
Yeast centrin Cdc31 is linked to the nuclear mRNA export machinery
10
ARTICLES 840 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004 Yeast centrin Cdc31 is linked to the nuclear mRNA export machinery Tamás Fischer 1 , Susana Rodríguez-Navarro 1 , Gislene Pereira 2,3 , Attila Rácz 1 , Elmar Schiebel 2 and Ed Hurt 1,4 Centrins are calmodulin-like proteins that function in the duplication of microtubule-organizing centres. Here we describe a new function of the yeast centrin Cdc31. We show that overproduction of a sequence, termed CID, in the carboxy-terminal domain of the nuclear export factor Sac3 titrates Cdc31, causing a dominant-lethal phenotype and a block in spindle pole body (SPB) duplication. Under normal conditions, the CID motif recruits Cdc31 and Sus1 (a subunit of the SAGA transcription complex) to the Sac3–Thp1 complex, which functions in mRNA export together with specific nucleoporins at the nuclear basket. A previously reported cdc31 temperature-sensitive allele, which is neither defective in SPB duplication nor Kic1 kinase activation, induces mRNA export defects. Thus, Cdc31 has an unexpected link to the mRNA export machinery. 1 Biochemie-Zentrum der Universität Heidelberg (BZH), Im Neuenheimer Feld 328, D-69120 Heidelberg, Germany. 2 Paterson Institute for Cancer Research, Christie Hospital NHS Trust, Manchester, UK. 3 Present address: School of Biological Sciences, University of Manchester, Oxford Road, Manchester M13 9PT, UK. 4 Correspondence should be addressed to E.H. (e-mail: [email protected]) Published online: 15 August 2004, DOI: 10.1038/ncb1163 Gene expression in eukaryotic cells requires bidirectional transport of molecules and macromolecules across the nuclear membrane. The exclusive transport machines mediating nucleocytoplasmic transport are the nuclear pore complexes (NPCs) in the nuclear membrane, which form the stationary phase, and the soluble and shuttling trans- port receptors, which cooperate with the FG-repeat-containing nucleo- porins during their passage through the pore channel 1 . Among the key molecules that must be exported to the cytoplasm are mRNAs, which are transcribed as pre-mRNAs by the RNA polymerase II machinery in the nucleus, before RNA processing and modification, ribonucleopro- tein particle (RNP) assembly and nuclear export. Biogenesis and export of mRNPs requires many factors that transiently interact with the evolving mRNPs 2 . An important feature of mRNA export that has emerged from recent studies is that the factors and mechanism of mRNA export are conserved in eukaryotic cells 3–6 . Among these con- served factors are the mRNA export receptor Mex67–Mtr2 in yeast and Tap–p15 in metazoans, and several components (for example,Yra1 and Sub2, and their respective metazoan counterparts, Aly and UAP56) that function upstream of the mRNA export receptor and couple intranu- clear steps in mRNP biogenesis with nuclear mRNA export 7–10 . Studies in yeast and mammals show that the coupling proteins Yra1 and Sub2 are part of a multi-subunit complex, the TREX complex 11 , which also contains components involved in transcription elongation (THO-complex members) 12 . As TREX members are co-transcription- ally recruited to active genes during transcription elongation 11,13–15 , it has been suggested that the TREX complex couples transcription elon- gation with mRNP assembly and nuclear export 3,4,6,11,16 . Recently, additional factors of the mRNA export machinery, Sac3 and Thp1 (refs 17–19), as well as Sus1 (ref. 20), were identified. Sac3 was dis- covered because its suppresses an actin mutant 21 , but it is also involved in cell-cycle progression 22 . Importantly, Sac3 interacts with Mex67 and nucleoporins of the nuclear basket (Nup60 and Nup1), suggesting a specific role in mRNA export at the nucleoplasmic side of the NPC 17 . In addition, Sac3 is reported to function on the cytoplasmic side of the nuclear pore complex 19 . Furthermore, Sac3 forms a complex with Thp1 (refs 17, 18) and Sus1 (ref. 20) in vivo. Both Thp1 and Sus1 have roles in transcription, and Sus1 is part of the SAGA (Spt/Ada/Gcn5 acetyltrans- ferase) histone acetyltransferase complex, which is involved in tran- scription initiation of a subset of genes 20,23 . Thus, the Sac3–Thp1–Sus1 complex could connect transcription and nuclear mRNA export 20 . Here, we identify Cdc31 as a new factor associated with the Sac3–Thp1–Sus1 complex. The yeast centrin Cdc31 is a member of the calmodulin superfamily that has a crucial function in SPB and centro- some duplication 24,25 . Thus, Cdc31 has an additional role in a nuclear- pore-associated complex that is involved in nuclear mRNA export. RESULTS Overproduction of the CID motif in Sac3 is dominant-lethal The mRNA export factor Sac3 is composed of several domains: the N- and M domains, which bind to Mex67–Mtr2 and Thp1, and a C-domain that mediates nuclear pore targeting 17 . Notably, overexpres- sion of the Sac3 C-domain is toxic and induces strong mRNA export defects 17 . Deletions within the Sac3 C-domain were created to assess which motif was responsible for the dominant-negative phenotype. Overexpressed Sac3 563–1301 and Sac3 733–1301 fragments still functioned as dominant-negatives, but the Sac3 851–1301 fragment, however, was not toxic (Fig. 1a). Thus, residues between 733 and 851 within Sac3 could induce the lethal phenotype. Indeed, overexpression of the Sac3 733–860 fragment was toxic to cells (Fig. 1a). We termed the Sac3 sequence between residue 733 and 860 the ‘CID’ motif (see below). Notably, the toxic effects of overexpressing Sac3 were not strictly coupled to the mRNA export defects. Whereas Sac3 563–1301 and ©2004 Nature Publishing Group
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Yeast centrin Cdc31 is linked to the nuclear mRNA export machinery

A RT I C L E S
840 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004
Yeast centrin Cdc31 is linked to the nuclear mRNAexport machineryTamás Fischer1, Susana Rodríguez-Navarro1, Gislene Pereira2,3, Attila Rácz1, Elmar Schiebel2 and Ed Hurt1,4
Centrins are calmodulin-like proteins that function in the duplication of microtubule-organizing centres. Here we describe a newfunction of the yeast centrin Cdc31. We show that overproduction of a sequence, termed CID, in the carboxy-terminal domain ofthe nuclear export factor Sac3 titrates Cdc31, causing a dominant-lethal phenotype and a block in spindle pole body (SPB)duplication. Under normal conditions, the CID motif recruits Cdc31 and Sus1 (a subunit of the SAGA transcription complex) tothe Sac3–Thp1 complex, which functions in mRNA export together with specific nucleoporins at the nuclear basket. A previouslyreported cdc31 temperature-sensitive allele, which is neither defective in SPB duplication nor Kic1 kinase activation, inducesmRNA export defects. Thus, Cdc31 has an unexpected link to the mRNA export machinery.
1Biochemie-Zentrum der Universität Heidelberg (BZH), Im Neuenheimer Feld 328, D-69120 Heidelberg, Germany. 2Paterson Institute for Cancer Research, ChristieHospital NHS Trust, Manchester, UK. 3Present address: School of Biological Sciences, University of Manchester, Oxford Road, Manchester M13 9PT, UK.4Correspondence should be addressed to E.H. (e-mail: [email protected])
Published online: 15 August 2004, DOI: 10.1038/ncb1163
Gene expression in eukaryotic cells requires bidirectional transport ofmolecules and macromolecules across the nuclear membrane. Theexclusive transport machines mediating nucleocytoplasmic transportare the nuclear pore complexes (NPCs) in the nuclear membrane,which form the stationary phase, and the soluble and shuttling trans-port receptors, which cooperate with the FG-repeat-containing nucleo-porins during their passage through the pore channel1. Among the keymolecules that must be exported to the cytoplasm are mRNAs, whichare transcribed as pre-mRNAs by the RNA polymerase II machinery inthe nucleus, before RNA processing and modification, ribonucleopro-tein particle (RNP) assembly and nuclear export. Biogenesis and exportof mRNPs requires many factors that transiently interact with theevolving mRNPs2. An important feature of mRNA export that hasemerged from recent studies is that the factors and mechanism ofmRNA export are conserved in eukaryotic cells3–6. Among these con-served factors are the mRNA export receptor Mex67–Mtr2 in yeast andTap–p15 in metazoans, and several components (for example, Yra1 andSub2, and their respective metazoan counterparts, Aly and UAP56) thatfunction upstream of the mRNA export receptor and couple intranu-clear steps in mRNP biogenesis with nuclear mRNA export7–10.
Studies in yeast and mammals show that the coupling proteins Yra1and Sub2 are part of a multi-subunit complex, the TREX complex11,which also contains components involved in transcription elongation(THO-complex members)12. As TREX members are co-transcription-ally recruited to active genes during transcription elongation11,13–15, ithas been suggested that the TREX complex couples transcription elon-gation with mRNP assembly and nuclear export3,4,6,11,16.
Recently, additional factors of the mRNA export machinery, Sac3 andThp1 (refs 17–19), as well as Sus1 (ref. 20), were identified. Sac3 was dis-covered because its suppresses an actin mutant21, but it is also involvedin cell-cycle progression22. Importantly, Sac3 interacts with Mex67 and
nucleoporins of the nuclear basket (Nup60 and Nup1), suggesting aspecific role in mRNA export at the nucleoplasmic side of the NPC17. Inaddition, Sac3 is reported to function on the cytoplasmic side of thenuclear pore complex19. Furthermore, Sac3 forms a complex with Thp1(refs 17, 18) and Sus1 (ref. 20) in vivo. Both Thp1 and Sus1 have roles intranscription, and Sus1 is part of the SAGA (Spt/Ada/Gcn5 acetyltrans-ferase) histone acetyltransferase complex, which is involved in tran-scription initiation of a subset of genes20,23. Thus, the Sac3–Thp1–Sus1complex could connect transcription and nuclear mRNA export20.
Here, we identify Cdc31 as a new factor associated with theSac3–Thp1–Sus1 complex. The yeast centrin Cdc31 is a member of thecalmodulin superfamily that has a crucial function in SPB and centro-some duplication24,25. Thus, Cdc31 has an additional role in a nuclear-pore-associated complex that is involved in nuclear mRNA export.
RESULTSOverproduction of the CID motif in Sac3 is dominant-lethal The mRNA export factor Sac3 is composed of several domains: the N-and M domains, which bind to Mex67–Mtr2 and Thp1, and a C-domain that mediates nuclear pore targeting17. Notably, overexpres-sion of the Sac3 C-domain is toxic and induces strong mRNA exportdefects17. Deletions within the Sac3 C-domain were created to assesswhich motif was responsible for the dominant-negative phenotype.Overexpressed Sac3563–1301 and Sac3733–1301 fragments still functionedas dominant-negatives, but the Sac3851–1301 fragment, however, wasnot toxic (Fig. 1a). Thus, residues between 733 and 851 within Sac3could induce the lethal phenotype. Indeed, overexpression of theSac3733–860 fragment was toxic to cells (Fig. 1a). We termed the Sac3sequence between residue 733 and 860 the ‘CID’ motif (see below).
Notably, the toxic effects of overexpressing Sac3 were not strictlycoupled to the mRNA export defects. Whereas Sac3563–1301 and
print ncb1163 13/8/04 4:33 PM Page 840
© 2004 Nature Publishing Group

A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004 841
Sac3733–1301 fragments induced both a lethal phenotype and nuclearmRNA accumulation, the SAC3CID construct was toxic for cells butfailed to inhibit mRNA export (Fig. 1b).
Moreover, we determined which sequences in the Sac3 C-domainmediate nuclear envelope targeting. We fused green fluorescent proteinto the Sac3 deletions and examined their localization. As with wild-typeSac3, the GFP-tagged Sac3563–1301 and Sac3733–1301 fragments localizedto the nuclear periphery, but the non-toxic Sac3851–1301 had a predomi-nantly intranuclear GFP fluorescence pattern with weak fluorescenceobserved in the nuclear envelope (Fig. 1c). Conversely, the toxic Sac3CID
fragment was concentrated at the nuclear periphery with somedetectable within the nucleus (Fig. 1c). Thus, in addition to the 130-residue-long CID being a major determinant for the NPC localization ofSac3, the C-domain also has other motifs (for example, the one presentin Sac3851–1301) that contribute to efficient NPC association (see below).
Cdc31 binds to overexpressed Sac3CID
The toxicity of overexpressed Sac3CID may be the result of titration andsequestration of an essential protein in yeast. To identify this factor, weaffinity purified the various tandem affinity purification (TAP)-taggedSac3 C-domain fragments and searched for associated proteins. Weapplied only the first affinity purification step of the TAP method (TEVelution from IgG–Sepharose); thus, the TEV-eluates were contami-nated by common impurities — for example, heat shock proteins, fattyacid synthetase, translation factors, ribosomal proteins and more —that often stick to IgG–Sepharose. However, as shown by SDS–PAGEand Coomassie staining, two bands with approximate relative molecu-lar masses (Mrs) of 16,000 and 9,000 were significantly co-enrichedwith the toxic Sac3 constructs, including Sac3CID, but were absent fromthe purification of the non-toxic Sac3851–1301 fragment (Fig. 2a).MALDI–TOF mass spectrometry analysis identified the yeast SPBcomponent Cdc31 as the 16K band, and the Sac3 complex component
Sus1 as the 9K band. Western blot analysis with anti-CBP antibodiesdetected the various TAP-tagged Sac3 bait proteins (Fig. 2b), and anti-bodies against Cdc31 and myc-tagged Sus1 confirmed that these pro-teins were only associated with the toxic SAC3 C-fragments (Fig. 2b).
To determine whether Sac3CID binds to Cdc31 independently of Sus1,we overexpressed and affinity purified Sac3733–1301 from yeast cells thatcarry a disrupted sus1 gene. In this strain background, Sac3733–1301 wasstill toxic (data not shown), and affinity purified Sac3733–1301 efficientlyrecruited Cdc31 (Fig. 1c). This data suggests that Sac3CID binds toCdc31 independently of Sus1. Hence, we termed this 130-amino-acid-long motif in the Sac3 C-domain ‘CID’, for Cdc31-interacting domain.
Overexpression of Sac3CID arrests yeast as large budded cellswith a seemingly single SPBOverexpressed Sac3CID is toxic without inhibiting mRNA export.Sac3CID toxicity may be caused by Cdc31 inhibition, because overex-pressed Sac3CID binds to the SPB component Cdc31, which is essentialfor SPB duplication24,26; this should inhibit SPB duplication andinduce an arrest in cell-cycle progression. To visualize the SPB in livingcells, the SPB marker Spc42–GFP27
, together with overexpressedSac3CID constructs, was expressed in α-factor-synchronized cells thatwere released into a new cell cycle. Overexpression of Sac3 fragmentscontaining the CID motif arrested yeasts as large budded cells with aseemingly single SPB (Fig. 3a). In contrast, overproduction of the non-toxic Sac3851–1301 did not cause such a cell-cycle phenotype.
If our assumption that overexpression of Sac3CID interferes withCdc31 function during the SPB duplication is true, then overproductionof Cdc31 should compensate for this toxicity. Notably, high copy expres-sion of CDC31 from a 2µ plasmid significantly diminished the domi-nant-negative phenotype exerted by the various overexpressedCID-containing Sac3-C fragments (Fig. 3b). Thus, overproduction ofSac3CID interferes with SPB duplication by titrating out Cdc31.
a b cSac3
N M C
GAL
GAL::SAC3563−1301
GAL::SAC3733−1301
GAL::SAC3851−1301
GAL::SAC3CID(733−860)
mRNA DNA
GFP Nomarski
Figure 1 Overexpression of the Sac3 CID motif produces a dominant-negative phenotype. (a) Schematic representation of the Sac3 domains andC-domain deletion constructs, which were under the control of the GAL1promoter and expressed in wild-type cells. Cells were spotted in 10−1
dilutions on galactose-containing media, and growth was analysed afterincubation for three days at 30 °C. (b) inhibition of mRNA export in cells
that express the indicated overexpressed Sac3 C-domain deletionconstructs. Poly(A)+ RNA localization was determined by in situhybridization 5 h after induction in galactose-containing medium. DNA wasstained with DAPI. (c) Intracellular localization of the indicated GFP-taggedSac3 C-domain deletion constructs in sac3∆ cells as determined byfluorescence microscopy analysis.
print ncb1163 13/8/04 4:33 PM Page 841
© 2004 Nature Publishing Group

A RT I C L E S
842 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004
Cdc31 is a genuine member of the Sac3–Thp1–Sus1 complexPreviously, Cdc31 was found in TAP purifications of Thp1 and Sus1(refs 18, 20). It was suggested that calmodulin-like Cdc31 may be a con-taminant, because it binds to the CBP-tag, which constitutes a calmod-ulin-binding peptide18. However, Cdc31 does not co-purify withTAP-tagged bait proteins in general (E.H., unpublished observations),and the data from this study suggest a specific association of Cdc31 withthe Sac3CID motif (see above). To obtain definitive evidence that Cdc31is a novel subunit of the Sac3–Thp1–Sus1 complex, we affinity purifiedthe Sac3 complex through a novel ‘split double-tag method’ (seeMethods section). In this approach, a yeast strain was generated that co-expressed chromosomally integrated TAP-tagged Sus1 and glutathione
S-transferase (GST)-tagged Thp1. In the first step, Sus1–TAP was affin-ity purified by IgG–Sepharose before TEV elution. In the second step,the TEV eluate was passed over a glutathione (GSH)–Sepharose columnto enrich for Thp1–GST. After elution with GSH, the final eluate wasanalysed by SDS–PAGE and Coomassie staining (Fig. 4a). Using thisnew purification scheme, the Sac3–Thp1–Sus1 complex was efficientlypurified, and Cdc31 (identified by mass spectrometry analysis) was theprominent band seen above the Sus1 band (Fig. 4a).
To further confirm that Cdc31 is a stably associated subunit of theSac3–Thp1–Sus1 complex, we performed gel filtration chromatogra-phy. Cdc31 co-fractionated with the other subunits of theSac3–Thp1–Sus1 complex (Fig. 4b), which all peaked in fraction 14(Mr, ~ 400K) of the Superose-6 column (also see ref. 20). Thus, co-frac-tionation of Cdc31 with Sac3, Thp1 and Sus1 suggests that Cdc31 is agenuine component of this mRNA export complex.
Finally, we sought to purify Cdc31 from yeast cells and test whether itassociates with members of the Sac3 complex. Cdc31 was TAP-taggedat the C-terminus by genomic integration (see Methods section) andaffinity purified from lysates by the TAP method. Several proteins co-enriched with Cdc31, including Sac3, Thp1 and Sfi1 (Fig. 4c). Theaffinity purification of Protein A (ProtA)-tagged Cdc31 was recentlyreported, along with the identification of Sfi1, a genuine SPB-compo-nent, as well as Thp1 (ref. 28). Thus, biochemical analyses have shownthat Cdc31 is not only associated with SPB components in vivo24,26, butalso with members of the Sac3–Thp1–Sus1 complex.
Cdc31 is involved in nuclear mRNA exportCdc31 has been reported to co-purify with nuclear pore complexes29.This observation agrees with our finding that Cdc31 is a component ofthe Sac3–Thp1–Sus1 complex that is concentrated at the nuclearpores20. However, indirect immunofluorescence with anti-Cdc31 anti-bodies detected Cdc31 at SPBs, but not at the nuclear envelope25. Were-investigated the sub-cellular localization of Cdc31 using a GFP-tagged CDC31 construct (under the control of its endogenous pro-moter) that was expressed in repressed tetO::cdc31 cells (also seebelow). In these living cells, Cdc31–GFP not only strongly labelled theSPBs, but also significantly labelled the nuclear envelope surroundingthe chromosomal DNA (Fig. 5). These data suggest that Cdc31 associ-ates with both the SPB and nuclear pore complexes.
In the course of dissecting the functions of Cdc31, cdc31 temperature-sensitive alleles were isolated that were grouped into three differentclasses30: first, temperature-sensitive alleles in the C-terminal region thataffected SPB function; second, temperature-sensitive alleles in the middleregion that caused cell lysis and reduced Kic1 kinase activity; and third,temperature-sensitive alleles in a third region of Cdc31 (for example,cdc31-115; L45S), the linkage of which remained unknown as they did notaffect cell lysis or SPB function. Notably, cdc31-115, as a member of thethird class, showed a significant accumulation of poly(A)+ RNA inside thenucleus after a 2 h shift to the restrictive temperature (Fig. 5b). In con-trast, cdc31-54, which was defective in Kic1 and Kar1 binding, did notinduce the same mRNA export defect (Fig. 5b). We conclude that Cdc31is involved in nuclear export of poly(A)+ RNA in an allele-specific way.
Moreover, we tested whether repression of CDC31 gene expressioncauses inhibition of mRNA export. Thus, the CDC31 gene was placedunder the control of the tetO promoter, which can be repressed by doxy-cycline31. Addition of doxycycline to yeast cells that carry the integratedtetO::CDC31 allele induced a significant accumulation of poly(A)+ RNAinside the nucleus during CDC31 gene repression (Fig. 5c). Finally, wetested whether the export of tRNA, ribosomal subunits and cargoes car-rying nuclear export signals is inhibited in the tetO::CDC31 mutant, butno defects were observed (see Supplementary Information, Fig. S1).
a
b c
SAC
356
3-13
01SA
C3
733-
1301
SAC
385
1-13
01SA
C3
CID
SAC
356
3−13
01SA
C3
733−
1301
SAC
385
1−13
01
SAC
373
3−13
01SA
C3
851−
1301
SAC
3C
ID
Tev
Cdc31
Sus1
Mr(K)
70 -
50 -
30 -
25 -
20 -
15 -
10 -
CBP
Sus1−myc
Cdc31
SUS1−MYC sus1∆
- 100
- 70
- 50
- 30
- 20
Mr(K)
Figure 2 Overexpressed Sac3CID recruits Cdc31. (a, b) Correlation betweentoxicity of overexpressed Sac3 C-fragments and binding to Cdc31 and Sus1.The indicated TAP-tagged Sac3 deletion constructs (under GAL1 promotercontrol) were overexpressed in SUS1 (a) or SUS1–MYC cells (b) and affinitypurified by IgG-Sepharose chromatography and TEV elution. Eluates wereresolved on a 12% SDS–PAGE gel and analysed by: Coomassie staining (a)and western blotting (b) with anti-CBP (to detect TAP-tagged Sac3 baitproteins), anti-myc (to detect Sus1–13myc) and anti-Cdc31 antibodies.Cdc31 and Sus1 bands were identified by mass spectrometry. (c) Sac3CIDbinds to Cdc31 independently of Sus1. The indicated TAP-tagged Sac3 C-domain fragments were overexpressed in sus1∆ cells, affinity purified andanalysed by SDS–PAGE and western blotting, as described above.
print ncb1163 13/8/04 4:33 PM Page 842
© 2004 Nature Publishing Group

A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004 843
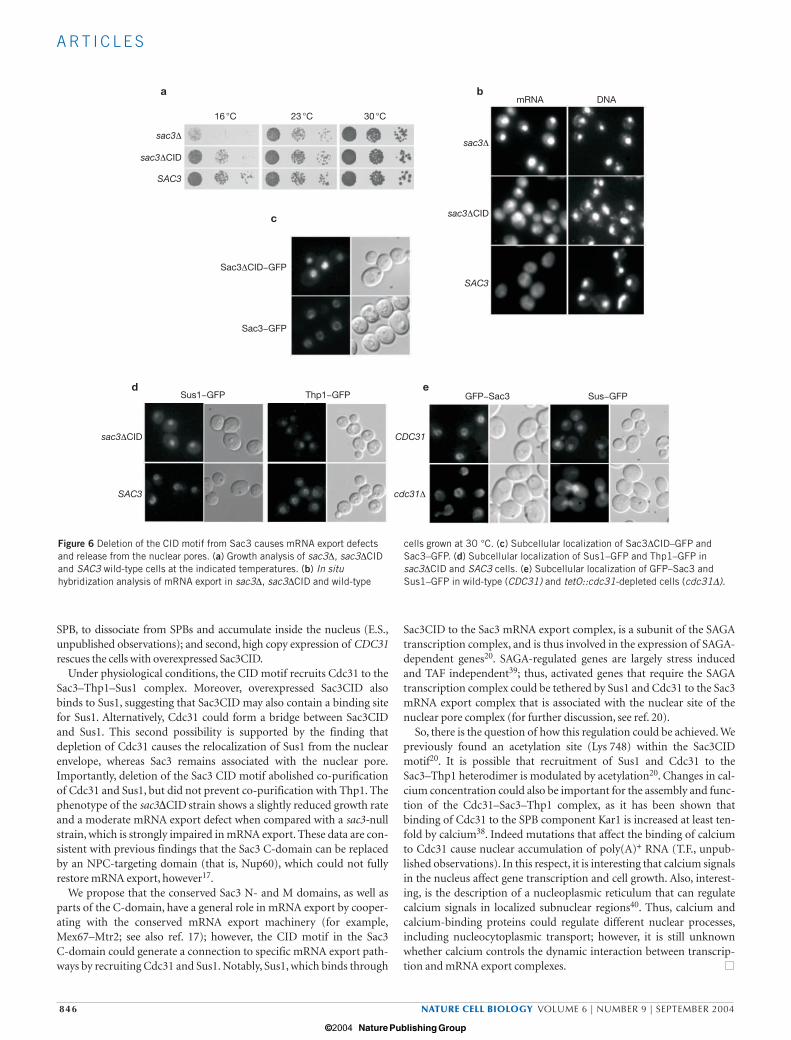
Sac3∆CID induces an mRNA export defect and is no longerassociated with Cdc31Our data had already shown that the Sac3 CID motif mediates the inter-action with Cdc31, and that GFP–Sac3CID is targeted to the nuclearenvelope. Next, we wanted to test the functional consequence of deletingthe CID domain from Sac3. Yeast cells expressing Sac3∆CID grewslightly slower than wild-type cells, particularly at lower temperatures(Fig. 6a). Moreover, sac3∆CID cells showed mRNA export defects (Fig.6b). Interestingly, a fusion of Sac3∆CID and GFP did not associate with
the nuclear periphery, but accumulated in the nucleus (Fig. 6c).Concomitantly, Sus1–GFP and Thp1–GFP were dissociated from thenuclear envelope in sac3∆CID cells (Fig. 6d). These data suggest that theCID motif is important for the steady state association of theSac3–Thp1–Sus1–Cdc31 complex with the nuclear pores.
Moreover, we analysed the localization of Sus1–GFP in thetetO::CDC31 repression mutant and found that it was no longer concen-trated at the nuclear periphery (Fig. 6e). In contrast, Sac3–GFP still local-ized to the nuclear envelope in the tetO::CDC31 repressed strain (Fig. 6e).
GAL GAL::SAC3
GAL::SAC3CID
GAL::SAC3563−1301
GAL::SAC3563−1301
GAL::SAC3733−1301
GAL::SAC3CID
GAL::SAC3851−1301
GAL
GAL::SAC3733−1301
GAL::SAC3851−1301
a
b
100
75
50
25
00 108642
Time (h)
Per
cent
age
of c
ells
SPC42−GFP100
75
50
25
00 108642
Time (h)
Per
cent
age
of c
ells
SPC42−GFP
100
75
50
25
00 108642
Time (h)
Per
cent
age
of c
ells
SPC42−GFP
100
75
50
25
00 108642
Time (h)
Per
cent
age
of c
ells
SPC42−GFP
100
75
50
25
00 108642
Time (h)
Per
cent
age
of c
ells
SPC42−GFP
100
75
50
25
00 108642
Time (h)
Per
cent
age
of c
ells
SPC42−GFP
pRS426 pRS426−CDC31
Figure 3 Overexpressed Sac3CID causes cell-cycle defects that can be rescuedby high copy CDC31. (a) α-factor synchronized cells overexpressing theindicated Sac3 C-fragments were released into a new cell cycle. Cellmorphology, SPB status and the SPB marker Spc42–GFP were monitored overtime. Note that SAC3, SAC3563–1301, SAC3733–1301 and SAC3CID cells are allarrested only in the second cell cycle after α-factor release. This phenotype isconsistent with defects in Cdc31 function33. Open squares, small budded
cells with a single SPB; open circles, medium and large budded cells withduplicated and separated SPBs; black triangles, large budded cells with asingle SPB. (b) Cdc31 overexpression rescues the dominant-negativephenotype of overexpressed Sac3CID. Cells expressing the indicated Sac3 C-domain constructs were transformed with the high-copy plasmid pRS426,with or without the CDC31 gene. Transformants were spotted in 10−1 dilutionsteps on galactose-containing plates and grown for four days at 30 °C.
print ncb1163 13/8/04 4:34 PM Page 843
© 2004 Nature Publishing Group

A RT I C L E S
844 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004
To obtain biochemical evidence that the CID motif is necessary torecruit Cdc31 to the Sac3 complex, we sought to further characterizethese interactions. Thus, we affinity purified the Sac3 complex fromSAC3 and sac3∆CID cells (see Methods section). Cdc31 was seem-ingly absent from the Sac3 complex that lacked the CID motif(Fig. 7a, lane 2). Moreover, Sus1 was no longer associated with aSac3∆CID–Thp1 core complex. However, the absence of Sus1 from aSac3∆CID–Thp1 complex did not affect the binding of Sus1 to theSAGA complex (Fig. 7a, lane 4). Taken together, these data show thatthe CID motif in Sac3 is necessary and sufficient to recruit Cdc31and Sus1 to a Sac3–Thp1 heterodimer, and the Sac3–Thp1–Sus1complex to the nuclear periphery.
Finally, crosslinking studies were performed to identify physical con-tacts within the Sac3–Thp1–Sus1–Cdc31 complex, using the bivalentchemical crosslinker BS3 (see Methods section). As shown by westernblot analysis with antibodies against Cdc31, Thp1 and Sus1, thecrosslinker generated several discrete crosslinked products (Fig. 7b).Bands in the lower molecular weight range of the gel were interpretedto be products crosslinked between Cdc31 and Sus1, Cdc31 and Thp1,and Cdc31 with itself. The higher molecular weight bands were moredifficult to assign, but were consistent with crosslinks between Cdc31and Sac3. We conclude from these studies that Cdc31 is tightly embed-ded within the Sac3–Thp1–Sus1 complex, and that it directly binds toseveral members of the complex.
DISCUSSIONSo far, the best characterized role of centrins is their participation inthe cell-cycle-dependent duplication of microtubule-organizingcentres (MTOCs), such as centrosomes and spindle pole bodies(SPBs)25,32,33. Recent evidence, however, suggests that centrins have
additional functions, for example, in nucleotide excision repair(human centrin 2)34, photoreceptor signal transduction (humancentrin 1)35 or regulation of cell integrity through kinase activation(yeast centrin Cdc31)36.
In this study, we have extended the role of centrins with the identifi-cation of the Sac3–Thp1–Sus1 complex as a novel Cdc31 target. Wepreviously showed that the Sac3–Thp1–Sus1 complex associates withnuclear pores and is involved in transcription-coupled mRNAexport17,20. Cdc31, in association with Sac3–Thp1–Sus1, could eitherfunction in SPB duplication or mRNA export. However, the analysis ofsac3-null cells did not identify an obvious SPB defect (E.S., unpub-lished observations). Instead, our data suggest that Cdc31 has a specificrole in nuclear mRNA export.
It seems common that proteins with an important role in distinctcellular pathways are physically linked to the nuclear pore complex.Examples include the following: first, Sec13, which is a subunit of theCOPII complex that is involved in vesicular transport from the endo-plasmic reticulum to the Golgi and is also a member of the Nup84complex, which plays a crucial role in mRNA export37; second, Ndc1,which is both a subunit of the spindle pole body and of the nuclearpore complex29; and third, Sus1, which is part of an NPC-associatedmRNA export complex and a subunit of the SAGA-transcription com-plex20. In this way, major cellular pathways may be able to influenceeach other or can be coordinately regulated.
Our data suggest that Cdc31 binds to a 130-amino-acid-long sequencewithin the Sac3 C-domain, termed CID. Small peptides that mediateCdc31 binding have been identified in Kar1, Kic1 and Sfi1 (refs 26, 28, 36).These binding motifs in Kar1 and Kic1 have the following in common: aseries of positively charged amino acids, a negatively charged amino acid,a bulky hydrophobic amino acid (W/Y) in the centre, followed by
a b c
Sus1−TAPThp−GST
Cdc31−TAP
Mr(K) Mr(K) Mr(K)
200 -
120 -
100 -
70 -
50 -
30 -
20 -
15 -
- Tra1
- Spt7
Sac3
Thp1
Cdc31Sus1
L 9 16151413121110 17 232221201918 24 Fraction
- 200
- 70
- 50
- 30
- 15
120 -
100 -
70 -
50 -
30 -
20 -
15 -
- Sac3
- Sfi1- Sac3
- Thp1
- Porin (E. coli)
- Cdc31
- Cdc31
Figure 4 Cdc31 is a component of the Sac3–Thp1–Sus1 complex.(a) Split-double-tag affinity purification of the Sac3–Thp1–Sus1–Cdc31complex. Yeast cells expressing Sus1–TAP and Thp1–GST were lysed,and Sus1–TAP was affinity purified on IgG–Sepharose. After TEVrelease, the eluate was affinity purified by Thp1–GST onGSH–Sepharose. The final eluate was analysed on a 4–12% gradientSDS–PAGE gel and visualized by Coomassie staining. Co-purifyingproteins were identified by mass spectrometry and are indicated. (b) Gel
filtration chromatography of the purified Sac3–Thp1–Sus1–Cdc31complex isolated by the split-double-tag method (see above). Load (L)and fractions 9–24 from the Superose-6 column are shown. Asterisksdenote the position of the Sac3 doublet band, Thp1, Sus1 and Cdc31.(c) Affinity purification of TAP-tagged Cdc31 highlights an associationwith Sac3 and Thp1. The final EGTA eluate from calmodulin–Sepharosewas analysed by SDS–PAGE and Coomassie staining, and the indicatedbands were identified by mass spectrometry.
print ncb1163 13/8/04 4:34 PM Page 844
© 2004 Nature Publishing Group

A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004 845
hydrophobic amino acids and terminated by a cluster of positivelycharged amino acids. Moreover, these peptides can form an amphipathicα-helix, which is a structural element common to many calmodulin- andcentrin-binding motifs38. Notably, such a putative amphipathic Cdc31-binding sequence (N-795KKKFFEKWQASYSQAKKNR813-C) is presentwithin the CID motif of Sac3 (E.S., unpublished observations).
The finding that Cdc31 binds to Sac3CID explains why overexpressionof Sac3CID is toxic and why cells arrest in the second cell cycle as largebudded cells with a single apparent SPB, a morphology that is typical formutants defective in Cdc31 function at SPBs32. Excess Sac3CID with-draws Cdc31 from the SPB, which agrees with the observations that: first,overexpression of SAC3CID causes Cdc31–GFP, which localizes to the
a
b
c
Cdc31−GFP + DNA + Nomarski
tet7−C
DC
31 C
DC
31−G
FP
23 °C 37 °C
cdc31−115
cdc31−54
mRNA
mRNA
DNA DNAmRNA
DNA
tetO::CDC31
0 h 3 h 5 h 7 h
Figure 5 Cdc31 associates with the nuclear envelope and cdc31 mutantsaccumulate poly(A)+ RNA inside the nucleus. (a) Intracellular localization ofCdc31–GFP. Endogenous CDC31 was depleted in tetO::CDC31 cellscarrying pRS316–CDC31–GFP by treatment with doxycycline. DNA wasstained with Hoechst 33342 and cells were inspected without fixation.
Scale bar represents 5 µm. (b) Nuclear accumulation of poly(A)+ RNA incdc31-115, but not cdc31-54, cells after shifting the temperature to 37 °Cfor 2 h. (c) Nuclear accumulation of poly(A)+ RNA in tetO::CDC31 cells afterthe addition of doxycycline. The localization of poly(A)+ RNA was analysedby in situ hybridization and DNA was stained with DAPI.
print ncb1163 13/8/04 4:34 PM Page 845
© 2004 Nature Publishing Group
A RT I C L E S
846 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004
SPB, to dissociate from SPBs and accumulate inside the nucleus (E.S.,unpublished observations); and second, high copy expression of CDC31rescues the cells with overexpressed Sac3CID.
Under physiological conditions, the CID motif recruits Cdc31 to theSac3–Thp1–Sus1 complex. Moreover, overexpressed Sac3CID alsobinds to Sus1, suggesting that Sac3CID may also contain a binding sitefor Sus1. Alternatively, Cdc31 could form a bridge between Sac3CIDand Sus1. This second possibility is supported by the finding thatdepletion of Cdc31 causes the relocalization of Sus1 from the nuclearenvelope, whereas Sac3 remains associated with the nuclear pore.Importantly, deletion of the Sac3 CID motif abolished co-purificationof Cdc31 and Sus1, but did not prevent co-purification with Thp1. Thephenotype of the sac3∆CID strain shows a slightly reduced growth rateand a moderate mRNA export defect when compared with a sac3-nullstrain, which is strongly impaired in mRNA export. These data are con-sistent with previous findings that the Sac3 C-domain can be replacedby an NPC-targeting domain (that is, Nup60), which could not fullyrestore mRNA export, however17.
We propose that the conserved Sac3 N- and M domains, as well asparts of the C-domain, have a general role in mRNA export by cooper-ating with the conserved mRNA export machinery (for example,Mex67–Mtr2; see also ref. 17); however, the CID motif in the Sac3 C-domain could generate a connection to specific mRNA export path-ways by recruiting Cdc31 and Sus1. Notably, Sus1, which binds through
Sac3CID to the Sac3 mRNA export complex, is a subunit of the SAGAtranscription complex, and is thus involved in the expression of SAGA-dependent genes20. SAGA-regulated genes are largely stress inducedand TAF independent39; thus, activated genes that require the SAGAtranscription complex could be tethered by Sus1 and Cdc31 to the Sac3mRNA export complex that is associated with the nuclear site of thenuclear pore complex (for further discussion, see ref. 20).
So, there is the question of how this regulation could be achieved. Wepreviously found an acetylation site (Lys 748) within the Sac3CIDmotif20. It is possible that recruitment of Sus1 and Cdc31 to theSac3–Thp1 heterodimer is modulated by acetylation20. Changes in cal-cium concentration could also be important for the assembly and func-tion of the Cdc31–Sac3–Thp1 complex, as it has been shown thatbinding of Cdc31 to the SPB component Kar1 is increased at least ten-fold by calcium38. Indeed mutations that affect the binding of calciumto Cdc31 cause nuclear accumulation of poly(A)+ RNA (T.F., unpub-lished observations). In this respect, it is interesting that calcium signalsin the nucleus affect gene transcription and cell growth. Also, interest-ing, is the description of a nucleoplasmic reticulum that can regulatecalcium signals in localized subnuclear regions40. Thus, calcium andcalcium-binding proteins could regulate different nuclear processes,including nucleocytoplasmic transport; however, it is still unknownwhether calcium controls the dynamic interaction between transcrip-tion and mRNA export complexes.
a
c
d e
b
16 °C 23 °C 30 °C
sac3∆sac3∆
sac3∆CID
sac3∆CID
sac3∆CID
SAC3
SAC3
SAC3
CDC31
cdc31∆
mRNA DNA
Sac3∆CID−GFP
Sac3−GFP
Sus1−GFP GFP−Sac3 Sus−GFPThp1−GFP
Figure 6 Deletion of the CID motif from Sac3 causes mRNA export defectsand release from the nuclear pores. (a) Growth analysis of sac3∆, sac3∆CIDand SAC3 wild-type cells at the indicated temperatures. (b) In situhybridization analysis of mRNA export in sac3∆, sac3∆CID and wild-type
cells grown at 30 °C. (c) Subcellular localization of Sac3∆CID–GFP andSac3–GFP. (d) Subcellular localization of Sus1–GFP and Thp1–GFP insac3∆CID and SAC3 cells. (e) Subcellular localization of GFP–Sac3 andSus1–GFP in wild-type (CDC31) and tetO::cdc31-depleted cells (cdc31∆).
print ncb1163 13/8/04 4:34 PM Page 846
© 2004 Nature Publishing Group

A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004 847
METHODSStrains, plasmids and growth conditions. The yeast strains and plasmids used inthis study are listed in the Supplementary Information, Table 1 and 2, respec-tively. Chromosomal integration of GFP, GST, the 13myc or TAP as C-terminaltags was performed as described20. Integration of the tetracycline-regulatabletetO promoter was also performed as described31. For synchronization, cells were
incubated with 10 µg ml−1 α-factor (Sigma, St Louis, MO) for 3 h at 30 °C. Afterwashing twice with medium to remove α-factor, cells progressed synchronouslythrough the cell cycle. For galactose induction, yeast cells carrying genes underthe control of the GAL1 promoter were grown to early logarithmic phase in syn-thetic complete medium containing 3% raffinose as the sole carbon source. TheGAL1 promoter was induced after addition of 2% galactose. For depletion of
Sac3−Thp1 purification Sus1 purification
SAC3 SAC3∆CID SAC3 SAC3∆CID
Sac3 Sac3∆CID
- ?
Tra1 -
Spt7 -
Cdc31 -
Cdc31 -
Sus1 -
Thp1 -
SA
GA
- 150
- 100
- 70
- 50
- 30
- 25
- 20
- 15
Thp1 -
Cdc31 -Sus1 -
Cdc31 -Sus1 -
1 2 3 4
Coomassie
Western
a
b
200 -150 -
100 -
70 -
50 -
30 -
20 -
Mr(K)− + + − + + − + + BS3
Cdc31−Thp1 Cdc31−Thp1
Cdc31−Sus1 Cdc31−Sus1
Sus1
Cdc31−Cdc31
Cdc31
Thp1
Thp11 2 3
Cdc311 2 3
Sus11 2 3
Sac3
Figure 7 The CID motif of Sac3 is necessary to recruit Cdc31 and Sus1 toSac3–Thp1. (a) Sus1 and Cdc31 no longer co-purify with Sac3∆CID. Strainsused for affinity purification were Thp1–GST/Sus1–TAP/ProtA–TEV–Sac3(lanes 1 and 3) and Thp1–GST/Sus1–TAP/ProtA–TEV–Sac3∆CID (lanes 2 and4). Sac3–Thp purification: ProtA–TEV–Sac3 (lane 1) or ProtA–TEV–Sac3∆CID(lane 2), respectively, were first affinity purified on IgG–Sepharose and TEVeluted (note that Sus1–TAP also enriches at this step). In a second step, theeluates were passed over GSH–Sepharose to enrich for Thp1–GST. The finaleluates were analysed by SDS–PAGE and Coomassie staining (top) or westernblotting with anti-Cdc31 and anti-CBP antibodies (bottom). Labelled bands
were identified by mass spectrometry. Asterisk indicates Sac3 breakdownproducts. Sus1 purification: flow-through fractions from the GSH–Sepharose(see above) were passed over Calmodulin–Sepharose. The final EGTA eluates,which contained purified Sus1–CBP, were analysed by SDS–PAGE andCoomassie staining or western blotting. (b) Crosslinking analysis of theSac3–Thp1–Sus1–Cdc31 complex. The purified Thp1–Sac3–Sus1–Cdc31complex was incubated with 50 µM (lane 2), 100 µM (lane 3) or no (lane 1)BS3 chemical crosslinker. Products were analysed by SDS–PAGE and westernblotting with anti-CBP (Thp1–CBP), anti-Cdc31 or anti-myc (Sus1–myc)antibodies. Potential cross-linked products are indicated.
print ncb1163 13/8/04 4:34 PM Page 847
© 2004 Nature Publishing Group

A RT I C L E S
848 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 9 | SEPTEMBER 2004
Cdc31, the expression of CDC31 under the control of the tetO promoter was sup-pressed in YPD medium supplemented with 2 µg ml−1 doxycycline31.
Fluorescence microscopy. Yeast cells expressing GFP-tagged proteins wereanalysed with or without fixation by 4% paraformaldehyde in 150 mM phos-phate buffer at pH 6.5, for 10 min at 25 °C. The DNA of fixed cells was stainedwith 4,6-diamidino-2-phenylindole (DAPI). For staining of the DNA in livecells, yeast cell cultures were incubated with 0.5 µg ml−1 bisbenzimide H33342(Hoechst 33342; Sigma) for 20–45 min. Cells were examined by fluorescencemicroscopy with a Zeiss Axiophot microscope with a Xillix Microimager CCDcamera. In situ hybridization of poly(A)+ RNA was performed as described17.
Protein purification, chemical crosslinking and mass spectrometry. Affinitypurification of TAP-tagged proteins was performed as described41. TEV orEGTA eluates were analysed by SDS–PAGE and Coomassie staining, usingNovex 12% or 4–12% gradient gels (Invitrogen, Carlsbad, CA). GST purifica-tion was performed in TAP lysis buffer and elution was performed in the pres-ence of 10 mM glutathione. For gel-filtration chromatography, Sus1–TAP andThp1–GST were affinity purified as described above. The final eluate was treatedwith 20 U ml−1 of DNAse I (RNase free), concentrated by ultrafiltration andapplied on a Superose-6 column. Gel filtration analysis was performed on anEttan LC (Amersham, Piscataway, NJ). For chemical crosslinking, the TAP-puri-fied Sac3 complex in PBS and 5 mM EGTA was mixed with freshly prepared BS3
(Pierce, Rockford, IL) in 50 µM or 100 µM concentration, incubated for 1 h onice, quenched by adding 25 mM Tris buffer at pH 8.0, TCA precipitated andanalysed by SDS–PAGE and western blotting with anti-myc, anti-CBP and anti-Cdc31 antibodies. Mass spectrometry analysis of tryptic digests fromCoomassie stained bands was performed as described42.
BIND identifiers. Six BIND identifiers (www.BIND.ca) are associated with thismanuscript: 151162, 151163, 151164, 151165, 151166 and 151167.
Note: Supplementary Information is available on the Nature Cell Biology website.
ACKNOWLEDGEMENTSWe would like to thank the following for technical assistance: E.-M. Lammer, M.Schneider and S. Brettschneider in the Hurt laboratory and S. Merker and P. Ihrigin the Lechner laboratory (Mass Spectrometry Unit, BZH, Heidelberg). E.C.H. wassupported by grants from the Deutsche Forschungsgemeinschaft (SFB638, Leibniz-Programm) and Fonds der Chemischen Industrie.
COMPETING FINANCIAL INTERESTSThe authors declare that they have no competing financial interests.
Received 21 June 2004; accepted 27 July 2004Published online at http://www.nature.com/naturecellbiology.
1. Fahrenkrog, B. & Aebi, U. The nuclear pore complex: nucleocytoplasmic transport andbeyond. Nature Rev. Mol. Cell Biol. 4, 757–766 (2003).
2. Maniatis, T. & Reed, R. An extensive network of coupling between gene expressionmachines. Nature 416, 499–506 (2002).
3. Reed, R. Coupling transcription, splicing and mRNA export. Curr. Opin. Cell Biol. 15,326–331 (2003).
4. Stutz, F. & Izaurralde, E. The interplay of nuclear mRNP assembly, mRNA surveillanceand export. Trends Cell Biol. 13, 319–327 (2003).
5. Reed, R. & Hurt, E. C. A conserved mRNA export machinery coupled to pre-mRNAsplicing. Cell 108, 523–531 (2002).
6. Jensen, T. H., Dower, K., Libri, D. & Rosbash, M. Early formation of mRNP. License forexport or quality control? Mol. Cell 11, 1129–1138 (2003).
7. Zenklusen, D., Vinciguerra, P., Strahm, Y. & Stutz, F. The yeast hnRNP-like proteinsYra1p and Yra2p participate in mRNA export through interaction with Mex67p. Mol.Cell. Biol. 21, 4219–4232 (2001).
8. Sträßer, K. & Hurt, E. C. Yra1p, a conserved nuclear RNA binding protein, interactsdirectly with Mex67p and is required for mRNA export. EMBO J. 19, 410–420(2000).
9. Stutz, F. et al. REF, an evolutionarily conserved family of hnRNP-like proteins, inter-acts with TAP/Mex67p and participates in mRNA nuclear export. RNA 6, 638–650(2000).
10. Rodrigues, J. P. et al. REF proteins mediate the export of spliced and unsplicedmRNAs from the nucleus. Proc. Natl Acad. Sci. USA 98, 1030–1035 (2001).
11. Sträßer, K. et al. TREX is a conserved complex coupling transcription with messengerRNA export. Nature 417, 304–308 (2002).
12. Chávez, S. et al. A protein complex containing Tho2, Hpr1, Mft1 and a novel protein,
Thp2, connects transcription elongation with mitotic recombination in Saccharomycescerevisiae. EMBO J. 19, 5824–5834 (2000).
13. Hurt, E., Luo, M. J., Rother, S., Reed, R. & Strasser, K. Cotranscriptional recruitmentof the serine-arginine-rich (SR)-like proteins Gbp2 and Hrb1 to nascent mRNA via theTREX complex. Proc. Natl Acad. Sci. USA 101, 1858–1862 (2004).
14. Kim, M., Ahn, S. H., Krogan, N. J., Greenblatt, J. F. & Buratowski, S. Transitions inRNA polymerase II elongation complexes at the 3′ ends of genes. EMBO J. 23,354–364 (2004).
15. Zenklusen, D., Vinciguerra, P., Wyss, J. C. & Stutz, F. Stable mRNP formation andexport require cotranscriptional recruitment of the mRNA export factors Yra1p andSub2p by Hpr1p. Mol. Cell Biol. 22, 8241–8253 (2002).
16. Huertas, P. & Aguilera, A. Cotranscriptionally formed DNA:RNA hybrids mediate tran-scription elongation impairment and transcription-associated recombination. Mol. Cell12, 711–721 (2003).
17. Fischer, T. et al. The mRNA export machinery requires the novel Sac3p–Thp1p com-plex to dock at the nucleoplasmic entrance of the nuclear pores. EMBO J. 21,5843–5852 (2002).
18. Gallardo, M., Luna, R., Erdjument-Bromage, H., Tempst, P. & Aguilera, A. The Nab2pand the Thp1p–Sac3p complex functionally interact at the interface between tran-scription and mRNA metabolism. J. Biol. Chem. 278, 24225–24232 (2003).
19. Lei, P. et al. Sac3 is an mRNA export factor that localizes to cytoplasmic fibrils ofnuclear pore complex. Mol. Biol. Cell 14, 836–847 (2003).
20. Rodriguez-Navarro, S. et al. Sus1, a functional component of the SAGA histone acety-lase complex and the nuclear pore-associated mRNA export machinery. Cell 116,75–86 (2004).
21. Novick, P., Osmond, B. C. & Botstein, D. Suppressors of yeast actin mutations.Genetics 121, 659–674 (1989).
22. Jones, A. L. et al. SAC3 may link nuclear protein export to cell cycle progression. Proc.Natl Acad. Sci. USA 97, 3224–3229 (2000).
23. Gallardo, M. & Aguilera, A. A new hyperrecombination mutation identifies a novelyeast gene, THP1, connecting transcription elongation with mitotic recombination.Genetics 157, 79–89 (2001).
24. Biggins, S. & Rose, M. D. Direct interaction between yeast spindle pole body compo-nents: Kar1p is required for Cdc31p localization to the spindle pole body. J. Cell Biol.125, 843–852 (1994).
25. Spang, A., Courtney, I., Fackler, U., Matzner, M. & Schiebel, E. The calcium-bindingprotein cell division cycle 31 of Saccharomyces cerevisiae is a component of the halfbridge of the spindle pole body. J. Cell Biol. 123, 405–416 (1993).
26. Spang, A., Courtney, I., Grein, K., Matzner, M. & Schiebel, E. The cdc31p-bindingprotein Kar1p is a component of the half bridge of the yeast spindle pole body. J. CellBiol. 128, 863–878 (1995).
27. Donaldson, A. D. & Kilmartin, J. V. Spc42p: A phosphorylated component of the S.cerevisiae spindle pole body (SPB) with an essential function during SPB duplication.J. Cell Biol. 132, 887–901 (1996).
28. Kilmartin, J. V. Sfi1p has conserved centrin-binding sites and an essential function inbudding yeast spindle pole body duplication. J. Cell Biol. 162, 1211–1221 (2003).
29. Rout, M. P. et al. The yeast nuclear pore complex: composition, architecture, andtransport mechanism. J. Cell Biol. 148, 635–651 (2000).
30. Ivanovska, I. & Rose, M. D. Fine structure analysis of the yeast centrin, Cdc31p, iden-tifies residues specific for cell morphology and spindle pole body duplication.Genetics 157, 503–518 (2001).
31. Belli, G., Gari, E., Piedrafita, L., Aldea, M. & Herrero, E. An activator/repressor dualsystem allows tight tetracycline-regulated gene expression in budding yeast. NucleicAcids Res. 26, 942–947 (1998).
32. Baum, P., Furlong, C. & Byers, B. Yeast gene required for spindle pole body duplica-tion: Homology of its product with calcium binding proteins. Proc. Natl Acad. Sci.USA 83, 5512–5516 (1986).
33. Paoletti, A. et al. Fission yeast cdc31p is a component of the half-bridge and controlsSPB duplication. Mol. Biol. Cell 14, 2793–2808 (2003).
34. Araki, M. et al. Centrosome protein centrin 2/caltractin 1 is part of the xeroderma pig-mentosum group C complex that initiates global genome nucleotide excision repair. J.Biol. Chem. 276, 18665–18672 (2001).
35. Pulvermuller, A. et al. Calcium-dependent assembly of centrin-G-protein complex inphotoreceptor cells. Mol. Cell. Biol. 22, 2194–2203 (2002).
36. Sullivan, D. S., Biggins, S. & Rose, M. D. The yeast centrin, cdc31p, and the interactingprotein kinase, Kic1p, are required for cell integrity. J. Cell Biol. 143, 751–765 (1998).
37. Siniossoglou, S. et al. A novel complex of nucleoporins, which includes Sec13p and aSec13p homolog, is essential for normal nuclear pores. Cell 84, 265–275 (1996).
38. Geier, B. M., Wiech, H. & Schiebel, E. Binding of centrins and yeast calmodulin tosynthetic peptides corresponding to binding sites in the spindle pole body componentsKar1p and Spc110p. J. Biol. Chem. 271, 28366–28374 (1996).
39. Huisinga, K. L. & Pugh, B. F. A genome-wide housekeeping role for TFIID and a highlyregulated stress-related role for SAGA in Saccharomyces cerevisiae. Mol. Cell 13,573–585 (2004).
40. Echevarria, W., Leite, M. F., Guerra, M. T., Zipfel, W. R. & Nathanson, M. H.Regulation of calcium signals in the nucleus by a nucleoplasmic reticulum. NatureCell Biol. 5, 440–446 (2003).
41. Gavin, A.-C. et al. Functional organization of the yeast proteome by systematic analy-sis of protein complexes. Nature 415, 141–147 (2002).
42. Baßler, J. et al. Identification of a 60S pre-ribosomal particle that is closely linked tonuclear export. Mol. Cell 8, 517–529 (2001).
print ncb1163 13/8/04 4:34 PM Page 848
© 2004 Nature Publishing Group

S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
Figure S1 Nuclear export of tRNA, ribosomal subunits and NES-carrying cargoes is not affected in the tetO::cdc31 repression mutant. Analysis of nuclear export of tRNA (a), ribosomal subunits using the Rpl25-GFP (large
subunit) and Rps2-GFP (small subunit) reporters, and of a NLS-NES-GFP construct (b) in tetO::CDC31 cells after addition of doxycycline according to published methods (Fischer et al., 2002; Stade et al., 1997).




















