kD�-zein::GFP and the 16-kD �-zein; lane 8, yeast expressing con-struct pGPD-r (see Figure 8A); lanes 9, 10, and 11, yeast coexpress-ing construct pGPD-r and the 15-kD �-zein, 16-kD �-zein, and 22-kD�-zein, respectably. The molecular masses (kD) of protein standardsare shown at left.
Figure 8. Confocal Laser Scanning Microscopy Images of YeastCells Producing Maize Zein Proteins as GFP Fusions.
(A) Yeast cells synthesizing GFP alone.(B) Yeast cells synthesizing 22-kD �-zein::GFP.(C) Yeast cells synthesizing 22-kD �-zein::GFP and the 15-kD �-zein.(D) Yeast cells synthesizing 22-kD �-zein::GFP and the 16-kD �-zein.(E) Yeast cells synthesizing construct pGPD-r.(F) Yeast cells synthesizing construct pGPD-r and the 15-kD �-zein.Yeast cells expressing zein::GFP fusion proteins contained relativelysmall, fluorescent structures (arrows) and fluorescent clumps (ar-rowheads) of various shapes and sizes, whereas fluorescence wasdiffuse in the cytoplasm of yeast cells expressing GFP alone. Thefluorescence was excluded from the large central vacuole (v). All im-ages are at a same magnification; bar in (A) � 10 �m.

10 of 18 The Plant Cell
the cytoskeleton for electron microscopic observation usinghigh-pressure freezing, but these were not successful.Therefore, it is possible that the zein mRNAs, which mayhave been anchored at certain locations in vivo by cytoskel-etal elements, were redistributed artifactually during samplepreparation.
The method we used for mRNA localization is similar tothat described by Li et al. (1993a) to determine mRNA spa-tial distribution in rice endosperm. In the rice study, it wasconcluded that an asymmetric distribution of glutelin versusprolamin mRNAs on the ER occurred despite the absence ofcytoskeletal structures. Therefore, the random distributionof prolamin mRNAs on the ER of maize endosperm, com-pared with their asymmetric distribution in rice endosperm,may indicate a true difference between the two cereals. It ispossible that in rice, the segregation of prolamin and glutelinmRNAs is functionally important, because the prolamins areretained in the ER, whereas the glutelins are transported toprotein storage vacuoles. The segregation of zein mRNAsmay not be necessary, because once the proteins are in-serted into the lumen of the ER, they may simply diffuse andcoalesce based on their biochemical and structural proper-ties (see below). Regardless of whether zein mRNAs are in-serted exclusively at sites in which protein bodies are forming,the cytoskeleton may play a role in transporting them to theER membrane.
If zein mRNAs are targeted more or less randomly to ERmembranes, by what mechanism do the proteins associatewith one another and assemble into a discrete proteinbody? There is evidence that ER chaperonins, such as Bipand PDI, play a role in zein protein folding (Fontes et al.,
1991; Zhang and Boston, 1992; Li and Larkins, 1996), butwhether they are components of a macromolecular complexthat assembles zeins simultaneously into a protein body, assuggested for rice prolamins (Li et al., 1993b), is unclear.The observation that zeins appear to be secreted more orless randomly into the ER suggests the existence of sometype of assembly complex or an autoassembly processbased on the structural features of the zein proteins them-selves.
Interactions between Zein Proteins Can Be Identified with the Yeast Two-Hybrid System
Expressing zeins as Gal4 fusions in the yeast two-hybridsystem appears to be an effective approach to investigateinteractions between these proteins. In interpreting the re-sults of these experiments, we made several hypotheses.First, protein associations in the nucleus are comparable tothose occurring within the lumen of the ER. This hypothesisis implicit in many other experiments in which interactionsbetween nonnuclear proteins are examined with the yeasttwo-hybrid system (Shaywitz et al., 1997; Takatsu et al.,2001). On the basis of the authenticity of the results fromsuch experiments, we believe that this is a valid hypothesis.Second, we assume that the intensity of the �-galactosidasereaction, as measured by the hydrolysis of X-Gal (Figure 3)or ONPG, reflects the amount of functional �-galactosidaseenzyme produced, and hence the affinity between two pro-teins. We also assume that the affinities between interactingzeins are such that differences in the transcription and nu-
Figure 9. Immunolocalization of Zeins in Yeast Cells.
(A) Yeast cells synthesizing 22-kD �-zein::GFP.(B) Yeast cells synthesizing 15-kD �-zein::GFP.(C) Yeast cells synthesizing 22-kD �-zein::GFP and the 15-kD �-zein.(D) Yeast cells synthesizing construct pGPD-r.Arrowheads indicate accretions decorated with multiple gold particles. The most prominent protein bodies found from another cell of the corre-sponding yeast population are shown in the insets. For the single immunolabeling shown in (A), (B), and (D), 10-nm gold particles were used. In(C), 22-kD �-zein::GFP was labeled with 10-nm gold particles, and the 15-kD �-zein was labeled with 5-nm gold particles. Bars � 200 nm.

Zein Interactions in Protein Bodies 11 of 18
clear import of the fusion proteins were negligible. Althoughwe did not measure the level of transcription between yeaststrains expressing the various zein constructs, based on theobservation of consistent levels of �-galactosidase expres-sion in replicated experiments, we believe that this was thecase. Each assay of a two-hybrid interaction was dupli-cated, and the experiments were replicated many times.Based on filter and 3-aminotriazole assays of �-galactosi-dase activity, a functional transcription factor was formedregardless of which vector was used to express zein codingsequences. Cells expressing the zeins with the pACT2 andpAS2 plasmids showed comparable rates of growth and ex-hibited normal phenotypes, in contrast to the expression ofnative zeins in yeast (see below). The addition of a C-termi-nal GFP fusion protein to the mature 22-kD �-zein or dele-tion mutants of this construct did not influence the yeasttwo-hybrid interaction significantly, and GFP was a valuabletool for detecting the proteins when they were made assecretory proteins.
The strong yeast two-hybrid interactions between the �- and�-zeins are consistent with the colocalization of these pro-teins in developing protein bodies (Lending and Larkins,1989; Woo et al., 2001). Although we do not know the struc-ture of the domains that interact, based on the unique fea-tures of the N termini of the 50- and 27-kD �-zein proteins,we hypothesize that it is most likely the C-terminal 130amino acids that are highly conserved among them. We cre-ated deletion mutants of this region with the 27-kD �-zein,but nearly all of these modifications eliminated interactionsin the yeast two-hybrid system. Consequently, it appearsthat this sequence creates a highly ordered secondarystructure that is disrupted easily.
In view of previous experiments showing that the 27-kD�-zein associates with the 22-kD �-zein and promotes itsretention in the ER (Coleman et al., 1996), we were surprisednot to detect a stronger association between the 27-kD �-zeinand the 22- and 19-kD �-zein proteins in the yeast two-hybridsystem (Figures 3 and 5). Nevertheless, we did observe astrong association between the 16-kD �-zein and the 15-kD�-zein and between the 22-kD �-zein and the 10-kD �-zein.This finding suggests that the C-terminal 130 amino acids ofthe �- and �-zein proteins also interact with the �- and �-zeins.If the Pro-rich repeats of the 27-kD �-zein are directed at thesurface of the protein body, as suggested by Geli et al.(1994), then the C-terminal regions of the �-zeins would be in-ternal, and they could be in contact with the �- and �-zeins.
We expected that the �-zeins, which are structurallyclosely related (Argos et al., 1982), would have a greatertendency for self-interaction than for binding with other zeinproteins. However, the self-interaction of the 22-kD �-zein inthe yeast two-hybrid system was weaker than that of the19-kD �-zein (Figure 3), and there was a surprisingly weakinteraction between the 22- and 19-kD �-zeins. A C-terminalGFP fusion to the 22-kD �-zein enhanced its interaction withitself and with the 15-kD �-zein somewhat (as measured inthe 3-aminotriazole assay; Figure 3B); however, this influ-
ence was not obvious in the X-Gal filter assay (Figure 3A).Because yeast cells expressing mature zeins as GFP fu-sions generally grew faster than those without them, thisstronger interaction may be a consequence. The predictedstructure of the �-zein proteins suggests that they associatethrough hydrogen bonds and hydrophobic interactions (Argoset al., 1982; Garratt et al., 1993). It is possible that these areauthentic interactions, but they are too weak to promote theformation of an effective GAL4 transcription factor. Althoughyeast cells grew faster when the zein proteins were made asGFP fusions, the GFP protein did not appear to alter the na-ture of the interactions between these proteins. The C-ter-minal GFP fusion appeared to enhance the 22-kD �-zeinassociation with itself, but it did not alter qualitatively the in-teractions with the native 22- and 19-kD �-zeins. Neither didthe addition of GFP to the 22-kD �-zein appear to influencethe interactions with other types of zein proteins in theONPG assay, which is the most sensitive and quantitativemeasure of yeast two-hybrid interactions (Figure 5).
To identify regions in the 22-kD �-zein that interact withthe �-zeins, we constructed a set of deletion mutants inwhich the N terminus, the C terminus, and variable numbersof the repeated peptides were removed. Because equivalentinteractions were obtained regardless of which yeast two-hybrid vector expressed the native proteins, we tested onlythe mutants in the pACT2 plasmid (Figure 4). Yeast cells ex-pressing the deletion mutants grew twice as fast as thoseexpressing the full-length 22-kD �-zein, but the nature of theprotein interactions was qualitatively the same (cf. Figure 5Awith Figures 5B to 5E). The results of these experimentssuggest that interactions between the 22-kD �-zein andother zein proteins involve more than just the repeated pep-tide domain. The 39 amino acids of the N terminus and thelast 31 amino acids of the C terminus clearly influence theassociation of the 22-kD �-zein with other zein proteins. In-terestingly, the N terminus and the first two repeated pep-tides appear to have an affinity for the 22-kD �-zein and the10-kD �-zein, whereas the last two repeats and the C termi-nus have a greater affinity for the 15-kD �-zein and the 16-kD�-zein (cf. Figures 5B and 5C). It may not be valid to com-pare the strength of the affinities determined by this assay;however, the data in Figures 5B to 5E were obtained withcultures grown simultaneously for the same length of time.
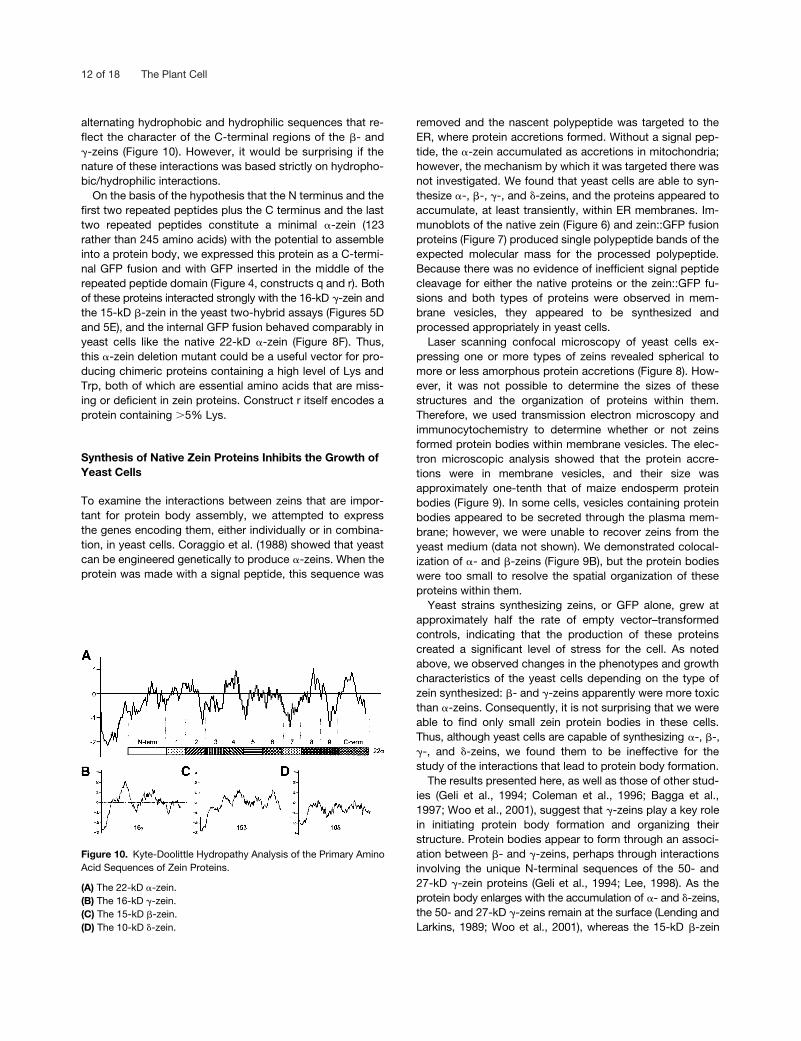
Unfortunately, little is known about the three-dimensionalstructure of zein proteins, and models describing the confor-mation of �-zeins make no predictions regarding the N- andC-terminal regions (Argos et al., 1982; Garratt et al., 1993).Consequently, it is difficult to explain the basis of the affini-ties of the N- and C-terminal regions of the 22-kD �-zein forthe �-/�-zeins and �-/�-zeins, respectively. The hydropathyplots of these regions suggest that the N terminus and thefirst two repeats are more hydrophobic than the last two re-peats and the C terminus (Figure 10). This might explain thegreater affinity of the 10-kD �-zein for the N terminus, be-cause the �-zein is by far the most hydrophobic protein ofthe group. The C terminus and the last two repeats contain
12 of 18 The Plant Cell
alternating hydrophobic and hydrophilic sequences that re-flect the character of the C-terminal regions of the �- and�-zeins (Figure 10). However, it would be surprising if thenature of these interactions was based strictly on hydropho-bic/hydrophilic interactions.
On the basis of the hypothesis that the N terminus and thefirst two repeated peptides plus the C terminus and the lasttwo repeated peptides constitute a minimal �-zein (123rather than 245 amino acids) with the potential to assembleinto a protein body, we expressed this protein as a C-termi-nal GFP fusion and with GFP inserted in the middle of therepeated peptide domain (Figure 4, constructs q and r). Bothof these proteins interacted strongly with the 16-kD �-zein andthe 15-kD �-zein in the yeast two-hybrid assays (Figures 5Dand 5E), and the internal GFP fusion behaved comparably inyeast cells like the native 22-kD �-zein (Figure 8F). Thus,this �-zein deletion mutant could be a useful vector for pro-ducing chimeric proteins containing a high level of Lys andTrp, both of which are essential amino acids that are miss-ing or deficient in zein proteins. Construct r itself encodes aprotein containing �5% Lys.
Synthesis of Native Zein Proteins Inhibits the Growth of Yeast Cells
To examine the interactions between zeins that are impor-tant for protein body assembly, we attempted to expressthe genes encoding them, either individually or in combina-tion, in yeast cells. Coraggio et al. (1988) showed that yeastcan be engineered genetically to produce �-zeins. When theprotein was made with a signal peptide, this sequence was
removed and the nascent polypeptide was targeted to theER, where protein accretions formed. Without a signal pep-tide, the �-zein accumulated as accretions in mitochondria;however, the mechanism by which it was targeted there wasnot investigated. We found that yeast cells are able to syn-thesize �-, �-, �-, and �-zeins, and the proteins appeared toaccumulate, at least transiently, within ER membranes. Im-munoblots of the native zein (Figure 6) and zein::GFP fusionproteins (Figure 7) produced single polypeptide bands of theexpected molecular mass for the processed polypeptide.Because there was no evidence of inefficient signal peptidecleavage for either the native proteins or the zein::GFP fu-sions and both types of proteins were observed in mem-brane vesicles, they appeared to be synthesized andprocessed appropriately in yeast cells.
Laser scanning confocal microscopy of yeast cells ex-pressing one or more types of zeins revealed spherical tomore or less amorphous protein accretions (Figure 8). How-ever, it was not possible to determine the sizes of thesestructures and the organization of proteins within them.Therefore, we used transmission electron microscopy andimmunocytochemistry to determine whether or not zeinsformed protein bodies within membrane vesicles. The elec-tron microscopic analysis showed that the protein accre-tions were in membrane vesicles, and their size wasapproximately one-tenth that of maize endosperm proteinbodies (Figure 9). In some cells, vesicles containing proteinbodies appeared to be secreted through the plasma mem-brane; however, we were unable to recover zeins from theyeast medium (data not shown). We demonstrated colocal-ization of �- and �-zeins (Figure 9B), but the protein bodieswere too small to resolve the spatial organization of theseproteins within them.
Yeast strains synthesizing zeins, or GFP alone, grew atapproximately half the rate of empty vector–transformedcontrols, indicating that the production of these proteinscreated a significant level of stress for the cell. As notedabove, we observed changes in the phenotypes and growthcharacteristics of the yeast cells depending on the type ofzein synthesized: �- and �-zeins apparently were more toxicthan �-zeins. Consequently, it is not surprising that we wereable to find only small zein protein bodies in these cells.Thus, although yeast cells are capable of synthesizing �-, �-,�-, and �-zeins, we found them to be ineffective for thestudy of the interactions that lead to protein body formation.
The results presented here, as well as those of other stud-ies (Geli et al., 1994; Coleman et al., 1996; Bagga et al.,1997; Woo et al., 2001), suggest that �-zeins play a key rolein initiating protein body formation and organizing theirstructure. Protein bodies appear to form through an associ-ation between �- and �-zeins, perhaps through interactionsinvolving the unique N-terminal sequences of the 50- and27-kD �-zein proteins (Geli et al., 1994; Lee, 1998). As theprotein body enlarges with the accumulation of �- and �-zeins,the 50- and 27-kD �-zeins remain at the surface (Lending andLarkins, 1989; Woo et al., 2001), whereas the 15-kD �-zein
Figure 10. Kyte-Doolittle Hydropathy Analysis of the Primary AminoAcid Sequences of Zein Proteins.
(A) The 22-kD �-zein.(B) The 16-kD �-zein.(C) The 15-kD �-zein.(D) The 10-kD �-zein.

Zein Interactions in Protein Bodies 13 of 18
and the 16-kD �-zein proteins may become displaced to-ward the interior through association with the �- and �-zeins.Lending and Larkins (1989) showed that as protein bodiesincrease in diameter, immunolabeling of the �- and �-zeinsresulted in a number of gold particles scattered across thecenter of the protein body, suggesting that these proteinsare not restricted to the surface.
Because the �-zein antiserum used in this study (Lendinget al., 1988) reacted with the 27-kD �-zein, but not with the50- and 16-kD �-zeins, and the �-zein antiserum cross-reacted with the 15-kD �-zein and the 16-kD �-zein pro-teins, it is possible that the proteins detected in the center ofthe protein body are the 15-kD �-zein and 16-kD �-zeinpolypeptides. This hypothesis is consistent with the obser-vation that the center of the protein body stains relativelylightly with uranyl acetate. The outer surface of the proteinbody has a dark staining intensity, consistent with this re-gion binding a higher concentration of this heavy metal(Lending and Larkins, 1989). Because the N-terminal regionsof the 50- and 27-kD �-zein proteins contain 13 to 18%His, in contrast to the N-terminal regions of the 16-kD�-zein and the 15-kD �-zein, which contain 4% and no His,respectively, it is possible that the dark appearance of theprotein body surface results from uranyl acetate binding toHis, although it also could be influenced by the high Cyscontent of these proteins (Woo et al., 2001).
This model of zein protein body structure suggests an im-portant role for the 15-kD �-zein and the 16-kD �-zein inbinding and organizing �- and �-zeins in the center of theprotein body. Because of the conserved nature of the C-ter-minal domains of the �- and �-zeins, a mutant deficient inone of these proteins might not show an altered kernel phe-notype. However, mutations that disrupt the structure ofthese proteins could affect protein body organization, re-sulting in an altered kernel phenotype, similar to floury2(Coleman et al., 1997). We determined recently that the Mu-curonate mutant contains a defective 16-kD �-zein, resultingfrom an altered reading frame toward the C terminus of thepolypeptide (R. Wrobel, J. Gillikan, and R.S. Boston, unpub-lished data; C.S. Kim, B.C. Gibbon, A. Tikhonov, R. Jung,and B.A. Larkins, unpublished data), and protein bodies inthis opaque mutant have an irregular, warty appearance.Experiments are in progress to further characterize thestructure of the C-terminal domain of the 16-kD �-zein anddetermine its interaction with �-zein proteins.
METHODS
In Situ Hybridization
Maize (Zea mays) plants (W64A�) were grown at the University of Ar-izona West Agricultural Research Center in Tucson. Kernels wereharvested at 14 to 16 days after pollination, and approximately2-mm3 pieces of endosperm were placed in fixative (4% formalde-hyde and 0.5% glutaraldehyde in 50 mM KPO4, pH 7.0) for 2 hr at
room temperature. The tissue was embedded in LR Gold resin (Lon-don Resin Company Ltd., Ft. Washington, PA) polymerized at roomtemperature using 1% (w/v) benzoyl peroxide paste. Sections 70 to90 nm thick were cut onto formvar-coated nickel grids.
Single-stranded, digoxigenin-labeled RNA probes were made byin vitro transcription. cDNA clones encoding the 22-kD �-zein andthe 27-kD �-zein (Marks et al., 1985; Woo et al., 2001) were clonedinto pBluescript SK� (Statagene, La Jolla, CA) or pT7/T3-18 (Be-thesda Research Laboratories, Bethesda, MD) vectors. These plas-mids have T3 and T7 promoters in opposite orientation flanking thecDNA inserts, so both sense and antisense transcripts can be syn-thesized. Plasmids were linearized with appropriate restriction enzymes,extracted with phenol/chloroform and ethanol, and precipitated. Invitro transcription was performed for 2 to 4 hr at 37 C using 1 to 2 �gof linearized plasmid, 1 unit of RNA polymerase, 1.0 mM ATP, CTP,and GTP, 0.65 mM UTP, 0.35 mM digoxygenein-11-UTP, and 10 mMDTT in the appropriate transcription buffer. Subsequently, plasmid inthe reaction was removed by digestion with 1 unit of DNase, and thereaction was stopped by the addition of EDTA (20 mM final concen-tration).
RNAs were extracted with phenol/chloroform, and the labeledprobe was separated from unincorporated nucleotides during etha-nol precipitation via the addition of glycogen. To ensure that theprobes were of appropriate size and labeled with digoxigenin, theywere separated by electrophoresis on agarose gels and blotted ontonitrocellulose membranes. Probes were visualized by incubating theblot with anti-digoxigenin antibodies conjugated to alkaline phos-phatase, followed by reaction with 5-bromo-4-chloro-3-indolyl phos-phate and nitroblue tetrazolium (Sambrook et al., 1989). Antisensebut not sense probes were confirmed to recognize single bands inRNA gel blots of endosperm transcripts. The concentration of probewas estimated after gel electrophoresis and ethidium bromide stain-ing by visually comparing serial dilutions of in vitro transcripts withstandards of known RNA concentration. Similarly, dot blots of serialdilutions were made, and the digoxigenin was visualized with anti-digoxigenin antibody conjugated to alkaline phosphatase, as de-scribed above. The intensities of these spots were compared withthose of an RNA standard containing a known amount of digoxi-genin. Probes were lyophilized and resuspended in hybridization so-lution (50% formamide, 4 � SSC [1 � SSC is 0.15 M NaCl and 0.015 Msodium citrate], 1 � Denhardt’s solution [1 � Denhardt’s solution is0.02% Ficoll, 0.02% polyvinylpyrrolidone, and 0.02% BSA], 0.5mg/mL heat-denatured salmon sperm DNA, 0.25 mg/mL Escherichiacoli tRNA, 5% dextran sulfate, and 10 mM DTT) at a concentration of37.5 ng/�L.
The pretreatment and hybridization of endosperm tissue sectionswere performed by a combination of protocols described by Li et al.(1993a), McFadden (1991), and Bostwick et al. (1992), with modifica-tions. All incubations took place on baked glass spot plates linedwith Parafilm. Unless noted otherwise, for rinses, grids were dippedinto appropriate solutions in baked glass beakers. All solutions weretreated with diethylpyrocarbonate (DEPC), when possible, or weremade with DEPC-treated water. Tissue sections were incubated inprehybridization solution by floating each grid, tissue side down, on a4-�L droplet for 15 min at 42 C. The grids were floated on 4-�L drop-lets of heat-denatured probe solution overnight at 42 C in a humiditychamber sealed with autoclave (heat-resistant) tape. After hybridiza-tion, the sections were rinsed in 4 � SSC and 2 � SSC for severalminutes and finally incubated in 0.1 � SSC (by floating grids on 15-�Ldroplets) for 2 hr at 48 C. The grids then were floated on 15-�L drop-lets of blocking solution containing 5% normal rabbit serum and 2%BSA in buffer I (150 mM NaCl in 100 mM Tris, pH 7.5).

14 of 18 The Plant Cell
For the detection of the digoxigenin-labeled probes, immunocy-tochemistry was performed as follows. The grids were floated on asolution of anti-digoxigenin Fab fragments (diluted 1:100 in an anti-body solution containing 5% normal rabbit serum and 1% BSA inbuffer I) at room temperature for 1 hr followed by rinsing in buffer I.Subsequently, the grids were floated on a solution of rabbit anti-sheep antibodies conjugated to 15-nm colloidal gold particles (di-luted 1:100 in antibody solution) for 1 hr at room temperature. Thegrids were rinsed in buffer I and then with DEPC-treated water beforepoststaining with 2% uranyl acetate and 2% lead citrate.
Sections were viewed on a Japanese Electron Optical Laborato-ries 100 CX transmission electron microscope (JEOL USA, Peabody,MA) at 80 kV. Micrographs were produced at a variety of magnifica-tions, and the subcellular distributions of gold particles were as-sessed.
A gold particle was determined to be associated with the endo-plasmic reticulum (ER) membrane (either cisternal or protein body) ifit was found within 50 nm of that membrane (Li et al., 1993a). In thecase of protein body ER, when the number of gold particles per mi-crometer of ER was calculated, the circumference of the proteinbody was taken into account; for cisternal ER, both sides of the dou-ble membrane were measured. Measurements were made by firstplacing marks on a string, and the distance between the marks wasmeasured with a ruler. These values were converted to micrometersbased on the micrograph magnification. The two-dimensional cellu-lar surface area represented by each micrograph was calculatedbased on magnification. Microsoft Excel (Microsoft, Redmond, WA)was used for statistical analyses.
Construction of Plasmid Vectors for the Synthesis of Zein Proteins in Yeast
The following plasmid constructs were made with zein cDNAs (Table1) for the synthesis of native proteins and green fluorescent protein(GFP) in yeast.
pGPD22-kD �-zein. A 22-kD �-zein cDNA was amplified by poly-merase chain reaction (PCR) with primers 5�-CGCGGATCCGC-GATGGCTACCAAGATATTATC-3� and 5�-CGGAATTCCGCTAAAAGA-
TGGCACCTCC-3� and cloned into the pGPD414 and pGPD426plasmid vectors between the BamHI and EcoRI cloning sites.
pGPD27-kD �-zein. The 27-kD �-zein cDNA was amplified by PCRwith primers 5�-CGCGGATCCGCGATGAGGGTGTTGCTCGTTGC-3�
and 5�-CGGAATTCCGTCAGTGGGGGACACCGCCGGC-3� and in-serted into the pGPD414 and pGPD426 plasmid vectors between theBamHI and EcoRI cloning sites.
pGPD16-kD �-zein. The 16-kD �-zein cDNA was amplified by PCRwith primers 5�-TCCCCCGGGGGAATGAAGGTGCTGATCGTTGC-3�
and 5�-CGGAATTCCGTCAGTAGTAGACACCGCC-3� and insertedinto the pGPD414 and pGPD426 plasmid vectors between the SmaIand EcoRI cloning sites.
pGPD15-kD �-zein. The 15-kD �-zein cDNA was amplified by PCRwith primers 5�-CGCGGATCCGCGATGAAGATGGTCATCGTTC-3�
and 5�-CGGAATTCCGTCAGTAGTAGGGCGGAATGG-3� and insertedinto the pGPD414 and pGPD426 plasmid vectors between theBamHI and EcoRI cloning sites.
pGPDGFP. The GFP gene was amplified by PCR with primers 5�-TCCCCCGGGGATGGTGAGCAAGGGC-3� and 5�-CGGAATTCCGT-TACTTGTACAGCTCGTC-3� and inserted into the pGPD414 plasmidvector between the SmaI and EcoRI cloning sites.
pGPD-r. The signal peptide of the 22-kD �-zein fragment was am-plified with primers 5�-CGCGGATCCGCGATGGCTACCAAGATAT-TATC-3� and 5�-TGAGCATTGTGGAATAATGGACGCATTTGTTGC-GCTCGCA-3�, and 22��sp-zein::GFP fusion construct (Figure 8A,construct r) sequences were amplified with primers 5�-TGCGAG-CGCAACAAATGCGTCCATTATTCCACAATGCTCA-3� and 5�-CCG-GAATTCCGGCTAGACAGGATTCATCAAAGAGAA-3�. The two PCRproducts were annealed, reamplified with primers 5�-CGCGGATCC-GCGATGGCTACCAAGATATTATC-3� and 5�-CCGGAATTCCGGCTA-GACAGGATTCATCAAAGAGAA-3�, and inserted into pGPD414 atthe BamHI and EcoRI cloning sites.
pGPD22-kD::GFP. The signal peptide of the 22-kD �-zein frag-ment was amplified with primers 5�-CGCGGATCCGCGATGGCTAC-CAAGATATTATC-3� and 5�-TGAGCATTGTGGAATAATGGACGC-ATTTGTTGCGCTCGCA-3�, and 22��sp-zein::GFP fusion construct(Figure 8A, construct a) sequences were amplified with primers 5�-TGCGAGCGCAACAAATGCGTCCATTATTCCACAATGCTCA-3� and5�-CGGAATTCCGTTACTTGTACAGCTCGTC-3�. The two PCR prod-
Table 1. Genes and Oligonucleotide Sequences Used to Make Zein Expression Constructs
Gene Accession Number Upstream and Downstream Primer Sequences
22��sp J01246 5�-TCCCCCGGGGATCTCCATTATTCCACAATGCTCACTTGCTCCT-3� and 5�- GTCGACGGATCCCTAAAAGATGGCACCTCC-3�
19��sp M12146 5�-TCCCCCGGGGATCACAATTTTCCCTCAATGC-3� and5�-GTCGACGGATCCCTAAAAGAGGGCACCACC-3�
50��sp AF371279 5�-AACCATGGGTACACAGACAGGCGGCTGC-3� and5�-ATGGATCCATTGCAAACACCACTAATG-3�
27��sp AF371261 5�-TCCCCCGGGGATCACGCATACAAGCGGCGGC-3� and5�-GTCGACGGATCCTCAGTGGGGGACACCG-3�
16��sp AF371262 5�-TCCCCCGGGGATCAGTACAAGCGGCGGCTGT-3� and5�-CCGCTCGAGCGGTCAGTAGTAGACACCGCC-3�
15��sp M12147 5�-TCCCCCGGGGATCATGCAGATGCCCTGCCCCTGC-3� and5�-GTCGACGGATCCTCAGTAGTAGGGCGGAATGGC-3�
10��sp AF371266 5�-TCCCCCGGGGATCACCCATATTCCAGGGCAC-3� and5�-GTCGACGGATCCCTAGAATGCAGCACCAAC-3�

Zein Interactions in Protein Bodies 15 of 18
ucts were annealed, reamplified with primers 5�-CGCGGATCCGC-GATGGCTACCAAGATATTATC-3� and 5�-CGGAATTCCGTTACTTGT-ACAGCTCGTC-3�, and inserted into pGPD414 at the BamHI andEcoRI cloning sites.
pGPD16-kD �-zein::GFP. The 16-kD �-zein cDNA fragment wasamplified with primers 5�-TCCCCCGGGGGAATGAAGGTGCTG-ATCGTTGC-3� and 5�-CTCGCCCTTGCTCACCATGTAGTAGACAC-CGCCGGC-3�, and GFP gene sequences were amplified with prim-ers 5�-GCCGGCGGTGTCTACTACATGGTGAGCAAGGGCGAG-3� and5�-CGGAATTCCGTTACTTGTACAGCTCGTC-3�. The two PCR prod-ucts were annealed, reamplified with primers 5�-TCCCCCGGG-GGAATGAAGGTGCTGATCGTTGC-3� and 5�-CGGAATTCCGTT-ACTTGTACAGCTCGTC-3�, and inserted into pGPD414 at the SmaIand EcoRI cloning sites.
pGPD15-kD::GFP. The 15-kD gene fragment was amplified withprimers 5�-CGCGGATCCGCGATGAAGATGGTCATCGTTC-3� and 5�-CTCGCCCTTGCTCACCATGTAGTAGGGCGGAATGGC-3�, and GFPgene sequences were amplified with primer 5�-GCCATTCCGCCC-TACTACATGGTGAGCAAGGGCGAG-3� and 5�-CGGAATTCCGTT-ACTTGTACAGCTCGTC-3�. These two PCR products were fused byreamplification with primers 5�-CGCGGATCCGCGATGAAGATGGTC-ATCGTTC-3� and 5�-CGGAATTCCGTTACTTGTACAGCTCGTC-3�
and cloned into pGPD414 at the BamHI and EcoRI sites.The PCR products were inserted into the yeast expression vectors
pGPD414 and pGPD426 between the glyceraldehyde-3-phosphatedehydrogenase promoter (Bitter and Egan, 1984) and the cyto-chrome c oxidase terminator (Guarente et al., 1984). Amplification ofthe construct to fuse DNA fragments or to add restriction enzymesites was performed by PCR with Pfu-Turbo DNA polymerase (Strat-agene, La Jolla, CA). The nucleotide sequences of newly constructedclones were verified by DNA sequencing.
SDS-PAGE and Immunoblot Analysis
The W303 (MAT� ade2-1 his3-11 his3-15 trp1-1 ura3-1 leu2-3 leu2-112) yeast strain was used for the expression of heterologous proteins.Protein was extracted from yeast cells as described (Yaffe and Schatz,1984). A zein extract from maize endosperm (Wallace et al., 1990) wasused as a positive control in immunoblots. Immunoblotting was per-formed according to Sambrook et al. (1989). �-Zein, �-zein, and �-zeinprimary antisera were used at a dilution of 1:5000 (Lending and Larkins,1989). Goat anti-rabbit alkaline phosphatase–conjugated secondaryantibodies (Sigma, St. Louis, MO) were used at a dilution of 1:30,000. Amouse monoclonal anti-GFP antibody was obtained from Zymed Lab-oratories (South San Francisco, CA) and used at a dilution of 1:5000.
Confocal Laser Scanning Microscopy of Yeast Cells Expressing GFP and Zein::GFP Fusion Proteins
Yeast cells were transformed with GFP or zein::GFP fusion genes andcultured in liquid selection medium (Clonetech, Pablo Alto, CA) to anOD600 of 1.0. One milliliter of culture was centrifuged, and the cells wereresuspended in 50 �L of PBS, pH 7.0. Approximately 3 to 5 �L of thecells was placed on a glass microscope slide and examined with aMRC-1024 laser scanning confocal microscope (Bio-Rad, Hercules, CA)at �1000 magnification with a fluorescein isothiocyanate excitation andemission filter set. Usually, clusters of two to six cells with a relativelyhigh level of green fluorescence were selected for imaging. The cellswere scanned optically at 1-�m increments to produce four to six image
sections that were stacked into a composite. All images were treated inthe same manner to adjust for brightness and contrast with the AdobePhotoshop 5.0 software program (Adobe Systems, San Jose, CA).
Immunolocalization of Zein::GFP Fusion Proteins in Yeast Cells
Transformed yeast cells were fixed for 1 hr in 3.7% paraformalde-hyde, 0.1% glutaraldehyde, and 0.2% picric acid prepared in 50 mMpotassium phosphate and 5 mM EGTA buffer, pH 6.8. Further fixa-tion in 0.1% osmium tetroxide for 1 hr was followed by dehydrationin graded ethanol concentrations. The cells were infiltrated with LRWhite resin (London Resin Company Ltd., Ft. Washington, PA) andcured under UV light at 10 C for 2 days. Plastic tissue blocks werecut into 80- to 120-nm-thick sections and collected on formvar-coated nickel grids. The grids were incubated for 30 min in blockingsolution (pH 8.2) containing 0.2% BSA, 0.06% Tween 20, 20 mMTris-HCl, and 500 mM NaCl. They were labeled subsequently withprimary antibodies overnight at 4 C, whereas labeling with second-ary antibodies conjugated with gold particles was performed for 1 hrat room temperature. For immunolabeling of yeast cells producingtwo different zeins, the sections were incubated in primary anti-body solution containing monoclonal GFP antibody (Zymed Labo-ratories) to detect the zein-GFP fusion and rabbit antibody againstthe other type of zein protein. The primary antibodies were detectedwith anti-mouse antibodies or anti-rabbit antibodies conjugated with10- and 5-nm gold particles, respectively. The grids were stainedwith 2.5% uranyl acetate for 15 min followed by three rinses in dou-ble-distilled water. The grids were examined with a JEOL 100CX2transmission electron microscope at 80 kV.
Analysis of Zein Interactions in Yeast Two-Hybrid Vectors
Two-hybrid constructs were transformed into yeast strain Y190[MATa gal4 gal80 his3 trp1-901 ade2-101 ura3-52 leu2-3 leu2-112 �URA3::GAL��lacZ LYS2::GAL(UAS)��HIS3 cycr] by the lithiummethod (Schiestl and Gietz, 1989), and cells were selected on –Leumedium (pACT2) or –Trp medium (pAS2). A subclone of each type ofzein cDNA (Woo et al., 2001) was constructed without its signal pep-tide (�sp) by PCR amplification using primers containing SmaI andBamHI restriction endonuclease sites (Table 1, sequences in bold-face). The coding sequences were inserted into the pACT2 and pAS2vectors of the yeast two-hybrid system.
Each of the pACT2 constructs was introduced into zein-express-ing strains, which were produced from pAS2 transformations, andvice versa; the double transformants were selected on –Trp/Leu me-dium. Colonies were transferred from –Trp/Leu plates to –Trp/Leuplates lacking His and containing 35 mM 3-aminotriazole. The samecolonies were replated on –Trp/Leu medium and used for X-Gal filterassays according to the procedure of Bai and Elledge (1996). To mea-sure the strength of the yeast two-hybrid interaction between selectedzein constructs, a liquid �-galactosidase assay, using o-nitrophenyl�-D-galactopyranoside (ONPG) as substrate, was performed as de-scribed by the manufacturer (Clontech, Palo Alto, CA).
Construction of 22��sp-Zein Deletion Mutants for the Analysis of Yeast Two-Hybrid Interactions
The 22��sp-zein constructs listed in Table 2 were inserted betweenthe SmaI and BamHI sites (shown in boldface) in the pACT2 vector,

16 of 18 The Plant Cell
Table 2. Clones and Primers for 22��sp-Zein Deletion Mutants
Clone Primer
b 5�-TCCCCCGGGGATGCTACAACAAGCGATTGCGGCGA-3� and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
c 5�-TCCCCCGGGGATGGCACTGAGCCACCTAGCCATGG-3� and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
d 5�-TCCCCCGGGGATGTTTCTGCCAGCGCTTAGTCAAC-3� and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
e 5�-TCCCCCGGGGATGTTGTTGCAACAGATTGTACCAG-3� and5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
f 5�-TCCCCCGGGGATGTTGCAACAGCTGCTTCCATTCA-3� and5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
g 5�-TCCCCCGGGGATGCAGCGACAACAGTTACTTAATC-3� and5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
h 5�-TCCCCCGGGGATGCAGCAGCAACAATTGCTGCCAT-3� and5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
i 5�-TCCCCCGGGGATGGTGAGCAAGGGC-3� and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
j 5�-TCCCCCGGGGATCTCCATTATTCCACAATGCTCACTTGCTCCT-3�, 5�-CAATTGTTGCTGCTGCCTATAGGCTTGCAC-3�, 5�-GTGCAAGCCTATAGGCAGCAGCAACAATTG-3�, and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
k 5�-TCCCCCGGGGATGGCACTGAGCCACCTAGCCATGG-3�, 5�-AGCTGGTACAATCTGTTGCAACAACCTATAGGCTTGCACAGC -3�,5�-GCTGTGCAAGCCTATAGGTTGTTGCAACAGATTGTACCAGCT-3�, and5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
l 5�-TCCCCCGGGGATGTTGTTGCAACAGATTGTACCAG-3�,5�-CTCGCCCTTGCTCACCATTAGGAAGGCAGCGACCAATGG-3�, 5�-CCATTGGTCGCTGCCTTCCTAATGGTGAGCAAGGGCGAG-3�, and5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
m 5�-TCCCCCGGGGATGTTGCAACAGCTGCTTCCATTCA-3�, 5�-CTCGCCCTTGCTCACCATGACAGGATTCATCAAAGAGAA-3�,5�-TTCTCTTTGATGAATCCTGTCATGGTGAGCAAGGGCGAG-3�, and5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
n 5�-TCCCCCGGGGATCTCCATTATTCCACAATGCTCACTTGCTC CT-3�, 5�-CTCGCCCTTGCTCACCATTTCTTGTTGCAGGTATGT-3�,5�-ACATACCTGCAACAAGAAATGGTGAGCAAGGGCGAG-3�, and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
o 5�-TCCCCCGGGGATCTCCATTATTCCACAATGCTCACTTGCTCCT-3�, 5�-CTCGCCCTTGCTCACCATTGGTAGGAACTGTTGTTGCTG-3�, 5�-CAGCAACAACAGTTCCTACCAATGGTGAGCAAGGGCGAG-3�, and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
p 5�-TCCCCCGGGGATGCTACAACAAGCGATTGCGGCGA-3�, 5�-CTCGCCCTTGCTCACCATTGGTAGGAACTGTTGTTGCTG-3�, 5�-CAGCAACAACAGTTCCTACCAATGGTGAGCAAGGGCGAG-3�, and5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
q 5�-TCCCCCGGGGATCTCCATTATTCCACAATGCTCACTTGCTCCT-3�, 5�-GAATGGAAGCAGCTGTTGCAATGGTAGGAACTGTTGTTGCTG-3�, 5�-CAGCAACAACAGTTCCTACCATTGCAACAGCTGCTTCCATTC-3�, and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�
r 5�-TCCCCCGGGGATCTCCATTATTCCACAATGCTCACTTGCTCCT-3�,5�-CTCCTCGCCCTTGCTCACCATTGGTAGGAACTGTTGTTGCTG-3�, 5�-CAGCAACAACAGTTCCTACCAATGGTGAGCAAGGGCGAGGAG-3�, 5�-TGAATGGAAGCAGCTGTTGCAACTTGTACAGCTCGTCCATGCC-3�,5�-GGCATGGACGAGCTGTACAAGTTGCAACAGCTGCTTCCATTCA-3�, and 5�-CGCGGATCCGCGCTAGACAGGATTCATCAAAGAGAA-3�

Zein Interactions in Protein Bodies 17 of 18
and the clones obtained were verified by DNA sequence analysis.Deletion mutants of the 22��sp-zein were constructed with 3� GFPfusions to facilitate their subsequent analysis in yeast cells. We foundno evidence that GFP interfered with the interaction between zeinproteins. A superglow GFP variant (Sheen et al., 1995) was insertedbetween the SmaI and BamHI sites of pACT2 (Figure 8A, construct i)using oligonucleotide primers 5�-TCCCCCGGGGATGGTGAGCAA-GGGC-3� and 5�-GTCGACGGATCCTTACTTGTACAGCTCGTC-3�.The 22��sp-zein::GFP fusion construct (Figure 8A, construct a) wasmade as follows. The 22�sp gene fragment was amplified with prim-ers 5�-TCCCCCGGGGATCTCCATTATTCCACAATGCTCACTTGCTC-CT-3� and 5�-GCCCTTGCTCACCATAAAGATGGCACCTCC-3�, andGFP sequences were amplified with primers 5�-GGAGGTGCC-ATCTTTATGGTGAGCAAGGGC-3� and 5�-GTCGACGGATCCTTA-CTTGTACAGCTCGTC-3�. The two PCR products were annealed andreamplified with the following primers: 5�-TCCCCCGGGGATCTC-CATTATTCCACAATGCTCACTTGCTCCT-3� and 5�-GTCGACGGA-TCCTTACTTGTACAGCTCGTC-3�; they were cloned into pACT2 af-ter restriction enzyme digestion with SmaI and BamHI.
Hydropathy Analysis of Zein Amino Acid Sequences
Hydropathy characteristics of the 22-kD �-zein, the 15-kD �-zein, the16-kD �-zein, and the 10-kD �-zein were determined by the method ofKyte and Doolittle (1982) (http://searchlauncher.bcm.tmc.edu).
ACKNOWLEDGMENTS
We are grateful to the colleagues who made many useful sugges-tions for the improvement of the manuscript. This research wassupported by a grant from the Department of Energy (No. DE-FG03-95ER20183) to B.A.L.
Received October 3, 2001; accepted December 13, 2001.
REFERENCES
Abe, S., You, W., and Davies, E. (1991). Protein bodies in cornendosperm are enclosed by and enmeshed in F-actin. Proto-plasma 165, 139–149.
Argos, P., Pedersen, K., Marks, M.D., and Larkins, B.A. (1982). Astructural model for maize zein proteins. J. Biol. Chem. 257,9984–9990.
Bagga, S., Adams, H.P., Rodriguez, F.D., Kemp, J.D., andSengupta-Gopalan, C. (1997). Coexpression of the maize delta-zein and beta-zein genes results in stable accumulation of delta-zein in endoplasmic reticulum-derived protein bodies formed bybeta-zein. Plant Cell 9, 1683–1696.
Bai, C., and Elledge, S.J. (1996). Gene identification using the yeasttwo-hybrid system. Methods Enzymol. 273, 331–347.
Bar-Peled, M., Bassham, D.C., and Raikhel, N.V. (1996). Transportof proteins in eukaryotic cells: More questions ahead. Plant Mol.Biol. 32, 223–249.
Bitter, G.A., and Egan, K.M. (1984). Expression of heterologous
genes in Saccharomyces cerevisiae from vectors utilizing the glyc-eraldehydes-3-phosphate dehydrogenase gene promoter. Gene 32,263–274.
Bostwick, D.E., Dannenhoffer, J.M., Skaggs, M.I., Lister, R.M.,Larkins, B.A., and Thompson, G.A. (1992). Pumpkin phloem lec-tin genes are specifically expressed in companion cells. Plant Cell4, 1539–1548.
Choi, S.B., Wang, C.L., Muench, D.G., Ozawa, K., Franceschi,V.R., Wu, Y.J., and Okita, T.W. (2000). Messenger RNA targetingof rice seed storage proteins to specific ER subdomains. Nature407, 765–767.
Chui, C.F., and Falco, S.C. (1995). A new methionine-rich seedstorage protein from maize. Plant Physiol. 107, 291.
Clore, A.M., Dannenhoffer, J.M., and Larkins, B.A. (1996). EF-1�
is associated with a cytoskeletal network surrounding proteinbodies in maize endosperm cells. Plant Cell 8, 2003–2014.
Coleman, C.E., Herman, E.M., Takasaki, K., and Larkins, B.A.(1996). The maize gamma-zein sequesters alpha-zein and stabi-lizes its accumulation in protein bodies of transgenic tobaccoendosperm. Plant Cell 8, 2335–2345.
Coleman, C.E., Clore, A.C., Ranch, J.P., Higgins, R., Lopes, M.A.,and Larkins, B.A. (1997). Expression of a mutant �-zein createsthe floury2 phenotype in transgenic maize. Proc. Natl. Acad. Sci.USA 94, 7094–7097.
Coraggio, I., Martegani, E., Compagno, C., Porro, D., Alberghina,L., Bernard, L., Faoro, F., and Viotti, A. (1988). Differential tar-geting and accumulation of normal and modified zein polypep-tides in transformed yeast. Eur. J. Cell Biol. 47, 165–172.
Esen, A., and Stetler, D.A. (1992). Immunocytochemical localizationof �-zein in the protein bodies of maize endosperm cells. Am. J.Bot. 79, 243–248.
Fontes, E.B.P., Shank, B.B., Wrobel, R.L., Moose, S.P., Obrian,G.R., Wurtzel, E.T., and Boston, R.S. (1991). Characterization ofan immunoglobulin binding protein homolog in the maize floury2endosperm mutant. Plant Cell 3, 483–496.
Garratt, R., Oliva, G., Caracelli, I., Leite, A., and Arruda, P. (1993).Studies of the zein-like alpha-prolamins based on an analysis ofamino acid sequences: Implications for their evolution and three-dimensional structure. Proteins 15, 88–99.
Geli, M.I., Torrent, M., and Ludevid, D. (1994). Two structuraldomains mediate two sequential events in �-zein targeting: Pro-tein endoplasmic reticulum retention and protein body formation.Plant Cell 6, 1911–1922.
Guarente, L., Lalonde, B., Gifford, P., and Alani, E. (1984). Distinctlyregulated tandem upstream activation sites mediate cataboliterepression of the CYC1 gene of S. cerevisiae. Cell 36, 503–511.
Keegstra, K., and Cline, K. (1999). Protein import and routing sys-tems of chloroplasts. Plant Cell 11, 557–570.
Kirihara, J.A., Petri, J.B., and Messing, J. (1988). Isolation andsequence of a gene encoding a methionine-rich 10-kDa zein pro-tein from maize. Gene 71, 359–370.
Kyte, J., and Doolittle, R.F. (1982). A simple method for displayingthe hydrophobic nature of a protein. J. Mol. Biol. 157, 105–132.
Larkins, B.A., Wallace, J.C., Galili, G., Lending, C.R., andKawata, E.E. (1989). Structural analysis and modification ofmaize storage proteins. J. Ind. Microbiol. 30, 203–209.

18 of 18 The Plant Cell
Lee, D.H. (1998). Characterization of 27K zein as a transmembraneprotein. J. Biochem. Mol. Biol. 31, 196–200.
Lending, C.R., and Larkins, B.A. (1989). Changes in the zein com-position of protein bodies during maize endosperm development.Plant Cell 1, 1011–1023.
Lending, C.R., Kriz, A.R., Larkins, B.A., and Bracker, C.E. (1988).Structure of maize protein bodies and immunocytochemical local-ization of zeins. Protoplasma 143, 51–62.
Li, C.P., and Larkins, B.A. (1996). Expression of protein disulfideisomerase is elevated in the endosperm of the maize floury2mutant. Plant Mol. Biol. 30, 873–882.
Li, X., Franceschi, V.R., and Okita, T.W. (1993a). Segregation ofstorage protein mRNAs on the rough endoplasmic reticulummembranes of rice endosperm cells. Cell 72, 869–879.
Li, X.X., Wu, Y.J., Zhang, D.Z., Gillikin, J.W., Boston, R.S.,Franceschi, V.R., and Okita, T.W. (1993b). Rice prolamine pro-tein body biogenesis: A Bip-mediated process. Science 262,1054–1056.
Lopes, M.A., and Larkins, B.A. (1993). Endosperm origin, develop-ment, and function. Plant Cell 5, 1383–1399.
Marks, M.D., Lindell, J.S., and Larkins, B.A. (1985). Nucleotidesequence analysis of zein mRNAs from maize endosperm. J. Biol.Chem. 260 16451–16459.
McFadden, G. (1991). Electron Microscopy of Plant Cells. (SanDiego, CA: Academic Press).
Mumberg, D., Muller, R., and Funk, M. (1995). Yeast vectors forthe controlled expression of heterologous proteins in differentgenetic backgrounds. Gene 156, 119–122.
Pedersen, K., Argos, P., Naravana, S.V., and Larkins, B.A. (1986).Sequence analysis and characterization of a maize gene encodinga high-sulfur zein protein of Mr 15,000. J. Biol. Chem. 261, 6279–6284.
Pomeroy, M.E., Lawrence, J.B., Singer, R.H., and Billings-Gagliardi, S. (1991). Distribution of myosin heavy chain mRNA inembryonic muscle tissue visualized by ultrastructural in situhybridization. Dev. Biol. 143, 58–67.
Prat, S., Cortadas, J., Puigdomenech, P., and Palau, J. (1985).Nucleic acid (cDNA) and amino acid sequences of the maizeendosperm protein glutelin-2. Nucleic Acids Res. 13, 1493–1504.
Prat, S., Perez-Grau, L., and Puigdomenech, P. (1987). Multiplevariability in the sequence of a family of maize endosperm pro-teins. Gene 52, 41–49.
Rabanal, R., Ludevid, D., Pons, M., and Giralt, E. (1993). CD ofproline rich polypeptides: Application to the study of the repetitivedomain of maize glutelin-2. Biopolymers 33, 1019–1028.
Rings, E.H., Buller, H.A., Neele, A.M., and Dekker, J. (1994). Pro-tein sorting versus messenger RNA sorting? Eur. J. Cell Biol. 63,161–171.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). MolecularCloning: A Laboratory Manual, 2nd ed. (Cold Spring Harbor, NY:Cold Spring Harbor Laboratory Press).
Schiestl, R.H., and Gietz, R.D. (1989). High efficiency transforma-tion of intact yeast cells using single stranded nucleic acids as acarrier. Curr. Genet. 16, 339–346.
Shaywitz, D.A., Espenshade, P.J., Gimeno, R.E., and Kaiser, C.A.(1997). COPII subunit interactions in the assembly of the vesiclecoat. J. Biol. Chem. 272, 25413–25416.
Sheen, J., Hwang, S., Niwa, Y., Kobayashi, H., and Galbraith,D.W. (1995). Green-fluorescent protein as a new vital marker inplant cells. Plant J. 8, 777–784.
Singer, R.H., Langevin, G.L., and Lawrence, J.B. (1989). Ultra-structural visualization of cytoskeletal mRNAs and their associ-ated proteins using double-label in situ hybridization. J. Cell Biol.108, 2343–2353.
Stankovic, B., Abe, S., and Davies, E. (1993). Co-localization ofpolysomes, cytoskeleton, and membranes with protein bodiesfrom corn endosperm. Protoplasma 177, 66–72.
St. Johnston, D. (1995). The intracellular localization of messengerRNAs. Cell 81, 161–170.
Takatsu, H., Katoh, Y., Shiba, Y., and Nakayama, K. (2001). Golgi-localizing, �-adaptin ear homology domain, ADP-ribosylation factor-binding (GGA) proteins interact with acidic dileucine sequenceswithin the cytoplasmic domains of sorting receptors through theirVps27p/Hrs/STAM (VHS) domains. J. Biol. Chem. 276, 28541–28545.
Vitale, A., and Denecke, J. (1999). The endoplasmic reticulum:Gateway of the secretory pathway. Plant Cell 11, 615–628.
Wallace, J.C., Lopes, M.A., Paiva, E., and Larkins, B.A. (1990).New methods for extraction and quantitation of zeins reveal ahigh content of �-zein in modified opaque-2 maize. Plant Physiol.92, 191–196.
Woo, Y.-M., Hu, D.W.-N., Larkins, B.A., and Jung, R. (2001).Genomics analysis of genes expressed in maize endosperm iden-tifies novel storage proteins and clarifies patterns of zein geneexpression. Plant Cell 13, 2297–2317.
Yaffe, M.P., and Schatz, G. (1984). Two nuclear mutations thatblock mitochondrial protein import in yeast. Proc. Natl. Acad. Sci.USA 81, 4819–4823.
Zhang, F., and Boston, R.S. (1992). Increases in binding protein(BiP) accompany changes in protein body morphology in threehigh-lysine mutants in maize. Protoplasma 171, 142–152.
Copyright © 2022 FDOKUMEN




















