
Pyramiding different alien chromosome segments in durum wheat: Feasibility and breeding potential
10
© 2008 Science From Israel / LPPLtd., Jerusalem Israel Journal of Plant Sciences Vol. 55 2007 pp. 267–276 *Author to whom correspondence should be addressed. E-mail: [email protected] Pyramiding different alien chromosome segments in durum wheat: Feasibility and breeding potential ANDREA GENNARO, a PAOLA FORTE, a ROBERTA CAROZZA, a MARIA L. SAVO SARDARO, a DANIELA FERRI, a ALESSANDRA BITTI, a GRAZIA M. BORRELLI, b MARIA G. D’EGIDIO, c AND CARLA CEOLONI a, * a Department of Agrobiology and Agrochemistry, University of Tuscia, Viterbo 01100, Italy b C.R.A.—Cereal Research Centre, Foggia 71100, Italy c C.R.A.—Research Unit for Cereal Quality, Rome 00191, Italy (Received 7 August 2007; accepted in revised form 7 February 2008) ABSTRACT Wheat chromosome engineering, i.e., the transfer of alien chromosome segments from various Triticeae species into cultivated wheats, is greatly benefiting from the recent advancements in molecular genetics, cytogenetics, and genomics. Powerful tools are currently available that make selection of desired genotypes far more pre- cise and effective than in the past, thus giving this transfer strategy considerable po- tential for meaningful practical achievements. Such tools were successfully applied to engineer the durum wheat genome with small alien segments containing genes for disease resistance and quality traits, including Lr19 (leaf rust resistance) and Yp (yellow endosperm pigmentation) from Thinopyrum ponticum, Pm13 (powdery mildew resistance) from Aegilops longissima, and the Glu-D3 and Glu-D1 glutenin subunit genes from Triticum aestivum. Targeted segments with such genes, all being of minimal size, were first separately incorporated into durum wheat by ph1-induced homoeologous recombination. The positive performance of selected recombinant lines from each transfer project encouraged attempts to develop multiple alien seg- ment combinations. The results obtained with double and also triple recombinants indicate a good tolerance of the durum wheat genome even toward such complex manipulations, which thus appear to offer good prospects for simultaneously enrich- ing durum wheat germplasm with several valuable traits from related Triticeae. Keywords: chromosome engineering, multiple alien transfers, disease resistance, quality, Triticum spp. INTRODUCTION Manipulation of multiple genes to achieve a multifac- eted improvement has always been the ultimate goal of any breeding strategy, from conventional intraspecific crossing activities, aimed at combining the best features of divergent parental varieties, to gene stacking via transformation technology. The former approach, par- ticularly for species such as the Triticum spp., finds an inherent impediment for further and diversified goals in the currently restricted genetic variation of the cultivat- ed gene pool, with many potentially useful genes “left behind” in the course of domestication and of intensive selection during the last century’s “Green Revolutions”. On the other hand, progress toward genetically modified crops with numerous novel genes can only be expected once the bottleneck of developing technologies for the coordinated manipulation of multiple genes or traits is removed (Halpin, 2005). Lying somewhere between traditional breeding and genetic engineering, chromosome engineering allows
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Pyramiding different alien chromosome segments in durum wheat: Feasibility and breeding potential

© 2008 Science From Israel / LPPLtd., Jerusalem
IsraelJournalofPlantSciences Vol.55 2007 pp.267–276
*Author to whom correspondence should be addressed.E-mail: [email protected]
Pyramiding different alien chromosome segments in durum wheat: Feasibility and breeding potential
AndreA GennAro,a PAolA Forte,a robertA CArozzA,a MAriA l. SAvo SArdAro,a dAnielA Ferri,a AleSSAndrA bitti,a GrAziA M. borrelli,b MAriA G. d’eGidio,c And CArlA Ceolonia,*
aDepartmentofAgrobiologyandAgrochemistry,UniversityofTuscia,Viterbo01100,ItalybC.R.A.—CerealResearchCentre,Foggia71100,Italy
cC.R.A.—ResearchUnitforCerealQuality,Rome00191,Italy
(Received7August2007;acceptedinrevisedform7February2008)
ABSTRACT
Wheat chromosome engineering, i.e., the transfer of alien chromosome segments from various Triticeae species into cultivated wheats, is greatly benefiting from the recent advancements in molecular genetics, cytogenetics, and genomics. Powerful tools are currently available that make selection of desired genotypes far more pre-cise and effective than in the past, thus giving this transfer strategy considerable po-tential for meaningful practical achievements. Such tools were successfully applied to engineer the durum wheat genome with small alien segments containing genes for disease resistance and quality traits, including Lr19 (leaf rust resistance) and Yp (yellow endosperm pigmentation) from Thinopyrumponticum, Pm13 (powdery mildew resistance) from Aegilops longissima, and the Glu-D3 and Glu-D1 glutenin subunit genes from Triticumaestivum. Targeted segments with such genes, all being of minimal size, were first separately incorporated into durum wheat by ph1-induced homoeologous recombination. The positive performance of selected recombinant lines from each transfer project encouraged attempts to develop multiple alien seg-ment combinations. The results obtained with double and also triple recombinants indicate a good tolerance of the durum wheat genome even toward such complex manipulations, which thus appear to offer good prospects for simultaneously enrich-ing durum wheat germplasm with several valuable traits from related Triticeae.
Keywords: chromosome engineering, multiple alien transfers, disease resistance, quality, Triticum spp.
INTRODUCTION
Manipulation of multiple genes to achieve a multifac-eted improvement has always been the ultimate goal of any breeding strategy, from conventional intraspecific crossing activities, aimed at combining the best features of divergent parental varieties, to gene stacking via transformation technology. The former approach, par-ticularly for species such as the Triticum spp., finds an inherent impediment for further and diversified goals in the currently restricted genetic variation of the cultivat-ed gene pool, with many potentially useful genes “left
behind” in the course of domestication and of intensive selection during the last century’s “Green Revolutions”. On the other hand, progress toward genetically modified crops with numerous novel genes can only be expected once the bottleneck of developing technologies for the coordinated manipulation of multiple genes or traits is removed (Halpin, 2005).
Lying somewhere between traditional breeding and genetic engineering, chromosome engineering allows

IsraelJournalofPlantSciences 55 2007
268
the transfer, in principle genome-wide, of genes from the wild or anyway crossable gene pool of the crop rela-tives via meiotic recombination between corresponding chromosomes of the recipient and donor species.
Crop species of paramount importance to mankind, such as the polyploid wheats, not only benefit from a wealth of natural genetic resources of alien Triticeae, but also a unique richness of cytogenetic materials that have greatly facilitated knowledge of the wheat and related Triticeae genomes and hence their controlled manipulation via chromosome engineering (Feldman and Sears, 1981). It is mainly thanks to the mind and work of E.R. Sears that such materials were conceived and developed, and that the main procedures for wheat chromosome engineering, i.e., the “transferring of seg-ments of alien chromosomes carrying particular desired genes to wheat chromosomes” (Sears, 1972), were originally delineated. In this framework, a refinement to the “toolkit” of wheat chromosome engineers came from isolation of mutants for the Ph1 gene, both in com-mon and in durum (reviewed in Ceoloni et al., 2005b; Qi et al., 2007). By removal of normally occurring suppression of recombination between wheat and ho-moeologous Triticeae chromosomes, largely due to the action of the wheat Ph1 gene, the way has been opened to transfers also from distantly related gene pools. These, although never as frequently obtained as when homologous exchanges are involved, result in largely balanced products in which the size of the alien seg-ment, and consequent linkage drag, can be conveniently minimized (e.g., Ceoloni et al., 2005a).
Until recently, these propitious conditions have been somewhat in contrast with the relatively modest success of chromosome engineering in making alien genetic variation effectively available to wheat breed-ers. Luckily, concomitantly with the increased need for enlarging the crop genetic basis that is pervading the breeding perspectives of this millennium (see, e.g., McCouch, 2004), the genomics revolution is offering unrivalled opportunities for understanding structure and function and hence for targeted manipulation of even complex plant genomes. No doubt it is greatly impacting all breeding strategies, including chromo-some engineering. In fact, molecular marker technol-ogy, as well as molecular cytogenetic techniques, such non-radioactive in situ hybridization (e.g., FISH and GISH), effectively complementing classical methods of analysis, are making detection and characterization of wheat–alien introgression products much more efficient and accurate than ever (see, e.g., Ceoloni and Jauhar, 2006, for a review). Such a “smart breeding” strategy is thus expected to accelerate the development of breeder-friendly germplasm, even in the form of more ambitious
products to be delivered, such as those where different valuable alien traits are pyramided. Here we will il-lustrate how this goal has been successfully pursued in durum wheat.
CHROMOSOME ENGINEERING AT THE DURUM WHEAT LEVEL: AN OVERVIEW
OF THE PERFORMANCE OF SINGLE ALIEN TRANSFERS
Durum wheat (Triticum turgidum subsp. durum, 2n = 4x = 28, AABB) is a species of traditional economic importance for Italy and other world areas with similar environmental conditions. We have exerted a major effort in its chromosome manipulation with the aim of stably introducing novel attributes enhancing the crop’s response to relevant diseases, as well as its qualitative performance. Our early attempts (Ceoloni et al., 1996) clearly confirmed the lower buffering ability of the T.durum genome as compared to that of T. aestivum (2n = 6x = 42, AABBDD) toward genic and chromo-somal alterations (reviewed in Ceoloni et al., 2005b), and hence the greater difficulties inherently associated with its manipulation (Ceoloni and Jauhar, 2006). None-theless, by taking advantage of currently available diag-nostic and selection tools, we succeeded in fulfilling an essential requisite for stability and consequent practical usefulness of introgression products, particularly those involving durum wheat, i.e., to obtain lines that incorpo-rate minimal amounts of alien chromatin. Materials of this kind could be precisely characterized and selected by molecular linkage maps (Donini et al., 1995; Cenci et al., 1999), meiotic pairing analyses (Donini et al., 1995; Carozza et al., 2003), and FISH/GISH-based physical maps (e.g., Biagetti et al., 1999; Carozza et al., 2003; Ceoloni et al., 2005a; see also Fig. 1) of the wheat–alien chromosomes resulting from each specific transfer pro-cess. Such integrated cytogenetic maps allowed marker and gene assignment to wheat and alien target chromo-some regions of defined length and position, as well as precise line ranking.
This could be achieved for each of three different transfer projects, through which the Pm13 gene from the diploid Aegilops longissima, the Lr19+Yp genes from the decaploid Thinopyrum ponticum, and the Glu-D1 or Gli-D1/Glu-D3 genes from T.aestivum were separately introduced into durum wheat. In all cases, lines could be isolated in which the size of the target alien segment did not exceed 23% of the engineered arm (Table 1). Even at pre-breeding stages such transfers appeared to be genetically well compensated, as indeed expected since they were obtained by resorting to the manipu-lation of homoeologous recombination. In particular,

Gennaroetal./Pyramidingalienchromosomesegmentsinwheat
269
free seasons (Gennaro et al., 2003, and unpublished). Yield stability over years and different environments, accompanied by the high carotenoid pigment content determined by the alien Yp gene (Table 1; Gennaro et al., 2003), and by good values of other relevant grain and semolina traits (Gennaro et al., 2003, and unpublished), led to the recent decision to submit one RC line for reg-istration to the Italian National Variety List.
Similarly advanced breeding work has not been performed for recombinant lines OR, PS, and PL (see Table 1), whose novel traits, however, may have consider-able breeding value. On the one hand, the Pm13 gene has been able so far to confer a complete resistance to pow-dery mildew pathotypes spread in Italy, as well as in other European countries (Chelkowski, pers. comm.) and in China (Zhou et al., 2005). On the other hand, introduction of the high- (HMW) and low-molecular-weight (LMW) glutenin subunit genes from common wheat chromosome 1D (Glu-D1d in line PL and Glu-D3 in line PS, Table 1), known to have a major impact on bread-making proper-ties, in place of their durum wheat 1A orthologs, signifi-cantly affects gluten properties and indicates the actual possibility to widen the spectrum of potential end uses of the recipient species (Ceoloni et al., 2000).
The observations reported above encouraged at-tempts aimed at simultaneously enriching a durum wheat genotype with different valuable traits of alien or-igin, and these have been undertaken adopting a strategy of “segment pyramiding”. In fact, while a few examples of stable stacking of multiple alien translocations with breeding potential exist in common wheat (e.g., Singh et al., 1998), no case history is available to our knowledge in durum wheat.
transfer of the Pm13 gene was first obtained at the 6´ level (Ceoloni et al., 1992) by use of the ph1b Chinese Spring mutant (Sears, 1977) and then moved into a 4´ background by homologous recombination (Ceoloni et al., 1996), while for the other transfers illustrated in Table 1 (Ceoloni et al., 1996, 2005a; Vitellozzi et al., 1997), wheat–alien chromosome recombination was induced exploiting ph1c durum wheat mutants (Giorgi, 1983; Ceoloni et al., 2005b).
The selected recombinant chromosomes (Table 1), named RC (Lr19+Yp), OR (Pm13), PS (Gli-D1/Glu-D3), and PL (Glu-D1d), were proved to be normally transmitted through both germlines (Ceoloni et al., 1996, 2005a; Vitellozzi et al., 1997), and not to impair relevant agronomic parameters of the recipient lines, as shown by evaluations of F3 progeny of the different recombinant lines following 2–3 backcrosses to adapted varieties (Ceoloni et al., 2000).
Also in later generations, particularly remarkable was the overall performance of one of the durum wheat–Th.ponticum recombinant lines, named R5-2-10, pos-sessing both the Lr19 and Yp genes in a 7AgL segment distally located on 7AL and spanning 23% of the arm (Ceoloni et al., 2005a). Starting from BC2F4, derivatives from the cross of R5-2-10 with the leaf-rust-susceptible Italian variety Simeto (named RC lines, Table 1) were subjected for several years to multi-location field tests. These provided consistent evidence of the efficacy of the Lr19-dependent leaf-rust-resistant response, as well as a remarkably positive performance of RC lines. They in fact yielded significantly more than varietal controls following heavy leaf-rust epidemics, but also confirmed their intrinsically good yield potential in virtually rust-
Table 1Characteristics of selected durum wheat recombinant lines carrying alien chromosome segments transferred from
wild and cultivated relatives
Rec. Recombinant Donor Alien segment Target genes Effectsline chromosome species (% arm)
RC Thinopyrum 23 Lr19+Yp Res. leaf rust + ponticum > carotenoid pigment content
OR Aegilops 20 Pm13 Res. powdery mildew longissima
PS Triticumaestivum 17 Gli-D1/ Glu-D3 Enhanced gluten qualityPL Triticumaestivum 16 Glu-D1d
IsraelJournalofPlantSciences 55 2007
270
A STEP FORWARD IN MANIPULATING THE DURUM WHEAT GENOME: ALIEN
CHROMOSOME SEGMENT PYRAMIDING
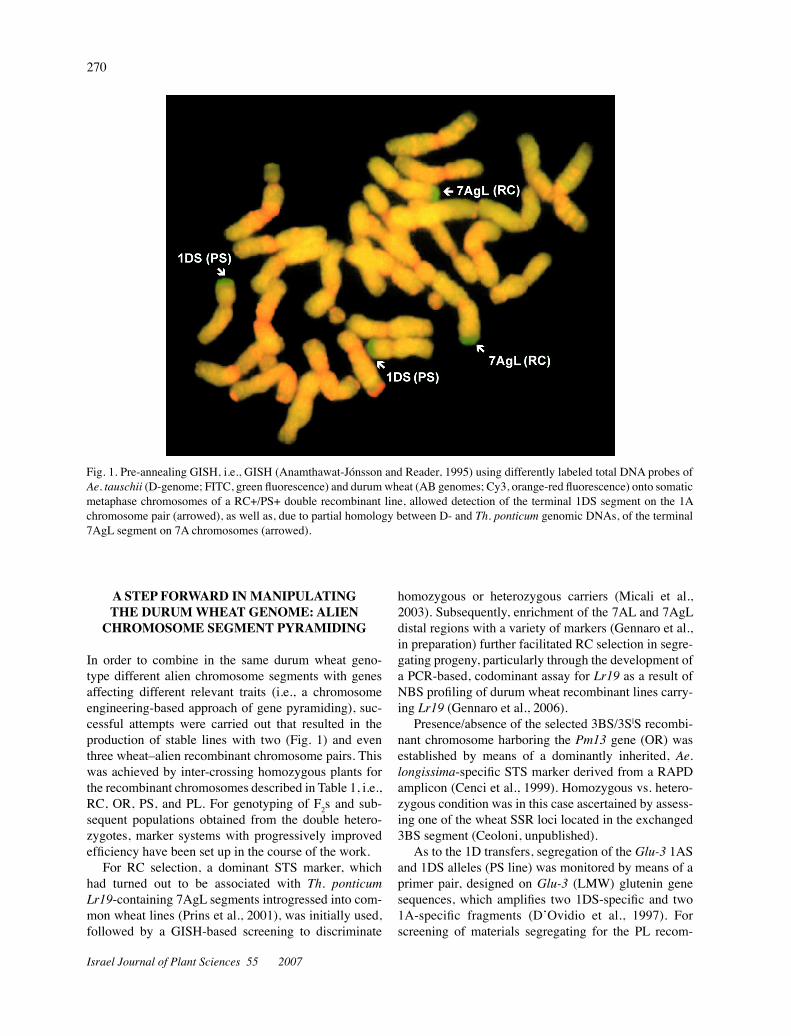
In order to combine in the same durum wheat geno-type different alien chromosome segments with genes affecting different relevant traits (i.e., a chromosome engineering-based approach of gene pyramiding), suc-cessful attempts were carried out that resulted in the production of stable lines with two (Fig. 1) and even three wheat–alien recombinant chromosome pairs. This was achieved by inter-crossing homozygous plants for the recombinant chromosomes described in Table 1, i.e., RC, OR, PS, and PL. For genotyping of F2s and sub-sequent populations obtained from the double hetero-zygotes, marker systems with progressively improved efficiency have been set up in the course of the work.
For RC selection, a dominant STS marker, which had turned out to be associated with Th. ponticum Lr19-containing 7AgL segments introgressed into com-mon wheat lines (Prins et al., 2001), was initially used, followed by a GISH-based screening to discriminate
homozygous or heterozygous carriers (Micali et al., 2003). Subsequently, enrichment of the 7AL and 7AgL distal regions with a variety of markers (Gennaro et al., in preparation) further facilitated RC selection in segre-gating progeny, particularly through the development of a PCR-based, codominant assay for Lr19 as a result of NBS profiling of durum wheat recombinant lines carry-ing Lr19 (Gennaro et al., 2006).
Presence/absence of the selected 3BS/3SlS recombi-nant chromosome harboring the Pm13 gene (OR) was established by means of a dominantly inherited, Ae.longissima-specific STS marker derived from a RAPD amplicon (Cenci et al., 1999). Homozygous vs. hetero-zygous condition was in this case ascertained by assess-ing one of the wheat SSR loci located in the exchanged 3BS segment (Ceoloni, unpublished).
As to the 1D transfers, segregation of theGlu-3 1AS and 1DS alleles (PS line) was monitored by means of a primer pair, designed on Glu-3 (LMW) glutenin gene sequences, which amplifies two 1DS-specific and two 1A-specific fragments (D’Ovidio et al., 1997). For screening of materials segregating for the PL recom-
Fig. 1. Pre-annealing GISH, i.e., GISH (Anamthawat-Jónsson and Reader, 1995) using differently labeled total DNA probes of Ae.tauschii (D-genome; FITC, green fluorescence) and durum wheat (AB genomes; Cy3, orange-red fluorescence) onto somatic metaphase chromosomes of a RC+/PS+ double recombinant line, allowed detection of the terminal 1DS segment on the 1A chromosome pair (arrowed), as well as, due to partial homology between D- and Th.ponticum genomic DNAs, of the terminal 7AgL segment on 7A chromosomes (arrowed).

Gennaroetal./Pyramidingalienchromosomesegmentsinwheat
271
binant, two primer pairs, one specific for the 1D ´ 5 component at the Glu-D1-1d locus and the other specific for the Glu-A1-2a (=1Ay) allele, were initially employed (Micali et al., 2003). More recently, development of detailed molecular maps of the PS and PL recombinants (Carozza et al., 2003; Ceoloni et al., in preparation), and, as a result of this, use of the WMC93 codominant SSR marker (Somers et al., 2004), which turned out to be associated with the exchanged 1AL vs. 1DL seg-ments containing Glu-1 loci, allowed their discrimina-tion through one PCR assay only.
Marker-driven analysis of F2 progeny of double heterozygous combinations (in OR/PS/RC combination the RC chromosome was homozygous) indicated simul-taneous transmission of the two recombinant chromo-somes to be normal through both germlines in all cases (Micali et al., 2003, and data not shown).
EVALUATION OF AGRONOMIC AND GRAIN QUALITY CHARACTERISTICS OF MULTIPLE
RECOMBINANT LINES
Agronomic traits
To test the effect of the multiple combinations on parameters of agronomic relevance, small-scale field tests were carried out for two years (2003–2004 and 2004–2005) in the same locality (Viterbo, central Italy) on F3 and F4 progeny of genotypes homozygous for ei-ther one or for all (2 or 3) recombinant chromosomes, or for their absence (Table 2). A mild leaf-rust attack was observed in both years; thus, a possible selective advan-tage for RC carriers (Lr19 gene) could be considered negligible. Characters including number of grains/ear (G/E), number of grains/spikelet (G/S), and 1000 grain weight (TGW) were evaluated, and data of the various genotypes within each combination were compared by ANOVA (Table 2). For all combinations, though with some between-years differences, no major difference was observed for ear fertility parameters, except for a significant increase of G/E shown in the second year by RC homozygotes compared to the other genotypes of the RC/OR F4 progeny. Also noticeable, albeit not sta-tistically significant, are the higher G/E and G/S values of OR/PS/RC triple homozygotes, which were consis-tently detected over the two generations and were not at the expense of their TGW (Table 2). Since both grain size and grain number are important yield-contributing traits, this evidence gives good hopes for considerable yield potential of such a complex genotype, as indeed initial yield trials seem to highlight (see ahead and Table 3).
Also for TGW most inter-genotype comparisons re-
sulted in nonsignificant differences. However, remark-ably higher values were associated with the presence of the RC chromosome, either alone or in combination with PS, in F3 progeny of the RC+PS combination, though among F4 genotypes only minor differences were ob-served. On the other hand, significantly lower TGW was detected in RC+/PL+ F3 plants and in their RC–/PL+ sibs, but the difference became nonsignificant when F4 genotypes of the RC/PL combination were compared. Reduced TGW was also observed in 1AS.1AL-1DL translocation lines carrying the same Glu-D1d allele (HMW glutenin subunits 5 + 10) as present in our PL line (Klindworth et al., 2005), although contained in a longer 1DL segment (Carozza et al., 2003; Ceoloni et al., 2005a; Xu et al., 2005). Major and minor QTLs for grain yield and yield components, including TGW, are widely distributed around the wheat genome, but no major QTL specifically related to TGW appears to be associated with distal 1AL or 1DL chromosome regions (Quarrie et al., 2005). Nonetheless, also hypothesizing that a weak genetic factor was involved in the different 1AL-1DL exchanges, compensation for the depressing effect could be exerted by positive alleles at other TGW-related QTLs present in variable genetic backgrounds, resulting in inconsistent and, probably, environmentally dependent results.
This possibility is in line with variable TGW values shown by F5 progeny of RC/PL lines subjected to yield trials in two rather different Italian environments (Ta-ble 3). In one of them (Po Valley = PV), the RC+/PL+ plots showed a lower TGW compared to control variet-ies, while the RC–/PL– materials had a higher value; on the other hand, the reverse was observed in the Adriatic Coast (AC) location, where the RC+/PL+ genotype per-formed better than the commercial varieties, whereas the opposite was true for the RC–/PL– genotype. As to the RC/PS combination, in both AC and PV locations RC+/PS+ progeny had higher TGW than control variet-ies, while the RC–/PS– sib progeny outperformed the control varieties in one location (PV) but had a lower value in the other (AC).
As evidently offset by other yield-contributing fac-tors, the observed variations in TGW did not seem to have a direct correspondence with yield performances (Table 3). In this regard, it has to be underlined that in the 2005–2006 season in both PV and AC environments a mild leaf-rust attack occurred; therefore, the inher-ent yield potential of contrasting genotypes (sib lines carrying or lacking the RC chromosome pair with the Lr19gene) could be assessed and compared to similarly rust-free (but otherwise susceptible) control varieties. Overall, very good yield performance was exhibited by both RC+/PS+ and RC+/PL+ recombinant genotypes,

IsraelJournalofPlantSciences 55 2007
272
Tabl
e 2
Mea
ns a
nd A
NO
VA p
val
ues o
f yie
ld-r
elat
ed tr
aits
of d
iffer
ent g
enot
ypes
sele
cted
from
var
ious
reco
mbi
nant
com
bina
tions
. Dat
a de
rive
from
5–1
0 sp
ikes
/row
(trip
licat
e, ra
ndom
-iz
ed, 1
.2-m
-long
row
s/ge
noty
pe) o
f F3 (
uppe
r lin
e) a
nd F
4 (lo
wer
line
) pla
nts (
2–5
prog
eny/
geno
type
)R
ecom
b.
Gra
ins/
spik
elet
G
rain
s/ea
r 10
00 g
rain
wei
ght
com
bina
tion1
Gen
otyp
es2
Gen
otyp
es2
Gen
otyp
es2
+/
+ +/
– –/
+ –/
– p
+/+
+/–
–/+
–/–
p +/
+ +/
– –/
+ –/
– p
RC
/ PS
3.
7 3.
7 3.
9 3.
8 0.
17
63.6
64
.2
64.5
68
.4
0.14
56
.4
57.8
52
.9
53.5
0.
03
3.5
3.4
3.5
3.4
0.57
68
.9
65.1
66
.1
66.3
0.
24
45.7
46
.9
45.1
45
.5
0.82
RC
/ PL
3.
5 3.
7 3.
8 3.
7 0.
07
62.0
69
.3
70.1
67
.2
0.36
41
.0
53.9
41
.6
51.6
0.
01
3.5
3.6
3.6
3.7
0.15
66
.3
69.1
66
.4
70.8
0.
19
42.0
44
.8
45.3
46
.2
0.06
RC
/ O
R
2.5
2.9
2.7
2.9
0.13
41
.4
49.2
47
.7
49.5
0.
48
52.0
48
.1
48.6
46
.5
0.25
3.
4 3.
7 3.
4 3.
4 0.
06
65.4
73
.5
65.4
68
.4
0.03
43
.6
44.2
46
.1
43.7
0.
51O
R /
PS /
RC
3 3.
8 —
3.
5 3.
6 0.
33
77.0
-
70.2
71
.9
0.08
42
.8
—
43.8
43
.1
0.90
3.
6 —
3.
5 3.
3 0.
17
73.0
-
72.1
67
.5
0.14
42
.7
—
41.6
41
.7
0.41
1 See
Tab
le 1
for a
bbre
viat
ions
and
des
crip
tion
of re
com
bina
nt c
hrom
osom
es. 2 “
+” a
nd “
–” in
dica
te h
omoz
ygos
ity fo
r pre
senc
e an
d ab
senc
e, re
spec
tivel
y, o
f the
firs
t and
seco
nd
reco
mbi
nant
chr
omos
ome
as in
the
sequ
ence
indi
cate
d in
the
first
col
umn.
3 The
RC
chr
omos
ome
is a
lway
s hom
ozyg
ous i
n th
is tr
iple
com
bina
tion.
Tabl
e 3
Prel
imin
ary
field
eva
luat
ions
of F
5 dur
um w
heat
mul
tiple
reco
mbi
nant
line
s and
thei
r sib
s (2
prog
eny
per
geno
type
) com
pare
d to
ada
pted
var
ietie
s use
d as
con
trols
(2–3
per
loca
lity;
con
trol m
eans
= 1
00) i
n di
f-fe
rent
Ital
ian
envi
ronm
ents
(AC
= A
dria
tic S
ea C
oast
, cen
tral-e
ast;
PV=
Po
Valle
y, c
entra
l-nor
th; T
C=
Ty
rrhe
nian
Sea
Coa
st, c
entra
l-wes
t)1,2
Line
Te
st w
eigh
t 10
00 g
rain
wei
ght
Yie
ld
AC
PV
TC
AC
PV
AC
PV
TCR
C+
/ PS+
10
1 99
—
10
9 10
5 98
10
3 —
RC
– / P
S–
102
97
—
95
110
104
104
—R
C+
/ PL+
10
3 99
—
10
3 87
* 10
3 10
0 —
RC
– / P
L–
101
95
—
95
109*
91
10
3 —
OR
+ / P
S+ /
RC
+ —
—
11
1*
—
—
—
—
160*
*O
R–
/ PS–
/ R
C+
—
—
110*
—
—
—
—
17
2**
1 Lea
f-ru
st a
ttack
was
ver
y m
ild i
n AC
and
PV
loca
litie
s (2
005–
2006
sea
son)
and
ver
y se
vere
in
TC
(200
6–20
07 se
ason
). 2
repl
icat
es o
f 7.2
m2 pl
ots i
n AC
and
PV;
3 re
plic
ates
of 4
.2 m
2 plo
ts in
TC
.2 *, **
Indi
cate
sig
nific
ant d
iffer
ence
s vs
. con
trol m
eans
at p
= 0
.05
and
0.01
, res
pect
ivel
y, a
ccor
ding
to
Stud
ent’s
t-te
st. A
ll ot
her d
iffer
ence
s wer
e no
nsig
nific
ant.
Gennaroetal./Pyramidingalienchromosomesegmentsinwheat
273
which in both areas had a seed production close to or even better than the controls. On the other side, presence of the Lr19 gene determined a tremendous increase in yield (average 66%) of OR/PS/RC recombinant geno-types over the three susceptible control varieties in a field test carried out in 2006–2007 near the Tyrrhenian Sea Coast (TC, Table 3). This is an area where durum wheat is widely and traditionally cultivated, but where the crop is consistently and severely damaged by the leaf-rust disease. Thus, materials of this kind may well provide an effective alternative to conventional, largely susceptible varieties, currently representing the majority of the cultivated durum wheat germplasm.
Quality characteristics
The overall good prospects that the multiple recombi-nant lines we developed seem to hold in terms of ag-ronomic performance are further substantiated by their quality attributes. As previously shown for the separate recombinants (Ceoloni et al., 2000; Gennaro et al., 2003), in double and triple combinations carrying the alien genes Yp (RC chromosome, Table 1), Glu-D1, and Glu-D3 (PL and PS chromosome, respectively), such genes determined considerable increases of indexes used to measure quality characteristics that, besides protein content, are of major interest for durum wheat improvement, i.e., gluten quality and color.
Gluten quality, as expressed by SDS-sedimentation and Gluten Index, was appreciably affected by the pres-ence of 1D-encoded proteins in RC+/PL+, RC+/PS+, and OR+/PS+/RC+ lines (Table 4). Compared to the controls, increments in SDS values were around 30% in double recombinants with either PL or PS lines chromo-some pairs, and about 15% in the triple combination.
Differences in Gluten Index were smaller (around 15% for PS- and PL-containing double recombinants and very minor in the triple one). As expected, quality parameters were not genotype-dependent in the RC/OR combina-tion. Quantitative variations in the effect of 1D-encoded proteins even among recombinants carrying the same 1D transfer (i.e., RC+/PS+ and OR+/PS+/RC+) may be due to residual differences in their background seed storage protein composition. In fact, the two to three backcrosses to the same durum wheat cultivar ‘Simeto’, performed with all separate recombinants prior to their use as parents of multiple combinations, are not expected to homogenize their background genotypes, although they seem to possess the same major protein determinants of good pasta quality, including an LMW-2 type of pattern and the Glu-B1-encoded 7+8 HMW glutenin subunits, as the recurrent variety (not shown). Preliminary results from alveograph measurements on F4 progeny of multiple recombinants with either PS or PL segments indicate that semolina rheological proper-ties are considerably but differently influenced by Gli-D1/Glu-D3- and Glu-D1d-encoded proteins, the former mainly determining a substantial increment in dough strength, the latter having a stronger increasing effect on tenacity.
Semolina of the same recombinant combinations was also used to assess color parameters (Table 4). As a whole, the observed values confirmed earlier obser-vations on RC-type recombinant lines carrying the Yp gene from Th.ponticum (Gennaro et al., 2003). In terms of Yellow Index, RC+ F3 plants exhibited a 10–16% increment with respect to RC–. Spectrophotometric determination of total carotenoid pigments (Borrelli et al., 1999) and evaluation of their lutein and b-carotene
Table 4Quality parameters of durum wheat multiple recombinant lines (mean values of 2–4 F3 progeny plants per genotype)
Genotype1 SDS Gluten Yellow Carotenoid pigments LOX (mm) index index Total Lutein b-carotene (EU/g) (μg/g) (μg/g) (μg/g)RC+ / PS+ 47 98 23.4 9.3 4.4 0.10 1.9RC– / PS– 36 84 21.3 7.8 2.7 0.08 2.3RC+ / PL+ 55 100 24.3 11.0 5.9 0.10 2.3RC– / PL– 43 87 21.6 8.5 4.0 0.08 2.1RC+ / OR+ 38 72 24.1 9.9 4.5 0.15 1.2RC– / OR– 37 90 20.8 6.8 2.6 0.09 1.3OR+ / PS+ / RC+ 55 99 24.4 9.3 4.4 0.14 1.1OR– / PS– / RC+ 48 96 23.6 8.9 4.2 0.13 1.91See Table 1 for abbreviations and description of recombinant chromosomes. “+” and “–” indicate homozygosity for presence or absence, respectively, of the given recombinant chromosome.2LOX = lipoxygenase activity.

IsraelJournalofPlantSciences 55 2007
274
fractions by high-performance liquid chromatography (Adom et al., 2003) showed more clear-cut differences. In particular, total pigment content was about 20%, 30%, and over 40% higher in RC+/PS+, RC+/PL+, and RC+/OR+, respectively, than in their sib lines lacking the alien Yp gene. As expected, the main contribution to these increments was due to lutein, the major com-ponent of total pigments in the wheat grain (Adom et al., 2003): lutein was almost 50% more abundant in RC+/PL+ lines as compared to RC–/PL– controls, and the increases were over 60% and 70%, respectively, in the case of RC/PS and RC/OR combinations. Accord-ing to expectation, comparison between the OR/PS/RC triple recombinant genotypes, all being homozygous for the presence of the RC chromosome, revealed no ap-preciable difference in total pigment content, nor in that of lutein and b-carotene (Table 4). On the other hand, amounts of the latter compound were considerably high-er (+66%) in RC+/OR+ plants than in RC–/OR– con-trols, while in both RC+/PS+ and RC+/PL+ genotypes a 25% increment over the corresponding –/– genotypes of the same progeny was detected. In semolina extracted from the various recombinant combinations, lipoxygen-ase (LOX) activity, the main determinant of oxidative degradation of pigments during processing, was also assessed. In line with the known independent location of wheat LOX genes (Hart and Langston, 1977) and of major QTLs affecting endosperm color pigments (e.g., Parker et al., 1998), no genotype-dependent relation be-tween LOX activity and pigment content was detected (Table 4). Indeed, among RC+ genotypes, a number were found to express relatively low LOX activity, and these are expected to cause a minimum reduction of color in pasta products (Borrelli et al., 1999).
CONCLUDING REMARKS
In the breeding perspective of this millennium, we are assisting the fortunate transition of what was until re-cently just a largely shared conviction, i.e., that of the need for widening the crop genetic basis as a key to meaningful breeding advancements, to its translation into the actual possibility to develop effectively im-proved varieties, capable of meeting an array of constant as well as newly arisen socioeconomic and environmen-tal challenges.
Our results demonstrate how this definitely applies to the “smart” or “precision” chromosome engineer-ing-based breeding approach, even when the target is pyramiding multiple alien segments, and the crop species, as in the durum wheat case, has a genome relatively “sensitive” to extensive manipulations. We have in fact succeeded in stably assembling into durum
wheat chromosomal segments from up to three related genomes, two from the wild gene pool (Ae. longis-sima and Th.ponticum) and one from common wheat, contributing to the simultaneous enhancement of traits that can positively affect environment safety and pro-duction incomes and costs, but also the technological and nutritional quality of the end product. The multiple recombinant lines have so far quite brilliantly passed the most critical test for a breeding application, i.e., yield performance, as seems to be particularly the case for RC (Lr19+Yp genes)-containing lines. Moreover, their quality attributes represent an important added value. Actually, the feasible combination of high carotenoid pigment content and low LOX activity mentioned above can enhance the aesthetic appeal and also the health benefits of food products derived from RC genotypes (Adom et al., 2003). Also, a possible expansion of the durum wheat market can be envisaged if a “dual-purpose” gluten can be realized, and good prospects seem to be particularly associated with lines bearing the PS (Gli-D1/Glu-D3 genes) recombinant chromosome (Ceoloni et al., 2000, and in preparation).
While larger-scale field and quality tests are currently underway to better evaluate the overall potential of the materials described here, they are being employed in marker-assisted breeding to simultaneously introduce the novel traits into an array of adapted varieties. This will allow the development of novel germplasm in which the effects of the segment introgressions can be conveniently optimized, in view of suiting dif-ferent environments and market appreciation. In the meantime, additional alien segment combinations are being developed, in which, for example, various alien resistance genes toward the same or different pathogens are brought together, or where both PS and PL transfers are introduced into the same 1A chromosome. The lat-ter “reshuffling”, whether practically useful or not, will allow one to delineate a more complete picture of the effects of 1D gene products on durum wheat gluten properties.
Although, as earlier recalled, the case history is very limited for wheat (for other crop species see, e.g., Mc-Couch, 2004, and references therein), it is likely that the successful chromosome segment pyramiding described here will be reproducible with a variety of alien trans-fers, and it will certainly be more amenable if addressed to the hexaploid genome of common wheat.
However, the recent accomplishments with either single or multiple introgressions already provide ample evidence of the actual possibility of a finely-tuned and breeding-responsive makeup of the wheat genome through a “smart” chromosome engineering approach (see, e.g., Ceoloni et al., 2005a; Ceoloni and Jauhar,

Gennaroetal./Pyramidingalienchromosomesegmentsinwheat
275
2006; Qi et al., 2007, and references therein). This should be considered in the present and future perspec-tives of invigorating cultivars with “new blood”, for which a policy should be judiciously followed that contemplates the resort, at proper times, to all efficiently working strategies, refraining from roughly-made eval-uations (Jacobsen and Schouten, 2007), but just aspiring to effectively achievable benefits.
REFERENCES
Adom, K.K., Sorrells, M.E., Hai, L.R. 2003. Phytochemical profiles and antioxidant activity of wheat varieties. J. Ag-ric. Food Chem. 51: 7825–7834.
Anamthawat-Jónsson, K., Reader, S.M. 1995. Pre-annealing of total genomic DNA probes for simultaneous genomic in situ hybridization. Genome 38: 814–816.
Biagetti, M., Vitellozzi, F., Ceoloni, C. 1999. Physical map-ping of the wheat-Aegilops longissima breakpoints in mildew resistant recombinant lines using FISH with highly repeated and low-copy DNA probes. Genome 42: 1013–1019.
Borrelli, G.M., Troccoli, A., Di Fonzo, N., Fares, C. 1999. Du-rum wheat lipoxigenase activity and other quality param-eters that affect pasta color. Cereal Chem. 76: 335–340.
Carozza, R., Pagnotta, M.A., Ceoloni, C. 2003. Cytogenetic characterization of chromosomally engineered wheat chro-mosomes by FISH-based physical maps, molecular linkage maps and meiotic pairing analysis. Proc. 4th European Cytogenetics Conf., Bologna, 6–9 September 2003. Ann. Génét. 46-n° 2–3: 301.
Cenci, A., D’Ovidio, R., Tanzarella, O.A., Ceoloni, C., Porceddu, E. 1999. Identification of molecular markers linked to Pm13, an Aegilops longissima gene conferring resistance to powdery mildew of wheat. Theor. Appl. Genet. 98: 448–454.
Ceoloni, C., Jauhar, P.P. 2006. Chromosome engineering of the durum wheat genome: Strategies and applications of potential breeding value. In: Singh, R.J., Jauhar, P.P., eds. Genetic resources, chromosome engineering, and crop improvement: cereals. CRC Press, Boca Raton, FL, pp. 27–59.
Ceoloni, C., Del Signore, G., Ercoli, L., Donini, P. 1992. Locating the alien chromatin segment in common wheat-Aegilops longissima mildew resistant transfers. Hereditas 116: 239–245.
Ceoloni, C., Biagetti, M., Ciaffi, M., Forte, P., Pasquini, M. 1996. Wheat chromosome engineering at the 4´ level: the potential of different alien gene transfers into durum wheat. Euphytica 89: 87–97.
Ceoloni, C., Forte, P., Ciaffi, M., Nenno, M., Bitti, A., De Vita, P., D’Egidio, M.G. 2000. Chromosomally engineered durum wheat: the potential of alien gene introgressions affecting disease resistance and quality. Options Médit. A-40: 363–371.
Ceoloni, C., Forte, P., Gennaro, A., Micali, S., Carozza, R., Bitti, A. 2005a. Recent developments in durum wheat
chromosome engineering. Cytogenet. Genome. Res. 109: 328–334.
Ceoloni, C., Pasquini, M., Simeone, R. 2005b. The cytogenet-ic contribution to the analysis and manipulation of the du-rum wheat genome. In: Royo, C., Nachit, M.N., Di Fonzo, N., Araus, J.L., Pfeiffer, W.H., Slafer, G.A., eds. Durum wheat breeding: current approaches and future strategies. Haworth Press, New York, pp. 165–196.
Donini, P., Koebner, R.M.D., Ceoloni, C. 1995. Cytogenetic and molecular mapping of the wheat-Aegilopslongissima chromatin breakpoints in powdery mildew resistant intro-gression lines. Theor. Appl. Genet. 91: 738–743.
D’Ovidio, R., Simeone, M., Masci, S., Porceddu, E. 1997. Molecular characterization of a LMW-GS gene located on chromosome 1B and the development of primers specific for the Glu-B3 complex locus in durum wheat. Theor. Appl. Genet. 95: 1119–1126.
Feldman, M., Sears, E.R. 1981. The wild gene resources of wheat. Sci. Am. 244: 98–109.
Gennaro, A., Borrelli, G.M., D’Egidio, M.G., De Vita, P., Rav-aglia, S., Ceoloni, C. 2003. A chromosomally engineered durum wheat-Thinopyrumponticum recombinant line with novel and promising attributes for varietal development. In: Pogna, N.E., Romanò, M., Pogna, E.A., Galterio, G., eds. Proc. 10th Int. Wheat Genetics Symp., Paestum, Italy, S.I.M.I., Rome, pp. 881–883.
Gennaro, A., Ceoloni, C., Koebner, R.M.D. 2006. Application of NBS profiling to durum wheat–Th.ponticum recombi-nant lines carrying the Lr19 leaf-rust resistance gene. In: Börner, A., Pankova, K., Snape, J.W., eds. EWAC Newslet-ter, pp. 127–129.
Giorgi, B. 1983. Origin, behaviour and utilization of a Ph1 mutant of durum wheat, Triticumturgidum (L.) var. durum. In: Sakamoto, S., ed. Proc. 6th Int. Wheat Genetics Symp., Kyoto, Japan, Plant Germplasm Inst., Kyoto Univ., pp. 1033–1040.
Halpin, C. 2005. Gene stacking in transgenic plants—the chal-lenge for 21st century plant biotechnology. Plant Biotech. J. 3: 141–155.
Hart, G.E., Langston, P.J. 1977. Chromosomal location and evolution of isozyme structural genes in hexaploid wheat. Heredity 39: 263–277.
Jacobsen, E., Schouten, H.J. 2007. Cisgenesis strongly im-proves introgression breeding and induced translocation breeding of plants. Trends Biotechnol. 25: 219–23.
Klindworth, D.L., Hareland, G.A., Elias, E.M., Xu, S.S. 2005. Agronomic and quality characteristics of 1AS.1AL-1DL translocation lines of durum wheat carrying glutenin allele Glu-D1d. Crop Sci. 45: 77–84.
McCouch, S. 2004. Diversifying selection in plant breeding. PloS Biol. 2: e347.
Micali, S., Forte, P., Bitti, A., D’Ovidio, R., Ceoloni, C. 2003. Chromosome engineering as a tool for effectively intro-gressing multiple useful genes from alien Triticeae into durum wheat. In: Pogna, N.E., Romanò, M., Pogna, E.A., Galterio, G., eds. Proc. 10th Int. Wheat Genetics Symp., Paestum, Italy, S.I.M.I., Rome, pp. 896–898.
Parker, G.D., Chalmers, K.J., Rathjen, A.J., Langridge, P.

IsraelJournalofPlantSciences 55 2007
276
1998. Mapping loci associated with flour colour in wheat (Triticumaestivum L.). Theor. Appl. Genet. 97: 238–245.
Prins, R., Groenewald, J.Z., Marais, G.F., Snape, J.W., Koeb-ner, R.M.D. 2001. AFLP and STS tagging of Lr19, a gene conferring resistance to leaf rust in wheat. Theor. Appl. Genet. 103: 618–624.
Qi, L., Friebe, B., Zhang, P., Gill, B.S. 2007. Homoeologous recombination, chromosome engineering and crop im-provement. Chromos. Res. 15: 3–19.
Quarrie, S.A., Steed, A., Calestani, C., et al. 2005. A high-den-sity genetic map of hexaploid wheat (TriticumaestivumL.) from the cross Chinese Spring x SQ1 and its use to com-pare QTLs for grain yield across a range of environments. Theor. Appl. Genet. 110: 865–880.
Sears, E.R. 1972. Chromosome engineering in wheat. Proc. Stadler Genet. Symp. 4: 23–38.
Sears, E.R. 1977. An induced mutant with homoeologous pair-ing in common wheat. Can. J. Genet. Cytol. 19: 585–593.
Singh, R.P., Huerta-Espino, J., Rajaram, S., Crossa, J. 1998. Ag-ronomic effects from chromosome translocations 7DL.7Ag and 1BL.1RS in spring wheat. Crop Sci. 38: 27–33.
Somers, D.J., Isaac, P., Edwards, K. 2004. A high-density microsatellite consensus map for bread wheat (Triticumaestivum L.). Theor. Appl. Genet. 109: 1105–1114.
Vitellozzi, F., Ciaffi, M., Dominici, L., Ceoloni, C. 1997. Iso-lation of a chromosomally engineered durum wheat line carrying the common wheat Glu-D1d allele. Agronomie 17: 413–417.
Xu, S.S., Faris, J.D., Cai, X., Klindworth, D.L. 2005. Molecu-lar cytogenetic characterization and seed storage protein analysis of 1A/1D translocation lines of durum wheat. Chromos. Res. 13: 559–568.
Zhou, R., Zhu, Z., Kong, X., Huo, N., Tian, Q., Li, P., Jin, C., Dong, Y., Jia, J. 2005. Development of wheat near-isogenic lines for powdery mildew resistance. Theor. Appl. Genet. 110: 640–648.




















