![t] x| f} + of] hgf -cf=j](https://static.fdokumen.com/doc/165x107/631922cab41f9c8c6e099861/t-x-f-of-hgf-cfj.jpg)
x^ Bottor of $litlo!£opi)p t^ ,s^^ - CORE
293
x^ COORDINATION OF SULFUR AND NITROGEN IN THE PROTECTION OF PHOTOSYNTHESIS, GROWTH AND YIELD OF CADMIUM-TREATED MUSTARD (Brassicajuncea) \. ' ABSTRACT THESIS / / ' SUBMITTED FOR THE AWARD OF THE DEGREE OF , r Bottor of $litlo!£opi)p BOTANY ') SARVAJEET SINGH t^ ,s^^ DEPARTMENT OF BOTANY ALIGARH MUSLIM UNIVERSITY ALIGARH (INDIA) 2008
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of x^ Bottor of $litlo!£opi)p t^ ,s^^ - CORE

x^
COORDINATION OF SULFUR AND NITROGEN IN THE PROTECTION OF PHOTOSYNTHESIS, GROWTH AND YIELD OF CADMIUM-TREATED MUSTARD (Brassicajuncea)
\. '
ABSTRACT
THESIS / / '
SUBMITTED FOR THE AWARD OF THE DEGREE OF
, r
Bottor of $litlo!£opi)p
BOTANY
' )
SARVAJEET SINGH
t^ ,s ^
DEPARTMENT OF BOTANY ALIGARH MUSLIM UNIVERSITY
ALIGARH (INDIA)
2008

# . '>^
. ^

Coordination of sulfur and nitrogen in ttie protection of
photosynthesis, growth and yield of cadmium-treated
mustard (Brassicajuncea)
Abstract
Sarvajeet Singh
Abstract of the thesis submitted to the Aligarh Muslim University, Aligarh, India for
the degree of Doctor of Philosophy in Botany.
The present thesis comprises of six chapters.
The importance of the problem and justifications for the present work undertaken
were emphasized in Chapter 1.
Chapter 2 is the review of literature. It includes literature available on the
aspects of the physiological analysis of various growth-, photosynthetic- and,
biochemical-characteristics, antioxidant studies and yield of various crop plants under
cadmium stress. The importance of sulfur and nitrogen nutrients in the alleviation of
stress and in the regulation of plant growth and development under stress condition
were also reviewed. The chapter has been divided in to sections and sub-sections for
better understanding of the work of other research reports in this field of study. In
addition, the critical appraisal of the review of literature has also been included to
identify the gap in the field of study.
The details of the material used in the study and methodology adopted to
determine various characteristics recorded in the three experiments have been
described in the Chapter 3. In addition the Chapter 3 also mentions the relevant
information on the location of the study and the environmental conditions during the
data sampling times.
Chapter 4 includes the results on the response of mustard to cadmium, sulfur
and nitrogen treatments in three Experiments. The data obtained were statistically
analyzed and the significance was determined at/7<0.05.
The results obtained in the Experiments have been discussed in Chapter 5 in
the light of observations recorded and supported with the earlier findings, if available.
This chapter also presents the possible explanations of the data obtained to reach a
conclusion and possible future prospects.

Chapter 6 present the summary of the work reported in this thesis.
In the following pages a brief account of the importance of the study
undertaken, results obtained and conclusion are given.
Importance of the study undertaken
Abiotic stresses such as salinity, heavy metals, cold, drought and heat are the primary
cause of crop loss worldwide and the relative decreases in potential maximum yields
vary between 50-80% (Bray et al. 2000). Among abiotic stresses, heavy metal
contamination is a serious environmental problem that limits plant productivity and
threatens human health (Wagner 1993, Sanita di Toppi and Gabbrielli 1999, Khan and
Singh 2008). Cadmium (Cd) is of special concern due to its wide availability and
potential toxicity to biota even at low concentrations (Das et al. 1997, Sanita di Toppi
and Gabbrielli 1999). It is released into the environment by power stations, heating
systems, metal working industries, waste incinerators, urban traffic, cement factories
and as a byproduct of sewage sludge and phosphatic fertilizers (Sanita di Toppi and
Gabbrielli 1999, Lombi et al 2000, Nolan et al. 2003, McLaughlin et al. 2006,
Sharma and Dudey 2006, Xie et al. 2006, Verma et al. 2007, Singh et al. 2008).
For sustainable crop production, it is important to develop methods or
techniques for alleviating the Cd-induced growth inhibition and reducing its
accumulation in plants. Increasing evidence suggests that mineral-nutrient status of
plants plays a critical role in increasing plant resistance to environmental stresses
(Marschner 1995, Vassilev et al. 2005, Anjum et al. 2008a,b, Hassan et al. 2008,
Khan et al. 2008). The use of sulfur (S) and nitrogen (N) fertilizers opens potential
option which can be used to alleviate Cd toxicity. Sulfiir is a structural constituent of
several coenzymes and prosthetic groups, such as ferredoxin, which are also
important for N assimilation. Thus, S plays an important role in plant growth and in
the regulation of plant development (Ernst et al. 2008). It has been found that S
nutrition is a critical factor for the alleviation of Cd toxicity (Popovic et al. 1996,
Chen and Huerta 1997, Hassan et al. 2005a, Vassilev et al. 2005, Anjum et al. 2008a,
b). Further, N is an important component of many structural, genetic and metabolic
compounds in plant cells. Plant N status is highly dependent on N fertilization
(Hernandez et al. 1997). Nitrogen is also a major component of chlorophyll and
amino acids, the building blocks of proteins. Increase in N supply can stimulate plant
growth and productivity (Joel et al. 1997) as well as photosynthetic activity (Makino
et al. 1992) through increased amounts of stromal and thylakoid proteins in leaves
f U 31?

(Bungard et al. 1997). Pankovic et al. (2000) have shown that optimal N supply
decreased the inhibitory effects of Cd on photosynthesis of Helianthus annuus. A
proper N supply has been shown to have a positive effect in overcoming the adverse
effects caused by Cd toxicity in Oryza sativa (Hassan et al. 2005b). The assimilation
pathways of S and N are well coordinated (Brunold 1993, Takahashi and Saito 1996,
Abdin et al. 2003, Hawkesford et al. 2006). Therefore, coordination of S and N could
play a prominent role in alleviating the Cd-induced toxicity in crop plants.
The oleiferous Brassica is the third most important source of vegetable oil in
the world after palm and soybean oil and grown as an edible or an industrial oil crop
which is used as a source of edible protein, in much the same way as soybean protein
(Ahmad and Abdin 2000, Zhang et al. 2003). The major mustard oil-producing
countries include Canada, China, France, Germany, India and UK. According to a
report of United States Department of Agriculture (USDA), the world oilseed
production is 397metric tons in 2006-07. This total production is an increase of
9metric tons, or 2% of the last season. Indian agriculture contributes about 15% and
8% to the world total acreage under oilseed cultivation and production, respectively.
However, the productivity in India is only 791kg ha"' as compared to the world
average of 1718kg ha'' (Damodaran and Hedge 2002). About 90% of the total land
under oilseed cultivation in India is occupied by Brassica juncea (Khan et al. 2007).
Despite a large area under cultivation of mustard, the productivity of the crop has
dropped because of anthropogenic activities emitting heavy metals in the
environment.
Discussed the importance of sulftir and nitrogen in plant metabolism and their
role in stress tolerance it was assumed that mustard (Brassica juncea L. Czem &
Coss.) may serve as a model crop in the study of the influence of S and N in plant
metabolism under Cd stress. Further, the fertilization of crop with balanced S and N
may prove fruitftil in alleviating Cd stress and improving crop productivity. Studies
on crops with a view of alleviating Cd stress with balanced S and N application have
not been carried out. Therefore, the reported research was undertaken with the
following objectives:
• To screen and select mustard cultivars as Cd tolerant and Cd non-tolerant
grown under Cd treatments.

• To study the influence of S application in the alleviation of Cd-induced effects
in Cd tolerant and Cd non-tolerant cultivars of mustard.
• To study the combined influence of S and N in the alleviation of Cd-induced
effects in Cd tolerant and Cd non-tolerant cultivars of mustard.
Experimental Results
The results of the experiments are summarized below: Experiment 1
The Experiment was conducted to study the effect of five concentrations of Cd viz., 0,
25, 50, 100 and I50mg kg"' soil on Cd accumulation in root and leaf, growth,
photosynthetic and yield characteristics of five mustard cultivars {Brassica juncea L.)
namely, Alankar, Varuna, Pusa Bold, Sakha and RH30. The treatments in this
Experiment were arranged in a factorial randomized block design. Cadmium
accumulation, growth and photosynthetic characteristics were studied at pre-flowering
(30DAS), flowering (60DAS) and post-flowering (90DAS) growth stages, while yield
characteristics were noted at harvest (120DAS). Tolerance index of the five cultivars
of mustard was calculated and the cultivars were designated as Cd tolerant and Cd
non-tolerant on the basis of their performance under Cd stress. The effect of Cd on
growth, photosynthetic and yield characteristics was found significant at all sampling
times. The interaction of Cd treatments and cultivars was also found significant. For
growth characteristics, the interaction effect was found non-significant at 30 DAS.
The increase in Cd levels decreased the growth, photosynthetic and yield
characteristics of all the five cultivars at all sampling stages. The observations
recorded at the three sampling stages showed similar pattern of cultivar response to
Cd treatments. Maximum reduction in growth, photosynthesis and yield
characterisfics was noted with 150mg Cd kg'' soil followed by 100, 50 and 25mg Cd
kg"' soil.
Among cultivars, Alankar showed lesser decrease in growth, photosynthetic
and yield characteristics followed by Varuna and Pusa Bold, whereas, RH30 and
Sakha exhibited greater reduction in growth characteristics under Cd stress. The
tolerance index of cultivars was: Alankar >Varuna >Pusa Bold >Sakha >RH30.
Experiment 2
Experiment 2 was conducted on the basis of findings of Experiments 1. As observed
in Experiment 1, Alankar emerged as Cd-tolerant and R_H30 as Cd non-tolerant
cuhivar. Among Cd levels, 150mg Cd kg"' soil was found most toxic and caused
4 ' ti.'.*" *'

maximum reductions in the characteristics studied. In the present Experiment, the aim
was to study the alleviation potential of elemental S levels (50 and lOOmg S kg' soil)
on the effects of 50 and 150mg Cd kg"' soil. The treatments in this Experiment were
arranged in a factorial randomized block design. The assessment was done by
studying the changes in Cd accvmiulation in root and leaf, growth and photosynthetic
characteristics, S and N assimilation, components of antioxidant defense system and
yield characteristics of tolerant (Alankar) and non-tolerant (RH30) cultivars. The
sampling stages for Cd accumulation, growth and photosynthetic characteristics, S
and N assimilation, components of antioxidant defense system were pre-flowering
(30DAS), flowering (60DAS) and post-flowering (90DAS). The yield characteristics
were noted at harvest (120DAS).
The growth and photosynthetic characteristics and N assimilation decreased
maximally in both the cultivars treated with 150mg Cd kg"' soil. Contrarily, there was
significant increase in the Cd accumulation, S assimilation, H2O2 and TBARS content
and components of enzymatic antioxidant system and glutathione content was found.
Non-tolerant cultivar RH30 exhibited greater Cd effects than tolerant cultivar
Alankar. In comparison to lOOmg S kg"' soil, the application of 50mg S kg"' soil
maximally alleviated the toxic effect of 50mg Cd kg"' soil and improved growth,
photosynthesis, S and N assimilation and components of enzymatic and non-
enzymatic antioxidant system thus yield characteristics of both the cultivars. Further,
no ameliorative effect of lOOmg S kg"' soil was observed in plants treated with 150mg
Cd kg"' soil. Alleviation effects of S were higher in Alankar than RH30.
Experiment 3
Experiment 3 was conducted on the basis of the findings of Experiments 2. In
Experiment 2, it was observed that the application of 50mg S kg"' soil to plants treated
with 50mg Cd kg"' soil alleviated Cd-induced toxicity in both the culdvars.
Supplementafion of 50mg S kg"' soil maximally overcome the toxic effects of 50mg
Cd kg"' soil in Alankar (tolerant cultivar), while, the same combination lowered the
Cd-induced toxic effects in RH30 to some extent. The present experiment was aimed
to study the effect of supplementation of different N levels (40, 80 and 120mg N kg"'
soil) to 50mg S kg"' soil for alleviation of adverse effects of 50mg Cd kg"' soil. The
treatments in this Experiment were arranged in a factorial randomized block design.
The assessment was done by analyzing the changes in Cd accumulation in root and

leaf, growth and photosynthetic characteristics, S and N assimilation, components of
antioxidant defense system at 30, 60 and 90DAS and yield characteristics at harvest.
The alterations in growth, photosynthesis, S and N assimilation, components
of antioxidant system and yield characteristics caused by 50mg Cd kg" soil were
alleviated by 50mg S kg"' soil in tolerant (Alankar) and non-tolerant (RH30) cultivars,
but the alleviation potential of S varied between the cultivars. Application of different
levels of N (40, 80 and 120mg N kg"' soil) further enhanced the alleviation effect of S
(50mg S kg"' soil) in both the cultivars. Among the N levels used, the application of
80mg N kg'' soil proved superior than the other N levels. A combined application of S
and N (50mg S kg"' soil plus SOmg N kg"' soil) not only ameliorated the Cd-induced
(50mg Cd kg"' soil) effects but also increased the growth, photosynthesis, S and N
assimilation and components of enzymatic and non-enzymatic antioxidant system
thus yield characteristics in Alankar. In RH30, this combination (SOmg S kg"' soil
plus SOmg N kg"' soil) only lowered the adverse effects of Cd.
Conclusion
Conclusively, nutrients are known to decipher essential role in plant metabolism and
augmenting growth and productivity of crops. However, a nutrient package is
required for sustainable agriculture under varied environmental conditions. Crop
cultivars display their inherent potential. The cultivar Alankar surpassed other
cultivars tested and tolerated Cd stress to a significant degree. The cultivar RH30 was
weak in performance and least tolerant to Cd among the cultivars tested. Therefore,
Alankar exhibited lesser decreases in growth, photosynthetic and yield characteristics
under Cd stress. Correspondingly, Alankar also showed lesser oxidative stress and
increased antioxidant system than RH30 to protect photosynthetic machinery and
consequent effects on other attributes. Sulfur proved significant potential in the
alleviation of Cd stress in both the cultivars. The decreases in the characteristics
observed due to Cd stress were lowered by S application and SOmg S kg"' soil proved
effective in alleviation of Cd stress than the high doses of S. The coordination of S
and N proved most effective in nullifying the Cd stress effects. The potential of SOmg
S kg'' soil in the alleviation of Cd stress was substantially enhanced by the
simultaneous application of SOmg N kg"' soil. The combined application of SOmg S
kg" soil plus SOmg N kg"' soil not only resulted in restricting the decrease caused by
Cd but also nullified the Cd effects and even enhanced the characteristics values over
the control. A package of SOmg S kg"' soil plus SOmg N kg"' soil appears to be most
6

effective in the cultivation of mustard under Cd stress. The effect of this combination
was due to coordination of S and N in maintaining plant metabolism and alleviating
Cd stress.
New Report in the Thesis and Future Prospects
Mineral nutrient status of plants plays a critical role in increasing plant resistance to
environmental stress factors. Of the mineral nutrients, S and N are major
macronutrients necessary for the plant life cycle. The uptake and assimilation of S and
N in higher plants are the crucial factors determining plant growth and vigor and crop
yield. The present study showed that a balanced S and N supply can mitigate the
inhibitory effects of Cd on mustard metabolism. The individual effects of S and N and
other nutrients have been studied. However, no effort has been made to study
combined application of S and N in tolerance of plants, although coordinated
functions of these nutrients are well known. For sustainable agriculture a nutrient
protocol with balanced S and N is necessary in the growth and development of crops
under Cd stress. The results reported in this thesis have confirmed this. The effect of
combined application of S and N on oxidative stress and antioxidants under Cd stress
has been reported for the first time. Efforts have been made for the first time to
develop S and N protocol for alleviation of Cd stress in mustard.
Further, the efforts should be focused to dissect the mechanism(s) of
regulation of sulfate and nitrate assimilation at the molecular level in crop plants
under varied environmental conditions. Strategies should also aim to manipulate steps
of pathways leading to the production of thiols and their products in plants through
overexpressing Ser acetyl transferase (SAT), y-ECS and GSHS enzymes of S and N
nutrition under Cd stress. More detailed studies are required to understand the Cd-
induced stress response modulated by S and N metabolism at molecular and
mechanistic levels. This would help to develop an effective strategy to raise
transgenic species for stress resistance.
Bibliography
Abdin MZ, Ahmad A, Khan N, Khan I, Jamal A, Iqbal M (2003) Sulphur interaction
with other nutrients. In: Abrol YP, Ahmad A (Eds) Sulphur in plants. Kluwer
Academic Publishers, The Netherlands, pp359-374.
Ahmad A, Abdin MZ (2000) Effect of sulphur application on lipid, RNA, protein
content and fatty acid composition in developing seeds of rapeseed {Brassica
campesths L). Plant Sci 150:71-75.
7

Anjum NA, Umar S, Ahmad A, Iqbal M, Khan NA (2008a) Sulphur protects mustard
(Brassica campestris L.) from cadmium toxicity by improving leaf ascorbate and
glutathione. Plant Growth Regul 54:271-279.
Anjum NA, Umar S, Singh S, Nazar R, Khan NA (2008b) Sulfur assimilation and
cadmium tolerance in plants. In: Khan NA, Singh S, Umar S (Eds) Sulfur
assimilation and abiotic stresses in plants. Springer-Verlag, The Netherlands,
pp271-302.
Bray EA, Bailey-Serres J, Weretilnyk E (2000) Responses to abiotic stresses. In:
Gruissem W, Bucharman B, Jones R (Eds) Biochemistry jmd molecular biology of
plants. Am Soc Plant Biol, Rockville, MD, ppl 158-1249.
Brunold C (1993) Regulatory interactions between sulfate and nitrate assimilation. In:
de Kok LJ, Stulen I, Renenberg H, Brunold C, Rauser W (Eds) Sulfur nutrition
and sulfur assimilation in higher plants: regulatory, agricultural and environmental
aspects, SPB Academic PubHshing, The Hague, ppl25-138.
Bungard RA, McNeil D, Morton JD (1997) Effects of nitrogen on the
photosynthetic apparatus of Clematis vitalba grown at several irradiances.
Austr J Plant Physiol 24:205-214.
Chen Y, Huerta AJ (1997) Effects of sulfur nutrition on photosynthesis in
cadmium-treated barley seedlings. J Plant Nutr 20:845-856.
Damodaran T, Hegde DM (2002) Oilseeds situation: A statistical compendium,
Directorate of oilseeds research, Indian council of agricultural research,
Hyderabad, India.
Das P, Samantaray S, Rout GR (1997) Studies on cadmium toxicity in plants: A
review. Environ Pollut 98:29-36.
Ernst WHO, Krauss G-J, Verkleij JAC, Wesenberg D (2008) Interaction of heavy
metals with the sulphur metabolism in angiosperms from an ecological point
of view. Plant Cell Environ 31:123-143.
Hassan MJ, Wang Z, Zhang G (2005a) Sulfur alleviates growth inhibition and
oxidative stress caused by cadmium toxicity in rice. J Plant Nutr 28:1785-1800.
Hassan MJ, Wang F, Ali S, Zhang G (2005b) Toxic effect of cadmium on rice as
affected by nitrogen fertilizer form. Plant Soil 277:359-365.
^ ^

Hassan MJ, ShafiM, Zhang G, Zhu Z, Qaisar M (2008) The growth and some
physiological responses of rice to Cd toxicity as affected by nitrogen form. Plant
Growth Regul 54:125-132.
Hawkesford MJ, Howarth JR, Buchner P (2006) Control of sulfur uptake,
assimilation and metabolism. In: WC Plaxton WC, McManus MT (Eds)
Control of primary metabolism in plants. Annual Plant Reviews, Vol 22,
Blackwell Publishing, Oxford, pp348-372.
Hernandez E, Olguin E, Trujillo SY, Vivanco J (1997) Recycling and treatment of
anaerobic effluents from pig waste using Lemna sp. under temperate climatic
conditions. In: Wise DL (Ed) Global enviroimiental biotechnology. Elsevier, The
Netherlands, pp293-304.
Joel G, Gamon JA, Field CB (1997) Production efficiency in sunflower: The role of
water and nitrogen stress. Remote Sensing Environ 62:176-188.
Khan NA, SamiuUah, Singh S, Nazar R (2007) Activities of antioxidative enzymes,
sulphur assimilation, photosynthetic activity and growth of wheat {Triticum
aestivum) cultivars differing in yield potential under cadmium stress. J Agron
Crop Sci 193:435-444.
Khan NA, Singh S (2008) Abiotic stress and plant responses. IK international
publishing house, New Delhi, India.
Khan NA, Singh S, Umar S (2008) Sulfur assimilation and abiotic stress in plants.
Springer-Verlag, Berlin, Heidelberg, Germany.
Lombi E, Zhao FJ, Dunham S, McGrath SP (2000) Cadmium accumulation in
populations of Thlaspi caerulescens and Thlaspi goesingense. New Phytol 145:11-
20.
Makino A, Sakashita H, Hidema J, Mae T, Ojima K, Osmond B (1992) Distinctive
responses of ribulose-1,5- bisphosphate carboxylase and carbonic anhydrase in
wheat leaves to nitrogen nutrition and their possible relationships to CO2 transfer
resistance. Plant Physiol 100:1737-1743.
Marschner H (1995) Mineral nutrition of higher plants. Academic Press, New York.
McLaughlin MJ, Hamon RE, McLaren RG, Speir TW, Rogers SL (2000)
Review: A bioavailability-based rationale for controlling metal and metalloid
contamination of agricultural land in Australia and New Zealand. Austr J Soil
Res 38:1037-1086.

Nolan AL, McLaughlin MJ, Mason SD (2003) Chemical speciation of Zn, Cd, Cu,
and Pb in pore waters of agricultural and contaminated soils using Donnan
dialysis. Environ Sci Technol 37:90-98.
Pankovic D, Plesnicar M, Arsenijevic-Maksimovic I, Petrovic N, Sakac Z, Kastori R
(2000) Effects of nitrogen nutrition on photosynthesis in Cd-treated sunflower
plants. Ann Bot 86:841-847.
Popovic M, Kevresan S, Kandrac J, Nicolic J, Petrovic N, Kastori R (1996) The role
of sulphur in detoxification of cadmium in young sugar beet plants. Biol Plant
38:281-287.
Sanita di Toppi L, Gabbrielli R (1999) Response to cadmium in higher plants.
Environ Exp Bot 41:105-130.
Sharma P, Dubey RS (2006) Cadmium uptake and its toxicity in higher plants.
In: Khan NA, Samiullah (Eds) Cadmium toxicity and tolerance in plants.
Narosa Publishing House, New Delhi, pp64-86.
Singh S, Anjum NA, Khan NA, Nazar R (2008) Metal-binding peptides and
antioxidant defence system in plants: significance in cadmium tolerance. In: Khan
NA, Singh S (Eds) Abiotic stress and plant responses. IK International, New
Delhi, pp 159-189.
Takahashi H, Saito K (1996) Subcellular localization of spinach cysteine synthase
isoforms and regulation of their gene expression by nitrogen and sulfur. Plant
Physiol 112:273-280.
Vassilev A, Malgozata B, Nevena S, Zlatko Z (2005) Chronic Cd toxicity of bean
plants can be partially reduced by supply of ammonium sulphate. J Central Euro
Agric 6:389-396.
Verma VK, Singh YP, Rai JPN (2007) Biogas production fi-om plant biomass used for
phytoremediation of industrial waste. Biores Technol 98:1664-1669.
Wagner GJ (1993) Accumulation of cadmium in crop plants and its consequences to
human health. Adv Agron 51:173-212.
Xie RK, Seip HM, Wibetoe G, Nori S, McLeod CM (2006) Heavy coal
combustion as the dominant source of particulate pollution in Taiyuan,
China, corroborated by high concentrations of arsenic and selenium in PMio-
Sci Total Environ 370:409-415.
Zhang FQ, Shi WY, Jin ZX, Shen ZG (2003) Response of antioxidative enzymes
in cucumber chloroplast to cadmium toxicity. J Plant Nutr 26:1779-1788.
10 , . . - . .

X rr <;\S
COORDINATION OF SULFUR AND NITROGEN IN THE PROTECTION OF PHOTOSYNTHESIS, GROWTH AND YIELD OF CADMIUM-TREATED MUSTARD (Brassi'ca juiicea)
i • \
/ / THESIS
SUBMITTED FOR THE AWARD OF THE DEGREE OF
II Bottor of $t)ilosiop[)p IN / /
\ \ A < BOTANY
SARVAJEET SINGH
iS^ DEPARTMENT OF BOTANY
ALIGARH MUSLIM UNIVERSITY ALIGARH (INDIA)
2008

< : #
f e d U. CoraiJUi C<8it:x
2 7 JAN d
T6547

Dedicated to my father.
< ^ s\5

<Dr. fNafeesA- Xfian ^Pmfessof of (plant Hysiotogy
Department of Botany Aligarh Muslim University Aligarli-202 002, INDIA Off: +91-571-2702016 Mobile:+91-9411488881 E-mail: naf9(^lycos.com
Dated: ^t)- Z> ^OO^
CERTIFICATE
This is to certify that the thesis entitled, '"Coordination of sulfur and
nitrogen in the protection of photosynthesis, growth and yield of
cadmium-treated mustard (Brassica juncea)" submitted for the degree
of Doctor of Philosophy in Botany is a faithful record of bonafide
research work carried out at the Department of Botany, Aligarh
Muslim University, Aligarh by Mr. Sarvajeet Singh under my
guidance and supervision and that no part of it has been submitted for any
other degree or diploma.
i ^ : . \ ^
(P fafees A. Khan)

ACKNOWLEDGEMENTS
I bow in reverence to GOD the Almighty, Who blessed me with an innumerable
favour of academic work.
Like all creative works, completing a thesis in any discipline is really a hard
work to accomplish. I have no words to express my heartfelt gratitude to my
supervisor Professor Nafees A. Khan, Department of Botany, Aligarh Muslim
University, Aligarh for his excellent guidance, valuable and timely suggestions,
stimulating discussions and academic alertness, sense of perfection and precision
which enabled me to complete this work. His affectionate instinct and constant
encouragement were always been boon to me. Without his unending support and
encouragement this thesis would have never been completed.
I am extremely obliged to Professor Ainul H. Khan, Chaimian, Department of
Botany for providing necessary research facilities. Thanks in this regard are also due
to Ex-Chairman Professor SamiuUah.
I have always been inspired by my respected teachers. I duly acknowledge
their moral support. I am also thankful to my friend Ms. Priya and lab colleagues
Ms. Rabat Nazar and Ms. Noushina Iqbal for the help they extended whenever
required.
I also place on record the assistance rendered by Dr. Naser A. Anjum,
Dr. Moinudddin Khan, Dr. Asgar Ali Shah, Dr. Lamabam Peter Singh, Dr. Sandeep
Sharma, Dr. Naeem, Dr. Jarrar Ahmad, Dr. Shuaib Khan, Dr. Riyasat Ali and
Mr. Rais A. Khan during the gruelling task.
I am also thankful to the Council of Scientific and Industrial Research,
Government of India, New Delhi for providing financial assistance as Senior
Research Fellowship to me.
Finally, I would like to express my greatest gratitude to people who gave me
the greatest encouragement and motivation to carry out my research work; my
parents, paternal aunt Surjit, brothers Gurvinder, Dev and Ranjeet and sister Jasbeer.
(Sarvajeet Singh)

ABBREVIATIONS
%
^g Ag ALA AsA As ATP ATP-S CA Car CAT Cd CdCb CdBP Chi a Chlb Chi cm Co CO2 Cr Cu/Zn-SOD cv. Cys d DAS DHAR DMSO DW EDTA FAD Fe Fe-SOD Fv/Fm FW g GDH Gly Glu GPOX GR GS GSH GSSG GST h H2O2
Percent Gravity Silver 5-aminolaevulinic acid Ascorbic acid Arsenic Adenosine triphosphate ATP-sulfiirylase Carbonic anhydrase Carotenoids Catalase Cadmium Cadmium chloride Cd-binding peptides Chlorophyll a Chlorophyll b Chlorophyll Centimeter Cobalt Carbon dioxide Cromium Copper/zinc superoxide dismutases Cultivar Cysteine Days Days after sowing Dehydroascorbate reductase Dimethyl sulphoxide Dry weight Ethylenediamine tetraacetic acid Flavin adenin dinucleotide Iron Iron superoxide dismutase Photosynthetic quantum yield Fresh weight Gram Glutamate dehydrogenase Glycine Glutamate Guaiacol peroxidase Glutathione reductase Glutamine synthetase Reduced glutathione Oxidized glutathione Glutathione S-transferase Hour Hydrogen peroxide

HEPES Hg. HMW hPC LMW M MDA MDHAR Mg mg Met min mM Mn-SOD Mo mt MT N NaOCl NADH-GDH NADPH NBT Ni NiR ng NPT NR O2 'O2 O2-°C OH-Pb PC PDM pH ppm PSI PSII ROS rpm Rubisco Sb -SH sec SOD TBARS Va liM
^g
Hydroxyethylenepiperazine ethanesulfonic acid Mercury High molecular weight Homophytochelatin Low molecular weight Molar Malondialdehyde Monodehydroascorbate reductase Magnecium Miligram Methionine Minute Millimolar Manganese superoxide dismutase Molybdenum Metric ton Metallothionein Nitrogen Sodium hypochlorite Glutamate dehydrogenase Nicotinamide adenine dinucleotide phosphate Nitroblue tetrazolium Nickel Nitrite reductase Nano gram Non-protein thiol Nitrate reductase Molecular oxygen Singlet oxygen Superoxide anion Degree celcius Hydroxyl radical Lead Phytochelatin Plant dry mass Hydrogen ion potential Parts per million Photosystem I Photosystem II Reactive oxygen species Revolutions per minute Ribulose 1,5 bisphosphate carboxylase Antimony Sulfhydryl group Second Superoxide dismutase Thiobarbituric acid reactive substances Vanadium Micro mol Micro gram

Chapters
1. Introduction
2. Review of Literature
3. Material and Methods
4. Experimental Results
5. Discussion
6. Summary
Bibliography
CONTENTS
Page No.
1
7
48
65
85
117
121

INTRODUCTION

INTRODUCTION Chapter 1
Environmental stress is referred to the extreme environmental conditions that lead to
alterations in plant metabolism resulting in decreased rate of plants growth processes,
loss in productivity or inducing damaging effect in any of plant's organ/part,
alteration in anatomical, biochemical or molecular regulation.
Abiotic stress such as salinity, heavy metal, cold, drought or heat is the
primary cause of crop loss worldwide. The relative decrease in potential maximum
yields caused by abiotic stress varies between 50-80% (Bray et al. 2000, Mahajan and
Tuteja 2005, Khan et al. 2006). Among the abiotic factors, heavy metals are
ubiquitous in the enviroimient, depending on both geological origin and
anthropogenic activities. Metal contamination to soil is considered as major threat
because of their acute toxicity, non-biodegradable nature and gradual build up of high
concentrations (Prasad 1995, Prasad and Hagemeyer 1999, Khan and Singh 2008).
Although, heavy metals at minute quantities are natural components of soil, but the
anthropogenic activities such as mining, industry, and localized agriculture have
contributed to undesirable accumulations of toxic metals in the environment (Alloway
1995, Kabata-Pendias and Pendias 2001). Heavy metal concentration in soil ranges
from traces to as high as 100,000mg kg"' soil (Blaylock and Huang 2000), depending
on the location and the type of metal.
Among heavy metals, cadmium (Cd) is of special concern due to its wide
availability and potential toxicity to biota at low concentrations (Das et al. 1997,
Sanita di Toppi and Gabbrielli 1999). It is released into the enviroimient by power
stations, heating systems, metal working industries, waste incinerators, urban traffic,
cement factories, and as a byproduct of sewage sludge and phosphatic fertilizers
(Sanita di Toppi and Gabbrielli 1999, Lombi et al. 2000, McLaughlin et al. 2000,
Nolan et al. 2003, Sharma and Dubey 2006, Xie et al. 2006, Verma et al. 2007, Singh
et al. 2008b). Cadmium in unpolluted soil is mainly originated from soil parent ores.
Globally, Cd content in soil is about 0.01~2mg kg'' soil, with an average of 0.35mg
kg'' soil (Xu and Yang 1995). The global production of Cd has increased steadily
since 1910 when Cd electroplating was developed commercially. The average armual
global production of Cd was 12000tons in 1960-1969 which increased to 15000-
20000tons in 1980-1985 (Sandra 1986).
t^^

Cadmium contamination may pose a serious hazard to plants and human
health through its uptake in food chain (Jackson and Alloway 1992, Wagner 1993,
Hail 2002). The Food and Agriculture OrganizationAVorld Health Organization
recommended 400-500^g Cd per week as a maximum tolerable intake (Fassett 1980).
Average dietary intakes of Cd around the world range between 25 and 75^g per day
(Page et al. 1981). Although Cd is not essential for plant growth, but it is readily
taken up by roots and translocated into the leaves in m ^ y plant species (Prasad
1995). Root uptake of Cd from soil depends on the Cd concentration in soil, soil pH,
level of organic matter and Zn concentration in soil (Eriksson et al. 1996, Ciecko et
al. 2001, Yu et al. 2005, Singh et al. 2008b). Soil factors affect chemical availability
of soil Cd to plant roots (Oliver et al. 1995). Most of the Cd absorbed is accumulated
in the roots and very little amount of Cd is transported from roots to shoots. Retention
of Cd in roots is advantageous as the entry of Cd into human food chain is reduced.
According to Grant et al. (1998) Cd concentration in grains reduced when Cd
translocation from roots to shoots is restricted. It has been shown that Cd toxicity is a
major factor limiting plant growth in many soils (Sandalio et al. 2001, Artetxe et al.
2002, Fediuc and Erdei 2002). The most characteristic symptoms of Cd toxicity
are brown and short roots, chlorosis, fewer tillers, senescence and reduced plant
growth and biomass (Greger e/^/. 1991, Arduini et al. 1994, Boussama et al. 1999,
Wu and Zhang 2002b, Wu et al. 2003, Drazkiewicz and Baszynski 2005, Khan et
al. 2006, 2007a, Gianazza et al. 2007, Anjum et al. 2008a,c,d, Singh et al. 2008a) and
plant death in extreme cases (Sanita di Toppi and Gabbrielli 1999). It causes
inhibition of shoot and root growth (Schutzendubel et al. 2001), disorganization of the
grana structures and reduction in the biosynthesis of chlorophyll (Padmaja et al. 1990,
Somashekaraiah et al. 1992, Siedlecka and Krupa 1995^. Cadmiimi can also interfere
with photosynthesis (Baszynski 1986, Prasad 1995, Sandalio et al. 2001, Fargasova
2004, Ben Ammar et al. 2005, Khan et al. 2006, 2007a, 2008b, Samiullah et al. 2007,
Mobin and Khan 2007, Singh et al. 2008a), respiration, water relations and
reproduction (Barcelo and Poschenreider 1990, Prasad 1997). It causes changes in
organelles by disruption of membrane structure and fiinction (Sandalio et al. 2001,
Perfiis-Barbeoch et al. 2002, Gratao et al. 2006). Moreover, Cd can also inhibit the
activity of several groups of enzymes such as those of the Calvin cycle (Sandalio et
al. 2001), nitrogen metabolism (Boussama et al. 1999, Gouia et al. 2000, 2003,
Chaffei et al. 2003, Ghnaya et al. 2005, Anjana et al. 2006, Hasan et al. 2008),
2

carbohydrate metabolism (Sanita di Toppi and Gabbrielli 1999, Verma and Dubey
2001) and phosphorus metabolism (Shah and Dubey 1998, Sharma and Dubey 2006).
It also limits the uptake of both cations (K"", Ca "", Mg "", Mn " , Zn "", Fe "") and anions
like NO3". The toxicity symptoms observed in plants in the presence of excessive
amounts of Cd are due to a range of interactions at the cellular level (Hall 2002). It
enhances lipid peroxidation and alters functionality of membranes, and causes
changes in enzymatic and non-enzymatic antioxidant systems (De Knecht et al. 1995,
Piqueras et al. 1999, Chien and Kao 2000, Dixit et al. 2001, Olmos et al. 2003, Kuo
and Kao 2004, Ben Youssef er al. 2005, Cho and Seo 2005, Gratao et al. 2006, Mobin
and Khan 2007, Khan et al. 2007a, Anjum et al. 2008a,c,d, Singh et al. 2008a,b).
It has also been reported that excess of Cd stimulates the formation of free
radicals and reactive oxygen species (ROS) (Hendry et al. 1992, Groppa et al. 2001,
Sandalio et al. 2001, Fomazier et al. 2002a,b, lannelli et al. 2002, Milone et al. 2003,
Romero-Puertas et al. 2004, Ben Youssefet al. 2005, Gratao et al. 2006, Mobin and
Khan 2007, Singh et al. 2008a,b), that triggers activation of plant antioxidative
defense system which involves several enzymes and low molecular weight quenchers,
present in plant cells. Among antioxidant enzymes, superoxide dismutase (SOD; EC
1.15.1.1) constitutes the primary step of cellular defense and dismutates superoxide
radicals (O2") to H2O2 and O2. Accumulation of H2O2, a strong oxidant, is prevented
in the cell either by catalase (CAT; EC 1.11.1.6) or by the Ascorbate-Glutathione
cycle, where ascorbate peroxidase (APX; EC 1.1.11.1) converts it to H2O. Finally,
glutathione reductase (GR; EC 1.6.4.2) catalyzes the NADPH-dependent reduction of
oxidized glutathione (GSSG) to the reduced glutathione (GSH) (Noctor et al. 2002).
Ascorbate (AsA) and glutathione (GSH) are important components of Ascorbate-
Glutathione cycle responsible for the removal of H2O2 in different cellular
compartments (Foyer et al. 2002). Ascorbate is the major, probably the only,
antioxidant buffer in the apoplast and is an essential metabolite involved with vital
cell functions. It is a key primary antioxidant that reacts directly with OH- radicals,
O2", and O2 (Chen and Gallic 2004). Glutathione, present in plant cell is a major
non-enzymatic scavenger of ROS (Mishra et al. 2006). In particular, GSH play a very
important role in plants exposed to Cd and sequester it into the vacuoles by the
contribution of a protein with properties similar to those of glutathione-S-transferase
(GST; EC 2.5.1.18) (Marrs and Walbot 1997). Glutathione in the reduced form
protects the thiol (-SH) groups of many enzymes from oxidative conditions. For this
3

reason, its concentration is controlled by a complex homeostatic mechanism where
the availability of sulfiir (S) seems to be required (May et al. 1998a,b). Both AsA and
GSH can also react directly and scavenge certain ROS. However, a general
conclusion on the regulation of the antioxidant system by S assimilation has not been
drawn. Cadmium has a high affinity to metabolic processes of S assimilation by its
effect on the activity of ATP-sulfurylase (ATP-S; EC 2.7.7.4) (Sanita di Toppi and
Gabbrielli 1999, Khan et al. 2007a), and the enzymes involved in the activation of
sulfur (Dominguez-Solis et al. 2001, Harada et al. 2002, Hassan et al. 2006). The
entry of Cd into the cytosol promptly activates S metabolism for the accumulation of
reduced S-containing compound, GSH, which serves as substrates for the biosynthesis
of phytochelatins (PCs) (Rauser 2000, Cobbett 2000b, Leustek et al. 2000, Cobbett
and Goldsbrough 2002), and cysteine (Cys) is a precursor for the biosynthesis of GSH
and its derivatives (Rauser 1990, Inouhe 2005). The rate-limiting step in PC
biosynthesis is the provision of Cys. Thus, the availability of reduced S is considered
as of prime importance (Lee and Leustek 1999, Goldsbrough 2000). Both Cys and.
GSH hence are said to be involved in heavy metal detoxification (Inouhe 2005).
As GSH contains three moles of nitrogen (N) per mole of S, it may be
assumed that GSH biosynthesis is also dependent on the availability of N precursors
and thus N nutrition of plants. However, the sink strength of GSH biosynthesis for N
may be low compared with the other major N sinks such as the synthesis of proteins
or nucleotides (Kopriva and Rennenberg 2004). The GSH biosynthesis is, therefore,
regulated not only by the Cys availability (S, N and carbon metabolism) but also on
the availability of Glutamate (Glu) and Glycine (Gly). Glutamate the primary product
of the GS-GOGAT pathway of N assimilation is produced in photoautotrophic cells
(Gebler et al. 1998). Glycine the C-terminal amino acid of GSH, is an intermediate of
photorespiration in photoautotrophic cells. The availability of Gly hence is also
dependent on N assimilation and the Citric Acid cycle in mitochondria. Thus, GSH
biosynthesis has a tighter relationship with S and N, and also to carbon metabolism
(Kopriva and Rennenberg 2004).
Increasing evidence suggests that mineral-nutrient status of plants plays a
critical role in increasing plant resistance to environmental stress (Marschner 1995,
Vassilev et al. 2005, Hassan et al. 2006, 2008a,b, Anjum et al. 2008a,b, Khan et al.
2008a). The use of S and N fertilizers is a potential option which can be used to
alleviate Cd toxicity. The plant nutrients, S and N are of great importance. Sulfur is an
4

essential macronutrient of plants that plays a vital role in the regulation of plant
growth and development (Ernst et al. 2008). In addition, it is a structural constituent
of several coenzymes and prosthetic groups, such as ferredoxine, which is also
important for N assimilation. Nitrogen is an important component of several
important structural, genetic and metabolic compounds in plant cells. Plant N status is
highly dependent on N fertilization (Hernandez et al. 1997). N is also a major
component of chlorophyll and amino acids, the building blocks of proteins. Increase
in N supply can stimulate plant growth and productivity (Joel et al. 1997), as well as
photosynthetic activity (Makino et al. 1992) through increased amounts of stromal
and thylakoid proteins in leaves (Bungard et al. 1997).
Several studies have established regulatory interactions between assimilatory
sulfate and nitrate reduction in plants (Rennenberg et al. 1979, Reuveny et al. 1980,
Brunold and Suter 1984, Haller et al. 1986, Takahashi and Saito 1996, Abdin et al.
2003, Hawkesford et al. 2006). Most of the reduced S and N in a plant are utilized in
protein synthesis. As the nitrate reductase (NR; EC 1.6.6.1) (Abrol et al. 1976,
Ahmad et al. 1999, Abdin et al. 2003, Abrol 2007) and ATP-sulfiirylase (Brunold
1990, Kopriva et al. 2002, Saito 2004, Anjum et al. 2008b) enzymes catalyze the first
steps of the nitrate and sulfate assimilation pathways, respectively, they can be used
as indicators of the states of regulation of these pathways (Barney and Bush 1985).
Also, the assimilatory pathways of these elements have been considered convergent
and well coordinated (Brunold 1993, Abdin et al. 2003, Hawkesford et al. 2006). A
positive role of sulfate in regulating nitrate reductase was found by Pal et al. (1976)
and Friedrich and Schrader (1978), and role of N in the regulation of sulfate
assimilation at the ATP-sulfurylase step was observed by Smith (1975). Absence of
readily available external N source decreases ATP-sulfurylase activity (Brunold and
Suter 1984). The interrelationship of the regulation of nitrate and sulfate assimilation
has been found to be an effective mechanism to coordinate and meet the requirement
of net protein synthesis (Reuveny et al. 1980). It has also been established through
genetic studies that sulfate reduction is regulated by N nutrition at the transcriptional
level (Koprivova et al. 2000).
The oleiferous Brassica is the third most important source of vegetable oil in
the world after palm and soybean oil and grown as an edible or an industrial oil crop
which is used as a source of edible protein, in much the same way as soybean protein
(Zhang et al. 2003). Brassica is the Latin name of a genus that is taxonomically
5

placed within the Brassicaceae (Cruciferae), which is one of the tenth most
economically important plant families in the world. The major mustard oil-producing
countries include Canada, China, France, Germany, India and UK. According to a
report of United States Department of Agriculture (USDA), the world oilseed
production was 397metric tons in 2006-07. Indian agriculture contributes about 15%
and 8% to the world total acreage under oilseed cultivation and production,
respectively. However, the productivity in India is only 791kg ha"' as compared to the
world average of 1718kg ha'' (Damodaran and Hedge 2002). About 90% of the total
land under oilseed cultivation in India is occupied by Brassica juncea (Khan et al.
2007b). Despite a large area under cultivation of mustard, the productivity of the crop
has dropped because plant growth and development are affected by heavy metals
introduced into soil by anthropogenic activities.
Discussed the importance of S and N in plant metabolism and their role in
stress tolerance it was assumed that mustard {Brassica juncea L. Czem & Coss.) may
serve as a model crop in the study of the influence of S and N on plant metabolism
under Cd stress. Further, the fertilization of crop with balanced S and N may prove
fruitful in alleviating Cd stress and improving crop productivity. Studies on crops
with a view of alleviating Cd stress with balanced S and N application have not been
carried out. Therefore, the reported research was undertaken with the following
objectives:
• To screen and select mustard cuhivars as Cd tolerant and Cd non-tolerant
grown vmder Cd treatments.
• To study the influence of S application in the alleviation of Cd-induced effects
in Cd tolerant and Cd non-tolerant cultivars of mustard.
• To study the influence of combined application of S and N in the alleviation of
Cd-induced effects in Cd tolerant and Cd non-tolerant cultivars of mustard.

REVIEW OF LITERATURE

Contents
REVIEW OF LITERATURE
2.1 2.2 2.2.1 2.2.2 2.3 2.4 2.4.1 2.5 2.5.1 2.5.1.1 2.5.1.2 2.5.2 2.6 2.7 2.8 2.9 2.9.1 2.9.2 2.10 2.10.1 2.10.1.1 2.10.1.2 2.10.1.3 2.10.1.4 2.10.1.5 2.10.1.6 2.10.1.7 2.10.1.8 2.10.2 2.10.2.1 2.10.2.2 2.11
2.12 2.13 2.14
Physical and Chemical Characteristics of Cd Sources of Cd Contamination in Soil Natural sources Anthropogenic sources Uptake, Accumulation, Transport and Localization of Cd Morphological Alterations under Cd Stress Plant growth Physiochemical Alterations under Cd Stress Photosynthetic pigments Chlorophyll Carotenoids Photosynthesis Sulfur Metabolism imder Cd Stress Nitrogen Metabolism under Cd Stress Yield Characteristics under Cd Stress Cadmium-induced Oxidative Stress Lipid peroxidation Hydrogen peroxide content Cadmium-tolerance Mechanism in Plants Enzymatic antioxidant system Superoxide dismutases Catalase Ascorbate peroxidase Glutathione reductase Monodehydroascorbate reductase Glutathione S-transferase Glutathione peroxidase Guaiacol peroxidase Non-enzymatic antioxidant system Ascorbic acid Glutathione Coordination of Sulfur and Nitrogen in Plant Growth and Productivity Coordination of Sulfur and Nitrogen under Environmental Stress Research Work Reported from Our Laboratory Critical Appraisal of Review of Literature
7 8 8 9 9
12 12 14 14 14 15 16 18 20 21 23 25 26 27 28 28 30 31 32 33 34 34 34 35 35 36
38 39 42 46

REVIEW OF LITERATURE Chapter 2
Heavy metal contamination is a major environmental problem worldwide.
Environmental pollution by metals increased with the increase in mining and
industrial activities in the late 19* and early 20'*' century. The current worldwide mine
production of Cu, Cd, Pb and Hg is considerably large (Pinto et al. 2004). They are
released into the environment through various means including natural and
anthropogenic sources. The metal pollutants, derived from a growing number of
diverse anthropogenic sources, have had enormous impact on different ecosystems
(Macfarlane and Burchett 2001). In fact, heavy metals enlist a relatively large series
of elements with specific density over 5g cm" and relative atomic mass above 40.
Many of these metals manifest high affinity for sulfur-containing ligands and strongly
bind to the latter.
Fifty-three of the ninety naturally occurring elements are heavy metals (Weast
1984). Of these, Fe, Mo and Mn are important as micronutrients, while Zn, Ni, Cu,
Co, Cr and Va are toxic element, but have importance as trace elements. On the other
hand, Ag, As, Cd, Hg, Pb and Sb have no known function as nutrients, and seem to be
toxic to plants and microorganisms (Nies 1999). The presence of heavy metals in the
atmosphere, soil and water in excess amount, can cause serious problems to all
organisms. Among heavy metals, Cd is considered as an extremely potent pollutant
affecting all life forms due to its high toxicity, long half life and greater solubility in
water (Pinto et al. 2004). It is readily taken up by plants from soil via the root system
(Sharma and Dubey 2006). The level of Cd in soil is continuously increasing over
time (Jones et al. 1992). Soils contaminated with Cd show a sharp decline in crop
productivity and therefore, Cd serves as a serious problem for agriculture (Shah and
Dubey 1997).
2.1 Physical and Chemical Characteristics of Cd
Pure cadmium is a soft, silver-white, malleable, ductile and flexible metal. It is not
usually found in the environment as a metal, but occurs normally as a mineral
combined with other elements such as oxygen (cadmium oxide), chlorine (cadmium
chloride) or sulfur (cadmium sulfate, cadmium sulfide).

Atomic number
Relative atomic mass ('^C=12.000)
Melting point/°C
Boiling point/°C
Density/g cm'
Electron affinity (M-M-)/kJ mol"'
48
112.41
321
765
8.64
+26
Source: Chatterjee and Dube (2006)
Cadmium is the second member of Class II Group b of the Periodic Table. The
stable state of Cd in the natural environment is Cd" . It has a medium Class b
character compared to Zn and Hg. This imparts moderate covalency in bonds and high
affinity for sulfhydryl groups, leading to increased lipid solubility, bioaccumulation
and toxicity.
2.2 Sources of Cd Contamination in Soil
Some of the heavy metals are abundant in serpentine soils (Ni, Cr, Co etc.), some
others in calamine soils (Zn, Pb, Cd etc.). As they cannot be degraded or destroyed,
they are persistent in all parts of the environment. Humans also affect the natural,
geological and biological redistribution of heavy metals by altering the chemical form
of heavy metals released into the environment. Among heavy metals, Cd inputs have
also natural as well as anthropogenic sources. Cadmium is naturally occurring
metallic element, one of the components of earth's crust and present everywhere in
the environment (Chatterjee and Dube 2006). Different types of sources are given
below.
2.2.1 Natural sources
The natural occurrence of Cd in the environment results mainly from gradual
phenomena, such as rock erosion/abrasion, that estimates for 15,000mt Cd per annum
(OECD 1994, WHO 1992). Natural calamities such as volcanic eruptions, add 820mt
Cd per annum to the environment (Nriagu 1980, 1989, OECD 1994, WHO 1992). The
naturally existing concentration of Cd in the atmosphere is 0.1-0.5ng m"''. In the
earth's crust, its concentration varies from 0.1 to 0.5|ag g"', but there are reports of a
much higher accumulation in the sedimentary rocks and marine phosphates.
Phosphorites have been reported to contain Cd levels as high as a SOOppm (Cook and
Morrow 1995). Forest fires are also regarded as a natural source of Cd emissions in
the air that range from l-70mt per annum (Nriagu 1990).
8

Atmosphere Earth's crust Marine sediment Sea-water
0.1 toSngm"^* O.ltoO.S^gg"'
- l^gg' ' ~o.l^g^'
* no — ng - nanograms = 10" g
Source: McLaughlin et al. (2000), Chaney and Homick (1978)
2.2.2 Anthropogenic sources
Cadmium contamination of soils may be categorized as i) inputs to agricultural soils,
ii) inputs to non-agricultural soils, and iii) depositions in controlled landfills. In the
first case, the main chunk of Cd comes from atmospheric deposition, sewage sludge
application, and phosphate fertilizers application (Jensen and Bro-Rasmussen 1992,
Van Assche and Ciarletta 1993, Chen et al. 2007). In the second case, inputs arise
mainly from the iron and steel industry, non-ferrous metals production, fossil fuel
combustion, and cement manufacture (ERL 1990, Jackson and MacGillivray 1993,
OECD 1994). In fact, Cd is discharged into the soil to the tune of 22,000ton per year
globally (Nriagu and Pacyna 1988). In the case of controlled landfills, presence of Cd
is mainly due to the disposal of products, which may contain Cd impurities, and
naturally-occurring wastes such as grass, food and soil which inherently contain trace
levels of Cd (Chandler 1996).
2.3 Uptake, Accumulation, Transport and Localization of Cd
Cadmium is taken up by plants from soil \ia the root system and to a lesser extent
through leaves. As soon as Cd enters the roots, it can reach the xylem through an
apoplastic and/or symplastic pathway. Plants show a differing metal distribution and
accumulation pattern among different parts. Most of Cd that enters the plant system
accumulates in the roots and only a small portion is translocated to the above ground
parts (Schutzendubel et al. 2001, Vitoria et al. 2001, Pereira et al. 2002, Ramos et al.
2002, Patel et al. 2005, Kovacik et al. 2006, Sharma and Agrawal 2006, Mobin and
Khan 2007, Ekmekci et al. 2008, Liu et al. 2007, Singh et al. 2008a,b). The actual
accumulation of Cd in plant parts depends on the plant species and soil properties
(Fediuc and Erdei 2002, Arao et al. 2003). Substantial variability among 99 Pisum
sativum genotypes in tolerance to Cd and uptake of different heavy metals was
reported by Belimov et al. (2003). Zhang et al. (2002) reported significant differences
among Triticum aestivum genotypes in shoot Cd concentration. Significant
differences in Cd accumulation and tolerance were found in Sedum alfredii

populations (Deng et al. 2007). Ten times higtier Cd accumulation in the roots than
above ground parts was reported in Hordeum vulgare plants by Vassilev et al. (1998).
Cadmium was accumulated 1826 times more in the roots of Allium sativum than the
control with the application of lO' M Cd and very low amount was transported to the
bulbs and shoots (Jiang et al. 2001). Kovacik et al. (2006) reported that Cd
accumulation was seven- (60^M Cd) to eleven- (120^M Cd) fold higher in the roots
than in the leaves of Matricaria chamomilla, whereas, only 6% of Cd was
accumulated in the leaves of Crotalaria juncea compared to roots with 2mM CdCb
(Pereira et al. 2002). The Milyang 23 rice accumulated 10-15% of the total soil Cd in
its shoot (Murakami et al. 2007). Djebali et al. (2008) reported that the roots of Cd
treated Solarium lycopersicum plants accumulated four to five fold Cd as much as
mature leaves. Liu et al. (2007) suggested that the root tissue was a barrier to Cd
uptake and translocation within rice plants. Retention of Cd in roots might be due to
its cross linking with carboxyl groups of cell wall proteins (Barcelo and Poschenreider
1990) and/or an interaction with the thiol groups of soluble proteins and non-protein
thiols operating as a tolerance mechanism in root cells (Chaoui et al. 1997a). Low
temperature inhibits the uptake of Cd as well as its transport across plasma membrane
(Hart era/. 1998).
The uptake of Cd by plants varies not only among plant species but also
among cultivars (Salt et al. 1995, Arao et al. 2003, Metwally et al. 2005, Grant et al.
2008). Li et al. (1997) reported significant variation in the grain Cd level of
Helianthus annuus, Triticum aestivum and Linum usitatissimum. Arao et al. (2003)
found that lower level of Cd in the seeds of certain soybean varieties was due to the
lower initial uptake. Studies of Cd uptake by plants have indicated that at lower Cd
concentrations (2.5-90nM), its transport across membrane is an active, energy
requiring H" -ATPase mediated process, whereas at high concentrations of Cd the
uptake is a non metabolic (passive) process, involving diffusion coupled with
sequestration (Grant et al. 1998). Although the mechanism of transport of Cd across
the plasmalemma of root cells is not well understood, the electrochemical potential
across the membrane appears to be an important factor (Grant et al. 1998). It is
suggested that the uptake of cationic solutes is likely to be driven largely by the
negative membrane potential across the plasma membrane, which is generated in part
by metabolically dependent processes such as proton extrusion via the plasma
membrane H''-ATPase (Kochian 1991). Following uptake by plant roots, Cd gets
10

accumulated in cytosol, cytosolic organelles and vacuoles. At low level of exposure,
Cd forms complexes in the cytosol with glutathione whereas, at higher Cd exposure
levels, it is transported into the vacuoles, where it forms complexes with organic acids
and phytochelatins (Grant et al. 1998). hi most of the plant species much of the Cd
taken up by plants is retained in root and its translocation to aerial portion is low. In
Glycine max plants, about 98% of the accumulated Cd is strongly retained by roots
and only 2% is transported to the shoot system (Cataldo et al. 1983). Rice plants
absorb Cd from the rooting medium against the concentration gradient and the
localization of absorbed Cd in rice is greater in roots than in shoots. When rice
seedlings were raised in sand cultures for 20d in nutrient medium containing 500|iM
Cd, localization of absorbed Cd was about 3 times in roots compared to its level in
shoots (Shah and Dubey 1998). Movement of Cd from root to shoot is also likely to
occur via the xylem and is driven by the transpiration stream from the leaves that
include metal uploading into root xylem cells, long distance transport from root to
shoot within xylem and reabsorption of metal from the xylem stream by leaf
mesophyll cells (Raskin and Ensley 2000). Evidence for this was provided by Salt et
al. (1995) who showed that ABA-induced stomatal closure reduced Cd accumulation
in shoots of Brassicajuncea. Reduced movement of Cd from roots to shoots in plants
is believed to result from barrier fimction of root endodermis and mechanism
involving sequestration and decreased xylem loading of Cd (Hart et al. 1998). The
casparian strips of root endodermis retard the entrance of Cd into the central cylinder
(Seregin et al. 2004), vascular compartmentation of Cd tends to limit symplastic
movement of Cd. Movement of Cd across the tonoplast occurs via Cd^VH' -antiport
system (Salt and Wagner 1993) as well as by phytochelatin-Cd transporter (Vogeli-
Lange and Wagner 1990) that appears to be Mg-ATP dependent (Salt and Rauser
1995). The absorption of Cd by green microalgae, Chlorella vulgaris, Ankistrodesmus
braunii and Eremosphera viridis revealed that Cd is mainly absorbed in the cell wall.
The binding sites seem to be an acidic group in the cell wall structure (Geisweid and
Urbach 1983). In Eichhornia crassipes, Cd was found to accumulate throughout the
roots (Hosayama et al. 1994). Ammar et al. (2007) reported that Cd was found to be
mainly accumulated in roots, but a severe inhibition of biomass production occurred
in Lycopersicon esculentum leaves, even at its low concentration (l|uM). In Solidago
altissima, Cd was found in most parts of the plant, but located mainly in the cambium,
cortex and phloem tissues (Hosayama et al. 1994).
11

Most of the Cd accumulates in the vacuole within a cell. The fact that Cd is
found in golgi apparatus and endoplasmic reticulum is apparently related to metal
secretion through the cell surface and into the vacuole. A small quantity of Cd reaches
nuclei, chloroplast, and mitochondria and exerts toxic effects on these organelles
(Miller et al. 1973, Malik et al. 1992a). Localization of Cd in different parts of a plant
appears to be maximum in roots (Hart et al. 1998).
2.4 Morphological Alterations under Cd Stress
2.4.1 Plant growth
The growth of whole plant or of plant parts is frequently used as an easily
measurable parameter to monitor the effects of various stressors. The root system
of plants acts usually as the first barrier or acceptor to heavy metals in soil.
Reduction in growth and biomass yield with increased levels of Cd in growth
media arises because of altered physiological phenomena (Demirevska-Kepova
et al. 2006). The most characteristic symptoms of Cd stress are brown and short
roots, chlorosis, fewer tillers, senescence and reduced plant growth and biomass
(Arduini et al. 1994, Wu and Zhang 2002b, Wu et al. 2003, Cosio et al. 2006). In
Elodea canadensis, a thiimer stem, less expanded leaves with partial bleaching of
green tissues and 40% internode shortening were observed in response Cd
treatments when compared with control plants (Vecchia et al. 2005). Bachir et
al. (2004) found decrease in root length, plant height and fruiting branch number
in cotton with increasing Cd concentration in a pot experiment. Cadmium
treatments inhibited the shoot and root growth and dry weight of two wheat
cultivars, C-1252 and Balcali-85. The decrease was more distinct in C-1252
(Ozturk et al. 2003). Sandalio et al. (2001) reported significant reduction in root
grov^h in Pisum sativum plants. Roots of Pisum sativum plants were more
sensitive to Cd toxicity than shoot (Metwally et al. 2005). Increasing
concentration of Cd in liquid culture and pot experiment decreased the
germination and root growth of carrot and radish plants (Chen et al. 2003). Rai et
al. (2005) observed significant decrease in root and shoot length and fresh and
dry weight of Phyllanthus amarus under Cd stress. Cadmium inhibited root dry
mass and induced changes in biomass allocation pattern without any effect on
biomass accumulation at the whole plant level in Hordeum vulgare (Vassilev et
al. 2004). Leaf expansion and root growth were inhibited significantly at high Cd
concentrations in Sedum alfredii, and Cd was suggested to suppress cell
12

expansion and induced senescence (Zhou and Qiu 2005). Exposure to increasing
Cd concentrations reduced the fresh weight of the upper part
(hypocotyis+cotyledons) of the seedlings of Brassica napus more strongly than
that of the root system, which was accompanied by higher Cd accumulation in
these tissues (Filek et al. 2007). Ekmekci et al. (2008) reported that increasing Cd
concentration significantly reduced the leaf and root dry weight of Zea mays cultivars.
Prince et al. (2002) reported significant decrease in plant height, number of
leaves and leaf yield of Cd exposed Morus alba plants. Root and shoot growth of
Crotalaria juncea seedlings was strongly inhibited by Cd concentrations above
0.2mM CdCb (Pereira et al. 2002). Di Cagno et al. (1999) observed Cd-induced
inhibition of growth parameters in Halianthus annuus seedlings in both mature
and young leaves after 7d of exposure. The presence of Cd decreased the
seedling growth of Zea mays, Triticum aestivum, Cucumis sativus and Sorghum
bicolor in terms of root and shoot growth (An 2004). Ghnaya et al. (2005)
reported that Cd severely inhibited Mesembryanthemum crystallinum growth
even at low concentration. Cadmium caused gradual decrease in plant growth of
Lemna minor (Razinger et al. 2007). Patel et al. (2005) observed Cd-induced
decrease in dry matter production of Colocassia esculentum plants. Increasing Cd
concentration in the solution significantly decreased the shoot and root dry
weight of two selected Brassica napus and Brassica juncea plants (Su and Wong
2004). Wahid et al. (2007) observed increased shoot Cd accumulation and leaf
chlorosis with a concomitant reduction in shoot dry weight, leaf area, relative
growth rate, net assimilation rate and relative leaf expansion rate in Vigna
radiata seedlings under Cd stress. Krantev et al. (2007) reported that exposure of
Zea mays plants to Cd caused a gradual decrease in the shoot and root dry weight
accumulation. Ahsan et al. (2007) reported that increasing Cd concentration in
the medium significantly decreased the germination rate, shoot elongation and
biomass of Oryza sativa. Kuriakose and Prasad (2008) reported that at
concentrations above 3mM Cd, seed germination of Sorghum bicolor was
adversely affected with a complete cessation of seedling growth.
The over all reduction in growth has been attributed to repression of the
elongation and the growth rates of cells because of an irreversible inhibition
exerted by Cd on proton pump (Aidid and Okamoto 1993). However, slight
increase in the growth was noticed when the plants were exposed to very low Cd
13

concentrations. Stimulatory effect of lower doses of Cd on growth has also been
observed in Lycopersicon esculentum and Solanum melongena (Khan and Khan
1983), Beta vulgaris (Greger and Lindberg 1986), Medicago saliva seedlings
(Peralta et al. 2000), Hordeum vulgare (Wu et al. 2003), Miscanthus sinensis
(Arduini et al. 2004) and Lycopersicon esculentum seedlings (Dong et al. 2005).
2.5 Physiochemical Alterations under Cd Stress
2.5.1 Photosynthetic pigments
Chlorophyll (Chi) and carotenoids (Car) are the most abundant biological pigments in
plants (Hall and Rao 1999). The presence of these pigments in plants reflects the
nutritional status, growth as well as crop productivity (Seyyedi et al. 1999). Chlorosis
and retardation of plant growth that is frequently observed in metal polluted
enviroimient indicate that an impairment of photosynthetic pigment biosynthetic
pathway is among the earlier targets of heavy metal influence on plant metabolism.
Lower content of photosynthetic pigments induces changes in plastid development,
photosynthetic efficiency as well as in general metabolism. Better is the status of
these pigments, better will be the light harvesting capability and the capability of
plants to fix CO2.
2.5.1.1 Chlorophyll
Chlorophylls play a fundamental role in the process of photosynthesis because of its
ability to absorb light. Baszynski et al. (1980) reported about the negative effect of Cd
on Lycopersicon esculentum plants. Lately, Cd-induced decrease in Chi content was
observed in Triticum aestivum and Cucumis sativus (Buszek 1984, Malik et al.
1992a,b), Zea mays (El-Enany 1995, Stiborova et al. 1986, Krantev et al. 2007,
Ekmekci et al. 2008), Phaeolus vulgaris (Barcelo et al. 1988a,b, Siedlecka and Krupa
1996), Hordeum vulgare genotypes (Wu et al. 2003), Oryza sativa (Kuo and Kao
2004), Glycine max seedlings (Drazic et al. 2004), Matricaria chamomilla (Kovacik
et al. 2006), Brassica juncea (Mobin and Khan 2007), Bechmeria nivea (Liu et al.
IQQl), Brassica campestris (Anjum et al. 2008a) and Vigna mungo (Singh et al.
2008a). Following Cd treatment concentration of chlorophyll a (Chi a) is reduced
more than chlorophyll b (Chi b) in Phyllanthus amarus (Rai et al. 2005), Brassica
juncea plants (Mobin and Khan 2007) and Vigna mungo (Singh et al. 2008a). The
increased Chi a:b ratio was linked with the change in pigment composition of
photosynthetic apparatus which posses lower level of light harvesting Chi proteins
(Loggini et al. 1999). Skorzynska-Polit and Baszynski (1997) reported that Cd-
14

induced necrosis of leaf tissues might be the reason of Cd mobilization and its
transport to above ground plant parts.
Most researchers connect the reduction of Chi in Cd-treated plants with
inhibition of its biosynthesis. Cadmium-induced inhibition of Chi biosynthesis was
even suggested to be a primary event as compared to the inhibition of photosynthesis
(Baszynski et al. 1980). Stobart et al. (1985) established that Cd inhibited Chi
biosynthesis at two levels: in the synthesis of 5-aminolaevulinic acid (ALA) and in
the formation of photoactive protochlorophyllide reductase complex. Horvath et al.
(1996) reported that the photoconversion of protochlorophyllide was not inhibited, but
Cd disturbed Chi molecules integration in stable complexes. On the other hand,
Greger and Lindberg (1986) suggested that the lower Chi concentrations in plants
were a result of the deficiency of Mg and Fe in the leaves of Cd-treated Beta vulgaris
plants. The decrease in Mg and Fe content in leaves as a response to Cd treatment has
been established in other plant species also (Breckle and Kahle 1992, Rubio et al.
1994). More convincing data about the interactions between Fe deficiency (this
element is a co-factor of an enzyme taking a part in Chi biosynthesis) and Chi
concentrations were reported by Krupa and Baszynski (1995). They showed that
50^M Cd induced Fe deficiency and decreased Chi by 55% in bean plants. Lang et al.
(1995) confirmed that translocation of the labeled Fe to the over-ground organs of Cd-
treated Cucumis sativus plants was inhibited and decreased Chi. Somashekaraiah et
al. (1992) showed that treatment of 100|aM Cd for 6d increased the lipoxygenase
activity and decreased Chi concentration in Phaseolus vulgaris plants.
2.5.1.2 Carotenoids
Carotenoids carry out three major functions in plants. First, they absorb light at
wavelength between 400 and 550nm and transfer it to the Chi (an accessory light-
harvesting role) (Siefermann-Harms 1987). Second, they protect the photosynthetic
apparatus by quenching a triplet sensitizer (ChP), singlet oxygen and other harmful
free radicals which are naturally formed during photosynthesis (an antioxidant
ftmction) (Oelmuller 1989, Thiele et al. 1996, Havaux et al. 2000, Collins 2001).
Third, they are important for the photosystem (PS) 1 assembly and the stability of
light harvesting complex proteins as well as thylakoid membrane stabilization (a
structural role) (Mayfield and Taylor 1984, Siefermann-Harms 1987, Niyogi et al.
2001).
15

The influence of Cd on Car has been investigated in different plants.
Cadmium-induced decrease in Car content in the seedlings of Raphanus sativus, Ulva
lactuca, Triticum aestivum and Oryza sativa was reported by Naguib et al. (1982).
Baszynski et al. (1980) observed Cd induced decrease in Car content in Lycopersicon
esculentum. The toxic effect of Cd was partially reversed by the addition of Mn;
however, the Car content never reached the value of control plants. Chi and Car
contents were increased initially and decreased thereafter in Zea mays leaves under
Cd stress (Prochazkova et al. 2001). Baryla et al. (2001) reported reduced Car content
in Brassica napus plants imder Cd stress. Rai et al. (2005) and Singh et al. (2008a)
reported decreased Car and Chi contents in Phyllanthus amarus and Vigna mungo
plants with increasing Cd concentration, respectively. In Brassica napus, Cd reduced
the total Chi and Car contents and increased the non-photochemical quenching
(Larsson et al. 1998). Ekmekci et al. (2008) reported that the increase in Cd
concentration caused loss of Car in Zea mays cultivars. Collin et al. (2008) also
reported decreased concentration of Car in Arabidopsis plants. Car content of
Hordeum vulgare seedlings decreased under Cd-stress (Demirevska-Kepova et al.
2006) or remained less affected (Clijsters and Van Assche 1985). An increase in Car
content was also reported following Cd stress (Foyer and Harbinson 1994).
2.5.2 Photosynthesis
Cadmium has been shown to affect the photosynthetic functions through interacting
with photosynthetic apparatus at various levels of organization and architecture viz.,
accumulation of metal in leaf (main photosynthetic organ), partitioning in leaf tissues
like stomata, mesophyll and bundle sheath cells, interaction with cytosolic enzymes
and alteration of the functions of chloroplast membranes. Cadmium has been shown
to be the most effective inhibitor of photosynthetic activity (Bazzaz et al. 1974,
Huang et al. 1974, Hampp et al. 1976), particularly the oxygen evolving reactions of
photosystem II (Bazzaz and Govindjee 1974, Baszynski et al. 1980, Atal et al. 1991).
With only small amount of Cd in chloroplasts, many direct and indirect effects are
observed, resulting in strong inhibition of photosynthesis. Earlier investigations have
demonstrated a marked reduction in the rate of photosynthesis by Cd in different plant
species (Baszynski et al. 1980, Sawhney et al. 1990, Sheoran et al. 1990a,b, Chugh
and Sawhney 1999, Arduini et al. 2004, Wojcik and Tukiendorf 2005, Jing et al.
2005, Mobin and Khan 2007, Singh et al. 2008a, Khan et al. 2006, 2007a, Anjum et
al. 2008a). Deleterious effects of Cd on various facets of photosynthesis, such as Chi
16

metabolism (Padmaja et al. 1990, Stobart et al. 1985, Sandalio et al. 2001, Chaffei et
al. 2004, Hsu and Kao 2004, Vassilev et al. 2005), functioning of photochemical
reactions (Li and Miles 1975, Skorzynska-Polit and Baszynski 1995) and the activities
of the Calvin cycle enzymes (Ascencio and Cedeno-Maldonado 1979, Weigel 1985,
Krupa 1999) have also been reported. Baszynski et al. (1980) observed that the
decline in chlorophyll content preceded that of CO2 fixation and proposed this to be
the primary cause of diminished photosynthetic activity. Cadmium-induced reduction
in the activity of ribulose 1,5 bisphosphate carboxylase (Rubisco) has been reported in
Hordeum vulgare (Stiborova et al. 1986, Vassilev et al. 2004), Cajanus cajan
(Sheoran et al. 1990b), Triticum aestivum (Malik et al. 1992b), Pisum sativum
seedlings (Chugh and Sawhney 1999), Zea mays (Krantev et al. 2007) and Brassica
juncea (Mobin and Khan 2007). Wahid et al. (2007) reported Cd-induced reduction in
transpiration rate, stomatal conductance and net photosynthesis due to reduced CO2
fixation by Rubisco in Vigna radiata plants. Di Cagno et al. (2001) also observed
significant reduction in CO2 fixation rate and Rubisco activity while stomatal
conductance and Fv:Fm ratio remain unchanged in Cd treated sunflower plants.
Carbonic anhydrase (CA), a zinc metalloenzyme related to photosynthesis in higher
plants (Khan 1994), was significantly reduced by Cd in Brassica juncea (Mobin and
Khan 2007) and Vigna mungo (Singh et al. 2008a). The loss of water from crop plants
is controlled mainly by the stomatal movement on leaves that has been shown to be
sensitive to various environmental factors including Cd stress (Barcelo et al. 1986a,b).
Stomatal closure to minimize water loss has been identified as an early event in plant
response to Cd-induced water deficiency leading to limitations in carbon uptake by
leaves (Barcelo et al. 1986a,b, Chaves 1991, Poschenreider et al. 1989). In addition,
conductance and index of stomata, transpiration and net CO2 uptake are greatly
reduced with elevated Cd levels in the growth media (Bindhu and Bera 2001,
Balakhnina et al. 2005). Siedlecka and Krupa (1996) found that leaf chlorophyll
content and Hill activity decreased with increased chlorophyllase activity under Cd
stress. Cadmium interacts with the water balance (Costa and Morel 1994) and
damages the photosystem apparatus, in particularly the PSI and PSII (Siedlecka and
Krupa 1996). In Medicago sativa, Becerril et al. (1989) found that Cd inhibited
transpiration and CO2 assimilation in a drastic manner, whereas, in Picea abies
seedlings decreased CO2 assimilation was mainly due to stomatal closure (Schlegel et
al. 1987). Cd-induced reduction in photosynthesis and transpiration was attributed to
17

stomatal closure in Beta vulgaris (Greger and Johansson 1992). Mobin and Khan
(2007) found that the Cd-induced decrease in net photosynthetic rate in non-tolerant
cultivar (RH30) of Brassica juncea was accompanied by an increased transpiration
rate and stomatal conductance but in tolerant cultivar (Varuna), it remained unaltered.
2.6 Sulfur Metabolism under Cd Stress
Plants can manage in a wide range of environmental stress conditions using
appropriate physiological responses. In the case of heavy metals stress, the processes
consist of modulation of the activity of plasma membrane and/or vacuolar transporters
and biosynthesis of intracellular metal-chelators (Clemens 2006). In this context,
sulfur plays a pivotal role because sulfate transporters can mediate the entry of
sulfate-analogues into the cells (Maruyama-Nakashita et al. 2007), and S-containing
compounds like GSH, PCs and metallothioneifis (MTs) often improve the tolerance of
plants to several metals and metalloids through complexation and/or further
sequestration of toxic forms inside cellular vacuoles (Xiang and Oliver 1998, Cobbett
and Goldsbrough 2002, Hall 2002).
Sulfur is an essential macronutrient that plays a vital role in the regulation of
plant growth and development (Ernst 1998, Singh 2004, Anjum et al. 2008b,
Bimbraw 2008, Ernst et al. 2008, Kumaran et al. 2008, Ratti and Giordano 2008). It is
constituent of amino acids, Cys and Met, GSH and of varieties of other plant
metabolites involved in stress tolerance (Saito 2000, Rausch and Wachter 2005,
Bouranis et al. 2008, Burritt 2008). A significant induction in S assimilation has been
reported in heavy metal-exposed higher plants (Tukiendorf and Rauser 1990).
Exposure of plants to Cd induces enzymes involved in the sulfate assimilation
pathway (Herbette et al. 2006, Khan et al. 2007a). It has been noted that genes
involved in S assimilation pathway are rapidly up-regulated such as Sultrl;l and
Sultr2;l encoding two sulfate transporters which are up-regulated after 2 or 6h of Cd-
treatment and 12-24h after sulfate-depletion (Takahashi et al. 2000, Herbette et al.
2006, Sarry et al. 2006). Nussbaum et al. (1988) reported that Cd accumulation
increased ATP-sulftorylase (ATP-S) and adenosine 5-phosphosulfate reductase (AR)
activities in Zea mays seedlings. Ruegsegger et al. (1990) showed that AR activity is
induced coordinately with glutathione synthetase in Cd-treated Pisum sativum plants.
In fact the capacity of plants to survive in a polluted environment is partially linked to
the efficiency of their reductive sulfate assimilation pathway. Upon heavy metal
stress, some genes involved in the S assimilation pathway are known to be
18

transcriptionally activated, resulting in an elevation of enzymatic activity (Xiang and
Oliver 1988, Schafer et al. 1998, Heiss et al. 1999, Lee and Leustek 1999, Barroso et
al. 2006). The involvement of S in Cd-tolerance mechanism was reported in
Arahidopsis thaliana (Dominguez-Solis et al. 2001, Harada et al. 2002), Brassica
juncea (Zhu et al. 1999a,b), Nicotiana tabacum (Harada et al. 2001), Triticum
aestivum (Khan et al. 2007a) and Brassica campestris (Anjum et al. 2008a).
A good part of S incorporated into organic molecules in plants is located in
thiol (-SH) groups in proteins (cys-residues) or non-protein thiols, GSH (Noji and
Saito 2003, Tausz et al. 2003, De Kok et al. 2005, Anjum et al. 2008b). Sulfur being a
component of PC may play an important role in their synthesis and ultimately in
detoxification of Cd through the formation of Cd-binding peptides (CdBP) (Cobbett
2000a,b, Harada et al. 2002, Cobbett and Goldsbrough 2002). In many plants, PC
synthesis is reported to be activated by heavy metal treatment and a supply of
precursors, including Cys, y-glutamylcysteine and GSH (Zenk 1996, Chen et al. 1997,
Goldsbrough 2000, Xiang et al. 2001, Nocito et al. 2002, 2006). Studies have
demonstrated the critical role of PCs in Cd detoxification and tolerance of plants to
Cd (Kuboi et al. 1987, Howden et al. 1995a,b, Inouhe et al. 2000, Harada et al. 2001,
2002, Ebbs et al. 2002, Srivastava et al. 2004, Wojcik et al. 2005, Zhang et al. 2008).
It has been showed that PC formation was induced in the leaf, stem and root tissues of
Sedum alfredii upon exposure to 400|aM Cd (Zhang et al. 2008). PCs rapidly form a
'low molecular weight' (LMW) complex with Cd (Sanita di Toppi and Gabbrielli
1999). These complexes acquire acid labile S at the tonoplast and form a 'high
molecular weight' (HMW) complex with a higher affinity for Cd ions (Hu et al.
2001). Gupta et al. (2002) reported an increase in Cys and GSH in the shoots of
Arabidopsis and suggested that these changes can be expected to have a positive
effect via transport on the PC/hPC synthesis and/or the other stress responses in roots
in a Cd-contaminated environment.
Moreover, evidence suggests that CdS solids may be formed and coated by
CdBP-Cd complexes after exposure of cultured suspension cells to high levels of Cd
(Rauser 1990) and the co-occurrence of CdS and CdBP-Cd has been thought to play a
role in stabilizing CdBP-Cd complexes (Rauser 1990, Wang and Evangelou 1995). As
CdBP synthesis and CdS solids formation are closely dependent on S metabolism and
increase the need for thiol compounds by cells, there is a need for maintaining high S
19

ions in the roots to sustain a high S-assimilation rate during phytochelatin
biosynthesis to detoxify Cd.
2.7 Nitrogen Metabolism under Cd Stress
The presence of Cd in plants results in many physiological alterations affecting both
nitrogen and carbohydrate metabolism (Greger and Bertell 1992, Hernandez et al.
1997, Chaffei et al. 2003). The nitrate assimilation process consumes about 25% of
energy produced by photosynthesis (Solomonson and Barber 1990). In most of the
plants nitrate reduction takes place in leaves where major part of the reducing power
arises directly from light via ferredoxin. A high rate of CO2 assimilation favours an
efficient N assimilation and vice versa (Ferrario et al. 1998). Cadmium as a potential
inhibitor of photosynthetic process considerably inhibits nitrate assimilation. It has
been shown that enzymes of N metabolism are differentially affected by Cd stress
(Petrovic et al. 1990, Chugh et al. 1992, Singh et al. 1994, Syntichaki et al. 1996,
Boussama et al. 1999). Among various enzymes nitrate reductase (NR) is a key
enzyme in the conversion of nitrate to nitrite, and its sustained activity is crucial to N
assimilation (Srivastava 1992, Gouia et al. 2000, Ghnaya et al. 2005, 2007). Nitrate
reductase and nitrite reductase (NiR) activities are significantly decreased by Cd,
leading to reduced nitrate assimilation by plants (Ferretti et al. 1993, Chaffei et al.
2004, Rai et al. 1998, Khudsar et al. 2001, Balestrasse et al. 2004, Anjana et al. 2006,
Wahid et al. 2007, Wang et al. 2008). Cadmium reduces absorption of nitrate and its
transport from root to shoot by inhibiting the NR activity in shoots (Hernandez et al.
1996). Cadmium treatment can also result in endogenous ammonium increase through
deamination of some free amino acids and other N forms. Chaffei et al. (2003)
demonstrated that Cd treatment produced ammonium accumulation through an
increase in protease activity. Glutamine synthetase (GS) is one of the key enzymes in
the main pathway of ammonium assimilation in higher plants (Lea and Miflin 2004),
and deleterious effects of Cd on its activity have been observed in several species
(Astolfi et al. 2004, Balestrasse et al. 2006a, Chaffei et al. 2004). In contrast,
glutamate dehydrogenase (GDH), an important enzyme in the "shunt" of N
metabolism, was induced under Cd stress, but the physiological role of GDH and the
increase in its activity are still controversial (Hernandez et al. 1997, Astolfi et al.
2004, Chaffei et al. 2004, Lea and Miflin 2004, Balestrasse et al. 2006a, Hsu et al.
2006). Nitrate assimilation also requires carbon skeletons, especially in the form of
2-oxoglutarate, which is produced via the anaplerotic pathway in the cytosol. 2-
20

Table 1. Effect of Cd on activities of enzymes of different metabolic processes. '+' and '- ' signs denote stimulatory and inhibitory effects of Cd on enzymes, respectively.
Chlorophyll Synthesis
Photosynthesis
ALA synthase
ALA dehydratase
NADPH:protochlorophyll -ide oxidoreductase
RUB? Carboxylase
PEP Carboxylase
Carbonic anhydrase
Phaseolus vulgaris Hordeum vulgare Phaseolus vulgaris Hordeum vulgare Triticum aestivum Phaseolus vulgaris Brassica juncea Triticum aestivum Cajanus cajan Zea mays
Vigna radiata Phaseolus vulgaris Cajanus cajan Zea mays
Phaseolus vulgaris Ceratophyllu m demersum Brassica juncea Vigna mungo Triticum aestivum Brassica juncea Cicer arietinum Cicer arietinum
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
Padmaja et al. 1990 Stobart et al. 1985 Padmaja et al. 1990 Stobart et al. 1985 Boddi et al. 1995 Siedlecka et al. 1997 Mobin and Khan 2007 Malik et al. 1992b Sheoran et al. 1990b Krantev et al. 2007 Anjum et al. 2008c Siedlecka et al. 1997 Sheoran et al. 1990b Krantev et al. 2007 Gouia et al. 2003 Aravind and Prasad 2004 Mobin and Khan 2007 Singh et al. 2008a Khan et al. 2007a Hayat et al. 2007 Hasan et al. 2007 Hasan et al. 2008

N metabolism
S metabolism
Nitrate reductase
Glutamine synthatase
Glutamate dehydrogenase ATP-sulfurylase
ATP-sulfurylase, APS reductase, Sulfite reductase ATP-sulfurylase, APS reductase OASTL
Glutamate dehydrogenase, Serine acetyltransferase and 0-acetyl-L-serine (thiol)lyase iS-nitrosoglutathione reductase
Sesamum indicum Vigna radiata Brassica juncea Cicer arietinum Phaseolus vulgaris Pisum sativum Phaseolus vulgaris Triticum aestivum Arabidopsis thaliana
Brassica juncea
Chlamydom o-nas reinhardtii
Pisum sativum
—
—
—
—
—
—
+ +
+ +
+ +
+ +
+ +
+ +
—
Singh et a/. 1994 Wahid et al. 2007 Hayat et al. 2007 Hasan et al. 2008 Gouia et al. 2000 Chugh et al. 1992 Gouia et al. 2000,2003 Khan et al. 2007a Harada et al. 2002
Lee and Leustekl999 Barroso et al. 1999 Dominguez et al. 2003
Barroso et al. 2006

Oxoglutarate is imported into the chloroplasts (Lancien et al. 2000) where it serves as
the NH4 acceptor by the coupled reaction of giutamine synthase and glutamate
synthase under physiological conditions (Oaks 1994, Ouariti et al. 1997, Gouia et al.
2000). Wahid et al. (2007) reported that Cd-induced inhibition of growth was mainly
due to the damaged photosynthetic apparatus and disruption of the coordination
between C and N metabolism in Vigna radiata. A high rate of CO2 assimilation
favours an efficient N assimilation and vice-versa (Ferrario et al. 1998). Nitrate
reductase and giutamine synthetase enzymes activities are sensitive to Cd stress
whereas, NAD-dependent glutamate dehydrogenase (GDH) shows a substantial rise
under the influence of Cd with dramatic build-up of ammonium pool (Gouia et al.
2003, Boussama et al. 1999). The induction of GDH activity by Cd results from de
novo synthesis and/or activation of specific isoenzymes that removes excess
ammonium (Syntichaki et al. 1996). GDH isoenzymes appear to remove in part the
excess of ammonium under Cd toxicity conditions. Under physiological conditions
the incorporation of ammonium into organic compounds occurs mainly via the
GS/GOGAT cycle (Gouia et al. 2000). The most striking change in Cd treated plants
appears to be a rapid decay of GS/Fd-GOGAT and NADH-GOGAT activities and the
accumulation of ammonia (Boussama et al. 1999, Balestrasse et al. 2006a). This
implies a reduced capacity of GS/GOGAT cycle and induced activity of the
alternative mode of ammonium assimilation by NADH-GDH pathway under Cd
treatment.
Upon exposure to Cd, plants often synthesize a set of N-containing
metabolites through N metabolism, such as proline, glutathione and phytochelatins,
which play a significant role in Cd tolerance of plants (Sharma and Dietz 2006).
Accordingly, plants might exhibit a higher Cd tolerance by the maintenance of normal
N metabolism levels under Cd stress (Gussarsson et al. 1996).
2.8 Yield Characteristics under Cd Stress
Stress factors prevent the plants from reaching their fiill genetic potential and limit the
crop productivity worldwide (Mahajan and Tuteja 2005). A comparison of record
yields and average yields for various crop plants indicates that crops mainly attain
only 20% of their genetic potential for yield due to various biotic and abiotic stress
factors. Abiotic stress is in fact the principal cause of crop failure worldwide, dipping
the average yield for most major crops by more than 50% (Boyer 1982, Bray et al.
2000, Mahajan and Tuteja 2005, Khan et al. 2006).
2 ^ l ^ ^ l f ^

Cadmium affects growth and yield through disturbances in several morpho-
physiological processes and nutrient uptake. Reduction in growth and yield with
increased levels of Cd in growth media arises because of increased leaf rolling and
chlorosis of leaf and stem (Ghani and Wahid 2007) and reduced photosynthetic rate
(Chugh and Sawhney 1999). Wahid and Ghani (2007) reported significant reduction
in number of pods per plant and seeds per pod, 100 seed weight, seed yield and
harvest index of Vigna radiata genotypes as a result of Cd toxicity. It has been
suggested that although varietal difference exists, the accumulated Cd is mainly toxic
to the mesophyll tissue, most probably by interfering with the uptake of essential
nutrients, thereby reducing growth and yield at various stages. Cadmium significantly
reduced the number of ear, ear weight, ear length, spikelet number, grains per ear,
1000-grain weight and grain yield of Triticum aestivum cultivars and the decrease was
correlated with the reduced photosynthetic capacity of the cultivars (Khan et al. 2006,
2007a).
It has been shown that Cd stress significantly reduced grain yield, panicle
number per plant, spikelets per panicle, filled spikelet rate and grain weight of Oryza
sativa genotypes and reduction in yield components of rice genotypes was
proportional to grain Cd content (Cheng et al. 2006). Genotypic differences on the
basis of biomass production, yield and yield components were observed in Triticum
aestivum and it was noted that Cd significantly reduced the root and stem biomass and
spikes per plant but the grains per ear and grain weight were not significantly reduced
(Zhang et al. 2002). They found that the inconsistent decrease in number of grains per
ear and single grain weight among wheat genotypes were because of the
compensation among yield components.
Wu et al. (2004) reported reduction in yield of three Gossypium hirsutum
genotypes under Cd stress and found that the reduction in yield was proportional to
Cd accumulation. The Gossypium hirsutum genotype Simian 3 showed higher Cd
concentration and greater decrease in yield than the other two genotypes (Zhongmian
16 and Zhonmian 16-2). Liu et al. (2007) noted great variation among Oryza sativa
cultivars in their tolerance to soil Cd stress with respect to tillering, plant height, leaf
area, dry matter accumulation and grain yield. The relative change in the number of
grains per panicle showed a strong positive correlation with relative change in grain
yield. Of the four grain yield components measured (panicles per pot, grains per
22

panicle, filled grain percentage, weight per grain), the reduction of grains per panicle
was the main cause of grain yield loss under Cd stress.
2.9 Cadmium-induced Oxidative Stress
Plant cells are continuously exposed to various environmental and biotic stresses
which lead to the increased production of reactive oxygen species (ROS). The
responses of plants to these excess ROS have recently been analyzed extensively at
biochemical and molecular levels (Kreps et al. 2002, Gachomo et al. 2003, Kouril et
al. 2003, Rizhsky et al. 2004, Kotchoni and Gachomo 2006). Under normal
physiological conditions, ROS are continuously produced in the chloroplasts,
mitochondria, peroxisomes as byproducts of aerobic metabolic processes like
photosynthesis, respiration and photorespiration. These ROS are scavenged by both
enzymatic and non-enzymatic antioxidant pathways for the maintenance of normal
plant growth (Jimenez et al. 1998, del Rio et al. 2002, Mittler et al. 2004, Davletova
et al. 2005, Asada 2006, Kotchoni and Gachomo 2006, Nathawat et al. IQQl, Mobin
and Khan 2007, Chalapathi Rao and Reddy 2008). The production of ROS through
Cd-exposed oxidative stress is common in a variety of plants species (Sandalio et al.
2001, Ali et al. 2002, Ranieri et al. 2005, Smeets et al. 2005, Singh et al. 2008a,b).
Oxidative stress occurs when there is a serious imbalance in any cell compartment
between the production of ROS and antioxidant defense, leading to significant
physiological challenges (Foyer and Noctor 2000). These excess ROS cause damage
to proteins, lipids, carbohydrates, DNA and ultimately may result in cell death
(Fadzilla et al. 1997, Gueta-Dahan et al. 1997, Fahmy et al. 1998, Mittler et al. 2004,
Foyer and Noctor 2005, Sairam et al. 2005, Shulaev and Oliver 2006, Djebali et al.
2008).
Photosynthesizing plants are naturally prone to oxidative stress because they
have an array of photosensitizing pigments. These pigments produce and consume
oxygen which can easily donate electrons to form ROS. The chlorophyll pigments
associated with the electron transport system are the primary source of singlet oxygen
('O2). It may also arise as a by product of lipoxygenase activity and is highly
destructive, reacting with most biological molecules at near diffusion-controlled rates.
Superoxides, produced by the transport of electron to oxygen, are not compatible with
metabolism and are required to be eliminated by the antioxidative defense system
while recycling of phosphoglycolate to phosphoglycerate (re-enter the Bassam-Calvin
cycle) results in a considerable loss of assimilated carbon.
23

Cadmium has been found to induce oxidative stress in plants (Hendry et al.
1992, Somashekaraiah et al. 1992, Piqueras et al. 1999, Chien et al. 2001, Dixit et al.
2001, Sandalio et al. 2001, Okamoto et al. 2001, Pereira et al. 2002, Milone et al.
2003, Wu et al. 2003, Kuo and Kao 2004, Skorzynska-Polit et al. 2003/04, Liu et al.
2007, Djebali et al. 2008, Singh et al. 2008a,b). But in contrast with other heavy
metals, such as Cu, it does not seem to act directly on the production of ROS through
Fenton type reactions (Salin 1988). Evidence that Cd causes the production of ROS in
plants (Foyer et al. 1997) arised from the observations that new isozymes of
peroxidases were detectable in both root and leaves of Phaseolus vulgaris (Van
Assche and Clijsters 1990). Further evidence of the Cd-induced oxidative stress arised
from the detection of lipid peroxidation, increased lipoxygenase activity, chlorophyll
degradation and inhibition or stimulation of the activity of several antioxidant
enzymes (Padmaja et al. 1990, Stochs and Bagchi 1995, Shaw 1995a,b, Dixit et al.
2001, lannelli et al. 2002, Leon et al. 2002, Skorzynska-Polit et al. 2003/04, Mobin
and Khan 2007, Agrawal and Mishra 2008, Anjum et al. 2008a). Cadmium provokes
significant disturbances in the structural organization and fiinctional activity of
photosynthetic apparatus (Baszynski 1986, Krupa and Baszynski 1995, Vassilev et al.
1995, Dahlin et al. 2000). The main targets of toxic Cd effects are the pigment
apparatus and photosynthetic gas exchange system (Clijsters and Van Assche 1985,
Tukiendrof and Baszynski 1991, Lang etal. 1995).
Heavy metal exposed plants adopt the process of avoidance of the production
of ROS as the first line of defense against oxidative stress. Once formed, ROS must
be detoxified as efficiently as possible to minimize eventual damage. Thus, the
detoxification mechanisms constitute the second line of defense against the
detrimental effects of ROS (Moller 2001). In fact, compounds having the property of
quenching the ROS without undergoing conversion to a destructive radical can be
described as 'antioxidant'. Antioxidant enzymes are considered as those that either
catalyses such reactions, or are involved in the direct processing of ROS (Medici et al.
2004). Hence, antioxidants (enzymatic and non-enzymatic) function to interrupt the
cascades of imcontrolled oxidation (Noctor and Foyer 1998). Though, the expression
for antioxidant enzymes is altered under stress conditions, their up regulation has a
key role in combating the abiotic stress-induced oxidative stress. However, the level
of up regulation is subject to type and magnitude of the stress. Superoxide dismutase
(SOD), catalase (CAT), ascorbate peroxidase (APX), glutathione reductase (GR),
24

monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR),
guaiacol peroxidase (GOPX) and glutathione S-transferase (GST) showed great
variations in their activities depending on the Cd concentration and the plant species
used.
2.9.1 Lipid peroxidation
Photosynthesizing plants are especially at the risk of oxidative damage, because of
their oxygenic conditions and the abundance of the photosensitizers and
polyunsaturated fatty acids in the chloroplast envelope. It is reported that 1% of the
oxygen consumed by plants is diverted to produce activated oxygen species like
hydroxyl radical (0H-), singlet oxygen ('O2) and superoxide radicals (O2") (Asada
and Takahashi 1987). Free radicals and other derivatives of oxygen are inevitable by
products of biological redox reactions. Their production is considered to be a
universal and common feature of living world under natural conditions as a by
product of respiration and photosynthesis during electron transport systems of
mitochondria and chloroplast. Their concentration increases under unfavorable
conditions. Litracellular structures like membranes and biomolecules like proteins,
enzymes, lipids and DNA have a high degree of organization that is at the risk of
being destructed by these oxidative radicals.
Lipids are more prone to oxidative damage. The peroxidation of lipids is
considered as the most damaging process known to occur in every living organism
(Zhang and Kirkham 1996, Hung and Kao 1998). Membrane damage is sometimes
taken as a single parameter to determine the level of lipid destruction (i.e., lipid
peroxidation). Now, it has been recognized that during the lipid peroxidation,
products are formed from polyunsaturated precursors that include small hydrocarbon
fragments such as ketones, malondialdehyde (MDA), etc. and compounds related to
them (de Vos et al. 1993, Weckx and Clijsters 1996). Some of these compounds react
with thiobarbituric acid (TBA) to form coloured products called thiobarbituric acid
reactive substances (TBARS) that can be measured by monitoring their absorption at
around 530nm (Gray 1978).
Plants exposed to heavy metal stress exhibited an increase in lipid
peroxidation due to the generation of free radicals (Van Assche and Clijsters 1990,
Shaw 1995b, Chaoui et al. 1997b, Lozano-Rodriguez et al. 1997, Vanaja et al. 2000).
Increase in TBARS with increasing Cd concentration has been reported in
germinating Phaseolus vulgaris seedlings (Somashekaraiah et al. 1992). This was
25

related to blockage of electron flow in PSII by metal ions that lead to the formation of
excited Chi which in turn causes the production of free radicals (Kato and Shimizu
1985). Treatment with Cd notably increased the accumulation of lipid peroxides in
Phaseolus vulgaris (Chaoui et al. 1997b), Pisum sativum (Lozano-Rodriguez et al.
1997, Dixit et al. 2001, Metwally et al. 2005), Oryza sativa (Chien et al. 2001, Guo et
al. 2007, Ahsan et al. 2007), different Hordeum vulgare genotypes (Wu et al. 2003),
Helianthus annuus (Gallego et al. 1996a,b, Groppa et al. 2001), Arabidopsis seedlings
(Cho and Seo 2005), Lemna minor (Razinger et al. 2007), Brassica juncea (Mobin
and Khan 2007), Glycine max (Balestrasse et al. 2004, Noriega et al. 10()1),
Bechmeria nivea (Liu et al. 2007), Lycopersicon esculentum (Ammar et al. 2007),
Brassica napus (Filek et al. 2007) and Vigna mungo (Singh et al. 2008a). Contrarily,
decreased rate of lipid peroxidation in peroxisomes of Pisum sativum plants has been
reported due to Cd (Romero-Puertas et al. 1999). However, no lipid peroxidation was
observed in the hairy roots of Daucus carota under Cd stress (Sanita di Toppi et al.
1998). Piqueras et al. (1999) reported an increased lipid peroxidation in BY2 cell
cultures of Lycopersicon esculentum plants exposed to 5mM Cd. The increase was
related to a rapid generation of H2O2 followed by an alteration of the antioxidant
system. Dixit et al. (2001) reported an increase in lipid peroxidation in roots and
leaves of Pisum sativum plants exposed to varying range of Cd in hydroponic system.
Okamoto et al. (2001) observed oxidative damage to lipids in isolated chloroplasts of
unicellular alga, Ganyaulax polyendra under Cd stress.
2.9.2 Hydrogen peroxide content
H2O2 is produced at high flux rates by two processes associated with photosynthesis,
the Mehler reaction and the Glycolate oxidase reaction of photorespiration (Foyer and
Noctor 2000). In addition, there are a number of other enzymes in leaves that are
capable of producing significant amounts of H2O2, including peroxidases, NADPH
oxidases and oxalate oxidase (Bema and Bemier 1999, Bolwell et al. 1999, Sagi and
Fluhr2001).
The accumulation of H2O2 after Cd exposure has been detected in the leaf of
different plant species such as Pisum sativum (Dixit et al. 2001, Romero-Puertas et al.
2004), Arabidopsis thaliana (Cho and Seo 2005), Brassica juncea (Mobin and Khan
2007), Vigna mungo (Singh et al. 2008a). The formation of H2O2 after Cd exposure
has also been detected in Solanum tuberosum tuber discs (Stroinski and Zielezinska
1997), suspension cultures of Nicotiana tabacum cells (Piqueras et al. 1999), and
26

Pinus sylvesths and Pisum sativum roots (Schutzendubel et al. 2001, Romero-Puertas
et al. 2003). Balestrasse et al. (2006b) also reported that Cd increased concentrations
and in situ accumulation of H2O2 and O2" in soybean leaves. Exposure to increasing
Cd concentration increased the H2O2 content in the upper part
(hypocotyls+cotyledons) and roots of Brassica napus (Filek et al. 2007). Guo et al.
(2007) reported that exposure to 50mM Cd significantly increased the H2O2 content in
the roots of Oryza sativa. Zawoznik et al. (2007) reported that leaves of wild type
Arabidopsis thaliana plants exposed to Cd showed accumulation of H2O2. It has also
been reported that Cd increased the accumulation of H2O2 in soybean root tips (Yang
era/. 2007).
2.10 Cadmium-tolerance Mechanism in Plants
Plants adopt several strategies such as physically avoiding the metal-contaminated
environment. These include making exudates of complexing agents into rhizosphere
region, binding metal ions in the cell wall, effluxing the metal ions from the
symplasm, preventing the upward transport of metal ions to the above ground parts,
transporting metal-peptide/ligand complexes into the vacuole, storing metal ions in
the vacuoles by complexation with vacuolar peptides/ligands and or forming metal-
resistant enzymes or metabolites to minimize metal-induced severe internal metabolic
injuries (Hall 2002, McGrath and Zhao 2003, Gratao et al. 2005, Zhang et al. 2006).
Plants have evolved a complex array of mechanisms to maintain optimal metal
levels and avoid the detrimental effects of excessively high concentrations (Clemens
2001). When these homeostatic mechanisms are overwhelmed, plants suffer metal-
induced damage and pro-oxidant conditions within cells. However, higher plants are
very well equipped with antioxidant mechanisms (Mittler et al. 2004, Gratao et al.
2006, Singh et al. 2008b). Plant cells display an antioxidant network including
numerous soluble and membrane compounds, particularly in mitochondria and in
chloroplasts where respiratory and photosynthetic electron transfer chains take place.
Antioxidant enzymes are considered as those that either catalyze such reactions, or are
involved in the direct processing of ROS (Medici et al. 2004). Plants possess very
efficient enzymatic (SOD, CAT, APX, GR, MDHAR, DHAR, GPX, GOPX and GST)
and non-enzymatic (AsA, GSH, phenolic compounds, alkaloids, non-protein amino
acids and a-tocopherols) antioxidant defense systems. Components of antioxidant
defense systems control the cascades of uncontrolled oxidation (Noctor and Foyer
1998) and protect plant cells from oxidative damage by scavenging of ROS.
27 ^^^iV-::-^-^

2.10.1 Enzymatic antioxidant system
2.10.1.1 Superoxide dismutase
Superoxide dismutase (SOD; EC 1.15.1.1) was first isolated by Mann and Keilin
(1938) and thought to be a copper-storage protein. Subsequently, it was identified by
different names, erythrocuprein, indophenol oxidase, and tetrazolium oxidase until its
catalytic function was discovered by McCord and Fridovich (1969). SOD catalyzes
the disproportionation of superoxide (02~) to hydrogen peroxide and molecular
oxygen. It removes Or' and hence decreases the risk of hydroxyl radical formation
fi-om 02~ via the metal catalyzed Haber-Weiss-type reaction.
Sensitive to SOD isozymes
Fe-SOD
Mn-SOD
Cu/Zn-SOD
Location
Chloroplast
Mitochondria and
Peroxisomes
Resistant to
KCN
KCN and H2O2
Chloroplast and Cytosol
H202
H2O2 and KCN
There are three distinct types of SOD classified on the basis of the metal
cofactor: the copper/zinc (Cu/Zn-SOD), the manganese (Mn-SOD) and the iron (Fe-
SOD) isozymes have been reported in various plant species (Bannister et al. 1987,
Alscher et al. 2002). These isozymes can be separated by native polyacrylamide gel
electrophoresis. Their activity is detected by negative staining and identified on the
basis of their sensitivity to KCN and H2O2. The Mn-SOD is resistant to both
inhibitors; Cu/Zn-SOD is sensitive to both inhibitors whereas; Fe-SOD is resistant to
KCN and sensitive to H2O2. The subcellular distribution of these isozymes is also
distinctive. The Mn-SOD is found in the mitochondria of eukaryotic cells and in
peroxisomes (del Rio et al. 2003); some Cu/Zn-SOD isozymes are found in the
cytosolic fractions, and also in chloroplasts of higher plants (del Rio et al. 2002). The
Fe-SOD isozymes, often not detected in plants (Ferreira et al. 2002) are usually
associated with the chloroplast compartment when present (Bowler et al. 1992,
Alscher et al. 2002). The prokaryotic Mn-SOD and Fe-SOD, and the eukaryotic
Cu/Zn-SOD enzymes are dimers, whereas Mn-SOD of mitochondria are tetramers
(Scandalios 1993). Peroxisomes and glyoxysomes of Citrillus vulgaris have been
shown to contain both Cu/Zn- and Mn-SOD activity (Sandalio and del Rio 1988), but
there are no reports of extracellular SOD enzymes in plants. All forms of SOD are
28

nuclear-encoded and targeted to their respective subcellular compartments by an
amino terminal targeting sequence. Several forms of SOD have been cloned from a
variety of plants (Scandalios 1990, Bowler et al. 1992). The response of SOD to
heavy metal stress varies considerably depending upon plant species, stage of the
plant development, metal in the experiment and the exposure time. SOD activity in
leaves exhibited increase in response to Cd, whereas, in roots there was no significant
variation in its activity was noted (Vitoria et al. 2001). Activity staining for SOD in
Glycine max revealed seven isozymes in leaves and eight in roots, corresponding to
Mn-SOD and Cu/Zn SOD isozymes. Although a clear effect of Cd on plant growth
was observed, the activities of the SOD isozymes were unaltered (Ferreira et al.
2002). In Saccharum officinarum seedlings several isozymes have been observed, but
growth in the presence of Cd did not result in any significant alteration in SOD
activity (Fomazier et al. 2002a). In Pisum sativum plants, a strong reduction in
chloroplastic and cytosolic Cu/Zn SODs by Cd was reported and to a lesser extent for
Fe-SOD, while Mn-SOD was only affected by the highest Cd concentration tested.
This showed that Mn-SOD was the isozyme more resistant to Cd (Sandalio et al.
2001). In Pisum sativum leaf peroxisomes, the Mn-SOD activity did not change in
response to Cd treatment (Romero-Puertas et al. 1999). Contrarily, increases in total
SOD activity were detected following the application of Cd in Pisum sativum
(Dalurzo et al. 1997), Solarium tuberosum (Stroinski and Kozlowska 1997), Hordeum
vulgare (Guo et al. 2004), Arabidopsis thaliana (Skorzynska-Polit et al. 2003/04),
Oryza sativa (Hsu and Kao 2004), Triticum aestivum (Khan et al. 2007a), Zea mays
(Krantev et al. 2007), Brassica juncea (Mobin and Khan 2007), Vigna mungo (Singh
et al. 2008a), Cicer arietinum (Hasan et al. 2008), Vigna radiata (Anjum et al.
2008c), Brassica campestris (Anjum et al. 2008d) and hyperaccumulator plants of the
genus Alyssum (Schickler and Caspi 1999). SOD activity remained unaltered in
Helianthus annuus (Gallego et al. 1996b, 1999) and declined in Amaranthus lividus
(Bhattacharjee 1998), Phragmites australis (lannelli et al. 2002), Capsicum annuum
plants (Leon et al. 2002), Glycine max (Noriega et al. 2007) under Cd stress. In
clonal, hydroponically grown poplar plants {Populus x canescens, a hybrid of
Populus termula x Populus alba) (Schutzendubel et al. 2002) and Arabidopsis lividus
(Bhattachagee 1998) exposure to Cd resulted in inhibition of SOD activity. Romero-
Puertas et al. (2004) studied the involvement of H2O2 and O2" in the signaling events
29

that lead to the variation of the transcript levels of Cu/Zn-SOD in Pisum sativum
plants under Cd stress.
2.10.1.2 Catalase
Catalase (CAT; EC 1.11.1.6) is a heme-containing enzyme that catalyzes the
dismutation of hydrogen peroxide into water and oxygen (Frugoli et al. 1996). Present
in all aerobic eukaryotes, this is important in the removal of hydrogen peroxide
generated in peroxisomes (microbodies) by oxidases involved in P-oxidation of fatty
acids, the glyoxylate cycle (photorespiration) and purine catabolism. CAT is one of
the first enzymes isolated in a highly purified state. The isozymes of catalase have
been studied extensively in higher plants (Polidoros and Scandalios 1999). Scandalios
et al. (2000) characterized three genetically distinct CAT isozymes in maize plants.
All forms of the enzyme are tetramers in excess of 220,000 molecular weight.
Multiple forms of catalase have been described in many plants. Zea mays has 3
isoforms termed as CAT 1, CAT 2 and CAT 3, and are found on separate
chromosomes and are differentially expressed and independently regulated
(Scandalios 1990). CAT 1 and CAT 2 aie localized in peroxisomes and the cytosol,
whereas CAT 3 is mitochondrial. Plants contain multiple CAT isozymes e.g., 2 in
Hordeum vulgare (Azevedo et al. 1998), 4 in Helianthus annuus cotyledons
(Azpilicueta et al. 2007) and as many as 12 isozymes in Brassica (Frugoli et al.
1996). Catalase isozymes have been shown to be regulated temporally and spatially
and may respond differentially to light (Willekens et al. 1994, Skadsen et al. 1995).
The variable response of CAT activity has been observed under Cd stress.
CAT activity declined in Helianthus annuus leaves (Gallego et al. 1996b), Phaseolus
vulgaris (Chaoui et al. 1997b), Phaseolus aureus (Shaw 1995b), Pisum sativum
(Daliu^o et al. 1997), Lemna minor (Mohan and Hossetti 1997), Amaranthus lividus
(Bhattacharjee 1998), Glycine max roots (Balestrasse et al. 2001), Phragmites
australis (lannelli et al. 2002), Capsicum annuum (Leon et al. 2002) and Arabidopsis
thaliana (Cho and Seo 2005) under Cd stress conditions. A significant decline in CAT
activity was reported after 50|aM Cd applications for 48h in the roots and shoots of
Bacopa monnieri (Singh et al. 2006). However, CAT activity increased in Agropyron
repens (Brej 1998), Helianthus annuus (Gallego et al. 1999), Glycine max nodules
(Balestrasse et al. 2001), Oryza sativa leaves (Hsu and Kao 2004), in tolerant varieties
oiSolanum tuberosum (Stroinski and Kozlowska 1997), in roots of Raphanus sativus
seedlings (Vitoria et al. 2001), Brassica juncea (Mobin and Khan 2007), Triticum
30

aestivum (Khan et al. 2007a), Vigna mungo roots (Singh et al. 2008a), Vigna radiata
(Anjum et al. 2008c), Brassica campestris (Anjum et al. 2008d) and Cicer arietinum
(Hasan et al. 2008). Azpihcueta et al. (2007) reported that incubation of Helianthus
annuus leaf discs with 300 and 500|iM CdCh under light conditions increased CATA3
transcript level but this transcript was not induced by Cd in etiolated plants.
Moreover, in roots of the transgenic CAT-deficient tobacco lines {CAT IAS), the
DNA damage induced by Cd was higher than in wild type tobacco (SR 1) roots
(Gichner et al. 2004). Furthermore, CAT activity remained unaltered under Cd stress
in Glycine max leaves (Ferreira et al. 2002).
2.10.1.3 Ascorbate peroxidase
Ascorbate peroxidase (APX; EC 1.11.1.11) is a heme protein, and its primary function
is the rapid removal of H2O2 at the site of generation (Asada 1992). APX isozymes
are distributed in at least four distinct cell compartments, the stroma (sAPX),
thylakoid membrane (tAPX), the mitochondria (mAPX), and the cytosol (cAPX)
(Asada 1992, Miyake and Asada 1992, Ishikawa et al. 1998). The various isoforms of
APX respond differentially to metabolic and environmental signals (Kubo et al.
1995). Thylakoid membrane-bound APX is limiting factor of antioxidant systems
under photoxidative stress in chloroplasts and the enhanced tAPX activity maintain
the redox status of ascorbate under stress conditions (Yabuta et al. 2002).
Chloroplasts contain APX in two isoforms, thylakoid-bound and soluble stromal
enzymes. At least one half of the chloroplastic APX is tAPX, but the ratio of
tAPX/sAPX varies according to the plant species, possibly, leaf age, but the
biosynthetic ratio of the two APXs is controlled by alternative splicing (Asada 1999).
tAPX bind to the stroma thylakoids where the PSI complex is located, while sAPX is
thought to be localized in the stroma (Asada 1999). Plants also contain the cytosolic
isoforms of APX (cAPX), which has a different amino acid sequence in comparison
to chloroplastic APXs, but participate in the scavenging of H2O2 in compartments
other than chloroplasts. cAPX is a homodimer and its electron donor is not so specific
for ascorbate, unlike tAPX and sAPX (Asada 1999).
APX has an important role in the scavenging of H2O2 under stressed
conditions but its activity depends on the Cd concentration applied. Increased leaf
APX activity under Cd stress has been reported in Ceratophyllum demersum (Aravind
and Prasad 2003), Brassica juncea (Mobin and Khan 2007), Pisum sativum (Romero-
Puertas et al. 1999), Phaseolus aureus (Shaw 1995b), Phaseolus vulgaris (Chaoui et
31

al. 1997b), Zea mays (Krantev et al. 2007), Triticum aestivum (Khan et al. 2007a),
Vigna radiata (Anjum et al. 2008c), Brassica campestris (Anjum et al. 2008d) and
Vigna mungo (Singh et al. 2008a), however, in Hordeum vulgare roots, the APX
activity was reduced at high concentration of Cd (Hegedus et al. 2001). Balestrasse et
al. (2001) reported that low Cd levels led to an increased APX activity in Glycine max
roots and nodules, but the activity decreased with high Cd concentration. The lower
APX activity was also noted in Cucumis sativus chloroplasts with increasing Cd
concentration (Zhang et al. 2003). Whereas, Cd induced inhibition of APX activity
was also observed in clonal, hydroponically grown Populus x canescens.
(Schutzendubel et al. 2002) and Helianthus annuus plants (Gallego et al. 1996b).
APX activity in Ceratophyllum demersum showed a very high increase in its activity
in Cd+Zn treated plants as compared to Cd or Zn alone, indicating efficient
antioxidant and ROS scavenging activities by Zn against Cd induced free radicals and
oxidative stress (Aravind and Prasad 2003). Khan et al. (2008b) also reported
increased APX activity in Triticum aestivum plants treated with Cd under low Zn
levels.
2.10.1.4 Glutathione reductase
Glutathione reductase (GR; EC 1.6.4.2) is a flavo-protein oxidoreductase, found in
both prokaryotes and eukaryotes (Mullineaux and Creissen 1997, Romero-Puertas et
al. 2006). This enzyme was first reported in eukaryotes and yeast (Meldrum and Tarr
1935) as well as in plants (Corm and Vennesland 1951, Mapson and Goddord 1951).
GR maintains the balance between reduced glutathione (GSH) and ascorbate pools,
which in turn maintain cellular redox state (Lascano et al. 1999, 2001, Ansel et al.
2006, Reddy and Raghavendra 2006, Romero-Puertas et al. 2006, Chalapathi Rao and
Reddy 2008). The enzyme protein, although synthesized in the cytoplasm, can be
targeted to both chloroplast and mitochondria (Mullineaux and Creissen 1997). In
higher plants, GR is involved in defense against oxidative stress, whereas, GSH plays
an important role within the cell system, which includes participation in the
Ascorbate-GIutathione cycle, maintenance of the sulfhydryl (-SH) group and a
substrate for glutathione-S-transferases (Noctor et al. 2002, Reddy and Raghavendra
2006). Glutathione reductase and GSH play a crucial role in determining the tolerance
of a plant during environmental stresses (Chalapathi Rao and Reddy 2008). In almost
all the biological functions, GSH is oxidized to GSSG which should be converted
back to GSH in plant cell to perform normal physiological functions. Hence, rapid
32

recycling of GSH is more essential rather than synthesis of GSH, which is a highly
regulated and ATP requiring process. GR activity increases as part of the defense
against Cd-exposure, which is dose-dependent and variable over time (Fomazier et al.
2002a). GR activity increased in the presence of Cd in Phaseolus vulgaris (Chaoui et
al. 1997b), Solarium tuberosum (Stroinski and Kozlowska 1997), Raphanus sativus
(Vitoria et al. 2001), Crotolaria juncea (Pereira et al. 2002), Glycine max (Ferreira et
al. 2002), Saccharum ojficinarum (Fomazier et al. 2002b), Capsicum annuum (Leon
et al. 2002), Arabidopsis thaliana (Skorzynska-Polit et al. 2003/4), Triticum aestivum
(Khan et al. 2007a), Brassica juncea (Mobin and Khan 2007), Brassica campestris
(Anjum et al. 2008d) and Vigna mungo (Singh et al. 2008a). In Raphanus sativus, it
exhibited very little variation in the roots and leaves of control plants, indicating a
direct correlation with Cd accumulation (Vitoria et al. 2001). In Pisum sativum, GR
activity was enhanced more with 40nM than with 4)4,M Cd (Dixit et al. 2001).
However, a decrease in GR activity after application of Cd has also been reported for
a few plant species such as Helianthus annuus (Gallego et al. 1996a,b), Pisum
sativum (Dalurzo et al. 1997) and Solanum tuberosum (Stroinski and Kozlowska
1997).
2.10.1.5 Monodehydroascorbate reductase
Monodehydroascorbate reductase (MDHAR; EC 1.6.5.4) is a flavin adenin
dinucleotide (FAD) enzyme that is present as chloroplastic and cytosolic isozymes
which share similar properties. Monodehydroascorbate reductase exhibits a high
specificity for monodehydro asorbate (MDHA) as the electron acceptor, preferring
NADH rather than NADPH as the electron donor. Asada (1999) studied the multi-step
reduction of FAD in detail. The first step is the reduction of the enzyme-FAD to form
a charge transfer complex. The reduced enzyme donates electrons successively to
MDHA, producing two molecules of ascorbate via a semiquinone form [E-FAD-
NADP(P)' ]. The disproportionation by photoreduced ferredoxin (redFd) in the
thylakoids is of great importance. Since redFd (reduced ferredoxin) can reduce
MDHA more effectively than NADP" , MDHAR caimot participate in the reduction of
MDHA in the thylakoidal scavenging system. Therefore, MDHAR would function at
a site where NAD(P)H is available, but redFd is not (Asada 1999).
MDHAR is also located in peroxisomes and mitochondria accompanying
APX. MDHAR could remove and scavenge H2O2 in these organelles similar to that in
chloroplasts, which has escaped from break down by peroxisomal CAT (del Rio et al.
33

2002). Schutzendubel et al. (2001) have noted enhanced MDHAR activity in Cd-
exposed Pinus sylvestris and a declined MDHAR activity in Cd-exposed poplar
hybrids {Populus x canescens).
2.10.1.6 Glutathione S-transferase
Glutathione S-transferases (GSTs; EC 2.5.1.18) catalyze the conjugation of tripeptide
GSH into a variety of hydrophobic, electrophylic and cytotoxic substrates (Marrs
1996). Noctor et al. (2002) observed that GSTs can remove genotoxic or cytotoxic
compounds that have potential to damage or react with genetic material (DNAs and
RNAs) and protein. In fact, glutathione-S-transferases can reduce peroxides with the
help of GSH and produce scavengers of cytotoxic and genotoxic compounds. An
increased GST activity was found in leaves and roots of Cd-exposed Pisum sativum
plants by Dixit et al. (2001) and in roots of Oryza sativa and Phragmites australis
plants (lannelli et al. 2002, Moons 2003).
2.10.1.7 Glutathione peroxidase
H2O2 can be metabolized by another plant peroxidase-scavenging enzyme called
glutathione peroxidase (GPX; EC 1.11.1.9) (Noctor et al. 2002). Millar et al. (2003)
identified a family of seven related proteins in cytosol, chloroplast, mitochondria and
endoplasmic reticulum, named AtGPXl-AtGPX7 in Arabidopsis. Stress increases
GPX activity in cultivars of Capsicum annuum plants (Leon et al. 2002) but decreases
in roots and causes no significant change in the leaves of Cd-exposed Pisum sativum
plants (Dixit e/a/. 2001).
2.10.1.8 Guaiacol peroxidase
Ascorbate peroxidase (APX) can be distinguished fi-om plant-isolated guaiacol
peroxidase (GPOX; EC 1.11.1.7) in terms of differences in sequences and
physiological fiinctions (Chen et al. 1992). GPOX decomposes indole acetic acids
(lAAs) and has a role in the biosynthesis of lignin and defense against pathogens by
consuming H2O2. GPOX prefers aromatic electron donors such as guaiacol and
pyragallol usually oxidizing ascorbate at the rate of around 1% that of guaiacol
(Asada 1999).
The activity of GPOX varies considerably depending upon plant species and
the concentration of Cd used. It increased in Cd-exposed plants of Triticum aestivum
(Milone et al. 2003), Arabidopsis thaliana (Cho and Seo 2005) and Ceratophyllum
demersum (Aravind and Prasad 2003) and decreased in Cd-exposed Pisum sativum
plants (Sandalio et al. 2001). Radotic et al. (2000) noted an initial increase in GPOX
34

activity in spruce needles subjected to Cd stress and subsequent Cd-treatments caused
a decline in the activity.
2.10.2 Non-enzymatic antioxidant system
2.10.2.1 Ascorbic acid
All plants can synthesize ascorbate, which can accumulate to millimolar
concentrations in both photosynthetic and non-photosynthetic tissues (Foyer et al.
1983). Ascorbate is one of the most powerful antioxidants (Noctor and Foyer 1998,
Smirnoff et al. 2001), reacts directly with hydroxyl radicals, superoxide and singlet
oxygen and reduces H2O2 to water via ascorbate peroxide reaction (Noctor and Foyer
1998). Ascorbate also acts as an electron donor in the regeneration of a-tocopherol.
Under physiological conditions it exists mostly in reduced form in leaves and
chloroplast and its intracellular concentration can build up to millimolar range (viz.,
20mM in the cytosol and 20-300niM in the chloroplast stroma) (Foyer and Lelandais
1996). The ability to donate electrons in a wide range of enzymatic and non-
enzymatic reactions makes ascorbate the main ROS-detoxifying compound in the
aqueous phase. In addition to the importance of ascorbate in the Ascorbate-
Glutathione cycle, it plays a role in preserving the activities of enzymes that contain
prosthetic transition metal ions (Noctor and Foyer 1998). The ascorbate redox system
consists of L-ascorbic acid, MDHA and DHA. Both oxidized forms of ascorbate are
relatively unstable in aqueous environments while DHA can be chemically reduced
by GSH to ascorbate (Foyer and Halliwell 1976). Evidence to support the actual role
of DHAR, GSH and GR in maintaining the foliar ascorbate pool has been observed in
transformed plants overexpressing GR (Foyer et al. 1995). Nicotiana tabacum and
Populus X canescens plants have higher foliar ascorbate contents and improved
tolerance to oxidative stress (Aono et al. 1993, Foyer et al. 1995). Demirevska-
Kepova et al. (2006) reported that the content of oxidized ascorbate increased during
Cd exposure in Hordeum vulgare plants. A decrease in the ascorbate content in the
roots and nodules of Glycine max under Cd stress has been observed (Balestrasse et
al. 2001). Cadmium also decreases ascorbate content in Cucumis sativus chloroplast
and in the leaves of Arabidopsis thaliana, Pisum sativum and Brassica campestris
(Zhang et al. 2003, Skorzynska-Polit et al. 2003/04, Romero-Puertas et al. 2007,
Anjum et al. 2008a,d), respectively, whereas, it remained unaffected in Populus x
canescens roots (Schutzendubel et al. 2002).
35

2.10.2.2 Glutathione
The tripeptide (y-GluCysGly) glutathione (GSH) plays a central role in several
physiological processes, including regulation of sulfate transport, signal transduction,
conjugation of metabolites, detoxification of xenobiotics (Xiang et al. 2001) and the
expression of stress-responsive genes (Muliineaux and Rausch 2005). The reduced
form of glutathione, GSH, is an abundant compound in plant tissue that exists
interchangeably with the oxidized form, GSSG. GSH is abundant (3-lOmM) in
cytoplasm, nuclei and mitochondria and is the major soluble antioxidant in these cell
compartments. GSH has been associated with several growth and development related
events in plants, including cell differentiation, cell death and senescence, pathogen
resistance and enzymatic regulation (Ogawa 2005, Rausch and Wachter 2005) and its
content is affected by S nutrition (Blake-Kalff e/ al. 2000). GSH is the major reservoir
of non-protein sulfur. It is the major redox buffer in most aerobic cells, and plays an
important role in physiological functions, including redox regulation, conjugation of
metabolites, detoxification of xenobiotics and homeostasis and cellular signaling that
triggers adaptive responses (Noctor et al. 2002, Kopriva and Koprivova 2005). It also
plays an indirect role in protecting membranes by maintaining a-tocopherol and
zeaxanthin in the reduced state. It can also function directly as a free radical
scavenger by reacting with superoxide, singlet oxygen and hydroxyl radicals. GSH
protects proteins against denaturation caused by the oxidation of protein thiol groups
under stress. In addition, GSH is a substrate for glutathione peroxidase (GPX) and
glutathione-S-transferases (GST), which are also involved in the removal of ROS
(Noctor et al. 2002). GSH is a precursor of PCs, which are crucial in controlling
cellular heavy metal concentrations. GSH and its oxidized form, GSSG maintains a
redox balance in the cellular compartments. This property of glutathione is of great
biological importance since it allows fine-tuning of the cellular redox environment
under normal conditions and upon onset of stress, and provides the basis for GSH
stress signaling. A central nucleophilic Cys residue is responsible for higher reductive
potential of GSH. It scavenges cytotoxic H2O2, and reacts non-enzymatically with
other ROS i.e., O2", OH- and 'O2 (Larson 1988). The central role of GSH in the
antioxidative defense system is due to its ability to regenerate another water soluble
antioxidant, ascorbate, in Ascorbate-Glutathione cycle (Foyer and Halliwell 1976,
Noctor and Foyer 1998). The role of GSH in the antioxidant defense system provides
a strong basis for its use as a stress marker. However, the concentration of cellular
36

GSH has a major effect on its antioxidant function and it varies considerably under Cd
stress. Furthermore, strong evidence has indicated that an elevated GSH concentration
is correlated with the ability of plants to withstand metal-induced oxidative stress
(Freeman et al. 2004). Xiang et al. (2001) observed that plants with low levels of
glutathione were highly sensitive to even low levels of Cd due to limited PC
synthesis. The increased demand for GSH can be met by the activation of pathways
involved in S assimilation and Cys biosynthesis. Its concentration is controlled by a
complex homeostatic mechanism where the availability of S seems to be required
(May et al. 1998a). Manipulation of GSH biosynthesis increases resistance to
oxidative stress (Youssefian et al. 2001, Sirko et al. 2004). It has been observed that
upon Cd exposure, one of the mam responses observed was the induction of genes
involved in S assimilation-reduction and GSH metabolism in the roots of Arabidopsis
(Herbette et al. 2006).
Feed back inhibition of y-glutamylcysteine synthase (y-ECS) by GSH has been
considered as a fundamental central point for GSH synthesis. In vitro studies with the
enzymes from tobacco and parsley cells showed that the plant y-ECS was inhibited by
GSH (Noctor and Foyer 1998). Oxidation of GSH to GSSG decreases GSH levels and
allows increased y-ECS activity under stressed conditions (Noctor and Foyer 1998).
Environmental stresses trigger an increase in ROS levels in plants and the
response of GSH can be crucial for adaptive responses. Antioxidant activity in the
leaves and chloroplast of Phragmites australis Trin. (cav.) ex Steudel was associated
with a large pool of GSH, protecting the activity of many photosynthetic enzymes
against the thiophilic bursting of Cd exerting a direct important protective role in the
presence of Cd (Pietrini et al. 2003). Increased concentration of GSH has been
observed wdth the increasing Cd concentration in Pisum sativum (Gupta et al. 2002),
Lactuca sativa (Maier et al. 2003), Phragmites australis (Pietrini et al. 2003),
Brassica juncea (Qadir et al. 2004), Pisum sativum (Metwally et al. 2005), Sedum
alfredii (Sun et al. 2007), Oryza sativa (Hassan et al. 2008). However, decay in GSH
content in Glycine max roots (Balestrasse et al. 2001), Helianthus annuus leaves
(Gallego et al. 1996b), Zea mays seedlings (Rauser 1990), Pisum sativum
(Ruegsegger et al. 1990), Pinus sylvestris roots (Schutzendubel et al. 2001), Cucumis
sativus chloroplast (Zhang et al. 2003), Populus x canescens roots (Schutzendubel et
al. 2002) and Oryza sativa leaves (Hsu and Kao 2004) has been reported under Cd
stress. Furthermore, unaltered GSH content was observed in the nodules of
37

Table 2. Antioxidative enzymes in economically important crop plants grown under cadmium stress. Symbols used: - - Decrease; + + Increase/overexpressed; x x Unaltered
Antioxidants
Superoxide dismutase (SOD)
Ascorbate peroxidase (APX)
Plant species/ part studied
Raphanus sativus leaf
Raphanus sativus root Pisum sativum leaf Solarium tuberosum
Hordeum vulgare Arabidopsis thaliana
Oryza sativa leaves Alyssum spp
Helianthus annuus Amaranthus lividus Phragmites australis Helianthus annuus leaves Capsicum annuum plant Bacopa monnieri Vigna mungo roots Vigna mungo leaves Triticum aestivum leaves Brassicajuncea leaves
Brassica campestris leaves Zea mays Cicer arietinum Pisum sativum
Oryza sativa Arabidopsis thaliana Oryza sativa shoots and roots Oryza sativa roots Zea mays Glycine max Brassica napus Ceratophyllum demersum Pisum sativum leaves
Phaseolus aureus
Response
+ +
X X
+ + + +
+ + + +
+ + + +
X X
— —
—
+ +
+ + + + + +
+ +
+ + + + + +
+ + + + + +
+ + — —
+ +
+ +
+ +
Reference (s)
VitoriaeM/. 2001
Vitoriae/a/. 2001 DalurzoeM/. 1997 Stroinski and Kozlowska 1997 Guo et al. 2004 Skorzynska-Polit et al. 2003/04 Hsu and Kao 2004 Schickler and Caspi 1999 Gallegoera/. 1999 Bhattacharjee 1998 lannelli et al. 2002 Gallego et al. 1996b Leon et al. 2002 Mishra^/a/. 2006 Singh et al. 2008a Singh et al. 2008a Khan et al. 2007a Mobin and Khan 2007 Anjum et al. 2008d
Ekmekci et al. 2008 Hasan et al. 2008 Agrawal and Mishra 2008 Shah e/a/. 2001 Cho and Seo 2005 Wu et al. 2006
Guo et al. 2007 Krantev et al. 2007 Noriega et al. 2007 Filek et al. 2007 Aravind and Prasad 2003 Romero-Puertas et al. 1999 Shaw 1995b

Catalase (CAT)
Hordeum vulgare roots Ceratophyllum demersum Glycine max roots and nodules Cucumis chloroplast Populus plant
Vigna mungo leaves and roots Triticum aestivum leaves Zea mays
Helianthus annuus leaves Brassicajuncea leaves
Brassica campestris leaves Arabidopsis thaliana Bacopa monnieri Brassica napus Helianthus annuus leaves Phaseolus vulgaris Phaseolus aureus Pisum sativum Lemna minor
Amaranthus lividus Glycine max leaves
Phargmitis australis Capsicum annuum Agropyron repens Glycine max nodules
Helianthus annuus
Oryza sativa leaves Solanum tuberosum
Raphanus sativus roots Saccharum officinarum In vitro callus Glycine max leaves Phaseolus vulgaris roots and leaves Bacopa monnieri
—
+ +
+ +
—
—
+ +
+ + + +
—
+ +
+ +
+ + + + + + —
—
—
—
—
—
—
—
—
+ + + +
+ +
+ + + +
+ + + +
X X
—
—
Hegeduse/a/. 2001 Aravind and Prasad 2003 Balestrasse et al. 2001 Zhang et al. 2003 Schutzendubel et al. 2002 Singh et al. 2008a
Khan et al. 2007a Krantev et al. 2007, Ekmekci et al. 2008 Gallego et al. 1996b, 2002 Mobin and Khan 2007 Anjum et al. 2008d
Cho and Seo 2005 Mishra et al. 2006 Filek et al. 2007 Gallego et al. 1996b Chaouie/a/. 1997b Shaw 1995b Dalurzoe/a/. 1997 Mohan and Hossetti 1997 Bhattacharjee 1998 Balestrasse et al. 2001 lannelli et al. 2002 Leon et al. 2002 Brej 1998 Balestrasse et al. 2001 Gallego e/a/. 1999, 2002 Hsu and Kao 2004 Stroinski and Kozlowska 1997 Vitoriae^a/. 2001 Fomazier et al. 2002b Ferreira et al. 2002 Chaouie/a/. 1997b
Mishra et al. 2006
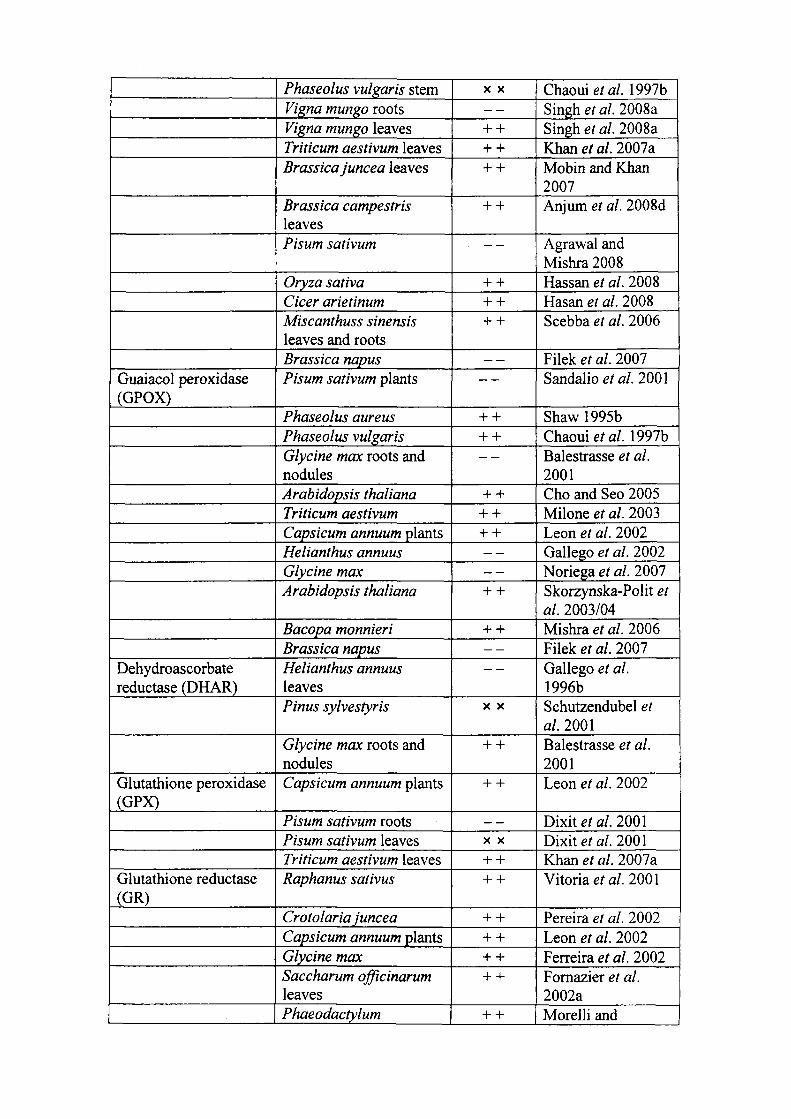
Guaiacol peroxidase (GPOX)
Dehydroascorbate reductase (DHAR)
Glutathione peroxidase (GPX)
Glutathione reductase (GR)
•
Phaseolus vulgaris stem Vigna mungo roots Vigna mungo leaves Triticum aestivum leaves Brassicajuncea leaves
Brassica campestris leaves Pisum sativum
Oryza sativa Cicer arietinum Miscanthuss sinensis leaves and roots Brassica napus Pisum sativum plants
Phaseolus aureus Phaseolus vulgaris Glycine max roots and nodules Arabidopsis thaliana Triticum aestivum Capsicum annuum plants Helianthus annuus Glycine max Arabidopsis thaliana
Bacopa monnieri Brassica napus Helianthus annuus leaves Pinus sylvestyris
Glycine max roots and nodules Capsicum annuum plants
Pisum sativum roots Pisum sativum leaves Triticum aestivum leaves Raphanus sativus
Crotolaria juncea Capsicum annuum plants Glycine max Saccharum officinarum leaves Phaeodactylum
X X
—
+ + + + + +
+ +
—
+ + + + + +
—
—
+ + + + —
+ + + + + + — —
+ +
+ + —
—
X X
+ +
+ +
X X
+ + + +
+ + + + + + + +
+ +
Chaouiera/. 1997b Singh et al. 2008a Singh et al. 2008a Khan e/a/. 2007a Mobin and Khan 2007 Anjumera/. 2008d
Agrawal and Mishra 2008 Hassan et al. 2008 Hasan et al. 2008 Scebba et al. 2006
Filek et al. 2001 Sandalio^/flf/. 2001
Shaw 1995b Chaouie/a/. 1997b Balestrasse et al. 2001 Cho and Seo 2005 Milone et al. 2003 Leon et al. 2002 Gallegoe/a/. 2002 Noriega et al. 2007 Skorzynska-Polit et al. 2003/04 Mishra et al. 2006 Filek et al. 2007 Gallego et al. 1996b Schutzendubel et al. 2001 Balestrasse et al. 2001 Leon et al. 2002
Dixit era/. 2001 Dixit era/. 2001 Khan et al. 2007a Vitoriaera/. 2001
Pereira et al. 2002 Leon et al. 2002 Ferreira et al. 2002 Fomazier et al. 2002a Morelli and

Glutathione S-transferase (GST)
Monodehydroascorbate reductase (MDHAR)
Ascorbic acid (AsA)
Glutathione (GSH)
tricomutum Pisum sativum roots and leaves Alyssum sp.
Cucumis chloroplast Pisum sativum Solarium tuberosum
Helanthus annuus
Viciafaba shoots Bacopa monnieri Viciafaba roots Vigna mungo leaves and roots Triticum aestivum leaves Brassicajuncea leaves
Brassica campestris leaves Zea mays Arabidopsis thaliana Pisum sativum plants
Oryza sativa roots Phragmites australis Pinus sylvestris
Populus canescens (poplar hybrid roots) Glycine max roots and nodules Arabidopsis thaliana leaves Cucumis chloroplast Populus canescens roots
Brassica juncea leaves Pisum sativum
Brassica campestris leaves Oryza sativa roots Sedum alfredii
Pisum sativum
+ +
—
—
— —
—
+ + + + —
+ +
+ + + +
+ +
+ + + + + +
+ + + + + +
—
—
—
— X X
—
—
+ + + +
+ +
Scarano 2004 Dixit e/a/. 2001
Schickler and Caspi 1999 Zhang et al. 2003 DaluTzo etal. 1997 Stroinski and Kozlowska 1997 Gallego et al. 1999b Rosa et al. 2003 Mishra et al. 2006 Rosa et al. 2003 Singh et al. 2008a
Khan et al. 2007a Mobin and Khan 2007 Anjum et al. 2008d
Ekmekci et al. 2008 Cho and Seo 2005 Dixit etal. 2001
Moons 2003 lannelli et al. 2002 Schutzendubel et al. 2001 Schutzendubel et al. 2001 Balestrasse et al. 2001 Skorzynska-Polit et al. 2003/04 Zhang et al. 2003 Schutzendubel et al. 2002 Qadir et al. 2004 Romero-Puertas et al. 2007, Agrawal and Mishra 2008 Anjum et al. 2008a,d Guo et al. 2007 Sun et al. 2007
Metwally et al. 2005

Brassica juncea leaves Phragmites australis Lactuca sativa Oryza sativa Oryia sativa roots Glycine max roots
Helianthus annuus leaves Zea mays seedlings Pisum sativum
Pinus sylvestris roots
Cucumis chloroplast Populus canescens roots
Oryza sativa leaves Glycine max Brassica campestris
Glycine max nodules
Pisum sativum
+ + + + + + + + + + —
—
—
—
—
—
—
—
—
—
X X
+ +
Qadir eM/. 2004 Pietrini et al. 2003 Maier era/. 2003 Hassan et al. 2008 Guo et al. 2007 Balestrasse et al. 2001 Gallego et al. 1996b Rauser 1990 Ruegsegger et al. 1990 Schutzendubel et al. 2001 Zhang et al. 2003 Schutzendubel et al. 2002 Hsu and Kao 2004 Noriega et al. 2007 Anjum et al. 2008a,d Balestrasse et al. 2001 Gupta et al. 2002

Glycine max (Balestrasse et al. 2001). Cadmium-induced depletion of GSH has been
mainly attributed to phytochelatin synthesis (Grill et al. 1985). Phytochelatin-heavy
metal complexes have been reported to be accumulated in the vacuole of Nicotiana
tabacum leaves (Vogeli-Lange and Wagner 1990) and in Avena sativa. These
complexes have been shown to be transported across the tonoplast (Salt and Rauser
1995). The decline in the levels of GSH might also be attributed to an increased
utilization for ascorbate synthesis or for a direct interaction with Cd (Pietrini et al.
2003). The variety of response to Cd-induced oxidative stress is probably related not
only to the levels of Cd supplied, but also to the plant species, the age of the plant and
duration of the treatment.
2.11 Coordination of Sulfur and Nitrogen in Plant Growth and Productivity
The improvement in crop production and quality depend upon a number of plant- soil-
and nutrients-associated factors i.e. balanced fertilization, optimization of nutrient
replenishment, minunization of nutrient losses to the enviromnent and the concept of
co-ordination in action among plant nutrients. Among the plant nutrients, S and N are
of great importance and are required by plants for maintaining normal growth and
development (Scherer 2008, Bimbraw 2008). Several studies have established
regulatory interactions between assimilatory sulfate and nitrate reduction in plants
(Rennenberg et al. 1979, Reuveny et al. 1980, Brunold and Suter 1984, Haller et al.
1986, Takahashi and Saito 1996). The two pathways are very well coordinated
(Brunold 1993, Hawkesford et al. 2006). A deprivation of one leads to a reduction of
the metabolism of the other (Reuveny et al. 1980, Prosser et al. 2001, Hesse et al.
2004, Scherer 2008). It has also been established through genetic studies that sulfate
reduction is regulated by N nutrition at the transcriptional level (Koprivova et al.
2000).
The S and N affect enzyme activity in their respective assimilatory pathways
(Reuveny et al. 1980, Brunold and Suter 1984, Barney and Bush 1985, Bell et al.
1995, Ahmad et al. 1999). While N directly affects the photosynthesis efficiency of
the plant, S affects the photosynthesis efficiency indirectly by improving the N
utilization efficiency of the plants as evident from the relationship between N content
and photosynthetic rate in the leaves of S-treated and S-untreated plants (Ahmad and
Abdin 2000a). It has also been shown in Hordeum vulgare that high levels of nitrate
and ammonium can induce a high affinity sulfate transporter gene and hence sulfate
38

uptake in N-fed plants suggesting that a N metabolite may affect sulfate transporter
gene expression (Vidmar et al. 1999).
Positive interaction between S and N results in increased crop productivity
(Podlesna and Cacak-Pietrzak 2008). Brassica genotypes require higher amounts of S
in addition to N for optimum growth and yield (Aulakh et al. 1980, Lakkineni and
Abrol 1992, 1994, Abdin et al. 2003, Aulakh 2003). Application of S along with N
lead to enhanced biomass production and increased leaf area as both nutrients are
involved in biosynthesis of proteins and several other molecules. Nitrogen is a basic
constituent of proteins and with the increase in the rate of N application, the N
availability increases. Similarly, increased S supply increases seed yield with higher
protein content. Combined application of S and N promotes the uptake of S and N,
which leads to significant enhancement in seed protein and oil content in Brassica
juncea and Brassica campestris (McGrath and Zhao 1996, Ahmad and Abdin 2000b,
Abdin et al. 2003). Sulfur and nitrogen relationship in terms of crop yield and quality
has been established in many studies (Singh and Bairathi 1980, Sachdev and Deb
1990, Lakkineni and Abrol 1992, McGrath and Zhao 1996, Ahmad et al. 1998,
Fismes et al. 2000).
2.12 Coordination of Sulfur and Nitrogen under Environmental Stress
Plants have developed strategies resulting in optimal adaptation to environmental
resources needed for growth, respiration, and propagation (Rennenberg and Brunold
1994, Brunold et al. 1996). Among these adaptations, strategic co-ordination of
macronutrient assimilation, i.e., carbon, nitrogen, phosphorous, water, and sulfur
['CHONSP'] is especially important, as these pathways are dependent on the varying
availability of mineral nutrients and CO2 and changes in light energy (Hesse et al.
2004). The assimilatory pathways interact in the formation of products and are
regulated in a coordinated maimer to balance macro- and micronutrients in possible
synergistic or antagonistic effects (Brunold 1993, Leustek et al. 2000, Saito 2000).
As heavy metals can induce essential nutrient deficiency and even decrease
the concentration of several macronutrients in plants (Siedlecka 1995), it may be
possible to reverse or reduce (at least partly) some of the metal induced negative
effects on the plants by optimization of mineral nutrition. Chen and Huerta (1997)
showed that S is a critical nutritional factor for reduction of Cd toxicity. These authors
observed that negative effects of Cd on growth and photosynthesis are stronger in
Hordeum vulgare plants supplied with 0.1 mM S than in plants receiving ImM S. A
39

positive effect of S nutrition on Cd detoxification in Beta vulgaris plants has also
been established (Popovic et al. 1996). It has been found that at sub-optimal S
nutrition Cd exposed plants preferably allocate S to PCs synthesis, which may
provoke S deficiency (McMahon and Anderson 1998). Probably, the improved S
nutrition allows a more adequate plant defense response to Cd and also prevents S
deficiency. Good S nutrition diminishes the toxicity of Cd by restoring a new steady
state of the GSH level earlier than in plants grown at low S supply (McMahon and
Anderson 1998). Glutathione represents the major pool of non-protein reduced S in
plants (Kunert and Foyer 1993) and it accounts only for 2% of the total organic S
content in plants ranging from 120 to 380mmol S kg"' dry matter (McMahon and
Anderson 1998, Hawkesford and De Kok 2006, Sun et al. 2007, Ernst 2008).
Glutathione has critical functions aside from its role as a redox buffer. It serves as the
first line of defense against the products of oxygen metabolism, ROS, and heavy
metals (May et al. 1998a,b). Recently, El-Shintinawy (1999) showed that exogenous
GSH counteracted all retardation effects in Glycine max seedlings induced by Cd. He
provided data to conclude that the dual fimction of GSH; first, as being an antioxidant
and oxy-radicals scavenger, it protects chloroplast from oxidative damage by trapping
the hydroxyl radicals, secondly as a substrate for PCs synthesis that mainly sequesters
and detoxifies excess Cd ions. When both functions are realized, repairing membrane
damage and sequestration of Cd are achieved by GSH. The electron transport rate as
well as photosynthetic parameters is not disturbed and this results in a photosynthetic
performance similar to that in the control plants. The crucial role of plant PCs in Cd
detoxification was strongly supported by the investigations on two mutants from
Arabidopsis, cadi and cad2, which were deficient in PC and GSH biosynthesis,
respectively, and are consequently more sensitive to Cd (Howden et al. 1995a,b,
Cobbett et al. 1998, Cobbett and Goldsbrough 2002). Inouhe et al. (2000) found a
close relationship between high Cd sensitivity and lack of phytochelatin synthase in
the cultured cells of Vigna angularis. In a recent study, Ranieri et al. (2005) reported
that the leaves of Triticum aestivum exposed to Cd stress were coimteracted by PC
biosynthesis. Furthermore, Anjum et al. (2008a) reported that S supplementation
increased the production of GSH content under low level of Cd which protects the
Brassica campestris plants by improving the growth and photosynthesis.
Pankovic et al. (2000) have shown that optimal N supply decreased the
inhibitory effects of Cd on photosynthesis of Helianthus annuus plants. The authors
40

studied the effects of Cd on Helianthus annuus plants grown at optimal, sub-optimal
and supra-optimal N supplies. They found the lowest inhibition of photosynthetic
activity by Cd at optimal N supply, when an investment in soluble proteins and
Rubisco were at their maximum. Higher N supplies did not alleviate the toxic Cd
effects; therefore the authors concluded that N nutrition can be manipulated as a
means of decreasing Cd phytotoxicity. A proper N supply has positive effects in
overcoming the adverse effects caused by Cd toxicity in Oryza sativa (Hassan et al.
2005a). An exhaustive research by Hassan et al. (2005a) has revealed that different
forms of N show differential response in the alleviation of Cd toxicity in Oryza sativa.
They concluded that among the N forms that unproved growth and photosynthetic
traits to a large extent was Ca(N03)2 followed by NH4NO3 and (NH4)2S04. Beside, a
substantial difference was noted among N forms in their effect on Cd and N uptake,
(NH4)2S04-fed plants had less Cd and more N uptake than both Ca(N03)2- and
NHtNOs-fed plants, suggesting potential antagonist effect between Cd and
ammonium-N, and synergist effect between Cd and nitrate-N.
Phytochelatin biosynthesis is closely dependent on S metabolism (Cobbett
2000a,b, Leustek et al. 2000), synthesized from different isoforms of GSH (Grill et al.
1985). It is important to note here that GSH with the help of Cys takes part in the
biosynthesis of PCs. In plants PCs play a crucial role in Cd detoxification (Howden et
al. 1995b). It complexes cations (Cd " ), and makes them less harmful to plant cells
(Weigel and Jager 1980). Expression of genes involved in reductive sulphate
assimilation pathway and enzyme activities are stimulated by Cd (Herbette et al.
2006, Ernst et al. 2008). Cadmium exposure induces the activity of enzymes (y-
glutamyl-Cys synthatase, yECS and glutathione synthatase, GS) involved in the
biosynthesis of GSH. Herbette et al. (2006) suggested that plants activate the S
assimilation pathway by increasing transcription of related genes to provide an
enhanced supply of GSH for PC biosynthesis to cope with Cd toxicity. In addition,
when plants encounter reactive oxygen species, GSH is a direct source of electrons for
stress mitigation by the enzyme glutathione peroxidase (GPX) or an indirect means to
maintain a reduced pool of ascorbate, another antioxidant. In fact, the biosynthesis of
y-glutamylcysteinylglycine (GSH) depends on the linking and the availability of its
constituent amino acids Cys, Glu and Gly. The biosynthesis of GSH in plants is well
established: two sequential ATP-dependent reactions allow the synthesis of y-
glutamylcysteine (y-EC) from L-Glu and L-Cys, followed by the formation of Gly to
41

the C-terminal end of y-EC (Meister 1988). These reactions are catalyzed by y-ECS
and GS (May et al. 1998a, Noctor et al. 1998b). Glutathione contains three moles of
N per mole of S and its GSH biosynthesis may depend on the availability of N
precursors and thus N nutrition of plants. However, the sink strength of GSH
biosynthesis for N may be low compared with the other major N sinks such as the
synthesis of proteins or nucleotides (Kopriva and Rennenberg 2004). The GSH
biosynthesis is, therefore, regulated not only by the dependency of Cys availability on
S, N and C metabolism, but also on the availability of Glu and Gly. In this way, the
coordinative functions of S and N may strengthen the stress tolerance ability of plants
growing under abiotic stresses.
2.13 Research Work Reported from Our Laboratory
We have conducted research to study the effects of Cd on growth, photosynthesis and
response of the components of antioxidant defense system in different plants such as
Triticum aestivum, Vigna mungo, Brassica juncea, Brassica campestris and Lepidium
sativum. Following is the summary of the published work.
Wheat (Triticum aestivum L.)
Khan et aL (2006) World J Agric Sci 2: 223-226
The variation in growth, photosynthesis and yield characteristics of five Triticum
aestivum cultivars, namely PBW343, HT2329, PBW373, UP2338 and WH542 at
three plant growth stages, treated with 0, 25, 50 and lOOmg Cd kg'' soil was studied.
Maximal significant reductions in the growth characteristics were observed with
lOOmg Cd kg"' soil in all the cultivars at all the sampling times. Among cultivars,
PBW343 proved tolerant and showed lesser decrease in the characteristics, whereas,
WH542 emerged as non-tolerant and suffered maximum decrease.
Khan et aL (2007a) J Agron Crop Sci 193:435-444
The activities of antioxidative enzymes and S assimilation were studied in Cd-treated
Triticum aestivum cultivars to assess their involvement in determining yield potential.
The cultivar WH542 (low yielding type) accumulated Cd to a greater amount in both
root and leaf, and also exhibited higher contents of H2O2 and TEARS and the activity
of SOD than cultivar PBW343 (high yielding type). The activities of other
antioxidative enzymes, CAT, APX, GR and GPOX, activity of ATP-sulfurylase, S
content, photosynthetic, growth and yield characteristics were higher in PBW343 than
in WH542 in Cd treatment compared to the control. The results suggest that the
efficient fimctioning of enzymes of the antioxidative system and S assimilation helped
42

in alleviating the effects of Cd in PBW343, protected photosynthetic ability and
maintained high yielding potential of the cultivar.
Khan et aL (2008b) J Plant Interactions 3:31-37.
Leaf carbonic anhydrase (CA) activity, net photosynthetic rate (PN), stomatal
conductance (gs) and plant dry mass (DM) exhibited a dose-dependent response to Cd
in Triticum aestivum grown under low and sufficient Zn levels at 30 d after seedling
emergence in sand culture. Cadmium at lOmM concentration enhanced CA, PN and
dry mass at low Zn level but these remain unaltered at sufficient Zn. Stomatal
conductance decreased with increasing CdCb concentration at both the Zn levels.
Cadmium higher than lOmM inhibited the characteristics irrespective of Zn level. The
activity of SOD was not saturated with the highest Cd concentration (50mM) at both
the Zn levels. However, the activities of APX and GR increased up to 25mM Cd and
decreased at 50mM Cd at both the Zn levels. It is suggested that CA activity was
induced at low Cd (lOmM) under low Zn level mcreasing the photosynthesis. The
synergies among the activities of antioxidative enzymes helped to maintain CA and
thus photosynthesis and plant dry mass at low Cd concentration under low Zn level.
Blackgram (Vigna mungo L.)
Singh et aL (2008a) Amer J Plant Physiol 3:25-32
Vigna mungo plants were treated with 0, 25, 50 and lOOmg Cd kg"' soil as CdCb for
30d. The changes in total chlorophyll (Chi), Chi a^, net photosynthetic rate (PN),
stomatal conductance (gs), water use efficiency (WUE) and carbonic anhydrase (CA)
activity were noted. The activities of antioxidative enzymes in root and leaf were also
assayed together with the content of TBARS and H2O2. The concentration of Cd in
root and leaf increased with the increasing Cd concentration. Greatest decrease in
photosynthetic traits was observed with lOOmg Cd kg"' soil. The activity of SOD
increased in leaf but decreased in root, whereas, the activity of CAT decreased in both
root and leaf By contrast to CAT, the activity of APX increased in root and leaf
However, GR activity increased in root and decreased in leaf The results suggest that
the antioxidative enzymes showed differential pattern in root and leaf and the
decrease in photosynthesis with lOOmg Cd kg"' soil was associated with the
accumulation of TBARS and H2O2 content and reduction in Chi content, stomatal
conductance and CA activity.
43

Mustard {Brassica juncea L.)
Mobin and Khan (2007) J Plant Physiol 164:601-610
Photosynthetic performance, contents of chlorophyll and associated pigments, cellular
damage and activities of antioxidative enzymes were investigated in two Brassica
juncea cultivars differing in photosynthetic capacity subjected to Cd stress. Exposure
to Cd severely restricted the PN of RH30 (low photosynthetic potential) compared to
Varuna (high photosynthetic potential). This corresponded to the reductions in the
activities of CA and ribulose-l,5-bisphosphate carboxylase (Rubisco) in both the
cultivars. Decline in Chi (a+b) and Chi a content was observed but decrease in Chi b
was more conspicuous in Varuna under Cd treatments, which was responsible for
higher Chi a:b ratio. Additionally, the relative amount of anthocyanin remained higher
in Varuna compared to RH-30 even in the presence of high Cd concentration, while
percent pheophytin content increased in RH-30 at low Cd concentration. A higher
concentration of Cd (lOOmg Cd kg'' soil) resulted in elevated H2O2 content in both
the cultivars. However, Varuna exhibited lower content of H2O2 in comparison to
RH30. This was reflected in the increased cellular damage in RH30, expressed by
greater TBARS content and electrolyte leakage. The enhanced activities of
antioxidative enzymes, APX, CAT and GR and also lower activity of SOD in Varuna
alleviated Cd stress and protected the photosynthetic activity.
Anjum et aL (2007) XXXth Indian Botanical Conference pp. 148
The differential response of two Brassica juncea cultivars, Varuna and RH30
differing in photosynthetic potential to 0, 50, 100, 150 and 200mg Cd kg"' soil were
investigated. Varuna (high photosynthetic potential) showed higher rate of
photosynthesis, greater production of Cys, non-protein thiol and PC and the activity of
ATP-sulfurylase than the cultivar RH30 (low photosynthetic potential). In both the
cultivars the response was greatest at lOOmg Cd kg"' soil and decreased at higher Cd
concentration. The activities of antioxidative enzymes, SOD, GR, CAT and APX
showed a dose-dependent response to Cd in both the cultivars. But the cultivar Varuna
showed greater increase in the activities of enzymes such as APX, CAT and GR with
respect to control than the cultivar RH30 at lOOmg Cd kg'' soil. However, the
repression in the activities of enzymes at 150 and 200mg Cd kg"' soil was greater in
RH30 than Varuna compared to control. The cultivar RH30 exhibited a higher degree
of oxidative stress than Varuna and therefore, the activity of SOD, the contents of
TBARS, H2O2 and electrolyte leakage were found greater in RH30 cultivar than
44

Varuna with increasing Cd concentrations. The results show that high photosynthetic
mustard cultivar Varuna protected photosynthesis through enhanced production of
cysteine, non-protein thiol and phytochelatins and simultaneously by reducing
oxidative stress through efficient antioxidative enzyme system.
Rapeseed {Brassica campestris L.)
Anjum et aL (2008a) Plant Growth Regul 54:271-279.
The ameliorative effect of S (0 or 40mg S kg"' soil) on Cd (0, 25, 50 and lOOmg Cd
kg' soil)-induced growth inhibition and oxidative stress in Brassica campestris
cultivar Pusa Gold was studied. Cadmium at lOOmg kg'' soil caused maximum
increase in the contents of Cd and TBARS in leaves. Maximum reductions in growth
(plant dry mass, leaf area), Chi content, PN and the contents of ascorbate and
glutathione were observed with lOOmg Cd kg"' soil compared to control. The
application of S helped in reducing Cd toxicity, which was greater for 25 and 50mg
Cd kg"' soil compared tolOOmg Cd kg"' soil. Addition of S to Cd-treated plants
showed decrease in Cd and TBARS content in leaves and restoration of growth and
photosynthesis through increase in the contents of ascorbate and glutathione. Net
photosynthetic rate and plant dry mass were strongly and positively correlated with
the contents of ascorbate and glutathione. It is suggested that S may ameliorate Cd
toxicity and protects growth and photosynthesis of mustard involving ascorbate and
glutathione.
Anjum etaL (2008b) J Plant Interactions DOI 10.1080/17429140701823164
Brassica campestris cultivar Pusa Gold plants, exposed to different Cd levels (0, 25,
50 and lOOmg kg"' soil), were analyzed for the distribution of metal, biomass and the
degree of growth stage to Cd-sensitivity. A significant maximum decrease in plant
biomass was observed at flowering stage followed by pre-flowering and post-
flowering stages. Activities of enzymatic antioxidants such as SOD, CAT, APX and
GR differentially increased; while, the concentrations of non-enzymatic antioxidants
such as ascorbate and glutathione drastically decreased in plants exposed to Cd at
various growth stages. However, the concentrations of glutathione and ascorbate
decreased maximally in plant groups exposed to Cd at flowering stage. Maximum Cd-
accumulation occurred in roots followed by leaves and stem. Various Cd levels
inhibited also the contents of plant nutrients such as N, P, K and S in leaves. The
study concludes the existence of close relationships among growth parameters, Cd-
45

sensitivity of phenological stages of the crop and the components of antioxidant
system in rapeseed plants.
Garden cress (Lepidium sativum L.)
Singh etaL (2007) XXXth Indian Botanical Conference pp. 161
Lepidium sativum is a medicinally and economically important plant which contains
significant amounts of iron, calcium and folic acid, in addition to vitamins A and C.
An experiment was conducted to examine the effect of different concentrations of
cadmium (0, 25, 50 and lOOmg kg'' soil) on the performance of garden cress plants.
Cadmium accumulation in roots and leaves increased with the increasing Cd
concentration in soil, and roots accumulated more Cd than leaves. Cadmium
treatments inhibited leaf area and plant dry mass. Dose dependent increase in TEARS,
H2O2 content and the activity of antioxidant enzymes (viz. SOD, CAT, APX and GR)
was observed under Cd stress. A progressive decline in relative water content,
photosynthetic rate, stomatal conductance, intercellular CO2, Chi content (Chi a, Chi
b, total Chi), CA activity, Rubisco, NR activity and N content was also observed with
the increasing Cd concentration. However, ATP-sulfiirylase activity, S content and
activities of antioxidant enzymes were increased. The decrease in NR activity and N
content showed that Cd affects nitrogen metabolism; whereas, the increase in ATP-
sulfurylase activity and S content suggests the up-regulation of S assimilation
pathway for possible Cd tolerance.
2.14 Critical Appraisal of Review of Literature
The literature reviewed above includes studies on the physiological analysis of
various growth-, photosynthetic- and biochemical-traits, antioxidant studies and yield
of various crop plants xmder Cd stress. It appears that there are few reports concerning
the effect of fertilization (especially S and N) under abiotic stress on various
physiological processes, including productivity of mustard, an important oilseed crop.
In fact, the work on various crop plants is invariably aimed at establishing
mechanisms of antioxidant machinery of plants exposed to various abiotic stresses.
However, in-depth understanding of the response of various mineral nutrients in
enhancing and/or strengthening the antioxidant machinery to coimteract the effects of
abiotic stress-induced ROS production has not been done.
Only few reports are available on the individual effects of S and N in the
amelioration of Cd toxicity, and that no work has been taken to study the interactive
effects of S and N in crop plants grown under Cd stress. The reported study was
46

undertaken on Brassica juncea (S requiring crop) to explore the possibility of
ameliorating the adverse effects of Cd through the balanced application of S and N.
Following the application of S and N to Cd-treated Brassica juncea plants, concurrent
changes in growth, physiological and biochemical traits, components of antioxidant
defense system and the productivity oi Brassica juncea were studied.
47

MATERIAL AND METHODS

Contents
MATERIAL AND METHODS
3.1 3.1.1 3.1.1.1 3.1.1.2 3.1.1.3 3.2 3.3 3.3.1 3.3.2 3.3.3 3.4 3.4.1 3.4.2 3.4.3 3.5 3.6 3.7 3.8 3.8.1 3.8.2 3.8.2.1 3.8.2.2 3.8.2.2.1 3.8.2.2.2 3.8.2.2.3 3.8.2.3 3.8.2.3.1 3.8.2.4 3.8.2.4.1 3.8.2.5 3.8.2.5.1 3.8.2.5.2 3.8.2.6 3.8.2.6.1 3.8.2.6.2 3.8.2.7 3.8.2.7.1 3.8.2.7.2 3.8.2.8 3.8.2.9 3.8.2.10 3.8.2.11 3.8.2.11.1 3.8.2.11.2 3.8.2.11.2.1 3.8.2.11.2.1.:
Plant Material Botanical description of mustard Nomenclature Botanical description Genomic relationships of Brassica juncea Agro-Climatic Conditions of Aligarh Experimentation Preparation and analysis of soil Pot preparation and treatments Sowing of seeds Experimental Set-up Experiment 1 Experiment 2 Experiment 3 Sampling Technique Chemicals Parameters Studied Methodology Growth characteristics Physiological and biochemical characteristics Net photosynthetic rate and stomatal conductance Chlorophyll and carotenoid contents Extraction Estimation Calculation for chlorophyll and carotenoids content Assay of carbonic anhydrase activity Estimation Assay of nitrate reductase activity Estimation Assay of ATP-sulfurylase activity Extraction Estimation Determination of nitrogen Digestion of leaf powder Estimation of nitrogen Determination of sulfur Digestion of leaf powder Estimation of sulfur Determination of cadmium Determination of thiobarbituric acid reactive substances Determination of hydrogen peroxide content Components of en2ymatic and non-enzymatic antioxidant system Activities of antioxidant enzymes En2yme extraction Superoxide dismutase activity
1 Enzyme assay
48 48 48 48 49 50 50 50 51 51 52 52 52 53 53 54 54 55 55 55 55 55 55 55 55 56 56 56 57 57 57 57 58 58 58 59 59 59 59 60 60 60 60 60 60 61

3.8.2.11.2.2 Catalase activity 61 3.8.2.11.2.2.1 Enzyme assay 61 3.8.2.11.2.3 Ascorbate peroxidase activity 61 3.8.2.11.2.3.1 Enzyme assay 61 3.8.2.11.2.4 Glutathione reductase activity 62 3.8.2.11.2.4.1 Enzyme assay 62 3.8.2.11.2.5 Soluble protein content 62 3.8.2.11.2 Non-enzymatic antioxidants 62 3.8.2.11.2.1 Ascorbic acid content 62 3.8.2.11.2.1.1 Procedure 63 3.8.2.11.2.2 Glutathione content 63 3.8.2.11.2.2.1 Procedure 63 3.8.3 Yield characteristics 63 3.8.4 Statistical analysis 64

MATERIAL AND METHODS Chapter 3
The chapter deals with the description of material used for the study and methods
adopted for the experimentation and determination of various traits during the course
of the investigation. The details of agro-climatic conditions, soil analyses,
experimentation and the techniques and procedures employed in this regard are given
below.
3.1 Plant Material
The Experimental material selected for the present study was mustard {Brassica
juncea L. Czem & Coss.) cultivars. Healthy and authentic seeds of mustard cultivars
namely, Alankar, Varuna, Pusa Bold, Sakha and RH30, were obtained from National
Research Centre on Plant Biotechnology (NRCPB) of the Indian Agricultural
Research Institute (lARI), New Delhi.
3.1.1 Botanical description of mustard
3.1.1.1 Nomenclature
The oleiferous Brassica grown in India are divided into four groups:
1. Brown mustard: Commonly known as rai, raya or laha {Brassica juncea L.
Czem & Coss.)
2. Sarson
a. Yellow sarson: Brassica campestris L. var. Sarson Prain
b. Brown sarson: Brassica campestris L. var. Dichotoma Watt
3. Toria: lahi or maghi lahi Brassica campestris L. var. Toria Duth
4. Taramira or Tara {Eruca sativa Mill.)
In addition, there are two other species, namely Brassica nigra Koch. (Banarasi
rai) and Brassica juncea var. Rugosa (Pahadi rai). These two species do not fall under
any of the four groups. These are, moreover, grown to a limited extent. Mustard
{Brassica juncea L. Czem & Coss.) is the dominant species grown in India (Prakash
1980).
3.1.1.2 Botanical description
Rape and mustard include annual herbs. Roots, in general, are long and tapering.
Toria is more or less a surface feeder but Brown sarson bears long roots with limited
lateral spread enabling its successful cultivation under drier conditions. The height of
the stem varies from 45cm (in some varieties of Toria) to 190cm (in Yellow sarson).
In Toria and Brown sarson, the branches arise at an angle of 30° to 40°. In Yellow
48

sarson, the branches arise laterally at an angle of about 10° to 20° and give the plant a
narrow and pyramidal shape. The inflorescence is a corymbose raceme. In the case of
Yellow sarson, the four petals are spread apart, whereas, in Brown sarson and Toria,
the petals overlap or may be placed apart, depending upon the cultivar. The flowers
bear a hypogynous ovary. In Brown sarson and Toria, the ovary is bicarpellary,
whereas, in Yellow sarson, it may also be tri-or tetra-carpellary. The fruit is sliliqua.
The sliliqua are two-valved, three-valved or four-valved, depending upon the number
of carpels in the ovary. The flowers begin to open from 8:00h and continue up to
12:00hnoon.
3.1.1.3 Genomic relationships of Brassica juncea
There are six species of Br assica that merit attention for their economic importance.
Among the six species, three are diploid: B. campestris, B. nigra and B. oleraceae,
and the other three are amphidiploids: B. juncea, B. napus and B. carinata. The
botanical and genomic relationships between these six species may be represented in
the form of atriangle usually knovm as triangle of U (U 1935) (Figure 1).
Fig 1. The "Triangle of U" representing the genomic relationships among Brassica
species.
Brassica juncea is generally thought to have originated in the Middle East,
whereas, B. rapa and B. nigra species overlapped in the wild, but central Asia and
China are suggested as the sites of primary origin (Prakash 1980). Hemingway
49

(1976), however, considers that B. juncea L. may also have arisen by independent
hybridization at secondary centers in India, China and Caucasus, as B. nigra was
widely used as the commercial spice from early times.
3.2 Agro-Climatic Conditions of Aiigarh
Aligarh has an area of about 5,024sq kms and is situated at 27°52'N latitude, 78°5rE
longitude, and 187.45m altitude above sea level. It has semi-arid and subtropical
climate, with hot dry summers and cold winters. The winter extends from the middle
of October to the end of March. The mean temperatures for December and January,
the coldest months, are about 15°C and 13°C and minimum recorded for any single
day is 2°C and 0.5°C, respectively. The simimer season extends from April to June
and the average temperature for May is 34.5°C and 45.5°C, respectively. The
monsoon extends from the end of June to middle of October. The mean annual
rainfall is about 847.3mm.
More than 85% of the total down pour is delivered during a short span of four
months from Jime to September. The remaining showers are received during winter.
Winter rainfall is especially usefiil for 'Rabi' (winter) crops. The district of Aligarh
has the same soil composition and appearances as those found generally in the plains
of Uttar Pradesh. Different types of soils, such as sandy, loamy, sandy-loam and clay-
loam are found in the district.
3.3 Experimentation
Experiments were conducted in the earthen pots during the winter season of 2003,
2004 and 2005 on mustard (Brassica juncea L. Czem & Coss.) in the green house of
the Department of Botany, Aligarh Muslim University, Aligarh, India under natural
day/night conditions. The Experimental period varied from October to March.
3.3.1 Preparation and analysis of soil
To ensure maximum soil-aeration, thoroughly ploughed soil was collected from
University Agriculture Farm, Aligarh Muslim University, Aligarh for the filling of
earthen pots. Before sowing, soil samples were collected randomly from different pots
for the analysis of the selected soil-characteristics. The physicochemical properties of
soil are given in Table 3.
50

Sandy loam 7.80 90.54 8.89 112.37 15.52 0.310
Sandy loam 8.10 100.16 8.75 110.55 16.00 0.300
Sandy loam 8.00 98.72 8.58 108.65 16.31 0.323
Table 3. Some physicochemical properties of the soil used in the present study.
Experiment 1 Experiment 2 Experiment 3 Soil characteristics (2003-2004) (2004-2005) (2005-2006) Texture pH Available N (mg kg"' soil) Available P (mg kg"' soil) Available K (mg kg"' soil) Available S (mg kg"' soil) Total Cd (mg kg"'soil)
3.3.2 Pot preparation and treatments
Before the start of each Experiment, earthen pots of 23-cm diameter were filled with
soil thoroughly mixed with farmyard manure in 3:1 ratio making a total of 4.0kg soil
pot"'. Appropriate amount i.e., 0, 163.20, 326.16, 652.28, 978.44mg CdCb was
mixed thoroughly with 4.0kg soil to achieve 0, 25, 50, 100 and 150mg Cd
kg"'soil, respectively. In Experiments where application of sulfur was done at the
rate of 0, 50 and lOOmg S kg"'soiI, elemental S was applied to the soil 15 days
before sowing for complete mobilization by microbial activity. For nitrogen
application, it was given at the rate of 0, 40, 80 and 120mg N kg"'soil before
sowing as urea. In Experiment 1 different mustard cultivars were treated with Cd
alone, whereas, in Experiment 2 tolerant and non-tolerant mustard cultivars were
treated with Cd alone or Cd in combination with S. Further, in Experiment 3,
combined doses of S and N were given to Cd-treated tolerant and non-tolerant
mustard cultivars. Plants grown without any treatment (no Cd, S or N) served as
control.
3.3.3 Sowing of seeds
Healthy seeds of mustard cultivars were surface sterilized with 0.5% (v/v) sodium
hypochlorite for 15min and then rinsed three times with deionized water. Before
sowing light application of deionized water was given in each pot to provide
necessary moisture for germination of the seeds. Ten seeds per pot were sown to
avoid germination failure and after the establishment of seedlings, thiiming was done
to retain only two healthy plants of nearly equal size in each pot. In Experiment 1,
twelve pots for each treatment were maintained. Four sets of pots with three pots
(each pot with two plants) in each set were maintained. Each set of pots served as
sampling material at pre-flowering (30days after sowing, DAS), flowering (60DAS),
51

post-flowering (90DAS) stages and harvest (120DAS). Thus, total number of pots
was sixty for each cultivar in Experiment 1. One plant in a pot of each set was
considered as one replicate. Thus, there were three replicates. Diagrammatic
representation of the arrangement of pots for Experiment 1 is shown in Scheme 1. In
Experiment 2, similar distribution of pots was made, but due to seven treatments, the
total number of pots was eighty four for each cultivar. The distribution of pots for
Experiment 2 for determining various characteristics at 30, 60, 90 and 120DAS is
described in Scheme 2. Distribution of pots for Experiment 3 was same as in
Experiments 2 but due to six treatments, total number of pots was sixty for each
cultivar. The distribution of pots for Experiment 3 for determining various
characteristics at 30, 60, 90 and 120DAS is described in Scheme 3.
Crop was irrigated with tap water as and when required. In order to check the
aphid contagion, if any, insecticidal spray of Dimecron-100 was done.
3.4 Experimental Set-up
The Experiments were carried out according to the standard agricultural practices.
Calendar of various operations in each Experiment is given in Table 4, which shows
time schedule for experimentation, treatment and the data collection.
3.4.1 Experiment 1
Experiment 1 was conducted in the winter season of 2003-2004. Surface sterilized
seeds of five mustard cultivars namely, Alankar, Varuna, Pusa Bold, Sakha and RH30
were sovm on 15* October 2003 and the crop was harvested on 13* February 2004.
The treatments in this Experiment were arranged in a factorial randomized block
design. The aim of the Experiment 1 was to study the effects of five levels of Cd (0,
25, 50, 100 and 150mg Cd kg'' soil) on growth, photosynthetic and yield
characteristics and select tolerant and non-tolerant cultivars. Samplings for growth
and photosynthetic characteristics were done at pre-flowering (30DAS), flowering
(60DAS), post-flowering stage (90DAS) and yield characteristics were recorded at
harvest (120DAS). The scheme of treatments for Experiment 1 is given in Table 5.
3.4.2 Experiment 2
Experiment 2 was conducted on the basis of the findings of Experiment 1 in the
winter season of 2004-2005. The sovdng of the seeds of Alankar (tolerant) and RH30
(non-tolerant) cultivars of mustard was done on 20* October 2004 and the crop was
harvested on 18* February 2005. The treatments in this Experiment were arranged in
a factorial randomized block design. The aim of the Experiment 2 was to assess the
52

effect of S application in the alleviation of Cd stress in Cd-tolerant and non-tolerant
mustard cultivars. Sulfur was given at the rate of 0, 50 and lOOmg S kg'' soil prior to
15 days of sowing to plants treated with 0, 50 and 150mg Cd kg'' soil. The response
of Cd tolerant and Cd non-tolerant cultivars to S application under Cd stress was
assessed in terms of the growth, photosynthetic and S and N assimilation, components
of antioxidant system and the yield characteristics. The timing of sampling was same
as for Experiment 1. The scheme of treatments for Experiment 2 is given in Table 6.
3.4.3 Experiment 3
Experiment 3 was conducted on the basis of the findings of Experiments 2 in the
winter season of 2005-2006. The sowing of mustard seeds (tolerant and non-tolerant
mustard cultivars as in Experiment 2) was done on 17* October 2005 and the crop
was harvested on 15* February 2006. The treatments in this Experiment were
arranged in a factorial randomized block design. The aim of the Experiment 3 was to
assess the effect of N application on the effectiveness of S in the alleviation of Cd
stress in Cd-tolerant and non-tolerant mustard cultivars. Sulfur was given at the rate of
0 and 50mg kg"' soil along with 0, 40, 80 and 120mg N kg"' soil to plants treated with
0 and 50mg Cd kg'' soil. The timing of sampling was same as for Experiment 1 and 2.
The scheme of treatments for Experiment 3 is given in Table 7.
3.5 Sampling Technique
To study the growth and photosynthetic characteristics, Cd accumulation in root and
leaf, S and N assimilation and components of antioxidant system, samplings were
done at 30, 60 and 90DAS corresponding to pre-flowering, flowering and post-
flowering stages. For the determination of growth one plant from each pot of a set
corresponding to the sampling time in each cultivar was taken. In a set of pots, there
were three pots for each sampling time. Therefore, three plants served as three
replicates. The remaining one plant in each pot at various sampling stages was used
for photosynthetic measurements and other analyses. There was a separate set of pots
for the determination of yield characteristics. For growth, destructive sampling was
done and plzints were taken to the laboratory. Photosynthetic measurements were
made on intact plants and the plants on which photosynthetic measurements were
made also taken to laboratory for further analyses. For recording yield, plants were
cut at the base, siliqua were plucked and thrashed manually. Seeds were cleaned and
collected separately from each treatment for the measurement of seed yield and its
attributes.
53

3 plants were 3 plants were 3 plants were Yield used for growth used for growth used for growth characteristics & 3 for & 3 for & 3 for were recorded photosynthesis at photosynthesis at photosynthesis at at 120DAS 30DAS 60DAS 90DAS
,.....»„„W,W>W nffM MffM MffM Treatments
.,-"^T^/v/ \^nU \7UU V/V/U
25Cd
50Cd
lOOCd
150Cd
WWW WWW www www UUU V/UV/ UUV UUU
www www www www UUU UUU UUU UUU
www www www www UUU UUU UUU UUU
www www www www ^ ^ ^ ^ ^ ^ ^ ^ ^ ^ ^ ^
Scheme 1. Diagrammatic representation of the arrangement of pots for Experiment 1. Scheme shows the arrangement of pots for one cultivar. The other cuhivars also had similar arrangement of pots.

3 plants were used for growth & 3 for photosynthesis, S, N & antioxidants analysis at 30DAS
3 plants were used for growth & 3 for photosynthesis, S, N & antioxidants analysis at 60DAS
3 plants were used for growth & 3 for photosynthesis, S, N & antioxidants analysis at 90DAS
Yield characteristics were recorded at 120DAS
Treatments f l f t f (mgkg-'soil)^y/ ^ 1 / ^
0
50Cd
l50Cd
50Cd +50S
50Cd
150Cd +50S
150Cd
ft ft ft Y y Y y Y y
ft ft ft
M/ Mf M,
ft ft ft
wv YY WW
ft ft ft
ft ft ft M/ M/ %
ft ft ft Y y TV Yy ft ft ft Y y Y y Y y ft ft ft Y y T y Y y ft ft ft Y y Y y Y y
ft ft ft Y y Y y Y y ft ft ft Y y Y y Y y ft ft ft Y y Y y Y y ft ft ft Y y Y y \ y ft ft ft Y y Y y \ y
ft ft ft Y y Y y T 7 ft ft ft Y y Y y Y y ft ft ft Y y Y y T 7 ft ft ft Y y Y y Y y ft ft ft Y y Y y \ y
f t f t ft ft ft ft ft ft ft ft ft ft
m/ M, M, M, M/ M/ M/ Mi w% M, M, M/
ft ft ft ft ft ft ft
V t Tt Tt Tt ft Tt Tt Tt ft ft ft ml f^^xy ^ ^ ^ ^ ^ ^ \ [ j / ^ ^
Scheme 2. Digramatic representation of the arrangement of pots for Experiment 2. Scheme shows the arrangement of pots for one cultivar. The other cultivar also had similar arrangement of pots.

3 plants were used for growth & 3 for photosynthesis, S, N & antioxidants analysis at 30DAS
3 plants were used for growth & 3 for photosynthesis, S, N & antioxidants analysis at 60DAS
3 plants were used for growth & 3 for photosynthesis, S, N & antioxidants analysis at 90DAS
Yield characteristics were recorded at 120DAS
Treatments ( m g k g - ' s o i l ) ^ ^ ^ ^ ^ ^
50Cd
50Cd +50S
fl ft ft ft ft ft ft ft ft vvYy Y y \ 7 \ y \ y \v \ y \ y
ft ft ft ft ft ft ft ft ft M, M, M, M, M, M, M, M, M,
ft ft ft Y y Y y \ y
ft ft ft ft ft ft Y y Y y Yy Yy \v \v
ft ft ft Y y Y y Y t ft ft ft Y y Y y Y /
ft ft ft Yy YV Yy
50Cd +50S +40N
50Cd +50S +80N
ft ft ft ft ft ft ft ft ft ft ft ft Y y Y y Yy \ y Yy Yy Yy \v \ t \ y \ t Yy
ft ft ft ft ft ft ft ft ft ft ft ft Y y Y y \ y \ y Y y \ y \ y \ y \ y T y Y y Y y
50Cd +50S +120N
ft ft ft ft ft ft ft ft ft ft ft ft Y y Y y Y y Y y Y y \ y Y y Y y Y t Y y Y t Y /
Scheme 3. Digramatic representation of the arrangement of pots for Experiment 3. Scheme shows the arrangement of pots for one cultivar. The other cultivar also had similar arrangement of pots.

Table 4. Experimental calendar.
Preparation of pots Sowing Treatments Cadmium
Sulfur
Nitrogen
Replications
Cultivars
Samplings
Pre-flowering (SODAS) Flowering (60DAS) Post-flowering (90DAS) Harvest (120D AS)
Experiment 1 (2003-2004)
11.10.2003 15.10.2003
19.11.2003 (0,25,50, 100 or 150mgCdkg"'soil)
Three
Alankar, Vanma, Pusa Bold, Sakha, RH30
14.11.2003 14.12.2003 13.01.2004 13.02.2004
Experiment 2 (2004-2005)
04.10.2004 20.10.2004
17.11.2004 (0, 50or lOOmg Cd kg-' soil)
05.10.2004 (0,50orl00mg S kg ' soil)
Three
Alankar (Tolerant) RH30 (Non-tolerant)
19.11.2004 19.12.2004 19.01.2005 18.02.2005
Experiment 3 (2005-2006)
02.10.2005 17.10.2005
18.11.2005 (0 or 50mg Cd kg-' soil)
03 10 2005 (0 or 50mg S kg-' soil)
15.10.2005 (0, 40, 80 or 120mgkg-' soil)
Three
Alankar (Tolerant) RH30 (Non-tolerant)
16.11.2005 16.12.2004 16.01.2005 15.02.2006
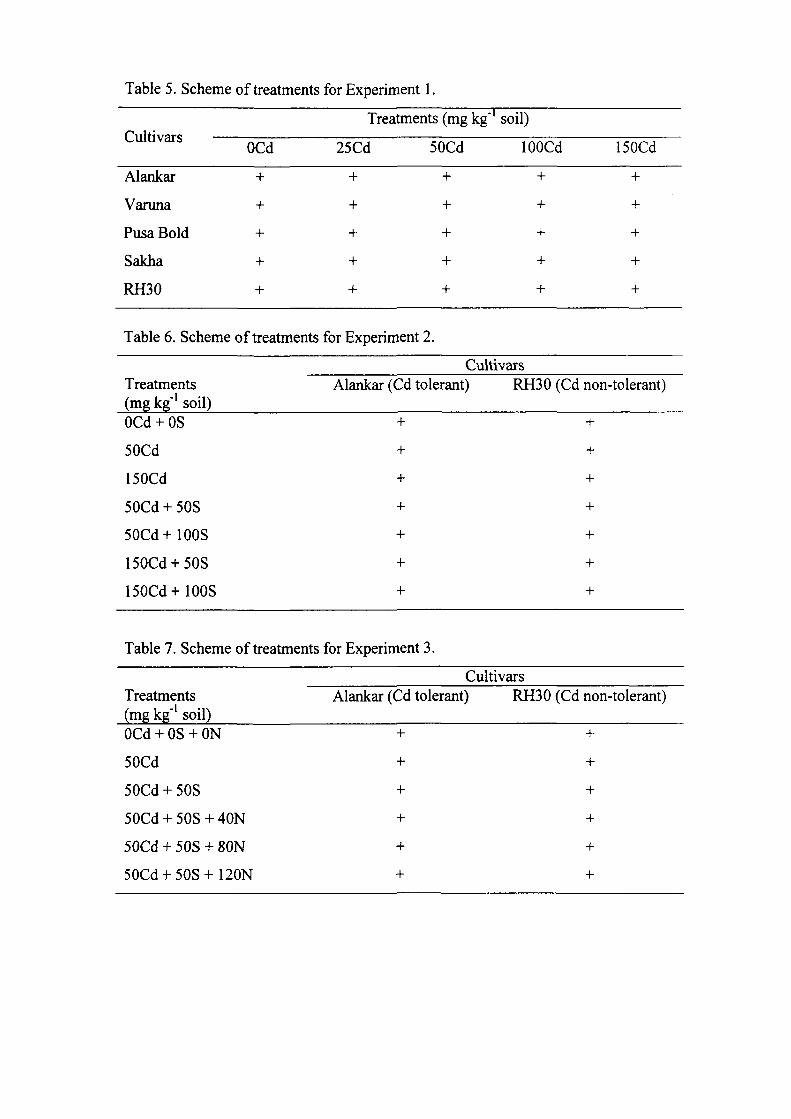
Table 5. Scheme of treatments for Experiment 1.
v_.uiiivurs
Alankar
Vanma
Pusa Bold
Sakha
RH30
3Cd
+
+
+
+
+
Treatments (mg kg''
25Cd
+
+
+
+
+
SOCd
Table 6. Scheme of treatments for Experiment 2.
Treatments (mg kg'' soil) OCd + OS
50Cd
I50Cd
50Cd + SOS
50Cd+100S
150Cd + 50S
150Cd+100S
+
+
+
+
+
soil)
lOOCd
Cultivars Alankar (Cd tolerant)
+
+
+
+
+
+
+
Table 7. Scheme of treatments for Experiment 3.
Treatments (mg kg'' soil) OCd + OS + ON
50Cd
50Cd + SOS
SOCd + SOS + 40N
SOCd + SOS + SON
SOCd + SOS+ 120N
+
+
+
+
+
RH30 (Cd
Cultivars Alankar (Cd tolerant)
+
+
+
+
+
+
RH30 (Cd
ISOCd
+
+
+
+
+
non-tolerant)
+
+
+
+
+
+
+
non-tolerant)
+
+
+
+
+
+

3.6 Chemicals
For the analysis of various enzymes activities, chemicals were obtained from Sigma-
Aldrich (St. Louis Mo, USA). Other major and minor salts, buffer components were
procured from MERK and/or SRL. All chemicals were obtained in highest purity
available commercially.
3.7 Parameters Studied
Following parameters were studied at different sampling times.
> Growth characteristics
Root length per plant
Shoot length per plant
Leaf area per plant
Plant dry mass per plant
Physiological and biochemical characteristics
Net photosynthetic rate
Stomatal conductance
Total chlorophyll content
Carotenoids content
Carbonic anhydrase activity
Nitrate reductase activity
ATP-sulfurylase activity
N, S and Cd content
Superoxide dismutase activity
Catalase activity
Ascorbate peroxidase activity
Glutathione reductase activity
Soluble protein content
Glutathione content
Ascorbic acid content
Yield characteristics
Nimiber of siliqua per plant
Number of seeds per siliqua
1000 seeds weight
Seed yield per plant
54

' MlS ' *''^'^ t.y*
3.8 Methodology
3.8.1 Growth characteristics \
Plants were uprooted and length of the main tap root and shoot length wa§,ga3^sured
on a meter scale. For dry mass accumulation, plants were dried in a hot air oven at
80°C till constant weight. Weights of the samples were recorded with the help of an
electronic balance (CY204, Scalteo Ins., Germany).
After separating all the leaves at petiole and stem junction, leaf area per plant
was recorded vnih the help of a leaf area meter (LA21, Systronics, Ahmedabad,
India).
3.8.2 Physiological and biochemical characteristics
3.8.2.1 Net photosynthetic rate and stomatal conductance
Net photosynthetic rate (PN) and stomatal conductance (gs) were measured on fully
expanded uppermost leaves of plants using close chamber infra-red gas analyzer
(IRGA, LiCOR, 6200, Lincoln, NE, USA) at light saturating intensity between 11:00-
12:00h. The capacity of leaf chamber was 1 litre. During the measurements, the air
relative humidity, photosynthetically active radiation, ambient temperature and CO2
concentration were 68±4%, 785±22|amol photons m^s"', 24±3°C and 350±5^mol
mol"', respectively.
3.8.2.2 Chlorophyll and carotenoids content
Total chlorophyll (Chi) and carotenoids (Car) were extracted using the method of
Hiscox and Israelstam (1979) by using dimethyl sulphoxide (DMSO) as an extraction
medium, and estimated and calculated by the method of Amon (1949).
3.8.2.2.1 Extraction
Fresh leaves (lOOmg) were cut into small pieces and collected in test tubes containing
7.0ml of DMSO. The test tubes were covered with black paper and incubated at 45°C
for 40min for the extraction. The content was transferred to a graduated tube and the
final volume was made to 10.0ml with DMSO.
3.8.2.2.2 Estimation
Extract measuring 3.0ml was transferred to a cuvette and the absorbance was read at
645 and 663nm for Chi content and at 480 and 510nm for Car content on UV-VIS
spectrophotometer (SL164, Elico, Hyderabad, India).
3.8.2.2.3 Calculation for chlorophyll and carotenoids content
Total Chi and Car content were calculated according to the following equations.
55

Total Chi content (mg g ' FW) = [(20.2 x OD645) + (8.02 x OD663)] x
Car content (mg g"' FW) = [(7.6 x OD480) - (1.49 x OD510)] x
V dxlOOOxW
dxlOOOxW
Where, V = volume of the extract
W = mass of the leaf tissue taken
d = distance travelled by the light path
OD = optical density at the given wave lengths viz. 645, 663,480 and 5 lOnm
3.8.2.3 Assay of carbonic anhydrase activity
Carbonic anhydrase (CA) facilitates the supply of CO2 to the carboxylation sites. It
catalyzes the reversible hydration of CO2 (Raven 1995, Khan et al. 2004).
H2O + CO2 ^ ^ ' ^ H^ + HCOs"
3.8.2.3.1 Estimation
Carbonic anhydrase activity was measured by adopting the method of Dwivedi and
Randhava (1974). Leaves selected for photosynthetic measurements were used for the
enzyme assay. Leaves were cut into small pieces (2-3mm) in 10.0ml of 0.2M cysteine
in petri dish at 0-4°C. The solution adhering to the leaf surface was removed with the
help of blotting paper followed by the immediate transfer of leaves to test tube having
4.0ml of 0.2M Na-phosphate buffer (pH 6.8). To this, 4.0ml of 0.2M sodium
bicarbonate in 0.02M sodium hydroxide solution and 0.2ml 0.002% bromothymol
blue indicator were added to the tubes. The tubes were kept at 4°C for 20min.
CO2 librated during catalytic action of the enzyme on sodium bicarbonate was
estimated by titration of the reaction mixture against 0.05N hydrochloric acid, using
methyl red as an indicator. The control reaction mixture was also titrated against
0.05N hydrochloric acid. The difference of the sample and the control readings was
noted for the calculation of the enzyme activity.
3.8.2.4 Assay of nitrate reductase activity
The nitrate reductase (NR) activity was estimated by the method of Jaworski (1971),
which is based on the reduction of nitrate to nitrite.
56

NOs' + NADH^ H^ ^ ^ > NO2' + NAD^ + H2O
3.8.2.4.1 Estimation
Fresh leaves (200mg) were cut into small pieces and transferred to plastic vial. To the
vial, 2.5ml of O.IM K-phosphate buffer (pH 7.5) and 0.5ml of 0.2M potassium nitrate
solution were added, followed by the addition of 2.5ml of 5% isopropanol. Later, two
drops of chloramphenicol solution were added to avoid bacterial growth in the
medium. Each vial was incubated for 2h in the dark at 30°C.
From the incubated mixture, 0.4ml was taken in a test tube and 0.3ml each of
1% sulphanilamide and 0.02% N-(l-Naphthyl) ethylenediamine dihydrochloride
(NED-HCL) was added. The test tube was kept for 20min at room temperature for
colour development. The mixture was diluted to 5.0ml by adding sufficient amount of
deionized water. The optical density was read at 540rmi. A blank consisting of 4.4ml
of deionized water and 0.3ml each of sulphanilamide and NED-HCl was also run side
by side.
Standard curve was plotted by taking known graded dilutions of sodium nitrite
from a standard aqueous solution of this salt. The optical density of the samples was
compared with the calibrated curve and nitrate reductase activity was calculated.
3.8.2.5 Assay of ATP-sulfurylase activity
The method of Wilson and Bandurski (1958) was followed for the assay of ATP-
sulfurylase (ATP-S) activity. The details of the procedure are given below.
3.8.2.5.1 Extraction
Extract was prepared by grinding 500mg of fresh leaves in 5.0ml of extraction buffer
[O.IM Tris-HCl (pH 8.0) containing 2mM MgCh, lOOmM KCl and lOmM DTE] in a
chilled glass homogenizer and centrifuged at 5,000x rpm for 15min at 4°C.
3.8.2.5.2 Estimation
The assay was started by adding 0.1ml of extract and 0.4ml of reaction mixture
[consists of 8.0ml deionized water (phosphate and sulphate free), 4.0ml 40mM
MgCl2.6H20, 3.0ml 0.4mM Tris buffer (pH 8.0), 40mg Na2Mo04.2H20, 45mg
Na2ATP and 20^L of inorganic pyrophosphate]. The resultant solution was incubated
at 30°C for lOmin. The reaction was stopped by adding 1.0ml of stop solution
[consists of 18.0ml 0.5M sodium acetate and 100ml 0.5M acetic acid]. Control
was also prepared by substituting NaCl in place of sodium molybdate.
57

ATP + SO4 ^TP-sulfiuylase ^ ^ ^ ^ pp.
pp. Inorganic Pyrophosphatase. .
The inorganic phosphate was estimated by the procedure of Fiske and
SubbaRow (1925). In this procedure, 1.0ml of 5N H2SO4, 0.5ml of 2.5% ammonium
molybdate and 0.1ml of reducing agent [Ig of l-amino-2-naphthol-4-sulphonic acid,
3mg of Na2S03.7H20 and 6mg of Na2S205 in a mortar and pestle and stored in dark at
4°C. 0.25g of this mixture was dissolved in 10.0ml of deionized water] were added to
the aliquot. The volume was made up to 10.0ml by adding deionized water. After a
lapse of 20min, the absorbance was measured at 660nm. The calibration curve was
dravm by using KH2PO4 solution as standard.
3.8.2.6 Determination of nitrogen
The leaves were dried in hot air oven at 80°C for 24h. Oven-dried leaves were
powdered with mortar and pestle and meshed through a 72mm mesh sieve.
3.8.2.6.1 Digestion of leaf powder
Oven-dried leaf powder (lOOmg) was carefully transferred to a digestion tube and
2.0ml of concentrated H2SO4 was added to it. It was heated on a temperature
controlled assembly for about 2h to allow complete reduction of nitrates present in the
plant material. After heating, the contents of the tube turned black. It was cooled for
about 15min at room temperature and then 0.5ml 30% H2O2 was added drop by drop.
The solution was heated again till the colour of the solution changed from black to
light yellow. Again after cooling for about BOmin, additional 3 to 4 drops of 30%
H2O2 were added, followed by reheating for another 15min. The addition of H2O2
followed by careful heating was repeated until the contents of the tube turned
colourless. The peroxide-digested-material was transferred from the tube to a 100ml
volumetric flask with three washings of deionized water. The volume of the
volumetric flask was then made up to the mark (100ml) with deionized water. This
aliquot was used to estimate nitrogen content.
3.8.2.6.2 Estimation of nitrogen
Nitrogen was estimated according to the method of Lindner (1944). A lO.OmI aliquot
of the digested material was taken in a 50.0ml volumetric flask. To this, 2.0ml of 2.5N
sodium hydroxide and 1.0ml of 10% sodium silicate solutions were added to
neutralize the excess of acid and to prevent turbidity, respectively. The volume was
58

made up to the mark with deionized water. In a 10.0ml graduated test tube, 5.0ml
aliquot of this solution was taken and 0.5ml Nessler's reagent was added. The
contents of the test tubes were allowed to stand for 5min for maximum colour
development. The optical density of the solution was read at 525nm.
A standard curve was plotted using different concentrations of ammonium
sulphate solution versus optical density of the solution. The reading of each sample
was compared with the standard calibration curve to estimate the nitrogen content.
3.8.2.7 Determination of sulfur
Oven-dried leaves were powdered with mortar and pestle and meshed through a
72mm mesh sieve.
3.8.2.7.1 Digestion of leaf powder
Oven-dried leaf powder (lOOmg) was taken in digestion tube of 75ml capacity. In
digestion tube, 4.0ml acid mixture (consists of concentrated nitric acid and perchloric
acid in the ratio of 1:1) and 0.0075g of selenium dioxide as catalyst was added. The
digestion was carried out till the digested solution become colourless. Subsequent to
digestion, the volume was made up to 75.0ml with deionized water. The interference
of silica was checked by filtering the contents of the tube.
3.8.2.7.2 Estimation of sulfur
Total sulfur in plant samples was estimated according to turbidimetric method of
Chesnin and Yien (1951). A 5.0ml aliquot was pippeted out from the digested
solution for turbidity development in 25.0ml volumetric flasks. Turbidity was
developed by adding 2.5ml gum acacia (0.25%) solution, l.Og barium chloride
(BaCh, sieved through 40-60mm mesh) and the volume was made up to the mark,
with deionized water. Contents of 25.0ml volumetric flask were thoroughly shaken till
BaCl2 completely dissolved. Turbidity was allowed to develop for 2min. The values
were recorded at 415imi with in lOmin after the turbidity development. A blank was
also run simultaneously after each set of determination.
The amount of sulfate was calculated with the help of a calibration curve
drawn afresh using a series of K2SO4 solutions, prepared following the procedure
similar to aliquot, described as above.
3.8.2.8 Determination of cadmium
Cadmium content was determined in root and leaf samples. Leaves were washed with
deionized water, and the roots were washed with an ice-cold 5mM CaCb solution for
1 Omin to displace extracellular Cd (Rauser 1987). The root/leaf samples were dried
59

for 48h at 80°C, weighed and ground to fine powder, and digested with concentrated
HNO3-HCIO4 (3:1). Cadmium concentration was determined by atomic absorption
spectrophotometer (GBG, 932 plus, Australia).
3.8.2.9 Determination of thiobarbituric acid reactive substances
The level of lipid peroxidation products in the leaves was determined by
thiobarbituric acid reactive substances (TBARS) as described by Dhindsa et al.
(1981). Fresh leaf tissue (200mg) was ground in 0.25% 2-thiobarbituric acid (TBA) in
10% trichloroacetic acid (TCA) using mortar and pestle. After heating at 95°C for
30min, the mixture was quickly cooled on ice bath and centrifuged at 10,000 xg for
lOmin. The absorbance of the supematant was read at 532nm and corrected for non
specific turbidity by subtracting the absorbance of the same at 600nm. The blank was
0.25% TBA in 10% TCA. The TBARS content was calculated using the extinction
coefficient (155 mM"' cm'').
3.8.2.10 Determination of hydrogen peroxide content
The content of H2O2 in leaves was measured as described by Jana and Chaudhuri
(1981). Fresh leaves (lOOmg) were homogenized with 6.0ml of K-phosphate buffer
(50mM, pH 6.5). The homogenate was centrifiiged at 6,000xg for 25min. To 3.0ml of
the extracted solution, 1.0ml of 0.1% titanium sulfate in 20% H2SO4 was added, and
then centrifiiged at 6,000xg for 15min. The colour intensity of the supematant was
measured at 410nm. The H2O2 content was calculated using the extinction coefficient
(0.28^imor'cm'').
3.8.2.11 Components of enzymatic and non-enzymatic antioxidant system
3.8.2.11.1 Activities of antioxidant enzymes
3.8.2.11.2 Enzyme extraction
Fresh leaf tissue (200mg) was homogenized with an extraction buffer containing
lOOmM K-phosphate buffer (pH 7.0), 0.5% Triton X-100 and 1%
polyvinylpyrrolidone (PVP) using chilled mortar and pestle. The homogenate
was centrifuged at 15,000xg for 20min at 4°C. The supernatant obtained was
used for the enzymatic assays. For ascorbate peroxidase, extraction buffer was
supplemented with 2mM ascorbate. Enzyme extract was used for the assay of
superoxide dismutase, catalase, ascorbate peroxidase and glutathione reductase.
3.8.2.11.2.1 Superoxide dismutase activity
The method described by Dhindsa et a/. (1981) was followed with slight modification
for the estimation of superoxide dismutase activity.
60

3.8.2.11.2.1.1 Enzyme assay
Superoxide dismutase activity in the supernatant was assayed by its ability to inhibit
the photochemical reduction. The assay mixture, consisting of 1.5ml O.IM K-
phosphate buffer (pH 7.8), 0.2ml of methionine, 0.1ml enzyme extract with equal
amount of IM NaCOs, 2.25mM NBT solution, 3mM EDTA, 60^M riboflavin and
1.0ml of deionized water, was taken in test tubes which were incubated under light of
15W inflorescent lamp for lOmin at 25°C. Blank 'A' containing all the above
substances of the reaction mixture, along with the enzyme extract, was placed in the
dark. Blank 'B' containing all the above substances of reaction mixture except the
enzyme was placed in light along with the sample. The reaction was terminated by
switching off the light, and the tubes were covered with black cloth. The non-
irradiated reaction mixture containing enzyme extract did not develop light blue
colour. Absorbance of the samples along with Blank 'B' was read at 560nm against
the Blank 'A'. The difference of percent reduction in color between Blank 'B' and the
sample was then calculated.
A reduction of 50% in the colour was considered as one Unit of enzyme
activity.
3.8.2.11.2.2 Catalase activity
Catalase activity in leaves was determined by the method of Aebi (1984) with slight
modification.
3.8.2.11.2.2.1 Enzyme assay
Catalase activity was determined by monitoring the disappearance of H2O2 by
measuring the absorbance at 240run. Reaction was carried in a final volume of 2.0ml
of reaction mixture containing 0.5M K-phosphate buffer (pH 7.2) with 0.1ml 3niM
EDTA, 0.1ml of enzyme extract and 0.1ml of 3niM H2O2. The reaction was allowed
to run for 5min. Activity was calculated by using extinction coefficient 0.036mM"'
cm" . One Unit of enzyme activity is defined as the amount necessary to decompose
1 mol of H2O2 per min at 25°C.
3.8.2.11.2.3 Ascorbate peroxidase activity
Ascorbate peroxidase activity was estimated by the method used by Nakano and
Asada(1981).
3.8.2.11.2.3.1 Enzyme assay
Ascorbate peroxidase activity was determined by the decrease in absorbance of
ascorbate at 290nm, due to its enzymatic breakdown. 1.0ml of 50mM K-phosphate
61

buffer (pH 7.2) contained 0.5mM ascorbate, 0.1 mM H2O2, 0.1 mM EDTA and 0.1ml
enzyme extract. The reaction was allowed to run for 5min at 25°C. Ascorbate
peroxidase activity was calculated by using extinction coefficient 2.8 mM" cm" . One
Unit of enzyme activity is defined as the amount necessary to decompose 1 fimol of
substrate consumed per min at 25°C.
3.8.2.11.2.4 Glutathione reductase activity
Glutathione reductase activity was determined by the method of Foyer and Halliwell
(1976) as modified by Rao (1992).
3.8.2.11.2.4.1 Enzyme assay
Glutathione reductase activity was determined by monitoring the glutathione-
dependent oxidation of NADPH at its absorption maxima of wavelength 340nm.
Reaction mixture (1.0ml) contained 0.2mM NADPH, 0.5mM GSSG and 50|al of the
enzyme extract. The reaction was allowed to run for 5min at 25°C. Corrections were
made for any GSSG oxidation in the absence of NADPH. The activity was calculated
by using extinction coefficient (6.2 mNT cm"'). One Unit of enzyme activity is
defined as the amount necessary to decompose 1 famol of NADPH per min at 25 °C.
3.8.2.11.2.5 Soluble protein content
Soluble protein content of leaves was estimated following the method of Bradford
(1976).
Fresh leaf tissue (200mg) was homogenized in 2.0ml of extraction buffer with
the help of pre-chilled mortar and pestle at 4°C. The homogenate was transferred to
2.0ml tube and centrifiiged at 5000x rpm at 4°C for lOmin. An equal volume of 10%
trichloroacetic acid (TCA) was added to 1.0ml of the crude extract, and allowed to
centrifuge at 3300x rpm for lOmin. The precipitate was then discarded and the pellet
was dissolved completely in O.IN NaOH. To 0.1ml of aliquot, 5.0ml Bradford's
reagent was added and mixed vigorously. The intensity of blue colour developed
within 2min was read at 595nm. Calibration curve was drawn using different
concentrations of Bovine Serum Albumin (BSA) processed similarly as that of
aliquots to calculate protein content.
3.8.2.11.2 Non-enzymatic antioxidants
3.8.2.11.2.1 Ascorbic acid content
Estimation of ascorbic acid (AsA) content was done by the method of Law et al.
(1983) with slight modifications.
62

3.8.2.11.2.1.1 Procedure
Fresh leaf tissue (200mg) was ground in 2.0ml of lOOmM K-phosphate buffer (pH
7.0) with ImM EDTA and centrifuged at 10,000xg for lOmin. The supernatant was
collected and 200nl of 10% TCA was added. After mixing the solution by vortex, it
was allowed to stand for 5min. Following mixing, 10|al of 5M NaOH was added and
centriftjged for 2min. To 200^1 of supernatant, 200(J1 of 150mM K-phosphate buffer
(pH 7.4), 100^1 of lOmM 5',5'-dithiobis-2-nitrobenzoic acid (DTT) were added,
mixed thoroughly and left at room temperature for 15min. After mixing 100^1 of
0.5% N'N-ethylemaleimide (NEM) was added. Samples were mixed by vortex and
incubated at 24°C for 30sec. To each sample 400^1 of 10% TCA, 400^1 H3PO4,400^1
of 4% bipyridyl dye (N'N-dimethyl bipyridyl) and 200^1 of 3% FeCb were added.
After vortex-mixing, samples were incubated at 37°C for Ih and the absorbance was
recorded at 525imi. A standard curve in the range of 10-lOOnmol of ascorbic acid was
used for calibration. Values in both cases were corrected for the absorbance
eliminating the supernatant in the blank prepared separately for AsA and AsA+DHAs.
3.8.2.11.2.2 Glutathione content
Reduced (GSH) content was determined by the glutathione recycling method of
Anderson (1985). The details are as follows.
3.8.2.11.2.2.1 Procedure
Fresh leaf tissue (200mg) was homogenized in 2.0ml of 5%) sulphosalicylic acid under
cold conditions. The homogenate was centriftaged at 10,000xg for lOmin. To 0.5ml of
supernatant, 0.6ml of lOOmM K-phosphate buffer (pH 7.0) and 40|al of 5',5'-dithiobis-
2-nitrobenzoic acid (DTNB) were added. After 2min the absorbance was read at
412mn. Standard curve for calculations was prepared from GSH, covering a range of
10-lOOnmol.
3.8.3 Yield characteristics
Yield is the final manifestation of morphological, physiological and
biochemical traits of a crop.
At harvest (120DAS) following parameters were recorded.
1. Number of siliqua per plant
2. Number of seeds per siliqua
3. 1000 seed weight
4. Seed yield per plant
63

At harvest, siliqua were collected and counted. The number of seeds from each
siliqua was counted. From the produce of the pot, a sample of thousand seeds was
randomly drawn and the weight was recorded using electronic balance. The total
seeds from a plant in each treatment were cleared, sun-dried and weighed to compute
seed yield per plant.
3.8.4 Statistical analysis
The data were analyzed statistically using two way analysis of variance (ANOVA) in
each Experiment. F-value was calculated and the level of significance was determined
at p<0.05 using the SPSS statistical program (ver. 12.0 Inc., Chicago, USA). Least
significant difference (LSD) was calculated for the significant data at/7<0.05. Model
of ANOVA for the Experiments is given in Tables 8, 9 and 10.
64

Table 8. Model of analysis of variance (ANOVA) of Experiment 1.
Source of Variation df SS MSS F. value Sig.
Replication 2
Cultivar 4
Treatment 4
Interaction 16
Error 48
Total 74
Table 9. Model of analysis of variance (ANOVA) of Experiment 2.
Source of Variation df SS MSS F. value Sig
Replication
Cultivar
Treatment
Interaction
Error
Total
2
1
6
6
26
41
Table 10. Model of analysis of variance (ANOVA) of Experiment 3.
Source of Variation df SS MSS F. value SigT
Replication
Cultivar
Treatment
Interaction
Error
Total
2
1
5
5
22
35

EXPERIMENTAL RESULTS

Contents
EXPERIMENTAL RESULTS
4.1 Experiment 1: Screening of Mustard Cultivars Treated with Different Levels of Cadmium 65
4.1.1 Growth characteristics 65 4.1.2 Photosynthetic characteristics 66 4.1.3 Cadmium accumulation 68 4.1.4 Yield characteristics 68 4.2 Summary of Experiment 1 69 4.3 Experiment 2: Study of Alleviation of Cd-induced Effects With
Sulfur Application Cd-tolerant and non-tolerant Mustard Cultivars 69 4.3.1 Growth characteristics 69 4.3.2 Photosynthetic characteristics 70 4.3.3 Sulfur and nitrogen assimilation 71 4.3.4 Cadmivmi accumulation 72 4.3.5 Thiobarbituric acid reactive substances and hydrogen peroxide
content 73 4.3.6 Components of antioxidant system 74 4.3.6.1 Enzymatic antioxidants 74 4.3.6.2 Non-en2ymatic antioxidants 75 4.3.7 Yield characteristics 76 4.4 Summary of Experiment 2 77 4.5 Experiment 3: Coordination of Sulfur and Nitrogen Application in
the Alleviation of Cd-toxicity in Cd tolerant and non-tolerant Mustard Cultivars 78
4.5.1 Growth characteristics 78 4.5.2 Photosynthetic characteristics 78 4.5.3 Sulfur and nitrogen assimilation 79 4.5.4 Cadmium accumulation 80 4.5.5 Thiobarbitxiric acid reactive substances and hydrogen peroxide
content 80 4.5.6 Components of antioxidant system 81 4.5.6.1 Enzymatic antioxidants 81 4.5.6.2 Non-enzymatic antioxidants 82 4.5.7 Yield characteristics 83 4.6 Summary of Experiment 3 83

EXPERIMENTAL RESULTS Chapter 4
Experimental results are presented herein mainly as changes in growth and
photosynthetic characteristics, sulfur and nitrogen assimilation, components of
antioxidant system and yield characteristics of mustard cultivars as influenced by
different concentrations of cadmium, sulfur and nitrogen.
4.1 Experiment 1: Screening of Mustard Cultivars Treated with Different Levels
of Cadmium
The Experiment was conducted to study the effect of five concentrations of Cd viz., 0,
25, 50, 100 and 150mg kg'' soil on Cd accumulation, growth, photosynthetic and
yield characteristics of five mustard cultivars, namely Alankar, Varuna, Pusa Bold,
Sakha and RH30. Tolerance index of the five cultivars of mustard was calculated and
cultivars were designated as Cd tolerant and Cd non-tolerant on the basis of their
performance under Cd stress. Results are described below in detail.
4.1.1 Growth characteristics
The effect of Cd on growth characteristics was found significant at the three sampling
stages. The interaction of Cd treatments and cultivars was also found significant at 60
and 90DAS but was non-significant at SODAS for all the growth characteristics
studied (Figures 2-5). The increase in Cd concentration decreased the growth
characteristics of all the five cultivars at all sampling stages. The observations
recorded at all the sampling stages showed similar pattern of cultivar response to Cd
treatments. The pattern of decrease in growth characteristics for the cultivars was
RH30 >Sakha >Pusa Bold >Varuna >Alankar. Maximum reduction in growth
characteristics was noted with 150mg Cd kg'' soil followed by 100, 50 and 25mg Cd
kg'' soil at all growth stages.
Among cultivars, Alankar showed lesser decrease in growth characteristics
followed by Varuna and Pusa Bold, whereas, RH30 and Sakha exhibited greater
reduction in growth characteristics under Cd stress.
In Alankar, root length was decreased by 9.1, 15.1, 24.2, 37.1% at 30DAS;
7.1, 11.9, 20.2, 32.1% at 60DAS and 5.2, 10.4, 18.0, 27.7% at 90DAS due to 25, 50,
100, and 150mg Cd kg'' soil, respectively, over their respective control. Contrarily, a
higher decrease of 17.0, 27.0, 40.0, 58.0% at 30DAS; 16.4, 30.3, 45.0, 60.6% at
60DAS and 13.0, 23.7, 33.6, 42.6% at 90DAS in root length of RH30 was noted with
65

1
I I
30DAS LSDoo5(CxT) = NS
60DAS ,0.05 (CxT) = 0.89
18
16 ^
14
12 -I
10
8
6 18
16 •
14 •
12
10
8
6 18
16
14
12 H
10
8
6 16
14
12
10
8
6
4 14 1
12
10
8
6 ^
4
2
90DAS LSDo„5(CxT) = NS
Varuna
Pusa Bold
Sakha
RH30
25 50 100 150
-1 Treatments (mg kg" soil)
Fig. 2. Effect of Cd treatments (T) on root length (cm plant'') of five cultivars (C) of mustard {Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

SODAS
100 90 80 70 60 50 40 30 20
100 90 80 70 60 50 40 30
y-N
•4-'
c SJ O.
F u x: tt, c _a) ^-i
o o C/1
20 90 80
70
60
50 40
30
20 80
70
60
50
40
30
20
10 70
60
50
40
30
20
10
60DAS LSDoo5(CxT) = 2.32
Aiankar
90DAS LSD„o3 (CxT) = 2.08
Varuna
Pusa
Sakha
RH30
25 50 100 150
Treatments (mg kg soil)
Fig. 3. Effect of Cd treatments (T) on shoot length (cm plant"') of five cultivars (C) of mustard {Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

- • - 30DAS LSDoo5(CxT) = NS
1000
800
600
400 -
200 •
0 1000
800
600 ^
400
200
0 800 1
§ 600
1 400 i to
200 •
0 800
600
400
200
0 700
600
500
400
300
200
100
0
60DAS LSDoo5(CxT)= 10.78
Alankar
90DAS LSDoo5(CxT) = 7.80
Varuna
Pusa Bold
Sakha
RH30
25 50 100
Treatments (mg kg soil)
150
Fig. 4. Effect of Cd treatments (T) on leaf area (cm^ plant"') of five cultivars (C) of mustard (Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

SODAS LSDoo5(CxT) = NS
60DAS LSDo<,5(CxT) = 0.28
Alankar
90DAS LSD„o3(CxT) = 0.57
Treatments (mg kg" soil)
Fig. 5. Effect of Cd treatments (T) on plant dry mass (g plant'') of five cultivars (C) of mustard {Brassica juncea L.) at 30, 60 and 90days afler sowing (DAS).

25, 50, 100 and 150mg Cd kg"' soil, respectively, over their respective control
(Figures 2,6).
The decrease in shoot length followed the same pattern as observed in root
length. In comparison to control, shoot length of Alankar decreased by 5.4, 10.2, 17.4,
26.2% at 30DAS; 4.6, 8.9, 14.7, 24.8% at 60DAS and 3.4, 6.5, 12.5, 20.4% at 90DAS
with 25, 50, 100 and 150mg Cd kg'' soil, respectively. In RH30, reductions of 10.3,
22.7, 29.6, 47.0% at 30DAS; 14.3, 26.8, 30.6, 50.2% at 60DAS and 8.4, 20.3, 25.6,
42.2% at 90DAS were noted with 25, 50, 100 and 150mg Cd kg'' soil, respectively,
over their respective control (Figures 3,6).
When compared to control, leaf area of Alankar decreased by 8.6, 15.5, 29.3,
37.9% at 30DAS; 9.5, 15.5, 30.5, 40.0% at 60DAS and 5.0, 7.7, 16.9, 26.5% at
90DAS with 25, 50, 100 and 150mg Cd kg'' soil, respectively. Whereas, RH30
showed reductions of 16.7, 26.2, 38.1, 54.7% at 30DAS; 21.9, 30.9, 40.8, 58.1% at
60DAS and 15.9, 21.6, 32.7, 42.5% at 90DAS with 25, 50, 100 and 150mg Cd kg"'
soil, respectively, over their respective control (Figures 4,7).
The trend of plant response to Cd treatments in terms of plant dry mass was
almost similar to those observed for the individual growth characteristics. Plant dry
mass of Alankar decreased by 6.0, 11.3, 25.3, 33.3% at 30DAS; 5.4, 9.5, 21.2, 29.8%
at 60DAS and 3.4, 7.1, 20.3, 26.9% at 90DAS due to 25, 50, 100 and 150mg Cd kg"'
soil, respectively, in comparison to their respective control. While, in RH30, plant dry
mass was decreased by 13.2, 21.5, 39.6, 54.5% at 30DAS; 15.5, 25.3, 41.4, 60.3% at
60DAS and 9.6, 18.1, 31.6, 43.7% at 90DAS with 25, 50, 100 and 150mg Cd kg"'
soil, respectively, over their respective control (Figures 5, 7). The trend of sensitivity
of mustard cultivars to Cd toxicity in terms of tolerance index (calculated on the basis
of plant dry mass) was: RH30 >Sakha >Pusa Bold >Varuna >Alankar (Figure 8).
4.1.2 Photosynthetic characteristics
Photosynthetic characteristics (total chlorophyll content, carotenoids content, carbonic
anhydrase activity, net photosynthetic rate and stomatal conductance) of the five
mustard cultivars decreased significantly with the increasing Cd concentration at all
growth stages (Figures 9-13). The interaction of Cd treatments and cultivars was
significant at all the sampling stages except for carotenoids content which remained
non-significant at 30DAS. The greatest significant reduction in photosynthetic
characteristics was recorded with highest Cd level (150mg Cd kg"' soil) in all the
66

Cultivars
Fig. 6. Per cent change in root and shoot length of five cultivars of mustard {Brassica juncea L.) due to 150mg Cd kg"' soil over control at 30, 60 and 90days after sowing (DAS).

40 -a
N \ - ^ ' ^ 0 ° ' ' S ' " ^ N X - ^ ' % 0 ° ' ' S * " ^ Cultivars
Fig. 7. Per cent change in leaf area and plant dry mass of five cultivars of mustard (Brassicajuncea L.) due to 150mg Cd kg"' soil over control at 30, 60 and 90days after sowing (DAS).

X V
u o B (8
O
v\j -
80 -
60 -
40 -
20 -
0 -
T ,
I I ^ d I 1 I I ^^^9H^^^I ^^^SM^^H ^^^M^^^I ^^BSJ^^B
^^^^H ^ H ^ H ^H^^H ^ l ^ ^ l ^ ^ ^ ^ 1 ^ ^ ^ ^ 1 ^ ^ ^ ^ 1 ^ I' S I
• ^ 30DAS g ^ l bODAb
• • 90DAS
1 1 m
1 ^H *(l l 1 I ^^H
1 19 Alankar Varuna Pusa Bold
Cultivars
Sakha RH30
Fig. 8. Tolerance index of five cultivars of mustard (Brassica juncea L.) treated with 150mg Cd kg"' soil. Tolerance index was calculated as percentage of the plant dry mass obtained in 150mg Cd kg"' soil and control at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates ± S.E.

cultivars. The pattern of decrease in photosynthetic characteristics was: RH30 >Sakha
>Pusa Bold >Varuna >Alankar.
Maximum reduction in total chlorophyll and carotenoids content was observed
in RH30 and least in Alankar under Cd stress at all sampling stages. In Alankar, total
chlorophyll content was decreased by 13.7, 21.6, 32.7, 44.4% at 30DAS; 10.9, 17.4,
22.3, 35.3% at 60DAS; 13.0, 20.0, 28.0, 41.0% at 90DAS and in carotenoids content
by 5.3, 17.4, 21.1, 32.6% at 30DAS; 4.3, 17.1, 25.2, 34.2% at 60DAS; 4.8, 18.6,25.7,
38.3% at 90DAS with 25, 50, 100 and 150mg Cd kg"' soil, respectively, over their
respective control. RH30 showed greater reduction in total chlorophyll content, which
was 20.3, 31.6, 43.6, 60.1% at 30DAS; 26.3, 41.3, 52.6, 67.7% at 60DAS; 26.9, 46.1,
53.8, 69.2% at 90DAS and carotenoids content decreased by 16.0, 33.3, 40.7, 49.3%
at 30DAS; 19.7, 34.3, 44.5, 54.0% at 60DAS; 18.7, 34.9, 45.5, 55.3% at 90DAS with
25, 50, 100 and 150mg Cd kg'' soil, respectively, over their respective control
(Figures 9,10,14).
The decrease in carbonic anhydrase activity in Alankar was 8.6, 14.5, 32.7,
40.7% at 30DAS; 9.1, 16.4, 36.8, 44.7% at 60DAS and 9.3, 20.3, 39.4, 45.2% at
90DAS with 25, 50, 100 and 150mg Cd mg"' soil, respectively, over their respective
control. Carbonic anhydrase activity in RH30 was decreased by 22.1, 35.0, 55.4,
67.5% at 30DAS; 26.6, 46.9, 58.3, 70.6% at 60DAS and 29.4, 48.8, 63.5, 78.9% at
90DAS with 25, 50, 100 and 150mg Cd mg'' soil, respectively, over their respective
control (Figures 11,15).
In comparison to control, net photosynthetic rate of Alankar was decreased by
7.9, 14.3, 32.5, 37.7% at 30DAS; 7.1,13.2, 30.8, 33.8% at 60DAS and 5.9, 12.5, 27.6,
29.9% at 90DAS with 25, 50, 100 and 150mg Cd kg'' soil, respectively, over their
respective control. Whereas, in RH30, it was decreased by 12.4, 27.9, 46.1, 53.5% at
30DAS; 14.3, 32.0,47.4, 54.7% at 60DAS and 14.1, 33.3,47.8, 60.7% at 90DAS with
25, 50, 100 and 150mg Cd kg'' soil, respectively, over their respective control
(Figures 12,16).
The decrease in stomatal conductance followed the same pattern as observed
in net photosynthetic rate. When compared with the respective control, reduction in
stomatal conductance in Alankar was 9.7, 16.1, 25.2, 40.3% at 30DAS; 11.3, 17.7,
30.0, 42.3% at 60DAS and 9.8, 20.2, 30.4, 45.5% at 90DAS with 25, 50, 100 and
150mg Cd kg"' soil, respectively. RH30 showed decrease in stomatal conductance by
16.4, 26.5, 49.8, 57.8% at 30DAS; 18.9, 30.4, 52.6, 60.3% at 60DAS and 21.0, 34.9,
67
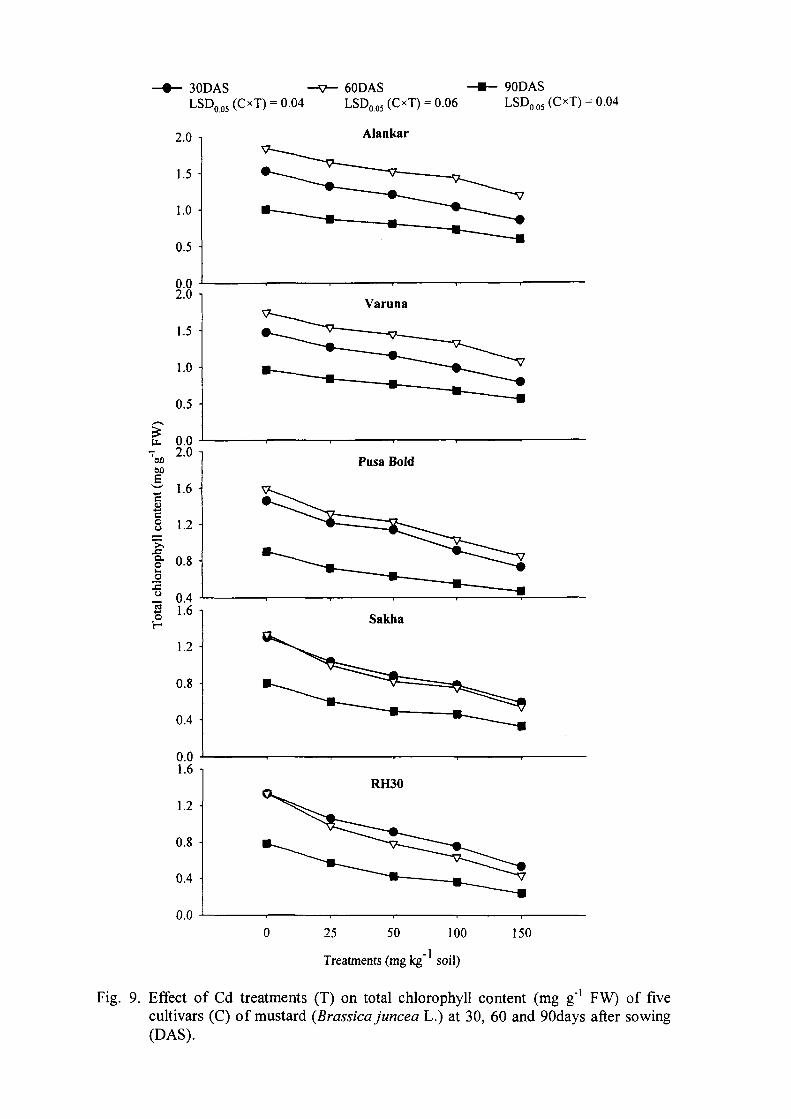
30DAS LSD„o5(CxT) = 0.04
2.0
1.5 ^
1.0
0.5 ^
0.0 2.0
1.5 -
1.0 -
0.5
u.
au 00
e N w /
c (1>
e o u • > .
J: a. o t^
o JS u 4-<
0.0 ?,.o
1.6
1.2
0.8
0.4 1.6
1.2
0.8
0.4 •
0.0 1.6 1
1.2
0.8 -
0.4
0.0
60DAS LSDooj (CxT) = 0.06
Alankar
Varuna
Pusa Bold
Sakha
RH30
25 50 100
Treatments (mg kg soil)
90DAS LSDo.05 (CxT) = 0.04
150
Fig. 9. Effect of Cd treatments (T) on total chlorophyll content (mg g'' FW) of five cultivars (C) of mustard (Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

30DAS LSD,o5(C'<T) = NS
0.20
0.15
0.10
0.05 •
to
o u CO
-a '5 c <u o u ca
U
0.00 0.25
0.20
0.15
0.10
0.05
0.00 0.20
0.18
0.16
0.14
0.12
0.10
0.08 0.20
0.15
0.10
0.05
0.00 0.16
0.12 -
0.08
0.04
0.00
60DAS LSDoo5 (CxT) = 0.007
Alankar
90DAS LSDQQ5 (CXT) = 0.007
Varuna
Pusa Bold
Sakha
RH30
25 50 100 150
Treatments (mg kg' soil)
Fig. 10. Effect of Cd treatments (T) on carotenoids content (mg g"' FW) of five cultivars (C) of mustard {Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

O U
>
< u
SODAS LSDoo5(CxT) = 0.21
10
8^
6
4
2
0 10
8
6
4
2
0 10
8
6
4
2
0
4 ^
2
60DAS LSDg jj (CxT) = 0.22
Aiankar
90DAS LSDooj (CxT) = 0.26
Varuna
Pusa Bold
Sakha
RH30
25 50 100
Treatments (mg kg' ' soil)
150
Fig. 11. Effect of Cd treatments (T) on carbonic anhydrase activity (mmol CO2 g"' FW) of five cultivars (C) of mustard (Brassica Juncea L.) at 30, 60 and 90days after sowing (DAS).

o u "o e
c
o o.
Z
30DAS LSDo„5(CxT) = 0.40
25
20
15
10
5
0 20
15 -
10 -
5 -
0 20
18
16
14
12
10
8
6 18 16 14 12 10 8 6 4 2 0
16 14 12 10 8 6 •
4 •
2 0
60DAS LSDoo5(CxT) = 0.51
Alankar
90DAS LSDo<,3(CxT) = 0.33
Pusa Bold
Sakha
RH3G
25 50 100 150
-1 Treatments (mg kg" soil)
Fig. 12. Effect of Cd treatments (T) on net photosynthetic rate ( trnol CO2 m" s'') of five cultivars (C) of mustard {Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

30DAS LSD(,o5(CxT)= 11.02
25
20 -i
15
10 -
5 •
0 20
15
10
5 •
l/l
's "o
g ^•m^
0)
u s CQ
u 3
o u BJ
o on
0 20
18
16
14
12 10
X 6
18
14 12 10 8 6 4 2 0
16 14 12 10 8
6 4 -I 2 0
60DAS LSDo,„(CxT)= 12.21
Alankar
90DAS LSDoo5 (CxT) = 48.40
Varuna
Pusa Bold
Sakha
RH30
25 50 100 150
Treatments (mg kg soil)
Fig. 13. Effect of Cd treatments (T) on stomatal conductance (mmol m' s'') of five cultivars (C) of mustard {Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

N\-^' '^<vv^^°' ' S-^" ^ ' ^ ' N X - ^ ' ^ < ^ . . ^ ° ' ^ ^-^ ' ^ ' '
Cultivars
Fig. 14. Per cent change in total chlorophyll content and carotenoids content of five cultivars of mustard {Brassica juncea L.) due to 150mg Cd kg"' soil over control at 30, 60 and 90days after sowing (DAS).

Alankar Varuna PusaBold Sakha RH30
Cultivars
Fig. 15. Per cent change in carbonic anhydrase activity of five cultivars of mustard {Brassica juncea L.) due to 150mg Cd kg'' soil over control at 30, 60 and 90days after sowing (DAS).

^X.^''\^^,M' S- ' ^ ' ^ ' N X ^ ^ ' % < ^ S . ^ ° ' ' ^-^ ' ^ ' '
Cultivars
Fig. 16. Per cent change in net photosynthetic rate and stomatal conductance of five cultivars of mustard {Brassica juncea L.) due to 150mg Cd kg"' soil over control at 30, 60 and 90days after sowing (DAS).

54.2, 68.1% at 90DAS with 25, 50, 100 and 150mg Cd mg'' soil, respectively, in
comparison to their respective control (Figures 13,16).
4.1.3 Cadmium accumulation
Root and leaf Cd accumulation increased with the increase in soil Cd concentration
and was significant at 30, 60 and 90DAS (Figures 17-18). The effect of Cd treatment
and cultivar interaction was also found significant at all the growth stages. For every
concentration used in the study, Cd concentration was more pronounced in the roots
than the leaves in all the five cultivars. The trend of Cd accumulation in root and leaf
of five cultivars was: RH30 >Sakha >Pusa Bold >Varuna >Alankar at all growth
stages.
Among cultivars, RH30 accumulated highest amount of Cd in root and leaf,
whereas, Alankar accumulated least. Maximum accumulation of Cd in the root of
RH30 was 354.2, 403.7, 452.8^g Cd g"' DW and in the leaf the accumulation was
137.1, 205.6, 130.2^g Cd g"' DW at 30, 60 and 90DAS, respectively of plants treated
with 150mg Cd kg"' soil. Root and leaf of Alankar accumulated 273.1, 330.8,
381.2^g Cd g-' DW and 70.7, 95.3, 51.2^g Cd g- DW at 30, 60 and 90DAS,
respectively, when treated with 150mg Cd kg'' soil (Figures 17-19).
4.1.4 Yield characteristics
The increasing concentration of Cd significantly decreased the yield characteristics of
all the five cultivars (Figure 20). The interaction of Cd treatments and cultivars was
also significant. Maximum decrease in yield characteristics was noted with 150mg Cd
kg'' soil. The cultivars RH30 and Sakha exhibited similar and greatest decrease in
yield characteristics, whereas, Alankar showed lowest decrease followed by Varuna
and Pusa Bold. The order of performance for yield characteristics of cultivars was
Alankar >Varuna >Pusa Bold >Sakha >RH30.
In Alankar, the decrease in number of siliqua per plant was 11.5, 22.7, 30.4,
35.7%; number of seeds per siliqua was 10.6, 22.7, 32.2, 33.8%; 1000 seed weight
was 13.7, 25.0, 35.8, 41.4% and seed yield was 13.7, 21.6, 28.1, 32.5% due to 25, 50,
100 and 150mg Cd kg" soil, respectively, over their respective control. In contrast,
RH30 showed greater reduction in number of siliqua per plant which was 24.5, 35.5,
43.5, 53.8%, number of seeds per siliqua was decreased by 25.2, 39.3, 51.1, 55.9%,
1000 seed weight by 25.6, 37.4, 53.2, 60.0% and seed yield by 22.4, 35.3, 45.4,
52.7% with to 25, 50, 100 and 150mg Cd kg"' soil, respectively (Figures 20-21).
68

SODAS LSD„o3(CxT) = 6.21
Q
'oo 60
B
c o o
U 100 i o o
60DAS LSDo_o3(CxT)= 13.79
Alankar
90DAS LSDoo5(CxT) = 6.33
25 50 100 150
Treatments (mg kg" soil)
Fig. 17. Effect of Cd treatments (T) on root Cd content (i^g g"' DW) of five cultivars (C) of mustard (Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

SODAS LSDoo5(CxT) = 2.92
60DAS LSDo,o5(CxT) = 2.37
Alankar
90DAS LSDoos (CxT) = 2.42
25 50 100 150
Treatments (mg kg soil)
Fig. 18. Effect of Cd treatments (T) on leaf Cd content (^g g"' DW) of five cultivars (C) of mustard {Brassica juncea L.) at 30, 60 and 90days after sowing (DAS).

N \ - ^ ^ 0 ° ' ' S ' ^^' N\- '%<,s. °' ^-^' ^ ' ' ^j-o>'
Cultivars
Fig. 19. Per cent change in root Cd content and leaf Cd content of five cultivars of mustard {Brassica juncea L.) due to 150mg Cd kg"' soil over control at 30, 60 and 90days after sowing (DAS).

100
80
60 ^
40
20
0 100
80
60
40 -
20 -
Number of siliqua per plant LSDoo5(CxT)=1.83
Number of seeds per siliqua LSDoo5(CxT) = 0.31
Atankar
1000 seed weight (g) LSD„o3(CxT) = 0.12
Seed yield (g plant'') LSDo„5(CxT) = 0,14
-8 S- ^
0 80
60
40
20
Varuna
60
50
40
30
20
10
0
Pusa Bold
s ^ ^
25 50 100 150
-1 Treatments (mg kg soil)
Fig. 20. Effect of Cd treatments (T) on number of siliqua per plant, number of seeds per siliqua, 1000 seed weight (g) and seed yield (g plant'') of five cultivars (C) of mustard {Brassica juncea L.) at harvest i.e., 120days after sowing (DAS).

Alankar Vanina PusaBold Sakha RH30 Alankar Vanina PusaBold Saidia RH30
Cultivars
Fig. 21. Per cent change in number of siliqua per plant, number of seeds per siliqua, 1000 seed weight and seed yield of five cultivars of mustard (Brassica juncea L.) due to 150mg Cd kg"' soil over control at harvest i.e., 120days after sowing (DAS).

4.2 Summary of Experiment 1
• The accumulation of Cd in root and leaf was found Cd dose dependent. The
accumulation was maximum in RH30 followed by Sakha, Pusa Bold, Varuna
and Alankar.
• Cadmium treatments resulted in an overall reduction in growth, photosynthetic
and yield characteristics in mustard cultivars with more pronounced decrease
with the 150mg Cd kg'' soil treatment.
• Among cultivars, RH30 showed greater reduction in growth, photosynthetic
and yield characteristics, whereas, least decrease in these characteristics was
found in Alankar.
• The trend of sensitivity of mustard cultivars to Cd toxicity was: RH30 >Sakha
>Pusa Bold >Varuna >Alankar.
• On the basis of overall performance of the cultivars under Cd stress, Alankar
emerged as Cd tolerant and RH30 as Cd non-tolerant.
4.3 Experiment 2: Study of Alleviation of Cd-induced Effects with Sulfur
Application in Cd-tolerant and non-tolerant Mustard Cultivars
Experiment 2 was conducted on the basis of the findings of Experiments 1. As
observed in Experiment 1, Alankar emerged as Cd-tolerant and RH30 as Cd non-
tolerant cultivar. The treatment 150mg Cd kg"' soil caused maximum decrease in the
observed characteristics and 50mg Cd kg"' soil was moderate toxic in effects.
Therefore, the present Experiment was aimed to study the alleviation potential of
elemental S (0, 50 and lOOmg S kg'' soil) under the moderate (50mg Cd kg'' soil) and
extreme (150mg Cd kg'' soil) Cd levels by studying the changes in Cd accumulation,
growth and photosynthetic characteristics, sulfur and nitrogen assimilation,
components of antioxidant system and yield characteristics of tolerant (Alankar) and
non-tolerant (RH30) mustard cultivars.
Detailed results of this Experiment are presented in the following section.
4.3.1 Growth characteristics
Growth characteristics (root length, shoot length, leaf area and plant dry mass)
decreased in both the cultivars treated with Cd and the decrease was proportional to
the concentration of Cd (50 and 150mg Cd kg'' soil). Sulfur application nullified the
effects of 50mg Cd kg"', but could not ameliorate the effects of 150mg Cd kg' soil.
Ironically, the application of 50mg S kg"' soil proved most effective in improving
growth characteristics of both the cultivars treated with 50mg Cd kg'' soil. Whereas,
69

no such effect was observed for the supplementation of lOOmg S kg"' soil (Figures 22-
23).
Both the levels of Cd (50 and 150mg Cd kg'' soil) significantly reduced the
growth characteristics of both the cultivars. The extent of decrease was greater in non-
tolerant cultivar RH30 than the tolerant cultivar Alankar. In Alankar, the variation in
the reduction of plant dry mass was 10.5 and 35.7% at 30DAS; 7.9 and 27.9% at
60DAS and 7.1 and 27.9% at 90DAS due to 50 and 150mg Cd kg"' soil, respectively,
in comparison to their respective control. In RH30, the decrease in plant dry mass due
to 50mg Cd kg"' soil was about 25% and 50% due to 150mg Cd kg"' soil at all
sampling times (Figures 24-25).
The application of 50mg S kg"' soil completely overcome the effects of 50mg
Cd kg"' soil and also increased the growth characteristics. This increasing effect of S
was seen in Alankar. Root length, shoot length, leaf area and plant dry mass of
Alankar treated with 50mg Cd kg"' soil were increased by 1.5, 3.9, 1.6, 1.4% at
30DAS; 3.3, 4.7, 4.3, 2.6% at 60DAS and 2.6, 3.6, 3.9, 1.9% at 90DAS, respectively,
over their respective control due to 50mg S kg"' soil (Figures 22-25).
In RH30, supplementation of 50mg S kg"' soil reduced the decrease in growth
characteristics caused by 50mg Cd kg"' soil (Figures 22-25). The effect of
supplementation of lOOmg S kg"' soil in the alleviation of the effects of 50mg Cd kg"'
soil was lesser than 50mg S kg"' soil. Furthermore, no ameliorative effect of lOOmg S
kg'' soil was observed in plants treated with 150mg Cd kg"' soil (Figures 22-25).
4.3.2 Photosynthetic characteristics
Significant reductions in photosynthetic characteristics (total chlorophyll content,
carotenoids content, carbonic anhydrase activity, net photosynthetic rate and stomatal
conductance) were noted with 50 and 150mg Cd kg'' soil in both the cultivars.
However, the extent of decrease was higher in non-tolerant cultivar RH30 than
Alankar (tolerant). The application of 50mg S kg"' soil alleviated and improved the
photosynthetic characteristics of both the cultivars treated with 50mg Cd kg" soil.
The application of lOOmg S kg"' soil proved ineffective in the alleviation of 150mg
Cd kg"' soil-induced toxicity (Figures 26-28, 29-31).
Lesser decrease in photosynthetic characteristics was noted when plants were
treated with 50mg S kg"' soil plus 50mg Cd kg"' soil in comparison to SOmg Cd kg"'
soil alone. In Alankar, supplementation of 50mg Cd kg"' soil treated plants with 50mg
S kg"' soil limited the decrease in total chlorophyll and carotenoids content and
70

— n.
OS ^ - '
ri II
16
14
12
10
8
6
4
2
0
25
20
15 •
10
LSD„o5(CxT) = 0.50
oc
OS > - ^
0
25
20 •
15 -
10
5 ]
•— Alankar RH30
LSD„„3(CxT) = 0.32
30DAS I • • I — - — T
LSDoo5 (CxT) = 0.40 LSD„„(CxT) = 0.48
60DAS T ' I-" ' T . f • — - . -^ -
LSD„„, (CxT) = 0.39 LSD<,03 (CxT) =1.61
90DAS
r 50
• 40
30 f,-r
20
10
0
100
80
60 t C w a .
h40 ^ 1
- 20
0
120
100
80
0 0 ' * j
c c 60 -^ "=
40
20
Control 50Cd 150Cd 50Cd 50Cd 150Cd ISOCd Control 50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ 50S + 1 0 0 S + 5 0 S + lOOS + SOS + lOOS + 50S + lOOS
Treatments (mg kg'' soil)
Fig. 22. Effect of sulfur (S) application on root and shoot length (cm plant'') of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

80
60
<*- a. 40
20
0
1000
800
o -^ 600 u c
-I i 400-1
200 -
0 800
600 -
« c ;2 -E. 400
200
LSD„„3(CxT) = 2.99 Alankar RH30 LSDoo5 (CxT) = 0.04
30DAS
LSDooj (CxT) = 20.41 LSDoo, (CxT) = 0.10
60DAS
LSD„„3 (CxT) =19.40 LSD„„, (CxT) = 0.79
90DAS
r 2.0
1.6
1 2 =3 -
0.8 I ^
0.4
0.0
r 14
12
I- 10
6
• 4
• 2
0
40
30
20
10
Control 50Cd l50Cd 50Cd 50Cd 150Cd 150Cd
+ SOS + lOOS + SOS + IOCS
Control SOCd ISOCd SOCd SOCd ISOCd ISOCd
+ 50S + IOCS + 5 0 S + IOCS
•^ a . c oc a '—-
to •—'
Treatments (mg kg" soil)
Fig. 23. Effect of sulfur (S) application on leaf area (cm^ plant"') and plant dry mass (g plant'') of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

50Cd ISOCd 50Cd
+ 50S
50Cd
+ lOOS
150Cd
+ 50S
ISOCd
+ 100S
SOCd ISOCd SOCd
+ 50S
SOCd
+ IOCS
ISOCd
+ 50S
ISOCd
+ lOOS
Treatments (mg kg" soil)
Fig. 24. Per cent change in root and shoot length of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

50Cd 150Cd 50Cd
+ 50S
50Cd
+ lOOS
ISOCd
+ 50S
150Cd
+ lOOS
50Cd ISOCd 50Cd
+ 50S
50Cd
+ lOOS
ISOCd
+ 50S
ISOCd
+ lOOS
Treatments (mg kg' soil)
Fig. 25. Per cent change in leaf area and plant dry mass of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) imder Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

2.0
1,5 •
1.0 •
0.5
0.0
2.5
2.0 -
1.5 •
1.0
0.5
0.0
1.2
1.0 -
0.8 -
0.6
0.4 -
0.2
0.0
LSDoo5(CxT) = 0.05 Alankar RH30 LSDoo5(CxT) = 0.002
30DAS r r
LSDoo5 (CxT) = 0.05 LSD„o5(CxT) = 0.003
60DAS
LSDo.o3(CxT) = 0.03 LSDoo5 (CxT) = 0.003
90DAS
Control 50Cd 150Cd 50Cd 50Cd I50Cd l50Cd Conlroi 50Cd l50Cd 50Cd 50Cd l50Cd 150Cd
+ SOS + lOOS + SOS + lOOS + SOS + lOOS + SOS + lOOS
0.25
0.20
0.15
0 10
o o V5
•o o c (L>
o
u. ~p
0/J OJJ E "•—'
0.05
0.00
0.25
0.20
c
0.15 I i~ T3 T •5 M
\ 0.10 B. E o --'
0.05
0.00
r 0.20
0.15
0.10 Y'^ C 00
£ E
• 0.05
0.00
Treatments (mg kg" soil)
Fig. 26. Effect of sulfur (S) application on total chlorophyll content (mg g"' FW) and carotenoids content (mg g'' FW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

10 2 -
0 I
^1
• > ofi
0
12
10
8 -I
3 -
1 -
LSDQ 05 (CxT) = 0.03 Alankar RH30
30DAS
LSD„„ (CxT) = 0.09
60DAS
LSDo„5 (CxT) = 0.04
90DAS
Control 50Cd 150Cd 50Cd + 50S
50Cd + lOOS
!50Cd + 50S
150Cd + 100S
Treatments (mg kg" soil)
Fig. 27. Effect of sulfur (S) application on carbonic anhydrase activity (nmol CO2 g"' FW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

20 1
16
12
4 -
0
24
20 -
16 -
12 -
0
16
12
8 -
LSD„„3(CxT) = 0.45 -•— Alankar - O - RH30 LSDoo5(CxT) = 8.03
400
• 300
- 200
100
30DAS
LSD<,<,3(CxT) = 0.92 LSD„o3 (CxT) = 8.99
0
500
400
- 300
- 200
100
60DAS
LSDoo3(CxT) = 0.37 LSDoo5 (CxT) = 8.29
0
400
- 300
• 200
100
90DAS
Control 50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ 505 + lOOS + 505 + lOOS
Control 50Cd 150Cd 50Cd 50Cd l50Cd 150Cd
+ 505 + lOOS + 505 + lOOS
Treatments (mg kg" soil)
Fig. 28. Effect of sulfur (S) application on net photosynthetic rate (fjmol CO2 m" s"') and stomatal conductance (mmol m" s"') of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.
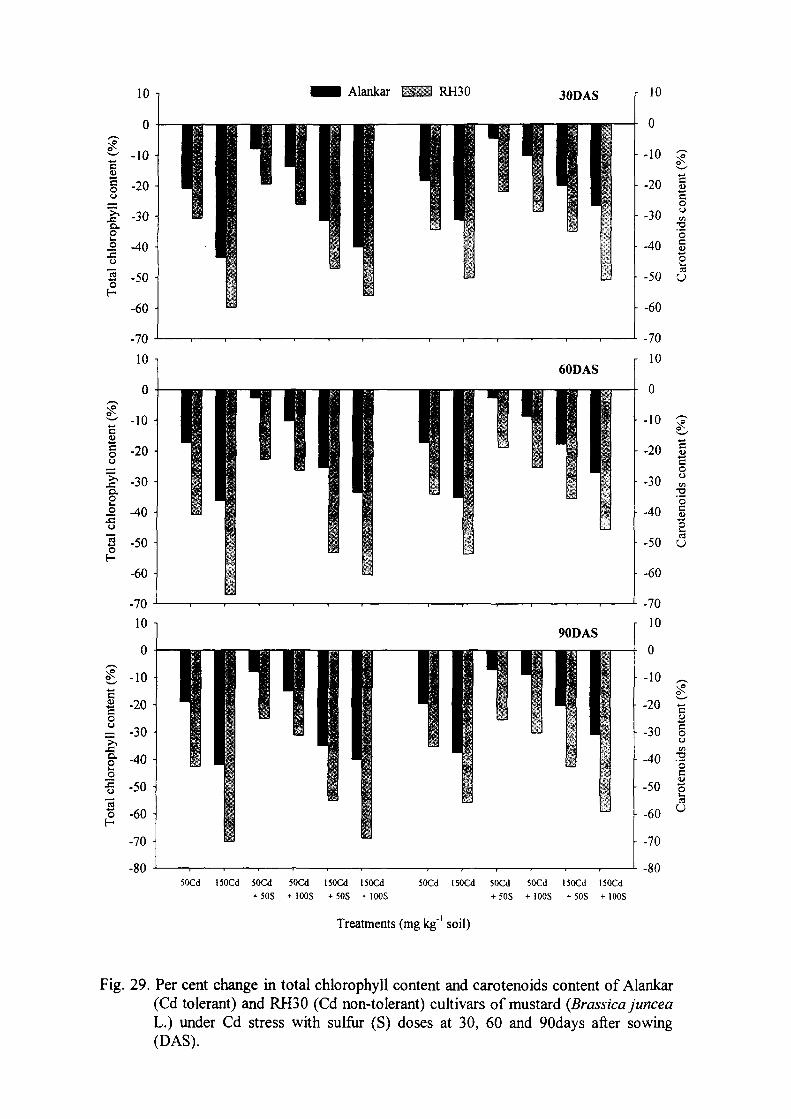
Alankar ^ O RH30
50Cd ISOCd 50Cd
+ 50S
50Cd
+ lOOS
150Cd
+ 50S
150Cd
+ lOOS
50Cd ISOCd 50Cd
+ 50S
50Cd
+ lOOS
150Cd
+ 50S
ISOCd
+ lOOS
Treatments (mg kg"' soil)
Fig. 29. Per cent change in total chlorophyll content and carotenoids content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard {Brassica juncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

-80
10
0
-10 ^
-20
.^ -30 -> 1 -40 <
U -50
-60
-70
-80 50Cd 150Cd 50Cd + 50S
50Cd + lOOS
150Cd + 50S
150Cd + lOOS
90DAS
Treatments (mg kg'' soil)
Fig. 30. Per cent change in carbonic anhydrase activity of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress with sulflir (S) doses at 30, 60 and 90days after sowing (DAS).

10 1
« -30
S -40
Alankar ^ ^ RH30 30DAS
-70
10
0
-10 •
-20
-50 -
-60
-70
1 r-
60DAS
-10
-20
-30
• -40
-50
50Cd rSOCd 50Cd
+ 50S
50Cd l50Cd
+ lOOS + SOS
ISOCd
+ lOOS
50Cd ISOCd SOCd
+ 50S
50Cd ISOCd ISOCd
+ lOOS + SOS + lOOS
-60
10
0
-10
-20
-30
-40
-50
a C8
E o
-60
-70
-70
Treatments (mg kg' soil)
Fig. 31. Per cent change in net photosynthetic rate and stomatal conductance of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard {Brassica juncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

stomatal conductance caused by 50mg Cd kg"' soil alone. Finally, the decrease noted
in total chlorophyll and carotenoids content and stomatal conductance was 7.9, 2.7
and 8.0% at 30DAS; 4.6, 2.7 and 7.1% at 60DAS and 3.3, 0.7 and 5.3% at 90DAS,
respectively. Similar effect was noted for carbonic anhydrase activity and net
photosynthetic rate, which was decreased by 1.7 and 0.6% at 30DAS and 1.5 and
0.4% at 90DAS, respectively. At 60DAS, the application of 50mg S kg'' soil not only
reduced the adverse effects of Cd (50mg Cd kg'' soil) but overcome the effect and
increased the carbonic anhydrase activity and net photosynthetic rate. The increase
was found to be 0.3 and 1.5% at 60DAS (Figures 29-31).
In RH30, the application of 50mg S kg'' soil only reduced the adverse effects
of 50mg Cd kg'' soil at all the growth stages. The decrease in total chlorophyll
content, carotenoids content, carbonic anhydrase activity, net photosynthetic rate and
stomatal conductance was 19.4, 21.8, 20.0, 14.6, 15.0% at 30DAS; 22.6, 18.8, 21.5,
13.6, 16.2% at 60DAS and 25.0, 25.4, 24.6, 15.9, 20.7% at 90DAS, respectively, over
their respective control (Figures 26-31). Whereas, no ameliorative effect of lOOmg S
kg'' soil was observed in plants treated with 150mg Cd kg'' soil in both the cultivars
(Figures 29-31).
4.3.3 Sulfur and nitrogen assimilation
Sulfur and nitrogen assimilation varied differentially under Cd stress. Cadmium stress
increased the ATP-sulfurylase activity and S content, whereas, nitrate reductase
activity and N content were decreased in both cultivars at all growth stages (Figures
32-33). ATP-sulfurylase activity and S content increased in Cd treated plants and the
increases were higher in Alankar than RH30. In Alankar, the increase in ATP-
sulfurylase activity and S content due to 50 and 150mg Cd kg'' soil was 39.2, 44.5%
and 21.9, 25.5% at 30DAS; 35.4, 45.5% and 21.3, 24.7% at 60DAS and 27.3, 36.1%
and 15.0, 20.0% at 90DAS, respectively, in comparison to their respective control.
This increase in RH30 was 22.6, 30.3% and 7.4, 12.0% at 30DAS; 23.4, 33.8% and
10.4, 13.3% at 60DAS and 20.0, 25.5% and 5.0, 10.1% at 90DAS, respectively, in
comparison to their respective control (Figure 34).
The application of 50mg S kg'' soil further increased the ATP-sulfurylase
activity and S content of Cd treated plants of both the cultivars. Maximum significant
increase was noted with 50mg S kg'' soil and 50mg Cd kg"' soil treated plants. The
effect of lOOmg S kg'' soil plus 50mg Cd kg'' soil was almost similar to 50mg S kg''
71
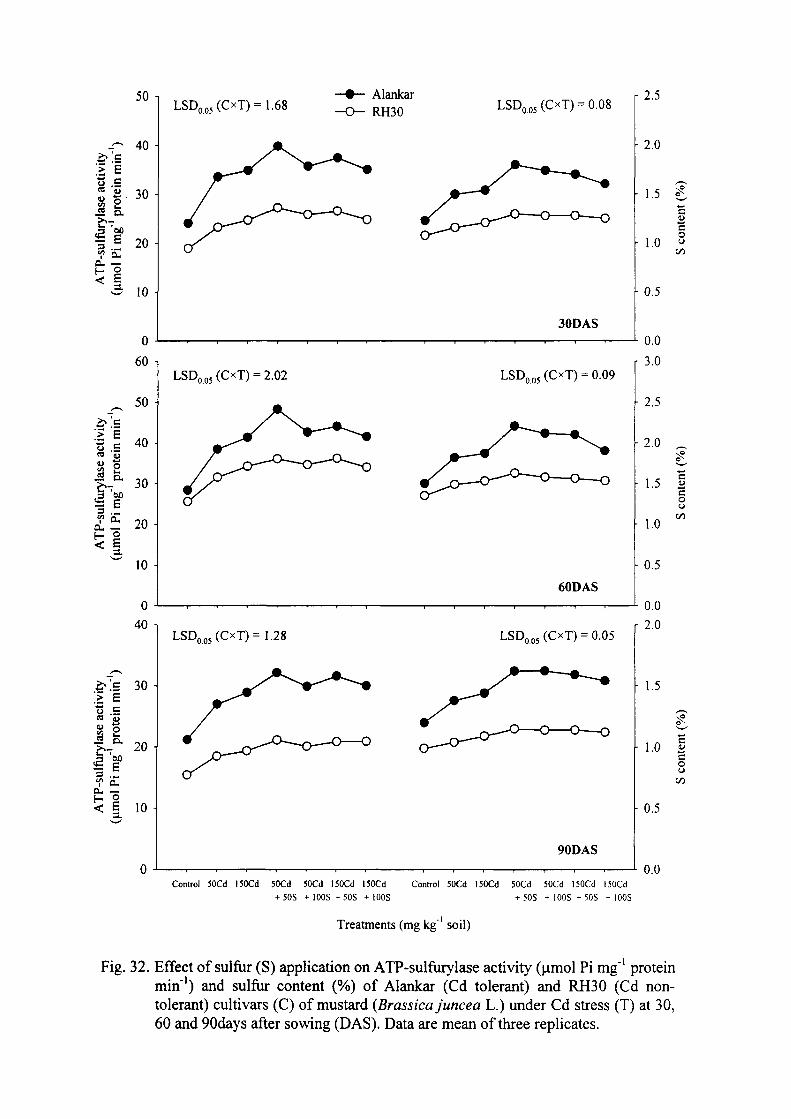
50 1
40
B. 30
20
10 -
0
60
50
40 -
30 -
20
10
0
40
Z^'.S 30
20 •
10 -
LSDoo5(CxT)=1.68 -•— Alankar
- O - RH30 LSD„o5(CxT) = 0.08
30DAS T " 1 I
LSDooj (CxT) = 2.02 LSDoo5(CxT) = 0.09
60DAS
•7 ' I -
LSD„,o5 (CxT) =1.28 LSDoo5 (CxT) = 0.05
90DAS
Control 50Cd l50Cd 50Cd 50Cd 150Cd l50Cd
+ SOS + lOOS + SOS + lOOS
Control SOCd ISOCd SOCd SOCd ISOCd ISOCd
+ SOS + lOOS +503 - lOOS
2.5
2.0
- 1.5 S
c o
1.0 " (/2
• 0.5
0.0
r 3,0
2.5
20 _
• 1.5 S c o
• 1.0
0.5
0.0
r 2.0
1.5
1.0
0.5
0.0
Treatments (mg kg" soil)
Fig. 32. Effect of sulfur (S) application on ATP-suifurylase activity (|imol Pi mg"' protein min'') and sulfur content (%) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

500
400
300
Z 200 -
100 •
600 1
500
400
300 -
200 -
100 -
0
400
300
200
100
LSDoo3 (CxT) = 7.25 -•— Alankar
-O- RH30 LSDoo5 (CxT) = 0.09 r 5
30DAS ' I - T
LSDooj (CxT) =10.55 LSDo,o5 (CxT) = 0.07
60DAS • • f ^ ~ - ' 1 • • , • • •—• . . , . 11
LSD„„5 (CxT) = 8.16 LSDooj (CxT) = 0.08
9GDAS
3 e c 1) B O
2 o
0
r 6
3 I c o o
[2 ^
• 1
0
5
• 4
• 3 c o c o o
Control 50Cd 150Cd 50Cd 50Cd ISOCd 150Cd
+ SOS + lOOS + 50S + lOOS Control 50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ 50S + lOOS +50S + lOOS
Treatments (mg kg"' soil)
Fig. 33. Effect of sulfur (S) application on nitrate reductase activity (nmol NO2 g"' FW h'') and nitrogen content (%) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are the mean of three replicates.

Alankar P ? ^ RH30
U 50Cd 150Cd 50Cd 50Cd
+ 50S
150Cd 150Cd
lOOS + SOS + lOOS 50Cd 150Cd 50Cd 50Cd ISOCd ISOCd
+ SOS + lOOS + SOS + IOCS
Treatments(mgkg' soil)
Fig. 34. Per cent change in ATP-sulfuryiase activity and sulfur content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard {Brassica juncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

soil plus 50mg Cd kg'' soil. Further, no ameliorative effect of lOOmg S kg'' soil was
observed in plants treated with 150mg Cd kg'' soil.
Cadmium treatments significantly decreased NR activity and N content of
both the cultivars but the decrease was greater in RH30 than Alankar. The activity of
NR and N content of Alankar was decreased by 14.6, 11.0% and 27.2, 25.4% at
30DAS; 15.5, 10.6% and 29.9, 25.6% at 60DAS and 21.0, 17.2% and 34.4, 30.4% at
90DAS with 50 and 150mg Cd kg''soil, respectively, in comparison to their respective
control. Whereas, in RH30 the decrease in NR activity and N content was 26.4,19.4%
and 48.1, 38.6% at 30DAS; 34.7, 22.5% and 50.0, 39.4% at 60DAS and 37.7, 27.7%
and 58.6, 45.7% at 90DAS with 50 and 150mg Cd kg''soil, respectively, in
comparison to their respective control (Figure 35).
The reductions in NR activity and N content due to Cd were lowered by S
supplementation. Lesser decrease in NR activity and N content was observed when
50mg Cd kg"' soil treated plants were given 50mg S kg'' soil in comparison to 50mg
Cd kg'' soil alone, hi Alankar, the decrease in NR activity and N content due to 50mg
Cd kg'' soil was limited to 1.8 and 1.7% at 30DAS; 1.1 and 1.4% at 60DAS and 8.8
and 10.1% at 90DAS with 50mg S kg"' soil. In RH30, these decreases were restricted
to 15.1 and 7.9% at 30DAS, 18.6 and 13.2% at 60DAS and 20.9 and 16.5% at 90DAS
with 50mg S kg"' soil (Figure 35).
4.3.4 Cadmium accumulation
Cadmivmi content in the root and leaf of both the cultivars increased with the
increasing Cd concentration in soil. Non-tolerant cultivar (RH30) accumulated more
Cd in root and leaf than tolerant cultivar Alankar (Figure 36). In comparison to root,
the content of Cd was less in the leaf of both the cultivars due to S supplementation.
The application of 50mg S kg"' soil maximally lowered the Cd content in the root and
leaf of both the cultivars treated with 50mg Cd kg"' soil. The application of lOOmg S
kg"' soil also lowered the Cd content in root and leaf of 50mg Cd kg"' soil treated
plants but the values obtained were almost similar to that of 50mg S kg"' soil plus
50mg Cd kg"' soil. Application of lOOmg S kg"' soil did not decrease the Cd content
in root and leaf of 150mg Cd kg"' soil treated plants.
Cadmium content in the root and leaf of Alankar was 110.8, 27.7 and 274.3,
70.0^g g"' DW at 30DAS; 141.3, 40.7 and 331.5, 95.8^g g"' DW at 60DAS and
184.4, 24.5 and 380.5, SOA^g g'' DW at 90DAS due to 50 and 150mg Cd kg'' soil,
respectively. The addition of 50mg S kg'' soil maximally lowered the Cd content in
72

Alankar felMM RH30 30DAS
a: 2
-20
-40
-60
10
0
-10
^ -20
:> - 30 -» o
oi -40
-50
-60
-70
T T -
-1 1 f • -•} r -
60DAS
- 1 p— I • I
90DAS
10
0
-10
F -20 r c u
-30 § Z
-40
-50
-60
10
• - 20
-10
-20 r c u
-30 § Z
-40
-50
-60
0
C
c o o
-40 2
-60
50Cd 150Cd 50Cd 50Cd 150Cd ISOCd
+ SOS + lOOS + SOS + lOOS
SOCd ISOCd 50Cd SOCd ISOCd ISOCd
+ SOS + lOOS +50S ^ IOCS
Treatments (mg kg" soil)
Fig. 35. Per cent change in nitrate reductase activity and nitrogen content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

400
300
200 •
100 •
LSD„„,(CxT)= 10.66
Alankar RH-30
LSD„„5(CxT) = 3.68
0
600
500
400
300
200
100
0
700
600
500
400 -I
Z. op 300 o =1.
200
100
LSDoo5(CxT)= 14.03 LSD„„5(CxT) = 5.40
60DAS
LSDo 05 (CxT)= 16.95 LSDo_o5(CxT) = 3.48
200
150
100
50
0
250
200
50
00
c o u
U
^ Q
'ofi
a. -
50
0
200
150
c a ^ 8 Q
50
Control 50Cd 150Cd 50Cd 50Cd 150Cd 150Cd Control 50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ SOS + lOOS + SOS + lOOS + SOS + IOCS + SOS + lOOS
Treatments (mg kg' soil)
Fig. 36. Effect of sulfur (S) application on root Cd content and leaf Cd content (|ig g"' DW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

root and leaf of plants treated with 50mg Cd kg'' soil and the values were 85.5 and
18.8^g g"' DW at SODAS; 106.7 and 25.4^g g"' DW at 60DAS and 154.6 and 18.3^g
g"' DW at 90DAS, respectively (Figure 37).
In RH30, Cd content in the root and leaf was 135.8, 63.3 and 357.5, 138.1^g
g"' DW at 30DAS; 190.7, 92.6 and 488.3, 206.2^g g"' DW at 60DAS and 286.1, 62.4
and 556.4, 132.3^g g'' DW at 90DAS due to 50 and 150mg Cd kg"' soil, respectively.
The combined application of 50mg S kg'' soil and 50mg Cd kg'' soil lowered the Cd
content in root and leaf and the values were 115.3 and 49.8|ig g'' DW at 30DAS;
161.6 and 72.1^g g'' DW at 60DAS and 260.2 and 53.3|ag g"' DW at 90DAS,
respectively (Figure 37).
4.3.5 Thiobarbituric acid reactive substances and hydrogen peroxide content
Significant Cd-dose dependent increase in TBARS and H2O2 content was observed in
both cultivars and the extent of increase was found greater in RH30 than Alankar.
Maximum increase in TBARS and H2O2 content was noted with 150mg Cd kg'' soil
in both the cultivars at all growth stages. Application of 50mg S kg'' soil maximally
lowered the TBARS and H2O2 content of 50mg Cd kg"' soil treated plants at all
growth stages. The values obtained for TBARS and H2O2 content with lOOmg S kg''
soil plus 50mg Cd kg'' soil were almost similar to the values obtained in plants treated
with 50mg S kg'' soil plus 50mg Cd kg'' soil. Application of lOOmg S kg'' soil did
not lower the TBARS and H2O2 content of 150mg Cd kg'' soil treated plants (Figure
38).
In Alankar, the increase in TBARS and H2O2 content was 146.3, 317.6% and
74.8, 142.6% at 30DAS; 168.1, 324.4% and 85.1, 145.2% at 60DAS and 173.3,
363.7% and 86.4, 150.5% at 90DAS due to 50 and 150mg Cd kg'' soil, respectively,
over their respective control. Supplementation of 50mg S kg'' soil to plants treated
with 50mg Cd kg"' soil significantly lowered the TBARS and H2O2 content by 41.4
and 28.4% at 30DAS; 34.9 and 25.9% at 60DAS and 62.4 and 33.1% at 90DAS,
respectively, in Alankar (Figure 39).
The content of TBARS and H2O2 in RH30 was increased by 224.8, 412.4%
and 144.2, 197.7% at 30DAS; 339.4, 427.8% and 156.0, 206.5% at 60DAS and 403.5,
445.1% and 161.5, 228.0% at 90DAS due to 50 and 150mg Cd kg"' soil, respectively,
over their respective control. Whereas, supplementation with 50mg S kg' soil
lowered the TBARS and H2O2 content by 162.9 and 99.7% at 30DAS; 180.2 and
73

7000 Alankar ^ ^ RH30 30DAS r 5000
4->
c o u •a U o o
5000
4000
3000
2000
0 8000
«3^
o u T3 U o o ai
c <D * - » B O
^ 4000 o o
50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ 50S + lOOS +50S +100S
50Cd l50Cd 50Cd 50Cd ISOCd ISOCd
+ 50S + lOOS +50S + lOOS
Treatments (mg kg' soil)
Fig. 37. Per cent change in root Cd content and leaf Cd content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard {Brassica juncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

100 1
80 •
60 -
40 -
20 -
LSDo„5(CxT)=1.85 Alankar RH30
LSDoo5(CxT) = 7.97
0
100
80
60 ^
40
20
0
100
80 •
60
40 -
20
30DAS
LSDQOS (CXT) = 2.08 LSD„„,(CxT) = 9.56
60DAS I I
LSD„„, (CxT) = 3.49 LSDoo5 (CxT) =10.23
90DAS
r 300
250
200 „ ^
150 2 '60
100
50
0
400
300
c _ 200 S 'ej)
100
0
400
300
200 o _M
- 100
Control 50Cd 150Cd 50Cd 50Cd 150Cd 150Cd Control 50Cd I50Cd 50Cd 50Cd 150Cd ISOCd
+ 50S +100S+50S + lOOS + SOS + lOOS + SOS + lOOS
Treatments (mg kg' soil)
Fig. 38. Effect of sulfur (S) application on thiobarbituric acid reactive substances (nmol g"' FW) and hydrogen peroxide content (^mol g"' FW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

50Cd 150Cd 50Cd
+ 50S
50Cd 150Cd 150Cd
+ lOOS + SOS + lOOS 50Cd ISOCd 50Cd
+ 50S
50Cd ISOCd ISOCd
+ lOOS +50S + lOOS
Treatments (mg kg" soil)
Fig. 39. Per cent change in thiobarbituric acid reactive substances and hydrogen peroxide content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica j'uncea L.) under Cd stress with sulftir (S) doses at 30, 60 and 90days after sowing (DAS).

100.9% at 60DAS and 250.3 and 112.5% at 90DAS, respectively, in plants treated
with 50mg Cd kg"' soil (Figure 39).
4.3.6 Components of antioxidant system
4.3.6.1 Enzymatic antioxidants
The response of antioxidant enzymes was differential to Cd stress in both the
cultivars. In general, S application (50mg S kg'' soil) maximally increased the
activities of catalase (CAT), ascorbate peroxidase (APX) and glutathione reductase
(GR) under Cd stress (50mg Cd kg'' soil). Whereas, the application of 50mg S kg"'
soil lowered the increase in SOD activity caused by 50mg Cd kg'' soil alone (Figures
40-41).
The activity of SOD increased with increasing Cd concentration in soil and
extent of increase was greater in RH30 than Alankar. Sulfur supplementation lowered
the Cd induced increase in SOD activity of both the cultivars. In Alankar, significant
increase in SOD activity was 51.3, 71.6% at 30DAS; 43.4, 57.2% at 60DAS and 42.7,
33.9% at 90DAS due to 50 and 150mg Cd kg'' soil, respectively, over the control.
However, the activity of SOD was reduced to 24.5, 27.5 and 26.8% at 30, 60 and
90DAS, respectively, when the plants were given 50mg S kg'' soil in the presence of
50mg Cd kg"' soil (Figure 42).
The activity of SOD was increased by 71.4, 116.1% at 30DAS; 76.4, 100.0%
at 60DAS and 50.3, 69.2% at 90DAS in RH30 due to 50 and 150mg Cd kg"' soil,
respectively, over the control. Its activity was restricted to 40.3, 33.8 and 28.5% at 30,
60 and 90DAS, respectively, with the addition of 50mg S kg"' soil to 50mg Cd kg"'
soil-treated RH30 plants (Figure 42).
The activity of CAT was increased with the increasing Cd concentration in the
soil at 30, 60 and 90DAS in Alankar. Whereas, in RH30, the increase in CAT activity
was noted only at 30DAS. At later stages (60 and 90DAS), Cd significantly decreased
the CAT activity. Sulfur supplementation further increased the Cd induced increase in
CAT activity at all growth stages in tolerant cultivar Alankar. In RH30, addition of S
lowered the decrease in CAT activity only at 60 and 90DAS, whereas, slight increase
in its activity was noted at 30DAS.
Significant increase in CAT activity of Alankar due to 50 and 150mg Cd kg"'
soil was 33.3, 56.4% at 30DAS; 24.1, 42.1% at 60DAS and 23.6, 28.3% at 90DAS,
respectively, in comparison to their respective control. Addition of 50mg S kg"' soil
74

significantly increased the CAT activity by 40.2, 48.3 and 30.2% at 30, 60 and
90DAS, respectively; in Alankar treated with 50mg Cd kg"' soil (Figure 42).
In RH30, the increase in CAT activity was 10.7 and 13.9% at 30DAS,
whereas, its activity was decreased by 20.4, 36.7% at 60DAS and 18.1, 35.3% at
90DAS due to 50 and 150mg Cd kg'' soil, respectively. S supplementation (50mg S
kg'' soil) increased the CAT activity by 16.1% at 30DAS, while, at 60 and 90DAS its
application only lowered the decrease in CAT activity by 9.2 and 10.6%, respectively,
in plants treated with 50mg Cd kg'' soil (Figure 42).
Supplementation of 50mg S kg'' soil significantly and maximally increased
the APX activity of both the cultivars treated with 50mg Cd kg'" soil at all growth
stages.
The increase in APX activity of Alankar due to 50 and 150mg Cd kg' soil was
70.4, 128.4% at 30DAS; 158.4, 193.1% at 60DAS and 109.7, 144.7% at 90DAS,
respectively, over control. Further, the addition of 50mg S kg'' soil caused maximal
significant increase of 200.7, 231.8 and 175.0% in APX activity at 30, 60 and 90DAS,
respectively, of plants treated with 50mg Cd kg'' soil (Figure 43). However, in RH30
lesser increase was observed in plants treated with 50mg Cd kg"' soil or these plants
supplemented with 50mg S kg'' soil (Figure 43).
Glutathione reductase activity of both the cultivars was increased under Cd
stress. Maximal significant increase in GR activity was noted in both the cultivars
treated with 50mg Cd kg"' soil. Treatment of 150mg Cd kg"' soil significantly
decreased the GR activity in non-tolerant cultivar RH30. Addition of S (50mg S kg"'
soil) further increased the GR activity of both the cultivars at all growth stages and
extent of increase was higher in Alankar than RH30.
The increase in GR activity of Alankar due to 50 and 150mg Cd kg"' soil was
61.4, 40.6% at 30DAS; 55.3, 38.3% at 60DAS and 39.5, 26.3% at 90DAS,
respectively, over the control. Treating these plants with 50mg S kg"' soil further
increased the activity, which was maximum compared to other S treatments. Similar
response of RH30 was noted with respect to Cd and S treatments (Figure 43).
4.3.6.2 Non-enzymatic antioxidants
Cadmium treatments significantly decreased the ascorbic acid content at all growth
stages and greater decrease was found in RH30 than Alankar. Supplementation of Cd-
treated plants with S lowered the Cd-induced reduction in ascorbic acid content of
both the cultivars at all growth stages and 50mg S kg"' soil maximally lowered the
75

18 1
15
•S 12-
6 -
3 •
0
21
18 •
15 -
12
9 -
3 •
0
15
12
9 -
6 -
LSDooj (CxT) = 0.24 Alankar RH30
LSDoos (CxT) =1.44
30DAS
LSDoo5 (CxT) = 0.15 LSDoo5 (CxT) =1.32
60DAS
LSD005 (CxT) = 0.03 LSD005 (CxT) =1.09
90DAS
250
• 200
150
100
• 50
0
300
I- 250
200
1- 150
100
(- 50
0
200
150
100
50
Control 50Cd 150Cd 50Cd 50Cd 150Cd I50Cd Control 50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ 50S +100S+50S + lOOS + SOS + lOOS + SOS + lOOS
Treatments (mg kg" soil)
Fig. 40. Effect of sulfur (S) application on superoxide dismutase activity (Units mg"' protein) and catalase activity (Units mg"' protein) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

2.5
2.0 -
o -
0- a 1.0
0.5
/—V
.s .^^ > t.
^ - » * - * •
w so X E O N ^
< •? D N — ^
-—N
c
> t . * - » *- o — « 'OO X E tt^ « < c
3
0.0
4.0
3.5
3.0
2.5
2.0
1.5
1.0
0.5
0.0 4.0
3.5
3.0
2.5
2.0
1.5
1.0
0.5
0.0
LSDooj (CxT) = 0.008 -•— Alankar - O - RH30
LSD„o5 (CxT) = 0.008
30DAS
LSD„„5 (CxT) = 0.092 LSD„„, (CxT) = 0.004
60DAS
" I I I
LSD, LSD„„5 (CxT) = 0.004
90DAS
0.5
- 0.4
0.3 2:> £
E 0.2 Q S
c
0.1
0.0
0.6
0.5
• 0.4
0.3
• 0.2
£-> t j
a: a
'33 o D.
'nn f t /1
-t—t
c
0.1
0.0 r 0.5
0.4
0.3 ^ e
0-2 o .-s
T •" I
- 0.1
0.0 Control 50Cd 150Cd 50Cd 50Cd ISOCd ISOCd Control 50Cd 150Cd 50Cd 50Cd l50Cd ISOCd
+ 50S +100S+50S + lOOS + SOS +100S+50S + lOOS
Treatments (mg kg"' soil)
Fig. 41. Effect of sulfur (S) application on ascorbate peroxidase activity (Units mg"' protein) and glutathione reductase activity (Units mg'' protein) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

Alankar C2E3 RH30 30DAS
m
- 70
' 60
50
40
• 30
• 20
10
. n
> t> ta (-< U
50Cd 150Cd 50Cd
+ 50S
50Cd
+ IOOS
!50Cd
+ 50S
I50Cd
+ lOOS
50Cd l50Cd 50Cd
+ 50S
50Cd
+ lOOS
ISOCd
+ 50S
150Cd
+ lOOS
Treatments (mg kg"' soil)
Fig. 42. Per cent change in superoxide dismutases activity and catalase activity of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica j'uncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ 50S + lOOS +50S +100S
50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ 50S + lOOS +50S + lOOS
Treatments (mg kg" soil)
Fig. 43. Per cent change in ascorbate peroxidase activity and glutathione reductase activity of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard {Brassica juncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

ascorbic acid content. Treatment of lOOmg S kg"' soil to 50mg Cd kg'' soil-treated
plants also lowered the reductions but was less effective compared to 50mg S kg"
soil. Application of lOOmg S kg"' soil did not improve the ascorbic acid content of
150mg Cd kg"' soil treated plants (Figure 44)
Ascorbic acid content of Alankar was decreased by 27.9, 44.2% at 30DAS;
24.2, 49.1% at 60DAS and 27.2, 46.0% at 90DAS with 50 and 150mg Cd kg"'soil,
respectively, over the control. Addition of S (50mg S kg"' soil) maximally lowered the
reduction in ascorbic acid content of 50mg Cd kg"' soil treated plants by 11.9, 5.3 and
10.6% at 30, 60 and 90DAS, respectively, over the control (Figure 45).
In RH30, the decrease in ascorbic acid content was 44.6, 61.2% at 30DAS;
50.0, 64.0% at 60DAS and 57.5, 68.7% at 90DAS with 50 and 150mg Cd kg"'soil,
respectively, over the control. Supplementation of 50mg S kg"' soil maximally
lowered the reduction in ascorbic acid content of 50mg Cd kg"' soil treated plants by
27.3,25.8 and 30.3% at 30, 60 and 90DAS, respectively, over the control (Figure 45).
Glutathione content increased in Cd-treated both the cultivars. The increase
was higher in Alankar than RH30. Sulfur supplementation further increased the
glutathione content of both the cultivars and maximum significant increase in its
content was noted in plants treated with 50mg S kg"' soil plus 50mg Cd kg"' soil at all
growth stages.
The increase in glutathione content of Alankar due to 50 and 150mg Cd kg"'
soil was 19.7, 24.4% at 30DAS; 25.3, 36.0% at 60DAS and 25.3, 31.9% at 90DAS,
respectively, over the control. Further maximal significant increase in its content
occurred with the addition of 50mg S kg"' soil to 50mg Cd kg"' soil treated plants.
RH30 also responded similarly to Cd and S treatments (Figure 45).
4.3.7 Yield characteristics
Cadmium treatments significantly decreased the yield characteristics (expressed as
number of siliqua per plant, number of seeds per siliqua, 1000 seed weight and seed
yield per plant) at harvest and the extent of decrease was greater in RH30 than
Alankar. Sulfiir supplementation (50mg S kg"' soil) lowered the Cd (50mg Cd kg"'
soil)-induced reduction in yield characteristics of both the cultivars (Figure 46). The
application of lOOmg S kg"' soil also lowered the reductions in yield characteristics
caused by 50mg Cd kg"' soil but was less effective than 50mg S kg"' soil. Further, the
application of lOOmg S kg"' soil did not alleviate the reduction in yield characteristics
caused by 150mg Cd kg"' soil.
76

400 1
300 •
200 •
100
0
500
400 •
300 •
200
100
0
500
400
300
200
100
LSDo,o5(CxT) = 2.81 Alankar
RH30 LSDoo, (CxT) = 5.76
30DAS I I
LSDoo5 (CxT) = 2.25 LSDQOS ( C X T ) = 2.60
1000
800
600
400 g I
200
0
800
600
• 400
• 200
60DAS
LSDoo3 (CxT) = 2.18 LSDoo, (CxT) =1.15
90DAS
0
600
400
• 200
Control 50Cd 150Cd 50Cd 50Cd 150Cd 150Cd
+ 505 + lOOS + 505 + 1005
Control 50Cd 150Cd 50Cd 50Cd 150Cd l50Cd
+ 505 + 1005 +505 +1005
Treatments (mg kg" soil)
Fig. 44. Effect of sulfur (S) application on ascorbic acid content (nmol g"' FW) and glutathione content (nmol g"' FW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

-70
10
0
-10
-20
-30
-40
-50
-60
It l l lMII 1 -
in i i 1 ii ! i
w
60DAS
50Cd 150Cd 50Cd
+ 50S
50Cd
+ lOOS
150Cd
+ 50S
150Cd
+ lOOS
50Cd ISOCd 50Cd
+ 50S
50Cd
+ lOOS
130Cd
+ 50S
ISOCd
+ lOOS
Treatments (mg kg' soil)
Fig. 45. Per cent change in ascorbic acid content and glutathione content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress with sulfur (S) doses at 30, 60 and 90days after sowing (DAS).

100
c (8 O. u u O.
3 O"
crt t«-o k-
<u £l
B 3
z
8U
60
40
20
0 8
OD
.SP '5 ? <u
o o o
LSDoo3(CxT)=1.29
Alankar RH30
LSDQOS (CXT) = 6.69
I I I " " I
LSDoo5 (CxT) = 0.45 LSDoo5 (CxT) = 0.52
r 20
15§-
o.
i o |
5 3 Z
• 6
• 4 -^
Control 50Cd 150Cd 50Cd 50Cd 150Cd ISOCd
+ SOS + lOOS + SOS + IOCS
Control SOCd ISOCd SOCd SOCd ISOCd ISOCd
+ 50S + lOOS +SOS + lOOS
Treatments (mg kg"' soil)
2 u
(U aj
C/D
Fig. 46. Effect of sulfur (S) application on number of siliqua per plant, number of seeds per siliqua, 1000 seed weight (g) and seed yield (g plant'') of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at harvest i.e., 120days after sowing (DAS). Data are mean of three replicates.

c as
a. (8 3
X)
E 3
eo
u
u
o o o
Control 50Cd ISOCd 50Cd 50Cd 150Cd I50Cd Control 50Cd 150Cd 50Cd 50Cd ISOCd 150Cd
+ 50S +1005+505 +1005 +505 +1005+505 +1005
Treatments (mg kg' soil)
Fig. 47. Per cent change in number of siliqua per plant, number of seeds per siliqua, 1000 seed weight and seed yield of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress with sulfur (S) doses at harvest i.e., 120days after sowing (DAS).

Number of siliqua per plant, number of seeds per siliqua, \1
and seed yield per plant of Alankar due to 50 and 150mg Cd M'*5pil was decreased
by 22.7, 22.8, 26.2, 21.6% and 35.7, 33.8, 42.4, 32.5%, respecti>^^ver the control.
Application of 50mg S kg' soil maximally lowered the 50mg Cd kg'^oil-induced
reductions in these characteristics, which were 8.3, 13.3, 12.7 and 10.8%,
respectively, over the control. RH30 also responded similarly to Alankar for Cd and S
treatments but the cultivar was less responsive (Figure 47).
4.4 Summary of Experiment 2
• Application of 50mg S kg'' soil proved most effective in alleviating the effects
of 50mg Cd kg'' soil on growth, photosynthetic, biochemical and yield
characteristics of Alankar and RH30 cultivars.
• Cd treatments up-regulated the S assimilation pathway and increased ATP-
sulfurylase activity and S content. Whereas, N assimilation pathway was
down-regulated and decrease in nitrate reductase activity and N content were
observed. Sulfur supplementation (50mg S kg'' soil) to plants treated with
50mg Cd kg'' soil further increased the ATP-sulflirylase activity and S content
and also lowered the decrease in nitrate reductase activity and N content.
• Application of 50mg S kg'' soil decreased the Cd content maximally in root
and leaf of both the cultivars treated with 50mg Cd kg'' soil.
• Cd-induced accimiulation of TBARS and H2O2 was lowered by the application
of50mgSkg''soil.
• Responses of antioxidant enzyme activities were found differential in both the
cultivars under Cd stress. S application (50mg S kg'' soil) increased the
activities of CAT, APX and GR of 50mg Cd kg'' soil treated plants.
• Application of 50mg S kg'' soil maximally lowered the 50mg Cd kg'' soil-
induced decrease in ascorbic acid content and increased the glutathione
content in both the cultivars.
• Application of 50mg S kg'' soil also lowered the Cd-induced decrease in yield
characteristics of both the cultivars.
• The application of 50mg S kg'' soil was found most effective in mitigating the
50mg Cd kg'' soil induced reductions in growth, photosynthetic, biochemical
and yield characteristics in Alankar, whereas, reductions in these
characteristics were only lowered in RH30.
77 ^V^v

4.5 Experiment 3: Coordination of Sulfur and Nitrogen Application in the
Alleviation of Cd-induced Effects on Cd tolerant and non-tolerant Mustard
Cultivars
Experiment 3 was conducted on the basis of the findings of Experiments 2. The
present Experiment was aimed to study the effectiveness of addition of N levels (40,
80 and 120mg N kg"' soil) to the alleviation potential of 50mg S kg'' soil on 50mg Cd
kg'' soil-induced reductions in Alankar (Cd tolerant) and RH30 (Cd non-tolerant)
cultivars.
Results of this Experiment are presented in the following section.
4.5.1 Growth characteristics
Application of different N levels (40, 80 and 120mg N kg'' soil) further enhanced the
alleviation effect of 50mg S kg'' soil in Alankar and RH30 cultivars treated with
50mg Cd kg'' soil. Among the N levels used, the addition of 80mg N kg'' soil to
50mg S kg"' soil nulhfied the adverse effects of 50mg Cd kg'' soil and also maximally
increased the growth characteristics of Alankar. Other N levels proved less effective
(Figures 48-49).
The treatment 80mg N kg'' soil plus 50mg S kg"' soil resulted in maximal
increase in shoot length, root length, leaf area and plant dry mass by 6.5, 5.7, 5.6,
4.1% at 30DAS; 7.3, 6.9, 5.9, 5.3% at 60DAS and 5.2, 5.6, 4.8, 4.8% at 90DAS of
50mg Cd kg"' soil treated Alankar plants, respectively, over their respective control
(Figures 50-51).
Similar effect of the treatment (80mg N kg"' soil plus 50mg S kg'' soil) was
observed in RH30. Cd-induced reductions in growth characteristics were lowered by
all N levels applied in combination with 50mg S kg'' soil. However, application of
80mg N kg'' soil with 50mg S kg'' soil maximally lowered the reductions in shoot
length, root length, leaf area and plant dry mass and these were limited to 8.4, 8.4, 8.2,
8.5% at 30DAS; 6.6, 6.4, 8.6, 6.4% at 60DAS and 7.0, 7.7, 9.7, 6.1% at 90DAS of
50mg Cd kg"' soil treated plants, respectively, over their respective control (Figures
50-51).
4.5.2 Photosynthetic characteristics
The reductions caused by 50mg Cd kg'' soil in photosynthetic characteristics (total
chlorophyll content, carotenoids content, carbonic anhydrase activity, net
photosynthetic rate and stomatal conductance) were reversed by the application of
50mg S kg' soil at all the growth stages in both the cultivars. Ameliorative effect of
78

20
1 5 •
5 7 60 g
l l 10
LSD„„5(CxT) = 0.08
0
25
20 -
15 -
10
0
25
20
15
10
5 ]
Alankar RH30
LSD„o,(CxT) = 0.57
30DAS
LSDo(,5(CxT) = 0.12 LSDo,o5(CxT) = 0.72
60DAS
LSDo,„3(CxT) = 0.17 LSDoo5(CxT) = 0.44
« • •
90DAS
60
50
40
30 - -5.
20
10
0
120
100
80
60
40
20
0
140
120
100
I- 80
60
40
20
Control 50Cd OCd +50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+120N
Control 50Cd 50Cd + 50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ I20N
Treatments (mg kg' soil)
Fig. 48. Effect of sulfur (S) and nitrogen (N) application on root and shoot length (cm plant'') of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.
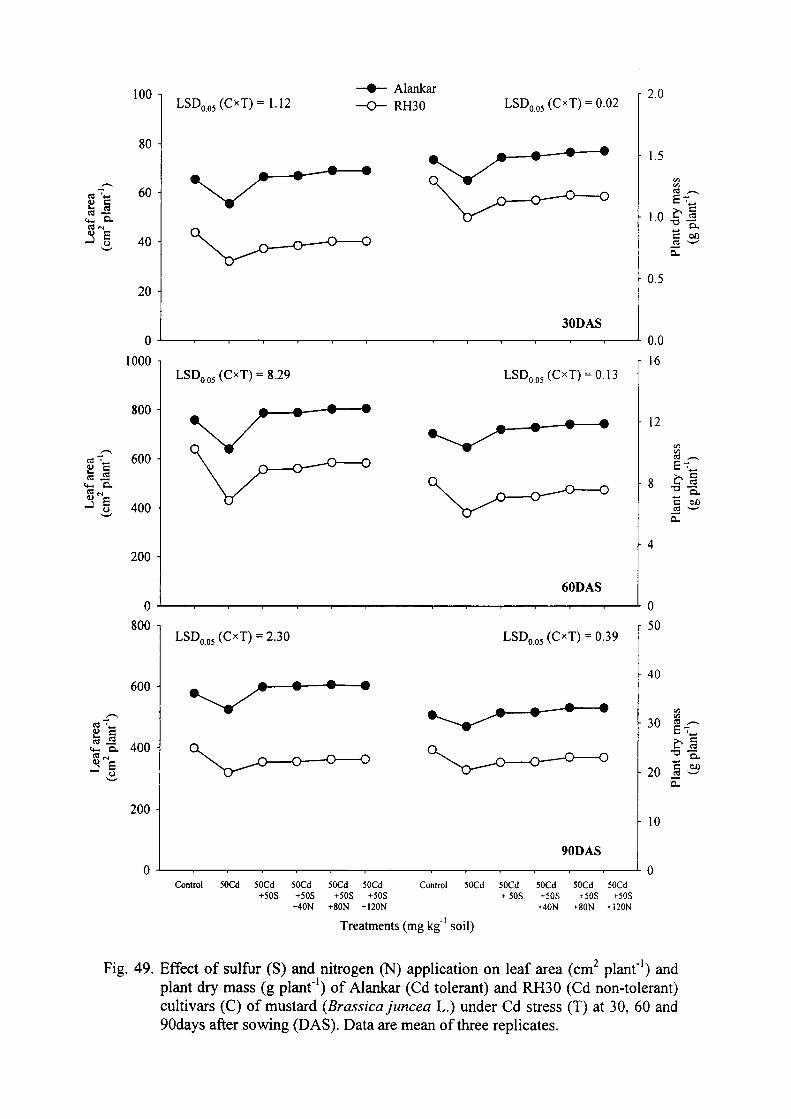
(0 ' •«
CO <N
-J o
100 1
80 •
60
40 ^
20
0
1000
800
CO -^ 600 u c c3 i2
( i i a ,
S " -• i , 400
200 ^
0
800
LSDoo5(CxT)=1.12 -•— Alankar - O - RH30
r I • T I r
LSD„„5(CxT) = 8.29
600
<2 -Q. 400
"1
200 -
LSDo„5(CxT) = 2.30
- • •
LSD„„,(CxT) = 0.02
30DAS
LSD„„(CxT) = 0.13
60DAS
LSDoo5(CxT) = 0.39
90DAS
2.0
- 1.5
1.0
« .-^ i=-„
C 6 0 CO ^ - -
• 0.5
0.0
16
12
s | l c 00
r 50
40
30
20
i £
T3
1 § Q.
3
10
Control 50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
Control 50Cd 50Cd 50Cd 50Cd 50Cd + 50S +50S +50S +50S
+40N +80N -^nON
Treatments (mg kg"' soil)
Fig. 49. Effect of sulfur (S) and nitrogen (N) application on leaf area (cm^ plant'') and plant dry mass (g plant"') of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

10 1
•10
-20
-30
1 • • • Alankar \":
H ^1 ^1 Hi ^1
111 1
^ ^ RH30
1 '
L
S ''
30DAS
r" 1 1
•
•
50Cd 50Cd +50S +50S +80N +120N
-10
-20
-30
Treatments (mg kg' soil)
Fig. 50. Per cent chaise in root and shoot length of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress with sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +I20N
Treatments (mg kg'' soil)
Fig. 51. Per cent change in leaf area and plant dry mass of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

different levels of N along with 50mg S kg"' soil was almost similar as observed for
growth characteristics. Among the N levels used, 80mg N kg'' soil along with 50mg S
kg"' soil appeared as the most effective combination for reversing the Cd-induced
(50mg Cd kg'' soil) reductions in photosynthetic characteristics (Figures 52-54).
Application of 50mg S kg"' soil together with 80mg N kg'' soil increased the
total chlorophyll and carotenoids content in Alankar by 3.2 and 3.1% at 30DAS and
5.3 and 4.8% at 60DAS but at 90DAS, this combination only lowered the reductions,
which limited to 3.9 and 2.4% in plants treated with 50mg Cd kg'' soil, respectively,
over their respective control. The activity of carbonic anhydrase, net photosynthetic
rate and stomatal conductance in Alankar were increased by 4.6, 6.2, 4.0% at 30DAS;
5.7, 6.6, 5.7% at 60DAS and 3.5, 4.2, 2.3% at 90DAS, respectively, over their
respective control due to 80mg N kg'' soil plus 50mg S kg'' soil of 50mg Cd kg'' soil-
treated plants (Figures 55-57).
Cadmium-caused reductions in photosynthetic characteristics in RH30 were
only lowered by all N levels in combination with 50mg S kg'' soil. The application of
80mg N kg"' soil along with 50mg S kg'' soil maximally lowered the reductions in
photosynthetic characteristics of RH30 plants treated with 50mg Cd kg'' soil (Figures
55-57).
4.5.3 Sulfur and nitrogen assimilation
Application of 50mg S kg'' soil together with 80mg N kg"' soil substantially increased
the ATP-sulfurylase activity and S content in both the cultivars treated with 50mg Cd
kg"' soil at all growth stages. Among the combinations, 80mg N kg"' soil and 50mg S
kg"' soil given to 50mg Cd kg"' soil treated plants resulted in maximum ATP-
sulfurylase activity and S content at all growth stages. Whereas, the same combination
maximally lowered the reductions in nitrate reductase activity and N content in both
the cultivars at all growth stages. Application of other N levels with 50mg S kg"' soil
proved less effective (Figures 58-59).
In Alankar, application of 80mg N kg"' soil and 50mg S kg ' soil maximally
increased ATP-sulfurylase activity and S content by 82.1, 54.4% at 30DAS; 85.5,
55.5% at 60DAS and 72.6, 42.1% at 90DAS of plants treated with 50mg Cd kg"' soil.
In RH30, ATP-sulfurylase activity and S content were increased by 60.0, 29.9% at
30DAS; 61.6% at 60DAS and 47.5, 21.4% at 90DAS over their respective control
(Figure 60).
79
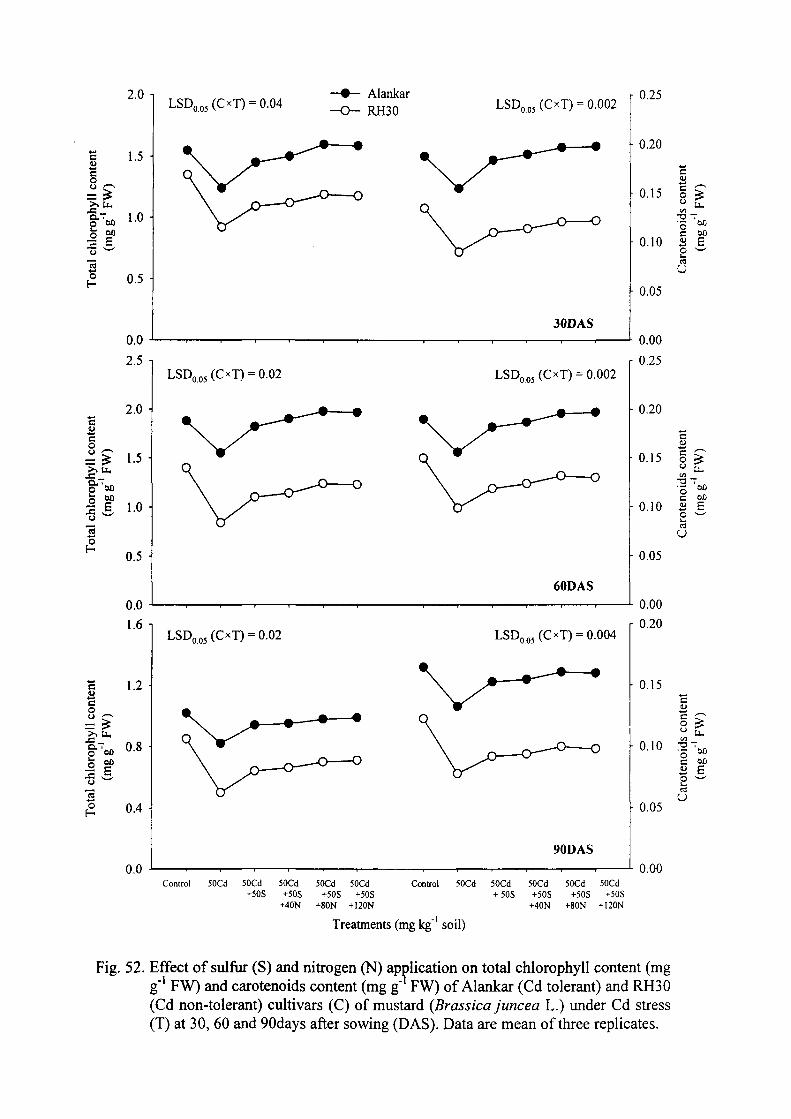
2.0
1.5
1.0 •
0.5
0.0
2.5
2.0 •
1.5 -
1.0
0.5
0.0
1.6
1.2
0.8
0.4 -
0.0
LSDoo5(CxT) = 0.04 -•— Alankar - O - RH30
LSDoo5(CxT) = 0.002
30DAS T I
LSD„„5(CxT) = 0.02 LSDoos (CxT) = 0.002
— •
60DAS r I
LSD(,o5 (CxT) = 0.02 LSD„„, (CxT) = 0.004
90DAS
Control 50Cd 50Cd +50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ I20N
Control 50Cd 50Cd + 50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ I20N
0.25
0.20
0.15
0.10
o c/l
•V
O c u o
^ u. , OJJ 01)
E N — i ^
• 0.05
0.00
0.25
0.20
c 1)
0.15 I ^
C 00
0.10 B E o ~-
0.05
0.00
0.20
0.15
• 0.10
- 0.05
0.00
Treatments (mg kg" soil)
Fig. 52. Effect of sulfur (S) and nitrogen (N) application on total chlorophyll content (mg g"' FW) and carotenoids content (mg g FW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.
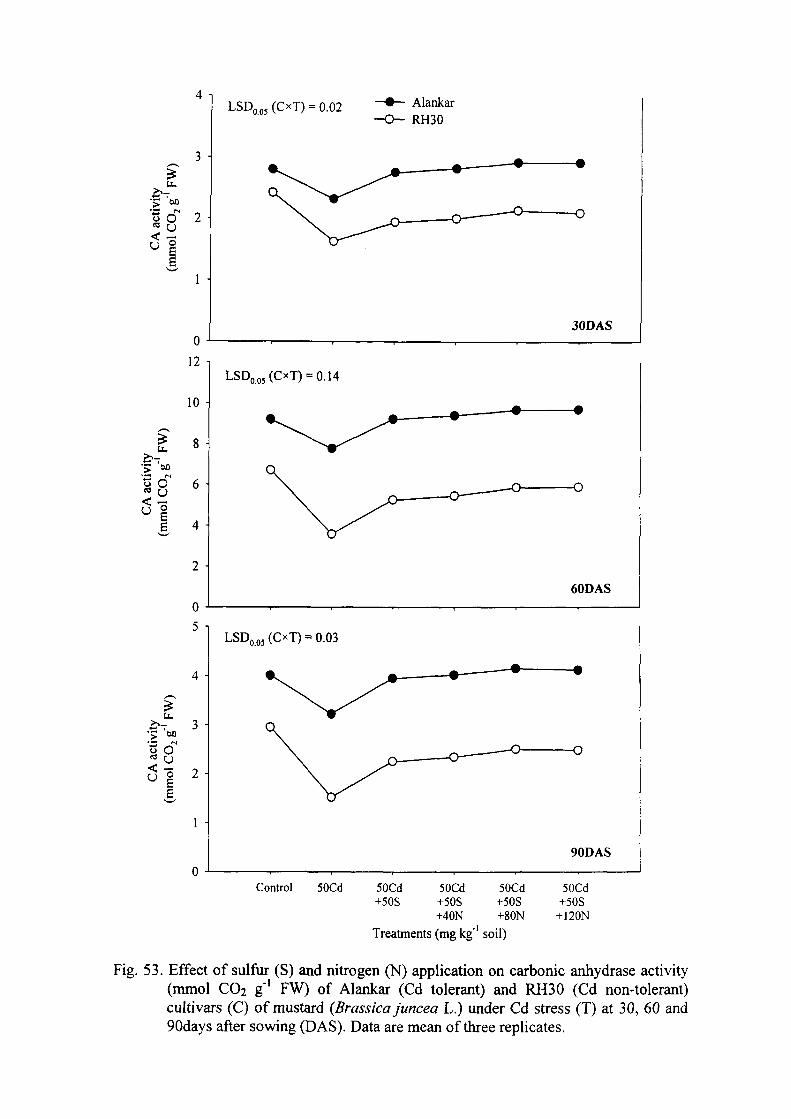
•5 60 ••3 A"
u 1
3 •
•> SO
CO JJ
61
1 •
0
12
10
LSDo„(CxT) = 0.02
8
2 ^
0
5
3 -
-•— Alankar • O - RH30
30DAS
LSDoo5(CxT) = 0.14
•
^ \
m m
0
60DAS
LSD005 (CxT) = 0.03
90DAS
Control 50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N Treatments (mg kg"' soil)
Fig. 53. Effect of sulfur (S) and nitrogen (N) application on carbonic anhydrase activity (mmol CO2 g"* FW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica Juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

25
20 •
15
10 -
5 -
0
25
20
15
10 ]
LSDoo3(CxT) = 0.17 -#— Alankar - C ^ RH30 LSDoo3(CxT) = 3.56
30DAS
0 16
12
LSDoo5(CxT) = 0.41
r T I
LSDoo3 (CxT) = 7.26
60DAS
8
4 -
LSD„o5 (CxT) = 0.18 LSD„„, (CxT) = 4.27
500
400 u u c
300 i T3 ' C o u
200 1 E o
uri
100
0
600
500
400 5
300
200 I
100
0 500
400 1) <j c
300 i ' • a ' c o u
^ 2 0 0 1 E o
100
90DAS
Control 50Cd 50Cd +50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ I20N
Control 50Cd 50Cd 50Cd 50Cd 50Cd + 50S +50S +50S +50S
+40N +80N +I20N
Treatments (mg kg" soil)
Fig. 54. Effect of sulfur (S) and nitrogen (N) application on net photosynthetic rate (^mol CO2 m' s"') and stomatal conductance (mmol m" s"') of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica Juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

-25
-30
-35 -
-40
10
Alankar ^ ^ RH30
-30
-40 -
-50 10
I -10 s o u
-20
-30
-40
-50
30DAS
I
^
90DAS
50Cd 50Cd +50S
50Cd +50S +40N
50Cd +50S +80N
50Cd +50S +120N
50Cd 50Cd +50S
50Cd +50S +40N
50Cd +50S +80N
50Cd +50S + 120N
5
0
-5
10 ^
-15
-20 ^
u -25 o
-30
-35
-40
10
•10 s
-20
-30
-40
-50
10
•10 ^
-20
-30
-40
-50
Treatments (mg kg' soil)
Fig. 55. Per cent change in total chlorophyll content and carotenoids content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard {Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).
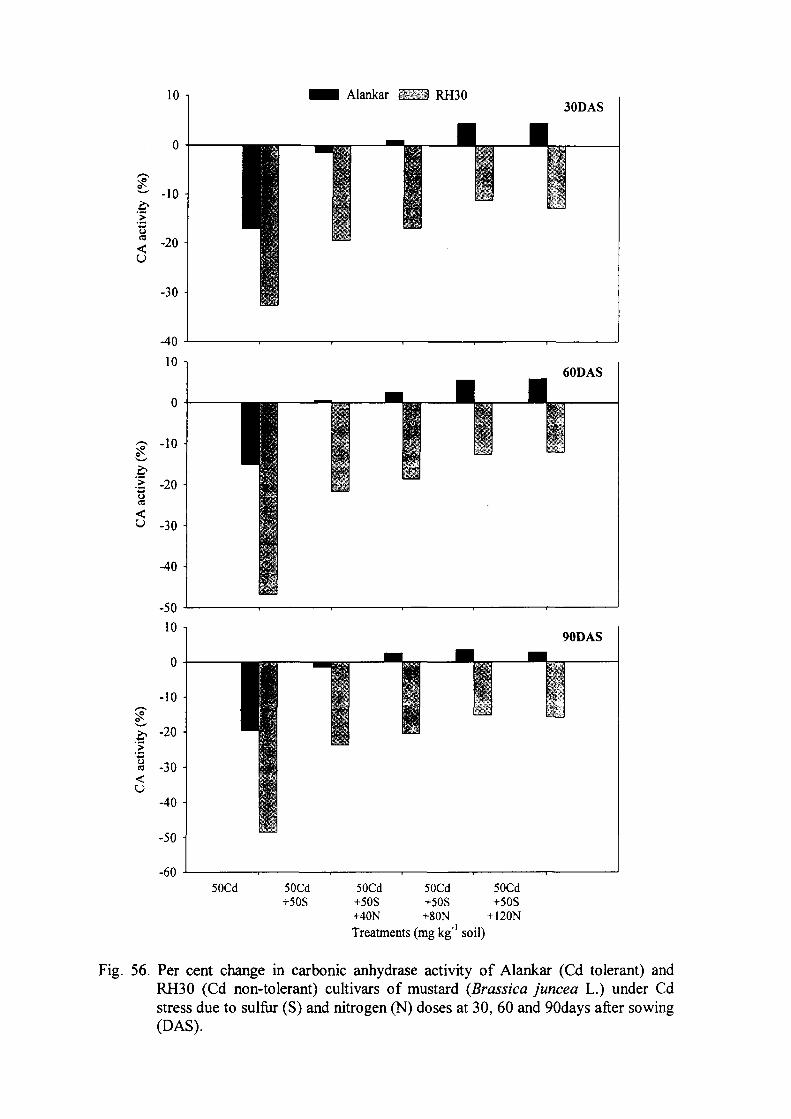
Alankar f^PTl RH30
lU -
0 -
^ -10 • 5? ^ > -20 • U
< U -30 •
-40 •
<n .
^ ^ • n k
:|!%
1 p ^
'-<•
•fC
>^.
* • ? ! '
l..» 1
_
w u V
b'iH •
I »>;;.
:.f
60DAS • -i
. 1 ! •
? ^
Ik,'*.
'r * J "
90DAS
50Cd 50Cd 50Cd +50S +50S
+40N
50Cd 50Cd +50S +50S +80N +120N
-1 Treatments (mg kg' soil)
Fig. 56. Per cent change in carbonic anhydrase activity of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

10
-40 10
-40
.a -10 -
2 -20
-30
90DAS
I % ^
50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
• - 5
-10
-15 B
-20 I • -25
-30
10
-10 5
-20 -=
• -30
-40 10
-10 B
-20
-30
-40
Treatments (mg kg" soil)
Fig. 57. Per cent change in net photosynthetic rate and stomatal conductance of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cuitivars of mustard {Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

60
50 -
40
30
20
10
0
50
40
B 30
20
10
0
70
60 •
50 -
40 -
30
20 ^
10
LSDo„5(CxT) = 2.99 -•— Alankar - O - RH30
LSDoo5(CxT) = 0.04
30DAS
LSDoo,(CxT)=1.44 LSDoo5(CxT) = 0.04
60DAS r t
LSDo,o5(CxT) = 0.22 LSDoo5 (CxT) = 0.03
90DAS
2.5
• 2.0
1.5 ^ .
c I- 1.0 S
0.5
0.0
3.0
2.5
I- 2.0
- 1.5 I c o o
Control 50Cd 50Cd +50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ 120N
Control 50Cd 50Cd + 50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ 120N
1.0
0.5
0.0
r 2.5
• 2.0
1.5
• 1.0
• 0.5
0.0
Treatments (mg kg' soil)
Fig. 58. Effect of sulfur (S) and nitrogen (N) application on ATP-sulfurylase activity (nmol Pi mg"' protein min"*) and sulfur content (%) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

600 1
500
400
300 •
200
100 -
LSDpoj (CxT) = 0.95 -•— Alankar - O - RH30
0
600
500 ^
400
• | ' « ^ 300
200
100
0
400
300
200
100
I I
LSDoo5(CxT) = 2.58
I • ' " • I
LSDoo5(CxT) = 2.43
LSDo„(CxT) = 0.03
30DAS
LSD„.,(CxT) = 0.04
60DAS
LSD„„(CxT) = 0.02
90DAS
r 6
• 5
4 ^
3 I c o u
• 1
c 3 B
c o u
• 4
• 3
c o
2 " Z
Control 50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
Treatments (mg kg" soil)
Control 50Cd 50Cd 50Cd 50Cd 50Cd + 50S +503 +50S +50S
+40N +80N +120N -1
Fig. 59. Effect of sulfur (S) and nitrogen (N) application on nitrate reductase activity (nmol NO2 g' FW h'') and nitrogen content (%) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

SOCd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
SOCd SOCd SOCd +50S +SOS
+40N
SOCd +S0S +80N
SOCd ••-SOS +120N
Treatments (mg kg" soil)
Fig. 60. Per cent change in ATP-sulfiirylase activity and sulfiir content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

The reductions in nitrate reductase activity and N content were reversed by S
supplementation in Cd stressed plants. Lesser decrease in nitrate reductase activity
and N content was observed when plants were given 50mg S kg'' soil plus 50mg Cd
kg"' soil in comparison to 50mg Cd kg"' soil alone. In Alankar, application of 80mg N
kg"' soil and 50mg S kg"' soil maximally increased the nitrate reductase activity and N
content by 5.2, 3.8% at SODAS; 5.2, 4.2% at 60DAS but at 90DAS, this combination
only lowered the reductions by 2.8, 5.4% in plants treated with 50mg Cd kg" soil,
respectively, over their respective control. In RH30, 50mg S kg"' soil alone and in
combination with 80mg N kg"' soil lowered the reductions in nitrate reductase activity
and N content of 50mg Cd kg"' soil treated plants (Figure 61).
4.5.4 Cadmium accumulation
In both the cultivars, increased accumulation of Cd in root and leaf due to 50mg Cd
kg"' soil was lowered by the application of 50mg S kg"' soil alone and in combination
with 40, 80 and 120mg N kg"' soil at all growth stages. Application of 40mg N kg"'
soil plus 50mg S kg"' soil also lowered the Cd accumulation in root and leaf in both
the cultivars but to a lesser extent than 80mg N kg"' soil. The effect of 120mg N kg"'
soil was equal to 80mg N kg"' soil on Cd accumulation. In Alankar, combined
application of 50mg S kg"' soil and 80mg N kg"' soil to 50mg Cd kg"' soil treated
plants maximally lowered the Cd content in root and leaf, which was 73.1, 14.6|ig g"'
DW at 30DAS; 88.5, 20.6^g g"' DW at 60DAS and 145.6, 15.4^g g"' DW at 90DAS,
respectively. Cadmium content in the root and leaf of RH30 with the same
combination was 104.3, 42.9^g g"' DW at 30DAS; 145.0, 61.9^g g"' DW at 60DAS
and 244.3, 50.3^g g"' DW at 90DAS, respectively (Figures 62-63).
4.5.5 Thiobarbituric acid reactive substances and hydrogen peroxide content
Application of 50mg S kg"' soil lowered the TEARS and H2O2 content of 50mg Cd
kg"' soil treated plants at all growth stages. Supplementation of 50mg S kg"' soil with
different levels of N (40, 80 and 120mg N kg"' soil) again lowered the contents of
TEARS and H2O2 of both the cultivars treated with 50mg Cd kg"' soil at all growth
stages. Combined application of 40mg N kg"' soil plus 50mg S kg"' soil also lowered
the TEARS and H2O2 content in both the cultivars but to a lesser extent than 80mg N
kg"' soil. Whereas, the effect of 120mg N kg"' soil plus 50mg S kg"' soil was equal to
80mg N kg"' soil plus 50mg S kg"' soil on TEARS and H2O2 content. In particular,
combined application of 80mg N kg"' soil plus 50mg S kg"' soil maximally lowered
80

10
5
0
? -5
1 -10 <J
(8
oi -15 Z
-20
-25
-30
10
Alankar RH30 30DAS
-10
-20
-30
-40
5
0
-5 ^
-10
-15
-20
-25
-30
-35
-40
—1 1—
60DAS
• T • 1 r~
90DAS
50Cd 50Cd +50S
50Cd +50S +40N
50Cd +50S +80N
50Cd +50S +120N
50Cd 50Cd +50S
50Cd +50S +40N
50Cd +50S +80N
50Cd +50S + 120N
10
5
0
-10 I c o
•15 z
-20
-25
-30
10
•10
c o
-20 " Z
-30
-40
5
0
-5
-10
-15
-20
-25
-30
-35
-40
Treatments (mg kg' soil)
Fig. 61. Per cent change in nitrate reductase activity and nitrogen content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

160
140
120
100 -
80 •
60 -
40
20
LSDoo3(CxT)= 11.19
Alankar RH-30 LSD„o5(CxT) = 4,14
0
250
200
150
100 -
50 •
0 350
300
250
200
150
100
50
0
LSDoo5(CxT)= 15.02 LSDoo5(CxT) = 6.08
LSD„„5(CxT) = 23.24 LSD„,o5(CxT) = 4.49
80
60
40
20
0
100
80
(- 60
40
20
0
80
60
40
20
Control 50Cd 50Cd +50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ 120N
Control 50Cd 50Cd + 50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ I20N
Treatments (mg kg'' soil)
Fig. 62. Effect of sulfur (S) and nitrogen (N) application on root Cd content and leaf Cd content (ng g'' DW) of Alankar (Cd tolerant) and RH30 (Cd non-toierant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

2500 1 Alankar F T ^ RH30
50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
1000
Treatments (mg kg' soil)
Fig. 63. Per cent change in root Cd content and leaf Cd content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

the TBARS and H2O2 contents by 27.1, 15.2% at 30DAS; 21.8, 11.6% at 60DAS and
45.2, 26.5% at 90DAS, respectively, over their respective control, in 50mg Cd kg'
soil treated Alankar plants (Figures 64-65).
In RH30, combined application of 80mg N kg'' soil plus 50mg S kg'' soil
lowered the TBARS and H2O2 contents by 97.1, 80.2% at 30DAS; 90.5, 77.6% at
60DAS and 149.2, 97.6% at 90DAS, respectively, over their respective control, in
plants treated with 50mg Cd kg"' soil (Figure 65).
4.5.6 Components of antioxidant system
4.5.6.1 Enzymatic antioxidants
The increase in SOD activity in plants treated with 50mg Cd kg"' soil was lowered by
the application ofSOmg S kg"' soil alone and in combination with 40, SO and 120mg
N kg"' soil in both the cultivars at all growth stages. Application of 40mg N kg"' soil
plus 50mg S kg"' soil also lowered the SOD activity in both the cultivars but to a
lesser extent than 80mg N kg"' soil. Whereas, the effect of 120mg N kg"' soil plus
50mg S kg"' soil was equal to 80mg N kg'' soil plus 50mg S kg"' soil on SOD activity
when treated with 50mg Cd kg'' soil. In Alankar, combination of 50mg S kg"' soil and
80mg N kg"' soil to 50mg Cd kg"' soil fed plants maximally lowered the SOD activity
by 20.6, 19.5 and 25.3% at 30, 60 and 90DAS, respectively, over their respective
control. Same combination lowered the SOD activity of RH30 plants treated with
50mg Cd kg"' soil by 36.7, 30.8 and 28.3% at 30, 60 and 90DAS, respectively, over
their respective control (Figure 66,68).
In Alankar, the increase in CAT activity due to 50mg Cd kg ' soil was further
increased by the application of 50mg S kg"' soil alone and in combination with 40, 80
and 120mg N kg'' soil at all growth stages. Combined application of 50mg S kg'' soil
and 80mg N kg"' soil to 50mg Cd kg"' soil treated plants maximally increased the
CAT activity by 52.2, 58.9, 34.3% at 30, 60 and 90DAS, respectively, over their
respective control. In RH30, the application of 50mg S kg"' soil and 80mg N kg"' soil
increased the CAT activity by 23.9% at 30DAS, while, at 60 and 90DAS its
application only limited the decrease in CAT activity to 1.02 and 2.30%, respectively,
in plants treated with 50mg Cd kg"' soil (Figure 66,68).
The increase in APX activity due to 50mg Cd kg"' soil was further increased
by the application of 50mg S kg"' soil alone and in combination with 40, 80 and
120mg N kg"' soil in both the cultivars at all growth stages. In Alankar, combined
application of 50mg S kg'' soil and SOmg N kg"' soil to 50mg Cd kg"' soil treated
81

50
40 •
30
20
10
LSD„„5(CxT) = 0.90
0
100
80
60
40
20
0
100
80
60
40
20 -I
-•— Alankar
- O - RH30 LSDo„5(CxT)= 14.60 r 250
200
• 150
- 100
- 50
30DAS
LSDo.o3(CxT) = 0.90 LSDoo5(CxT)= 16.86
60DAS
LSD„„,(CxT) = 0.J LSDoo5(CxT)= 18.63
90DAS
0
300
• 250
200
150
100
50
0
r 300
250
200
150
100
50
Control 50Cd 50Cd +50S
50Cd +50S
+40N
50Cd 50Cd +50S +50S
+80N +120N
Control 50Cd 50Cd + 50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ 120N
Treatments (mg kg" soil)
Fig. 64. Effect of sulfur (S) and nitrogen (N) application on thiobarbituric acid reactive substances (nmol g"' FW) and hydrogen peroxide content (|imol g'' FW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

SOCd SOCd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
SOCd SOCd +50S
SOCd SOCd +SOS +SOS +40N ^80N
SOCd +50S + I20N
Treatments (mg kg" soil)
Fig. 65. Per cent change in thiobarbituric acid reactive substances and hydrogen peroxide content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard {Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

15
12
D C OS 6
D
3 •
0
15
12
3 •
0
15
12 •
LSDoo5(CxT) = 0.95 Alankar RH30 LSDoo3(CxT) = 2.75
30DAS -T 1 r— t —I ~r—
LSDo„(CxT)=1.00 LSDo„5(CxT) = 3.12
60DAS
LSD„„,(CxT) = 0.95 LSDoo5 (CxT) = 0.63
-O O-
90DAS
250
200
150 .?
I- 100
50
0
350
300
250
- 200
150
- 100
- 50
0
250
- 200
- 150 .e
100
Control 50Cd 50Cd SOCd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
Control SOCd SOCd + SOS
SOCd +50S
+40N
SOCd +SOS
+80N
SOCd +50S
+ 120N
Treatments (mg kg"' soil)
50
Fig. 66. Effect of sulfur (S) and nitrogen (N) application on superoxide dismutases activity (Units mg'' protein) and catalase activity (Units mg'' protein) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

3.5
3.0
I §. 2.0
X ^ 15 a, =2 < c
B 1.0 0.5 -
LSDQOJ (CXT) = 0.03
0.0
5.0
4.0 -
.S •f S 3.0 o -
X 6 0- a 2.0
1.0
0.0 4.0
3.0
2.0 -
1.0
0.0
Alankar RH30 LSDoo5 (CxT) = 0.006
30DAS
LSDooj (CxT) = 0.28 LSD(,o5 (CxT) = 0.005
60DAS
LSD„„5 (CxT) = 0.03 LSDooj (CxT) = 0.003
90DAS
r 0.6
0.5
• 0.4
Control 50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
Control 50Cd 50Cd + 50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+ 120N
.E
0.3 o 'oo
a .- 0.2 5
0.1
0.0
0.8
0.6 ^ _c '53
0.4 5 'oo
0.2
0.0
0.5
0.4
0.3
0.2
0.1
0.0
Treatments (mg kg" soil)
Fig. 67. Effect of sulfur (S) and nitrogen (N) application on ascorbate peroxidase activity (Units mg"' protein) and glutathione reductase activity (Units mg'' protein) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

50Cd 50Cd 50Cd SOCd 50Cd +50S +50S +50S +50S
+40N +80N +120N
SOCd SOCd SOCd SOCd SOCd +50S +50S +50S +50S
+40N +80N +120N
Treatments (mg kg" soil)
Fig. 68. Per cent change in superoxide dismutases activity and catalase activity of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

-•""s
^
^ > 4 ^
u ea X O. < •
350
300
250
200
150
100
50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
50Cd 50Cd +50S
50Cd +50S +40N
50Cd +50S +80N
50Cd +50S + 120N
Treatments (mg kg" soil)
Fig. 69. Per cent change in ascorbate peroxidase activity and glutathione reductase activity of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30,60 and 90days after sowing (DAS).

plants maximally increased the APX activity by 297.3, 301.2, 209.1% at 30, 60 and
90DAS, respectively, over their respective control. Whereas, the increase in APX
activity of RH30 by this combination was 135.0, 175.5 and 120.4% at 30, 60 and
90DAS, respectively, over their respective control (Figure 67,69).
The increase in GR activity was same as observed in the case of APX activity.
In Alankar, combined application of 50mg S kg"' soil and 80mg N kg'' soil to 50mg
Cd kg'' soil treated plants maximally increased the GR activity by 150.8, 157.3,
92.1% at 30, 60 and 90DAS, respectively, over their respective control. Whereas, the
increase in APX activity of RH30 by this combination was 85.5,94.0,47.8% at 30, 60
and 90DAS, respectively, over their respective control (Figure 67,69).
4.5.6.2 Non-enzymatic antioxidants
The decrease in ascorbic acid content due to 50mg Cd kg"' soil was lowered by the
application of 50mg S kg'' soil alone and in combination with 40, 80 and 120mg N
kg'' soil in both the cultivars at all growth stages. Combined application of 40mg N
kg"' soil plus 50mg S kg'' soil also lowered the decrease in ascorbic acid content of
both the cultivars but to a lesser extent than 80mg N kg"' soil. The effect of 120mg N
kg"' soil plus 50mg S kg'' soil was equal to 80mg N kg'' soil plus 50mg S kg"' soil on
ascorbic acid content when treated with 50mg Cd kg"' soil. In Alankar, combined
application of 50mg S kg"' soil and 80mg N kg'' soil to 50mg Cd kg'' soil treated
plants lowered the decrease in ascorbic acid content by 5.4, 4.3, 7.5%) at 30, 60 and
90DAS, respectively, over their respective control. Whereas, in RH30, this
combination lowered the decrease in ascorbate content by 15.2, 10.1, 18.1%) at 30, 60
and 90DAS, respectively, over their respective control (Figures 70-71).
The increase in glutathione content due to 50mg Cd kg'' soil was further
increased by the application of 50mg S kg'' soil alone and in combination with 40, 80
and 120mg N kg'' soil in both the cultivars at all growth stages. Combination of 40mg
N kg'' soil plus 50mg S kg"' soil also increased the glutathione content of both the
cultivars but to a lesser extent than 80mg N kg'' soil plus 50mg S kg'' soil. Whereas,
the effect of combined application of 120mg N kg'' soil plus 50mg S kg'' soil was
almost equal to 80mg N kg'' soil plus 50mg S kg"' soil on glutathione content when
treated with 50mg Cd kg"' soil. In Alankar, combined application of 50mg S kg"' soil
and 80mg N kg"' soil to 50mg Cd kg"' soil fed plants maximally increased the
glutathione content by 58.8, 60.7, 47.3% at 30, 60 and 90DAS, respectively, over
their respective control. While, the same combination increased the glutathione
82

400
300 •
200 -
100 -
0
500
400
300
200 ^
100
400
300
200 -
100 -
LSD„„,(CxT) = 0.63 Alankar RH30 LSDoo5 (CxT) = 2.03
30DAS I I
LSDo„ (CxT) = 0.74 LSD„„ (CxT) = 4.49
60DAS
LSD„o5 (CxT) = 0.85 LSD„„ (CxT) = 2.17
90DAS
800
700
600
500 c > "J rT
S 'wi • 400
- 300
200
100
0
r 1200
• 1000
• 800
• 600
• 400
- 200
0
700
600
500
400 ^ ta-
300 ^ I
200
100
Control 50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
Treatments (mg kg' soil)
Control 50Cd 50Cd 50Cd 50Cd 50Cd + 50S +50S +50S +50S
+40N +80N +120N
1
Fig. 70. Effect of sulfur (S) and nitrogen (N) application on ascorbic acid content (nmol g' FW) and glutathione content (nmol g FW) of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard {Brassica juncea L.) under Cd stress (T) at 30, 60 and 90days after sowing (DAS). Data are mean of three replicates.

50Cd 50Cd +50S
50Cd +50S +40N
50Cd +50S +80N
50Cd +50S + 120N
50Cd 50Cd 50Cd +50S +50S
+40N
50Cd +50S +80N
50Cd +50S + 120N
Treatments(mgkg' soil)
Fig. 71. Per cent change in ascorbic acid content and glutathione content of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard {Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at 30, 60 and 90days after sowing (DAS).

M
.2P ' S
-a u <u o o o
100
e « o. u u a. a 3 CT-
'« O Urn
<u £ 3
:^
80
60
40
20
0
8
LSDoo3(CxT) = 3.32
Alankar RH30
LSD„„,(CxT)=1.21
r I
LSD„.o5(CxT) = 0.28 LSD„„5(CxT) = 0.14
-O
16
• 14
12 §•
• 10
• 6
• 4
Control 50Cd 50Cd +50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+I20N
Control 50Cd 50Cd + 50S
50Cd +50S
+40N
50Cd +50S
+80N
50Cd +50S
+120N
Treatments (mg kg' soil)
a. • a
u Xi
3
u c/1
Fig. 72. Effect of sulfur (S) and nitrogen (N) application on number of siliqua per plant, number of seeds per siliqua, 1000 seed weight (g) and seed yield (g plant"') of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars (C) of mustard (Brassica j'uncea L.) under Cd stress (T) at harvest i.e., 120days after sowing (DAS). Data are mean of three replicates.

Alankar
3
o.
u s> B 3
2
00
.2P ' S
• o <u (U
o o o
50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
50Cd 50Cd 50Cd 50Cd 50Cd +50S +50S +50S +50S
+40N +80N +120N
C
D. a 3 a-
x> E 3
z
2 u
'?> T3
on
Treatments (mg kg' soil)
Fig. 73. Per cent change in number of siliqua per plant, number of seeds per siliqua, 1000 seed weight and seed yield of Alankar (Cd tolerant) and RH30 (Cd non-tolerant) cultivars of mustard (Brassica juncea L.) under Cd stress due to sulfur (S) and nitrogen (N) doses at harvest i.e., 120days after sowing (DAS).

content in RH30 by 37.1, 40.5, 33.8% at 30, 60 and 90DAS, respectively, over their
respective control (Figures 70-71).
4.5.7 Yield characteristics
The reductions in yield characteristics (number of siliqua per plant, number of seeds
per siliqua, 1000 seed weight and seed yield per plant) due to 50mg Cd kg'' soil were
lowered by the application of 50mg S kg'' soil alone and in combination with 40, 80
and 120mg N kg'' soil in both the tolerant (Alankar) and non-tolerant (RH30)
cultivars. A combination of 50mg S kg'' soil and 80mg N kg'' soil maximally lowered
the reductions in yield characteristics. In Alankar, lesser decrease in number of siliqua
per plant, number of seeds per siliqua, 1000 seed weight and seed yield per plant due
to 50mg S kg'' soil and 80mg N kg"' soil was 1.9, 5.0, 4.8 and 1.9%, respectively,
over their respective control. Whereas, in RH30, the decrease remained to 19.2, 23.0,
21.0 and 20.4% due to 80mg N kg'' soil plus 50mg S kg'' soil (Figures 72-73).
4.6 Summary of Experiment 3
• Coordination of sulfiir and nitrogen proved effective in mustard cultivation. A
package of these nutrients not only helped in increasing crop productivity but
also exhibited its potential in the alleviation of Cd effects.
• Among the N levels applied, 80mg N kg'' soil strengthened the effectiveness
of 50mg S kg'' soil in the alleviation of 50mg Cd kg'' soil induced reductions
in both the cultivars at all growth stages. The effectiveness of the combined
application of 50mg S kg'' soil plus 80mg N kg'' soil was different in tolerant
and non-tolerant cultivars. In Alankar (tolerant cultivar), the Cd-induced
reductions in growth, photosynthetic, biochemical and yield characteristics
were completely overcome, whereas, the reductions in these characteristics
were only lowered by this combination in RH30 (non-tolerant cultivar).
• Application of 50mg S kg'' soil and 80mg N kg'' soil to 50mg Cd kg'' soil
treated plants increased the ATP-sulfurylase activity and S content and also
improved the NR activity and N content to a greater extent.
• Cadmium-induced increase in root and leaf Cd content was lowered by the
combined application of 50mg S kg'' soil and 80mg N kg'' soil to 50mg Cd
kg'' soil treated plants in both the cultivars at all growth stages. The extent of
decrease in Cd content was more in Alankar than RH30.
• The accumulation of TBARS and H2O2 due to 50mg Cd kg'' soil was lowered
by the combined application of 50mg S kg'' soil and 80mg N kg'' soil in both
83

the cultivars at all growth stages. The extent of decrease in TBARS and H2O2
content was greater in Alankar than RH30.
The activities of antioxidant enzymes (catalase, ascorbate peroxidase and
glutathione reductase) were maximally increased by the combined application
of 50mg S kg'' soil and 80mg N kg'' soil in 50mg Cd kg"' soil treated plants.
Whereas, activity of SOD was lowered by this combination of sulfur and
nitrogen in both the cultivars at all growth stages.
Combined application of 50mg S kg'' soil and 80mg N kg'' soil lowered the
Cd-induced decrease in ascorbic acid content. This combination, however,
increased the glutathione content in both the cultivars at all sampling stages.
Application of 50mg S kg'' soil plus 80mg N kg'' soil reversed the reduction
in yield characteristics of Alankar and values reached on par to control.
Whereas, this combination only lowered the reduction in yield characteristics
ofRH30.
84

DISCUSSION

Contents
DISCUSSION
5.1 Screening of Mustard Cultivars Treated with Different Levels of Cadmium 86 Growth characteristics 86 Photosynthetic characteristics 88 Cadmium accumulation 92 Yield characteristics 93 Alleviation of Cadmium-toxicity by Sulfur and Nitrogen in Alankar (Cd tolerant) and RH30 (Cd non-tolerant) Cultivars of Mustard 95 Growth characteristics 96 Photosynthetic characteristics 97 Sulfur and nitrogen metabolism 100 Cadmium accumulation 103 Cadmium-induced oxidative stress 104 Components of antioxidant system 105 Enzymatic antioxidants 105 Non-enzymatic antioxidants 110 Yield characteristics 113 New Report in the Thesis and Future Prospects 115
5.1.1 5.1.2 5.1.3 5.1.4 5.2
5.2.1 5.2.2 5.2.3 5.2.4 5.2.5 5.2.6 5.2.6.1 5.2.6.2 5.2.7 5.3

DISCUSSION Chapter 5
Every year India loses hundreds of millions of rupees from reductions in crop
productivity caused by abiotic stresses (Mahajan and Tuteja 2005). According to Bray
et al. (2000), the relative decreases in potential maximum yields associated with
abiotic stress factors vary between 50-80%. Among abiotic stresses, heavy metal
contamination is a serious envirormiental problem that limits plant productivity and
threatens human health (Wagner 1993, Sanita di Toppi and Gabbrielli 1999). The
agricultural soil may have toxic levels of heavy metals due to agricultural and
industrial practices such as application of pesticides and chemical fertilizers, waste
water irrigation, precipitation from heavy coal combustion, and smelter wastes and
residues from metalliferous mining (Lombi et al. 2000, Vassilev et al. 2006, Xie et al.
2006, Verma et al. 2007).
For sustainable crop production, it is important to develop methods or
techniques for alleviating the Cd-induced growth inhibition and reducing its
accumulation in plants. One option is the use of balanced S and N fertilizers for
alleviating Cd stress and maintaining productivity of cultivated soils. As Cd induces
essential nutrient deficiency and even decreased concentration of several
macronutrients in plants (Jiang et al. 2004), it seems possible to reverse or at least
partly reduce the Cd-induced growth inhibition by optimization of S and N.
Increasing evidence suggests that mineral-nutrient status of plants plays a critical role
in increasing plant resistance to environmental stresses (Marschner 1995, Vassilev et
al. 2005, Hassan et al. 2005a,b, 2008a,b, Anjum et al. 2008a,b, Khan et al. 2008a).
Sulfur and N are essential nutrients for normal growth and development of plants.
Their assimilation pathways are well coordinated (Brunold 1993, Takahashi and Saito
1996, Abdin et al. 2003, Hawkesford et al. 2006). Deficiency of one element was
shown to repress the other pathway (Neuenschwander et al. 1991, Kim et al. 1999,
Koprivova et al. 2000, Migge et al. 2000, Prosser et al. 2001, Hesse et al. 2004,
Scherer 2008). S is a structural constituent of several coenzymes and prosthetic
groups, such as ferredoxin, which are also important for N assimilation. Thus, S plays
an important role in plant growth and in the regulation of plant development. It has
also been found that S nutrition is a critical factor for the alleviation of Cd toxicity
(Popovic et al. 1996, Chen and Huerta 1997, Hassan et al. 2005b, 2006, Vassilev et
al. 2005, Anjum et al. 2008a,b). A positive effect of S nutrition on Cd detoxification
85

in Beta vulgaris plants has also been established (Popovic et al. 1996). It has been
found that at suboptimal S nutrition Cd exposed plants preferably allocate S to
phytochelatin synthesis (McMahon and Anderson 1998). Anjum et al. (2008a) also
reported that S supplementation increased the production of glutathione content under
low level of Cd which protects the Brassica campestris plants by improving the
growth and photosynthesis. Pankovic et al. (2000) have shown that optimal N supply
decreased the inhibitory effects of Cd on photosynthesis of Helianthus annuus plants.
A proper N supply has been shown to have a positive effect in overcoming the
adverse effects caused by Cd toxicity in various crop species (Hassan et al. 2005a).
Therefore, coordination of S and N may alleviate the Cd-induced toxicity in crop
plants.
The present study was undertaken to understand the response of Brassica
juncea cultivars to Cd stress and the use of S nutrition alone and in combination with
N in the amelioration of Cd toxicity in Brassica juncea cultivars.
5.1 Screening of Mustard Cultivars Treated with Different Levels of Cadmium
The order of decrease in growth, photosynthetic and yield characteristics for all the
cultivars was RH30 >Sakha >Pusa Bold >Varuna >Alankar. RH30 accumulated
greatest amount of Cd and Alankar accumulated lowest Cd in root and leaf (Figures 2-
21).
5.1.1 Growth characteristics
Plant growth and development are an outcome of coordination of the main biological
processes in plants (Vassilev et al. 1998). Plant growth and development are
susceptible to stresses of all kinds including those of heavy metals. Cd adversely
affects plant growth and metabolism. The most common effect of Cd toxicity in plants
is stunted growth, leaf chlorosis and alteration in the activity of many key enzymes of
various metabolic pathways (Godbold and Hutterman 1985, Arduini et al. 1994, Wu
and Zhang 2002b, Chen et al. 2003, Wu et al. 2003, Rai et al. 2005). Bachir et
al. (2004) found decrease in root length, plant height and fruiting branch number
in cotton plants with increasing Cd concentration in a pot experiment. Growth
reduction in Cd-treated plants has been described (Ouzounidou et al. 1997, Wu and
Zhang 2002b, Wu et al. 2003, Demirevska-Kepova et al. 2006), due to the higher
accumulation of Cd and reductions in the availability of other nutrients resulting in
disturbed metabolism (Jalil et al. 1994, Wu et al. 2006). In my studies shoot length,
root length, leaf area and plant dry mass of all the Brassica Juncea cultivars were
86

decreased with different concentrations of Cd (0, 25, 50, 100 and 150mg Cd kg"' soil).
Root growth was found more susceptible to Cd toxicity compared to the shoot.
Metwally et al. (2005) also reported that the roots of Pisum sativum plants were
more sensitive to Cd toxicity than shoot. Increasing concentration of Cd in liquid
culture and pot experiment decreased the germination and root growth of
Raphanus sativus and Daucus carota plants (Chen et al. 2003). In the present
study, greater impact of Cd observed on root growth as compared to shoot in all
Brassica juncea cultivars, could be due to higher accumulation of Cd in root, leading
to impaired growth metabolism. A greater reduction in the root length could also be
an adaptation of the cultivars studied in response to high external Cd concentration, so
as to reduce the uptake of Cd along with water and their subsequent transport to the
shoot. Parameters such as root length, shoot length, leaf area as well as plant dry mass
have been used as indicators of metal toxicity in plants (Ouzounidou et al. 1997). In
my study, Alankar cultivar showed minimum decrease in shoot length, root length,
leaf area and plant dry mass, while RH30 showed maximum decrease in these
parameters (Figiires 2-7). The reduction in the growth of these cultivars could also be
due to suppression of the elongation growth rate of cells, because of an irreversible
inhibition exerted by Cd on the proton pump responsible for the process (Aidid and
Okamoto 1993). It has also been reported previously that species and cultivars display
marked differences for Cd tolerance in Triticum aestivum (Zhang et al. 2002, Khan et
al. 2006), Hordeum vulgare (Wu et al. 2003), Gossypium hirsutum (Wu et al. 2004),
Brassica juncea (Qadir et al. 2004), Pisum sativum (Metwally et al. 2005), Oryza
sativa (Wu et al. 2006, He et al. 2006), Vigna radiata (Wahid and Ghani 2007,
Anjum et al. 2008c) and Brassica campestris (Anjum et al. 2008d). Cadmium
treatments inhibited shoot and root growth and dry weight of two Triticum
aestivum cultivars, C-1252 and Balcali-85. The decrease was more distinct in
cultivar C-1252 (Ozturk et al. 2003). The presence of Cd decreased the seedling
growth of Zea mays, Triticum aestivum, Cucumis sativus and Sorghum bicolor in
terms of root and shoot growth (An 2004). Ghnaya et al. (2005) reported that Cd
severely inhibited Mesembryanthemum crystallinum growth even at low
concentration. The process of tolerance in plants is exhibited at different structural
and functional levels, viz., molecular, biochemical, cellular and the whole plant. At
the whole plant level it is possible that the tolerance mechanism is connected to the
87

pattern of Cd distribution as well as the response of components of antioxidant
system.
Analyzing the effect of Cd on many plant species, Vassilev and Yordanov
(1997) concluded that Cd diminishes relative growth rate through inhibiting mainly
net assimilation rate, and to a lesser extent, by restricting leaf area ratio. Inhibition of
net assimilation rate is caused by disturbances of dark respiration and photosynthesis.
Decreased leaf area ratio in Cd-treated plants is a consequence of the decrease of
turgor potential and cell wall elasticity, resulting in smaller size of leaf cells formed
with smaller intercellular space area,
5.1.2 Photosynthetic characteristics
Cd induces changes in the physiological processes such as respiration, photosynthesis
and gas exchange of the plants (Lagriffoul et al. 1998, Hegedus et al. 2001, Mobin
and Khan 2007). Chloroplasts comprise only about one percent of the total Cd
accumulated by a plant. Despite its very low relative concentration in chloroplast, it
seriously blocks the activity of photosynthetic processes at different routes
(Ghoshrony and Nadakavukaren 1990, Siedlecka and Krupa 1999). As a visible
symptom the preferential degradation of chlorophyll and carotenoids content can be
used to monitor the metal-induced damage in leaves (Horvath et al. 1996, Sanita di
Toppi et al. 1998, Oncel et al. 2000).
In the reported study, chlorophyll and carotenoids content decreased with the
increase in Cd levels at all growth stages in all Brassica juncea cultivars. The extent
of decrease in chlorophyll and carotenoids content was maximum in RH30 while it
was minimum in Alankar (Figures 9-10,14). It has been reported earlier that Cd
decreased the chlorophyll and carotenoids content in Helianthus annuus (Di Cagno et
al. 2001), Hordeum vulgare (Wu et al. 2003, Vassilev et al. 2004), Oryza sativa (Kuo
and Kao 2004), Glycine max (Drazic et al. 2004), Sedum alfredii (Zhou and Qiu
2005), Matricaria chamomilla (Kovacik et al. 2006), Brassica juncea (Mobin and
Khan 2007), Triticum aestivum (Khan et al. 2007a), Brassica campestris (Anjum et
al. 2008a) and Vigna mungo (Singh et al. 2008a). Decreased photosynthesis can be
caused by the decrease in the level of photosynthetic pigments, breakdown or the
inhibition of its synthesis (Padmaja et al. 1990, Mobin and Khan 2007). Cadmium
interferes with chlorophyll formation, with functional SH-group of sulfhydryl
requiring enzymes such as ALA synthetase, ALA dehydratase and
protochlorophyllide reductase. Apart from inhibition of biosynthetic enzymes of
88

chlorophyll formation, the increased level of free radicals of fatty acids produced
from polyunsaturated fatty acids due to the higher activity of lipooxygenase may also
contribute to the decreased level of chlorophyll with Cd treatments (Klein et al. 1984,
Somashekaraiah et al. 1992). Cd might have replaced the central Mg from chlorophyll
molecules and thereby reduced the photosynthetic light harvesting ability of plants
(Kupper et al. 1996). The decrease in chlorophyll content may also be correlated with
the adverse effect of Cd on the uptake and accumulation of other essential nutrients in
plants viz. Fe, Mg, Ca and K (Greger and Ogrer 1991, Greger and Lindberg 1987,
Ouzounidou et al. 1997), since both Fe and Mg are essential elements required for the
formation of chlorophyll. Earlier reports also suggest that the change in Fe:Zn may be
responsible for the reduced chlorophyll content in plants (Root et al. 1975).
Carotenoids are synthesized in the chloroplast of plants. Carotenoids are
important as they absorb light at wavelength between 400 and 550nm and transfer it
to the chlorophylls (an accessory light-harvesting role) (Siefermann-Harms 1987) and
protect the photosynthetic apparatus by quenching a triplet sensitizer (Chi ), singlet
oxygen and other harmfiil free radicals which are naturally formed during
photosynthesis (an antioxidant function) (OelmuUer 1989, Thiele et al. 1996, Havaux
et al. 2000). Moreover, they are important for the photosystem (PSl) assembly and
the stability of light harvesting complex proteins as well as thylakoid membrane
stabilization (a structural role) (Mayfield and Taylor 1984, Siefermann-Harms 1987,
Wrischer et al. 1998, Niyogi et al. 2001, Kim et al. 2004). The decrease of
carotenoids content in seedlings of Raphanus sativus, Ulva lactuca, Triticum aestivum
and Oryza sativa growing in the presence of Cd was reported by Naguib et al. (1982).
Panda and Khan (2003) reported decrease in carotenoids content due to heavy metal
treatments. In my experiments, carotenoids content decreased with the increase in Cd
concentration in all the cultivars at all growth stages. When the level of carotenoids
was decreased by Cd treatments (Khudsar et al. 2001, Khan et al. 2007a, Singh et al.
2008a), the protective functions of carotenoids could not be maintained.
Consequently, the oxidative degradation of chlorophyll and rapid destruction of
photosynthetic membranes occurred.
Photosynthesis is an integral process with a high degree of self regulation.
Inhibition of photosynthetic rate could be due to structural and ftmctional disorders.
Cadmium may interact with the photosynthetic apparatus at various levels of
organization and architecture (Mysliwa-Kurdziel and Strzalka 2002). Besides
89

affecting the functions of chloroplast membranes, Cd can also alter the components of
photosynthetic electron transport chain, particularly the PSI and PSII (Siedlecka and
Krupa 1996).
In the present study, increasing concentration of Cd in the soil leads to reduced
photosynthetic characteristics in all Brassica juncea cultivars at all growth stages.
Maximum decrease in these characteristics was noted with 150mg Cd kg"' soil. The
decrease in net photosynthetic rate, stomatal conductance and carbonic anhydrase
activity varied among cultivars and RH30 exhibited maximum decrease while
Alankar showed minimum decrease (Figures 11-13,15-16). Earlier investigations have
also demonstrated a marked reduction in the overall rate of photosynthesis by Cd in
different plant species (Arduini et a/. 2004, Wojcik and Tukiendorf 2005, Jing et al.
2005, Khan et al. 2006, 2007, Hayat et al. 2007, Mobin and Khan 2007, Anjum et al.
2008a, Singh et al. 2008a). Deleterious effects of Cd on various facets of
photosynthesis such as, gas exchange, biosynthesis of chlorophyll, functioning of
photochemical reactions and the activities of the enzymes of Calvin cycle have been
studied (Stobart et al. 1985, Weigel 1985, Padmaja et al. 1990, de Fillippis and
Zeigler 1993, Chugh and Sawhney 1999, Verma and Dubey 2002, Vassilev et al.
2005, Mobin and Khan 2007). In addition, conductance and index of stomata,
transpiration and net CO2 uptake are greatly reduced with elevated Cd levels in the
growth media (Bindhu and Bera 2001, Balakhnina et al. 2005). Both the PSI and PSII
have been shown to be affected by Cd. Photosystem II is highly sensitive to
deleterious effect of Cd and its functioning is inhibited to a greater extent than that of
PSI (Chugh and Sawhney 1999). Inhibition of PSII might be due to Cd-induced
alterations in the level of magno-protein (Mg-protein) in the water splitting system.
Cd restricts the PSII-related electron transport (Baszinsky et al. 1980) probably as a
result of the structural and functional changes in thylakoid membranes, the reduced
ferredoxin-NADP* oxidoreductase activity, and the arrested plastoquinine synthesis
(Krupa and Baszynski 1995). The Cd stress mostly causes metal inhibition of
Rubisco, an important enzyme of the Calvin cycle (Vassilev et al. 2005, Mobin and
Khan 2007). As observed in the study, the decrease in net photosynthetic rate has also
been attributed to the reduction in carbonic anhydrase activity under Cd stress
(Siedlecka and Krupa 1999, Mobin and Khan 2007, Singh et al. 2008a). The
inhibitory effects of Cd might be either due to its interaction with -SH group, as two
molecules of -SH group are found in enzyme center (on cys 173 and cys 456)
90

essential for normal activity of Rubisco (Lorimer 1981), and/or metal substitution
property of Cd leading to the decrease in ratio of combined activity of
carboxylation/oxygenation.
In Brassica juncea cultivars, 150mg Cd kg"' soil maximally reduced the
stomatal conductance and the maximum decrease was noted in RH30 and minimum in
Alankar (Figure 16). The loss of water from crop plants is controlled mainly by the
stomata on leaves that have been shown sensitive to Cd (Barcelo et al. 1986a,b,). To
minimize the water loss the process of stomatal closure is adopted by plants exposed
to various stresses affecting plant-water status. It has been identified as an early event
in plant response to Cd-induced water deficiency leading to limitations to C uptake by
leaves (Barcelo et al. 1986a,b, Poschenreider et al. 1989). Cd-induced alteration in
mineral nutrients may also contribute to the reductions in stomatal conductance which
in turn lead to suppression of major physiological processes and metabolic activities
as well.
Carbonic anhydrase activity in all Brassica juncea cultivars also decreased in
the similar manner as photosynthesis and stomatal conductance. Maximum decrease
was noted with 150mg Cd kg"' soil. Among cultivars the decrease in its activity was
maximum in RH30, whereas, Alankar experienced minimum decrease (Figure 15). As
observed in this study, the decrease in carbonic anhydrase activity was also reported
previously in other Cd treated plants (Aravind and Prasad 2004, Hasan et al. 2007,
Khan et al. 2007a, Mobin and Khan 2007, Singh et al. 2008a). Siedlecka and Krupa
(1999) also reported Cd-induced decrease in carbonic anhydrase activity and other
elements of Rubisco activation system. Cadmiimi (like several other heavy metals) is
known to affect the structure and functioning of enzymes (Van Assche and Clijsters
1990) through peroxidation and production of ROS (Dietz et al. 1999) which would
affect the functioning of the enzymes and proteins. ROS induce fragmentation of
protein and impose oxidative modification, rendering cells susceptible to enzymatic
proteolysis and hydrolysis. Carbonic anhydrase is in fact a Zn metalloprotein
catalyzing reversible inter-conversion of HCO3" and CO2 (Xin Bin et al. 2001, Khan
et al. 2004); represents 1-2% of total soluble protein in leaf (Okabe et al. 1984) and is
related to photosynthesis in higher plants (Khan 1994, Henry 1996). Cd can readily
inhibit most of the Zn-dependent processes, either by displacement or by occupying
the active sites of the Zn metalloproteins (Nieboer and Richardson 1980, Siedlecka
1995). Aravind and Prasad (2004) noted toxic effect of Cd on carbonic anhydrase
91

activity in Ceratophyllum demersum which proved the concept that Cd occupies the
active sites of important Zn-metalloproteins. The study also showed a marked
reduction in Zn content and its substitution by Cd impaired the structure as well as the
activity of carbonic anhydrase in Ceratophyllum demersum. Consequently, this leads
to nonfunctioning of carbonic anhydrase and hence the associated photosynthetic
processes were impaired.
5.1.3 Cadmium accumulation
In the present study, plants grown with increasing levels of Cd exhibited an increase
in Cd-concentrations in root and leaf of all Brassica juncea cultivars at all growth
stages. Among cultivars, RH30 accumulated maximum Cd in root and leaf, whereas,
Alankar accumulated less (Figures 17-19). The results obtained in this study suggest
that roots are efficient barriers to Cd translocation to the aboveground plant parts.
Accumulation of heavy metals by plants and their restriction at root level is a
widespread phenomenon. The retention of Cd in roots may be due to cross linking of
Cd to carboxyl groups of cell wall proteins (Barcelo and Poschenreider 1990) and/or
an interaction of Cd ions with the thiol groups of soluble proteins and non-protein
thiols operating as a tolerance mechanism in root cells (Chaoui et al. 1997a). In some
cases, Cd ions may also be bound by pectic sites and hystidyl groups of cell wall
(Leita et al. 1995). Ranieri et al. (2005) suggested that retention/immobilization of
high amount of Cd in root tissues typical of several plants can be regarded as an
important protection mechanism against the diffusion of the metal in plants. For this
reason, Cd-concentration in roots can reach to an average of 75-80% of the total metal
taken up (Wojcik and Tukiendorf 1999). The movement of Cd fi-om roots to shoots is
also likely to occur via the xylem and is driven by the transpiration from the leaves. In
this regard the closure of stomata may reduce the accumulation or retention of Cd in
root (Sah et al. 1995). In addition, reduced movement of Cd from roots to shoots in
plants is believed to result from barrier function of root endodermis and mechanism
involving sequestration and decreased xylem loading of Cd (Hart et al. 1998).
However, the immobilization of Cd at root level is primarily dependent on the
concentration of Cd supplied and the plant species (Sanita di Toppi and Gabbrielli
1999). Due to disturbed root function(s) and/or the reduction in the selectivity of
plasma membrane root-Cd may be translocated to the aboveground parts (Vassilev et
al. 1998). In the present study, the translocation of Cd from root to leaf was found Cd-
dose dependent and higher in cultivar RH30 than in Alankar (Figuresl7-18). In fact,
92

restriction of heavy-metal transport from root to shoot has been thought as a
mechanism of plant tolerance to Cd. The lower translocation of Cd from root to leaf
may, therefore, be a strategy of Alankar to protect its photosynthetic function from
Cd-induced oxidative stress which is in close conformity with Dixit et al. (2001),
Anjum et al. (2008a) and Singh et al. (2008a).
The uptake of Cd by plants varies not only among plant species but also
among cultivars (Salt et al. 1995, Arao et al. 2003, Metwally et al. 2005, Khan et al.
2006, Anjum et al. 2008c). The difference in the ability of cultivars to accumulate Cd
may also be related to differences in their root morphology (An 2004). Das et al.
(1997) suggested that plant with numerous thin roots would accumulate more metals
than one with few thick roots. The transport of Cd in plants has been found highly
species dependent. Cucumis sativus retained greatest amount of Cd in roots followed
by Triticum aestivum and Zea mays (An 2004). Differences in the net uptake of Cd in
these cultivars may be the result of differences in the electrochemical gradient that is
created by ion exchange reactions in the membrane structures as reported in the foliar
uptake of metals by Martin and Juniper (1970). A lower electrochemical gradient in
Alankar (tolerant cultivar) than in the RH30 (non-tolerant) could, therefore, be one of
the reasons for lesser uptake of Cd in Alankar.
5.1.4 Yield characteristics
Cadmium stress affects growth and yield through disturbances in several morpho-
physiological processes and nutrient uptake. Reduction in growth and yield with
increased levels of Cd in growth media arises because of increased leaf rolling and
chlorosis of leaf and stem (Ghani and Wahid 2007) and reduced photosynthetic rate
(Chugh and Sawhney 1999, Khan et al. 2006, 2007a). Like growth and photosynthetic
characteristics, yield and its attributes were decreased in mustard cultivars with Cd
treatments. The maximum reduction was noted with 150mg kg'' soil in all the
cultivars. The tolerant cultivar, Alankar exhibited minimum decrease in yield
characteristics (number of siliqua per plant, number of seeds per siliqua, seed yield
per plant and 1000 seed weight) while maximum decrease in these characteristics was
observed in non-tolerant cultivar RH30 (Figures 20-21). In the present study, decrease
in yield and its attributes in Brassica juncea cultivars treated with Cd is possibly the
fall out of the poor growth and photosynthesis. The decrease in photosynthesis may be
considered as one of the important factors responsible for reduced plant growth and
productivity under Cd stress (Khan et al. 2007a). Inhibition of photosynthetic pigment
93

and its biosynthesis is one of the primary events in plants under Cd stress condition.
As a consequence of delay in the assembly of the photosynthetic apparatus, lower
photosynthetic efficiency, slower plant growth and decreased biomass production
occurs that leads to the reduced yield.
Wahid and Ghani (2007) reported significant reduction in number of pods per
plant and seeds per pod, 100 seed weight, seed yield and harvest index of Vigna
radiata genotypes as a result of Cd toxicity. It has been suggested that although
varietal difference exists, the accumulated Cd is mainly toxic to the mesophyll tissue,
most probably by interfering with the uptake of essential nutrients, thereby reducing
growth and yield at various stages. Cadmium significantly reduced number of ear, ear
weight, ear length, spikelet number, grains per ear, 1000 grain weight and grain yield
of Triticum aestivum cultivars and the decrease was correlated with photosynthetic
capacity of the cultivars (Khan et al. 2006, 2007a). Genotypic differences on the basis
of biomass production, yield and yield components were observed in Triticum
aestivum and it was noted that Cd significantly reduced the root and stem biomass and
spikes per plant but grains per ear and grain weight were not significantly reduced
(Zhang et al. 2002). Liu et al. (2007) noted variation among Oryza sativa cultivars in
their tolerance to soil Cd stress with respect to tillering, plant height, leaf area, dry
matter accumulation and grain yield. The relative change in the number of grains per
panicle showed a strong positive correlation with relative change in grain yield and, of
the grain yield components measured (panicles per pot, grains per panicle, filled grain
percentage, weight per grain) and the reduction of grains per panicle was noticed as
the main cause of grain yield loss under soil Cd stress. Wu et al. (2004) reported
reduction in yield of three Gossypium hirsutum genotypes under Cd stress and found
that the reduction in yield was proportional to Cd accumulation. Among the
Gossypium hirsutum cultivars tested, the cultivar Simian 3 showed higher Cd
concentration and greater decrease in lint yield than the other two genotypes
(Zhongmian 16 and Zhonmian 16-2).
Taken together, the present study revealed that all the Brassica juncea
cultivars responded differentially to Cd stress and the severity of Cd stress in terms of
decrease in the growth, photosynthetic and yield characteristics was minimum in
cultivar Alankar followed by Varuna, Pusa Bold, Sakha and RH30. Contrarily, in
terms of the accumulation of Cd in root and leaf, RH30 accumulated maximum
94

followed by Sakha, Pusa Bold, Varuna and Alankar. On the basis of overall
performance of all Brassica juncea cultivars under Cd stress, Alankar proved as Cd-
tolerant and showed lesser decrease in the characteristics, whereas, RH30 emerged as
Cd non-tolerant and suffered maximum decrease.
5.2 Alleviation of Cadmium-toxicity by Sulfur and Nitrogen in Alankar (Cd
tolerant) and RH30 (Cd non-tolerant) Cultivars of Mustard
Increasing evidences suggest that mineral nutrient status of plants plays significant
role in increasing plant resistance to heavy metal stress (Marschner 1995, Astolfi et
al. 2004, Ahmad et al. 2005, Vassilev et al. 2006, Hassan et al. 2005a,b, 2008a,b,
Bouranis et al. 2008, Anjum et al. 2008a,b). Among the plant nutrients, S and N are
of great importance and are required by plants for maintaining normal growth and
development (Marschner 1995). Several studies have established regulatory
interactions between assimilatory sulfate and nitrate reduction in plants (Takahashi
and Saito 1996, Ahmad et al. 1999). The two pathways are very well coordinated
(Brunold 1993, Koprivova et al. 2000) and the deprivation of one leads to reduction in
the metabolism of the other (Reuveny et al. 1980, Prosser et al. 2001).
Positive interaction between S and N results in increased crop productivity
(Abdin et al. 2003). Brassica genotypes require higher amounts of S in addition to N
for optimum growth and yield (Abdin et al. 2003, Aulakh 2003, Khan et al. 2005).
Application of S along with N leads to enhanced biomass production and increased
leaf area as both nutrients are involved in biosynthesis of proteins and several other
molecules. Nitrogen is a basic constituent of proteins and with the increase in the rate
of N application, the N availability increases. Similarly, increased S supply increases
seed yield with higher protein content. Combined application of S and N promotes the
uptake of S and N, which leads to significant enhancement in seed protein and oil
content in Brassica juncea and Brassica campestris (McGrath and Zhao 1996,
Kachroo and Kumar 1997, Ahmad and Abdin 2000a,b, Abdin et al. 2003). Sulfur and
N relationship in terms of crop productivity has also been established in many studies
(Singh and Bairathi 1980, Sachdev and Deb 1990, Lakkineni and Abrol 1992,
McGrath and Zhao 1996, Ahmad et al. 1998, Fismes et al. 2000).
Experiments 2 and 3 were conducted where two cultivars of Brassica Juncea
namely, Alankar (Cd tolerant) and RH30 (Cd non-tolerant) (screened out from the
Experiment 1) were given S alone and in combination with N to ameliorate Cd-
95

induced toxicity. In Experiment 2 both the cuhivars of Brassica juncea were treated
with 0, 50 and 150mg Cd kg'' soil and supplemented with 0, 50 and lOOmg S kg"'
soil. Application of 50mg S kg'' soil to 50mg Cd kg'' soil-treated Alankar plants
overcome the Cd-induced toxicity whereas, in RH30, S-supplementation only lowered
the severity of Cd toxicity. Therefore, in Experiment 3, 50mg Cd kg'' soil-treated
cultivars were given 50mg S kg'' soil along with different levels of N (0, 40, 80 or
120mg N kg'' soil) to strengthen the S induced tolerance. In Experiment 3, it was
found that combined application of 50mg S kg'' soil and 80mg N kg'' soil completely
overcome the ill effects of 50mg Cd kg'' soil in Alankar (tolerant) whereas, in RH30
(non-tolerant), this combination only lowered the Cd-induced toxicity. The following
section discuses major results of the Experiments 2 and 3 with relevant supporting
citations. In addition, the participation of S and N in plant tolerance mechanisms has
also been discussed.
5.2.1 Growth characteristics
In Experiment 2, application of 50mg S kg'' soil ameliorated the Cd-induced toxicity
and improved the growth characteristics of tolerant cultivar Alankar but lowered the
reductions in non-tolerant cultivar RH30. The application of 50mg S kg'' soil nullified
the effects of Cd, to a lesser extent, in the plants fed with higher concentrations of Cd
(150mg Cd kg'' soil) but completely, if given low level of Cd (50mg Cd kg''). In
particular, the application of 50mg S kg'' soil proved most effective in improving
growth characteristics of both the Brassica juncea cultivars treated with 50mg Cd kg''
soil. A significant increase in root length, shoot length, plant dry mass and leaf area
was observed with 50mg S kg'' soil which varied greatly between cultivars grown
with 50mg Cd kg"' soil (Figures 22-25). Thus, application of S was instrumental in
mitigating to some extent the toxic effects of Cd on plant growth characteristics. This
indicates that S application reduces the toxic effects of Cd on plant through
improvement in the tolerance capacity of the plant. S supplementation to plants has
been shown to result in greater biomass production under normal (Ahmad et al. 2005)
and Cd stress conditions (Hassan et al. 2005b, Anjum et al. 2008a). However, the
mitigating effect by S nutrition was found dependent on Cd dose and on Brassica
juncea cultivar. Between the cultivars, the ameliorative effect of S nutrition was found
maximum in Alankar while minimum effect was noted in RH30. Our findings on
ameliorative effect of S in Cd-induced growth reductions are in agreement with
Anjum et al. (2008a). Anjana et al. (2006) reported that S could alleviate the Cd
96

induced impairment of biochemical and anatomical features of the Brassica
campestris plant. Hassan et al. (2005b) also showed that S application ameliorated the
Cd-induced inhibition of growth parameters in two Oryza saliva cultivars and the
effect of S treatment on these characteristics varied greatly with Cd level and between
cultivars.
In the present study, combined application of S and N further increased the
growth characteristics of Cd-treated Brassica juncea cultivars Alankar (tolerant) and
RH30 (non-tolerant) (Figures 48-51). A combination of 50mg S kg'' soil plus 80mg N
kg'' soil proved superior in enhancing the growth characteristics of both the cultivars
treated with 50mg Cd kg'' soil (Figures 50-51). S and N are essential macro nutrients
required for normal growth and development of plants (Marschner 1995) and have
been found to play a pivotal role in the protection of plants against environmental
stresses, including Cd toxicity (Pankovic et al. 2000, Hassan et al. 2005a,b, 2006,
2008a,b, Anjum et al. 2008a,b). It has also been found in previous studies that the
coordination of S and N results in enhanced crop yield and quality of S-requiring
Brassica (Aulakh et al. 1980, Lakkineni and Abrol 1992, 1994, Abdin et al. 2003,
Aulakh 2003). In the present study, the application of S along with N led to enhanced
biomass production and increased leaf area of Alankar than RH30 under Cd stress. It
is suggested that combined application of S and N promotes the uptake of S and N,
which leads to significant enhancement in growth of plants (Ahmad and Abdin
2000a,b, Abdin et al. 2003). Another explanation is that both nutrients are involved in
the synthesis of amino acids, proteins and various other cellular components,
including thiol compounds and the so-called secondary S compounds which have a
significant role in the protection of plants under stressed conditions. Moreover, N is a
basic constituent of proteins and the N availability increases with the increase in the
rate of N application. It has been shown in Hordeum vulgare that high levels of nitrate
and ammonium can induce a high affinity sulfate transporter gene and hence sulfate
uptake in N-fed plants suggesting that N metabolite may affect sulfate transporter
gene expression (Vidmar et al. 1999) and hence leads to enhancement in growth
characteristics.
5.2.2 Photosynthetic characteristics
All the Cd levels used in the present study significantly reduced the photosynthetic
characteristics. The effect of Cd stress on these characteristics also varied between
cultivars. Non-tolerant cultivar RH30 suffered maximum decrease in photosynthetic
97

characteristics, whereas, tolerant cultivar Alankar showed minimum decrease (Figures
26-31). More severe decreases of photosynthetic characteristics in RH30 than in
Alankar under Cd stress may partially account for the differential responses of growth
parameters between these two cultivars grown under Cd stress condition. Earlier
studies have also showed that photosynthetic functions are highly sensitive to heavy
metals including Cd (Zhang et al. 2003). Cd inhibits the enzymes of the Calvin cycle,
biosynthesis of chlorophyll and accessory pigments and declines the activity of
Rubisco (Siedlecka et al. 1997, Mobin and Khan 2007). In the present study, Cd
treatment declined the net photosynthetic rate, stomatal conductance and carbonic
anhydrase activity with the increase in the concentrations of Cd in the soil (Figures
27-28, 30-31). Moreover, Cd-induced reductions in photosynthetic responses have
been found to involve both stomatal and non-stomatal limitations (Mobin and Khan
2007). Contrarily, Di Cagno etal. (2001) observed no change in stomatal conductance
but noted significant reductions in CO2 assimilation rate and Rubisco activity in Cd-
exposed Helianthus annuus. Ali et al. (2000) reported that the decrease in the rate of
photosynthesis was associated with decreased stomatal conductance and transpiration
in in vitro grown Bacopa monniera plants treated with Cd. In the present
investigation, Cd significantly decreased both the net photosynthetic rate and stomatal
conductance in tolerant (Alankar) and non-tolerant (RH30) cultivars; suggests that
stomatal functions limit the photosynthesis.
Application of 50mg S kg"' soil ameliorated the Cd-induced toxicity and
improved the photosynthetic characteristics of both the tolerant (Alankar) and non-
tolerant (RH30) cultivars (Figures 26-28). In comparison to lOOmg S kg'' soil,
application of 50mg S kg'' soil ameliorated the Cd-induced (50mg Cd kg'' soil)
reductions in photosynthetic characteristics of both the cultivars (Figures 29-31). In
Triticum aestivum, an efficient S assimilation and antioxidative system was helpful in
protecting the photosynthetic ability and maintaining high yield potential under Cd
stress (Khan et al. 2007a). Resurreccion et al. (2001) reported that S application
increased the chlorophyll and Rubisco content which led to increased photosynthesis
of Oryza sativa. Khan et al. (2005) has also reported that S application increased the
relative growth rate, plant growth rate, net assimilation rate and carbon dioxide
exchange rate of Brassica juncea plants. Anjana et al. (2006) reported that the
reductions caused by lower level of Cd in leaf dry weight, chlorophyll, sugar and
protein content were reversed by S treatment in Brassica campestris. At the cellular
98

level S starvation reduces the mesophyll cell number per cm^ and the chlorophyll
content per chloroplast. The photosynthetic CO2 uptake by intact leaves,
photoreduction of ferricyanide, cyclic and non-cyclic photophosphorylation of
isolated chloroplast and the rate of CO2 assimilation by Rubisco may decrease with
the decreased total S content (Terry 1976), showing the importance of S in plant
metabolism. Chloroplasts contain proteins rich in S (Hanson et al. 1941), and
chloroplast morphology is considerably affected by S (Repka et al. 1971, Whatley
1971, Hall et al. 1972). The decreased sink strength in turn could lead to a feedback
inhibition of photosynthetic rate and a decrease in Rubisco synthesis (Krapp and Stitt
1995, Sexton et al. 1997). Further, the stresses usually were shown to enhance the
process of degradation of cellular proteins, also reduce Rubisco (Sheoran et al.
1990a,b, Stiborova et al. 1986), the major source of S-concentration in the chloroplast
(Ferreira and Teixeira 1992). Thus, Cd usually interacts with -SH group of Rubisco
and degrades them (Majumdar et al. 1991). Ferreira and Teixeira (1992) have shown
lesser number of-SH groups, the major form of reduced S in Rubisco, isolated from
S-deprived plants. Further, Cd-induced reductions in the rate of photosynthesis in S-
deprived plants might also be due to restricted use of ATP and NADPH by the Calvin
cycle enzymes (Dietz and Heilos 1990) and decrease in phosphorylation capacity
(Becerril et al. 1989). Nitrogen is one of the essential nutrient elements with a
remarkable effect on crop growth and productivity (Marshner 1995). Sugiharto et al.
(1990) found a significant positive correlation between the photosynthetic capacity of
leaves and their leaf N concentration suggesting that most of the N is used for the
synthesis of components of the photosynthetic apparatus. In particular, Rubisco, the
leaf protein playing the major role in carbon assimilation, was strongly affected by N
deficiency (Seemann et al. 1987).
The decrease in stomatal conductance was possibly due to the fact that cell
growth depends primarily on turgor potential as the driving force of expansion, and
Cd affects both turgor potential and cell wall elasticity (Barcelo et al. 1986b). On the
other hand, S may help in maintaining the water status of the plant by enhancing the
root growth and thereby reducing the stress condition imposed by Cd. S availability
regulates N utilization efficiency of plants and, thus photosynthesis, growth and dry
mass accumulation of crops since photosynthates accumulation has a close
relationship with N and S assimilation (Tandon 1995, Kopriva et al. 2002, Khan et al.
2005).
99

5.2.3 Sulfur and nitrogen metabolism
In the present study the parameters of S and N assimilation responded differentially to
Cd stress. Cd stress increased the ATP-sulftirylase activity and S content, whereas,
NR activity and N content were decreased in both tolerant (Alankar) and non-tolerant
(RH30) cultivars at all growth stages (Figures 32-35). ATP-sulfurylase activity and S
content were higher in Alankar than in RH30. Cd-induced increase in ATP-
sulfurylase could be a possible consequence of an increased requirement of
glutathione for the biosynthesis of phytochelatins. The capacity of plants to survive in
a polluted environment is partially linked to the efficiency of their reductive sulfate
assimilation pathway and its induction has been reported in several plants exposed to
heavy metals (Tukiendorf and Rauser 1990). Expression of genes involved in
reductive sulfate assimilation pathway and enzyme activities are stimulated by Cd
(Ernst et al. 2008). Cd-induced induction of enzymes of sulfate assimilation pathway
has been observed in Arabidopsis thaliana (Harada et al. 2002, Herbette et al. 2006),
Chlamydomonas reinhardtii (Dominguez et al. 2003) and Triticum aestivum (Khan et
al. 2007a). Nussbaum et al. (1988) reported that the accumulation of Cd increased
ATP-sulfurylase activity in Zea mays seedlings. Ruegsegger et al. (1990) also showed
that APS reductase activity is induced coordinately with glutathione synthetase in Cd-
treated Pisum sativum plants. Cd-induced decrease in the NR activity of both the
cultivars is in conformity with Gouia et al. (2000), Chaffei et al. (2003), Anjana et al.
(2006), Ghnaya et al. (2007), Wahid et al. (2007) and Wang et al. (2008). Cd reduces
absorption of nitrate and its transport from root to shoot by inhibiting NR activity in
shoots (Hernandez et al. 1996). Wahid et al. (2007) reported that Cd-induced
inhibition of growth was mainly due to the damaged photosynthetic apparatus and
disruption of the coordination between C and N metabolism in Vigna radiata. In the
present study a consistent relationship between photosynthesis and NR activity was
observed in both the cultivars. Since deprivation of CO2 causes inactivation of NR
(Kaiser and Behnisch 1991), it is not surprising that Cd decreased the NR activation
state. On the other hand, Gouia et al. (2000) have reported that Cd not only decrease
the uptake and transport of water and nitrate but also cause a decrease in NR
activation state and 80% decrease in NR protein level in short term exposure (24h) in
Phaseolus vulgaris plants. Barcelo and Poschenrieder (1990) provided direct reasons
for Cd-induced changes in N-metabolism showing that Cd inhibits the transport and
uptake of water. They proved that the accumulation of Cd in roots alters the process
100

of uptake and transport of water and nitrate to the shoot. Furthermore, the uptake of
nitrate declined under Cd stress (Hernandez et al. 1996, 1997, Ouariti et al. 1997) that
possibly lowered NR activity and disturbed N metabolism.
The application of S further increased the ATP-sulfurylase activity and S
content of Cd-treated plants but the extent of increase was greater in the tolerant
cultivar Alankar than non-tolerant cultivar RH30. Maximum significant increase was
noted in plants supplemented with 50mg S kg'' soil in 50mg Cd kg'' soil treated
plants (Figures 32-35). Application of 50mg S kg'' soil with different levels of N (40,
80 and 120mg N kg"' soil) again increased the ATP-sulfurylase activity and S content
in both the cultivars treated with 50mg Cd kg'' soil at all growth stages (Figures
58,60). Our results suggest that maximum activity of ATP-sulfurylase can only be
achieved at a suitable S and N supply (50mg S kg'' soil + 80mg N kg'' soil) to the
plants. The involvement of S in Cd-tolerance mechanism was reported in Arabidopsis
thaliana (Dominguez-Solis et al. 2001, Harada et al. 2002), Brassica Juncea (Zhu et
al. 1999a,b), Nicotiana tabacum (Harada et al. 2001), Triticum aestivum (Khan et al.
2007a) and Brassica campestris (Anjana et al. 2006, Anjum et al. 2008a). A good part
of S incorporated into organic molecules in plants is located in thiol (-SH) groups in
proteins (cys-residues) or non-protein thiols, glutathione (Noji and Saito 2003, Tausz
et al. 2003, De Kok et al. 2005, Anjum et al. 2008a). Chen and Huerta (1997) showed
that S is a critical nutritional factor for reduction of Cd toxicity. Popovic et al. (1996)
reported a positive effect of S nutrition on Cd-detoxification in Beta vulgaris. In fact
Cd stress provokes plants to enhance the biosynthesis of glutathione for the synthesis
of phytochelatins (Herbette et al. 2006). Besides, it has been previously studied that
good S nutrition diminished the toxicity of Cd by restoring a new steady state of the
glutathione level earlier than in plants grown at low S supply (McMahon and
Anderson 1998). Hence, the improved S nutrition allows a more adequate plant
defense response to Cd toxicity and also prevents S deficiency. It has been found that
at sub-optimal S nutrition, Cd exposed plants preferably allocate S to phytochelatins
synthesis. Sulfur being a component of phytochelatins may play an important role in
their synthesis and ultimately in detoxification of Cd through the formation of Cd-
binding peptides (CdBP) (Cobbett 2000a,b, Harada et al. 2002, Cobbett and
Goldsbrough 2002). Exposing the heavy metal-accumulator Brassica juncea to Cd
was reported to induce a rapid synthesis of phytochelatins, which appeared to be
sufficient to chelate all the Cd taken up, presumably resulting in a complete
101

preservation of the photosynthetic activities (Haag-Kerwer et al. 1999). Several
studies have also demonstrated the critical role of phytochelatins in Cd detoxification
and tolerance of plants to Cd (Inouhe et al. 2000, Harada et al. 2001, 2002, Ebbs et al.
2002, Srivastava et al. 2004, Wojcik et al. 2005, Zhang et al. 2008, Anjum et al.
2008b).
The reductions in NR activity and N content were reversed by S application in
Cd treated Alankar (tolerant) and RH30 (non-tolerant) cultivars (Figures 33-35).
Lesser decrease in NR activity and N content was observed when plants were given
50mg S kg"' soil plus 50mg Cd kg'' soil in comparison to 50mg Cd kg'' soil alone.
The combined application of 80mg N kg'' soil plus 50mg S kg"' soil to 50mg Cd kg''
soil-treated plants maximally lowered the reductions in NR activity and N content in
both the cultivars at all growth stages (Figures 59,61). The negative effects of heavy
metals on NR activity may be due to interaction of metals with thiol/histidyl groups of
proteins, thereby slowing down their catalytic activity and/or completely inhibiting
their functions (Wang and Evangelou 1995). As a resuh, enzymatic reactions may be
blocked, leading to accumulation of nitrate in plants and disturbing the normal plant
metabolism. Also, it may be due to interference with the uptake of nutrients and/or
induction of leakage of nutrients by damaging plasma membrane and protein
alterations due to heavy metal toxicity in leaves (Brune et al. 1994). However, the
beneficial effect of S on NR activity is primarily due to its being a component of
various enzyme proteins, cofactors, and metabolites (Sairam et al. 1995). S
application results in adequate N uptake and in high protein yields in Brassica juncea
and Triticum aestivum (Purakayastha and Nad 1997). Therefore, plants supplied with
S showed an increased NR activity and improved N assimilation as a result of
balanced supply of both N and S, ultimately leading to increased protein formation. It
may also be due to the role of S in increasing S-amino acids (Met and Cys) (Bapet et
al. 1986). N metabolism, therefore, may act as a marker for the study of the response
of plants to Cd toxicity. Upon exposure to Cd, plants often synthesize a set of N-
containing metabolites through N metabolism, such as proline, glutathione and
phytochelatins, which are well known for their significant roles in Cd tolerance of
plants (Sharma and Dietz 2006). Accordingly, plants might exhibit a higher Cd
tolerance by the maintaining the normaJ N metabolism levels under Cd stress
(Gussarsson et al. 1996). The ameliorative effect of S in recovering the NR activity as
102

obtained in our results, are in agreement with the findings of Friedrich and Schrader
(1978), Prosser et al. (2001) and Astolfi et al. (2004).
Combined application of S and N increased the activity of ATP-sulfurylase
while improved the NR activity of Cd treated plants.
5.2.4 Cadmium accumulation
In the present study, Cd content in the root and leaf of both the cultivars increased
with the increasing Cd concentration in soil. Non-tolerant cultivar RH30 accumulated
more Cd in root and leaf than tolerant cultivar Alankar. In both the cultivars,
increased accumulation of Cd in root and leaf due to 50mg Cd kg"' soil was lowered
with the application of 50mg S kg'' soil alone and in combination with 40, 80 and
120mg N kg"' soil at all growth stages. However, combined application of 50mg S kg"
' soil and 80mg N kg'' soil to 50mg Cd kg"' soil-treated plants maximally lowered the
Cd content in root and leaf of both the tolerant (Alankar) and non-tolerant (RH30)
cultivars at all growth stages (Figures 36-37,62-63).
The differences in Cd accumulation among cultivars have previously been
reported (Salt et al. 1995, Arao et al. 2003, Metwally et al. 2005, Khan et al. 2006,
Anjum et al. 2008c). In the present study, roots of both the cultivars accumulated
more Cd than leaves. The retention of high amount of Cd in the root tissues typical of
several plants can be regarded as an important protection mechanism against the
diffusion of the metal (Ranieri et al. 2005). For this reason, Cd-concentration in roots
can reach an average of 75-80% of the total metal taken up (Wojcik and Tukiendorf
1999). Ranieri et al. (2005) have reported that the highest level of Cd was retained in
roots. In the present study, S application reduced the uptake and accumulation of Cd
in both the plant parts. A combined application of S and N also reduced the
concentration of Cd in root and leaf of both the cultivars. Pankovic et al. (2000) found
the lowest inhibition of photosynthetic activity by Cd at optimal N supply, when an
investment in soluble proteins and Rubisco were at their maximum and that higher N
supplies did not alleviate the toxic Cd effects. A proper N supply has positive effects
in overcoming the adverse effects caused by Cd toxicity in Oryza sativa (Hassan et al.
2005a, Pankovic et al. 2000). In addition, S may reduce Cd availability in nutrient
solution through binding with Cd and detoxify Cd in plant cells through enhancing
synthesis of glutathione, which is considered as a first line of defense against Cd
toxicity (Hassan et al. 2005b, Anjum et al. 2008a).
103

5.2.5 Cadmium-induced oxidative stress
Heavy metals, in general, cause toxicity and cellular disruption in plant species
through induction of oxidative stress via increased generation of ROS, like O2, O2 ,
OH' and H2O2 (Gratao et al. 2005, Singh et al. 2008b). In the present study, Cd-
induced oxidative stress in Brassica juncea cultivars is evident from the significant
increases in TBARS and H2O2 contents with increasing Cd concentration. However,
compared to the non-tolerant cultivar RH30, tolerant cultivar Alankar showed a much
lower H2O2 content in leaves and TBARS production, which are indicative of lower
oxidative stress in Alankar. Less Cd accumulation and lower level of H2O2 content,
together with the reduced formation of TBARS, thus explain the higher potential of
Alankar than RH30 (Figures 36-39). Malondialdehyde, as the decomposition product
of polyxmsaturated fatty acids of bio-membranes showed greater accumulation under
Cd stress in other studies also (Qadir et al. 2004, Metwally et al. 2005, Liu et al.
2007, Anjum et al. 2008a, Singh et al. 2008a). Cell membrane stability has been
widely used to differentiate stress tolerant and susceptible cultivars of some crops and
that higher membrane stability can be correlated with Cd stress tolerance (Mobin and
Khan 2007). The enhanced cellular damage in RH30 seems to reflect the deterioration
on the equilibrium between generation of ROS and defense mechanisms towards
removal of ROS. Increase in TBARS with increasing Cd concentration has also been
reported in germinating Phaseolus vulgaris seedlings (Somashekaraiah et al. 1992).
This was related to restriction of electron flow in PSII by metal ions that led to the
formation of excited chlorophyll which in tum caused the production of free radicals
(Kato and Shimizu 1985). Cd-induced increase in lipid peroxidation has also been
found earlier in different plants by Dixit et al. (2001), Chien et al. (2001), Groppa et
al. (2001), Wu et al. (2003), Balestrasse et al. (2004), Metwally et al. (2005), Guo et
al. (2007), Ahsan et al. (2007), Razinger et al. (2007), Mobin and Khan (2007),
Noriega et al. (2007), Liu et al. (2007), Ammar et al. (2007), Filek et al. (2007),
Anjum et al. (2008a) and Singh et al. (2008a).
H2O2 is produced at high flux rates by two processes associated with
photosynthesis, the Mehler reaction and the Glycolate oxidase reaction of
photorespiration (Foyer and Noctor 2000). In addition, there are number of other
enzymes in leaves that are capable of producing significant amounts of H2O2,
including peroxidases, NADPH oxidases and oxalate oxidase (Bema and Bernier
1999, Bolwell 1999, Sagi and Fluhr 2001). H2O2 is a relatively stable molecule that
104

can pass freely across the membranes and is capable of spreading damage
(Azpilicueta et al. 2007). Metabolic modeling has also suggested that the loss in
antioxidative defense was due to H2O2 accumulation (Polle 2001, Schutzendubel and
PoUe 2002). H2O2 generation is induced in plants following exposure to a wide
variety of stresses including Cd (Dixit et al. 2001, Schutzendubel et al. 2002,
Romero-Puertas et al. 2004, Cho and Seo 2005, Balestrasse et al. 2006b, Zawoznik et
al. 2007, Yang et al. 2007, Filek et al IQQl, Mobin and Khan 2007, Singh et al.
2008a). Also a pronounced increase in the activity of superoxide dismutase in RH30
with the increase in Cd levels possibly generated higher level of superoxide radicals
and resulted in higher cellular damage in comparison to Alankar. Gossett et al. (1994)
reported that higher superoxide dismutase activity without complementary increase in
the ability to scavenge the formed H2O2 can result in the increased cellular damage.
In the present study, application of 50mg S kg'' soil maximally lowered the
TBARS and H2O2 content of 50mg Cd kg'' soil-treated plants. Application of 50mg S
kg"' soil with different levels of N (40, 80 and 120mg N kg'' soil) further lowered the
contents of TBARS and H2O2 of both the cultivars grown with 50mg Cd kg'' soil and
enhanced the effectiveness of S application (Figures 38-39,64-65). These results on
the significant reduction in TBARS content with S application is in conformity with
the findings of Anjum et al. (2008a) in Brassica campestris leaves. In addition, net
photosynthetic rate and plant dry mass were strongly and positively correlated with
the contents of ascorbate and glutathione. It was concluded that S may ameliorate Cd
toxicity and protects growth and photosynthesis of mustard involving ascorbate and
glutathione. Vassilev et al. (2005) also reported that supply of (NH4)2S04, a well
knovm fertilizer consisting both S and N nutrients, could reduce Cd toxicity in
Hordeum vulgare plants by lowering the TBARS content.
5.2.6 Components of antioxidant system
5.2.6.1 Enzymatic antioxidants
Oxidative stress is a central factor in abiotic stress phenomena that occurs when there
is a serious imbalance in any cell compartment between the production of ROS and
antioxidant defense, leading to physiological changes (Foyer and Noctor 2000,
Agrawal and Mishra 2008). The presence of high concentration of ROS causes
oxidative damage to photosynthetic pigments, bio-molecules such as lipids, proteins
and nucleic acids, leakage of electrolytes via lipid peroxidation resulting in the
disruption of cellular metabolism (Asada 1994, Mobin and Khan 2007). Thus, it is
105

important for cells to have tight control on the concentration of ROS (Schutzendubel
and PoUe 2002). Plants are in fact endowed with a wide array of defense strategies to
protect the photosynthetic apparatus and cellular functions from ROS (Foyer et al.
1994). Plant antioxidant defense system comprises antioxidant enzymes such as
superoxide dismutase, catalase, ascorbate peroxidase, glutathione reductase and low
molecular weight antioxidants (ascorbate; glutathione; proline; carotenoids, a-
tocopherols and phenolics etc.) (Foyer and Noctor 2003). These components help
plants to cope with the menace of ROS, thus minimizing the oxidative damages,
during exposure to stress factors including Cd (Foyer et al. 1994, Mobin and Khan
2007, Anjum et al. 2008a, Singh et al. 2008a,b). Changes in antioxidant enzyme
activities play an important role in metal tolerance. Though, the expression for
antioxidant enzymes is altered under stress conditions, their upregulation has a key
role in combating the abiotic stress-induced oxidative stress.
In my study, Cd treatments increased superoxide dismutase activity in both
cultivars, the extent of increase was higher in RH30 than Alankar (Figures 40,42).
Mobin and Khan (2007) also reported that higher superoxide dismutase activity in
Brassica juncea cv. RH30 was responsible for higher cellular damage due to
excessive accumulation of H2O2. It has been suggested that high SOD activity may be
harmful for plants due to high H2O2 production, which in turn might inhibit other
enzymes such as ascorbate peroxidase and catalase (Asada 1994, Agrawal and Mishra
2008). Cd-dependent induction of superoxide dismutase activity has also been
observed in earlier studies in other plant species (Dixit et al. 2001, Schutzendubel et
al. 2001, Vitoria et al. 2001, Qadir et al. 2004, Cho and Seo 2005, Metwally et al.
2005, Wu et al. 2006, Krantev et al. 2007, Ekmekci et al. 2008, Hasan et al. 2008,
Agrawal and Mishra 2008, Singh et al. 2008a). Increased superoxide dismutase
activity has also been shown to confer increased protection from oxidative damage in
transgenic plants (Allen et al. 1997). The activity of superoxide dismutase is of great
relevance in metal stress studies for the maintenance of overall defense system of
plants subjected to oxidative damage (Slooten et al. 1995).
Sulfur supplementation to Alankar (tolerant) and RH30 (non-tolerant) treated
with Cd lowered the superoxide dismutase activity (Figures 40,42). The application of
50mg S kg"' soil plus 80mg N kg"' soil further lowered the Cd-induced (50mg Cd kg"'
soil) increase in superoxide dismutase activity (Figures 66-68). The decrease in 50mg
Cd kg"' soil activity due to S and N application conferred to less formation of ROS.
106

The function of S in alleviating the stress may be attributed to its participation in the
synthesis of glutathione, an important ROS scavenger. Sulfur helped the plants to
maintain CO2 (whose absence is the major cause of 02"" production in the process of
photosynthesis) and major reductants (ascorbate, glutathione) in chloroplasts,
maintenance of the sulf-hydryl status of proteins; and hence diminished the chances of
O2' formation and ultimately provided protection from 02~. In a study conducted on
Oryza sativa cvs. Bing 97252 and Xiushui 63, Hassan et al. (2005b) found that higher
S levels caused significant reduction in superoxide dismutase activity in Cd treated
plants. They found that the reduction in superoxide dismutase activity was more
pronounced in Bing 97252 than in Xiushui 63. They concluded that high Cd and
MDA content were consistently accompanied by higher superoxide dismutase activity
and higher S levels caused a marked increase in GSH content and a reduction in SOD
activity, indicating a positive effect of S in alleviating Cd-induced oxidative stress.
Ajnum et al. (2008a) also reported that S supplementation alleviates the Cd-induced
oxidative stress in Brassica campestris plants. Hassan et al. (2008a) suggested that the
oxidative stress of Oryza sativa plants exposed to Cd toxicity could be alleviated
when (NH4)2S04 was supplied as N fertilizer. Moreover, it was also found that
(NH4)2S04-fed plants had higher GSH content under Cd stress.
Catalase activity increased with the increasing Cd concentration at all growth
stages in Alankar, whereas, in RH30, the increase was noted only at 30DAS (Figures
40-42). At later stages (60 and 90DAS), Cd significantly decreased the catalase
activity. In the present study the inhibition of catalase activity in non-tolerant cultivar
RH30 may be due to its inactivation by excess H2O2 produced by superoxide
dismutase under Cd stress. Agrawal and Rathore (2007) also reported that the decline
in catalase activity was due to more consumption of catalase to detoxify H2O2 or its
inactivation. The variable response of catalase activity has been observed under Cd
stress in different plant species. Catalase activity increased in Glycine max nodules
(Balestrasse et al. 2001), Oryza sativa leaves (Hsu and Kao 2004), Brassica
campestris leaves (Ajnum et al. 2008a), in tolerant varieties of Solarium tuberosum
(Stroinski and Kozlowska 1997), in roots ofRaphanus sativus seedlings (Vitoria et al.
2001), Brassica juncea (Mobin and Khan 2007), Triticum aestivum (Khan et al.
2007a) and Cicer arietinum (Hasan et al. 2008). However, catalase activity decreased
in Amaranthus lividus (Bhattacharjee 1998), Glycine max roots (Balestrasse et al.
2001), Phragmites australis (lannelli et al. 2002), Capsicum annuum (Leon et al.
107

2002) and Arabidopsis thaliana (Cho and Seo 2005) under Cd stress conditions.
Furthermore, catalase activity has also been found to remain unaltered under Cd stress
in Glycine max leaves (Ferreira et al. 2002).
Sulfur supplementation further increased the Cd-induced increase in catalase
activity in Alankar. In RH30, addition of S lowered the decrease in catalase activity
only at 60 and 90DAS, whereas, slight increase in its activity was noted at 30DAS. In
Alankar, the increase in catalase activity due to 50mg Cd kg'' soil was further
increased with the application of 50mg S kg'' soil alone and in combination with
80mg N kg'' soil at all growth stages (Figures 40,42,66,68). A combined application
of S and N increased the Cd-induced catalase activity in Alankar which is in close
conformity with the findings of Hassan et al. (2008a). They noted that (NH4)2S04 the
source of both S and N also increased the catalase activity of Cd treated Oryza sativa
plants.
In our study, Cd-dose dependent increase in ascorbate peroxidase activity was
observed in both the cultivars but its activity was greater in Alankar (tolerant) than
RH30 (non-tolerant) at all growth stages (Figures 41,43). Ascorbate peroxidase has an
important role in the scavenging of H2O2 under stressed conditions but its activity
depends on the Cd concentration applied (Mobin and Khan 2007, Singh et al.
2008a,b). Increased leaf ascorbate peroxidase activity under Cd stress has been
reported in Phaseolus aureus (Shaw 1995b), Phaseolus vulgaris (Chaoui et al.
1997b), Pisum sativum (Romero-Puertas et al. 1999), Ceratophyllum demersum
(Aravind and Prasad 2003), Brassica juncea (Mobin and Khan 2007), Zea mays
(Krantev et al. 2007), Triticum aestivum (Khan et al. 2007a), Brassica campestris
(Anjum et al. 2008a) and Vigna mungo (Singh et al. 2008a). The action of ascorbate
peroxidase may be correlated with the reductions in the total ascorbate content i.e.,
greater ascorbate peroxidase activity needed more substrate and hence consumed
more ascorbate as electron donor, thus causing a decline in the ascorbate
concentration in the present study. Similar variations in the ascorbate content were
observed by Schutzendubel et al. (2001) and Qadir et al. (2004) in Cd-exposed Pinus
sylvestyris and Brassica genotypes, respectively.
Sulfur supplementation further increased the ascorbate peroxidase activity of
both the cultivars and maximum significant increase in its activity was noted in plants
treated with 50mg Cd kg"' soil. Combination of 50mg S kg"' soil and 50mg Cd kg'
soil proved best in enhancing the ascorbate peroxidase activity. The increase in
108

ascorbate peroxidase activity due to 50mg Cd kg"' soil was further increased by the
application of 50mg S kg'' soil alone and in combination with 80mg N kg'' soil in
both the cultivars at all growth stages (Figures 41-42,67-68). The demand-driven
increase in the ascorbate peroxidase activity implies the existence of stress sensors
and signal transduction cascades that elicit the response of ascorbate peroxidase
activity using ascorbate as reductant. In fact, S supplementation to the Cd-exposed
plants helped to maintain the synthesis of electron providers (ascorbate and
glutathione), that improved the regeneration of ascorbate (Anjum et al. 2008a), an
electron source for ascorbate peroxidase. Thus, S alleviated the Cd stress alterations
and helped plants to continue the ascorbate peroxidase activity to maintain the
conversion of H2O2 into H2O and O2 by regenerating ascorbate and glutathione
efficiently. Role of glutathione as a signal intermediate in increasing ascorbate
peroxidase expression under metal stress has also been reported by Pekker et al.
(2002). The addition of S and N to Cd-fed plants increased the ascorbate peroxidase
activity in both the cultivars which may be correlated with the improvement in
ascorbate content by S and N.
Glutathione reductase activity of both the cultivars was increased under Cd
stress. Maximal significant increase in glutathione reductase activity was noted in
both the cultivars treated with 50mg Cd kg'' soil. Treatment of 150mg Cd kg ' soil
significantly decreased the glutathione reductase activity in non-tolerant cultivar
RH30 (Figures 41-42). The majority of the studies determining the response of
glutathione reductase to Cd exposure have shown that glutathione reductase activity
increases as part of the defense against the Cd-exposure, an alteration that has often
been dose-dependent and variable over time (Fomazier et al. 2002a,b).
The up regulation of glutathione reductase activity may resuh in the
improvement of abiotic stress tolerance by reducing oxidized glutathione (GSSG)
produced in Ascorbate-Glutathione cycle. Besides, S plays a crucial role in the
synthesis of Cys, a precursor molecule for the production of glutathione (Suter et al.
2000). Addition of 50mg S kg'' soil plus 80mg N kg"' soil further increased the
glutathione reductase activity of both the cultivars at all growth stages and extent of
increase was higher in Alankar than RH30 (Figures 41-42, 67-68). The enhancement
in glutathione reductase activity might be due to enhancement in the overexpression
of enzymes (y-ECS and GSH synthetase) in the chloroplasts (Noctor et al. I998a,b).
In almost all the biological functions, glutathione is oxidized to GSSG which should
109

be converted back to glutathione in plant cell to perform normal physiological
ftinctions. Hence, rapid recycling of glutathione is more essential rather than synthesis
of glutathione, which is a highly regulated and ATP requiring process, glutathione is
regenerated by glutathione reductase in a NADPH-dependent reaction. In this way
glutathione reductase activity maintains cellular GSH:GSSG ratio which is essential
for the control of gene expression and normal protein functions (Foyer and Noctor
2001, Noctor et al. 2002). Hence, the S-induced upregulation of glutathione reductase
activity maintained one of the important hydrophilic antioxidants, the glutathione.
Another explanation of the higher glutathione reductase activity in the present work
may be that as a component of Ascorbate-Glutathione cycle, glutathione maintains the
normal operation of Ascorbate-Glutathione cycle at a high rate in order to detoxify the
ROS; it is also essential to keep glutathione in a reduced form prior to its
incorporation into the synthesis of phytochelatins (Cobbett 2000a,b).
5.2.6.2 Non-enzymatic antioxidants
All plants can synthesize ascorbate, which can accumulate to millimolar
concentrations in both photosynthetic and non-photosynthetic tissues (Foyer et al.
1983). Ascorbate is one of the most powerful antioxidants (Noctor and Foyer 1998,
Smirnoff e/ al. 2001), reacts directly with OH', O2" and ^Oj and reduces H2O2 to H2O
via ascorbate peroxidase (Noctor and Foyer 1998, Chen and Gallie 2004). Ascorbate
is the major, probably the only, antioxidant buffer in the apoplast and is an essential
metabolite involved with vital cell ftinctions. The ascorbate content of plant tissues is
modulated by a large number of factors internal or external to plants.
In the present study, Cd treatments significantly decreased the ascorbate
content at all growth stages and the decrease was greater in non-tolerant cultivar
RH30 than tolerant cultivar Alankar (Figures 44-45). Ascorbate together with
glutathione affects plant tolerance to ROS by participation in the detoxification of
ROS in plant cells (Noctor and Foyer 1998, Wu and Zhang 2002a). The ascorbate
peroxidase activity, in the present investigation, was found enhanced in both the
cultivars. Maximum utilization of ascorbate by up-regulated ascorbate peroxidase
activity, might be one of the major reasons of reduction in ascorbate content.
Secondly, the reduction of MDHA/or DHA into the reduced ascorbate (regeneration)
form might be delayed by Cd. Other workers have also observed that Cd toxicity
reduces ascorbate content (Ozturk et al. 2003, Hsu and Kao 2004, Kuo and Kao 2004,
110

Qadir et al. 2004, Aravind and Prasad 2005, Romero-Puertas et al. l^Ql, Agrawal and
Mishra 2008, Anjum et al. 2008a).
Sulfur supplementation lowered the Cd-induced reduction in ascorbate content
of both the cultivars at all growth stages. The decrease in ascorbate content due to
50mg Cd kg"' soil was lowered by the application of 50mg S kg'' soil alone and in
combination with 80mg N kg"' soil in both the cultivars at all growth stages (Figures
44-45,70-71). Sulfur application under Cd stress has shown important role in the
alleviation of Cd-induced oxidative stress (Hassan et al. 2005b, Anjum et al. 2008a,
Khan et al. 2008a). In the present study, S application improved the ascorbate content
which probably reacted with Oj', 'O2 (directly), H2O2 (enzymatically through
ascorbate peroxidase) and thereby assisted in maintaining these potential toxicants.
The Cd-induced changes in the levels of cellular antioxidants severely affected and
impaired the functioning of the Ascorbate-Glutathione cycle (Zhang and Kirkham
1996). The Ascorbate-Glutathione cycle is a major H2O2 scavenging antioxidant
pathway that operates both in chloroplasts as well in cytosol (Zhang and Kirkham
1996).
The tripeptide (y-GluCysGly) glutathione (GSH) plays a central role in several
physiological processes, including regulation of sulfate transport, signal transduction,
conjugation of metabolites, detoxification of xenobiotics (Xiang et al. 2001) and the
expression of stress-responsive genes (MuUineaux and Rausch 2005). The reduced
form of glutathione, GSH, is an abundant compound in plant tissue that exists
interchangeably with the oxidized form, GSSG. GSH has been associated with several
growth and development related events in plants, including cell differentiation, cell
death and senescence, pathogen resistance and enzymatic regulation (Ogawa 2005,
Rausch and Wachter 2005) and its content is affected by S nutrition (Blake-Kalff et
al. 2000, Anjum et al. 2008a,b, Khan et al. 2008a). In addition, GSH is a substrate for
glutathione peroxidase (GPX) and glutathione-S-transferases (GST), which are also
involved in the removal of ROS (Noctor et al. 2002). GSH is a precursor of
phytochelatins, which are crucial in controlling cellular heavy metal concentrations in
plants (Wojcik et al. 2005, Herbette et al. 2006, Zhang et al. 2008, Anjum et al.
2008a,b).
Cd dose-dependent increase in GSH content was observed in both the cultivars
but its content was greater in Alankar (tolerant) than RH30 (non-tolerant) at all the
growth stages (Figures 44-45). Environmental stresses trigger an increase in ROS
111

levels in plants and the response of GSH can be crucial for adaptive responses.
Antioxidant activity in the leaves and chloroplast of Phragmites australis Trin. (cav.)
ex Steudel was associated with a large pool of GSH, protecting the activity of many
photosynthetic enzymes against the thiophilic bursting of Cd and exerting a direct
important protective role in the presence of Cd (Pietrini et al. 2003). Increased
concentration of GSH has been observed with the increasing Cd concentration in
Pisum sativum (Gupta et al. 2002), Lactuca sativa (Maier et al. 2003), Phragmites
australis (Pietrini et al. 2003), Brassica juncea (Qadir et al. 2004), Pisum sativum
(Metwally et al. 2005), Sedum alfredii (Sun et al. 2007), Oryza sativa (Hassan et al.
2008a).
Sulfur supplementation further increased the glutathione content of both the
cultivars and maximum significant increase in its content was noted in plants grown
with 50mg S kg'' soil at all growth stages. The increase in glutathione content due to
50mg Cd kg"' soil was further increased by the application of 50mg S kg'' soil alone
and in combination vvith 80mg N kg'' soil in both the cultivars at all growth stages
(Figures 44-45,70-71). S is required for the synthesis of various compounds, such as
thiols (GSH), sulpholipids and secondary S compounds, which play an important role
in the metabolism of plants, and in the protection and adaptation of plants against
stresses. The content of the secondary S compounds is strongly dependent on the
stage of development of the plant, temperature, water availability, and the level of S
and N nutrition (Randle et al. 1993, 1995, Randle 2000, Randle and Lancaster 2002,
Coolong and Randle 2003a,b). Fitzgerald et al. (1999) reported that when plants
receive adequate S and N, they develop substantial reserves of sulfate in the root and
glutathione in the leaves. Glutathione besides performing critical functions in
regulating plant growth and adaptation to abiotic stresses, acts as an important S sink
in the plant system (Leustek et al. 2000, Maughan and Foyer 2006). The synthesis of
glutathione is mainly regulated by the availability of its constituent amino acids Cys,
Glu and Gly along with transcriptional regulation of enzymes of glutathione
biosynthesis, y-glutamylcysteine synthase and GSH synthetase (Tomaszewska 2002).
Further, the S-supplementation might help plants to improve the content of GSH by
enhancing y-ECS enzymes as shown by Schneider and Bergmaim (1995) and Strohm
et al. (1995). Cadmium is known to rapidly induce the synthesis of phytochelatins,
thiol-based complexing substances through the up-regulation of glutathione
biosynthesis (Xiang and Oliver 1998, Rauser 2000). As phytochelatins are Glu- and
112

Cys-rich peptides, S and N assimilation pathways are most likely involved in their
synthesis: enzymes of the S metabolism are required for the synthesis of Cys,
enzymes of N metabolism are required for the synthesis of Glu. The glutathione
synthesis in plant cells is dependent on and regulated by the S-supply of the plants. In
addition, glutathione contains three moles of N per mole of S and glutathione
biosynthesis may depend on the availability of N precursors and thus N nutrition of
plants. However, the sink strength of GSH biosynthesis for N may be low compared
with the other major N sinks such as the synthesis of proteins or nucleotides (Kopriva
and Rennenberg 2004). Our results are in close conformity with the observations of
Hassan et al. (2005a,b, 2008a) and Anjum et al. (2008a). Hassan et al. (2005b)
reported that the higher S level in the growing medium alleviate the oxidative stress of
the Oryza sativa plants exposed to Cd. Anjum et al. (2008a) found strong and positive
correlation between net photosynthetic rate and plant dry mass with the contents of
ascorbate and glutathione. In addition, application of S to Cd-treated plants showed
increase in the contents of ascorbate and glutathione which reduced the Cd and
TBARS content in leaves and restored the growth and photosynthesis of Brassica
campestris. Hassan et al. (2008a) reported that addition of (NH4)2S04 to Cd-fed plants
showed increase in glutathione content which reduces the oxidative stress in Oryza
sativa plants that was related to more S supply for this N form. It has also been
suggested that (NH4)2S04 is a better fertilizer because of the presence of both S and N
for use in Cd-contaminated soil (Hassan et al. 2008a,b).
5.2.7 Yield characteristics
Yield is the final manifestation of growth, photosynthesis and biochemical traits of a
plant which are strongly regulated by several environmental factors. In our study, Cd
treatments alone significantly decreased the yield characteristics and the extent of
decrease was greater in RH30 than Alankar. The reductions in yield characteristics
due to 50mg Cd kg"' soil were lowered with the application of 50mg S kg"' soil alone
and in combination with 40, 80 and 120mg N kg"' soil in both the cultivars. The
application of 50mg S kg"' soil plus 80mg N kg"' soil maximally lowered the Cd-
induced (50mg Cd kg"' soil) reduction in yield characteristics of both the cultivars
(Figures 46-47,72-73). The improvement in yield characteristics due to S and N
application to Cd-treated plants can be correlated with the enhancement in growth,
photosynthesis, and biochemical characteristics. Furthermore, S and N application
reduced the Cd uptake and H2O2 and TEARS content along with significant increase
U3

in the enzymatic and non-enzymatic components of antioxidant defense system which
in turn reduced the Cd-induced oxidative stress and hence provided tolerance to plants
and maintained the yield. S and N supplementation to the Cd-stressed plants
ameliorated the ill effects of Cd on yield and its components by greater increase in
siliqua per plant, seeds per siliqua, 1000 seed weight and seed yield per plant of
Alankar (tolerant) than RH30 (non-tolerant) cultivars. Sulfur supplementation to
plants has also been shown as a result of greater biomass production under normal
(Abdin et al. 2003, Ahmad et al. 2005, Bouranis et al. 2008, Bimbraw 2008) and Cd
stress conditions (Hassan et al. 2005b, Anjum et al. 2008a).
Studies have shown a marked influence of applied S on the yield of pulses,
oilseeds and other crops (Pasricha and Aulakh 1991, Tandon 1991, Ahmad et al.
1998, Aulakh and Pasricha 1986, 1998, Singh 2001, Abdin et al. 2003, Bimbraw
2008). Sulfur fertilization is useful not only for increasing crop production, oil
content, and protein content, but also in improving soil conditions for crop growth
(Abdin et al. 2003). Besides it was suggested that an adequate S fertilization to
Brassica is a feasible technique to suppress the uptake of Cd by plants, and to enhance
the uptake efficiency of several essential elements which resulted in increased crop
production and improved quality. Nitrogen is critical in plant growth and development
and is an essential component of amino acids, proteins, nucleic acids, and many
enzymes. Plants grown under limited N levels have reduced Chi a and Chi b
pigments, resulting in stimted plants and characteristic leaf chlorosis (Marschner
1995). As observed also in the present study, increased additions of N usually resuh in
increased yield of crop plants (Mills and Jones 1996, Hochmuth et al. 1999). N
increases yield by influencing a number of growth parameters such as branches and
flowers per plant and by producing more vigorous growth and development (Allen
and Morgan 1972, Taylor et al. 1991). Wright et al. (1988) reported that N prolongs
the life of leaves, improves leaf area duration after flowering and increases overall
crop assimilation, thus contributing to increased seed yield. Therefore, the combined
application of S and N will improve the yield of the crop.
Hassan et al. (2008a) reported that the addition of NH4(S04)2 to Cd-fed plants
improved the yield characteristics of Oryza sativa. They suggested that the
improvement in yield characteristics was due to the ameliorative action of N and S in
the NH4(S04)2 on growth and components of antioxidant defense system of Cd-treated
plants.
114

Conclusively, nutrients are known to decipher essential role in plant
metabolism and augmenting growth and productivity of crops. However, a nutrient
package is required for sustainable agriculture under varied environmental conditions.
Crop cultivars display their inherent potential. The cultivar Alankar surpassed other
cultivars tested and tolerated Cd stress to a significant degree. The cultivar RH30 was
weak in performance and least tolerant to Cd among the cultivars tested. Therefore,
Alankar exhibited lesser decreases in growth, photosynthetic and yield characteristics
under Cd stress. Correspondingly, Alankar also showed lesser oxidative stress and
increased antioxidant system than RH3Q to protect photosynthetic machinery and
consequent effects on other attributes. Sulfur proved significant potential in the
alleviation of Cd stress in both the cultivars. The decreases in the characteristics
observed due to Cd stress were lowered by S application and 50mg S kg'' soil proved
effective in alleviation of Cd stress than the high doses of S. The coordination of S
and N proved most effective in nullifying the Cd stress effects. The potential of 50mg
S kg"' soil in the alleviation of Cd stress was substantially enhanced by the
simultaneous application of 80mg N kg"' soil. The combined application of 50mg S
kg"' soil plus 80mg N kg"' soil not only resulted in restricting the decrease caused by
Cd but also nullified the Cd effects and even enhanced the characteristics values over
the control. A package of 50mg S kg"' soil plus 80mg N kg"' soil appears to be most
effective in the cultivation of mustard under Cd stress. The effect of this combination
was due to coordination of S and N in maintaining plant metabolism and alleviating
Cd stress.
5.3 New Report in the Thesis and Future Prospects
Mineral nutrient status of plants plays a critical role in increasing plant resistance to
enviroimiental stress factors. Of the mineral nutrients, S and N are major
macronutrients necessary for the plant life cycle. The uptake and assimilation of S and
N in higher plants are the crucial factors determining plant growth and vigor and crop
yield. The present study showed that a balanced S and N supply can mitigate the
inhibitory effects of Cd on mustard metabolism. The individual effects of S and N and
other nutrients have been studied. However, no effort has been made to study
combined application of S and N in tolerance of plants, although coordinated
functions of these nutrients are well known. For sustainable agriculture a nutrient
protocol with balanced S and N is necessary in the growth and development of crops
under Cd stress. The results reported in this thesis have confirmed this. The effect of
115

combined application of S and N on oxidative stress and antioxidants under Cd stress
has been reported for the first time. Efforts have been made for the first time to
develop S and N protocol for alleviation of Cd stress in mustard.
Further, the efforts should be focused to dissect the mechanism(s) of
regulation of sulfate and nitrate assimilation at the molecular level in crop plants
under varied environmental conditions. Strategies should also aim to manipulate steps
of pathways leading to the production of thiols and their products in plants through
overexpressing Ser acetyl transferase (SAT), y-ECS and GSHS enzymes of S and N
nutrition under Cd stress. More detailed studies are required to understand the Cd-
induced stress response modulated by S and N metabolism at molecular and
mechanistic levels. This would help to develop an effective strategy to raise
transgenic species for stress resistance.
116

SUMMARY

SUMMARY Chapter 6
The present thesis entitled "Coordination of sulfur and nitrogen in the protection of
photosynthesis, growth and yield of cadmium-treated mustard {Brassica juncea)"
comprises of six chapters.
The importance of the problem and justifications for the present work
imdertaken were emphasized in Chapter 1.
Chapter 2 is the review of literature. It includes literature available on the
aspects of the physiological analysis of various growth-, photosynthetic- and,
biochemical-characteristics, antioxidant studies and yield of various crop plants under
Cd stress. The importance of S and N nutrients in the alleviation of stress and in the
regulation of plant growth and development under stress condition were also
reviewed. The chapter has been divided in to sections and sub-sections for better
understanding of the work of other research reports in this field of study. In addition,
the critical appraisal of the review of literature has also been included to identify the
gap in the field of study.
The details of the material used in the study and methodology adopted to
determine various characteristics recorded in the three experiments have been
described in the Chapter 3. In addition the Chapter 3 also mentions the relevant
information on the location of the study and the environmental conditions during the
data sampling times.
Chapter 4 includes the results on the response of mustard to cadmium, sulfur
and nitrogen treatments in three Experiments. The data obtained were statistically
analyzed and the significance was determined at;7<0.05.
The results obtained in the Experiments have been discussed in Chapter 5 in
the light of observations recorded and supported with the earlier findings, if available.
This chapter also presents the possible explanations of the data obtained to reach a
conclusion and possible future prospects.
Chapter 6 present the summary of the work reported in this thesis.
The results of the Experiments are summarized below:
Experiment 1
The Experiment was conducted to study the effect of five concentrations of Cd viz., 0,
25, 50, 100 and 150mg kg'' soil on Cd accumulation in root and leaf, growth,
photosynthetic and yield characteristics of five mustard cultivars {Brassica juncea L.)
117

namely, Alankar, Varuna, Pusa Bold, Sakha and RH30. The treatments in this
Experiment were arranged in a factorial randomized block design. Cadmium
accumulation, growth and photosynthetic characteristics were studied at pre-flowering
(30DAS), flowering (60DAS) and post-flowering (90DAS) growth stages, while yield
characteristics were noted at harvest (120DAS). Tolerance index of the five cultivars
of mustard was calculated and the cultivars were designated as Cd tolerant and Cd
non-tolerant on the basis of their performance under Cd stress. The effect of Cd on
growth, photosynthetic and yield characteristics was found significant at all sampling
times. The interaction of Cd treatments and cultivars was also found significant. For
growth characteristics, the interaction effect was found non-significant at 30 DAS.
The increase in Cd levels decreased the growth, photosynthetic and yield
characteristics of all the five cultivars at all sampling stages. The observations
recorded at the three sampling stages showed similar pattern of cultivar response to
Cd treatments. Maximum reduction in growth, photosynthesis and yield
characteristics was noted with 150mg Cd kg"' soil followed by 100, 50 and 25mg Cd
kg'' soil.
Among cultivars, Alankar showed lesser decrease in growth, photosynthetic
and yield characteristics followed by Varuna and Pusa Bold, whereas, RH30 and
Sakha exhibited greater reduction in growth characteristics under Cd stress. The
tolerance index of cultivars was: Alankar >Varuna >Pusa Bold >Sakha >RH30.
Experiment 2
Experiment 2 was conducted on the basis of findings of Experiments 1. As observed
in Experiment 1, Alankar emerged as Cd-tolerant and RH30 as Cd non-tolerant
cultivar. Among Cd levels, 150mg Cd kg"' soil was found most toxic and caused
maximum reductions in the characteristics studied. In the present Experiment, the aim
was to study the alleviation potential of elemental S levels (50 and lOOmg S kg"' soil)
on the effects of 50 and 150mg Cd kg"' soil. The treatments in this Experiment were
arranged in a factorial randomized block design. The assessment was done by
studying the changes in Cd accumulation in root and leaf, growth and photosynthetic
characteristics, S and N assimilation, components of antioxidant defense system and
yield characteristics of tolerant (Alankar) and non-tolerant (RH30) cultivars. The
sampling stages for Cd accumulation, growth and photosynthetic characteristics, S
and N assimilation, components of antioxidant defense system were pre-flowering
118

(SODAS), flowering (60DAS) and post-flowering (90DAS). The yield characteristics
were noted at harvest (120DAS).
The growth and photosynthetic characteristics and N assimilation decreased
maximally in both the cultivars treated with 150mg Cd kg'' soil. Contrarily, there was
significant increase in the Cd accumulation, S assimilation, H2O2 and TBARS content
and components of enzymatic antioxidant system and glutathione content was found.
Non-tolerant cultivar RH30 exhibited greater Cd effects than tolerant cultivar
Alankar. In comparison to lOOmg S kg'' soil, the application of 50mg S kg'' soil
maximally alleviated the toxic effect of 50mg Cd kg'' soil and improved growth,
photosynthesis, S and N assimilation and components of enzymatic and non-
enzymatic antioxidant system thus yield characteristics of both the cultivars. Further,
no ameliorative effect of lOOmg S kg'' soil was observed in plants treated with 150mg
Cd kg"' soil. Alleviation effects of S were higher in Alankar than RH30.
Experiment 3
Experiment 3 was conducted on the basis of the findings of Experiments 2. In
Experiment 2, it was observed that the application of 50mg S kg"' soil to plants treated
with 50mg Cd kg"' soil alleviated Cd-induced toxicity in both the cultivars.
Supplementation of 50mg S kg"' soil maximally overcome the toxic effects of 50mg
Cd kg"' soil in Alankar (tolerant cultivar), while, the same combination lowered the
Cd-induced toxic effects in RH30 to some extent. The present experiment was aimed
to study the effect of supplementation of different N levels (40, 80 and 120mg N kg"'
soil) to 50mg S kg"' soil for alleviation of adverse effects of 50mg Cd kg"' soil. The
treatments in this Experiment were arranged in a factorial randomized block design.
The assessment was done by analyzing the changes in Cd accumulation in root and
leaf, growth and photosynthetic characteristics, S and N assimilation, components of
antioxidant defense system at 30, 60 and 90DAS and yield characteristics at harvest.
The alterations in growth, photosynthesis, S and N assimilation, components
of antioxidant system and yield characteristics caused by 50mg Cd kg"' soil were
alleviated by 50mg S kg"' soil in tolerant (Alankar) and non-tolerant (RH30) cultivars,
but the alleviation potential of S varied between the cultivars. Application of different
levels of N (40, 80 and 120mg N kg"' soil) further enhanced the alleviation effect of S
(50mg S kg"' soil) in both the cultivars. Among the N levels used, the application of
80mg N kg"' soil proved superior to the other N levels. A combined application of S
119

and N (50mg S kg'' soil plus 80mg N kg"' soil) not only ameliorated the Cd-induced
(50mg Cd kg"' soil) effects but also increased the growth, photosynthesis, S and N
assimilation and components of enzymatic and non-enzymatic antioxidant system
thus yield characteristics in Alankar. In RH30, this combination (50mg S kg"' soil
plus 80mg N kg"' soil) only lowered the adverse effects of Cd.
The present chapter is followed by an up-to-date bibliography of the literature
cited in the text.
120

BIBLIOGRAPHY

BIBLIOGRAPHY
Abdin MZ, Ahmad A, Khan N, Khan I, Jamal A, Iqbal M (2003) Sulphur interaction
with other nutrients. In: Abrol YP, Ahmad A (Eds) Sulphur in plants. Kluwer
Academic Publishers, The Netherlands, pp359-374.
Abrol YP (2007) Agricultural nitrogen use and its environmental implications. IK
International, New Delhi, pp400.
Abrol YP, Kaim MS, Nair TVR (1976) Nitrogen assimilation, its mobilization and
accumulation in wheat (Triticum aestivum L.) grains. Cereal Res Commun 4:431-
440.
Aebi H (1984) Catalase in vitro. Meth Enzymol 105:121-126.
Agrawal SB, Mishra S (2008) Effects of supplemental ultraviolet-B and cadmium on
growth, antioxidants and yield of Pisum sativum L. Ecotoxicol Environ Saf
doi:10.1016/j.ecoenv.2007.10.007
Agrawal SB, Rathore D (2007) Changes in oxidative stress defense in wheat
(Triticum aestivum L.) and mung bean (Vigna radiata L.) cultivars grown with
and without mineral nutrients and irradiated by supplemental ultraviolet-B.
Environ Exp Hot 59:21-33.
Ahmad A, Abdin MZ (2000a) Photosynthesis and its related physiological
variables in the leaves of Brassica genotypes as influenced by sulphur
fertilization. Physiol Plant 110:144-149.
Ahmad A, Abdin MZ (2000b) Effect of sulphur application on lipid, RNA,
protein content and fatty acid composition in developing seeds of rapeseed
(Brassica campestris L.). Plant Sci 150:71-75.
Ahmad A, Abraham G, Abdin MZ (1999) Physiological investigation on the
impact of nitrogen and sulphur application on seed and oil yield of rapeseed
(Brassica campestris L.) and mustard (Brassica juncea L. Czern and Coss.)
genotypes. J Agron Crop Sci 183:10-25.
Ahmad A, Abraham G, Gandotra N, Abrol YP, Abdin MZ (1998) Interactive
effect of nitrogen and sulphur on growth and yield of rapeseed-mustard
(Brassica juncea L. Czern and Coss. and Brassica campestris L.) genotypes.
J Agron Crop Sci 181:193-199.
121

Ahmad A, Khan I, Anjum NA, Abrol YP, Iqbal M (2005) Role of sulphate
transporter systems in sulphur efficiency of mustard genotypes. Plant Sci
169:842-846.
Ahsan N, Lee S-H, Lee D-G, Lee H, Lee SW, Bahk JD, Lee B-H (2007)
Physiological and protein profiles alternation of germinating rice seedlings
exposed to acute cadmium toxicity. C R Biol 330:735-746.
Aidid SB, Okamoto H (1993) Effect of lead, cadmium and zinc on the electric
membrane potential at the xylem/symplast interface and cell elongation of
Impatiens balsamina. Environ Exp Bot 32:439-448.
Ali G, Srivastava PS, Iqbal M (2000) Influence of cadmium and zinc on growth
and photosynthesis of Bacopa monniera cultivated in vitro. Biol Plant
43:599-601.
Ali MB, Chun HS, Kim BK, Lee CB (2002) Cadmium-induced changes in
antioxidant enzyme activities in rice {Oryza sativa L. cv. Dongjin). J Plant
Biol 45:134-140.
Allen EJ, Morgan DG (1972) A quantitative analysis of the effects of nitrogen on
the growth, development and yield of oilseed rape. J Agric Sci (Cambridge)
78:315-324.
Allen RD, Webb RP, Schake SA (1997) Use of transgenic plants to study
antioxidant defenses. Free Rad Biol Med 23:473-479.
Alloway BJ (1995) Heavy Metals in Soils, 2" * Ed. Blackie Academic & Professional
Publishers, London, pp368.
Alscher RG, Erturk N, Heatrh LS (2002) Role of superoxide dismutases (SODs)
in controlling oxidative stress in plants. J Exp Bot 53:1331-1341.
Ammar WB, Nauairi I, Zarrouk M, Jamel F (2007) Cadmium stress induces
changes in the lipid composition and biosynthesis in tomato {Lycopersicon
esculentum Mill.) leaves. Plant Growth Regul 53:75-85.
An YJ (2004) Soil ecotoxicity assessment using cadmium sensitive plants.
Environ Pollut 127:21-26.
Anderson ME (1985) Determination of glutathione and glutathione disulfides in
biological samples. Meth Enzymol 113:548-570.
Anjana, Umar S, Iqbal M (2006) Functional and structural changes associated with
cadmium in mustard plant: effect of applied sulphur. Commun Soil Sci Plant Anal
37:1205-1217.
122

Anjum NA, Umar S, Ahmad A, Iqbal M, Khan NA (2008a) Sulphur protects mustard
{Brassica campestris L.) from cadmium toxicity by improving leaf ascorbate and
glutathione. Plant Growth Regul 54:271-279.
Anjum NA, Umar S, Singh S, Nazar R, Khan NA (2008b) Sulfur assimilation and
cadmium tolerance in plants. In: Khan NA, Singh S, Umar S (Eds) Sulfur
assimilation and abiotic stresses in plants. Springer-Verlag, The Netherlands,
pp271-302.
Anjum NA, Umar S, Ahmad A, Iqbal M (2008c) Responses of components of
antioxidant system in moongbean [Vigna radiata (L.) Wilczek] genotypes to
cadmium stress. Commun Soil Sci Plant Anal (In Press)
Anjum NA, Umar S, Ahmad A, Iqbal M, Khan NA (2008d) Ontogenic variation in
response oi Brassica campestris L. to cadmium toxicity. J Plant Inter 1-11. doi:
10.1080/17429140701823164
Anjum NA, Khan NA, Singh S, Nazar R (2007) Overproduction of cysteine in high
photosynthetic potential mustard {Brassica juncea L.) cultivar reduces cadmium-
induced oxidative stress. XXXth Botanical Conference, Gwalior, MP, 28-30
November, ppl48.
Ansel DC, Franklin MLT, De Carvalho MHC, Lameta ADA, Fodil YZ (2006)
Glutathione reductase in leaves of cowpea: cloning of two cDNAs, expression and
enzymatic activity under progressive drought stress desiccation and abscisic acid
treatment. Ann Bot 98:1279-1287.
Aono M, Kubo A, Saji H, Tanaka K, Kondo N (1993) Enhanced tolerance to
photooxidative stress of transgenic Nicotiana tabacum with high
chloroplastic glutathione reductase activity. Plant Cell Physiol 34:129-135.
Arao T, Ae N, Sugiyama M, Takahashi M (2003) Genotypic differences in
cadmium uptake and distribution in soybeans. Plant Soil 251:247-253.
Arduini I, Godbold D, Onnis A (1994) Cadmium and copper change root growth
and morphology of Pinus pinea and Pinus piaster seedlings. Physiol Plant
92:675-680.
Arduini I, Masoni A, Mariotti M, Ercoli L (2004) Low cadmium application
increase miscanthus growth and cadmium translocation. Environ Exp Bot
52:89-100.
Amon DI (1949) Copper enzymes in isolated chloroplasts polyphenoloxidase in Beta
vulgaris. Plant Physiol 24:1-5.
123

Artetxe U, Garcia-Plazaola JI, Hernandez A, Becerril JM (2002) Low light grown
duckweed plants are more protected against the toxicity induced by Zn and Cd.
Plant Physiol Biochem 40:859-863.
Aravind P, Prasad MNV (2003) Zinc alleviates cadmium-induced oxidative
stress in Ceratophyllum demersum L.: a free-floating freshwater macrophyte.
Plant Physiol Biochem 41:391-397.
Aravind P, Prasad MNV (2004) Carbonic anhydrase impairment in cadmium-
treated Ceratophyllum demersum L. (free floating freshwater macrophyte):
toxicity reversal by zinc. J Anal At Spectrom 19:52-57.
Aravind P, Prasad MNV (2005) Cadmium and zinc interactions in a hydroponic
system using Ceratophylum demersum L.: adaptive ecophysiology,
biochemistry and molecular toxicology. Braz J Plant Physiol 17:3-20.
Asada K (1992) Ascorbate peroxidase: a hydrogen peroxide scavenging enzyme
in plants. Physiol Plant 85:235-241.
Asada K (1994) Production and action of active oxygen in photosynthetic tissue.
In: Foyer CH, MuUineaux PM (Eds) Causes of photooxidative stress and
amelioration of defense system in plants. CRC Press, Boca Raton, FL, USA,
pp77-104.
Asada K (1999) The water-water cycle in chloroplasts: scavenging of active
oxygen and dissipation of excess photons. Annu Rev Plant Physiol Plant Mol
Biol 50:601-639.
Asada K (2006) Production and scavenging of reactive oxygen species in
chloroplasts and their functions. Plant Physiol 141:391-396.
Asada K, Takahashi M (1987) Production and scavenging of active oxygen
species in photosynthesis. In: Kyle DJ, Osmond CB, Arntzen CJ (Eds)
Photoinhibition. Elsevier, Amsterdam, pp227-287.
Ascencio CL, Cedeno-Maldonado A (1979) Effects of cadmium on carbonic
anhydrase and activities dependent on electron transport of isolated
chloroplasts. J Agric Univ Puerto Rico 63:195-201.
Astolfi S, Zuchi S, Passera C (2004) Role of sulphur availability on cadmium-
induced changes of nitrogen and sulphur metabolism in maize {Zea mays L.)
leaves. J Plant Physiol 161:795-802.
Atal N, Pardhasaradhi P, Mohanty P (1991) Inhibition of chloroplast
photochemical reactions by treatment of wheat seedlings with low
124

concentrations of cadmium: Analysis of electron transport activities and
changes in fluorescence yield. Plant Cell Physiol 32:943-951.
Aulakh MS (2003) Crop responses to sulfur nutrition. In: Abrol YP, Ahmad A (Eds)
Sulphur in Plants. Kluwer Academic Publishers, The Netherlands, pp341-358.
Aulakh MS, Pasricha NS (1986) Role of sulphur in the production of the grain
legumes. Pert News 3:31-35.
Aulakh MS, Pasricha NS (1998) The effect of green manuring and fertilizer N
application on enhancing crop productivity in mustard-rice rotation in semiarid
subtropical regions. Euro J Agron 8:51-58.
Aulakh MS, Pasricha NS, Sahota NS (1980) Yield, nutrient concentration and
quality of mustard crop as influenced by nitrogen and sulfur fertilization. J
Agr Sci Cambridge 84:545-549.
Azevedo RA, Alas RM, Smith RJ, Lea PA (1998) Response of antioxidant
enzymes to transfer from elevated carbon dioxide to air and ozone
fumigation, in leaves and roots of wild-type and catalase-deficient mutant of
barley. Physiol Plant 104:280-292.
Azpilicueta CE, Benavides MP, Tomaro ML, Gallego SM (2007) Mechanism of
CATA3 induction by cadmium in sunflower leaves. Plant Physiol Biochem
45:589-595.
Bachir MLD, Wu FB, Zhang GP, Wu HX (2004) Genotypic differences in effect
of cadmium on the development and mineral concentrations of cotton.
Commun Soil Sci Plant Anal 35:285-299.
Balakhnina T, Kosobryukhov A, Ivanov A, Kreslavskii V (2005) The effect of
cadmium on CO2 exchange, variable fluorescence of chlorophyll, and the
level of antioxidant enzymes in pea leaves. Russ J Plant Physiol 52:15-20.
Balestrasse KB, Gallego SM and Tomaro ML (2006a) Oxidation of the enzymes
involved in nitrogen assimilation plays an important role in the cadmium-
induced toxicity in soybean plants. Plant Soil 284:187-194.
Balestrasse KB, Gallego SM, Tomaro ML (2004) Cadmium-induced senescence
in nodules of soybean (Glycine max L.) plants. Plant Soil 262:373-381.
Balestrasse KB, Gardey L, Gallego SM, Tomaro ML (2001) Response of
antioxidant defence system in soybean nodules and roots subjected to
cadmium stress. Austr J Plant Physiol 28:497-504.
125

Balestrasse KB, Noriega GO, BatUe A, Tomaro ML (2006b) Heme oxygenase
activity and oxidative stress signaling in soybean leaves. Plant Sci 170:339-
346.
Bannister JV, Bannister WH, Rotilio G (1987) Aspects of the structure, function
and applications of superoxide dismutase. CRC Crit Rev Biochem 22:111-
180.
Bapet PN, Sinha SB, Bhinde DA (1986) Effect of S and P on yield and nutrient
content of blackgram. J Ind Soc Soil Sci 34:82-86.
Barcelo J, Cobet C, Poschenreider Ch (1986a) Cadmium induced decrease of
water stress resistance in bush bean plants {Phaseolus vulgaris L. cv.
Conterder). II. Effects of Cd on endogenous abscisic acid levels. J Plant
Physiol 125:27-34.
Barcelo J, Poschenreider C, Andreu I, Gunse B (1986b) Cadmium induced
decrease of water stress resistance in Bush bean plants {Phaseolus vulgaris L.
cv. Contender) I. Effects of cadmium on water potential, relative water
content and cell wall elasticity. J Plant Physiol 125:17-25.
Barcelo J, Poschenreider Ch (1990) Plant water relations as affected by heavy
metal stress: a review. J Plant Nutr 13:1-37.
Barcelo J, Vazques MD, Poschenreider Ch (1988a) Structural and ultrastructural
disorders in cadmium-treated bush bean plants {Phaseolus vulgaris L.). New
Phytol 108:37-49.
Barcelo J, Vazquez MD, Poschenreider Ch (1988b) Cadmium-induced structural
and ultra-stuctural changes in the vascular system of bush bean stems.
Botanica Acta 101:254-261.
Barney Jr PE, Bush LP (1985) Interaction of nitrate and sulphate reduction in
tobacco. I. Influence of availability of nitrate and sulphate. J Plant Nutr
8:507-515.
Barroso JB, Corpas FJ, Carreras A, Rodriguez-Serrano M, Esteban FJ, Fernandez-
Ocana A, Chaki M, Romero-Puertas MC, Valderrama R, Sandalio LM, del Rio
LA (2006) Localization of S-nitrosogluthathione and expression of S-
nitrosoglutathione reductase in pea plants under cadmium stress. J Exp Bot
57:1785-1793.
126

Baryla A, Carrier P, Franck F, Coulomb C, Sahut C, Havaux M (2001) Leaf chlorosis
in oilseed rape plants {Brassica napus) grown on cadmium-polluted soils: causes
and consequences for photosynthesis and growth. Planta 212:696-709.
Baszynski T (1986) Interference of Cd " in functioning of the photosynthetic
apparatus of higher plants. Acta Soc Hot Pol 55:291-304.
Baszynski T, Wajda L, Krol M, Wolinska D, Krupa Z, Tukendor A (1980)
Photosynthetic activities of cadmium-treated tomato plants. Physiol Plant
48:365-370.
Bazzaz FA, Govindjee (1974) Effects of cadmium nitrate on spectral
characteristics and light reactions of chloroplasts. Environ Sci Lett 6:1-12.
Bazzaz FA, Rolfe GL, Carlson RW (1974) Effect of cadmium on photosynthesis
and transpiration of excised leaves of corn and sunflower. Plant Physiol
32:373-376.
Becerril JM, Munoz-Rueda A, Aparicio-Tejo P, Gonzalez-Murua C (1989) The
effects of cadmium and lead on photosynthetic electron transport in clover
and lucerne. Plant Physiol Biochem 26 913-918.
Belimov AA, Safronova VI, Tsyganov VE, Borisov AY, Kozhemyakov AP,
Stepanok VV, Martenson AM, Gianinazzi-Pearson V, Tikhonovich lA (2003)
Genetic variability in tolerance to cadmium and accumulation of heavy
metals in pea {Pisum sativum L.). Euphytica 131:25-35.
Bell CI, Clarkson DT, Cram WJ (1995) Partitioning and redistribution of sulphur
during S-stress in Macroptilium atropurpureum cv. Siratro. J Exp Bot 46:73-
81.
Ben Ammar W, Nouairi I, Tray B, Zarrouk M, Jemal F, Ghorbel MH (2005)
Effets du cadmium sur 1'accumulation ionique et les teneurs en lipides dans
les feuilles de tomate (Lycopersicon esculentum). J Soc Biol 199:157-163.
Ben Youssef N, Nouairi I, Ben Temime S, Taamalli W, Zarrouk M, Ghorbel MH,
Ben Miled Daoud D (2005) Effets du cadmium sur le metabolisme des lipides
de plantules de colza {Brassica napus L.). C R Biol 328:745-757.
Benavides MP, Gallego SM, Tomaro ML (2005) Cadmium toxicity in plants.
Braz J Plant Physiol 17:21-34.
Bema B, Bemier F (1999) Regulation by biotic and abiotic stress of a wheat germin
gene encoding oxalate oxidase, a H202-producing enzyme. Plant Mol Biol 39:539-
549.
127

Bhattacharjee S (1998) Membrane lipid peroxidation, free radical scavengers and
ethylene evolution in Amaranthus as affected by lead and cadmium. Biol
Plant 40:131-135.
Bimbraw AS (2008) Sulfur nutrition and assimilation in crop plants. In: Khan
NA, Singh S, Umar S (Eds) Sulfur assimilation and abiotic stresses in plants.
Springer-Verlag, The Netherlands, pp55-95.
Bindhu SJ, Bera AK (2001) Impact of cadmium toxicity on leaf area, stomatal
frequency, stomatal index and pigment content in mungbean seedlings. J Environ
Biol 22:307-309.
Blake-Kalff MMA, Hawkesford MJ, Zhao FJ, McGrath SP (2000) Diagnosing sulfur
deficiency in field-grown oilseed rape (Brassica napus L.) and wheat (Triticum
aestivum L.). Plant Soil 225:95-107.
Blaylock MJ, Huang JW (2000) Phytoextraction of metals. In: Raskin I, Ensley BD
(Eds) Phytoremediation of toxic metals: using plants to clean up the environment.
John Wiley and Sons, Inc, Toronto, Canada, pp303.
Boddi B, Oravecz AR, Lehoczki E (1995) Effect of cadmium on organization and
photoreduction of protochlorophyllide in dark-grown leaves and etioplast inner
membrane preparations of wheat. Photosynthetica 31:411-420.
Bolwell GP, Blee KA, Butt VS, Davies DR, Gardner SL, Gerrish C, Minibayeba F,
Rowntree EG, Wojtaszek P (1999) Recent advances in understanding the origin of
the apoplastic oxidative burst in plant cells. Free Radical Res 31:S137-S145.
Bouranis DL, Buchner P, Chorianopoulou SN, Hopkins L, Protonotarios VE,
Siyiannis VF, Hawkesford MJ (2008) Responses to sulfur limitation in maize. In:
Khan NA, Singh S, Umar S (Eds) Sulfur assimilation and abiotic stresses in
plants. Springer-Verlag, The Netherlands, ppl-19.
Boussama N, Ouariti O, Suzuki A, Ghorbal MH (1999) Cd-stress on nitrogen
assimilation. J Plant Physiol 155:310-317.
Bowler C, Van Montagu M, Inze D (1992) Superoxide dismutase and stress
tolerance. Annu Rev Plant Physiol Plant Mol Biol 43:83-116.
Boyer JS (1982) Plant productivity and environment potential for increasing crop
plant productivity, genotypic selection. Science 218:443-448.
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal Biochem
72:248-254.
128

Bray EA, Bailey-Serres J, Weretilnyk E (2000) Responses to abiotic stresses. In:
Gruissem W, Buchannan B, Jones R (Eds) Biochemistry and molecular
biology of plants. Am Soc Plant Biol, Rockville, MD, ppl 158-1249.
Breckle S, Kahle H (1992) Effects of toxic heavy metals (Cd, Pb) on growth and
mineral nutrition of beech (Fagus syhatica L.). Vegetatio 101:45-53.
Brej T (1998) Heavy metal tolerance in Agropyron repens (L.) p. Bauv.
Populations from the Legnica copper smelter area, Lower Silesia. Acta Soc
Bot Pol 67:325-333.
Brune A, Urbach W, Dietz KJ (1994) Compartmentation and transport of zinc in
barley primary leaves as basic mechanisms involved in zinc tolerance. Plant
Cell Environ 17:153-162.
Brunold C (1990) Regulation of sulfate sulfide. In: Renenberg H, Brunold C, De
Kok LJ, Stulen I (Eds) Sulfur nutrition and sulfur assimilation in higher
plants: fundamental, environmental and agricultural aspects, SPB Academic
Publishing, The Hague, ppl3-31.
Brunold C (1993) Regulatory interactions between sulfate and nitrate
assimilation. In: de Kok LJ, Stulen I, Renenberg H, Brunold C, Rauser W
(Eds) Sulfur nutrition and sulfur assimilation in higher plants: regulatory,
agricultural and enviroimiental aspects, SPB Academic Publishing, The
Hague, ppl25-138.
Brunold C, Ruegsegger A, Brandle R (1996) Stress bei Pflanzen. Bern,
Switzerland, Haupt.
Brunold C, Suter M (1984) Regulation of sulphate assimilation by nitrogen
nutrition in the duckweek Lemna minor L. Plant Physiol 76:579-583.
Bungard RA, McNeil D, Morton JD (1997) Effects of nitrogen on the
photosynthetic apparatus of Clematis vitalba grown at several irradiances.
Austr J Plant Physiol 24:205-214.
Burritt DJ (2008) Glutathione metabolism in bryophytes under abiotic stress. In:
Khan NA, Singh S, Umar S (Eds) Sulfur assimilation and abiotic stresses in
plants. Springer-Verlag, The Netherlands, pp303-316.
Buszek J (1984) Effect of cadmium on chlorophyll content and dry matter yield
in wheat and cucumber plants supplied either with nitrate or ammonium. Acta
Univ Bratislaviensis 31:11-21.
129

Cataldo CD, Garland TR, Wildung RE (1983) Cadmium uptake, kinetics in intact
soybean plants. Plant Physiol 73:844-848.
Chaffei C, Gouia H, Gorbel MH (2003) Nitrogen metabolism of tomato under
cadmium stress conditions. J Plant Nutr 26:1617-1634.
Chaffei C, Pageau K, Suzuki A, Gouia H, Ghorbel MH, Masclaux-Daubresse C
(2004) Cadmium toxicity induced changes in nitrogen management in
Lycopersicon esculentum leading to a metabolic safeguard through an amino
acid storage strategy. Plant Cell Physiol 45:1681-1693.
Chalapathi Rao ASV, Reddy AR (2008) Glutathione reductase: a putative redox
regulatory system in plant cells. In: Khan NA, Singh S, Umar S (Eds) Sulfur
assimilation and abiotic stresses in plants. Springer-Verlag, The Netherlands,
ppl 11-147.
Chandler AJ (1996) Characterising cadmium in municipal solid waste. Sources
of cadmium in the environment, Inter-Organisation Programme for the Sound
Management of Chemicals (lOMC), Organisation for Economic Co-operation
and Development (OECD), Pads, France.
Chaney RL, Hornick SB (1978) Accumulation and effects of cadmium on crops.
Cadmium 77: Proc 1st Int Cd Conference San Francisco. Metal Bulletin Ltd,
London.
Chaoui A, Ghorbal MH, El-Ferjani E (1997a) Effect of cadmium-zinc on
hydrogenically grown bean (Phaseolus vulgaris L.). Plant Sci 126:21-28.
Chaoui A, Mazhoudi S, Ghorbal MH, El-Ferjani E (1997b) Cadmium and zinc
induction of lipid peroxidation and effects on antioxidant enzyme activities in
bean (Phaseolus vulgaris L.). Plant Sci 127:139-147.
Chatterjee C, Dube BK (2006) Cadmium-a metal-an enigma: An overview. In:
Khan NA, Samiullah (Eds) Cadmium toxicity and tolerance in plants. Narosa
Publishing House, New Delhi, ppl59-177.
Chaves MM (1991) Effects of water deficits on carbon assimilation. J Exp Bot
42:1-16.
Chen GX, Sano S, Asada K (1992) The amino acid sequence of ascorbate
peroxidase from tea has high degree of homology to that of cytochrome c
peroxidase from yeast. Plant Cell Physiol 33:109-116.
Chen J, Zhou J, Goldsbrough PB (1997) Characterization of phytochelatin
synthase from tobacco. Physiol Plant 101:165-172.
130

Chen W, Chang AC, Wu L (2007) Assessing long-term environmental risks of
trace elements in phosphate fertilizers. Ecotoxicol Environ Saf 67:48-58.
Chen Y, Huerta AJ (1997) Effects of sulfur nutrition on photosynthesis in
cadmium-treated barley seedlings. J Plant Nutr 20:845-856.
Chen Z, Gallic DR (2004) The ascorbic acid redox state controls guard cell
signaling and stomatal movement. The Plant Cell 16:1143-1162.
Chen Z, Young TE, Ling J, Chang S, Gallic D R (2003) Increasing vitamin C
content of plants through enhanced ascorbate recycling. Proc Natl Acad Sci
USA 100:3525-3530.
Cheng W-D, Zhang G-P, Yao H-G, Wu W, Xu M (2006) Genotypic and
environmental variation in cadmium, chromium, arsenic, nickel, and lead
concentration in rice grains. J Zhejiang Univ SCI 87:565-571.
Chesnin L, Yien C H (1951) Turbidimetric determination of available sulfates. Soil
Sci Soc Am J 15:149-151.
Chien H-F, Kao CH (2000) Accumulation of ammonium in rice leaves is response to
excess cadmium. Plant Sci 156:111-115.
Chien H-F, Wang J-W, Lin CC, Kao CH (2001) Cadmium toxicity of rice leaves is
mediated through lipid peroxidation. Plant Growth Regul 33:205-213.
Cho U, Seo N (2005) Oxidative stress in Arahidopsis thaliana exposed to cadmium is
due to hydrogen peroxide accumulation. Plant Sci 168:113-120.
Chugh LK, Gupta VK, Sawhney SK (1992) Effect of cadmium on enzymes of
nitrogen metabolism in pea seedlings. Phytochem 31:395-400.
Chugh LK, Sawhney SK (1999) Photosynthetic activities of Pisum sativum
seedlings grown in presence of cadmium. Plant Physiol Biochem 37:297-303.
Ciecko Z, Kalembasa S, Wyszkowski M, Rolka E (2001) The effect of elevated
cadmium content in soil on the uptake of nitrogen by plants. Plant Soil
Environ 50:283-294.
Clemens S (2001) Molecular mechanisms of plant metal tolerance and
homeostasis. Planta 212:475-486.
Clemens S (2006) Toxic metal accumulation, responses to exposure and
mechanisms of tolerance in plants. Biochimie 88:1707-1719.
Clijsters H, Van Assche F (1985) Inhibition of photosynthesis by heavy metals.
Photosynth Res 7:40-41.
131

Cobbett C, May M, Howden R, Rolls B (1998) The glutathione-deficient,
cadmium-sensitive mutant, cad2-l, of Arabidopsis thaliana is deficient in
gamma-glutamylcysteine synthetase. Plant J 16:73-78.
Cobbett CS (2000a) Phytochelatins and their roles in heavy metal detoxification.
Plant Physiol 123:825-832.
Cobbett CS (2000b) Phytochelatin biosynthesis and function in heavy metal
detoxification. Curr Opin Plant Biol 3:211-216.
Cobbett CS, Goldsbrough P (2002) Phytochelatins and metallothioneins: roles in
heavy metal detoxification and homeostasis. Annu Rev Plant Physiol Plant
Mol Biol 53:159-182.
Collin VC, Eymery F, Genty B, Rey P, Havaux M (2008) Vitamin E is essential
for the tolerance of Arabidopsis thaliana to metal-induced oxidative stress.
Plant Cell Environ 31:244-257.
Collins A (2001) Carotenoids and genomic stability. Mutat Res 475:1-28.
Conn EE, Vermesland B (1951) Glutathione reductase in wheat germ. J Biol Chem
192:17-28.
Cook M, Morrow H (1995) Anthropogenic sources of cadmium to the Canadian
environment. Workshop proceedings. National workshop on cadmium transport
into plants, Canadian network of toxicology centres, June 20-21, Ottawa, Canada.
Coolong TW, Randle WM (2003a) Ammonium nitrate fertility levels influence flavor
development in hydroponically grown 'Granex 33' onion. J Sci Food Agric
83:477-482.
Coolong TW, Randle WM (2003b) Temperature influences flavor intensity and
quality in 'Granex 33' onion. J Amer Soc Hort Sci 128:176-181.
Cosio C, Vollenweider P, Keller C (2006) Localization and effects of cadmium in
leaves of a cadmium-tolerant willow {Salix viminalis L.) I. Macrolocalization and
phytotoxic effects of cadmium. Environ Exp Bot 58:64-74.
Costa G, Morel J (1994) Water relations, gas exchange and amino acid content in
Cd-treated lettuce. Plant Physiol Biochem 32:561-570.
Dahlin C, Aronsson H, Almkvist J, Sundqvist C (2000) Protochlorophyllide-
independent import of two NADPH: Pchlide oxidoreductase proteins (PORA
and PORB) from barley into isolated plastids. Physiol Plant 109:298-303.
132

Damodaran T, Hegde DM (2002) Oilseeds situation: A statistical compendium,
Directorate of oilseeds research, Indian council of agricultural research,
Hyderabad, India.
Dalurzo HC, Sandalio LM, Gomez M, del Rio LA (1997) Cadmium infiltration
of detached pea leaves: effect on its activated oxygen metabolism. Phyton
(Austria) 37:59-64.
Das P, Samantaray S, Rout GR (1997) Studies on cadmium toxicity in plants: a
review. Environ Pollut 98:29-36.
Davletova S, Rizhsky L, Liang H, Shengqiang Z, Oliver DJ, Coutu J, Shulaev V,
Schlauch K, Mittler R (2005) Cytosolic ascorbate peroxidase 1 is a central
component of the reactive oxygen gene network of Arabidopsis. Plant Cell
17:268-281.
de Fillippis LF, Zeigler H (1993) Effect of sublethal concentration of zinc,
cadmium and mercury on the photosynthetic reduction cycle of Euglena. J
Plant Physiol \Al:\ei-\ll.
De Knecht JA, Van Baren N, ten Bookum WM, Wong Pong Sang HW, Koevoets
PLM, Schat H, Verkleij JAC (1995) Synthesis and degradation of
phytochelatins in cadmium sensitive and cadmium-tolerant Silene vulgaris.
Plant Sci 106:9-18.
De Kok LJ, Castro A, Durenkamp M, Kralewska A, Posthumus FS, Elisabeth,
Stuiver E, Yang L, Stulen I (2005) Pathways of plant sulphur uptake and
metabolism-an overview. Landbauforschung Volkenrode 283:5-13.
De Vos CHR, Ten Boukum WM, Vooijs R, Schat H, De Kok LJ (1993) Effect of
copper on fatty acid composition and peroxidation of lipids in the roots of
copper tolerant and sensitive Silene cucbalus. Plant Physiol Biochem 31:151-
158.
del Rio LA, Corpas FJ, Sandalio LM, Palma JM, Gomez M, Barroso JB (2002)
Reactive oxygen species, antioxidant systems and nitric oxide in
peroxisomes. J Exp Bot 53:1255-1272.
del Rio LA, Sandalio LM, Altomare DA, Zilinskas BA (2003) Mitochondrial and
peroxisomal magnese superoxide dismutase: differential expression during
leaf senescence. J Exp Bot 54:923-933.
133

Demirevska-Kepova K, Simova-Stoilova L, Stoyanova ZP, Feller U (2006)
Cadmium stress in barley: growth, leaf pigment, and protein composition and
detoxification of reactive oxygen species. J Plant Nutr 29:451-468.
Deng DM, Shu WS, Zhang J, Zou HL, Lin Z, Ye ZH, Wong MH (2007) Zinc and
cadmium accumulation and tolerance in populations of Sedum alfredii.
Environ Pollut 147:381-386.
Dhindsa RH, Plumb-Dhindsa P, Thorpe TA (1981) Leaf senescence correlated with
increased level of membrane permeability, lipid peroxidation and decreased level
of SOD and CAT. J Exp Bot 32:93-101.
Di Cagno R, Guidi L, De Gara L, Soldatini GF (2001) Combined cadmium and
ozone treatments affect photosynthesis and ascorbate-dependent defences in
sun flower. New Phytol 151:627-636.
Di Cagno R, Guidi L, Stefani A, Soldatini GF (1999) Effect of cadmium on
growth of Hellianthus annuus seedlings: physiological aspects. New Phytol
144:65-71.
Dietz KJ, Baier M, Kramer U (1999) Free radicals and reactive oxygen species
as mediators of heavy metal toxicity in plants. In: Prasad MNV, Hagemeyer J
(Eds) Heavy metal stress in plants: from molecules to ecosystems. Springer-
Verlag, Berlin, pp73-97.
Dietz KJ, Heilos L (1990) Carbon metabolism in spinach leaves as affected by
leafage and phosphorous and sulphur nutrition. Plant Physiol 93:1219-1225.
Dixit V, Pandey V, Shyam R (2001) Differential oxidative responses to cadmium
in roots and leaves of pea (Pisum sativum L. cv. Azad). J Exp Bot 52:1101-
1109.
Djebali W, Gallusci P, Polge C, Boulila L, Galtier N, Raymond P, Chaibi W,
Brouquisse R (2008) Modifications in endopeptidase and 20S proteasome
expression and activities in cadmium treated tomato (Solanum lycopersicum
L.) plants. Planta 227:625-639.
Dominguez MJ, Gutierrez F, Leon R, Vilchez C, Vega JM, Vigara J (2003)
Cadmium increases the activity levels of glutamate dehydrogenase and
cysteine synthase in Chlamydomonas reinhardtii. Plant Physiol Biochem
41:828-832.
134

Dominguez-Solis JR, Gutierrez-Alcala G, Romero LC, Gotor C (2001) The
cytosolic 0-acetylserine (thiol) lyase gene is regulated by heavy-metals and
can function in cadmium tolerance. J Biol Chem 276:9297-9302.
Dong J, Wu F-B, Zhang GP (2005) Effect of cadmium on growth and
photosynthesis of tomato seedlings. J Zhejiang Univ SCI 68:974-980.
Drazic G, Mihailovic N, Stojanovic Z (2004) Cadmium toxicity: the effect on
macro- and micronutrients contents in soybean seedlings. Biol Plant 48:605-
507.
Drazkiewicz M, Baszynski T (2005) Growth parameters and photosynthetic
pigments in leaf segments of Zea mays exposed to cadmium, as related to
protection mechanisms. J Plant Physiol 162:1013-1021.
Dwivedi RS, Randhava NS (1974) Evaluation of a rapid test for the hidden hunger of
zinc in plants. Plant Soil 40:445-451.
Ebbs S, Lau I, Ahner B, Kochian L (2002) Phytochelatin synthesis is not
responsible for Cd tolerance in the Zn/Cd hyperaccumulator Thlaspi
caerulescens (J. & C. Presl). Planta 214:635-640.
Ekmekci Y, Tanyolac D, Ayhana B (2008) Effects of cadmium on antioxidant
enzyme and photosynthetic activities in leaves of two maize cultivars. J Plant
Physiol doi:10.1016/j.jplph.2007.01.017
El-Enany A (1995) Alleviation of cadmium toxicity on maize seedlings by
calcium. Biol Plant 37:93-99.
El-Shintinawy F (1999) Glutathione counteract the inhibitory effect induced by
cadmium on photosynthetic process in soybean. Photosynthetica 36:171-179.
Environmental Resources Limited (ERL) (1990) Evaluation of the sources of
human and environmental contamination by cadmium. Prepared for the
commission of the European community, Directorate general for
environment, consumer protection and nuclear safety, London.
Eriksson J, Oborn I, Jansson G, Andersson A (1996) Factors influencing Cd-
content in crops. Swed J Agric Res 26:125-133.
Ernst WHO (1998) Sulfur metabolism in higher plants: potential for
phytoremediation. Biodegradation 9:311-318.
Ernst WHO, Krauss G-J, Verkleij JAC, Wesenberg D (2008) Interaction of heavy
metals with the sulphur metabolism in angiosperms from an ecological point
of view. Plant Cell Environ 31:123-143.
135

Fadzilla NM, Finch RP, Burdon RH (1997) Salinity, oxidative stress and
antioxidant responses in shoot cultures of rice. J Exp Bot 48:325-331.
Fahmy AS, Mohamed TM, Mohamed SA, Saker MM (1998) Effect of salt stress
on antioxidant activities in cell suspension cultures of cantaloupe {Cucumis
melo). Egypt J Physiol Sci 22:315-326.
Fargasova A (2004) Toxicity comparison of some possible toxic metals (Cd, Cu,
Pb, Se, Zn) on young seedlings of Sinapis alba L. Plant Soil Environ 50:33-
38.
Fassett DW (1980) Cadmium. In: Waldron HA (Ed) Metals in the environment.
Academic Press, London, UK, pp61-l 10.
Fediuc E, Erdei L (2002) Physiological and molecular aspects of cadmium
toxicity and protective mechanisms induced in Phragmites australis and
Typha latifolia. J Plant Physiol 159:265-271.
Ferrario S, Valadier MH, Foyer CH (1998) Overexpression of nitrate reductase
in tobacco delays drought-induced decreases in nitrate reductase activity and
mRNA. Plant Physiol 117:239-302.
Ferreira MBR, Teixeira RNA (1992) Sulphur starvation in Lemna leads to
degradation of ribulose biphosphate carboxylase without plant death. The J
Biol Chem 267:7253-7257.
Ferreira RR, Fomazier RF, Vitoria AP, Lea PJ, Azevedo RA (2002) Changes in
antioxidant enzyme activities in soybean under cadmium stress. J Plant Nutr
25:327-342.
Ferretti M, Ghisi R, Merlo L, Dalla Vecchia F, Passera C (1993) Effect of
cadmium on photosynthetic sulphate and nitrate assimilation. Photosynthetica
29:49-54.
Filek M, Keskinen R, Hartikainen H, Szarejko I, Janiak A, Miszalski Z, Golda A
(2007) The protective role of selenium in rape seedlings subjected to
cadmium stress. J Plant Physiol doi:10.1016/j.jplph.2007.06.006
Fiske CH, SubbaRow Y (1925) Colorimetric determination of phosphorus. J Biol
Chem 66:375-400.
Fismes J, Vong PC, Guckert A, Frossard E (2000) Influence of sulphur on
apparent N-use efficiency, yield and quality of oilseed rape (Brassica napus
L.) grown on a calcareous soil. Europ J Agron 12:127-141.
136

Fitzgerald MA, Ugalde TD, Anderson JW (1999) Sulphur nutrition changes the
source of S in vegetative tissues of wheat during generative growth. J Exp
Bot 50:499-508.
Fomazier RF, Ferreira RR, Pereira GJG, Molina SMG, Smith RJ, Lea PJ, Azevedo
RA (2002b) Cadmium stress in sugarcane callus cultures: effects on antioxidant
enzymes. Plant Cell Tissue Organ Cult 71:125-131.
Fornazier RF, Ferreira RR, Vitoria AP, Molina SMG, Lea PJ, Azevedo RA
(2002a) Effects of cadmium on antioxidant enzyme activities in sugar cane.
Biol Plant 45:91-97.
Foyer CH (2001) Prospects for enhancement of the soluble antioxidants,
ascorbate and glutathione. BioFactors 15:75-78.
Foyer CH, Halliwell B (1976) The presence of glutathione and glutathione reductase
in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 133:21-25.
Foyer CH, Harbinson J (1994) Oxygen metabolism and the regulation of
photosynthetic electron transport. In: Foyer CH, MuUineaux P (Eds) Causes
of photooxidative stresses and amelioration of defense systems in plants.
CRC Press, Boca Raton, FL, USA, ppl-42.
Foyer CH, Lelandais M (1996) A comparison of the relative rates of transport of
ascorbate and glucose across the thylakoid, chloroplast and plasmalemma
membranes of pea leaf mesophyll cells. J Plant Physiol 148:391-398.
Foyer CH, Lelandais M, Kunert KJ (1994) Photooxidative stress in plants.
Physiol Plant 92:696-717.
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen peroxide and
glutathione associated mechanisms of acclimatory stress tolerance and
signaling. Physiol Plant 100:241-254.
Foyer CH, MuUineaux PM (1994) Causes of photooxidative stress and
amelioration of defense systems in plants. CRC Press, Boca Raton, FL, USA.
Foyer CH, Noctor G (2000) Oxygen processing in photosynthesis: regulation and
signaling. New Phytol 146:359-388.
Foyer CH, Noctor G (2001) The molecular biology and metabolism of
glutathione. In: Grill D, Tausz M, De Kok LJ (Eds) Significance of
glutathione to plant adaptation to the environment. Kluwer Academic
Publishers, The Netherlands, pp27-57.
137

Foyer CH, Noctor G (2003) Redox sensing and signaling associated witli reactive
oxygen in chloroplasts, peroxisomes and mitochondria. Piiysiol Plant
119:355-364.
Foyer CH, Noctor G (2005) Oxidant and antioxidant signaling in plants: a re-
evaluation of the concept of oxidative stress in a physiological context. Plant
Cell Environ 28:1056-1071.
Foyer CH, Rowell J, Walker D (1983) Measurements of the ascorbate content of
spinach leaf protoplasts and chloroplasts during illumination. Planta 157:239-
244.
Foyer CH, Souriau N, Perret S, Lelandais M, Kunert KJ, Pruvost C, Jouanin L
(1995) Overexpression of glutathione reductase but not glutathione
synthetase leads to increases in antioxidant capacity and resistance to
photoinhibition in poplar trees. Plant Physiol 109:1047-1057.
Foyer CH, Vanacker H, Gomez LD, Harbinson J (2002) Regulation of
photosynthesis and antioxidant metabolism in maize leaves at optimal and
chilling temperatures: review. Plant Physiol Biochem 40:659-668.
Freeman JL, Persan MW, Nieman K, Albrecht C, Peer W, Pickering IJ, Salt DE
(2004) Increased glutathione biosynthesis plays a role in nickel tolerance in
Thlaspi nickel hyperaccumulators. The Plant Cell 16:2176-2191.
Friedrich JW, Schrader LE (1978) Sulfur deprivation and nitrogen metabolism in
maize seedlings. Plant Physiol 61:900-903.
Frugoli JA, Zhong HH, Nuccio ML, McCourt P, McPeek MA, Thomas TL,
McClung CR (1996) Catalase is encoded by a multigene family in
Arabidopsis thaliana (L.). Plant Physiol 112:327-336.
Gachomo WE, Shonukan 0 0 , Kotchoni OS (2003) The molecular initiation and
subsequent acquisition of disease resistance in plants. Afr J Biotechnol 2:26-
32.
Gallego SM, Benavides MP, Tomaro ML (1996a) Effect of heavy metal ion
excess on sunflower leaves: evidence for involvement of oxidative stress.
Plant Sci 121:151-159.
Gallego SM, Benavides MP, Tomaro ML (1996b) Oxidative damage caused by
cadmium chloride in sunflower. Phyton 58:41-52.
Gallego SM, Benavides MP, Tomaro ML (1999) Effect of cadmium ions on
antioxidant defense system in sunflower cotyledons. Biol Plant 42:49-55.
138

Gallego SM, Benavides MP, Tomaro ML (2002) Involvement of an antioxidant
defence system in the adaptive response to heavy metal ions in Helianthus
annuus L. cells. Plant Growth Regul 36:267-273.
Gebler A, Schneider S, Weber P, Hanemann U, Rennenberg H (1998) Soluble N
compounds in trees exposed to high load of N: a comparison between the
roots of Norway spruce (Picea abies) and beech (Fagus sysvatica) trees
grown under field conditions. New Phytol 138:385-399.
Geisweid HJ, Urbach W (1983) Absorbtion of cadmium by the green microalgae
Chlorella vulgaris, Ankistrodesmus braunii and Eremosphaera viridis. Z
Pflanzenphysiol 109:127-142.
Ghani A, Wahid A (2007) Varietal difference for cadmium induced seedling mortality
and foliar-toxicity symptoms in mungbean (Vigna radiata). Int J Agric Biol
9:555-558.
Ghnaya T, Nouairi I, Slama I, Messedi D, Grignon C, Abdelly C, Ghorbel MH (2005)
Cadmium effects on growth and mineral nutrition of two halophytes: Sesuvium
portulacastrum and Mesembryanthemum crystallinum. J Plant Physiol 162:1133-
1140.
Ghnaya T, Slama I, Messedi, D, Grignon C, Ghorbel MH, Abdelly C (2007)
Effects of cadmium on activity of nitrate reductase and on other enzymes of
the nitrate assimilation pathway in bean. Plant Physiol Biochem 38:629-638.
Ghoshrony S, Nadakavukaren KJ (1990) Influence of cadmium on the ultrastructure
of developing chloroplasts of soybean and com. Environ Exp Bot 30:187-192.
Gianazza E, Wait R, Sozzi A, Regondi S, Saco D, Labra M, Agradi E (2007)
Growth and protein profile changes in Lepidium sativum L. plantlets exposed
to cadmium. Environ Exp Bot 59:179-187.
Gichner T, Patkova Z, Szakova J, Demnerova K (2004) Cadmium induces DNA
damages in tobacco roots, but no DNA damage, somatic mutations or
homologous recombinations in tobacco leaves. Mut Res Genetic Toxic
Environ Mut 559:49-57.
Godbold DL, Hutterman A (1985) Effect of zinc, cadmium and mercury on root
elongation on Picea abies (Karst.) seedlings and the significance of these
metals to forest die-back. Environ Pollut 38:375-381.
139

Goldsbrough P (2000) Metal tolerance in plants: the role of phytochelatins and
metallothioneins. In: Terry N, Banuelos G (Eds) Phytoremediation of
contaminated soil and water. CRC Press, LLC, pp221-233.
Gossett DR, Millhollon EP, Lucas MC, Banks SW, Marney MM (1994) The
effects of NaCl on antioxidant enzyme activities in callus tissue of salt-
tolerant and salt-sensitive cotton cultivars (Gossypium hirsutum L.). Plant
Cell Rep 13:498-503.
Gouia H, Ghobal MH, Meyer C (2000) Effects of cadmium on activity of nitrate
reductase and on other enzymes of the nitrate assimilation pathway in bean.
Plant Physiol Biochem 38:629-638.
Gouia H, Suzuki A, Brulfert J, Ghorbal MH (2003) Effects of cadmium on the
co-ordination of nitrogen and carbon metabolism in bean seedlings. J Plant
Physiol 160:367-376.
Grant CA, Buckley WT, Bailey LD, Selles F (1998) Cadmium accumulation in
crops. Can J Plant Sci 78:1-17.
Grant CA, Clarke JM, Duguid S, Chaney RL (2008) Selection and breeding of
plant cultivars to minimize cadmium accumulation. Sci Total Environ
doi:10.1016/j.scitotenv.2007.10.038
Gratao PL, Gomes-Junior RA, Delite FS, Lea PJ, Azevedo RA (2006)
Antioxidant stress responses of plants to cadmium. In: Khan NA, Samiullah
(Eds) Cadmium toxicity and tolerance in plants. Narosa Publishing House,
New Delhi, ppl-34.
Gratao PL, Polle A, Lea PJ, Azevedo RA (2005) Making the life of heavy metal-
stressed plants a little easier. Funct Plant Biol 32:481-494.
Gray JI (1978) Measurement of lipid oxidation: A review. J Am Oil Chem Soc
55:539-546.
Greger M, Bertell G (1992) Effects of Ca^^ and Cd^^ on the carbohydrate
metabolism in sugar beet {Beta vulgaris). J Exp Bot 43:167-173.
Greger M, Brammer E, Lindberg S, Larsson G, Idestam-Almquist J (1991) Uptake
and physiological effects of cadmium in sugar beet {Beta vulgaris) related to
mineral provision. J Exp Bot 42:729-737.
Greger M, Johansson M (1992) Cadmium effects on leaf transpiration of sugar
beet {Beta vulgaris). Physiol Plant 86:465-473.
140

Greger M, Lindberg S (1986) Effects of Cd •*" and EDTA on young sugar beets
(Beta vulgaris). I. Cd " uptake and sugar accumulation. Physiol Plant 66:69-
74.
Greger M, Lindberg S (1987) Effects of Cd^^ and EDTA on young sugar beets
(Beta vulgaris). II. Net uptake and distribution of Mg " , Ca " and Fe^^/Fe .
Physiol Plant 69:81-86.
Greger M, Ogrer E (1991) Direct and indirect effect of Cd^^ on photosynthesis in
sugar beet (Beta vulgaris). Physiol Plant 83:129-135.
Grill E, Winnacker E-L, Zenk MH (1985) Phytochelatins: The principal heavy-
metal complexing peptides of higher plants. Science 230:674-676.
Groppa MD, Tomaro ML, Benavides MP (2001) Polyamines as protectors
against cadmium or copper-induced oxidative damage in sunflower leaf discs.
Plant Sci 161:481-488.
Gueta-Dahan Y, Yaniv Z, Zilinskas BA, Ben-Hayyim G (1997) Salt and
oxidative stress: similar and specific responses and their relation to salt
tolerance in citrus. Planta 203:460-469.
Guo B, Liang YC, Zhu YG, Zhao FJ (2007) Role of salicylic acid in alleviating
oxidative damage in rice roots (Oryza sativa) subjected to cadmium stress.
Environ Pollut 146:743-749.
Guo T, Zhang G, Zhou M, Wu F, Chen J (2004) Effects of aluminum and cadmium
toxicity on growth and antioxidant enzyme activities of two barley genotypes with
different Al resistance. Plant Soil 258:241-248.
Gupta DK, Tohoyama H, Joho M, Inouhe M (2002) Possible role of
phytochelatins and glutathione metabolism in cadmium tolerance in chickpea
roots. J Plant Res 115:429-437.
Gussarsson M, Asp H, Adalsteinsson S (1996) Enhancement of cadmium effects on
growth and nutrient composition of birch (Betula pendula) by buthionine
sulphoximine (BSO). J Exp Dot 47:211-215.
Haag-Kerwer A, Schafer HJ, Heiss S, Walter C, Rausch T (1999) Cadmium exposure
in Brassica juncea causes a decline in transpiration rate and leaf expansion
without effect on photosynthesis. J Exp Bot 50:1827-1835.
Hall DO, Rao KK (1999) Photosynthesis, 6" Ed. Cambridge University Press,
Cambridge, pp214.
141

Hall JD, Barr R, Al-Abbas AH, Crane FL (1972) The ultrastructure of chloroplasts in
mineral deficient maize leaves. Plant Physiol 50:404-409.
Hall JL (2002) Cellular mechanisms for heavy metal detoxification and
tolerance. J Exp Bot 53:1-11.
Haller E, Suter M, Brunold C (1986) Regulation of ATP-sulfurylase and
adenosine 5'-phosphosulfate sulfotransferase by the sulfur and the nitrogen
source in heterotrophic cell suspension cultures of Paul's scarlet rose. J Plant
Physiol 125:275-286.
Hampp R, Beulich K, Ziegler H (1976) Effect of zinc and cadmium on
photosynthetic C02-fixation and Hill activity of isolated spinach chloroplasts.
Z Pflanzenphysiol 77:336-344.
Hanson EA, Barrien BS, Wood JG (1941) Relations between protein-nitrogen,
protein-sulphur and chlorophyll in leaves of Sudan grass. Aust J Exp Biol
19:231-234.
Harada E, Eui Choi Y, Tsuchisaka A, Obata H, Sano H (2001) Transgenic
tobacco plants expressing a rice cysteine synthase gene are tolerant to toxic
levels of cadmium. J Plant Physiol 158:655-661.
Harada E, Yamaguchi Y, Koizumi N, Sano H (2002) Cadmium stress induces
production of thiol compounds and transcripts for enzymes involved in sulfur
assimilation pathway in Arabidopsis. J Plant Physiol 159:445-448.
Hart JJ, Welch RM, Norvell WA, Sullivan LA, Kochian LV (1998)
Characterization of cadmium binding, uptake, and translocation in intact
seedlings of bread and durum wheat cultivars. Plant Physiol 116:1413-1420.
Hasan SA, Ali B, Hayat S, Ahmad A (2007) Cadmium-induced changes in the
growth and carbonic anhydrase activity of chickpea. Turk J Biol 31:137-140.
Hasan SA, Hayat S, Ali B, Ahmad A (2008) 28-Homobrassinolide protects
chickpea (Cicer arietinum) from cadmium toxicity by stimulating
antioxidants. Environ Pollut 151:60-66.
Hassan MJ, Shafi M, Zhang G, Zhu Z, Qaisar M (2008a) The growth and some
physiological responses of rice to Cd toxicity as affected by nitrogen form. Plant
Growth Regul 54:125-132.
Hassan MJ, Wang F, Ali S, Zhang G (2005a) Toxic effect of cadmium on rice as
affected by nitrogen fertilizer form. Plant Soil 277:359-365.
142

Hassan MJ, Wang Z, Zhang G (2005b) Sulfur alleviates growth inhibition and
oxidative stress caused by cadmium toxicity in rice. J Plant Nutr 28:1785-1800.
Hassan MJ, Zhang G, Zhu Z (2008b) Influence of cadmium toxicity on plant growth
and nitrogen uptake in rice as affected by nitrogen form. J Plant Nutr 31:251-262.
Hassan MJ, Zhu Z, Ahmad B, Mahmood Q (2006) Influence of cadmium toxicity on
rice genotypes as affected by zinc, sulfur and nitrogen fertilizers. Casp J Env Sci
4:1-8.
Havaux M, Bonfils J-P, Lutz C, Niyogi KK (2000) Photodamage of the
photosynthetic apparatus and its dependence on the leaf developmental stage in
the npql Arabidopsis mutant deficient in the xanthophyll cycle enzyme
violaxanthin de-epoxidase. Plant Physiol 124:273-284.
Hawkesford MJ, De Kok LJ (2006) Managing sulfur metabolism in plants. Plant Cell
Environ 29:282-295.
Hawkesford MJ, Howarth JR, Buchner P (2006) Control of sulfur uptake, assimilation
and metabolism. In: WC Plaxton WC, McManus MT (Eds) Control of primary
metabolism in plants, Armual Plant Reviews, Vol 22, Blackwell Publishing,
Oxford, pp348-372.
Hayat S, Ali B, Hasan SA, Ahmad A (2007) Brassinosteroids enhanced the level of
antioxidants imder cadmium stress in Brassicajuncea. Environ Exp Bot 60:33-41.
He J, Zhu C, Ren Y, Yan Y, Jiang D (2006) Genotypic variation in grain cadmium
concentration of lowland rice. J Plant Nutr Soil Sci 169:711-716.
Hegedus A, Erdei S, Horvath G (2001) Comparative studies of H2O2 detoxifying
enzymes in green and greening barley seedlings under cadmium stress. Plant
Sci 160:1085-1093.
Heiss S, Schafer H, Haag-Kerwer A, Rausch T (1999) Cloning sulfur
assimilation genes of Brassica juncea L.: cadmium differentially affects the
expression of a putative low affinity sulfate transporter and isoforms of ATP
sulfurylase and APS reductase. Plant Mol Biol 39:847-857.
Hemingway JS (1976) Mustards. In: Simmonds NW (Ed) Evolution of crop
plants. Longman Group, London, pp56-59.
Hendry GAP, Baker AJM, Ewart CF (1992) Cadmium tolerance and toxicity,
oxygen radical processes and molecular damage in cadmium-tolerant and
cadmium-sensitive clones of Holcus lanatus. Acta Bot Neerl 41:271-281.
143

Henry RP (1996) Multiple roles of carbonic anhydrase in cellular transport and
metabolism. Annu Rev Physiol 58:523-538.
Herbette S, Taconnat L, Hugouvieux V, Piette L, Magniette M-LM, Cuine S,
Auroy P, Richuad P, Forestier C, Bourguignon J, Renou J-P (2006) Genome-
wide transcriptome profiling of the early cadmium response of Arabidopsis
roots and shoots. Biochimie 88:1751-1765.
Hernandez LE, Olguin E, Trujillo SY, Vivanco J (1997) Recycling and treatment
of anaerobic effluents from pig waste, using Lemna sp. under temperate
climatic conditions. In: Wise DL (Ed) Global environmental biotechnology.
Elsevier, The Netherlands, pp293-304.
Hernandez LE, Carpena-Ruiz R, Garate A (1996) Alterations in the mineral
nutrition of pea seedlings exposed to cadmium. J Plant Nutr 19:1581-1598.
Hesse H, Nikiforova V, Gskieare B, Hoefgen R (2004) Molecular analysis and
control of cysteine biosynthesis: integration of nitrogen and sulfur
metabolism. J Exp Bot 55:1283-1292.
Hiscox JD, Israelstam GF (1979) A method for the extraction of chlorophyll from leaf
tissue without maceration. Can J Bot 57:1132-1134.
Hochmuth G J, Brecht JK, Bassett MJ (1999) Nitrogen fertilization to maximize
carrot yield and quality on a sandy soil. HortSci 34:641-645.
Horvath G, Droppa M, Oravecz A, Raskin VI, Marder JB (1996) Formation of
photosynthetic apparatus during greening of cadmium-poisoned barley
leaves. Planta 199:238-243.
Hosayama Y, Daiten S, Sakai MI (1994) Histochemical demonstration of
cadmium in plant tissues. Kenkyu Kiyo-Nihon daigaku Bunrigakabu shizen
Kagaku Kenkyusho 29:107-110.
Howden R, Anderson C, Goldsbrough P, Cobbett C (1995a) A cadmium-sensitive,
glutathione-deficient mutant oi Arabidopsis thaliana. Plant Physiol 107:1067-
1073.
Howden R, Goldsbrough P, Anderson C, Cobbett C (1995b) Cadmivmi-sensitive, cadi
mutants of Arabidopsis thaliana are phytchelatin deficient. Plant Physiol
107:1059-1066.
Hsu YT, Kao CH (2004) Cadmium toxicity is reduced by nitric oxide in rice
leaves. Plant Growth Regul 42:227-238.
144

Hsu YT, Kuo MC and Kao CH (2006) Cadmium-induced ammonium ion
accumulation of rice seedlings at high temperature is mediated through abscisic
acid. Plant Soil 287:267-277.
Hu SX, Lau KWK, Wu M (2001) Cadmium sequestration in Chalamydomonas
reinhardtii. Plant Sci 161:987-996.
Huang CY, Bazzaz FA, Vanderhoef LN (1974) The inhibition of soybean
metabolism by cadmium and lead. Plant Physiol 54:122-124.
Hung KT, Kao CH (1998) Involvement of lipid peroxidation in methyl
jasmonate-promoted senescence in detached rice leaves. Plant Growth Regul
24:17-21.
lannelli MA, Pietrini F, Fiore L, Petrilli L, Massacci A (2002) Antioxidant
response to cadmium in Phragmites australis plants. Plant Physiol Biochem
40:977-982.
Inouhe M (2005) Phytochelatins. Braz J Plant Physiol 17:65-65.
Inouhe M, Ito R, Ito S, Sasada N, Tohoyama H, Joho M (2000) Azuki bean cells
are hypersensitive to cadmium and do not synthesize phytochelatins. Plant
Physiol 123:1029-1036.
Ishikawa T, Yoshimura K, Sakai K, Tamoi M, Takeda T, Shigeoka S (1998)
Molecular characterization and physiological role of a glyoxysome-bound
ascorbate peroxidase from spinach. Plant Cell Physiol 39:23-34.
Jackson AP, Alio way BJ (1992) Transfer of cadmium from soils to the human food
chain. In: Adriano DC (Ed) Biogeochemistry of trace metals. Lewis Publisher,
Baton Rouge, pp 109-158.
Jackson T, MacGillivray A (1993) Accounting for cadmium. Stockholm
Environment Institute, London.
Jalil A, Selles F and Clarke JM (1994) Effect of cadmium on growth and uptake
of cadmium and other elements by durum wheat. J Plant Nutr 17:1839-1858.
Jana S, Choudhuri MA (1981) Glycolate metabolism of three submerged aquatic
angiosperms during aging. Aquat Bot 12:345-354.
Jaworski EG (1971) Nitrate reductase assay in intact plant tissues. Biochem Biophy
Res Commun 43:1274-1279.
Jensen A, Bro-Rasmussen F (1992) Environmental contamination in Europe. Rev
Environ Contam Toxicol 125:101-181.
145

Jiang W, Liu D, Hou W (2001) Hyperaccumulation of cadmium by roots, bulbs and
shoots of garlic {Allium sativum L.). Biores Technol 76:9-13.
Jiang XJ, Luo YM, Liu Q, Liu SL, Zhao QG (2004) Effects of cadmium on nutrient
uptake and translocation by Indian mustard. Environ Geochem Health 26:319-
324.
Jimenez A, Hernandez J A, Pastori G, Del Rio LA, Sevilla F (1998) Role of the
ascorbate-glutathione cycle of mitochondria and peroxisomes in the senescence of
pea leaves. Plant Physiol 118:1327-1335.
Jing D, Wu F-B, Zhang G-P (2005) Effect of cadmium on growth and photosynthesis
of tomato seedlings. J Zhejiang Univ SCI 68:974-980.
Joel G, Gamon JA, Field CB (1997) Production efficiency in sunflower: The role of
water and nitrogen stress. Remote Sensing Environ 62:176-188.
Jones KC, Jackson A, Johnston AE (1992) Evidence for an increase in the Cd
content of herbage since 1860's. Environ Sci Technol 26:834-836.
Kabata-Pendias A, Pendias H (2001) Trace elements in soils and plants. 3 '' edition.
CRC Press, Boca Raton, FL, USA, pp413.
Kachroo D, Kumar A (1997) Nitrogen and sulphur fertilization in relation to yield
attributes and seed yield of Indian mustard {Brassica juncea). Ind J Agron 42:145-
147.
Kaiser WM, Behnisch EB (1991) Rapid modulation of spinach leaf nitrate reductase
by photosynthesis. I. Modulation in vivo by CO2 availability. Plant Physiol
96:363-367.
Kato M, Shimizu S (1985) Chlorophyll metabolism in higher plants. VI.
Involvement of peroxidase in chlorophyll degradation. Plant Cell Physiol
26:1291-1301.
Khan NA (1994) Variation in carbonic anhydrase activity and its relationship
with dry mass of mustard. Photosynthetica 30:317-320.
Khan NA, Ahmad I, Singh S, Nazar R (2006) Variation in growth,
photosynthesis and yield of five wheat cultivars exposed to cadmium stress.
World J Agric Sci 2:223-226.
Khan NA, Javid S, Samiullah (2004) Physiological role of carbonic anhydrase in CO2
fixation and carbon partitioning. Physiol Mol Biol Plants 10:153-166.
146

Khan NA, Mobin M, SamiuUah (2005) The influence of gibberelHc acid and sulfur
fertilization rate on growth and S-use efficiency of mustard {Brassica juncea).
Plant Soil 270:269-274.
Khan NA, SamiuUah, Singh S, Nazar R (2007a) Activities of antioxidative
enzymes, sulphur assimilation, photosynthetic activity and growth of wheat
{Triticum aestivum) cultivars differing in yield potential under cadmium
stress. J Agron Crop Sci 193:435-444.
Khan NA, Singh S (2008) Abiotic stress and plant responses. IK international
publishing house. New Delhi, India.
Khan NA, Singh S, Anjum NA, Nazar R (2008b) Cadmium effects on carbonic
anhydrase, photosynthesis, dry mass and antioxidative enzymes in wheat
{Triticum aestivum) under low and sufficient zinc. J Plant Inter 3:31-37.
Khan NA, Singh S, Nazar R, Lone PM (2007b) The source-sink relationship in
mustard. The Asian Aust J Plant Sci Biotechnol 1:10-18.
Khan NA, Singh S, Umar S (2008a) Sulfur assimilation and abiotic stress in plants.
Springer-Verlag, Berlin, Heidelberg, Germany.
Khan S, Khan NN (1983) Influence of lead and cadmium on the growth and
nutrient concentration of tomato and egg plant. Plant Soil 74:387-394.
Khudsar T, Mahmooduzaffar, Iqbal M (2001) Cadmium induced changes in leaf
epidermis, net photosynthetic rate and pigment concentration in Cajanus cajan.
Biol Plant 44:59-64.
Kim J-S, Jung S, Hwang IT, Cho K-Y (1999) Characteristics of chlorophyll a
fluorescence induction in cucumber cotyledons treated with diuron, norflurazon,
and sulcotrione. Pesticide Biochem Physiol 65:73-81.
Kim J-S, Yun B-W, Choi JS, Kim T-J, Kwak S-S, Cho K-Y (2004) Death
mechanisms caused by carotenoid biosynthesis inhibitors in green and in
undeveloped plant tissues. Pesticide Biochem Physiol 78:127-139.
Klein BP, Grossmann S, King D, Cohen BS, Pinskuy A (1984) Pigment bleaching,
carbonyl production and antioxidant effects during anaerobic lipoxygenase
reaction. Biochim Biophys Acta 793:72-79.
Kochian LV (1991) Mechanisms of micronutrient uptake and translocation in
plants. In: Mortvedt JJ (Ed) Micronutrients in agriculture. Soil Sci Soc Am,
Madison, Wisconsin, pp251-270.
147

Kopriva S, Koprivova A (2005) Sulfate assimilation and glutathione synthesis in
C4 plants. Photosyn Res 86:363-372.
Kopriva S, Rennenberg H (2004) Control of sulphate assimilation and
glutathione synthesis: interaction with N and C metabolism. J Exp Bot
55:1831-1842.
Kopriva S, Suter M, von Ballmoos P, Hesse H, Krahenbuhl U, Rennenberg H,
Brunold C (2002) Interaction of sulfate assimilation with carbon and nitrogen
metabolism in Lemna minor. Plant Physiol 130:1406-1413.
Koprivova A, Suter M, Op den Camp R, Brunold C, Kopriva S (2000)
Regulation of sulphate assimilation by nitrogen in Arabidopsis. Plant Physiol
122:737-746.
Kotchoni OS, Gachomo EW (2006) The reactive oxygen species network
pathways: an essential prerequisite for perception of pathogen attack and the
acquired disease resistance in plants. J Biosci 31:389-404.
Kouril R, Lazar D, Lee H, Jo J, Naus J (2003) Moderately elevated temperature
eliminates resistance of rice plants with enhanced expression of glutathione
reductase to intensive photooxidative stress. Photosynthetica 41:571-580.
Kovacik J, Tomko J, Backor M, Repcak M (2006) Matricaria chamomilla is not
a hyperaccumulator, but tolerant to cadmium stress. Plant Growth Regul
50:239-247.
Krantev A, Yordanova R, Janda T, Szalai G, Popova L (2007) Treatment with
salicylic acid decreases the effect of cadmium on photosynthesis in maize
plants. J Plant Physiol doi:10.1016/j.jplph.2006.11.014
Krapp A, Stitt M (1995) An evaluation of direct and indirect mechanisms for the
"sink regulation" of photosynthesis in spinach: Changes in gas exchange,
carbohydrates, metabolites, enzyme activities and steady-state transcript
levels after cold-girdling source leaves. Planta 195:313-323.
Kreps JA, Wu Y, Chang HS, Zhu T, Wang X, Harper JF (2002) Transcriptome
changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant
Physiol 130:2129-2141.
Krupa Z (1999) Cadmium against higher plants photosynthesis- A variety of
effects and where do they possibly come from? Zeitschrift fur
Naturforschung 54:723-729.
148

Krupa Z, Baszynski T (1995) Some aspects of heavy metals toxicity towards
photosynthetic apparatus direct and indirect effects on light and dark
reactions. Acta Physiol Plant 17:177-190.
Kubo A, Saji H, Tanaka K, Kondo N (1995) Expression of Arabidopsis cytosolic
ascorbate peroxidase gene in response to ozone or sulfur dioxide. Plant Mol
Biol 29:479-489.
Kuboi T, Noguchi A, Yazaki J (1987) Relationship between tolerance and
accumulation characteristics of cadmium in higher plants. Plant Soil 104:275-
280.
Kumaran S, Francois JA, Krishnan HB, Jez JM (2008) Regulatory protein-
protein interactions in primary metabolism: the case of the cysteine synthase
complex. In: Khan NA, Singh S, Umar S (Eds) Sulfur assimilation and
abiotic stresses in plants. Springer-Verlag, The Netherlands, pp97-109.
Kunert KJ, Foyer C (1993) Thiol/disulfide exchange in plants. In: De Kok LJ,
Stulen I, Rennenberg H, Brunold C, Rauser WE (Eds) Sulfur nutrition and
assimilation in higher plants: regulatory, agricultural and environmental
aspects. SPB Academic Publishing, The Hague, pp 139-151.
Kuo MC, Kao CH (2004) Antioxidant enzyme activities are upregulated in
response to cadmium in sensitive, but not in tolerant, rice {Oryza sativa L.)
seedlings. Bot Bull Acad Sin 45:291-299.
Kupper H, Kupper F, Spiller M (1996) Environmental relevance of heavy metal
substituted chlorophylls using the example of water plants. J Exp Bot 47:259-
266.
Kuriakose SV, Prasad MNV (2008) Cadmium stress affects seed germination and
seedling growth in Sorghum bicolor (L.) Moench by changing the activities of
hydrolyzing enzymes. Plant Growth Regul 54:143-156.
Lagriffoul A, Mocquot B, Mench M, Vangronsveld J (1998) Cadmium toxicity effects
on growth, mineral and chlorophyll contents, and activities of stress related
enzymes in young maize plants {Zea mays L.). Plant Soil 200:241-250.
Lakkineni KC, Abrol YP (1992) Sulphur requirement of rapeseed-mustard,
groundnut and wheat: a comparative assessment. J Agron Crop Sci 169:281-
285.
Lakkineni KC, Abrol YP (1994) Sulfur requirement of crop plants: physiological
analysis. Fert News 39:11-18.
149

Lancien M, Gadal P, Hodges M (2000) Enzyme redundancy and the importance
of 2-oxoglutarate in higher plant ammonium assimilation. Plant Physiol
123:817-824.
Lang F, Sarvari E, Szigeti Z, Fodor F, Cseh E (1995) Effects of heavy metals on
the photosynthetic apparatus in cucumber. In: Mathis P (Ed) Photosynthesis:
from light to biosphere IV. Kluwer Academic Publishers, The Netherlands,
pp533-536.
Larson RA (1988) The antioxidants of higher plants. Phytochem 27:969-978.
Larsson EH, Bornman JF, Asp H (1998) Influence of UV-B radiation and Cd^^
on chlorophyll fluorescence, growth and nutrient content in Brassica napus. J
Exp Bot 323:1031-1039.
Lascano HR, Casano LM, Melchiorre MN, Trippi VS (2001) Biochemical and
molecular characterization of wheat chloroplastic glutathione reductase. Biol
Plant 44:509-516.
Lascano HR, Gomez LD, Casano LM, Trippi VS (1999) Wheat chloroplastic
glutathione reductase activity is regulated by the combined deffect of pH,
NADPH and GSSG. Plant Cell Physiol 40:683-690.
Law ME, Charles SA, Halliwell B (1983) Glutathione and ascorbic acid in spinach
{Spinacia oleracea) chloroplasts: the effect of hydrogen peroxide and of paraquat.
Biochem J 210:899-903.
Lea PJ, Miflin BJ (2004) Glutamate synthase and the synthesis of glutamate in plants.
Plant Physiol. Biochem 41:555-564.
Lee S, Leustek T (1999) The affect of cadmium on sulfate assimilation enzymes
in Brassica juncea. Plant Sci 141:201-207.
Leita L, Marchiol L, Martin M, Peressotti A, Delle Vedove G, Zerbi G (1995)
Transpiration dynamics in cadmium-treated soybean {Glycine max L.) plants.
J Agron Crop Sci 175:153-156.
Leon AM, Palma JM, Corpas FJ, Gomez M, Romero-Puertas MC, Chatterjee D,
Mateos RM, del Rio LA, Sandalio LM (2002) Antioxidant enzymes in
cultivars of pepper plants with different sensitivity to cadmium. Plant Physiol
Biochem 40:813-820.
Leustek T, Martin MN, Bick J-A, Davies JP (2000) Pathways and regulation of
sulfur metabolism revealed through molecular and genetic studies. Annu Rev
Plant Physiol Plant Mol Biol 51:141-165.
150

Li EH, Miles CD (1975) Effects of cadmium on photoreactions of chloroplasts.
Plant Sci Lett 5:33-70.
Li Y-M, Scaney R, Schneiter A, Miller J, Elias E, Hammond J (1997) Screening
for low grain cadmium phenotypes in sunflower, durum wheat and flax.
Euphytica 94:23-30.
Lindner RC (1944) Rapid analytical methods for some of the more common inorganic
constituents of plant tissues. Plant Physiol 19:70-89.
Liu CP, Shen ZG, Li XD (2007) Accumulation and detoxification of cadmium in
Brassicapekinensis and B. chinensis. Biol Plant 51:116-120.
Liu HJ, Zhang JL, Zhang FS (2007) Role of iron plaque in Cd uptake by and
translocation within rice {Oryza sativa L.) seedlings grown in solution culture.
Environ Exp Bot 59-314-320.
Liu X, Zhang S, Shan XQ, Christie P (2007) Combined toxicity of cadmium and
arsenate to wheat seedlings and plant uptake and antioxidative enzyme responses
to cadmiimi and arsenate co-contamination. Ecotoxicol Environ Saf 68:305-313.
Liu Y, Wang X, Zeng G, Qu D, Gu J, Zhou M, Chai L (2007) Cadmium-induced
oxidative stress and response of the ascorbate-glutathione cycle in Bechmeria
nivea (L.) Gaud. Chemosphere 69:99-107.
Loggini B, Scartazza A, Brugnoli E, Navari-izzo F (1999) Antioxidant defense
system, pigment composition and photosynthetic efficiency in two wheat
cultivars subjected to drought. J Plant Physiol 119:1091-1099.
Lombi E, Zhao FJ, Dunham S, McGrath SP (2000) Cadmium accumulation in
populations of Thlaspi caerulescens and Thlaspi goesingense. New Phytol 145:11-
20.
Lorimer GH (1981) The carboxylation and oxygenation of ribulose-l-5-bisphosphate:
the primary events in photosynthesis and photorespiration. Annu Rev Plant
Physiol 32:349-383.
Lozano-Rodriguez E, Hernandez LE, Bonay P, Carpena-Ruiz RO (1997)
Distribution of cadmium in shoot and root tissues of maize and pea plants:
physiological disturbances. J Exp Bot 306:123-128.
Macfarlane GR, Burchett MD (2001) Photosynthetic pigments and peroxidase
activity as indicators of heavy metal stress in the grey mangrove, Avicennia
marina (Forsk.). Vierh Mar Pollut Bull 42:233-240.
151

Mahajan S, Tuteja N (2005) Cold, salinity and drought stresses: An overview. Arch
Biochem Biophy 444:139-158.
Maier EA, Rosalyn DM, Jennifer AMD, Rebecca RW, Beth AA (2003)
Environmental cadmium levels increase phytochelatin and glutathione in
lettuce grown in a chelator-buffered nutrient solution. J Environ Qua!
32:1356-1364.
Majumdar S, Ghosh S, Glick BR, Dumbroff EB (1991) Activities of
chlorophyllase, phosphoenolpyruvate carboxylase and ribulose-1,5-
bisphosphate carboxylase in the primary leaves of soybean during senescence
and drought. Physiol Plant 81:473-480.
Makino A, Sakashita H, Hidema J, Mae T, Ojima K, Osmond B (1992) Distinctive
responses of ribulose-1,5- bisphosphate carboxylase and carbonic anhydrase in
wheat leaves to nitrogen nutrition and their possible relationships to CO2 transfer
resistance. Plant Physiol 100:1737-1743.
Malik D, Sheoran IS, Singh R (1992a) Lipid composition of thylakoid
membranes of cadmium treated wheat seedlings. Ind J Biochem Biophys
29:350-354.
Malik D, Sheoran IS, Singh R (1992b) Carbon metabolism in leaves of cadmium
treated wheat seedlings. Plant Physiol Biochem 30:223-229.
Mann T, Keilin D (1938) Homocuprein and heptacuprein, copper-protein
compounds of blood and liver in mammals. Proc R Soc Lond B 126:303-315.
Mapson LW, Goddord DR (1951) Reduction of glutathione by coenzyme II.
Nature 167:975-977.
Marrs KA (1996) The functions and regulation of glutathione S-transferases in
plants. Annu Rev Plant Physiol Plant Mol Biol 47:127-158.
Marrs KA, Walbot V (1997) Expression and RNA splicing of the maize glutathione
S-transferase Bronze 2 is regulated by cadmiimi and other sources. Plant Physiol
113:93-102.
Marschner H (1995) Mineral nutrition of higher plants. Academic Press, New York.
Martin TJ, Juniper EB (1970) The cuticles of plants. Edward Arnolds Ltd, Edinburgh.
Maruyama-Nakashita A, Inoue E, Saito K, Takahashi H (2007) Sulfur responsive
promoter of sulfate transporter gene is potentialy useful to detect and
quantify selenate and chromate. Plant Biotechnol 24:261-273.
152

Maughan S, Foyer CH (2006) Engineering and genetic approaches to modulating
the glutathione network in plants. Physiol Plant 126:382-397.
May MJ, Vernoux T, Leaver C, Van-Montagu M, Inze D (1998a) Glutathione
homeostasis in plants: Implications for environmental sensing and plant
development. J Exp Bot 49:649-667.
May MJ, Vernoux T, Sanchez-Fernandez R, Van Montagu M, Inze D (1998b)
Evidence for posttranscriptional activation of gamma-glutamylcysteine
synthetase during plant stress responses. Proc Natl Acad Sci USA 95:12049-
12054.
Mayfield SP, Taylor WC (1984) Carotenoid-deficient maize seedlings fail to
accumulate light-harvesting chlorophyll a/b binding protein (LHCP) mRNA.
Eur JBiochem 144:79-84.
McCord JM, Fridovich I (1969) Superoxide dismutase, an enzymatic function for
erythrocuprein. J Biol Chem 244:6049-6055.
McGrath SP, Zhao FJ (1996) Sulphur uptake, yield response and the interactions
between N and S in winter oilseed rape {Brassica napus). J Agric Sci
Cambridge 126:53-62.
McGrath SP, Zhao FJ (2003) Phytoextraction of metals and metalloids from
contaminated soils. Curr Opin Biotech 14:277-282.
McLaughlin MJ, Hamon RE, McLaren RG, Speir TW, Rogers SL (2000)
Review: A bioavailability-based rationale for controlling metal and metalloid
contamination of agricultural land in Australia and New Zealand. Austr J Soil
Res 38:1037-1086.
McMahon PJ, Anderson JW (1998) Preferential allocation of sulphur into Y-
glutamylcysteinyl peptides in wheat plants grown at low sulphur nutrition in
the presence of cadmium. Physiol Plant 104:440-448.
Medici LO, Azevedo RA, Smith RJ, Lea PJ (2004) The influence of nitrogen
supply on antioxidant enzymes in plant roots. Funct Plant Biol 31:1-9.
Meister A (1988) Glutathione metabolism and its selective modification. J Biol
Chem 263:17205-17208.
Meldrum NU, Tarr HLA (1935) The reduction of glutathione by the Warburg-
Christian system. Biochem J 29:108-115.
Metwally A, Safronova VI, Belimov AA, Dietz KJ (2005) Genotypic variation of
the response to cadmium toxicity in Pisum sativum L. J Exp Bot 56:167-178.
153

Migge A, Bork C, Hell R, Becker TW (2000) Negative regulation of nitrate
reductase gene expression by glutamine or asparagine accumulating in leaves
of sulfur-deprived tobacco. Planta 211:587-595.
Millar AH, Mittova V, Kiddle G, Heazlewood JL, Bartoli CG, Theodoulou FL,
Foyer CH (2003) Control of ascorbate synthesis by respiration and its
implication for stress responses. Plant Physiol 133:443-447.
Miller RJ, Biuell JE, Koeppe DE (1973) The effect of cadmium on electron and
energy transfer reactions in corn mitochondria. Physiol Plant 28:166-171.
Mills HA, Jones Jr JB (1996) Plant analysis handbook II. MicroMacro
Publishing, Athens, GA.
Milone MT, Sgherri C, Clijters H, Navari-Izzo F (2003) Antioxidative responses
of wheat treated with realistic concentrations of cadmium. Environ Exp Bot
50:265-273.
Mishra S, Srivastava S, Tripathi RD, Govindarajan R, Kuriakose SV, Prasad
MNV (2006) Phytochelatin synthesis and response of antioxidants during
cadmium stress in Bacopa monnieri. Plant Physiol Biochem 44:25-37.
Mittler R, Vanderauwers S, Gollery M, Van Breusegem F (2004) Reactive
oxygen gene network of plants. Trends Plant Sci 9:490-498.
Miyake C, Asada K (1992) Thylakoid-bound ascorbate peroxidant in spinach
chloroplasts and photoreduction of its primary oxidation product
monodehydro ascorbate radicals in thylakoid. Plant Cell Physiol 33:541-553.
Mobin M, Khan NA (2007) Photosynthetic activity, pigment composition and
antioxidative response of two mustard (Brassica juncea) cultivars differing in
photosynthetic capacity subjected to cadmium stress. J Plant Physiol
164:601-610.
Mohan BS, Hossetti BB (1997) Potential phytotoxicity of lead and cadmium to
Lemna minor grown in sewage stabilization ponds. Environ Pollut 98:233-
238.
Moller IM (2001) Plant mitochondria and oxidative stress: electron transport,
NADPH turnover, and metabolism of reactive oxygen species. Annu Rev
Plant Physiol Plant Mol Biol 52:561-591.
Moons A (2003) Osgstu3 and osgstu4, encoding tau class glutathione S-
transferase, are heavy metal- and hypoxic stress-induced and differetially salt
stress-responsive in rice roots. FEBS Let 553:427-432.
154

Morelli E, Scarano G (2004) Copper-induced changes of non-protein thiols and
antioxidant enzymes in the marine microalga Phaeodactylum tricomutum.
Plant Sci 167:289-296.
Mullineaux PM, Creissen GP (1997) Glutathione reductase: regulation and role
in oxidative stress. In: Scandalios JC (Ed) Oxidative stress and the molecular
biology of antioxidant denfenses. Cold Spring Harbor Laboratory Press, Cold
Spring Harbor, NY, pp667-713.
Mullineaux PM, Rausch T (2005) Glutathione, photosynthesis and the redox
regulation of stress-responsive gene expression. Photosynth Res 86:459-474.
Murakami M, Ae N, Ishikawa S (2007) Phytoextraction of cadmium by rice {Oryza
sativa L.), soybean {Glycine max (L.) Merr.), and maize {Zea mays L.). Environ
Pollut 145:96-103.
Mysliwa-Kurdziel B, Strzalka K (2002) Influence of metals on the biosynthesis of
photosynthetic pigments. In: Prasad MNV, Strzalka K (Eds) Physiology and
biochemistry of metal toxicity and tolerance in plants. Kluwer Academic
Publisher, Dordrecht, Netherlands, pp201-228.
Naguib MI, Hamed AA, Al-Wakeel SA (1982) Effect of cadmium on growth
criteria of some crop plants. Egypt J Bot 25:1-12.
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific
peroxidase in spinach chloroplasts. Plant Cell Physiol 22:867-880.
Nandi DL, Shemin D (1968) 5-aminolaevulinic acid dehydratase of
Rhodopseudomonas sphaaeroides. J Biol Chem 243:1236-1242.
Nathawat NS, Nair JS, Kumawat SM, Yadava NS, Singh G, Ramaswamy NK,
Sahu MP, D'Souza SF (2007) Effect of seed soaking with thiols on the
antioxidant enzymes and photosystem activities in wheat subjected to water
stress. Biol Plant 51:93-97.
Neuenschwander U, Suter M, Brunold C (1991) Regulation of sulfate
assimilation by light and 0-acetyl-L-serine in Lemna minor L. Plant Physiol
97:253-258.
Nieboer E, Richardson DHS (1980) The replacement of the nondescript term
'heavy metals' by a biologically and chemically significant classification of
metal ions. Environ Pollut (Series B) 1:2-26.
Nies DH (1999) Microbial heavy-metal resistance. Appl Microbiol Biotech
51:730-750.
155

Niyogi KK, Shih C, Chow WS, Pogson BJ, DellaPenna D, Bjorkman 0 (2001)
Photoprotection in a zeaxanthin- and lutein-deficient double mutant of
Arabidopsis. Photosynth Res 67:139-145.
Nocito FF, Lancilli C, Crema B, Fourcoy P, Davidian J-C, Sacchi GA (2006)
Heavy metal stress and sulfate uptake in maize roots. Plant Physiol 141:1138-
1148.
Nocito FF, Pirovano L, Cocucci M, Sacchi GA (2002) Cadmium-induced sulfate
uptake in maize roots. Plant Physiol 129:1872-1879.
Noctor G, Arisi ACM, Jouanin L, Foyer CH (1998a) Manipulation of glutathione
and amino acid biosynthesis in the chloroplast. Plant Physiol 118:471-482.
Noctor G, Arisi ACM, Jouanin L, Kunert KJ, Rennenberg H, Foyer CH (1998b)
Glutathione: biosynthesis, metabolism and relationship to stress tolerance
explored in transgenic plants. J Exp Bot 49:623-647.
Noctor G, Foyer CH (1998) Ascorbate and glutathione: Keeping active oxygen
under control. Annu Rev Plant Physiol Plant Mol Biol 49:249-279.
Noctor G, Gomez LA, Vanacker H, Foyer CH (2002) Interactions between
biosynthesis, comparmentation and transport in the control of glutathione
homeostasis and signaling. J Exp Bot 53:1283-1304.
Noji M, Saito K (2003) Sulfur amino acids: biosynthesis of cysteine and
methionine. In: Abrol YP, Ahmad A (Eds) Sulphur in plants. Kluwer
Academic Publishers, The Netherlands, pp 135-144.
Nolan AL, McLaughlin MJ, Mason SD (2003) Chemical speciation of Zn, Cd, Cu,
and Pb in pore waters of agricultural and contaminated soils using Donnan
dialysis. Environ Sci Technol 37:90-98.
Noriega GO, Balestrasse KB, BatUe A, Tomaro ML (2007) Cadmium induced
oxidative stress in soybean plants also by the accumulation of 6-
aminolaevulinic acid. Biometals 20:841-851.
Nriagu JO (1980) Production, uses and properties of cadmium. In: Nriagu JO
(Ed) Cadmium in the environment. Part I Wiley, New York, pp35-70.
Nriagu JO (1989) A global assessment of natural sources of atmospheric trace
metals. Nature 338:47-49.
Nriagu JO (1990) Global metal pollution. Poisoning the biosphere? Environ
32:7-33.
156

Nriagu JO, Pacyna JM (1988) Quantitative assessment of worldwide
contamination of air, water and soils by trace metals. Nature 333:134-139.
Nussbaum S, Schmutz D, Brunold C (1988) Regulation of assimilatory sulfate
reduction by cadmium in Zea mays L. Plant Physiol 88:1407-1410.
Oaks A (1994) Primary nitrogen assimilation in higher plants and its regulation.
Can J Bot 72:739-750.
OelmuUer R (1989) Photooxidative destruction of chloroplasts and its effect on
nuclear gene excpression and extraplastidic enzyme level. Photochem
Photobiol 49:229-239.
Ogawa K (2005) Glutathione-associated regulation of plant growth and stress
responses. Antiox Red Signal 7:973-981.
Okabe K, Yang S, Tsuzuki M, Miyachi S (1984) Carbonic anhydrase: Its content
in spinach leaves and its taxonomic diversity studied with anti-spinach leaf
carbonic anhydrase antibody. Plant Sci Lett 33:145-153.
Okamoto OK, Pinto E, Latorie LR, Becharie EJH, Colepicola P (2001)
Antioxidative modulation in response to metal-induced oxidative stress in
algal chloroplasts. Arch Environ Contam Toxicol 40:18-24.
Oliver DP, Gartrell JW, Tiller KG, Correll R, Cozens GD, Youngberg BL (1995)
Differential responses of Australian wheat cultivars to cadmium
concentration in wheat grain. Aust J Agric Res 46:873-886.
Olmos EO, Martinez-Solano JR, Piqueras A, Hellin E (2003) Early steps in the
oxidative burst induced by cadmium in cultured tobacco cell (BY-2 line). J Exp
Bot 54: 291-301.
Oncel I, Kele Y, Ustim AS (2000) Interactive effects of temperature and heavy metal
stress on the growth and some biochemical compounds in wheat seedlings.
Environ PoUut 107:315-320.
Organisation for Economic Cooperation and Development (OECD) (1994) Risk
reduction monograph No. 5: Cadmium OECD Environment Directorate,
Paris, France.
Ouariti O, Boussama N, Zarrouk M, Cherif A, Ghorbel MH (1997) Cadmium and
copper- induced changes in tomato membranes lipids. Phytochem 45:1343-
1350.
157

Ouzounidou G, Moustakas M, Eleftheriou EP (1997) Physiological and
ultrastructural effects of cadmium on wheat {Triticum aestivum L.) leaves.
Arch Environ Contam Toxicol 32:154-160.
Ozturk L, Eker S, Ozkutlu F, Cakmak I (2003) Effect of cadmium on growth and
concentrations of cadmium, ascorbic acid and sulfhydryl groups in durum
wheat cultivars. Turk J Agric For 27:161-168.
Padmaja K, Prasad DDK, Prasad ARK (1990) Inhibition of chlorophyll synthesis
Phaseolus vulgaris L. seedlings by cadmium acetate. Photosynthetica 24:399-
405.
Page AL, Bingham FT, Chang AC (1981) Cadmium. In: Lepp NW (Ed) Effect of
heavy metal pollution on plants. Applied Science, London, pp72-109.
Pal UR, Gossett DR, Sims JL, Leggett JE (1976) Molybdenimi and sulfur nutrition
effects on nitrate reduction in Burley tobacco. Can J Hot 54:2014-2022.
Panda SK, Khan MH (2003) Antioxidant efficiency in Oryza sativa leaves under
heavy metal toxicity. J Plant Biol 30:23-29.
Pankovic D, Plesnicar M, Arsenijevic-Maksimovic I, Petrovic N, Sakac Z, Kastori R
(2000) Effects of nitrogen nutrition on photosynthesis in Cd-treated sunflower
plants. Ann Bot 86:841-847.
Pasricha NS, Aulakh MS (1991) Twenty years of sulphur research and oilseed
production in Punjab, India. Sulphur Agric 15:17-23.
Patel MJ, Patel JN, Subramanian RB (2005) Effect of cadmium on growth and the
activity of H2O2 scavenging enzymes in Colocassia esculentum. Plant Soil
273:183-188.
Pekker I, Elisha TO, Mittler R (2002) Reactive oxygen intermediates and glutathione
regulate the expression of cytosolic ascorbate peroxidase during iron mediated
oxidative stress in bean. Plant Mol Biol 49:429-438.
Peralta JR, Gardea-Torresdey JL, Tiemann KJ, Gomez E, Arteaga S, Rascon E,
Parsons JG (2000) Study of the effects of heavy metals on seed germination and
plant growth on alfalfa plant {Medicago sativa) grown in solid media. Proceedings
of the 2000 conference on Hazardous Waste Research, Denver, Colorado, 23-25
May, ppl35-140.
Pereira GJG, Molina SMG, Lea PJ, Azevedo RA (2002) Activity of antioxidant
enzymes in response to cadmium in Crotalaria juncea. Plant Soil 239:123-
132.
158

Perfus-Barbeoch L, Leonhardt N, Vavasseur A, Forestier C (2002) Heavy metal
toxicity: cadmium permeates through calcium channels and disturbs the plant
water status. Plant J 32:548-639.
Petrovic N, Kastori R, Rajean I (1990) The effect of cadmium on nitrate
reductase activity in sugar beet (Beta vulgaris). In: van Beusichem ML (Ed)
Plant nutrition-physiology and applications. Kluwer Academic Publishers,
Dordrecht, pp 107-109.
Pietrini F, lannelli MA, Pasqualini S, Massacci A (2003) Interaction of camium
with glutathione and photosynthesis in developing leaves and chloroplasts of
Phragmites australis (Cav.) Trin. ex Steudel. Plant Physiol 133:829-837.
Pinto AP, Mota AM, de Varennes A, Pinto FC (2004) Influence of organic
matter on the uptake of cadmium, zinc, copper and iron by sorghum plants.
Sci Total Environ 326:239-247.
Piqueras A, Olmos E, Martines-Solano JR, Hellin E (1999) Cd-induced oxidative
burst in tabacco BY2 cells: time course, subcellular localisation and
antioxidative response. Free Rad Res 31:33-38.
Podlesna A, Cacak-Pietrzak G (2008) Effects of fertilization with sulfur on
quality of winter wheat: A case study of nitrogen deprivation. In: Khan NA,
Singh S, Umar S (Eds) Sulfur assimilation and abiotic stresses in plants.
Springer-Verlag, The Netherlands, pp355-365.
Polidoros NA, Scandalios JG (1999) Role of hydrogen peroxide and different
classes of antioxidants in the regulation of catalase and glutathione S-
transferase gene expression in maize {Zea mays L.). Physiol Plant 106:112-
120.
Polle A (2001) Dissecting the superoxide dismutases-ascorbate peroxidase-
glutathione pathway in chloroplasts by metabolic modeling. Computer
simulations as a step towards flux analysis. Plant Physiol 126:445-462.
Popovic M, Kevresan S, Kandrac J, Nicolic J, Petrovic N, Kastori R (1996) The role
of sulphur in detoxification of cadmium in young sugar beet plants. Biol Plant
38:281-287.
Poschenreider Ch, Gunse B, Barcelo J (1989) Influence of cadmium on water
relations, stomatal resistance and abscisic acid content in expanding bean
leaves. Plant Physiol 90:1365-1371.
159

Prakash S (1980) Cruciferous oilseeds in India. In: Tsumoda S, Hinata K, Gomes
Campo C (Eds) Brassica crops and wild allies: biology and breeding. Japan
Science Society Press, Tokyo, Japan, ppl51-163.
Prasad MNV (1995) Cadmium toxicity and tolerance to vascular plants. Environ Exp
Bot 35:525-545.
Prasad MNV (1997) Plant Ecophysiology. John Wiley and Sons, NY.
Prasad MNV, Hagemeyer J (1999) Heavy metal stress in plants from molecules
to ecosystems. Springer-Verlag, Heidelberg, pp 107-109.
Prince WSPM, Kumar PS, Doberschutz KD, Subburam V (2002) Cadmium
toxicity in mulberry plants with special reference to the nutritional quality of
leaves. J Plant Nutr 25:689-700.
Prochazkova D, Sairam RK, Srivastava GC, Singh DV (2001) Oxidative stress
and antioxidant activity as the basis of senescence in maize leaves. Plant Sci
161:765-771.
Prosser IM, Purves JV, Saker LR, Clarkson DT (2001) Rapid disruption of
nitrogen metabolism and nitrate transport in spinach plants deprived of
sulphate. J Exp Bot 52:113-121.
Purakayastha TJ, Nad BK (1997) Effect of sulphur, magnesium and molybdenum
on mustard {Brassica juncea L.) and wheat {Triticum aestivum L.): Nitrate
nitrogen accumulation and protein yield. Ind J Plant Physiol 2:110-113.
Qadir S, Qureshi MI, Javed S, Abdin MZ (2004) Genotypic variation in
phytoremediation potential of Brassica juncea cultivars exposed to Cd stress.
Plant Sci 167:1171-1181.
Radotic K, Ducic T, Mutavdzic D (2000) Changes in perxidase activity and
isoenzymes in spruce needles after exposure to different concentrations of
cadmium. Environ Exp Bot 44:105-113.
Rai UN, Gupta M, Tripathi RD, Chanda P (1998) Cadmium regulated nitrate
reductase activity in Hydrilla verticillatta (l.f.) Royale. Water Air Soil Pollut
106:171-177.
Rai V, Khatoon S, Bisht SS, Mehrotra S (2005) Effect of cadmium on growth,
ultramorphology of leaf and secondary metabolites of Phyllanthus amarus
Schum. and Thonn. Chemosphere 61:1644-1650.
160

Ramos I, Esteban E, Lucena JJ, Garate A (2002) Cadmium uptake and
subcellular distribution in plants of Lactuca sp. Cd-Mn interaction. Plant Sci
162:761-767.
Randle WM (2000) Increasing nitrogen concentration in hydroponic solutions
affects onion flavor and bulb quality. J Amer Soc Horti Sci 125:254-259.
Randle WM, Bussard ML, Warnock DF (1993) Ontogeny and sulphur fertility
affect leaf sulphur in short-day onions. J Amer Soc Horti Sci 118:762-765.
Randle WM, Lancaster JE (2002) Sulphur compounds in Alliums in relation to
flavour quality. In: Rabinowitch HD, Currah L (Eds) Allium crop science:
recent advances. CAB International, Wallingford, UK, pp329-356.
Randle WM, Lancaster JE, Shaw ML, Sutton KH, Hay RL, Bussard ML (1995)
Quantifying onion flavor compounds responding to sulphur fertility. Sulphur
increases levels of alk(en)ylcysteine sulphoxides and biosynthetic
intermediates. J Amer Soc Horti Sci 120:1075-1081.
Ranieri A, Castagna A, Scebba F, Careri M, Zagnoni I, Predieri G, Pagliari M,
Sanita di Toppi L (2005) Oxidative stress and phytochelatin characterisation
in bread wheat exposed to cadmium excess. Plant Physiol Biochem 43:45-54.
Rao MV (1992) Cellular detoxification mechanisms to determine age dependent
injury in tropical plant exposed to SO2. J Plant Physiol 140:733-740.
Raskin I, Ensley BD (2000) Phytoremediation of toxic metals: Using plants to
clean up the envirormient. John Wiley &. Sons, New York, pp304.
Ratti S, Giordano M (2008) Allocation of S to sulfonium compound in
microalgae. In: Khan NA, Singh S, Umar S (Eds) Sulfur assimilation and
abiotic stresses in plants. Springer-Verlag, The Netherlands, pp317-333.
Rausch T, Wachter A (2005) Sulfur metabolism: a versatile platform for
launching defence operations. Trends Plant Sci 10:503-509.
Rauser WE (1987) Compartmental efflux analysis and removal of extracellular
cadmium from roots. Plant Physiol 85:62-65.
Rauser WE (1990) Phytochelatins. Annu Rev Biochem 59:61-86.
Rauser WE (2000) Roots of maize seedlings retain most of their cadmium through
two complexes. J Plant Physiol 156: 545-551.
Raven J A (1995) Photosynthetic and non-photosynthetic roles of carbonic anhydrase
in algae and cyanobacteria. Phycol 34:93-101.
161

Razinger J, Dermastia M, Koce JD, Zrimec A (2007) Oxidative stress in
duckweed (Lemna minor L.) caused by short-term cadmium exposure.
Environ Pollut doi:10.1016/j.envpol.2007.08.018
Reddy AR, Raghavendra AS (2006) Photooxidative stress. In: Madhava Rao KV,
Raghavendra AS, Reddy KJ (Eds) Physiology and molecular biology of stress
tolerance in plants. Springer-Verlag, The Netherlands, ppl57-186.
Rennenberg H, Brunold C (1994) Significance of glutathione metabolism in
plants under stress. Prog Bot 55:142-156.
Rennenberg H, Schmitz K, Bergmann L (1979) Long distance transport of sulfur
in Nicotiana tabacum. Planta 147:57-62.
Repka J, Saric M, Marek M, Zima M (1971) Effect of macroelement deficiency
on structure of chloroplasts and productivity of photosynthesis in maize
plants. Soviet Plant Physiol 18:938-943.
Resurreccion AP, Makino A, Bennett J, Mae T (2001) Effect of sulfur nutrition
on the growth and photosynthesis of rice. Soil Sci Plant Nutr 47:611-620.
Reuveny Z, Dougall DK, Trinity PM (1980) Regulatory coupling of nitrate and
sulphate assimilation pathways in cultured tobacco cells. Proc Natl Acad Sci
USA 77:6670-6672.
Rizhsky L, Davletova S, Liang H, Mittler R (2004) The zinc finger protein Zatl2
is required for cytosolic ascorbate peroxidase 1 expression during oxidative
stress in Arabidopsis. J Biol Chem 279:11736-11743.
Romero-Puertas MC, Corpas FJ, Rodriguez-Serrano M, Gomez M, del Rio LA,
Sandalio LM (2007) Differential expression and regulation of antioxidative
enzymes by cadmium in pea plants. J Plant Physiol 164:1346-1357.
Romero-Puertas MC, Corpas FJ, Sandalio LM, Leterrier M, Rodriguez-Serrano
M, del Rio LA, Palma JM (2006) Glutathione reductase from pea leaves:
response to abiotic stress and characterization of the peroxisomal isozyme.
NewPhytol 170:43-52.
Romero-Puertas MC, McCarthy I, Sandalio LM, Palma JM, Corpas FJ, Gomez
M, del Rio LA (1999) Cadmium toxicity and oxidative metabolism of pea
leaf peroxisomes. Free Rad Res 31(Suppl):25-32.
Romero-Puertas MC, Rodriguez-Serrano M, Corpas FJ, Gomez M, del Rio LA,
Sandalio LM (2004) Cadmium-induced subcellular accumulation of O2' and
H2O2 in pea leaves. Plant Cell Environ 27:1122-1134.
162

Romero-Puertas MC, Zabalza A, Rodriguez-Serrano M, Gomez M, del Rio LA,
Sandalio LM (2003) Antioxidative response to cadmium in pea roots. Free Rad
Res 37:44-48.
Root RA, Miller RJ, Koeppe DE (1975) Uptake of cadmium its toxicity, and effect on
the iron ratio in hydroponically grown corn. J Environ Qual 4:473-476.
Rosa EVC, Valgas C, Souza-Sierra MM, Correa AXR, Redetski CM (2003) Biomass
growth, micronucleus induction, and antioxidant stress enzyme responses in Vicia
faba exposed to cadmium in solution. Environ Toxic Chem 22:645-649.
Rubio M, Escrig I, Martinez-Cortina C, Lopez-Benet F, Sanz A (1994) Cadmium
and nickel accumulation in rice plants: Effects on mineral nutrition and
possible interactions of abscisic and gibberellic acids. Plant Growth Regul
14:151-157.
Ruegsegger A, Schmutz D, Brunold C (1990) Regulation of glutathione synthesis
by cadmium in Pisum sativum L. Plant Physiol 93:1579-1584.
Sachdev MS, Deb DL (1990) Nitrogen and S uptake and efficiency in the
mustard-moong-maize cropping systems. FertNews 35:49-55.
Sagi M, Fluhr R (2001) Superoxide production by plant homologues of the gp91phox
NADPH oxidase: Modulation of activity by calcium and tobacco mosaic virus
infection. Plant Physiol 126:1281-1290.
Sairam RK, Srivastava GC, Agarwal S, Meena RC (2005) Differences in antioxidant
activity in response to salinity stress in tolerant and susceptible wheat genotypes.
Biol Plant 49:85-91.
Sairam RK, Till AR, Blair GJ (1995) Effect of sulphur and molybdenum on growth,
nitrate assimilation and nutrient content of Phalaris. J Plant Nutr 18:2093-2103.
Saito K (2000) Regulation of sulfate transport and synthesis of sulfur-containing
amino acids. Curr Opin Plant Biol 3:188-195.
Saito K (2004) Sulfur assimilatory metabolism. The long and smelling road.
Plant Physiol 136:2443-2450.
Salin ML (1988) Toxic oxygen species and protective systems of the
chloroplasts. Physiol Plant 72:681-689.
Salt DE, Blaylock M, Kumar NPBA, Dushenkov V, Ensley BD, Chet I, Raskin I
(1995) Phytoremediation: a novel strategy for the removal of toxic metals
from the environment using plants. Biotechnol 13:468-474.
163

Salt DE, Rauser WE (1995) Mg-ATP-dependent transport of phytochelatins
across the tonoplast of oat roots. PlantPhysiol 107:1293-1301.
Salt DE, Wagner GJ (1993) Cadmium transport across tonoplast of vesicles from
oat roots: evidence for a Cd^VH^antiport activity. J Biol Chem 268:12297-
12302.
Samiullah, Khan NA, Nazar R, Ahmad I (2007) Physiological basis for reduced
photosynthesis and growth of cadmium-treated wheat cultivars differing in
yield potential. J Food Agric Environ 5:375-377.
Sandalio LM, Dalurzo HC, Gomez M, Romero-Puertas MC, del Rio LA (2001)
Cadmium-induced changes in the growth and oxidative metabolism of pea
plant. J Exp Bot 52:2115-2126.
Sandalio LM, del Rio LA (1988) Intra-organellar distribution of superoxide
dismutase in plant peroxisomes (glyoxysomes and leaf peroxisomes). Plant
Physiol 88:1215-18.
Sandra P (1986) Altering the earths chemistry assessing and risk. World Watch Paper.
Wold Watch Institute.
Sanita di Toppi L, Gabbrielli R (1999) Response to cadmium in higher plants.
Environ Exp Bot 41:105-130.
Sanita di Toppi L, Lambard M, Paazzali L, Cappugi G, Durat M, Gabbrielli R
(1998) Response to cadmium in carrot in vitro plant and cell suspension
cultures. Plant Sci 137:119-129.
Sarry JE, Kuhn L, Ducruix C, Lafaye A, Junot C, Hugouvieux V, Jourdain A,
Bastien O, Fievet J, Vaihen D, Amekraz B, Moulin C, Ezan E, Garin J,
Bourguignon J (2006) The early responses of Arabidopsis thaliana cells to
cadmium exposure explored by protein and metabolite profiling analyses.
Proteomics 7:2180-2198.
Sawhney V, Sheoran IS, Singh R (1990) Nitrogen fixation, photosynthesis and
enzymes of ammonia assimilation and ureide biosynthesis in nodules of
mungbean {Vigna radiata) grown in presence of cadmium. Ind J Exp Biol
28:883-886.
Scandalios JG (1990) Response of plant antioxidant defense genes to
environmental stress. Adv Genet 28:1-41.
Scandalios JG (1993) Oxygen stress and superoxide dismutases. Plant Physiol
101:7-12.
164

Scandalios JG, Acevedo A, Ruzsa S (2000) Catalase gene expression in response
to chronic high temperature stress in maize. Plant Sci 156:103-110.
Scebba F, Arduini I, Ercoli L, Sebastiani L (2006) Cadmium effects on growth
and antioxidant enzymes activities in Miscanthus sinensis. Biol Plant 50:688-
692.
Schafer HJ, Haag-Kerwer A, Rausch T (1998) cDNA cloning and expression
analysis of genes encoding GSH synthesis in roots of the heavy metal
accumulator Brassica juncea L.: evidence or Cd-induction of a putative
mitochondrial y-glutamylcysteine synthetase isoform. Plant Mol Biol 37:87-
97.
Scherer HW (2008) Impact of sulfur on N2 fixation of legumes. In: Khan NA,
Singh S, Umar S (Eds) Sulfur assimilation and abiotic stresses in plants.
Springer-Verlag, The Netherlands, pp43-54.
Schickler H, Caspi H (1999) Response of antioxidative enzymes to nickel and
cadmium stress in hyperaccumulator plants of the genus Alyssum. Physiol
Plant 105:39-44.
Schlegel H, Godbold DL, Huttermann A (1987) Whole plant aspect of heavy
metal induced changes in CO2 uptake and water relation of spruce {Picea
abies) seedlings. Physiol Plant 69:265-270.
Schneider S, Bergmann L (1995) Regulation of glutathione synthesis in
suspension cultures of parsley and tobacco. Botanical Acta 108:34-40.
Schutzendubel A, Nikolova P, Rudolf C, PoUe A (2002) Cadmium and H2O2
induced oxidative stress in Populas x canescens roots. Plant Physiol Biochem
40:577-584.
Schutzendubel A, Polle A (2002) Cadmium and H202-induced oxidative stress in
Populus X canescens roots. Plant Physiol Biochem 40:577-584.
Schutzendubel A, Schwanz P, Teichmann T, Gross K, Langenfeld-Heyser R,
Godbold DL, Polle A (2001) Cadmium-induced changes in antioxidative
systems, H2O2 content and differentiation in pine (Pinus sylvestris) roots.
Plant Physiol 127:887-898.
Seemann R, Sharkey TD, Wang J, Osmond CB (1987) Environmental effects on
photosynthesis, nitrogen use efficiency and metabolite pools in leaves of sun
and shade plants. Plant Physiol 84:796-802.
165

Seregin IV, Shpigun LK, Ivaniov VB (2004) Distribution and toxic effects of
cadmium and lead on maize roots. Russ J Plant Physiol 51:525-533.
Sexton PJ, Batchelor WD, Shibles R (1997) Sulphur availability, rubisco content
and photosynthetic rate of soybean. Crop Sci 37:1801-1806.
Seyyedi M, Timko MP, Sundqvist C (1999) Protochlorophyllide, NADPH-
protochlorophyllide oxidoreductase and chlorophyll formation in the lipJ
mutant of pea. Physiol Plant 106:344-354.
Shah K, Dubey RS (1997) Cadmium alters phosphate level and suppresses
activity of phosphorolytic enzymes in germinating rice seeds. J Agron Crop
Sci 179:35-45.
Shah K, Dubey RS (1998) A 18 kDa Cd inducible protein complex: its isolation
and characterization from rice (Oryza sativa L.) seedlings. J Plant Physiol
152:448-454.
Shah K, Kumar RG, Verma S, Dubey RS (2001) Effect of cadmium on lipid
peroxidation, superoxide anion generation and activities of antioxidant
enzymes in growing rice seedlings. Plant Sci 161:1135-1144.
Sharma P, Dubey RS (2006) Cadmium uptake and its toxicity in higher plants.
In: Khan NA, Samiullah (Eds) Cadmium toxicity and tolerance in plants.
Narosa Publishing House, New Delhi, pp64-86.
Sharma RK, Agrawal M (2006) Single and combined effects of cadmium and
zinc on carrots: Uptake and bioaccumulation. J Plant Nutr 29:1791-1804.
Sharma SS, Dietz KJ (2006) The significance of amino acids and amino acid-
derived molecules in plant responses and adaptation to heavy metal stress. J
Exp Hot 57:711-726.
Shaw BP (1995a) Changes in the levels of photosynthetic pigments in Phaseolus
aureus Roxb. exposed to Hg and Cd at two stages of development: A
comparative study. Bull Environ Contam Toxicol 55:574-580.
Shaw BP (1995b) Effects of mercury and cadmium on the activities of
antioxidative enzymes in the seedlings of Phaseolus aureus. Biol Plant
37:587-596.
Sheoran IS, Agarwal N, Singh R (1990a) Effect of cadmium and nickel on in
vivo carbon dioxide exchange rate of pigeon pea (Cajanus cajan L.), Plant
Soil 129:243-249.
166

Sheoran IS, Singal HR, Singh R (1990b) Effect of cadmium and nickel on
photosynthesis and the enzymes of photosynthetic carbon reduction cycle in
pigeon pea (Cajanus cajan). Photosynth Res 23:345-351.
Shulaev V, Oliver DJ (2006) Metabolic and proteomic markers for oxidative
stress: new tools for reactive oxygen species research. Plant Physiol 141:367-
372.
Siedlecka A (1995) Some aspects of interactions between heavy metals and plant
mineral nutrients. Acta Soc Bot Pol 64:265-272.
Siedlecka A, Krupa Z (1996) Interaction between cadmium and iron and its
effects on photo-synthetic capacity of primary leaves of Phaseolus vulgaris.
Plant Physiol Biochem 34:833-841.
Siedlecka A, Krupa Z (1999) Cd/Fe interaction in higher plants - its consequences for
the photosynthetic apparatus. Photosynthetica 36: 321-331.
Siedlecka A, Krupa Z, Samuelsson G, Oquist G, Gardestrom P (1997) Primary
carbon metabolism in Phaseolus vulgaris plants under Cd/Fe interaction.
Plant Physiol Biochem 35:951-957.
Sieferman-Harms D (1987) The light harvesting function of carotenoids in
photosynthetic membrane. Plant Physiol 69:561-568.
Singh KS, Bairathi RC (1980) A study on the sulphur fertilization of mustard in
the semi-arid tract of Rajasthan. Aim Arid Zone 19:197-202.
Singh MV (2001) Importance of sulphur in balanced fertilizer use in India. Pert
News 46:13-35.
Singh PK, Tewari RK (2003) Cadmium toxicity induced changes in plant water
relations and oxidative metabolism of Brassica juncea L. plants. J Environ
Biol 24:107-112.
Singh RP, Bharti N, Kumar G (1994) Differential toxicity of heavy metals to
growth and nitrate reductase activity of Sesamum indicum seedlings.
Phytochem 35:1153-1156.
Singh S, Anjum NA, Khan NA, Nazar R (2008b) Metal-binding peptides and
antioxidant defence system in plants: significance in cadmium tolerance. In: Khan
NA, Singh S (Eds) Abiotic stress and plant responses. IK International, New
Delhi, pp 159-189.
167

Singh S, Anjum NA, Nazar R, Khan NA (2007) Influence of cadmium on growth
nitrogen and sulfur assimilation in garden cress {Lepidium sativum L.). XXXth
Botanical Conference, Gwalior, MP, 28-30 November, ppl61.
Singh S, Eapen S, D'Souza SF (2006) Cadmium accumulation and its influence on
lipid peroxidation and antioxidative system in an aquatic plant, Bacopa monnieri
L. Chemosphere 62:233-246.
Singh S, Khan NA, Nazar R, Anjum NA (2008a) Photosynthetic traits and activities
of antioxidant enzymes in blackgram (Vigna mungo L. Hepper) under cadmium
stress. Am J Plant Physiol 3:25-32.
Singh Y P (2004) Role of sulphur and phosphorus in black gram production. Pert
News 49:33-36.
Sirko A, Blaszczyk A, Liszewska F (2004) Overproduction of SAT and/or OASTL in
transgenic plants: a survey of effects. J Exp Hot 55:1881-1888.
Skadsen RW, Schulz-Lefert P, Herbt JM (1995) Molecular cloning,
characterization and expression analysis of two classes of catalase isozyme
genes in barley. Plant Mol Biol 29:1005-1014.
Skorzynska-Polit E, Baszynski T (1995) Photochemical activity of primary
leaves in cadmium stressed Phaseolus coccineus depends on their growth
stages. Acta Soc Bot Pol 64:273-279.
Skorzynska-Polit E, Baszynski T (1997) Differences in sensitivity of the
photosynthetic apparatus in Cd-stressed runner bean plants in relation to their
age. Plant Sci 128:11-21.
Skorzynska-Polit E, Drazkiewicz M, Krupa Z (2003/04) The activity of the
antioxidative system in cadmium-treated Arabidopsis thaliana. Biol Plant
47:71-78.
Slooten L, Capiau K, Van Camp W, Van Montagu M, Sybesma C, Inze D (1995)
Factors affecting the enhancement of oxidativ stress tolerance in transgenic
tobacco overexpressing mangnese overexpressing superoxide dismutase in
the chloroplasts. Plant Physiol 107:737-750.
Smeets K, Cuypers A, Lambrechts A, Semane B, Hoet P, Laere A, Vangronsveid
J (2005) Induction of oxidative stress and antioxidative mechanisms in
Phaseolus vulgaris after Cd application. Plant Physiol Biochem A^-All-AAA.
Smirnoff N, Conklin PL, Loewus FA (2001) Biosynthesis of ascorbic acid in
plants: a renaissance. Annu Rev Plant Physiol Plant Mol Biol 52:437-467.
168

Smith IK (1975) Sulfate transport in cultured tobacco cells. Plant Physiol
55:303-307.
Solomonson L, Barber MJ (1990) Assimilatory nitrate reductase: functional
properties and regulation. Annu Rev Plant Physiol Plant Mol Biol 41:225-
253.
Somashekaraiah BV, Padmaja K, Prasad ARK (1992) Phytotoxicity of cadmium
ions on germinating seedlings of mung bean (Phaseolus vulgaris):
involvement of lipid peroxides in chlorophyll degradation. Physiol Plant.
85:85-89.
Srivastava HS (1992) Multiple functions and forms of higher plant nitrate
reductase. Phytochem 31:2941-2947.
Srivastava S, Tripathi RD, Dwivedi UN (2004) Synthesis of phytochelatins and
modulation of antioxidants in responses to cadmium stress in Cuscuta reflexa
- an angiospermic parasite. J Plant Physiol 161:665-674.
Stiborova M, Doubravova M, Leblova S (1986) A comparative study of the
effect of heavy metal ions on ribulose-l,5-bisphosphate carboxylase and
phosphoenolpyruvate carboxylase. Biochemie und Physiologic der Pflanzen
181:373-379.
Stobart AK, Griffiths WT, Ameen-Bukhari I, Sherwood RP (1985) The effect of
Cd " on biosynthesis of chlorophyll in leaves of barley. Physiol Plant 63:293-
298.
Stochs SJ, Bagchi D (1995) Oxidative mechanism in the toxicity of metal ions.
Free Rad Biol Med 18:321-336.
Strohm M, Jouanin L, Kunert KJ, Pruvost C, Polle A, Foyer CH, Rennenberg H
(1995) Regulation of glutathione synthesis in leaves of transgenic poplar
(Populus tremula x P. alba) overexpressing glutathione synthetase. The Plant
J 7:141-145.
Stroinski A, Kozlowska M (1997) Cadmium induced oxidative-stress in potato
tuber. Acta Soc Bot Pol 66:189-195.
Stroinski A, Zielezinska M (1997) Cadmium effect on hydrogen peroxide, glutathione
and phytochelatin levels in potato tuber. Acta Physiol Plant 19:127-135.
Su DC, Wong JWC (2004) Selection of oilseed rapes (Brassica juncea) from
China for phytoremediation of cadmium contaminated soil. Bull Environ
Contam Toxicol 72:991-998.
169

Sugiharto B, Miyata K, Nakamoto H, Sasakawa H, Sugiyama T (1990)
Regulation of expression of carbon-assimilating enzymes by nitrogen in
maize leaf. Plant Physiol 92:963-969.
Sun Q, Yec ZH, Wang XR, Wong MH (2007) Cadmium hyperaccumulation
leads to an increase of glutathione rather than phytochelatins in the cadmium
hyperaccumulator 5e£/M/w alfredii. J Plant Physiol 164:1489-1498.
Suter M, Von Ballmoos P, Kupriva S, Opdencamp R, Schaller J, Kuhlemeier C,
Scurmann P, Brunold C (2000) Adenosine 5-sulphosulphate sulpho
transferease and adenosine 5-phospho sulpho reductase are identical
enzymes. J Biol Chem 275:930-936.
Syntichaki KM, Loulakakis KA, Loulakakis-Roubelakis KA (1996) The amino-
acid sequence similarity of plant glutamate dehydrogenase to the
extremophilic archaeal enzyme conforms to its stress-related function. Gene
68:87-92.
Takahashi H, Saito K (1996) Subcellular localization of spinach cysteine
synthase isoforms and regulation of their gene expression by nitrogen and
sulfur. Plant Physiol 112:273-280.
Takahashi H, Watanabe-Takahashi A, Smith F, Blake-Kalff M, Hawkesford M,
Daito K (2000) The role of three functional sulfate transporters involved in
uptake and translocation of sulphate in Arabidopsis thaliana. Plant J 23:171-
182.
Tandon HLS (1991) Sulphur research and agricultural production in India, 3 ''
Ed. The Sulphur Institute, Washington, DC.
Tandon HLS (1995) Sulphur fertilisers for Indian agriculture, 2^^ Ed. Fertiliser
Development and Consultation Organisation, New Delhi.
Tausz M, Gullner G, Komives T, Grill D (2003) The role of thiols in plant
adaptation to environmental stress. In: Abrol YP, Ahmad A (Eds) Sulphur in
plants. Kluwer Academic Publishers, The Netherlands, pp221-244.
Taylor AJ, Smith CJ, Wilson IB (1991) Effect of irrigation and nitrogen fertilizer
on yield, oil content, nitrogen accumulation and water use of canola
{Brassica napus L.). Pert Res 29:249-260.
Terry N (1976) Effects of sulphur on the photosynthesis of intact leaves and
isolated chloroplasts of sugar beets. Plant Physiol 57:477-479.
170

Thiele A, Schirwitz K, Winter K, Krause GH (1996) Increased xanthophyll cycle
activity and reduced Dl protein inactivation related to photoinhibition in two
plant systems acclimated to excess light. Plant Sci 115:237-250.
Tomaszewska B (2002) Glutathione and thiol metabolism in metal exposed
plants. In: Prasad MNV, Strzalka K (Eds) Physiology and biochemistry of
metal toxicity and tolerance in plants. Kluwer Academic Publisher,
Dordrecht, Netherlands, pp36-58.
Tukiendorf A, Baszynski T (1991) The in vivo effect of cadmium on
photochemical activities in chloroplasts of runner bean plants. Acta Physiol
Plant 13:51-57.
Tukiendorf A, Rauser WE (1990) Changes in glutathione and phytochelatins in
roots of maize seedlings exposed to cadmium. Plant Sci 70:155-166.
U N (1935) Genome analysis in Brassica with special reference to the experimental
formation of Brassica napus and peculiar mode of fertilization. Japan J Bot 7:389-
452.
Van Assche F, Ciarletta P (1993) Envirorunental exposure to cadmium in
Belgium: Decreasing trends during the 1980s. Heavy metals in the
environment 1:34-37.
Van Assche F, Clijsters H (1990) Effects of metals on enzyme activity in plants.
Plant Cell Environ 13:195-206.
Vanaja M, Charyulu NVN, Rao KVN (2000) Effect of some organic acids and
calcium on growth toxic effect and accumulation of cadmium in
Stigeoclonium tenue Kutz. Ind J Plant Physiol 7:163-167.
Vassilev A, Malgozata B, Nevena S, Zlatko Z (2005) Chronic Cd toxicity of bean
plants can be partially reduced by supply of ammonium sulphate. J Central Euro
Agric 6:389-396.
Vassilev A, Schwitzguebel J-P, Thewys T, van der Lelie D, Vangronsveld J (2004)
The use of plants for remediation of metal contaminated soils. The Scientific
World J 4:9-34.
Vassilev A, Tsonev T, Yordanov 1 (1998) Physiological response of barley plants
{Hordeum vulgare) to cadmium contamination in soil during ontogenesis. Environ
Pollut 103:287-293.
Vassilev A, Yordanov I (1997) Reductive analysis of factors limiting growth of Cd-
exposed plants: a review. Bulg J Plant Physiol 23:114-133.
171

Vassilev A, Yordanov I, Chakalova E, Kerin V (1995) Effect of cadmium stress
on growth and photosynthesis of young barley (//. vulgare L.) plants. 2.
Structural and functional changes in photosynthetic apparatus. Bulg J Plant
Physiol 21:12-21.
Vassilev A, Yordanov I, Vangronsveld J (2006) Cadmium phytoextraction from
contaminate soils. In: Khan NA, Samiullah (Eds) Cadmium toxicity and
tolerance in plants. Narosa Publishing House, New Delhi, ppl03-l 14.
Vecchia FC, Rocca NL, Moro I, Faveri S, Andreoli C, Rascio N (2005)
Morphogenetic, ultrastructural and physiological damages suffered by
submerged leaves of Elodea canadensis exposed to cadmium. Plant Sci
168:329-338.
Verma S, Dubey RS (2001) Effect of cadmium on soluble sugars and enzymes of
their metabolism in rice. Biol Plant 44:117-123.
Verma S, Dubey RS (2002) Influence of lead toxicity on photosynthetic pigments,
lipid peroxidation and activities of antioxidant enzymes in rice plants. Ind J Agri
Biochem 15:17-22.
Verma VK, Singh YP, Rai JPN (2007) Biogas production from plant biomass used for
phytoremediation of industrial waste. Biores Technol 98:1664-1669.
Vidmar JJ, Schjoerring JK, Touraine B, Glass ADM (1999) Regulation of the
hvstl gene encoding a high-affinity sulfate transporter from Hordeum
vulgare. Plant Mol Biol 40:883-892.
Vitoria AP, Lea PJ, Azevedo RA (2001) Antioxidant enzyme responses to
cadmium in radish tissue. Phytochem 57:701-710.
Vogeli-Lange R, Wagner GJ (1990) Subcellular localization of cadmium and
cadmium-binding peptides in tobacco leaves-implication of a transport
function for cadmium-binding peptide. Plant Physiol 92:1086-1093.
Wagner GJ (1993) Accumulation of cadmium in crop plants and its consequences to
human health. Adv Agron 51:173-212.
Wahid A, Ghani A (2007) Varietal differences in mungbean (Vigna radiata) for
growth, yield, toxicity symptoms and cadmium accumulation. Ann applied Biol
152:59-69.
Wahid A, Ghani A, Ali I, Ashraf MY (2007) Effects of cadmium on carbon and
nitrogen assimilation in shoots of mungbean [Vigna radiata (L.) Wilczek]
seedlings. J Agron Crop Sci 193:357-365.
172

Wang J, Evangelou VP (1995) Metal tolerance aspects of plant cell wall and
vacuole. In: Perssarkali M (Ed) Handbook of plant and crop physiology.
Marcel Dekker, New York, pp697-717.
Wang L, Zhou Q, Ding L, Sun Y (2008) Effect of cadmium toxicity on nitrogen
metabolism in leaves of Solarium nigrum L. as a newly found cadmium
hyperaccumulator. J Hazardous Materials doi: 10.10l6/j.jhazmat.2007.10.097
Weast RC (1984) CRC Handbook of chemistry and physics, 64 ^ Ed. CRC Press,
Boca Raton, FL, USA.
Weckx JEJ, Clijsters HMM (1996) Oxidative damage and defense mechanisms in
primary leaves of Phaseolus vulgaris as a result of root assimilation of toxic
amounts of copper. Physiol Plant 96:506-512.
Weigel HJ (1985) Inhibition of photosynthetic reactions of isolated intact
chloroplasts by cadmium. J Plant Physiol 119:179-189.
Weigel HJ, Jager HJ (1980) Subcellular distribution and chemical form of
cadmium in bean plants. Plant Physiol 65:480-482.
Whatley JM (1971) Ultrastructural changes in chloroplasts of P. vulgaris during
development under conditions of nutrient deficiency. New Phytol 70:725-
742.
Willekens H, Langebartels C, Tire C, Van Montagu M, Inze D, Van Camp W
(1994) Differential expression of catalase genes in Nicotiana plumbaginifolia
(L.). Proc Natl Acad Sci USA 91:10450-10454.
Wilson LG, Bandurski RS (1958) Enzymatic reactions involving sulfate, sulfite,
selenate and molybdate. J Biol Chem 233:975-981.
Wojcik M, Tukiendorf A (1999) Cd-tolerance of maize, rye and wheat seedlings. Acta
Physiol Plant 21:99-107.
Wojcik M, Tukiendorf A (2005) Cadmium uptake, localization and detoxification in
Zea mays. Biol Plant 49:237-245.
Wojcik M, Vangronsveld J, Tukiendorf A (2005) Cadmium tolerance in Thlaspi
caerulescens I. Growth parameters, metal accumulation and phytochelatin
synthesis in response to cadmium. Environ Exp Bot 53:151-161.
World Health Organization (WHO) (1992) Environmental health criteria 134
Cadmium International Programme on Chemical Safety (IPCS) Monograph.
173

Wright GC, Smith CJ, Woodroffe MR (1988) The effect of irrigation and
nitrogen fertilizer on rapeseed (Brassica napus): Production in South-Eastern
Australia I. Growth and seed yield. Irrig Sci 9:1-13.
Wrischer M, Ljubesic N, Salopek B (1998) The role of carotenoids in the
structural and functional stability of thylkoids in plastids of dark-grown
spruce seedlings. J Plant Physiol 153:46-52.
Wu FB, Dong J, Jia GX, Zheng SJ, Zhang GP (2006) Genotypic differences in
the responses of seedling growth and Cd toxicity in rice (Oryza sativa L.).
Agric Sci China 5:68-76.
Wu FB, Wu HG, Zhang GP, Bachir MLD (2004) Differences in growth and yield
in response to cadmium toxicity in cotton genotypes. J Plant Nutr Soil Sci
167:85-90.
Wu FB, Zhang G-P (2002a) Alleviation of cadmium-toxicity by application of zinc
and ascorbic acid in barley. J Plant Nutr 25:2745-2761.
Wu FB, Zhang G-P (2002b) Genotypic differences in effect of Cd on growth and
mineral concentration in barley seedlings. Bull Environ Contam Toxicol
69:219-227.
Wu FB, Zhang GP, Dominy P (2003) Four barley genotypes respond differently
to cadmium: lipid peroxidation and activities of antioxidant capacity. Environ
Exp Bot 50:67-77.
Xiang C, Oliver DJ (1998) Glutathione metabolic genes co-ordinately respond to
heavy metals and jasmonic acid in Arabidopsis. The Plant Cell 10:1539-1550.
Xiang C, Werner BL, Christensen EM, Oliver DJ (2001) The biological
functions of glutathione revisited in Arabidopsis transgenic plants with
altered glutathione levels. Plant Physiol 126:564-574.
Xie RK, Seip HM, Wibetoe G, Nori S, McLeod CM (2006) Heavy coal
combustion as the dominant source of particulate pollution in Taiyuan,
China, corroborated by high concentrations of arsenic and selenium in PMIQ.
Sci Total Environ 370:409-415.
Xin Bin D, Rongxian Z, Wei L, Xiaming X, Schuqing C (2001) Effects of
carbonic anhydrase in wheat leaf on photosynthetic function under low CO2
concentration. Sci Agric Sinica 34:97-100.
Xu JL, Yang JR (1995) Heavy metals in land ecosystem. China Environ Sci Press,
Beijing 1:24-36.
174

Yabuta Y, Motoki T, Yoshimura K, Takeda T, Ishikawa T, Shigeoka S (2002)
Thylakoid membrane-bound ascorbate peroxidase is a limiting factor of
antioxidative systems under photo-oxidative stress. The Plant J 32:915-925.
Yang Y-J, Cheng L-M, Liu Z-H (2007) Rapid effect of cadmium on lignin
biosynthesis in soybean roots. Plant Sci 172:632-639.
Youssefian S, Nakamura M, Orudgev E, Kondo N (2001) Increased cysteine
biosynthesis capacity of transgenic tobacco overexpressing an O-
acetylserine(thiol) lyase modifies plant responses to oxidative stress. Plant Physiol
126:1001-1011.
Yu XZ, Cheng JM, Wong MH (2005) Earthworm-mycorrhiza interaction on Cd
uptake and growth of ryegrass. Soil Biol Biochem 37:195-201.
Zawoznik MS, Groppa MD, Tomaro ML, Benavides MP (2007) Endogenous salicylic
acid potentiates cadmium-induced oxidative stress in Arabidopsis thaliana. Plant
Sci 173:190-197.
Zenk MH (1996) Heavy metal detoxification in higher plants- A review. Gene
179:21-30.
Zhang FQ, Shi WY, Jin ZX, Shen ZG (2003) Response of antioxidative enzymes
in cucumber chloroplast to cadmium toxicity. J Plant Nutr 26:1779-1788.
Zhang G, Wu F, Wei K, Ding Q, Dai F, Chen F, Yang J (2006) Cadmium stress
in higher plants. In: Khan NA, SamiuUah (Eds) Cadmium toxicity and
tolerance in plants. Narosa Publishing House, New Delhi, pp87-101.
Zhang G-P, Motohiro F, Hitoshi S (2002) Influence of cadmium on mineral
concentrations and yield components in wheat genotypes differing in Cd
tolerance at seedling stage. Field Crops Res 77:93-99.
Zhang J, Kirkham MB (1996) Enzymatic responses of ascorbate-glutathione
cycle to drought stress in sorghum and sunflower plants. Plant Sci 113:139-
147.
Zhang Z, Gao X, Qui B (2008) Detection of phytochelatins in the
hyperaccumulator Sedum alfredii exposed to cadmium and lead. Phytochem
69:911-918.
Zhou W, Qiu B (2005) Effects of cadmium hyperaccumulation on physiological
characteristics of Sedum alfredii Hance (Crassulaceae). Plant Sci 169:737-
745.
175

Zhu YL, Pilon-Smits EAH, Jouanin L, Terry N (1999a) Overexpression of
glutathione synthetase in Indian mustard enhances cadmium accumulation
and tolerance. Plant Physiol 119:3-79.
Zhu YL, Pilon-Smits EAH, Jouanin L, Terry N (1999b) Cadmium tolerance and
accumulation in Indian mustard is enhanced by overexpressing y
glutamylcysteine synthetase. Plant Physiol 121:1169-1177.
176




















