
Antelope, Boone, Boyd, Brown, Burt, Cedar, Cherry, Colfax ...
Upload
khangminh22Category
view
2download
0
Louisiana State University Louisiana State University
LSU Digital Commons LSU Digital Commons
LSU Doctoral Dissertations Graduate School
2004
Developing embryo technologies for the eland antelope Developing embryo technologies for the eland antelope
(Taurotragus oryx) (Taurotragus oryx)
Gemechu G. Wirtu Louisiana State University and Agricultural and Mechanical College
Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_dissertations
Part of the Veterinary Medicine Commons
Recommended Citation Recommended Citation Wirtu, Gemechu G., "Developing embryo technologies for the eland antelope (Taurotragus oryx)" (2004). LSU Doctoral Dissertations. 505. https://digitalcommons.lsu.edu/gradschool_dissertations/505
This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Doctoral Dissertations by an authorized graduate school editor of LSU Digital Commons. For more information, please [email protected].

DEVELOPING EMBRYO TECHNOLOGIES FOR THE ELAND ANTELOPE(TAUROTRAGUS ORYX)
A DissertationSubmitted to the Graduate Faculty of the
Louisiana State University andAgricultural and Mechanical College
in partial fulfillment of therequirements for the degree of
Doctor of Philosophy
In
The Interdepartmental Program inVeterinary Medical Sciencesthrough the Department of
Comparative Biomedical Sciences
byGemechu Wirtu
D.V.M., Addis Ababa University, 1992M.S., Virginia Polytechnic Institute and State University, 1999
May 2004

ii
Acknowledgments
My dream to pursue graduate studies in the USA became a reality when my application for
the Fulbright Scholarship was miraculously approved in 1997. I am deeply indebted to all involved at
that critical stage. At Louisiana State University (LSU) and Audubon Center for Research of
Endangered Species (ACRES), I am grateful to those who were helpful in my getting admission to
LSU graduate school.
I am grateful to my graduate Committee Members: Drs. Charles Short, Earle Pope and
Robert Godke (Committee Co-chairs) and Drs. Barry Bavister, Leonard Kappel and Changaram
Venugopal (Committee members). I also thank Dr. James Cronin for valuable comments on my
dissertation. Drs. Pope, Bavister and Godke closely supervised aspects of my research. I am deeply
indebted to Dr. Betsy Dresser, Director of ACRES, for continued support and confidence in me.
Many people at ACRES had important inputs in my project. From the Veterinary Research
Unit, Dr. Susan Mikota (made me feel at home and had valuable inputs at the inception of the project
idea); Dr. Chris Janssen assisted with designing custom-made adaptor for the ultrasound transducer;
Barbara Vincent and Contessa Knight were always ready to offer their help to me. Dr. Alexander
Cole attended most of the eland procedures and I am thankful for his full collaboration. I also had
valuable discussions with Dr. Ellen Boyd, a fellow graduate student. Drs. Roberto Aguilar and Steve
Miller of the Audubon Institute also attended some of the oocyte retrieval procedures.
From Species Survival Center, I am especially grateful to Jeff Vaccaro and Erin Sarrat who
have been cooperative in all aspects of my project that required their involvement. I could not have
done the eland handling part of my project without their creative inputs and facilitative roles. Thank
you very much. I should also mention the valuable support I received from Carol Allen, Lola Curtis,
Dierdre Havnen and many interns and volunteers who assisted me over the last four years.
I received invaluable assistance from personnel from Dr. Pope and Dr. Bavister laboratories.
From Dr. Pope’s laboratory I would like to thank Amy King, Rebecca Harris, Dr. Martha Gomez,

iii
Lola Curtis, Angelica Giraldo, Cherie Dumas, Kia Gray and Liesl Nel-Themaat; and from Dr.
Bavister’s laboratory, Kimberly Poole, Dr. Jane Andrews, Dr. Byeongchun Lee, Inger Carlson,
Denise Kinsey and Stephanie Nichols. Dr. Philip Damiani also played crucial roles by doing the
nuclear transfer and intracytoplasmic sperm injection. He also was responsible for acquiring most of
the bovine oocytes that were used for the bovine embryo culture experiments.
Gwen Alleman, Celestine Washington, Josie Wolfe, Alicia Benoit, Stella Sullivan and Jackie
Coulon of ACRES are acknowledged for administrative and moral support. I also thank Dr. Stanley
Leibo of ACRES/University of New Orleans for allowing me access to his large collection of
reference materials.
From the Department of Comparative Biomedical Sciences (previously Veterinary
Physiology, Pharmacology and Toxicology), I thank Dr. Steven Barker, Coordinator of Graduate
Programs, and Mona Busby, Shelia Rigby and Jeanie Tyler for their administrative support. I am also
grateful to Dr. Ramaswamy for the friendship and helping me in many ways. Similarly, Dr. Asfaw
Bekele of Tarleton State University helped me with statistics and by sending many articles online.
Dr. Tadele Kiros of the University of Saskatchewan (Canada) also offered a similar assistance.
The work presented here was partly supported by grants from Johnston Science Foundation
and the Coypu foundation.
Many more people had extended their full support to me and the absence of your names from
list does not mean I am not grateful…Thank you all.
Finally, to my family and children: thank you for your patience and understanding during my
prolonged disappearances from your vicinity. In addition, I will always remain grateful to my
parents, who took the initial step of sending me to school by breaking the tradition of their parents
and community.

iv
Table of Contents
ACKNOWLEDGMENTS..................................................................................................................... ii
LIST OF TABLES ................................................................................................................................v
LIST OF FIGURES............................................................................................................................ v i i
ABSTRACT...................................................................................................................................... .viii
CHAPTER 1. INTRODUCTION ..........................................................................................................1
CHAPTER 2. LITERATURE REVIEW................................................................................................42.1. THE ROLE OF REPRODUCTIVE TECHNOLOGIES IN BIODIVERSITY CONSERVATION .........................42.2. GAMETE RETRIEVAL AND IN VITRO EMBRYO PRODUCTION IN FARM AND NONDOMESTIC
MAMMALS ...........................................................................................................................................62.3. ASSISTED REPRODUCTIVE TECHNOLOGIES IN THE COMMON ELAND ...........................................18
CHAPTER 3. DEVELOPMENT OF IN VITRO-DERIVED BOVINE EMBRYOS IN DEFINED(PROTEIN-FREE) MEDIA .................................................................................................................38
3.1. INTRODUCTION ...........................................................................................................................383.2. MATERIAL AND METHODS ..........................................................................................................393.3. RESULTS .....................................................................................................................................453.4. DISCUSSION ................................................................................................................................53
CHAPTER 4. HANDLING ELANDS FOR REPRODUCTIVE TECHNOLOGY PROCEDURESWITHOUT INDUCING GENERAL ANESTHESIA: ROLES OF BEHAVIORAL TRAININGAND HYDRAULIC CHUTE ..............................................................................................................60
4.1. INTRODUCTION ...........................................................................................................................604.2. MATERIALS AND METHODS ........................................................................................................624.3. RESULTS .....................................................................................................................................694.4. DISCUSSION ................................................................................................................................80
CHAPTER 5. TRANSVAGINAL ULTRASOUND-GUIDED OOCYTE COLLECTION, INVITRO OOCYTE MATURATION AND EMBRYO PRODUCTION IN THE ELANDANTELOPE .........................................................................................................................................88
5.1. INTRODUCTION ...........................................................................................................................885.2. MATERIALS AND METHODS ........................................................................................................905.3. RESULTS ...................................................................................................................................1035.4. DISCUSSION ..............................................................................................................................115
CHAPTER 6. SUMMARY AND CONCLUSIONS .........................................................................122
REFERENCES...................................................................................................................................126
APPENDIXA: ATTRIBUTES OF SPECIES OF TRAGELAPHINE ANTELOPES...............................................................155
B: ABBREVIATIONS..........................................................................................................................157
VITA ..................................................................................................................................................159

v
List of Tables
Table 2.1. List of mammalian species in which ART has been successful with offspring produced ..19
Table 2.2. Attempts to hybridize tragelaphine antelopes and the results .............................................27
Table 3.1. Design of Experiment 3.1....................................................................................................42
Table 3.2. Components and concentrations of modified KSOM, BM-3 and three groups of aminoacids...............................................................................................................................................43
Table 3.3. Development frequencies and cell count of in vitro-derived bovine embryos cultured inprotein-free BM-3 or mKSOM supplemented with two groups of amino acids ...........................46
Table 3.4. Blastocyst expansion and hatching frequencies of in vitro-derived bovine embryoscultured in protein-free BM-3 or mKSOM supplemented with two groups of amino acids.........46
Table 3.5. Effects of osmotic pressure of modified BM-3 and supplements on the development of ..48
Table 3.6. Effects of glucose, pyruvate, lactate and phosphate in high or low osmotic pressuremodified BM-3 on the frequency of cleavage (day 2) and development to morula (day 7) andblastocyst (day 9) stages of in vitro-derived bovine embryos cultured in protein-free media ......49
Table 3.7. Effects of glucose, pyruvate, lactate and phosphate in high or low osmotic pressuremodified BM-3 on the frequency (%) of blastocyst expansion (days 7 and 8) and hatching (day9) in vitro-derived bovine embryos ...............................................................................................50
Table 4.1. Mean interval (min) from conditioned sound cue to acceptance of handheld treats duringmorning and afternoon training sessions in elands........................................................................70
Table 4.2. Effect of sedative treatments on onset of sedation, frequency of recumbency or a need forsecond dosing of sedative agents in elands during handling and oocyte retrieval in a hydraulicchute. .............................................................................................................................................71
Table 4.3. Some physiological parameters of female elands during sedation and handling in ahydraulic chute for oocyte retrieval...............................................................................................75
Table 4.4. Some variations between elands with good (n = 5) or poor (n = 5) taming potential .........75
Table 5.1. Ovarian response and oocyte recovery rates in elands (n = 4) treated with Folltropin®-Vgiven 2 (single) or 4 and 2 (double) days before ultrasound-guided oocyte retrieval.................105
Table 5.2. Morphological distribution of cumulus oocyte complexes recovered from eland donors(n = 4) treated with Folltropin®-V for 2 (single) or 4 and 2 (double) days before oocyteretrieval........................................................................................................................................105
Table 5.3. Effect of targeting the preovulatory or postovulatory follicular wave on ovarianresponse to FSH treatment and oocyte recovery rates in elands (n = 6). ....................................106
Table 5.4. Effect of three factors on the in vitro maturation (meiotic status) of eland oocytesrecovered after Folltropin®-V-induced ovarian stimulation........................................................112

vi
Table 5.5. Development of eland oocytes subjected to three methods of in vitro production (IVP)of embryos...................................................................................................................................114

vii
List of Figures
Figure 3.1. Blastocyst development of in vitro-derived bovine embryos cultured in high osmoticpressure BM-3-20aa with different supplements. .........................................................................51
Figure 3.2. Blastocyst development of in vitro-derived bovine embryos cultured in low osmoticpressure BM-3-20aa with different supplements. .........................................................................52
Figure 3.3. Pathways of glucose metabolism & related factors influencing embryonic development 57
Figure 4.1. Sketch of the eland holding area and associated structures ...............................................63
Figure 4.2. Blood glucose (mean ± SD) and hematocrit (mean or single sample values) in sedatedelands handled in a hydraulic chute for oocyte collection (n = 18)...............................................73
Figure 4.3. Mean blood CPK level in sedated elands handled for oocyte retrieval (n = 18). ..............74
Figure 4.4. Heart rate (mean ± SD) profile during xylazine-butorphanol induced sedation in elandshandled for oocyte retrieval (n = 25).............................................................................................76
Figure 4.5. Blood oxygen saturation (mean ± SD) profile during xylazine-butorphanol-inducedsedation in elands handled for oocyte retrieval (n = 23). ..............................................................77
Figure 4.6. Respiratory rate (mean ± SD) profile during xylazine-butorphanol induced sedation inelands handled for oocyte retrieval (n = 23)..................................................................................78
Figure 4.7. Peripheral body temperature profile during xylazine-butorphanol induced sedation inelands handled for oocyte retrieval (n = 13)..................................................................................79
Figure 5.1. Estrous synchronization and ovarian stimulation treatment schedule before ultrasound-guided oocyte collection in elands in Experiment 5.1...................................................................92
Figure 5.2. Treatment schedules to target preovulatory or postovulatory follicular waves in elandsbefore conducting ultrasound-guided oocyte retrieval in Experiment 5.2a...................................94
Figure 5.3. Ultrasound transducer and a needle guide enclosed in the custom-made adaptor.............96
Figure 5.4. Effect of animal on follicles observed and oocytes recovered from eland donors. .........107
Figure 5.5. Mean number of follicles observed and oocytes recovered from eland donors duringeach month of the year. ...............................................................................................................108
Figure 5.6. Correlation between the number of follicles observed and oocytes recovered fromeland donors. ...............................................................................................................................109
Figure 5.7. Morphological appearance of eland oocytes recovered by ultrasound-guidedtransvaginal follicular aspiration .................................................................................................111
Figure 5.8. (A-J) Status of eland oocytes after applying IVM, IVF (A-H) or SCNT with gianteland somatic cells (I, J) ..............................................................................................................113
Figure 5.9. Day 7 blastocyst (400x) that developed after SCNT into an eland oocyte using acumulus cell from the same oocyte donor...................................................................................114

viii
Abstract
Assisted reproductive technologies developed in domestic cattle serve as a starting point in
similar studies on nondomestic bovids. The common eland is a useful model species for studies on
rare tragelaphine antelopes. In Chapter 3 of the present study, effects of components/attributes of
protein-free embryo culture media on the in vitro development of in vitro-derived bovine embryos
were evaluated. A 2 x 2 factorial study comparing effects of groups of amino acids (20aa or 11aa) in
two base media (modified KSOM or BM-3) demonstrated that amino acids and base medium
affected embryonic development. A subsequent 7 x 2 factorial experiment to evaluate effects of
osmotic pressure and supplement type in BM-3-20aa showed that embryonic development was
largely affected by supplements and identified glucose (0.2 mM) as a crucial supplement.
In Chapter 4, the use of behavioral training and handling of elands in a hydraulic chute to
perform transvaginal ultrasound-guided oocyte retrieval without inducing general anesthesia were
evaluated. Nine of 10 females associated specific sound cues with food treats. Females varied in their
response interval to audio cues and to training for voluntary entry into the chute. Handling elands for
oocyte retrieval required sedation and increased blood glucose levels.
In Chapter 5, type of estrous synchronization or ovarian stimulation protocol did not affect
ovarian response. Animals, but not month of the year, affected ovarian response. In 37 oocyte
retrieval procedures using seven females, an average of 12.8 follicles yielded 9.8 oocytes, of which
up to 73% matured to metaphase II. In vitro fertilization, intracytoplasmic sperm injection and
nuclear transfer resulted in embryonic development. In conclusion, the bovine embryo culture study
suggests that the beneficial effects of amino acids are influenced by the base medium and glucose
plays more important roles in non-ATP producing pathways. Behavioral training and handling of
sedated females in a hydraulic chute is a reliable method for collecting eland oocytes, which can
undergo in vitro maturation and some in vitro embryonic development.

1
Chapter 1. Introduction
Many of the fauna and flora of the earth are under increasing risk of extinction, mainly due to
loss of habitat because of human activities. According to the International Union for Conservation of
Nature and Natural Resources (IUCN), one-fourth of the approximately 5,000 species of mammals
are at risk of extinction. Extinction of mammals continues to be recorded as recently as the year
2000. For example, the last Pyrenean ibex (Capra pyrenaica pyrenaica) died in January 2000. In
spite of improved awareness about the need for protection of biodiversity by some sectors of the
society, the number of threatened species has continued to increase since 1996, when IUCN started
an organized collection of data. For mammals, the number of threatened species increased from
1,096 to 1,137 between 1996 and 2002 (IUCN 2002). With the ever-increasing human population
and its subsequent effect on the environment, more and more species are likely to suffer the
imminent threat of extinction.
General measures involving the protection of habitat are key to the conservation of
biodiversity; additionally, captive breeding is an important tool in the conservation of specific
species. Not only can captive breeding be used to re-establish wild populations of rare and
endangered species, it can also serve as “insurance” against extinction in case of natural disasters
(Sillero-Zubiri et al. 1997; Holt and Pickard 1999; Hildebrandt et al. 2002). However, several factors
could hinder captive breeding by natural mating (Lasley et al. 1994), including behavioral and
genetic incompatibility of mating pairs.
Assisted reproductive technologies (ART) offer viable alternatives for alleviating some
limitations encountered with natural breeding, including increased possibility for out crossing
different individuals beyond behavioral, spatial (geographical) and temporal restrictions (Ptak et al.
2002). In spite of the huge potential of ART in the propagation of threatened species and also the

2
maintenance of genetic diversity, the use of such technologies has not been investigated for most of
the species.
The limited application of ART can be attributed to a number of factors. First, except for
artificial insemination, the efficiency of most ART is low even in domestic animals, and results are
even lower when applying the technologies to nondomestic animals, for which basic information on
reproductive physiology is lacking or limited. Second, the numbers of endangered nondomestic
animals in captivity are scant; therefore, biological samples used to develop or apply ART are scarce.
Third, the behavioural predisposition of most large nondomestic mammals requires inducing general
anaesthesia, even for minor procedures. However, it is stressful to animals and technically not
feasible to induce general anaesthesia at short/frequent intervals, as required for ART procedures.
Thus, for ART to have a positive impact on the conservation of biodiversity, available
techniques in domestic animals should be improved. This will subsequently improve the prospect of
success when the techniques are applied to nondomestic species, although similar efficiencies should
not be expected. Furthermore, to effectively use nondomestic animals available in captivity, methods
should be developed that minimize the stress of handling while performing ART procedures.
The common eland (Taurotragus oryx), one of the two largest African antelopes, is closely
related to eight species of spiral-horned antelopes belonging to two genera: Tragelaphus and
Taurotragus. Both genera contain at least 18 subspecies, of which three are endangered. Even the
common eland itself may be threatened with extinction in the near future as many experts have
expressed concern that the population in their natural habitat is declining rapidly. Nevertheless,
compared with other spiral-horned antelopes, the common eland is well represented in captivity.
Moreover, baseline information is available to demonstrate the feasibility of developing ART
in this species for application in other spiral-horned antelopes. The eland has been shown to be
capable of maintaining pregnancy and producing offspring following interspecies transfer of bongo
embryos (Dresser et al. 1985) and may be able to serve as a recipient of embryos from the

3
endangered giant eland. The reproductive patterns of tragelaphine antelopes also seem to be similar,
as demonstrated by inter-specific breeding and subsequent live births of hybrid offspring involving at
least five species (Benirschke et al. 1980).
Thus, the common eland can serve as a useful model species in which to study ART in
tragelaphine antelopes. Additionally, the domestic bovid is a valuable general model species for
development of ART such as in vitro embryo production in nondomestic bovids because economic
impetus and animal availability has allowed the technology to advance rather quickly. It is hoped that
eventually, ART developed in bovine can be applied successfully to the eland, and then to rare
tragelaphine antelopes. Accordingly, the present study consists of two primary sections. In the first
section (Chapter 3), domestic bovid embryos were used to optimize simple (defined) culture media
for their in vitro development. In the second part (Chapters 4 and 5), investigations were carried to
develop a minimally stressful and repeatable method of gamete (oocyte) collection in the eland. In
addition, aspects of eland behavior and the in vitro developmental potential of eland oocytes were
evaluated.

4
Chapter 2. Literature Review
2.1. The Role of Reproductive Technologies in Biodiversity Conservation
The definition and scope of ART varies among species. For example, the Centers for Disease
Control and Prevention define human ART as “all fertility treatments in which both egg and sperm
are handled”, including in vitro fertilization (IVF), gamete intra-fallopian transfer, zygote
intrafallopian transfer and intracytoplasmic sperm injection (ICSI). However, artificial insemination
(AI) and the use of superovulation treatments without a concomitant plan for retrieval of oocytes are
not defined as ART (SART-CDC 2003). In nondomestic animals and species of agricultural
importance, the term ART is used in a broader sense. For example, in the first two series of
international symposia on “ART and Genetic Management of Wildlife” that met in 2001 and 2002
(Omaha, Nebraska, USA), frequent reference to ART was made when referring to artificial
insemination, simple embryo transfer, in vitro maturation of oocytes or culture and preservation of
somatic cells for potential use in cloning. This disparity in the definition of ART in the human and
animals is a reflection of the different purposes served by the technology and the legal implications
of human ART.
Human interest in assisting the breeding of plants and animals is probably as old as the
history of domestication. However, the use and development of modern age assisted reproductive
technologies emerged subsequent to the invention of the microscope that enabled visualization of
gametes and embryos. Artificial insemination and embryo transfer are the first generation of such
technologies.
Spallanzani documented the first artificial insemination using birds and dogs in the 1780s
(Heape 1897; Foote 2002). Embryo transfer was first reported in 1890 in the rabbit (Heape 1890).
Subsequently, these two technologies have been successfully applied in a wide range of species. The
development of methods for cryopreservation of spermatozoa and, subsequently, of embryos and

5
oocytes starting in the 1940s has also enhanced the potential application of these technologies (see
(Betteridge 2003); and Table 2.1).
The applications and benefits of ART in domestic or nondomestic mammals are numerous.
Some examples are outlined below:
1. ability to salvage and propagate genetic material from postmortem animals and those
with physical or physiological conditions/problems, including individuals that are
geriatric, prepubertal, pregnant, incompatible with mate, seasonally anestrus, physically
disabled, or have reproductive tract pathologies;
2. capability of transporting gametes or embryos across national or international boundaries
for alleviating genetic depression and repopulating the natural habitat;
3. more efficient use of i) spermatozoa, including low quality samples, by using IVF or
micro-assisted fertilization (ICSI, zona drilling) and ii) oocytes from genetically valuable
females by in vivo collection and rescue of oocytes from the pool undergoing follicular
atresia;
4. to reduce risk of disease transmission, that could occur during natural breeding;
5. ability to control sex ratio of offspring; and
6. to improve knowledge of the biology of gametes/early embryos.
For nondomestic mammals, regulations restricting the shipment of live animals across
international borders make it difficult to maintain genetically viable populations of rare species at
facilities carrying out captive breeding. Even when legal shipment of live animals is possible, the
transport causes stress and deaths occur during transit. Behavioral and genetic incompatibility can
also preclude safe mating. The use of ART, such as artificial insemination and embryo transfer,
combined with cryopreservation of spermatozoa and embryos, “eliminates the problems of distance

6
and time as major obstacles in captive breeding programs” (Lasley et al. 1994; Rott 1996;
Hildebrandt et al. 2002).
In addition, when combined with in vitro production (IVP) of embryos, ART would
maximize the use of the large pool of male and female gametes that, otherwise, are “wasted” during a
lifespan of natural breeding. Thus, ART would economize the use of gametes for production of
offspring from endangered species. For example, theoretically, hundreds of calves can be produced
from a single domestic cow by using a combination of technologies such as ultrasound-guided oocyte
retrieval and in vitro maturation (IVM), IVF, embryo culture and embryo transfer (Hansel, 2003).
Similarly, an enormous number of offspring could be produced from a single individual using
somatic cell nuclear transfer (SCNT) and embryo transfer.
Early stage antral follicles (<1 mm) can be cultured in vitro to yield mature oocytes capable
of undergoing fertilization and producing offspring [see (Miyano 2003)]. Activation of primordial
follicles and understanding the mechanisms responsible for the atresia (apoptosis) of most precursor
germ cells [see (Alonso-Pozos et al. 2003)] may also enable use of the large pool of female germ
cells for ART. The possibility of in vitro production of both male and female germ cells/gametes
from embryonic stem cells (Hubner et al. 2003; Toyooka et al. 2003) also suggests that additional
tools for the propagation of rare species may become available.
2.2. Gamete Retrieval and In Vitro Embryo Production in Farm and NondomesticMammals
2.2.1. Methods of Gamete (Oocyte) Retrieval
Studies on in vitro gamete biology and the development of ART for mammalian species are
dependent on the availability of both male and female gametes. Gametes (spermatozoa and oocytes)
can be colleted postmortem or ante-mortem. The methods used in cattle have been reviewed (Gordon
1994). For domestic animals such as cattle, sheep, goats and pigs, slaughterhouse ovaries and testes
are readily available and this resource has been used extensively in the development of ART. In

7
addition, gonads from small companion animals, especially cats and dogs, are easily obtained from
local veterinary clinics and some animal shelters.
Semen can be collected from males of large domestic and nondomestic animals by using an
artificial vagina, masturbation or automasturbation, electroejaculation, transrectal massage of the
ampullae and accessory sex glands, collection during or after mating from the vagina or via a fistula
made in the region of the penile urethra (Crump and Crump 1994; Pope et al. 1997; Schmitt and
Hildebrandt 1998; Skidmore et al. 2001). Semen samples are also commonly collected from the
epididimydes and vasa deferentia of postmortem or castrated animals. Generally, spermatozoa are
more readily collected than oocytes in many mammalian species. This accessibility has facilitated the
propagation of selected paternal genotypes in economically important species, such as cattle.
Due to the visceral location of the ovaries, the in vivo recovery of follicular stage oocytes
requires approaches that allow for visualization of follicles. Thus, invasive methods such as
laparotomy, needle puncturing of ovaries that are positioned in the flank area by rectal manipulation,
or transvaginally by surgical approaches (colpotomy) have been used [reviewed in (Gordon 1994)].
In smaller animals, laparoscopic methods of collecting oocytes in situ may be the most efficient
method.
In large mammals, recent development of methods for real-time visualization of ovarian
follicles (and other structures) using ultrasonography has led to a relatively non-invasive approach
for collecting oocytes from live animals. Ultrasound-guided oocyte retrieval in domestic animals
(cattle) was first reported in the 1980s (Callesen et al. 1987; Pieterse et al. 1988). The methods were
based on similar developments in human reproductive technology (Gleicher et al. 1983). The
feasibility of applying transvaginal ultrasound guided aspiration to recover oocytes has subsequently
been demonstrated in other domestic species, including horses (Brück et al. 1992) and goats (Graff et
al. 2000).

8
The potential of ultrasound-guided oocyte retrieval, when combined with IVP, for
propagating large mammals can be demonstrated by the story of a cow from Auburn University:
“A 13-year-old Limousin cow ...was admitted to the IVF program in 1996.Her last calf had been born 4-years prior to her arrival. Over a 2-year period,284 eggs were collected in 53 attempts…(5.3 eggs per collection). ...IVFtechniques using semen from 13 different bulls...produced 81 transferableembryos ....resulting in 24 pregnancies. Twenty-one calves were born overthe 2-year-period.”(http://www.vetmed.auburn.edu/art/Description_IVF_Text.htm,accessed October 31, 2003).
This is in contrast to a maximum of two calves that can be produced from a healthy and fertile
domestic cow during two years by using AI or natural mating methods.
Transvaginal ultrasound-guided oocyte collection has also been applied in several
nondomestic large mammals. The list includes the addax, mountain bongo, African buffalo, common
eland, gaur, lowland gorilla, red hartebeest, llama, red deer, sable, tsessebe, wapiti, and Burchell’s
and Hartmann’s zebra (Armstrong et al. 1995; Loskutoff et al. 1995; Meintjes et al. 1997; Pope et al.
1998b; Asa et al. 1998; Brogliatti et al. 2000; Wirtu et al. 2002a; Berg and Asher 2003; Loskutoff et
al. 2004). This method has particular advantages as a method of in vivo oocyte recovery. It is rapid,
simple, accurate, relatively non-invasive and can be repeated frequently (Brück et al. 1992; Pieterse
et al. 1991). Using an ultrasound-guided approach, oocytes can also be recovered from pregnant
females (Meintjes et al. 1995; Cochran et al. 1998) or postpartum animals as early as day 5 after
parturition (Perez 2003). Moreover, it may be useful to ease restrictions on the import/export of
embryos derived from IVF, as the donors can be tested for diseases over an extended period of
gamete/embryo importation (Loskutoff et al. 1995). Other applications of ultrasound examination in
ART include early pregnancy diagnosis, in vivo intra-follicular gamete transfer and determination of
the physiological status of gonads and reproductive tracts.
The utility of ultrasound-guided method as a practical method for recovery of oocytes has
been demonstrated in several nondomestic artiodactyls (see above). However, only in the gaur and
wapiti have pregnancies been established and offspring born after IVF and transfer of embryos to

9
recipient females (Armstrong et al. 1995; Hammer et al. 2001; Berg and Asher 2003). While a few
other nondomestic embryos been produced in vitro from oocytes recovered by ultrasound-guided
approach, the efficiency is reduced, as compared with that seen in more, well-studied species,
because of the lack of information about in vitro requirements for oocyte maturation/embryo culture
and cryopreservation/processing of spermatozoa. Only improving the availability of nondomestic
animal gametes and embryos for basic and applied studies can narrow the information gap. Greater
accessibility to increased numbers of oocytes of nondomestic species is essential for accelerating
progress in this area.
While ultrasound-guided oocyte retrieval does have several advantages over other methods of
oocyte recovery in nondomestic artiodactyls, the ability to do multiple retrievals has been limited
because of the necessity to use general anesthesia. Not only is there an inherent risk to general
anesthesia, there is additional risk of physical injury during the induction. Furthermore, stress
associated with the induction of general anesthesia (additional handling, pen shifting, restricting
space) can compromise many aspects of animal physiology including reproduction (Haigh 2001).
Therefore, alternative approaches that allow for multiple oocyte collections with minimal stress and
reduced risk to animals and personnel need to be developed to allow more effective use of captive
nondomestic artiodactyls for developing methods for in vitro production of embryos.
2.2.2. Approaches to In Vitro Embryo Production
Subfertility due to reproductive tract pathologies can occur in animals with normal gonadal
function. Although artificial insemination and/or embryo transfer can be applied to treat some
subfertility conditions, other problems may be managed using the more recently developed embryo
technologies.
Moreover, during natural breeding, the male ejaculate contains millions of spermatozoa, of
which only one is required to fertilize each ovulated oocyte. In the absence of mating, spermatozoa in

10
the testis and accessory glands are lysed and phagocytized or lost in urine. Similarly, the ovaries of
fetal cattle contain about 3 million germ cells. Most of these undergo atresia, with about 200,000
(7%) remaining in the newborn calf. Still less than 1% of the germ cells present at birth are ovulated
[(Erickson 1966); see also (Gordon 1994; Hansel 2003)]. Even among the gametes that are
released/ovulated, only a fraction participates in fertilization and subsequent development.
Thus, the application of in vivo gamete recovery methods in combination with in vitro
embryo production techniques and/or cryopreservation would theoretically maximize the efficiency
of gamete use from an individual animal of interest. This is of special interest for basic and applied
studies of ART in rare and endangered species, in which the supply of gametes is scarce.
2.2.2.1. In Vitro Fertilization
In vitro fertilization is a simple form of IVP, in which male and female gametes are co-
incubated under conditions that would allow for sperm penetration and subsequent activation of the
oocyte, formation of pronuclei, syngamy and subsequent cleavage. Recorded attempts at in vitro
fertilization date back to 1878. However, M.C. Chang (Chang 1959) was the first to unequivocally
demonstrate successful in vitro fertilization followed by the birth of live offspring [see (Bavister
2002a)]. Since then, similar successes have been reported in a number of species. In vitro
fertilization, with subsequent production of offspring after the transfer of resulting embryos to
recipient females has been reported in at least 30 domestic and nondomestic mammalian species
(Table 2.1).
Although offspring have yet to be produced, the potential of in vitro fertilization has also
been demonstrated in many other species. These include artiodactyls such as greater kudu (Loskutoff
et al. 1995), mountain bongo (Pope et al. 1998b), addax (Hall-Woods et al. 1999), klipspringer
(Raphael et al. 1991) and common eland (Wirtu et al. 2002a), and non-artiodactylids such as leopard,
puma, cheetah, clouded leopard, jaguarondi, lion, black footed cat, snow leopard, jaguar, jungle cat

11
and zebra (Goodrowe et al. 1989; Miller et al. 1990; Meintjes et al. 1997; Pope 2000). However, low
frequencies of fertilization in nondomestic species using standard domestic animal IVF protocols
[e.g., (Winger et al. 1997; Kidson et al. 2000; Wirtu et al. 2002a)], indicates the critical need for
further investigations and the refinement of these technologies.
2.2.2.2. Intracytoplasmic Sperm Injection (ICSI)
Various approaches have been adopted to maximize fertilization rates in vitro. For example,
when quality or quantity of spermatozoa are low, micro-assisted fertilization by intracytoplasmic
sperm injection (ICSI) is often used. Offspring have been produced after transferring ICSI-produced
embryos in several species including cattle, sheep, horse, pig and cat (Goto et al. 1990; Catt 1996;
Cochran et al. 1998; Pope et al. 1998a; Martin 2000). This approach circumvents the inability of
spermatozoa to fertilize oocytes due to low sperm numbers or poor motility (Goto et al. 1990; Pope
et al. 1998a; Lacham-Kaplan et al. 2003). Moreover, complete structural integrity of the
spermatozoon is not required to achieve fertilization after ICSI (Ward et al. 1999).
Given that spermatozoa from most nondomestic artiodactylids have low viability after
cryopreservation and that large numbers of ejaculates are required to develop reliable
cryopreservation methods, ICSI appears to have higher potential than conventional IVF protocols for
propagation of rare artiodactylids. The success of IVF also depends on the ability to induce sperm
capacitation, which is considered a limiting factor for successful IVF in some species (example,
horses). However, sperm capacitation is not an absolute requirement to achieve successful
fertilization after ICSI (Galli et al. 2003b). ICSI could also be used to circumvent polyspermic
fertilization seen after IVF in species such as pigs (Kren et al. 2003).
Regardless of the potential, the application of ICSI in nondomestic mammals has so far been
limited, partly, due to the technical skill and expensive micromanipulation equipment that is
required. Nevertheless, three normal rhesus monkey offspring have been produced after ICSI (Chan

12
et al. 2000). Also, transferable jaguarundi (Herpailurus yaguarondi) embryos have been produced
using ICSI (Pope et al. 1998a).
2.2.2.3. Cloning
The basis for the current nuclear transfer developments in various mammalian species can be
traced back to the basic scientific questions raised by early investigators in the mid-1800s and early
1900s. The German zoologist, August Weismann (1834-1914) is credited for proposing the simple
and testable hypothesis that in vertebrates “the zygote contains all the genetic determinants to form a
complete individual”, and also “that genetic determinants of the zygote are divided when the egg
divides” (McKinnel 1985). Subsequently, other German scientists including Wilhelm Roux (1850-
1924) and Adolph Eduard Driesch (1867-1941) performed experiments to further test Weismann’s
hypothesis.
However, it was Hans Spemann (1869-1941) who conclusively demonstrated for the first
time that two blastomeres from a single vertebrate (salamander) embryo are capable of developing
into two complete larvae. Spemann constricted a zygote using a loop of hair such that one side of the
loop contained anuclear ooplasm and the other side contained nucleated ooplasm. After the nucleated
ooplasm developed to a 16-cell stage embryo, he loosened the loop and allowed a single blastomere
to pass to the anucleated side. Subsequently twin whole organisms developed. Thus, Weismann’s
theory that the genetic determinants for forming a complete individual are divided during subsequent
divisions of the zygote was disproved. Rather, totipotency was not only an attribute of the zygote but
is also a characteristic of individual blastomeres; nevertheless, Weismann’s hypothesis served as a
basis for the subsequent experiments.
With subsequent development of micromanipulation equipment, it became possible to
introduce a single cell (nucleus) into an oocyte (nuclear transfer). This led to the first cloning of a
higher vertebrate species (the frog) from embryonic cells (Briggs and King 1952). That individual

13
embryonic cells (blastomeres) are capable of developing into a whole organism has subsequently
been demonstrated in various species including lower vertebrates and mammals [reviewed in (Di
Berardino 1997) (see also Table 2.1 for a list of mammals cloned from embryonic cells)].
Further studies on totipotency using somatic cells have also led to the demonstration that
differentiated (adult) cells can be reprogrammed to direct embryonic development. Since the
groundbreaking report by the Roslin Institute (University of Edinburgh) group on the birth of lambs
after SCNT using cells from adult and fetal (day 26 old) cell lines (Wilmut et al. 1997), the
totipotency of mammalian somatic cells has been demonstrated in several species of domestic and
laboratory animals. The list, which is still growing, includes cattle, goat, pig, horse, mule, cat, rabbit,
mice and rat (see Table 2.1). It is interesting to note that some of the pioneers working in the area of
nuclear transfer believed in the “lack of totipotency of nuclei from adult organisms” until 1997
(McKinnel 1985; Di Berardino 1997). Nevertheless, in less than 10 years of the demonstration of the
totipotency of somatic cells, SCNT is already affecting many aspects of agriculture and biomedical
research.
Potential applications of SCNT in the propagation of endangered species are also emerging,
as demonstrated by offspring production in four nondomestic mammalian species: gaur, mouflon
sheep, banteng and African wildcat [(Lanza et al. 2000a; Ptak et al. 2002; Holden 2003)
(http://www.auduboninstitute.org/rcenter/res_cloning.htm, accessed February 22, 2004)]. In each of
these studies, a closely related domestic species served as the source of oocytes used for nuclear
transfer (interspecies somatic cell nuclear transfer, iSCNT) and as recipients of the resulting embryos
(interspecies embryo transfer, iET). These approaches are an extension of studies on the possibility
of embryonic development after iSCNT (Dominko et al. 1999) and of pregnancy and live births after
iET (Table 2.1). Some hope that iSCNT and iET will be utilized not only for the propagation of rare
species but also for reviving extinct species (http://www.chl.ca/cnewsscience0001/10_clone2.html,
accessed February 22, 2004).

14
However, the low success rates after SCNT, <4 % live birth for each micromanipulated
oocyte (Wilmut et al. 2002), or after iET (Loskutoff 1999) appear to be somewhat discouraging for
using these approaches in the propagation and genetic management of rare mammalian species. The
problem with iET could partly be due to maternal immune response to the embryo from the other
species (Kraemer 1983; Loskutoff 1999). Factors negatively affecting the success of SCNT include
failure of complete genomic reprogramming of the somatic cell or lack of its synchrony with the cell
cycle stage of the cytoplast ooplasm, and abnormal placentation and pregnancies after the transfer of
nuclear transfer embryos to recipient females (Wilmut et al. 2002). The presence of heterogeneous
cytoplasm and organelles from the cytoplast and the somatic cell further complicates interspecies
SCNT; however, steady improvements in the success rate of SCNT are being recorded.
Another form of cloning is embryo bisection. As described earlier, blastomere(s) of
vertebrate embryos are totipotent. This developmental capability has been applied to produce two or
more genetically identical siblings from a single embryo in several species including mice, sheep,
cattle, horse and monkeys [see (McKinnel 1985; Di Berardino 1997)]. In mammals, the first evidence
for the developmental potential of isolated blastomeres was provided in the rat (Nicholas and Hall
1942). Among nondomestic artiodactylids, embryo bisection has been successful in the common
eland (Gelwicks et al. 1989). The latter investigators recovered seven embryos from a single female
and non-surgically transferred bisected embryo-pairs to five female eland recipients, one of which
delivered a live calf. However, the application of this technology has been limited, in both domestic
and nondomestic mammals, mainly due to the less than expected number of offspring resulting
following the transfer of manipulated embryos to recipient females.
2.2.2.4. Gamete Transfer
Besides natural mating and artificial insemination, transferring oocytes into the oviduct and
subsequently inseminating the female (oocyte transfer), or transferring both gametes into the oviduct

15
(gamete intrafallopian transfer, GIFT) facilitates embryo production. Oocyte transfer and gamete
intrafallopian transfer have been successful in horses [see (Squires et al. 2003)] and pigs (Rath et al.
1994).
Direct deposition of both gametes into follicles before ovulation (intrafollicular gametes
transfer) or spermatozoa into dominant follicles have also led to the birth of human babies (Werner-
von der et al. 1993; Nuojua-Huttunen et al. 1995). Intrafollicular oocyte transfer can be done in cattle
(Bergfelt et al. 1998) and has led to the establishment of pregnancies in the horse [see (Hinrichs
1998)].
Thus, gamete transfer technologies have potential applications in propagation and genetic
management of rare mammalian species; however, to date, this potential has not yet been
investigated in nondomestic species.
2.2.2.5. Embryo Culture
Zygotes or early embryos produced by standard IVF or techniques involving
micromanipulation (ICSI, nuclear transfer) require a period of in vitro culture before transfer to a
recipient female for continued development. In most ungulates in which embryo transfer has been
successful, non-surgical uterine transfer of embryos is feasible and can be accomplished during
simple physical restraint. For successful uterine transfer, IVP embryos should be cultured to the
morula or blastocyst stage. Thus, in vitro culture (IVC) of embryos is an integral component of ART.
In domestic cattle, in vitro-cultured embryos display marked morphological, biochemical and
functional deviations when compared with those produced in vivo (Massip et al. 1995; Holm and
Callesen 1998; Niemann and Wrenzycki 2000; Thompson 2000). These anomalies are due, partly, to
inappropriate in vitro gamete/embryo handling and culture systems (Massip et al. 1995; Abe et al.
2002). Embryos cultured in media containing complex components, such as serum, display severe
abnormalities (Massip et al. 1995; Niemann and Wrenzycki 2000; Jacobsen et al. 2000; van

16
Wagtendonk-de Leeuw et al. 2000; Cho et al. 2002). Thus, many investigators are advocating the
development and use of simple and defined or semi-defined embryo culture media.
Many aspects of IVP require improvements. The longest phase of IVP is the culture of
embryos after IVF or SCNT, usually five to seven days. This long duration of IVC presents a primary
opportunity for improvements of the IVP system. Moreover, in addition to cell cleavage, key events
take place during IVC, including the maternal-embryonic transition of genetic control of
development, compaction and blastocyst differentiation. Each of these processes can easily be
disrupted in vitro.
Aspects of IVC that influence embryonic development include the lack of proper nutrients
and/or presence of potentially toxic products of external or embryonic origin (Bavister 1995;
Thompson 1996; Thompson 2000). Different laboratories use different protocols, even when working
on the same species, leading to confusion about what methods to use for IVC (Yang et al. 1993;
Bavister 1995; Gardner et al. 2000). The choice of a culture method(s) is often made empirically
rather than being based on scientific evidence. Evaluation of different culture systems under the same
conditions could provide additional information on requirements (or non-requirements) of the
embryo and on the method(s) to use when working on a new species for which information is not
available.
Undefined media containing serum or bovine serum albumin (BSA) as external sources of
protein are still widely used even though similar embryo development is seen in some species with
protein-free media containing amino acids (Bavister 1995; Biggers et al. 2000). Although it is well
established that serum and BSA can provide stimulatory factors that promote embryo development in
vitro, there are negative aspects to their use, including pathogen risk, lot-to-lot variation (Kane 1983;
Batt and Miller 1988; McKiernan and Bavister 1992), inhibition of oocyte maturation (Goodrowe et
al. 1991) or early cleavage (Pinyopummintr and Bavister 1991; Bredbacka and Bredbacka 1995) and
fetal/neonatal abnormalities (Young et al. 1998). In addition, the risk of pathogens of agricultural

17
and public health importance is minimized when using defined media than those containing
undefined components such as serum albumin or serum. Another important advantage to the use of a
defined medium is that the effect of each component on embryo metabolism and development can be
assessed. Then that information can be used to design media that will improve embryos
developmental potential.
Thus, there is a growing interest among many laboratories to use simple/defined media for
IVP in different species, including rodents (Whitten and Biggers 1968; Kane and Bavister 1988; Goh
et al. 2000), rabbits (Carney and Foote 1991), sheep (Ledda et al. 1992), goats (Keskintepe et al.
1997), primates (Schramm and Bavister 1996; Ali et al. 2000; Zheng et al. 2001), pigs (Yoshioka et
al. 2002) and cattle (Lonergan et al. 1994; Liu and Foote 1997; Caamano et al. 1998; Olson and
Seidel, Jr. 2000; Wrenzycki et al. 2001; Hernandez-Fonseca et al. 2002; Holm et al. 2002).
Amino acids have beneficial effects during in vitro culture of embryos of various mammalian
species. They play important roles as energy substrates, osmolytes, regulators of pH and enzyme
activity, scavengers of free radicals, chelators of heavy metals, precursors of macromolecules such as
proteins and nucleic acids, and mediators of transport across the cell membrane (Bavister 1995; Liu
and Foote 1995a; Lee and Fukui 1996). However, the beneficial effects may be restricted to certain
amino acids (Bavister 1995; Steeves and Gardner 1999) and may be affected by the base medium
(Van Winkle and Campione 1996). For example, Eagle’s essential amino acids inhibit the
development of early-stage cattle embryos (Keskintepe and Brackett 1996), possibly due to toxic
effects of ammonia production (Lane et al. 2001). Nevertheless, most laboratories use the mixture of
20 amino acids as originally developed for somatic cell culture. The total concentration in this system
is less than 6 mM, whereas free amino acids constitute ~32 and 44 mM in bovine oviductal and
uterine fluids, respectively (Elhassan et al. 2001).
Some studies have evaluated the effects of individual or groups of few amino acids during in
vitro culture [(see (Bavister 1995; Gardner et al. 2000; Thompson et al. 2000)]. It would be difficult

18
to conclusively identify the role(s) of each of the 20 amino acids and the effect of their interactions
and different concentrations on in vitro development of bovine embryos. A logical step would be to
evaluate the feasibility of protocols already developed in laboratory animals. For example, after
extensive evaluations of the effects of all amino acids during in vitro culture of hamster embryos in
chemically defined system, 11 beneficial amino acids were identified (McKiernan et al. 1995).
Osmotic pressure and/or high NaCl concentration of a medium can affect the development of
embryos in vitro. Lower osmotic pressure and a concomitant reduction in NaCl concentration is
partially responsible for the improved development of mouse embryos in the potassium simplex
optimized medium, KSOM (Summers et al. 1995). In most instances, increased osmotic pressure of
culture media is synonymous with a high concentration of NaCl. However, extracellular Na+ can
affect intracellular pH via its effect on the Na+/H+ anti-porter (Lane and Bavister 1999). Reducing
extracellular NaCl also alters gene expression (Ho et al. 1994) and stimulates protein synthesis (Liu
and Foote 1997). Osmotic stress could also activate the polyol pathway, thereby inducing apoptosis
(Galvez et al. 2003).
Culturing bovine embryos in media with >270 mOsmol was detrimental (Lim et al. 1994; Liu
and Foote 1996). Thus, base medium influences a number of embryonic attributes as well as the
effects of supplements. Moreover, the mechanisms by which cattle embryos produce energy at each
stage of preimplantation development could well be affected by the base medium.
2.3. Assisted Reproductive Technologies in the Common Eland
2.3.1. Overview of Tragelaphine Antelopes
Antelopes are a diverse group of species in the bovidae family. The term “antelope” does not
have taxonomic significance; however, about 100 of some 160 species of bovidae are traditionally
known by this name. Tragelaphine (spiral-horned) antelopes consist of two genera with nine species

19
Table 2.1. List of mammalian species in which ART has been successful with offspring produced________________________________________________________________________________________________________________Species Date ART typea Referencesb Comments________________________________________________________________________________________________________________
ArtiodactylaAddax 1987 CPs/AI (Densmore et al. 1987) AI done under manual restraintAlpaca 1966 AI Fernandez-Baca & Calderon [see (Pugh and Montes 1994)]
1974 ET Sumar and Franco [see (Del Campo et al. 1995)] Surgical collection and transfer2000 iET (Taylor et al. 2001) Llama recipient
Banteng 1983 iET Wiesner et al. (www.medicine.ucsd.edu/cpa/indxfs.html) Domestic cow recipient2002 CPs/AI (Johnston et al. 2002) Domestic cow recipient2003 iSCNT/iET Lanza et al. [see (Holden 2003)] Domestic cow recipient
Blackbuck 1988 AI (Holt et al. 1988) Under anesthesia1988 CPs/AI (Holt et al. 1988)
Bison (American) 1994 CPs/AI (Dorn 1995)1993 ET Foxworth [see (Dorn 1995)]
Bison (European) 1993 CPs/AI Sipko et al. [see (Rott 1995)]Buffalo (Asian) 1983 ET (Drost et al. 1983)
1990 CPs/ET (Misra et al. 1990)1991 IVF/ET see (Suzuki et al. 1992; Madan et al. 1994) After IVM/IVF/IVC
Bongo 1984 ET (Dresser et al. 1985)1984 iET (Dresser et al. 1985) Common eland recipient
Camel (Bactrian) 1990 CPs/AI Chen et al. [see (Loskutoff and Betteridge 1992)]Camel (Dromedary) 1992 ET (McKinnon et al. 1994)
1992 AI Annouassi et al. [see (Loskutoff and Betteridge 1992)]Cattle (domestic) ~1900 AIC Ivanow [see (Foote 2002)]
1951 CPs/AI (Stewart 1951)1951 ET (Willett et al. 1951)1973 CPe/ET (Wilmut and Rowson 1973)1982 IVF/ET (Brackett et al. 1982)1987 ECNT/ET (Robl et al. 1987; Prather et al. 1987)1990 ICSI/ET (Goto et al. 1990)1993 IVF/CP/ET (Zhang et al. 1993)1998 CPo/IVF/ET (Vajta et al. 1998)1998 SCNT/ET (Cibelli et al. 1998; Kato et al. 1998)
(table continued)
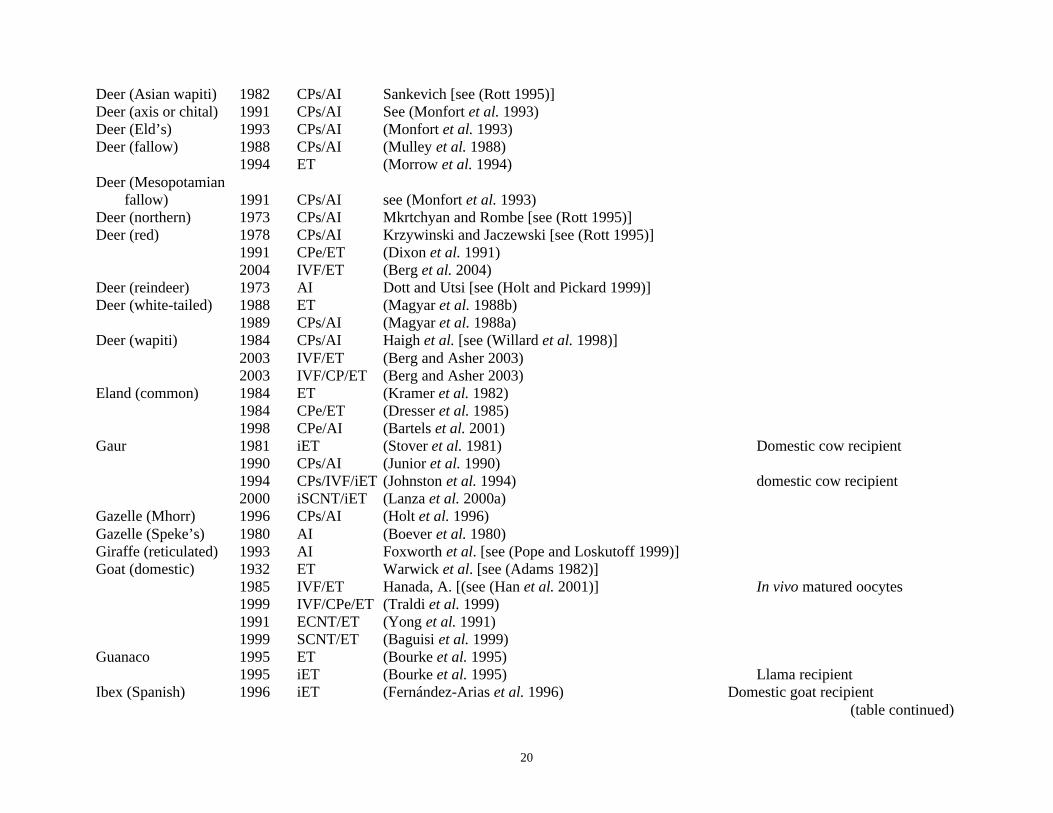
20
Deer (Asian wapiti) 1982 CPs/AI Sankevich [see (Rott 1995)]Deer (axis or chital) 1991 CPs/AI See (Monfort et al. 1993)Deer (Eld’s) 1993 CPs/AI (Monfort et al. 1993)Deer (fallow) 1988 CPs/AI (Mulley et al. 1988)
1994 ET (Morrow et al. 1994)Deer (Mesopotamian
fallow) 1991 CPs/AI see (Monfort et al. 1993)Deer (northern) 1973 CPs/AI Mkrtchyan and Rombe [see (Rott 1995)]Deer (red) 1978 CPs/AI Krzywinski and Jaczewski [see (Rott 1995)]
1991 CPe/ET (Dixon et al. 1991)2004 IVF/ET (Berg et al. 2004)
Deer (reindeer) 1973 AI Dott and Utsi [see (Holt and Pickard 1999)]Deer (white-tailed) 1988 ET (Magyar et al. 1988b)
1989 CPs/AI (Magyar et al. 1988a)Deer (wapiti) 1984 CPs/AI Haigh et al. [see (Willard et al. 1998)]
2003 IVF/ET (Berg and Asher 2003)2003 IVF/CP/ET (Berg and Asher 2003)
Eland (common) 1984 ET (Kramer et al. 1982)1984 CPe/ET (Dresser et al. 1985)1998 CPe/AI (Bartels et al. 2001)
Gaur 1981 iET (Stover et al. 1981) Domestic cow recipient1990 CPs/AI (Junior et al. 1990)1994 CPs/IVF/iET (Johnston et al. 1994) domestic cow recipient2000 iSCNT/iET (Lanza et al. 2000a)
Gazelle (Mhorr) 1996 CPs/AI (Holt et al. 1996)Gazelle (Speke’s) 1980 AI (Boever et al. 1980)Giraffe (reticulated) 1993 AI Foxworth et al. [see (Pope and Loskutoff 1999)]Goat (domestic) 1932 ET Warwick et al. [see (Adams 1982)]
1985 IVF/ET Hanada, A. [(see (Han et al. 2001)] In vivo matured oocytes1999 IVF/CPe/ET (Traldi et al. 1999)1991 ECNT/ET (Yong et al. 1991)1999 SCNT/ET (Baguisi et al. 1999)
Guanaco 1995 ET (Bourke et al. 1995)1995 iET (Bourke et al. 1995) Llama recipient
Ibex (Spanish) 1996 iET (Fernández-Arias et al. 1996) Domestic goat recipient (table continued)

21
Llama 1985 ET (Wiepz and Chapman 1985) Non-surgical collection and ET1977? AI Leyva et al. [see (Pugh and Montes 1994)]
Oryx (Scimitar horned) 1986 CPe/ET Schmitt, DL [see (Pope and Loskutoff 1999)] Calf stillborn1989 CPs/AI (Garland et al. 1992) Under general anesthesia1991 ET (Pope et al. 1991) Under general anesthesia
Pig 1950 ET Kvasnitski [see (Kvasnitski 2001)]1957 CPs/AI Hess et al. [see (Curry 2000)]1983 IVF/ET (Cheng et al. 1986)1989 CPe/ET (Hayashi et al. 1989)1989 ECNT/ET (Prather et al. 1989)2000 ICSI/ET (Martin 2000)2000 SCNT/ET (Onishi et al. 2000; Polejaeva et al. 2000)
Sheep (Armenian red) 1995 iET (Coonrod et al. 1994; Flores-Foxworth et al. 1995) Laparoscopic ET to domestic sheep1995 IVF/iET (Coonrod et al. 1994; Flores-Foxworth et al. 1995) Laparoscopic ET to domestic sheep
Sheep (big horn) 1986 CPs/AI Shaidulin and Roldugin [see (Rott 1995)]Sheep (Dall’s) 1990 ET (Buckrell et al. 1990)Sheep (domestic) 1933 ET Warwick et al [see (Adams 1982)]
1967 CPs/AI Salmon and Lightfoot [see (Curry 2000)]1984 IVF/ET (Cheng et al. 1986)1987 CP/ET (Heyman et al. 1987)1986 ECNT/ET (Willadsen 1986)1996 ICSI/ET (Catt 1996)1999 IVF/CPe/ET (Traldi et al. 1999)1997 SCNT/ET (Wilmut et al. 1997)
Sheep (mouflon) 1977 iET (Bunch et al. 1977) Domestic sheep recipient2001 CPe/iET (Naitana et al. 2000) Domestic sheep recipient2001 iSCNT/iET (Loi et al. 2001) Domestic sheep recipient2002 IVF/iET (Ptak et al. 2002) Domestic sheep recipient
Spring buck 1995 AI (Rott 1995)Suni antelope 1989 CPs/AI (Raphael et al. 1989) Transcervical AI; general anesthesia
1990 ET (Raphael et al. 1989; Loskutoff et al. 1990) Laparoscopic ET
CarnivoraAmerican black bear 1997 ET (Boone et al. 1997)Caracal 2000 IVF/ET (Pope et al. 2001)
2003 IVF/CP/ET Pope et al. (http://www.auduboninstitute.org/rcenter/res_cloning.htm)
(table continued)

22
Cat (African wild) 2000 IVF/CP/iET (Pope et al. 2000) Domestic cat recipient2003 iSCNT/iET Gomez et al. (www.auduboninstitute.org/rcenter/res_cloning.htm) Domestic cat recipient
Cat (Domestic) 1976 AI Platz et al. (Platz et al. 1978)1978 ET (Schriver and Kraemer 1978);1988 CPe/ET (Dresser et al. 1988)1988 IVF/ET (Goodrowe et al. 1988)1994 IVF/CP/ET (Pope et al. 1994)1995 ICSI/ET (Pope et al. 1998a)2002 SCNT (Shin et al. 2002)
Cat (Fishing) 2002 IVF/ET Dr. Earle Pope (Unpublished)Cat (Indian desert) 1989 IVF/iET (Pope et al. 1989) Domestic cat recipientCat (Serval) 2003 IVF/ET Dr. Earle Pope (Personal communication)Cat (Linking) 1989 iET Liansheng et al. [(see (Rott 1996)] Domestic cat recipientCheetah 1992 AI (Howard et al. 1992)
2000 CPs/AI (Howard et al. 1997)Dog (domestic) 1770 AI Spallanzani [see (Heape 1897)]
1979 ET (Kinney et al. 1979)Ferret (black footed) 1991 CPs/AI (Howard et al. 1991)Ferret (common) 1968 ET (Chang 1968)
1991 CPs/AI see (Wildt et al. 1992)Ferret (Siberian) 1991 CPs/AI see (Wildt et al. 1992)Fox (arctic or blue) 1972 CPs/AI Aamdal et al. [see (Rott 1995)] Both surgical and non-surgical AILeopard cat 1991 AI Howard and Doherty, [see (Swanson 2001)]
? CPs/AI See (Swanson 2001)Leopard (clouded) 1996 AI Howard et al. [see (Swanson 2001)]Mink 1975 ET (Adams 1982)Ocelot 1997 CPs/AI See (Swanson 2001)
2000 IVF/CPe/ET See (Swanson 2001)Panda (Giant) 1978 AI Liu et al. [see (Huang et al. 2002)]
1980 CP/AI See (Huang et al. 2002)]Polecat (European) 2002 ET (Lindeberg et al. 2002)
2003 CPe/ET (Lindeberg et al. 2003)Puma 1981 AI (Moore et al. 1981)Tigrina 1997 CPs/AI See (Swanson 2001)Tiger (Panthera tigris) 1990 IVF/ET (Donoghue et al. 1990)
(table continued)

23
Tiger (Siberian) 1993 AI Donoghue et al. [see (Donoghue et al. 1996)]Wolf 1975 CPs/AI Seager et al. [see (Rott 1995)]
CetaceaBottlenose dolphin 2000 AI [see (Robeck 2001), http://www.seaworld.org/whats-new/znn/2003/july/worlds-first.htm]
2002 CPs/AI http://www.seaworld.org/whats-new/znn/2003/july/worlds-first.htmKiller whale 2001 AI (Robeck 2001)(http://news.bbc.co.uk/1/hi/sci/tech/1523214.stm)
LogmorphaRabbit 1882 AI Ott [see (Heape 1897)]
1890 ET (Heape 1890)1959 IVF/ET (Chang 1959)1988 ECNT/ET (Stice and Robl 1988)1988 ICSI/ET (Hosoi et al. 1988)2002 SCNT/ET (Chesne et al. 2002)
MarsupialiaKoala 1999 AI (Johnston et al. 1999)Quokka 1963 ET (Tyndale-Biscoe 1963) Species name: Setonix brachyurs
Wallaby (tammar) 1970 ET (Tyndale-Biscoe 1970) Laparoscopic ET2002 AI (Paris et al. 2002)
PerissodactylaDonkey 1980 ET (Allen 1982) Domestic mare recipient
1985 ET (Davies et al. 1985) Mule recipientDonkey (Pouke) 2002 AI/iET Mckinnon (http://news.bbc.co.uk/1/hi/sci/tech/1804807.stm) Domestic mare recipientHorse (domestic) 1894 AI see (Heape 1897)
1957 CPs/AI (Barker and Gandier 1957)1974 ET (Oguri and Tsutsumi 1974)1980 ET Allen [see (Allen 1982)] Domestic donkey recipients1981 ET Salazar [see (Davies et al. 1985)] Mule recipient1982 CP/ET (Yamamoto et al. 1982)1991 IVF/ET (Palmer et al. 1991)1996 ICSI/ET (Squires et al. 1996; Cochran et al. 1998)2002 Oocyte transfer (Maclellan et al. 2002) Cryopreserved oocyte (CPo)
(table continued)

24
2003 SCNT/ET (Galli et al. 2003a)Horse (Przewalski’s ) 1985 ET (Kydd et al. 1985) Domestic mare recipientMule 1972 ET Allen and Rowson [see (Allen 1982)]
1972 ET Allen and Rowson [see (Allen 1982)] Mule recipient2003 SCNT/ET (Woods et al. 2003) Domestic mare recipient
Hinny 1972 ET Allen and Rowson [see (Allen 1982)]Zebra (Burchell’s) 1985 iET (Bennet and Foster 1985) Domestic mare recipientZebra (Grant’s) 1985 iET (Kydd et al. 1985) Pony mare recipient
ProboscideaElephant (Asian) 1999 AI (Schmitt et al. 2001)Elephant (African) 2000 AI (Schmitt et al. 2001)
RodentiaGerbil (Mongolian) 1981 ET Norris [see (Adams 1982)]Hamster 1964 ET (Blaha 1964)
1992 IVF/ET (Barnett and Bavister 1992)1999 CPe/ET (Lane et al. 1999)2002 ICSI/ET (Yamauchi et al. 2002)
Mastomys 1998 IVF/ET See (Ogonuki et al. 2003)2002 ICSI/ET (Ogonuki et al. 2003)
Mouse 1935 ET (Fekete and Little 1942)1968 IVF/ET (Whittingham 1968)1972 CPe/ET (Whittingham et al. 1972)1977 CPo/IVF/ET (Whittingham 1977)1981 ECNT/ET (Illmensee and Hoppe 1981; McGrath and Solter 1983)1997 CPs/IVF/ET (Songsasen et al. 1997)1995 ICSI/ET (Kimura and Yanagimachi 1995; Lacham-Kaplan and Trounson 1995)1998 SCNT/ET (Wakayama et al. 1998)
Rat 1933 ET (Nicholas 1933)1974 IVF/ET (Toyoda and Chang 1974)1998 ICSI/ET (Dozortsev et al. 1998)2003 SCNT/ET (Zhou et al. 2003)
PrimatesBaboon 1975 ET (Kraemer et al. 1976)
(table continued)

25
1984 CPe/ET (Pope et al. 1984)1984 IVF/ET (Clayton and Kuehl 1984)
Chimpanzee (common)1978 AI (Martin et al. 1978)Chimpanzee 1989 CPs/AI (Gould and Styperek 1989)Chimpanzee (bonobo) 2001 CPs/AI (Kusunoki et al. 2001)Gorilla (lowland) 1981 CPs/AI Douglass & Gould [see (Pope et al. 1997)]
1995 CPs/IVF/ET (Pope et al. 1997)Human 1799 AI Hunter [see (Heape 1897)]
1953 CPs/AI Bunge and Sherman [see (Curry 2000)]1978 IVF/ET (Steptoe and Edwards 1978)1984 IVF/CPe/ET (Downing et al. 1985)1992 ICSI/ET (Palermo et al. 1992)
Monkey (Cynomolgus) 1990 CPs/AI (Tollner et al. 1990)1984 IVF/ET (Balmaceda et al. 1984)1984 iET (Balmaceda et al. 1984) Rhesus recipient1986 IVF/CPe/ET (Balmaceda et al. 1986)
Monkey (Rhesus) 1977 ET (Marston et al. 1977)1983 IVF/ET (Bavister et al. 1984)1989 IVF/CPe/ET (Wolf et al. 1989)1997 ECNT/ET (Meng et al. 1997)2000 ICSI/ET (Chan et al. 2000)2000 AI (Gabriel Sanchez-Partida et al. 2000)2000 CPs/AI (Gabriel Sanchez-Partida et al. 2000)
Monkey (Marmoset) 1984 CPe/ET (Hearn and Summers 1986)1988 IVF/ET (Lopata et al. 1988)1997 ET (Marshall et al. 1997)
________________________________________________________________________________________________________________aAI = artificial insemination; ET = embryo transfer; iET = interspecies ET; CP = cryopreserved; CPs = CP spermatozoa; CPe = CP embryo;CPo = CP oocyte; ECNT = embryonic cell nuclear transfer; SCNT = somatic cell nuclear transfer; iSCNT = interspecies SCNT; IVF = invitro fertilization;bAttempts were made to cite the first successful report for each species.cAI in most farm animals was initiated in the late 1800 or early 1900’s [see (Foote 2002)].

26
Tragelaphus (bongo, bushbuck, greater kudu, lesser kudu, lowland nyala, mountain nyala, sitatunga)
and Taurotragus (common eland, giant eland). Each species has at least two subspecies or races;
thus, there are at least 18 species and subspecies.
Tragelaphine antelopes first appeared about 6 million years ago in sub-Saharan Africa, which
is also their current native habitat (Pappas 2002). They are “medium to large antelopes with deep
bodies, long necks and legs, narrow heads with big ears and twisted or spiral horns” (Kingdon 1982)
and have “variously developed face and body pattern of white spots and stripes”(Nowak 1999).
There is clear sexual dimorphism, males being larger than females and having horns in all species;
however, only female elands (common and giant) and bongo have horns, while females of the
remaining species are polled. All tragelaphine antelopes are non-territorial. They are also gregarious,
with the exception of bushbuck and sitatunga, which are solitary (Nowak 1999). Average body
weight of adult females ranges from 30 kg in the bushbuck to 450 kg in eland (Appendix A).
The beauty of tragelaphine antelopes has impressed many writers. Crandall (1964) described
male kudus as “the handsomest of antelopes” and bongos “among the most beautiful” of antelopes,
while Nowak (1999) asserted bongo as “the most beautiful of bovids”. The giant eland has been
given royal status as “regal and delighting to the eye” [see (East 1999)]. The common eland has been
hailed as the “apotheosis of antelope evolution” [see (Spinage 1986)]. Besides ecological importance,
there is increasing interest in the economic use of tragelaphine antelopes through game farming or
domestication (Treus and Lobanov 1971; Madzingira et al. 2002). In a recent study by the latter
authors (Madzingira et al. 2002), four tragelaphine antelope species (greater kudu, common eland,
bushbuck and lowland nyala) were among the seventeen species of antelopes kept by farmers
practicing mixed cattle: antelope farming.
The IUCN classifies the threat to extinction of eight tragelaphine species as “lower risk”; but
specifically three species (bongo, giant eland and sitatunga) are “near threatened” and the remaining
five species are assigned “conservation dependent” status. Mountain nyala (T. buxtoni) and two

27
subspecies, the western giant eland (T. derbianus derbianus) and mountain bongo (T. eurycerus
isaaci) are “endangered” [see (East 1999; IUCN 2002)].
With the exception of the mountain nyala, all species are represented in captivity in zoos. The
only record of keeping the mountain nyala in captivity ended when a pair at the Zoological Garden of
Berlin was killed during an air raid in 1944 (Crandall 1964). Additional descriptions of the general
characteristics and biological aspects of the tribe and specific species are described elsewhere (Ralls
1978; Kingdon 1982; Spinage 1986; Nowak 1999).
The closeness of tragelaphine antelopes to each other is exemplified by the “unusual
readiness to hybridize in captivity” (Kingdon 1982). To date, six hybrid pregnancies have been
reported (Table 2.2), including a fertile bongo sitatunga hybrid (Koulischer et al. 1973) and a
bushbuck sitatunga hybrid that survived for years (Kingdon 1982).
Tragelaphine antelopes are peculiar among mammals in having normal translocation of their
Y-chromosomes to autosomes. Karyotype studies are not available for giant eland and mountain
nyala; however, among the seven species, all except sitatunga and lesser kudu have the unusual
translocation of the Y-chromosome (see Appendix A). The similarity among the species has also
been demonstrated by analyzing loci of mitochondrial DNA (Essop et al. 1997). Accordingly, there
is a tendency to classify all the species in the Tragelaphus genus (East 1999; IUCN 2002).
Table 2.2. Attempts to hybridize tragelaphine antelopes and the results_____________________________________________________________________________________________Male Female Offspring Comment Reference_____________________________________________________________________________________________
Bongo Sitatunga Female, “Bongsi” Fertile (Koulischer et al. 1973)
Bushbuck Sitatunga Aborted See (Benirschke et al. 1980)
Eland Greater kudu Males Sterile See (Benirschke et al. 1980)
Eland Sitatunga Females Died See (Benirschke et al. 1980)
Lesser kudu Bushbuck ? ? (Kingdon 1982)
Lesser kudu Sitatunga Male Died See (Benirschke et al. 1980)
____________________________________________________________________________________________________________________________

28
The possibility of hybridization suggests that interspecies embryo transfer may be successful
among several tragelaphine species. This has already been demonstrated by the successful transfer of
bongo embryos to the common eland (Dresser et al. 1985). Hybrid females (e.g., mule) can support
pregnancy after the transfer of embryos from either of the parental species (Davies et al. 1985).
Similarly, hybrid female tragelaphine antelopes could probably support embryos from either parent.
Immune rejection (Loskutoff 1999) would possibly be reduced in the hybrid surrogates in
comparison to regular interspecies embryo transfer recipients.
2.3.2. Common Eland
The common eland is the largest African antelope. Three subspecies of the common eland
are: T. o. pattersonianus (East African race), T. o. livingstonii (Zambezi race) and T. o. oryx (Cape or
South African race). Skin color and the number of lateral body stripes are the most frequently
mentioned criteria for differentiating the three subspecies. Stripes are more numerous and marked in
the East African subspecies and may be absent in southern species (Crandall 1964; Spinage 1986;
Pappas 2002). Body weights of up to 595 kg (females) and 942 kg (males) have been recorded
(Crandall 1964; Kingdon 1982). At the Askanya-Nova Zoo, Ukraine, the average longevity of 23
females was 11 (maximum of 23) years (Treus and Lobanov 1971). In the United States, longevity of
up to 25 years has been recorded (Crandall 1964).
The “ox-like appearance” of the eland appears to be the primary source of interest for the
domestication of this species. Documented interest in eland domestication dates back to 1591 [see
(Spinage 1986)]. The most successful effort to produce a fully domesticated herd of eland, leading to
females that could be hand-milked, was carried out at the Askanya-Nova Zoo, Ukraine (Treus and
Lobanov 1971). Domestication attempts have also been made in Kenya, Zimbabwe, Britain, France
and Brazil (Kingdon 1982).

29
Advantages of domestication of the eland include their high tolerance to water shortage. The
eland can reduce sweating and hence conserve water by raising its body temperature from 39°C to
42°C during increased environmental temperatures (Kyle 1972). Elands also consume more diverse
forage species and are less competitive with conventional livestock (Abdullahi 1981). As a result,
they have been recommended as a more viable alternative species (better than domestic cattle) for
introduction and meat production in the Southwestern United States where invasive woody trees are
a problem (Humphries, Jr. 1972). They are also more efficient in feed conversion of the diverse
forage consumed. For example, eland gained 0.33 kg in body weight per day as compared with 0.14
kg in cattle managed under similar conditions (Kyle 1972). Also, they are resistant to diseases such
as trypanosomiasis and theileriosis (Kingdon 1982) that hinder livestock production in many parts of
Africa. Moreover, eland cows produce high quality meat and milk (Spinage 1986). With calving
intervals as short as 10 months (Kingdon 1982), reproductive performance in elands is more efficient
than most tropical cattle breeds.
Although the requirements by eland for large area and high quality feed that may not be
fulfilled by regular grazing have been discouraging in many domestication attempts (Kingdon 1982),
the eland could offer opportunities for agricultural diversification in such areas where
trypanosomiasis and East Coast Fever (theileriosis) are prevalent. Large numbers of elands are kept
in game ranches in southern Africa (East 1999; Madzingira et al. 2002). For example, eight of 11
farms practicing cattle-antelope mixed farming near Harare, Zimbabwe, kept an average of 33 elands
per farm (Madzingira et al. 2002). Since economics is the primary driving force behind most
technologies, ART included, the promotion of eland domestication could help advance antelope
ART.

30
2.3.3. Reproduction and ART in Tragelaphine Antelopes
Ovarian cyclicity in tragelaphine antelopes occurs throughout the year. However, in their
natural habitat, breeding and calving may be seasonally dependent on feed availability. Among
tragelaphines, only the greater kudu shows marked seasonality in calving pattern, both in the wild and
in captivity (Skinner et al. 2002). The estrous cycle length (Appendix A) is similar to that of domestic
bovids. Like the mare, tragelaphine females show a fertile postpartum estrus (Kingdon 1982).
Reported gestation lengths vary between 6 to 9 months (Appendix A). Members of the tribe reproduce
well, both in captivity and in the wild, with calving intervals of a year or less (Crandall 1964; Kingdon
1982). However, detailed information on reproductive performance is available only for the eland.
Like other tragelaphines, common eland cows cycle throughout the year [(Treus and
Lobanov 1971) and references cited there]. The estrous cycle ranges between 21-26 days (Atkinson
et al. 1999; Nowak 1999) and estrus lasts 1-3 days (Nowak 1999). Age at puberty ranges from 1-3 yr
in the female (Dittrich 1972), but may be delayed in the male (Nowak 1999). In North America, the
median age at first calving of captive eland was about 40 months (Lydon 2001). The mean age at first
calving was 36.4 months (n = 42) at the Askanya-Nova Zoo, Ukraine (Treus and Lobanov 1971).
However, female elands have given birth as early as 16 months of age (Lydon 2001), suggesting that
ovarian cyclicity can start before 10 months of age. Calves are produced after gestation length of 254
to 284 days (Dittrich 1972; Pope and Loskutoff 1999). Of 1,214 eland deliveries, a single calf was
born in 99% of the calvings and the remaining 1% delivered twins (Lydon 2001). Domestic eland
cows at the Askanya-Nova Zoo produced an average of eight calves (maximum 16) during their
lifetime (Treus and Lobanov 1971).
Assisted reproduction studies in tragelaphine antelopes are scant; however, the common
eland is the most studied. Treus and Lobanov (1971) described unsuccessful experiments of artificial
insemination of domestic cows using eland semen. Although details are not readily available, Treus’

31
group may have been the first to collect eland semen and perform artificial insemination. The
cryopreservation of eland spermatozoa was first reported in 1978 (Merilan et al. 1978).
Merilan and colleagues collected semen using electroejaculation from an eland bull under
general anesthesia. They cooled the sample to 4º C and froze it in 0.5-mL straws in liquid nitrogen
vapor. Post-thaw motility was 35% (fresh sample = 65%) after freezing semen in egg yolk-citrate-
extender and 6% glycerol (Merilan et al. 1978). There are few additional studies evaluating
cryopreservation of epididymal or ejaculated eland spermatozoa (Bartels et al. 2001; Herrick et al.
2002; Wirtu et al. 2002b). One study reported the birth of an eland calf after artificial insemination
using frozen-thawed epididymal spermatozoa (Bartels et al. 2001). In that report, the semen was
diluted in Triladyl® extender, cooled to 4ºC and loaded into 0.25-mL straws and frozen in liquid
nitrogen vapor. One of four eland cows inseminated with the frozen-thawed sample (55% motility)
delivered a calf after a gestation period of 11 months. However, the reported gestation period is at
least two months longer than the average for the eland (255 - 284 days, Appendix A).
In greater kudu, among seven cryoprotectant extenders that were evaluated (Schiewe et al.
1991a), EQ and BF5F supported the highest post-thaw motility of 51% and 40%, respectively. In
another study (Schmid et al. 1997), cryopreservation of greater kudu epididymal spermatozoa in five
extenders were evaluated, along with three thawing temperatures. Although results varied between
bulls, samples frozen in Tris-citrate-based extender containing 20% egg yolk and 7% glycerol had
the highest post-thaw motility (up to 65%), with slightly higher motility after thawing at room
temperature. In four sitatunga bulls, the post-thaw motility of ejaculated spermatozoa frozen in
Triladyl® extender ranged between 22% and 42% of the initial motility of 40 to 80% (Perez-Garnelo
et al. 2000). In the bongo antelope, the post thaw motility of spermatozoa frozen in TEST-yolk buffer
containing 6% glycerol was less than 40% (Dr. Earle Pope, personal communication). Similarly,
ejaculated giant eland spermatozoa frozen at White Oak Conservation Center, Florida, had post-thaw
motility of 30% or less after freezing in BF5F extender (personal observation). Thus, with the

32
exception of the two studies (Schmid et al. 1997; Bartels et al. 2001), the survival of tragelaphine
spermatozoa frozen using standard bovine semen freezing protocols was unsatisfactory.
Successful ART involving tragelaphine embryos are available for the common eland and
bongo. The transfer of fresh (unfrozen) embryos recovered after natural mating of superovulated
eland and bongo females has resulted in normal offspring in both species. Dresser and colleagues
recovered seven embryos from a superovulated bongo female 8 days after mating and transferred
single embryos to four eland and a bongo at ~12 hr after embryo collection. The bongo and one eland
recipient delivered normal bongo calves (Dresser et al. 1985). The same group reported the first
tragelaphine pregnancies carried to term after the transfer of frozen-thawed eland embryos to the
eland (Dresser et al. 1984; Pope and Loskutoff 1999). The transfer of bisected embryos to eland
cows also resulted in a normal eland calf (Gelwicks et al. 1989). To date, four eland and two bongo
calves have been produced after the transfer of in vivo produced embryos to recipient females (Pope
and Loskutoff 1999).
The above pioneering tragelaphine ART studies demonstrated the feasibility of estrous cycle
synchronization, ovarian stimulation and non-surgical embryo recovery and transfer in the two
species. Non-surgical embryo recovery has also been reported in the greater kudu (Schiewe et al.
1991b). In the latter study, two embryos were recovered after uterine flushing of two females
subjected to ovarian stimulation treatments and natural mating. However, details were not given on
the quality/stage of the embryos.
In vitro embryo production, using standard IVF or iSCNT, has been reported in the mountain
bongo, greater kudu, common eland and giant eland. Pope and colleagues (Pope et al. 1998b)
collected 76 oocytes (average = 5 per animal) from bongo antelope using transvaginal ultrasound-
guided follicular aspiration after ovarian stimulation treatments. IVF with frozen-thawed
spermatozoa led to cleavage frequency of 35 % (14 of 40 evaluated oocytes), with one embryo
developing to the blastocyst stage. A similar frequency of cleavage (35%, 117/330 oocytes) after

33
IVM-IVF-IVC was observed in greater kudu oocytes recovered from ovaries collected postmortem
(Loskutoff et al. 1995). Fresh or cooled epididymal spermatozoa were used for IVF. Four (1%) kudu
blastocysts developed.
As part of the current study, our group reported the use of transvaginal ultrasound-guided
follicular aspiration to recover common eland oocytes for IVP. We recovered an average of nine
oocytes per female after ovarian stimulation treatments. Two of 26 metaphase II stage oocytes
cleaved after IVF using frozen-thawed eland spermatozoa (Wirtu et al. 2002a). Obviously, IVF has
been rather inefficient in the three tragelaphine antelopes (bongo, greater kudu and common eland)
studied to date.
After interspecies somatic cell nuclear transfer to enucleated bovine oocytes, somatic cell
nuclei of mountain bongo (Lee et al. 2003) and giant eland (Damiani et al. 2003) can support
development to blastocysts. The average blastocyst development ranged from 19 - 27% (giant eland)
and 11 - 24% (bongo) of activated SCNT couplets. Another group (Matshikiza et al. 2004) also
reported the development of a blastocyst (2%) after “hand-made” nuclear transfer of common eland
somatic cells into bovine oocytes. Enucleated common eland oocytes have also supported embryonic
development up to the 8-cell stage after iSCNT with giant eland cells (Damiani et al. 2003).
Therefore, development of blastocysts has been reported after iSCNT of bongo, giant eland and
common eland somatic cells into bovine oocytes.
In applying or developing ART in a new species, the use of information developed in a
closely related domestic and nondomestic species as a starting point is recommended (Loskutoff et
al. 1995; Comizzoli et al. 2000; Pope 2000; Watson and Holt 2001; Leibo and Songsasen 2002).
Considering the close relationship of common eland to other tragelaphine antelopes (discussed
above), its availability and prolificacy in captivity, this species can serve as a sound model species to
study tragelaphine ART. Interest in the use of the common eland for game ranching and

34
domestication could also provide additional economic incentive for developing and applying ART in
the species.
2.3.4. Role of Behavioral Training and Advanced Handling Systems to Facilitate ARTProcedures
As discussed previously, obtaining gametes is critical to the development of ART, especially,
in rare and endangered species. Ultrasound-guided oocyte retrieval provides a safe and relatively
atraumatic approach for collection of oocytes from live animals. In cattle, the procedure can be
repeated on a single animal as frequently as twice per week without adverse effects (Goodhand et al.
1999; Chastant-Maillard et al. 2003; Petyim et al. 2003). Oocytes obtained by this method are
capable of developing into viable embryos after IVP.
Oocytes have been collected using the ultrasound-guided method in several nondomestic
species (reviewed in section 2.2). In most cases, general anesthesia was induced to facilitate oocyte
collection. However, ART procedures require frequent handling to synchronize the estrous cycle,
administer ovarian stimulation treatments, perform oocyte retrieval and transfer embryos.
The standard ovarian stimulation protocol in domestic cattle involves twice-daily injections
of FSH for 4 or 5 days. Applying such a regimen in nondomestic ungulates such as the eland
(Schiewe et al. 1991b) and bison (Dorn 1995) is stressful. Moreover, as with other remotely
delivered medications, it is difficult to ensure complete delivery of the required medications, some of
which are very expensive (e.g., FSH).
Immobilization or general anesthesia also poses multiple risks, such as cardiac and
respiratory arrest or even death. In a study on four species of immobilized antelopes (oryx, bongo,
kudu and common eland), assessment of blood cortisol levels indicated handling and immobilization
procedures were most stressful to elands. Also, maintenance of general anesthesia was more difficult
in the elands (Schiewe et al. 1991b). Individual variations to ovarian stimulation treatments among
the four species of antelopes were attributed to handling-related stress. Handling-induced stress may

35
have a negative effect on in vitro and in vivo development of oocytes and embryos, and can affect the
lifetime reproductive performance of the animal. Moreover, it is technically difficult to incorporate
immobilization procedures in studies (for example, reproductive endocrinology) requiring frequent
blood sampling or examinations.
Thus, because of the immediate and long-term risks associated with conventional
nondomestic animal handling/immobilization practices and the fact that such practices are technically
difficult to apply in routine ART procedures, there is a need to develop alternative methods of animal
handling. Studies on domestic cattle indicate that dissolving FSH in carrier solutions, such as
polyvinyl pyrrolidone (PVP), instead of saline, can prolong hormone action. This approach has been
used to reduce the number of injections of gonadotropins without affecting ovarian response
(Yamamoto et al. 1994; Takedomi et al. 1995).
A regimen involving two or three injections of prostaglandin F2α or its analogs is frequently
used in estrous synchronization protocols for domestic and nondomestic ungulates. Although
progestogens may be more effective for synchronization, application of progestogen pessaries can be
more stressful to nondomestic ungulates than the use of prostaglandins (Haigh 2001). Orally
administered progestogens could minimize handling stress since they can be added to the regular
diet. For example, altrenogest has been used to control estrous cycles in pigs and cattle (Estienne et
al. 2001; Ferguson et al. 2002) and to support pregnancy in cases of luteal insufficiency in horses
(Daels et al. 1998) and okapi (Schwarzenberger et al. 1999). However, its effectiveness in
controlling the estrous cycle of antelopes has not been evaluated.
Behavioral training, combined with improved handling systems, has been used to perform
procedures without immobilizing nondomestic ungulate species. Phillips and co-workers (Phillips et
al. 1998) have indicated that conditioning can often replace chemical immobilization with
subsequent reductions in potential risks to the animal and the handlers. This group successfully

36
conditioned lowland nyala and bongo in 97 and 118 days, respectively (Grandin 2000). Animals
cooperated with procedures such as blood sampling, injections, milking and treatment of wounds
without chemical or manual restraint. The method of conditioning involved acclimation of animals to
the presence of a shipping crate, habituation to walking through the crate, providing treats in the crate
and gradual training (conditioning) to veterinary procedures over a period of time.
Other species have been trained to comply with various tasks by applying the principles of
positive reinforcement or operant conditioning (Stevens 1978; Bloomsmith et al. 2003). Ungulates,
including giraffe, eland, wildebeest and Thompson’s gazelle have been trained to associate audio
cues with a food reward at the Disney’s Animal Kingdom (Joe Kalla and Marty Sevenich, personal
communication). However, there are no reports on the use of behavioral training to facilitate ART
procedures in nondomestic ungulates.
Advanced handling chutes have been used to do clinical procedures in many large mammals,
including the eland (Read et al. 1993). A prototype device is the hydraulic chute, The TamerTM,
originally developed for handling deer and later modified for larger hoofstock (Citino 1995; Atkinson
et al. 1999). The latter investigators have used the device to perform many veterinary procedures that
would have otherwise required immobilization in eland, bongo and kudu antelopes. In addition,
another type of device, the drop-floor chute, has been used to restrain addax for ART procedures.
After handling females in the chute, ultrasound-guided oocyte retrieval was done under
acepromazine-induced tranquilization and epidural analgesia (Junge 1998). Mechanical or manual
restraint has also been used to perform artificial insemination, with subsequent production of
offspring, in the giraffe, gaur and addax [(Densmore et al. 1987; Godfrey et al. 1990) see (Pope and
Loskutoff 1999)]. However, there is no report on the use of such restraint devices for ART
procedures in tragelaphine antelopes.
In the present study, we evaluated the development of in vitro-derived bovine embryos in
protein-free media and identified several factors affecting embryonic development in vitro (Chapter

37
3). Moreover, behavioral training and handling of female elands in a hydraulic chute were used to
facilitate ultrasound-guided oocyte retrieval. It is demonstrated that ART procedures can be carried
out in elands without inducing general anesthesia. Some parameters associated with handling of
female elands, and factors affecting ovarian response after gonadotropin treatment and the in vitro
development of eland oocytes after IVP are also presented (Chapters 4 and 5).

38
Chapter 3. Development of In Vitro-Derived Bovine Embryos in Defined(Protein-free) Media
3.1. Introduction
In vitro-produced cattle embryos display morphological, biochemical and functional
deviations when compared with those derived in vivo. These anomalies are more severe in embryos
produced in media containing complex components, such as serum. Thus, many laboratories are
attempting to develop and use simple and defined or semi-defined embryo culture media (Chapter 2).
The use of chemically defined culture media has multiple benefits (Bavister 1995). It reduces
variations among different lots of media and minimizes risks of contamination with pathogens.
Complex additives such as serum or serum protein preparations often contain ill-defined inhibitory or
stimulatory modifiers of in vitro embryo development. Testing the effects of such modifiers, or
eliminating them, can be achieved only by using chemically defined culture systems. Information
generated using chemically defined media (without serum, BSA or co-culture) is thus helpful for
improving our understanding of the requirements of embryos and subsequently for optimizing culture
systems (Bavister 1995; Keskintepe et al. 1995; Liu and Foote 1995b; Holm et al. 1999; Hernandez-
Fonseca et al. 2002).
Results of recent studies of cattle IVP using chemically defined culture systems are
encouraging. In vitro embryo developmental frequencies similar to those achieved in complex culture
media have been reported (Abe et al. 1999; Holm et al. 1999). Moreover, pregnancies have been
established (Keskintepe et al. 1995; Hernandez-Fonseca et al. 2002) and healthy calves born (Holm
et al. 1999; Hernandez-Fonseca et al. 2002) after transferring embryos cultured in chemically-
defined media to recipient cows. Even so further refinement of chemically defined media is needed
to increase the efficiency of embryo production and improve our understanding of early embryo
development.

39
Amino acids, energy substrates and osmotic pressure and/or NaCl concentration are among
the main factors influencing in vitro development of mammalian pre-implantation embryos (Bavister
1995; Liu and Foote 1995a; Lee and Fukui 1996; Steeves and Gardner 1999) (also see Chapter 2).
Thus, the hypothesis of the present study is: the in vitro development of bovine embryos in protein-
free media is affected by groups of amino acids, base media, supplement type and/or osmotic
pressure. Specific objective were to:
1. compare the effects of supplementing two base media (modified KSOM, BM-3) with two
groups of amino acids (20aa, 11aa) on the in vitro development of in vitro-derived bovine
embryos; and
2. determine the effects of osmotic pressure of BM-3-20aa and four selected supplements on the
in vitro development of in vitro-derived bovine embryos.
3.2. Material and Methods
3.2.1. General
All chemicals were purchased from Sigma Chemicals Co. (St. Louis, MO, USA) unless
indicated otherwise. All media were prepared in ultrapure (Milli-Q) water fortified with 1%
penicillin-streptomycin-amphotericin solution and passed through a syringe filter with 0.22 µm pore-
size before use. During IVF and IVC, a group of 10 to 15 oocytes/embryos were incubated in 50-µL
droplets in 60 x 15 mm Falcon® polystyrene petri dishes (Becton Dickinson Labware, Franklin
Lakes, NJ, USA). The droplets in the dishes were covered with embryo-tested mineral oil and
equilibrated for ≥2 hr in the incubator before placing gametes or embryos in them. Incubation
temperature was maintained at 39ºC for IVM, IVF and IVC.

40
3.2.2. In Vitro Maturation
Cattle oocytes were collected by aspirating follicles (<10 mm in diameter) from ovaries
obtained at a slaughterhouse. The commercial supplier (Cyagra, Inc., Gibbons, NE, USA) collected
and shipped the oocytes to our laboratory. Oocytes underwent IVM for 23 to 25 hr while en-route to
our laboratory. For IVM, 50 to 100 oocytes were placed in a 2-mL glass tube containing IVM
medium and gassed with 5% CO2 and shipped in a battery-operated portable incubator (Minitüb,
Tiefenbach, Germany). The temperature of the incubator was maintained at 39°C. The IVM medium
was bicarbonate-buffered TCM-199 with 10% fetal calf serum, bovine LH (0.01 U/mL) and estradiol
(1 µg/mL).
3.2.3. In Vitro Fertilization
At 23 to 25 hr of IVM, oocytes with homogeneous ooplasm and three or more expanded
cumulus layers were selected and washed at least three times in modified Tyrode’s solution
supplemented with lactate, HEPES and PVA (TL-HEPES-PVA, Bavister 1989) supplemented with
1.25 mM pyruvic acid and 1% Minimal Essential Medium (MEM) non-essential amino acids before
undergoing IVF. The latter was done in TALP [modified Tyrode’s solution + bovine serum albumin
(6 mg/mL) + DL-lactic acid (10 mM) + pyruvic acid (0.25 mM)] (Bavister and Yanagimachi 1977)
containing 20-µg/mL heparin (Parrish et al. 1988), PHE [penicillamine (20 µM) + hypotaurine (10
µM) + epinephrine (1 µM)] (Ball et al. 1983; Susko-Parrish et al. 1990) and 1% MEM amino acids.
Cryopreserved bull spermatozoa, which had been previously validated for IVF, were used. A
0.5 mL straw of semen was thawed in a water bath (37°C, 30 sec) and its contents were placed in a
35-mm dish. The semen was layered gently over a 1 mL of PureSperm (GenX, Guilford, CT, USA)
gradient in a 15-mL centrifuge tube consisting of 45% (top) and 90% (bottom) solutions in TL-
HEPES-PVA. The tube was then centrifuged (500 x g, 15 min). Then the pellet was recovered and
diluted in TL-HEPES-PVA to give a final insemination concentration of ~1.5 x 106 motile

41
spermatozoa/mL and volume of 1 µL. An atmosphere of 5% CO2 in air with high humidity was used
for IVF.
3.2.4. In Vitro Culture and Experimental Design
At 16 to 20 hr of sperm-oocyte co-incubation, presumptive zygotes were vortexed (1800 rpm,
2 min) to remove cumulus cell layers and attached spermatozoa, washed (≥3x), and randomly
allocated to in vitro culture treatments. Oil covered microdrops of IVC media were prepared as
described above (Section 3.2.1). The same dish contained all treatments. To prevent cross-
contamination of microdrops during the allocation of ova to IVC treatments, at least three extra
microdrops of the simplest treatment (i.e., HECM-6 in Experiment 3.1 and BM-3-20aa in Experiment
3.2) were used for additional washings. Immediately before placing the ova in the IVC drop, a final
washing was done in one of the extra microdrops of the IVC treatment.
In vitro culture was conducted in an atmosphere of 5% CO2, 10% O2 and 85% N2 with high
humidity in culture medium as specified in the experimental designs below. Embryos were moved to
fresh culture medium on day 4 post-insemination.
3.2.4.1. Experiment 3.1 (Effects of Amino Acids and Base Media)
This experiment was a 2 x 2 factorial arrangement (Table 3.1) designed to compare the
effects of two groups of amino acids and two base media on the development of in vitro-derived
embryos. Treatment 1 (HECM-6) was used as a control. The two base media were BM-3 (McKiernan
et al. 1995), and KSOM (Erbach et al. 1994) as modified previously (Yang et al. 1995). The first
group of 20 amino acids (20aa) consisted of glutamine and commercial preparations of Basal
Medium Eagle (BME) essential and MEM non-essential amino acids. [The classification of amino
acids as essential and non-essential is based on nutrient requirements in rats and “is not relevant to
embryos” (Liu and Foote 1997)]. However, it is helpful to use this nomenclature as a simple way to
identify these groups of amino acids.

42
The BME and MEM amino acids, purchased as 50x and 100x solutions, respectively, were
supplemented at 1% in the IVC media. The second group was the 11 amino acids developed
specifically for hamster IVC [HECM-6, McKiernan et al. 1995). Modified 11 amino acids had the
same components as HECM-6 but concentrations were increased to equal those in BME or MEM.
Glutamine, taurine and cysteine, which are not components of BME/MEM, were used at 1.0, 0.5 and
0.05 mM, respectively (Table 3.2).
Table 3.1. Design of Experiment 3.1_________________________________________________________________________________Treatment Base medium Amino acid (aa) supplements Designated as_________________________________________________________________________________
1 BM-3 11 (HECM-6) aa HECM-6
2 BM-3 modified 11 aa BM-3-m11aa
3 BM-3 20 aa (BME + MEM + glutamine) BM-3-20aa
4 modified KSOM modified 11 aa mKSOM-m11aa
5 modified KSOM 20 aa (BME + MEM + glutamine) mKSOM-20aa_________________________________________________________________________________
3.2.4.2. Experiment 3.2 (Effects of Glucose, Pyruvate, Lactate and Phosphate in BM-3Having Two Osmolalities)
In Experiment 3.1, interaction between base medium and group of amino acids was apparent,
with 20aa supplemented in modified KSOM supporting the highest frequency blastocyst
development. Therefore, Experiment 3.2 was designed to evaluate the differential effects of four
selected components of KSOM and the 20aa in BM-3 having two osmolality levels. The experiment
was a 7 x 2 factorial design with the following supplement treatments and two osmotic pressures:
1. KSOM-PVA + 20 amino acids (= KSOM-20aa)
2. BM-3 + 20 amino acids (= BM-3-20aa)
3. BM-3 + 20 amino acids + 0.2 mM glucose (= BM-3-20aa-gluc)
4. BM-3 + 20 amino acids + 0.2 mM pyruvic acid (= BM-3-20aa-pyr)

43
Table 3.2. Components and concentrations of modified KSOM, BM-3 and three groups of aminoacids_________________________________________________________________________________Base mediaa components and concentration (mM) Amino acids, final concentration (mM)
Component mKSOM BM-3 Component Twentyb Elevenc mEleven_________________________________________________________________________________
PVA (mg/mL) 0.1 0.1 MEM
NaCl 95.0 113.6 L-Alanine 0.10 -- --
KCl 2.5 3.0 L-Asparagine 0.11 0.01 0.11
KH2PO4 0.35 -- L-Aspartic acid 0.10 0.01 0.10
MgSO4 0.2 -- L-Glutamic acid 0.09 0.01 0.09
MgCl2 -- 0.5 Glycine 0.10 0.01 0.10
CaCl2.2H2O 1.7 1.9 L-Proline 0.10 0.01 0.10
HEPES 2.5 -- L-Serine 0.10 0.01 0.10
EDTA (Na salt) 0.01 -- BME
Pyruvic acid (Na salt) 0.2 -- L-Arginine HCl 0.06 -- --
Glutamine 1.0 0.2 L-Cystine 0.03 -- --
Glucose 0.2 -- L-Histidine 0.03 0.01 0.03
DL-Lactic acid (Na salt) 10.0 4.5 L-Isoleucine 0.10 -- --
NaHCO3 25.0 25.0 L-Leucine 0.10 -- --
HCl (1 mol/L) -- (1.4 µL/mL) L-Lysine HCl 0.10 0.01 0.10
L-Methionine 0.03 -- --
L-Phenylalanine 0.05 -- --
L-Threonine 0.10 -- --
L-Tryptophan 0.01 -- --
L-Tyrosine HCl 0.04 -- --
L-Valine 0.10 -- --
Other
Taurine -- 0.5 0.5
L-Glutamine 1.0 0.2 1.0
L-Cysteine.HCl.H2O -- 0.01 0.05
Osmolality 255 ± 5 275 ± 5_________________________________________________________________________________aBase media were prepared with glutamine and stored at 4°C for ≤1 week. bPremixed MEM/BMEamino acids were from Sigma Chemicals Co. cEleven amino acids were prepared (100x) and storedat -80°C before use.

44
5. BM-3 + 20 amino acids + 5.5 mM+ lactic acid (= BM-3-20aa-lact)
6. BM-3 + 20aa + 0.35 mM potassium bisphosphate (= BM-3-20aa-phos)
7. BM-3 + 20aa + 0.2 mM gluc + 0.2 mM pyr + 5.5 mM lact + 0.35 mM phos (= BM-3 +
All).
The two osmotic pressure groups of BM-3 were ~255 (designated “low”) and ~275
(designated “high”). The “high” osmotic pressure BM-3 was the regular BM-3 formulation
(McKiernan et al. 1995). However, treatments containing additional lactate required adjustments for
osmotic pressure. This was achieved by replacing the medium with ultrapure water (~7.3%, vol/vol).
In “low” BM-3, 95 mM NaCl (as in KSOM) was used and the osmolality in Treatments 2, 3, 4 and 6
was adjusted using sodium sulfate (Na2SO4). The latter has no known toxic or beneficial effects on
embryo development in vitro (Monis and Bavister 1990). Lactate-supplemented treatments in low
BM-3 did not require further adjustments. HEPES was excluded from modified KSOM (Control) in
Experiment 3.2.
3.2.5. Evaluation of Embryonic Development and Cell Counts
Frequencies of cleavage and development to 8-cell were evaluated on day 2 and day 3 post-
insemination. Advanced development was evaluated on day 7, day 8 and day 9. Blastocysts were
subjectively classified as “Expanded” when increment in diameter was noticeable during microscopic
examination. Blastocysts were fixed on day 9 and subjected to nuclear staining using a combination
of propidium iodide and Hoechst stains (Thouas et al. 2001) to achieve differential cell counts on
trophectoderm and inner cell mass. However, reliable differentiation could be achieved on only a
small fraction of embryos, so the values were combined and used as total cell counts. In the latter
part of the experiment, Hoechst staining alone was used for nuclear staining. Cells were counted
under a fluorescent microscope equipped with Nomarski differential interference optics.

45
3.2.6. Data Analyses
Percentage values for embryonic development were arcsine transformed before analyses.
Data were analyzed using the SAS software (SAS 2001). In Experiment 3.1, the general linear
model was used. In Experiment 3.2, the mixed model was used with supplement and osmotic
pressure groups included in the model as fixed effects and replicate and its interaction as random
variables. Tukey’s test was used for pair-wise comparisons of significantly different means. Data on
cell counts in Experiment 3.1 were not normally distributed and were analyzed using the Kruskal-
Wallis test. For the analyses, blastocysts observed escaping from their zonae pellucidae and those
that had completed this process were considered “hatched”. Differences in treatment means were
considered significant at P ≤ 0.05.
3.3. Results
3.3.1. Experiment 3.1 (Effects of Amino Acids and Base Media)
A total of 1,109 IVM oocytes were subjected to IVF in five replicates across days. Cleavage
frequency ranged from 61% (mKSOM-m11aa) to 75% (BM-3 + 20aa) (P>0.05; Table 3.3). The
frequency of blastocyst development continued to increase from days 7 to 9 in all treatments (Table
3.3). However, the percentage of total and expanded blastocysts was significantly higher in
mKSOM-20aa than in the other four treatments during these three days (P<0.05; Tables 3.3; 3.4).
The frequency of hatched blastocysts on day 7 was virtually zero in all treatments. On days 8
and 9, hatching frequency was significantly higher in mKSOM-20aa than in the other treatments
(Table 3.4). Blastocysts from this treatment also had the numerically highest cell numbers; however,
this was not significantly different from the three other treatments (Tables 3.3).

46
Table 3.3. Development frequencies and cell count of in vitro-derived bovine embryos cultured inprotein-free BM-3 or mKSOM supplemented with two groups of amino acids____________________________________________________________________________Treatment Blastocysts* Cell countgroup n ≥2-cell* day 7 day 8 day 9 (n) median____________________________________________________________________________
HECM-6 223 70.8 ± 7.4 5.9 ± 2.0a 13.6 ± 1.8a 16.9 ± 1.7a (23) 55.0ab
BM-3-m11aa 218 64.9 ± 8.8 1.7 ± 0.8a 7.6 ± 0.8a 10.2 ± 2.1a (12) 43.5a
BM-3-20aa 230 75.0 ± 8.7 1.6 ± 1.1a 10.2 ± 1.6a 12.6 ± 3.4a (25) 60.0ab
mKSOM-m11aa 203 60.9 ± 8.6 2.8 ± 1.8a 7.9 ± 1.6a 12.8 ± 3.3a (16) 53.0ab
mKSOM-20aa 235 73.9 ± 7.8 15.9 ± 6.5b 31.1 ± 7.1b 42.1 ± 8.5b (48) 82.0b
____________________________________________________________________________a,bValues in the same column with different superscripts are significantly different (P<0.05). *Figuresindicate % of oocytes ± SE.
Table 3.4. Blastocyst expansion and hatching frequencies of in vitro-derived bovine embryos culturedin protein-free BM-3 or mKSOM supplemented with two groups of amino acids
________________________________________________________________________________Treatment Expanded blastocysts* Hatched blastocysts* _group day 7 day 8 day 9 day 7 day 8 day 9________________________________________________________________________________
HECM-6 1.1 ± 1.1a 7.0 ± 1.1a 7.6 ± 1.2a 0 0a 0a
BM-3-m11aa 0.4 ± 0.4a 3.4 ± 0.9a 6.7 ± 2.1a 0 0a 0.4 ± 0.4a
BM-3-20aa 0a 4.2 ± 1.7a 9.5 ± 3.4a 0 0a 0.4 ± 0.4a
mKSOM-m11aa 0.9 ± 0.6a 4.4 ± 1.3a 6.3 ± 3.1a 0 0a 0a
mKSOM-20aa 8.3 ± 4.4b 19.7 ± 6.8b 27.8 ± 6.0b 0.4 ± 0.4 3.4 ± 1.7b 15.0 ± 5.6b
________________________________________________________________________________a,bValues in the same column with different superscripts are significantly different (P<0.05). *Figuresindicate % of oocytes ± SE.

47
3.3.2. Experiment 3.2 (Effects of Glucose, Pyruvate, Lactate and Phosphate in BM-3Having Two Osmolalities)
A total of 1,921 oocytes were used in six replicates. Neither supplement type nor osmotic
pressure affected the frequencies of initial cleavage and development to the 8-cell stage (Table
3.5). Supplement but not osmotic pressure affected the frequency development to at the least the
morula stage (day 7). The latter was lowest (36 to 38%) in pyruvate-supplemented media and highest
(68%)in glucose-supplemented media (Tables 3.5; 3.6). Similarly, supplement treatment affected the
frequency of total (day 9), expanded (day 8) and hatched (day 9) blastocysts. Effects of supplement
on the frequencies of total blastocysts on days 7 and 8, and expanded blastocysts on day 9 also
approached statistical significance (Table 3.5).
Regardless of treatment type, blastocyst development frequency peaked on day 8 (Figures
3.1; 3.2). The frequency of hatching continued to increase through day 9 (Tables 3.5; 3.6). Blastocyst
cell count was affected by both supplement type and osmotic pressure; however, supplement exerted
a more pronounced effect (Tables 3.5; 3.6). Osmotic pressure affected the frequency of blastocyst
expansion on day 7. Its effect on the frequencies of day 8 blastocyst expansion and day 9 hatching
also approached statistical significance. Generally, low osmotic pressure BM-3 treatments tended to
increase frequency of blastocyst expansion and hatching as well as cell counts. However, a
significant difference was detected only between the frequencies of day-7 expanded blastocysts in
BM-3-20aa treatments and between blastocyst cell counts in BM-3-20aa-phos treatments (Tables 3.6;
3.7).
Among the BM-3-based treatments, glucose-supplemented media supported (at least
numerically) superior embryonic development (Tables 3.6; 3.7; Figures 3.1; 3.2). In high BM-3, the
frequency of day-9 hatched blastocysts in glucose-supplemented media was significantly greater than
in all non-glucose supplemented treatments. Similarly, among low BM-3 treatments, the frequency of

48
day 9 hatching was significantly higher in glucose-supplemented medium than in all other BM-3
treatments (Table 3.7). Blastocysts produced in glucose-supplemented treatments also had greater
cell numbers than those in other treatments (Table 3.6). The effect of interaction between osmotic
pressure and supplement groups was not significant for any of the parameters evaluated (Table 3.5).
Table 3.5. Effects of osmotic pressure of modified BM-3 and supplements on the development ofin vitro-derived bovine embryos at different stages_________________________________________________________________________________
P-value Variables Osmotic pressure Supplement Interaction
_________________________________________________________________________________
≥2-cell (day 2) 0.8761 0.3870 0.8111
≥8-cell (day 3) 0.2518 0.3362 0.8638
≥Morula (day 7) 0.2237 0.0317 0.4047
Blastocysts
- Total (day 7) 0.3507 0.0978 0.5743
- Total (day 8) 0.1536 0.0592 0.6531
- Total (day 9) 0.4766 0.0414 0.8967
- Expanded (day 7) 0.0137 0.4699 0.5066
- Expanded (day 8) 0.0953 0.0360 0.1677
- Expanded (day 9) 0.3879 0.0904 0.7504
- Hatched (day 7) 0.1790 0.5017 0.6030
- Hatched (day 8) 0.3879 0.0904 0.7504
- Hatched (day 9) 0.0904 0.0003 0.8615
- Cell count (day 9) 0.0279 <0.0001 0.6769________________________________________________________________________________

49
Table 3.6. Effects of glucose, pyruvate, lactate and phosphate in high or low osmotic pressure modified BM-3 on the frequency of cleavage(day 2) and development to morula (day 7) and blastocyst (day 9) stages of in vitro-derived bovine embryos cultured in protein-free media______________________________________________________________________________________________________________Treatment High Low _group n (≥2-cell) ≥Morula Blastocyst Cell count n (≥2-cell) ≥Morula Blastocyst Cell count______________________________________________________________________________________________________________________________
KSOM-20aa 154 (84.7 ± 2.1) 63.2 ± 8.5ac 29.2 ± 2.3ab 81.6ac 127 (79.6 ± 2.3) 53.6 ± 6.1ab 27.1 ± 4.1 93.1ac
BM-3-20aa 140 (80.9 ± 2.4) 43.0 ± 7.9ab 20.1 ± 4.2bc 57.3b 126 (82.7 ± 2.5) 63.6 ± 12.9b 25.1 ± 6.3 66.2b
BM-3-20aa-gluc 154 (85.9 ± 4.1) 68.3 ± 9.2c 32.6 ± 2.5a 94.7c 113 (85.5 ± 3.0) 68.0 ± 5.0b 29.8 ± 3.8 113.0c
BM-3-20aa-pyr 153 (76.0 ± 3.2) 37.5 ± 6.1b 18.2 ± 2.0c 58.2b 124 (80.3 ± 3.1) 35.5 ± 9.7a 23.9 ± 3.1 69.2bd
BM-3-20aa-lact 149 (80.1 ± 3.1) 45.3 ± 12.1abc 24.6 ± 3.7abc 69.2a 131 (79.6 ± 3.4) 52.3 ± 9.0ab 27.6 ± 2.6 81.1ad
BM-3-20aa-phos 138 (80.6 ± 7.1) 34.8 ± 10.5b 22.7 ± 4.1abc 59.3b 134 (79.0 ± 4.1) 50.8 ± 11.8ab 22.9 ± 4.1 84.6a
BM-3aa + All 155 (82.0 ± 3.0) 51.3 ± 9.3abc 19.9 ± 3.9bc 109.7c 123 (84.2 ± 4.5) 57.5 ± 2.4ab 20.3 ± 5.5 102.4ac
_____________________________________________________________________________________________________________Values under 2-cell, Morula and Blastocyst headings represent mean (%) of oocytes inseminated ± SE. abcdValues in the same column withdifferent superscripts are significantly different (P<0.05). There was no statistical difference within a supplement treatment across osmoticpressure group except for cell count values of in BM-3-20aa-phos.

50
Table 3.7. Effects of glucose, pyruvate, lactate and phosphate in high or low osmotic pressure modified BM-3 on the frequency (%) ofblastocyst expansion (days 7 and 8) and hatching (day 9) in vitro-derived bovine embryos
_________________________________________________________________________________________________________Treatment High Low group day 7 expanded day 8 expanded day 9 hatched day 7 expanded day 8 expanded day 9 hatched_________________________________________________________________________________________________________
KSOM-20aa 8.4 18.9a 5.3a,b 8.3 14.5a,b 8.2b,c
BM-3-20aa 2.2 5.8bc 0.0a 10.5 20.5b 3.6a,b
BM-3-20aa-gluc 5.3 19.1a 10.6b 10.0 19.0a,b 11.9c
BM-3-20aa-pyr 3.6 6.6bc 0.0a 1.6 9.5a 1.4a
BM-3-20aa-lact 2.6 7.2bc 3.1a 9.7 12.2a,b 5.1a,b
BM-3-20aa-phos 2.9 8.7bc 0.0a 13.8 16.5a,b 1.3a
BM-3 + All 3.8 11.5ac 5.2a,b 9.9 10.2a 4.1a,b
_________________________________________________________________________________________________________a,b,cValues in the same column with different superscripts are significantly different (P<0.05). There was no statistical differencewithin a supplement treatment across osmotic pressure group except for frequencies of day 7 expanded blastocysts in BM-3-20aa.

51
Figure 3.1. Blastocyst development of in vitro-derived bovine embryos cultured in high osmoticpressure BM-3-20aa with different supplements (see Table 3.6 for day 9 development data).
0
5
10
15
20
25
30
35
40
KSOM-20aa
BM-3-20aa BM-3-20aa-gluc
BM-3-20aa-pyr
BM-3-20aa-lact
BM-3-20aa-phos
BM-3 + all
Mea
n (%
of o
ocyt
es)
± S
E
Blastocyst (day 7)
Blastocyst (day 8)

52
Figure 3.2. Blastocyst development of in vitro-derived bovine embryos cultured in low osmoticpressure BM-3-20aa with different supplements (see Table 3.6 for day 9 development data).
0
5
10
15
20
25
30
35
KSOM-20aa
BM-3-20aa BM-3-20aa-gluc
BM-3-20aa-pyr
BM-3-20aa-lact
BM-3-20aa-phos
BM-3 + all
Mea
n (%
of o
ocyt
es)
± S
E
Blastocyst (day 7)
Blastocyst (day 8)

53
3.4. Discussion
In the present study, effects on in vitro embryonic development of selected supplements and
osmotic pressure of culture media were evaluated. Protein-free HECM-6 (i.e., BM-3 + 11 amino
acids) supports complete preimplantation development of hamster embryos that are capable of
developing into normal offspring after embryo transfer; however, HECM-6 components have not
been systematically evaluated in bovine IVC. In Experiment 3.1, the much higher blastocyst
development on day 9 in modified KSOM-20aa than in the other treatments (Table 3.3) was
puzzling; hence, Experiment 3.2 was designed. We demonstrated that 20 amino acids and glucose
(0.2 mM) are crucial for culturing bovine embryos in BM-3.
In Experiment 3.1, mKSOM-20aa supported blastocyst development (42%) at similar
frequencies to other commonly used culture media [e.g., M199 or SOF plus serum or BSA, G1/G2 or
coculture: see (Holm et al. 1999; Krisher et al. 1999; Steeves and Gardner 1999; Lane et al. 2003)].
The frequency of day 7 blastocysts was relatively low (16%) indicating slow embryonic development
in mKSOM-20aa; but the frequency increased on subsequent days and reached 42% on day 9 (Table
3.3). The slow development indicates reduced developmental competence as reported for bovine and
other species (Massip et al. 1995; Bavister 2002b).
KSOM has been used to culture bovine embryos under defined (Liu and Foote 1995a; Liu
and Foote 1997) or complex (Al Katanani et al. 2002) conditions. In vitro development in protein-
free KSOM in previous studies (Liu and Foote 1995a; Liu and Foote 1997) was lower than in the
present study, possibly due to variations in the concentration of amino acids used or the stage at
which they were supplemented. For example, at concentrations employed for tissue culture (2%),
BME amino acids inhibit early-stage development of bovine embryos (Keskintepe et al. 1995; Liu
and Foote 1995b; Steeves and Gardner 1999).
Increasing the concentration of HECM-6 amino acids to that of BME/MEM did not improve
embryonic development beyond that seen in HECM-6 (Table 3.3). The greater frequency of embryo

54
development in the presence of 20 amino acids as compared with 11 amino acids suggests that
bovine and hamster embryos differ in their amino acid requirements. Other investigators (Liu and
Foote 1997) also did not observe further improvements in the in vitro development of bovine
embryos when the concentration of non-essential amino acids was increased up to five times. This is
paradoxical, considering that the concentration of free amino acids in the reproductive tract of cows
is >30 mM (Elhassan et al. 2001). The total concentration of amino acids used in the present study
was only 2.5 mM (20 amino acids), 0.8 mM (11 amino acids) and 2.3 mM (modified 11 amino
acids). Methionine, an amino acid that is essential to initiate protein synthesis is not one of the 11
(HECM-6) amino acids. Blastocyst expansion and, even at times, hatching occurred in media
supplemented with HECM-6 amino acids, suggesting that protein synthesis did take place without
the addition of external methionine. Thus, like mouse oocytes (Wassarman and Albertini 1994),
bovine oocytes probably store methionine.
In Experiment 3.1, blastocyst hatching was rare (<1%) in treatments other than mKSOM-
20aa (15% on day 9; Table 3.4). Others have also observed reduced hatching frequencies in bovine
embryos cultured in protein-free media (Krisher et al. 1999). Hatching is also reduced in bovine
blastocysts resulting from cumulus denuded oocytes matured in defined media (Holm et al. 1999;
Landim-Alvarenga et al. 2002). Nevertheless, modifications of defined media can improve hatching
frequencies [(Holm et al. 1999); the present study]. For example, using low osmotic pressure BM-3-
20aa with glucose increased the hatching frequency more than threefold (Table 3.7).
In Experiment 3.2, BM-3-20aa supplemented with glucose alone stood out in supporting a
high frequency of development comparable to KSOM-20aain (Table 3.6; Figures 3.1; 3.2). This is in
contrast to the widely held dogma that glucose inhibits early embryonic development, especially at
concentrations around physiologically normal blood levels and/or in the presence of phosphate
(Pinyopummintr and Bavister 1991; Takahashi and First 1992; Kim et al. 1993).
Most investigators agree on the need for glucose during culture of bovine morula and
blastocyst stage embryos. In the present study, 0.2 mM glucose during the entire IVC period

55
(including the pre-cleavage stage) was also beneficial. Although data on temporal metabolism of
glucose during the entire pre-elongation development of bovine embryos are limited, gene transcripts
and proteins involved in glucose transport and metabolism are present during all stages. Hence, it has
been suggested that substrate availability and not proteins are determinants of glucose metabolism in
early stage mammalian embryos (Gardner et al. 2000). The incubation environment or condition
could also modify the effect of glucose (Yamashita et al. 1999).
In the present study, the higher frequency of in vitro embryonic development in pyruvate-free
but glucose-supplemented medium, as compared with pyruvate-containing medium, suggests a more
important role of glucose in pathways other than glycolysis and/or the Krebs cycle. Moreover,
endogenous pyruvate could be adequate for energy metabolism. In fact, cleavage stage mammalian
embryos have a high ATP/ADP ratio and additional ATP is probably not required until after
embryonic genome activation (Gardner et al. 2000). Bovine embryos with reduced glycolytic activity
are developmentally more competent than those with high glycolysis (Rieger et al. 1995) and
embryos cultured in defined medium have low glycolytic activity that is similar to in vivo-derived
embryos (Krisher et al. 1999). Similarly, the gene expression pattern of bovine embryos cultured in
chemically defined medium is more similar to that of in vivo derived cattle embryos when compared
with embryos cultured in serum-supplemented medium (Wrenzycki et al. 1999). Furthermore,
morphologically, bovine embryos produced in defined or semi-defined media are more similar to
those derived in vivo than those embryos produced in complex media (Abe et al. 2002).
Alternative routes of glucose utilization include the pentose-phosphate pathway, which
generates intermediates for nucleic acid and lipid biosynthesis, and the hexosamine pathways which
are required for post-translational glycosylation of proteins [(Gardner et al. 2000; Ludwig et al.
2001); Figure 3.3]. During fertilization, oxidation of glucose is more important than that of pyruvate
(Pantaleon et al. 2001). The beneficial effects of glucose during mouse embryo culture also led to a
recommendation for increasing its concentration in KSOM from 0.2 to 3.4 mM (Biggers and
McGinnis 2001).

56
Pyruvate, phosphate and glucose are not components of HECM-6 (i.e., BM-3 + 11aa), which
has been shown to support bovine embryo development during the first 2 to 4 or 7 days of culture
(Pinyopummintr and Bavister 1996; Krisher et al. 1999). In the latter study (Krisher et al. 1999),
21% of inseminated oocytes developed to blastocysts with an average cell number of 73 after IVF,
and subsequent IVC in HECM-6 supplemented with pyruvate (0.25 mM) and glucose (2 mM). The
blastocyst frequency and cell count of embryos cultured in BM-3-20aa containing 0.2 mM glucose in
the present study were higher than in the previous report by Krisher et al. (1999), when HECM-6 was
supplemented with higher concentrations of glucose and pyruvate. Furthermore, in the present study,
pyruvate appeared to be inhibiting blastocyst development in high BM-3-20aa compared with the
same medium with glucose (Table 3.6).
Others (Yoshioka et al. 1993) also reported improved development of in vitro-derived bovine
embryos in a semi-defined medium when the concentrations of lactate and pyruvate were reduced
(from 20.1 to 3.0 mM and from 0.5 to 0.3 mM, respectively). Pyruvate was also reported to be either
unnecessary or toxic to bovine embryos in a semi-defined medium (Rosenkrans, Jr. et al. 1993) and it
is not a crucial supplement in sheep IVC medium (Thompson et al. 1992). Taken together, these data
underscore the uncertainty surrounding the beneficial role of external pyruvate, although it is
believed to be the preferred energy substrate for bovine embryos (Massip et al. 1995; Khurana and
Niemann 2000). Anti-oxidant properties of pyruvate have also been reported (Gardner et al. 2000);
however, such a role of pyruvate in defined IVC medium awaits confirmation.
Glycolysis and Krebs cycle are closely related such that imbalances in key substrates could
affect either pathway. The “Crabtree effect” is a prototype example in which upregulation of
glycolysis inhibits oxidative phosphorylation (Seshagiri and Bavister 1991). Data are scant on the
kinetics of the Krebs cycle enzymes during pre-elongation embryonic development; however, the
regulatory enzyme of glycolysis (phosphofructokinase) has low activity prior to mouse embryonic
genome activation (Barbehenn et al. 1974). High glucose (5.6 mM) in IVC media could prematurely
up-regulate glycolysis and/or activate the polyol pathway (Moley et al. 1996). The latter depletes

57
Glucose G-6-P F-6-P PFKF-1,6-P ... Pyruvate
Lactate
Krebs Cycle
PentosePhosphatePathway
F-2,6-P
Glyceraldehyde-3-P
Ribose-5-P Nucleic acidsynthesis
Ald
ose
redu
ctas
e
Sorbitol
NADPH
NADP
NADPHGSH
Fructose
Apoptosis
Macromolecules
Osmotic stress
High glucoseNAD
NADH
Figure 3.3. Pathways of glucose metabolism and related factors influencing embryonic development:The regulatory enzyme of glycolysis, phosphofructokinase (PFK), has low activity in early cleavage-stage mammalian embryos. High glucose could lead to premature activation of PFK and/or up-regulation of aldose reductase and subsequent accumulation of sorbitol in the cytoplasm. Osmoticstress may also stimulate the conversion of glucose to sorbitol, the formation of which depletesglutathione (GSH, an antioxidant) and up-regulates apoptotic pathways. Removal of sorbitol fromcells requires its conversion to fructose, a step that produces NADH that can subsequently inhibitoxidative phosphorylation in the mitochondria. The Pentose Phosphate Pathway produces severalimportant intermediates, including fructose-2,6-bisphosphate- the key allosteric activator of PFK[(Roskoski, Jr. 1996) and see also text for relevant references].

58
NADPH and could stimulate apoptotic pathways [(Galvez et al. 2003); Figure 3.3]
Compared with observations in Experiment 3.1, embryonic development in KSOM was
reduced in Experiment 3.2. The reason for this is not clear. The exclusion of HEPES was an unlikely
reason because adding it back in a sideline trial did not improve embryonic development during
Experiment 3.2 (personal observation). A possible reason is seasonal variation in oocyte quality,
which is poor during the hot (Summer) season (Zeron et al. 2001; Al Katanani et al. 2002).
(Experiment 3.1 was done in Fall and Experiment 3.2 was performed in Spring/Summer seasons).
Nevertheless, embryonic development in the control KSOM-20aa treatment was similar during
replicates run with high or low osmotic pressure BM-3 treatments.
Previous studies (Lim et al. 1994; Liu and Foote 1996) recommended reducing the osmotic
pressure of bovine IVC media to <270 but the most widely used media (SOF, TCM199, CR1) have
higher osmotic pressures than this. Reduced osmotic pressure with a concomitant reduction in [NaCl]
is among the factors responsible for improved development of mouse embryos in KSOM (Summers
et al. 1995). The use of low osmotic pressure BM-3 appeared to minimize or eliminate treatment
differences (Tables 3.6; 3.7) observed in the frequency of embryonic development in high BM-3.
Moreover, although there were large variations in embryonic development, low osmotic pressure
BM-3-20aa supported blastocyst development at a similar frequency as the same medium containing
glucose (Table 3.6; Figure 3.2). This may indicate the beneficial role of low osmotic pressure for
culturing bovine embryos as demonstrated previously (Lim et al. 1994; Liu and Foote 1996).
In most instances, increased osmotic pressure of culture media is synonymous with high
[NaCl]; however, extracellular Na+ affects intracellular pH via its effect on the Na+/H+ anti-porter
(Lane and Bavister 1999). Reducing extracellular [NaCl] also alters gene expression (Ho et al. 1994)
and protein synthesis (Liu and Foote 1997). Osmotic stress could activate the polyol pathway and
apoptosis [(Galvez et al. 2003); Figure 3.3]. The higher frequency of blastocyst expansion and
hatching, and mean cell numbers per blastocyst, in low BM-3 treatments suggests a similar
mechanism is operating.

59
In the present study, 20aa and glucose (0.2 mM) used during the entire IVC period were
beneficial. Glucose supported superior development in medium with low lactate (4.5 mM)
concentration and without pyruvate or phosphate. Although the mechanism by which glucose exerts
its effects during embryo development is not fully elucidated, its effects are evident as early as the
fertilization stage in mice and bovine (Pantaleon et al. 2001; Comizzoli et al. 2003). Moreover,
glucose metabolism via the pentose phosphate pathway provides intermediates for the synthesis of
key macromolecules and is important for redox regulation (Figure 3.3).
In another study (Wirtu et al. 2004), further supplementation of protein-free mKSOM-20aa
with insulin improved cleavage and blastocyst formation frequencies of in vitro-derived bovine
embryos. In addition, it increased cell numbers per blastocyst. With additional serum
supplementation during IVC of bovine embryos, BM-3-20aa containing glucose supported up to 41%
blastocyst formation (Wirtu et al. 2003). Similarly, up to 27% blastocysts developed in this medium
after culturing interspecies SCNT couplets reconstructed by using giant eland cells and bovine
oocytes (Damiani et al., 2003).
The respectable levels of in vitro development of bovine embryos in medium with reduced
substrates (i.e., no pyruvate and phosphate, low lactate and glucose concentrations; Tables 3.6; 3.7)
are also consistent with the hypothesis that reducing embryonic metabolism by restricting medium
supplements improves viability (Leese 2002). In conclusion, the beneficial effects of amino acids on
the in vitro development of bovine embryos are influenced by the base medium. Low osmotic
pressure facilitates blastocyst expansion, while medium supplements are more important in
determining the frequency of total blastocysts and cell number per blastocyst. Further evaluation of
the viability of such embryos by using additional end points, such as development following embryo
transfer, is needed.

60
Chapter 4. Handling Elands for Reproductive Technology Procedures withoutInducing General Anesthesia: Roles of Behavioral Training and Hydraulic
Chute
4.1. Introduction
Handling of most nondomestic ungulates for clinical or other procedures has traditionally
been carried out using complete chemical immobilization. Most assisted reproductive technology
studies in tragelaphine (spiral-horned) antelopes also used general anesthesia to perform semen
collection (Merilan et al. 1977; Merilan et al. 1978; Schiewe et al. 1991a; Schiewe et al. 1991b),
embryo recovery (Dresser et al. 1985; Gelwicks et al. 1989; Schiewe et al. 1991b), embryo transfer
(Dresser et al. 1985; Gelwicks et al. 1989) and artificial insemination (Bartels et al. 2001). However,
several risk factors limit the routine use of general anesthesia for the development and application of
reproductive technologies.
General anesthesia predisposes animals to the risk of cardiac and respiratory arrest, and long-
term cell/organ injuries. It can also interfere with normal endocrine responses and gamete and/or
embryo transport (Loskutoff and Betteridge 1992). Moreover, narcotic agents used for
immobilization pose a potential health hazard to the personnel participating in procedures. Animals
have also been known to renarcotize and die subsequent to narcotic general anesthesia (Domínguez
and Aguilar 2001). Elands (Taurotragus oryx) are among the bovid species considered difficult to
immobilize (Citino 2003). In addition, their athletic ability and flighty behavior complicates the
induction phase and could affect the outcome of anesthesia event.
Behavioral training and habituation (“conditioning and desensitization”) of animals has been
used to avoid the need for inducing general anesthesia while conducting selected procedures in many
mammalian species including ungulates, carnivores, primates and marine mammals (Dumonceaux et
al. 1998; Grandin et al. 1995; Phillips et al. 1998). Conditioning of common eland to sound cues has

61
been incorporated into ungulate management at Disney’s Animal Kingdom (Joe Kalla and Marty
Sevenich, personal communication).
While the use of advanced mechanical handling devices has enabled clinicians and scientists to
perform certain procedures without inducing general anesthesia in several ungulate species (Read et
al. 1993; Citino 1995; Atkinson et al. 1999), the use of these devices combined with behavioral
training has not been evaluated for performing reproductive technology procedures in tragelaphine
antelopes. Data on the safety or stressful effect of such handling are not available. Moreover,
although it has been reported that individual elands have unique identifying physical and behavioral
attributes (Kiley-Worthington 1978), it is unknown if physical characteristics influence the taming
potential or the manner in which specific eland respond to training or handling. Thus, the hypotheses
of the current study were:
1. female elands in a small group can be conditioned to associate specific sound cues with
food treats; and
2. sedated female elands will tolerate handling in a hydraulic chute during assisted
reproductive technology procedures.
Specific objectives were to:
1. evaluate the feasibility of conditioning individual female elands in a group of 10 to
specific sound cues;
2. assess the use of conditioned stimuli to move female eland into a hydraulic chute;
3. evaluate the physiological responses of elands during sedation and handling in a
hydraulic chute; and
4. determine, retrospectively, if the degree of cooperation with training and handling
(taming potential) is associated with two selected physical attributes of the elands.

62
4.2. Materials and Methods
4.2.1. Study Animals and Management
Animals were maintained at the Freeport-McMoran Species Survival Center, a facility of the
Audubon Nature Institute, located on the outskirts of New Orleans, Louisiana. The area has a semi-
tropical climate. The study was carried out over a period of three years (June 2000 to June 2003)
using a group of 10 adult elands. Animals were housed on an open yard (45 m x 56 m, plot A; Figure
4.1) surrounded by a 2.4 m high woven wire fence.
The yard has a feeding area (10 m x 12 m, see plot B) with a concrete floor. Feed was placed
in a trough on both sides of a shade structure (7.6 m x 2.5 m) built in the center of the feeding area.
The herd was fed daily as a single group with a bale of grass hay and 41 kg of pelleted concentrate
(Mazuri® CU ADF-16, PMI Nutrition International, Brentwood, MO, USA). The concentrate
contained a minimum of 17 % crude protein and 3 % crude fat, and a maximum of 15 % crude and
16 % acid detergent fiber. Water and mineral lick block were provided ad libitum.
Routine veterinary care included anthelminthic treatments twice a year by alternating among
fenbendazole (Panacur®, DPT Laboratories, San Antonio, TX, USA), pyrantel pamoate (Strongid®-T,
Pfizer Animal Health, Exton, PA, USA) and ivermectin (Ivomec®, Merial Ltd, Iselin, NJ, USA).
Vitamin E and selenium (BO-SE®, Schering Plough Animal Health Corp, Union, NJ, USA) were
also administered twice yearly. Animals received annual vaccinations against rabies (RabvacTM-3,
Fort Dodge Animal Health, Fort Dodge, IA, USA), Clostridia (Fortress®-7, Pfizer Animal Health,
Exton, Pennsylvania, USA) and Leptospira (Leptoferm-5®, Pfizer Animal Health). Fecal samples
were examined quarterly for parasite ova, and insecticides/repellents were applied as needed. Animal
procedures were carried out with the approval of the Institutional Animal Care and Use Committee of
the Audubon Center for Research of Endangered Species, New Orleans, Louisiana 70131, USA.

63
Figure 4.1. Sketch of the eland holding area and associated structures:
Plots include the main holding area for females (A), feeding area with a shade (B), temporaryholding area with a shade (C), holding and feeding areas for a male eland (D, E), keepers area (F)and barns (G, H) leading into a hydraulic chute (I). Gates are numbered from No. 1 to No. 27 anddoors inside the structure are remotely operable. Gates No. 25 and No. 26 lead to loading andunloading areas. Barn H is equipped with a remotely operable push-wall that can be moved betweenthe areas of Gate No. 14 and Gate No. 16. Sketch not drawn to scale.
D
EH
I
Hydraulic pump
Side accessSide access
1 2
3
5
72120
4
6
8910
11
13
14
16 17 18 19
22
2324
1227
15
26
25
45 m
16 m
29 m
16 m 6 m
56 m
A
B
C
F
G

64
4.2.2. Conditioning to Sound Cues
During animal training, classical or Pavlovian conditioning is often used to “bridge” primary
reinforcers (e.g., feed) with secondary stimuli such as clickers (www.wagntrain.com/OC/#Operant,
accessed 23 November 2003). Thus, feed rewards (treats) other than the regular diet were used to
condition animals to specific sound cues. Elands readily ate treats such as tree branches, spinach,
carrots, alfalfa hay and bananas; however, alfalfa and banana were preferred the most. We chose
bananas as a treat item because they can be hand-fed or by throwing it into the holding area; and
medications can be embedded in them for feeding them to a specific animal. Bananas are also stable
for several weeks when stored at 4°C, which makes them desirable.
The two criteria used to select sound producing tools were: 1) no two tools should produce
similar sounds to prevent confusion among animals of sound cues and 2) at least one hand should be
free for providing treats and/or recording observations. Examples of sound cue tools matched to each
female (tool: eland ID) were: clicker (KP-600R, Sunshine Books, Inc, Waltham, MA, USA): 125;
tapping pant leg with a hand: 140; coach’s whistle: 124; mouth whistling: 139; etc. We did not
attempt to condition one female (eland 128) because it was not possible to train her to accept
handheld treats; however, in an attempt to get treats, she would always approach the training area and
chase away subordinate females. She was also the most aggressive female and often attempted to
charge people standing outside of the fenced pen.
Conditioning to sound cues was conducted daily between June and August 2000 in sessions
of up to 4 hr each, in the vicinity of Gate No. 1 (Figure 4.1), while monitoring the response of
animals through the fence. To condition each animal to sound cues, treats were thrown into the
holding area near an animal or hand-fed to those females (elands 125, 123, 129, 139 and 140) already
accepting hand-fed treats. The specific sound cue was then produced continuously while the animal
was eating the treat. To avoid competition for treats and confusion over the conditioned sound cues, a

65
second person kept other females as far away as possible from the animal being conditioned.
Most training was conducted in the morning (before animals were fed). We also determined,
in a pilot study, the interval from producing conditioned audio cues to acceptance of handheld treats
by females. This response was evaluated in the morning (before feeding) and afternoon (3 to 4 hr
after feeding) by inviting each female, using the conditioned cue, on at least three different days. The
interval was recorded in whole number minutes.
4.2.3. Chute Training
After achieving positive association between sound cues and treats, the cues were then used
to move females, initially around the holding area (plots A and B, Figure 4.1) and then to entice each
female to the chute area, and subsequently, into the chute (plot I, Figure 4.1). The chute (The
TamerTM, Fauna Research, Inc., Red Hook, NY, USA) was padded internally and could be operated
using a hydraulic pump to restrain (squeeze) and lift an animal. Atkinson and colleagues have
provided a more detailed description of this device (Atkinson et al. 1999).
Animals were “called” or “invited” into the vicinity of Gates No. 15 to No. 18 via Gates No.
8 and No. 20. Invitation of females and observation of their responses was done through one of the
two sliding doors on either side of the chute. After repeated and consistent voluntary entry into the
chute, females were allowed to remain in the chute during its activation. Chute use was gradually
increased from simply turning on the hydraulic pump to partially squeezing animals. The response of
animals to chute training was graded from 0 to 6 scores as described next:
0 = does not approach the chute/does not enter the barn via Gate No. 18 (Figure 4.1);
1 = enters the barn but does not come close to the chute gate (Plot I, Figure 4.1);
2 = approaches the chute gate, takes hand-held treat at the gate before retreating back;
3 = approaches the chute gate and stays taking handheld treat;
4 = partially enters the chute, gets treat and stays for a prolonged period;

66
5 = enters the chute voluntarily; and,
6 = fulfills criterion No. 5 and allows being touched and/or tolerates closure of chute gate,
noise of hydraulic pump and partial squeezing of sides of the chute.
4.2.4. Use of Sedatives and Physiological Effects of Handling in a Hydraulic Chute
Tranquilization protocols and the safety of carrying out procedures in the chute were initially
established from trials done on the three most cooperative females (elands 123, 125, 140). Body
weights, taken by placing a flatbed AltraliteTM scale (Rice Lake Weighing Systems, PA, USA) on the
floor in the hydraulic chute, were used for calculating dosage of sedative agents. Two separate
neuroleptic agents (haloperidol, acepromazine), an α-2 agonist (xylazine) alone, and a combination
of xylazine with the opioid analgesic (butorphanol) were evaluated. The regular feed of the group
was reduced by one half on the day before sedation trials.
Each female was treated with 1 mg/kg of haloperidol lactate (Geneva Pharmaceuticals, Inc.
Broomfield, CO, USA) given orally embedded in bananas. Animals were monitored both inside and
outside the chute for approximately 6 hr after treatment and for the following three days. Attempts
were made to partially squeeze the animals in the chute. Due to inadequate sedation with the initial
regimen, the dosage was increased to a single treatment of 1.5 mg/kg or three consecutive daily
treatments of 1.25 mg/kg.
One animal was hand-injected in the chute with 10 mg of acepromazine maleate (Boehringer
Ingelheim, St. Joseph, MO, USA), i.m. and monitored during the first hour after injection. Sufficient
sedation to allow animal handling for reproductive procedures was not achieved with either
haloperidol or acepromazine. Thus, xylazine HCl (Xyla-Ject®, Phoenix Pharmaceutical, Inc., St.
Joseph, MO, USA) was chosen as the next sedative agent for evaluation.
Literature information on the use of xylazine alone for sedating eland was not available;

67
however, up to 150 mg of xylazine (~0.4 mg/kg) is often used to supplement other drugs when
inducing general anesthesia in the eland (Citino 1995; Nielsen 1996). We began with a dosage of 0.2
mg/kg i.m., which was increased based on individual response to the highest dose, 0.7 mg/kg.
Xylazine alone was used during the first five procedures. In subsequent procedures (n = 36), xylazine
was combined with a total dose of 25 mg of butorphanol tartrate (Torbugesic®, Fort Dodge Animal
Health, Fort Dodge, IA, USA) to achieve complementary sedative and analgesic effects.
Medications were hand-injected after positioning and slight squeezing of animals in the chute.
Time from injection to onset of sedation (unresponsiveness to hand movements in frontal area, loss
of prehensile ability when treats were provided, salivation, loss of ocular stare) and events
surrounding the sedation were recorded. Once sedated, animals were positioned by squeezing and
lifting with the chute. A bale of hay was placed across the brisket to restrict back and forth movement
of the animal. An epidural block was induced for doing transvaginal ultrasound-guided follicular
aspiration to recover oocytes (Chapter 6).
Heart rate, blood oxygen saturation and body temperature were monitored using a pulse
oximeter (Model 340V, Palco Labs, Inc, Santa Cruz, CA, USA) probe introduced into the nasal
cavity. Respiratory rate was determined by hearing and feeling each breath at the muzzle. After
completion of each oocyte retrieval procedure, blood samples were collected from the tail vein,
prophylactic treatment of 3.6 million U penicillin-G (G.C. Hanford Mfg. Co, Syracuse, NY, USA)
was administered and xylazine sedation was reversed using 600 mg of tolazoline HCl (TolazineTM,
Lloyd Laboratories, Shenandoah, IA, USA).
Blood gas analyses were performed on samples (n = 4) collected from coccygeal vessels
using a 20 g needle and 5-mL heparinized syringe. Analyses were completed within 10 min of
collection using an IRMA blood analysis system (Diametrics Medical, Inc. St. Paul, MN, USA).
Blood glucose (n = 18), hematocrit (n = 14) and creatine phosphokinase, CPK (n = 18) levels were
analyzed by a commercial laboratory (Antec Diagnostics, Memphis, TN, USA).

68
4.2.5. Taming Potential
Certain physical attributes influence the potential to tame/domesticate wild animals (Hemmer
1976; Hemmer 1990; Price 2002). Determination of such features in the eland or for related
antelopes has direct relevance to captive management or future domestication endeavors. Thus, an
attempt was made to determine if specific physical attributes of female eland are associated with
taming potential.
Animals that were well conditioned to sound cues and consistently moved at least to the
entrance of the chute gate (Figure 4.1: Gate No. 16) when invited were categorized as ‘Cooperating’
or having “Good” taming potential. The remaining animals were classified as having “Poor” taming
potential (“Not cooperating”).
Preliminary observations revealed two physical attributes that varied among the female eland.
These characteristics were the pattern of horn growth (generally, either splayed or straight) and the
number of vertical stripes on the sides of the body (ranged from 2 - 13). Thus, the association
between the taming potential and the pattern of horn growth and/or number of body stripes was
evaluated.
4.2.6. Data Analyses
Intervals from production of conditioned stimuli to acquisition of handheld treats in the
morning and afternoon sessions were compared using two-way ANOVA and Duncan’s test. Data on
the interval from injection of either xylazine or xylazine-butorphanol combination to onset of
sedation were compared using an unpaired t-test. Count data were analyzed using the Chi-square test
and/or Fisher’s Exact test. Averages were presented as mean ± SD. Variations in heart rate,
respiratory rate, body temperature and oxygen saturation during sedation were tested using repeated
measures ANOVA after excluding data points with missing values. Hematological values of animals
that underwent oocyte retrieval were also compared using t-test with values obtained in three females

69
opportunistically handled similarly but without doing oocyte retrieval.
All analyses were done using the GraphPad Instat® version 3.00 for Windows 95 (GraphPad
Software, San Diego, CA, USA) except for two-way ANOVA using SAS (SAS Software Release
8.2, Cary, NC, USA). Descriptive statistics were also used to present data. A P-value of �0.05 was
considered statistically significant.
4.3. Results
4.3.1. Conditioning to Sound Cues
All nine females recognized their specific sound cues and accepted handheld treats. Visible
indications of response were increased alertness, attempting to locate food treats and moving toward
the sound cue. However, the extent of acceptance of a handheld treat varied among the females.
Elands 126, 124 and 131 were more cautious and did not take handheld treats consistently. Some
degree of non-specific response to sound cues, which was more marked during training before
feeding was also observed. However, two females often stopped eating their regular diet and came to
the trainer upon hearing their specific sound cue. Dominant animals usually interfered with
conditioning by displacing subordinate females.
Conditioning to sound cues was done over a period of seven weeks. Animals seemed to
associate the sound cues with treats on the first session of training. Intervals from producing
conditioned audio cue to acquiring handheld treats varied from immediate (<0.5 min) to over an hour
(65 min). Mean intervals from activating audio cue to animals accepting handheld treat, during
morning or afternoon sessions, are presented in Table 4.1. The model effect was highly significant
(P <0.0001), but animals had a more marked (P <0.0001) effect than time of training (P = 0.058).
The interaction between animal and time of day was not significant (P = 0.867). The mean (±
SD) interval in the morning and afternoon was 13.6 ± 17.1 and 17.7 ± 19.5 min (overall mean =
15.7), respectively, and ranged from less than 2 min (eland 125) to almost 1 hr (eland 124; Table

70
Table 4.1. Mean interval (min) from conditioned sound cue to acceptance of handheld treats duringmorning and afternoon training sessions in elands_________________________________________________________________________________
Interval Eland ID Morning Afternoon Overall_________________________________________________________________________________
123 3.2 4.2 3.7a
124 53.3 58.3 55.8e
125 1.8 2.2 2.0a
126 23.3 28.7 26.0c
129 2.4 3.1 2.8a
131 35.0 42.7 38.8d
132 8.5 14.0 11.3ab
139 11.7 27.0 19.3bc
140 14.5 18.8 16.6b
_________________________________________________________________________________abcdeValues in the same column with different superscripts are significantly different (P<0.05).
4.1). Although the effect of time of day was not significant, mean intervals were consistently shorter
in the morning sessions, suggesting the fasting overnight was beneficial for training.
4.3.2. Chute Training
All females, including eland 124, which was not conditioned to associate specific sound cue
with treat, came into the barn and within close proximity of the chute during the chute-training
period. Five females voluntarily entered the chute when invited between the 10th and 16th training
sessions over a period of 2 months. The eventual scores of response to chute training varied from 1 -
6 (scale 0 to 6). Four females (eland 123, 125, 129, 140) attained the highest score of 6; one female
(eland 139) was at score 5 and another was at 4 (eland 132). The remaining four elands were more
difficult to train to the chute and were assigned scores of 2 (elands 124, 28, 131) and 1 (eland 126).

71
4.3.3. Use of Sedatives and Physiological Effects of Handling in a Hydraulic Chute
At the doses used, neither haloperidol (1.0 - 1.5 mg/kg) nor acepromazine (0.036 mg/kg)
produced adequate sedation for handling the eland in the chute. Xylazine induced sedation at all dose
ranges evaluated. Intervals from i.m. injection of xylazine or xylazine/butorphanol to the observation
of sedation ranged from 5 to 20 min, and did not differ between the two treatments (P = 0.612; Table
4.2). Excitement immediately before sedative treatment tended to prolong the interval to onset of
sedation.
The frequency of recumbency after administration of sedatives and before positioning them in
the chute was not different between the two treatments (P = 0.362; Table 4.2). A single sedative dose
was adequate to handle females during most of the procedures (34/41, 83%). The frequency of
procedures requiring a second dose to maintain sedation did not differ between the two treatments
(P = 1.00; Table 4.2). After prompting, all recumbent animals returned to standing and were able to
be repositioned to continue with the procedure.
Individual behavior influenced the dose of xylazine required for sedation. Eland 128 required
as much as 250 mg (0.7 mg/kg) of xylazine, while 100 to 150 mg was adequate for the other females.
Table 4.2. Effect of sedative treatments on onset of sedation, frequency of recumbency or a need forsecond dosing of sedative agents in elands during handling and oocyte retrieval in a hydraulic chute_________________________________________________________________________________
Number Dose, Onset of sedation, Recumbent RedosingDrug(s) treated mg/kg mean ± SD, min females, n (%) needed, n (%)_________________________________________________________________________________
Xylazine 5 0.2-0.7 11.0 ± 2.2 3 (60) 1 (25)
Xylazine/Butorphanol* 36 0.2-0.7/0.07 11.9 ± 3.9 13 (36) 6 (17)_________________________________________________________________________________*This protocol plus atropine was used for sedating eland in most oocyte retrieval procedures; initialtrials (n = 10) using haloperidol or acepromazine did not produce adequate sedation.
Some vital physiological parameters observed during sedation and handling for oocyte
retrieval are shown in Table 4.3. Animals subjected to sedation and oocyte retrieval had a

72
significantly greater blood glucose concentration (14.4 ± 3.1 vs. 9.3 ± 2.7 mmol/L; P = 0.0067) than
females opportunistically sedated and handled without oocyte collection. Other parameters, including
CPK activity (1084 ± 972 vs. 354 ± 133 U/L; P = 0.156), hematocrit (36 ± 4 vs. 42 ± 13 %; P =
0.121) and plasma osmotic pressure (290 ± 7 vs. 284 ± 5 mmol/kg; P = 0.298) were not different
between the two groups.
Mean blood glucose levels (mmol/L) per eland during sedation and handling for oocyte
retrieval varied between 10.9 (eland 125) and 19.1 (eland 128). Each female showed glucose
response that was typical for herself (Figure 4.2). For example, glucose levels in eland 125 were
close to 11 mmol/L (range: 10.3 to 11.4) while eland 124 and 128 consistently had values over 16.5
mmol/L (range: 16.7 to 20.6). Moreover, when animals were classified into two cooperative groups
(good or poor), there was a significant difference in blood glucose levels with the higher levels in
elands in which handling was more difficult (Table 4.4). The CPK profile also reflected the pattern of
glucose profile (Figures 4.2; 4.3) in all females except one animal (eland 139).
During the sedation and handling for oocyte retrieval, the mean intranasal temperature, heart
and respiratory rates and oxygen saturation of blood did not differ significantly among the time
points evaluated. However, slight increases in heart rate toward the end of the procedure and
reduction in oxygen saturation during the procedure were observed (Figures 4.4 – 4.7). Recovery
after reversal of sedation was uneventful in all handling procedures and occurred without
complication.
4.3.4. Taming Potential
It was possible to divide the animals into an equal number of cooperating (elands 123, 125,
129, 132, 140) and non-cooperating (elands 124, 126, 128, 131, 139) females. [Note that this
classification does not necessarily relate to the possibility of performing reproductive procedures on a
female or her chute training score; see Discussion].

73
0
5
10
15
20
25
30
35
40
45
#125 #140 #129 #123 #139 #124 #128
Animal ID number (#) in order from most to least cooperative (left to right)
Perc
ent o
r m
mol
/L
Glucose (mmol/L)
Hematocrit (%)
Figure 4.2. Blood glucose (mean ± SD) and hematocrit (mean or single sample values) in sedatedelands handled in a hydraulic chute for oocyte collection (n = 18).

74
0
200
400
600
800
1000
1200
1400
1600
1800
#125 #140 #129 #123 #139 #124 #128
Animal ID number (#) in order from most to least cooperative (left to right)
CPK
U/L
Figure 4.3. Mean blood CPK level in sedated elands handled for oocyte retrieval (n = 18).

75
Table 4.3. Some physiological parameters of female elands during sedation and handling in ahydraulic chute for oocyte retrieval._________________________________________________________________________________Variable* Mean ± SD Range n_________________________________________________________________________________
Blood glucose (mmol/L) 14.4 ± 3.1 9.8 - 20.6 18
Hematocrit 35.8 ± 4.3 28.3 - 42.7 14
Blood CPK (U/L) 1084.3 ± 972 232 - 3766 18
Blood pH 7.44 ± 0.05 7.37 - 7.49 4
Blood osmolality (mmol/kg) 290.0 ± 7.3 278 - 306 16
Heart rate/min 58.8 ± 16 32 - 99 20
Breathing rate/min 30.6 ± 10 12 - 55 18
Rectal temperature 37.6 ± 0.3 37.4 -37.8 2_________________________________________________________________________________*Temperature was measured 15 min after administering sedative agent; other data were collected atthe end of oocyte retrieval.
Table 4.4. Some variations between female elands with good (n = 5) or poor (n = 5) taming potential._________________________________________________________________________________
Taming potential Parameters Good (cooperating) Poor (non-cooperating) P-value_________________________________________________________________________________
Stripes, mean ± SD (range) 6.4 ± 2.3 (4 - 10) 11.2 ± 1.6 (9 - 13) 0.005
Straight-horned (n/group total) 4/5 1/5 0.21
Blood glucose (mmol/L)* 12.0 ± 2.8 16.0 ± 3.6 0.009_________________________________________________________________________________*Collected during handling.

76
Figure 4.4. Heart rate (mean ± SD) profile during xylazine-butorphanol-induced sedation in elandshandled for oocyte retrieval (n = 25).
50.2
47.7
58.0
0
10
20
30
40
50
60
70
80
15 30 45Interval from treatment (min)
Hea
rt r
ate/
min
P = 0.091

77
Figure 4.5. Blood oxygen saturation (mean ± SD) profile during xylazine-butorphanol-inducedsedation in elands handled for oocyte retrieval (n = 23).
90.1
86.4
90.8
0
20
40
60
80
100
120
15 30 45
Interval from treatment (min)
Blo
od o
xyge
n sa
tura
tion
(%)
P = 0.052

78
Figure 4.6. Respiratory rate (mean ± SD) profile during xylazine-butorphanol-induced sedation inelands handled for oocyte retrieval (n = 23).
30.6
27.3
28.8
0
5
10
15
20
25
30
35
40
45
15 30 45
Interval from treatment (min)
Res
pira
tory
rat
e/m
in
P = 0.497

79
Figure 4.7. Peripheral body temperature profile during xylazine-butorphanol induced sedation inelands handled for oocyte retrieval (n = 13).
36.2 36.0
35.2
32
33
34
35
36
37
38
15 30 45Interval from treatment (min)
Intr
anas
al te
mpe
ratu
re (
0C
)
P = 0.232

80
The number of lateral stripes varied from 2 (left side of eland 129) to 13 (right side of eland 124).
The mean of the higher number of stripes on either side of the animal was 8.8 ± 3.2 (range: 4 - 13).
Non-cooperating females had almost twice as many stripes as the cooperative ones. The cooperative
group females had horn growth pattern that was straighter, but the association between this factor and
cooperation by the eland was not significant (Table 4.4). Nevertheless, there was only one female
with splayed-horn appearance that was cooperative and only one female with straight horns that was
not cooperative (Table 4.4).
Blood glucose levels of all elands during handling was significantly lower in cooperating than
in non-cooperating females (Table 4.4). However, there was no correlation between the number of
stripes and mean blood glucose level of all females (r = 0.068; P = 0.851) or for females subjected to
oocyte retrieval (r = 0.3541; P = 0.436). When comparing all 10 females, blood glucose level
(mmol/L) was higher in elands with splayed horns than in females with straight horns (15.9 ± 3.2 vs.
11.5 ± 2.6; P = 0.002). Similarly, among females subjected to oocyte retrieval, glucose levels were
higher in elands with splayed horns than females with straight horns (16.3 ± 3.2 vs. 12.6 ± 1.7; P =
0.008) and in non-cooperating elands than in the cooperating females (16.5 ± 3.6 vs. 13.1 ± 1.9; P =
0.018).
Over time, handling of the eland for oocyte retrieval reduced their readiness to voluntarily
enter the chute. During the early phases of the study, five females readily entered the chute. After
repeated procedures (sedation, handling in the chute and oocyte retrievals), all five animals were
hesitant to enter the chute and required the use of mild enforcements such as water sprayed from a
garden hose and push walls to encourage entry into the chute. However, there was no indication of
reduced response to conditioned audio cues by the females when they were away from the chute area.
4.4. Discussion
The present study demonstrated that individual elands in a group can be conditioned to

81
identify specific sound cues. Subsequently, the conditioned cue was used to move animals from one
location to another, including into the handling chute, thus confirming the potential benefits that
classical and operant conditioning can offer to improve the management and handling of
nondomestic mammals in captivity (Stevens 1978; Fridman 2001; Bloomsmith et al. 2003). We have
continued to effectively use conditioned audio cues to separate individual females from the group for
various purposes.
There were marked variations in the time taken from audio cue to accepting handheld treats
[ranging from < 2 min to > 60 min] and in the response to the chute training. One female (eland 125)
often ran to the trainer in response to cues. This female and another (eland 129) also stopped eating
their regular diet and come to the trainer when prompted. It was also possible to get the attention of
other females by producing specific sounds during feeding. Thus, elands can discriminate between
two or more audio cues. Bongo and nyala antelopes have also been reported to recognize and become
agitated by the voice of the veterinarian who administers medications by darting (Grandin 2000). The
individual variation in the responses of eland to conditioning could be due to previous experiences
with people and/or may have a genetic basis.
Animals maintained excellent memory of conditioned stimuli even after reinforcement was
interrupted for several months. Maintenance of memory of conditioned stimuli after interrupting
reinforcement for a long time has been reported in other mammalian species. For example, an
elephant performed conditioned tasks after 8 yr without reinforcement (Stevens 1978). Bongo and
nyala antelopes also responded in a similar manner as in preceding sessions during monthly
reinforcements with conditioned stimuli (Grandin 2000), indicating animals maintained good
memory of the conditioning. Such memory in ungulates has implications for handling since animals
can remember, for a prolonged period, the manner in which they were handled.
Interference by dominant animals during training of subordinates has been observed in other
species, including bongo, but social rank did not interfere with training the nyala. In the nyala,

82
handlers could safely enter the room in which animals were kept to facilitate conditioning; however,
it was not safe to enter the bongo pen (Grandin et al. 1995; Phillips et al. 1998). Eland in the present
study are similar to the bongo in this respect because two females (elands 128 and 139) made active
attempts to charge people. However, during training, two persons could easily keep animals in
different corners of the holding area so that a single female could be trained in a separate area,
without interference by dominant females.
Training of the five eland to enter the chute required 10 to 16 training sessions (days) over a
period of 2 months. Bongo and nyala have been trained to enter a handling crate in 21 days (Phillips
et al. 1998). The aggregation of multiple females at the area of the gate of the chute, when training a
single female to the chute, was a problem because subordinates had to retreat when dominant females
approached the area. Also, the elevated entrance into the chute delayed voluntary entry. Thus,
considering the active pursuit of treats (e.g., more so than members of our bongo antelope) by eland,
a shorter interval of training them to enter the chute can be expected. However, they are more prone
to flight and more active than bongos and may not readily accept desensitization for handlings.
Initially, excessive physical movement by the three most cooperative females during restraint
in the Tamer chute precluded performing clinical exams and other procedures on non-sedated
animals. Oral treatment with 1 mg/kg of haloperidol has been used to sedate female elands for
manual handling (Boyd et al. 2000). Intramuscular or i.v. administration of 20 to 40 mg of
haloperidol tranquilized eland for 8 to 18 hr (Citino 1995). The reason for the failure of haloperidol
treatment to successfully sedate eland in the present study is unclear. In the holding area, only one of
the females treated with haloperidol displayed mild sedation (associated with signs of dizziness),
which was not apparent after she was moved into the chute. Individual animal variation, or possibly
the noise of the electric motor of the hydraulic pump may have affected the sedative effects of
haloperidol. Unpredictable effects of haloperidol have also been reported in the eland (Ebedes and
Raath 1995).

83
The eland is more tolerant to xylazine (up to 0.7 mg/kg in the present study) as compared
with domestic ruminants (≤0.3 mg/kg, www.merckvetmanual.com/mvm/index.jsp, accessed
December 31, 2003). Although we used butorphanol tartrate with xylazine to improve
analgesia/sedation, the onset of satisfactory sedation was not different, nor were there other
noticeable differences in response when using xylazine alone or in combination with butorphanol
tartrate. Thus, the actual benefit of butorphanol at the dosage used remains to be determined. The
duration of sedation (~1 hr) allowed sufficient time to perform ultrasound-guided oocyte retrieval
and other procedures.
Xylazine suppresses heart and respiratory rates in domestic ruminants (Mogoa et al. 2000;
Ndeereh et al. 2001). In the present study, heart rates and respiratory rates recorded at three intervals
during sedation were not significantly different (Figures 4.4; 4.6). Whether these differences are
attributable to species differences or the method of data collection remains to be determined. Blood
oxygen saturation declined early during sedation (e.g., up to 63% in one procedure) but oxygen
supplementation allowed a return to normal. Thus, cardiopulmonary parameters during sedation and
handling of the eland were within manageable ranges.
Body temperature, heart and respiratory rates, hematocrit, blood glucose, CPK and cortisol
levels are often used to assess acute stress during animal transport or handling (Knowles and Warris
2000). Establishment of baseline values for most of these variables in the eland is difficult because
data/sample collection requires either immobilization or vigorous physical restraint, both of which
alter normal parameters. Remote assessment of heart rate and respiratory rates on alert (non-
restrained) animals may be possible; however, we could count jugular pulse only on two of the
elands in the present study (mean rates were 35 and 51 for each female) and we could not remotely
count breathing movements. The good body condition of the animals may have reduced the remote
visibility of jugular pulse and breathing movements.

84
Thus, it is difficult to compare the present cardiopulmonary parameters and body temperature
with baseline values. Nevertheless, body temperatures ranging from 39.4 - 42.9 (mean = 41.0) have
been reported in immobilized females (Denney 1970). In the present study, mean intranasal
temperature varied by less than 2ºC. During two procedures, simultaneous recordings of rectal
temperature (37.4 to 37.8ºC) in the eland were as much as 2.7ºC higher than the intranasal
temperature.
The mean blood glucose level (14.4 mmol/L, 260 mg/dl) of elands during oocyte retrieval
was higher than in females handled similarly by us but without oocyte retrieval (9.3 mmol/L). It was
also higher than the mean value of multi-institute data (10.4 mmol/L) gathered by ISIS/MedARKS
for adult female elands in North America. Similarly, eland immobilized by darting from a helicopter
or those manually restrained after chasing into nets in South Africa (Ganhao et al. 1988) had lower
blood glucose levels (7.9 and 8.7 mmol/L, respectively) than values found in the present study.
Bongo antelope subjected to traditional immobilization methods had a blood glucose level of 9.3
mmol/L, which was 2.7 times higher than bongos conditioned to handling (Phillips et al. 1998).
In domestic cattle, the normal blood glucose level is <4.5 mmol/L and levels similar to those
recorded in the present study are observed only in animals subjected to i.v. glucose tolerance tests or
animals receiving glucocorticoid therapy (Abdel-Fattah et al. 1999; Panicke et al. 2002). The
numerically higher CPK activity in elands handled for oocyte retrieval as compared with those
handled without oocyte retrieval is also reflective of greater muscle exertion in the former group.
These results indicate that elands have a large pool of glucose and/or glucose precursors and
possess the ability to mobilize them during physical exertion. However, since high blood glucose
level is an indicator of stress during handling, and we noted a tendency for a higher glucose level in
eland that were more difficult to handle, alternative methods of minimizing the elevated glucose
response should be investigated. The higher-than-physiological blood glucose level observed during
oocyte collection may also lead to an increased follicular glucose level, due to increased

85
vascularization and permeability of preovulatory follicles (Larsen et al. 1996; Barboni et al. 2000).
Moreover, bleeding caused by aspiration of follicles occurs during ultrasound-guided oocyte retrieval
and may expose oocytes to supra-physiological glucose levels that could possibly affect the
developmental potential of the oocytes.
Initially, five females readily entered the hydraulic chute, but sedation followed by oocyte
retrieval negatively affected the readiness of voluntarily entry into the chute during subsequent
procedures. Whether a more intense habituation and desensitization or strict adherence to principles
of operant conditioning during training could prevent or minimize this negative effect remains to be
determined. Nevertheless, the response of elands to sound cues away from the chute area did not
show any sign of subsidence. Thus, the conditioned sound cue could be used throughout the study to
relocate animals outside the barn area and to direct them toward the chute.
More recently, after separating individual animals from the group, a combination of sprayed
water from a garden hose and push walls have been used to move animals into the chute. Guidance of
animals toward the chute was facilitated by additional modifications made to the holding area,
including the addition of two sliding doors in Plot B and the construction of an alley between Gates
No. 8 and No. 13. The first modification enabled us to separate an animal on one side of the feeding
area for subsequent movement through the alley into the barn. A push wall was then used to
encourage entry of non-compliant females into the chute. Thus, it was possible to consistently guide
animals into the chute and handle them, and oocyte retrievals could be done not only on the animals
trained to enter the chute but also on two additional untrained females.
The fact that eland 128 could be used for ultrasound-guided oocyte retrieval without
conditioning to audio cue and without training to voluntarily enter the chute underscores the
importance of knowing the “personality” profile of each eland. Because of her voracious appetite,
she was among the first to come to the feeding yard and hence it was easy to separate her for
providing estrous synchronization treatments. Eland 124 could also be treated similarly but isolating

86
her often required more time. Because of the tendency of elands 128 and 139 to charge people,
increased caution was required when around them. Elands 126 and 131 were excluded from oocyte
retrievals because of their excessive flighty behavior and since it was not possible to consistently
provide hormone treatments. Eland 132 was not used for oocyte retrieval because of her geriatric
state and poor body condition. Nevertheless, the previously described modifications to the holding
area enabled handling of all 10 animals in the chute at least once.
“Genetic and experiential” factors determine the degree of tameness of an animal (Price
2002). Behavioral modifications leading to tameness and domestication are associated with certain
anatomical and biochemical changes. For example, domestic counterparts of some wild species (e.g.,
domestic cats vs. wildcats or dogs vs. wolves) have evolved to have a smaller (up to 30%) brain.
Agouti rats and deer mice are tamer (easier to handle) than those with non-agouti phenotypes
(Hemmer 1990; Price 2002). Selection and subsequent breeding of animals with certain phenotypes,
is therefore, a viable approach to establish a tame or even domesticated line of wild animals (Cottle
and Price 1987; Hemmer 1990; Prasolova and Jing 1999; Trut et al. 2000).
Observations in the present study suggest that females with fewer vertical stripes, and
possibly, straighter horns may be good candidates for taming. Similarly, among another group of
eight female elands at our facility that were not included in the present study, an approach by
strangers induced shorter flight distance in females with fewer stripes and straighter horn growth
pattern (personal observation). The characteristics described in the present study may serve as a
starting point for characterizing additional phenotypic and genetic factors influencing eland
tameness. We have also demonstrated that social rank did not affect taming potential (Wirtu et al., In
Press).
Stripes are the main criteria for differentiating Northern (East African) and Southern
(Zambezi and Cape) strains of eland, with the Southern strains having fewer stripes. Thus, the
variation in stripe number among adult female eland in the present study may have a genetic basis.

87
Horns are understood as instruments for defense against predators, for establishing dominance
hierarchy and for breaking tree branches by tragelaphine antelopes (Kingdon 1982). They may also
have thermoregulatory functions (Picard et al. 1999). The behavioral variation among individuals
with different patterns of skin color may be explained by the close association of some genes and
pathways influencing both skin color and certain behavior of animals (Hemmer 1990; Price 2002). It
is unknown if horn growth pattern is also affected similarly.
Data are not readily available on whether the Northern and Southern strains of eland vary in
their horn growth pattern or whether they differ in their tameability. Interestingly, a successful eland
domestication endeavor at the Askanya-Nova Zoo in Ukraine was achieved with the southern (Cape
and Zambezi) strains (Treus and Lobanov 1971). On the other hand, a well-funded project designed
by international team members to domesticate eland in East Africa (Kenya) led to the conclusion that
eland were not a good candidate for domestication (Spinage 1986). An obvious question is whether
the strain of eland used in the Kenya study influenced the conclusions reached by the team.
In conclusion, a combination of behavioral conditioning/training and restraint of sedated
eland in a hydraulic chute was a reliable and repeatable method for performing minimally invasive
assisted reproductive techniques. Moreover, targeted audio conditioning of individual elands among
a group of females can be used to facilitate the administration of medications to specific animals and
can be helpful in the management of captive eland, such as fast translocation or separation of specific
animals from a herd without using expensive alley systems. During sedation and handling for oocyte
retrieval, several physiological parameters were either normal or within clinically manageable range.
However, blood glucose levels were elevated and may be used as a marker in reducing and/or
evaluating handling-induced physical exertion or stress in elands.

88
Chapter 5. Transvaginal Ultrasound-guided Oocyte Collection, In VitroOocyte Maturation and Embryo Production in the Eland Antelope
5.1. Introduction
There is a growing interest in using reproductive technologies, such as artificial insemination,
embryo transfer, and the more recently developed methods of in vitro embryo production in the
captive breeding of rare and endangered species of mammals. Moreover, many investigators believe
that these technologies can facilitate genetic transfer between captive and free-ranging populations of
nondomestic species (Lasley et al. 1994; Loskutoff et al. 1995; Wildt et al. 1997; Holt and Pickard
1999; Lanza et al. 2000b; Pope 2000; Berg et al. 2002; Ptak et al. 2002). Compared with natural
mating or the first generation of ART (i.e., AI and ET), in vitro embryo production, combined with
cryopreservation and ET, offers more flexibility in sire-dam selection and, theoretically, would
maximize the number of offspring that can be produced per male or female gamete donor.
Techniques for in vitro production of domestic ungulate (cattle, sheep, goats, pigs) embryos
have advanced greatly during the last three decades, such that viable embryos can now be produced
consistently and repeatably. The ease with which gametes or embryos can be collected in vivo and
the availability of reproductive organs from abattoirs have contributed immensely to this progress.
Direct extrapolation of techniques developed in domestic species (e.g., cryopreservation of
spermatozoa or embryos, IVF, embryo culture) to nondomestic ungulates has mostly resulted in low
success as evaluated by production of offspring (Hammer et al. 2001; Berg and Asher 2003),
suggesting the need for development of species-specific methods. Application of ART to large
nondomestic ungulates using captive animals can be facilitated by developing minimally stressful
handling methods for collecting biological samples (blood, feces, urine, gametes, embryos etc),
administering treatments and monitoring responses.
Applying traditional approaches of handling (e.g., frequent darting and complete chemical
immobilization) to ART procedures is stressful to large nondomestic ungulates. For example,

89
subjecting four species of antelopes (oryx, bongo, eland and kudu) to ART procedures by
confinement, daily treatment with hormones and multiple administration of anesthetics led to weight
losses of up to 0.4 kg/day (Schiewe et al. 1988) and a probable reduction in superovulatory response
and fertilization rate (Schiewe et al. 1988; Schiewe et al. 1991b). The multiple hormone treatments
and associated handling for estrous synchronization and ovarian stimulation are a major cause of
stress in nondomestic ungulates [e.g., the bison (Dorn 1995)] and among the factors that need
improvement for increasing the efficiency of commercial cattle embryo transfer (Hasler 2003).
The limitations of traditional handling methods for ART in wild ungulates has been
emphasized by Solti et al. as follows: “...there have been more losses of nondomesticated animals in
attempts to collect embryos than there have been offspring produced by the transfer of in vivo-
derived embryos…” (Solti et al. 2000). Thus, besides the possibility of using behavioral training and
modern handling devices to reduce the need for general anesthesia (Chapter 4), there is also a need to
develop approaches to reduce the number of hormone injections required for ovarian stimulation.
Ultrasound-guided oocyte collection offers a safe, relatively non-invasive, repeatable (twice
per week in domestic cattle) approach for recovering oocytes from large ungulates. In nondomestic
ungulates, the technique, in conjunction with in vitro embryo production and subsequent transfer of
the embryos to recipient females, has resulted in guar (Armstrong et al. 1995; Hammer et al. 2001)
and wapiti (Berg and Asher 2003) offspring. In vitro embryo development after IVF of oocytes
recovered by ultrasound-guided methods were also reported in several large nondomestic species
including the bongo (Pope et al. 1998b), addax (Asa et al. 1998), Burchell’s and Hartmann’s zebra
(Meintjes et al. 1997), red hartebeest, tsessebe, sable and African buffalo (Loskutoff et al. 1995).
The eland antelope is a useful model species for the study and application of ART in rare
tragelaphine antelopes. It is a large bovid and hence amenable to non-surgical, transrectal and/or
transcervical accessibility and manipulation of the female reproductive tract (Chapter 2). In vitro
embryo production has been reported in at least 13 species of bovidae (Hall-Woods 2001) and in

90
vitro-derived embryos have resulted in offspring after transfer to recipient females in eight species of
bovidae (Table 2.1). Recently, we reported the initial demonstration of ultrasound-guided oocyte
retrieval and some embryonic development in vitro in the eland (Wirtu et al. 2002a; Damiani et al.
2003). Thus, the hypotheses of the present study were:
1. transvaginal ultrasound-guided follicular aspiration can be used to collect oocytes in
sedated eland antelope; and
2. eland oocytes recovered by an ultrasound-guided approach are capable of in vitro
maturation and subsequent embryonic development after in vitro fertilization,
intracytoplasmic sperm injection or somatic cell nuclear transfer.
Specific objective were to:
1. determine ovarian response and oocyte recovery rates following treatment with pituitary-
derived FSH dissolved in a polyvinyl pyrrolidone carrier given four and/or two days
before oocyte retrieval,
2. compare effects of targeting preovulatory or postovulatory follicular waves on ovarian
response and oocyte recovery after incorporating an oral progestogen (altrenogest) in the
estrous synchronization and ovarian stimulation protocol; and
3. evaluate the in vitro developmental competence of eland oocytes after in vitro
maturation, and subsequent in vitro fertilization, intracytoplasmic sperm injection or
somatic cell nuclear transfer.
5.2. Materials and Methods
5.2.1. Study Animals
Seven adult eland females among a group of 10 animals were used for oocyte collection.
Animals lived in a 45 m x 56 m open yard fenced with woven wire. The holding area was equipped
with a hydraulic handling chute (TamerTM, Fauna Research, Inc., Red Hook, NY; USA) and gates

91
with remotely operable doors (see Chapters 4 for detailed descriptions of the holding area, herd
management and history). The seven females were selected based on the ability to consistently
administer estrous synchronization and ovarian stimulation treatments and move them to the
handling chute using conditioned audio cues and/or the gate and push-wall systems (Chapter 4).
5.2.2. Estrous Synchronization and Ovarian Stimulation
5.2.2.1. Experiment 5.1 [Single (day –2) vs. Double (day –4 and day –2) FSH Injections]
Before conducting oocyte retrieval procedures, the elands were treated to synchronize their
estrous cycle and to induce ovarian stimulation. Figure 5.1 shows the treatment schedule.
Starting at unknown stages of the estrous cycle, three consecutive doses of prostaglandin F2α,
(PGF2α, 25 mg of dinoprost tromethamine, Lutalyse, Pharmacia and Upjohn Company, Kalamazoo,
MI, USA), were administered i.m. The first (day 0), second (day 11) and third (day 22) injections
were given 11 days apart, with the third injection given two days (~48 hr) before the expected day of
oocyte retrieval. Thus, assuming ovulation occurs between 2 and 3 days after treatment with
prostaglandin, females were at days 8 to 9 of the estrous cycle during the last two prostaglandin
treatments. All parenteral treatments were hand-injected while females were in the hydraulic chute.
For ovarian stimulation, a porcine pituitary folltropin extract (Folltropin®-V, 400 mg NIH-
FSH-P1 per vial, Bioniche Animal Health Canada, Inc., Belleville, Ontario, Canada) was used. A
40% solution of polyvinyl pyrrolidone (PVP, mol. wt = 40,000) was prepared in ultrapure
(NANOpure) water. Aliquots of the PVP solution were sterilized by autoclaving in glass vials, and
were stored at 4ºC. On the day of use, the powdered contents of a vial of Folltropin®-V was dissolved
in 1.25 mL of saline and aspirated into a 3-mL syringe. The Folltropin®-V vial was then rinsed with
3.75 mL of 40% PVP solution, aspirated into a 5-mL syringe and then mixed with the 1.25 mL
solution of Folltropin®-V.

92
Figure 5.1. Estrous synchronization and ovarian stimulation treatment schedule before ultrasound-guided oocyte collection in elands in experiment 5.1.
Day 0 Day 11 Day 22 Day 24
Prostaglandin
FSHSingle400 mg
Day 20
Double133 mg267 mg
Oocyteretrieval
Treatment type Treatment day Treatmentgroup

93
After thorough mixing of the solution, animals received either a single (5.0 mL) injection or
two injections of two-thirds (3.3 mL) and one-third (1.7 mL) of the total volume, depending on the
experimental design (see below). The remaining quantity of re-constituted FSH was stored at –20ºC.
Thus, the final concentration of PVP used as a gonadotropin carrier was 30%.
In Treatment 1 (single injection), animals were treated i.m. with 400 mg of FSH (Folltropin®-
V) on day 22 of treatment (day 0 = first prostaglandin injection), 2 days before oocyte retrieval
(Figure 5.1). In Treatment 2 (double injections), 267 mg (two-thirds) and 133 mg (one-third) of FSH
were administered on days 20 and 22 of treatment 4 and 2 days before oocyte retrieval, respectively
(Figure 5.1). In both treatments (n = 13), prostaglandin administration was similar.
5.2.2.2. Experiment 5.2a (Targeting Preovulatory or Postovulatory Follicular Wave)
In this experiment, the effects of an oral progestogen (altrenogest) combined with a single
injection of prostaglandin F2α on estrous synchronization and subsequent ovarian response to two
Folltropin®-V treatments were evaluated. The two follicular waves (preovulatory and postovulatory)
that are universally found in ruminants studied to date (Adams 1999) were targeted. In Treatment 1,
the follicular wave that leads to ovulation (preovulatory) immediately following ovarian stimulation
and prostaglandin treatments was the target. In Treatment 2, the expected ovulation following estrous
synchronization using altrenogest and prostaglandin treatments was bypassed and ovarian stimulation
treatment was initiated one day after the expected day of ovulation. Thus, the immediate
postovulatory follicular wave was the target in Treatment 2.
Figure 5.2 shows the treatment schedules targeting each follicular wave. In Treatment 1
(preovulatory), females were fed 6 mL of 2.2% altrenogest (Regu-mate®, DPT Laboratories, San
Antonio, TX, USA) embedded in banana for 7 days (day 0 = day of first treatment). Animals
received 267 mg and 133 mg of FSH in 30% PVP on days 4 and 6, respectively. Prostaglandin was
also given on day 6 and ultrasound-guided oocyte retrieval was performed on day 8.

94
Figure 5.2. Treatment schedules to target preovulatory or postovulatory follicular waves in elandsbefore conducting ultrasound-guided oocyte retrieval in Experiment 5.2a.
FSH
Day8
133 mg267 mg
Oocyteretrieval
Prostaglandin
Altrenogest (day 0 - 6)
Day 10
133 mg267 mg
Day 12
Day 14
Altrenogest (day 0 - 6)
Preovulatory Postovulatory
Oocyteretrieval

95
In Treatment 2 (postovulatory), altrenogest and prostaglandin were given as in Treatment 1.
Day 9 was the expected day of ovulation and emergence of a new postovulatory follicular wave.
Thus, 267 mg and 133 mg of FSH were given on days 10 and 12, respectively, and oocytes were
recovered on day 14 (Figure 5.2). Eight procedures were conducted and compared in Experiment
5.2a. Subsequently, an additional 16 procedures were conducted using Treatment 1 (preovulatory) for
estrous synchronization and ovarian stimulation.
5.2.2.3. Experiment 5.2b (Individual and Seasonal Variation in Response to OvarianStimulation)
Experiments 5.1 and 5.2a produced results that were suggestive of individual variation among
the elands to ovarian stimulation treatments. To further explore this possibility, data from
Experiments 5.1 and 5.2a were combined with data from 16 additional ovarian stimulation/oocyte
retrieval procedures done as in Treatment 1 (preovulatory) of Experiment 5.2a. Data were combined
because there were no treatment effects on the number of follicles and oocytes recovered. These data
were used to test the effect of animals or months (season) on ovarian response to gonadotropin
treatment (follicle development) and oocyte production.
5.2.3. Ultrasound-guided Follicular Aspiration for Oocyte Retrieval and Evaluation ofOocytes
5.2.3.1. Custom-made Adaptor for Ultrasound Transducer
The first two attempts to do oocyte retrievals were not successful because the commercially
made adaptor of a 5-MHz, curvilinear ultrasound transducer (model UST-9111-5, Aloka Co. Ltd,
Tokyo, Japan) could not be passed through the vestibule of the two eland. Also, the adaptor for a
human transducer was not long enough to be able to scan the ovaries of either of the two females.
Thus, an in-house custom-molded plastic adaptor (see Figure 5.3) was constructed for
mounting the transducer. The design was based on the original adaptor but was made with smaller

96
Figure 5.3. Ultrasound transducer and a needle guide enclosed in the custom-made adaptor (A);B: Close-up views of the transducer tip with the custom-made (left) and original (right) adaptors.Note the ~1 cm reduction in the vertical dimension of the tip of the device in the custom-madeadaptor as compared with the original adaptor.
A
B

97
dimensions. After encasing the transducer and the needle guide in the modified adaptor, the device
was wrapped in self-fusing elastic tape (Tommy Tape, Manco, Inc Avon, OH, USA). Thus, an Aloka
SSD-500V real time ultrasound scanner (Corometrics Medical Systems, Inc., Wallingford, CT, USA)
and the 5 MHz transducer in a custom-molded plastic adaptor were used for ultrasound scanning of
ovaries and follicular aspiration to collect oocytes.
5.2.3.2. Oocyte Collection and Evaluation
For oocyte retrieval, females entered a hydraulic chute (Chapter 4) and were positioned by
adjusting the sides to prevent animals from turning around. Sedation was induced using 100 to 250
mg of xylazine with or without butorphanol tartrate. After sedation, females were further positioned
and lifted using the hydraulic chute (Chapter 4). The sacrococcygeal area was prepared for
administering epidural analgesia by clipping the hair and applying disinfectants. For the epidural
block, 4 mL of 2% lidocaine HCl (Biomeda Inc, Riverside, MO, USA) was injected into the epidural
space at the sacrococcygeal space or between the first and second coccygeal vertebrae. Paresis of the
tail was used as an indication of a successful epidural block.
After manually cleaning the rectum and the perineal region of the animal, the vagina was
rinsed with 30 mL of saline solution containing 1% lidocaine. Ultrasound transmission gel (Universal
Medical Systems, Inc., Bedford Hills, New York, USA) was applied to the tip of the transducer. The
latter, attached to the ultrasound unit, was lubricated (Lubogel-VTM, Agtech, Inc, Manhattan, KS,
USA) and covered with a condom before introducing into the vagina.
During rectal palpation, the dimensions of ovaries were estimated by using pre-calibrated
distances on the index and thumb fingers of the technician performing the palpation. The number and
size of ovarian follicles were recorded during ultrasound scanning.
An 18-g, 64-cm needle connected to a 50-mL centrifuge tube with a 65-cm long Teflon
tubing was used for follicular aspiration, The needle and tubing were rinsed with aspiration medium
(see later) and introduced through the needle guide attached to the transducer. All ovarian follicles

98
(�3 mm) visible on the monitor of the ultrasound unit were punctured and aspirated using a negative
vacuum pressure of 70 mm Hg, generated using a regulated pump (Model V-MAR-5000B, Cook
Veterinary Products, Eight Mile Plains, Australia).
Follicular contents from each ovary were aspirated into separate tubes and held at ~39ºC. The
aspiration medium was TL-HEPES solution (04-616F, BioWhittaker Co, Walkersville, MD, USA)
supplemented with 50 µg/mL of gentamicin and 10 U/mL of heparin. After each oocyte retrieval
procedure, blood samples were collected, prophylactic penicillin-G was administered and sedation
was reversed using tolazoline (Chapter 4). Initially, the transducer and needle guide were dismantled
from the adaptor for cleaning; however, removing several centimeters of tape from the scanning
portion of the transducer was found to provide adequate exposure of the device for cleaning.
After transporting aspirated fluid to the laboratory, oocytes were located using a
stereomicroscope and graded as previously described (de Loos et al. 1989), but with slight
modifications. The four quality grades were; A: multilayered, uniformly translucent cumulus
investment and homogenous ooplasm, B: multilayered, less translucent cumulus investment and
mostly homogenous ooplasm with some dark cortical areas, C: darker cumulus investment and
irregular ooplasm, and D: dark and irregular cumulus investment and nonhomogenous ooplasm.
Also, the degree of cumulus expansion was graded on a scale of 0 to 3: 0 (compact), 1 (slightly
expanded), 2 (moderately expanded) and 3 (fully expanded)(Hinrichs and Williams 1997); however,
expansion grades of 0 and 1 were categorized as “Compact” and 2 and 3 as “Expanded” for data
analysis. Diameter of oocytes was measured on an Olympus inverted microscope equipped with a
pre-calibrated eyepiece micrometer.

99
5.2.4. Oocyte Maturation and Embryonic Development in Vitro
5.2.4.1. Experiment 5.3 (In Vitro Maturation of Oocytes)
Initially (first portion of the study), IVM was done in medium 199 (TCM199, Gibco, Grand
Island, NY, USA) supplemented with 10% heat inactivated bovine calf serum (BCS, Hyclone
Laboratories, Logan, UT, USA), FSH, LH, estradiol, pyruvate and EGF (Krisher et al. 1999). Later
(second portion of the study), the IVM medium was modified because of unsatisfactory maturation
rates when using the previous protocol.
The modifications included use of TCM 199 (cat. # 9102, Irvine Scientific, Santa Ana, CA,
USA), 1 mM L-glutamine, 1.2 mM L of cysteine and 5 µg/mL of insulin with LH, estradiol, pyruvate
and EGF, as described above, but without FSH. In addition, either heat inactivated female eland
serum collected during the oocyte retrieval procedures, or BCS were used. The modified IVM
medium was used for oocytes obtained after incorporating altrenogest into the estrous
synchronization protocol.
For IVM, ~10 oocytes were put into each 50 µL drop of medium under mineral oil in a 60 x
15 mm Falcon® polystyrene petri dish. Incubation was conducted at 38.5ºC in an atmosphere of 5%
CO2 in humidified air.
In most experiments (n = 34 procedures), oocytes underwent IVM for ~24 hr before IVF,
ICSI or SCNT (intraspecies, or interspecies using giant eland somatic cells). However, during four
procedures, all or a sub-set of oocytes (n = 22) with fully expanded cumulus cell layers were selected
and used for in vitro embryo production on the day of oocyte collection (~ 6 hr post collection) after
visually determining their nuclear maturation status.
5.2.4.2. Experiment 5.4 (In Vitro Embryo Production and Embryo Culture)
In vitro fertilization was done in Tyrode’s-albumin-pyruvate (TALP) medium supplemented
with heparin and PHE (Chapter 3) using frozen-thawed eland semen. The semen samples were

100
collected from two eland bulls using either electroejaculation or rectal massage. The sample collected
by electroejaculation was cooled to 40C in TEST yolk buffer. Four to 6% glycerol in TEST yolk
buffer (Irvine Scientific) was added and the sample was loaded in 0.25 ml straws. Samples were then
frozen on dry ice and then plunged into liquid nitrogen for storage.
Samples collected by rectal massage were extended in Tris-citrate based extender (Bilady®,
Minitüb, Tiefenbach, Germany) containing 20% egg yolk. After cooling to 40C (3 to 12 hr), the same
extender containing 14% glycerol (final concentration =7%) was sequentially (10%, 20%, 30% and
40%) added at 10 min intervals and samples were loaded in 0.5 ml straws. Freezing was done by
placing straws 8-cm above the surface of liquid nitrogen for 20 min and then plunging the straws in
liquid nitrogen for storage.
The frozen-thawed eland spermatozoa were generally characterized by low motility (<40%)
and reduced longevity of progressive motility during incubation at 38.5°C. Accordingly, the IVF
procedures were modified by 1) supplementing the IVF microdrop with spermatozoa that were
maintained at room temperature for 3 to 4 hr after initial IVF; 2) centrifugation (400 x g/15 min) for
concentrating/cleaning semen samples, with a less frequent use of gradient centrifugation columns
for sperm separation.
Moreover, during the latter part of the study, heparin was excluded due to preliminary
observations suggesting a negative effect on the longevity of progressive motility of eland
spermatozoa during IVF. Attempts were also made to increase the percentage of motile spermatozoa
in IVF microdrops by depositing concentrated spermatozoa in two to three peripheral microdrops
connected to the oocyte-containing drop via channels of medium under oil.
Nuclear transfer was carried out using cumulus cells of the oocyte donor or frozen-thawed
epithelial cells of common eland bull (collected during rectal massage) or frozen-thawed skin
fibroblast cells of a giant eland bull. Enucleation of the oocyte chromatin and injection of somatic

101
cells was carried out as described previously (Lanza et al. 2000a). Micromanipulation was done
using a Nikon Eclipse (TE2000-S, Nikon Corp., Japan) inverted microscope with a Nikon UV light
source (C-SHG1, Nikon Corp., Japan). Couplets were fused electrically (Lee et al. 2003) and
chemically activated using 5 or 10 µM ionomycin and 2 mM 6-dimethylaminopurine (Lanza et al.
2000a; Lee et al. 2003).
Micromanipulation for intracytoplasmic sperm injection (ICSI) was carried out using the
Nikon Eclipse inverted microscope described above for nuclear transfer. Frozen-thawed common
eland spermatozoa were subjected to swim-up in a 15-mL centrifuge tube and a 5 to 10 µL aliquot
from the top layer was placed in a 100-µL microdrop of 10% PVP. A single spermatozoon was
immobilized using the injection pipette and injected directly into the ooplasm. After sperm injection,
oocytes were activated using ionomycin and 6-DMAP as described above for nuclear transfer.
Embryo culture after IVF, ICSI or SCNT was carried out at 38.5ºC in a humidified
atmosphere of 5%CO2, 5% O2 and 90% N2. Initially, HECM-6 was used for the first 2 to 3 days with
further IVC in TCM-199 supplemented with pyruvate and 10% BCS (Krisher and Bavister 1999).
However, due to poor embryonic development in the eland, different media were used during the
course of the study with the goal of improving in vitro development. Additional media used for IVC
included KSOM-20aa or BM-3-20aa-glucose supplemented with either bovine or eland serum
(Chapter 3), modified Tyrode’s solution (Gomez et al. 2000), CR1aa, deer SOF (Berg and Asher
2003) and alpha minimum essential medium (Gibco, Grand Island, NY, USA) supplemented with
5% BCS.
5.2.4.3. Evaluation of Oocyte Maturation and Embryonic Development
Determination of in vitro maturation was made using Hoechst staining (before nuclear
transfer) or by microscopic evaluation of the first polar body extrusion (during ICSI). Meiotic status
of oocytes subjected IVF was retrospectively determined based on cleavage and/or by staining �48 hr

102
post insemination. Oocytes were stained using Hoechst 33342 (Lee et al. 2003) or Giemsa after a
two-step fixation (Pope et al. 1993).
The nuclear maturation status of oocytes subjected to nuclear transfer was determined during
the routine Hoechst staining done to identify the recipient oocyte chromatin. Determination of
embryonic development after nuclear transfer was based on morphological evaluation for cleavage
and on Hoechst staining. Staining involved fixing couplets or embryos in a paraformaldehyde-Triton
X-100 solution, washing in PBS-BSA and mounting them in Hoechst-glycerol solution, as described
previously (Lee et al. 2003).
Chromatin structure was examined and photographed using bright (Giemsa stain) or UV
(Hoechst stain) light on an Olympus BX60F-3 (Olympus Optical Co. Ltd, Japan) connected to a
Sony color video printer (Model UP-1800MD, Sony Corporation, Japan). An Olympus UV light
source (Model BH2-RFL-T3) was used during florescent microscopy. Images were also taken using
an Olympus digital camera (model DP11-N) attached to an inverted Olympus microscope (model
IX70-S8F2).
The chromatin structure of oocytes was classified as germinal vesicle (GV), metaphase I or
metaphase II. When the chromatin structure was difficult to determine, it was classified as “Not
determined”. Embryonic development was evaluated both morphologically and by counting the
number of nuclei after staining by one of the two methods described above.
5.2.5. Data Analyses
Effects of Folltropin®-V treatment (single vs. double injection) on ovary size were tested
using the Mann-Whitney test while effects on the total number of follicles or oocytes were compared
using the t-test. Treatment effects on the distribution of follicles by size, the morphological
appearance of oocytes at recovery and the meiotic status of oocytes after IVM were tested using Chi-
square analysis. The effects of animal and month on ovarian response and number of oocytes

103
recovered were compared using one-way ANOVA and the Kruskal-Wallis test (nonparametric
ANOVA), respectively. Simple regression analysis was used to test the correlation between number
of follicles observed and oocytes recovered. Analyses were done using the GraphPad Instat® version
3.00 for Windows 95 (GraphPad Software, San Diego, CA, USA). A P-value of �0.05 was
considered statistically significant. Descriptive statistics were also used to present data.
5.3. Results
5.3.1. Experiment 5.1 [Single (day –2) vs. Double (day –4 and day –2) FSH Injections]
Ovaries were ovoid to spherical in shape. The size (= longest diameter) of the left and right
ovaries were not different after ovarian stimulation when compared within single (1.7 ± 1.2 cm vs.
1.8 ± 0.8 cm; P = 0.8892) or double (2.7 ± 1.0 cm vs. 3.4 ± 1.0 cm; P = 0.2576) Folltropin®-V
treatments, respectively. Thus, data on the size of both ovaries were combined to evaluate effects of
treatment.
The type of Folltropin®-V treatment affected the size of ovaries, with the ovaries in the
double treatment being nearly twice the size of ovaries from the single treatment (3.2 ± 1.0 cm vs.
1.7 ± 0.9 cm; P = 0.0013). Similarly, treatment affected the size distribution of follicles; however,
there was no effect on the mean number of follicles or oocytes recovered or the oocyte recovery rate
(oocytes recovered/follicles aspirated) (Table 5.1). After gonadotropin treatment, the number of
follicles observed on either ovary ranged from 4 to 17.
Most of the recovered oocytes (89%, 98/110) in both treatments had intact zona pellucidae,
homogenous ooplasm and multiple layers of cumulus cells (Table 5.2). The number of oocytes
recovered ranged from 3 to 17 with an average of 8.0 and 8.9 oocytes recovered per procedure in
single and double injection treatments, respectively (Table 5.1). Oocyte recovery rates ranged from
44 to 100% (mean = 73 to 78%). FSH treatment also affected (P = 0.0497) the morphological
distribution of oocytes, with more degenerate/damaged oocytes in the single than in the double

104
injection treatment group (Table 5.2). Analysis of data after excluding degenerate/damaged oocytes
showed that treatment type did not influence the degree of cumulus expansion (P = 0.5259).
5.3.2. Experiment 5.2a (Targeting the Preovulatory or Postovulatory Follicular Wave)
Whether gonadotropin treatment targeted the preovulatory (Treatment 1) or postovulatory
(Treatment 2) follicular wave did not affect ovarian size, number of follicles or number and
percentage of oocytes recovered. However, there was a tendency for more small-sized follicles and
lower oocyte recovery rates when targeting postovulatory follicular waves (Table 5.3). From 1 to 14
and 3 to 9 oocytes were recovered per procedure in Treatments 1 and 2, respectively. The average
number of oocytes recovered was numerically higher when targeting the preovulatory follicular
wave, but the difference was not statistically significant.
Moreover, when the postovulatory wave was targeted, ovarian bleeding tended to increase
during follicular aspiration, presumably due to the presence of corpora hemorrhagica, causing
blockage of the aspiration tubing. Treatment affected the percentage of follicles >10 mm in diameter,
but did not affect other parameters (Table 5.3).
In the 16 oocyte retrieval procedures subsequently done on seven elands, the preovulatory
follicular wave was targeted for stimulation by gonadotropin treatment. The overall mean oocyte
recovery rate in the 16 procedures was 80.7% (total oocytes = 192). The mean numbers of follicles
observed and oocytes recovered were 14.9 ± 6.9 (range = 6 to 28) and 12.0 ± 8.0 (range: 3 to 28),
respectively. The mean ovary size was 3.34 ± 0.95 cm (range = 1.5 to 5.0). When evaluated by the
degree of expansion of cumulus cell layers, cumulus oocytes complexes recovered after estrous
synchronization with altrenogest were more morphologically uniform in appearance than those
recovered after estrous synchronization with 3x prostaglandin treatment.
In Experiments 5.1 and 5.2a, since treatment did not affect ovarian response (i.e., number of
follicles) or the number of oocytes recovered, data from both experiments were combined to evaluate

105
effects of animal and month (season) in Experiment 5.2b. These factors were evaluated because of an
apparent pattern of individual variation in ovarian response to gonadotropin treatment.
Table 5.1. Ovarian response and oocyte recovery rates in elands (n = 4) treated with Folltropin®-Vgiven 2 (single) or 4 and 2 (double) days before ultrasound-guided oocyte retrieval_________________________________________________________________________________________FSH Procedures, Follicles, n (%) by size (mm) Total follicles, Oocytes recovered treatment n <5 mm 5-10 mm >10 mm n (mean ± SD) Total, n (Mean ± SD) %*
_________________________________________________________________________________________
Single 6 13 (21)a 48 (76) 2 (3)a 63 (10.5 ± 3.7) 48 (8.0 ± 3.2) 78
Double 7 7 (8)b 62 (74) 15 (18)b 84 (12.0 ± 4.6) 62 (8.9 ± 4.1) 73
P-value - 0.0498 0.8483 0.0076 0.3337 0.5338 0.6277_________________________________________________________________________________________*Values indicate percentage of oocytes recovered among the follicles detected.a,bValues in the same column with different superscripts are significantly different (P<0.05).
Table 5.2. Morphological distribution of cumulus oocyte complexes recovered from eland donors (n= 4) treated with Folltropin®-V for 2 (single) or 4 and 2 (double) days before oocyte retrieval_________________________________________________________________________________FSH Procedures, Cumulus oocyte complex morphology, n (%)* treatment n Degenerate/damaged Compact cumulus Expanded cumulus_________________________________________________________________________________
Single 6 9 (19)a 22 (46) 17 (35)
Double 7 3 (5)b 38 (61) 21 (34)
P-value - 0.0295 0.2562 1.00_________________________________________________________________________________*Oocytes classified as having compact and expanded cumulus had uniformly homogenous ooplasm.a,bValues in the same column with different superscripts are significantly different (P<0.05).

106
Table 5.3. Effect of targeting the preovulatory or postovulatory follicular wave on ovarian responseto FSH treatment and oocyte recovery rates in elands (n = 6)_________________________________________________________________________________
Procedures, Ovarian Follicles, n (%)a by size, mm Oocytes recovered Treatment n size, cm <5 5 to 10 >10 Total, mean ± SD Total, n (mean±SD) %b
_____________________________________________________________________________________________
Preovulatory 5 3.2 28 (50) 24 (43) 4 (7)c 56 (11.2 ± 4.5) 42 (8.4 ± 5.7) 75
Postovulatory 3 3.4 23 (68) 11 (32) 0 (0)d 34 (11.3 ± 1.5) 19 (6.3 ± 3.0) 56
P-value - 0.875 0.127 0.377 0.045 0.786 0.786 0.0677
_____________________________________________________________________________________________aValues in parentheses indicate percentage of total follicles per treatment. bOf total follicles/treatment.c,dValues in the same column with different superscripts are significantly different (P<0.05).
5.3.3. Experiment 5.2b (Individual and Monthly Variations in Response to OvarianStimulation)
There was a significant animal effect (P = 0.0105) on the number of follicles observed. This
was mainly due to one female (eland 123) having more follicles than three other females (eland 128,
129 and 139; Figure 5.4). Similarly, there was a slight animal effect on the mean number of oocytes
recovered (P = 0.0532).
Three to 10 oocyte retrieval procedures were done on each of the seven females. Overall, an
average of 12.8 follicles was observed and a mean of 9.8 oocytes was recovered (363/475 or 76.4%).
There was a strong correlation between the number of follicles observed and the number of oocytes
recovered (r = 0.9079; Figure 5.6; P< 0.0001). Also, there was a significant correlation between
percentage of oocyte recovery and the number of follicles observed (r = 0.4025; P = 0.0135);
however, the latter correlation may be an artifact because of the difficulty of accurately estimating
the number of follicles in highly responsive females.
Oocyte retrieval procedures were done in all months of the year, except December. There was
no effect of month on the number of follicles observed (P = 0.4417) or number of oocytes recovered
(P = 0.7692). The monthly distribution of procedures, follicles observed and oocytes recovered are
shown in Figure 5.5.

107
Figure 5.4. Effect of animal on follicles observed and oocytes recovered from eland donors.a,bMeans with different letters are significantly different (P<0.05).
0
5
10
15
20
25
30
123 124 125 128 129 139 140Animal ID
Cou
nt o
r m
ean
± S
D
Procedures, n
Follicles, mean
Oocytes, mean
b
b
a
b

108
Figure 5.5. Mean number of follicles observed and oocytes recovered from eland donors during eachmonth of the year.
0
5
10
15
20
25
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov
Cou
nt o
r m
ean
± S
D
Procedures, n
Follicles, mean
Oocytes, mean

109
Figure 5.6. Correlation between the number of follicles observed and oocytes recovered from elanddonors.

110
5.3.4. Morphology of Eland Oocytes
Most eland oocytes (91.7% or 333/363) were morphologically normal in appearance as
judged by criteria used for domestic ungulate oocytes (uniform or non-degenerate ooplasm, intact
zona pellucida and multiple layers of cumulus cells). A small proportion had degenerate ooplasm
(3.9%), lacked a cumulus cell layer (3.3%) or had damaged zonae pellucidae (1.1%; Figure 5.7).
The mean diameter of eland oocytes (n = 14) was 134.6 ± 10.3 µM. The mean thickness of
the zona pellucida was 16.9 ± 5.3 µM; and the mean total diameter, including the zona pellucida, was
172.5 ± 22.4 µM. The only effect of estrous synchronization and ovarian stimulation treatments on
cumulus oocyte complex morphology was on the degree of cumulus cell layer expansion.
5.3.5. Experiment 5.3 (In Vitro Maturation of Eland Oocytes)
The frequency of oocytes maturing to the metaphase-II stage varied between 56% and
73%. None of the three variables evaluated (course of the study, serum type or interval post-
oocyte recovery) affected the meiotic status of oocytes (P > 0.05; Table 5.4).
Analysis of the data after excluding oocytes in which chromatin structures could not be
determined indicated that the post-oocyte recovery interval affected the distribution of oocytes by
meiotic status (P = 0.0446) with a tendency for a higher percentage of metaphase II oocytes at 24 hr
post-recovery. The other two variables, course of study and serum type, did not affect the distribution
of oocytes by meiotic status.
5.3.5. Experiment 5.4 (In Vitro Embryonic Development of Eland Oocytes)
When three methods (IVF, ICSI and somatic cell nuclear transfer, SCNT) of in vitro embryo
production were evaluated, from 18 % to 55 % of the metaphase II oocytes developed to �2-cell
stage, most of which stopped development at ≤8-cell stage (Table 5.5; Figure 5.8). The only

111
blastocysts were derived after SCNT of common eland cells into enucleated eland oocytes. Three of
the four blastocysts were produced using male epithelial cells and the other blastocyst developed
after SCNT using cumulus cells from the oocyte donor.
Initially, the consistently poor in vitro development after IVP of eland oocytes was
considered to be due to an inappropriate culture medium. Therefore, in an effort to improve cleavage
frequency and embryo development in vitro, several different media were examined during the
course of the study (See Material and Methods). Even so, no further improvement in in vitro
development was observed in any single medium or combination of media.
Accordingly, it is inappropriate to discuss the effects of media, other than to say that, under
our culture conditions, none of the media used in the present study were able to adequately and
reliably support high rates of cleavage and subsequent development in vitro of eland embryos.
Nonetheless, development of one SCNT embryo to the blastocyst stage occurred during culture in
CR1aa medium supplemented with 5% eland serum (Figure 5.9). The other three blastocysts
developed during culture in alpha-MEM supplemented with pyruvate (0.25 mM) and 5% bovine calf
serum. Some of the developmental stages of eland oocytes after IVP are shown in Figures 5.8 and
5.9.
Figure 5.7. Morphological appearance of eland cumulus-oocyte complexes recovered byultrasound-guided transvaginal follicular aspiration:A: fully expanded cumulus cell massB: expanded cumulus cell mass with most cells dissociatedC: few cumulus cells, a damaged zona pellucida and ruptured oolemmaBar = 50 µm
A B C

112
Table 5.4. Effect of three factors on the in vitro maturation (meiotic status) of eland oocytesrecovered after Folltropin®-V-induced ovarian stimulation._____________________________________________________________________________________________
Procedures, Oocytesc GV, Metaphase I, Metaphase II, NotFactors* n evaluated, n n (%) n (%) n (%) determined,n(%)d
_____________________________________________________________________________________________
Portion of the study
First 13 81 4 (4.9) 11 (13.5) 45 (55.5) 21 (25.9)
Seconda 20 228 13 (5.7) 25 (11.0) 157 (68.9) 33 (14.5)
Serum type
Eland 14 161 10 (6.2) 14 (8.7) 113 (70.2) 24 (14.9)
Bovine 6 67 3 (4.5) 11 (16.4) 44 (65.7) 9 (13.4)
Duration post recoveryb
~6 hr 4 22 3 (13.6) 4 (18.2) 12 (54.5) 3 (13.6)
~24 hr 16 139 7 (5.0) 10 (7.2) 101 (72.7) 21 (15.1)_____________________________________________________________________________________________*Factors did not affect the distribution of oocytes by each chromatin structure; however, the duration post recovery affected the distribution when data were analyzed after excluding “not determined” oocytes.aThe IVM medium was modified from that used in the first portion of the study (see text).bSome oocytes with fully expanded cumulus cell layer were selected for evaluation on the day ofrecovery (~ 6 hr).
cExcludes degenerate or damaged oocytes or those lost during handling.dIncludes oocytes for which the meiotic status could not be determined after staining.

113
Figure 5.8. (A-J) Status of eland oocytes after applying IVM, IVF (A-H) or SCNT with giant elandsomatic cells (I, J):
A-H: note the presence of one (A, E, F, G) or two (B, H) polar bodies; 0 (A, E, H) or ≥ 1 (B, C, D, F,G) spermatozoa (arrows) attached; and a 3-cell (C) and a 5-cell (D) IVF/IVC-derived eland embryos.
I, J: 3- and ~8- Hoechst-stained nuclei, respectively, present after SCNT with giant eland cells andIVC.
K. Day-9 blastocyst that developed after SCNT of a giant eland cell into enucleated bovine oocyte.
Bar = 50 µm (all except H).H = 400x.
C DB
E G
HF
FC
I J K
A

114
Table 5.5. Development of eland oocytes subjected to three methods of in vitro production (IVP) ofembryos._____________________________________________________________________________________IVP Procedures, Metaphase II Embryonic development, n (%) type n oocytes, n* ≥2-cell 3- to 8-cell >8-cell Morula Blastocyst____________________________________________________________________________________
IVF 15 55 10 (18.2) 6 (10.9) 1 (1.8) 0 0
ICSI 9 44 19 (43.1) 9 (20.5) 1 (2.3) 0 0
SCNT
Common eland 9 48 26 (54.1) 22 (45.8) 8 (16.7) 7 (14.5) 4 (8.3)
Giant eland 2 11 6 (54.5) 5 (45.5) 0 0 0_____________________________________________________________________________________*The remaining MII oocytes were used for activation experiments (data not shown), which indicated thateland oocytes were activated at a higher rate in 10 µM of ionomycin than in 5 µM of ionomycin (Dr.Philip Damiani, personal communication).
Figure 5.9. Day-7 blastocyst (400x) that developed after SCNT into an eland oocyte using a cumuluscell from the same oocyte donor.

115
5.4. Discussion
In Chapter 4, the feasibility of handling female eland in a hydraulic chute for the purpose of
conducting assisted reproductive procedures was demonstrated. However, initially, it was not
possible to access the eland vaginal vault for ultrasound-guided oocyte retrieval with the
commercially available adaptor widely used in domestic cattle. A similar instrument has been
successfully used for transvaginal oocyte retrieval in several ungulate species that are smaller in size
than common eland, including mountain bongo, red deer and wapiti (Pope et al. 1998b; Berg and
Asher 2003).
It was not possible to determine if the difficulty in accessing the vaginal vault was related to
parity status because that information was not known for the eland used in the present study. Since
considerable body size variation does exist among female eland, it is certainly possible that the
commercially available adaptor may be usable on some females; however, we were not able to use it
with 5 females. The custom-molded modified adaptor we designed/built proved to be a reliable
instrument and was used throughout the course of the study. Unfortunately, several weeks were
required to design, build and validate the modified adaptor, which delayed the start of the ovarian
stimulation/oocyte retrieval experiments. Therefore, based on our experience, for similar experiments
in nondomestic ungulates, it is recommended that the size and patency of the vestibular/vulvar
passage be determined before initiating treatments/procedures.
In Experiment 5.1, it was demonstrated that one (single) or two (double) injections of porcine
FSH induce ovarian stimulation and that both treatments produce a similar yield of oocytes after
ultrasound-guided follicular aspiration. The significant increase in the size of ovaries in the double
FSH injection treatment group was clearly indicative of the ovarian response to treatment. Although
ultrasound-guided oocyte retrieval in domestic cows is commonly done without inducing ovarian
stimulation using exogenous gonadotropins, an attempt to collect oocytes in an eland female without

116
gonadotropin treatment was unsuccessful because there were no follicles of sufficient size for
aspiration. Moreover, a maximum of 4 follicles were detected during 4 weeks of twice-weekly
ultrasound scanning of the ovaries of an eland that did not receive gonadotropin treatment (personal
observation).
In domestic cattle, oocyte production without ovarian stimulation is maximized by aspirating
follicles at 2 to 3 day intervals because removal of the dominant follicle leads to the growth of a new
cohort of small follicles. While it was demonstrated that eland can be conditioned for handling in a
chute, they are still nondomesticated animals; therefore, it may not be possible to perform multiple
oocyte retrievals at frequent intervals as practiced in domestic cattle. Thus, it appears that inducing
follicular development with exogenous gonadotropins is an essential component of the treatment
protocol for successful ultrasound-guided retrieval of oocytes in the eland.
Previous studies on FSH-induced ovarian stimulation in the common eland indicate that the
average ovulation rate (number of corpora lutea) or number of ova/embryos recovered have ranged
from 8.4 to 12.3 after twice-daily injections of FSH over a period of 5 days (Schiewe et al. 1991b;
Pope and Loskutoff 1999). In the present study, after single or double injections of FSH, an average
of 12.8 follicles was detected and an average of 9.8 oocytes was recovered, suggesting that ovarian
response after single or double injections of FSH may be similar to that obtained after twice daily
FSH injections for 5 days. However, the proportion of eland follicles that would eventually ovulate
after ovarian stimulation with a reduced number of FSH injections, as described in the present study,
remains to be determined. In domestic cows, ovulation frequencies were reported to be similar when
FSH dissolved in saline was administered in multiple injections or it was dissolved in 30% PVP and
given as single injection (Yamamoto et al. 1994).
In ungulates, cumulus cell layer expansion and progression of meiosis from the germinal
vesicle to the metaphase II stage are mediated by LH. In domestic cows, cumulus cell expansion after
FSH (Folltropin®-V) treatment is minimal when pulsatile release of endogenous LH is suppressed by

117
concomitant administration of a GnRH agonist (Lindsey et al. 2002). In Experiment 5.1 of the
present study, regardless of the treatment type, ~35% of eland oocytes had expanded cumulus cell
layers at the time of recovery (Table 5.2). Oocytes with similar morphology were also frequently
observed during Experiment 5.2. The cumulus cell expansion observed in eland oocytes after
gonadotropin treatment may be a result of increased frequency of the endogenous pulsatile secretion
of LH, a surge of endogenous LH or a direct consequence of the LH present in the exogenous FSH
used for injection.
In Experiment 5.1, we anticipated that the longer exposure of follicles to FSH (96 hr vs. 48
hr) in the double FSH treatment group would produce oocytes with more cumulus cell expansion.
However, cumulus cell expansion was similar in both the single and double FSH injection treatment
groups. A possible explanation for the lack of difference between treatments may be that the extent
of cumulus expansion is determined mainly by the length of time between the preovulatory
administration of prostaglandin and oocyte collection. This interval was ~48 hr in both treatment
groups.
In Experiment 5.2, the stimulation of follicular development obtained by targeting the
follicular wave that emerges immediately after ovulation provided indirect evidence for the
emergence of a follicular wave following ovulation. This evidence of a postovulatory follicular wave,
along with the known preovulatory follicular wave that leads to ovulation, is indicative that the
common eland has at least two follicular waves, as seen in other ruminants (Adams 1999).
Confirmation of this will require frequent monitoring of follicular dynamics throughout the estrous
cycle. Nevertheless, there was no specific advantage of targeting the first (postovulatory) follicular
wave in terms of oocyte yield. Moreover, this treatment required a longer treatment period (15 days)
as compared with targeting the preovulatory wave (9 days).
Oral administration of altrenogest, combined with a single injection of prostaglandin, was
effective for estrous synchronization in the eland. Altrenogest treatment required fewer handling

118
events than needed for estrous cycle synchronization using multiple (3x) prostaglandin injections.
Furthermore, its use appeared to produce a more uniform group of oocytes, although its effects on
oocyte development are not known. Another indication that the estrous synchronization protocols
were effective was the basal levels of serum progesterone found in samples (n = 30, data not shown)
taken at the time of oocyte recovery.
The oocyte recovery rate obtained in the present study (76%) was relatively higher than most
rates reported in domestic cattle. In most of the eland procedures with the lowest oocyte recovery
rates, technical problems occurred, such as blockage of the aspiration tubing. Since it appears that
almost all aspirated follicles can produce oocytes, the high recovery rate suggests that the cumulus
oocyte complex is loosely attached to the follicular wall. Cell-to-cell contact may be loosened by
ovarian stimulation treatment, as previously suggested to occur in gonadotropin-treated domestic
cattle (Gordon 1994).
In domestic cattle, the mean number of oocytes recovered by ultrasound-guided follicular
aspiration in heifers was 9.1 after twice daily treatment with FSH (20 mg divided into six equal
doses) for three consecutive days or 7.1 oocytes when FSH was administered in decreasing doses
(Sirard et al. 1999). In another study using crossbred domestic cows (Goodhand et al. 2000), it was
reported that the average number of oocytes recovered after single (5.3 oocytes) or multiple (5.9
oocytes) FSH injections were similar and did not differ from that of nontreated females (4.1 oocytes);
however, more follicles were observed in FSH treated cows.
One of the highest numbers of oocytes recovered using ultrasonography in an ungulate
species was achieved after a single FSH injection in gaur cows primed with Synchromate-B® for 5 to
7 days (Armstrong et al. 1995). The females received a single injection of 200 mg of Folltropin®-V
dissolved in 1% carboxymethylcellulose and 2% Tween-80. An average of 34.3 (n = 4) oocytes/cow
were recovered on the third day after gonadotropin treatment and 15 to 17 % of the oocytes
developed to the blastocyst stage after IVF/IVC. A pregnancy was produced after transferring one of

119
the blastocysts to a domestic cow. Perez (2003) reported an average of 17.6 follicles per treatment
after administering a single injection of FSH dissolved in 30% PVP to postpartum beef cows. After
IVF/IVC, ~30 % of the oocytes developed to blastocysts. These studies indicate that gaur and bovine
oocytes recovered after a single injection of FSH are developmentally competent.
Several factors influence ovarian response and oocyte recovery rate, including season,
individual animal variation and ovarian status at the initiation of gonadotropin treatment (Adams and
Pierson 1995). Another factor affecting oocyte recovery rate is technician efficiency; however, this
effect was not evaluated in the present study because one person (the author) did all of the oocyte
recovery procedures. In the present study, month of the year (or season) did not have a detectable
influence on ovarian response to FSH treatment; however, there was a significant animal effect on
the number of follicles observed after gonadotropin treatment (Figure 5.4). Moreover, the effect of
animal on the number of oocytes recovered approached statistical significance (P = 0.0532).
Although the cause for such variation is unknown, similar individual differences in embryo
yield among eland donors after gonadotropin-induced ovarian stimulation have been documented
(Dresser et al. 1984). In the latter report, transcervical flushing of the uterus produced an average of
8 to 10 ova/embryos, including one female from which, 31 embryos were recovered. Thus, individual
differences in superovulatory response, rather than disparate fertilization rates, may have been the
primary cause of the variation in embryo yield.
The mean diameter of eland oocytes (135 µm) was larger than that of domestic cattle oocytes
[(114 µm, (Otoi et al. 1997)] collected from antral follicles, but smaller than that of Asian buffalo
oocytes [146 µm, (Raghu et al. 2002)]. Most domestic bovine oocytes (82%) recovered from
slaughterhouse ovaries had diameters ranging raging between 110 and 125 µm, and only 5% were
larger in size than 130 µm (Otoi et al. 1997).

120
Some embryonic development occurred after IVF, ICSI and SCNT of eland oocytes. Even
though in vitro maturation rates were satisfactory, embryonic development was very poor after
homologous IVF of eland oocytes. The almost complete lack of development after IVF was, to some
extent, due to the low motility of the eland spermatozoa. Alternatively, standard domestic bovid IVF
methods may not produce satisfactory results in the eland.
Cleavage frequency was similar after nuclear transfer into enucleated common eland
oocytes of either common eland or giant eland somatic cell nuclei. The development of
blastocysts in the former group may be because more replicates were attempted. This poor
development is in contrast to the more acceptable in vitro development (up to 77% cleavage, and
27% blastocyst formation) found when giant eland nuclei were transferred into enucleated
bovine oocytes and cultured under similar conditions (Damiani et al. 2003). These results
demonstrate that the ooplasm environment is a critical factor influencing the development of
eland nuclear transfer couplets.
Clearly, methods for activation of couplets and subsequent in vitro culture of eland embryos
are suboptimal. Nevertheless, the present study presents the initial demonstration that in vitro derived
eland embryos can be produced and provides the framework for further research. Additional studies
focusing on ultrastructural, molecular and biochemical events will provide further insights into the
biology of eland gametes/embryos and such information may be used to improve methods of in vitro
embryo production in this species.
To conclude, the present study has demonstrated that an average of 9.8 oocytes can be
collected in sedated common elands after a single or double injection(s) of FSH. The number of
follicles available for oocyte collection after one or two injections of FSH was similar to observations
in other eland studies in which FSH was given as multiple (~10) injections over several days. Eland
oocytes are capable of undergoing in vitro maturation and some embryonic development in vitro.

121
Future work to define and understand factors that influence in vitro developmental
competence of oocytes is warranted, including studies on basic aspects of eland gamete physiology,
cryobiology of eland spermatozoa and molecular/cytological analysis of embryos to verify
fertilization and embryonic genome activity. Moreover, endocrine profiles associated with ovarian
stimulation and their effect on the yield and developmental competence (in vitro) of eland oocytes
require characterization.

122
Chapter 6. Summary and Conclusions
The first part of the study (Chapter 3) evaluated several key components and attributes of
embryo culture media to determine the requirements of bovine embryos in protein-free media. In
Experiment 1, the effects of a group of either 20 (glutamine + essential + non-essential) or 11
(HECM-6) amino acids were evaluated in modified KSOM or Basic Medium (BM)-3. Results
showed that modified KSOM containing 20 amino acids (mKSOM-20aa) supported the highest
frequency of total, expanded (days 7, 8, and 9) and hatched blastocysts. This experiment
demonstrated that the beneficial effects of amino acids on the in vitro development of bovine
embryos are influenced by the base medium. The huge difference in embryonic development
between two base media (BM-3 and KSOM) was puzzling. Thus, a second experiment was designed
to identify the factors responsible for the developmental disparity.
In Experiment 2, the effects of glucose, pyruvate, lactate, phosphate or all four supplements
were evaluated in low (255) or high (275) osmotic pressure BM-3 containing 20 amino acids (BM-3-
20aa). Supplement type affected the frequency of development to at least the morula stage (day 7),
expanded (day 8), hatched (day 9) or total blastocysts and cell number per blastocyst. Osmotic
pressure affected the frequency of expanded blastocysts (day 7) and blastocyst cell number, with a
tendency for higher frequencies and more cell numbers in the low osmotic pressure treatments.
Regardless of the osmotic pressure, BM-3-20aa containing glucose (0.2 mM) supported the
highest frequency of blastocyst development. The interaction between supplement type and osmotic
pressure was not significant; however, treatment mean differences were more discernible in high than
in low osmotic pressure medium. Glucose was identified as the most crucial supplement of BM-3-
20aa and its addition led to embryonic development equivalent to that observed in protein-free
KSOM. The higher frequency of embryonic development in glucose containing media as compared

123
with pyruvate- and/or phosphate-supplemented media suggests that glucose plays more important
roles in non-energy generating pathways.
Difficulties and risks associated with restraining large nondomestic ungulates are limiting
factors to the development and application of assisted reproductive technologies, such as artificial
insemination and embryo transfer. Thus, the next studies evaluated the use of behavioral training and
handling of common eland females in a hydraulic chute on the possibility of doing transvaginal
ultrasound-guided oocyte retrieval without inducing general anesthesia. Some physiological
responses of elands to the handling and factors influencing them were assessed. The in vitro
developmental potential of eland oocytes was also evaluated.
In Chapter 4, 9 of 10 females were conditioned to associate specific sound cues with food
treats. The interval from the audio cues until acceptance of handheld treats varied among females.
Animals also differed in their response to training for voluntary entry into the chute.
Handling eland for doing oocyte retrievals in the hydraulic chute required sedation, which
was achieved using xylazine, either alone or in combination with butorphanol. During sedation and
handling, elands undergoing oocyte retrieval procedures had higher blood glucose levels than
females handled similarly but without doing oocyte retrieval. Females that were more difficult to
train to the chute had higher blood glucose levels than the more cooperative animals.
Other procedures including embryo transfer, ultrasound examination of the reproductive
tract, hoof trimming, body weight measurement and surgical correction of a persistent vaginal hymen
were also done without inducing general anesthesia and, in some cases, without sedation. There were
no complications, injuries or capture myopathy in more than 40 handling procedures. However, the
high blood glucose (and slightly elevated CPK) levels occurring during sedation and handling for
oocyte retrieval suggest the need to minimize the level of physical exertion and the associated stress.
Preliminary data suggested that certain physical attributes (number of lateral stripes) might be

124
associated with taming potential of elands; however, the confirmation of actual associations requires
the study of larger sample sizes.
In Chapter 5, data on ovarian response to gonadotropin treatments, the number and rate of
oocytes recovered and the in vitro developmental potential of eland oocytes were presented.
Ultrasound-guided oocyte retrieval in the eland required designing an adaptor for the transducer that
was smaller than the commercial adaptor. A single or double injection of FSH dissolved in 30%
polyvinyl pyrrolidone induced acceptable ovarian stimulation for oocyte retrieval. Modifications of
the estrous synchronization or ovarian stimulation protocol did not affect ovarian response or number
of oocyte recovered; however, daily oral administration of altrenogest combined with a single
injection of prostaglandin was effective for estrous synchronization and enabled oocyte collection
within 9 days of the start of synchronization/ovarian stimulation treatments.
In 37 oocyte retrieval procedures, an average of 9.8 oocytes were recovered (76% recovery
rate) by transvaginal ultrasound-guided follicular aspiration. Most of the oocytes appeared
morphologically normal and up to 73% matured to the metaphase II stage after 6 or 24 hr of in vitro
maturation. On applying three methods of in vitro embryo production (IVF, ICSI, SCNT) to the
eland oocytes, cleavage frequency varied between 18% after IVF to 55 % after nuclear transfer. Most
of the cleaved embryos stopped development at ≤ 8-cell stage. The only blastocyst development (n =
4; 8.3%) was observed after SCNT using common eland cells.
In addition to determining the role of key attributes and/or components of defined (protein-
free) embryo culture media, the experiments on bovine embryo culture served as a background
quality control for the eland IVP studies. Many of the eland embryo production experiments were
simultaneously carried out either on the same day or during the same week as the domestic bovine
studies, using similar embryo culture components.

125
The poor in vitro embryonic development in elands using protocols that produced acceptable
bovine embryo development in the same laboratory by the same technician suggests that either 1)
factors other than embryo culture media are more important in supporting the development of eland
embryos in vitro, or 2) eland oocytes/embryos differ in their in vitro culture requirements. Thus,
additional studies are required to improve the development of eland oocytes in vitro.

126
References
Abdel-Fattah M, Amer H, Ghoneim MA, Warda M, Megahed Y (1999) Response of one-humpedcamel (Camelus dromedarius) to intravenous glucagon injection and to infusion of glucose andvolatile fatty acids, and the kinetics of glucagon disappearance from the blood. J Vet.Med.Ser.A 46,473-481.
Abdullahi AN (1981) Forage preference of cattle and eland on a desert grassland in southernArizona. Dissert.Abstr.Int.B: Sci Engineer. 41, 2424 (Abstr).
Abe H, Otoi T, Tachikawa S, Yamashita S, Satoh T, Hoshi H (1999) Fine structure of bovinemorulae and blastocysts in vivo and in vitro. Anat.Embryol.(Berl) 199, 519-527.
Abe H, Yamashita S, Satoh T, Hoshi H (2002) Accumulation of cytoplasmic lipid droplets in bovineembryos and cryotolerance of embryos developed in different culture systems using serum-free orserum-containing media. Mol.Reprod.Dev. 61, 57-66.
Adams CE (1982) Egg transfer: Historical aspects. In 'Mammalian Egg Transfer'. (Ed. CE Adams)pp. 1-17. (CRC Press: Boca Raton, Florida)
Adams GP (1999) Comparative patterns of follicle development and selection in ruminants. JReprod.Fertil.Suppl 54, 17-32.
Adams GP, Pierson RA (1995) Bovine model for study of ovarian follicular dynamics in humans.Theriogenology 43, 113-120.
Al Katanani YM, Paula-Lopes FF, Hansen PJ (2002) Effect of season and exposure to heat stress onoocyte competence in Holstein cows. J Dairy Sci 85, 390-396.
Ali J, Shahata MA, Al Natsha SD (2000) Formulation of a protein-free medium for human assistedreproduction. Hum.Reprod. 15, 145-156.
Allen WR (1982) Embryo transfer in the horse. In 'Mammalian Egg Transfer'. (Ed. CE Adams) pp.135-154. (CRC Press:Boca Raton, Florida).
Alonso-Pozos I, Rosales-Torres AM, Avalos-Rodriguez A, Vergara-Onofre M, Rosado-Garcia A(2003) Mechanism of granulosa cell death during follicular atresia depends on follicular size.Theriogenology 60, 1071-1081.
Armstrong DL, Looney CR, Lindsey BR, Gonseth CL, Johnson DL, Williams KR, Simmons LG,Loskutoff NM (1995) Transvaginal egg retrieval and in vitro embryo production in gaur (Bosgaurus) with establishment of interspecies pregnancies. Theriogenology 43, 162 (Abstr).
Asa CS, Bauman KL, Houston EW, Fischer MT, Junge RE, Krisher RL (1998) Transvaginal oocyteretrieval and in vitro maturation, fertilization and embryo culture in addax (Addax nasomaculatus).Theriogenology 49, 261 (Abstr).

127
Atkinson MW, Welsh TH, Blumer ES (1999) Evaluation of an advanced handling system forphysiological data collection, testing and medical treatment of large, nondomestic hoofstock. Proc.Am.Assoc.Zoo Vet, Columbus, OH, USA. pp. 154-157.
Baguisi A, Behboodi E, Melican DT, Pollock JS, Destrempes MM, Cammuso C, Williams JL, NimsSD, Porter CA, Midura P, Palacios MJ, Ayres SL, Denniston RS, Hayes ML, Ziomek CA, MeadeHM, Godke RA, Gavin WG, Overstrom EW, Echelard Y (1999) Production of goats by somatic cellnuclear transfer. Nat.Biotechnol. 17, 456-461.
Ball GD, Leibfried ML, Lenz RW, Ax RL, Bavister BD, First NL (1983) Factors affecting successfulin vitro fertilization of bovine follicular oocytes. Biol Reprod. 28, 717-725.
Balmaceda JP, Heitman TO, Garcia MR, Pauerstein CJ, Pool TB (1986) Embryo cryopreservation incynomolgus monkeys. Fertil.Steril. 45, 403-406.
Balmaceda JP, Pool TB, Arana JB, Heitman TS, Asch RH (1984) Successful in vitro fertilization andembryo transfer in cynomolgus monkeys. Fertil.Steril. 42, 791-795.
Barbehenn EK, Wales RG, Lowry OH (1974) The explanation for the blockade of glycolysis in earlymouse embryos. Proc.Natl.Acad.Sci U.S.A 71, 1056-1060.
Barboni B, Turriani M, Galeati G, Spinaci M, Bacci ML, Forni M, Mattioli M (2000) Vascularendothelial growth factor production in growing pig antral follicles. Biol Reprod. 63, 858-864.
Barker CA, Gandier JC (1957) Pregnancy in a mare resulted from frozen epididymal spermatozoa.Can.J Comp Med.Vet.Sci 21, 45-51.
Barnett DK, Bavister BD (1992) Hypotaurine requirement for in vitro development of goldenhamster one- cell embryos into morulae and blastocysts, and production of term offspring from invitro-fertilized ova. Biol Reprod. 47, 297-304.
Bartels P, Friedmann Y, Lubbe K, Mortimer D, Rasmussen LA, Godke RA (2001) The live birth ofan eland (Taurotragus oryx) calf following estrous synchronization and artificial insemination usingfrozen-thawed epididymal sperm. Theriogenology 55, 381 (Abstr).
Batt PA, Miller BG (1988) Development of sheep embryos in vitro in a medium supplemented withdifferent batches of serum albumin. Aust.J Biol Sci 41, 371-376.
Bavister BD (1989) A consistently successful procedure for in vitro fertilization of golden hamstereggs. Gamete Res. 23, 139-158.
Bavister BD (1995) Culture of preimplantation embryos: Facts and artifacts. Hum.Reprod.Update 1,91-148.
Bavister BD (2002a) Early history of in vitro fertilization. Reproduction. 124, 181-196.
Bavister BD (2002b) Timing of embryo development. In 'Assessment of mammalian embryo quality:Invasive and non-invasive techniques'. (Eds. A Van Soom and ML Boerjan) pp. 139-155. (KluwerAcademic Publishers: Dordrecht, The Netherlands).

128
Bavister BD, Boatman DE, Collins K, Dierschke DJ, Eisele SG (1984) Birth of rhesus monkey infantafter in vitro fertilization and nonsurgical embryo transfer. Proc.Natl.Acad.Sci U.S.A 81, 2218-2222.
Bavister BD, Yanagimachi R (1977) The effects of sperm extracts and energy sources on the motilityand acrosome reaction of hamster spermatozoa in vitro. Biol Reprod. 16, 228-237.
Benirschke K, Ruedi D, Muller H, Kumamoto AT, Wagner KL (1980) The unusual karyotype of thelesser kudu, Tragelaphus imberbis. Cytogenet.Cell Genet. 26, 85-92.
Bennet SD, Foster WR (1985) Successful transfer of zebra embryo to a domestic horse. Equine Vet.JSuppl 3, 53-62.
Berg DK, Asher GW (2003) New developments reproductive technologies in deer. Theriogenology59, 189-205.
Berg DK, Beaumont SE, Berg MC, Asher GW (2004) Red deer (Cervus elaphus) calves born from invitro-produced blastocysts fertilized and cultured in deer synthetic oviduct fluid. Reprod.Fertil.Dev.16, 222 Abstr.
Berg DK, Pugh PA, Thompson JG, Asher GW (2002) Development of in vitro embryo productionsystems for red deer (Cervus elaphus). Part 3. In vitro fertilisation using sheep serum as acapacitating agent and the subsequent birth of calves. Anim Reprod.Sci 70, 85-98.
Bergfelt DR, Brogliatti GM, Adams GP (1998) Gamete recovery and follicular transfer (graft) usingtransvaginal ultrasonography in cattle. Theriogenology 50, 15-25.
Betteridge KJ (2003) A history of farm animal embryo transfer and some associated techniques.Anim Reprod.Sci 79, 203-244.
Biggers JD, McGinnis LK (2001) Evidence that glucose is not always an inhibitor of mousepreimplantation development in vitro. Hum.Reprod. 16, 153-163.
Biggers JD, McGinnis LK, Raffin M (2000) Amino acids and preimplantation development of themouse in protein- free potassium simplex optimized medium. Biol Reprod. 63, 281-293.
Blaha GC (1964) Effects of age of the donor and recipient on the development of transferred goldenhamster ova. Anat.Rec. 150, 413-416.
Bloomsmith MA, Jones ML, Snyder RJ, Singer RA, Gardner WA, Liu SC, Maple TL (2003) Positivereinforcement training to elicit voluntary movement of two giant pandas throughout their enclosure.Zoo Biol 22, 323-334.
Boever J, Knox D, Merilan CP, Read BW (1980) Estrus induction and artificial insemination withsuccessful pregnancy in Speke's gazelle. Proc.9th Int Cong Anim Reprod.AI, Madrid, Spain pp. 565-569.
Boone WR, Catlin JC, Casey KJ, Dye PS, Boone ET, Shuett RJ (1997) World's first bear cubproduced from a non-surgical embryo collection. 53rd Ann.Mtg.Am.Soc.Reprod. Med. Cincinnati,OH, USA. p. P-157 (Abstr).

129
Bourke DA, Kyle CE, McEvoy TG, Young P, Adam CL (1995) Superovulatory responses to eCG inllamas (Llama glama). Theriogenology 44, 255-268.
Boyd EH, Mikota S, Smith J, Aguilar R, deMaar T, Hunt D, Hosgood G (2000) Blood gas analysis inbongo (Tragelaphus eurycerus) and eland (Tragelaphus oryx) antelope. Proc.Am.Assoc.Zoo Vet.Int.Assoc.Aquatic Anim Med.Joint Conf., New Orleans, LA, USA) pp. 106-110.
Brackett BG, Bousquet D, Boice ML, Donawick WJ, Evans JF, Dressel MA (1982) Normaldevelopment following in vitro fertilization in the cow. Biol Reprod. 27, 147-158.
Bredbacka K, Bredbacka P (1995) Effect of polyvinylpyrrolidone, serum albumin and fetal calfserum on early cleavage of bovine embryos. Theriogenology 43, 174 (Abstr).
Briggs R, King TJ (1952) Transplantation of living nuclei from blastula cells into enucleated frogs'eggs. Proc.Natl.Acad.Sci U.S A 38, 455-463.
Brogliatti GM, Palasz AT, Rodriguez-Martinez H, Mapletoft RJ, Adams GP (2000) Transvaginalcollection and ultrastructure of llama (Lama glama) oocytes. Theriogenology 54, 1269-1279.
Brück I, Raun K, Synnestvedt B, Greve T (1992) Follicle aspiration in the mare using a transvaginalultrasound-guided technique. Equine Vet.J 24, 58-59.
Buckrell BC, Gartley CJ, Mehren KG, Crawshaw GJ, Johnson WH, Barker IK, Balke J, Coghill C,Challis JR, Goodrowe KL (1990) Failure to maintain interspecific pregnancy after transfer of Dall'ssheep embryos to domestic ewes. J Reprod.Fertil. 90, 387-394.
Bunch TD, Foote WC, Whitaker B (1977) Interspecies ovum transfer to propagate wild sheep. JWildlife Mgmt. 41, 726-730.
Caamano JN, Ryoo ZY, Youngs CR (1998) Promotion of development of bovine embryos producedin vitro by addition of cysteine and beta-mercaptoethanol to a chemically defined culture system. JDairy Sci 81, 369-374.
Callesen H, Greve T, Christensen CR (1987) Ultrasonically guided follicular aspiration of bovinefollicular oocytes. Theriogenology 27, 217 (Abstr).
Carney EW, Foote RH (1991) Improved development of rabbit one-cell embryos to the hatchingblastocyst stage by culture in a defined, protein-free culture medium. J Reprod.Fertil. 91, 113-123.
Catt JW (1996) Intracytoplasmic sperm injection (ICSI) and related technology. Anim Reprod.Sci 12,239-250.
Chan AW, Luetjens CM, Dominko T, Ramalho-Santos J, Simerly CR, Hewitson L, Schatten G(2000) Foreign DNA transmission by ICSI: Injection of spermatozoa bound with exogenous DNAresults in embryonic GFP expression and live rhesus monkey births. Mol.Hum.Reprod. 6, 26-33.
Chang MC (1959) Fertilization of rabbit ova in vitro. Nature 179, 466-467.
Chang MC (1968) Reciprocal insemination and egg transfer between ferrets and mink. J Exp.Zool.168, 49-59.

130
Chastant-Maillard S, Quinton H, Lauffenburger J, Cordonnier-Lefort N, Richard C, Marchal J,Mormede P, Renard JP (2003) Consequences of transvaginal follicular puncture on well-being incows. Reproduction 125, 555-563.
Cheng WTK, Moor RM, Polge C (1986) In vitro fertilization of pig and sheep oocytes matured invivo and in vitro. Theriogenology 25, 146 (Abstr).
Chesne P, Adenot PG, Viglietta C, Baratte M, Boulanger L, Renard JP (2002) Cloned rabbitsproduced by nuclear transfer from adult somatic cells. Nat.Biotechnol. 20, 366-369.
Cho SR, Cho SK, Lee SL, Lee HJ, Choe SY, Rho GJ (2002) Enhanced cryosurvival of bovineblastocysts produced in vitro in serum- free medium. J Assist.Reprod.Genet. 19, 487-492.
Cibelli JB, Stice SL, Golueke PJ, Kane JJ, Jerry J, Blackwell C, Ponce de Leon FA, Robl JM (1998)Cloned transgenic calves produced from nonquiescent fetal fibroblasts. Science 280, 1256-1258.
Citino SB (1995) The use of a modified, large cervid hydraulic squeeze chute for restraint of exoticungulates. Proc.Am.Assoc.Zoo Vet., Wildl Dis Assoc.Am.Assoc.Wildl Vet.Joint Conf. pp. 297-298.
Citino SB (2003) Bovidae (except sheep and goats) and Anitlocapridae. In 'Zoo and Wild AnimalMedicine'. (Eds. ME Fowler and RE Miller) pp. 649-674. (Saunders: St. Louis, Missouri, USA)
Clayton O, Kuehl T (1984) The first successful in vitro fertilization and embryo transfer in a non-human primate. Theriogenology 21, 228 (Abstr).
Cochran R, Meintjes M, Reggio B, Carter J, Pinto C, Paccamonti D, Godke RA (1998) Live foalsproduced from sperm-injected oocytes derived from pregnant mares. J Equine Vet.Sci 18, 736-740.
Comizzoli P, Mermillod P, Mauget R (2000) Reproductive biotechnologies for endangeredmammalian species. Reprod.Nutr.Dev. 40, 493-504.
Comizzoli P, Urner F, Sakkas D, Renard JP (2003) Up-regulation of glucose metabolism during malepronucleus formation determines the early onset of the S phase in bovine zygotes. Biol Reprod. 68,1934-1940.
Coonrod SA, Flores-Foxworth G, Moreno JF, Westhusin M, Byrd SR, Kraemer DC (1994) Birth ofArmenian red sheep (Ovis orientalis) lambs to domestic sheep (Ovis aries) following interspecificembryo transfer of IVM-IVF derived embryos. Theriogenology 41, 182 (Abstr).
Cottle CA, Price EO (1987) Effects of the nonagouti pelage-color allele on the behavior of captivewild Norway rats (Rattus norvegicus). J Comp Psychol. 101, 390-394.
Crandall LS (1964) 'Management of Wild Mammals in Captivity.' (The University of Chicago Press:Chicago, IL, USA)
Crump JP, Crump JW (1994) Manual collection from a Grevy's zebra stallion (Equus grevyi), onsetof sperm production, semen characteristics, and cryopreservation of semen with a comparison to thesperm production from a Grant's zebra stallion (Equus burchelli boehmi). Theriogenology 41, 1011-1021.

131
Curry MR (2000) Cryopreservation of semen from domestic livestock. Rev.Reprod. 5, 46-52.
Daels PF, Albrecht BA, Mohammed HO (1998) Equine chorionic gonadotropin regulates lutealsteroidogenesis in pregnant mares. Biol Reprod. 59, 1062-1068.
Damiani P, Wirtu G, Miller F, Cole A, Pope C, Godke R, Dresser BL (2003) Development of gianteland antelope (Taurotragus derbianus) embryos following nuclear transfer with common eland(Taurotragus oryx) and bovine (Bos taurus) oocytes. Theriogenology 59, 390 (Abstr).
Davies CJ, Antczak DF, Allen WR (1985) Reproduction in mules: Embryo transfer using sterilerecipients. Equine Vet.J Suppl 3, 63-67.
de Loos F, van Vliet C, van Maurik P, Kruip TA (1989) Morphology of immature bovine oocytes.Gamete Res. 24, 197-204.
Del Campo MR, Del Campo CH, Adams GP, Mapletoft RJ (1995) The application of newreproductive technologies to South American camelids. Theriogenology 43, 23-30.
Denney RN (1970) Body temperatures of some wild East African ungulates. E.Afr.Wildl.J 8, 212-216.
Densmore MA, Bowen MJ, Mgyar SJ, Amoss MS, Robinson RM, Harms PG, Kraemer DC (1987)Artificial insemination with frozen, thawed semen and pregnancy diagnosis in addax (Addaxnasomaculatus). Zoo Biol 6, 21-29.
Di Berardino MA (1997) 'Genomic Potential of Differentiated Cells.' (Columbia University Press:New York, NY, USA)
Dittrich L (1972) Gestation periods and age of sexual maturity of some African antelopes. Int.ZooYb. 12, 184-187.
Dixon TE, Hunter IW, Beatson NS (1991) Pregnancies following the export of frozen red deerembryos from New Zealand to Australia. Theriogenology 35, 193 (Abstr).
Domínguez CS, Aguilar RF (2001) San Juan de Argon, Mexico: The challenge of relocating an entirezoo. Proc.Am.Assoc.Zoo Vet.Int. Assoc.Aquatic Anim Med.Joint Conf pp. 497-498.
Dominko T, Mitalipova M, Haley B, Beyhan Z, Memili E, McKusick B, First NL (1999) Bovineoocyte cytoplasm supports development of embryos produced by nuclear transfer of somatic cellnuclei from various mammalian species. Biol Reprod. 60, 1496-1502.
Donoghue AM, Byers AP, Johnston LA, Armstrong DL, Wildt DE (1996) Timing of ovulation aftergonadotrophin induction and its importance to successful intrauterine insemination in the tiger(Panthera tigris). J Reprod.Fertil. 107, 53-58.
Donoghue AM, Johnston LA, Seal US, Armstrong DL, Tilson RL, Wolf P, Petrini K, Simmons LG,Gross T, Wildt DE (1990) In vitro fertilization and embryo development in vitro and in vivo in thetiger (Panthera tigris). Biol Reprod. 43, 733-744.

132
Dorn CG (1995) Application of reproductive technologies in the North American bison (Bisonbison). Theriogenology 43, 13-20.
Downing BG, Mohr LR, Trounson AO, Freemann LE, Wood C (1985) Birth after transfer ofcryopreserved embryos. Med.J Aust. 142, 409-411.
Dozortsev D, Wakaiama T, Ermilov A, Yanagimachi R (1998) Intracytoplasmic sperm injection inthe rat. Zygote. 6, 143-147.
Dresser BL, Gelwicks EJ, Wachs KB, Keller GL (1988) First successful transfer of cryopreservedfeline (Felis catus) embryos resulting in live offspring. J Exp.Zool. 246, 180-186.
Dresser BL, Kramer L, Dahlhausen RD, Pope CE (1984) Cryopreservation followed by successfultransfer of African eland antelope (Tragelaphus oryx) embryos.. Proc.10th Int.Cong.Anim Reprod.Artif.Insemin. (Urbana, IL, USA). pp. 191-193
Dresser BL, Pope CE, Kramer L, Kuehn G, Dahlhausen RD, Maruska EJ, Reece B, Thomas WD(1985) Birth of bongo antelope (Tragelaphus euryceros) to eland antelope (Tragelaphus oryx) andcryopreservation of bongo antelopes. Theriogenology 23, 190 (Abstr).
Drost M, Wright RW, Jr., Cripe WS, Richter AR (1983) Embryo transfer in the water buffalo(Bubalus bubalis). Theriogenology 20, 79-584.
Dumonceaux GA, Burton MS, Ball RL, Demuth A (1998) Veterinary procedures facilitated bybehavioral conditioning and desensitization in reticulated giraffe (Giraffa camelopardalis) and NileHippopotamus (Hippopotamus amphibius). Proc.Am.Assoc.Zoo Vet.Am.Assoc.Wildl.Vet.Joint Conf.pp. 388-391.
East R (1999) 'African antelope database 1998. IUCN/SSC Antelope Specialist Group.' (IUCN:Gland, Switzerland and Cambridge, UK)
Ebedes H, Raath JP (1995) The use of long term neuroleptics in the confinement and transport ofwild animals. Proc.Am.Assoc.Zoo.Vet: East Lansing, MI, USA. pp. 173-176.
Elhassan YM, Wu G, Leanez AC, Tasca RJ, Watson AJ, Westhusin ME (2001) Amino acidconcentrations in fluids from the bovine oviduct and uterus and in KSOM-based culture media.Theriogenology 55, 1907-1918.
Erbach GT, Lawitts JA, Papaioannou VE, Biggers JD (1994) Differential growth of the mousepreimplantation embryo in chemically defined media. Biol Reprod. 50, 1027-1033.
Erickson BH (1966) Development and senescence of the postnatal bovine ovary. J Anim Sci 25, 800-805.
Essop MF, Harley EH, Baumgarten I (1997) A molecular phylogeny of some Bovidae based onrestriction-site mapping of mitochondrial DNA. J Mammal. 78, 377-386.
Estienne MJ, Harper AF, Horsley BR, Estienne CE, Knight JW (2001) Effects of P.G. 600 on theonset of estrus and ovulation rate in gilts treated with Regu-mate. J Anim Sci 79, 2757-2761.

133
Fekete E, Little CC (1942) Observations on the mammary tumor incidence of mice born fromtransplanted ova. Cancer Res. 2, 525-530.
Ferguson CE, Perez O, Cartmill J, Green H, Godke RA (2002) Synchronizing estrus in yearling beefheifers with melengestrol acetate and altrenogest. Theriogenology 57, 375 (Abstr).
Fernández-Arias A, Folch J, Alabart JL, Ramon JP (1996) Successful interspecific embryo transferbetween Spanish ibex (Capra pyrenaica) and domestic goat (Capra hircus) using micro-osmoticpumps for FSH administration. Theriogenology 45, 247 (Abstr).
Flores-Foxworth G, Coonrod SA, Moreno JF, Byrd SR, Kraemer DC, Westhusin M (1995)Interspecific transfer of IVM IVF-derived red sheep (Ovis orientalis gmelini) embryos to domesticsheep (Ovis aries). Theriogenology 44, 681-690.
Foote RH (2002) The history of artificial insemination: Selected notes and notables. J Anim Sci 80,E1-E10.
Fridman T (2001) Axis deer training at Disney's animal kingdom. Anim Keeper's Forum 28, 146-148.
Gabriel Sanchez-Partida L, Maginnis G, Dominko T, Martinovich C, McVay B, Fanton J, Schatten G(2000) Live rhesus offspring by artificial insemination using fresh sperm and cryopreserved sperm.Biol Reprod. 63, 1092-1097.
Galli C, Lagutina I, Crotti G, Colleoni S, Turini P, Ponderato N, Duchi R, Lazzari G (2003a)Pregnancy: A cloned horse born to its dam twin. Nature 424, 635.
Galli C, Vassiliev I, Lagutina I, Galli A, Lazzari G (2003b) Bovine embryo development followingICSI: Effect of activation, sperm capacitation and pre-treatment with dithiothreitol. Theriogenology60, 1467-1480.
Galvez AS, Ulloa JA, Chiong M, Criollo A, Eisner V, Barros LF, Lavandero S (2003) Aldosereductase induced by hyperosmotic stress mediates cardiomyocyte apoptosis: Differential effects ofsorbitol and mannitol. J Biol Chem. 278, 38484-38494.
Ganhao MF, Hattingh J, Pitts N, Raath C, de Klerk B, de Vos V (1988) Physiological responses ofblesbok, eland and red hartebeest to different capture methods. S.Afr.J Wildl.Res. 18, 134-136.
Gardner DK, Pool TB, Lane M (2000) Embryo nutrition and energy metabolism and its relationshipto embryo growth, differentiation, and viability. Semin.Reprod.Med. 18, 205-218.
Garland P, Frazer L, Sanderson N, Mehren N, Kroetsch T (1992) Artificial insemination of scimitar-horned oryx at Orana Park with frozen semen from Metro Toronto Zoo. Symp.Zool.Soc.Lond. 64, 37-43.
Gelwicks EJ, Pope CE, Gillespie DG, Burton MS, Dresser BL (1989) Embryo collection, splittingand transfer in the common eland (Tragelaphus oryx). Theriogenology 31, 196 (Abstr).
Gleicher N, Friberg J, Fullan N, Giglia RV, Mayden K, Kesky T, Siegel I (1983) Egg retrieval for invitro fertilization by sonographically controlled vaginal culdocentesis. Lancet 2, 508-509.

134
Godfrey RW, Lunstra DD, French JA, Schwartz J, Armstrong DL, Simmons LG (1990) Estroussynchronization in the gaur (Gaur gaurus): Behavior and fertility to artificial insemination afterprostaglandin treatment. Zoo Biol 10, 35-41.
Goh VH, Adiga SK, Tain CF, Tong TY, Ng SC (2000) Successful in vitro growth of rat two-cellembryos to blastocysts using a simple chemically defined medium. J Pharmacol.Toxicol.Methods 43,171-175.
Gomez MC, Pope CE, Harris R, Davis A, Mikota S, Dresser BL (2000) Births of kittens produced byintracytoplasmic sperm injection of domestic cat oocytes matured in vitro. Reprod.Fertil.Dev. 12,423-433.
Goodhand KL, Staines ME, Hutchinson JS, Broadbent PJ (2000) In vivo oocyte recovery and in vitroembryo production from bovine oocyte donors treated with progestagen, oestradiol and FSH. AnimReprod.Sci 63, 145-158.
Goodhand KL, Watt RG, Stallknecht DE, Hutchinson JS, Broadbent PJ (1999) In vivo oocyterecovery and in vitro embryo production from bovine donors aspirated at different frequencies orfollowing FSH treatment. Theriogenology 51, 951-961.
Goodrowe KL, Hay M, King WA (1991) Nuclear maturation of domestic cat ovarian oocytes invitro. Biol Reprod. 45, 466-470.
Goodrowe KL, Miller AM, Wildt DE (1989) In vitro fertilization of gonadotropin-stimulated leopardcat (Felis bengalensis) follicular oocytes. J Exp.Zool. 252, 89-95.
Goodrowe KL, Wall RJ, O'Brien SJ, Schmidt PM, Wildt DE (1988) Developmental competence ofdomestic cat follicular oocytes after fertilization in vitro. Biol Reprod. 39, 355-372.
Gordon I (1994) 'Laboratory production of cattle embryos.' (CAB International: Wallingford, UK)
Goto K, Kinoshita A, Takuma Y, Ogawa K (1990) Fertilization of bovine oocytes by the injection ofimmobilized, killed spermatozoa. Vet.Rec. 127, 517-520.
Gould KG, Styperek RP (1989) Improved method for freeze preservation of chimpanzee sperm. Am.JPrimatol. 18, 275-284.
Graff KJ, Meintjes M, Han Y, Reggio BC, Denniston RS, Gavin WG, Ziomek C, Godke RA (2000)Comparing follicle stimulating hormone from two commercial sources for oocyte production fromout-of-season dairy goats. J Dairy Sci 83, 484-487.
Grandin T (2000) Habituating antelope and bison to cooperate with veterinary procedures. J App.Anim Welfare Sci 3, 253-261.
Grandin T, Rooney MB, Phillips M, Cambre RC, Irlbeck NA, Graffam W (1995) Conditioning ofnyala (Tragelaphus angasi) to blood sampling in a crate with positive reinforcement. Zoo Biol 14,261-273.

135
Haigh JC (2001) Handling and successful reproduction of animals: Can successes (and failures) indeer be applied to other species? 1st Int.Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl.,Omaha, NE, USA.pp. 67-72.
Hall-Woods ML (2001) In vitro embryo production in exotic bovids. 1st Int.Symp.Assist.Reprod.Technol.Conserv. Genet.Mgmt.Wildl., Omaha, NE, USA. pp. 69-73.
Hall-Woods ML, Asa CS, Bauman KL, Houston EW, Fischer MT, Junge RE, Krisher RL (1999)In vitro embryo production in addax (Addax nasomaculatus), an endangered desert antelope. 32nd
Ann. Mtg.Soc.Study Reprod. Pullman, WA, USA.
Hammer CJ, Tyler HD, Loskutoff NM, Armstrong DL, Funk DJ, Lindsey BR, Simmons LG (2001)Compromised development of calves (Bos gaurus) derived from in vitro-generated embryos andtransferred interspecifically into domestic cattle (Bos taurus). Theriogenology 55, 1447-1455.
Han Y, Meintjes M, Graff K, Denniston R, Zhang L, Ziomek C, Godke R (2001) Caprine offspringborn from fresh and frozen-thawed in vitro-produced embryos. Vet.Rec. 149, 714-716.
Hansel W (2003) The potential for improving the growth and development of cultured farm animaloocytes. Anim Reprod.Sci 79, 191-201.
Hasler JF (2003) The current status and future of commercial embryo transfer in cattle. AnimReprod.Sci 79, 245-264.
Hayashi S, Kobayashi K, Mizuno J, Saitoh K, Hirano S (1989) Birth of piglets from frozen embryos.Vet.Rec. 125, 43-44.
Heape W (1890) Preliminary note on the transplantation and growth of mammalian ova within auterine foster mother. Proc.Royal Soc., London 48, 457-458.
Heape W (1897) The artificial insemination of mammals and subsequent possible fertilization orimpregnation of their ova. Proc.Royal Soc., London 61, 52-63.
Hearn JP, Summers PM (1986) Experimental manipulation of embryo implantation in the marmosetmonkey and exotic equids. Theriogenology 25, 3-11.
Hemmer H (1976) Man's strategy in domestication - a synthesis of new research trends. Experientia32, 663-666.
Hemmer H (1990) 'Domestication: The Decline of Environmental Appreciation.' (CambridgeUniversity Press: Cambridge)
Hernandez-Fonseca HJ, Sirisathien S, Bosch P, Cho HS, Lott JD, Hawkins LL, Hollett RB, ColeySL, Brackett BG (2002) Offspring resulting from direct transfer of cryopreserved bovine embryosproduced in vitro in chemically defined media. Anim Reprod.Sci 69, 151-158.
Herrick JR, Bartels P, Krisher RL (2002) Cryopreservation and post-thaw evaluation of epididymalspermatozoa recovered from wild African bovids. 2nd Int Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE, USA. pp. 292-293.

136
Heyman Y, Vincent C, Garnier V, Cognie Y (1987) Transfer of frozen-thawed embryos in sheep.Vet.Rec. 120, 83-85.
Hildebrandt TB, Hermes R, Göritz F (2002) Novel instrument designs to facilitate assistedreproduction in non-domestic animals. 2nd Int.Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE, USA. pp. 219-223.
Hinrichs K (1998) Production of embryos by assisted reproduction in the horse. Theriogenology 49,13-21.
Hinrichs K, Williams KA (1997) Relationships among oocyte-cumulus morphology, follicularatresia, initial chromatin configuration, and oocyte meiotic competence in the horse. Biol Reprod. 57,377-384.
Ho Y, Doherty AS, Schultz RM (1994) Mouse preimplantation embryo development in vitro: Effectof sodium concentration in culture media on RNA synthesis and accumulation and gene expression.Mol.Reprod.Dev. 38, 131-141.
Holden H (2003) Another endangered species cloned. Science 300, 421.
Holm P, Booth PJ, Callesen H (2002) Kinetics of early in vitro development of bovine in vivo- and invitro-derived zygotes produced and/or cultured in chemically defined or serum- containing media.Reproduction 123, 553-565.
Holm P, Booth PJ, Schmidt MH, Greve T, Callesen H (1999) High bovine blastocyst development ina static in vitro production system using SOFaa medium supplemented with sodium citrate and myo-inositol with or without serum-proteins. Theriogenology 52, 683-700.
Holm P, Callesen H (1998) In vivo versus in vitro produced bovine ova: Similarities and differencesrelevant for practical application. Reprod.Nutr.Dev. 38, 579-594.
Holt WV, Abaigar T, Jabbour HN (1996) Oestrous synchronization, semen preservation and artificialinsemination in the Mohor gazelle (Gazella dama mhorr) for the establishment of a genome resourcebank programme. Reprod.Fertil.Dev. 8, 1215-1222.
Holt WV, Moore HD, North RD, Hartman TD, Hodges JK (1988) Hormonal and behaviouraldetection of oestrus in blackbuck, Antilope cervicapra, and successful artificial insemination withfresh and frozen semen. J Reprod.Fertil. 82, 717-725.
Holt WV, Pickard AR (1999) Role of reproductive technologies and genetic resource banks in animalconservation. Rev.Reprod. 4, 143-150.
Hosoi Y, Miyake M, Utsumi K, Iritani A (1988) Development of rabbit oocytes after microinjectionof spermatozoa. 11th Int.Cong.Anim Reprod.Artif.Insem, Dublin, Ireland. p. 331 (Abstr).
Howard JG, Bush M, Morton C, Morton F, Wentzel K, Wildt DE (1991) Comparative semencryopreservation in ferrets (Mustela putorius furo) and pregnancies after laparoscopic intrauterineinsemination with frozen-thawed spermatozoa. J Reprod.Fertil. 92, 109-118.

137
Howard JG, Donoghue AM, Barone MA, Goodrowe KL, Blumer ES, Snodgrass K, Starnes D,Tucker M, Bush M, Wildt DE (1992) Successful induction of ovarian activity and laparoscopicintrauterine artificial insemination in the cheetah (Actinonyx jubatus). J Zoo Wildl.Med. 23, 288-300.
Howard JG, Roth TL, Byers AP, Swanson WF, Wildt DE (1997) Sensitivity to exogenousgonadotropins for ovulation induction and laparoscopic artificial insemination in the cheetah andclouded leopard. Biol Reprod. 56, 1059-1068.
Huang Y, Wang P, Zhang G, Zhang H, Li D, Du J, Wei R, Tang C, Spindler RE, Wildt DE, HowardJG (2002) Use of artificial insemination to enhance propagation of giant pandas at the WolongBreeding Center. 2nd Int.Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE,USA). pp. 172-179.
Hubner K, Fuhrmann G, Christenson LK, Kehler J, Reinbold R, De La FR, Wood J, Strauss JF, III,Boiani M, Scholer HR (2003) Derivation of oocytes from mouse embryonic stem cells. Science 300,1251-1256.
Humphries JE, Jr. (1972) The Eland as a Meat Producer in the American Southwest: A NewApproach to Land-use. 'Ph.D. Dissertation,' University of Oklahoma Graduate College.
Illmensee K, Hoppe PC (1981) Nuclear transplantation in Mus musculus: Developmental potential ofnuclei from preimplantation embryos. Cell 23, 9-18.
IUCN. 2002 IUCN Red List of Threatened Species. <http://www.redlist.org> downloaded 30October 2003 . 2002.
Jacobsen H, Schmidt M, Holm P, Sangild PT, Greve T, Callesen H (2000) Ease of calving, bloodchemistry, insulin and bovine growth hormone of newborn calves derived from embryos produced invitro in culture systems with serum and co-culture or with PVA. Theriogenology 54, 147-158.
Johnston LA, Parrish JJ, Monson R, Leibfried-Rutledge L, Susko-Parrish JL, Northey DL, RutledgeJJ, Simmons LG (1994) Oocyte maturation, fertilization and embryo development in vitro and in vivoin the gaur (Bos gaurus). J Reprod.Fertil. 100, 131-136.
Johnston SD, McGowan MR, Blyde D (2002) Birth of a Banteng (Bos javanicus) calf at WesternPlains Zoo after fixed time artificial insemination. Aust.Vet.J 80, 94-95.
Johnston SD, McGowan MR, O'Callaghan P, Cox R, Houlden B, Haig S, Taddeo G (1999) Birth ofkoala pouch young following artificial insemination. p. 135. Melbourne, Australia)
Junge RE (1998) Epidural analgesia in addax (Addax nasomaculatus). J Zoo Wildl.Med. 29, 285-287.
Junior SM, Armstrong DL, Hopkins SH, Simmons LG, Schiewe MC, Gross TS (1990) Semencryopreservation and the first successful artificial insemination of the gaur (Gaur gaurus).Theriogenology 33, 262 (Abstr).
Kane MT (1983) Variability in different lots of commercial bovine serum albumin affects cellmultiplication and hatching of rabbit blastocysts in culture. J Reprod.Fertil. 69, 555-558.

138
Kane MT, Bavister BD (1988) Protein-free culture medium containing polyvinylalcohol, vitamins,and amino acids supports development of eight-cell hamster embryos to hatching blastocysts. JExp.Zool. 247, 183-187.
Kato Y, Tani T, Sotomaru Y, Kurokawa K, Kato J, Doguchi H, Yasue H, Tsunoda Y (1998) Eightcalves cloned from somatic cells of a single adult. Science 282, 2095-2098.
Keskintepe L, Brackett BG (1996) In vitro developmental competence of in vitro-matured bovineoocytes fertilized and cultured in completely defined media. Biol Reprod. 55, 333-339.
Keskintepe L, Burnley CA, Brackett BG (1995) Production of viable bovine blastocysts in defined invitro conditions. Biol Reprod. 52, 1410-1417.
Keskintepe L, Morton PC, Smith SE, Tucker MJ, Simplicio AA, Brackett BG (1997) Caprineblastocyst formation following intracytoplasmic sperm injection and defined culture. Zygote 5, 261-265.
Khurana NK, Niemann H (2000) Energy metabolism in preimplantation bovine embryos derived invitro or in vivo. Biol Reprod. 62, 847-856.
Kidson A, Lambrechts H, Bartels P, Denniston RS, Barry DM (2000) In vitro embryo developmentin the Cape buffalo (Syncerus caffer): Age, parity and oocyte quality-dependent differencesfollowing IVF. Theriogenology 53, 335 (Abstr).
Kiley-Worthington M (1978) The social organization of small captive group of eland, oryx and roanantelope with an analysis of personality profiles. Behaviour 66, 32-55.
Kim JH, Niwa K, Lim JM, Okuda K (1993) Effects of phosphate, energy substrates, and amino acidson development of in vitro-matured, in vitro-fertilized bovine oocytes in a chemically defined,protein-free culture medium. Biol Reprod. 48, 1320-1325.
Kimura Y, Yanagimachi R (1995) Intracytoplasmic sperm injection in the mouse. Biol Reprod. 52,709-720.
Kingdon J (1982) 'East African Mammals: An Atlas of Evolution in Africa Volume III part C(Bovids).' (Academic Press: New York, NY, USA)
Kinney GM, Pennycook JW, Schriver MD, Templeton JW, Kraemer DC (1979) Surgical collectionand transfer of canine embryos. Biol Reprod. 20, 96A.
Knowles TG, Warris PD (2000) Stress physiology of animals during transport. In 'LivestockHandling and Transport'. (Ed. T Grandin) pp. 385-406. (CAB International: Wallingford, UK)
Koulischer L, Tijskens J, Mortelmans J (1973) Chromosome studies of a fertile mammalian hybrid:The offspring of the cross bongo x sitatunga (Bovoidea). Chromosoma 41, 265-270.
Kraemer DC (1983) Intra- and interspecific embryo transfer. J Exp.Zool. 228, 363-371.
Kraemer DC, Moore GT, Kramen MA (1976) Baboon infant produced by embryo transfer. Science192, 1246-1247.

139
Kramer L, Dresser BL, Pope CE, Dahlhausen RD, Reed G (1982) Collection, transfer andcryopreservation of African eland (Taurotragus oryx) embryos. Proc.Am.Assoc.Zoo Vet. pp. 129-131
Kren R, Kikuchi K, Nakai M, Miyano T, Ogushi S, Ngai T, Suzuki S, Fulka J, Fulka J, Jr. (2003)Intracytoplasmic sperm injection in the pig: Where is the problem? J Reprod.Dev. 49, 271-273.
Krisher RL, Bavister BD (1999) Enhanced glycolysis after maturation of bovine oocytes in vitro isassociated with increased developmental competence. Mol.Reprod.Dev. 53, 19-26.
Krisher RL, Lane M, Bavister BD (1999) Developmental competence and metabolism of bovineembryos cultured in semi-defined and defined culture media. Biol Reprod. 60, 1345-1352.
Kusunoki H, Daimaru H, Minami S, Nishimoto S, Yamane KI, Fukumoto Y (2001) Birth of achimpanzee (Pan troglodytes) after artificial insemination with cryopreserved epididymalspermatozoa collected postmortem. Zoo Biol 20, 135-143.
Kvasnitski AV (2001) Research on interbreed ova transfer in pigs. Theriogenology 56, 1285-1289.
Kydd J, Boyle MS, Allen WR, Shephard A, Summers PM (1985) Transfer of exotic equine embryosto domestic horses and donkeys. Equine Vet.J Suppl 3, 80-83.
Kyle R (1972) Will the antelope recapture Africa? New Scientist 53, 640-643.
Lacham-Kaplan O, Shaw J, Sanchez-Partida LG, Trounson A (2003) Oocyte activation afterintracytoplasmic injection with sperm frozen without cryoprotectants results in live offspring frominbred and hybrid mouse strains. Biol Reprod. 69, 1683-1689.
Lacham-Kaplan O, Trounson A (1995) Intracytoplasmic sperm injection in mice: Increasedfertilization and development to term after induction of the acrosome reaction. Hum.Reprod. 10,2642-2649.
Landim-Alvarenga FC, Boyazoglu SE, Carvalho LR, Choi YH, Squires EL, Seidel GE, Jr. (2002)Effects of fetuin on zona pellucida hardening, fertilization and embryo development in cattle. AnimReprod.Sci 71, 181-191.
Lane M, Bavister BD (1999) Regulation of intracellular pH in bovine oocytes and cleavage stageembryos. Mol.Reprod.Dev. 54, 396-401.
Lane M, Bavister BD, Lyons EA, Forest KT (1999) Containerless vitrification of mammalianoocytes and embryos. Nat.Biotechnol. 17, 1234-1236.
Lane M, Gardner DK, Hasler MJ, Hasler JF (2003) Use of G1.2/G2.2 media for commercial bovineembryo culture: Equivalent development and pregnancy rates compared to co-culture.Theriogenology 60, 407-419.
Lane M, Hooper K, Gardner DK (2001) Effect of essential amino acids on mouse embryo viabilityand ammonium production. J Assist.Reprod.Genet. 18, 519-525.

140
Lanza RP, Cibelli JB, Diaz F, Moraes CT, Farin PW, Farin CE, West MD, Damiani P (2000a)Cloning of an endangered species (Bos gaurus) using cross-species nuclear transfer. Cloning 2, 79-90.
Lanza RP, Dresser BL, Damiani P (2000b) Cloning Noah's ark. Sci Am. 283, 84-89.
Larsen WJ, Chen L, Powers R, Zhang H, Russell PT, Chambers C, Hess K, Flick R (1996) Cumulusexpansion initiates physical and developmental autonomy of the oocyte. Zygote 4, 335-341.
Lasley BL, Loskutoff NM, Anderson GB (1994) The limitation of conventional breeding programsand the need and promise of assisted reproduction in non-domestic species. Theriogenology 41, 119-132.
Ledda S, Loi P, Branca A, Cappai P, Dattena M, Naitana S (1992) Use of a protein-free medium forthe in vitro development of sheep embryos. Boll.Soc.Ital.Biol.Sper. 68, 305-309.
Lee B, Wirtu GG, Damiani P, Pope E, Dresser BL, Hwang W, Bavister BD (2003) Blastocystdevelopment after intergeneric nuclear transfer of Mountain bongo antelope somatic cells into bovineoocytes. Cloning Stem Cells 5, 25-33.
Lee ES, Fukui Y (1996) Synergistic effect of alanine and glycine on bovine embryos cultured in achemically defined medium and amino acid uptake by vitro-produced bovine morulae andblastocysts. Biol.Reprod. 55, 1383-1389.
Leese HJ (2002) Quiet please, do not disturb: A hypothesis of embryo metabolism and viability.Bioessays 24, 845-849.
Leibo SP, Songsasen N (2002) Cryopreservation of gametes and embryos of non-domestic species.Theriogenology 57, 303-326.
Lim JM, Kim JH, Okuda K, Niwa K (1994) The importance of NaCl concentration in a chemicallydefined medium for the development of bovine oocytes matured and fertilized in vitro.Theriogenology 42, 421-432.
Lindeberg H, Aalto J, Amstislavsky S, Piltti K, Jarvinen M, Valtonen M (2003) Surgical recoveryand successful surgical transfer of conventionally frozen-thawed embryos in the farmed Europeanpolecat (Mustela putorius). Theriogenology 60, 1515-1525.
Lindeberg H, Amstislavsky S, Jarvinen M, Aalto J, Valtonen M (2002) Surgical transfer of in vivoproduced farmed European polecat (Mustela putorius) embryos. Theriogenology 57, 2167-2177.
Lindsey BR, Maclellan LJ, Whyte TR, Kinder JE, D'Occhio MJ (2002) Differential requirement forpulsatile LH during the follicular phase and exposure to the preovulatory LH surge for oocytefertilization and embryo development in cattle. Theriogenology 58, 1651-1662.
Liu Z, Foote RH (1995a) Development of bovine embryos in KSOM with added superoxidedismutase and taurine and with five and twenty percent O2. Biol.Reprod. 53, 786-790.
Liu Z, Foote RH (1995b) Effects of amino acids on the development of in-vitro matured/in-vitrofertilization bovine embryos in a simple protein-free medium. Hum.Reprod. 10, 2985-2991.

141
Liu Z, Foote RH (1996) Sodium chloride, osmolyte, and osmolarity effects on blastocyst formationin bovine embryos produced by in vitro fertilization (IVF) and cultured in simple serum-free media. JAssist.Reprod.Genet. 13, 562-568.
Liu Z, Foote RH (1997) Effects of amino acids and alpha-amanitin on bovine embryo developmentin a simple protein-free medium. Mol.Reprod.Dev. 46, 278-285.
Loi P, Ptak G, Barboni B, Fulka J, Jr., Cappai P, Clinton M (2001) Genetic rescue of an endangeredmammal by cross-species nuclear transfer using post-mortem somatic cells. Nat.Biotechnol. 19, 962-964.
Lonergan P, Carolan C, Mermillod P (1994) Development of bovine embryos in vitro followingoocyte maturation under defined conditions. Reprod.Nutr.Dev. 34, 329-339.
Lopata A, Summers PM, Hearn JP (1988) Births following the transfer of cultured embryos obtainedby in vitro and in vivo fertilization in the marmoset monkey (Callithrix jacchus). Fertil.Steril. 50,503-509.
Loskutoff NM (1999) Interspecies embryo transfer-a feasible strategy for safeguarding endangeredspecies? Proc.Am.Assoc.Zoo.Vet, Columbus, OH, USA. pp.100-103.
Loskutoff NM, Bartels P, Meintjes J, Godke RA, Schiewe MC (1995) Assisted reproductivetechnology in nondomestic ungulates: A model approach to preserving and managing geneticdiversity. Theriogenology 43, 3-12.
Loskutoff NM, Betteridge KJ (1992) Embryo technology in pets and endangered species. In'Embryonic Development and Manipulation in Animal Production: Trends in Research andApplications'. (Eds. A Lauria and F Gandolfi) pp. 235-248. (Portland Press: London and Chapel Hill)
Loskutoff NM, Bowsher TR, Chatfield JA, Stones GA, Ramey JW, Zhang L, Putman M, Boland C,Wharton D, Gardner DK (2004) Ovarian stimulation, transvaginal, ultrasound-guided oocyteretrieval, ICSI and blastocyst production in sequential media in the western lowland gorilla (Gorillagorilla gorilla). Reprod.Fertil.Dev. 16, 225 (Abstr).
Loskutoff NM, Raphael BL, Nemec LA, Wolfe BA, Howard JG, Kraemer DC (1990) Reproductiveanatomy, manipulation of ovarian activity and non-surgical embryo recovery in suni (Neotragusmoschatus zuluensis). J Reprod.Fertil. 88, 521-532.
Ludwig TE, Lane M, Bavister BD (2001) Differential effect of hexoses on hamster embryodevelopment in culture. Biol.Reprod. 64, 1366-1374.
Lydon, SA (2001) North American Regional Studbook: Common eland, Taurotragus oryx. (Boston,MA, USA). pp. 5-29.
Maclellan LJ, Carnevale EM, Coutinho da Silva MA, Scoggin CF, Bruemmer JE, Squires EL (2002)Pregnancies from vitrified equine oocytes collected from super-stimulated and non-stimulated mares.Theriogenology 58, 911-919.
Madan ML, Singla SK, Chauhan MS, Manik RS (1994) In vitro production and transfer of embryosin buffaloes. Theriogenology 41, 139-143.

142
Madzingira O, Mukaratirwa S, Pandey VS, Dorny P (2002) A questionnaire survey of themanagement and use of anthelmintics in cattle and antelope in mixed farming systems in Zimbabwe.J S.Afr.Vet.Assoc. 73, 70-73.
Magyar SJ, Biediger T, Hodges C, Kraemer DC, Seager SW (1988a) A method of artificialinsemination in captive white-tailed deer (Odocoileus virginianus). Theriogenology 31, 1075-1079.
Magyar SJ, Hodges C, Seager SW, Kraemer DC (1988b) Successful nonsurgical embryo collectionwith surgical transfer in captive white-tailed deer. Theriogenology 29, 273 (Abstr).
Marshall VS, Kalishman J, Thomson JA (1997) Nonsurgical embryo transfer in the commonmarmoset monkey. J Med.Primatol. 26, 241-247.
Marston JH, Penn R, Sivelle PC (1977) Successful autotransfer of tubal eggs in the rhesus monkey(Macaca mulatta). J Reprod.Fertil. 49, 175-176.
Martin DE, Graham CE, Gould KG (1978) Successful artificial insemination in the chimpanzee.Symp.Zool.Soc.Lond. 43, 249-260.
Martin MJ (2000) Development of in vivo-matured porcine oocytes following intracytoplasmicsperm injection. Biol Reprod. 63, 109-112.
Massip A, Mermillod P, Dinnyes A (1995) Morphology and biochemistry of in-vitro producedbovine embryos: Implications for their cryopreservation. Hum.Reprod. 10, 3004-3011.
Matshikiza M, Bartels P, Vajta G, Olivier F, Spies T, Bartels GE, Harley EH, Baumgarten I, CallesenH (2004) Embryo development following interspecies nuclear transfer of African buffalo (Synceruscaffer), bontebok (Damaliscus dorcus dorcus), and eland (Taurotragus oryx). Reprod.Fertil.Dev. 16,150 (Abstr).
McGrath J, Solter D (1983) Nuclear transplantation in the mouse embryo by microsurgery and cellfusion. Science 220, 1300-1302.
McKiernan SH, Bavister BD (1992) Different lots of bovine serum albumin inhibit or stimulate invitro development of hamster embryos. In Vitro Cell Dev.Biol. 28A, 154-156.
McKiernan SH, Clayton MK, Bavister BD (1995) Analysis of stimulatory and inhibitory amino acidsfor development of hamster one-cell embryos in vitro. Mol.Reprod.Dev. 42, 188-199.
McKinnel RG (1985) 'Cloning of Frogs, Mice, and Other Animals.' (University of Minnesota Press:Minneapolis, MN, USA)
McKinnon AO, Tinson AH, Nation G (1994) Embryo transfer in Dromedary camels. Theriogenology41, 145-150.
Meintjes M, Bellow MS, Broussard JR, Paul JB, Godke RA (1995) Transvaginal aspiration ofoocytes from hormone-treated pregnant beef cattle for in vitro fertilization. J Anim Sci 73, 967-974.
Meintjes M, Bezuidenhout C, Bartels P, Visser DS, Meintjes J, Loskutoff NM, Fourie FL, Barry DM,Godke RA (1997) In vitro maturation and fertilization of oocytes recovered from free- ranging

143
Burchell's zebra (Equus burchelli) and Hartmann's zebra (Equus zebra hartmannae). J ZooWildl.Med. 28, 251-259.
Meng L, Ely JJ, Stouffer RL, Wolf DP (1997) Rhesus monkeys produced by nuclear transfer. BiolReprod. 57, 454-459.
Merilan CP, Read BW, Boever WJ, Knox D (1978) Eland semen collection and freezing.Theriogenology 10, 265-268.
Merilan CP, Sikes JD, Read BW, Boever WJ, Kane KK (1977) Comparative characteristics ofspermatozoa and semen from greater kudu, Speke's gazelle and springbuck antelope. J Zoo AnimMed. 84, 16-18.
Miller AM, Roelke ME, Goodrowe KL, Howard JG, Wildt DE (1990) Oocyte recovery, maturationand fertilization in vitro in the puma (Felis concolor). J Reprod.Fertil. 88, 249-258.
Misra AK, Joshi BV, Agrawala PL, Kasiraj R, Sivaian S, Rangereddi NS, Siddiqui MU (1990)Multiple ovulation and embryo transfer in Indian buffalo. Theriogenology 33, 1131-1141.
Miyano T (2003) Bringing up small oocytes to eggs in pigs and cows. Theriogenology 59, 61-72.
Mogoa EG, Stegmann GF, Guthrie AJ, Swan GE (2000) Clinical, cardiopulmonary andhaemocytological effects of xylazine in goats after acute exposure to different environmentaltemperature and humidity conditions. J S.Afr.Vet.Assoc. 71, 153-159.
Moley KH, Chi MM, Manchester JK, McDougal DB, Lowry OH (1996) Alterations ofintraembryonic metabolites in preimplantation mouse embryos exposed to elevated concentrations ofglucose: A metabolic explanation for the developmental retardation seen in preimplantation embryosfrom diabetic animals. Biol Reprod. 54, 1209-1216.
Monfort SL, Asher GW, Wildt DE, Wood TC, Schiewe MC, Williamson LR, Bush M, Rall WF(1993) Successful intrauterine insemination of Eld's deer (Cervus eldi thamin) with frozen-thawedspermatozoa. J Reprod.Fertil. 99, 459-465.
Monis H, Bavister BD (1990) Analysis of the inhibitory effect of inorganic phosphate ondevelopment of four-cell hamster embryos in vitro. J Exp.Zool. 256, 75-83.
Moore HD, Bonney RC, Jones DM (1981) Successful induced ovulation and artificial inseminationin the puma (Felis concolor). Vet.Rec. 108, 282-283.
Morrow CJ, Asher GW, Berg DK, Tervit HR, Pugh PA, McMillan WH, Beaumont SE, Hall DR, BellAC (1994) Embryo transfer in Fallow deer (Dama dama): Superovulation, embryo recovery, andlaparoscopic transfer of fresh and cryopreserved embryos. Theriogenology 42, 579-590.
Mulley RC, Moore NW, English AW (1988) Successful uterine insemination of fallow deer withfresh and frozen semen. Theriogenology 29, 459-466.
Naitana S, Bogliolo L, Ledda S, Leoni G, Madau L, Falchi S, Mezzeddu M (2000) Survival ofvitrified mouflon (Ovis g. musimon) blastocysts. Theriogenology 53, 340 (Abstr).

144
Ndeereh DR, Mbithi PM, Kihurani DO (2001) The reversal of xylazine hydrochloride by yohimbineand 4-aminopyridine in goats. J S.Afr.Vet.Assoc. 72, 64-67.
Nicholas JS (1933) Development of transplanted rat eggs. Proc.Soc.Exp.Biol.Med. 30, 1111-1113.
Nicholas JS, Hall BV (1942) Experiments on developing rats II: The development of isolatedblastomeres and fused eggs. J Exp.Zool. 90, 441.
Nielsen L (1996) Chemical immobilization of free-ranging terrestrial mammals. In 'Lumb and Jone'sVeterinary Anesthesia'. (Eds. JC Thurmon, WJ Tranquilli and GJ Benson). pp. 736-764. (Williamsand Wilkins: Baltimore, MD, USA)
Niemann H, Wrenzycki C (2000) Alterations of expression of developmentally important genes inpreimplantation bovine embryos by in vitro culture conditions: Implications for subsequentdevelopment. Theriogenology 53, 21-34.
Nowak RM (1999) 'Walker's Mammals of the World: Volume II.' (The John Hopkins UniversityPress: Baltimore and London).
Nuojua-Huttunen S, Tuomivaara L, Juntunen K, Tomas C, Kauppila A, Martikainen H (1995)Intrafollicular insemination for the treatment of infertility. Hum.Reprod. 10, 91-93.
Ogonuki N, Mochida K, Inoue K, Matsuda J, Yamamoto Y, Takano K, Ogura A (2003) Fertilizationof oocytes and birth of normal pups following intracytoplasmic injection with spermatids inmastomys (Praomys coucha). Biol.Reprod. 68, 1821-1827.
Oguri N, Tsutsumi Y (1974) Non-surgical egg transfer in mares. J Reprod.Fertil. 41, 313-320.
Olson SE, Seidel GE, Jr. (2000) Reduced oxygen tension and EDTA improve bovine zygotedevelopment in a chemically defined medium. J Anim Sci 78, 152-157.
Onishi A, Iwamoto M, Akita T, Mikawa S, Takeda K, Awata T, Hanada H, Perry AC (2000) Pigcloning by microinjection of fetal fibroblast nuclei. Science 289, 1188-1190.
Otoi T, Yamamoto K, Koyama N, Tachikawa S, Suzuki T (1997) Bovine oocyte diameter in relationto developmental competence. Theriogenology 48, 769-774.
Palermo G, Joris H, Devroey P, Van Steirteghem AC (1992) Pregnancies after intracytoplasmicinjection of single spermatozoon into an oocyte. Lancet 340, 17-18.
Palmer E, Bézard J, Magistrini M, Duchamp G (1991) In vitro fertilization in the horse. Aretrospective study. J Reprod.Fertil.Suppl 44, 375-384.
Panicke L, Fischer E, Staufenbiel R, Reklewski Z (2002) Variation of parameters of the glucosetolerance test (GTT) in growing cattle. Anim Sci Papers Rep. 20 (Suppl 1), 55-61.
Pantaleon M, Ryan JP, Gil M, Kaye PL (2001) An unusual subcellular localization of GLUT1 andlink with metabolism in oocytes and preimplantation mouse embryos. Biol Reprod. 64, 1247-1254.
Pappas LA (2002) Taurotragus oryx. Mammalian Species 689, 1-5.

145
Paris DB, Taggart DA, Temple-Smith PD, Shaw G, Renfree MB (2002) Intra-uterine artificialinsemination in the tammar wallaby (Macropus eugenii). 2nd Int.Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE, USA. pp. 80-84.
Parrish JJ, Susko-Parrish J, Winer MA, First NL (1988) Capacitation of bovine sperm by heparin.Biol.Reprod. 38, 1171-1180.
Perez O (2003) Oocyte Production in the Early Postpartum Cow. 'Ph.D. Dissertation,' Louisiana StateUniversity, Baton Rouge, LA, USA.
Perez-Garnelo S, Gutierrez-Adan A, Romero A, Delclaux M, Lopez M, de la FJ (2000) Seminalcharacteristics and cryopreservation of sitatunga (Tragelaphus spekei gratus) sperm obtained byelectroejaculation. Theriogenology 53, 342 (Abstr).
Petyim S, Båge R, Hallap T, Bergqvist A, Rodríegez-Martìnez H, Larsson B (2003) Two differentschemes of twice-weekly ovum pick-up in dairy heifers: Effect on oocyte recovery and ovarianfunction. Theriogenology 60, 175-188.
Phillips M, Grandin T, Graffam W, Irlbeck NA, Cambre RC (1998) Crate conditioning of bongo(Tragelaphus eurycerus) for veterinary and husbandry procedures at the Denver Zoological Gardens.Zoo Biol. 17, 25-32.
Picard K, Thomas DW, Festa-Bianchet M, Belleville F, Laneville A (1999) Differences in thethermal conductance of tropical and temperate bovid horns. Ecoscience 6, 148-158.
Pieterse MC, Kappen KA, Kruip TA, Taverne MA (1988) Aspiration of bovine oocytes duringtransvaginal ultrasound scanning of the ovaries. Theriogenology 30, 751-762.
Pieterse MC, Voss P, Kruip TA, Wurth YA, van Beneden TH, Willemse AH, Taverne MA (1991)Transvaginal ultrasound-guided follicular aspiration of bovine oocytes. Theriogenology 35, 857-862.
Pinyopummintr T, Bavister BD (1991) In vitro-matured/in vitro-fertilized bovine oocytes candevelop into morulae/blastocysts in chemically defined, protein-free culture media. Biol.Reprod. 45,736-742.
Pinyopummintr T, Bavister BD (1996) Effects of amino acids on development in vitro of cleavage-stage bovine embryos into blastocysts. Reprod.Fertil.Dev. 8, 835-841.
Platz CC, Wildt DE, Seager SW (1978) Pregnancy in the domestic cat after artificial inseminationwith previously frozen spermatozoa. J Reprod.Fertil. 52, 279-282.
Polejaeva IA, Chen SH, Vaught TD, Page RL, Mullins J, Ball S, Dai Y, Boone J, Walker S, AyaresDL, Colman A, Campbell KH (2000) Cloned pigs produced by nuclear transfer from adult somaticcells. Nature 407, 86-90.
Pope CE (2000) Embryo technology in conservation efforts for endangered felids. Theriogenology53, 163-174.
Pope CE, Dresser BL, Chin NW, Liu JH, Loskutoff NM, Behnke EJ, Brown C, McRae MA,Sinoway CE, Campbell MK, Cameron KN, Owens OM, Johnson CA, Evans RR, Cedars MI (1997)

146
Birth of a western lowland gorilla (Gorilla gorilla gorilla) following in vitro fertilization and embryotransfer. Am.J Primatol. 41, 247-260.
Pope CE, Gelwicks EJ, Burton M, Reece B, Dresser BL (1991) Nonsurgical embryo transfer in theScimitar-horned Oryx (Oryx dammah): Birth of a live offspring. Zoo Biol. 10, 43-51.
Pope CE, Gelwicks EJ, Wachs KB, Keller GL, Maruska EJ, Dresser BL (1989) Successfulinterspecies transfer of embryos from the Indian desert cat (Felis sylvestris ornata) to the domesticcat (Felis catus) following in vitro fertilization. Biol Reprod. Suppl 40, 61 (Abstr).
Pope CE, Gomez MC, Davis AM, Harris RF, Mikota SK, Boyd EH, Dresser BL (2001) Oocyteretrieval, in vitro fertilization and embryo tranfer in the caracal (Caracal caracal). Theriogenology55, 397 (Abstr.).
Pope CE, Gomez MC, Mikota S, Dresser BL (2000) Development of in vitro produced Africanwildcat (Felis silvestris) embryos after cryopreservation and transfer into domestic cat recipients.Biol Reprod. 62 (Suppl. 1): 321 (Abstr 544)
Pope CE, Johnson CA, McRae MA, Keller GL, Dresser BL (1998a) Development of embryosproduced by intracytoplasmic sperm injection of cat oocytes. Anim Reprod.Sci 53, 221-236.
Pope CE, Keller GL, Dresser BL (1993) In vitro fertilization in domestic and non-domestic catsincluding sequences of early nuclear events, development in vitro, cryopreservation and successfulintra- and interspecies embryo transfer. J Reprod.Fertil.Suppl 47, 189-201.
Pope CE, Lim J, Mikota S, Cochran R, Carter J, Godke R, Dresser B (1998b) Transvaginal oocyteretrieval and in vitro maturation, fertilization and culture in bongo antelope (Tragelaphus eurycerus).Biol.Reprod. 58 (Suppl 1), 102 (Abstr).
Pope CE, Loskutoff NM (1999) Embryo transfer and semen technology from cattle applied tonondomestic artiodactylids. In 'Zoo & Wild Animal Medicine: Current Therapy 4'. (Eds. ME Fowlerand RE Miller) pp. 597-604. (W.B. Saunders Co: Philadelphia, PA, USA)
Pope CE, McRae MA, Plair BL, Keller GL, Dresser BL (1994) Successful in vitro and in vivodevelopment of in vitro fertilized two to four-cell cat embryos following cryopreservation, cultureand transfer. Theriogenology 42, 513-525.
Pope CE, Pope VZ, Beck LR (1984) Live birth following cryopreservation and transfer of a baboonembryo. Fertil.Steril. 42, 143-145.
Prasolova LA, Jing S (1999) The coat color mutation in silver foxes (Vulpes vulpes): Morphology ofguard hairs. Genetika 35, 1269-1273.
Prather RS, Barnes FL, Sims MM, Robl JM, Eyestone WH, First NL (1987) Nuclear transplantationin the bovine embryo: Assessment of donor nuclei and recipient oocyte. Biol Reprod. 37, 859-866.
Prather RS, Sims MM, First NL (1989) Nuclear transplantation in early pig embryos. Biol Reprod.41, 414-418.

147
Price EO (2002) 'Animal Domestication and Behavior.' (CAB International: Wallingford,Oxfordshire, UK)
Ptak G, Clinton M, Barboni B, Muzzeddu M, Cappai P, Tischner M, Loi P (2002) Preservation of thewild European mouflon: The first example of genetic management using a complete program ofreproductive biotechnologies. Biol Reprod. 66, 796-801.
Pugh DG, Montes AJ (1994) Advanced reproductive technologies in South American camelids.Vet.Clin.North Am.Food Anim Pract. 10, 281-289.
Raghu HM, Nandi S, Reddy SM (2002) Follicle size and oocyte diameter in relation todevelopmental competence of buffalo oocytes in vitro. Reprod.Fertil.Dev. 14, 55-61.
Ralls K (1978) Tragelaphus eurycerus. Mammalian Species 111, 1-4.
Raphael BL, Loskutoff NM, Howard JG, Wolfe BA, Nemec LA, Schiewe MC, Kraemer DC (1989)Embryo transfer and artificial insemination in suni (Neotragus moschatus zuluensis). Theriogenology31, 244.
Raphael BL, Loskutoff NM, Huntress SL, Kraemer DC (1991) Postmortem recovery, in vitromaturation, and fertilization of klipspringer (Oreotragus oreotragus) ovarian oocytes. J Zoo.Wildl.Med. 22, 115-118.
Rath D, Johnson LA, Welch GR, Niemann H (1994) Successful gamete intrafallopian transfer(GIFT) in the porcine. Theriogenology 41, 1173-1179.
Read BW, Williams B, Christman J (1993) Restraint devices for management and researchprocedures for ungulates. Int.Zoo Yb. 32, 148-154.
Rieger D, Grisart B, Semple E, Van Langendonckt A, Betteridge KJ, Dessy F (1995) Comparison ofthe effects of oviductal cell co-culture and oviductal cell-conditioned medium on the developmentand metabolic activity of cattle embryos. J Reprod.Fertil. 105, 91-98.
Robeck TR (2001) Cetacean reproductive biology and ART. 1st Int.Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE, USA. pp. 179-185.
Robl JM, Prather R, Barnes F, Eyestone W, Northey D, Gilligan B, First NL (1987) Nucleartransplantation in bovine embryos. J Anim Sci 64, 642-647.
Rosenkrans CF, Jr., Zeng GQ, MCNamara GT, Schoff PK, First NL (1993) Development of bovineembryos in vitro as affected by energy substrates. Biol Reprod. 49, 459-462.
Roskoski R, Jr. (1996) 'Biochemistry.' (W.B. Saunders Company: Philadelphia)
Rott NN (1995) Organization of genetic cryobanks and use of developmental biology methods forpreservation of rare animals species I. Cryopreservation of sperm of wild animals. Ontogenez 26,270-281.

148
Rott NN (1996) The organization of genetic cryobanks and the use of developmental methods for theconservation of rare animals II: Obtaining and cryopreservation of the embryos of wild animals.Russian J Dev.Biol 27, 201-210.
SART-CDC (2003) '2000 Assisted Reproductive Technology Success Rates.'(http://www.cdc.gov/nccdphp/drh/art.htm., downloaded December 29, 2003) Society for AssistedReproductive Technologies (SART) and Center for Disease Control (CDC), Atlanta, GA, USA.
SAS. 2001. SAS Software Release 8.2 (TS2M0). SAS Institute, Inc., Cary, NC, USA.,
Schiewe MC, Bush M, De VV, Brown JL, Wildt DE (1991a) Semen characteristics, sperm freezing,and endocrine profiles in free-living wildebeest (Connochaetes taurinus) and greater kudu(Tragelaphus strepsiceros). J Zoo.Wildl.Med. 22, 58-72.
Schiewe MC, Bush M, Phillips LG, Citino S, Wildt DE (1991b) Comparative aspects of estrussynchronization, ovulation induction, and embryo cryopreservation in the scimitar-horned oryx,bongo, eland and greater kudu. J Exp.Zool. 258, 75-88.
Schiewe MC, Bush MC, Phillips LG, Wildt DE (1988) Variables influencing the collection andcryopreservation of embryos from nondomestic hoofed species. 11th Int.Cong.Anim.Reprod.Artif.Insemin. Vol. 2, Dublin, Ireland. P. 190
Schmid, R., Lanterna, J., Garcia, A., Tomsett, G., and Loskutoff, N (1997). A comparison of standardbovine and equine cryodiluents for cryopreserving epididymal sperm from greater kudu(Tragelaphus strepsiceros). Theriogenology 47, 407 (Abstr).
Schmitt DL, Hildebrandt TB (1998) Manual collection and characterization of semen from Asianelephants (Elephas maximus). Anim Reprod.Sci 53, 309-314.
Schmitt DL, Hildebrandt TB, Hermes R, Göritz F (2001) Assisted reproductive technology inelephants. 1st Int.Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE, USA. pp.32-34.
Schramm RD, Bavister BD (1996) Development of in-vitro-fertilized primate embryos intoblastocysts in a chemically defined, protein-free culture medium. Hum.Reprod. 11, 1690-1697.
Schriver MD, Kraemer DC (1978) Embryo transfer in the domestic cat. Am.Assoc.Lab.Anim Sci 78,12.
Schwarzenberger F, Rietschel W, Matern B, Schaftenaar W, Bircher P, Van Puijenbroeck B, Leus K(1999) Noninvasive reproductive monitoring in the okapi (Okapia johnstoni). J Zoo Wildl.Med. 30,497-503.
Seshagiri PB, Bavister BD (1991) Glucose and phosphate inhibit respiration and oxidativemetabolism in cultured hamster eight-cell embryos: Evidence for the "Crabtree effect".Mol.Reprod.Dev. 30, 105-111.
Shin T, Kraemer D, Pryor J, Liu L, Rugila J, Howe L, Buck S, Murphy K, Lyons L, Westhusin M(2002) A cat cloned by nuclear transplantation. Nature 415, 859.

149
Sillero-Zubiri C, Mcdonald DW, IUCN/SSC Specialist Group (1997) 'The Ethiopian Wolf- StatusSurvey and Conservation Action Plan.' (IUCN: Gland, Switzerland)
Sirard MA, Picard L, Dery M, Coenen K, Blondin P (1999) The time interval between FSHadministration and ovarian aspiration influences the development of cattle oocytes. Theriogenology51, 699-708.
Skidmore JA, Billah M, Short RV, Allen WR (2001) Assisted reproductive techniques forhybridization of camelids. Reprod.Fertil.Dev. 13, 647-652.
Skinner JD, Moss DG, Skinner DC (2002) Inherent seasonality in the breeding seasons of Africanmammals: Evidence from captive breeding. Trans.R.Soc.S.Afr. 57, 25-34.
Solti L, Crichton EG, Loskutoff NM, Cseh S (2000) Economical and ecological importance ofindigenous livestock and the application of assisted reproduction to their preservation.Theriogenology 53, 149-162.
Songsasen N, Betteridge KJ, Leibo SP (1997) Birth of live mice resulting from oocytes fertilized invitro with cryopreserved spermatozoa. Biol Reprod. 56, 143-152.
Spinage CA (1986) 'The Natural History of Antelopes.' (Facts on File Publications: New York)
Squires EL, Carnevale EM, McCue PM, Bruemmer JE (2003) Embryo technologies in the horse.Theriogenology 59, 151-170.
Squires EL, Wilson JM, Kato H, Blaszczyk A (1996) Pregnancy after intracytoplasmic sperminjection into equine oocytes matured in vitro. Theriogenology 45, 305 (Abstr).
Steeves TE, Gardner DK (1999) Temporal and differential effects of amino acids on bovine embryodevelopment in culture. Biol Reprod. 61, 731-740.
Steptoe PC, Edwards RG (1978) Birth after the reimplantation of a human embryo. Lancet 2, 366.
Stevens VJ (1978) Basic operant conditioning research in the zoo. In 'Behavior of Captive WildAnimals'. (Eds. H Markowitz and VJ Stevens) pp. 209-305. (Nelson-Hall: Chicago, IL, USA)
Stewart DL (1951) Storage of bull spermatozoa at low temperatures. Vet.Rec. 63, 65-66.
Stice SL, Robl JM (1988) Nuclear reprogramming in nuclear transplant rabbit embryos. Biol Reprod.39, 657-664.
Stover J, Evans J, Dolensk EP (1981) Interspecies embryo transfer from gaur to domestic Holstein.Proc.Am Assoc.Zoo Vet pp. 122-124.
Summers MC, Bhatnagar PR, Lawitts JA, Biggers JD (1995) Fertilization in vitro of mouse ova frominbred and outbred strains: Complete preimplantation embryo development in glucose-supplementedKSOM. Biol Reprod. 53, 431-437.

150
Susko-Parrish JL, Wheeler MB, Ax RL, First NL, Parrish JJ (1990) The effect of penicillamine,hypotuarine, epinephrine and sodium metabisulfite, on bovine in vitro fertilization. Theriogenology33, 333 (Abstr).
Suzuki T, Singla SK, Sujata J, Madan ML (1992) In vitro fertilization of water buffalo follicularoocytes and their ability to cleave in vitro. Theriogenology 38, 1187-1194.
Swanson WF (2001) Reproductive biotechnology and conservation of the forgotten felids - the smallcats. pp. 1st Int.Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE, USA. Pp. 99-119.
Takahashi Y, First NL (1992) In vitro development of bovine one-cell embryos: Influence ofglucose, lactate, pyruvate, amino acids and vitamins. Theriogenology 37, 963-978.
Takedomi T, Aoyagi Y, Konishi M, Kishi H, Taya K, Watanabe G, Sasamoto S (1995)Superovulation of holstein heifers by a single subcutaneous injection of FSH dissolved inpolyvinylpyrrolidone. Theriogenology 43, 1259-1268.
Taylor PJ, Taylor S, James AN, Denniston RS, Godke RA (2001) Alpaca offspring born after crossspecies embryo transfer to llama. Theriogenology 55, 401 (Abstr).
Thompson JG (1996) Defining the requirements for bovine embryo culture. Theriogenology 45, 27-40.
Thompson JG (2000) In vitro culture and embryo metabolism of cattle and sheep embryos - Adecade of achievement. Anim Reprod.Sci 60-61, 263-275.
Thompson JG, McNaughton C, Gasparrini B, McGowan LT, Tervit HR (2000) Effect of inhibitorsand uncouplers of oxidative phosphorylation during compaction and blastulation of bovine embryoscultured in vitro. J Reprod.Fertil. 118, 47-55.
Thompson JG, Simpson AC, Pugh PA, Tervit HR (1992) Requirement for glucose during in vitroculture of sheep preimplantation embryos. Mol.Reprod.Dev. 31, 253-257.
Thouas GA, Korfiatis NA, French AJ, Jones GM, Trounson AO (2001) Simplified technique fordifferential staining of inner cell mass and trophectoderm cells of mouse and bovine blastocysts.Reprod.BioMed.Online 3, 25-29.
Tollner TL, VandeVoort CA, Overstreet JW, Drobnis EZ (1990) Cryopreservation of spermatozoafrom cynomolgus monkeys (Macaca fascicularis). J Reprod.Fertil. 90, 347-352.
Toyoda Y, Chang MC (1974) Fertilization of rat eggs in vitro by epididymal spermatozoa and thedevelopment of eggs following transfer. J Reprod.Fertil. 36, 9-22.
Toyooka Y, Tsunekawa N, Akasu R, Noce T (2003) Embryonic stem cells can form germ cells invitro. Proc.Natl.Acad.Sci U.S A 100, 11457-11462.
Traldi AS, Leboeuf B, Cognié Y, Poulin N, Mermillod P (1999) Comparative results of in vitro andin vivo survival of vitrified in vitro produced goat and sheep embryos. Theriogenology 51, 175.

151
Treus VD, Lobanov NV (1971) Acclimatization and domestication of the eland Taurotragus oryx atAskanya-Nova Zoo. Int.Zoo Yearb. 11, 147-156.
Trut LN, Pliusnina IZ, Prasolova LA (2000) Mutations hooded and nonagouti in the gray rat (Rattusnorvegicus): Effect of selection on behavior and photoperiod. Genetika 36, 813-822.
Tyndale-Biscoe CH (1963) Blastocyst transfer in the marsupial Setonix brachyurus. J Reprod.Fertil.6, 41-48.
Tyndale-Biscoe CH (1970) Resumption of development by quiescent blastocysts transferred toprimed, ovariectomized recipients in the marsupial Macropus eugenii. J Reprod.Fertil. 23, 25-32.
Vajta G, Holm P, Kuwayama M, Booth PJ, Jacobsen H, Greve T, Callesen H (1998) Open pulledstraw (OPS) vitrification: A new way to reduce cryoinjuries of bovine ova and embryos.Mol.Reprod.Dev. 51, 53-58.
van Wagtendonk-de Leeuw AM, Mullaart E, de Roos AP, Merton JS, den Daas JH, Kemp B, deRuigh L (2000) Effects of different reproduction techniques: AI MOET or IVP, on health andwelfare of bovine offspring. Theriogenology 53, 575-597.
Van Winkle LJ, Campione AL (1996) Amino acid transport regulation in preimplantation mouseembryos: Effects on amino acid content and pre- and post-implantation development.Theriogenology 45, 69-80.
Wakayama T, Perry AC, Zuccotti M, Johnson KR, Yanagimachi R (1998) Full-term development ofmice from enucleated oocytes injected with cumulus cell nuclei. Nature 394, 369-374.
Ward WS, Kimura Y, Yanagimachi R (1999) An intact sperm nuclear matrix may be necessary forthe mouse paternal genome to participate in embryonic development. Biol Reprod. 60, 702-706.
Wassarman PM, Albertini DF (1994) The mammalian ovum. In 'The Physiology of Reproduction'.(Eds. E Knobil and JD Neill). pp. 79-122. (Raven Press, Ltd.: New York)
Watson PF, Holt WV (2001) 'Cryobanking the Genetic Resource: Wildlife Conservation for theFuture.' (Taylor & Francis: London)
Werner-von der BW, Coordes I, Hatzmann W (1993) Pregnancy following intrafollicular gametetransfer. Hum.Reprod. 8, 771-774.
Whitten WK, Biggers JD (1968) Complete development in vitro of the pre-implantation stages of themouse in a simple chemically defined medium. J Reprod.Fertil. 17, 399-401.
Whittingham DG (1968) Fertilization of mouse eggs in vitro. Nature 220, 592-593.
Whittingham DG (1977) Fertilization in vitro and development to term of unfertilized mouse oocytespreviously stored at -196 degrees C. J Reprod.Fertil. 49, 89-94.
Whittingham DG, Leibo SP, Mazur P (1972) Survival of mouse embryos frozen to -196 degrees and-269 degrees C. Science 178, 411-414.

152
Wiepz DW, Chapman RJ (1985) Non-surgical embryo transfer and live birth in a llama.Theriogenology 24, 251-256.
Wildt DE, Monfort SL, Donoghue AM, Johnson LA, Howard JG (1992) Embryogenesis inconservation biology –– Or, how to make an endangered species embryo. Theriogenology 37, 167-184.
Wildt DE, Rall WF, Crister JK, Monfort SL, Seal US (1997) Genome resource banks: Livingcollections for biodiversity conservation. BioScience 47, 689-698.
Willadsen SM (1986) Nuclear transplantation in sheep embryos. Nature 320, 63-65.
Willard ST, Flores-Foxworth G, Chapman S, Drew ML, Hughes DM, Neuendorff DA, Randel RD(1998) Hybridization between wapiti (Cervus elephus manitobensis) and sika deer (Cervus nippon):A comparison of two artificial insemination techniques. J Zoo Wildl.Med. 29, 295-299.
Willett EL, Black WG, Casida LE, Stone WH, Buckner PJ (1951) Successful transplantation of afertilized bovine ovum. Science 113, 247.
Wilmut I, Beaujean N, De Sousa PA, Dinnyes A, King TJ, Paterson LA, Wells DN, Young LE(2002) Somatic cell nuclear transfer. Nature 419, 583-586.
Wilmut I, Rowson LE (1973) The successful low-temperature preservation of mouse and cowembryos. J Reprod.Fertil. 33, 352-353.
Wilmut I, Schnieke AE, McWhir J, Kind AJ, Campbell KH (1997) Viable offspring derived fromfetal and adult mammalian cells. Nature 385, 810-813.
Winger Q, Damiani P, Loskutoff NM (1997) The application of standard bovine protocols for thematuration and fertilization of blesbok (Damaliscus dorcas phillipsi) oocytes using epididymal spermcryopreserved in glycerol or dimethyl sulfoxide. Theriogenology 47, 412 (Abstr).
Wirtu G, Bavister B, Andrews J, Perez O, Short C, Pope E (2002a) Transvaginal ultrasound-guidedoocyte retrieval in sedated eland (Taurotragus oryx) and developmental potential of the oocytes.Theriogenology 57, 595 (Abstr).
Wirtu G, Damiani P, Bavister B, Miller F, Dresser B, Godke RA, Pope E (2003) Effects of eland andbovine serum on the development of in vitro-derived bovine embryos. Theriogenology 59, 354(Abstr).
Wirtu G, Pope CE, Damiani P, Dresser BL, Godke RA, Bavister BD (2004) Insulin improvesfrequency of cleavage and in vitro development of bovine embryos in defined medium.Reprod.Fertil.Dev. 16, 203 (Abstr).
Wirtu G, Pope CE, Lee BC, Cole A, Vaccaro J, Sarrat E, Bavister BD, Godke RA, Dresser BL(2002b) Collection of semen by rectal massage from an eland (Taurotragus oryx) bull.2nd Int. Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE, USA. pp. 318-320.
Wirtu G, Pope CE, Vaccaro J, Sarrat E, Cole A, Godke RA, Dresser BL. Dominance hierarchy in aherd of female eland antelope (Taurotragus oryx) in captivity. Zoo Biol In press.

153
Wolf DP, VandeVoort CA, Meyer-Haas GR, Zelinski-Wooten MB, Hess DL, Baughman WL,Stouffer RL (1989) In vitro fertilization and embryo transfer in the rhesus monkey. Biol Reprod. 41,335-346.
Woods GL, White KL, Vanderwall DK, Li GP, Aston KI, Bunch TD, Meerdo LN, Pate BJ (2003) AMule cloned from fetal cells by nuclear transfer. Science, 301, 1063.
Wrenzycki C, Herrmann D, Carnwath JW, Niemann H (1999) Alterations in the relative abundanceof gene transcripts in preimplantation bovine embryos cultured in medium supplemented with eitherserum or PVA. Mol.Reprod.Dev. 53, 8-18.
Wrenzycki C, Herrmann D, Keskintepe L, Martins A, Jr., Sirisathien S, Brackett B, Niemann H(2001) Effects of culture system and protein supplementation on mRNA expression in pre-implantation bovine embryos. Hum.Reprod. 16, 893-901.
Yamamoto M, Ooe M, Kawaguchi M, Suzuki T (1994) Superovulation in the cow with a singleintramuscular injection of FSH dissolved in polyvinylpyrrolidone. Theriogenology 41, 747-755.
Yamamoto Y, Oguri N, Tsutsumi Y, Hachinohe Y (1982) Experiments in freezing and storage ofequine embryos. J Reprod.Fertil.Suppl 32, 399-403.
Yamashita S, Abe H, Itoh T, Satoh T, Hoshi H (1999) A serum-free culture system for efficient invitro production of bovine blastocysts with improved viability after freezing and thawing.Cytotechnology 31, 121-129.
Yamauchi Y, Yanagimachi R, Horiuchi T (2002) Full-term development of golden hamster oocytesfollowing intracytoplasmic sperm head injection. Biol Reprod. 67, 534-539.
Yang BK, Giles JR, Yang X, Foote RH (1995) Development of in vitro matured/in vitro fertilizedbovine oocytes in a simple defined (KSOM) Medium. J Reprod.Dev. 41, 213-218.
Yang X, Jiang S, Foote RH (1993) Bovine oocyte development following different oocyte maturationand sperm capacitation procedures. Mol.Reprod.Dev. 34, 94-100.
Yong Z, Jianchen W, Jufen Q, Zhiming H (1991) Nuclear transplantation in goats. Theriogenology35, 299 (Abstr).
Yoshioka K, Suzuki C, Tanaka A, Anas IM, Iwamura S (2002) Birth of piglets derived from porcinezygotes cultured in a chemically defined medium. Biol Reprod. 66, 112-119.
Yoshioka K, Takahashi Y, Hishinuma M, Kanagawa H (1993) In vitro culture of bovine one-cellembryos derived from in vitro fertilization using a semi-chemically defined medium. J Vet.Med.Sci55, 901-904.
Young LE, Sinclair KD, Wilmut I (1998) Large offspring syndrome in cattle and sheep. Rev.Reprod.3, 155-163.

154
Zeron Y, Ocheretny A, Kedar O, Borochov A, Sklan D, Arav A (2001) Seasonal changes in bovinefertility: Relation to developmental competence of oocytes, membrane properties and fatty acidcomposition of follicles. Reproduction 121, 447-454.
Zhang L, Barry DM, Denniston RS, Bunch TD, Godke RA (1993) Birth of live calves after transferof frozen-thawed bovine embryos fertilised in vitro. Vet.Rec. 132, 247-249.
Zheng P, Wang H, Bavister BD, Ji W (2001) Maturation of rhesus monkey oocytes in chemicallydefined culture media and their functional assessment by IVF and embryo development. Hum.Reprod. 16, 300-305.
Zhou Q, Renard JP, Le Friec G, Brochard V, Beaujean N, Cherifi Y, Fraichard A, Cozzi J (2003)Generation of fertile cloned rats by regulating oocyte activation. Science 302, 1179.

155
Appendix A: Attributes of Species of Tragelaphine Antelopes
Species/common nameGestationlength, dd
Chromosomes, 2nmale : female
Adult femaleweight, kg
Age atpuberty, moc
Estrous cyclelength, dd
Estrusduration, dd
Populationstatus (IUCN)e
Tragelaphus eurycerus,
Bongo
285 [1;2]a 33:34 [3] 200 [4]
240 [5]
≤27 [2] 21-22 [2;4] 3 d [2;4];
29 hr [6]
LR/NT
T. strepsiceros,
Greater kudu
~240 [2;4]
~270 [5]
31:32 [7] 170 [5] 17 [2] LR/CD
T. imberbis,
Lesser kudu
~210 [5];
244-253 [2]
38 [8] 60 [5] 15 [2] LR/ CD
T. spekei,
Sitatunga
~225 [5];
247 [2]
30 [9] 55 [5] 12 [2] LR/NT
T. scriptus,
Bushbuck
~225 [4];
180 [2;5];
165 [5]
33:34 [10] 30 [5] 10 [2] LR/CD
T. angasi,
Lowland nyala
220 [2] 55:56
see [8]
55-127 [2] 20 [2] 10-30 [2] 2-3 [2] LR/CD
T. buxtoni,
Mountain nyala
~200 [11] EN
Taurotragus oryx,
Common eland
255-284
[4;5;12;13]
31:32 [14;15] 450 [5];
477 [12];
356b
≤28 [12];
≤32 [13]
23 [16],
21-26 [2]
LR/CD
T. derbianus, Giant eland 440 [5] LR/NTaFigures in brackets, [ ], indicate the reference number; bOur observation; cmonths; ddays; eLower risk (LR), Near threatened (NT),Conservation dependent (CD), Endangered (EN).

156
References for Appendix A
[1] Dresser BL, Pope CE, Kramer L, Kuehn G, Dahlhausen RD, Maruska EJ, Reece B, ThomasWD (1985) Birth of bongo antelope (Tragelaphus euryceros) to eland antelope (Tragelaphusoryx) and cryopreservation of bongo antelopes. Theriogenology 23, 190 (Abstr).
[2] Nowak RM (1999). “Walker's Mammals of the World”: Volume II, 6th ed. (The John HopkinsUniversity Press, Baltimore and London). pp. 1135-1145.
[3] Wurster DH (1972) Sex-chromosome translocations and karyotypes in bovid tribes.Cytogenetics 11, 197-207.
[4] Crandall LS (1964) “Management of Wild Mammals in Captivity” (The University of ChicagoPress, Chicago, IL, USA).
[5] Kingdon J (1982) East African Mammals: An Atlas of Evolution in Africa Volume III part C(Bovids). (Academic Press New York, NY, USA).
[6] Pope CE (2001) Antelope reproductive biology and ART. 1st Int.Symp.Assist.Reprod.Technol.Conserv.Genet.Mgmt.Wildl., Omaha, NE, USA.. pp. 49-53.
[7] Wallace C, Fairall N (1967). The chromosomes of the kudu. S.Afr.J Med.Sci 32, 61.
[8] Benirschke K, Ruedi D, Muller H, Kumamoto AT, Wagner KL (1980) The unusual karyotypeof the lesser kudu, Tragelaphus imberbis. Cytogenet.Cell Genet.26, 85-92.
[9] Wurster DH, Benirschke K, Noelke H (1968) Unusually large sex chromosomes in thesitatunga (Tragelaphus spekei) and the blackbuck (Antilope cervicapra). Chromosoma; 23,317-323.
[10] Wallace C (1977) Chromosome analysis in the Kruger National Park: The chromosomes of thebushbuck (Tragelaphus scriptus). Cytogenet.Cell Genet. 18, 50-56.
[11] CAMERAPIX (1989) ‘Spectrum Guide to African Wildlife Safaris’. (Facts on File, New York,NY, USA).
[12] Treus VD, Lobanov NV (1971). Acclimatization and domestication of the eland Taurotragusoryx at Askanya-Nova Zoo. Int.Zoo Yearb. 11, 147-156.
[13] Lydon SA (2001) North American Regional Studbook: Common eland, Taurotragus oryx.Boston, MA, USA. pp.5-29.
[14] Koulischer L (1969) Concept of cellular clonal evolution of karyotypes applied to evolution ofspecies. Acta Zool.Pathol.Antverp. 48, 21-28.
[15] Taylor KM, Taylor BK (1970). The chromosome of the eland Taurotragus oryx Pallas.Mammal.Chrom.Newsl. 11, 123-125.
[16] Atkinson MW, Welsh TH, Blumer ES (1999). Evaluation of an advanced handling system forphysiological data collection, testing and medical treatment of large, nondomestic hoofstock.Proc.Am.Assoc.Zoo Vet, Columbus, OH, USA.: pp. 154-157.

157
Appendix B: Abbreviations
aa = amino acids
AI = Artificial insemination
ART = Assisted reproductive technology
BCS = Bovine calf serum
BM = Basic medium
BME = Basal medium Eagle
BSA = Bovine serum albumin
CPK = Creatine phosphokinase
d = day(s)
EGF = Epidermal growth factor
ET = Embryo transfer
FCS = Fetal calf serum
FSH = Follicle stimulating hormone
GIFT = Gamete intrafallopian transfer
GV = Germinal vesicle
HECM = Hamster embryo culture medium
hr = Hour(s)
i.m. = Intramuscular (injection)
i.v. = Intravenous (injection)
ICSI = Intracytoplasmic sperm injection
iET = interspecies embryo transfer
iSCNT = interspecies somatic cell nuclear transfer
ISIS = International Species Information System
IUCN = International Union for Conservation of Nature and Natural Resources
IVC = In vitro culture of embryos
IVF = In vitro fertilization
IVM = In vitro maturation of oocytes

158
IVP = In vitro production (of embryos)
KSOM = Potassium simplex optimized medium
LH = Luteinizing hormone
m (e.g., 56 m) = Meter
m (e.g., mKSOM) = modified
MI or MII = metaphase I or II stage oocyte
Medarks = Medical Animal Records Keeping Systems
MEM = Minimal essential medium
min = Minute(s)
mo = Month(s)
mol wt = Molecular weight
PBS = Phosphate buffered saline
PCR = Polymerase chain reaction
PHE = Penicillamine-hypotaurine-epinephrine
PVA = Polyvinyl alcohol
PVP = Polyvinyl pyrrolidone
SART = Society for Assisted Reproductive Technology
SCNT = Somatic cell nuclear transfer
sec = Second
TALP = Tyrode’s solution containing bovine serum albumin, lactic acid and pyruvic acid
TCM = Tissue culture medium
TL-HEPES-PVA = Tyrode’s solution supplemented with lactic acid, HEPES and PVA
yr = Year

159
Vita
Gemechu Wirtu was born in Ethiopia in 1968. He is the son of Wirtu Gerba Amba and
Degetie Waqjira Nono. He grew up in rural Ethiopia. In 1992, he graduated in veterinary medicine
from Addis Ababa University. Subsequently, he was employed by the same university where he
taught courses to veterinary students. He also held different administrative positions and was
involved in several research projects. In 1997, he won the Fulbright Scholarship to complete two
years of graduate studies in the USA and received a Master of Science degree from Virginia
Polytechnic Institute and State University in 1999. His thesis research work focused on xenogenous
transfer of equine gametes and was conducted at the Virginia-Maryland Regional College of
Veterinary Medicine.
In Fall 1999, Gemechu was accepted as a doctoral student through a joint program between
Louisiana State University School of Veterinary Medicine and Audubon Institute Center for
Research of Endangered Species. His dissertation focused on the development of methods of gamete
collection in the eland antelope and in vitro production of embryos in cattle and eland. Drs. Barry
Bavister, Robert Godke, Earle Pope and Charles Short supervised his doctoral research. Gemechu
will receive his doctoral degree in May 2004.
He is married to Enkutatash Tadesse. Together, they are parents of Fidu, Milki and Kulani.
Copyright © 2022 FDOKUMEN




















