
Xenogeneic immunization in mice using HER2 DNA delivered by an adenoviral vector
11
Xenogeneic Immunization in Mice Using HER2 DNA Delivered by an Adenoviral Vector Pasquale Gallo 1 , Sridhar Dharmapuri 1 , Maurizio Nuzzo 1 , Daniele Maldini 1 , Manuela Iezzi 2 , Federica Cavallo 3 , Piero Musiani 2 , Guido Forni 3 and Paolo Monaci 1 * 1 Molecular and Cell Biology Department, Istituto di Ricerche di Biologia Melecolare P. Angeletti, Pomezia, Italy 2 Aging Research Center, “G. d’Annunzio” Foundation, University of Chieti, Chieti, Italy 3 Department of Clinical and Biologic Sciences, University of Turin, Orbassano, Italy The protective efficacy of xenogeneic vaccination with DNA en- coding the HER2 oncogene was evaluated in BALB/c mice trans- genic for the transforming form of the neu oncogene, which spon- taneously develops carcinomas in all mammary glands. Intramuscular injection of either plasmid DNA followed by elec- trical stimulation (pVIJ-HER2 with ES) or an adenoviral vector (Ad5-HER2), both expressing the HER2 oncogene, was tested. Immunization using pVIJ-HER2 with ES elicited a cell-mediated response that was much lower than that elicited by the immuni- zation with Ad5-HER2, as measured by the frequency of IFN-- secreting spleen cells. The dominant T-cell epitope of the HER2 protein product (p185) in the BALB/c (H-2 d ) genetic background was identified. While the T-cell response elicited was only partially crossreactive with the corresponding rat epitopes because of se- quence variations (89% similarity), a cytotoxic T-lymphocyte ac- tivity against the rat immunodominant epitope was also evident. The Ad5-HER2 vaccination induced also antibodies against p185, which crossreacted with the rat protein homolog. Both T- and B-cell responses slowly declined with time. Vaccination with Ad5- HER2 at 6 and 9 weeks of age delayed incidence and reduced multiplicity of tumors in neu transgenic mice. © 2004 Wiley-Liss, Inc. Key words: antitumor vaccination; breast cancer; adenovirus; neu transgenic mice An increasing number of reports underline the critical role played by the cell-mediated immune (CMI) response to control tumor growth and spread of metastases. 1,2 This has been clearly demonstrated in mice studies where depletion of T cells correlated with the loss of tumor protection. 3 These findings prompted the development of genetic vaccine strategies where the gene coding for the target antigen is expressed intracellularly, thus having the potential of inducing a specific T-cell response. As the cell ma- chinery ensures correct folding and posttranslational modifica- tions, these approaches can induce antibodies against native anti- gen structures. Gene-based immunization protocols can be grouped into 2 main categories according to the gene delivery vehicle adopted. The first class includes nonviral gene delivery by plasmid DNA expressing the target antigen. The relatively low efficiency of naked DNA immunization has promoted the development of several formula- tions to enhance its immunogenic potential (e.g., electrical stimu- lation, microbeads, adjuvants, costimulatory proteins). 4 A second class of genetic immunization approaches uses a variety of viral vectors, including modified vaccinia virus ankara, canarypox and adenovirus, which have been tested as antigen delivery vehicles with encouraging results. 5 p185, the protein encoded by the HER2 oncogene, is a member of the epidermal growth factor receptor family with tyrosine kinase activity. 6 Several elements make this antigen an attractive target for active specific immunotherapy. 7 p185 is overexpressed in about 30 –50% of human breast and in other types of carcinomas. This receptor has a high oncogenic potential and its elevated expression in tumors correlates with tumor aggressiveness. By contrast, only very low level expression can be detected in normal adult tissues. p185 is a self-antigen that plays an important role during em- bryogenesis, though some tumor patients do develop specific an- tibodies and cellular immune response to this receptor. Thus, vaccine strategies directed against p185 as well as other tumor antigens must overcome immunologic tolerance to self-proteins. Xenogeneic immunization uses a nonself heterologous protein as an immunogen to induce a crossreactive response against the self-antigen (target) through shared epitopes. It has proven to be a valuable strategy for breaking tolerance against tumor antigens. 8 As an example, DNA vaccination against the human melanoma antigen gp100 has been shown to induce crossreactive CTL re- sponses against the murine homolog of gp100. 9 In this work, we investigated the efficiency of a xenogeneic vaccination in mice using an adenoviral vector expressing p185. Inbred BALB/c mice engineered to overexpress the activated (G664E) neu oncogene under the control of the mouse mammary tumor virus promoter (MMTV-LTR; from now on referred to as neuT mice) were used as experimental model system. 10 In neuT mice, the transgene is prevalently expressed in mammary glands. At 3 weeks of age, female neuT mice start a process of rapid development of tumors involving all the mammary glands. p185 is a xenogeneic protein in neuT mice since it differs in more than 11% of the amino acid residues when compared to both rat (r-p185) and endogenous mouse proteins. 11 The goals of this study were to assess the efficiency of adeno- virus vs. plasmid DNA-mediated immunization by quantitatively measuring the humoral and the cell-mediated immune-response; to evaluate the efficiency of xenogeneic immunization in breaking tolerance in the transgenic mouse model and to verify the impact of Ad5-HER2-induced immunity on mammary tumor develop- ment in neuT mice. Material and methods Cell lines Mouse liver NMuLi and target mouse P815 plasmacytoma cells were obtained from the American Type Culture Collection (ATCC, Rockville, MD), Per.C6 cells were from Crucell (Leiden, The Netherlands) and mouse muscle myoblasts C2C7 from G. Abbreviations: A, absorbance; Ad5, adenovirus 5; AP, alkaline phos- phatase; CMI, cell-mediated immune; ECD, extracellular domain; ES, electrical stimulation; HER2, human epidermal growth factor receptor 2; ICS, intracellular staining; MCF, mean channel fluorescence; MMTV- LTR, mouse mammary tumor virus long terminal repeat; MOI, multiplicity of infection; neu, rat homolog of HER2; p185, protein encoded by HER2; PCNA, proliferating cell nuclear antigen; r-p185, protein encoded by neu; SEB, staphylococcus enterotoxin B; SFC, spot forming cell; vp, viral particle. Grant sponsor: the Italian Association for Cancer Research; Grant spon- sor: the Italian Ministry for University; Grant sponsor: the Center of Excellence on Aging at the University of Chieti; Grant sponsor: Italian Ministero dell’Istruzione, dell’Universita ` e della Ricerca; Grant number: FIRB RBME017BC4. *Correspondence to: Molecular and Cell Biology Department, I.R.B.M. P. Angeletti, Via Pontina km 30.6, 00040, Pomezia (Roma), Italy. Fax: 39-06-91093-654. E-mail: [email protected] Received 22 October 2003; Accepted after revision 9 June 2004 DOI 10.1002/ijc.20536 Publishedonline25August2004inWileyInterScience(www.interscience. wiley.com). Int. J. Cancer: 113, 67–77 (2005) © 2004 Wiley-Liss, Inc. Publication of the International Union Against Cancer
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Xenogeneic immunization in mice using HER2 DNA delivered by an adenoviral vector

Xenogeneic Immunization in Mice Using HER2 DNA Delivered by anAdenoviral VectorPasquale Gallo1, Sridhar Dharmapuri1, Maurizio Nuzzo1, Daniele Maldini1, Manuela Iezzi2, Federica Cavallo3,Piero Musiani2, Guido Forni3 and Paolo Monaci1*1Molecular and Cell Biology Department, Istituto di Ricerche di Biologia Melecolare P. Angeletti, Pomezia, Italy2Aging Research Center, “G. d’Annunzio” Foundation, University of Chieti, Chieti, Italy3Department of Clinical and Biologic Sciences, University of Turin, Orbassano, Italy
The protective efficacy of xenogeneic vaccination with DNA en-coding the HER2 oncogene was evaluated in BALB/c mice trans-genic for the transforming form of the neu oncogene, which spon-taneously develops carcinomas in all mammary glands.Intramuscular injection of either plasmid DNA followed by elec-trical stimulation (pVIJ-HER2 with ES) or an adenoviral vector(Ad5-HER2), both expressing the HER2 oncogene, was tested.Immunization using pVIJ-HER2 with ES elicited a cell-mediatedresponse that was much lower than that elicited by the immuni-zation with Ad5-HER2, as measured by the frequency of IFN-�-secreting spleen cells. The dominant T-cell epitope of the HER2protein product (p185) in the BALB/c (H-2d) genetic backgroundwas identified. While the T-cell response elicited was only partiallycrossreactive with the corresponding rat epitopes because of se-quence variations (89% similarity), a cytotoxic T-lymphocyte ac-tivity against the rat immunodominant epitope was also evident.The Ad5-HER2 vaccination induced also antibodies against p185,which crossreacted with the rat protein homolog. Both T- andB-cell responses slowly declined with time. Vaccination with Ad5-HER2 at 6 and 9 weeks of age delayed incidence and reducedmultiplicity of tumors in neu transgenic mice.© 2004 Wiley-Liss, Inc.
Key words: antitumor vaccination; breast cancer; adenovirus; neutransgenic mice
An increasing number of reports underline the critical roleplayed by the cell-mediated immune (CMI) response to controltumor growth and spread of metastases.1,2 This has been clearlydemonstrated in mice studies where depletion of T cells correlatedwith the loss of tumor protection.3 These findings prompted thedevelopment of genetic vaccine strategies where the gene codingfor the target antigen is expressed intracellularly, thus having thepotential of inducing a specific T-cell response. As the cell ma-chinery ensures correct folding and posttranslational modifica-tions, these approaches can induce antibodies against native anti-gen structures.
Gene-based immunization protocols can be grouped into 2 maincategories according to the gene delivery vehicle adopted. The firstclass includes nonviral gene delivery by plasmid DNA expressingthe target antigen. The relatively low efficiency of naked DNAimmunization has promoted the development of several formula-tions to enhance its immunogenic potential (e.g., electrical stimu-lation, microbeads, adjuvants, costimulatory proteins).4 A secondclass of genetic immunization approaches uses a variety of viralvectors, including modified vaccinia virus ankara, canarypox andadenovirus, which have been tested as antigen delivery vehicleswith encouraging results.5
p185, the protein encoded by the HER2 oncogene, is a memberof the epidermal growth factor receptor family with tyrosine kinaseactivity.6 Several elements make this antigen an attractive targetfor active specific immunotherapy.7 p185 is overexpressed inabout 30–50% of human breast and in other types of carcinomas.This receptor has a high oncogenic potential and its elevatedexpression in tumors correlates with tumor aggressiveness. Bycontrast, only very low level expression can be detected in normaladult tissues.
p185 is a self-antigen that plays an important role during em-bryogenesis, though some tumor patients do develop specific an-
tibodies and cellular immune response to this receptor. Thus,vaccine strategies directed against p185 as well as other tumorantigens must overcome immunologic tolerance to self-proteins.Xenogeneic immunization uses a nonself heterologous protein asan immunogen to induce a crossreactive response against theself-antigen (target) through shared epitopes. It has proven to be avaluable strategy for breaking tolerance against tumor antigens.8As an example, DNA vaccination against the human melanomaantigen gp100 has been shown to induce crossreactive CTL re-sponses against the murine homolog of gp100.9
In this work, we investigated the efficiency of a xenogeneicvaccination in mice using an adenoviral vector expressing p185.Inbred BALB/c mice engineered to overexpress the activated(G664E) neu oncogene under the control of the mouse mammarytumor virus promoter (MMTV-LTR; from now on referred to asneuT mice) were used as experimental model system.10 In neuTmice, the transgene is prevalently expressed in mammary glands.At 3 weeks of age, female neuT mice start a process of rapiddevelopment of tumors involving all the mammary glands. p185 isa xenogeneic protein in neuT mice since it differs in more than11% of the amino acid residues when compared to both rat(r-p185) and endogenous mouse proteins.11
The goals of this study were to assess the efficiency of adeno-virus vs. plasmid DNA-mediated immunization by quantitativelymeasuring the humoral and the cell-mediated immune-response; toevaluate the efficiency of xenogeneic immunization in breakingtolerance in the transgenic mouse model and to verify the impactof Ad5-HER2-induced immunity on mammary tumor develop-ment in neuT mice.
Material and methodsCell lines
Mouse liver NMuLi and target mouse P815 plasmacytoma cellswere obtained from the American Type Culture Collection(ATCC, Rockville, MD), Per.C6 cells were from Crucell (Leiden,The Netherlands) and mouse muscle myoblasts C2C7 from G.
Abbreviations: A, absorbance; Ad5, adenovirus 5; AP, alkaline phos-phatase; CMI, cell-mediated immune; ECD, extracellular domain; ES,electrical stimulation; HER2, human epidermal growth factor receptor 2;ICS, intracellular staining; MCF, mean channel fluorescence; MMTV-LTR, mouse mammary tumor virus long terminal repeat; MOI, multiplicityof infection; neu, rat homolog of HER2; p185, protein encoded by HER2;PCNA, proliferating cell nuclear antigen; r-p185, protein encoded by neu;SEB, staphylococcus enterotoxin B; SFC, spot forming cell; vp, viralparticle.
Grant sponsor: the Italian Association for Cancer Research; Grant spon-sor: the Italian Ministry for University; Grant sponsor: the Center ofExcellence on Aging at the University of Chieti; Grant sponsor: ItalianMinistero dell’Istruzione, dell’Universita e della Ricerca; Grant number:FIRB RBME017BC4.
*Correspondence to: Molecular and Cell Biology Department,I.R.B.M. P. Angeletti, Via Pontina km 30.6, 00040, Pomezia (Roma), Italy.Fax: �39-06-91093-654. E-mail: [email protected]
Received 22 October 2003; Accepted after revision 9 June 2004DOI 10.1002/ijc.20536Publishedonline25August2004inWileyInterScience(www.interscience.
wiley.com).
Int. J. Cancer: 113, 67–77 (2005)© 2004 Wiley-Liss, Inc.
Publication of the International Union Against Cancer

Cossu (University of Rome, Rome, Italy). N202.1Aneu� is acloned cell line established in vitro from a lobular mammarycarcinoma that arose in FVB female mouse (H-2q) transgenic forthe neu proto-oncogene driven by the MMTV promoter.12
N202.1Aneu� cells express high levels of r-p185 on the membrane,whereas N202.1Eneu� do not show detectable levels of the sameprotein as measured by cytofluorimetric analysis. The human em-bryonic kidney HEK-293 cells secreting the dimeric fusion be-tween the extracellular domain of p185 and the Fc portion ofhuman IgG1
13 (indicated as HER2/ECD throughout this article)were kindly provided by Dr. Y. Yarden (Weizmann Institute ofScience, Rehovot, Israel)
PeptidesThree hundred and twelve 15-amino acid peptides, overlapping
by 11 amino acids, were designed to span the entire HER2 se-quence. These peptides, which also included 7 peptides designedto overcome insolubility problems, were synthesized by SynPep(Dublin, CA). The same company also synthesized the 9 aminoacid peptides described in this work. All peptides were shown tobe � 90% pure by HPLC and were used without HPLC purifica-tion. Peptides were reconstituted at a maximal concentration of35 mg/ml in DMSO and combined so that the final concentrationof each peptide in the mix was 1 mg/ml. Each mix was aliquotedand stored at �80°C.
ImmunogenspVIJ-HER2. HER2 coding sequence was amplified by PCR
from plasmid pLTR-2/erb-B211 (kindly provided by P. Di Fiore,European Institute of Oncology, Milan, Italy) by HER2.for1(5�-CCAGTTTAAACATTTAAATGCCGCCACCATGGAGCT-GGCGGCCT-3�; coding sequence is underlined) and HER2.rev2(5�-GCCGTCGACTTTACACTGGCACGTCCAGACCCA-3�)primers by Taq polymerase (TaKaRa LA Taq, TaKaRa Otsu,Shiga, Japan). The amplification product incorporating an opti-mized translation start site14,15 was digested with PmeI and SalIrestriction enzymes and cloned into the EcoRV and SalI sites ofmammalian expression plasmid pVijnsA.16 The plasmid pVIJ-HER2 thus generated contained the full-length HER2 sequenceunder the transcriptional control of the human cytomegalovirusimmediate-early promoter with its intron A sequence. The HER2coding sequence was followed by the bovine growth hormonepolyadenylation signal sequence. Sequence analysis of the codingregion in plasmid pVIJ-HER2 detected nucleotide changes A64Gand G1963A corresponding to amino acid changes Ser22Gly andVal655Ile compared to the reported sequence.11
pVIJ-neu. neu cDNA was obtained by PCR amplificationof template plasmid pneu-cDNA3/1717 by the neu.for-1(5�-CCGATATCGCCGCCGCCATGATCATCATGGAGC-TqGGCGGCC-3�) and neu.rev-1 (5�-CAGTCGACTCATACA-GGTACATCCAGGCCTAG-3�) primers. The resulting PCRproduct was EcoRV-SalI-digested and cloned into the correspond-ing sites of pVIJnsA to obtain pVIJ-neu.
In vitro expression from both plasmids was confirmed by tran-siently transfecting human embryonic kidney HEK-293 or mousemyoblasts C2C7 cell lines and detecting p185 or r-p185 expressionby flow cytometry. Supercoiled endotoxin-free plasmid DNAsused for immunization were purified from E. coli DH12S cells(Invitrogen, Groningen, The Netherlands) by Qiagen endo-freeplasmid Giga Kit (Qiagen, Hilden, Germany).
Ad5-HER2. A 3,785 bp DNA fragment obtained by SwaI-SalIrestriction of plasmid pVIJ-HER2 and containing the HER2 cod-ing sequence was cloned into the corresponding sites of theMRKAd5 shuttle vector.18 The resulting plasmid MRKAd5-HER2contained a human CMV promoter without the intron A sequence,which drives the expression of the HER2 cDNA, followed by thebovine growth hormone polyadenylation signal. This HER2 ex-pression cassette and the flanking Ad5 sequences were insertedinto the adenoviral backbone plasmid MRKpAd5 via bacterialhomologous recombination to generate the preadenoviral plasmid
MRKpAd5-HER2. This vector incorporates features aimed at in-creasing its genetic stability, maximizing the viral output andensuring efficient transgene expression (fully intact packagingsignal region, E1�E3� backbone, left-to-right transgene orienta-tion, etc.).18
Preadenoviral plasmid MRKpAd5-HER2 was linearized byPacI digestion and transfected into PerC6 cells to generate Ad5-HER2 recombinant adenovirus. The virus was grown in largequantities by multiple rounds of amplification and purified bycaesium chloride gradient ultracentrifugation.19 Viral DNA wasextracted by proteinase K digestion and genomic integrity wasconfirmed by restriction analysis. Virus particle (vp) titer wasdetermined spectrophotometrically considering that 1 U of opticaldensity at 260 nm corresponds to 1.1 � 1012 particles/ml.20 Invitro expression was confirmed by flow cytometric analysis ofinfected murine liver NMuli cells infected with Ad5-HER2 atdifferent multiplicity of infection (MOI; data not shown).
Ad5-neu. DNA sequence coding for r-p185 was generated byPCR assembly of synthetic oligonucleotides to contain codonspreferred by highly expressed human genes, in addition to K753Amutation abolishing tyrosine kinase activity. A 3,820 bp PmeI-SalIDNA fragment containing the neu cDNA optimized was cloned inthe corresponding sites of the MRKAd5 shuttle vector. neu ex-pression cassette and the flanking Ad5 sequences from plasmidMRKAd5-neu were inserted into the adenoviral backbone plasmidMRKpAd5 via bacterial homologous recombination to generatethe preadenoviral plasmid MRKpAd5-neu. Preadenoviral plasmidMRKpAd5-HER2 was linearized by ClaI digestion and transfectedinto PerC6 cells to generate Ad5-neu recombinant adenovirus asdescribed above.
Recombinant vaccinia virus. neu cDNA was obtained by PCRamplification of template plasmid pRatHER2/neu-cDNA3/1717 us-ing the neu.for-1 (5�-CCGATATCGCCGCCGCCATGATCA-TCATGGAGCTGGCGGCC-3�) and neu.rev-1 (5�-CAGTCG-ACTCATACAGGTACATCCAGGCCTAG-3�) primers. The re-sulting PCR product was EcoRV-SalI-digested and cloned into thecorresponding sites of pVIJnsA to obtain pVIJ-neu. In vitro ex-pression was confirmed in HEK-293 and mouse myoblasts C2C7cell lines by flow cytometry. A 3,810 bp KpnI/NotI DNA fragmentwas obtained from plasmid pVIJ-neu, which contained the entireneu coding sequence and cloned into the corresponding sites ofshuttle plasmid pSC11 (kindly provided by B. Moss; NationalInstitutes of Health, Bethesda, MD).21 neu cDNA was inserteddownstream of the viral P7.5 promoter by homologous recombi-nation between the shuttle plasmid pSC11-neu and the vacciniavv-wt (strain WR, obtained from ATCC). The recombinant vac-cinia virus thus generated (vv-neu) was propagated in HeLa cellsand purified as described.22 Flow cytometry analysis detectedr-p185 expression upon infection of HEK-293 and C2C7 cell.
MiceFemale BALB/c mice (H-2d) transgenic for the transforming rat
HER2/neu oncogene (neuT�/neuT�) under the transcriptionalcontrol of the MMTV promoter and transgene negative (neuT�/neuT�) were bred under specific pathogen-free conditions byCharles River Breeding Laboratories (Calco, Italy).10,23 Mice werescreened for the presence of the transgene by duplex PCR. Briefly,genomic DNA was extracted from the mouse tail using theDNeasy 96 tissue Kit (Qiagen). Multiplex PCR was performed onthis substrate using 2 pairs of primers. MMTV-for1 (5�-AAG-GAGATGGCGCCCAACAGTC-3�) and MMTV-rev1 (5�-GGC-TGCGACTAAGCGAAACAATTC-3�) amplified a 248 bp of theMMTV promoter region. An additional pair of primers (m�glo-bin-for1, 5�-AGGGCGATTCTCTGAAGGCAGTTATT-3�, andm�globin-rev1, 5�-TTTCTTTGTTTCCCAGTTTGAGTGCAT-3�) were used to amplify mouse m�globin sequences as positiveinternal control, generating a 310 bp product.
In all operations, mice were treated in accordance with Euro-pean guidelines. In particular, mice were fully anesthetized with
68 GALLO ET AL.

ketamine (Imalgene 500; Merial Italia, Milano, Italy) at 100 mg/kgof body weight and xylazine (Xilor, BIO 98; S. Lazzaro, Bologna,Italy) at 5.2 mg/kg.
Evaluation of tumor growthneuT females were inspected weekly and tumor masses mea-
sured with calipers in 2 perpendicular diameters. Progressivelygrowing masses � 3 mm in mean diameter were regarded astumors. Growth was monitored until all mammary glands dis-played a palpable tumor or until a tumor exceeded an averagediameter of 10 mm, at which time mice were sacrificed for humanereasons. Except where otherwise specified, surviving neuT micewere sacrificed at 33 weeks.
Immunization protocolsMice received a total of 109 vp of Ad5-HER2 into the left and
right quadriceps muscles (5 � 108 vp in 50 �l of physiologicsolution per leg) at 6 and 9 weeks of age. Alternatively, at thetimes indicated, a total amount of 50 or 100 �g of pVIJ-HER2plasmid DNA was injected into the quadriceps muscle (25 or50 �g in 50 �l of physiologic solution per leg) and electrostimu-lation (ES) was performed as previously described24 (10 trainswith 1,000 bipolar pulses of 130 V, 75 mA, 200 �sec/phase). Serawere collected at 5 (preimmune serum), 11 and 22 weeks of age.
ELISPOT assayMouse spleen cells secreting IFN-� in an antigen-specific man-
ner were detected using a standard enzyme-linked immunospot(ELISPOT) assay.25 Spleen cells were prepared from immunizedmice and resuspended in R10 medium (RPMI-1640 supplementedwith 10% fetal calf serum, 2 mM L-glutamine, 50 U/ml penicillin,50 �g/ml streptomycin, 10 mM HEPES, 50 �M 2-mercaptoetha-nol). Multiscreen 96-well filtration plates (catalog numberMAIPS4510; Millipore, Bedford, MA) were coated with an affin-ity-purified rat antimouse IFN-� antibody (catalog number18181D, PharMingen, San Diego, CA). After overnight incuba-tion, plates were washed with PBST (0.005% Tween in PBS) andincubated with R10 medium for 2 hr at 37°C to block nonspecificbinding. Cells from the spleen of 2 or more immunized mice werepurified as described above, mixed, resuspended in R10 mediumand incubated for 15 hr in the presence of 6 �g/ml of a single ora pool of peptides, at a density of 2.5–5 � 105 cells/well. Afterextensive washing with PBST, biotinylated rat antimouse IFN-�antibody (catalog number 18112D, PharMingen) was added. Theplates were incubated at 4°C overnight and then washed withPBST prior to the addition of streptavidin-alkaline phosphatase(catalog number 13043E, PharMingen). After incubation for 2 hrat room temperature, the plates were extensively washed withPBST and developed by incubating with a one-step nitrobluetetrazolium-5-bromo-4-chloro-3-indolylphosphate substrate (cata-log number 34042, Pierce, Rockford, IL) for 5–15 min for devel-opment of spots. Rinsing the plates in water stopped the reaction.DMSO and concanavalin A (10 �g/ml) were included as back-ground and positive control for each sample. Spots were countedby computer-assisted imaging analysis (AID ELR02 coupled withAID ELISPOT 2.6 Software, Strassberg, Germany).
Flow cytometric analysisPlasmids pVIJ-HER2 or pVIJ-neu were lipofectamine-trans-
fected (Gibco BRL Invitrogen, Groningen, The Netherlands) inmouse myoblasts C2C7 and HEK-293 cells, while murine liverNMuli cells were infected with Ad5-HER2 or vv-neu. After 48 hrof incubation, cells were collected using trypsin and stained by astandard indirect immunofluorescence procedure using an anti-p185 mAb Ab-2 or anti-r-p185 mAb Ab-4 (Oncogene, Boston,MA) as primary antibody, followed by PE-conjugated goat anti-mouse IgG1 and antimouse IgG (catalog number Star81PE andSTAR76, respectively, Diamedix, Miami, FL). All samples wereacquired within 24 hr of staining using a FACSCalibur flowcytometer and CellQuest software (BD Immunocytometry Sys-tems, San Diego, CA).
Intracellular IFN-� production was measured according to BDPharmingen standard protocol. Briefly, 2 � 106 spleen cells werecultured for 15 hr in R10 medium in the presence of 6 �g/ml ofsingle or pool of peptides and brefeldin A as protein transportinhibitor (Cytofix/Cytoperm Plus with GolgiPlug Kit; BD Pharm-ingen). Staphylococcus enterotoxin B (SEB) at 10 �g/ml (catalognumber S4881, Sigma, St. Louis, MI) and DMSO were tested withthe spleen cells as positive and background control, respectively.Before staining of surface antigens, Ab antimouse CD16/CD32was used to reduce nonspecific immunofluorescent signal (catalognumber 553142, BD PharMingen). The specific signal was ob-tained with APC-antimouse CD3e, PE-antimouse CD4 and PerCP-antimouse CD8a (catalog numbers 553066, 553653 and 553036,BD Pharmingen). The cells were then washed, fixed, permeabil-ized and stained for intracellular IFN-� using FITC-conjugatedmAb (catalog number 554411, BD Pharmingen).
MAb Ab-4 was used to establish quantitative measurement ofanti-r-p185 antibody by flow cytometry. Ab-4 was serially dilutedand used to stain mouse N202.1Aneu�. Briefly, cells were collectedand incubated with mAb anti-CD16/CD32 at 4°C for 20 min.Serial dilutions of mAb Ab-4 were then added and left at 4°C for45 min. After washing, FITC-conjugated rabbit antimouse Ig (cat-alog number F0313, Dako, Glostrup, Denmark) was used fordetection of binding to r-p185; finally, propidium iodide was usedto gate out dead cells. Mean channel fluorescence (MCF) wasrecorded and the background signal generated by the secondary Ab(typically around 10) was subtracted. The range of Ab-4 antibodyconcentration that correlated with MCF in a linear fashion wasdetermined by regression analysis and found to be 5–1,000 ng/ml(R2 0.998). Serum antibodies crossreacting with r-p185 weremeasured through the same procedure. For each serum, serialdilutions were measured and the value obtained from the preim-mune serum at the lowest dilution was subtracted. No signal wasdetected using control N202.1Eneu� cells. Data were expressed asAb-4 equivalent on the basis of the standard curve.
ELISAMultiwell plates (Immunoplate Maxisorp, Nunc, Roskilde, Den-
mark) were coated overnight at 4°C with goat antihuman IgGFc-specific (catalog number 31123, Pierce) at a concentration of2 �g/ml in 50 mM NaHCO3 (pH 9.6). After discarding the coatingsolution, plates were incubated at 37°C for 60 min with PBSMT(5% nonfat dry milk, 0.05% Tween-20 in PBS). Plates werewashed with washing buffer (0.05% Tween-20 in PBS) and su-pernatant of IgB2-cells containing the HER2/ECD fused to humanFc region was added in saturating condition and incubated at roomtemperature for 2 hr. Plates were washed and serial dilutions inPBST buffer of preimmune or immune serum were added to thewells. A standard curve was generated using anti-p185 mAb Ab-2(Oncogene). Plates were incubated overnight at 4°C. After wash-ing, plates were incubated with mAb antimouse IgG AP-conju-gated (catalog number A7434, Sigma) for 60 min at room temper-ature and alkaline phosphatase activity detected by incubation withSigma 106 phosphatase substrate (catalog number A106, Sigma) indiethanolamine. The plates were read by an automated ELISAreader (Labsystems Multiskan Bichromatic, Helsinki, Finland) andthe results were expressed as A A405 nm–A620 nm.
Cytotoxicity assayCTL assay was performed as previously described.26 Briefly,
P815 cells expressing the H-2d class I molecule were used as targetcells. A total of 4 � 106 spleen cells were resuspended in R10supplemented with 5 �g of CD8�-specific peptide (HER2-15.3,HER2-301, or neu-15.3) and 10 U/ml of recombinant humaninterleukin-2 (Boehringer Mannheim, Mannheim, Germany) andcultured in 24-well flat-bottom plates (Multiwell-Tissue CulturePlate, Falcon-BD, San Diego, CA). Alternatively, about 20% ofspleen cells were stimulated with vv-rHER2 infection at an MOI of5 for 2 hr at 37°C and then plated as above.
69ANTI-r-p185 XENOGENEIC IMMUNIZATION
After 7 days of in vitro restimulation, cultures were assayed forcytotoxic activity in a standard 51Cr release assay. Target cellswere labeled with Na51CrO4 (Amersham Pharmacia Biotech,Buckinghamshire, U.K.) and pulsed with either the CD8� peptideor DMSO alone in R10 medium for 2 hr at 37°C and 5% CO2.After an extensive washing with R10 medium, target cells weremixed with CTLs at designated effector/target ratios in 96-wellround-bottom plates (Cell Wells, Corning Glass Works, Corning,NY) and incubated for 4 hr at 37°C and 5% CO2. Thirty �l
samples of supernatants were transferred to a LumaPlate-96 (Pack-ard Instrument, Meridien, CT), allowed to dry overnight, thencounted (TopCount, Microplate Scintillation Counter;PerkinElmer Instrument, Shelton, CT) to determine the amount of51Cr released in each well.
Whole-mount analysisGroups of 3 untreated and Ad5-HER2-vaccinated neuT mice
were sacrificed at weeks 17 and 23. Pelt preparation for mammary
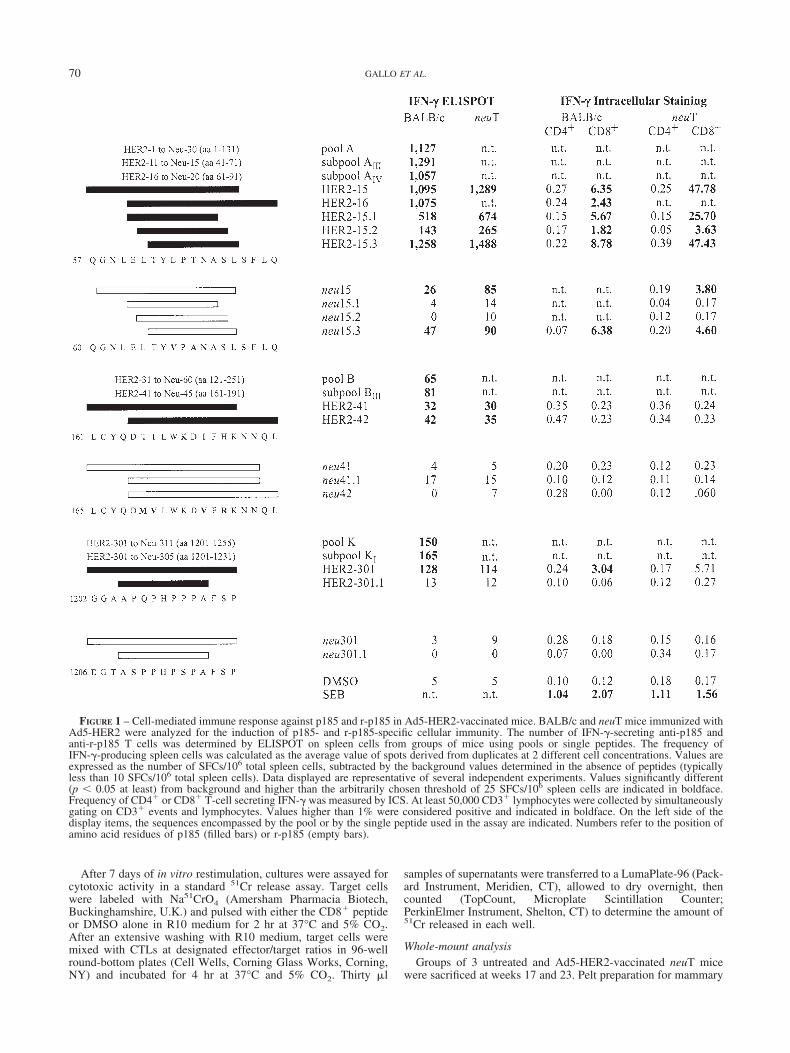
FIGURE 1 – Cell-mediated immune response against p185 and r-p185 in Ad5-HER2-vaccinated mice. BALB/c and neuT mice immunized withAd5-HER2 were analyzed for the induction of p185- and r-p185-specific cellular immunity. The number of IFN-�-secreting anti-p185 andanti-r-p185 T cells was determined by ELISPOT on spleen cells from groups of mice using pools or single peptides. The frequency ofIFN-�-producing spleen cells was calculated as the average value of spots derived from duplicates at 2 different cell concentrations. Values areexpressed as the number of SFCs/106 total spleen cells, subtracted by the background values determined in the absence of peptides (typicallyless than 10 SFCs/106 total spleen cells). Data displayed are representative of several independent experiments. Values significantly different(p 0.05 at least) from background and higher than the arbitrarily chosen threshold of 25 SFCs/106 spleen cells are indicated in boldface.Frequency of CD4� or CD8� T-cell secreting IFN-� was measured by ICS. At least 50,000 CD3� lymphocytes were collected by simultaneouslygating on CD3� events and lymphocytes. Values higher than 1% were considered positive and indicated in boldface. On the left side of thedisplay items, the sequences encompassed by the pool or by the single peptide used in the assay are indicated. Numbers refer to the position ofamino acid residues of p185 (filled bars) or r-p185 (empty bars).
70 GALLO ET AL.
gland whole mount was performed as described at http://ccm.ucdavis.edu/tgmouse/HistoLab/wholmt1.htm. Briefly, mice weresacrificed with anesthetic overdose and skinned. The pelts werefixed at least overnight in buffered formalin. Then mammaryglands were dissected, defatted in acetone, rehydrated and stainedwith ferric hematoxylin. Samples were then dehydrated andcleared with histolemon. Storing and image acquisitions of mam-mary glands were performed in methyl salicylate.
Histologic and immunohistochemical analysisAfter whole-mount image acquisition, 2 representative mam-
mary glands for each mouse were embedded in paraffin, sectionedat 4 �m and stained with hematoxylin and eosin. To evaluate theexpression of r-p185 protein and proliferating cell nuclear antigen(PCNA), paraffin-embedded sections were tested with polyclonalrabbit anti-r-p185 Ab (C-18, Santa Cruz Biotechnology, SantaCruz, CA) and anti-PCNA Ab (Ylem, Roma, Italy). After washing,they were overlaid with biotinylated goat antirabbit Ig (VectorLaboratories, Burlingame, CA) for 30 min. Unbound Ab wasremoved by washing and the slides were incubated with ABCcomplex/AP (Dako). Morphologic studies were conducted inde-pendently by 3 pathologists in a blind experiment.
Statistical analysisDifferences in tumor incidence were evaluated using the Man-
tel-Haenszel log-rank test and differences in tumor/mouse num-bers using Wilkoxon’s rank-sum test. Student’s t-test was used foridentifying statistically significant values in ELISPOT assay andwhen comparing antibody titers in the same or between differentgroups of mice.
ResultsIdentification of immunodominant T-cell epitopes inp185 protein
To identify the immunodominant T-cell epitopes of p185 inBALB/c mice (H-2d), 6-week-old female BALB/c mice wereimmunized by injecting 109 vp of Ad5-HER2 in the quadricepsmuscles. A second injection was performed after 3 weeks. Asecond group of mice was similarly injected with saline solution asnegative control. Three weeks after the second injection, the ani-mals were sacrificed and the frequency of IFN-� secreting T cellsin mouse spleen cells was evaluated by interferon-� ELISPOTassay.
Three hundred and eleven peptides, each 15 amino acids long,overlapping by 11 residues and spanning the entire p185 sequence,were combined into 11 pools indicated with alphabetical lettersfrom A to K, from N to C terminus. Each of these pools was testedfor its ability to stimulate IFN-� secretion by spleen T cells. Forpeptide pools A, B and K, ELISPOT measured a statisticallysignificant IFN-� production by mice immunized with Ad5-HER2as compared to control in the absence of peptide (Fig. 1). Toidentify the individual peptide responsible for this activity, pep-tides from pool A, B and K were divided into subpools, amongwhich AIII and AIV, BIII and KI scored positive. Single peptidesfrom these positive subpools were then tested for their ability totrigger IFN-� release. Overlapping peptides HER2-15 andHER2-16 exhibited high and comparable reactivity (Fig. 1). Amuch lower reactivity was exhibited by overlapping peptidesHER2-41 and HER2-42. Another peptide, HER2-301, was alsoshown to contain a T-cell epitope.
To confirm these data and identify the CD4� or CD8� T-cellsubset responsible for IFN-� production, we characterized IFN-�-
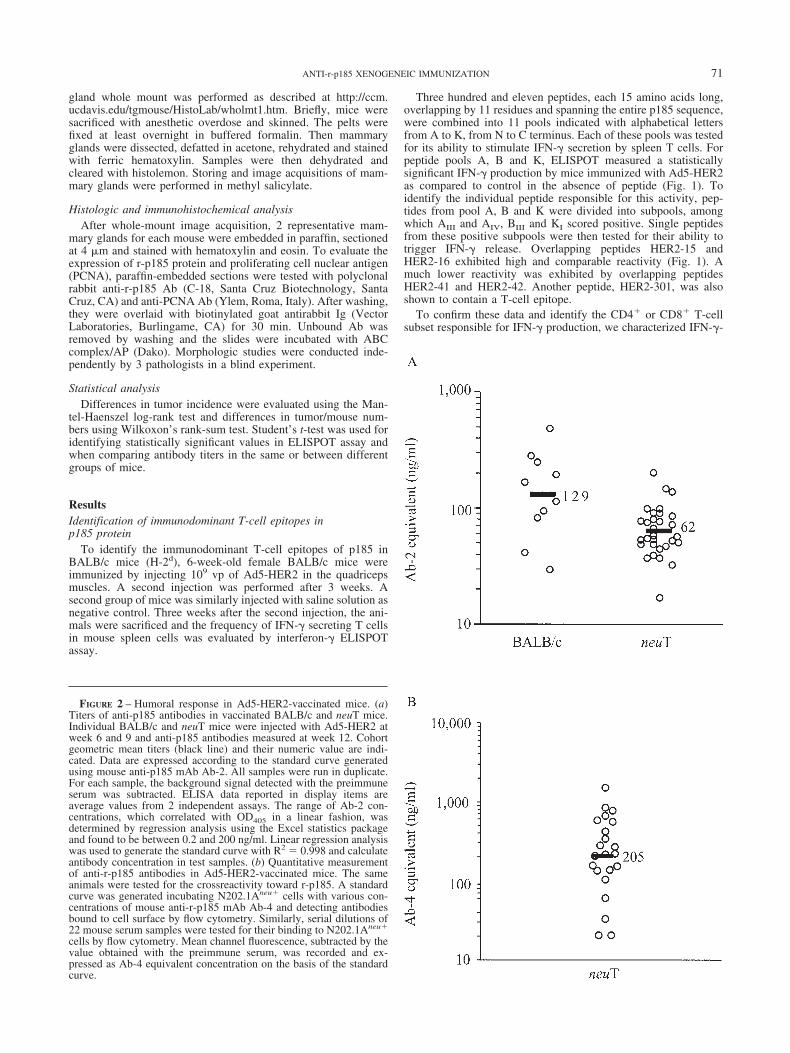
FIGURE 2 – Humoral response in Ad5-HER2-vaccinated mice. (a)Titers of anti-p185 antibodies in vaccinated BALB/c and neuT mice.Individual BALB/c and neuT mice were injected with Ad5-HER2 atweek 6 and 9 and anti-p185 antibodies measured at week 12. Cohortgeometric mean titers (black line) and their numeric value are indi-cated. Data are expressed according to the standard curve generatedusing mouse anti-p185 mAb Ab-2. All samples were run in duplicate.For each sample, the background signal detected with the preimmuneserum was subtracted. ELISA data reported in display items areaverage values from 2 independent assays. The range of Ab-2 con-centrations, which correlated with OD405 in a linear fashion, wasdetermined by regression analysis using the Excel statistics packageand found to be between 0.2 and 200 ng/ml. Linear regression analysiswas used to generate the standard curve with R2 0.998 and calculateantibody concentration in test samples. (b) Quantitative measurementof anti-r-p185 antibodies in Ad5-HER2-vaccinated mice. The sameanimals were tested for the crossreactivity toward r-p185. A standardcurve was generated incubating N202.1Aneu� cells with various con-centrations of mouse anti-r-p185 mAb Ab-4 and detecting antibodiesbound to cell surface by flow cytometry. Similarly, serial dilutions of22 mouse serum samples were tested for their binding to N202.1Aneu�
cells by flow cytometry. Mean channel fluorescence, subtracted by thevalue obtained with the preimmune serum, was recorded and ex-pressed as Ab-4 equivalent concentration on the basis of the standardcurve.
71ANTI-r-p185 XENOGENEIC IMMUNIZATION
secreting T cell by intracellular staining (ICS). IFN-� ICS con-firmed the reactivity of peptide HER2-15, identifying it as anepitope able to activate CD8� cells. Peptide HER2-15 andHER2-16 were equally reactive in ELISPOT analysis, suggestingthat the CD8� epitope should be comprised in the 11 amino acidresidues common to the 2 peptides. To identify the target nonamersequence, we tested 3 9-amino acid long peptides spanning theoverlapping region between HER2-15 and HER2-16. HER2-15.3proved the most reactive, displaying a slightly increased reactivitycompared to the 15 amino acid long peptides HER2-15 and HER2-16. Interestingly, about half of the reactivity was also detected withHER2-15.1, indicating that 2 overlapping but distinct CD8�
epitopes coexist in this 11 amino acid sequence.IFN-� ICS analysis also confirmed the reactivity of HER2-301
and typed it as a CD8� epitope. Analyzing these CD8� epitopesby IFN-� ELISPOT confirmed the results obtained by ICS (Fig. 1).Finally, a low reactivity was detected for HER2-41 and HER2-42peptides, whose low response was predominantly CD4�.
Identification of immunodominant T-cell epitopes inr-p185 protein
neuT mice overexpress r-p185 that has 89% amino acid simi-larity to the p185 human homolog. To test whether Ad5-HER2vaccination could induce a CMI response that crossreacted withr-p185, the frequency of IFN-�-secreting T cells triggered by ratpeptides corresponding to the immunodominant epitopes identifiedin the human protein was measured. As shown in Figure 1, thenonamer human CD8� epitope identified by peptide HER2-15.3has 2 amino acid residues different from r-p185, at position 3 and5. This sequence variation translates into about a 10-fold decreasein reactivity to the rat peptide, which however elicited a statisti-cally significant response in both ELISPOT and ICS assays. Thesame sequence variations completely abolished the reactivity ofthe overlapping epitope identified by the HER2-15.1 peptide.
Amino acid variations are also present between the sequencecovered by HER2-41/HER2-42 and the corresponding sequence inr-p185. These changes reduced the activity of the correspondingrat peptides to background levels in both ELISPOT and ICSassays. A similar situation was detected for the CD8� epitopedefined by the HER2-301 peptide. Five amino acid changes abol-ished the reactivity of the corresponding 15 amino acid rat peptidein both ELISPOT and ICS assays.
Data reported in Figure 1 show that all 3 epitopes analyzed,whether derived from the human or the rat sequence, display acomparable reactivity in BALB/c and neuT mice, as concomitantlymeasured by ELISPOT and ICS assays. As p185 is a nonself-protein in both BALB/c and neuT mice, transgene expression didnot affect the efficiency of the response induced by the Ad5-HER2vaccination.
Humoral response in Ad5-HER2-vaccinated miceWe evaluated by ELISA the titer of serum antibodies elicited in
BALB/c and neuT after vaccination with Ad5-HER2. We used thedimeric extracellular domain of p185 (HER2-ECD) as target an-tigen. A standard reference curve was obtained using an anti-p185mAb Ab-2 targeting an epitope in the HER2-ECD. Serum sampleswere taken from BALB/c and neuT mice 3 weeks after the secondinjection with Ad5-HER2 and individually tested by ELISA. An-tibody concentration was obtained through linear regression anal-ysis of serially diluted samples. Titers reported in Figure 2(a) areexpressed as mAb-2 concentration on the basis of the standardreference curve. High and comparable titers of anti-p185 ECDantibodies were measured in BALB/c and neuT mice (geometricmean 129 and 62 ng/ml, respectively). The difference betweenthe 2 distributions is not statistically significant.
N202.1Aneu� cells from neuT primary tumors constitutivelyexpress a high number of r-p185 membrane receptors.12 Thesecells were used to measure mouse antibodies elicited by Ad5-
TABLE I – COMPARISON OF p185- AND r-p185-SPECIFIC T-CELL RESPONSE ELICITED IN MICE BY INJECTING pVIJ-HER2 DNA AND pVIJ-neu WITH ES
Peptide
pVIJ-HER2 pVIJ-HER2 pVIJ-neu pVIJ-neu
50 �g week 6, 91 100 �g week 6, 9 50 �g week 6, 8,10, 12
50 �g week 6, 8,10, 122
50 �g week 6, 8,10, 12 50 �g week 6,9
BALB/c neuT BALB/c neuT BALB/c neuT BALB/c neuT BALB/c neuT BALB/c neuT
HER2-15.3 413 56 37 55 51 46 52 45 8 10 3 7neu-15.3 1 1 1 1 2 5 28 20 75 20 37 19HER2-41 1 2 4 2 3 4 18 10 19 12 12 13neu-41 3 3 2 2 2 2 30 20 38 16 40 19HER2-301 37 30 30 32 26 36 27 38 0 2 0 1neu-301 0 1 1 1 1 1 3 1 1 3 2 11Week of vaccination.–2pVIJ-HER2 was injected at week 6, 8 and 10, followed by pVIJ-neu at week 12.–3Values refer to the frequency of
IFN-�-secreting spleen cells as measured by ELISPOT. Data displayed are representative of several independent experiments. Values areexpressed as SFC/106 total spleen cells, subtracted by the background values determined in the absence of peptides (typically less than 5 SFC/106
spleen cells). Values significantly different (p 0.05 at least) from background measured in control experiments without antigenic peptides andhigher than the arbitrarily chosen threshold of 25 SFC/106 spleen cells are indicated in boldface.
TABLE II – COMPARISON OF p185- AND r-p185-SPECIFIC T-CELL RESPONSE ELICITED IN MICE BYIMMUNIZATION WITH Ad5-HER2 AND Ad5-neu
PeptideAd5-hHER2, 109 vp week 6, 91 Ad5-neu, 109 vp week 6, 9
BALB/c neuT BALB/c neuT
HER2-15.3 1,2582 1,488 53 11neu-15.3 47 90 1070 232HER2-41 32 30 n.t. n.t.neu-41 4 5 n.t. n.t.HER2-301 128 114 n.t. n.t.neu-301 3 9 n.t. n.t.1Week of vaccination.–2Values refer to the frequency of IFN-�-secreting spleen cells as measured by
ELISPOT. Data displayed are representative of several independent experiments. Values are expressed asSFC/106 total spleen cells, subtracted by the background values determined in the absence of peptides(typically less than 5 SFC/106 spleen cells). Values significantly different (p 0.05 at least) frombackground measured in control experiments without antigenic peptides and higher than the arbitrarilychosen threshold of 25 SFC/106 spleen cells are indicated in boldface.
72 GALLO ET AL.

HER2 vaccination that crossreacted with r-p185. The titers ofanti-r-p185 antibodies varied within a 10-fold range, from 21 to1,504 ng/ml, with a mean value of 205 ng/ml Ab-4 equivalent(Fig. 2b). The titer of antibodies against p185 paralleled thoseevaluated against r-p185, which is not surprising when we considerthe 90% similarity between the amino acid sequences of these 2extracellular domains.
Immunization efficiency of DNA injection withelectrical stimulation
To compare the immunization efficiency of adenovirus with thatof plasmid DNA associated with electrical stimulation, we con-structed plasmid pVIJ-HER2 harboring the CMV-HER2 expres-sion cassette. Fifty �g of plasmid pVIJ-HER2 were injected intoquadriceps muscles followed by ES at 6 and 9 weeks of age.Two weeks after boosting, animals were sacrificed and spleen cellscollected and stimulated with the peptides containing the immu-nodominant p185 and r-p185 epitopes. Very low spot forming cells(SFCs) were detected following stimulation with human peptidesboth in BALB/c and neuT mice (Table I). On average, the responsewas 50-fold lower than that induced by Ad5-HER2 immunization.On the same lines, crossreactivity with rat peptide epitopes wasbarely detectable with the neu-15.3 peptide containing a CD8�
epitope.To improve the efficiency of DNA immunization, we evaluated
a number of modifications of the standard protocol. A total amountof 100 �g of pVIJ-HER2 DNA was injected in BALB/c and neuTmice (50 �g in each quadriceps muscle, followed by ES). Thisinjection was repeated 3 weeks later. Two weeks after the lastinjection, mice were sacrificed and spleen cells collected. Increas-ing the amount of injected DNA did not raise the frequency of Tcells producing IFN-� compared to the standard protocol (Table I).We also tested the efficiency of an immunization protocol in whichmice were immunized with 50 �g of pVij-HER2 4 times with2-week intervals. This repeated immunization only slightly en-hanced the intensity of the response, as measured by ELISPOTanalysis (Table I). An additional protocol where the 4 immuniza-tions were administered weekly gave comparable results (data notshown). On the same line, testing different ES conditions did notincrease the intensity of the response (data not shown).
The above data show that immunizing with plasmid pVIJ-HER2induced a comparable response in BALB/c and neuT mice for eachprotocol. This response, however, was mostly directed toward thehuman rather than the rat epitopes. These findings led us toimmunize mice using plasmid pVIJ-neu according to the standardprotocol (injection of 50 �g plasmid DNA with ES at week 6 and9, analysis at week 14). In BALB/c mice, a response slightlyhigher than that induced by the human gene was detected. In thiscase, however, the response was mainly directed to the ratepitopes. In neuT mice, which are tolerant to r-p185, this reactivitywas still skewed in favor of the rat epitope, but clearly reducedcompared to that obtained in BALB/c mice.
These results indicate that xenogeneic immunization induced aCMI response to p185 epitopes, which poorly crossreacted withr-p185 epitopes. To increase the response to the rat proteinepitopes, we explored protocols combining xeno- and syngeneicvaccination. Mice were immunized with 50 �g of VIJ-HER2 atweek 6, 8 and 10, followed by injection of 50 �g of pVIJ-neu atweek 12. This protocol induced a response to the human and to therat epitopes, which mirrored that obtained by the control pVIJ-HER2 or pVIJ-neu immunization, respectively. It is worth notingthat the frequency of IFN-�-secreting T cells to the r-p185 immu-nodominant was clearly higher than that induced by pVIJ-HER2,but lower than that obtained by control pVIJ-neu immunization.Similarly, different combinations of pVIJ-HER2 and pVIJ-neuinjections did not induce a response against r-p185 higher than thatobtained by control pVIJ-neu immunization
As a whole, these results indicated that pVIJ-HER2 DNA im-munization has a lower efficiency in eliciting cell-mediated im-
munity against r-p185 as compared to pVIJ-neu. A higher responseto r-p185 epitopes is obtained by Ad5-HER2. Preliminary datashow that in neuT mice, an adenovirus expressing neu induces ananti-r-p185 T-cell response higher than that elicited by Ad-HER2immunization (Table II).
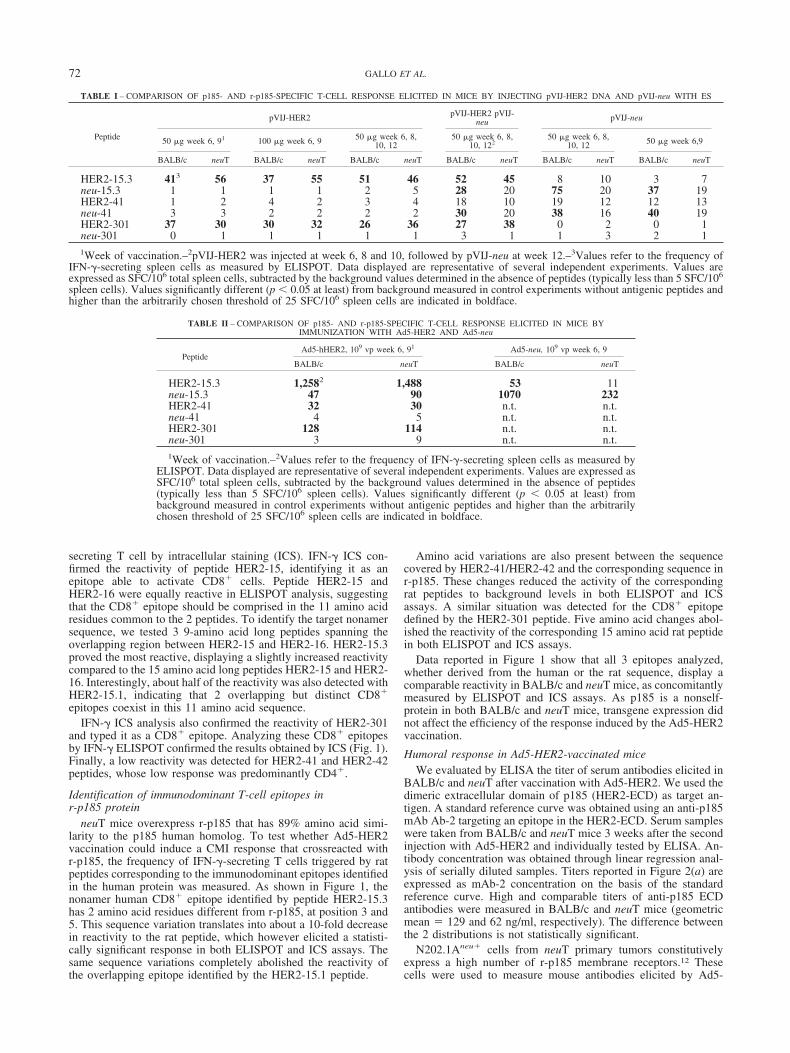
Cytotoxicity activityThe CTL activity was evaluated at week 11 in BALB/c and
neuT mice immunized with Ad5-HER2 injection at week 6 and 9.Spleen cells were restimulated in vitro with IL-2 and p185 peptidesfor 5–7 days. Target P815 cells (H-2d) were cultured in thepresence of human or rat CD8� peptides, labeled with 51Cr, thenmixed with effector cells at different ratios. Spleen cells fromBALB/c mice immunized with Ad5-HER2 and restimulated witheither HER2-15 or HER2-301 peptides displayed a significant lyticactivity against P815 target cells pulsed with the same p185peptide (Fig. 3). By contrast, restimulated spleen cells from im-munized neuT mice displayed a lower CTL activity. Followingrestimulation with HER2-15.3, their lytic activity against HER2-15.3-pulsed target cells was about 4-fold lower than in BALB/cmice. A similar decrease was observed when spleen cells restim-ulated with HER2-301 were tested against target cells pulsed withthe same peptide. Following restimulation with neu-15.3, neuTspleen cells showed a cytolytic activity against neu-15.3-pulsedtarget cells comparable to that displayed by HER2-15.3 peptide.
Finally, we evaluated the induction of CTL against naturallyprocessed r-p185 in neuT mice. To this end, we generated arecombinant vaccinia virus expressing the neu gene (vv-neu).About 20% of spleen cells from Ad5-HER2-immunized mice wereinfected with vv-neu, mixed with the remaining spleen cells andcultured as above (effector cells). P815 cells were pulsed with
FIGURE 3 – CTL response in Ad5-HER2-immunized mice. (a)Spleen cells from BALB/c mice vaccinated with Ad5-HER2 wererestimulated in vitro with HER2-15 or HER2-301 peptides and testedfor cytotoxic activity against P815 target cells pulsed with the samepeptides. As negative control, HER2-15- or HER2-301-restimulatedspleen cells were tested on DMSO-pulsed P815 target cells. Averagevalues are reported. (b) Spleen cells from neuT mice vaccinated withAd5-HER2 were restimulated in vitro with HER2-15.3, neu-15.3, orHER2-301 and tested for cytotoxic activity against P815 target cellspulsed with the same peptides or DMSO (negative control). Spleencells restimulated by infection with recombinant vaccinia virus ex-pressing r-p185 protein (vv-neu) were tested on target cells pulsedwith neu-15.3 peptide or DMSO (negative control). Spleen cells fromnonvaccinated mice reproducibly yielded background values ofcounts. The percentage of specific lysis was calculated by using thefollowing formula: % specific lysis 100 � [(experimental release �spontaneous release)/(maximum release � spontaneous release)],where the spontaneous and maximum release refer to the number ofcounts in DMSO or in 1% Triton X-100 target cell lysate, respectively.Effector cell/target cell ratios are indicated in the abscissa. The per-centage of specific killing is reported on the vertical axis. Averagevalue of negative controls is displayed. Reported data are representa-tive of several independent experiments.
73ANTI-r-p185 XENOGENEIC IMMUNIZATION
neu-15.3 peptide and labeled with Na51CrO4 (target cells). Effec-tor cells showed a detectable CTL activity when incubated withtarget cells at designated effector/target ratios (Fig. 3).
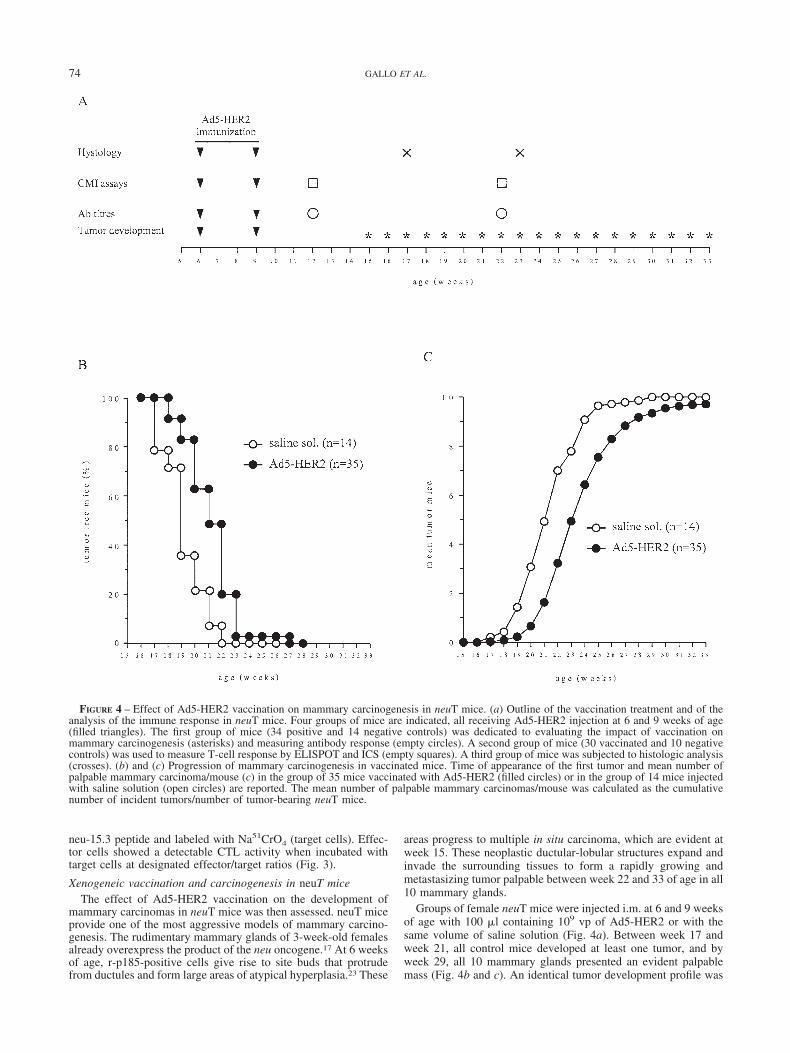
Xenogeneic vaccination and carcinogenesis in neuT miceThe effect of Ad5-HER2 vaccination on the development of
mammary carcinomas in neuT mice was then assessed. neuT miceprovide one of the most aggressive models of mammary carcino-genesis. The rudimentary mammary glands of 3-week-old femalesalready overexpress the product of the neu oncogene.17 At 6 weeksof age, r-p185-positive cells give rise to site buds that protrudefrom ductules and form large areas of atypical hyperplasia.23 These
areas progress to multiple in situ carcinoma, which are evident atweek 15. These neoplastic ductular-lobular structures expand andinvade the surrounding tissues to form a rapidly growing andmetastasizing tumor palpable between week 22 and 33 of age in all10 mammary glands.
Groups of female neuT mice were injected i.m. at 6 and 9 weeksof age with 100 �l containing 109 vp of Ad5-HER2 or with thesame volume of saline solution (Fig. 4a). Between week 17 andweek 21, all control mice developed at least one tumor, and byweek 29, all 10 mammary glands presented an evident palpablemass (Fig. 4b and c). An identical tumor development profile was
FIGURE 4 – Effect of Ad5-HER2 vaccination on mammary carcinogenesis in neuT mice. (a) Outline of the vaccination treatment and of theanalysis of the immune response in neuT mice. Four groups of mice are indicated, all receiving Ad5-HER2 injection at 6 and 9 weeks of age(filled triangles). The first group of mice (34 positive and 14 negative controls) was dedicated to evaluating the impact of vaccination onmammary carcinogenesis (asterisks) and measuring antibody response (empty circles). A second group of mice (30 vaccinated and 10 negativecontrols) was used to measure T-cell response by ELISPOT and ICS (empty squares). A third group of mice was subjected to histologic analysis(crosses). (b) and (c) Progression of mammary carcinogenesis in vaccinated mice. Time of appearance of the first tumor and mean number ofpalpable mammary carcinoma/mouse (c) in the group of 35 mice vaccinated with Ad5-HER2 (filled circles) or in the group of 14 mice injectedwith saline solution (open circles) are reported. The mean number of palpable mammary carcinomas/mouse was calculated as the cumulativenumber of incident tumors/number of tumor-bearing neuT mice.
74 GALLO ET AL.

detected when neuT mice were injected with Ad5-wt virus accord-ing to the same protocol, indicating that Ad5-wt immunization didnot have any protective effect (data not shown). In the vaccinatedgroup of mice, the first tumor appeared at week 18 and by week 27there were no tumor free mice. Following the same trend, at week33, 2 out of 35 mice presented palpable tumors in 4 or 6 of themammary glands. Statistical analysis confirmed a significant re-duction in tumor-free and mean tumor number in the Ad5-HER2-vaccinated group compared to the control group of mice (p 1.2 � 10�11 and p 1.5 � 10�5, respectively). This transientinhibition of carcinogenesis displayed its maximal activity aroundweek 19–22 and rapidly faded out a few weeks later. At week 19,the highest difference between the vaccinated and control popula-tion was detected in terms of tumor-free mice (35.7% vs. 82.9%).At week 22, the largest difference between the 2 groups in meantumor number was recorded (3.2 vs. 7). Whole-mount and histo-logic comparison of the mammary glands of 17-week-old un-treated (Fig. 5a) and vaccinated (Fig. 5b) mice showed that themammary gland of untreated mice were almost entirely invaded byneoplastic epithelial cell foci coalescing to form large nodularmasses, whereas those of immunized mice displayed atypicalhyperplastic foci and in situ carcinoma near the nipple. At23 weeks of age, the neoplastic lesions in the vaccinated mice weresimilar in size to those in the 17-week-old untreated mice, but stillless numerous and confined to the nipple area (Fig. 5c). In addi-tion, some glands were occupied by large and others by only smallneoplastic masses. Immunohistochemical examination at thesetime points showed that the expression of membrane r-p185 andPCNA was similar in both groups (data not shown) and there wereno clear differences between their reactive cell infiltrates.
Persistence of immune responseAnti-r-p185-ECD antibodies were measured for each mouse at
week 22. When compared to data obtained at week 12, the geo-metric mean of the titers measured at week 22 is reduced by 50%(Fig. 6; p 1.2 � 10�4). However, no correlation was foundbetween the antibody titers at week 12 or at week 22 and the timesof tumor appearance and the rate of tumor growth.
A distinct group of neuT mice was immunized at week 6 and 9with Ad5-HER2 and mice were sacrificed at week 22 and spleencells analyzed by ICS. Data reported in Table III showed that forboth human and rat CD8� peptides, a halved frequency of T cellwas detected at week 22.
FIGURE 5 – Evaluation of tumor growth by whole-mount analysis.Mammary glands obtained from groups of 3 untreated or immunizedwith Ad5-HER2. neuT mice were analyzed at 17 and 23 weeks of age.At 17 weeks of age, the glands of the vaccinated group (b) show a clearreduction of neoplastic lesions compared to those present in untreatedmice (a). At 23 weeks of age, the neoplastic lesions confined in thearea close to the nipple (arrow) in vaccinated mice (c) appear enlarged,reaching dimensions comparable to the nodular masses present in17-week-old untreated mice.
FIGURE 6 – Follow-up of anti-p185 humoral response in HER2-vaccinated neuT mice. Titers of individual animals (empty circles) andcohort geometric mean titers (black line) of neuT mice at 3 (12 weeksof age) and 11 weeks after boosting (22 weeks of age) are reported.Data are expressed according to the standard curve generated usingmouse anti-p185 mAb Ab-2. All samples were run in triplicate. Thebackground absorbance of secondary antibody was typically less than0.1 A405-A620 and was subtracted from each sample.
75ANTI-r-p185 XENOGENEIC IMMUNIZATION

Discussion
In this work, the immune response elicited in the neuT trans-genic mice by gene immunization was quantitatively character-ized. Through extensive peptide scanning analysis, we mapped theimmunodominant CD8� and CD4� epitopes within the p185protein in the H-2d background of neuT mice. A CD8� nonamersequence identified by IFN-� ELISPOT and IFN-� ICS was alsoacting as a target for CTL activity. This epitope coincides with thatdescribed by Nagata et al.27 for the human protein. This nonamersequence also contains an additional overlapping CD8� epitope.We identified an additional CD8� epitope in the intracellulardomain of the human protein as well as a weak CD4� epitope inthe ECD. These CD8� and CD4� epitopes have not been previ-ously reported.
As the human and the rat proteins share 89% sequence similar-ity, we tested whether the sequences of the r-p185 correspondingto the p185 epitopes also contain T-cell epitopes. The nonamersequence of the major CD8� epitope in p185 has 2 amino acidchanges in the r-p185. These variations reduce the crossreactivityof T cells elicited by the Ad5-HER2 vaccination toward the ratepitope. However, this epitope is still scored positive by bothELISPOT and ICS analysis and displays CTL activity in neuTmice. These data demonstrate that immunizing with Ad5-HER2elicited a remarkable response against the nonself human proteinand that the same response breaks tolerance in rat neuT mice. Theoverlapping CD8� epitope did not crossreact with the correspond-ing rat peptide. For the other r-p185 sequences corresponding tothe epitopes in the p185 protein, changes of 4 out of 9 (neu-301)or 4 out of 15 (neu-41) amino acids completely abolished reactiv-ity. We also tested with negative results a number of peptidescorresponding to epitopes described in literature27 or predicted bycomputer algorithm in r-p185 (data not shown).
In several cases, a xenogeneic immunization was reported to bevery effective in breaking tolerance and inducing T cells andantibodies crossreacting with the self-protein.8,28 We used theHER2 gene to target the r-p185 protein, which drives mammarycarcinogenesis in neuT mice. We show that immunizing with Ad5expressing the HER2 gene had a modest but significant inhibitoryeffect on spontaneous tumor development in mice, which peakedaround week 19–22. The morphologic data confirmed that vacci-nation greatly slowed tumor growth at week 17. At week 23, the
same analysis detected a delayed tumor development, resemblingthe morphologic scenario displayed by control mice at week 17. Asreported by other groups, at both times we did not detect a cleardifference in terms of tumor infiltrating reactive cells between thevaccinated and the control group.17,23
We measured a remarkable humoral response against the humanprotein, which crossreacted with the rat homolog. However, we didnot find any statistically significant correlation between the titer ofantibodies and the rate of tumor development for each mouse.Immunization also induced a weak r-p185-specific CMI response,as indicated by the frequency of IFN-� �, CD8� T cells and by theCTL activity. This response, which declined with time, was likelynot sufficient to exert any major effect on tumorigenesis. Correla-tion with protection is further hampered by the fact that CMIresponse and tumor progression were monitored on differentgroups of mice treated according to the same protocol.
Recent data in literature have demonstrated that intramuscularinjection of a plasmid encoding an antigen, followed by electricalstimulation treatment in the form of high-frequency, low-voltageelectric pulses, enhances B- and T-cell-immune responses inducedby genetic vaccination.26 We showed that i.m. injection of p185expression plasmid followed by ES induced a weak CMI response,as measured by ELISPOT analysis. The immunization efficiencywas not significantly improved by increasing the amount of in-jected plasmid, increasing the number or the frequency of theinjections or combining pVIJ-HER2 with pVIJ-neu injections.
Adenovirus-mediated immunization induced a much higherresponse than plasmid DNA followed by ES. This response,however, declined with time, indicating that additional immu-nizations are required to induce a long lasting immunity. How-ever, Ad5 is no longer effective because of neutralizing anti-bodies raised by previous injections (data not shown). Thus, amixed modality where Ad5-induced response is sustained by aseries of plasmid DNA injections is likely to be an effectivevaccination strategy.29,30
Acknowledgements
The authors thank their colleagues of the Cancer Vaccine Groupfor helpful discussion and J. Clench for revision of the text.
References
1. Zhang L, Conejo-Garcia JR, Katsaros D, Gimotty PA, MassobrioM, Regnani G, Makrigiannakis A, Gray H, Schlienger K, LiebmanMN, Rubin SC, Coukos G. Intratumoral T cells, recurrence, andsurvival in epithelial ovarian cancer. N Engl J Med 2003;348:203–13.
2. Knutson KL, Schiffman K, Disis ML. Immunization with a HER-2/neu helper peptide vaccine generates HER- 2/neu CD8 T-cell immu-nity in cancer patients. J Clin Invest 2001;107:477–84.
3. Hodge JW, Grosenbach DW, Aarts WM, Poole DJ, Schlom J. Vaccine
therapy of established tumors in the absence of autoimmunity. ClinCancer Res 2003;9:1837–49.
4. Dupuis M, Denis-Mize K, Woo C, Goldbeck C, Selby MJ, Chen M,Otten GR, Ulmer JB, Donnelly JJ, Ott G, McDonald DM. Distributionof DNA vaccines determines their immunogenicity after intramuscu-lar injection in mice. J Immunol 2000;165:2850–8.
5. Aarts WM, Schlom J, Hodge JW. Vector-based vaccine/cytokinecombination therapy to enhance induction of immune responses to aself-antigen and antitumor activity. Cancer Res 2002;62:5770–7.
TABLE III – PERSISTENCE OF ANTI-p185 AND r-p185 CELL-MEDIATED IMMUNE RESPONSE INAd5-HER2-VACCINATED MICE
Peptide
12 weeks of age1 22 weeks of age
BALB/c neuT BALB/c neuT
CD4� CD8� CD4� CD8� CD4� CD8� CD4� CD8�
HER2-15.3 0.22 48.78 0.39 47.43 0.57 22.35 0.64 38.18neu-15.3 0.07 6.38 0.20 4.60 0.42 3.69 0.30 2.59neu-41 0.35 0.23 0.36 0.24 0.56 0.45 0.32 0.28HER2-301 0.24 3.04 0.17 5.71 0.22 1.79 0.13 2.75DMSO 0.10 0.12 0.18 0.17 0.30 0.23 0.20 0.19SEB 1.04 2.07 1.11 1.56 1.98 2.89 1.10 2.551Values refer to the frequency of IFN-�-secreting, CD4� or CD8� T cells measured by ICS. Displayed
data are representative of several independent experiments. Values are expressed as 1,000 � [(IFN-��CD3� and CD4� or CD8�)/(CD3� and CD4� or CD8�)]. Values higher than 1‰ were consideredpositive and indicated in boldface.
76 GALLO ET AL.

6. Hung MC, Lau YK. Basic science of HER-2/neu: a review. SeminOncol 1999;26(4 Suppl 12):51–9.
7. Disis ML, Schiffman K. Cancer vaccines targeting the HER2/neuoncogenic protein. Semin Oncol 2001;28(6 Suppl 18):12–20.
8. Wei YQ. Immunotherapy of tumors with vaccines based on xenoge-neic homologous molecules. Anticancer Drugs 2002;13:229–35.
9. Hawkins WG, Gold JS, Dyall R, Wolchok JD, Hoos A, Bowne WB,Srinivasan R, Houghton AN, Lewis JJ. Immunization with DNAcoding for gp100 results in CD4 T-cell independent antitumor immu-nity. Surgery 2000;128:273–80.
10. Lucchini F, Sacco MG, Hu N, Villa A, Brown J, Cesano L, Mangia-rini L, Rindi G, Kindl S, Sessa F, et al. Early and multifocal tumorsin breast, salivary, harderian and epididymal tissues developed inMMTY-Neu transgenic mice. Cancer Lett 1992;64:203–9.
11. Di Fiore PP, Pierce JH, Kraus MH, Segatto O, King CR, AaronsonSA. erbB-2 is a potent oncogene when overexpressed in NIH/3T3cells. Science 1987;237:178–82.
12. Nanni P, Pupa SM, Nicoletti G, De Giovanni C, Landuzzi L, Rossi I,Astolfi A, Ricci C, De Vecchi R, Invernizzi AM, Di Carlo E, MusianiP, et al. p185(neu) protein is required for tumor and anchorage-independent growth, not for cell proliferation of transgenic mammarycarcinoma. Int J Cancer 2000;87:186–94.
13. Tzahar E, Pinkas-Kramarski R, Moyer JD, Klapper LN, Alroy I,Levkowitz G, Shelly M, Henis S, Eisenstein M, Ratzkin BJ, Sela M,Andrews GC, et al. Bivalence of EGF-like ligands drives the ErbBsignaling network. EMBO J 1997;16:4938–50.
14. Kozak M. An analysis of 5�-noncoding sequences from 699 vertebratemessenger RNAs. Nucl Acids Res 1987;15:8125–48.
15. Kozak M. At least six nucleotides preceding the AUG initiator codonenhance translation in mammalian cells. J Mol Biol 1987;196:947–50.
16. Montgomery DL, Shiver JW, Leander KR, Perry HC, Friedman A,Martinez D, Ulmer JB, Donnelly JJ, Liu MA. Heterologous andhomologous protection against influenza A by DNA vaccination:optimization of DNA vectors. DNA Cell Biol 1993;12:777–83.
17. Rovero S, Amici A, Carlo ED, Bei R, Nanni P, Quaglino E, PorceddaP, Boggio K, Smorlesi A, Lollini PL, Landuzzi L, Colombo MP, et al.DNA vaccination against rat her-2/Neu p185 more effectively inhibitscarcinogenesis than transplantable carcinomas in transgenic BALB/cmice. J Immunol 2000;165:5133–42.
18. Youil R, Toner TJ, Su Q, Casimiro D, Shiver JW, Chen L, Bett AJ,Rogers BM, Burden EC, Tang A, Chen M, Emini EA, et al. Com-parative analysis of the effects of packaging signal, transgene orien-tation, promoters, polyadenylation signals, and E3 region on growthproperties of first-generation adenoviruses. Hum Gene Ther 2003;14:1017–34.
19. Fallaux FJ, Bout A, van der Velde I, van den Wollenberg DJ, HehirKM, Keegan J, Auger C, Cramer SJ, van Ormondt H, van der Eb AJ,Valerio D, Hoeben RC. New helper cells and matched early region1-deleted adenovirus vectors prevent generation of replication-com-petent adenoviruses. Hum Gene Ther 1998;9:1909–17.
20. Ishibashi M, Maizel JV Jr. The polypeptides of adenovirus: V, young
virions, structural intermediate between top components and agedvirions. Virology 1974;57:409–24.
21. Chakrabarti S, Brechling K, Moss B. Vaccinia virus expression vec-tor: coexpression of beta-galactosidase provides visual screening ofrecombinant virus plaques. Mol Cell Biol 1985;5:3403–9.
22. Fu TM, Mylin LM, Schell TD, Bacik I, Russ G, Yewdell JW, BenninkJR, Tevethia SS. An endoplasmic reticulum-targeting signal sequenceenhances the immunogenicity of an immunorecessive simian virus 40large T antigen cytotoxic T-lymphocyte epitope. J Virol 1998;72:1469–81.
23. Boggio K, Nicoletti G, Di Carlo E, Cavallo F, Landuzzi L, Melani C,Giovarelli M, Rossi I, Nanni P, De Giovanni C, Bouchard P, Wolf S,et al. Interleukin 12-mediated prevention of spontaneous mammaryadenocarcinomas in two lines of Her-2/neu transgenic mice. J ExpMed 1998;188:589–96.
24. Rizzuto G, Cappelletti M, Maione D, Savino R, Lazzaro D, Costa P,Mathiesen I, Cortese R, Ciliberto G, Laufer R, La Monica N, FattoriE. Efficient and regulated erythropoietin production by naked DNAinjection and muscle electroporation. Proc Natl Acad Sci USA 1999;96:6417–22.
25. Miyahira Y, Murata K, Rodriguez D, Rodriguez JR, Esteban M,Rodrigues MM, Zavala F. Quantification of antigen specific CD8� Tcells using an ELISPOT assay. J Immunol Methods 1995;181:45–54.
26. Zucchelli S, Capone S, Fattori E, Folgori A, Di Marco A, Casimiro D,Simon AJ, Laufer R, La Monica N, Cortese R, Nicosia A. EnhancingB- and T-cell immune response to a hepatitis C virus E2 DNA vaccineby intramuscular electrical gene transfer. J Virol 2000;74:11598–607.
27. Nagata Y, Furugen R, Hiasa A, Ikeda H, Ohta N, Furukawa K,Nakamura H, Kanematsu T, Shiku H. Peptides derived from a wild-type murine proto-oncogene c-erbB-2/HER2/neu can induce CTL andtumor suppression in syngeneic hosts. J Immunol 1997;159:1336–43.
28. Sioud M, Sorensen D. Generation of an effective anti-tumor immunityafter immunization with xenogeneic antigens. Eur J Immunol 2003;33:38–45.
29. Casimiro DR, Chen L, Fu TM, Evans RK, Caulfield MJ, Davies ME,Tang A, Chen M, Huang L, Harris V, Freed DC, Wilson KA, et al.Comparative immunogenicity in rhesus monkeys of DNA plasmid,recombinant vaccinia virus, and replication-defective adenovirus vec-tors expressing a human immunodeficiency virus type 1 gag gene.J Virol 2003;77:6305–13.
30. Matsui M, Moriya O, Akatsuka T. Enhanced induction of hepatitis Cvirus-specific cytotoxic T lymphocytes and protective efficacy in miceby DNA vaccination followed by adenovirus boosting in combinationwith the interleukin-12 expression plasmid. Vaccine 2003;21:1629–39.
31. Wei YQHM, Yang L, Zhao X, Tian L, Lu Y, Shu JM, Lu CJ, Niu T,Kang BMY, Liu F, Wen YJ, Lei S, et al. Immunogene therapy oftumors with vaccine based on Xenopus homologous vascular endo-thelial growth factor as a model antigen. Proc Natl Acad Sci USA2001;98:11545–50.
77ANTI-r-p185 XENOGENEIC IMMUNIZATION




















