
Pyramiding of modified cry1Ab and cry1Ac genes of Bacillus thuringiensis in transgenic chickpea (...
16
Pyramiding of modified cry1Ab and cry1Ac genes of Bacillus thuringiensis in transgenic chickpea (Cicer arietinum L.) for improved resistance to pod borer insect Helicoverpa armigera Meenakshi Mehrotra • Aditya K. Singh • Indraneel Sanyal • Illimar Altosaar • D. V. Amla Received: 11 January 2010 / Accepted: 23 July 2011 / Published online: 11 August 2011 Ó Springer Science+Business Media B.V. 2011 Abstract The modified cry1Ab and cry1Ac insecti- cidal genes of Bacillus thuringiensis (Bt) under the control of two different constitutive promoters have been introduced into chickpea (Cicer arietinum L.) by Agrobacterium-mediated transformation of pre-condi- tioned cotyledonary nodes. 118 stable transformed T 0 plants as independent transformation events were obtained expressing individual cry1Ab, cry1Ac or both pyramided genes for their co-expression driven by either cauliflower mosaic virus 35S promoter with duplicated enhancer (CaMV35S) or synthetic constitutive promoter (Pcec) and their combinations. Integration and inheri- tance of transgenes in T 0 and T 1 population of transgenic chickpea plants were determined by PCR, RT-PCR and Southern hybridization. Results of Southern hybridiza- tion showed single copy integration of cry1Ab or cry1Ac genes in most of the transgenic plants developed with either single or pyramided genes and reflected Mendelian inheritance of transgenes in T 1 progeny. Real time PCR of pyramided transgenic plants clearly showed differential expression of transcripts for both the genes driven by CaMV35S and Pcec promoters. Quan- titative assessment of Bt Cry toxins by ELISA of T 0 transgenic chickpea plants showed expression of toxin ranging from 5 to 40 ng mg -1 of total soluble protein (TSP) in leaves of transgenic plants. Insect bioassay performed with transgenic plants showed relatively higher toxicity for plants expressing Cry1Ac protein as compared to Cry1Ab to Helicoverpa armigera. Pyram- ided transgenic plants with moderate expression levels (15–20 ng mg -1 of TSP) showed high-level of resis- tance and protection against pod borer larvae of H. armigera as compared to high level expression of a single toxin. These results have shown the significance of pyramiding and co-expression of two Cry toxins for efficient protection against lepidopteran pests of chickpea. Keywords Bacillus thuringiensis Transgenic chickpea plants Genetic transformation Gene pyramiding Helicoverpa armigera Bt-Cry toxin Introduction Chickpea (Cicer arietinum L.) is second important grain legume grown worldwide as a major source of dietary protein for human consumption and feed for livestock. In spite of its large demand, the global yield Electronic supplementary material The online version of this article (doi:10.1007/s10681-011-0501-3) contains supplementary material, which is available to authorized users. M. Mehrotra A. K. Singh I. Sanyal D. V. Amla (&) Plant Transgenic Lab, National Botanical Research Institute, Rana Pratap Marg, P.O. Box 436, Lucknow 226 001, India e-mail: [email protected] I. Altosaar Department of Biochemistry, Microbiology and Immunology, University of Ottawa, 451 Smyth Road, Ottawa, ON, Canada 123 Euphytica (2011) 182:87–102 DOI 10.1007/s10681-011-0501-3
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Pyramiding of modified cry1Ab and cry1Ac genes of Bacillus thuringiensis in transgenic chickpea (...

Pyramiding of modified cry1Ab and cry1Ac genesof Bacillus thuringiensis in transgenic chickpea(Cicer arietinum L.) for improved resistance to pod borerinsect Helicoverpa armigera
Meenakshi Mehrotra • Aditya K. Singh •
Indraneel Sanyal • Illimar Altosaar • D. V. Amla
Received: 11 January 2010 / Accepted: 23 July 2011 / Published online: 11 August 2011
� Springer Science+Business Media B.V. 2011
Abstract The modified cry1Ab and cry1Ac insecti-
cidal genes of Bacillus thuringiensis (Bt) under the
control of two different constitutive promoters have
been introduced into chickpea (Cicer arietinum L.) by
Agrobacterium-mediated transformation of pre-condi-
tioned cotyledonary nodes. 118 stable transformed T0
plants as independent transformation events were
obtained expressing individual cry1Ab, cry1Ac or both
pyramided genes for their co-expression driven by either
cauliflower mosaic virus 35S promoter with duplicated
enhancer (CaMV35S) or synthetic constitutive promoter
(Pcec) and their combinations. Integration and inheri-
tance of transgenes in T0 and T1 population of transgenic
chickpea plants were determined by PCR, RT-PCR and
Southern hybridization. Results of Southern hybridiza-
tion showed single copy integration of cry1Ab or cry1Ac
genes in most of the transgenic plants developed with
either single or pyramided genes and reflected
Mendelian inheritance of transgenes in T1 progeny.
Real time PCR of pyramided transgenic plants clearly
showed differential expression of transcripts for both the
genes driven by CaMV35S and Pcec promoters. Quan-
titative assessment of Bt Cry toxins by ELISA of T0
transgenic chickpea plants showed expression of toxin
ranging from 5 to 40 ng mg-1 of total soluble protein
(TSP) in leaves of transgenic plants. Insect bioassay
performed with transgenic plants showed relatively
higher toxicity for plants expressing Cry1Ac protein as
compared to Cry1Ab to Helicoverpa armigera. Pyram-
ided transgenic plants with moderate expression levels
(15–20 ng mg-1 of TSP) showed high-level of resis-
tance and protection against pod borer larvae of
H. armigera as compared to high level expression of a
single toxin. These results have shown the significance
of pyramiding and co-expression of two Cry toxins for
efficient protection against lepidopteran pests of
chickpea.
Keywords Bacillus thuringiensis � Transgenic
chickpea plants � Genetic transformation � Gene
pyramiding � Helicoverpa armigera � Bt-Cry toxin
Introduction
Chickpea (Cicer arietinum L.) is second important
grain legume grown worldwide as a major source of
dietary protein for human consumption and feed for
livestock. In spite of its large demand, the global yield
Electronic supplementary material The online version ofthis article (doi:10.1007/s10681-011-0501-3) containssupplementary material, which is available to authorized users.
M. Mehrotra � A. K. Singh � I. Sanyal � D. V. Amla (&)
Plant Transgenic Lab, National Botanical Research
Institute, Rana Pratap Marg, P.O. Box 436,
Lucknow 226 001, India
e-mail: [email protected]
I. Altosaar
Department of Biochemistry, Microbiology and
Immunology, University of Ottawa, 451 Smyth Road,
Ottawa, ON, Canada
123
Euphytica (2011) 182:87–102
DOI 10.1007/s10681-011-0501-3

of chickpea and some other large seeded grain legumes
is stagnating for last two decades primarily due to large
number of biotic and abiotic stresses and slow progress
of genetic improvements for yield parameters (Pop-
elka et al. 2004; Dita et al. 2006). Grain yield of
chickpea suffers massive loss due to field infestation of
lepidopteran pod borer Helicoverpa armigera and few
other insects (Romeis et al. 2004). Development of
improved varieties of grain legumes particularly of
chickpea for resistance to pathogens and insect pests
by conventional breeding has been slow and difficult
due to narrow genetic base, limited genetic diversity
for these traits, barriers for sexual incompatibility and
high degree of autogamy (Ahmad et al. 1988; Van
Rheenen et al. 1993; Somers et al. 2003). Considering
the limitations of conventional approaches, introduc-
tion and expression of insecticidal delta endotoxin
genes of Bacillus thuringiensis into chickpea, is a
potential option for developing resistance against pod
borer lepidopteran insects as demonstrated success-
fully into a wide variety of crop plants (Christou et al.
2006; Gatehouse 2008; James 2009).
Since most of the insect resistant transgenic plants
released for commercial cultivation harbour single
insecticidal Bt-cry gene and the target insects pop-
ulation are consistently being exposed to the single
toxin protein, therefore the possibility of insects
evolving resistance to single Bt toxin is high (Zhao
et al. 2005; Gunning et al. 2005). In recent years,
several Bt cotton hybrid lines expressing cry1Ac have
been approved for commercial cultivation in India
and due to small farm holdings, diverse cropping
system and immigration of insects to alternative
hosts, the possibility of developing heterogeneous
insect population are very high (James 2009). More-
over, pink bollworm resistant to Bt cotton harbouring
the Bt-cry1Ac gene has been reported in the fields in
India, where farmer compliance with refugia strategy
has been low (Tabashnik et al. 2010). Several
strategies have been proposed for the management
of resistance development in herbivorous field insects
including the application of diverse mixture of toxins,
high expression of Bt-toxin, weedy refugia, hybrid
and pyramiding of different Bt-toxin genes and use of
sterile insect (Gatehouse 2008; Tabashnik et al.
2010). In recent years, transgenic plants expressing
two dissimilar insect toxins have been developed and
the most successful example is Bt-cotton ‘‘Bollgard
II’’ expressing cry1Ac and cry2Ab2 genes (Perlak
et al. 2001; Zhao et al. 2005). The efficacy and
sustainability of transgenic plants towards develop-
ment of resistance in insects rely on the pyramiding
and co-expression of two or more diverse transgenes
without affecting the yield parameters (Zhao et al.
2003; Gatehouse 2008).
In the present study, we have obtained stable
transgenic plants of recalcitrant grain legume chick-
pea using Agrobacterium-mediated transformation of
cotyledonary nodes (CN) with dicot-preferred mod-
ified truncated synthetic cry1Ab and cry1Ac genes.
Our results of insect bioassay have indicated that
transgenic plants co-expressing both the Bt-cry genes
were highly toxic and showed relatively higher and
effective protection against H. armigera as compared
to plants expressing single cry genes.
Materials and methods
Bt insecticidal crystal protein genes, plasmids
and Agrobacterium culture
Codon optimized 1,845 bp sequences of modified,
Bt-cry1Ab and cry1Ac genes (Sardana et al. 1996)
with a 38 bp AMV (Alfalfa mosaic virus) 50 UTR
sequence flanking on 50 of the gene were cloned in
binary vector pBIN20 (Hennegan and Danna 1998)
under the control of 620 bp cauliflower mosaic virus
35S double enhancer promoter (CaMV35S), and nos
terminator to generate binary vectors pBIN200 and
pRD400 respectively along with nptII gene cassette
for kanamycin resistance as plant selection marker in
T-DNA region (Fig. 1a, b). Sub-cloning of 2,753 bp
HindIII fragment from pBIN200 containing
CaMV35S-cry1Ab-nos cassette into pRD400 vector
resulted into pRD401 vector harbouring both cry1Ab
and cry1Ac genes driven by independent CaMV35S
promoter in cis orientation (Fig. 1d). The 450 bp
synthetic constitutive expression promoter (Pcec)
comprised of 138 bp minimal expression cassette
(Pmec) and 312 bp activation module (Sawant et al.
2001) with cry1Ac gene and nos terminator was
cloned at SalI and EcoRI site in pBIN20 vector to
generate pCEC.Ac vector where cry1Ac is driven by
Pcec promoter and carry the nptII gene as the
selectable marker (Fig. 1c). The 2,753 bp HindIII
cassette containing CaMV35S-cry1Ab-nos from
pBIN200 was sub-cloned in pCEC.Ac vector to
88 Euphytica (2011) 182:87–102
123

result into pCEC.Ab.Ac vector having cry1Ab and
cry1Ac gene driven by CaMV35S and Pcec promoter
respectively (Fig. 1e). The binary vectors harbouring
cry1Ab (pBIN200), cry1Ac (pRD400 and pCEC.Ac)
and cry1Ab ? cry1Ac (pRD401 and pCEC.Ab.Ac)
were mobilized into competent Agrobacterium tum-
efaciens strain LBA4404 (Hoekema et al. 1983) via
electroporation and utilized for transformation of
chickpea cotyledonary nodes.
Agrobacterium co-cultivation and transgenic plant
regeneration
Mature breeder seeds of chickpea variety P-362 were
obtained from Indian Agricultural Research Institute,
New Delhi and used routinely for genetic transfor-
mation. The seeds were surface sterilized and soaked
overnight in sterile distilled water supplemented with
2.0 mg l-1 BAP. The seeds were incubated on basal
medium consisting of MS salts (Murashige and
Skoog 1962), B5 vitamins, 30 g l-1 (w/v) sucrose,
solidified with 0.8% (w/v) agar and supplemented
with 2.0 mg l-1 BAP for germination to obtain
multiple shoots (Sanyal et al. 2003). Excised non-
transformed CN on MS medium resulted into 8–10
adventitious shoots per explant after 4 weeks of
incubation. The CN explants were prepared and
co-cultivated as described earlier (Sanyal et al. 2005).
Excised 21-day-old chickpea CNs were pre-condi-
tioned for 24 h on MS basal medium supplemented
with sodium thiosulphate (100 mg l-1) and DTT
(100 lM) before using them for Agrobacterium co-
cultivation. Culture of A. tumefaciens strain
LBA4404, harbouring the individual binary vectors
pBIN200, pRD400, pCEC.Ac, pRD401 and pCE-
C.Ab.Ac were used for genetic transformation of the
pre-conditioned chickpea CNs under optimized con-
ditions as described earlier (Sanyal et al. 2005). The
primary transgenic shoots recovered after selection
on kanamycin were either transferred to root induc-
tion medium, for root development within a week of
incubation or micro-grafted onto 10-days-old scion of
same genotype followed by acclimatization and
hardening under controlled conditions. Complete
plantlets were transferred to pots for seed setting in
contained transgenic growth chambers. All cultures
were incubated in culture room maintained at
24 ± 1�C under cool white light of intensity
60 lmol m-2 s-1 for 16 h photoperiod while rooted
chickpea plants in pots were grown in contained
Fig. 1 Schematic diagram of T-DNA regions of different
binary vectors used for genetic transformation of chickpea. The
backbone of binary vector for all the constructs is pBIN20
harbouring nptII gene driven by nos promoter (Pnos) and nosterminator (Tnos). a, b pRD400 and pBIN200 containing
cry1Ac and cry1Ab gene respectively driven by CaMV35Spromoter; c pCEC.Ac containing cry1Ac gene driven by
synthetic promoter Pcec; d pRD401containing both cry1Aband cry1Ac genes driven by CaMV35S; e pCEC.Ab.Ac
containing cry1Ab and cry1Ac genes driven by CaMV35Sand Pcec promoters respectively. Different PCR primers used
in the study are shown with arrow marks at their respective
binding positions: nptII-F, nptII-R for amplification of nptII;Ab-F, Ab-R for cry1Ab and Ac–F, Ac-R for cry1Ac genes
respectively. Bold lines below nptII, cry1Ab and cry1Ac genes
represent 678 bp PstI fragment of nptII, 1.8 kb BamHI and
EcoRI fragment of cry1Ab and 1.8 kb BamHI and SacI
fragment of cry1Ac respectively, used for the preparation of
radiolabelled probes for Southern analysis in the present study.
The abbreviations denote; RB right border, LB left border,
AMV Alfalfa mosaic virus 50 UTR sequence
Euphytica (2011) 182:87–102 89
123

glasshouse under similar conditions. Percent trans-
formation efficiency in absolute terms was calculated
as the number of primary transgenic plants (T0)
recovered after three successive selection cycles on
kanamycin divided by total number of adventitious
shoots that would have been recovered on shoot
induction medium from an equivalent number of non-
transformed explants under similar conditions. Selfed
seeds of the primary transformants were sown in
solidified half-strength MS medium supplemented
with 50 mg l-1 kanamycin. The plants germinated on
selection medium were considered as T1 progeny and
used for further analysis.
Polymerase chain reaction (PCR) for screening
and characterization of transgenic plants
Genomic DNA from kanamycin-resistant chickpea
shoots was isolated by homogenizing 100 mg of leaf
tissue in liquid nitrogen followed by addition of
extraction buffer containing 50 mM Tris–Cl, 1%
sarcosyl, 0.25 M sucrose, 50 mM NaCl, 20 mM
EDTA (pH 8.0). PCR assays were performed with
the genomic DNA of T0 primary transformants and T1
transgenic plants using Gene Amp 9700 thermocycler
(Perkin Elmer, USA). Set of specific primers for
cry1Ac, cry1Ab and nptII genes were designed to
amplify 995, 800 and 678 bp amplicons for respective
genes as shown in Table 1. The 25 ll PCR mixture
was prepared containing 100 ng plant genomic DNA,
100 lM dNTPs mix, 25 ng of each primer, 2 mM
MgSO4 and 1 U Taq DNA polymerase (NEB, USA).
Amplification was performed with initial denaturation
at 95�C for 5 min followed by 30 cycles, each
comprising of denaturation at 94�C for 90 s, annealing
at 58�C (cry1Ac) or at 55�C (cry1Ab) for 1 min and
extension at 68�C for 3 min followed by final exten-
sion for 5 min at 68�C. Amplified DNA fragments of
PCR assays were electrophoresed on 1% agarose gels,
visualized, documented and analyzed on Gel Doc XR
(Bio-Rad, USA).
Southern blot analysis
Southern blot hybridization analysis was performed to
confirm the integration of T-DNA into transformants
according to Sambrook and Russell (2001). Approx-
imately 10 lg genomic DNA from chickpea transgenic
plants was digested overnight with SalI for transgenic
plants developed with pBIN200, while EcoRI for
transgenic plants developed with pRD400 and pCE-
C.Ac vectors, that would restrict the T-DNA at one site.
HindIII restriction of genomic DNA was performed for
transgenic plants developed with pRD401 and pCE-
C.Ab.Ac vectors that would restrict the T-DNA at two
places generating one 2.2 kb fragment bearing cry1Ab
cassette and another fragment having cry1Ac gene with
flanking genomic DNA from the transgenic plants. The
digested genomic DNA samples were electrophoresed
on 0.8% (w/v) agarose gel and transferred onto Zeta
probe GT nylon membrane (Bio-Rad, USA). The blots
were hybridized at 58�C for 24 h with a[32P] dCTP
radiolabelled probe, comprising of full-length 1.8 kb
cry1Ac and 678 bp fragment of nptII gene. The blots
were then exposed to Fuji screen for 48 h and scanned
on Molecular Imager FX (Bio-Rad, USA).
RT-PCR and quantitative real-time PCR
RT-PCR was performed with *20 ng total RNA
isolated from 100 mg fresh leaves of independent
transgenic and non-transgenic control chickpea plants
Table 1 PCR primers used for each gene and the expected size of the PCR products
Target gene Forward primer (50–30) Reverse primer (50–30) Product size (bp)
Primers for PCR and RT-PCR
cry1Ab TGGTACAACACTGGCTTGGA ATGGGATTTGGGTGATTTGA 800
cry1Ac ATTCCTGGTGCAAATTGAGC CGATTCCGCTCTTTCTGTAA 995
nptII TATTCGGCTATGACTTGGGC GCGAACGCTATGTCCTGATA 678
Primers for quantitative real-time PCR, designed by using DNASTAR software (Lasergene Inc.)
cry1Ab AAGGATTCTCCCACAGGTTG ATGGGATTTGGGTGATTTGAG 157
cry1Ac TCAGGGTGTCTACAGAACCT CGGTTCCGCTCTTTCTGTAA 155
b-actin GCTGGATTTGCTGGAGATGATGA TCCATGTCATCCCAATTGCTAAC 194
90 Euphytica (2011) 182:87–102
123

using TRI reagent (Sigma, USA). Two-step RT-PCR
was performed for cry1Ab and cry1Ac genes with
specific set of primers (Table 1) using the enhanced
avian HS RT-PCR kit (Sigma, USA). The relative
quantity of cry1Ab and cry1Ac transcripts in transgenic
chickpea plants developed with different vectors were
analysed by real-time quantitative RT-PCR. The
reactions were carried out in StepOne real-time PCR
system (Applied Biosystems, USA) using Quantifast
SYBR green RT-PCR kit (Qiagen, Germany). Chick-
pea b-actin gene was used as endogenous control in all
real-time RT-PCR. Total RNA extracted from 100 mg
of leaf tissues was reverse transcribed into cDNA and
used as template in real-time PCR with cry1Ab, cry1Ac
and b-actin gene specific primers (Table 1). Reverse
transcription reaction was performed at 50�C for
10 min and initial denatruation at 95�C for 5 min for
activation of hot-start Taq polymerase followed by 40
amplification cycles each comprising 10 s denatur-
ation at 95�C and combined annealing and extension
for 30 s at 60�C in 25 ll reaction mixture according to
manufacturer’s instructions. The specificity and iden-
tity of the reaction products were verified by agarose
gel electrophoresis and melt curve analyses. The
relative values obtained from the quantitation of
mRNA were expressed as 2-DDCt where DCt repre-
sents the difference between Ct (cycle threshold)
values of a target and the endogenous control (b-actin)
in the same sample and DDCt is the difference between
the DCt value of a particular sample and that of the
reference sample. The quantitative data of real-time
RT-PCR represent mean values and standards errors of
three independent experiments with three replicates of
the transgenic plants developed with different vectors.
Enzyme-linked immunosorbent assay (ELISA)
for quantitative estimation of Bt-toxin
Quantitative estimation of insecticidal Cry1A endo-
toxin proteins expressed by cry1Ac and cry1Ab genes
in leaves of transgenic chickpea plants was performed
by double antibody sandwich ELISA using the Cry1Ac
monoclonal antibodies (Agdia, USA). Cell-free
extracts of leaves from transgenic and control untrans-
formed plants were added into the wells of ELISA plate
pre-coated with primary monoclonal antibody and
detection of Cry1Ab and Cry1Ac toxins was monitored
with peroxidase labelled PathoScreen kit for Cry1Ab/
Cry1Ac protein according to the manufacturer’s
instructions (Agdia, USA). The optical density of the
reaction was monitored at 650 nm using Spectra Max
340PC spectrophotometer (Molecular Devices, USA).
Expression levels of Bt-toxins in transgenic plants are
shown as ng mg-1 total soluble protein (ng mg-1 TSP)
and were quantified on a linear standard curve plotted
with pure Bt-Cry protein (Agdia, USA). Each trans-
genic plant was assayed twice with three replicates and
the data was statistically analyzed.
Insect bioassays
Entomocidal activity of Cry1Ab and Cry1Ac toxins
expressed in transgenic chickpea plants harbouring
either cry1Ab, cry1Ac or both the genes were assayed
through no-choice test on detached leaflets, by larval
feeding bioassay using second instar larvae of H. ar-
migera. Larvae of H. armigera were reared on artificial
diet enriched with gram flour (Gupta et al. 2004)
followed by feeding on castor leaves to complete their
life cycle and to obtain the adult moths for egg laying.
About 200–250 mg fresh leaves of transgenic plants
were placed in plastic petri dishes on moist filter paper
and 10 neonate larvae from lab-reared moths were
infested. Plates were sealed with Parafilm to prevent
desiccation and kept in the insect rearing room at
26 ± 1�C for 16 h photoperiod and 70% relative
humidity. Feeding was allowed for 4 days with one
change of fresh leaves on alternate days and data were
taken on larval weight, percentage survival and
mortality. Each treatment was respected with three
replicates and data were analyzed statistically.
Statistical analysis
Values reported are mean ± SE of three replicates
and minimum of 40–50 CN explants and 10 insect
larvae per replicate were used in each experiment.
The data were analyzed by Bonferroni t-test using
statistical package for social sciences (SPSS).
Results
Agrobacterium-mediated transformation
and recovery of transgenic chickpea plants
Co-cultivation of total 991 cotyledonary node explants
of chickpea variety Pusa-362 with A. tumefaciens
Euphytica (2011) 182:87–102 91
123
strain LBA4404 harbouring different binary vectors
pRD400, pBIN200, pCEC.Ac, pRD401 and pCEC.A-
b.Ac shown in Fig. 1 were carried out in three different
experiments. A total of 118 T0 transgenic plants were
recovered after three cycles of kanamycin selection
and actual numbers of transgenic plants developed
with different vectors are summarized in Table 2.
Transformation frequency achieved was ranging
between 1.69 and 2.77% on the basis of total adven-
titious shoots that would have been recovered from
same number of untransformed cotyledonary nodes
(Table 2). T0 transgenic chickpea plants were suc-
cessfully grafted and grown to maturity for collection
of T1 seeds for further analysis.
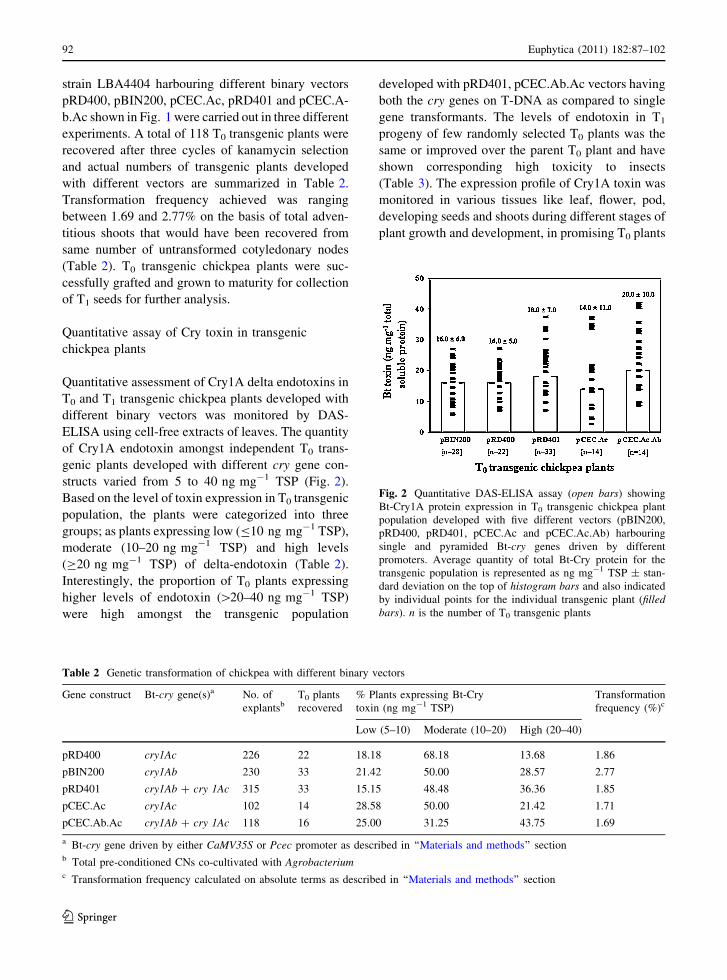
Quantitative assay of Cry toxin in transgenic
chickpea plants
Quantitative assessment of Cry1A delta endotoxins in
T0 and T1 transgenic chickpea plants developed with
different binary vectors was monitored by DAS-
ELISA using cell-free extracts of leaves. The quantity
of Cry1A endotoxin amongst independent T0 trans-
genic plants developed with different cry gene con-
structs varied from 5 to 40 ng mg-1 TSP (Fig. 2).
Based on the level of toxin expression in T0 transgenic
population, the plants were categorized into three
groups; as plants expressing low (B10 ng mg-1 TSP),
moderate (10–20 ng mg-1 TSP) and high levels
(C20 ng mg-1 TSP) of delta-endotoxin (Table 2).
Interestingly, the proportion of T0 plants expressing
higher levels of endotoxin ([20–40 ng mg-1 TSP)
were high amongst the transgenic population
developed with pRD401, pCEC.Ab.Ac vectors having
both the cry genes on T-DNA as compared to single
gene transformants. The levels of endotoxin in T1
progeny of few randomly selected T0 plants was the
same or improved over the parent T0 plant and have
shown corresponding high toxicity to insects
(Table 3). The expression profile of Cry1A toxin was
monitored in various tissues like leaf, flower, pod,
developing seeds and shoots during different stages of
plant growth and development, in promising T0 plants
Table 2 Genetic transformation of chickpea with different binary vectors
Gene construct Bt-cry gene(s)a No. of
explantsbT0 plants
recovered
% Plants expressing Bt-Cry
toxin (ng mg-1 TSP)
Transformation
frequency (%)c
Low (5–10) Moderate (10–20) High (20–40)
pRD400 cry1Ac 226 22 18.18 68.18 13.68 1.86
pBIN200 cry1Ab 230 33 21.42 50.00 28.57 2.77
pRD401 cry1Ab ? cry 1Ac 315 33 15.15 48.48 36.36 1.85
pCEC.Ac cry1Ac 102 14 28.58 50.00 21.42 1.71
pCEC.Ab.Ac cry1Ab ? cry 1Ac 118 16 25.00 31.25 43.75 1.69
a Bt-cry gene driven by either CaMV35S or Pcec promoter as described in ‘‘Materials and methods’’ sectionb Total pre-conditioned CNs co-cultivated with Agrobacteriumc Transformation frequency calculated on absolute terms as described in ‘‘Materials and methods’’ section
Fig. 2 Quantitative DAS-ELISA assay (open bars) showing
Bt-Cry1A protein expression in T0 transgenic chickpea plant
population developed with five different vectors (pBIN200,
pRD400, pRD401, pCEC.Ac and pCEC.Ac.Ab) harbouring
single and pyramided Bt-cry genes driven by different
promoters. Average quantity of total Bt-Cry protein for the
transgenic population is represented as ng mg-1 TSP ± stan-
dard deviation on the top of histogram bars and also indicated
by individual points for the individual transgenic plant (filledbars). n is the number of T0 transgenic plants
92 Euphytica (2011) 182:87–102
123

and in their T1 progeny which showed consistent level
of toxin till the stage of seed setting (data not shown).
Molecular characterization of transgenic chickpea
plants
Screening of putative T0 primary transformants
initially obtained after kanamycin selection was
performed through PCR assay for cry1Ab and cry1Ac
genes and Southern hybridization analyses and finally
118 T0 transgenic chickpea plants developed with
different vectors were selected. PCR analysis of the
transformants, using set of primers for amplification
of conserved internal sequences (amplicons) specific
to cry1Ac and cry1Ab showed amplification of
expected amplicons of 995 and 800 bp for cry1Ab
and cry1Ac genes respectively which were similar to
that of positive control DNA. The results of PCR
analysis of T0 transgenic plants developed with
different vectors have shown the amplification of
expected fragments for the corresponding Bt-cry
genes whereas, no such amplicon were observed in
non-transformed control chickpea plants under iden-
tical assay conditions (Fig. 3a–p). PCR analyses of
putative T0 transgenic plants after kanamycin selec-
tion showed that 98% of the plants were positive for
cry1Ab and cry1Ac genes. The results of PCR assay
of T0 transgenic plants developed with different
vectors for the corresponding gene(s) are shown in
Fig. 3.
The Southern blot analyses of genomic DNA from
independent transgenic plants (B1–B11 and B34–44)
developed with pBIN200 vector, digested with SalI
revealed hybridization of a single DNA fragment
ranging from 3.5 to 5.2 kb in comparison to 1.8 kb in
the positive control (Fig. 3a0, b0). Genomic DNA of
T0 transgenic plants (C1 to C22) developed with
pRD400 vector following digestion with EcoRI
showed distinct hybridization signals with single
fragment of DNA ranging between 3.6 and 4.4 kb
(Fig. 3c0, d0). Southern hybridization of seven ran-
domly selected independent T0 transgenic plants
PAc1 to 4, PAc7, 8 and 10 transformed with
pCEC.Ac vector harbouring cry1Ac gene driven by
constitutive synthetic promoter Pcec showed hybrid-
ization signals with single DNA fragment ranging
from 2.0 to 3.5 kb (Fig. 3l, m). Southern blots of
HindIII digested genomic DNA of T0 transgenic
plants developed with pRD401 and pCEC.Ab.Ac
vectors for pyramiding and co-expression of cry1Ab
and cry1Ac genes showed hybridization signals with
two fragments, one for cry1Ab of consistent size
*2.2 kb and second variable size fragment ranging
between 3.2 and 3.8 kb for cry1Ac gene (Fig. 3g, h, i,
p). Results of Southern hybridization showed inser-
tion of both the cry genes mostly as single copy
integration in transgenic plant pyramided for cry1Ab
and cry1Ac.
Transcript analysis by RT-PCR and quantitative
real-time PCR
Transcription of cry1Ab and cry1Ac genes in prom-
ising T0 transgenic chickpea plants selected on
kanamycin-supplemented medium was analyzed by
RT-PCR. Results of RT-PCR analysis of few ran-
domly selected T0 transgenic chickpea plants showed
the presence of desired transcript of 800 bp for
cry1Ab and 995 bp amplicon for cry1Ac respectively
with set of gene specific primers (Fig. 3q–v). Results
of RT-PCR assay of four T0 plants namely B1, B5,
B34 and B42 developed with vector pBIN200
showed amplification of expected transcript of
800 bp for cry1Ab whereas seven T0 plants devel-
oped with vector pRD400 designated as C3, C5, C7,
C11, C15, C16 and C20 and seven T0 plants
developed with vector pCEC.Ac namely PAc1–4,
PAc7, 8 and 10 showed amplification of expected
transcript of 995 bp for cry1Ac gene (Fig. 3r, u).
Results of RT-PCR performed with four T0 trans-
genic plants co-transformed with cry1Ac and cry1Ab
genes namely BC1, BC10, BC15 and BC20 devel-
oped with vector pRD401 and 11 plants designated as
pTBC1–11 developed with vector pCEC.Ab.Ac
showed the amplification of expected transcripts for
cry1Ac and cry1Ab genes (Fig. 3s, t, v). RT-PCR
results clearly depicted the formation of mRNA for
cry1Ac and cry1Ab genes and demonstrated the
expression of stable transcripts for both the genes in
transgenic chickpea plants transformed with both the
cry genes.
The relative levels of cry gene transcripts in the
leaves of promising transgenic chickpea plants
developed with different vectors such as pBIN200,
pRD400, pRD401, pCEC.Ac and pCEC.Ab.Ac was
analysed by real-time quantitative RT-PCR using
chickpea b-actin gene (accession No. EU 529707) as
endogenous control (Fig. 4). The relative values
Euphytica (2011) 182:87–102 93
123

Table 3 Expression of Bt-toxin in T0 and T1 chickpea transgenic plants and toxicity to H. armigera larvae
T0 plant (cry toxin
ng mg-1 TSP)aBt-gene
(construct)
Damage to
plant (%)bInsect
mortality
(%)b
T1 progeny
developed
Bt toxin expression
(ng mg-1 TSP)
Damage to
plant (%)
Insect
mortality
(%)
Control – 95–100 0.00 – – – –
B2 (22 ± 1) cry1Ab(pBIN200)
56–62 44 ± 2 B2-1 20 ± 1 58–64 40 ± 2
B2-2 23 ± 2 54–58 45 ± 2
B2-3 24 ± 2 54–56 46 ± 2
B2-4 19 ± 1 58–64 40 ± 2
B10 (17 ± 1) cry1Ab(pBIN200)
65–70 34 ± 2 B10-1 18 ± 1 65–70 36 ± 2
B10-2 20 ± 2 60–64 34 ± 2
B10-3 20 ± 1 60–64 34 ± 2
B10-4 16 ± 1 66–72 36 ± 2
C4 (23 ± 1) cry1Ac(pRD400)
22–26 84 ± 2 C4-1 25 ± 2 20–22 88 ± 2
C4-2 24 ± 1 22–24 86 ± 2
C4-3 24 ± 2 22–24 86 ± 2
C6 (29 ± 1) cry1Ac(pRD400)
18–24 89 ± 2 C6-1 30 ± 1 17–19 93 ± 2
C6-2 32 ± 2 15–17 96 ± 2
C6-3 28 ± 1 18–20 92 ± 2
C17 (16 ± 1) cry1Ac(pRD400)
28–32 70 ± 2 C17-1 16 ± 1 28–30 76 ± 2
C17-2 19 ± 2 24–26 78 ± 2
C17-3 18 ± 1 24–27 78 ± 2
PAc3 (16 ± 2) cry1Ac(pCEC.Ac)
60–65 30 ± 2 PAc3-1 20 ± 1 70–75 50 ± 1
PAc3-2 18 ± 2 60–65 40 ± 1
PAc3-3 14 ± 1 45–50 28 ± 2
BC1 (15 ± 1) cry1Ab ? Ac(pRD401)
26–30 81 ± 2 BC1-1 16 ± 1 26–28 86 ± 2
BC1-2 15 ± 1 26–29 86 ± 2
BC1-3 17 ± 1 25–28 90 ± 2
BC1-4 16 ± 1 26–30 88 ± 2
BC6 (36 ± 1) cry1Ab ? Ac(pRD401)
16–20 97 ± 2 BC6-1 35 ± 2 16–18 99 ± 1
BC6-2 39 ± 2 10–12 100.0
BC6-3 40 ± 2 10–14 98 ± 1
BC6-4 26 ± 1 12–14 98 ± 1
BC6-5 29 ± 2 18–20 98 ± 1
BC18 (26 ± 1) cry1Ab ? Ac(pRD401)
20–22 94 ± 2 BC18-1 28 ± 2 18–21 98 ± 1
BC18-2 26 ± 1 20–23 96 ± 1
BC18-3 26 ± 1 16–18 97 ± 1
BC18-4 24 ± 1 16–18 96 ± 1
TBC2 (18 ± 1) cry1Ab ? Ac(pCEC.Ab.Ac)
20–24 84 ± 2 TBC2-1 19 ± 1 18–20 92 ± 2
TBC2-2 20 ± 1 17–19 89 ± 2
TBC2-3 18 ± 1 20–22 91 ± 2
TBC10 (28 ± 1) cry1Ab ? Ac(pCEC.Ab.Ac)
16–20 95 ± 2 TBC10-1 30 ± 2 16–18 98 ± 1
TBC10-2 28 ± 1 15–18 98 ± 1
TBC10-3 31 ± 2 16–19 98 ± 1
a Determined by DAS-ELISA assayb Determined by insect mortality bioassay
94 Euphytica (2011) 182:87–102
123

obtained for transcript quantitation is expressed as
2-DDCt where DCt represents the differences between
the Ct values of Cry protein transcript and b-actin
among different transgenic plants and DDCt is the
difference between the DCt values in reference to that
of pRD400. Results of comparative qRT-PCR of 11
independent transgenic chickpea plants transformed
with different vectors showed almost 2.5-fold higher
transcript formation for both cry1Ab and cry1Ac
genes in transgenic plants developed either with
pRD401 or pCEC.Ab.Ac harbouring both the genes
driven by CaMV35S promoter or combination of
CaMV35S and Pcec for cry1Ab and cry1Ac respec-
tively (Fig. 4). These results clearly showed that
pyramiding of two genes with either similar or
different promoters has increased the transcript
formation and stability as compared to transcript in
transgenic plant developed with single gene con-
structs. The relative quantitation of the cry gene
transcripts among the T0 transgenic chickpea plants
transformed with single cry gene showed enhanced
level of transcript for cry1Ac gene driven either by
CaMV35S or Pcec promoter as compared to that of
cry1Ab gene driven by CaMV35S promoter. The
quantitative transcript in transgenic plants co-trans-
formed with both the cry genes have shown corre-
sponding increase in the transcript levels of both the
genes driven either by CaMV35S, Pcec or combina-
tion of both the promoters (Fig. 4). This difference in
transcript levels among transgenic plants could be
attributed to independent event for gene integration
site rather than the type of promoter.
Inheritance analysis of cry genes in T1 progeny
T0 transgenic chickpea plants showed normal flow-
ering pattern, except that the number of flowers were
less which could be due to physiological conditions
from culture room to contained glasshouse. Number
of pods and seeds in T0 transgenic plants was very
low as compared to tissue culture raised and field
grown untransformed plants (data not shown). The
inheritance pattern of cry1A and nptII genes in the T1
progeny of primary transformants was analyzed by
germinating the seeds on kanamycin-supplemented
medium (100 mg l-1). Antibiotic screening of T1
seeds followed by PCR analysis revealed segregation
according to Mendelian ratio 3:1 (resistant: suscep-
tible, P B 0.05, v2 = 3.841) for kanamycin tolerance
(Table 3). PCR and Southern analysis of the genomic
DNA from T1 progenies of selected T0 plants showed
amplification of expected amplicons of 800 and
995 bp for cry1Ab and cry1Ac genes respectively
similar to the positive controls (Fig. 5a–c). Result of
Southern blot hybridization showed single DNA
fragments of size ranging from 4.16 to 6.57 kb
hybridizing with 1.845 kb radiolabelled cry1Ab/
c probe in T1 transgenic plants developed with
pBIN200 and pRD400 vectors harbouring individual
cry1Ab or cry1Ac gene respectively and DNA
fragments of 2.82 to 8.62 kb in transgenic plants
developed with pRD401vector for co-expressing two
cry genes. Results of Southern hybridization also
showed insertion of cry1Ab and cry1Ac genes as
single copy integrations (Fig. 5a0–c0).
Insect bioassay of transgenic plants
The T0 and T1 chickpea transgenic plants expressing
moderate to high levels of Bt-toxins (Cry1Ab and
Cry1Ac) were evaluated for entomocidal activity by
insect feeding bioassays performed with second instar
larvae of H. armigera. Leaves from 30 days old T0 and
T1 transgenic chickpea plants were fed to the larvae
and response of their feeding on weight gain, life cycle
and mortality of insect was monitored. Data of insect
bioassay on leaves of transgenic plants expressing
Cry1Ac toxin (^20 ng mg-1 TSP) showed relatively
restricted damages to leaves while caused significant
weight loss and higher mortality to larvae compared to
the plants expressing similar levels of Cry1Ab toxin
(Table 3). Larvae challenged on leaves of trans-
genic plants with higher levels of Cry1Ac toxin
([20–40 ng mg-1 TSP) showed severely retarded
growth after 2 days of feeding and significantly high
rate of mortality ([90%) as compared to leaves with
Cry1Ab toxin. The pyramided transgenic plants co-
expressing both the toxins at moderate levels
(10–20 ng mg-1 TSP) showed 80–90% mortality of
insects after 4 days of feeding and leaves of these
plants suffered very little feeding damage (Table 3;
Fig. 6) whereas, transgenic chickpea plants co-
expressing higher levels of Cry1Ab and Cry1Ac
toxins ([20 ng mg-1 TSP) showed [90–95% mor-
tality in the challenged insects after 48 h of feeding
and very little damage to leaves. Bioassays performed
with T1 progenies of few promising T0 transgenic
plants reflected similar response to insect protection
Euphytica (2011) 182:87–102 95
123

96 Euphytica (2011) 182:87–102
123

corresponding to the level of expressed toxin(s). The
ratio of plants expressing higher levels of Bt-toxin was
increased in segregating population as per Mendelian
pattern for single dominant character. The larvae
exposed on non-transformed control plants continued
to feed and showed normal development of larvae with
average length of individual larvae up to 0.6 ± 0.1 cm
and body weight of *40 mg after 4 days of feeding
(Fig. 6). Insecticidal response and protection to insect
damages was relatively higher in transgenic chickpea
plants expressing Cry1Ac than Cry1Ab with equal
levels of toxins whereas transgenic plants co-express-
ing moderate levels of both the toxins (*20 ng mg-1
TSP) showed significant protection to insect damage
and mortality of H. armigera (Table 3).
Discussion
Transgenic crop plants expressing different insecti-
cidal proteins from B. thuringiensis have shown
significant resistance to important agricultural insect
pests in addition to reduced application of synthetic
pesticides and improved yield (Perlak et al. 2001;
James 2009). However, there is always a risk that
field insects could develop resistance to these toxins
after prolonged and consistent exposure (Bates et al.
2005). Adaptation of insect resistance to insecticidal
toxins of B. thuringiensis can reduce the efficacy of
toxins and thereby the benefits of the technology.
The predominant strategy for delaying pest resistance
to Bt crops requires either refuges of non-Bt host
plants to provide susceptible insects to mate with
resistant insects or the sufficient expression of
insecticidal toxins for efficient mortality of insects
(Tabashnik et al. 2010). The most practical
approaches for sustained efficacy of Bt-transgenics
are the expression of Bt-endotoxin at high level and
weedy refugia or stacking of two or more different
cry genes into the same plant (Zhao et al. 2005; Cao
et al. 2008). Cry1Ab and Cry1Ac are two most
effective larvicidal toxins against large number of
lepidopteran insects including H. armigera, infecting
several economically important crop plants in the
Fig. 4 Relative quantitation of cry1Ab (gray bar) and cry1Ac(black bar) transcripts in T0 transformed chickpea plantlets by
real time PCR assay using Quantifast SYBR green RT-PCR
kit. C4 and C6 plants were developed with pRD 400 vector
expressing cry1Ac driven by CaMV35S promoter; pAc.3 and
pAc.10 plants were transformed with pCEC.Ac vector express-
ing cry1Ac driven by Pcec promoter; B2 and B6 plants are
developed with pBIN 200 vector expressing cry1Ab driven by
CaMV35S promoter; BC1 and BC6 plants are developed with
pRD 401 vector showing transcript formation for both cry1Aband cry1Ac genes driven by CaMV35S promoter while TBC2,
TBC 10 and TBC 11 plants are developed with pCEC.Ab.Ac
vector showing transcript formation for both cry1Ab and
cry1Ac genes driven by CaMV35S and Pcec promoters
respectively. b-actin gene of chickpea served as the internal
control
Fig. 3 Molecular characterization of T0 transgenic chickpea
plants developed with different gene constructs by PCR,
Southern and RT-PCR analyses. a, b PCR amplification of
800 bp fragment of cry1Ab gene of transgenic plants developed
with pBIN200 vector and a0, b0 Southern blotting of 22
randomly selected plants following digestion of their genomic
DNA with SalI, while lane -C and ?C are DNA from non-
transformed control plant and plasmid as positive control
respectively; c, d PCR amplification of 995 bp for cry1Ac gene
of transgenic plants developed with pRD400 vector and c0, d0
the corresponding Southern blotting after digesting the
genomic DNA with EcoRI; e, e0 and f, f0 PCR amplification
of 995 bp for cry1Ac gene and 800 bp for cry1Ab gene
respectively of transgenic plants developed with pRD401
vector and g, h, i Southern blotting of T0 transgenic chickpea
plants and the genomic DNA was digested with HindIII;
j, k PCR amplification of 995 bp for cry1Ac and 678 bp for
nptII genes in T0 transgenic plants developed with pCEC.Ac
vector; l, m Southern blotting of T0 transgenic chickpea plants
digested with EcoRI for cry1Ac and nptII genes respectively; n,
o PCR amplification of 995 bp for cry1Ac and 800 bp for
cry1Ab gene respectively of transgenic chickpea plants
developed with pCEC.Ab.Ac vector; p Southern blotting of
T0 transgenic plants and their genomic DNA was digested with
HindIII; q–v RT-PCR analyses of few randomly selected T0
transgenic plants; q for cry1Ab gene (pBIN 200); r for cry1Acgene (pRD 400); s for cry1Ab and cry1Ac genes (pRD 401);
t for cry1Ac gene (pCEC.Ac); u, v for cry1Ac and cry1Abgenes with plants transformed with pCEC.Ab.Ac vector
respectively
b
Euphytica (2011) 182:87–102 97
123

field (Cheng et al. 1998; Homrich et al. 2008). In the
present study a recalcitrant grain legume chickpea
has been transformed with modified synthetic cry1Ab
and cry1Ac genes individually as well as pyramided
(co-expression) to assess the synergistic effect of both
the genes for toxicity to susceptible insects including
pod borer. Stacking of both the modified truncated
cry1Ab and cry1Ac genes in chickpea was performed
by Agrobacterium-mediated transformation of coty-
ledonary node explant with vectors having both the
genes in one replicon. Transformation efficiency
ranging from 1.69 to 2.77% was observed for co-
transformation of both the cry genes driven by
CaMV35S promoter or by two different constitutive
promoters. The observed transformation frequencies
for two Bt-genes are comparable to that observed for
other recalcitrant plants like cotton, rice and mustard
(Perlak et al. 2001; Riaz et al. 2006; Cao et al. 2008).
Results obtained from PCR, Southern blotting and
RT-PCR analyses of transgenic chickpea plants have
clearly confirmed the stable single copy integration
without any rearrangements of cry1Ab and cry1Ac
genes in transgenic plants and also into subsequent T1
generation. The different sizes of hybridizing genomic
DNA fragments of transgenic plants with the probes
indicated that they resulted from the independent stable
T-DNA integration event into the chickpea genome
and not from endophytic Agrobacterium contamina-
tion. However, presence of the expected hybridization
signals with genomic DNA fragments ([4.5 kb) in
majority of the transformed plants with two genes
showed that the probed genes cry1Ab and cry1Ac
Fig. 5 Molecular
characterization of T1
transgenic chickpea
progenies developed with
different vectors. a, a0 PCR
amplification of cry1Abgene and corresponding
Southern blots of plants
transformed with pBIN 200
vector; b, b0 T1 plants
transformed with pRD 400
vector and c, c0 for T1 plants
transformed with pRD 401
vector harbouring both
cry1Ab and cry1Ac genes
Fig. 6 Insect bioassay performed on Bt-transgenic chickpea
plants with 2nd instar larvae of Helicoverpa armigera.
a Control untransformed chickpea plants challenged with pod
borer larvae resulted into extensive consumption of leaves and
normal growth of the larvae. b Leaf of transgenic plant
expressing low levels of Bt-Cry toxin (\10 ng mg-1 TSP).
c Leaves of transgenic plant expressing moderate levels of Bt-
Cry toxin (^10–20 ng mg-1 TSP). d Leaves of transgenic
plant expressing higher levels of Bt-Cry toxin ([20 ng mg-1
TSP) surrounded by dead larvae
98 Euphytica (2011) 182:87–102
123

remained intact when integrated into the chickpea
genome. The T0 population of primary transgenic
chickpea plants have reflected independent pattern of
transgene expression due to complex and random
integration of foreign genes in the host genome
following Agrobacterium-mediated transformation.
Therefore, inheritance of foreign genes in transgenic
plants may display complex patterns for single as well
as two genes. Results obtained from Southern blot and
RT-PCR analyses have clearly confirmed the stable
integration and segregation of cry1Ab and cry1Ac
genes in T1 generation.
Quantitative analysis of transcript expression in
transgenic chickpea plants with cry1Ab ? cry1Ac
genes, independently driven by the constitutive
CaMV35S promoter, showed simultaneous co-expres-
sion of both the transcripts as found in transgenic
chickpea plants expressing cry1Ab and cry1Ac driven
by Pcec and CaMV35S promoters respectively. These
results override the assumptions of co-suppression due
to multiple copy integration of same promoter
sequences on single T-DNA associated with low level
of transgene expression (Wang and Waterhouse 2002).
The silencing of transgene expression, however, still
seems to be a puzzling issue and dependent on several
post-transcriptional factors including copy number of
transgene(s) and position effect of T-DNA insertion
particularly in recalcitrant plant species (Olhoft et al.
2004; Butaye et al. 2005). In order to avoid possible
silencing of cry gene(s) expression in chickpea trans-
genics, co-transformed with two genes, due to co-
suppression phenomenon of copy number of the same
promoter in expression vector, we considered the
option of using Pcec a different constitutive promoter
which is earlier reported for higher expression of gusA
and gfp reporter genes compared to CaMV35S in
tobacco transgenics (Sawant et al. 2001; Chaturvedi
et al. 2006). The CaMV35S promoter contains two
major enhancer domains and five sub-domains that
synergistically confer developmental and tissue-spe-
cific expression (Benfey and Chua 1990). Whereas
multifactorial expression cassette Pcec has been
designed and developed by deploying several cis
elements and conserved motifs commonly present in
promoter region of highly expressed plant genes. The
synergistic interaction of different motifs and archi-
tecture of Pcec seems to interact better with wide range
of native gene enhancer sequences for higher expres-
sion of transgenes compared to other natural promoters
(Sawant et al. 2001, 2005). Hence, CaMV35S and Pcec
were used to develop insect resistant transgenic plants
expressing cry1Ab or cry1Ac genes alone or in
combination within the same vector for increasing
the possibility of obtaining promising transgenic
events expressing very high levels of Bt-toxin(s). Our
results of ELISA for Bt-Cry toxin in T0 transgenic
population developed with either of the cry gene or
combination of promoters have revealed variation in
cry1A gene expression in T0 population, which
reflected corresponding level of resistance to insects.
These results are consistent with earlier observations
for differential expression of Bt-endotoxins in T0
plants (Perlak et al. 1991; Nayak et al. 1997; Husnain
et al. 2002). This variation in expression of heterolo-
gous protein like Bt-endotoxin amongst population of
primary T0 transgenics may be attributed to position
effect of gene integration, flanking sequences, chro-
matin context of the locus, increase in DNA methyl-
ation with increase in plant age and physiological
changes of the foreign protein in the plant tissues
(Peach and Velten 1991; Down et al. 2001; Husnain
et al. 2002).
The maximum co-expression of Cry1Ab and
Cry1Ac toxins in transgenic chickpea was up to
40 ng mg-1 TSP, irrespective of using identical or
two different promoters. The expression level of Bt-
toxins in the present study was higher than earlier
reports for other large-seeded grain legumes (Parrott
et al. 1994; Stewart et al. 1996; Sanyal et al. 2005)
except for transgenic peanut plants where expression
of Cry1A(c) protein up to 0.18% of TSP has been
documented (Singsit et al. 1997). High-level expres-
sion of Cry protein in transgenic chickpea co-
expressing both the genes may be attributed to some
or all the possible options such as specific modifica-
tions and codon optimization of the cry genes for
dicot plants, use of constitutive promoters, simulta-
neous co-expression of both the genes, AMV 50 UTR
element, optimum translation initiation and to the
integration of the transgene into transcriptionally
active region of the host genome (Sardana et al. 1996;
Cheng et al. 1998). The modified cry1Ab and cry1Ac
genes were designed for improved mRNA stability,
codon optimization for dicots, removal of polyade-
nylation sites, splicing sequences and optimization of
ATG consensus flanking nucleotides for proper
translation initiation (Perlak et al. 1991; Sardana
et al. 1996). The modified Bt-cry genes have been
Euphytica (2011) 182:87–102 99
123

extensively used for genetic transformation and
expression of insect-resistant trait in several plant
species (Ferry et al. 2004; Riaz et al. 2006). However,
the efficiency of Agrobacterium-mediated transfor-
mation and recovery of transgenics of grain legumes
are restricted due to several inherent limitations
including their complex genome (Eapen 2008; Dita
et al. 2006). To achieve higher expression of Cry
toxins we have used the multifactorial synthetic
expression cassette Pcec along with CaMV35S. The
designing of Pcec has several positive regulatory
elements and conserved motifs and has shown several
folds increased expression of gusA transgene in
tobacco and cotton compared to CaMV35S (Sawant
et al. 2001, 2005). Although we could not recover the
transgenic event of chickpea expressing very high-
level of Bt toxins from modified cry genes driven
either by CaMV35S, Pcec or their combinations but
we have obtained relatively high proportion of T0
transgenic plants with increased level of Cry1A
toxins ([20 ng mg -1 soluble protein) at population
level with combination of these promoters. The
overall performance of the synthetic expression
module Pcec in chickpea is comparable to CaMV35S
and our results are similar to earlier reports where
transcription factors binding to known sequence
motifs resulted in higher level of transgene expres-
sion (Rushton et al. 2002; Bhullar et al. 2003). The
wide range of cry gene expression in T0 transgenic
population of chickpea developed with different gene
constructs may be due to integration of the transgenes
at different positions in the genome of independently
selected event plants (Peach and Velten 1991; Perlak
et al. 2001). However, we could not obtain T0
transgenic chickpea event expressing higher Cry
toxin as ([1.0% of TSP) reported earlier for trans-
genic tobacco, maize, rice, cotton and tomato (Perlak
et al. 2001; James 2009). This suggests screening of a
larger population of primary transformants for
obtaining an event with higher expression of Bt-
toxin in recalcitrant plant species particularly the
grain legumes (Somers et al. 2003).
Entomocidal analysis and assessment of toxicity to
H. armigera have demonstrated that transgenic
chickpea plants co-expressing both the genes have
reflected higher toxicity and protection to insects as
compared to plants expressing single cry1Ab or
cry1Ac gene at relatively higher levels. Transgenic
chickpea plants harbouring both the genes have
reflected significant mortality ([95%) to second
instar larvae of H. armigera at toxin levels of
15 ng mg-1 TSP while plants expressing cry1Ac
gene could result similar toxicity at much higher
levels of expression ([22–25 ng mg-1 TSP). The
combined expression of Cry1Ab and Cry1Ac toxins
not only preserving the effectiveness of Bt-transgen-
ics for delaying the evolution of resistance but also
develop a more effective plant protection strategy
against the major Lepidopteran insect pests due to
combined synergistic action of two toxins, as
reported earlier (Zhao et al. 2005; Cao et al. 2008).
It is suggested that Cry1Ab and Cry1Ac bind to a
common receptor in H. armigera, but use different
epitopes and vary in their binding affinity to BBMV of
target insect, therefore co-expression of both the toxins
in same plant is complementary to enhance the
protection against H. armigera and delaying the
development of resistance in heterogenous field pop-
ulation of lepidopteran insects (Ferre and Van Rie
2002; Estela et al. 2004; James 2009). Therefore,
synergetic response of Cry1Ab and Cry1Ac toxins in
transgenic chickpea has reflected better and sustain-
able protection even at moderate level of expression
against the pod borer H. armigera and expected to
provide enhanced protection against several lepidop-
teran insects attacking the immature pods and foliage
of chickpea as reported earlier for pyramiding of
different Bt-cry genes (Maqbool et al. 2001; Zhao et al.
2003). In summary, we have reported the development
of stable transgenic plants of recalcitrant grain legume
chickpea expressing pyramided modified cry1Ab and
1Ac genes that are segregating in Mendelian fashion.
These transgenic plants have shown relatively better
and effective protection against H. armigera and
potential to delay the development of resistance in
target insect population more effectively than the
plants expressing the single cry gene.
Acknowledgments The authors are grateful to Director,
National Botanical Research Institute, Lucknow, India for
laboratory facilities. Thanks are due to Dr. Samir Sawant,
Scientist, National Botanical Research Institute, Lucknow,
India for providing the synthetic constitutive promoter Pcecand Prof. I. Altosaar, University of Ottawa, Canada for
providing modified synthetic truncated Bt-cry1Ab and cry1Acgenes. We thank Mr.S. M. H.Abidi for insect rearing and
bioassay studies. We acknowledge Council of Scientific and
Industrial Research, New Delhi for financial support and
fellowships to MM and AKS. This work was carried out under
the CSIR Network Project NWP003.
100 Euphytica (2011) 182:87–102
123

References
Ahmad F, Slinkard AE, Scoles GJ (1988) Investigations into
the barrier(s) to inter-specific hybridization between Cicerarietinum L. and eight other annual Cicer species. Plant
Breed 100:193–198
Bates SL, Zhao J, Roush RT, Shelton AM (2005) Insect
resistance management in GM crops: past, present and
future. Nat Biotech 23:57–62
Benfey PN, Chua N-H (1990) The cauliflower mosaic virus
35S promoter: combinatorial regulation of transcription in
plants. Science 250:959–966
Bhullar S, Chakravarthy S, Advani S, Datta S, Pental D, Burma
PK (2003) Strategies for development of functionally
equivalent promoters with minimum sequence homology
for transgene expression in plants: cis-elements in a novel
DNA context versus domain swapping. Plant Physiol
132:988–998
Butaye KMJ, Cammue BPA, Delaure SL, DeBolle MFC
(2005) Approaches to minimize variation of transgene
expression in plants. Mol Breed 16:79–91
Cao J, Shelton AM, Earle ED (2008) Sequential transformation
to pyramid two Bt genes in vegetable Indian mustard
(Brassica juncea L.) and its potential for control of dia-
mondback moth larvae. Plant Cell Rep 27:479–487
Chaturvedi CP, Sawant SV, Kiran K, Mehrotra R, Lodhi N,
Ansari SA, Tuli R (2006) Analysis of polarity in the
expression from a multifactorial bidirectional promoter
designed for high-level expression of transgenes in plants.
J Biotech 123:1–12
Cheng X, Sardana R, Kaplan H, Altosaar I (1998) Agro-bacterium-transformed rice plants expressing synthetic
cry1Ab and cry1Ac gene are highly toxic to striped stem
borer and yellow stem borer. Proc Natl Acad Sci USA
95:2767–2772
Christou P, Capell T, Kohli A, Gatehouse JA, Gatehouse AMR
(2006) Recent developments and future prospects in insect
pest control in transgenic crops. Trends Plant Sci 11:302–308
Dita MA, Rispail N, Prats E, Rubiales D, Singh KB (2006)
Biotechnology approaches to overcome biotic and abiotic
stress constraints in legumes. Euphytica 147:1–24
Down RE, Ford L, Bedford SJ, Gatehouse LN, Newell C,
Gatehouse JA, Gatehouse AMR (2001) Influence of plant
development and environment on transgene expression in
potato and consequences for insect resistance. Transgenic
Res 10:223–236
Eapen S (2008) Advances in development of transgenic pulse
crops. Biotech Adv 26:162–168
Estela A, Eschriche B, Ferre J (2004) Interaction of Bacillusthuringiensis toxins with larval midgut binding sites of
Helicoverpa armigera (Lepidoptera:Noctuidae). Appl
Environ Microbiol 70:1378–1384
Ferre J, Van Rie J (2002) Biochemistry and genetics of insect
resistance to Bacillus thuringiensis. Ann Rev Entomol
47:501–533
Ferry N, Edwards MG, Mulligan EA, Emami K, Petrova AS,
Frantescu M, Davison GM, Gatehouse AMR (2004)
Engineering resistance to insect pests. In: Christou P, Klee
H (eds) Handbook of plant biotechnology. John Wiley and
Sons Ltd, New York, pp 373–394
Gatehouse JA (2008) Biotechnological prospects for engi-
neering insect-resistant plants. Plant Physiol 146:881–887
Gunning RV, Dang HT, Kemp FC, Nicholson IC, Moores GD
(2005) New resistance mechanism in Helicoverpa armi-gera threatens transgenic crops expressing Bacillus thur-ingiensis Cry1Ac toxin. Appl Environ Microbiol
71:2558–2563
Gupta GP, Birah A, Ravi S (2004) Development of artificial
diet for mass rearing of American bollworms (Heli-coverpa armigera). Indian J Agri Sci 74:548–557
Hennegan KP, Danna KJ (1998) pBIN20: an improved binary
vector for Agrobacterium-mediated transformation. Plant
Mol Biol Rep 16:129–131
Hoekema A, Hirsch PR, Hooykaas PJJ, Schilperoort RA (1983)
A binary plant vector strategy based on separation of vir-
and T-region of the Agrobacterium tumefaciens Ti-plas-
mid. Nature 303:179–180
Homrich MS, Passaglia LMP, Pereira JF et al (2008) Resis-
tance to Anticarsia gemmatalis Hubner (Lepidoptera,
Noctuidae) in transgenic soybean [Glycine max (L.)
Merrill Fabales, Fabaceae] cultivar IAS5 expressing a
modified Cry1Ac endotoxin. Genet Mol Biol 31:522–531
Husnain T, Jan A, Maqbool SB, Datta SK, Riazuddin S (2002)
Variability in expression of insecticidal cry1Ab gene in
Indica basmati rice. Euphytica 128:121–128
James C (2009) Global status of commercialized biotech/GM
crops: briefs 41. ISAAA publication, Ithaca, New York
Maqbool SB, Riazuddin S, Loc NT, Gatehouse AMR, Gate-
house JA, Christou P (2001) Expression of multiple
insecticidal genes confers broad resistance against a range
of different rice pests. Mol Breed 7:85–93
Murashige T, Skoog F (1962) A revised medium for rapid
growth and bioassay with tobacco tissue cultures. Physiol
Plant 15:473–479
Nayak P, Basu D, Das S, Basu A, Ghosh D, Ramakrishnan NA,
Ghosh M, Sen SK (1997) Transgenic elite indica rice
plants expressing Cry1Ac d-endotoxin of Bacillus thur-ingiensis are resistant against yellow stem borer (Scrip-ophaga incertulas). Proc Natl Acad Sci USA 94:
2111–2116
Olhoft PM, Flagel LE, Somers DA (2004) T-DNA locus
structure in a large population of soybean plants trans-
formed using the Agrobacterium-mediated cotyledonary-
node method. Plant Biotech J 2:289–300
Parrott WA, All JN, Adang MJ, Bailey MA, Boerma HR,
Stewart CN Jr (1994) Recovery and evaluation of soybean
plants transgenic for a Bacillus thuringiensis var. kurstaki.insecticidal gene. In Vitro Cell Dev Biol Plant 30:
144–149
Peach C, Velten J (1991) Transgene expression variability
(position effect) of CAT and GUS reporter genes driven
by linked divergent T-DNA promoters. Plant Mol Biol
17:49–60
Perlak FJ, Fuchs RL, Dean DA, McPherson SL, Fischoff DA
(1991) Modification of the coding sequence enhances
plant expression of insect control protein genes. Proc Natl
Acad Sci USA 88:3324–3328
Perlak FJ, Oppenhuizen M, Gustafson K, Voth K, Sivasupr-
amaniam S, Heering D, Carey B, Ihrig RA, Roberts JK
(2001) Development and commercial use of Bollgard
Euphytica (2011) 182:87–102 101
123

cotton in the USA-early promises versus today’s reality.
Plant J 27:489–501
Popelka JC, Terryn N, Higgins TJV (2004) Gene technology
for grain legumes: can it contribute to the food challenge
in developing countries. Plant Sci 167:195–206
Riaz N, Husnain T, Fatima T, Makhdoom R, Bashir K, Masson
L, Altosaar I, Riazuddin S (2006) Development of indica
basmati rice harboring two insecticidal genes for sus-
tainable resistance against lepidopteran insects. South
African J Bot 72:217–223
Romeis J, Sharma HC, Sharma KK, Das S, Sarmah BK (2004)
The potential of transgenic chickpeas for pest control and
possible effects on non-target arthropods. Crop Prot
23:923–938
Rushton PJ, Reinstadler A, Lipka V, Lippok B, Somssich IE
(2002) Synthetic plant promoters containing defined reg-
ulatory elements provide novel insights into pathogen and
wound induced signaling. Plant Cell 14:749–762
Sambrook J, Russell DW (2001) Molecular cloning: a labora-
tory manual, 3rd edn. Cold Spring Harbor Press, NY
Sanyal I, Singh AK, Amla DV (2003) Agrobacterium tum-efaciens-mediated transformation of chickpea (Cicer ari-etinum L.) using mature embryo axes and cotyledonary
nodes. Indian J Biotech 2:524–532
Sanyal I, Singh AK, Kaushik M, Amla DV (2005) Agrobac-terium-mediated transformation of chickpea (Cicer ariet-inum L.) with Bacillus thuringiensis cry1Ac gene for
resistance against pod borer insect Helicoverpa armigera.
Plant Sci 168:1135–1146
Sardana R, Dukiandjiev S, Giband M, Cheng X, Cowan K,
Sauder C, Altosaar I (1996) Construction and rapid testing
of synthetic and modified toxin gene sequences CryIA (b
& c) by expression in maize endosperm culture. Plant Cell
Rep 15:677–681
Sawant S, Singh PK, Madanala R, Tuli R (2001) Designing of
an artificial expression cassette for the high-level
expression of transgenes in plants. Theor Appl Genet
102:635–644
Sawant SV, Kiran K, Mehrotra R, Chaturvedi CP, Ansari SA,
Singh P, Lodhi N, Tuli R (2005) A variety of synergistic
and antagonistic interactions mediated by cis-acting DNA
motifs regulate gene expression in plant cells and modu-
late stability of the transcription complex formed on a
basal promoter. J Exp Bot 56:2345–2353
Singsit C, Adang MJ, Lynch RE, Anderson WF, Wang A,
Cardineau G, Ozias-Akins P (1997) Expression of a
Bacillus thuringiensis cry1A(c) gene in transgenic peanut
plants and its efficacy against lesser cornstalk borer.
Transgenic Res 6:169–176
Somers DA, Samac DA, Olhoft PM (2003) Recent advances in
legume transformation. Plant Physiol 131:892–899
Stewart CN Jr, Adang MJ, All JN, Boerma HR, Cardineau G,
Tucker D, Parrott WA (1996) Genetic transformation,
recovery, and characterization of fertile soybean trans-
genic for a synthetic Bacillus thuringiensis cryIAc gene.
Plant Physiol 112:121–129
Tabashnik BE, Sisterson MS, Ellsworth PC, Dennehy TJ,
Antilla L, Liesner L, Whitlow M, Staten RT, Fabrick JA,
Unnithan GC, Yelich AJ, Ellers-Kirk C, Harpold VS, Li
X, Carriere Y (2010) Suppressing resistance to Bt cotton
with sterile insect release. Nature Biotech 28:1304–1307
Van Rheenen HR, Pundir RPS, Miranda JH (1993) How to
accelerate the genetic improvement of a recalcitrant crop
species such as chickpea. Curr Sci 654:414–417
Wang MB, Waterhouse PM (2002) Application of gene
silencing in plants. Curr Opin Plant Biol 5:146–150
Zhao J-Z, Cao J, Li Y, Collins HL, Roush RT, Earle ED,
Shelton AM (2003) Transgenic plants expressing two
Bacillus thuringiensis toxins delay insect resistance evo-
lution. Nature Biotech 21:1493–1497
Zhao J-Z, Cao J, Collins HL, Bates SL, Roush RT, Earle ED,
Shelton AM (2005) Concurrent use of transgenic plants
expressing a single and two Bacillus thuringiensis genes
speeds insect adaptation to pyramided plants. Proc Natl
Acad Sci USA 102:8426–8430
102 Euphytica (2011) 182:87–102
123








