
Xenopus eHAND: a marker for the developing cardiovascular system of the embryo that is regulated by...
13
Xenopus eHAND: a marker for the developing cardiovascular system of the embryo that is regulated by bone morphogenetic proteins Duncan B. Sparrow, Surendra Kotecha, Norma Towers, Timothy J. Mohun* Division of Developmental Biology, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 1AA, UK Received 17 November 1997; revised version received 7 January 1998; accepted 7 January 1998 Abstract The bHLH protein eHAND is a sensitive marker for cardiovascular precursors in the Xenopus embryo. The earliest site of expression is a broad domain within the lateral plate mesoderm of the tailbud embryo. This domain comprises precursors that contribute to the posterior cardinal veins in later stages. Surprisingly, expression is profoundly asymmetric at this stage and is random with respect to embryo side. XeHAND is also expressed in an anterior domain that encompasses the prospective heart region. Within the myocardium and pericardium, transcripts are also asymmetrically distributed, but in these tissues they are localised in a left-sided manner. Later in development XeHAND transcripts are largely restricted to the ventral aorta, aortic arches and venous inflow tract (sinus venosus) which flank the heart itself, but no expression is detected in neural crest derivatives at any stage. This demonstrates that patterns of XeHAND expression differ markedly amongst vertebrates and that in Xenopus, XeHAND expression identifies all of the earliest formed elements of the cardiovascular system. In animal cap explants, expression of XeHAND (but not other markers of cardiogenic differentiation) is strongly induced by ectopic expression of the TGFb family members, BMP-2 and BMP-4, but this can be blocked by coexpression of a dominant negative BMP receptor. This suggests that XeHAND expression in the embryo is regulated by the ventralising signals of bone morphogenetic proteins. High levels of expression are also detected in explants treated with high doses of activin A which induces cardiac muscle differentiation. No such effect is seen with lower doses of activin, indicating that a second pathway may regulate the XeHAND gene during cardiogenesis. 1998 Elsevier Science Ireland Ltd. Keywords: Xenopus laevis; Cardiovascular system; Heart; bHLH protein; eHAND; Bone morphogenetic protein 1. Introduction The basic helix-loop-helix (bHLH) family of transcrip- tion factors are involved in the regulation of differentiation in a diverse range of cell types. The best studied subfamilies of bHLH factors are those involved in skeletal myogenesis and neurogenesis. In both cases, tissue-specific members of the family play roles in fate determination and terminal differentiation. The central role played by these transcrip- tion factors has prompted a search for further members of the bHLH family which may perform similar, pivotal roles in the differentiation of other cell types. Two novel bHLH proteins, eHAND (Thing-1/HXT) and dHAND (Thing-2/HED), have recently been identified through their ability to from heterodimers with the ubiqui- tous bHLH protein, E12 (Cross et al., 1995; Cserjesi et al., 1995; Hollenberg et al., 1995; Srivastava et al., 1995) and comprise a distinct subfamily. In mouse embryos, their tran- scripts are expressed in complicated and overlapping pat- terns throughout early development. eHAND is strongly expressed in extraembryonic tissues (the early trophoblasts and differentiated giant cells) as well as the embryonic heart, elements of the cardiovascular system, the autonomic ganglia, the gut, and the pharyngeal arches. dHAND is expressed in many of the same embryonic tissues and in addition, is expressed in the maternally derived deciduum. Unlike eHAND, dHAND expression is absent from extraem- bryonic tissues. Little is known about the functional roles of the HAND proteins in early development, nor have gene targets for Mechanisms of Development 71 (1998) 151–163 0925-4773/98/$19.00 1998 Elsevier Science Ireland Ltd. All rights reserved PII S0925-4773(98)00010-0 * Corresponding author. Tel.: +44 181 9138621; fax: +44 181 9064477; e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Xenopus eHAND: a marker for the developing cardiovascular system of the embryo that is regulated by...

Xenopus eHAND:a marker for the developing cardiovascular systemof the embryo that is regulated by bone morphogenetic proteins
Duncan B. Sparrow, Surendra Kotecha, Norma Towers, Timothy J. Mohun*
Division of Developmental Biology, National Institute for Medical Research, The Ridgeway, Mill Hill, London NW7 1AA, UK
Received 17 November 1997; revised version received 7 January 1998; accepted 7 January 1998
Abstract
The bHLH protein eHAND is a sensitive marker for cardiovascular precursors in theXenopusembryo. The earliest site of expression is abroad domain within the lateral plate mesoderm of the tailbud embryo. This domain comprises precursors that contribute to the posteriorcardinal veins in later stages. Surprisingly, expression is profoundly asymmetric at this stage and is random with respect to embryo side.XeHANDis also expressed in an anterior domain that encompasses the prospective heart region. Within the myocardium and pericardium,transcripts are also asymmetrically distributed, but in these tissues they are localised in a left-sided manner. Later in developmentXeHANDtranscripts are largely restricted to the ventral aorta, aortic arches and venous inflow tract (sinus venosus) which flank the heart itself, but noexpression is detected in neural crest derivatives at any stage. This demonstrates that patterns ofXeHANDexpression differ markedlyamongst vertebrates and that inXenopus, XeHANDexpression identifies all of the earliest formed elements of the cardiovascular system. Inanimal cap explants, expression ofXeHAND(but not other markers of cardiogenic differentiation) is strongly induced by ectopic expressionof the TGFb family members, BMP-2 and BMP-4, but this can be blocked by coexpression of a dominant negative BMP receptor. Thissuggests thatXeHANDexpression in the embryo is regulated by the ventralising signals of bone morphogenetic proteins. High levels ofexpression are also detected in explants treated with high doses of activin A which induces cardiac muscle differentiation. No such effect isseen with lower doses of activin, indicating that a second pathway may regulate theXeHANDgene during cardiogenesis. 1998 ElsevierScience Ireland Ltd.
Keywords: Xenopus laevis; Cardiovascular system; Heart; bHLH protein; eHAND; Bone morphogenetic protein
1. Introduction
The basic helix-loop-helix (bHLH) family of transcrip-tion factors are involved in the regulation of differentiationin a diverse range of cell types. The best studied subfamiliesof bHLH factors are those involved in skeletal myogenesisand neurogenesis. In both cases, tissue-specific members ofthe family play roles in fate determination and terminaldifferentiation. The central role played by these transcrip-tion factors has prompted a search for further members ofthe bHLH family which may perform similar, pivotal rolesin the differentiation of other cell types.
Two novel bHLH proteins, eHAND (Thing-1/HXT) and
dHAND (Thing-2/HED), have recently been identifiedthrough their ability to from heterodimers with the ubiqui-tous bHLH protein, E12 (Cross et al., 1995; Cserjesi et al.,1995; Hollenberg et al., 1995; Srivastava et al., 1995) andcomprise a distinct subfamily. In mouse embryos, their tran-scripts are expressed in complicated and overlapping pat-terns throughout early development.eHAND is stronglyexpressed in extraembryonic tissues (the early trophoblastsand differentiated giant cells) as well as the embryonicheart, elements of the cardiovascular system, the autonomicganglia, the gut, and the pharyngeal arches.dHAND isexpressed in many of the same embryonic tissues and inaddition, is expressed in the maternally derived deciduum.Unlike eHAND, dHANDexpression is absent from extraem-bryonic tissues.
Little is known about the functional roles of the HANDproteins in early development, nor have gene targets for
Mechanisms of Development 71 (1998) 151–163
0925-4773/98/$19.00 1998 Elsevier Science Ireland Ltd. All rights reservedPII S0925-4773(98)00010-0
* Corresponding author. Tel.: +44 181 9138621; fax: +44 181 9064477;e-mail: [email protected]

these transcription factors been identified. In the mouse,embryos homozygous for a targeted interruption ofdHAND fail to complete cardiac looping, and appear tolack the right ventricle and aortic arch arteries (Srivastavaet al., 1997). No comparable knockout of theeHANDgenehas yet been reported buteHAND transcripts are detectedpredominantly on the left side of the myocardium prior toheart looping (Biben and Harvey, 1997) and this distributionis reversed in mutant embryos that show situs inversus.eHAND expression in the mouse thus appears to be regu-lated by the pathways that establish embryonic laterality. Afunctional role in cardiogenesis is suggested by the findingthat left-sidedeHAND expression is abolished inNkx2–5knock out embryos, which also fail to progress beyond thelinear heart tube stage (Lyons et al., 1995).
In contrast, studies of theHAND genes in chick develop-ment provide less clear cut evidence for the roles of thesetranscription factors. No asymmetry has been reported inthe distribution ofeHAND or dHAND transcripts withinthe heart tube, and attempts to inhibit the function ofeither using gene-specific antisense oligonucleotides haveproduced no apparent phenotype (Srivastava et al., 1995).However, simultaneous inhibition of bothHAND genesby antisense oligonucleotides does result in the arrestof cardiac development at the normal onset of cardiac loop-ing (HH stage 11–12). Despite the apparent differencesbetween chick and mouse, in each species theHANDgenes appear therefore to play an important role duringcardiac morphogenesis. The precise nature of this role hasyet to be established.
Here we show that in the amphibian,Xenopus laevis,theeHANDgene provides an early and sensitive marker for thedeveloping vasculature of the early embryo. Strikingly, theonset ofeHANDexpression in a broad domain of the lateralmesoderm is randomly asymmetric with respect to embryoside, despite the well established observation that left-rightlaterality is already determined by this stage. The lateraldomains apparently comprise mesenchymal vascular pre-cursor cells and subsequently condense to mark the newlyformed posterior cardinal veins. In the tailbud embryo,eHANDtranscripts are also localised in an anterior, ventraldomain that encompasses the heart primordium. Expressionis detected throughout the embryonic heart tube and subse-quently in both the developing heart, its major vessels andthe aortic arches. Within the heart, transcripts are distributedasymmetrically, marking the left side of the myocardiumand pericardium.
Using explant cultures, we also show that theXeHANDgene is activated in response to bone morphogenetic pro-teins, BMP-2 and BMP-4, which are believed to play a rolein dorso-ventral patterning in the embryo (Graff, 1997) andare implicated in the induction of cardiogenic fate in ante-rior mesoderm (Lough et al., 1996; Schultheiss et al., 1997).The explant assay provides a powerful assay to investigatethe mechanisms that mediate regulation ofXeHANDexpres-sion by the BMP signalling pathway. It may also facilitate
studies of the formation of vascular precursors in the earlyembryo.
2. Results
2.1. Cloning of Xenopus eHAND
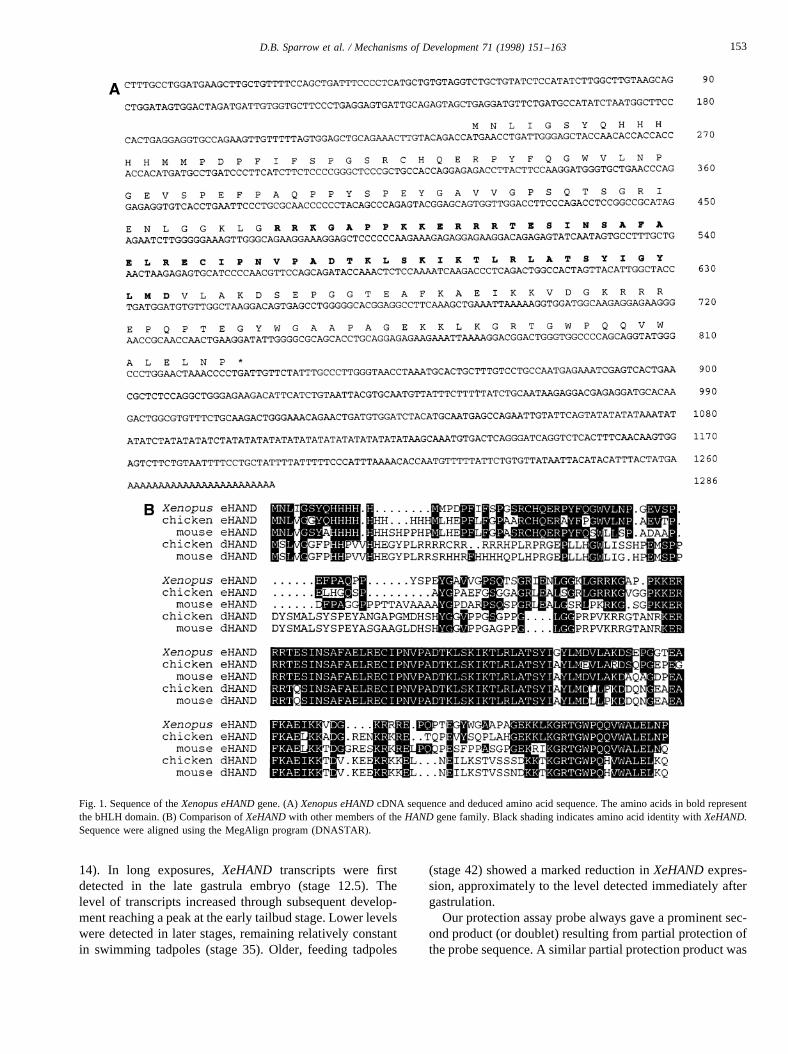
We screened aXenopusadult heart cDNA library usingthe yeast two-hybrid system to look for heart-specific mem-bers of the bHLH family and isolated two overlapping par-tial cDNAs derived from the same gene. The open readingframe contained a bHLH domain which most closelymatched that of eHAND/Thing-1/HXT, a bHLH transcrip-tion factor previously isolated from both mouse and chick(Cross et al., 1995; Cserjesi et al., 1995; Hollenberg et al.,1995). Overlapping clones representing the entire cDNAwere isolated from a random-primed tadpole cDNA library.The complete sequence and conceptual translation ofXeHANDare shown in Fig. 1A.
TheXenopusprotein shows extensive sequence similaritythroughout its length with the chick and mouse eHANDproteins (71% and 62% identity, respectively). The bHLHdomain of these proteins is much more highly conserved,the XeHAND bHLH region having 93% and 91% identitywith the equivalent portion of the mouse and chick proteins(Fig. 1B). In addition to the bHLH domain, there are twoother regions of highly conserved residues shared by theeHAND proteins. One contains 22 amino acids immediatelyC-terminal to the bHLH domain and the second comprisesthe C-terminal 22 amino acids. BLAST searches of thedatabase using these domains does not reveal any otherproteins containing such domains, and it remains to beseen whether these homologies have any functional signifi-cance.
In addition to the vertebrate eHANDs, theXenopussequence shows considerable similarity to the closelyrelated dHAND proteins (also termed Thing-2 or HED)which have been identified in chick and mouse (Cross etal., 1995; Hollenberg et al., 1995; Srivastava et al., 1995).However, apart from the 22 amino acid carboxy terminalmotif mentioned above which is shared by all the HANDproteins, the amino and carboxy terminal domains ofXeHAND can readily be distinguished from those ofchick or mouse dHAND (Fig. 1B). Homology betweenXeHAND and the dHAND sequences is therefore largelyrestricted to the bHLH domain (86% identity).
2.2. XeHAND expression commences during gastrulation
We first examinedXeHAND expression during earlydevelopment using an RNase protection assay (Fig. 2) anda probe comprised entirely of 5′ untranslated regionsequences. No maternalXeHANDtranscripts were detectedin the unfertilised egg but zygotic expression was readilydetected from the neurula stage onwards (Nieuwkoop stage
152 D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163
14). In long exposures,XeHAND transcripts were firstdetected in the late gastrula embryo (stage 12.5). Thelevel of transcripts increased through subsequent develop-ment reaching a peak at the early tailbud stage. Lower levelswere detected in later stages, remaining relatively constantin swimming tadpoles (stage 35). Older, feeding tadpoles
(stage 42) showed a marked reduction inXeHANDexpres-sion, approximately to the level detected immediately aftergastrulation.
Our protection assay probe always gave a prominent sec-ond product (or doublet) resulting from partial protection ofthe probe sequence. A similar partial protection product was
Fig. 1. Sequence of theXenopus eHANDgene. (A)Xenopus eHANDcDNA sequence and deduced amino acid sequence. The amino acids in bold representthe bHLH domain. (B) Comparison ofXeHANDwith other members of theHAND gene family. Black shading indicates amino acid identity withXeHAND.Sequence were aligned using the MegAlign program (DNASTAR).
153D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163

also obtained with a variety of probes from either the codingor 3′ untranslated regions (data not shown) indicating thepresence of transcripts from anotherXeHANDallele. (Coex-pression of divergent alleles is not uncommon forXenopuslaevis,since it is a pseudotetraploid species (Bisbee et al.,1977).)
2.3. XeHAND transcripts identify vascular precursors inthe embryo
RNA wholemount in situ hybridisation revealed that themajor domain ofXeHANDexpression comprised a broadwedge-shaped domain on the flank of the early tailbudembryos (Fig. 3A; stage 24). In contrast to the RNase pro-tection assay results, earlier expression ofXeHANDcouldnot be detected by RNA whole-mount in situ hybridisation,regardless of the length or region ofXeHANDused as theprobe. This presumably reflects a difference in the sensitiv-ity of the two techniques which we have frequentlyobserved. Transverse sections of embryos stained inwhole-mount showed that this expression was restricted tothe lateral plate mesoderm (Fig. 4A), its anterior ventraledge extending to the presumptive heart region of theembryo (Figs. 3A and 5E).
Surprisingly, this expression in the lateral mesoderm wasasymmetrical, with one side of the embryo having a consis-tently higher level of expression across a larger domain(compare the two sides of the same embryo in Fig. 3Aand B). As development proceeded through tailbud stages(stages 27–32) the domain became more defined andrestricted, but the initial asymmetry was retained (Fig. 3,compare C and D; E and F).
By the swimming tadpole stage (stage 33/34) this expres-sion was localised to a narrow band lying at the ventralmargin of the myotomes (Fig. 4F). The pronephric ductlies in a similar position but shows a distinct and irregularmorphology (revealed by the marker PaxB) quite differentfrom that ofXeHANDstaining (T. Carroll and P. Vize, pers.comm.). Furthermore, the pronephric duct lies in closeapposition with the myotomes. Embryos labelled for bothXeHAND and the myotomal muscle-specific markerXMLC1/3showed a significant gap between the ventral mar-gin of the myotomes and theXeHANDexpression domain(Fig. 5A) indicating that the latter stained the newly formingposterior cardinal veins.
Some patchy staining forXeHANDwas also detected inventral regions of late tailbud embryos (Fig. 3E,F; indicatedby the black arrow), which we presume corresponds to thevitelline veins, and both this and the staining of the posteriorcardinal veins was maintained in feeding tadpoles (stages42–46). By this stage it is notable that expression ofXeHANDon each side of the embryo was approximatelyequal.
2.4. The lateral domain of XeHAND shows randomasymmetry
The initial appearance ofXeHAND expression in thelateral mesoderm of early tailbud embryos was strikingfor its apparent asymmetry (Fig. 3). Identical results wereobtained with every batch of stained embryos, whether pig-mented or albino. To investigate this further, we scoredsome embryos for sidedness ofXeHAND expression atearly tailbud (stages 22–25) and others from the samebatch for the direction of gut looping at feeding tadpolestage (stage 42/46).XeHAND expression was randomlyasymmetric whilst 90% of the same embryo batch showeda normal direction of gut looping (Table 1). Danos and Yost(1996) have documented a 3–8% rate of spontaneous car-diac reversals in untreated batches ofXenopusembryos,which is similar to the 10% of embryos that we detectedwith reverse gut looping. These data confirm that the initialonset ofXeHANDexpression in lateral mesoderm was bothasymmetric and random with respect to the left–right axisof the embryo.
A number of trivial explanations for this pattern are pos-sible. Firstly, it could have been an artefact of the in situstaining technique, but this seems unlikely since embryosare agitated throughout the procedure. The staining patternmight also have resulted in some way from the randomdirection in which tailbud embryos bend within the vitellinemembrane, prior to hatching. To test this possibility, tailbudembryos were sorted for direction of bending within theirmembrane and analysed by in situ hybridisation eitherimmediately (stage 25) or at various stages after hatching.No correlation could be established between the direction ofbending and sidedness ofXeHAND expression in theembryo (Table 2).
Fig. 2. Expression ofXeHAND in Xenopusembryos. The distribution ofXeHAND transcripts was analysed by RNase protection assay. Lane 1,undigested probes; lane 2, tRNA control; lanes 3–12, unfertilised oocyte,stages 8, 10, 12.5, 14, 17, 20, 23, 34, 42 embryo RNA, respectively. Twoembryo equivalents of RNA (10mg) was used in each case. Full-lengthprotected fragments for each probe are indicated. As an internal control, aprobe forXMax2 (which is expressed at constant levels throughout earlydevelopment (Tonissen and Krieg, 1994)) was included in theXeHANDassay.
154 D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163
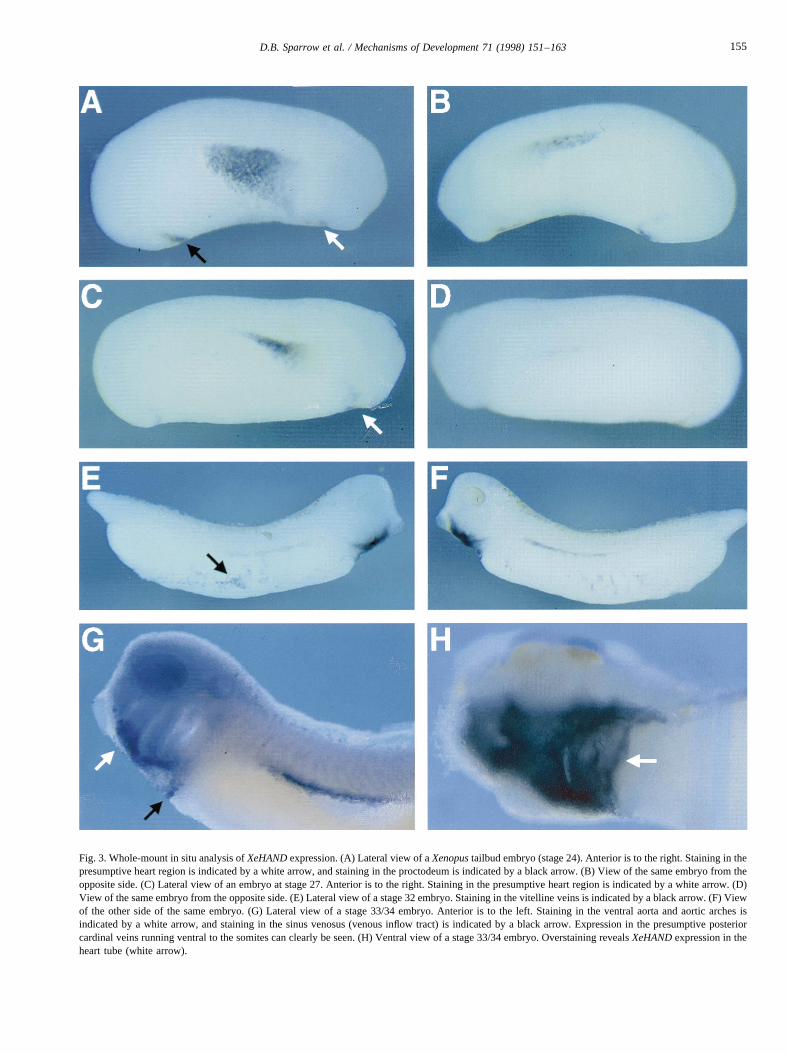
Fig. 3. Whole-mount in situ analysis ofXeHANDexpression. (A) Lateral view of aXenopustailbud embryo (stage 24). Anterior is to the right. Staining in thepresumptive heart region is indicated by a white arrow, and staining in the proctodeum is indicated by a black arrow. (B) View of the same embryo from theopposite side. (C) Lateral view of an embryo at stage 27. Anterior is to the right. Staining in the presumptive heart region is indicated by a white arrow. (D)View of the same embryo from the opposite side. (E) Lateral view of a stage 32 embryo. Staining in the vitelline veins is indicated by a black arrow. (F) Viewof the other side of the same embryo. (G) Lateral view of a stage 33/34 embryo. Anterior is to the left. Staining in the ventral aorta and aortic arches isindicated by a white arrow, and staining in the sinus venosus (venous inflow tract) is indicated by a black arrow. Expression in the presumptive posteriorcardinal veins running ventral to the somites can clearly be seen. (H) Ventral view of a stage 33/34 embryo. Overstaining revealsXeHANDexpression in theheart tube (white arrow).
155D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163
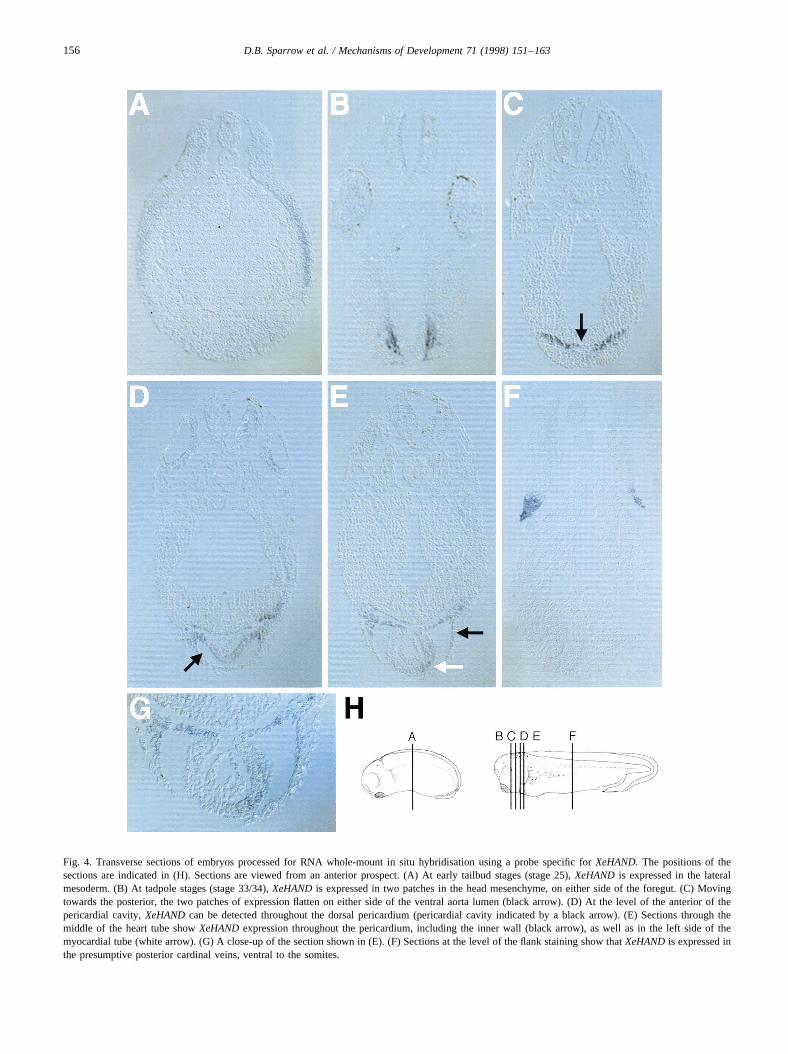
Fig. 4. Transverse sections of embryos processed for RNA whole-mount in situ hybridisation using a probe specific forXeHAND.The positions of thesections are indicated in (H). Sections are viewed from an anterior prospect. (A) At early tailbud stages (stage 25),XeHAND is expressed in the lateralmesoderm. (B) At tadpole stages (stage 33/34),XeHANDis expressed in two patches in the head mesenchyme, on either side of the foregut. (C) Movingtowards the posterior, the two patches of expression flatten on either side of the ventral aorta lumen (black arrow). (D) At the level of the anterior of thepericardial cavity,XeHANDcan be detected throughout the dorsal pericardium (pericardial cavity indicated by a black arrow). (E) Sections through themiddle of the heart tube showXeHANDexpression throughout the pericardium, including the inner wall (black arrow), as well as in the left side of themyocardial tube (white arrow). (G) A close-up of the section shown in (E). (F) Sections at the level of the flank staining show thatXeHANDis expressed inthe presumptive posterior cardinal veins, ventral to the somites.
156 D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163
2.5. XeHAND is expressed throughout the heartformingregion
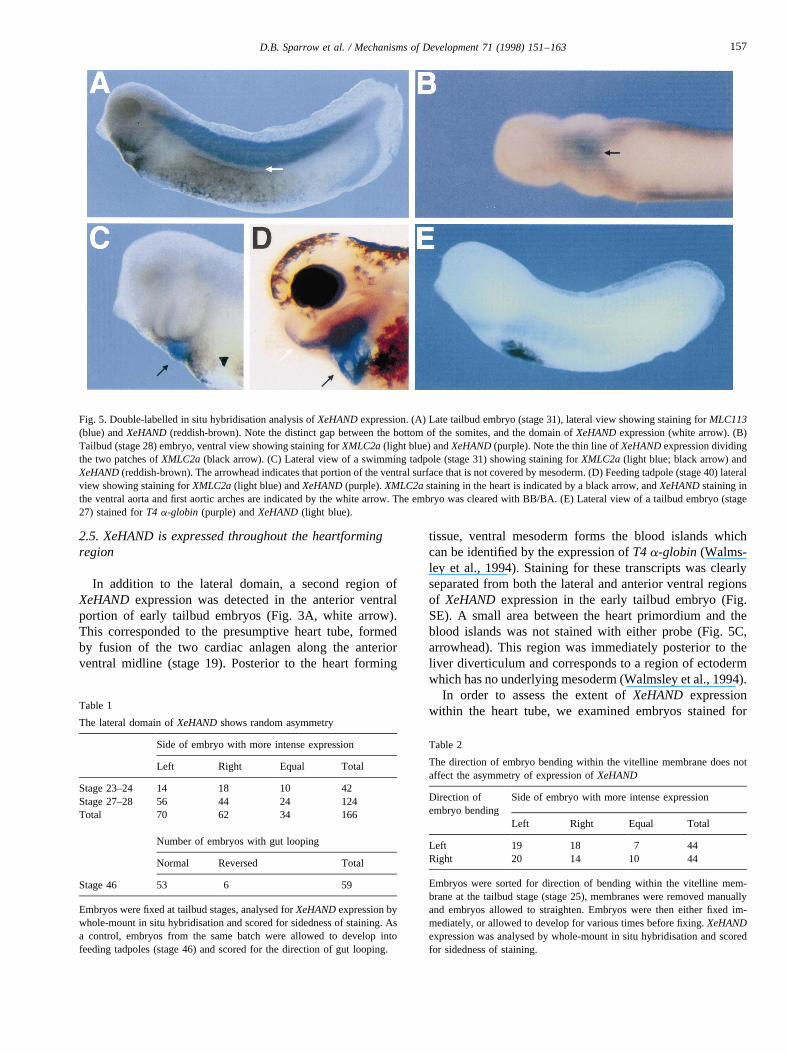
In addition to the lateral domain, a second region ofXeHAND expression was detected in the anterior ventralportion of early tailbud embryos (Fig. 3A, white arrow).This corresponded to the presumptive heart tube, formedby fusion of the two cardiac anlagen along the anteriorventral midline (stage 19). Posterior to the heart forming
tissue, ventral mesoderm forms the blood islands whichcan be identified by the expression ofT4 a-globin (Walms-ley et al., 1994). Staining for these transcripts was clearlyseparated from both the lateral and anterior ventral regionsof XeHAND expression in the early tailbud embryo (Fig.SE). A small area between the heart primordium and theblood islands was not stained with either probe (Fig. 5C,arrowhead). This region was immediately posterior to theliver diverticulum and corresponds to a region of ectodermwhich has no underlying mesoderm (Walmsley et al., 1994).
In order to assess the extent ofXeHAND expressionwithin the heart tube, we examined embryos stained for
Fig. 5. Double-labelled in situ hybridisation analysis ofXeHANDexpression. (A) Late tailbud embryo (stage 31), lateral view showing staining forMLC113(blue) andXeHAND(reddish-brown). Note the distinct gap between the bottom of the somites, and the domain ofXeHANDexpression (white arrow). (B)Tailbud (stage 28) embryo, ventral view showing staining forXMLC2a(light blue) andXeHAND(purple). Note the thin line ofXeHANDexpression dividingthe two patches ofXMLC2a(black arrow). (C) Lateral view of a swimming tadpole (stage 31) showing staining forXMLC2a(light blue; black arrow) andXeHAND(reddish-brown). The arrowhead indicates that portion of the ventral surface that is not covered by mesoderm. (D) Feeding tadpole (stage 40) lateralview showing staining forXMLC2a(light blue) andXeHAND(purple).XMLC2astaining in the heart is indicated by a black arrow, andXeHANDstaining inthe ventral aorta and first aortic arches are indicated by the white arrow. The embryo was cleared with BB/BA. (E) Lateral view of a tailbud embryo (stage27) stained forT4 a-globin (purple) andXeHAND(light blue).
Table 1
The lateral domain ofXeHANDshows random asymmetry
Side of embryo with more intense expression
Left Right Equal Total
Stage 23–24 14 18 10 42Stage 27–28 56 44 24 124Total 70 62 34 166
Number of embryos with gut looping
Normal Reversed Total
Stage 46 53 6 59
Embryos were fixed at tailbud stages, analysed forXeHANDexpression bywhole-mount in situ hybridisation and scored for sidedness of staining. Asa control, embryos from the same batch were allowed to develop intofeeding tadpoles (stage 46) and scored for the direction of gut looping.
Table 2
The direction of embryo bending within the vitelline membrane does notaffect the asymmetry of expression ofXeHAND
Direction ofembryo bending
Side of embryo with more intense expression
Left Right Equal Total
Left 19 18 7 44Right 20 14 10 44
Embryos were sorted for direction of bending within the vitelline mem-brane at the tailbud stage (stage 25), membranes were removed manuallyand embryos allowed to straighten. Embryos were then either fixed im-mediately, or allowed to develop for various times before fixing.XeHANDexpression was analysed by whole-mount in situ hybridisation and scoredfor sidedness of staining.
157D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163

both XeHANDand the myocardial muscle-specific marker,XMLC2a(Fig. 5B–D). At stage 28,XMLC2awas expressedin the two bilaterally symmetrical patches of the heart pri-mordium, on either side of the midline (light blue), sepa-rated by a thin stripe ofXeHAND expression (purple).XeHAND expression was also evident both anterior andposterior to theXMLC2a expression, in regions that formthe ventral aorta and sinus venosus.
By swimming tadpole stages, expression ofXeHANDwithin the central portion of the heart region was reduced(Fig. 3G) and staining was largely confined to tissue sur-rounding the developing heart (Fig. 5C). Staining anterior tothe heart primordium corresponded to the ventral aorta andthe aortic arches (Fig. 3G, white arrow), whilst the regionimmediately posterior to the heart was the sinus venosus(Fig. 3G, black arrow). Overstaining of embryos at thesestages also revealed a residual level ofXeHANDexpressionin the heart itself (Fig. 3H, white arrow). In later embryos,staining for XeHAND was detected in the ventral aorta,extending anterior from the heart (Fig. 5D, white arrow).
2.6. XeHAND expression is asymmetric in the formingheart
To determine the precise location of transcripts in thisregion, tailbud embryos (stage 33/34) stained in whole-mount were sectioned in the transverse plane (Fig. 4B–E).Most anteriorly, at the level of the eye vesicles, transcriptswere detected in two patches in the head mesenchyme oneither side of the foregut (Fig. 4B). This expression domaincorresponded to the first pair of aortic arches, which form atstage 31 from the ventral endocardial tube (Nieuwkoop andFaber, 1994). In sections moving progressively more poster-ior, the patches of expression appeared to move towards theventral surface of the embryo, and formed two horizontallines, separated by the lumen of the ventral aorta (Fig. 4C,the lumen of the ventral aorta indicated by the black arrow).At the level of the anterior pericardial cavity (Fig. 4D)expression levels were lower, and could be seen throughoutthe dorsal pericardium (black arrow). More posterior sec-tions (Fig. 4E) revealed that expression in the dorsal peri-cardium persisted (black arrow), but low levels ofexpression were also seen throughout the pericardium. Inaddition, prominent staining was evident in the left side ofthe myocardial layer surrounding the endocardial tube Fig.4E, white arrow; higher magnification shown in Fig. 4G).The restriction ofXeHANDexpression to the left-hand sideof the myocardium surrounding the endocardial tube and itsabsence from the endocardium is reminiscent of the expres-sion of eHAND in the mouse (Biben and Harvey, 1997;Srivastava et al., 1997) but contrasts with the distributionreported for chick embryos (Srivastava et al., 1995).
2.7. XeHAND expression is maintained in the adult heart
MouseeHAND has been detected in a wide variety of
adult tissues including liver, smooth muscle of the gut (Hol-lenberg et al., 1995), brain, testis, skeletal muscle (Cross etal., 1995); intestine and adrenal tissue (Cserjesi et al., 1995).Interestingly, despite its early expression in the embryonicheart tube, no transcripts have been detected in the adultheart. In contrast,XeHAND was highly expressed in theadult frog heart (Fig. 6, lane 9). Lower levels were alsofound in intestine and gall bladder (Fig. 6, lanes 4 and 6).Transcripts were undetectable in liver, spleen, stomach,lung or skeletal muscle, even after prolonged exposure ofthe autoradiograms.
2.8. XeHAND is activated by BMPs and FGF-2 in embryoexplants
The bone morphogenetic proteins, BMP-2 and BMP-4are thought to play an important role in dorso-ventral pat-terning in the early embryo. Both will induce ventral andposterior mesoderm markers in animal cap explants and inthe embryo their ventralising activity is antagonised by sig-nals from the organiser, such as noggin and chordin(reviewed in Graff, 1997).
In tailbud embryos, transcripts of bothBMPs are loca-
Fig. 6. Expression ofXeHAND in adult frog tissues. The distribution ofXeHANDmRNA in adult frog tissues was analysed by RNase protectionassay. Lane 1, undigested probes; lane 2, tRNA control; lanes 3–11, tad-pole embryo (stage 34), intestine, stomach, gall bladder, spleen, skeletal,heart, lung, liver RNA, respectively. Five micrograms of total RNA wasused in each assay. Full-length protected fragments for each probe areindicated. As an internal control, a probe for the highly abundantEF1-awas included in theXeHANDassay.
158 D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163
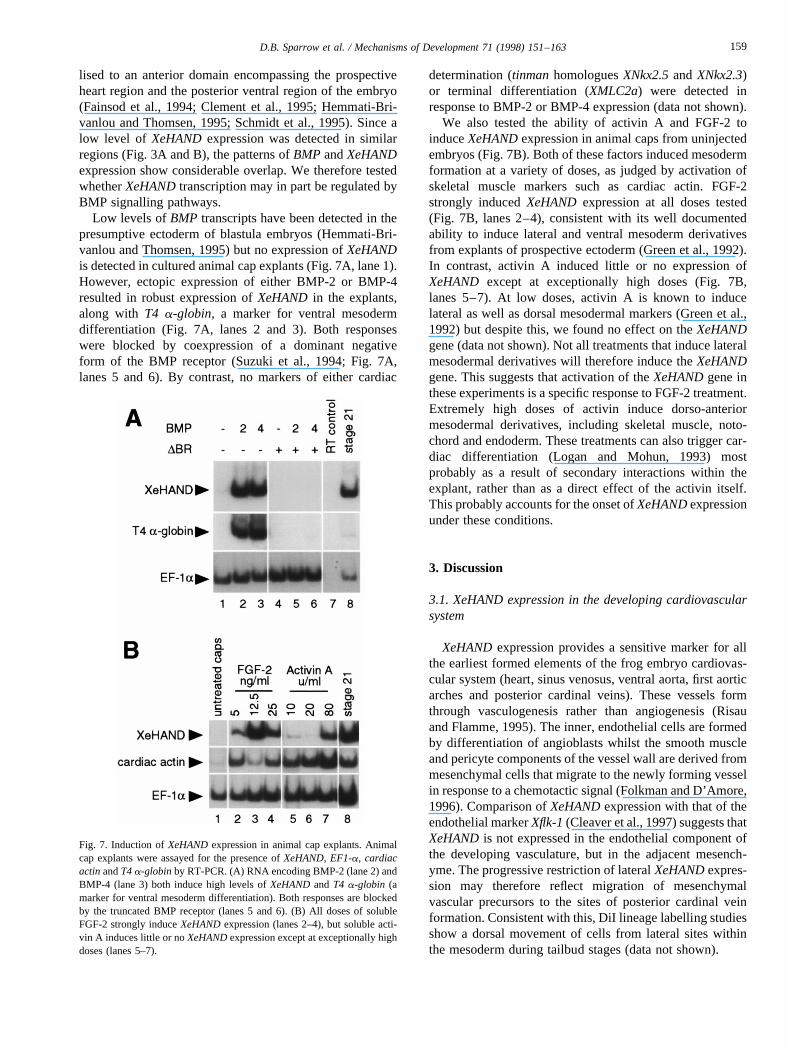
lised to an anterior domain encompassing the prospectiveheart region and the posterior ventral region of the embryo(Fainsod et al., 1994; Clement et al., 1995; Hemmati-Bri-vanlou and Thomsen, 1995; Schmidt et al., 1995). Since alow level of XeHANDexpression was detected in similarregions (Fig. 3A and B), the patterns ofBMP andXeHANDexpression show considerable overlap. We therefore testedwhetherXeHANDtranscription may in part be regulated byBMP signalling pathways.
Low levels ofBMP transcripts have been detected in thepresumptive ectoderm of blastula embryos (Hemmati-Bri-vanlou and Thomsen, 1995) but no expression ofXeHANDis detected in cultured animal cap explants (Fig. 7A, lane 1).However, ectopic expression of either BMP-2 or BMP-4resulted in robust expression ofXeHAND in the explants,along with T4 a-globin, a marker for ventral mesodermdifferentiation (Fig. 7A, lanes 2 and 3). Both responseswere blocked by coexpression of a dominant negativeform of the BMP receptor (Suzuki et al., 1994; Fig. 7A,lanes 5 and 6). By contrast, no markers of either cardiac
determination (tinmanhomologuesXNkx2.5andXNkx2.3)or terminal differentiation (XMLC2a) were detected inresponse to BMP-2 or BMP-4 expression (data not shown).
We also tested the ability of activin A and FGF-2 toinduceXeHANDexpression in animal caps from uninjectedembryos (Fig. 7B). Both of these factors induced mesodermformation at a variety of doses, as judged by activation ofskeletal muscle markers such as cardiac actin. FGF-2strongly inducedXeHAND expression at all doses tested(Fig. 7B, lanes 2–4), consistent with its well documentedability to induce lateral and ventral mesoderm derivativesfrom explants of prospective ectoderm (Green et al., 1992).In contrast, activin A induced little or no expression ofXeHAND except at exceptionally high doses (Fig. 7B,lanes 5–7). At low doses, activin A is known to inducelateral as well as dorsal mesodermal markers (Green et al.,1992) but despite this, we found no effect on theXeHANDgene (data not shown). Not all treatments that induce lateralmesodermal derivatives will therefore induce theXeHANDgene. This suggests that activation of theXeHANDgene inthese experiments is a specific response to FGF-2 treatment.Extremely high doses of activin induce dorso-anteriormesodermal derivatives, including skeletal muscle, noto-chord and endoderm. These treatments can also trigger car-diac differentiation (Logan and Mohun, 1993) mostprobably as a result of secondary interactions within theexplant, rather than as a direct effect of the activin itself.This probably accounts for the onset ofXeHANDexpressionunder these conditions.
3. Discussion
3.1. XeHAND expression in the developing cardiovascularsystem
XeHANDexpression provides a sensitive marker for allthe earliest formed elements of the frog embryo cardiovas-cular system (heart, sinus venosus, ventral aorta, first aorticarches and posterior cardinal veins). These vessels formthrough vasculogenesis rather than angiogenesis (Risauand Flamme, 1995). The inner, endothelial cells are formedby differentiation of angioblasts whilst the smooth muscleand pericyte components of the vessel wall are derived frommesenchymal cells that migrate to the newly forming vesselin response to a chemotactic signal (Folkman and D’Amore,1996). Comparison ofXeHANDexpression with that of theendothelial markerXflk-1(Cleaver et al., 1997) suggests thatXeHANDis not expressed in the endothelial component ofthe developing vasculature, but in the adjacent mesench-yme. The progressive restriction of lateralXeHANDexpres-sion may therefore reflect migration of mesenchymalvascular precursors to the sites of posterior cardinal veinformation. Consistent with this, DiI lineage labelling studiesshow a dorsal movement of cells from lateral sites withinthe mesoderm during tailbud stages (data not shown).
Fig. 7. Induction ofXeHANDexpression in animal cap explants. Animalcap explants were assayed for the presence ofXeHAND, EF1-a, cardiacactinandT4a-globinby RT-PCR. (A) RNA encoding BMP-2 (lane 2) andBMP-4 (lane 3) both induce high levels ofXeHANDandT4 a-globin (amarker for ventral mesoderm differentiation). Both responses are blockedby the truncated BMP receptor (lanes 5 and 6). (B) All doses of solubleFGF-2 strongly induceXeHANDexpression (lanes 2–4), but soluble acti-vin A induces little or noXeHANDexpression except at exceptionally highdoses (lanes 5–7).
159D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163

In the mouse embryo,eHAND expression has beenreported in a variety of neural crest derivatives, such asthe sympathetic trunk ganglia, pharyngeal arches, branchialarches, the septal region of the tongue and the adrenalmedulla. Interestingly, we have not detected by in situhybridisation any comparable expression ofXeHANDtran-scripts in neural crest-derived structures of theXenopusembryo. Taken together with the distribution ofeHANDtranscripts in the chick embryo, it is evident that there isconsiderable variation ineHAND expression patternsbetween vertebrates.
3.2. Asymmetry of XeHAND expression
Perhaps the most intriguing aspect ofXeHANDexpres-sion is the dramatic asymmetry in its initial expression in thetailbud embryo. In contrast to the later left-sidedness in themyocardium, the initial sidedness within lateral mesodermappears to be completely random. We know of no precedentfor such a pattern, which commences many hours after theleft/right signalling pathway is known to be established(Hyatt et al., 1996). One possible explanation is to invokea small, random disparity in the relative developmentalstage ofXeHAND expressing cells on each side of theembryo. If these cells are indeed vascular precursors, thena similar random asymmetry may be expected with othermolecular markers for this cell type. Furthermore, embryostransgenic for a lineage reporter (such as GFP) under thecontrol of theXeHAND gene regulatory elements shouldalso show a similar-sided asynchrony in transgene expres-sion.
3.3. Regulation of XeHAND expression
Embryo explant experiments demonstrate thatXeHANDcan be strongly activated in animal cap explants by theventralising growth factors BMP-2 and BMP-4. Is this anindirect consequence of the induction of lateral/ventralmesoderm derivatives, or does it indicate a more intimateregulatory relationship between theXeHANDgene and theBMP signalling pathway? Recent studies of mutant heartphenotypes in zebrafish strongly suggest that BMP-4 liesupstream ofeHAND in a regulatory hierarchy that controlsasymmetric cardiac morphogenesis (see below). In frogs,this relationship can now be explored using the explantassay. It is possible for example to examine whetherXeHAND isactivated in response to putative mediators ofBMP activity, such as the homeobox factor,Xom(Ladher etal., 1996).
In chick embryos, BMPs have also been shown to inducecardiogenic differentiation and it has been proposed that thecardiogenic crescent arises as a result of these and the antag-onistic effects of BMP regulators (such as noggin) on ante-rior mesoderm (Schultheiss et al., 1997). It is not yet knownif BMPs play a similar role in other vertebrates. InXenopus,cardiac fate is determined during gastrulation (Sater and
Jacobson, 1989, 1990a,b; Nascone and Mercola, 1995)and there is currently no evidence to indicate a role forBMPs in this process. After neurulation, BMP-2 andBMP-4 transcripts are expressed in overlapping domainsthat encompass the heart primordium (Fainsod et al.,1994; Clement et al., 1995; Hemmati-Brivanlou and Thom-sen, 1995; Schmidt et al., 1995), suggesting a role for thesefactors in heart morphogenesis. Their precise distributionshave not been compared in embryo sections and we do notknow if they have similar or distinct roles in cardiogenesis.Differences in expression become evident in tadpole stages,with BMP-4 expression becoming confined largely to theventral side of the branchial arches rather than the heart tubeitself (Hemmati-Brivanlou and Thomsen, 1995). This laterexpression pattern is remarkable similar to that ofXeHANDin the ventral aorta (compare Fig. 5D with Fig. 3J in Hem-mati-Brivanlou and Thomsen, 1995).
Cardiac muscle differentiation can be induced in animalpole explants by treatment with extremely high doses ofactivin A (Logan and Mohun, 1993), cardiogenic differen-tiation probably resulting from secondary inductive interac-tions between anterior character mesoderm and endodermformed within the explant (Wright et al., 1989; Sasai et al.,1996). Induction ofXeHANDunder these conditions is notdue to formation of lateral mesoderm since lower doses ofactivin fail to induce theXeHANDgene. Rather, it reflectsthe induction of cardiogenic differentiation in the explants.Simultaneous treatment of animal caps with activin andBMPs neither increases the extent of cardiogenic differen-tiation at high activin doses nor extends the activin doserange which will activate cardiac markers (Mohun andSparrow, unpublished data), indicating differentiation perse is not dependent on BMP signalling. If so, these datasuggest two conclusions. Firstly,XeHANDexpression dur-ing cardiogenesis is regulated by a second signalling path-way, independent of the BMPs; secondly, BMPs in the heartprimordium may regulate cardiac morphogenesis ratherthan terminal differentiation.
3.4. eHAND expression and cardiac laterality
Heart looping provides a convenient marker for the ana-lysis of laterality in the vertebrate embryo, but we do notknow how this asymmetric morphogenetic process is regu-lated. Antisense experiments in the chick indicate a role fortheHAND genes in the onset of heart tube looping but theirprecise function is unclear. Neither is apparently expressedasymmetrically within the heart tube and looping is onlyblocked by simultaneous inhibition of both genes (Srivas-tava et al., 1995). In contrast,eHAND expression in themouse embryo provides the first marker for the future direc-tion of heart looping. The caudal domain ofeHANDexpres-sion is initially bilateral and symmetrical within the linearheart tube, but it becomes predominantly left-sided in themyocardium and sinus venosus prior to the onset of right-ward looping. As a result,eHAND expression becomes
160 D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163

restricted largely to the atrium and left ventricle of thechambered heart. These results have prompted the sugges-tion that activity of theeHANDtranscription factor mediatesthe influence of embryo laterality on cardiac morphogenesis(Biben and Harvey, 1997; Srivastava et al., 1997).
Strikingly, in the zebrafishBMP-4 is expressed in a simi-lar fashion. Like in the mouse (Biben and Harvey, 1997), thezebrafish linear heart tube first ‘jogs’ to the left then loops tothe right (Chen et al., 1997). Although initially expressedsymmetrically in the myocardium,BMP-4 expressionbecomes predominantly left-sided immediately prior to theleftward ‘jog’. However, ifBMP-4 expression is renderedmore symmetric by overexpression, or if its signalling isinterrupted by injection of a truncated receptor, the leftward‘jog’ is blocked, and subsequent looping is disrupted. Like-wise in mutants where the heart laterality is reversed and theheart ‘jogs’ to the right and loops to the left,BMP-4 isexpressed predominantly on the opposite (right-hand) sideof the myocardium to the wild-type embryos. Furthermore,in embryos in which the heart fails to ‘jog’ at all,BMP-4expression is symmetrical throughout the myocardium.
Our findings of left sidedXeHANDexpression in the tad-pole myocardium and that BMPs can activateXeHANDexpression link these two observations and is consistentwith BMP-4being upstream ofHAND genes in the processof heart looping. If this is the case, it is interesting to notethat in the zebrafish other markers of cardiogenic determi-nation such asMEF-2and thetinmanhomologueNkx2.5aresymmetrically expressed throughout the heart looping pro-cess, a result consistent with our failure to induce thesemarkers by BMP expression inXenopus animal capexplants. The molecular cues that establish left/right pat-terning in the embryo appear to be common amongst verte-brates (King and Brown, 1997) and in amphibians, these canbe disrupted experimentally in several ways (Danos andYost, 1995, 1996). It should therefore be possible to testthe effect of such perturbations on the sidedness ofXeHAND expression within the tadpole myocardium. Inaddition, we are currently investigating whetherBMP-2and BMP-4 are asymmetrically expressed in theXenopusembryo.
4. Experimental procedures
4.1. Yeast two-hybrid screen
A cDNA library was made in yeast vector pGAD10 frompoly A+ RNA derived from adultXenopusheart tissue usingthe Clontech MATCHMAKER kit according to the manu-facturer’s instructions. A library of approximately 2× 107
independent clones with an average insert size of approxi-mately 1.0 kb was generated, using a combination of oligodT and random priming. A bait for the yeast two-hybridscreen (XE12-pGBT9) was constructed by fusing codons507–658 encompassing the bHLH and carboxy terminal
domains ofXenopusE 12 (Rashbass et al., 1992) in frameto the Gal4 DNA binding domain.
The yeast strain YN166 (MATa gal4 gal80 trpl-901ade2–101 leu2–3, –112+ URA::GAL → LacZ HIS3::GAL(UAS) → Ura3), a gift of Steven Elledge (Baylor Col-lege of Medicine), was transformed with XE12-pGBT9 andgrown in SD-Trp medium to an OD600 of 0.4–0.6. 250 ml ofculture was transformed with 200mg of Xenopusadult heartcDNA library and 10 mg of salmon sperm carrier DNA bythe lithium acetate method (Schiestl and Gietz, 1989) withdimethylsulfoxide addition to 10% final concentration (Hillet al., 1991). For maximum transformation efficiency, thetransformation mixture was divided into 10 aliquots of 1.5ml for the heat shock step. After 2 h recovery in SD-Trp-Leumedium, the transformants were plated on medium to selectfor uracil prototrophy (SD-Trp-Leu-Ura). Approximately7 × 105 independent cDNA clones were screened.
Transformants expressing theUra3 gene were tested forexpression of the second reporter gene (LacZ) by b-galac-tosidase filter assay (Breeden and Nasmyth, 1985). Libraryplasmids were recovered from double positive yeast colo-nies (Robzyk and Kassir, 1992) and sequenced. Two inde-pendent clones encoding theXenopus eHANDgene wereisolated. Both clones had the same 5′ end-point, and thelarger clone, pXeHAND-y1, contained a polyA tail.
4.2. Isolation of full-length XeHAND cDNA
A random-primed cDNA library was constructed usingRNA from stage 35Xenopusembryos and thelZapIIyvector (Stratagene). The library contained approximately106 independent clones with an average insert size ofapproximately 1.0 kb. This was screened with the entirecoding region from pXeHAND-y1PstD (nucleotides 257–761 of XeHAND cDNA sequence), at high stringency.Several positively hybridising cDNA clones were analysedand the entireXeHAND sequence was constructed as acomposite from one of these clones (pXeHAND-3.1)and pXeHAND-y1. The composite sequence was analysedusing DNA*y (DNASTAR) and BLAST (at the NCBIusing the BLAST network server: http://www.ncbi.nlm.-nih.gov/). TheXeHANDnucleotide sequence has been sub-mitted to the EMBL database under accession numberZ95080.
4.3. RNA preparation and assay
RNA was prepared from embryos and adult tissues asdescribed previously (Logan and Mohun, 1993).XeHANDtranscripts were detected with a probe consisting entirely of5′ UTR (nucleotides 1–220). This was synthesised usingEcoRI linearised template and T3 RNA polymerase.RNase protection assays were performed as described pre-viously (Chambers et al., 1994). Control probes forEF1a(Sargent and Bennett, 1990) andXMax2 (Tonissen andKrieg, 1994) were prepared as previously described.
161D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163

4.4. Whole-mount RNA in situ hybridisation
Albino embryos (stages 14–46) were used forRNA whole-mount in situ hybridisation as describedby Harland (1991), using digoxigenin-labelled probes.The antisenseXeHAND probe was derived fromNotIlinearised pXeHAND-y1PstID (nucleotides 257–761) byusing T3 RNA polymerase. A sense control probewas synthesised fromXhoI linearised template with T7RNA polymerase. A second anti-sense digoxygenin-labelled probe, consisting entirely of 5′ UTR sequence(nucleotides 1–220) was also used to confirm the specificityof the in situ expression pattern. Probes forT4 a-globin (Walmsley et al., 1994),XMLC1/3 (Theze et al.,1995), andXMLC2a (Chambers et al., 1994) were usedfor double-labelled in situ hybridisations. BCIP (Boehrin-ger-Mannheim), Magenta-Phos (Biosynth AG) or NBT/BCIP were employed for the chromogenic reactions. Forhistological analysis, fixed embryos were embedded in par-affin and 10mm sections photographed under Nomarskioptics.
4.5. Animal cap explant assays
Embryos were injected with 500–1000 pg of syntheticRNA at the one- or two-cell stage. Animal cap explantsfrom injected and uninjected embryos were dissected atthe blastula stage (stage 89) cultured overnight (Chamberset al., 1994) and harvested when sibling control embryosreached early tailbud stage (stage 20/21). Human recombi-nant activin A andXenopusFGF-2 were kindly provided byJim Smith (NIMR). Capped RNAs were transcribed (Cham-bers et al., 1994), using constructs previously described:BMP-4 (Dale et al., 1992); truncated BMP receptor (Suzukiet al., 1994) and aXenopusBMP-2 clone (Nishimatsu et al.,1992) in the transcription vector pSP64T (kindly providedby Les Dale).
4.6. Reverse transcription–polymerase chain reaction (RT-PCR)
RNA from 15–20 explants was treated with DNAse 1 andone-tenth used for random primed cDNA synthesis. One-tenth of each cDNA sample was used for PCR amplificationwith the following primer pairs:XeHAND: (28 cycles)ACCCCTGATTGTTCTATTTGCC and GAATACAAT-TCTGGCTCATTGC;EF1a: (25 cycles) CAGATTGGT-GCTGGATATGC and ACTGCCTTGATGACTCCTAG;T4 a-globin: (28 cycles) AGCTGCCAAGCACATCGAT-GAC and GTGAGCTGTCCTTGCTGATACC;cardiacactin: (28 cycles) CTGACAGAATGCAGAAGG andGTTGCTTGGAGGAGTGTG. Under these cycle condi-tions, positive signals fell within a 10–20-fold range ofreasonable linearity (data not shown).32P-labelled PCR pro-ducts were resolved on native 4% acrylamide gels andexposed overnight..
Acknowledgements
This work was supported by the Medical Research Coun-cil and the British Heart Foundation. We wish to thankSteve Sedgewick for help in setting up the yeast two-hybridscreen; Peter Vize, Ondine Cleaver, and Richard Harvey forhelpful discussions and communicating results before pub-lication; and Sally Dunwoodie for critical comments on thetext.
References
Biben, C., Harvey, R.P., 1997. Homeodomain factor Nkx2–5 controls left/right asymmetric expression of the bHLH geneeHandduring murineheart development. Genes Dev. 11, 1357–1369.
Bisbee, C.A., Baker, M.A., Wilson, A.C., Hadji-Azimi, I., Fischberg, M.,1977. Albumin phylogeny for clawed frogs (Xenopus). Science 195,785–787.
Breeden, L., Nasmyth, K., 1985. Regulation of the yeastHO gene. ColdSpring Harbor Symp. Quant. Biol. 50, 643–650.
Chambers, A.E., Logan, M., Kotecha, S., Towers, T., Sparrow, D.B.,Mohun, T.J., 1994. The RSRF/MEF2 protein SL1 regulates cardiacmuscle-specific transcription of a myosin lightchain gene inXenopusembryos. Genes Dev. 8, 1324–1334.
Chen, J.N., van Eeden, J.M., Warren, K.S., Chin, A., Nu¨sslein-Volhard, C.,Haffter, P., Fishman, M.C., 1997. Left-right pattern of cardiacBMP4may drive asymmetry of the heart in zebrafish. Development 124,4373–4382.
Cleaver, O., Tonissen, K.T., Saha, M.S., Krieg, P.A., 1997. Neovascular-ization of theXenopusembryo. Dev. Dyn. 210, 66–77.
Clement, J.H., Fettes, P., Kno¨chel, S., Lef, J., Kno¨chel, W., 1995. Bonemorphogenetic protein 2 in the early development ofXenopus laevis.Mech. Dev. 52, 357–370.
Cross, J.C., Flannery, M.L., Blanar, M.A., Steingrimsson, E., Jenkins,N.A., Copeland, N.G., Rutter, W.J., Werb, Z., 1995.Hxt encodes abasic helix-loop-helix transcription factor that regulates trophoblastcell development. Development 121, 2513–2523.
Cserjesi, P., Brown, D., Lyons, G.E., Olson, E.N., 1995. Expression of thenovel basic helix-loop-helix geneeHANDin neural crest derivatives andextraembryonic membranes during mouse development. Dev. Biol. 170,664–678.
Dale, L., Howes, G., Price, B.M., Smith, J.C., 1992. Bone morphogeneticprotein 4: a ventralizing factor in earlyXenopus development.Development 115, 573–585.
Danos, M.C., Yost, H.J., 1995. Linkage of left-right asymmetry and dor-sal-anterior development inXenopus. Development 121, 1467–1474.
Danos, M.C., Yost, H.J., 1996. Role of notochord in specification of car-diac left-right orientation in zebrafish andXenopus. Dev. Biol. 177, 96–103.
Fainsod, A., Steinbeisser, H., De Robertis, E.M., 1994. On the function ofBMP-4 in patterning the marginal zone of theXenopusembryo. EMBOJ. 13, 5015–5025.
Folkman, J., D’Amore, P.A., 1996. Blood vessel formation: What is itsmolecular basis?. Cell 87, 1153–1155.
Graff, J.M., 1997. Embryonic patterning: To BMP or not BMP, that is thequestion. Cell 89, 171–174.
Green, J.B., New, H.V., Smith, J.C., 1992. Responses of embryonicXeno-puscells to activin and FGF are separated by multiple dose thresholdsand correspond to distinct axes of the mesoderm. Cell 71, 731–739.
Harland, R.M., 1991. In situ hybridisation: An improved whole mountmethodforXenopzasembryos. In: Kay, B.K., Peng, H.B. (Eds.),Xeno-pus laevis:Practical Uses in Cell and Molecular Biology. AcademicPress, London, UK, pp. 658–695.
162 D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163

Hemmati-Brivanlou, A., Thomsen, G.H., 1995. Ventral mesodermal pat-terning inXenopusembryos: Expression patterns and activities of BMP-2 and BMP-4. Dev. Genet. 17, 78–89.
Hill, J., Donald, K.A., Griffiths, D.E., 1991. DMSO-enhanced whole cellyeast transformation. Nucleic Acids Res. 19, 5791.
Hollenberg, S.M., Sternglanz, R., Cheng, P.F., Weintraub, H., 1995. Iden-tification of a new family of tissue-specific basic helix-loop-helix pro-teins with a two-hybrid system. Mol. Cell. Biol. 15, 3813–3822.
Hyatt, B.A., Lohr, J.L., Yost, H.J., 1996. Initiation of vertebrate left-rightaxis formation by maternal Vg1. Nature 384, 62–65.
King, T., Brown, N.A., 1997. Embryonic asymmetry: Left TGFb at theright time?. Curr. Biol. 7, R212–R215.
Ladher, R., Mohun, T.J., Smith, J.C., Snape, A.M., 1996.Xom: a Xenopushomeobox gene that mediates the early effects of BMP-4. Development122, 2385–2394.
Logan, M., Mohun, T.J., 1993. Induction of cardiac muscle differentiationin isolated animal pole explants ofXenopus laevis embryos.Development 118, 865–875.
Lough, J., Barron, M., Brogley, M., Sugi, Y., Bolender, D.L., Zhu, X.,1996. Combined BMP-2 and FGF-4, but neither factor alone, inducescardiogenesis in non-precardiac embryonic mesoderm. Dev. Biol. 178,198–202.
Lyons, I., Parsons, L.M., Hartley, L., Li, R., Andrews, J.E., Robb, L.,Harvey, R.P., 1995. Myogenic and morphogenetic defects in the hearttubes of murine embryos lacking the homeobox geneNkx2–5. GenesDev. 9, 1654–1666.
Nascone, N., Mercola, M., 1995. An inductive role for the endoderm inXenopuscardiogenesis. Development 121, 515–523.
Nieuwkoop, P.D., Faber, J., 1994. Normal Table ofXenopus laevis(Dau-din). Garland, New York.
Nishimatsu, S., Suzuki, A., Shoda, A., Murakami, K., Ueno, N., 1992.Genes for bone morphogenetic proteins are differentially transcribedin early amphibian embryos. Biochem. Biophys. Res. Commun. 186,1487–1495.
Rashbass, J., Taylor, M.V., Gurdon, J.B., 1992. The DNA-binding proteinE12 co-operates with XMyoD in the activation of muscle-specific geneexpression inXenopusembryos. EMBO J. 11, 2981–2990.
Risau, W., Flamme, I., 1995. Vasculogenesis. Annu. Rev. Cell Dev. Biol.11, 73–91.
Robzyk, K., Kassir, Y., 1992. A simple and highly efficient procedure forrescuing autonomous plasmids from yeast. Nucleic Acids Res. 20, 3790.
Sargent, M.G., Bennett, M.F., 1990. Identification inXenopusof a struc-
tural homologue of theDrosophilagene snail. Development 109, 967–973.
Sasai, Y., Lu, B., Piccolo, S., De Robertis, E.M., 1996. Endoderm induc-tion by the organizer-secreted factors chordin and noggin inXenopusanimal caps. EMBO J. 15, 4547–4555.
Sater, A.K., Jacobson, A.G., 1989. The specification of heart mesodermoccurs during gastrulation inXenopus laevis. Development 105, 821–830.
Sater, A.K., Jacobson, A.G., 1990. The restriction of the heart morphoge-netic field inXenopus laevis. Dev. Biol. 140, 328–336.
Sater, A.K., Jacobson, A.G., 1990. The role of the dorsal lip in the induc-tion of heart mesoderm inXenopus laevis. Development 108, 461–470.
Schiestl, R.H., Gietz, R.D., 1989. High efficiency transformation of intactyeast cells using single-stranded nucleic acids as carrier. Curr. Genet.16, 339–346.
Schmidt, J.E., Suzuki, A., Ueno, N., Kimelman, D., 1995. Localized BMP-4 mediates dorsal/ventral patterning in the earlyXenopusembryo. Dev.Biol. 169, 37–50.
Schultheiss, T.M., Burch, J.B.E., Lassar, A.B., 1997. A role for bonemorphogenetic proteins in the induction of cardiac myogenesis. GenesDev. 11, 451–462.
Srivastava, D., Cserjesi, P., Olson, E.N., 1995. A subclass of bHLH pro-teins required for cardiac morphogenesis. Science 270, 1995–1999.
Srivastava, D., Thomas, T., Lin, Q., Kirby, M.L., Brown, D., Olson, E.N.,1997. Regulation of cardiac mesodermal and neural crest developmentby the bHLH transcription factor, dHAND. Nature Genet. 16, 154–160.
Suzuki, A., Thies, R.S., Yamaji, N., Song, J.L., Wozney, J.M., Murakami,K., Ueno, N., 1994. A truncated bone morphogenetic protein receptoraffects dorsal-ventral patterning in the earlyXenopusembryo. Proc.Natl. Acad. Sci. USA 91, 10255–10259.
Theze, N., Hardy, S., Wilson, R., Allo, M.R., Mohun, T., Thiebaud, P.,1995. The MLClf/3f gene is an early marker of somitic muscle differ-entiation inXenopus laevisembryo. Dev. Biol. 171, 352–362.
Tonissen, K.F., Krieg, P.A., 1994. Analysis of a variantMax sequenceexpressed inXenopus laevis. Oncogene 9, 33–38.
Walmsley, M., Guille, M.J., Bertwistle, D., Smith, J.C., Pizzey, J.A.,Patient, R.K., 1994. Negative control ofXenopusGATA-2 by activinand noggin with eventual expression in precursors of the ventral bloodislands. Development 120, 2519–2529.
Wright, C.V.E., Schnegelsberg, P., De Robertis, E.M., 1989. XlHbox 8: anovelXenopushomeo protein restricted to a narrow band of endoderm.Development 105, 787–794.
163D.B. Sparrow et al. / Mechanisms of Development 71 (1998) 151–163




















