
Virus morphogenesis of Helicoverpa armigera nucleopolyhedrovirus in Helicoverpa zea serum-free...
13
Downloaded from www.microbiologyresearch.org by IP: 54.90.36.180 On: Sat, 27 Aug 2016 08:10:00 Journal of General Virology (2000), 81, 2531–2543. Printed in Great Britain ................................................................................................................................................................................................................................................................................... Virus morphogenesis of Helicoverpa armigera nucleopolyhedrovirus in Helicoverpa zea serum-free suspension culture Linda H. L. Lua and Steven Reid Department of Chemical Engineering, The University of Queensland, Queensland 4072, Australia Helicoverpa armigera single nucleopolyhedrovirus (HaSNPV) replication in Helicoverpa zea serum- free suspension culture was studied in detail and the sequence of virus morphogenesis was determined by transmission electron microscopy. By 16 h post-infection (p.i.), virus replication was observed in the virogenic stroma by the appearance of nucleocapsids. Polyhedron formation was detected by 24 h p.i. and the polyhedron envelope (PE) was completely formed by 72 h p.i. PE morphogenesis of HaSNPV is significantly different compared to the extensively studied Autograph californica (Ac)MNPV. In AcMNPV-infected cells, fibrillar structures are found in both cytoplasm and nuclei, and the fibrillar structures in nuclei are in close association with maturing polyhedra during PE formation. Fibrillar structures that resemble the AcMNPV fibrillar structures were detected only in the cytoplasm of HaSNPV-infected cells and appeared to interact with calyx precursors there, but their role in PE formation is unclear. However, prominent calyx precursor structures of various shapes and sizes were observed in the nuclei of HaSNPV-infected cells as well, and they appeared to interact with polyhedra during PE formation. Both the calyx precursor structure and the cytoplasmic fibrillar structure were detected only after HaSNPV virion occlusion had started, indicating that they might have a role in formation of PE. Similar calyx precursor structures and cytoplasmic fibrillar structures were observed in both serum-supplemented and serum-free suspension cultures, as well as in HaSNPV-infected larval tissues, indicating that the structures observed are not cell culture artefacts. Introduction Insect pest control is one of the most important issues facing the 21st Century. Helicoverpa and Heliothis spp. (com- monly referred to as heliothis) are a major focus of pest control programmes throughout the world. They attack well over 60 crops (Christian, 1994). Use of chemical insecticides to control these pests has failed in South-East Asia and Australia, resulting in a major crisis with increasing and more widespread resistance (White et al., 1996). Increasing resistance of heliothis to existing chemical insecticides has also increased the cost of control and loss of production (Fitt, 1994 ; McGahan et al., 1991). Failure to control heliothis with chemical insecticides has hastened the development of technology for production of Author for correspondence : Linda Lua. Fax ›61 7 3365 4199. e-mail lindal!cheque.uq.edu.au biological pesticides that have the potential to control heliothis effectively. Helicoverpa armigera single nucleopolyhedrovirus (HaSNPV), a wild-type baculovirus, has the potential for use as a biopesticide for effective control of heliothis. HaSNPV is specific and highly virulent to its host (Hughes et al., 1983 ; Teakle et al., 1985). The development of in vitro production of HaSNPV is becoming increasingly important as the demand for effective heliothis control increases. Various factors relating to the replication of HaSNPV, such as virus infection kinetics, optimal time of infection (TOI), nutrient consumption and passage effect, have been studied previously (Chakraborty et al., 1995, 1996, 1999 ; Chakraborty & Reid, 1999). These studies were aimed primarily at optimizing production processes for this virus. Despite all the reported studies on HaSNPV, little is known about HaSNPV morphogenesis in Helicoverpa zea serum-free suspension cul- ture. In vitro production of high quality and biologically active 0001-7072 # 2000 SGM CFDB
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Virus morphogenesis of Helicoverpa armigera nucleopolyhedrovirus in Helicoverpa zea serum-free...

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
Journal of General Virology (2000), 81, 2531–2543. Printed in Great Britain. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Virus morphogenesis of Helicoverpa armigeranucleopolyhedrovirus in Helicoverpa zea serum-freesuspension culture
Linda H. L. Lua and Steven Reid
Department of Chemical Engineering, The University of Queensland, Queensland 4072, Australia
Helicoverpa armigera single nucleopolyhedrovirus (HaSNPV) replication in Helicoverpa zea serum-free suspension culture was studied in detail and the sequence of virus morphogenesis wasdetermined by transmission electron microscopy. By 16 h post-infection (p.i.), virus replication wasobserved in the virogenic stroma by the appearance of nucleocapsids. Polyhedron formation wasdetected by 24 h p.i. and the polyhedron envelope (PE) was completely formed by 72 h p.i. PEmorphogenesis of HaSNPV is significantly different compared to the extensively studied Autographcalifornica (Ac)MNPV. In AcMNPV-infected cells, fibrillar structures are found in both cytoplasm andnuclei, and the fibrillar structures in nuclei are in close association with maturing polyhedra duringPE formation. Fibrillar structures that resemble the AcMNPV fibrillar structures were detected onlyin the cytoplasm of HaSNPV-infected cells and appeared to interact with calyx precursors there, buttheir role in PE formation is unclear. However, prominent calyx precursor structures of variousshapes and sizes were observed in the nuclei of HaSNPV-infected cells as well, and they appearedto interact with polyhedra during PE formation. Both the calyx precursor structure and thecytoplasmic fibrillar structure were detected only after HaSNPV virion occlusion had started,indicating that they might have a role in formation of PE. Similar calyx precursor structures andcytoplasmic fibrillar structures were observed in both serum-supplemented and serum-freesuspension cultures, as well as in HaSNPV-infected larval tissues, indicating that the structuresobserved are not cell culture artefacts.
IntroductionInsect pest control is one of the most important issues
facing the 21st Century. Helicoverpa and Heliothis spp. (com-monly referred to as heliothis) are a major focus of pest controlprogrammes throughout the world. They attack well over 60crops (Christian, 1994). Use of chemical insecticides to controlthese pests has failed in South-East Asia and Australia, resultingin amajor crisis with increasing and morewidespread resistance(White et al., 1996). Increasing resistance of heliothis toexisting chemical insecticides has also increased the cost ofcontrol and loss of production (Fitt, 1994 ; McGahan et al.,1991). Failure to control heliothis with chemical insecticideshas hastened the development of technology for production of
Author for correspondence: Linda Lua.
Fax 61 7 3365 4199. e-mail lindal!cheque.uq.edu.au
biological pesticides that have the potential to control heliothiseffectively.
Helicoverpa armigera single nucleopolyhedrovirus(HaSNPV), a wild-type baculovirus, has the potential for use asa biopesticide for effective control of heliothis. HaSNPV isspecific and highly virulent to its host (Hughes et al.,1983 ; Teakle et al., 1985). The development of in vitroproduction of HaSNPV is becoming increasingly important asthe demand for effective heliothis control increases.
Various factors relating to the replication of HaSNPV, suchas virus infection kinetics, optimal time of infection (TOI),nutrient consumption and passage effect, have been studiedpreviously (Chakraborty et al., 1995, 1996, 1999 ; Chakraborty& Reid, 1999). These studies were aimed primarily atoptimizing production processes for this virus. Despite all thereported studies on HaSNPV, little is known about HaSNPVmorphogenesis in Helicoverpa zea serum-free suspension cul-ture. In vitro production of high quality and biologically active
0001-7072 # 2000 SGM CFDB

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
L. H. L. Lua and S. ReidL. H. L. Lua and S. Reid
Fig. 1. Viable cell density (E), % viability(_) and volumetric polyhedra yield (+)of Helicoverpa zea suspension cultures inSF900II medium infected with HaSNPV at5¬105 cells/ml with an m.o.i. of 2 p.f.u.per cell. The error bars indicate standarddeviations of the means of triplicatecounts of duplicate cultures.
HaSNPV necessitates an understanding of virus replication andassembly in this production system. Knowledge of themorphogenic events in this system will aid in optimizingproduction parameters such as the time of harvest for fullymature polyhedra, and in documenting any structuraldifferences between this system and other reported baculo-viruses.
Previous studies on Helicoverpa zea SNPV (Goodwin et al.,1973 ; Granados, 1978, 1981 ; Ignoffo et al., 1971 ; Lenz et al.,1991 ; McIntosh & Ignoffo, 1981 ; Rice et al., 1989 ; Yamada etal., 1982) revealed very little about the morphogenesis of thevirus. A brief sequence of morphogenesis was described byGranados et al. (1981). However, virion occlusion andpolyhedron envelope morphogenesis was not reported indetail. Ultrastructural studies on replication of other NPVshave been performed extensively over the last two decades.Most of these studies were done using either infected larvaltissues (Adams et al., 1977 ; Granados & Lawler, 1981 ; Harrap,1972b ; Hess & Falcon, 1978 ; Kawamoto et al., 1977 ; Tewari &Datta, 1996), or by using monolayers of cells grown in serum-containing media (Carstens et al., 1979 ; Fraser, 1986 ; Knudson& Harrap, 1976 ; MacKinnon et al., 1974 ; Young et al., 1993).The most well documented virus morphogenesis processavailable is for Autographa californica NPV (AcMNPV), whichwas reviewed by Williams & Faulkner (1997).
NPVs have a biphasic replication cycle that is localized inthe cell nucleus (Vlak, 1992). The first phase of replicationgenerates nucleocapsids and budded virus. The second phaseinvolves the development of large proteinaceous para-crystalline occlusion bodies, also known as polyhedra, whichare composed mainly of a polyhedrin protein of about 30 kDa,in which numerous enveloped virions are embedded. Varyingfrom 0±5 to 15 µm in diameter, the polyhedra stabilize virions,thus allowing them to remain viable for long periods in theenvironment (Bergold, 1963). Each polyhedron has an electron-
dense layer known as the polyhedron calyx or polyhedronenvelope (PE), the function of which is unknown. Gross et al.(1994) suggested that the PE may prevent virions fromdislodging from the polyhedron matrix and, in general, helpsseal the virus complex. The PE consists mainly of carbo-hydrates, with 60% hexose, 29% pentose and small amountsof uronic acids and hexosamines (Minion et al., 1979), and atleast one protein (Whitt & Manning, 1988).
This paper documents the morphogenic sequence ofHaSNPV in H. zea serum-free suspension cultures and furtherdiscusses prominent structural differences between the mor-phogenesis of HaSNPV and that of other baculoviruses. Thisstudy improves our understanding of HaSNPV replication inserum-free suspension cultures and will undoubtedly aid inthe development of an optimized production process ofHaSNPV as a commercial biopesticide.
Methods+ Cell line. Helicoverpa zea cells were a gift from CSIRO, Division ofEntomology, Canberra, Australia. The cell line was isolated from pupalovarian tissue, strain BCIRL-HZ-AM1 (McIntosh & Ignoffo, 1981). Theuncloned cell line was gradually adapted to SF900II (GIBCO BRL) serum-free medium in suspension culture (Chakraborty et al., 1995). Cultures aretypically grown in 250 ml Erlenmeyer flasks on an orbital shakeroperated at 120 r.p.m., inside a refrigerated incubator at 28 °C.
+ Virus. HaSNPV (uncloned) passage one stock was established in cellsgrown in SF900II plus 10% foetal bovine serum (CSL, Australia) withhaemolymph collected from larvae (obtained from the Department ofPrimary Industries, Long Pocket, Queensland). The haemolymph from 22infected larvae was pooled together into a total volume of 100 ml ofmedium. It was frozen in liquid nitrogen and later thawed and filteredthrough a 0±2 µm filter before use. 1±8 ml was used to infect a 20 mlHelicoverpa zea suspension culture at 3¬10& cells}ml. 80 ml fresh mediumwas added at 4 days p.i., to prevent over growth of cells and to providefresh nutrients. Budded virus of passage one was harvested at 80% cell
CFDC

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
Virus morphogenesis of H. armigera nucleopolyhedrovirusVirus morphogenesis of H. armigera nucleopolyhedrovirus
viability (8 days p.i.). The cell suspension was centrifuged at 1000 g for
5 min at room temperature. The cell pellet was discarded and the virus-
containing supernatant was stored at ®70 °C. Passage two stock was
made from passage one virus in the following way, for use in this study.
A culture with a 50 ml working volume was seeded at 3¬10& cells}ml
in SF900II medium and allowed to grow to 1¬10' cells}ml. It was
diluted back to 5¬10& cells}ml with fresh medium and infected with an
m.o.i. of 0±5 p.f.u. per cell. Passage two budded virus was harvested at
70% cell viability (5 days p.i.), stored at ®70 °C and titred before use.
+ Quantification of budded virus. The virus titres of passage one
and two stocks were determined by a plaque assay using Helicoverpa zea
cells in SF900II plus 10% foetal bovine serum cells. 60 mm Petri dishes
(Corning) were seeded with 3 ml Helicoverpa zea cells at 3¬10& cells}ml,
and incubated overnight at 28 °C. The medium was removed from the
cells and 0±5 ml of virus at an appropriate dilution was inoculated onto
the plates and rocked gently for 4 h. Virus inoculum was removed before
4 ml of medium–agarose mix was overlaid onto each plate. The overlay
consisted of an equal volume of 2¬ SF900II plus 10% foetal bovine
serum and 2% low melting point SeaPlaque agarose (FMC BioProducts).
+ Virus infection. Duplicate 50 ml cultures were seeded at
3¬10& cells}ml in SF900II medium and allowed to grow to
1¬10' cells}ml before being diluted to 5¬10& cells}ml with 50 ml fresh
medium. These cultures were infected at an m.o.i. of 2 p.f.u. per cell.
Samples were collected for transmission electron microscopy (TEM)
processing at 4 hourly intervals post-infection for the first 48 h p.i. and
subsequently every 24 h p.i. Samples from mock-infected cultures were
also collected and processed for TEM. Cell density and viability were
determined daily in triplicate using the 0±1% trypan blue exclusion
method (Nielsen et al., 1991). To determine polyhedra density, cells were
lysed with 0±5% SDS for 1 h at 28 °C before triplicate counts were done
in a haemocytometer counting chamber.
An infected Sf9 cell pellet was obtained from Matthew Rosinki
(University of Queensland). In brief, Sf9 cells were infected with a
recombinant AcMNPV expressing β-galactosidase (pAc-360 β-gal), at an
m.o.i. of 0±001 and harvested at 4 days p.i. Another HaSNPV infection in
SF900II supplemented with 10% serum was performed to determine that
some of the structures observed in the serum-free medium were not a
result of the lack of serum. Infections in the serum-supplemented cultures
were carried out under conditions similar to those used for serum-free
cultures and infected cell samples were collected at 2 and 3 days p.i. for
TEM. Fourth instar Helicoverpa armigera larvae were also infected with in
vivo- produced HaSNPV polyhedra, and the fat bodies of infected larvae
were harvested at 4 days p.i. for TEM.
+ Transmission electron microscopy. TEM processing was similar
to previous reports in the literature but was modified for cell culture
samples. The cells were harvested at different times post-infection,
pelleted and fixed with 3% glutaraldehyde in 0±1 M phosphate buffer
pH 7±2, for at least an hour at 4 °C. Cells were washed once with
phosphate buffer pH 7±2 at 4 °C and resuspended in 2% SeaPlaque
agarose. The cell–agarose block was cut into 1 mm squares and washed
twice again with phosphate buffer, before being fixed with 1% osmium
tetroxide in 0±1 M cacodylate buffer, pH 6±8, for 1 h at 4 °C. The samples
were then washed once with 0±1 M cacodylate buffer, pH 6±8, and once
with water at room temperature. After washing, they were sequentially
dehydrated once in 50, 70 and 90% acetone and twice in 100% acetone
for 5 min at each step. Samples were infiltrated with 50% Spurr’s low
viscosity resin for 1 h, followed by 100% Spurr’s resin for a further 1 h.
After one change in 100% Spurr’s resin, they were left in resin overnight
on a rotator. Samples were embedded in fresh resin and polymerized at60 °C for 3 days. Ultra-thin sections (60–80 nm) were cut with a LeicaUltracut UCT microtome. The sections on copper grids were stained for2 min in 5% uranyl acetate and 1 min in Reynolds ’ lead stain before beingviewed under a JEOL 1010 transmission electron microscope.
ResultsVirus infection
Polyhedra were first detectable at 48 h p.i. and the yield atthis time was 10% of the final peak volumetric yield (Fig. 1).The peak volumetric yield of polyhedra was obtained whencell viabilities were less than 5% (144 h p.i.). The volumetricyield remained level after 0% cell viability had been achieved,indicating that no disintegration of polyhedra was occurring.The specific yield of polyhedra was 161³3 polyhedra per cell.The size of polyhedra ranged from 0±7 to 1±2 µm, based onmeasurements of a small number of polyhedra photographedvia TEM. These data indicate that the cultures were rapidlyinfected at an m.o.i. of 2 p.f.u. per cell and achieved a highspecific cell yield, thus highlighting that the sequence of virusmorphogenesis documented is indeed based on culturesinfected under near-optimum conditions.
Sequence of virus morphogenesis
TEM examination of ultra-thin sections of infected cellsamples at intervals of 4 h p.i. allowed determination of thesequence of HaSNPV morphogenesis in Helicoverpa zeasuspension cultures. The sequence of infection was docu-mented based on viewing the rapidly infected and highproducing cells amongst the population of infected cells.
Mock-infected cells harvested at 24 and 48 h p.i. were alsoobserved under TEM as a control to the infection studies (Fig.2a, b). Infection with an m.o.i. of 2 p.f.u. per cell was insufficientto observe early infection events (such as virus attachment andentry into host cells) in the first 4 h p.i. Infection with a higherm.o.i. of 10 or 100 p.f.u. per cell could not be performed due tothe low virus stock titre (2¬10( p.f.u.}ml). Pictures wereobtained from the secondary infections (at 24 h p.i.) to illustratevirus attachment and entry at the plasma membrane (Fig.2 c). The first indication of virus infection was enlargement ofthe cell nucleus and formation of a loose network of granularmaterial in the middle of the nucleus (Fig. 2d). Development ofthis intranuclear virus replication centre, the virogenic stroma,was first detected at 12 h p.i. Clumps of heterochromatin,normally seen dispersed throughout the nucleoplasm of anuninfected cell (Fig. 2a, b), were reduced and displacedperipherally along the inner nuclear membrane by the formingstroma. Maturation of the virogenic stroma yielded a signifi-cant and morphologically distinct peristromal compartment ofnucleoplasm, called the ring zone (Williams & Faulkner, 1997).Examination of the 16 h p.i. sample revealed formation of shortprogeny nucleocapsids in the stromal network (Fig. 2 e). These
CFDD

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
L. H. L. Lua and S. ReidL. H. L. Lua and S. Reid
Fig. 2. For legend see facing page.
CFDE

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
Virus morphogenesis of H. armigera nucleopolyhedrovirusVirus morphogenesis of H. armigera nucleopolyhedrovirus
nucleocapsids appeared to associate very closely with thestromal edge. Short membrane profiles were observed in thering zone. These membrane profiles likely represent the sourceof virion envelopes. Some unidentified circular structures werealso seen near the inner nuclear membrane.
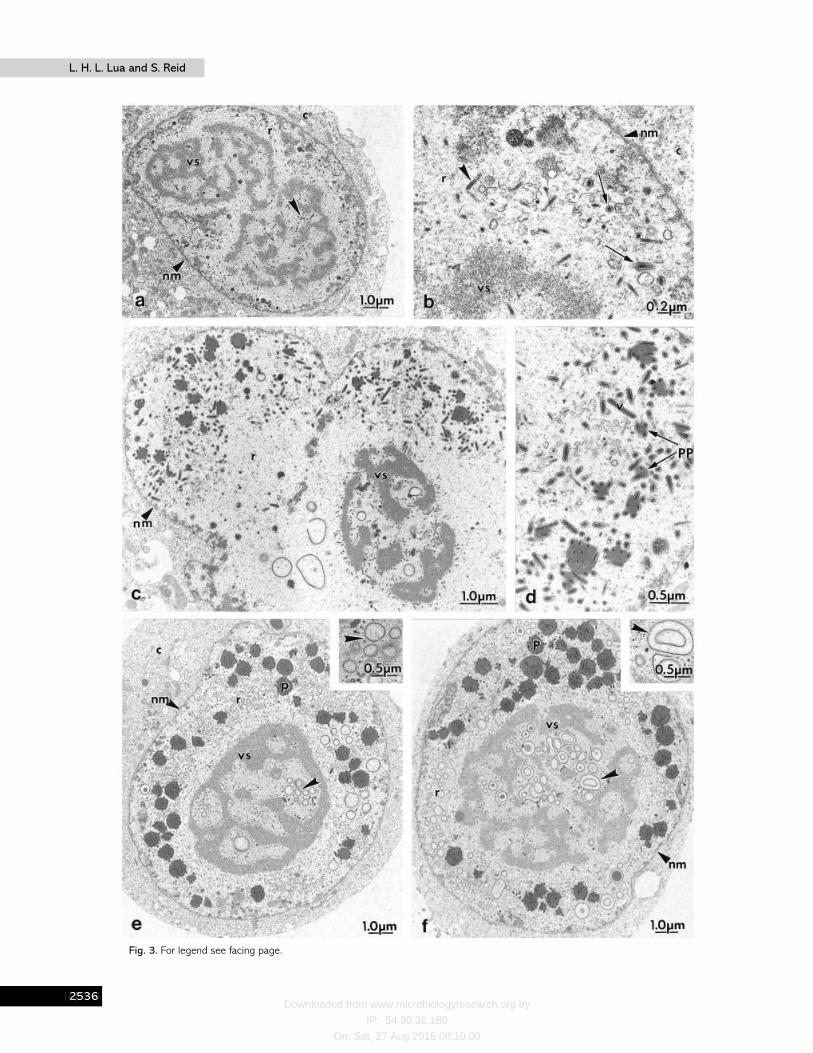
By 20 h p.i., the matured virogenic stroma had condensedinto a dense lattice-like structure, filling the nucleus almostcompletely, and the intrastromal spaces were enriched withnucleocapsids (Fig. 3a). At this time, intranuclear nucleocapsidenvelopment was also observed in the ring zone (Fig.3b). Nucleocapsids formed within the virogenic stroma hadmoved to the ring zone, specifically to the locale of themembrane profiles, presumably to obtain their envelopes. At24 h p.i., many virus particles were budding from the plasmamembrane, acquiring a loose-fitting envelope. Unenvelopedvirus was often detected in the cytoplasm and vacuoles.During this interval, there was also an increase in virusreplication and virion envelopment with the nucleus. Furtherenlargement of the cell nucleus had reduced the ‘ cytoplasmspace ’ between the plasma membrane and the nuclearmembrane. The virogenic stroma had also reduced in size,resulting in a wider ring zone (Fig. 3 c). Early virion occlusionwas detected in the ring zone. Polyhedrin proteins weredeposited around the enveloped virions, occluding the virionsprogressively (Fig. 3d). Only enveloped virions appeared to beoccluded. By 28 h p.i., the polyhedra had increased in size,occluding more virions and were approaching maturity. Someelectron-dense ring structures were first detected in thevirogenic stroma around this time (Fig. 3 e). These structuresappeared as multiple layers of circles (Fig. 3 f).
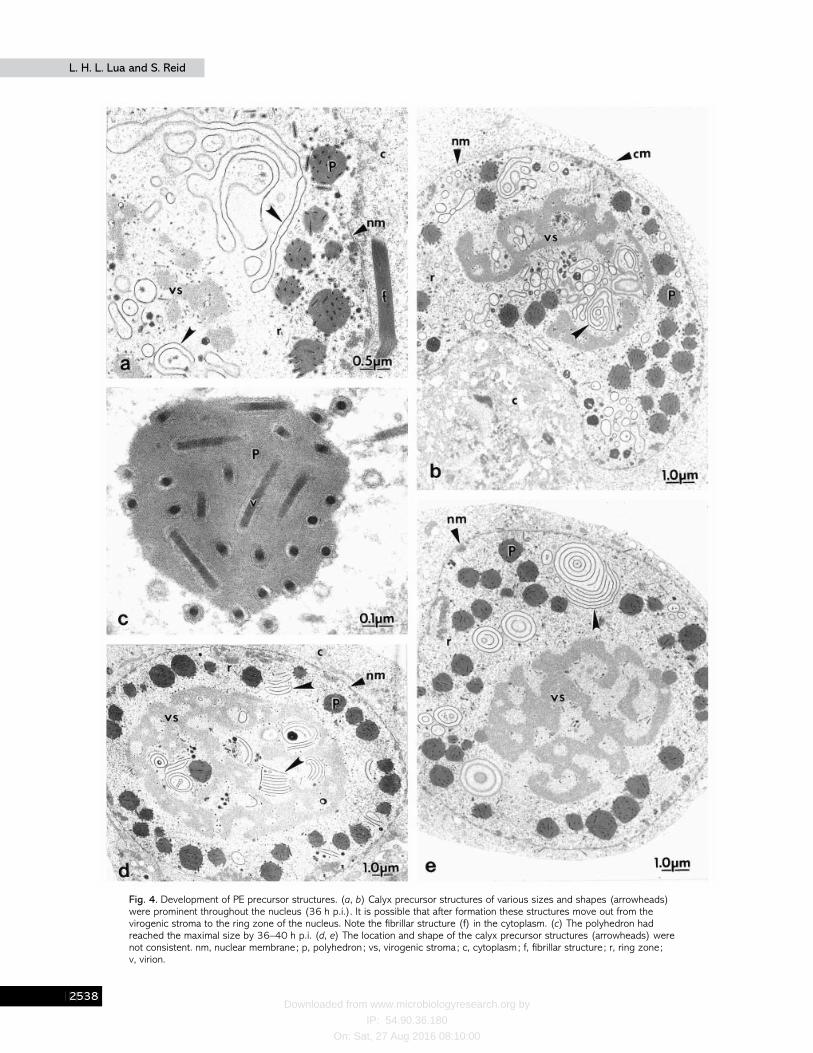
Fibrillar structures were also noted in the cytoplasm ofinfected cells (Fig. 4a). The electron-dense ring structures haddeveloped into irregular folds of various sizes that ultra-structurally appeared to be calyx precursors as observed byWilliams & Faulkner (1997). These structures were prominentin the virogenic stroma and ring zone by 32 h p.i. Theyappeared to have moved to the ring zone, in the close vicinityof the maturing polyhedra (Fig. 4b). Many polyhedra hadreached their maximal size by 36 h p.i. (Fig. 4 c). The calyxprecursor structures were not uniform in size or shape (Fig. 4d,e). They were made up of multiple electron-dense layers andappeared either as ‘ stacks ’, layers of concentric circles or swirlsof irregular shape.
Formation of a PE was first observed at around 48 h p.i. Thecalyx precursor structures appeared to encircle maturing
Fig. 2. Mock-infected cells and infected cells. (a) Mock-infected cell at 24 h p.i. (1¬106 cells/ml). Clumps of heterochromatinwere dispersed throughout the nucleus. (b) Mock-infected cell at 48 h p.i. (2¬106 cells/ml). Note the increase in vacuoles inthe cytoplasm. (c) A budded virus with loose fitting envelope (arrow) entering the cell by endocytosis. (d) Early virogenicstroma (12 h p.i.) forming within the infected cell nucleus, displacing clumps of heterochromatin (arrowheads) to the peripheryof the nucleus. (e) At 16 h p.i., the virogenic stroma had condensed into a dense structure and progeny nucleocapsids(arrowheads) appeared in the stromal network (see also insert I). Short membrane profiles (arrows) were observed in the ringzone (see also insert III). An unknown viral structure (insert II) also appeared near the inner nuclear membrane. BV, buddedvirus ; cm, cell membrane; vs, virogenic stroma; h, heterochromatin ; Nu, nucleus ; nm, nuclear membrane; c, cytoplasm; r, ringzone.
polyhedra (Fig. 5a). PE formed at this interval were not fullymatured, as they were not continuous around the polyhedron(Fig. 5b). By 72 h p.i., many polyhedra were fully mature, eachcontaining a complete electron-dense continuous PE (Fig. 5 c).Calyx precursor structures were no longer visible in the nucleithat contained mature polyhedra and cell plasma membraneswere deformed. By 96 h p.i., most cells observed had deformedor disintegrated cell plasma membranes but the nuclearmembranes remained intact (Fig. 5d). All cell nuclei viewedunder TEM at 144 h p.i. were lysed, with mature polyhedrabeing released into the medium.
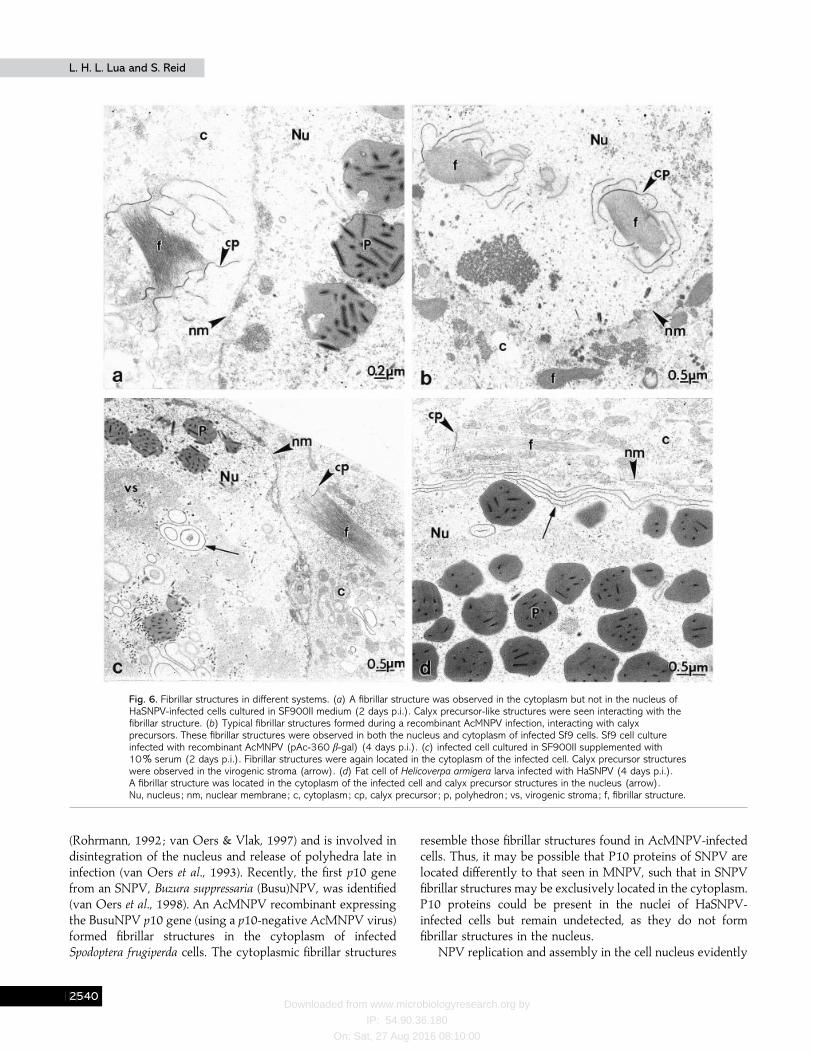
Fibrillar structures, presumably the homologues of fibrillarstructures observed in all AcMNPV infections, were foundonly in the cytoplasm of HaSNPV-infected cells cultured inSF900II medium (Fig. 6a). No structure similar to the calyxprecursor structures or the fibrillar structures was detected inthe cytoplasm of the mock-infected cells (Fig. 2a, b). Tounderstand the significance of these observations and to verifythat these structures observed in HaSNPV-infected serum-freesuspension cultures are not artefacts, virus morphogenesis inother cell-virus systems was studied. A control infection of Sf9cells with a recombinant AcMNPV expressing β-galactosidase(pAc-360 β-gal) clearly showed previously reported fibrillarstructures in both the nuclei and cytoplasm of infected cells(Fig. 6b). The fibrillar structures usually had calyx precursorsassociated with them. HaSNPV infection in Helicoverpa zea cellsgrown in serum-supplemented medium also showed fibrillarstructures, but located only in the cytoplasm of the infectedcells (Fig. 6 c). Fibrillar structures were again found located onlyin the cytoplasm of HaSNPV-infected larval fat cells (Fig. 6d).
In an effort to document that the HaSNPV morphogenicevents observed are representative of an infected cell popu-lation, cells were harvested at 16, 24, 32, 40, 48, 72 and 96 hp.i., and 50–100 infected cells were scored into six categoriesbased upon the most advanced event recognized in each cell.The categories are virogenic stroma formation, nucleocapsidproduction, virion envelopment, initial virion occlusion, latepolyhedron formation and mature polyhedra. From the samepopulation of scored cells, the percentage of infected cellsexhibiting calyx precursor structures and fibrillar structureswas also determined. Fig. 7 presents the typical morphogenicprocess of a HaSNPV-infected cell population. At 16 h p.i., allinfected cells viewed were either forming the virogenic stromaor producing nucleocapsids ; by 96 h p.i. all infected cellsshowed polyhedron morphogenesis. Fig. 7 clearly indicates
CFDF
Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
L. H. L. Lua and S. ReidL. H. L. Lua and S. Reid
Fig. 3. For legend see facing page.
CFDG

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
Virus morphogenesis of H. armigera nucleopolyhedrovirusVirus morphogenesis of H. armigera nucleopolyhedrovirus
that both calyx precursor structures and fibrillar structuresappeared after virion occlusion had started and increased aspolyhedron formation and PE morphogenesis increased.
DiscussionThis paper presents a detail electron-microscopy study on
the replication of HaSNPV in serum-free suspensionHelicoverpazea cell cultures, allowing further understanding of HaSNPVmorphogenesis in vitro. The morphogenesis of HaSNPV isgenerally similar to previous reports on MNPV such asAcMNPV (Williams & Faulkner, 1997), Trichoplusia ni(Tn)MNPV (MacKinnon et al., 1974) and Spodoptera frugi-perda (Sf)MNPV (Knudson & Harrap, 1976), yet significantdifferences were observed.
In brief, the main morphogenic events are replication ofprogeny virus, virion envelopment, virion occlusion and PEformation. The first sign of infection is formation of thevirogenic stroma, an electron-dense matte with electron-lucentintrastromal spaces. Nucleocapsids develop adjacent to thestroma and inside the intrastromal spaces, near the edge of thestromal matte. The virogenic stroma contains discrete DNA-and RNA-containing structures that are related to replicationand packaging of the progeny virus (Young et al., 1993). Invitro studies on AcMNPV-infected Sf21 cells have shown thatthe electron-dense matte is extremely sensitive to RNasewhereas areas adjacent to the intrastromal spaces are sensitiveto DNase.
Nucleocapsids made in the virogenic stroma are subse-quently enveloped by one of two processes. They are eitherenveloped during budding through the cell plasma membrane,acquiring a loosely fitting envelope (budded virus), orenveloped in the nucleus through de novo envelope mor-phogenesis to form preoccluded virions. In this study, bothmorphogenic events occurred at about the same time post-infection. These two forms of virus differ morphologically andbiochemically. Braunagel & Summers (1994) have demon-strated that the protein and lipid composition of the envelopesof budded virus and preoccluded virions is significantlydifferent.
The de novo intranuclear nucleocapsid envelopment processis clearly observed in this study, and is similar to such processesreported for other baculoviruses (Fraser, 1986 ; Stoltz et al.,1973). At 16 h p.i., short membrane profiles synthesized denovo in the nuclei of infected cells accumulate in areas of thering zone, and nucleocapsids that migrate out to the ring zone,
Fig. 3. Virion envelopment and polyhedron formation. (a, b) By 20 h p.i., the virogenic stroma contained a large quantity ofnucleocapsids (arrowheads) and intranuclear nucleocapsid envelopment was observed in the ring zone. A few envelopedvirions were seen at this time (arrows). (c) At 24 h p.i., the virogenic stroma had reduced in size and virion occlusion wasobserved in the ring zone. (d) Enveloped virions were progressively occluded by deposition of polyhedrin protein. (e) By 28 hp.i., polyhedra were increasing in size and approaching maturity. Calyx precursor structures (arrowheads) were first noticed inthe virogenic stroma at around this time. (f) The calyx precursor structures had increased in size and numbers by 32 h p.i.(insert). vs, virogenic stroma; pp, polyhedrin protein ; p, polyhedron ; r, ring zone; nm, nuclear membrane; c, cytoplasm;v, virion.
form preoccluded enveloped virions. These intranuclear mem-brane profiles were not observed in mock-infected cultures.
The initial stages of polyhedron formation can be detectedby deposition of polyhedrin protein between envelopedvirions, forming a typical lattice structure between the virions.Naked, unenveloped nucleocapsids are not occluded. Thevirion envelope may have a possible role as a site forpolyhedrin protein polymerization (Wood, 1980). The en-velope of occluded virions in the polyhedrin matrix fits snuglyaround the nucleocapsid, unlike the budded virus envelope.Enlargement of the polyhedra were observed over time,between 24 and 36 h p.i. It is unclear what factors determinethe size of polyhedra or why the deposition of polyhedrinprotein ceases at a certain stage. Harrap (1972a) suggested thatthe available pool of polyhedrin protein monomer may beexhausted at this time.
Early PE morphogenesis in HaSNPV appears to be markedby the appearance of calyx precursor structures in the virogenicstroma. These structures appear to move progressively to thering zone to interact with maturing polyhedra. Ultra-structurally, they appear similar to the calyx precursors thatinteract with fibrillar structures of AcMNPV (van Oers & Vlak,1997 ; Williams & Faulkner, 1997 ; Williams et al., 1989) andother baculoviruses (Knudson & Harrap, 1976 ; MacKinnon etal., 1974). However, in HaSNPV, these structures appeared toemerge and develop randomly in the nuclei, especially in thevirogenic stroma, and to have no definite form. The calyxprecursor structures often encircle the polyhedra duringacquisition of the PE. These calyx precursor structures are mostlikely the PE precursors, as they appear to be involved in theapplication of the PE to the surface of the polyhedra. Theirappearance coincided with polyhedron formation and theywere not observed in the nuclei of infected cells aftercompletion of PE formation, indicating that they have a role inPE morphogenesis.
Fibrillar structures enriched in P10 proteins are commonlyfound in the nuclei and cytoplasm of AcMNPV-infected cells(Lee et al., 1996 ; Rohrmann, 1992, Williams & Faulkner, 1997).In AcMNPV-infected cell nuclei, these fibrillar structuresusually associate with maturing polyhedra during PE mor-phogenesis. This phenomenon is frequently observed and welldocumented for other baculovirus infections such as TnMNPV(MacKinnon et al., 1974), SfMNPV (Knudson & Harrap, 1976)and Orgyia pseudotsugata (Op)MNPV (Gross et al., 1994).However, such a prominent fibrillar structure was not observedin the nuclei of HaSNPV-infected cells. Instead, fibrillar
CFDH
Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
L. H. L. Lua and S. ReidL. H. L. Lua and S. Reid
Fig. 4. Development of PE precursor structures. (a, b) Calyx precursor structures of various sizes and shapes (arrowheads)were prominent throughout the nucleus (36 h p.i.). It is possible that after formation these structures move out from thevirogenic stroma to the ring zone of the nucleus. Note the fibrillar structure (f) in the cytoplasm. (c) The polyhedron hadreached the maximal size by 36–40 h p.i. (d, e) The location and shape of the calyx precursor structures (arrowheads) werenot consistent. nm, nuclear membrane; p, polyhedron ; vs, virogenic stroma; c, cytoplasm; f, fibrillar structure ; r, ring zone;v, virion.
CFDI

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
Virus morphogenesis of H. armigera nucleopolyhedrovirusVirus morphogenesis of H. armigera nucleopolyhedrovirus
Fig. 5. Formation of polyhedron envelope. (a) A ‘cage-like ’ structure (arrowhead) formed around a polyhedron (48 h p.i.).(b) The polyhedron envelope was incomplete and not continuous around the polyhedron at this time. (c) By 72 h p.i.,polyhedra were fully matured and each had an electron-dense envelope. (d) Disintegration of the cell plasma membrane aftercompletion of PE formation but the nuclear membrane remained intact. p, polyhedron ; PE, polyhedron envelope; nm, nuclearmembrane; Nu, nucleus.
structures (presumptive homologues of AcMNPV fibrillarstructures), resembling to some extent the AcMNPV fibrillarstructures, were clearly detected only in the cytoplasm ofHaSNPV-infected cells. Mock-infected cells showed no fibrillarstructures, indicating that these cytoplasmic fibrillar structuresare associated with HaSNPV infection events. These cyto-plasmic fibrillar structures were first detected at around28–32 h p.i., just after initial virion occlusion events in thenuclei. Occasionally, calyx precursor-like structures were seeninteracting with these cytoplasmic fibrillar structures. It cannotbe ruled out that the absence of fibrillar structure in the nuclei
of infected cells and their location only in the cytoplasm is anartefact of serum-free cultures. However, HaSNPV infectionsin serum-supplemented medium produced similar results(appearance of fibrillar structures in the cytoplasm of infectedcells but not in the nuclei). This is further confirmed by theobservation that Helicoverpa armigera larva fat cells infectedwith HaSNPV also have cytoplasmic fibrillar structures only.
The P10 protein is known to be a structural component ofthe fibrillar structures, calyx precursors and PE in AcMNPV-infected cells (Lee et al., 1996). P10 is a poorly conservedprotein that is hyperexpressed very late in infection
CFDJ
Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
L. H. L. Lua and S. ReidL. H. L. Lua and S. Reid
Fig. 6. Fibrillar structures in different systems. (a) A fibrillar structure was observed in the cytoplasm but not in the nucleus ofHaSNPV-infected cells cultured in SF900II medium (2 days p.i.). Calyx precursor-like structures were seen interacting with thefibrillar structure. (b) Typical fibrillar structures formed during a recombinant AcMNPV infection, interacting with calyxprecursors. These fibrillar structures were observed in both the nucleus and cytoplasm of infected Sf9 cells. Sf9 cell cultureinfected with recombinant AcMNPV (pAc-360 β-gal) (4 days p.i.). (c) infected cell cultured in SF900II supplemented with10% serum (2 days p.i.). Fibrillar structures were again located in the cytoplasm of the infected cell. Calyx precursor structureswere observed in the virogenic stroma (arrow). (d) Fat cell of Helicoverpa armigera larva infected with HaSNPV (4 days p.i.).A fibrillar structure was located in the cytoplasm of the infected cell and calyx precursor structures in the nucleus (arrow).Nu, nucleus ; nm, nuclear membrane; c, cytoplasm; cp, calyx precursor ; p, polyhedron ; vs, virogenic stroma; f, fibrillar structure.
(Rohrmann, 1992 ; van Oers & Vlak, 1997) and is involved indisintegration of the nucleus and release of polyhedra late ininfection (van Oers et al., 1993). Recently, the first p10 genefrom an SNPV, Buzura suppressaria (Busu)NPV, was identified(van Oers et al., 1998). An AcMNPV recombinant expressingthe BusuNPV p10 gene (using a p10-negative AcMNPV virus)formed fibrillar structures in the cytoplasm of infectedSpodoptera frugiperda cells. The cytoplasmic fibrillar structures
resemble those fibrillar structures found in AcMNPV-infectedcells. Thus, it may be possible that P10 proteins of SNPV arelocated differently to that seen in MNPV, such that in SNPVfibrillar structures may be exclusively located in the cytoplasm.P10 proteins could be present in the nuclei of HaSNPV-infected cells but remain undetected, as they do not formfibrillar structures in the nucleus.
NPV replication and assembly in the cell nucleus evidently
CFEA

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
Virus morphogenesis of H. armigera nucleopolyhedrovirusVirus morphogenesis of H. armigera nucleopolyhedrovirus
Fig. 7. Percentage of an infected cell population exhibiting different stages of virus morphogenic events, at 16, 24, 32, 40, 48,72 and 96 h p.i. Infected cells (50–100) were viewed under TEM and based upon the most advanced event recognized ineach cell, they were scored into six categories : VS, virogenic stroma formation ; NC, nucleocapsid production ; V, virionenvelopment ; VO, virion occlusion ; PF, late polyhedron formation ; MP, mature polyhedra. From the same cells that werescored, the percentage of these cells exhibiting calyx precursor structures (CP) and fibrillar structures (F) were also determinedand are presented in the figure (continuous and dashed lines).
involves a complex but elegantly controlled synthetic process.This paper clearly documents the unique morphogenesis ofcalyx precursor structures and cytoplasmic fibrillar structuresduring the HaSNPV PE morphogenic process, which has notbeen reported previously in the literature. Calyx precursorstructures and the exclusive cytoplasmic location of fibrillarstructures could be unique to HaSNPV morphogenesis sincethey are observed in both serum and serum-free cultures andalso in the in vivo system. This ultrastructural study providesan outline of a general mode of HaSNPV replication inHelicoverpa zea cells, with time-scales for several morphogenicevents, which will facilitate further investigations on pro-duction of viral proteins specific to PE formation, such as P10and PE protein. Research to identify the nucleotide sequencesand level of expression of these critical viral proteins is underway.
This research was funded by the Australian Research Council and theGrains Research and Development Corporation, with scholarship supportfrom the University of Queensland (UQ).
We would like to thank Dr Robert E. Teakle (Queensland Departmentof Primary Industry) for the generous gift of HaSNPV, Dr ShuklaChakraborty (UQ) for her efforts in collection of the original virushaemolymph, Dr Dall (CSIRO) for the gift of the Helicoverpa zea cell lineand Matthew Rosinki (UQ) for his kind contribution of the rAcMNPV-
infected Sf9 cells. Dr Teakle’s assistance in the larvae EM work is alsogratefully acknowledged. We also acknowledge the valuable technicaladvice provided by the Centre for Microscopy and Microanalysis (UQ).
ReferencesAdams, J. R., Goodwin, R. H. & Wilcox, T. A. (1977). Electronmicroscopic investigations on invasion and replication of insect baculo-viruses in vivo and in vitro. Biologie Cellulaire 28, 261–268.
Bergold, G. H. (1963). The nature of nuclear polyhedrosis viruses. InInsect Pathology : An Advance Treatise, pp. 413–455. Edited by E. A.Steinhaus. New York : Academic Press.
Braunagel, S. C. & Summers, M. D. (1994). Autographa californicanuclear polyhedrosis virus, PDV, and ECV viral envelopes andnucleocapsids : structural proteins, antigens, lipids and fatty acid profiles.Virology 202, 315–328.
Carstens, E. B., Tjia, S. T. & Doerfler, W. (1979). Infection of Spodopterafrugiperda cells with Autographa californica nuclear polyhedrosis virus.Virology 99, 386–398.
Chakraborty, S., Kanhaisingh, A., Greenfield, P. F., Reid, S., Monsour,C. J. & Teakle, R. (1995). In vitro production of wild type Heliothisbaculoviruses for use as biopesticides. Australasian Biotechnology 5,82–86.
Chakraborty, S., Greenfield, P. F. & Reid, S. (1996). In vitro productionstudies with a wild-type Helicoverpa baculovirus. Cytotechnology 22,217–224.
CFEB

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
L. H. L. Lua and S. ReidL. H. L. Lua and S. Reid
Chakraborty, S., Monsour, C., Teakle, R. & Reid, S. (1999). Yield,biological activity, and field performance of a wild-type HelicoverpaNucleopolyhedrovirus produced in Helicoverpa zea cell cultures. Journal ofInvertebrate Pathology 73, 199–205.
Chakraborty, S. & Reid, S. (1999). Serial passage of a Helicoverpaarmigera nucleopolyhedrovirus in Helicoverpa zea cell cultures. Journal ofInvertebrate Pathology 73, 303–308.
Christian, P. (1994). Recombinant baculovirus insecticides : catalysts fora change of heart ? In Biopesticides : Opportunities for Australian Industry.Brisbane, Queensland : University of Queensland.
Fitt, G. P. (1994). Cotton pest management. Part 3. An Australianperspective. Annual Review of Entomology 39, 543–562.
Fraser, M. J. (1986). Ultrastructural observations of virion maturation inAutographa californica nuclear polyhedrosis virus infection Spodopterafrugiperda cell cultures. Journal of Ultrastructure and Molecular StructureResearch 95, 189–195.
Goodwin, R. H., Vaughn, J. L., Adams, J. R. & Louloudes, S. J. (1973).The influence of insect cell lines and tissue-culture media on baculoviruspolyhedra production. Miscellaneous Publication of the Entomological Societyof America 9, 66–72.
Granados, R. R. (1978). Early events in the infection of Heliothis zeamidgut cells by a baculovirus. Virology 90, 170–174.
Granados, R. R. & Lawler, K. A. (1981). In vivo pathway of Autographacalifornica baculovirus invasion and infection. Virology 108, 297–308.
Granados, R. R., Lawler, K. A. & Burand, J. P. (1981). Replication ofHeliothis zea baculovirus in an insect cell line. Intervirology 16, 71–79.
Gross, C. H., Russell, R. L. Q. & Rohrmann, G. F. (1994). Orgyiapseudotsugata baculovirus p10 and polyhedron envelope protein genes :analysis of their relative expression levels and role in polyhedronstructure. Journal of General Virology 75, 1115–1123.
Harrap, K. A. (1972a). The structure of nuclear polyhedrosis viruses. I.The inclusion body. Virology 50, 114–123.
Harrap, K. A. (1972b). The structure of nuclear polyhedrosis viruses. III.Virus assembly. Virology 50, 133–139.
Hess, R. T. & Falcon, L. A. (1978). Electron microscope observations ofthe membrane surrounding polyhedral inclusion bodies of insects.Archives of Virology 56, 169–176.
Hughes, P. R., Gettig, R. R. & McCarthy, W. J. (1983). Comparison ofthe time–mortality response of Heliothis zea to 14 isolates of Heliothisnuclear polyhedrosis virus. Journal of Invertebrate Pathology 41, 256–261.
Ignoffo, C. M., Shapiro, M. & Hink, W. F. (1971). Replication and serialpassage of infectious Heliothis nucleopolyhedrosis virus in an establishedline of Heliothis zea cells. Journal of Invertebrate Pathology 18, 131–134.
Kawamoto, F., Suto, C., Kumada, N. & Kobayashi, M. (1977).Cytoplasmic budding of a nuclear polyhedrosis virus and comparativeultrastructural studies of envelopes. Microbiology and Immunology 21,255–265.
Knudson, D. L. & Harrap, K. A. (1976). Replication of a nuclearpolyhedrosis virus in a continuous cell culture of Spodoptera frugiperda :microscopy study of the sequence of events of the virus infection. Journalof Virology 17, 254–268.
Lee, S. Y., Poloumienko, A., Belfry, S., Chen, W., MacAfree, N., Morin,B., Lucarotti, C. & Krause, M. (1996). A common pathway for p10 andcalyx proteins in progressive stages of polyhedron envelope assembly inAcMNPV infected Spodoptera frugiperda larvae. Archives of Virology 141,1247–1258.
Lenz, C. J., McIntosh, A. H., Mazzacano, C. & Monderloh, U. (1991).Replication of Heliothis zea nuclear polyhedrosis virus in cloned cell lines.Journal of Invertebrate Pathology 57, 227–233.
McGahan, P., Lloyd, R. J. & Rynne, K. P. (1991). The cost of Heliothis inQueensland crops. In QDAYS P. I. Conference and Workshop SeriesQC91006, pp. 11–28.
McIntosh, A. H. & Ignoffo, C. M. (1981). Replication and infectivity ofthe single-embedded nuclear polyhedrosis virus, baculovirus Heliothis, inhomologous cell line. Journal of Invertebrate Pathology 37, 258–264.
MacKinnon, E. A., Henderson, J. F., Stoltz, D. B. & Faulkner, P.(1974). Morphogenesis of nuclear polyhedrosis virus under conditionsof prolonged passage in vitro. Journal of Ultrastructural Research 49,419–435.
Minion, F. C., Coons, L. B. & Broome, J. R. (1979). Characterisation ofthe polyhedral envelope of nuclear polyhedrosis virus of Heliothisvirescens. Journal of Invertebrate Pathology 34, 303–307.
Nielsen, L. K., Smyth, G. K. & Greenfield, P. F. (1991). Hemacytometercell count distributions : implications of non-Poisson behavior. Bio-technology Progress 7, 560–563.
Rice, W. C., McIntosh, A. H. & Ignoffo, C. M. (1989). Yield and activityof the Heliothis zea single nuclear polyhedrosis virus propagated in clonedand uncloned lines of Heliothis cells. In Vitro Cellular and DevelopmentalBiology 25, 201–204.
Rohrmann, G. F. (1992). Baculovirus structural proteins. Journal ofGeneral Virology 73, 749–761.
Stoltz, D. B., Pavan, C. & da Cunha, A. B. (1973). Nuclear polyhedrosisvirus : a possible example of de novo intranuclear membrane mor-phogenesis. Journal of General Virology 19, 145–150.
Teakle, R. E., Jensen, J. M. & Mulder, J. C. (1985). Susceptibility ofHeliothis armigera (Lepidoptera : Noctuidea) on sorghum to a nuclearpolyhedrosis virus. Journal of Economic Entomology 78, 1373–1378.
Tewari, S. K. & Datta, R. K. (1996). Ultrastructural studies on nuclearpolyhedrosis virus infecting silkworm, Bombyx mori L. Indian Journal ofExperimental Biology 34, 141–150.
van Oers, M. M. & Vlak, J. M. (1997). The baculovirus 10 kDa protein.Journal of Invertebrate Pathology 70, 1–17.
van Oers, M. M., Flipsen, J. T., Reusken, C. B., Sliwinsky, E. L.,Goldbach, R. W. & Vlak, J. M. (1993). Functional domains of the p10protein of Autographa californica nuclear polyhedrosis virus. Journal ofGeneral Virology 74, 563–574.
van Oers, M. M., Hu, Z., Arif, B. M., van Strien, E. A., van Lent, J. W. &Vlak, J. M. (1998). The single-nucleocapsid nucleopolyhedrovirus ofBuzura suppressaria encodes a P10 protein. Journal of General Virology 79,1553–1562.
Vlak, J. M. (1992). The biology of baculovirus in vivo and in culturedinsect cells. In Baculovirus and Recombinant Protein Processes, pp. 2–9.Edited by J. M. Vlak, E.-J. Schlaeger & A. R. Bernard. Zu$ rich, Switzerland :Editions Roche.
White, G. G., Murray, D. A. H. & Walton, M. P. (1996). Workshop report :Heliothis management in Australia. Brisbane, Australia : CooperativeResearch Centre for Tropical Pest Management, Brisbane ; CooperativeResearch Centre for Sustainable Cotton Production, Narrabi.
Whitt, M. A. & Manning, J. S. (1988). A phosphorylated 34-kDa proteinand a subpopulation of polyhedrin are thiol linked to carbohydrate layersurrounding a baculovirus occlusion body. Virology 163, 33–42.
Williams, G. V. & Faulkner, P. (1997). Cytological changes and viralmorphogenesis during baculovirus infection. In The Baculoviruses, pp.61–108. Edited by L. K. Miller. New York : Plenum.
Williams, G. V., Rohel, D. Z., Kuzio, J. & Faulkner, P. (1989). Acytopathological investigation of Autographa californica nuclear poly-hedrosis virus p10 gene function using insertion}deletion mutants.Journal of General Virology 70, 187–202.
CFEC

Downloaded from www.microbiologyresearch.org by
IP: 54.90.36.180
On: Sat, 27 Aug 2016 08:10:00
Virus morphogenesis of H. armigera nucleopolyhedrovirusVirus morphogenesis of H. armigera nucleopolyhedrovirus
Wood, H. A. (1980). Isolation and replication of an occlusion body-deficient mutant of the Autographa californica nuclear polyhedrosis virus.Virology 105, 338–344.
Yamada, K., Sherman, K. E. & Maramorosch, K. (1982). Serial passageof Heliothis zea singly embedded nuclear polyhedrosis virus in ahomologous cell line. Journal of Invertebrate Pathology 39, 185–191.
Young, J. C., MacKinnon, E. A. & Faulkner, P. (1993). The architectureof the virogenic stroma in isolated nuclei of Spodoptera frugiperda cells invitro infected by Autographa californica nuclear polyhedrosis virus. Journalof Structural Biology 110, 141–153.
Received 30 March 2000; Accepted 26 June 2000
CFED




















