
转cry1Ac基因抗虫棉花DAS-21023-5进口用作加工原料的安全 ...
257
1 项目号: 项目类: 农业转基因生物安全评价 申 报 书 项目名称:转 cry1Ac 基因抗棉花 DAS-21023-5 进口用作工原料 的安证书 申请单位:科迪华业科技有限责任司 申 请 人: 地 址: 邮政码: 电 话: 传 真: E-mail: 填报期:2021 年 10 月 中华人和国业村部
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of 转cry1Ac基因抗虫棉花DAS-21023-5进口用作加工原料的安全 ...

1
项目编号: 项目类别:
农业转基因生物安全评价申 报 书
项目名称:转 cry1Ac 基因抗虫棉花 DAS-21023-5 进口用作加工原料
的安全证书
申请单位:科迪华农业科技有限责任公司
申 请 人:
地 址:
邮政编码:
电 话:
传 真:
E-mail:
填报日期:2021 年 10 月
中华人民共和国农业农村部制

2

3
目 录
商业保密资料(CBI)声明 ......................................................................................... 1
一、申请表.................................................................................................................... 2
二、项目内容摘要........................................................................................................ 4
三、工作目的和意义.................................................................................................... 8
四、国内外研究的相关背景材料................................................................................ 9
五、安全性评价.......................................................................................................... 11
1 受体植物的安全性评价...................................................................................... 11
2 基因操作的安全性评价...................................................................................... 21
3 转基因植物的安全性评价.................................................................................. 57
4 转基因植物产品的安全性评价........................................................................ 109
5 供体生物的安全性评价.................................................................................... 114
六、相关附件资料.................................................................................................... 119
七、本单位农业转基因生物安全小组审查意见.................................................... 233
八、本单位审查意见................................................................................................ 234
参考文献.................................................................................................................... 235

4

5
填 写 说 明
1.在填写申报书之前,应认真阅读《农业转基因生物安全管理条例》、《农
业转基因生物安全评价管理办法》、《农业转基因生物进口安全管理办法》等有
关法规,了解相关要求。
2.此申报书同样适用于法规规定的报告类实验研究和中间试验的报告,名
称分别改为“农业转基因生物实验研究报告书”和“农业转基因生物中间试验报告
书”。
3.申报书内容应当包括以下部分:申请表、项目内容摘要、工作目的和意
义、国内外研究的相关背景资料、安全性评价、试验方案、相关附件资料、本
单位农业转基因生物安全小组审查意见、本单位审查意见、所在省(市、自治
区)的农业行政主管部门的审查意见。
4.申请实验研究的,申请表按表 2 填写,项目名称应包含转基因性状、受
体生物名称、实验研究所在省(市、自治区)名称和实验研究阶段等内容,如
“转基因抗虫棉花在河北省的实验研究”。一份申报书只能包含同一物种的受体
生物和相同的转基因性状。“安全性评价”可不填写。“试验方案”包括研究目
标、外源基因的名称和来源、载体构建图谱、转化方法和规模、地点、安全控
制措施等。“相关附件资料”包括法人证书和营业执照的复印件,若涉及合作项
目,应提交双方合作或转让协议等。“所在省(市、自治区)的农业行政主管部
门的审查意见”可不填写,申报材料报送农业农村部时,抄送实验研究所在省
(市、自治区)的农业行政主管部门。
5.申请中间试验的,申请表按表 2 填写。申报书中“安全性评价”、“试验
方案”和“相关附件资料”依照《农业转基因生物安全评价管理办法》附录 I、II、
III 要求逐条填写。“所在省(市、自治区)级农业行政主管部门的审查意见”可
不填写,申报材料报送农业农村部时,抄送中间试验所在省(市、自治区)级
农业行政主管部门。
6.申请环境释放和生产性试验的,申请表按表 2 填写;申报书中“安全性
评价”、“试验方案”和“相关附件资料”依照《农业转基因生物安全评价管理办
法》附录 I、II、III 要求逐条填写。对于已批准过的环境释放或生产性试验,在
试验结束后,若同一转基因生物在原批准地点以相同规模重复试验,在申报时
“安全性评价”部分可省略。
7.申请农业转基因生物安全证书的,申请表按表 3 填写;申报书中“安全
性评价”和“相关附件资料”依照《农业转基因生物安全评价管理办法》附录 I、
II、III 要求逐条填写,“试验方案”不用填写;转基因植物的品种命名应符合
《农业植物品种命名规定》;首次申请农业转基因生物品种(或品系)生产应用
安全证书的,需提供《农业转基因生物品种(或品系)生产应用综合评价报
告》,主要包括对我国生产、贸易、社会等方面影响。

6
8.首次申请农业转基因生物生产性试验和安全证书的,在申请截止日 30 个
工作日前提供所申报转基因生物活性样品和技术资料。样品要求:符合要求
的,有活性的植物种子 2.5kg,动物 5ml 血样或 3g 组织各三份,微生物 3 批
次,各 10 管样品。技术资料要求:申请生产性试验提供外源片段整合进基因组
的 Southern 杂交结果、拷贝数及检测方法等;申请安全证书提供外源插入片段
信息及转化体特异性 PCR 检测方法等。
9.申报书应当用中文填写,一式十份,一律使用 A4 纸,正文用小四宋体
打印,标准字间距和单倍行距,并提供光盘。对于不符合要求的申报书,不予
受理。
10.申请者可以注明哪些资料需要保密并说明理由。
11.受理单位:农业农村部行政审批综合办公室科教窗口。地址:北京市
朝阳区农展馆南里 11 号。邮政编码:100125。

1
商业保密资料(CBI)声明
此申请包含商业保密资料并受版权法的保护。此文件仅限于科迪华农业科技
有限责任公司递交申请的法规和注册机构使用,并且仅支持本公司的申请。没有
本公司事先的书面同意, 所有其它目的使用将被严格禁止。递交此文件给相关法
规和注册机构并不等于本公司将此文件中的信息或知识产权授权或特许给任何组
织或实体。本公司花费了大量的人力物力开发研究本转化事件并攥写该申报资料,
保密资料的泄露会对本公司造成巨大的损失。现特依据《农业转基因生物安全评
价管理办法》第三章第二十七条的有关规定要求对本申请书、附件和检测报告中
涉及商业保密资料部分进行保密。
2
一、申请表
农业转基因生物安全证书申请表
项 目 概 况
项目名称 转 cry1Ac 基因抗虫棉花 DAS-21023-5 进口用作加工原料的安全证书
项目来源 科迪华农业科技有限责任公司
转
基
因
生
物
概
况
类 别 动物 植物√ 微生物 (选√)
转基因生物名称 DAS-21023-5
受体生物
中文名 棉花 学 名 Gossypium hirsutum L.
分类学 地位
锦葵科(Malvaceae); 棉属(Gossypium); 陆地棉种(Gossypium
hirsutum L.)
品种(品
系)名称 GC510 安全等级 Ⅰ
目的基因 1
名 称 cry1Ac 供体生物 土壤细菌苏云金芽孢杆
菌 (Bacillus thuringiensis) kurstaki 亚种
生物学 功能
使表达该基因的棉花对特定的鳞翅目害虫具有抗性。
启动子 ZmUbi1 启动子 终止子 AtuORF25/26 3' UTR
载 体 1 质粒载体 pMYC3006 供 体 生 物 根癌农杆菌菌株 LBA4404
﹡标记基因 1 名 称 pat 供体生物
绿色产色链霉菌 (Streptomyces
viridochromogenes)
启动子 AtuMas 启动子 终止子 AtuORF25/26 3' UTR
﹡报告基因 1 名 称 无 供体生物 无
启动子 无 终止子 无
转基因方法 农杆菌介导法 基因操作类型 2
转基因生物品系(株系)名
称 DAS-21023-5
转基因生物品系(株系)个
数 1
转基因生物安全等
级 Ⅰ 转基因生物产品安全等级 Ⅰ
试
验
概
况
中间 试验 情况
转基因生物 名称及编号
可以不填写
批准文号 可以不填写
试验的时间、地点和规模 可以不填写
环境 释放 情况
转基因生物名称及编号 可以不填写
批准文号 可以不填写
批准时间、地点和规模 可以不填写
3
生产 性试 验情 况
转基因生物名称及编号 可以不填写
批准文号 可以不填写
批准时间、地 点和规模
可以不填写
拟申请使用范围(省、自 治 区 、 直 辖
市 ) 境内
拟 申 请 使 用 年 限 5 年
申 请 单 位 概 况
单位名称 科迪华农业科技有限责任公
司 地 址
邮 编 电 话
传 真 电子邮件
单位性质 境内单位(事业 企业 中外合作 中外合资 外方独
资 )
境外单位(企业√ 其它 )(选√)
申请人姓名 电 话
传 真 电子邮箱
联系人姓名 电 话
传 真 电子邮箱
研
制
单
位
概
况
单位名称 法人代表
联系人姓名 电 话
传 真 电子邮箱
主 要 完 成 人
姓名 性别 出身年月
学历 专业技术职务
何时何地曾从事何种
基因工程工作
参 与 完 成 人
姓名 年龄 学历 职称 单 位 在本项目中的分工
注:1.申请农业转基因生物实验研究的,“受体生物品种(品系)名称、目的基因启动子和
终止子、载体、标记基因、报告基因、调控序列、转基因生物品系(株系)”栏目不用填写。
2.如果“标记基因”或“报告基因”已删除,应在表中标注。3.申请人指所申请项目的安全监
管具体负人。

DAS-21023-5 棉花入境审批书 – 种用棉籽 用于环境安全检测

DAS-21023-5 棉花入境审批书 – 谷物 用于食用安全检测(15 kg)

DAS-21023-5 棉花入境审批书 – 谷物 用于食用安全检测(9 kg)

DAS-21023-5 棉花入境审批书-阳性样品

4
二、项目内容摘要
本公司开发了转基因抗虫棉花 DAS-21023-5(OECD 名称为 DAS-21Ø23-5),
该棉花与传统非转基因棉花同样安全。本公司向中华人民共和国农业农村部申请
转基因棉花 DAS-21023-5 进口用作加工原料的安全证书。以下为本申报书摘要。
1. 开发转基因抗虫棉花 DAS-21023-5 的意义
采用农杆菌介导转化法将 cry1Ac 基因和 pat 基因转化至受体陆地棉 GC510
基因组中,然后通过传统育种方法选育出含转化事件 DAS-21023-5 的抗虫棉花。
cry1Ac 基因来自土壤细菌苏云金芽孢杆菌 kurstaki 亚种(Bacillus thuringiensis
subsp. kurstaki, Bt),编码杀虫蛋白质 Cry1Ac。表达 Cry1Ac 的 DAS-21023-5 棉
花对鳞翅目多种害虫具有抗性,包括棉铃虫、烟草夜蛾、棉红铃虫、甜菜夜蛾、
大豆尺蠖和南部粘虫等。pat 基因来自土壤细菌绿色产色链霉菌(Streptomyces
viridochromogenes),编码膦丝菌素乙酰转移酶(phosphinothricin acetyltransferase,
PAT),PAT 在 DAS-21023-5 棉花转化过程中作为选择标记。
DAS-21023-5 棉花可有效地抵抗数种鳞翅目害虫,在虫害猖獗的地区能降低
或代替杀虫剂,不仅可提高生产耕作效率、减少投入、增加产出,还可减少环境
污染,从而给棉花种植带来极大的经济效益,给环境带来生态效益。
2.DAS-21023-5 棉花对环境和人类健康无负面影响
本申报书对转化受体、基因操作、转化体本身、其棉花产品及外源基因或元
件的供体生物进行了综合评价,评估结果显示,DAS-21023-5 棉花和非转基因传
统棉花在环境安全方面实质等同,在食品安全方面实质等同,该转化体不会对动
物、植物、微生物及环境带来负面影响。评价结果简述如下。
分子特征方面:本转化体的分子特征清晰明确。1)转化质粒 pMYC3006 中
的 T-DNA 片段含有 cry1Ac 和 pat 两个基因表达盒。外源基因整合至第 8 染色
体,外源片段全序列及旁侧序列明确,DAS-21023-5 棉花基因组中有结构完整、
单一拷贝、单一插入位点的、来源于转化质粒的 T-DNA 片段,且转化体基因组
中未整合质粒的骨架序列(因此不含抗生素抗性基因)。2)转化体上除 T-DNA
整合造成了 16 bp 删除(无明确生物学功能)以外,无其它意外插入及意外删除。
遗传转化未在接合区引起表达新的致敏、致敏蛋白质。3)转化载体上各遗传元
件安全,演变出致病性的可能性极低。4)从大田取样,采用 ELISA 方法对不同
时期、不同组织中外源蛋白质进行定量,Cry1Ac 蛋白质在棉花植株上除了蜜腺
以外的各组织中(种子、叶、花、花蕾、花粉、棉铃、根和全株)均有不同水平
的表达,在籽粒中 Cry1Ac 表达量为 0.57 ng/mg 组织干重;PAT 蛋白质在各组织
器官中表达量普遍低,几乎检测不到,在籽粒中 PAT 表达量为 0.06 ng/mg 组织
干重。
遗传稳定性方面:跟踪检测了 DAS-21023-5 棉花代际间在 DNA 整合水平、

5
蛋白质表达水平和性状表现水平的遗传稳定性。采用 Southern blot 验证了 BC1F3、
BC3F3、BC3F4 和 BC4F1 四个世代植株的外源基因均为单位点且单拷贝插入、表
达框结构均完整且转化体不含载体骨架序列; ELISA 检测显示 BC4F2、F2 和
BC3F6 三个世代中 Cry1Ac 蛋白质在种子、根和叶片中表达量相对稳定,PAT 亦
然;2001-2004 年进行了多年、多代、多地点的田间试验,结果显示本转化体代
际间对鳞翅目昆虫表现出稳定的防治效果。
并且,本公司还检测了同一世代内(BC3F2 群体)个体间的稳定性,即使用
蛋白侧向层析试纸条和 DAS-21023-5 转化事件特异性 PCR 方法,检测了外源基
因在该群体的分离情况。结果显示,表达与不表达 Cry1Ac 蛋白质的个体比例与
3:1 无显著差异,含与不含 cry1Ac 基因的个体比例与 3:1 无显著差异,即插入基
因分离比例与预期的孟德尔单一位点分离模式一致,表明所插入外源基因可稳定
遗传。
环境安全性方面:在生存竞争能力方面、基因漂移方面、对非靶标生物及生
物多样性影响方面,转基因棉花 DAS-21023-5 均与非转基因棉花实质等同。
1)2002 年在美国产棉区内 20 个试验地点进行了大规模田间试验,转基因
棉花 DAS-21023-5 与非转基因棉花相比,种子的休眠和活力、传播能力、生长发
育、生殖率和产量等各项农艺学性状均等同,抗虫性状的引入没有改变该转化体
生存竞争能力,也未提高其杂草化倾向和侵蚀自然生态环境的能力。
2)基因漂移带来的环境风险极低。基因垂直漂移方面,DAS-21023-5 花粉
与非转基因对照花粉特征等同,因此二者向其它种属授粉的概率等同。且由于棉
花以自花授粉为主,,因生殖隔离及地理隔离等因素限制了 DAS-21023-5 棉花的
外源基因向其它属、种、品种漂移的概率极低,演变成杂草的可能性与非转基因
的棉花等同。基因水产漂移方面,遗传物质从植物向微生物水平转移概率极其微
小,迄今尚未见棉花的遗传物质向其它微生物转移的报导。另外本转化体中插入
的外源基因均来源于自然界中广泛存在,而且没有任何关于这些基因对环境的不
利影响报道,所以即便有基因漂移也不会对环境安全造成影响。
3)2001 至 2004 年间多年、多地点的田间试验和实验室研验证了 DAS-21023-
5 棉花可持续稳定地抗虫。
4)DAS-21023-5 棉花对生物多样性无负面影响。首先表现在该转化体与其
非转基因对照对土壤微生物的影响一致;且对非靶标生物如植食性生物(蚜虫)、
天敌(草蛉和寄生蜂)、禽类(美洲鹌鹑)、土壤生物(蚯蚓和跳虫)、水生生物
(大型溞和虹鳟鱼)、访花昆虫(蜜蜂)及哺乳动物(小鼠)无不利的影响,也
不会威胁到濒临灭绝的物种和有益昆虫。转基因棉花表达的 Cry1Ac 蛋白质在土
壤中的半衰期为 1.3 天,不会蓄积。
食品安全性方面:转基因棉花 DAS-21023-5 及其表达的外源蛋白质在食用
安全方面与非转基因传统棉花等同。二者营养成分无差异,DAS-21023-5 棉花

6
及其表达的外源蛋白质无潜在的毒性和致敏性,遗传操作未引入抗生素抗性基
因,外源蛋白质在人类膳食中的接触风险极低,加工过程不影响 DAS-21023-5
棉花的安全性,且未发现非预期效应。
1)外源蛋白质 Cry1Ac 以及 PAT 均来自安全的供体生物并且有长期的安全
应用历史;这两个蛋白分别存在许多商业化种植的转基因作物中,具有长期安全
使用历史;田间和实验室生物测定未发现其对非靶标生物有毒性;与已知毒性蛋
白质、已知抗营养因子、已知过敏原的氨基酸序列无较高的相似性;外源蛋白质
受热不稳定,在模拟胃液中易消化,因此不具备致毒和致敏物质的特征;对哺乳
动物无经口急性毒性,Cry1Ac 和 PAT 的 LD50均大于 5050 mg/(kgBW);采用极
为保守的方法估计,Cry1Ac 和 PAT 对中国人群膳食风险的接触界限 MoE 分别
达 108 和 109 量级,即接触风险极低。将含有 DAS-21023-5 转化体的棉粕掺入大
鼠基础饲料中(10%)饲喂大鼠 90 天,未见亚慢性急性毒性反应。
2)在大田取样,检测棉花植株、棉籽及其加工产品(棉仁、棉籽壳、熟棉
粕和精炼棉籽油)中的各类营养成分和抗营养因子的含量,各成分含量在 DAS-
21023-5 棉花和非转基因对照间或差别微小,或与文献报导值相近、或在文献报
导值范围内,即二者营养成分和抗营养因子实质等同。将含有 DAS-21023-5 转化
体的棉粕掺入肉鸡基础饲料中,饲喂肉鸡 42 天,未见实验动物营养学不良反应。
上述 90 天大鼠实验亦未见该转化体对实验动物带来营养学不良反应。因此,转
基因棉花及其加工品在营养成分及抗营养因子方面和非转基因传统棉花是实质
等同的,踪传操作未改变棉花的营养成分。
3)T-DNA 上无抗生素抗性基因,Southern blot 显示 DAS-21023-5 棉花基因
组中无载体骨架序列,即未整合抗生素抗性基因。
4)DAS-21023-5 的目标性状是抗特定的鳞翅目害虫。虽然其含有 pat 基因,
但是此功能仅用做遗传转化的选择性标记。一般不用于大田喷洒草铵膦。即便如
此,本公司仍对含有 DAS-21023-5 的转基因棉花进行除草剂残留检测。检测结果
显示,草铵膦及其代谢物在棉籽中的残留量为 0.042 µg/g,该值远低于美国 EPA
界定的草铵膦在棉花最大残留限量 4.0 µg/g,即本转化体不会带来农药残留危害。
3.DAS-21023-5 棉花在其它国家的审批状况
基于上述安全性评估,本公司向世界多个国家提交了种植、食用、饲用的申
请。自 2004 年起 DAS-21023-5 棉花陆续获得美国农业部(USDA)、美国环境
保护署(EPA)、美国食品与药物管理局(FDA)、欧盟委员会(EC)、巴西国
家生物安全技术委员会(CTNBio)、加拿大卫生部 (Health Canada)、加拿大
食品监督局(CFIA)、澳大利亚和新西兰食品标准局(FSANZ)、日本厚生劳动
省(MHLW)、日本农林水产省(MAFF)、日本农林水产省和环境省(MAFF
Env)、韩国食品药品管理局(MFDS)、韩国农村振兴厅(KRDA)、哥伦比亚
农业研究所(ICA)、哥伦比亚卫生和社会保护部(INVIMA)、南非农林渔业部
(DAFF)及中国台湾地区(TFDA,COA)等机构批准,并以相应用途投放市场。

7
含有本转化体的复合性状棉花自 2005 年开始在美国及其它国家种植,迄今未见
其对人类、动物、植物、微生物和环境带来任何不利影响的报导。
4. 小结
基于对 DAS-21023-5 棉花科学的安全评估结果以及全球审批情况,本公司
向中国农业农村部申请转基因抗虫棉花 DAS-21023-5 用作加工原料的安全证书,
盼农业农村部予以审核批准。

8
三、工作目的和意义
目的
棉花是重要的经济作物,可生产棉纤维、棉籽油、棉粕等产品,广泛用于被
服、食品、饲料、军工及医药等行业。世界各地棉花消费与生产的不对称性促使
世界棉花贸易蓬勃发展,棉产品从产量丰富的国家和地区流向需求巨大的国家和
地区。
世界主要棉花出口国有美国、印度、巴西、澳大利亚、乌兹别克斯坦等,
主要进口国有中国、土耳其和孟加拉国等。2012/13、2013/14、2014/15 年度,
世界棉花生产总量依次为 2697.6、2621.4 和 2593.1 万吨,中国为 762.0、714.1
和 653.2 万吨;2014/15 年度,印度、中国和美国产量分别为 574.8、653.2 和
354.9 万吨;2014/15 年度全球出口总量为 764.2 万吨,其中美国出口量为 243.9
万吨,印度出口量为 91.4 万吨,澳大利亚出口量为 52.3 万吨
(http://www.cottoninc.com/corporate/Market-Data/MonthlyEconomicLetter/pdfs/English-pdf-charts-and-tables/World-Cotton-Production-Bales.pdf)。2016-2020 年中国棉花进口量呈现逐年增加的趋势。随
着近年来国内棉花产量的下降,储备棉库存的减少,国内棉花存在明显的供需
缺口,所以中国棉花进口量逐年增加。2020 年棉花进口量在 215.45 万吨
(texnet.com.cn)。
本申报书的目的是申请转基因抗虫棉花 DAS-21023-5 用作加工原料的安全
证书,以便实现中国及出口国间 DAS-21023-5 棉花的国际贸易。
意义
DAS-21023-5 棉花能够表达杀虫晶体蛋白质 Cry1Ac,该蛋白质赋予 DAS-
21023-5 棉花对多种鳞翅目害虫的抗性,可以显著减少了杀虫剂的使用,不仅为
棉农带来巨大的经济效益,而且在全球范围内带来显著的社会经济效益和生态效
益。
1)社会经济效益:一方面,该抗虫棉花显著地减少了传统喷施杀虫剂所需
要的劳动力、时间、投入和设施,也减少人畜中毒事件,从而提高了生产耕作效
率并加强了生产和环境安全。另一方面,与外用杀虫剂相比,DAS-21023-5 棉花
持久表达抗虫蛋白质,抗虫性特异高效且衡定,不存在雨水冲洗和被风吹散农药
的问题,易于管理。由于减少投入且增加产出,种植者及消费者获得了更大的经
济效益。
2)生态效益:DAS-21023-5 棉花对靶标害虫具有高度的专一性,仅对靶标
害虫有效,不作用于非靶标生物;种植 DAS-21023-5 棉花可减少杀虫剂的喷施,
降低了对农田其它昆虫的影响,维护了生物多样性,从而保护了生态环境。

9
四、国内外研究的相关背景材料
自 1996 年转基因作物大规模商业化以来,转基因作物的种植已经超过了二
十年。有 26 个国家种植了转基因作物,44 个国家进口转基因作物。1996 至 2018
年间,全球转基因作物累计种植面积达到 25 亿公顷。二十多年的商业化证明,
转基因作物为农民乃至为全社会带来了农业、环境、经济、健康和社会效益
(ISAAA, 2018)。
转 Bt 基因抗虫棉花的研究与应用进展
转基因棉花 DAS-21023-5 中含有的 cry1Ac 基因来自苏云金芽孢杆菌
(Bacillus thuringiensis , Bt),Bt 菌广泛存在于土壤中。Bt 菌分布极广且能形成
芽孢。Bt 菌与常用的化学杀虫剂的杀虫机理不同,化学杀虫剂多为神经触杀性杀
虫剂,而Bt菌会产生一种Bt杀虫晶体蛋白质或称 δ-内毒素,包括Cry(Crystalline)
和 Cyt(cytolytic)两大类蛋白质(Schnepf et al., 1998)。其中,Cry1 类蛋白质约
130~140 kD,它进入敏感昆虫的中肠后,被胰蛋白酶消化,激活成具有杀虫活
性的核心毒素,该核心毒素可与敏感昆虫中肠的上皮细胞纹缘膜上的特异性受体
结合,使其构象发生改变后插入到细胞膜,致使细胞穿孔,从而使消化道细胞渗
透压失衡,引起细胞膨胀甚至裂解,导致消化道内物质流入淋巴,最终致使昆虫
死亡(Bravo, 1997; Glare 和 O'Callaghan, 2000b)。Bt 杀虫晶体蛋白具有极高的特
异性,比如 Cry1 类蛋白只对鳞翅目的少数几种昆虫起作用,对其他昆虫、节肢
动物、脊椎动物、禽类、水生类动物和人类无毒性。Bt 广泛存在于自然界,在害
虫防治中应用了几十年。已注册的 Bt 微生物农药对哺乳动物无毒,对非靶标生
物的毒性极微,且它在环境中的残留期较短(OECD, 2007)。
通过现代生物技术将编码 Bt 蛋白质的基因转化至植物中,Bt 蛋白质在植物
中持续表达,可使转基因植物免受靶标昆虫的危害,从而不再喷施或少喷施 Bt
微生物农药。试验表明,在长期栽培转 Bt 基因植物的土壤中检测不到 Bt 蛋白质
残留,因此即使长期积累也不会在土壤中产生毒性作用(OECD, 2007)。1986 年
美国环境保护署(EPA)首次批准了种植转 Bt 基因的烟草,截止 2014 年 4 月,
国际生命科学研究所(International Life Science Institute,ILSI)网站公布的全球
商业化转 Bt 基因作物(单价或多价)达上百种,涉及棉花、大豆、玉米、番茄
和马铃薯。人们不断发现新的 Bt 基因并加以利用,至今共命名了 300 多个 Bt 基
因(http://www.btnomenclature.info/)。其中 cry1F、cry1Ac、cry1B、cry1A10、
cry35Ab1/cry35Ab1、cry2Ab、cry2Ab2、cry3A、cry3Bb1 及 vip 已在全球商业化
推广应用(http://cera-gmc.org/index.php?action=gm_crop_database)。转 Bt 基因植
物的推广为植物虫害治理提供了有效方法,为保护生态环境、实现农业可持续发
展作出了积极的贡献。
转 Bt 基因作物的安全性评价
转 Bt 基因作物推广数十年的历史也伴随着对其安全性进行评价和研究的历

10
史。
1)毒理学方面,建立了苏云金芽孢杆菌的毒理学数据库,各项数据均表明
Bt 蛋白质对哺乳动物健康无负面影响。例如分别用 Cry1F、Cry1Ac、Cry2Ab2 及
Cry34Ab1/Cry35Ab1 蛋白质以 3760~5220 mg/(kgBW)高剂量经口灌胃小鼠,均
未出现急性毒性反应。此方法验证了转基因玉米 TC1507 和大豆 DAS-81419-2 中
的 Cry1F 蛋白质对哺乳动物无经口急性毒性(EFSA, 2005)。基于 Bt 蛋白质的安
全性,其残留不再属于监管范围,美国环保署(EPA)依据 Cry1F/Cry1Ac 蛋白质
的安全性综合评估结果(包括模拟胃液消化试验、与已知致敏原及毒性蛋白质序
列相似性比较、对哺乳动物有无毒性)明确了该蛋白质的安全性,并于 2004 年
最终决定不再对大豆、棉花和玉米中的 Cry1F/Cry1Ac 残留进行监管(40 CFR
§174.504)。
2)Bt 蛋白质食用及饲用安全性方面,迄今为止所测定的 Bt 蛋白质与已知
过敏原蛋白的序列无较高的相似性,且 Cry1F、Cry1Ac、Cry2Ab2、Cry34Ab1 及
Cry35Ab1 等蛋白质受热后不稳定;几乎所有商品化的 Bt 蛋白质均易在模拟消化
液中迅速降解。如转基因玉米 TC1507 和大豆 DAS-81419-2 中的 Cry1F 具有热不
稳定性且在模拟消化液中易失活,即不具备致敏物质热稳定和不易消化的特征
(OECD, 2007)。
3)转 Bt 基因植物在环境安全方面的研究评价也取得了显著成就。转基因与
非转基因对照棉花农艺学特性等同,对非靶标生物(哺乳动物、鸟类、淡水鱼、
淡水中无脊椎动物、河生和海生动物、蚯蚓及非靶标昆虫)的影响等同(OECD,
2007)。
4)在抗性持久性方面,目前主要的抗性治理策略有高剂量和庇护所策略、
新毒素策略和转多价基因策略(Bates et al., 2005)。Cry1F、Cry2A 及 Vip 类为第
二代 Bt 蛋白质,在增强杀虫活性、抗性治理、扩大杀虫谱等方面起到了重要的
作用(Gouffon et al., 2011;Yu et al., 2011)。目前已经商业化生产的转基因棉花
中,双价 Bt 基因组合主要有 cry1F/cry1A、cry1Ac/cry2Ab 和 Vip3A/cry1Ab 等(Yu
et al., 2011),双价抗虫棉有效减缓了靶标昆虫的抗性演化。经十余年的跟踪记录,
中国农业科学院科学家研究了棉铃虫种群地位演化与 Bt 棉花的关系,结果表明
Bt 棉花的大规模商业化种植破坏了棉铃虫在华北地区季节性多寄主转换的食物
链,压缩了棉铃虫的生态地位,不仅有效控制了棉铃虫对棉花的危害,而且高度
抑制了棉铃虫在玉米、大豆、花生和蔬菜等其它作物田间的发生与危害。并为转
基因抗虫作物对靶标害虫种群演化的调控机理提供了理论基础,为棉铃虫的可持
续控制奠定了基础(Wu K,2010)。
综上所述,转基因棉花已在全球广泛推广数十年,人们对 Bt 基因具有深入
的了解,且不断开发新的 Bt 基因家族成员并加以利用,转 Bt 基因棉花在全球广
泛种植和消费,人们对抗虫棉花的环境及食用安全性进行了科学的评价,积累了
丰富的经验。

11
五、安全性评价
1 受体植物的安全性评价
1.1 受体植物的背景资料:
1.1.1 学名、俗名和其他名称;
学名:陆地棉
俗名:棉花
1.1.2 分类学地位;
科:锦葵科(Malvaceae)
属:棉属(Gossypium)
种:陆地棉(Gossypium hirsutum L.)
1.1.3 试验用受体植物品种(或品系)名称;
陆地棉品种 GC510。
1.1.4 是野生种还是栽培种;
陆地棉品种 GC510 是栽培种。
1.1.5 原产地及引进时间;
起源中心: 从干旱到半干旱的热带和亚热带地区,棉属有多个起源中心,生
物多样性中心代表有非洲、阿拉伯半岛、澳大利亚和墨西哥。棉属大约有 50 多
个种(OECD,2008),其中有四个栽培种:两个是二倍体棉种,分别为非洲棉
(G.herbaceum L.,2n=26)和亚洲棉(G. arboretum L.,2n=26);两个是四倍体
棉种,分别为陆地棉(G. hirsutum L.,2n=52)和海岛棉(G. barbadense L.,2n=52)。
在以上 4 个栽培种中,陆地棉和海岛棉占棉花总产量的 95%(OECD, 2008)。
陆地棉及其衍生品种是世界棉纺工业的主要来源,普遍认为墨西哥和危地马拉接
壤的边境是陆地棉的起源中心与生物多样性中心。陆地棉起源于 100 万至 200 万
年前 A 染色体组二倍体种和 D 染色体组二倍体异交后进化的 4 倍体后代(Wendel
et al., 1992;Senchina et al., 2003;Wendel and Cronn, 2003;Kebede et al., 2007)。
经过人类长期栽培驯化,陆地棉逐渐成为世界范围栽培最广泛的棉种,并己经成
为典型的一年生且高度栽培化的纤维植物(Brubaker et al., 1999;Fryxell, 1979,
1984;叶国祥, 2011)。
陆地棉引进中国的时间:中国具有悠久的植棉历史,据文献记载和文物考证,
早在 2000 多年前中国南部沿海岛屿,福建和新疆等地已经开始种植棉花。约公
元 6、7 世纪,其它地方也开始种植棉花。公元 13 世纪以后,棉花成为重要的经

12
济作物。
1865 年中国开始引种陆地棉栽培种,最先从美国引入中国上海试种,1892、
1893 年又从美国选购两个品种在湖北省天门、孝感、武昌等州县广为试种,以后
又多次从美国、前苏联等国引入陆地棉品种在主要棉区试种。其中脱字棉、隆字
棉、斯字棉 4 号、德字棉 531、河字棉 100、斯字棉 2B、伤字棉 15 等在全国大
面积种植。至今,陆地棉已是中国主要的栽培棉种(叶国祥, 2011)。
1.1.6 用途;
棉花是重要的经济作物之一,其长绒、棉籽油、短绒、棉籽壳、棉杆和棉酚
等广泛应用于纺织、食品、饲料、军工及医药等领域。
首先,棉纤维成品含纤维素达 99%,棉花是世界上天然纤维的主要来源。其
纤维可作纺织工业的原材料,用作被服、军用帘子布。它具有吸湿、透气、保暖、
不产生静电、手感柔软等人造纤维难以取代的特点,是全世界人们衣着的主要原
料之一。低品质的纤维可以用来制造毛毡、床垫填充物和特殊用途的纸张,纤维
素经加工可以用于各种日用消费品,如牙膏、唇彩、冰淇淋和蛋黄酱。
棉籽含油份 14~18%。棉籽油在润滑油和生物燃料领域有广泛的应用前
景,脱酚的棉籽油是品质较高的食用油。
另外,棉籽壳可用于牛、羊粗饲料,是重要的饲料成分。
棉酚可作为男性避孕剂(Coutinho, 2002;OECD, 2009)。
1.1.7 在国内的应用情况;
陆地棉是中国的主要栽培棉种,除新疆地区种植部分海岛棉外,其余棉区均
种植陆地棉。
生产情况:中国是植棉大国,棉花是仅次于粮食的大宗农产品和最重要的经
济作物。植棉业是我国农业的支柱性产业,关系国计民生。自 20 世纪 80 年代中
期以来,我国成为世界上最大的棉花生产国、消费国和进口国之一,总产和单产
均居世界前列。2015/16 年,中国的棉花种植面积为 340 万公顷,产量达 1,524
kg/ha(USDA Cotton Outlook, 2016, www.usda.gov/oce/forum)。
我国棉花种植主要分布在黄淮流域、长江流域和西北内陆这三大主产区,其
中长江流域棉区占全国种植面积的 25%,占总产量的 22%;黄河流域棉区占全国
种植面积的 40%,占总产量的 37%;西北内陆棉区占全国种植面积的 35%,占
总产量的 41%(喻树迅, 2013)。据农业农村部调查统计,在我国黄淮棉区和长江
棉区,棉花收入占植棉农户总收入的 60%左右,新疆棉区植棉农户收入约 80%来
自棉花。
消费情况:自 20 世纪 60 年代末 70 年代初以来,中国棉花年均消费量达 200
万吨,此后中国一直居于世界棉花消费国首位。加入 WTO 以后,中国棉花消费

13
受纺纱业发展的影响,呈快速增长态势。据美国农业部统计,1981~1998 年,世
界棉花年均消费量为 1764 万吨,中国年均棉花消费量为 415 万吨,占世界消费
总量的 23.5%;2000 年以后,国内棉花需求进一步提高,2002~2006 年中国棉
花消费量增长速度最快,平均增速高达 13.47%;2007~2011 年由于金融危机爆
发,棉花消费冲高回落,年均下降 3.78%,但仍然是世界最大棉花消费国;截止
2011 年,棉花消费量占世界棉花消费总量的 39.0%。
在我国,棉纤维主要作为纺织工业用棉、军需和民用絮棉及其他用棉,其中
最主要的消费方式是纺纱用棉。据全国供销总社棉麻局的纺织工业数据显示:20
世纪 80 年代,纺纱用棉与军需和民用絮棉年均消费量分别占棉纤维消费比例的
85%和 13%,90 年代两者分别为 90%和 6%,21 世纪后两者分别约 95%和 2.5%
(冯献和詹玲, 2012)。
1.1.8 对人类健康和生态环境是否发生过不利影响;
全世界约有 75 个产棉国家,分布在南纬 32°到北纬 47°之间,棉花用于生产
纤维、脱酚棉籽油、棉籽蛋白质,具有长期安全使用和食用的历史,未见对人类
健康和生态环境发生过不利影响。棉花中的棉酚对非反刍类动物有一定的毒性,
但是食用棉籽油均经过脱酚处理后,其棉酚结构遭到破坏,对人类无毒无副作用
(OECD, 2009)。
棉花中没有已知的蛋白质过敏原。加工的棉花纤维 99%以上是纤维素,由于
它的刺激作用低而广泛用于医药业(OECD, 2008)。
1.1.9 从历史上看,受体植物演变成有害植物(如杂草等)的可能性;
杂草常指对人类行为或利益有害或有干扰且无应用价值的植物。杂草往往生
长迅速并且具有强大的生存竞争力,从而影响作物生长(如抢夺光、水及营养),
给农业生产造成损失。杂草具有更强的竞争能力、较高结实能力、较强抗逆能力,
如种子休眠逃避逆境、种子在土壤中持续存活,生活周期短,种子繁殖率高,长
距离散播种子能力高等(崔荣荣 et al., 2012)。而现代的棉花栽培种不具备上述
杂草的特性。
栽培作物经人类千百年驯化而来,主要对所需要的性状如产量、品质及对常
见胁迫条件的耐受性进行了选择,因此理论上栽培作物在没有人类养护的环境中
难以产生自己的群落,即不具有竞争性和入侵性。棉花本身不属于杂草,不具有
杂草化倾向和侵蚀自然生态环境的能力。
转化受体棉花 GC510 是栽培品种,高度栽培化引起的形态学改变使其离开
人类的养护难以存活。在传统栽培条件下,栽培棉花主要以种子为越冬器官作为
一年生作物种植,而残留在田间未收获的棉杆在来年不再生。在适宜的地区,棉
花可在自然状态下生长,但是千百年的种植史中,尚未出现其演变为入侵杂草或
恶性杂草的报道(Holm et al., 1997;Randall, 2002;Weber, 2003)。

14
1.1.10 是否有长期安全应用的记录。
受体植物陆地棉具有长期安全应用的记录。
人类对棉花的应用历史渊源,早在公元前 5000 年之前,中美洲已开始利用
棉花,在南亚次大陆也有 5000 年安全应用的历史(晓婷, 2010)。在我国,早在
2000 年以前,广西、云南、新疆等地区已采用棉纤维作为纺织原料。《梁书·高昌
传》对棉花的记载:其地有“草,实如茧,茧中丝如细纩,名为白叠子。”从明
代宋应星的《天工开物》中所记载的“棉布寸土皆有”,“织机十室必有”,可知
当时植棉和棉纺织已遍布全国。
长久以来,棉花应用于多种领域,未见棉花及其加工产品危害人畜健康及环
境安全的记录。
1.2 受体植物的生物学特性:
1.2.1 是一年生还是多年生;
转化受体 GC510 陆地棉为一年生作物。
1.2.2 对人及其他生物是否有毒,如有毒,应说明毒性存在的部位及
其毒性的性质;
棉花籽粒中含有抗营养物质或毒性物质,大量摄入会产生毒害,如棉酚(一
种萜烯醛类物质)、环丙烯脂肪酸和单宁酸。
棉酚最先在根、叶、花蕾和种子的色素腺体中发现,它对非反刍动物、鸟类、
昆虫和微生物具有毒性,可引起哺乳动物食欲降低、体重减轻、呼吸困难。棉酚
有两种不同的异构体,这两种异构体通常以游离态或结合态的形式存在。完整的
棉籽中通常存在游离态的棉子酚,在反刍动物的瘤胃中微生物群的作用下,游离
态的棉子酚转化成结合态的棉子酚,从而阻止其进入血管。在整个棉籽的加工过
程中,棉子酚进入棉籽油组分或粗碾组分,继而棉籽油脱酚后可安全食用;在粗
碾的组分中,棉子酚通常与蛋白质结合,其毒性很小(Santos et al., 2002)。酚失
去活性后或脱酚后,棉籽可用来饲喂猪、家禽和鲶鱼(Jones and Wedegaertner,
1985)。
环丙烯脂肪酸存在于棉籽中,是一种抗营养化合物,主要干扰饱和脂肪的代
谢,在棉籽油加工过程中(如人造奶油和色拉油),环丙烯脂肪酸被破坏,不会
引起人、畜毒性反应(OECD, 2008)。
在加工过程中,棉酚被脱去;经压榨和烘烤等高温处理,抗营养因子的结构
或功能破坏,因此不会因为棉花含上述抗营养因子引起人、畜、禽不良反应
(OECD, 2008)。
1.2.3 是否有致敏原,如有,应说明致敏原存在的部位及其致敏的特

15
性;
未见转化受体(陆地棉)含致敏原的报导(OECD, 2008)。
1.2.4 繁殖方式是有性繁殖还是无性繁殖,如为有性繁殖,是自花授
粉还是异花授粉或常异花授粉;是虫媒传粉还是风媒传粉;
陆地棉属于有性繁殖植物,通过授粉、受精、结实的方式进行繁殖。
在自然环境中棉花以自花授粉为主。有适宜的昆虫传粉时棉花可以进行异花
授粉。由于其花粉粒大、沉重且具有粘性,不适合风媒传播(McGregor, 1976;
Umbeck et al., 1991)。
1.2.5 在自然条件下与同种或近缘种的异交率;
棉花主要以自花授粉为主,同种内品种间可异交,并产生正常可育的后代。
在传粉昆虫活跃时,存在低水平的异花授粉。离花粉源越远,异交率越低,花粉
源 10 米处异交率小于 1%(OECD,2008)。
1.2.6 育性(可育还是不育,育性高低,如果不育,应说明属何种不
育类型);
转化受体为陆地棉品种 GC510,充分可育。
1.2.7 全生育期;
栽培棉花的全生育期包括播种、出苗、现蕾、开花和结实。陆地棉的正常发
育需要 180~200 天无霜期,150 天适宜温度(>15.5℃积温 1200 热量单位)。几
个生育期的基本特征如下。
1)播种到出苗:即从棉花播种到 50%的棉株第一片真叶展开。不同品种从
播种到出苗所需的时间基本相同。出苗快慢不受光照时长影响,而是取决于地温。
播种到出苗可分为三个生物学阶段,即种子萌发(5 cm 最低地温为 10℃),胚根
发生(5 cm 最低地温为 15℃)和子叶下轴伸长(最低温度为 15℃)。
2)出苗到现蕾:即从棉花第一片真叶展开到田间 50%的棉株第一果枝第一
果节现蕾。所需最低温度为 12℃,最适温度为 30℃,最高温度为 35℃。不同品
种所需时间有一定差异,取决于温度和日照时长的共同作用。
3)现蕾到开花:即从现蕾到田间 50%棉株第一果枝第一果节棉蕾开花。发
育最低温度为 12℃,最适温度为 30℃,最高温度为 35℃。多数棉花品种对光周
期反应不敏感。
4)开花到吐絮:即从开花期至田间 50%棉株吐絮。不同品种完成这一生育
阶段所需的生理发育时间有一定差异,主要取决于温度和品种的熟期。发育所需
最低温度为 12℃,最适温度为 30℃,最高温度为 35℃(张立祯和曹卫星, 2003)。

16
1.2.8 在自然界中生存繁殖的能力,包括越冬性、越夏性及抗逆性等。
栽培棉花在养护下生长,生长环境水肥充足,无杂草争夺光、养及水份,因
此高度驯化的栽培品种在自然界中适应能力、抗生物逆境(病虫害,杂草等)和
非生物逆境(旱、高温、低温、盐碱等)的能力降低。无人类养护时,栽培棉花
难以象野生棉种、近缘野生种及杂草一样在自然界中竞争。
棉花种子是其越冬器官。棉花种子有一定的休眠期(植物种子可以通过休眠
躲避逆境),但是通过驯化和选择育种,现代栽培品种的休眠特性已经降到了最
短或完全丧失。
夏天严重干旱会影响栽培棉花的开花结铃,从而影响种子活力(OECD,
2008)。
1.3 受体植物的生态环境:
1.3.1 在国内的地理分布和自然生境;
中国位于世界上最大的大陆—欧亚大陆的东南部,且东临世界上最大的海洋
—太平洋,气侯受海洋和陆地影响均非常明显。我国的棉区从东到西分属东部季
风区和西北干旱区。东部季风区分属北热带、北亚热带、中亚热带、南亚热带、
南温带和中温带。大体上,热量资源由南向北逐渐降低,降水量从东南向西北逐
渐减少,存在明显的地域性差异。根据积温多寡、纬度高低、降水量多少等自然
生态条件,将我国植棉区由南而北、从东向西依次划分为华南棉区(只有零星种
植)、长江流域棉区、黄河流域棉区、北部特早熟棉区(已缩减)和西北内陆棉
区共五大棉区(图 1)。
这五大棉区自南向北分布,热量、水分资源依次递减,各区之间生态类型、
耕作栽培特点、主要病、虫害发生与危害程度方面均呈现规律的变化。生产上常
把华南棉区和长江流域棉区统称为南方棉区;把黄河流域棉区、辽河流域棉区以
及西北内陆棉区统称为北方棉区(张海清, 2004)。
我国棉区北起新疆北部的玛纳斯河流域,南至海南岛,西起新疆的喀什地区,
东抵长江三角洲的沿海地带和东北的辽河流域(北纬 18°~46°,东经 76°~124°)
均有棉花种植。近年,全国有产棉省(市、自治区)24 个,棉田面积超过 40 万
公顷的有 7 个(新疆、河南、江苏、湖北、山东、河北和安徽),这 7 个省区的
棉花占全国种植面积的 70%,占总产量的 80%,其中新疆为特大产棉区;棉田面
积超过 10 万公顷的有 4 个省(湖南、江西、四川和山西);棉田面积超过 5 万公
顷的有 2 个省(浙江和陕西);棉田面积约 l 万公顷的有 3 个(天津、辽宁和甘
肃);分散产区有 8 个(重庆、广西、云南、贵州、内蒙古、吉林、北京和海南)。
1.3.2 生长发育所要求的生态环境条件,包括自然条件和栽培条件的
改变对其地理分布区域和范围影响的可能性;

17
图 1. 中国棉区的地理分布(《中国农业区划的理论与实践》, 1993 )
http://www.zhongnong.com/BingHai/11502.html
生态环境中,光、温、水、肥及盐碱度等条件均影响棉花生长发育。
尽管棉花起源于热带和亚热带地区,但是最终大多数棉花栽培于亚热带和温
带地区(如中国)。影响棉花生长发育的气候因素中,温度是决定棉花种植分布
的主要因素。通常,低于 15℃时棉花种子不萌发,但是高于 38℃时种子的萌发
和出苗受阻。对陆地棉而言,白天最佳温度范围为 30℃~35℃,高于 35℃时成
铃减少,低于 25℃时产量减少 50%(Reddy et al., 1992)。陆地棉的正常发育需要
180~200 天无霜期,平均需要 150 天适宜温度,即 15.5℃以上积温 1200 热量单
位(Duke, 1983)。地温也是决定棉花生长和结实的因素,通常在土壤 10 cm 深处
至少 3 天温度超过 14℃时才能播种。
棉花在灌溉良好、富含有机质和保湿性良好的深耕土壤中长势更好。在贫瘠
的土地中,通过灌溉也可以栽培棉花,但需提供必要的湿度和营养。旱作棉花在
其生长期至少需要 500 mm 的降水量(OECD, 2008)。
根据土质不同,应在棉花植株的根部适量施加磷肥和钾肥,棉花对过量的氮
肥较为敏感,过量的氮肥能够促进棉花保持更长时间的营养生长而延迟成熟。适
量的生长调节剂和除草剂可用来控制棉花的生长发育,提高产量。

18
种子所处的周边环境也影响其生长繁衍。在棉田旁边的自生苗通常会被家畜
啃食或当作杂草除去,从而限制其发展成群落。曾在澳大利亚运输棉籽的沿途进
行了调查,发现该棉花不在路边繁衍,即使成簇发芽生长,也只是暂时性群体,
其群落存在时间不会超过 3 年(Addison et al., 2007)。
1.3.3 是否为生态环境中的组成部分;
作为主要的农作物之一,棉花是农业生态环境中的组成部分。
1.3.4 与生态系统中其他植物的生态关系,包括生态环境的改变对这
种(些)关系的影响以及是否会因此而产生或增加对人类健康和生态
环境的不利影响;
首先,棉花不寄生于其它植物。
其次,杂草与陆地棉栽培种有竞争关系。
另外,栽培棉花本身不属于杂草,不具有杂草化倾向和侵袭自然生态环境的
能力。栽培棉花经过人类长期的驯化,在没有人类养护的环境中难以产生并繁
衍自己的群落,即对生态环境无入侵性。
而且,由于遗传隔离、地理隔离及时间隔离,陆地棉栽培种与其它棉属、棉
种、野生棉种、近缘野生棉种及杂草的天然异交的可能性极低,即交换遗传物
质的可能性极低(OECD, 2008)。
总之,栽培棉花悠久的安全使用历史结合生态环境的变迁,至今未见其对人
类健康和生态环境带来不利影响。
1.3.5 与生态系统中其他生物(动物和微生物)的生态关系,包括生
态环境的改变对这种(些)关系的影响以及是否会因此而产生或增加
对人类健康或生态环境的不利影响。
除杂草外,棉花还与生态环境中的微生物、昆虫之间存在着各种形式的相互
联系,主要表现为棉花与微生物共生、受微生物感染、受昆虫为害方面。
有益微生物:棉花与丛枝菌根真菌(Glomus mosseae)间具有共生的关系
(Youssef and Mankarios, 1974;Nehl et al., 2004)。该真菌生长于棉花根部皮层细
胞之间,在此区域与棉花进行矿物质和碳水化合物交换。丛枝菌根真菌可使棉花
少受其它微生物侵染(Liu, 1995)。
致病微生物:棉花病害在世界各产棉国普遍发生,据统计棉花病害有 60 余
种,在我国已造成不同程度危害的有 23 种,其中常见的有 10 多种,如棉花枯萎
病和黄萎病(大丽轮枝菌感染所致)等。我国每年因病害损失皮棉约 36 万吨,
达 70 亿元(田逢秀, 1996)。
虫害:有害的节肢动物能够影响棉花生长、棉铃的纤维品质和棉花产量。蚜
虫(棉蚜、黑豆蚜、桃蚜)和烟粉虱是影响棉纤维品质的主要害虫,如果在生长

19
季节后期不防治,会产生带有黑色斑点的黏性纤维。粉螟、各种半翅目害虫(如
草盲蝽)及各种螨类(如二斑叶螨)等也能减少纤维产量、降低纤维品质。影响
棉铃产量的主要害虫有棉铃虫、大豆尺蠖、棉斑实蛾、甜菜夜蛾和棉铃象甲等。
生态环境的改变(如病、虫害发生)有可能影响栽培棉花的产量和品质,但
作为栽培作物的棉花,其产量和品质的变化不会产生或增加其对人类健康和生态
环境的不利影响。
1.3.6 对生态环境的影响及其潜在危险程度;
在中国,棉花属于重要的经济作物,已经种植了数千年,是广泛种植、食用
和饲用的栽培作物。生态环境在棉花悠长的栽培历史中几经变迁,未见迹象表明
栽培棉花对生态环境的不利影响。
1.3.7 涉及到国内非通常种植的植物物种时,应描述该植物的自然生
境和有关其天然捕食者、寄生物、竞争物和共生物的资料。
在中国境内,棉花已有数千年的栽培使用历史,2015 年中国棉花种植面积
约 380 万公顷。棉花是重要的经济栽培作物,不属于“非通常种植物种”。
另,本申报书仅申请转基因棉花 DAS-21023-5 用于加工原料,不用于种
植。
1.4 受体植物的遗传变异:
1.4.1 遗传稳定性;
在漫长的进化过程中,变异、遗传及地理隔离形成了稳定的物种。栽培种棉
花具有高度的遗传稳定性。
陆地棉栽培棉花是四倍体,与二倍体棉种存在生殖隔离,这两个棉种极难自
发异交并产生可育后代(OECD, 2008)。由于生殖隔离,陆地棉与杂草有性杂交
产生可育后代的可能性更低(Holm et al., 1979;Holm et al., 1997;Randall, 2002;
Weber, 2003)。
中国的植棉区主要种植陆地棉,这些区域均无陆地棉近缘的野生棉种,由于
地理隔离,难以发生种间杂交。在精密的实验条件下,在锦葵科内,棉属和其他
属杂交可结实,所结杂交种可育(Mehetre et al., 1980), 但是自然条件下自发的
属间杂交几乎难以发生。
由于生殖隔离、地理隔离及其它因素,棉属间、种间的遗传物质交换极难发
生,保障了棉种的遗传稳定性。
1.4.2 是否有发生遗传变异而对人类健康或生态环境产生不利影响的
资料;

20
棉花在全球范围内广泛种植,具有悠久的、安全的使用和食用历史。至今未
见棉花因发生遗传变异而对人类健康或生态环境产生不利影响的报道。
1.4.3 在自然条件下与其他植物种属进行遗传物质交换的可能性;
种间杂交:根据棉属细胞遗传学和种间杂交试验可产生存活或可育杂种的能
力,将棉种分为 8 个二倍体组和一个四倍体组。通常同一组的棉种之间可异交形
成正常的后代,即部分或全部 F1 可育。陆地棉与海岛棉属于四倍体种,它们之
间可以相互授、受粉,且后代可育。不同组间杂交难以结实,即使结实,减数分
裂可能异常,或 F1 代不育(OECD, 2008)。
与转化受体陆地棉亲缘关系较近的四倍体野生棉种有 3 个:黄褐棉(G.
mustelinum)、达尔文氏棉(G. darwinii)和夏威夷棉(G.tomentosum)。黄褐棉分
布于巴西东北部半干旱地区,是极为稀有的地方种(Freire et al., 1998;Batista et
al., 2005),与原始异源四倍体的祖先最为接近;达尔文氏棉是一种广泛分布在加
拉帕戈斯群岛的地方物种,与海岛棉亲缘关系最近(Wendel and Percy, 1990;
Lacape et al., 2007);夏威夷棉是夏威夷群岛的特有种,与陆地棉的亲缘关系最
近。理论上陆地棉与这些棉花种之间可杂交,且遗传重组频率较高。但是在自然
环境中,中国境内不生长上述 3 个野生棉种,转化受体陆地棉与这些野生棉种地
缘上彼此分隔,与这些四倍体棉花难以发生天然杂交(Brubaker and Wendel, 1994)。
与锦葵科其他属的杂交:在精密的实验条件下,棉属和锦葵科其它属杂交产
生可育杂种植株已有报道(Mehetre et al., 1980),但是由于生殖隔离(遗传关系
远),自发的属间杂交极难发生(OECD, 2008)。
1.4.4 在自然条件下与其他生物(例如微生物)进行遗传物质交换的
可能性。
基因从植物中水平转移到人类和动物或其肠道微生物的概率极其微小。
在精密的实验条件下通过现代生物技术可以将微生物中的遗传物质转化至
棉花基因组中。但反过来,尚未见遗传物质从植物直接转移至微生物和动物的现
象(Bertolla 和 Simonet, 1999;Nielsen et al., 2000)。据世界粮农组织/世界卫生
组织报导,没有任何证据证明植物的完整基因可自发转移到哺乳动物细胞中并在
其中表达(FAO/WHO, 2000)。
1.5 受体植物的监测方法和监控的可能性。
本受体棉花(陆地棉)为锦葵科棉属的栽培物种,在植物分类学上已有明确
的界定,适合目视监测和监控。如有逃逸现象,也很容易用物理或化学方法予以
清除或拔除。
1.6 受体植物的其他资料。

21
尚无其他有关的资料。
1.7 根据上述评价,参照本办法第十一条有关标准划分受体植物的安
全等级。
受体棉花具有悠久的栽培和应用历史,对人类健康和生态环境未曾发生过不
良影响,依据《农业转基因生物安全评价管理办法》第二章第十一条,其安全等
级为 I 级。
2 基因操作的安全性评价
2.1 转基因植物中引入或修饰性状和特性的叙述。
DAS-21023-5 棉花中引入了 cry1Ac 和 pat 基因,使其表现抗鳞翅目害虫和
耐受草铵膦两个新性状,对草铵膦的耐受性作为转化过程的选择标记。
Cry1Ac 蛋白质引入性状的作用机理(特征叙述):
cry1Ac 基因来自于广泛存在于自然界的土壤细菌苏云金芽孢杆菌(Bacillus
thuringiensis, Bt),该基因编码杀虫晶体蛋白质 Cry1Ac,Cry1Ac 可以保护植株
免受鳞翅目多种害虫的取食。
Cry1 类杀虫蛋白质一般以晶体形式存在。该类蛋白质的作用机理为:被敏
感昆虫摄食后,Cry1 蛋白质在强碱性的昆虫中肠(~pH 11)被蛋白酶(如胰蛋白
酶类)酶解,释放出具有杀虫活性的核心毒素,激活后的毒素通过其结构域 II、
III 的特定氨基酸与昆虫中肠上皮细胞膜的特异性受体(cadherin 类膜蛋白质)结
合,核心毒素 N-末端随之形成四聚体并插入细胞膜。毒素四聚体形成跨膜的离
子通道或非特异性小孔,从而导致昆虫中肠上皮细胞裂解和坏死,以致昆虫死亡
(Bravo et al., 2004;Vadlamudi et al., 1993; Gahan et al., 2001; Morin et al., 2003;
Coates et al., 2005; Yang et al., 2011)。通常,该类蛋白质仅对少数几种鳞翅目幼
虫发挥毒性。Cry1 蛋白具有较高的特异性, 由于缺乏类似于昆虫中肠的强碱性激
活环境,或无靶标昆虫中肠上皮细胞膜的特异性受体,,其他生物不在其杀虫谱
中。
PAT 蛋白质引入性状的作用机理(特征叙述):
pat 基因来自于土壤细菌绿色产色链霉菌(Streptomyces viridochromogenes),
编码膦丝菌素乙酰转移酶(phosphinothricin acetyltransferase,PAT)。表达 PAT
的棉花可耐受草铵膦除草剂,pat 作为棉花 DAS-21023-5 转化体开发过程中的选
择标记基因。
PAT 的作用机理:草铵膦的活性成分左旋膦丝菌素(L-PPT)是植物体中谷
氨酸盐合成酶(GS)的潜在抑制剂,因此,PPT 可以用作非选择性的除草剂(Bayer

22
et al., 1972;Devine et al., 1993;Shah et al., 1986;Kishore and Shah, 1988)。PPT
对 GS 的抑制作用会引起细胞内氨的快速积累,从而引起光呼吸终断,最终导致
植物细胞死亡。植物体表达 PAT 可以催化乙酰 CoA 的乙酰基转移至 PPT 生成
乙酰化草铵膦和 CoASH(图 2),从而使草铵膦类除草剂失去活性,使植物体免
受伤害。
PAT 蛋白酶的理化特征及酶催化特征比较清晰。米氏动力学分析显示,PAT
蛋白质在 pH 6~11 时对底物草铵膦均有较高的特异性(刘洪艳,2007)。PAT 酶
活性受温度影响,当温度≥55℃时,PAT 丧失至少 99% 酶活性。
图 2. PAT 蛋白质的抗除草剂作用原理(Duke, 1996)
PAT 催化乙酰 CoA 的乙酰基转移至 PPT 生成乙酰化草铵膦和 CoASH,从而使 PPT 失去活
性。
2.2 实际插入或删除序列的以下资料:
2.2.1 插入序列的大小和结构,确定其特性的分析方法;
插入序列的大小和结构
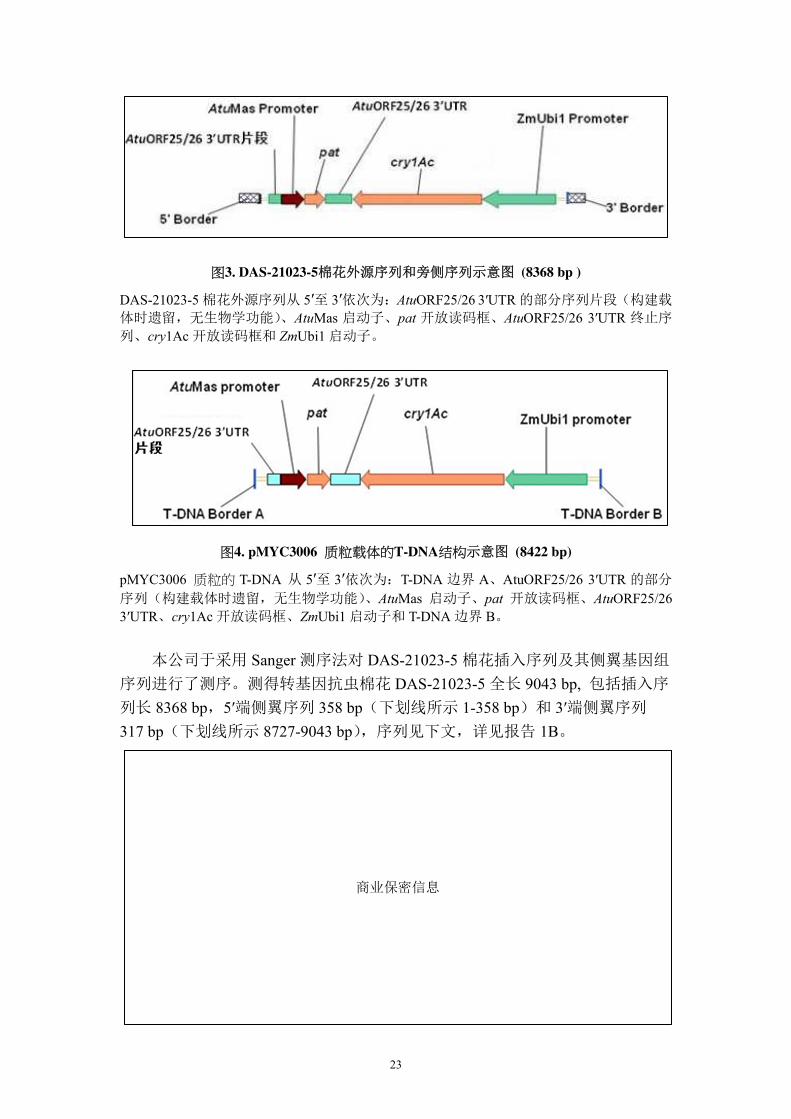
DAS-21023-5 棉花实际外源序列结构见图 3,外源片段全长 8368 bp (报告 1B),
从 5′至 3′依次为: AtuORF25/26 3′UTR 片段(构建载体时遗留,无功能)、AtuMas
启动子、pat 开放读码框、AtuORF25/26 3′UTR、cry1Ac 开放读码框和 ZmUbi1 启
动子。pat 与 cry1Ac 表达盒尾尾相连,共用一个双向终止子。pat 和 cry1Ac 表
达盒的结构与 T-DNA 上的结构一致(图 4)。DNA 测序结果显示转化体上外源
序列与载体上的一致,转化体上各遗传元件大小见表 1。
DAS-21023-5 棉花中未整合质粒 pMYC3006 的骨架序列
以覆盖整个 pMYC3006 质粒骨架区域的 4 个探针与内切酶消化的 DAS-
21023-5 基因组 DNA 进行 Southern blot。未见杂交信号,表明转化载体的骨架序
列没有插入到转基因抗虫棉花 DAS-21023-5 的基因组中。 详见后文 2.2.5 插入
序列的拷贝数部分(报告 1E)。
23
图3. DAS-21023-5棉花外源序列和旁侧序列示意图 (8368 bp )
DAS-21023-5 棉花外源序列从 5′至 3′依次为:AtuORF25/26 3′UTR 的部分序列片段(构建载
体时遗留,无生物学功能)、AtuMas 启动子、pat 开放读码框、AtuORF25/26 3′UTR 终止序
列、cry1Ac 开放读码框和 ZmUbi1 启动子。
图4. pMYC3006 质粒载体的T-DNA结构示意图 (8422 bp)
pMYC3006 质粒的 T-DNA 从 5′至 3′依次为:T-DNA 边界 A、AtuORF25/26 3′UTR 的部分
序列(构建载体时遗留,无生物学功能)、AtuMas 启动子、pat 开放读码框、AtuORF25/26 3′UTR、cry1Ac 开放读码框、ZmUbi1 启动子和 T-DNA 边界 B。
本公司于采用 Sanger 测序法对 DAS-21023-5 棉花插入序列及其侧翼基因组
序列进行了测序。测得转基因抗虫棉花 DAS-21023-5 全长 9043 bp, 包括插入序
列长 8368 bp,5′端侧翼序列 358 bp(下划线所示 1-358 bp)和 3′端侧翼序列
317 bp(下划线所示 8727-9043 bp),序列见下文,详见报告 1B。
商业保密信息
24
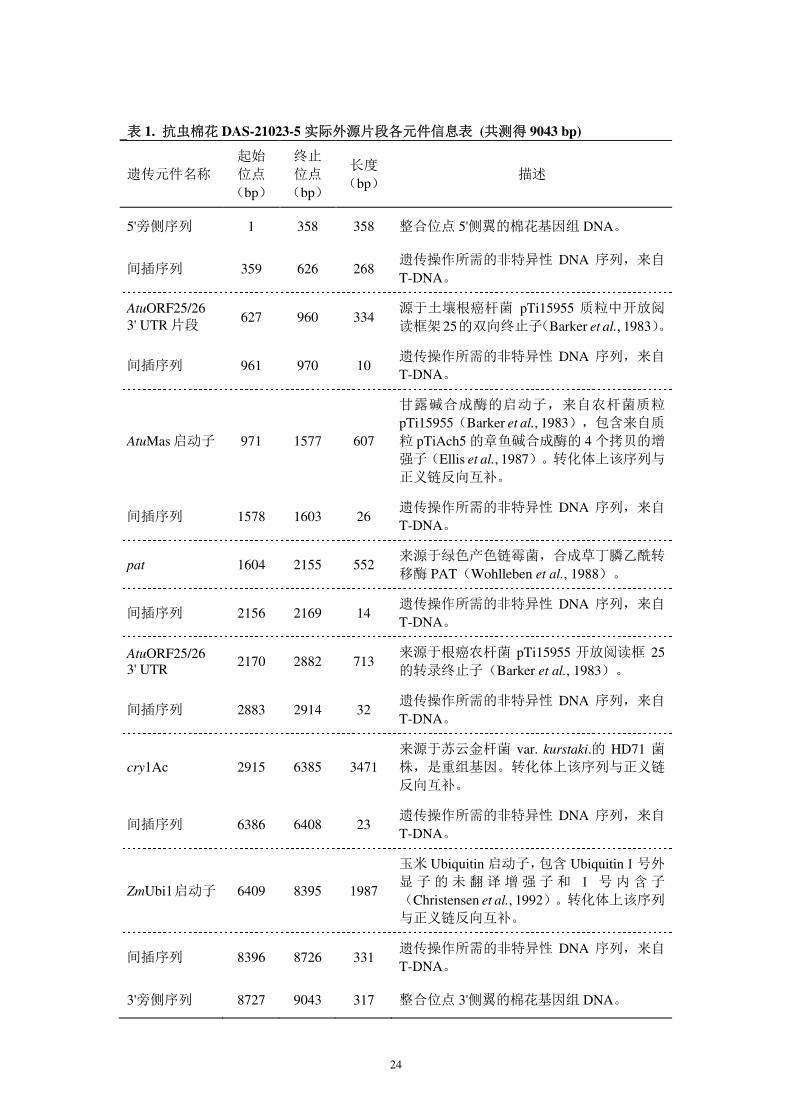
表 1. 抗虫棉花 DAS-21023-5 实际外源片段各元件信息表 (共测得 9043 bp)
遗传元件名称 起始 位点
(bp)
终止 位点
(bp)
长度 (bp)
描述
5'旁侧序列 1 358 358 整合位点 5'侧翼的棉花基因组 DNA。
间插序列 359 626 268 遗传操作所需的非特异性 DNA 序列,来自
T-DNA。
AtuORF25/26 3' UTR 片段
627 960 334 源于土壤根癌杆菌 pTi15955 质粒中开放阅
读框架25的双向终止子(Barker et al., 1983)。
间插序列 961 970 10 遗传操作所需的非特异性 DNA 序列,来自
T-DNA。
AtuMas 启动子 971 1577 607
甘露碱合成酶的启动子,来自农杆菌质粒
pTi15955(Barker et al., 1983),包含来自质
粒 pTiAch5 的章鱼碱合成酶的 4 个拷贝的增
强子(Ellis et al., 1987)。转化体上该序列与
正义链反向互补。
间插序列 1578 1603 26 遗传操作所需的非特异性 DNA 序列,来自
T-DNA。
pat 1604 2155 552 来源于绿色产色链霉菌,合成草丁膦乙酰转
移酶 PAT(Wohlleben et al., 1988)。
间插序列 2156 2169 14 遗传操作所需的非特异性 DNA 序列,来自
T-DNA。
AtuORF25/26 3' UTR
2170 2882 713 来源于根癌农杆菌 pTi15955 开放阅读框 25的转录终止子(Barker et al., 1983)。
间插序列 2883 2914 32 遗传操作所需的非特异性 DNA 序列,来自
T-DNA。
cry1Ac 2915 6385 3471 来源于苏云金杆菌 var. kurstaki.的 HD71 菌
株,是重组基因。转化体上该序列与正义链
反向互补。
间插序列 6386 6408 23 遗传操作所需的非特异性 DNA 序列,来自
T-DNA。
ZmUbi1启动子 6409 8395 1987
玉米 Ubiquitin 启动子,包含 Ubiquitin 1 号外
显 子 的 未 翻 译增 强 子和 1 号 内 含子
(Christensen et al., 1992)。转化体上该序列
与正义链反向互补。
间插序列 8396 8726 331 遗传操作所需的非特异性 DNA 序列,来自
T-DNA。
3'旁侧序列 8727 9043 317 整合位点 3'侧翼的棉花基因组 DNA。

25
确定插入序列的大小、结构及其特性的两种分析方法
通过以下两种方法分析 DAS-21023-5 棉花插入序列大小和结构:方法一,采
用反向 PCR 扩增旁侧序列、再用常规 PCR 扩增插入序列、然后将 PCR 产物拼
接、测序,进行生物信息学分析(序列比对及注释),最后确定了插入序列的大
小和结构(报告 1A);方法二,采用 Southern blot 确定并验证插入序列的整合位
点数、拷贝数和完整性,并证明该转基因棉花中未整合转化质粒 pMYC3006 的
骨架序列(报告 1E)。
方法一,克隆外源片段并测序确定其大小、结构和序列
以反向 PCR 的方法克隆了整合位点处 5′和 3′端旁侧棉花基因组序列。图 5
显示,用 HindIII 消化 DAS-21023-5 棉花基因组 DNA,消化产物使用 T4 连接酶
自连,采用巢式 PCR 进行扩增(引物信息见表 2),回收扩增产物后测序。以 T-
DNA 内侧两端序列为参照,比对分析测得转化位点 5′端和 3′端旁侧棉花基因组
序列。
接着,利用 5′和 3′端旁侧棉花基因组序列和质粒 pMYC3006 的 T-DNA 内部
序设计引物(引物组合 281-5/3006-6-6 和 281-4/3006-12b-3),扩增出 2 个片段,
即 5′端插入片段和 3′端插入片段(图 6),回收扩增产物并测序,测得序列与 T-
DNA 的一致(报告 1A)。
最后,将 5′端旁侧序列、5′端插入序列、3′端插入序列和 3′端旁侧序列四个
重叠片段拼接(图 6),获得了外源片段全序列及其旁侧序列。
表 2. DAS-21023-5 棉花插入序列及侧翼序列 PCR 扩增引物
扩增片段 引物名称 序列(5′-3′) 备注
3′旁侧序列
3006-3 商业保密信息 巢式 PCR 第一轮扩增
3006-4 商业保密信息
281-5 商业保密信息 巢式 PCR 第二轮扩增
3006-5 商业保密信息
3′外源序列 281-4 商业保密信息
扩增 3′插入序列 3006-12b 商业保密信息
5′外源序列 3006-6 商业保密信息
扩增 5′插入序列 281-5 商业保密信息
5′旁侧序列 3006-20 商业保密信息
扩增 5′边界序列 3006-21 商业保密信息
3′接合序列 3006-12 商业保密信息 扩增 3′转化事件
特异性序列 a 3006-9 商业保密信息
a:对 3′接合序列特异性扩增所用引物与后文(第六部分)DAS-24236-5 转化事件特异性检
测方法中的引物不同。
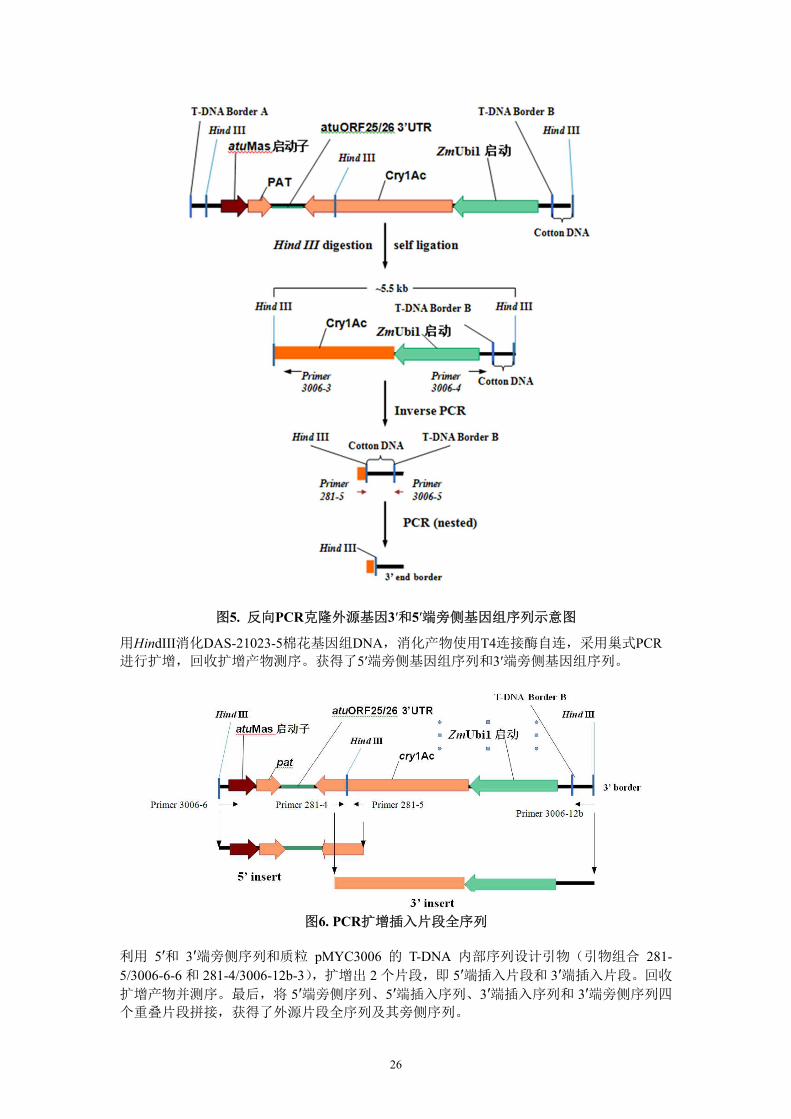
26
图5. 反向PCR克隆外源基因3′和5′端旁侧基因组序列示意图
用HindIII消化DAS-21023-5棉花基因组DNA,消化产物使用T4连接酶自连,采用巢式PCR进行扩增,回收扩增产物测序。获得了5′端旁侧基因组序列和3′端旁侧基因组序列。
图6. PCR扩增插入片段全序列
利用 5′和 3′端旁侧序列和质粒 pMYC3006 的 T-DNA 内部序列设计引物(引物组合 281-5/3006-6-6 和 281-4/3006-12b-3),扩增出 2 个片段,即 5′端插入片段和 3′端插入片段。回收
扩增产物并测序。最后,将 5′端旁侧序列、5′端插入序列、3′端插入序列和 3′端旁侧序列四
个重叠片段拼接,获得了外源片段全序列及其旁侧序列。

27
方法二,采用 Southern blot 确定插入序列拷贝数和完整性(详见后文 2.2.5
节)
Southern blot 显示 DAS-21023-5 棉花仅在单一位点整合了外源片段,外源片
段为单拷贝,其上含有结构完整的 cry1Ac 和 pat 基因表达盒;该转化体基因组
中未整合载体骨架序列,外源片段在 BC1F3、BC3F3、BC3F4 和 BC4F1四代间稳定
整合(报告 1E)。
插入位点数及拷贝数分析:DAS-21023-5 棉花基因组 DNA 分别经 BamHI、
HindIII、PacI、NcoI 和 Bgl II 消化(因整合位点不同而产生不同大小的片断),
与不同遗传元件探针杂交,均显示与预期相符的转化事件特异性杂交信号。不仅
证实了该转化体外源基因为单一拷贝,而且仅一个插入位点。
外源基因结构完整性分析:1)使用 BamHI 消化基因组 DNA,分别与
AtuORF25/26 3′UTR、cry1Ac、ZmUbi1 探针,均能显示与各遗传元件大小相同的
杂交信号条带。表明 cry1Ac 表达盒及其元件结构完整。2)使用 HindIII、XhoI 消
化基因组 DNA,用不同探针进行杂交,按完整的结构推测杂交结果,实际观察
结果与预期结果一致,不同酶与不同探针组合的 Southern blot 结果相互验证,证
实了 pat 表达盒及其元件结构完整。
DAS-21023-5 棉花中不含载体骨架序列:以覆盖整个 pMYC3006 质粒骨架
区域的 4 个探针与不同的内切酶消化的 DAS-21023-5 基因组 DNA 进行 Southern
blot,均未见杂交信号。表明转化载体的骨架序列没有插入到 DAS-21023-5 棉花
的基因组中。
2.2.2 删除区域的大小和功能;
1)删除序列分析
比较转化体 DAS-22023-5 与转化受体 GC510 在转化位点的序列,可知在 T-
DNA 整合过程中,在插入位点造成了 16 bp 的删除,删除序列为:CTTGGTTATT
CTATGA。
该段缺失序列不影响转化体生物学功能,原因如下:1)生物信息学比对
表明,整合位点上下游 1028 bp 片段内未发现开放阅读框(报告 1C),即未打
断具有特定功能的基因。2)转化受体陆地棉为四倍体,转化位点共 4 个等位基
因(allie),其中一个缺陷可由其它等位基因弥补。3)在后期农艺学调查、营
养组分分析、食用安全评价及环境安全评价中均未发现本转化体异常(报告 2
和报告 5)。4)在 DAS-21023-5 棉花分离群体中(BC4F2)转化位点的分离比例
未显著偏离 3:1,即符合单一位点的孟德尔遗传定律,从而进一步证明插入片
段位于染色体上(报告 1F)。因此,因此,该 16 bp 缺失不会对生物学功能带来
负面影响。
2)整合引起的接合区潜在新读码框分析

28
采用生物信息学的方法对 DAS-21023-5 转化事件整合区的推导蛋白进行分
析,这些推导蛋白与人、畜有害的蛋白质序列不相似,即生物信息学分析未见新
读码框的致敏、毒素风险。
将 2 个终止密码子之间定义为一个读码框,在 DAS-21023-5 棉花外源序列
及接合区共发现个 478 个长度≥ 8 aa 的潜在的读码框(推导蛋白)。2020 年,将
这些推导蛋白与公共的COMPARE(v4.0; http://comparedatabase.org)致敏数据库比
对,从而分析潜在的致敏性。结果显示:一方面,以≥ 80 aa 为一个滑动窗口,未
返回一致性>35%的匹配序列;另一方面,未见连续 8 aa 完全一致的匹配(蛋白
长度 8~29 aa 时)。即生物信息学分析未见 DAS-2102-5 棉花外源序列及接合区
致敏性风险。
推导蛋白和本公司内部毒素数据库进行比对,未返回 E ≤ 10-4 的匹配序列。
推导蛋白和 NCBI nr 数据库(ftp://ftp.ncbi.nlm.nih.gov/blast/db/)进行比对,返回了
9 个匹配(E ≤ 10-4 ),这些匹配序列均不属于毒性蛋白质。即生物信息学分析认
为推导蛋白不会带来毒性风险。
2.2.3 目的基因的核苷酸序列和推导的氨基酸序列;
本转化体含目的基因 cry1Ac 和选择标记基因 pat。
目的基因 cry1Ac 的核苷酸序列如下(含终止密码子):
目的基因cry1Ac推导氨基酸序列如下:
标记基因 pat 的核苷酸序列如下(含终止密码子):
标记基因 pat 推导的氨基酸序列如下:
2.2.4 插入序列在植物细胞中的定位(是否整合到染色体、叶绿体、
线粒体,或以非整合形式存在)及其确定方法;
商业保密信息
商业保密信息
商业保密信息
商业保密信息

29
外源基因整合至棉花受体基因组第 8 染色体。
采用反向 PCR(inverse PCR)、DNA 测序的方法获得了 DAS-21023-5 棉花
外源片段插入位点旁侧的基因组序列。将插入片段两侧的序列与转化受体上相应
转化位点区域的序列比对,从而确定转化位点在基因组第 8 染色体(报告 1C),该
插入未破坏内源基因。
将 DAS-21023-5 棉花 T-DNA 插入序列、旁侧边界序列和受体棉花转化位点
的序列与数据库中的序列进行了比较。使用 BLAST 算法搜索最新的 GenBank
非冗余核苷酸、非人类和非鼠类表达序列标签(EST)和非冗余蛋白质数据库(于
2018 年),结果显示,外源序列未插入棉花内源基因或调控元件。
报告 1A 采用 DAS-21023-5 转化事件特异性 PCR 的方法验证外源序列所在
位置。以 DAS-21023-5 棉花整合位点 3′端接合区(含旁侧基因组 DNA 及外源序
列)设计引物,对该接合区进行转化事件特异性 PCR 扩增,引物组合为 3006-
12/3006-9,引物序列见上文表 2。结果显示,除了在 DAS-21023-5 棉花中可有效
扩增之外,在非转基因棉花、其它棉花转化体、其它物种中均不扩增(图 7)。从
而验证了转化位点定位的正确性。
图 7. DAS-21023-5 棉花插入位点 3′端接合区特异性 PCR
(与第六部分检测方法中的引物不同)
1:分子量标准;2:水对照;3:非转基因玉米对照 B680;4:转基因棉花 DAS-24236-5;5 和
6:DAS-21023-5;7 和 8:棉花 3006-48-84;9 和 10:转基因棉花 3406;11 和 12:转基因
棉花 3408;13 和 14:转基因棉花 3412。
以引物组合 3006-12/3006-9 对基因组 DNA 进行扩增,仅在 DAS-21023-5 中可扩增出 661 bp的特异片段,在非转基因棉花、其它转基因棉花及其它物种玉米中无扩增产物。由此可证实
外源基因的整合位点。
2.2.5 插入序列的拷贝数。
采用 Southern blot 试验方法,以 DAS-21023-5 棉花的 BC1F3、BC3F3、BC3F4
和 BC4F1 多个世代单株为实验材料, 验证了外源序列为单一的插入位点、单一

30
拷贝、插入结构完整,并且转化体中未整合载体骨架序列,外源序列在代际间稳
定整合并遗传(报告 1E)。
根据旁侧序列及外源序列的限制性内切酶位点及探针位置(表 3),可预测杂
交信号片段个数和大小,比较预期结果与实际观察片段(表 4),从而判断外源基
因的拷贝数及表达盒完性。
1)插入位点数和拷贝数
不同转化体整合位点各异,整合位点邻近基因组序列不同。采用内切酶识别
位点分别在外源序列和基因组上的酶切片段,结合相应探针进行 Southern blot,
可以鉴定外源基因的插入位点数和拷贝数(图 8 和图 9)。本实验使用限制性内
切酶 BamHI、HindIII、PacI、NcoI 和 Bgl II 消化鉴定 DAS-21023-5 棉花各基因
和调控元件的拷贝数。
BamHI 酶切:基因组 DNA 经 BamHI 酶解后与 AtuORF25/26 3′UTR 探针杂
交,在 DAS-21023-5 棉花泳道,除了显示~700bp 信号条带与预期的 747 bp 的相
符,还有另外一个 12,000 bp 的特异信号,与预期的>905 bp 相符(图 18A)。表
明 AtuORF25/26 3′UTR 为单一拷贝。
HindIII 酶切:DAS-21023-5 基因组 DNA 经 HindIII 酶解后与 cry1Ac 探针
杂交,显示~3,000 bp 和~5,500 bp 的杂交带,与预期 3,010 bp 和>5,091 bp 一
致(图 10B);和 ZmUbi1 探针杂交,显示~5,500 bp 的杂交片段,与预期的>
5,091bp 相符(图 14B)。 表明 cry1Ac 为单一拷贝。
PacI 酶切:DAS-21023-5 基因组 DNA 经 PacI 酶解后与 cry1Ac 探针杂交,
显示~2,300 bp 和~14,500 bp 的杂交带,与预期 2,280 bp 和>5,778bp 一致(图
10A);和 pat 探针杂交,显示~14,500 bp 的杂交片段,与预期的>5,778 bp 相符
(图 12B);与 AtuMas 探针杂交,显示 14,500 bp 杂交信号,与预期的>5,778bp
一致(图 16B)。表明 cry1Ac、pat 和 AtuMas 为单一拷贝。
NcoI 酶切:DAS-21023-5 基因组 DNA 经 NcoI 酶解后与 pat 探针杂交,显
示一条~11,000 bp 的杂交条带,与表 4 中预期结果一致(>6,995 bp),见图 13A。
表明 pat 为单一拷贝。
BglII 酶切:DAS-21023-5 基因组 DNA 经 BglII 酶切后与 ZmUbi1 启动子探
针杂交,显示两个信号,一个在~4,000 bp 处,与预期的 3,360 bp 相符;另一个
在~3,400 处,与预期的>1,272 bp 相符(图 15)。表明 ZmUbi1 启动子为单一拷
贝;以 BglII 消化的 DAS-21023-5 基因组 DNA 与 AtuORF25/26 3′UTR 探针进
行 Southern blot,在~7500 bp 处观察到杂交条带,与预期的>604 bp 相符(图
19),表明外源基因为单一拷贝。
综合以上内切酶与遗传元件组合的 Southern blot 结果,外源插入序列在单一
位点整合,且为单拷贝。

31
Southern blot 观察结果与预测结果均一致,且不同酶切方式的 Southern blot
相互验证,表明棉花 DAS-21023-5 基因组中整合了一个拷贝的 cry1Ac 表达盒,
一个拷贝的 pat 表达盒。
2)插入序列的结构完整性
cry1Ac 表达盒的完整性
以 cry1Ac 序列设计探针(称为 cry1Ac 探针,以下类推),与 BamHI 消化的
DAS-21023-5 棉花基因组 DNA 进行 Southern blot,在~3,500 bp 处显示一个与
cry1Ac 基因大小相符的信号(图 11),且与预期 3,481 bp 相符。
以 ZmUbi1 启动子序列设计探针,与 BamHI 酶切的 DAS-21023-5 基因组
DNA 杂交,可见一条~2,100 bp 的与 ZmUbi1 启动子大小相符的信号条带(图
14A),与预期的 2,050 bp 相符。
以 AtuORF25/26 3′UTR 终止序列设计探针,与 HindIII 消化 DAS-21023-5
的 DNA 进行 Southern blot,在~3000 bp 处观察杂交信号,与 pat 表达盒大小
( 3013 bp)相附)。
不同的酶、不同的遗传元件作探针所观察到的杂交信号均与预期相符,且相
互印证,表明 DAS-21023-5 棉花包含一个完整的 cry1Ac 表达盒。
pat 表达盒的完整性
用 pat 开放读码框设计探针,与 HindIII 消化的 DAS-21023-5 基因组 DNA 进
行 Southern blot,在~3,000 bp 处观察到信号条带(图 12A),与预期 3,013 bp 相
符;与 XhoI 消化的基因组 DNA 进行杂交,显示~3,600 bp 杂交信号(图 13B),
与预期 3,581 bp 相符。
用 AtuMas 启动子设计探针,与 HindIII 消化的基因组 DNA 进行杂交,在~
3,000 bp 处观察到杂交信号(图 16A),与预期 3,031 bp 相符;与 XhoI 消化的基
因组 DNA 进行杂交,在~3,600 处显示了信号条带(图 17),与预期的 3581 bp
相符。
pat 与 cry1Ac 表达盒共用一个 AtuORF25/26 终止序列,上文验证了其完整
性。
不同的酶与遗传元件探针组合所观察到的杂交信号均与预期相符,相互印证,
表明 DAS-21023-5 棉花包含完整的 pat 表达盒。
3)DAS-21023-5 基因组未整合载体骨架序列
采用覆盖整个转化质粒 pMYC3006 载体骨架的 4 个探针与 NcoI 和 HindIII
酶切的基因组 DNA 进行 Southern blot,以证实 DAS-21023-5 基因组未整合载体
骨架序列。
以 NcoI 酶切的基因组 DNA 与第一组探针(BB-A 和 BB-D)进行杂交,阳
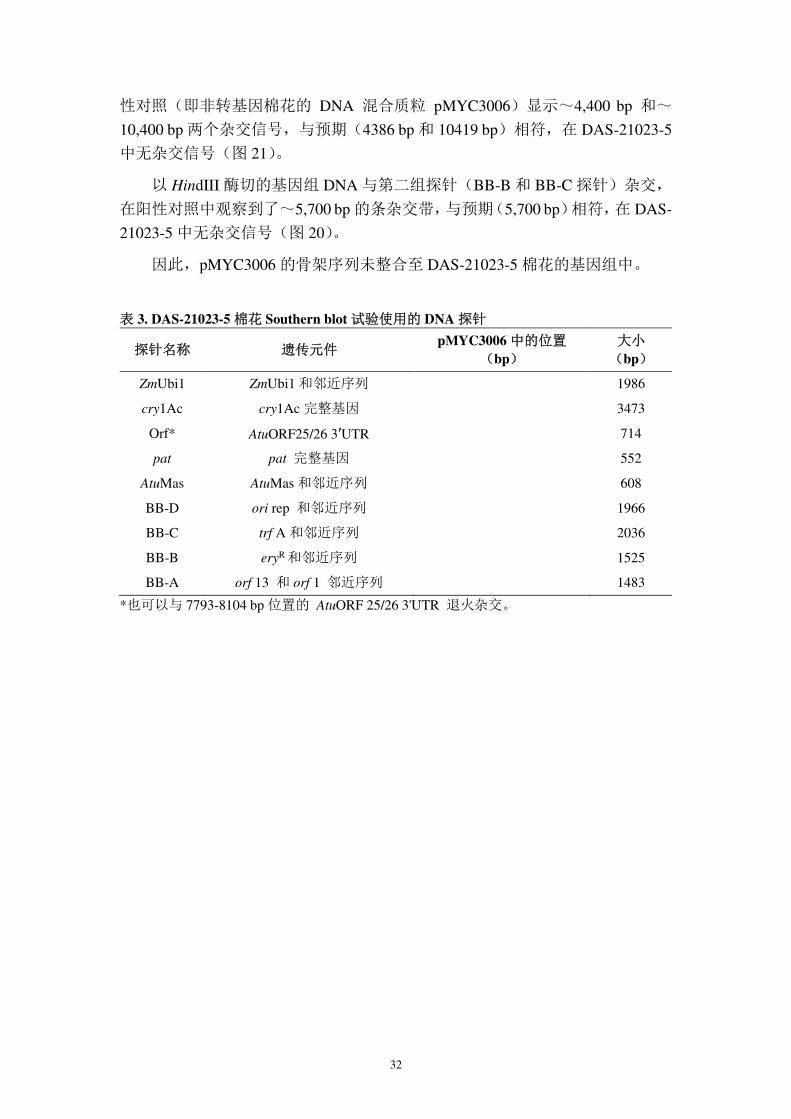
32
性对照(即非转基因棉花的 DNA 混合质粒 pMYC3006)显示~4,400 bp 和~
10,400 bp 两个杂交信号,与预期(4386 bp 和 10419 bp)相符,在 DAS-21023-5
中无杂交信号(图 21)。
以 HindIII 酶切的基因组 DNA 与第二组探针(BB-B 和 BB-C 探针)杂交,
在阳性对照中观察到了~5,700 bp 的条杂交带,与预期(5,700 bp)相符,在 DAS-
21023-5 中无杂交信号(图 20)。
因此,pMYC3006 的骨架序列未整合至 DAS-21023-5 棉花的基因组中。
表 3. DAS-21023-5 棉花 Southern blot 试验使用的 DNA 探针
探针名称 遗传元件 pMYC3006 中的位置
(bp)
大小
(bp)
ZmUbi1 ZmUbi1 和邻近序列 1986
cry1Ac cry1Ac 完整基因 3473
Orf* AtuORF25/26 3′UTR 714
pat pat 完整基因 552
AtuMas AtuMas 和邻近序列 608
BB-D ori rep 和邻近序列 1966
BB-C trf A 和邻近序列 2036
BB-B eryR 和邻近序列 1525
BB-A orf 13 和 orf 1 邻近序列 1483
*也可以与 7793-8104 bp 位置的 AtuORF 25/26 3'UTR 退火杂交。

33
图 8. Southern blot 探针在转化载体 pMYC3006 上的位置
34
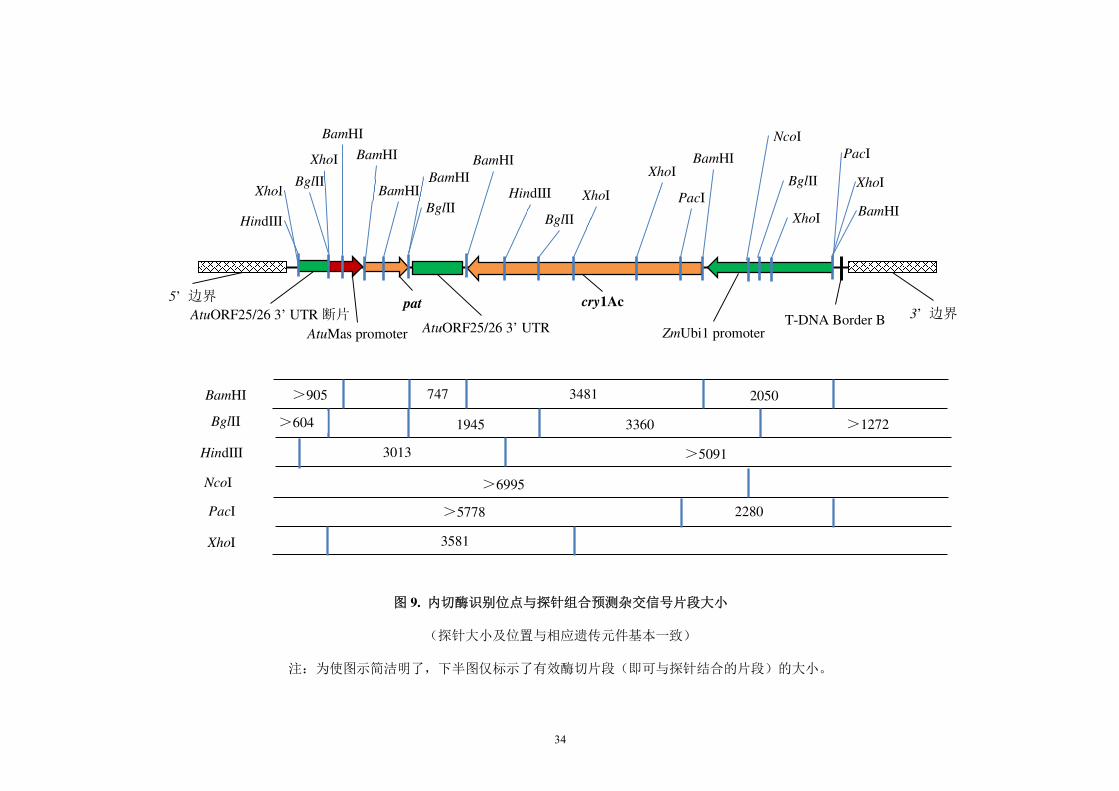
图 9. 内切酶识别位点与探针组合预测杂交信号片段大小
(探针大小及位置与相应遗传元件基本一致)
注:为使图示简洁明了,下半图仅标示了有效酶切片段(即可与探针结合的片段)的大小。
AtuMas promoter
5’ 边界 pat
AtuORF25/26 3’ UTR
cry1Ac
ZmUbi1 promoter T-DNA Border B 3’ 边界
BglII
HindIII
PacI
AtuORF25/26 3’ UTR 断片
BamHI
XhoI
HindIII
XhoI
BamHI
XhoI BglII
BamHI
BamHI
BglII
BamHI
HindIII
BglII
XhoI
XhoI
PacI
BamHI
BglII
XhoI
PacI
XhoI
BamHI BamHI
>905 747 3481 2050
>604 1945 3360 >1272
3013 >5091
>5778 2280
3581
NcoI
NcoI
>6995
35
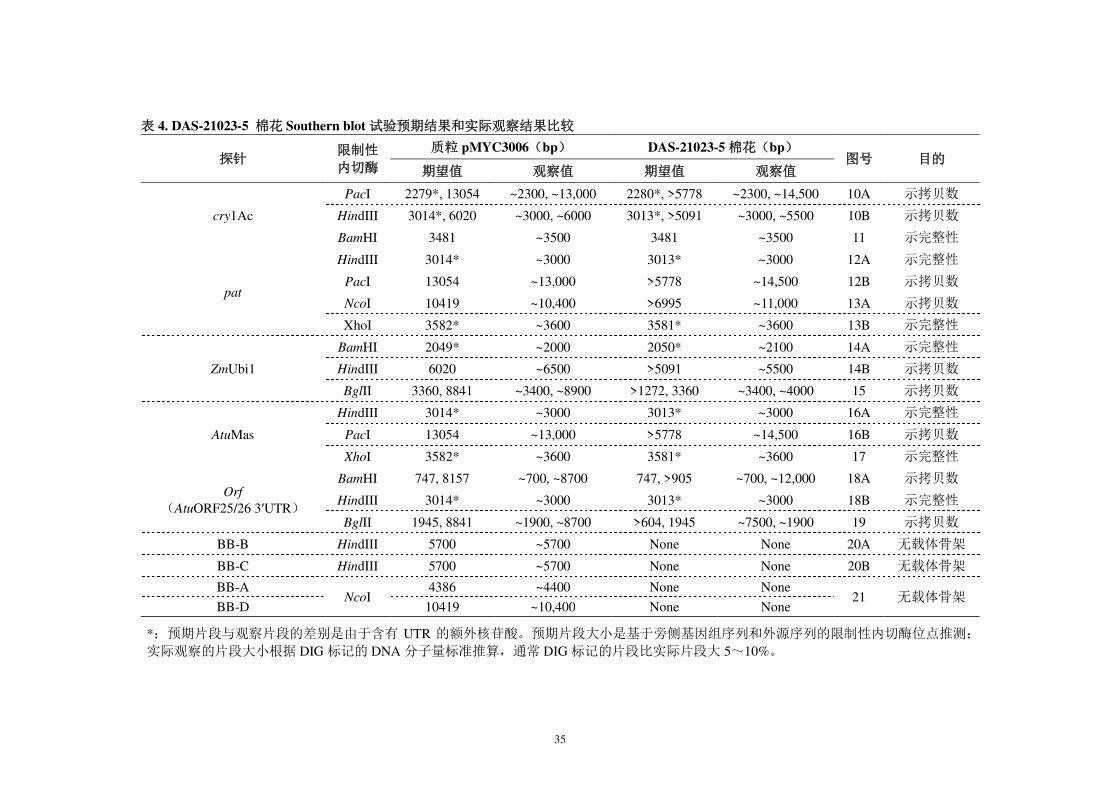
表 4. DAS-21023-5 棉花 Southern blot 试验预期结果和实际观察结果比较
探针 限制性
内切酶
质粒 pMYC3006(bp) DAS-21023-5 棉花(bp) 图号 目的
期望值 观察值 期望值 观察值
cry1Ac
PacI 2279*, 13054 ~2300, ~13,000 2280*, >5778 ~2300, ~14,500 10A 示拷贝数
HindIII 3014*, 6020 ~3000, ~6000 3013*, >5091 ~3000, ~5500 10B 示拷贝数
BamHI 3481 ~3500 3481 ~3500 11 示完整性
pat
HindIII 3014* ~3000 3013* ~3000 12A 示完整性
PacI 13054 ~13,000 >5778 ~14,500 12B 示拷贝数
NcoI 10419 ~10,400 >6995 ~11,000 13A 示拷贝数
XhoI 3582* ~3600 3581* ~3600 13B 示完整性
ZmUbi1
BamHI 2049* ~2000 2050* ~2100 14A 示完整性
HindIII 6020 ~6500 >5091 ~5500 14B 示拷贝数
BglII 3360, 8841 ~3400, ~8900 >1272, 3360 ~3400, ~4000 15 示拷贝数
AtuMas
HindIII 3014* ~3000 3013* ~3000 16A 示完整性
PacI 13054 ~13,000 >5778 ~14,500 16B 示拷贝数
XhoI 3582* ~3600 3581* ~3600 17 示完整性
Orf
(AtuORF25/26 3′UTR)
BamHI 747, 8157 ~700, ~8700 747, >905 ~700, ~12,000 18A 示拷贝数
HindIII 3014* ~3000 3013* ~3000 18B 示完整性
BglII 1945, 8841 ~1900, ~8700 >604, 1945 ~7500, ~1900 19 示拷贝数
BB-B HindIII 5700 ~5700 None None 20A 无载体骨架
BB-C HindIII 5700 ~5700 None None 20B 无载体骨架
BB-A NcoI
4386 ~4400 None None 21 无载体骨架
BB-D 10419 ~10,400 None None
*:预期片段与观察片段的差别是由于含有 UTR 的额外核苷酸。预期片段大小是基于旁侧基因组序列和外源序列的限制性内切酶位点推测;
实际观察的片段大小根据 DIG 标记的 DNA 分子量标准推算,通常 DIG 标记的片段比实际片段大 5~10%。
36
泳道 膜 A 膜 B
泳道 膜 A 膜 B
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII
1 PSC355-2+pMYC3006 PSC355-2+pMYC3006 9 3006-BC3F4-1 3006-BC3F4-1
2 PSC355-2 PSC355-2 10 3006-BC3F4-2 3006-BC3F4-2
3 3006-BC1F3-1 3006-BC1F3-1 11 3006-BC3F4-3 3006-BC3F4-3
4 3006-BC1F3-2 3006-BC1F3-2 12 3006-BC4F1-3 3006-BC4F1-3
5 3006-BC1F3-4 3006-BC1F3-4 13 3006-BC4F1-4 3006-BC4F1-4
6 3006-BC3F3-1 3006-BC3F3-1 14 3006-BC4F1-5 3006-BC4F1-5
7 3006-BC3F3-4 3006-BC3F3-4 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5 3006-BC3F3-5
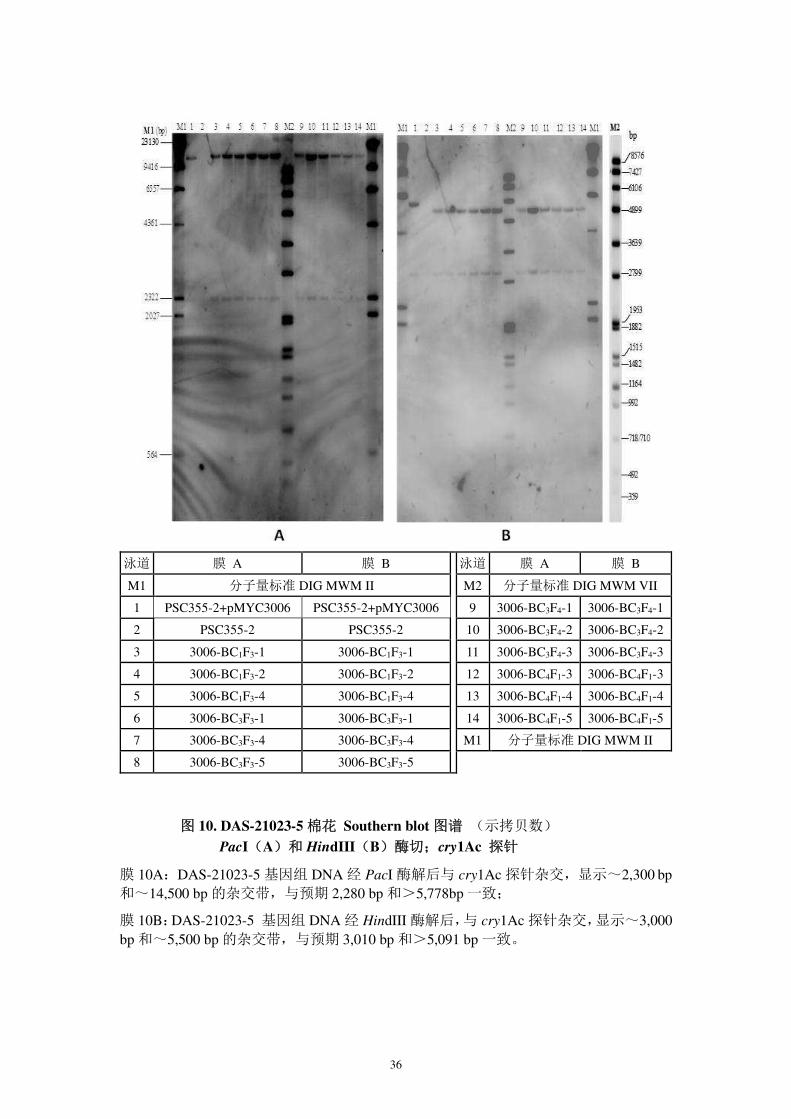
图 10. DAS-21023-5 棉花 Southern blot 图谱 (示拷贝数)
PacI(A)和 HindIII(B)酶切;cry1Ac 探针
膜 10A:DAS-21023-5 基因组 DNA 经 PacI 酶解后与 cry1Ac 探针杂交,显示~2,300 bp和~14,500 bp 的杂交带,与预期 2,280 bp 和>5,778bp 一致;
膜 10B:DAS-21023-5 基因组 DNA 经 HindIII 酶解后,与 cry1Ac 探针杂交,显示~3,000 bp 和~5,500 bp 的杂交带,与预期 3,010 bp 和>5,091 bp 一致。

37
图 11. DAS-21023-5 棉花的 Southern blot 图谱 (示完整性)
BamHI 酶切;cry1Ac 探针
以 cry1Ac 序列设计探针,与 BamHI 消化的 DAS-21023-5 棉花基因组 DNA 进行 Southern blot,在~3,500 bp 处显示一个与 cry1Ac 基因大小相符的信号,且与预期 3,481 bp 相符。
泳道 试样 泳道 试样
M1 分子量标准 DIG MWM II
M2 分子量标准 DIG MWM VII
1 PSC355-8+pMYC3006 9 3006-BC3F4-1
2 PSC355-8 10 3006-BC3F4-2
3 3006-BC1F3-1 11 3006-BC3F4-5
4 3006-BC1F3-2 12 3006-BC4F1-1
5 3006-BC1F3-4 13 3006-BC4F1-2
6 3006-BC3F3-2 14 3006-BC4F1-3
7 3006-BC3F3-3 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5
38
泳道 膜 A 膜 B
泳道 膜 A 膜 B
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII
1 PSC355-2+pMYC3006 PSC355-2+pMYC3006 9 3006-BC3F4-1 3006-BC3F4-1
2 PSC355-2 PSC355-2 10 3006-BC3F4-2 3006-BC3F4-2
3 3006-BC1F3-1 3006-BC1F3-1 11 3006-BC3F4-3 3006-BC3F4-3
4 3006-BC1F3-2 3006-BC1F3-2 12 3006-BC4F1-3 3006-BC4F1-3
5 3006-BC1F3-4 3006-BC1F3-4 13 3006-BC4F1-4 3006-BC4F1-4
6 3006-BC3F3-1 3006-BC3F3-1 14 3006-BC4F1-5 3006-BC4F1-5
7 3006-BC3F3-4 3006-BC3F3-4 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5 3006-BC3F3-5
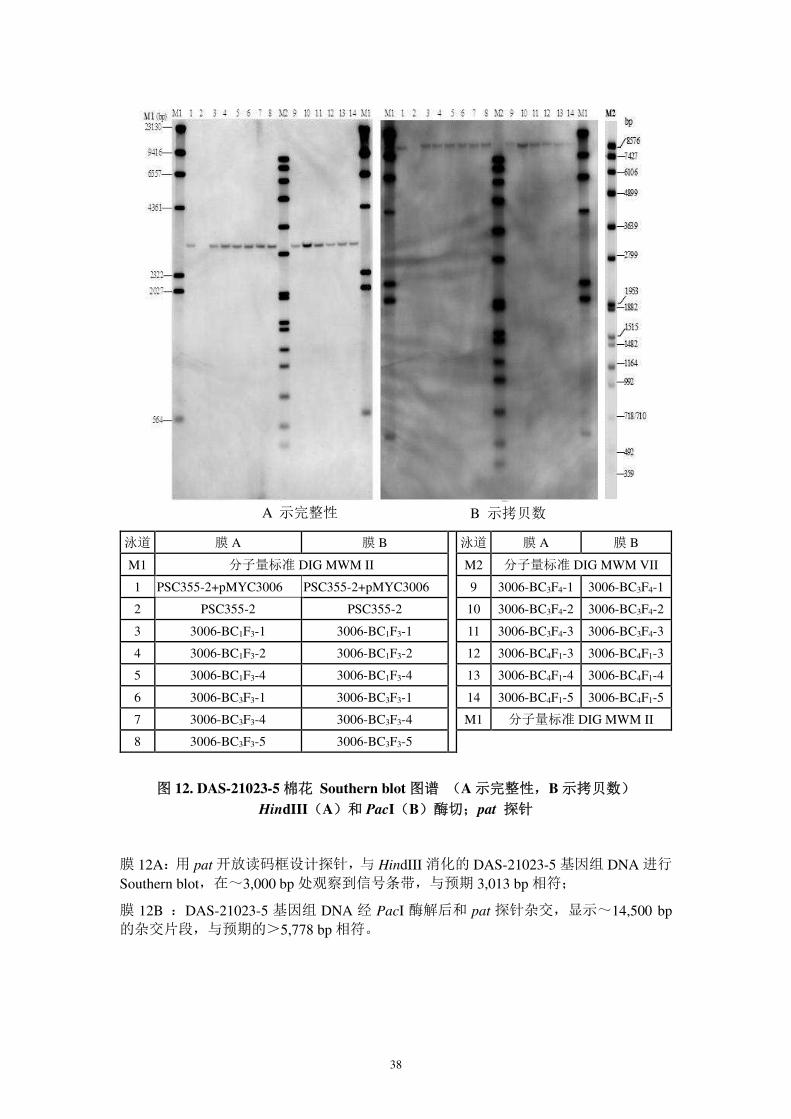
图 12. DAS-21023-5 棉花 Southern blot 图谱 (A 示完整性,B 示拷贝数)
HindIII(A)和 PacI(B)酶切;pat 探针
膜 12A:用 pat 开放读码框设计探针,与 HindIII 消化的 DAS-21023-5 基因组 DNA 进行
Southern blot,在~3,000 bp 处观察到信号条带,与预期 3,013 bp 相符;
膜 12B :DAS-21023-5 基因组 DNA 经 PacI 酶解后和 pat 探针杂交,显示~14,500 bp的杂交片段,与预期的>5,778 bp 相符。
A 示完整性 B 示拷贝数
39
泳道 膜 A 膜 B
泳道 膜 A 膜 B
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII 1 PSC355-8+pMYC3006 PSC355-3+pMYC3006 9 3006-BC3F4-1 3006-BC3F4-1 2 PSC355-8 PSC355-3 10 3006-BC3F4-2 3006-BC3F4-2 3 3006-BC1F3-1 3006-BC1F3-1 11 3006-BC3F4-5 3006-BC3F4-3 4 3006-BC1F3-2 3006-BC1F3-2 12 3006-BC4F1-1 3006-BC4F1-2 5 3006-BC1F3-3 3006-BC1F3-4 13 3006-BC4F1-2 3006-BC4F1-3 6 3006-BC3F3-2 3006-BC3F3-1 14 3006-BC4F1-4 3006-BC4F1-5 7 3006-BC3F3-3 3006-BC3F3-4 M1 分子量标准 DIG MWM II 8 3006-BC3F3-5 3006-BC3F3-5
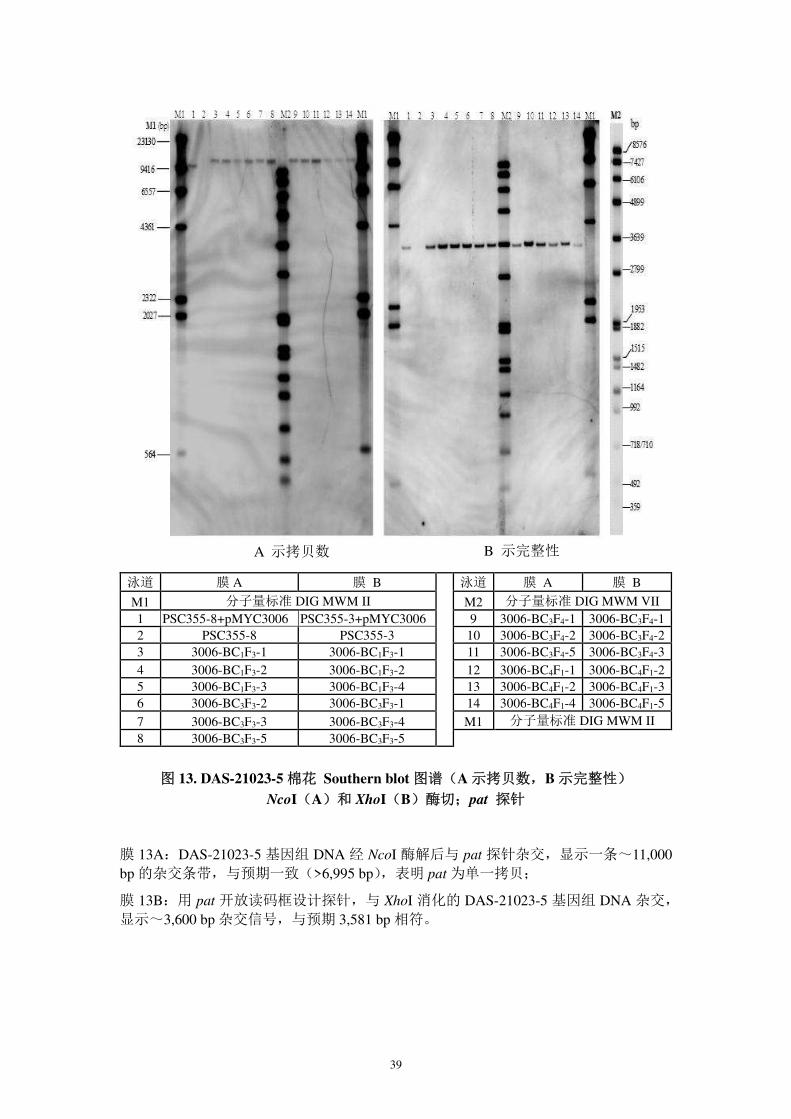
图 13. DAS-21023-5 棉花 Southern blot 图谱(A 示拷贝数,B 示完整性)
NcoI(A)和 XhoI(B)酶切;pat 探针
膜 13A:DAS-21023-5 基因组 DNA 经 NcoI 酶解后与 pat 探针杂交,显示一条~11,000 bp 的杂交条带,与预期一致(>6,995 bp),表明 pat 为单一拷贝;
膜 13B:用 pat 开放读码框设计探针,与 XhoI 消化的 DAS-21023-5 基因组 DNA 杂交,
显示~3,600 bp 杂交信号,与预期 3,581 bp 相符。
B 示完整性 A 示拷贝数

40
泳道 膜 A 膜 B
泳道 膜 A 膜 B
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII
1 PSC355-8+pMYC3006 PSC355-2+pMYC3006 9 3006-BC3F4-1 3006-BC3F4-1
2 PSC355-8 PSC355-2 10 3006-BC3F4-2 3006-BC3F4-2
3 3006-BC1F3-1 3006-BC1F3-1 11 3006-BC3F4-5 3006-BC3F4-3
4 3006-BC1F3-2 3006-BC1F3-2 12 3006-BC4F1-1 3006-BC4F1-3
5 3006-BC1F3-4 3006-BC1F3-4 13 3006-BC4F1-2 3006-BC4F1-4
6 3006-BC3F3-2 3006-BC3F3-1 14 3006-BC4F1-3 3006-BC4F1-5
7 3006-BC3F3-3 3006-BC3F3-4 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5 3006-BC3F3-5
图 14. DAS-21023-5 棉花 Southern blot 图谱(A 示完整性,B 示拷贝数)
BamHI(A)和 HindIII(B)酶切;ZmUbi 1 探针
膜 14A:以 ZmUbi1 启动子序列设计探针,与 BamHI 酶切的 DAS-21023-5 基因组 DNA杂交,可见一条~2,100 bp 的与 ZmUbi1 启动子大小相符的信号条带,与预期 2,050 bp相符;
膜 14B:DAS-21023-5 基因组 DNA 经 HindIII 酶解后与 ZmUbi1 探针杂交,显示~5,500 bp 的杂交片段,与预期的>5,091bp 相符。 表明 cry1Ac 为单一拷贝。
A 示完整性 B 示拷贝数
41
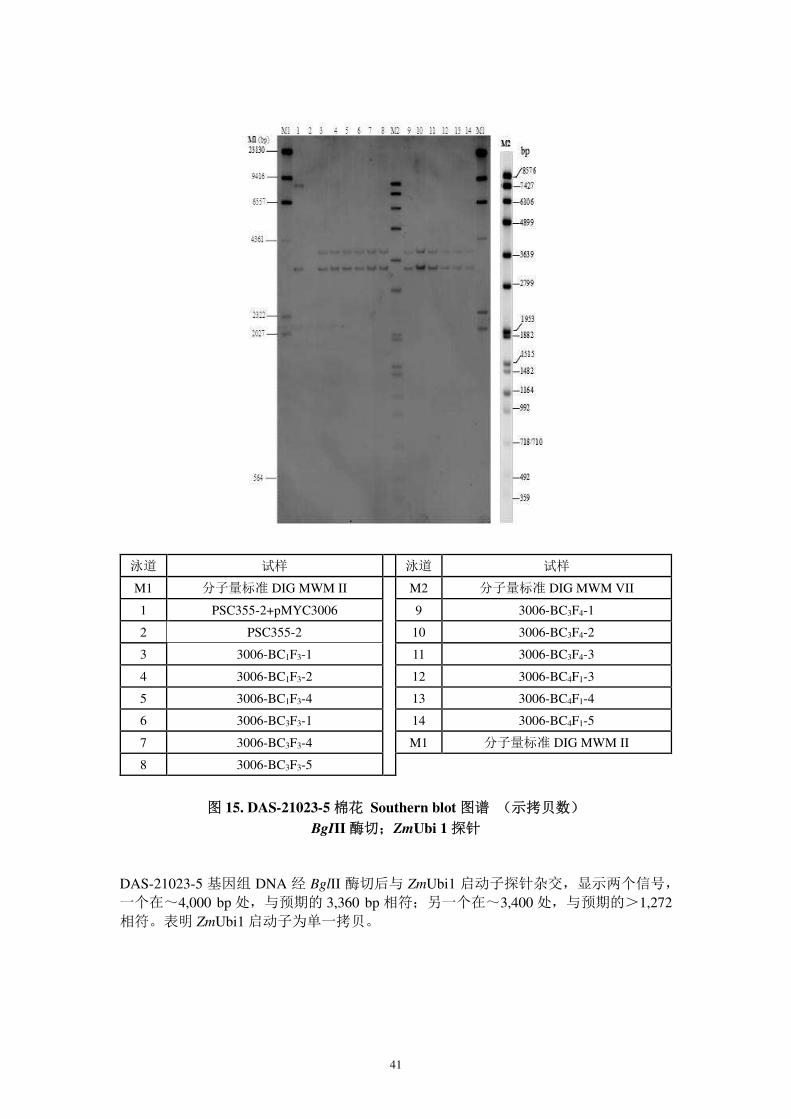
图 15. DAS-21023-5 棉花 Southern blot 图谱 (示拷贝数)
BgIII 酶切;ZmUbi 1 探针
DAS-21023-5 基因组 DNA 经 BglII 酶切后与 ZmUbi1 启动子探针杂交,显示两个信号,
一个在~4,000 bp 处,与预期的 3,360 bp 相符;另一个在~3,400 处,与预期的>1,272相符。表明 ZmUbi1 启动子为单一拷贝。
泳道 试样 泳道 试样
M1 分子量标准 DIG MWM II
M2 分子量标准 DIG MWM VII
1 PSC355-2+pMYC3006 9 3006-BC3F4-1
2 PSC355-2 10 3006-BC3F4-2
3 3006-BC1F3-1 11 3006-BC3F4-3
4 3006-BC1F3-2 12 3006-BC4F1-3
5 3006-BC1F3-4 13 3006-BC4F1-4
6 3006-BC3F3-1 14 3006-BC4F1-5
7 3006-BC3F3-4 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5
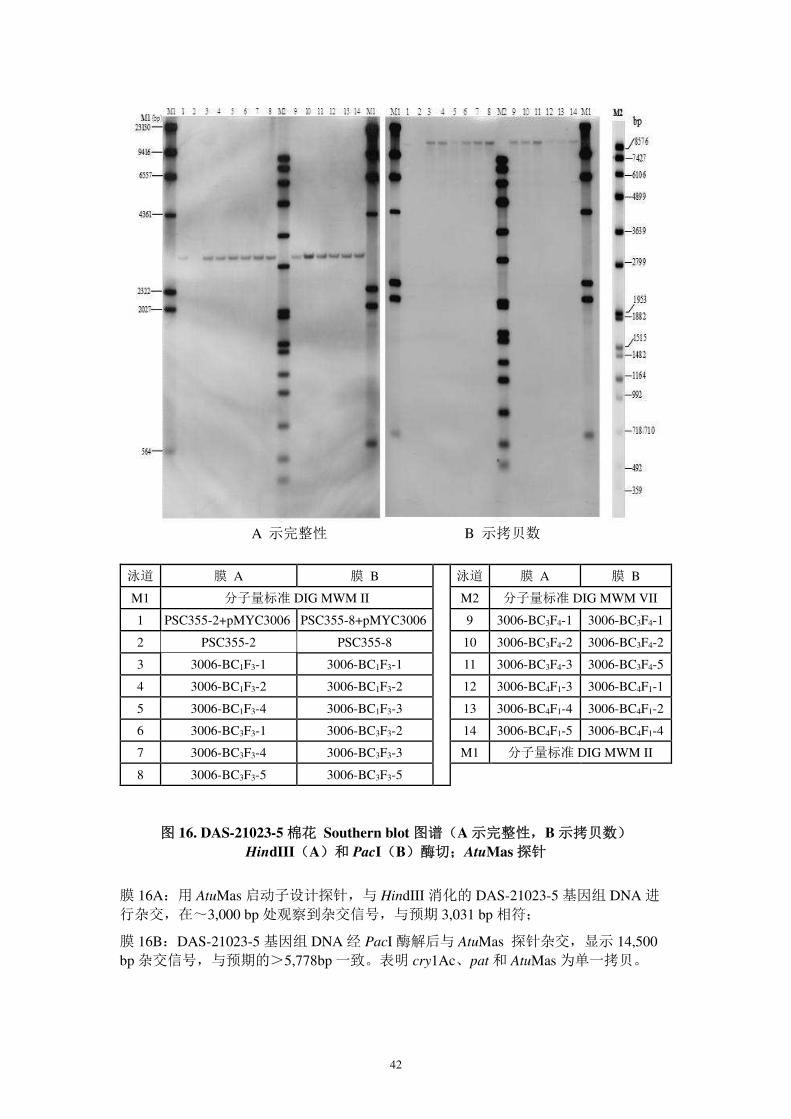
42
图 16. DAS-21023-5 棉花 Southern blot 图谱(A 示完整性,B 示拷贝数)
HindIII(A)和 PacI(B)酶切;AtuMas 探针
膜 16A:用 AtuMas 启动子设计探针,与 HindIII 消化的 DAS-21023-5 基因组 DNA 进
行杂交,在~3,000 bp 处观察到杂交信号,与预期 3,031 bp 相符;
膜 16B:DAS-21023-5 基因组 DNA 经 PacI 酶解后与 AtuMas 探针杂交,显示 14,500 bp 杂交信号,与预期的>5,778bp 一致。表明 cry1Ac、pat 和 AtuMas 为单一拷贝。
泳道 膜 A 膜 B
泳道 膜 A 膜 B
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII
1 PSC355-2+pMYC3006 PSC355-8+pMYC3006 9 3006-BC3F4-1 3006-BC3F4-1
2 PSC355-2 PSC355-8 10 3006-BC3F4-2 3006-BC3F4-2
3 3006-BC1F3-1 3006-BC1F3-1 11 3006-BC3F4-3 3006-BC3F4-5
4 3006-BC1F3-2 3006-BC1F3-2 12 3006-BC4F1-3 3006-BC4F1-1
5 3006-BC1F3-4 3006-BC1F3-3 13 3006-BC4F1-4 3006-BC4F1-2
6 3006-BC3F3-1 3006-BC3F3-2 14 3006-BC4F1-5 3006-BC4F1-4
7 3006-BC3F3-4 3006-BC3F3-3 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5 3006-BC3F3-5
A 示完整性 B 示拷贝数
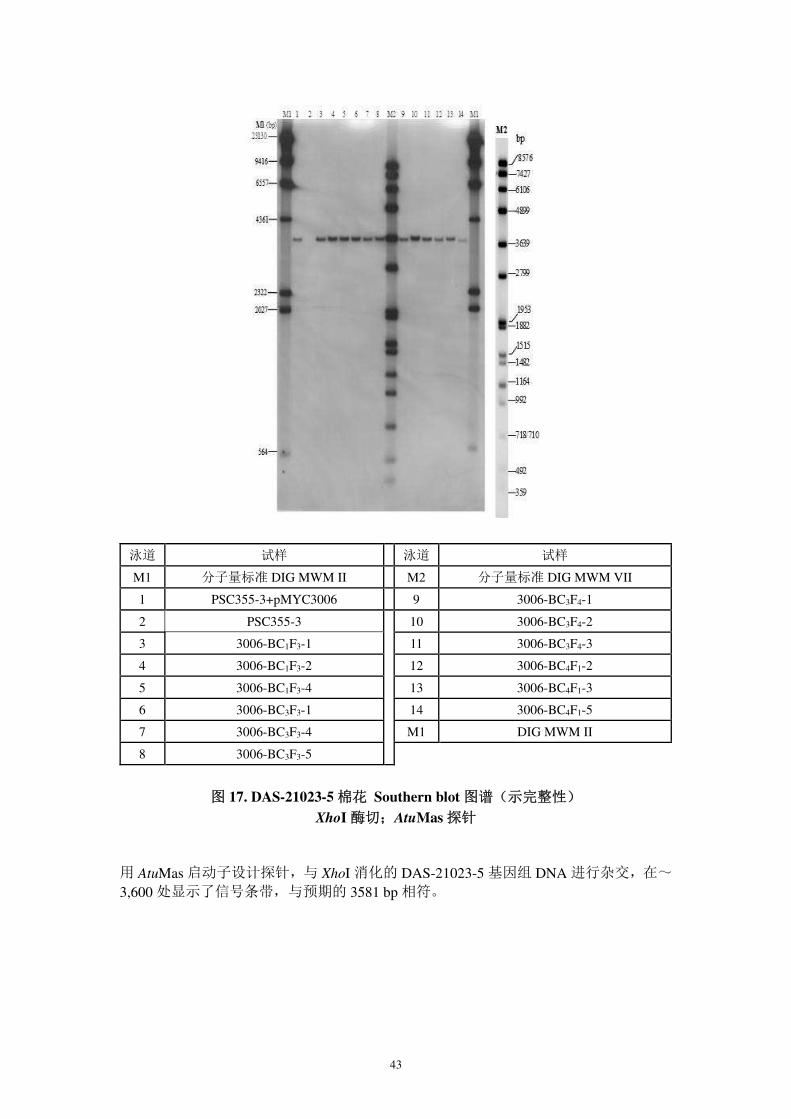
43
图 17. DAS-21023-5 棉花 Southern blot 图谱(示完整性)
XhoI 酶切;AtuMas 探针
用 AtuMas 启动子设计探针,与 XhoI 消化的 DAS-21023-5 基因组 DNA 进行杂交,在~
3,600 处显示了信号条带,与预期的 3581 bp 相符。
泳道 试样 泳道 试样
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII
1 PSC355-3+pMYC3006 9 3006-BC3F4-1
2 PSC355-3
10 3006-BC3F4-2
3 3006-BC1F3-1 11 3006-BC3F4-3
4 3006-BC1F3-2 12 3006-BC4F1-2
5 3006-BC1F3-4 13 3006-BC4F1-3
6 3006-BC3F3-1 14 3006-BC4F1-5
7 3006-BC3F3-4 M1 DIG MWM II
8 3006-BC3F3-5

44
泳道 膜 A 膜 B
泳道 膜 A 膜 B
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII
1 PSC355-8+pMYC3006 PSC355-2+pMYC3006 9 3006-BC3F4-1 3006-BC3F4-1
2 PSC355-8 PSC355-2 10 3006-BC3F4-2 3006-BC3F4-2
3 3006-BC1F3-1 3006-BC1F3-1 11 3006-BC3F4-5 3006-BC3F4-3
4 3006-BC1F3-2 3006-BC1F3-2 12 3006-BC4F1-1 3006-BC4F1-3
5 3006-BC1F3-4 3006-BC1F3-4 13 3006-BC4F1-2 3006-BC4F1-4
6 3006-BC3F3-2 3006-BC3F3-1 14 3006-BC4F1-3 3006-BC4F1-5
7 3006-BC3F3-3 3006-BC3F3-4 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5 3006-BC3F3-5
图 18. DAS-21023-5 棉花 Southern blot 图谱
BamHI(A)和 HindIII(B)酶切;orf 探针
膜 18A:DAS-21023-5 基因组 DNA 经 BamHI 酶解后与 orf 探针(依 AtuORF25/26 3′UTR 设计)杂交,除了显示~700bp 信号条带与预期的 747 bp 的相符,还有另外一个
12,000 bp 的特异信号,与预期的>905 bp 相符,表明 AtuORF25/26 3′UTR 为单一拷
贝;
膜 18B:另该杂交还观察到~3000 bp 片段(与 3013 相附),以 HindIII 消化 DAS-21023-5 基因组 DNA 与 AtuORF25/26 3′UTR 探针进行 Southern blot,显示了的 pat 表达盒完
整性。
A 示完整性和拷贝数 B 示完整性

45
图 19. DAS-21023-5 棉花 Southern blot 图谱 (示完整性)
Bgl II 酶切;orf 探针
以 BglII 消化的 DAS-21023-5 基因组 DNA 与 AtuORF25/26 3′UTR 探针进行 Southern blot,在~7500 bp 处观察到目标杂交条带,与预期的>604 bp 相符(图 19),表明外源
基因为单一拷贝。
泳道 试样 泳道 试样
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII
1 PSC355-2+pMYC3006
9 3006-BC3F4-1
2 PSC355-2 10 3006-BC3F4-2
3 3006-BC1F3-1 11 3006-BC3F4-3
4 3006-BC1F3-2 12 3006-BC4F1-3
5 3006-BC1F3-4 13 3006-BC4F1-4
6 3006-BC3F3-1 14 3006-BC4F1-5
7 3006-BC3F3-4 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5
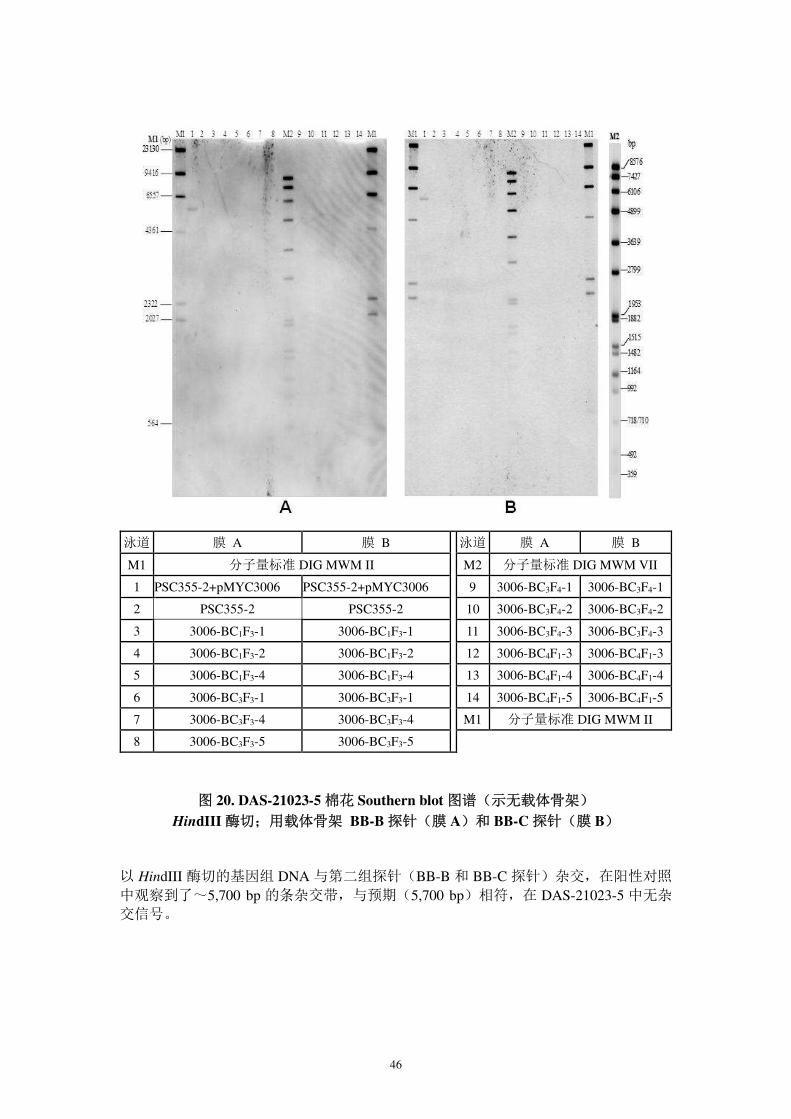
46
泳道 膜 A 膜 B
泳道 膜 A 膜 B
M1 分子量标准 DIG MWM II M2 分子量标准 DIG MWM VII
1 PSC355-2+pMYC3006 PSC355-2+pMYC3006 9 3006-BC3F4-1 3006-BC3F4-1
2 PSC355-2 PSC355-2 10 3006-BC3F4-2 3006-BC3F4-2
3 3006-BC1F3-1 3006-BC1F3-1 11 3006-BC3F4-3 3006-BC3F4-3
4 3006-BC1F3-2 3006-BC1F3-2 12 3006-BC4F1-3 3006-BC4F1-3
5 3006-BC1F3-4 3006-BC1F3-4 13 3006-BC4F1-4 3006-BC4F1-4
6 3006-BC3F3-1 3006-BC3F3-1 14 3006-BC4F1-5 3006-BC4F1-5
7 3006-BC3F3-4 3006-BC3F3-4 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5 3006-BC3F3-5
图 20. DAS-21023-5 棉花 Southern blot 图谱(示无载体骨架)
HindIII 酶切;用载体骨架 BB-B 探针(膜 A)和 BB-C 探针(膜 B)
以 HindIII 酶切的基因组 DNA 与第二组探针(BB-B 和 BB-C 探针)杂交,在阳性对照
中观察到了~5,700 bp 的条杂交带,与预期(5,700 bp)相符,在 DAS-21023-5 中无杂
交信号。

47
泳道 试样 泳道 试样
M1 分子量标准 DIG MWM II
M2 分子量标准 DIG MWM VII
1 PSC355-2+pMYC3006 9 3006-BC3F4-1
2 PSC355-2 10 3006-BC3F4-2
3 3006-BC1F3-1 11 3006-BC3F4-3
4 3006-BC1F3-2 12 3006-BC4F1-3
5 3006-BC1F3-4 13 3006-BC4F1-4
6 3006-BC3F3-1 14 3006-BC4F1-5
7 3006-BC3F3-4 M1 分子量标准 DIG MWM II
8 3006-BC3F3-5
图 21. DAS-21023-5 棉花 Southern blot 图谱 (示无载体骨架)
NcoI 酶切;载体骨架 BB-A 和 BB-D 探针
以 NcoI 酶切的基因组 DNA 与第一组探针(BB-A 和 BB-D)进行杂交,阳性对照显示~
4,400 bp 和~10,400 bp 两个杂交信号,与预期(4386 bp 和 10419 bp)相符,在 DAS-21023-5 中无杂交信号。
48
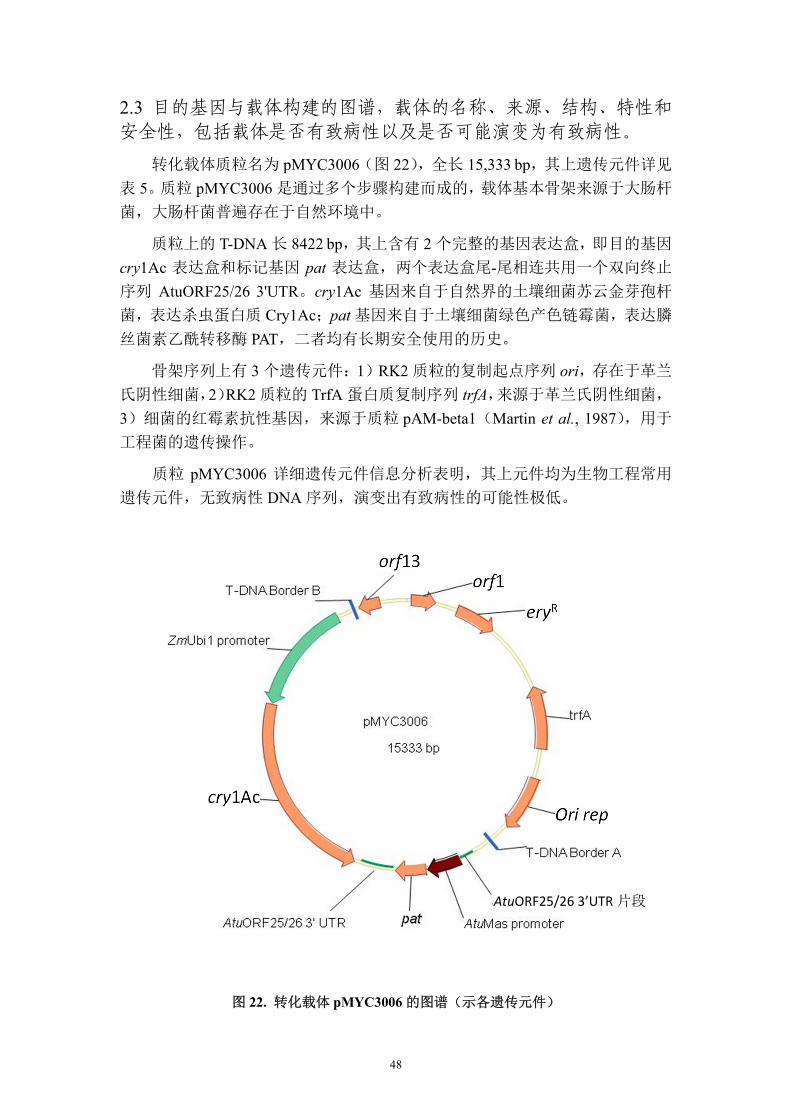
2.3 目的基因与载体构建的图谱,载体的名称、来源、结构、特性和
安全性,包括载体是否有致病性以及是否可能演变为有致病性。
转化载体质粒名为 pMYC3006(图 22),全长 15,333 bp,其上遗传元件详见
表 5。质粒 pMYC3006 是通过多个步骤构建而成的,载体基本骨架来源于大肠杆
菌,大肠杆菌普遍存在于自然环境中。
质粒上的 T-DNA 长 8422 bp,其上含有 2 个完整的基因表达盒,即目的基因
cry1Ac 表达盒和标记基因 pat 表达盒,两个表达盒尾-尾相连共用一个双向终止
序列 AtuORF25/26 3'UTR。cry1Ac 基因来自于自然界的土壤细菌苏云金芽孢杆
菌,表达杀虫蛋白质 Cry1Ac;pat 基因来自于土壤细菌绿色产色链霉菌,表达膦
丝菌素乙酰转移酶 PAT,二者均有长期安全使用的历史。
骨架序列上有 3 个遗传元件:1)RK2 质粒的复制起点序列 ori,存在于革兰
氏阴性细菌,2)RK2 质粒的 TrfA 蛋白质复制序列 trfA,来源于革兰氏阴性细菌,
3)细菌的红霉素抗性基因,来源于质粒 pAM-beta1(Martin et al., 1987),用于
工程菌的遗传操作。
质粒 pMYC3006 详细遗传元件信息分析表明,其上元件均为生物工程常用
遗传元件,无致病性 DNA 序列,演变出有致病性的可能性极低。
图 22. 转化载体 pMYC3006 的图谱(示各遗传元件)
AtuORF25/26 3’UTR 片段
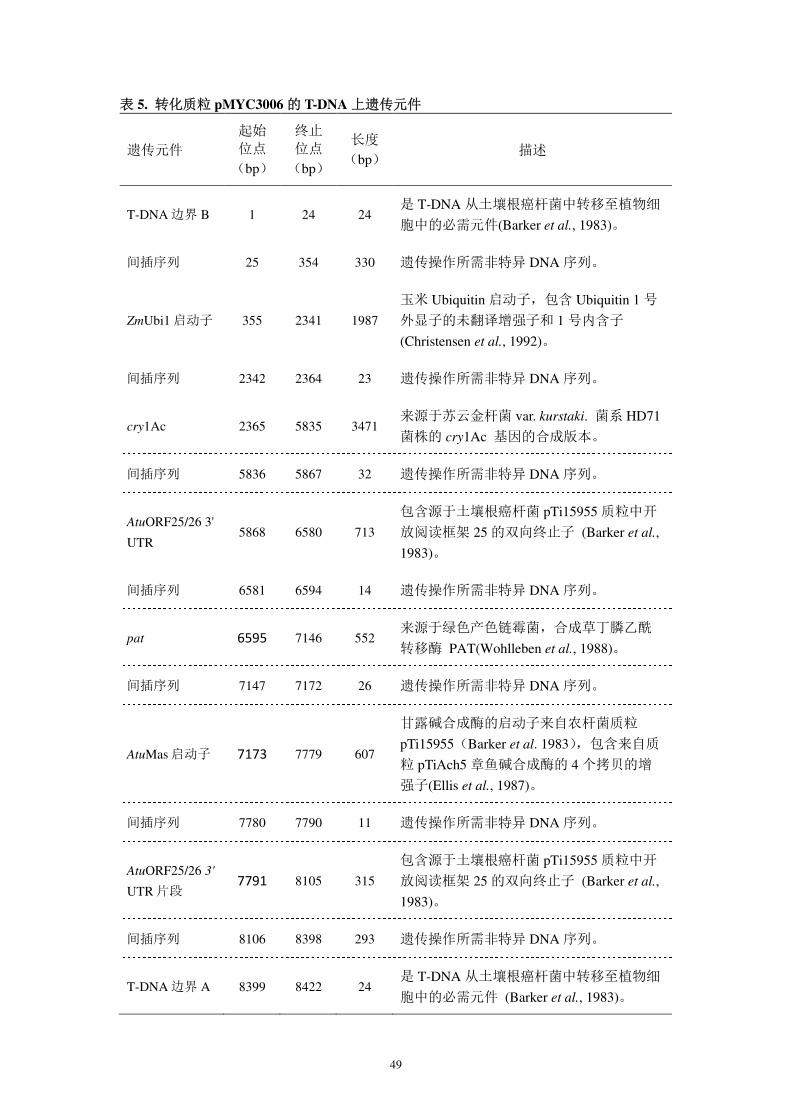
49
表 5. 转化质粒 pMYC3006 的 T-DNA 上遗传元件
遗传元件
起始 位点
(bp)
终止 位点
(bp)
长度 (bp)
描述
T-DNA 边界 B 1 24 24 是 T-DNA 从土壤根癌杆菌中转移至植物细
胞中的必需元件(Barker et al., 1983)。
间插序列 25 354 330 遗传操作所需非特异 DNA 序列。
ZmUbi1 启动子 355 2341 1987
玉米 Ubiquitin 启动子,包含 Ubiquitin 1 号
外显子的未翻译增强子和 1 号内含子
(Christensen et al., 1992)。
间插序列 2342 2364 23 遗传操作所需非特异 DNA 序列。
cry1Ac 2365 5835 3471 来源于苏云金杆菌 var. kurstaki. 菌系 HD71
菌株的 cry1Ac 基因的合成版本。
间插序列 5836 5867 32 遗传操作所需非特异 DNA 序列。
AtuORF25/26 3'
UTR 5868 6580 713
包含源于土壤根癌杆菌 pTi15955 质粒中开
放阅读框架 25 的双向终止子 (Barker et al.,
1983)。
间插序列 6581 6594 14 遗传操作所需非特异 DNA 序列。
pat 6595 7146 552 来源于绿色产色链霉菌,合成草丁膦乙酰
转移酶 PAT(Wohlleben et al., 1988)。
间插序列 7147 7172 26 遗传操作所需非特异 DNA 序列。
AtuMas 启动子 7173 7779 607
甘露碱合成酶的启动子来自农杆菌质粒
pTi15955(Barker et al. 1983),包含来自质
粒 pTiAch5 章鱼碱合成酶的 4 个拷贝的增
强子(Ellis et al., 1987)。
间插序列 7780 7790 11 遗传操作所需非特异 DNA 序列。
AtuORF25/26 3'
UTR 片段 7791 8105 315
包含源于土壤根癌杆菌 pTi15955 质粒中开
放阅读框架 25 的双向终止子 (Barker et al.,
1983)。
间插序列 8106 8398 293 遗传操作所需非特异 DNA 序列。
T-DNA 边界 A 8399 8422 24 是 T-DNA 从土壤根癌杆菌中转移至植物细
胞中的必需元件 (Barker et al., 1983)。

50
2.4 载体中插入区域各片段的资料:
转化载体 pMYC3006 的 T-DNA 上有 cry1Ac 基因表达盒(ZmUbi1 启动子、
cry1Ac 编码区、AtuORF25/26 3′UTR)和 pat 基因表达盒(AtuMas 启动子、pat
编码区、AtuORF25/26 3′UTR)。两个表达盒尾-尾相连共用一个双向终止序列,
即 T-DNA 上仅一个 AtuORF25/26 3′UTR。详见上表 5。
2.4.1 启动子和终止子的大小、功能及其供体生物的名称;
cry1Ac 表达框:
ZmUbi1 的启动子长 1987 bp,功能为启动 cry1Ac 转录,来源于玉米(Zea mays
L.)的泛素蛋白基因。
AtuORF25/26 3′UTR 序列长 713 bp,其序列含有土壤根癌杆菌 pTi15955 质
粒的开放阅读框架 25 的转录终止子和多乙酰化位点。AtuORF25/26 3′UTR 是一
个双向转录终止子,用来终止 cry1Ac 和 pat 基因的转录。
pat 基表达框:
AtuMas 启动子长 607 bp,功能为启动 pat 转录,AtuMas 启动子是农杆菌质
粒 pTi15955 中编码甘露碱(Mannopine)合成酶基因的启动子,其序列包含来自
质粒 pTiAch5 上编码章鱼碱(Octopine)合成酶基因的增强子(4 个拷贝)。
pat 与 cry1Ac 共用同一个双向终止子 AtuORF25/26 3′UTR。
2.4.2 标记基因和报告基因的大小、功能及其供体生物的名称;
选择标记基因为 pat,表达膦丝菌素乙酰转移酶 PAT。pat 基因的开放读码框
长 552 bp。PAT 蛋白质可以使植物耐受草铵膦除草剂,因此在 DAS-21023-5 棉花
开发过程中 pat 用作选择标记,PAT 的作用机理见上文第 2.1 节。
载体的 T-DNA 中无报告基因。
2.4.3 其他表达调控序列的名称及其来源(如人工合成或供体生物名
称)。
载体的 T-DNA 上无其他表达调控序列。
在构建载体的遗传操作过程中,在 T-DNA 的边界 A 内侧遗留下了
AtuORF25/26 3'UTR 片段,本片段无调控功能。
2.5 转基因方法。
采用农杆菌介导的转化方法获得了棉花转化体 DAS-21023-5。转化受体为品
系 GC510,以子叶作为外殖体,农杆菌菌株为去除致瘤性序列的 LBA4404,转
化质粒为 pMYC3006。转化流程如下所述:
从无菌条件下生长 7~10 天的 GC510 棉花幼苗上分离出子叶,切成片段;

51
子叶片段与含有 pMYC3006 质粒的农杆菌株系 LBA4404 共培养;将共培养的子
叶片段转移到含有膦丝菌素的愈伤组织诱导培养基上,选择存活的愈伤组织,筛
选转化体。愈伤组织诱导培养基还含有羧苄青霉素,可以杀死残留的农杆菌;将
存活的愈伤组织转移到胚诱导培养基上分化出胚,之后分化出转基因幼苗;将转
基因幼苗移至土壤中,在生长箱中驯化适应,最终移栽到温室。采用 Southern blot
法初步证明 cry1Ac 和 pat 基因整合至转化受体基因组。从转基因植株的叶片上
取小圆片,测试其对棉铃虫的抗性,进一步筛选获得转基因棉花转化体 DAS-
21023-5。转化、筛选过程见图 23。
将转化体 DAS-21023-5 与棉花优良品系 PCS355 回交,收获种子供进一步安
全性评价和育种使用。转基因棉花 DAS-21023-5 的选育过程见图 24。
图 23. 抗虫棉花 DAS-21023-5 转化筛选过程
构建含有cry1Ac和pat表达盒的载体pMYC3006,将其转化至脱毒的农杆菌菌株
LBA4404中
以GC510棉花为转化受体,以其子叶为外殖体,与含有pMYC3006的LBA4404菌
株共培养
培养基中加入膦丝菌素和羧苄青霉素,筛选转化细胞,并抑制农杆菌生长
评价T1代植株及其自交后代,评价内容包括分子鉴定、蛋白表达、农艺学表现、
对鳞翅目昆虫的抗性
筛选转化位点纯合体,与 PCS355 回交选育出含有转化事件 DAS-21023-5 的优良
品系
对T0代进行初步评价(不含载体骨架序列、外源基因拷贝数、耐受草铵膦)。并
自交收获T1代种子
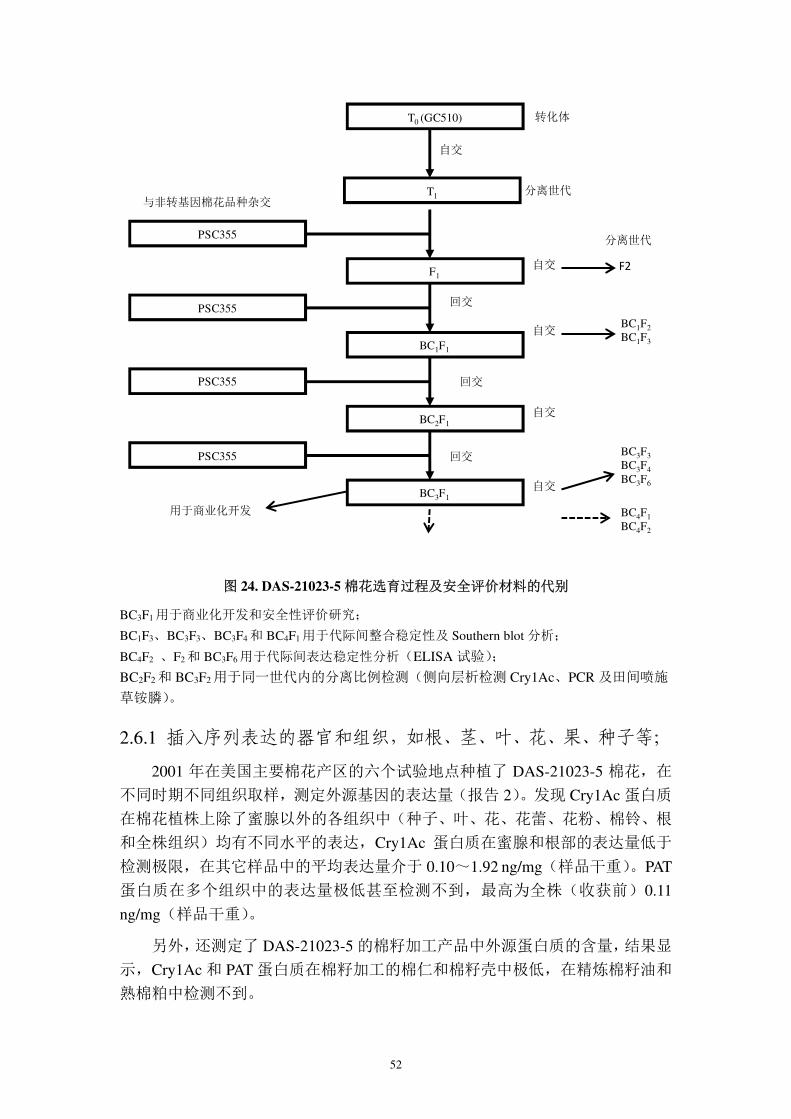
52
图 24. DAS-21023-5 棉花选育过程及安全评价材料的代别
BC3F1 用于商业化开发和安全性评价研究;
BC1F3、BC3F3、BC3F4 和 BC4F1 用于代际间整合稳定性及 Southern blot 分析;
BC4F2 、F2 和 BC3F6 用于代际间表达稳定性分析(ELISA 试验);
BC2F2 和 BC3F2 用于同一世代内的分离比例检测(侧向层析检测 Cry1Ac、PCR 及田间喷施
草铵膦)。
2.6.1 插入序列表达的器官和组织,如根、茎、叶、花、果、种子等;
2001 年在美国主要棉花产区的六个试验地点种植了 DAS-21023-5 棉花,在
不同时期不同组织取样,测定外源基因的表达量(报告 2)。发现 Cry1Ac 蛋白质
在棉花植株上除了蜜腺以外的各组织中(种子、叶、花、花蕾、花粉、棉铃、根
和全株组织)均有不同水平的表达,Cry1Ac 蛋白质在蜜腺和根部的表达量低于
检测极限,在其它样品中的平均表达量介于 0.10~1.92 ng/mg(样品干重)。PAT
蛋白质在多个组织中的表达量极低甚至检测不到,最高为全株(收获前)0.11
ng/mg(样品干重)。
另外,还测定了 DAS-21023-5 的棉籽加工产品中外源蛋白质的含量,结果显
示,Cry1Ac 和 PAT 蛋白质在棉籽加工的棉仁和棉籽壳中极低,在精炼棉籽油和
熟棉粕中检测不到。
T0 (GC510)
F1
BC1F1
BC2F1
PSC355
PSC355
PSC355
BC3F1
PSC355
转化体
自交
回交
回交
回交
自交BC1F2BC1F3
自交
自交
BC3F3BC3F4BC3F6
BC4F1BC4F2
与非转基因棉花品种杂交
分离世代
T1 分离世代
自交
用于商业化开发
F2

53
2.6.2 插入序列的表达量及其分析方法;
1)未加工产品中 Cry1Ac 蛋白质的表达量(报告 2):
Cry1Ac 在不同生长期不同组织中的表达量高低不同:在棉籽中含量为 0.57
±0.09 ng/mg 组织干重,以该值进行后续的接触风险分析;在蜜腺中检测不到;
在收获前的根部表达量为 0.05 ng/mg 组织干重,低于定量极限;在花器官和 3~
6 周的幼叶中表达量最高,均为 1.92 ng/mg 组织干重(表 6)。
表 6. DAS-21023-5 棉花中 Cry1Ac 蛋白质在不同时期不同组织的表达量
棉花组织(生长期) Cry 1Ac(ng/mg a) 定量极限
LOQ (ng/mg) 平 均 值 b 标 准 差 最小值~最大值
幼叶(3~6 周) 1.92 0.7 0.46~3.5 0.1
顶端叶 1.44 0.5 0.24~2.4
花 1.92 0.3 1.3~2.4 0.1
棉蕾 1.84 0.5 1.0~3.1 0.1
棉铃 0.77 0.2 0.46~1.1 0.1
全株(苗期) 1.59 0.4 0.8~2.2 0.1
全株(授粉期) 1.15 0.5 0.57~2.1
全株(收获前) 0.81 0.3 0.31~1.3 0.1
根(苗期) 0.20 0.1 (0.09)~0.44
0.25 根(授粉期) 0.10 0.07 ND~0.23
根(收获前) (0.05) 0.04 ND~0.11
花粉 1.44 0.5 0.9~2.4 0.1
蜜腺 ND NA NA -
棉籽 0.57 0.09 (0.33)~0.78 0.375
a:花粉、蜜腺、棉籽中用鲜重表示,其它组织中表达量以干重表示。 b:6 个试验地点样品的平均值。 ():括号中的值处于检测极限(LOD)和定量极限(LOQ)之间。 ND(not detectable):低于检测极限。 NA(not applicable):不适用。
2)未加工产品中 PAT 蛋白质的表达量(报告 2):
PAT 在不同时期不同组织中的表达量普遍较低甚至检测不到,在棉籽中表达
量为 0.06±0.06 ng/mg 组织干重,以该值进行后续的接触风险分析,在全株(收
获前)表达量为 0.11±0.05 ng/mg 组织鲜重;其他组织如花、棉蕾、棉铃、开花
授粉期的全株中均检测不到,即低于检测极限,见表 7。
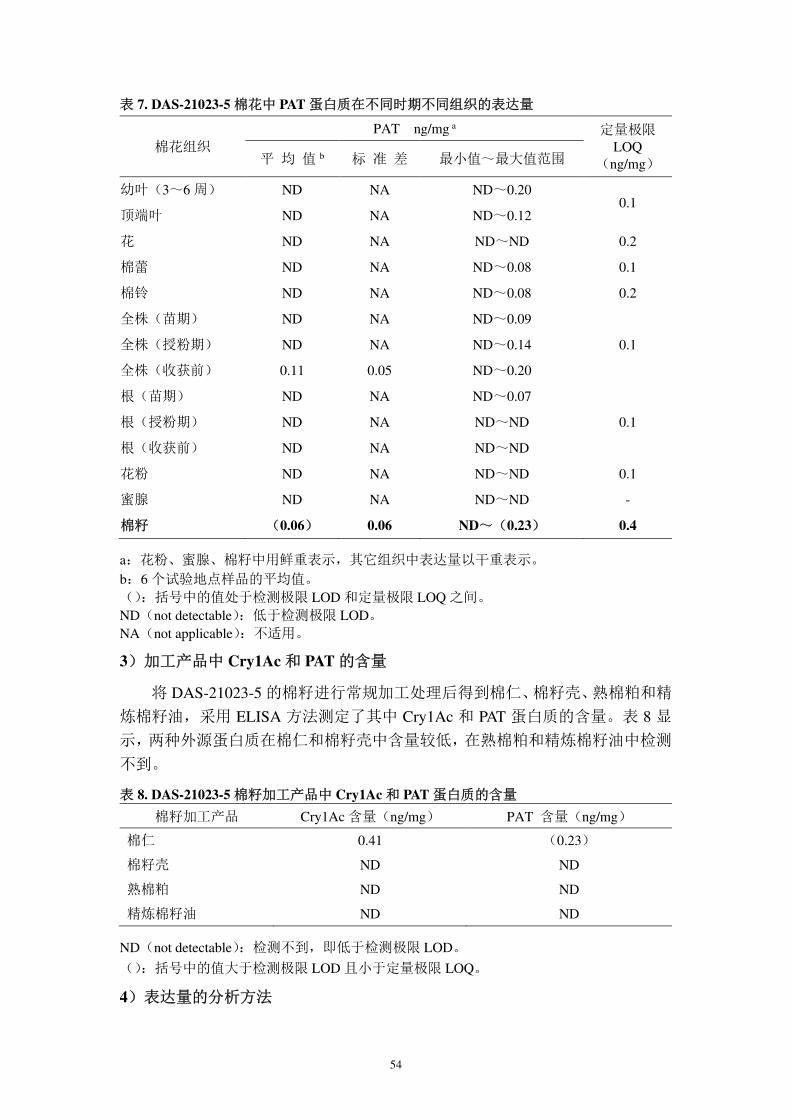
54
表 7. DAS-21023-5 棉花中 PAT 蛋白质在不同时期不同组织的表达量
棉花组织 PAT ng/mg a 定量极限
LOQ (ng/mg) 平 均 值 b 标 准 差 最小值~最大值范围
幼叶(3~6 周) ND NA ND~0.20 0.1
顶端叶 ND NA ND~0.12
花 ND NA ND~ND 0.2
棉蕾 ND NA ND~0.08 0.1
棉铃 ND NA ND~0.08 0.2
全株(苗期) ND NA ND~0.09
0.1 全株(授粉期) ND NA ND~0.14
全株(收获前) 0.11 0.05 ND~0.20
根(苗期) ND NA ND~0.07
0.1 根(授粉期) ND NA ND~ND
根(收获前) ND NA ND~ND
花粉 ND NA ND~ND 0.1
蜜腺 ND NA ND~ND -
棉籽 (0.06) 0.06 ND~(0.23) 0.4
a:花粉、蜜腺、棉籽中用鲜重表示,其它组织中表达量以干重表示。 b:6 个试验地点样品的平均值。 ():括号中的值处于检测极限 LOD 和定量极限 LOQ 之间。 ND(not detectable):低于检测极限 LOD。 NA(not applicable):不适用。
3)加工产品中 Cry1Ac 和 PAT 的含量
将 DAS-21023-5 的棉籽进行常规加工处理后得到棉仁、棉籽壳、熟棉粕和精
炼棉籽油,采用 ELISA 方法测定了其中 Cry1Ac 和 PAT 蛋白质的含量。表 8 显
示,两种外源蛋白质在棉仁和棉籽壳中含量较低,在熟棉粕和精炼棉籽油中检测
不到。
表 8. DAS-21023-5 棉籽加工产品中 Cry1Ac 和 PAT 蛋白质的含量
棉籽加工产品 Cry1Ac 含量(ng/mg) PAT 含量(ng/mg)
棉仁 0.41 (0.23)
棉籽壳 ND ND
熟棉粕 ND ND
精炼棉籽油 ND ND
ND(not detectable):检测不到,即低于检测极限 LOD。
():括号中的值大于检测极限 LOD 且小于定量极限 LOQ。
4)表达量的分析方法

55
从不同地点取样,采用 ELISA 方法检测不同时期、不同组织中外源蛋白质
的含量。
种植地点及田间设计
2001年经美国农业部批准在美国主要棉花产区的六个试验地点种植了DAS-
21023-5 及对照棉花。试验地点位于美国亚利桑那州(AZ)、加利福尼亚州(CA)、
密西西比州(MS)、北卡罗来纳州(NC)和得克萨斯州(TX1 和 TX2 两处),
这些地点代表了美国棉花多样化的农事操作类型与生态环境。每试验点设 3 次重
复。播种转基因和对照棉花的小区与缓冲区棉田间隔至少 200 米,用界桩区分
(报告 2)。
取样和制样
在不同生长期取不同组织试样,时期及取样部位为:幼叶(3~6 周)、顶叶、
棉蕾、花、全株(苗期)、全株(授粉期)、全株(收获前)、根(苗期)、根(授
粉期)、根(收获前)、棉铃、棉籽、花粉及花蜜,详见表 6 和表 7。
在田间取样后将样品冻存于-80℃冰箱。分析前将样品粗磨、冻干、细磨制
样(棉籽油中提取目标蛋白质的方法略有不同)。使用含有 0.05% Tween-20 和 1%
聚乙烯吡咯烷酮(PVP)的磷酸缓冲液(PBST/PVP)从棉花组织中提取 Cry1Ac
和 PAT 蛋白质,萃取液用 PBST/PVP 稀释后待用。
用已知表达 Cry1Ac 或 PAT 蛋白质的棉花组织提取液体系作为阳性对照;用
已知不表达 Cry1Ac 或 PAT 蛋白质的棉花组织提取液体系作为阴性对照;以不含
上述蛋白质的缓冲液体系作空白对照。在试验开展之前,采用侧向层析试纸条或
Western blot 方法验证试样是否含有 Cry1Ac 或 PAT 蛋白质。
ELISA 体系建立
采用“三明治式”ELISA 测定转基因棉花 DAS-21023-5 提取液可溶物中
Cry1Ac 和 PAT 蛋白质的含量。
纯化 Cry1Ac 特异的单克隆抗体,将一个单克隆抗体包被在反应孔内,另一
个单克隆抗体与酶偶联作为检测抗体。Cry1Ac 蛋白质 ELISA 试剂盒购自美国
Strategic Diagnostic Inc 公司。
同理,纯化 PAT 特异抗体,将多克隆抗体包被在反应孔内,单克隆抗体与酶
偶联作为检测抗体。PAT 蛋白质 ELISA 试剂盒购自美国 Envirologix, Inc 公司。
ELISA 检测方法详见第六部分:转基因性状及产物的检测和鉴定技术。
标准曲线及 ELISA 操作步骤
绘制标准曲线时,Cry1Ac 标准蛋白质的浓度梯度设为 0.5~10.0 ng/mL,PAT
标准蛋白质的浓度梯度设为 0.375~10.0 ng/mL。按不同浓度及其吸光度绘制标
准曲线。分析时,在 ELISA 微量滴定板的各孔中加入辣根过氧化酶标记的检测
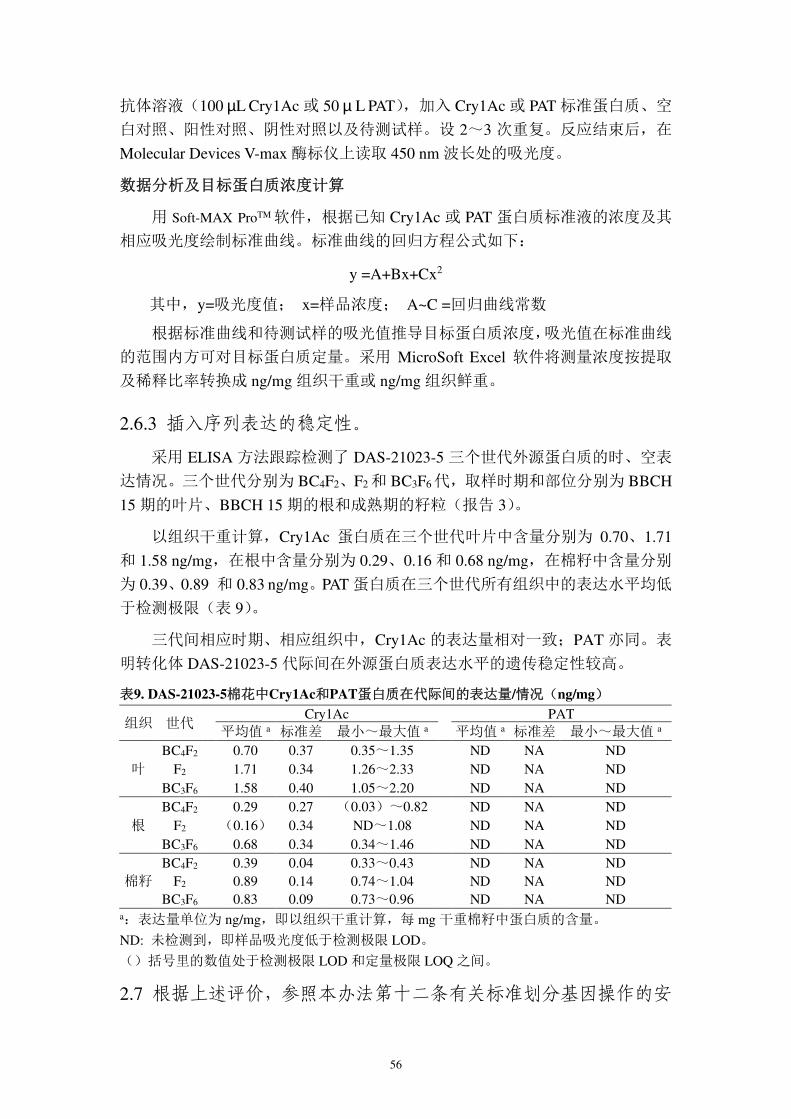
56
抗体溶液(100 µL Cry1Ac 或 50 µ L PAT),加入 Cry1Ac 或 PAT 标准蛋白质、空
白对照、阳性对照、阴性对照以及待测试样。设 2~3 次重复。反应结束后,在
Molecular Devices V-max 酶标仪上读取 450 nm 波长处的吸光度。
数据分析及目标蛋白质浓度计算
用 Soft-MAX ProTM 软件,根据已知 Cry1Ac 或 PAT 蛋白质标准液的浓度及其
相应吸光度绘制标准曲线。标准曲线的回归方程公式如下:
y =A+Bx+Cx2
其中,y=吸光度值; x=样品浓度; A~C =回归曲线常数
根据标准曲线和待测试样的吸光值推导目标蛋白质浓度,吸光值在标准曲线
的范围内方可对目标蛋白质定量。采用 MicroSoft Excel 软件将测量浓度按提取
及稀释比率转换成 ng/mg 组织干重或 ng/mg 组织鲜重。
2.6.3 插入序列表达的稳定性。
采用 ELISA 方法跟踪检测了 DAS-21023-5 三个世代外源蛋白质的时、空表
达情况。三个世代分别为 BC4F2、F2 和 BC3F6代,取样时期和部位分别为 BBCH
15 期的叶片、BBCH 15 期的根和成熟期的籽粒(报告 3)。
以组织干重计算,Cry1Ac 蛋白质在三个世代叶片中含量分别为 0.70、1.71
和 1.58 ng/mg,在根中含量分别为 0.29、0.16 和 0.68 ng/mg,在棉籽中含量分别
为 0.39、0.89 和 0.83 ng/mg。PAT 蛋白质在三个世代所有组织中的表达水平均低
于检测极限(表 9)。
三代间相应时期、相应组织中,Cry1Ac 的表达量相对一致;PAT 亦同。表
明转化体 DAS-21023-5 代际间在外源蛋白质表达水平的遗传稳定性较高。
表9. DAS-21023-5棉花中Cry1Ac和PAT蛋白质在代际间的表达量/情况(ng/mg)
a:表达量单位为 ng/mg,即以组织干重计算,每 mg 干重棉籽中蛋白质的含量。
ND: 未检测到,即样品吸光度低于检测极限 LOD。
()括号里的数值处于检测极限 LOD 和定量极限 LOQ 之间。
2.7 根据上述评价,参照本办法第十二条有关标准划分基因操作的安
组织 世代 Cry1Ac PAT 平均值 a 标准差 最小~最大值 a 平均值 a 标准差 最小~最大值 a
叶 BC4F2 0.70 0.37 0.35~1.35 ND NA ND
F2 1.71 0.34 1.26~2.33 ND NA ND BC3F6 1.58 0.40 1.05~2.20 ND NA ND
根 BC4F2 0.29 0.27 (0.03)~0.82 ND NA ND
F2 (0.16) 0.34 ND~1.08 ND NA ND BC3F6 0.68 0.34 0.34~1.46 ND NA ND
棉籽 BC4F2 0.39 0.04 0.33~0.43 ND NA ND
F2 0.89 0.14 0.74~1.04 ND NA ND BC3F6 0.83 0.09 0.73~0.96 ND NA ND

57
全类型。
上述遗传操作未增加或降低生物体的安全性,依《农业转基因生物安全评价
管理办法》第二章第十二条,本遗传操作属于类型 2。
3 转基因植物的安全性评价
3.1 转基因植物的遗传稳定性
转化体的遗传稳定性是其生物安全稳定性的基础。代际间 DAS-21023-5 可
在 DNA 整合水平、蛋白质表达水平和性状表现水平稳定遗传。在同一世代的个
体间,外源基因整合位点符合单一位点孟德尔遗传分离比例,在个体间证明了遗
传稳定性。
代际间 DNA 整合水平的遗传稳定性
为了证明 DAS-21023-5 棉花中外源序列在不同代别间稳定整合,对 BC1F3、
BC3F3、BC3F4和 BC4F1 四个世代植株进行了 Southern blot 跟踪验证。以 T-DNA
转化片段所有遗传元件序列设计探针(以及质粒 pMYC3006骨架序列设计探针),
与多种内切酶消化的DAS-21023-5棉花基因组DNA进行Southern blot(如HindIII、
NcoI、PacI、XhoI 和 BamHI)。四个世代的杂交结果相同,外源片段均为单位点、
单拷贝、结构完整地整合至棉花基因组中,且转化体中无转化质粒的骨架序列。
表明外源序列在 DAS-21023-5 棉花的多个世代中可稳定遗传(详见前文 2.2.5 节
或报告 1E)。
代际间蛋白质表达水平的遗传稳定性
跟踪检测了 DAS-21023-5 三个世代 BC4F2、F2 和 BC3F6 外源蛋白质的时、
空表达情况。取样时期和部位分别为 BBCH 15 期的叶片、BBCH 15 期的根和成
熟期籽粒,采用 ELISA 方法测定 Cry1Ac 和 PAT 蛋白质的表达量。三代间不同
时期的相应组织中,无论是 Cry1Ac 还是 PAT 的表达量均相对一致。表明转化体
DAS-21023-5 代际间在外源蛋白质表达水平可稳定遗传(详见前文第 2.6.3 节或
报告 3)。
代际间性状表现水平的遗传稳定性
在 2001-2004 年进行了多年、多世代、多地点的田间试验和实验室研究,验
证了 DAS-21023-5 抗虫效果的稳定性。
以 DAS-21023-5 与 PSC355 回交第 BC3F4 代为实验材料,2001 年六个田间
试验点调查了对主要鳞翅目害虫防治效果。结果显示,与非转基因对照 PSC355
相比,在重复人工接种情况和自然发生情,DAS-21023-5 棉花对棉铃虫和烟草夜
蛾具有高度的抗性;对棉红铃虫有符合商业要求的抗性;对甜菜夜蛾表现出中度
的抗性(报告 4A)。2002 年在亚利桑那州和新墨西哥州的实验结果显示,与非转
基因对照 PSC355 相比,人工接种条件下,DAS-21023-5 可以有效抵抗棉红铃虫。
58
在人工接种甜菜夜蛾并在接种后 3~6 天(种植后 75 天)调查,发现在 DAS-
21023-5 棉花叶片上的甜菜夜蛾数量比 PSC355 上的数量显著减少(报告 4B)。
同一世代个体间的遗传稳定性
在 DNA 整合水平、蛋白质表达水平及性状表现水平验证了同一世代个体间
的遗传稳定性,试验方法有:DAS-21023-5 转化事件特异性 PCR,侧向层析试纸
条白质检测方法(Lateral Flow Strip)及苗期喷施草铵膦法。以上 3 种方法均测
得插入基因分离比例符合单一位点的孟德尔分离比例,即转化体中外源插入基因
可稳定遗传。
DAS-21023-5 棉花 T1 代与非转基因棉花 PSC355 杂交获得 BC1F1,继续回交
和自交获得 BC4F2。以 BC4F2 为实验群体,采用 DAS-21023-5 转化事件特异性
PCR,来判断该群体整合位点的分离情况。结果显示,所测 121 植株中,有 84 株
为 PCR 阳性(有外源基因),37 株为阴性(无外源基因)。比例与 3:1 差异不显
著,χ2 值为 2.008,小于临界值 3.841(表 10),该位点符合单一位点的孟德尔遗
传模式(报告 1F)。
使用上述 121 个单株采用侧向层析试纸条检测了 BC4F2 代群体在 Cry1Ac 蛋
白质表达水平的分离情况。其中 84 株为 Cry1Ac 蛋白质阳性(表达),37 株为阴
性(不表达),比例与 3:1 差异不显著,统计量 χ2 为 2.008,小于临界值 3.841(表
10),因此该位点符合单一位点的孟德尔遗传模式。三株 PSC355 对照全为阴性
(不表达),表明本检测体系有效(报告 1F)。
以 BC3F2 为实验材料,以回交受体 PSC355 为非转基因对照,在苗期喷施草
铵膦,统计抗感比例。共调查了 110 株,其中 88 株耐受草铵膦,22 株对该除草
剂敏感,抗感比例与 3:1 差异不显著,统计量 χ2 为 0.142,小于临界值 3.841(表
10),因此,该位点符合单一位点的孟德尔遗传模式。
上述三个层次的试验结果均显示,分离世代中插入序列的遗传模式符合单一
位点的孟德尔分离比例(3:1)。证明了外源基因在同一群体内个体间的遗传稳定
性。
表 10. 外源基因在分离世代个体间的遗传稳定性(示分离比例)
分析方法 分离世代 观察个数 期望个数
χ2 值 阳性频数 阴性频数 阳性频数 阴性频数
转化事件特异性 PCR BC4F2 84 37 91 30 2.008
Cry1Ac 侧向层析试纸条 BC4F2 84 37 91 30 2.008
苗期喷施草铵膦 BC3F2 88(抗) 27(感) 86(抗) 29(感) 0.142
χ20.05(1)=3.841。
3.2 转基因植物与受体或亲本植物在环境安全性方面的差异:
大量田间试验和实验室试验表明,DAS-21023-5 棉花仅引入了抵抗鳞翅目昆

59
虫和耐草铵膦(筛选标记)的特性,在生殖方式与生殖率、传播方式和传播能力、
休眠期、适应性、农艺学性状、对环境中有益或有害生物(非靶标生物)的影响
等方面,该转基因棉花与其非转基因对照实质等同;外源基因从 DAS-21023-5 转
移至其它生物的可能性极低,转移的概率与非转基因对照等同;DAS-21023-5 演
变为杂草的可能性极低,且演变为杂草的可能性与其非转基因对照等同。尚未发
现该转化体对生态环境有负面作用,因此本转化体与其非转基因对照在环境安全
方面等同。
在转基因棉花品系 DAS-21023-5 选育过程中,以非转基因传统对照棉花
PSC355 为轮回亲本将该转化事件回交转育至 PSC355 背景中,育成转基因棉花
DAS-21023-5 品系。PSC355 与 DAS-21023-5 具有相似的(可比的)遗传背景,
因此以 PSC355 作为安全评价的非转基因对照。
3.2.1 生殖方式和生殖率;
DAS-21023-5 棉花中仅引入了抗虫和耐受除草剂(筛选标记)的性状,无预
期改变其生殖方式和生殖率。
通常栽培棉花为一年生作物,生殖方式是以自花授粉为主,有昆虫时可异花
授粉,受精后发育成种子,种子是其繁殖和传播的载体(McGregor, 1976; Umbeck
et al., 1991)。与传统非转基因棉花品系一样,转基因棉花 DAS-21023-5 仍按照
授粉-受精-结实的生殖方式繁殖。
实验表明 DAS-21023-5 棉花的生殖率(即繁育系数,由单铃籽棉重、百粒重
等指标指示)没有变化。2002 年在美国主要棉花带地区设置 21 个田间试验点,
调查了转基因棉花的农艺性状,发现 DAS-21023-5 棉花与其非转基因对照的繁
育系数无明显区别(报告 5)。本实验按完全随机区组排列,设置 4 次重复。整个
试验均采用地区性一致的的农艺措施。以轮回本 PSC355 为非转基因对照。从表
14 可见,单铃籽棉重、百粒重在转基因棉花 DAS-21023-5 与非转基因对照间差
异微小,即二者产生后代的数量无明显区别。表明生殖率并未因外源基因的引入
而变化。
理论依据和实证数据均表明,该转化体不会通过生殖方式改变或生殖率提高
而提高生存竞争力,从而对环境造成负面影响。
3.2.2 传播方式和传播能力;
与传统棉花一样,转基因棉花可能会通过植物体、种子或花粉传播。经理论
分析和试验数据支持,转基因棉花DAS-21023-5及其非转基因对照的传播方式和
传播能力实质等同。
植物体传播能力无变化。首先,高度驯化导致栽培棉花离开人类的养护就难
以存活。栽培棉花是一年生植物,在常规栽培条件下,主要以种子形式越冬并在
来年发芽。残留在田间未消毁或未收获的棉杆不能再生。另外,转化体如果获得

60
了更强的生存竞争能力,其通过植物体传播的可能性将提高,但2002年的田间试
验显示,转化体与其非转基因对照的农艺学性状大体等同(表14),该田间试验
后连续12个月的田间观察也没有检测到任何自生苗。可见,引入外源基因并在转
化体中表达并未改变本转化体通过植物体传播的能力。
种子传播能力无变化。本公司检测了DAS-21023-5棉花种子的发芽率,比较
了种子低温活力、暖温4天时的发芽率、暖温7天时的发芽率、总发芽率及休眠种
子比例(表13),结果显示,除DAS-21023-5的低温活力比非转基因的低以外,上
述指标在二者间无显著差异,表明转基因与非转基因棉籽对照相比,并没有提高
其发芽率和休眠特性。另一方面,表14显示转基因棉花与非转基因对照的产量(由
结铃率、百粒重等指示)无明显差异,表明二者繁殖系数相同。
由于DAS-21023-5种子休眠特征没有改变,因此不会提高逃避逆境并在适宜
环境中萌发的能力;繁殖系数没有提高,不会散布更多种子到环境中;种子萌发
率没有提高,不会有更多的后代存活于环境中。因此,DAS-21023-5棉花通过种
子传播的能力与非转基因对照等同。
花粉传播能力无变化。棉花以自花授粉为主,品种间异交概率较低,
McGregor曾在蜜蜂聚集的棉田周围追踪过棉花花粉的漂移,发现在距离棉田(花
粉源)46~61米处仅有1.6%的花粉(McGregor,1976)。由于生殖隔离和地理隔
离,DAS-21023-5的遗传物质通过花粉转移到其它棉种、野生近缘种、野生种及
杂草中的概率极低。
花粉与转化体的传播密切相关,因此花粉特征成为评价传播方式和传播能力
的重要指标。本公司比较了 DAS-21023-5 棉花和非转基因对照的花粉活力和形
状大小,结果显示二者的花粉特征无显著差异,可推测出遗传转化并未改变本转
化体的传播方式和传播能力(报告 6),实验主要内容如下所述。
收集 10 个单株上的花粉,采用改良的 Alexander 染液(Bonhomme et al.,
EMBO, Alexander, 1969; Alexander, 1980)染色花粉 24 小时。Alexander 染液含有
酸性品红,可将花粉粒的细胞质染成红色,并可将细胞壁染成孔雀绿(Atlagić et
al., 2012)。有活性的花粉细胞含有健康的细胞壁和细胞质,因此会被染成深红色
并包围一层绿色薄边;无活性的花粉粒没有完整的细胞质,孔雀绿进入细胞壁,
仅呈现一个中空的绿色圆环。采用 Leica DM5000B 直立复式显微镜放大 100 倍
观察染色的花粉,使用 Leica DCF 7000 T 数码照相机和 Leica 系列软件(4.6 版)
成像。转基因和非转基因对照均取多个视野。运用 ImageJ 图像处理软件对光学
显微图中的花粉活性打分。
观察了 DAS-21023-5 花粉(1503 粒)和非转基因对照花粉(1465 粒),二者
有活力花粉比例分别占 98.7%和 99.9%(表 11),差异极微小。表 12 比较了花粉
大小,抽取至少 1200 粒花粉,放大 100 倍,用 ImageJ 软件标注花粉大小。转基
因棉花与非转基因对照的平均 Feret 径极近,分别为 118.83 μm 和 118.82 μm。图
25 可直观得看到,DAS-21023-5 与其对照花粉型态相同,二者花粉粒均可染,且
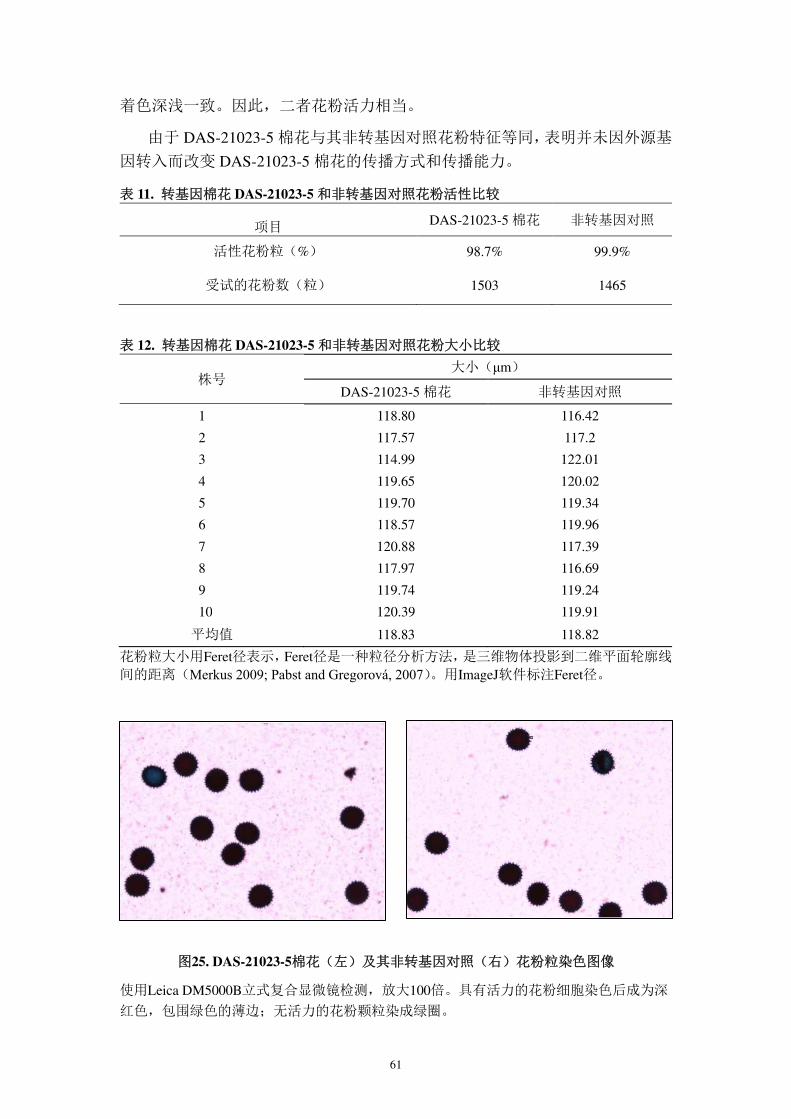
61
着色深浅一致。因此,二者花粉活力相当。
由于 DAS-21023-5 棉花与其非转基因对照花粉特征等同,表明并未因外源基
因转入而改变 DAS-21023-5 棉花的传播方式和传播能力。
表 11. 转基因棉花 DAS-21023-5 和非转基因对照花粉活性比较
项目 DAS-21023-5 棉花 非转基因对照
活性花粉粒(%) 98.7% 99.9%
受试的花粉数(粒) 1503 1465
表 12. 转基因棉花 DAS-21023-5 和非转基因对照花粉大小比较
株号 大小(μm)
DAS-21023-5 棉花 非转基因对照
1 118.80 116.42
2 117.57 117.2
3 114.99 122.01
4 119.65 120.02
5 119.70 119.34
6 118.57 119.96
7 120.88 117.39
8 117.97 116.69
9 119.74 119.24
10 120.39 119.91
平均值 118.83 118.82
花粉粒大小用Feret径表示,Feret径是一种粒径分析方法,是三维物体投影到二维平面轮廓线
间的距离(Merkus 2009; Pabst and Gregorová, 2007)。用ImageJ软件标注Feret径。
图25. DAS-21023-5棉花(左)及其非转基因对照(右)花粉粒染色图像
使用Leica DM5000B立式复合显微镜检测,放大100倍。具有活力的花粉细胞染色后成为深
红色,包围绿色的薄边;无活力的花粉颗粒染成绿圈。
62
3.2.3 休眠期;
种子改变休眠期,可以逃避逆境并在适宜环境中萌发,本转化体休眠特性与
其非转基因对照实质等同。本公司于 2002 年棉花结实后,测试并比较了子代种
子的发芽率,设置低温与暖温两种处理,在暖温 4 天和 7 天时计数、统计发芽
率,并计算休眠种子的比例(报告 5)。方差分析显示,在暖温处理条件下,DAS-
21023-5 棉花与非转基因对照的发芽率无显著差异,即 P 值均大于 0.05;在低温
条件下,DAS-21023-5 棉花相对于非转基因对照的发芽率虽有差异,但转化体萌
发率低于对照; DAS-21023-5 棉花与非转基因对照间休眠种子比例无显著差异
(表 13)。因此,DAS-21023-5 棉花中插入序列及其表达并没有提高棉花的发芽
率,即本转化体不会通过改变休眠特征的方式来逃避逆境,不会因此提高生存竞
争能力。
表 13. DAS-21023-5 与其非转基因对照棉花的发芽率比较
条件 DAS-21023-5
发芽能力(%) 非转基因对照 发芽能力(%)
P 值
冷温 32 38 <0.05
暖温发芽率(4 天时) 64 65 >0.05
暖温发芽率(7 天时) 80 82 >0.05
总发芽率 85 87 >0.05
休眠种子 0.6 0.3 >0.05
3.2.4 适应性;
植物体对环境的适应性是综合表现,体现在生物体能够适应当前的环境并正
常生长、繁殖。
2002 年以棉花 DAS-21023-5 及其非转基因对照 PSC355 为材料进行了大规
模的田间试验(报告 5),对出苗率、开花期、生殖潜力及皮棉产量进行了调查
(表 14),并对发芽能力进行了分析(表 13),上述各项指标在转基因和其非转
基因对照之间差异微小,即二者农艺学特征实质等同。又因为二者生殖方式和生
殖率等同(前文 3.2.1),传播方式和传播能力无差异(前文 3.2.2),休眠特征相
同(前文 3.2.3),因此,转基因棉花 DAS-21023-5 的适应性并未高过传统非转基
因棉花。
3.2.5 生存竞争能力;
悠久的栽培历史证明,栽培棉花在自然生态环境下无竞争优势,不具有生物
入侵性,未见其自然演变成优势物种。即使在运输过程中无意遗撒,只能在适宜
的环境中萌发形成自已的群落。
2002 年在美国的田间试验显示,DAS-21023-5 的生存竞争性不高于其非转
基因棉花的生存竞争性(报告 5)。具体体现在种子活力、生长习性、开花习性及

63
植株活力、繁殖特性及产量、休眠期、抗病和抗虫能力等方面。
种子活力方面:田间出苗率和种子低温活力两个指标体现了种子活力,DAS-
21023-5 的两个指标均低于非转基因对照(表 13 和表 14)。即 DAS-21023-5 的种
子活力不高于其非转基因对照,转基因棉花不会通过提高种子活力使更多个体存
活在环境中从而提高竞争能力。
生长习性方面:试验调查了株高、总节数、株高与果节数比例、第一果枝上
的果节数、果枝、成铃位置总数及营养铃数。DAS-21023-5 与其非转基因对照就
上述指标差异微小(表 14)。因此,转基因棉花的生长势并未高于非转基因棉花。
开花习性方面:调查了初花期、15 天时白花苤节数和 30 天时白花苤节数,
这些参数在转基因棉花 DAS-21023-5 与非转基因对照间差异微小,二者生育期
无明显差别(表 14)。另一方面,DAS-21023-5 与其非转基因对照的花粉特征无
差异,即二者传播方式和传播能力等同(表 11 和表 12)。上述结果均表明转基因
棉花不会提前或推后开花期和成熟期而逃避逆境,亦不会提高花粉的传播能力来
散布更多后代,并以此提高其在自然界的竞争性。
繁殖特性及产量方面:结铃率、第一位置结铃率、棉铃开裂率、单铃籽棉重、
衣分、百粒重、及皮棉产量指示了繁殖及产量特征,从表 14 可见,就上述指标,
在转基因棉花与其非转基因对照间差异极微小。表明转基因棉花不会产生更多的
种子从而提高竞争性。
种子休眠性状:种子休眠是一种有效的生存机制,可使个体避开极端环境并
在适宜环境中发芽,发芽率是植物休眠性状的一个重要指标。不同温度条件的发
芽试验显示,DAS-21023-5 并未提高棉花的发芽率(表 13),即 DAS-21023-5 棉
花并未通过休眠来逃避逆境从而提高其生存竞争的能力。
抗病、虫能力方面:自 1999 年起在美国和波多黎哥棉花主产区调查了
DAS-21023-5 的病、虫害发生情况。育种家在田间目测并比较了 DAS-21023-5
与其非转基因对照间病、虫害发生情况,发现除靶标昆虫外,未见二者有明显
差别(USDA, 2003,
https://www.aphis.usda.gov/biotechnology/petitions_table_pending.shtml)。
自生苗监测:2003 年, 在美国六个棉花的主产区(亚利桑那州、加利福尼亚、
密西西比州,北卡罗莱纳和德克萨斯的 2 个地点)种植了 DAS-21023-5 及其非
转基因对照棉花,并调查了自生苗发生情况。于收获后的下一个生长季对所有小
区的自生苗进行观察,每月定期监测,持续 12 个月或者连续两次监测未发现有
自生苗则停止监测。如若发现自生苗,则采取喷洒有效除草剂、犁地或者手动拔
苗等方式去除。6 个地点历时 12 个月监测数据显示并未发现自生苗。
综上所述,DAS-21023-5 棉花在环境中的生存竞争能力并未高于其非转基因
对照。

64
表 14. 转基因抗虫棉花与非转基因对照棉花间农艺学特征比较
主要农艺 性状
指标(单位) 转基因棉 DAS-21023-5
非转基因对照 PSC355
试验地点数目
出苗率 田间出苗率% 63.6 82.3 19
种子低温活力% 32 38 20
生长习性
株高(英寸) 39.9 41.5 15
总节数(节/株) 17.4 17.6 16
株高与果节数比例(英寸/节) 2.29 2.35 15
第一果枝上的果节数(节) 6.7 6.6 16
果枝(个/株) 11.7 12.1 16
成铃位置总数(个/株) 25.6 26.6 16
营养铃数(个/株) 2.3 1.6 15
营养活力 营养枝(个/株) 2.9 2.6 15
开花习性
初花期(天) 61.9 60.6 18
15 天时白花苤节数(节) 12.9 12.9 17
30 天时白花苤节数(节) 17.0 16.8 15
繁殖潜力
结铃率(%) 49.0 44.4 16
第一位置结铃率(%) 58.5 54.3 16
单株棉铃开裂率(%/株) 73.5 75.4 17
单铃籽棉重(g) 5.5 5.1 19
衣分(%) 37.9 37.3 19
百粒重(g ) 11.0 10.7 18
皮棉产量(英磅/英亩) 1005 993 17
3.2.6 转基因植物的遗传物质向其它植物、动物和微生物发生转移的
可能性;
科学资料和实验数据均表明,DAS-21023-5 棉花中遗传物质向其他植物、动
物和微生物发生转移的可能性极低,且转移概率与非转基因棉花的等同。
1)转基因与非转基因棉花向其它植物授粉的概率相同
通过实验比较了 DAS-21023-5 棉花及其非转基因对照花粉的活力和形状大
小,结果显示二者的花粉特征无显著差异(报告 6),因此二者通过花粉传播的概
率等同。即使 DAS-21023-5 棉花可向其它品种、种、野生近缘种、锦葵科其它属
授粉,其概率与非转基因的相同。
2)向其它种、野生近缘种、锦葵科其它属垂直转移的可能性极低
在自然界,四倍体陆地棉种(DAS-21023-5 的转化受体)向其它植物转移遗

65
传物质仅限于通过雌、雄配子结合的有性方式。棉花以自花授粉为主,可与同种
内其它品种授、受粉,距花粉源 1 m 处异交率约为 1%(Van Deynze et al., 2005),
由于生殖隔离和地理隔离,DAS-21023-5 棉花与其它棉种、野生近缘种、锦葵科
其它属间遗传物质的交流概率极低。
地理隔离:DAS-21023-5 栽培棉花与 3 个野生陆地棉种亲缘关系较近(四倍
体),这 3 个野生棉种为黄褐棉(G. mustelinum L.)、达尔文氏棉(G. darwinii L.)
和夏威夷棉(G.tomentosum L.)。分布于巴西东北部半干旱地区(Freire et al., 1998;
Batista et al., 2005)、加拉帕戈斯群岛(Wendel and Percy, 1990;Lacape et al., 2007)
和夏威夷群岛,与中国大陆地缘相隔。因此,棉花 DAS-21023-5 的遗传物质无法
自发地转移到野生棉种中。
生殖隔离:在自然界,棉花(四倍体陆地棉种)向其它植物转移遗传物质仅
限于通过雌、雄配子的有性方式。1)品种间,棉花以自花授粉为主,可与同种
内其它品种授、受粉,花粉源 10 米处异交率极低,约为 1%(Van Deynze et al.,
2005)。2)新疆棉区的海岛棉虽然与陆地棉遗传亲和,之间异交通常需要人工辅
助授粉(如杂交选育新品种),而这种交换在自然条件下发生频率极低。3)棉属
内其它棉种均为二倍体,与陆地棉(DAS-21023-5 的背景)有很大的生殖隔离,
异交当代或减数分裂紊乱,或难以结实,或 F1 发育不良,或 F1 不育。4)DAS-
21023-5 棉花与锦葵科其它属间遗传隔离更远,通过有性的方式传播遗传物质的
可能性比棉属内更低。
3)向动物和微生物水平转移的可能性极低
微生物之间存在遗传物质交流的现象,在自然界转移的频率约为 10-8~10-9
(Paul, 1999)。消化道内的 DNA 片段水平转移到消化道细菌或直接转移至动物
和人类组织的概率同样微小。FAO/WHO 经广泛审查有关吸收、固定哺乳动物消
化系统的外源 DNA 归趋的文献后得出结论:虽然人类通过饮食摄入 RNA 和
DNA 的量为每天 0.1~1.0 克,但没有任何证据表明从植物中提取的完整基因能
够转移到哺乳动物细胞中并在其中表达(FAO/WHO, 2000)。另外,外源基因来
源于自然环境中常见的微生物,即使转多也不会带来环境风险。
转基因棉花中缺乏遗传迁移的辅助元件:外源基因整合进植物基因组后,T-
DNA 边界序列往往会有部分丢失。由于缺少边界序列的协助,外源 DNA 转移到
其它动、植物的可能性极低(Bakkeren et al.,1989)。
转基因棉花的外源基因表达盒在加工过程受损:在自然界尚未见基因从植物
转移至人或动物基因组的报导(Conner et al., 2003)。人畜接触转基因的途径主要
是通过摄入 DAS-21023-5 棉籽的加工产品。基因组 DNA 是食品和饲料的组成成
份,在加工过程中、在人类和动物消化过程中,DNA 充分降解。
外源基因从棉花的转移率低于背景转移率:自然界中基因的水平转移为宿主
或基因本身提供了选择性优势,从而水平基因转移成为了微生物基因组进化的重

66
要机制 (Dobrindt et al., 2004; Gogarten and Townsend, 2005)。然而,从植物到原
核生物的基因水平转移发生率极低,从转基因作物到其他生物的基因水平转移发
生率低于背景转移率(Keese, 2008)。DAS-21023-5 棉花中 cry1Ac 和 pat 基因的供
体微生物苏云金芽孢杆菌和绿色产色链霉菌广泛存在于环境中,即使外源基因能
以极低的概率从 DAS-21023-5 棉花中转移至其它生物,但与环境中的供体相比,
其概率更低(即低于从微生物向其它生物转移的背景转移率)。早期人们研究了
转基因马铃薯品系与细菌病原体 Erwinia chrysanthemi 之间的基因水平转移,发
现在模拟“自然”感染的条件下,检测不出基因水平转移(Schlüter et al., 1995)。之
后,人们又利用不同的转基因植物在不同的环境条件下深入研究了外源基因向细
菌中水平转移的潜力 (Nielsen et al., 1998; Kay et al., 2002; Tepfer et al., 2003;
Demanèche et al., 2008; Kim et al., 2010; Ma et al., 2011),均检测不到水平转移,
并认为序列相似性是基因水平转移的前提条件。
序列一致性是转移的必要条件:由于相邻两次同源重组是基因水平转移发
生的必要步骤,因此外源基因与微生物基因组之间的序列一致性可以评估外源
基因从转基因植物到微生物基因组的转移潜力。EFSA(2009)报道,当相关基
因两侧的序列相似性长度分别从 1000 bp 减少到 300 bp 和 100 bp 时,质粒
DNA 向巴氏不动杆菌(Acinetobacter baylyi)高频转化的效率显著降低,分别低至
10-4 和 10-7 量级。同源重组的频率随着供体和受体之间的序列差异的增加而降
低((Majewski et al., 2000)。将序列一致性降低到 90%,肺炎链球菌的重组效率
降低一个数量级,枯草芽孢杆菌的重组效率降低三个数量级(Kung et al.,
2013)。外源基因来源于微生物,理论上与棉花基因的同源性低,该转化体中的
外源基因转移至其它微物的概率低。考虑到植物和微生物之间的水平转移极为
罕见,并且 DAS-21023-5 棉花的外源序列对人类、动物、环境均无害,认为外
源基因从 DAS-21023-5 棉花水平转移至微生物从而对生态、动物健康、人类健
康带来负面影响的可能性几乎为零。
3.2.7 转变成杂草的可能性;
DAS-21023-5 棉花转变成杂草的可能性极低,且与非转基因棉花转变成杂草
的可能性相同。
杂草无利用价值,且有较强的生存竞争优势。棉花本身不属于杂草,不具有
杂草化倾向和侵蚀自然生态环境的能力。棉花转变成杂草可能的途径可归为两种,
一是提高生存竞争能力,从而在环境中能够自生繁衍,二是与杂草连续授、受粉
且产生稳定的、可育的后代,从而获得杂草的特征。理论依据和实证数据均表明,
DAS-21023-5 棉花转变成杂草的可能性极低。
一、自生繁衍能力是指该植物不需要经过人类播种、病虫害防控、肥水管理
等干预措施就可完成生活周期的能力(强胜, 2000)。种子休眠期的改变、种子萌
发率的提高以及较高结实能力等都可能导致转基因植物生存和繁殖能力提高,使
转基因植物具有竞争优势,从而入侵生态环境,导致杂草化(刘谦等, 2002;崔

67
荣荣 et al., 2012;强胜, 2000)。实验表明 DAS-21023-5 棉花生存竞争能力不比
其非转基因常规对照高,难以演变为有害植物:1)表 14 显示,出苗率、种子休
眠特性、生长习性、营养活力、开花期、生殖潜力及皮棉产量等农艺学性状在转
基因棉花 DAS-21023-5 与其非转基因对照之间实质等同;2)表 11 和 12 显示,
转基因与非转基因对照花粉的数量、大小、活力均无差异。3)表 13 显示,转基
因未提高其发芽率。因此,DAS-21023-5 棉花竞争能力不比非转基因对照高,演
变为杂草的可能性极低。
二、由于生殖隔离,DAS-21023-5 棉花与杂草间连续授、受粉产生可育后代
的可能性极低(OECD,2008),通过这种方式获得杂草特性(更强的适应性和竞
争能力、较高的结实能力、更强的种子传播能力、较强的抗逆境能力)的可能性
极低。
3.2.8 抗病虫转基因植物对靶标生物及非靶标生物的影响,包括对环
境中有益和有害生物的影响;
对靶标生物的影响方面,本公司于 2001 和 2002 年分别调查了 DAS-21023-5
对靶标昆虫的抗性(报告 4A 和报告 4B)。结果显示,该转化事件对鳞翅目多种
昆虫具有有效的控制效果。通过科学的种植、及时的抗性监测可有效进行抗性治
理。
对非靶标生物的影响方面,分析测试了转基因棉花表达的外源蛋白质对非靶
标生物的影响(报告 7A-7K)。结果表明,转基因抗虫棉花及其外源蛋白质对非
靶标生物无不良影响。
1 )对靶标昆虫的影响
DAS-21023-5 棉花可防治特定的鳞翅目害虫幼虫。如:烟草夜蛾(TBW)、
棉红铃虫(PBW)、棉铃虫(CBW)、粉纹夜蛾(CL)、盐泽灯蛾(SC)、棉叶
潜蛾(CLP)、大豆尺蠖(SL)、甜菜夜蛾(BAW)、草地贪夜蛾(FAW)、黄纹
夜蛾(YAW)和欧洲玉米螟(ECB)等。
2001 年对靶标昆虫的抗性试验
实验设在十个地点,分别位于密西西比州(5 处)、阿拉巴马州、北卡罗来纳
州、加利福尼亚州、亚利桑那州和路易斯安那州。靶标昆虫包括烟草夜蛾、棉铃
虫、棉红铃虫、甜菜夜蛾和草地贪夜蛾。在某些试验地点评价了对次要害虫粉纹
夜蛾、大豆尺蠖、南方粘虫和盐泽灯蛾等的抗虫效果。试验田分为“防治区”和
“不防治区”,在“防治区”按常规虫害管理,在“不防治区”只控制非鳞翅目
昆虫(不控制鳞翅目昆虫)。设 4 次重复。采用方差分析比较转基因与非转基因
棉花间的差异,采取最小显著差异(LSD)或 Tukey’s HSD 方法确定差异的显著
性(p=0.05)。结果显示,与非转基因对照 PSC355 相比,在重复人工接种试验中,
DAS-21023-5 棉花对棉铃虫和烟草夜蛾具有高度的抗性;对棉红铃虫有符合商业
要求的抗性;对甜菜夜蛾表现出中度的抗性,对大豆尺蠖和南部粘虫有一定的防

68
效(报告 4A)。
2002年对靶标昆虫的抗性试验
试验地点在亚利桑那州和新墨西哥州。整个生长季节,在“防治区”喷施传
统的杀虫剂即正常虫害管理,在“不防治区”只控制非鳞翅目昆虫不控制鳞翅目
昆虫。设4次重复。实验结果显示,与非转基因对照PSC355相比,人工接种条件
下,DAS-21023-5可以有效抵抗棉红铃虫。在人工接种甜菜夜蛾并在之后3~6天
(种植后75天)调查,发现在DAS-21023-5棉花叶片上的甜菜夜蛾数量显著少于
PSC355上的(报告4B)。
2)抗性风险管理
WideStrike(DAS-21023-5×DAS-24236-5)棉花同时表达 Cry1F 和 Cry1Ac
杀虫蛋白(EPA, 2005),是本申请转化体的最终商业化推广产品。单转化体 DAS-
21023-5 或 DAS-24236-5 棉花没有在任何区域单独商业化推广,所以本申请书中
对 WideStrike 的抗性风险管理阐述适合于本转化体的实际应用情况。包含本转化
体的 WideStrike 棉花对数种重要的取食叶片及棉铃的鳞翅目害虫具有高效的杀
虫效果。为了使这一重要的杀虫管理工具得以持续应用,必须执行抗性风险管理
计划。
交互作用分析:了解不同性状的杀虫机制相似性和差异性有利于预测这些杀
虫蛋白之间的交互作用模式。这反过来又有助于了解对于特定的产品在特定的环
境中的抗性风险。人们对Cry1蛋白的作用模式进行了广泛研究,Cry1蛋白的杀虫
活性是由于该毒素蛋白能在中肠细胞膜上与特异受体蛋白结合并最终在中肠膜
上形成气孔引起的。位于敏感昆虫的中肠顶端微绒毛膜上的受体蛋白决定了Cry1
蛋白的特异作用机制以及对靶标昆虫的特异性(Bravo et al.,2007)。
Bt蛋白靶位点(受体蛋白)分析:WideStrike(DAS-21023-5×DAS-24236-5)
棉花聚合了两种具有相同关键靶标害虫的杀虫蛋白Cry1Ac和Cry1F,增加了Bt作
物应用持久性(Storer et al.,2012)。Cry1Ac和Cry1F杀虫蛋白在同一种靶标害虫
中与不完全相同的受体蛋白结合而起到类似的杀虫效果,从而导致不完全的交互
抗性。在鳞翅目昆虫中,已确定至少有四种结合Cry1的受体蛋白:类钙黏蛋白、
糖基磷脂酰-肌醇(GPI)-锚定氨肽酶N(APN),GPI-锚定碱性磷酸酶(ALP)和
P252蛋白(Bravo et al.,2007)。据报道,Cry1Ac毒素在至少六种鳞翅目物种中
结合钙粘蛋白受体(Bravo et al.,2011)。Cry1毒素在几种鳞翅目昆虫还结合APN
受体。在棉铃虫中,Cry1Ac结合ALP受体。也有报道研究了Cry1Ac和Cry1F蛋白
在烟草夜蛾和棉铃虫(Jurat-Fuentes and Adang, 2001;Sheets and Storer, 2001)的
受体,Cry1F和Cry1Ac蛋白均对烟草夜蛾和棉铃虫有效。Cry1Ac蛋白和Cry1F蛋
白在烟草夜蛾中,很少或根本没有共同的受体结合位点(Jurat Fuentes and Adang,
2006)。在棉铃虫中,Cry1Ac和Cry1F蛋白至少结合4组受体,其中两组受体是通
用的(Storer, 2002;Hernández and Ferré,2005)。但对棉红铃虫只有Cry1Ac有效,
而Cry1F效果不显著。Pereira等人使用抗Cry1F欧洲玉米螟证明了Cry1Ac和Cry1F

69
之间低水平的交互抗性(Pereira et al.,2008)。也有报道使用抗Cry1F草地贪夜蛾
(Vélez et al., 2013)和亚洲玉米螟(Zhang et al., 2014)证明Cry1Ac和Cry1F之间
低水平的交互抗性。
高剂量表达杀虫蛋白及庇护所效应:高剂量表达杀虫蛋白的 Bt 作物对杂合
基因型抗性昆虫具有高致死率。通过三种 EPA 方法的研究,表明 WideStrike 对
烟草夜蛾满足高剂量的定义,即 Bt 作物中蛋白浓度造成至少 99.9%敏感昆虫死
亡或杀死所有敏感昆虫所需剂量的 25 倍(EPA,1998; EPA,2001)。WideStrike
造成了杂合抗性基因型害虫后代的低存活率。另外,采用庇护所策略可以减少纯
和抗性基因型害虫产生的频率。一个由非 Bt 寄主植物组成的庇护所,提供足够
数量的敏感鳞翅目害虫用以与罕见的在 Bt 作物存活的抗性害虫交配。这些交配
产生的杂合抗性基因型害虫后代由于 Bt 作物的高剂量而致死,所以不会产生抗
性等位基因的纯和体后代。对于烟草夜蛾和棉铃虫,庇护所包括非棉作物和野生
寄主植物(“天然庇护所”)。在美国,对于棉红铃虫,不提供庇护所,用以彻底消
除该害虫。
抗性基线设定:同时,WideStrike 产品在商业化种植前对靶标害虫进行了基
线研究。为了了解可能对 Cry1F 和 Cry1Ac 蛋白产生抗性的靶标害虫与转基因作
物的生物相关性,例如变化是否由于 WideStrike 棉花而引起,就必须了解区域内
抗 Cry1F 和 Cry1Ac 种群内及群间变化的天然背景。因此,在 WideStrike 商业化
种植之前使用 15 个烟草夜蛾和棉铃虫的种群评估了烟草夜蛾、棉铃虫和棉红铃
虫种群对 Bt 蛋白的剂量敏感反应关系。该研究确定了商业化种植后年度监测剂
量,并再次证实 Cry1F 和 Cry1Ac 蛋白对目标害虫分别有效。
抗性监测:抗性监测包括收集该 Bt 作物种植领域多个田间试验点的关键靶
标害虫用以蛋白生测,并使用已经建立的基线敏感水平进行对比。任何明显的敏
感性变化将通过收集田间昆虫并进一步饲养此可能的抗性种群,从而进行蛋白质
和/或 Bt 作物生物测定进一步调查验证。2005 年至今,在美国每年都进行广泛的
年度敏感性监测(田间种群及害虫),针对 WideStrike 棉花表达的 Cry1F 和
Cry1Ac 蛋白对靶标害虫烟草夜蛾及棉铃虫的敏感性进行验证调查。迄今为止实
验室的试验显示无任何害虫对 Bt 蛋白的敏感性发生改变。同样,自 2005 年起,
棉红铃虫对 Bt 毒素的敏感性监测未发现 Cry1Ac 抗性棉红铃虫群体。
考虑到本转化体申请仅用于进口加工原料,因此该产品在中国产生抗性风险
可能性极微小。
3)对非靶标生物的影响
鉴于Cry1蛋白的作用机制、作用特异性和其宿主及蛋白的长期安全使用历
史,可以认为DAS-21023-5的靶标昆虫仅限于少数几种鳞翅目类害虫,其他生
物应视为非靶标生物。根据转基因安全评价指南要求,对非靶标生物的选择应
考虑植食性生物、有益生物(如天敌昆虫、资源昆虫和传粉昆虫等)及受保护
的物种等。依据此原则,本公司以微生物来源的Cry1Ac蛋白为测试物对植食性

70
昆虫蚜虫、天敌昆虫草蛉、瓢虫、寄生蜂、土壤腐生昆虫蚯蚓、跳虫、水生动
物大型溞、虹鳟鱼、禽类美洲鹑、哺乳动物小鼠、经济传粉昆虫蜜蜂及保护昆
虫帝王斑蝶等12种非靶标生物进行了一系列毒性评估。
采用高端曝露评估值(High End Exposure Estimates, HEEE)描述直接摄入
DAS-21023-5的暴露水平,用环境浓度估计值(EstimatedEnvironmental
Concentrations, EEC)来描述间接摄入该转基因棉花的暴露水平。HEEE = 90%上
限 = 蛋白平均表达量 + 1.96×标准差。依此可计算出种子中Cry1Ac的HEEE值
为0.69 ng/mg,花粉中Cry1Ac的HEEE值为2.43 ng/mg。以预测的生物学产量和
1999-2001年美国平均棉花产量(每英亩1.35 bales)分区为基础,将HEEE转换
成土壤和水基质中的环境浓度估计值EEC (NASS, 2002)。土壤中Cry1Ac蛋白的
EEC是 0.0196 mg ai/kg。使用GENEEC农场池塘评估了表层水中Cry1Ac蛋白的
EEC值(USEP A, 2000),Cry1Ac蛋白在水中的EEC是107 ng/L(表15)。
表15. 环境中DAS-21023-5表达的Cry1Ac实际暴露水平
直接/间接摄入 组织 暴露水平
直接摄入 DAS-24236-5 时-高端曝露评估值
(HEEE)
种子 6.31 ng/mg
花粉 0.35 ng/mg
间接摄入 DAS-24236-5 时-环境浓度估计值
(EEC)
土壤 0.317 mg/kg
水体 1710 ng/L
a. Cry1Ac 蛋白对蚜虫的影响
Cry1Ac 蛋白(剂量为 14 µg /mL)与液体饲料混合后,覆上一层薄膜对蚜虫
进行生测。蚜虫幼虫可以通过薄膜吸食含有 Cry1Ac 蛋白的液体饲料。3 天后统
计死亡率。生测结果表明,处理组与对照组死亡率无显著差异,LC50 >14 µg/mL,
认为蚜虫不是 Cry1Ac 蛋白的敏感群体(报告 7B)。Cry1Ac 对靶标生物烟草夜蛾
和粉纹夜蛾的 LC50 分别为 0.0012 和 0.0044 µg/ cm2,与蚜虫相比相差 104 量级,
进一步表明 Cry1Ac 对非靶标蚜虫无作用(告 7B)。
b. Cry1Ac 蛋白对草蛉的影响
将 Cry1Ac 蛋白掺入蛾卵,饲喂孵化后 0 天、7 天和 14 天的草蛉幼虫,剂
量为 46.8 µg/g,对照与处理组各有 30 头幼虫。饲喂后两小时开始观察,之后的
每一天都统计死亡率,直至第 16 天,对照组中超过 20%的草蛉死亡。生测结果
表明,处理组与对照组死亡率无显著差异,无害作用量 NOEC >46.8 µg/g。草蛉
直接接触花粉中 Cry1Ac 的 HEEE 值为 2.43 ng/mg(表 15),得出暴露界限
MOE>46.8÷2.43≈19,由于 MOE 大于 10,Cry1Ac 蛋白对草蛉的风险极低(报告
7A 和报告 7C)。
c. Cry1Ac 蛋白对瓢虫的影响
将 Cry1Ac 蛋白与蔗糖溶液适量混合(0.161 mg:1 mL),Cry1Ac 剂量为 20.5
ng/mg。设置对照与处理组各 25 头瓢虫成虫,4 次重复。每周至少饲喂 2 次新鲜

71
的饲料。饲喂后 1-2 小时内开始观察,之后的每一天都统计死亡率及观察临床症
状,直至第 15 天,对照组中超过 20%的瓢虫死亡。生测结果表明,处理组与对
照组死亡率无显著差异,临床症状也无区别,无害作用量 NOEC >20.5 ng/mg。
瓢虫直接接触花粉中 Cry1Ac 的 HEEE 值为 2.43 ng/mg(表 15),得出暴露界限
MOE>20.5÷2.43≈8(报告 7A 和报告 7D)。此外,2002 年陶氏益农公司在美国
多地对表达 Cry1Ac 蛋白的转基因棉花的田间试验统计也未发现对瓢虫有不良的
影响(报告 7L)。文献报道中也没有关于 Cry1Ac 蛋白对瓢虫的作用。因此,Cry1Ac
蛋白对瓢虫的风险极微小。
d. Cry1Ac 蛋白对寄生蜂的影响
将 Cry1Ac 蛋白与蔗糖溶液混合,饲喂寄生蜂成虫 10 天,Cry1Ac 的剂量为
46.8 µg/mL(相当于 42.5 ng/mg),设置对照与处理组各 25 头成虫,3 次重复。
每天提供新鲜的饲料。饲喂后 0.5-1 小时内开始观察,之后的每一天都统计死亡
率及观察临床症状,直至第 10 天,对照组中超过 20%的寄生蜂死亡。生测结果
表明,处理组与对照组死亡率无显著差异,临床症状也无区别。由 Cry1Ac 蛋白
对寄生蜂的无害作用量 NOEC>42.5 ng/mg,寄生蜂直接接触花粉中 Cry1Ac 的由
HEEE 值为 2.43 ng/mg(表 15),得出暴露界限 MOE > 42.5÷2.43≈17,由于 MOE
大于 10 ,可知 Cry1Ac 蛋白对寄生蜂的风险极低(报告 7A 和报告 7E)。上述评
估比寄生蜂捕食鳞翅目幼虫(摄食转基因植物棉花)的安全因性更高,即实际
Cry1Ac 蛋白对寄生蜂的风险更低。
e. Cry1Ac 蛋白对蚯蚓的影响
将成年蚯蚓暴露于含有 Cry1Ac 蛋白(剂量为 107 mg/kg)的土壤中,设置对
照与处理组各 10 头,4 次重复。测试期间不饲喂任何饲料。暴露 4 小时内观察
两次,之后的第 7 天和第 14 天分别统计死亡率及观察临床症状。生测结果表明,
处理组与对照组死亡率无显著差异,体重和行为也无差异。无害作用量
NOEC >107 mg/kg。蚯蚓对土壤中 Cry1Ac 间接接触的 EEC 值为 0.0196 mg/kg
(表 15),得出暴露界限 MOE>107÷0.0196≈5,459,远大于 10,因此 Cry1Ac 蛋
白对土壤无脊椎动物蚯蚓的风险极微小(报告 7A 和报告 7F)。
f. Cry1Ac 蛋白对跳虫的影响
将 Cry1Ac 掺入酵母(含 22.6 mg/kg),饲喂 9-11 天的跳虫幼虫。设置对照
与处理组各 10 头,4 次重复。阴性对照为酵母,阳性对照为灭多威酵母梯度混
合物 1.0 mg/kg, 100 mg/kg 及 1,000 mg/kg。28 天后观察存活率及后代指标发现,
阳性对照组为存活率分别为 88%、33%和 0%,处理组与阴性对照组的存活率无
显著差异,均为 98%。处理组与阴性对照组的后代指标亦无显著差异,
NOEC >22.6 mg/kg。跳虫对土壤中 Cry1Ac 间接接触的 EEC 值为 0.0196 mg/kg
(表 15),得出暴露界限 MOE>22.6÷0.0196≈1153,由于 MOE 远大于 10,故
Cry1Ac 蛋白对土壤腐生昆虫跳虫的风险极微小(报告 7A 和报告 7G)。

72
另外,进行了冻干棉花叶片暴露试验。结果证明,与暴露于试验对照的生物
相比,暴露于包含冻干 Cry1Ac 棉花叶片和冻干 PSC3355 对照棉花叶片的饲料
(5.0% 和 50% 浓度)对于跳虫幼虫的繁殖没有不良影响。存活率也没有影响。
以上所述混合试验和转基因棉花冷冻叶片试验证明在单独 Cry1Ac 蛋白测试中观
察到的后代昆虫数量减少是由于蛋白或配方中的杂质导致,与 Cry1Ac 蛋白无关
(报告 7A 和报告 7G)。
g. Cry1Ac 蛋白对大型溞的影响
将大型溞暴露于含有 Cry1Ac 蛋白(剂量为 2,500 µg/L)的水中,设置对照
(水)与处理组各 10 头,三次重复。暴露 48 小时,测试期间不饲喂任何饲料。
于 24 小时及 48 小时各观察一次,未发现处理组与对照组中有不良效应。无害作
用量 NOEC ≥2,500 µg/L,大型溞对水体中 Cry1Ac 间接接触的 EEC 值为 107 ng/L
(表 15),得出暴露界限 MOE>2,500÷0.107≈23,364,远大于 10,因此 Cry1Ac
蛋白对水生生物大型溞的风险极微小(报告 7A 和报告 7H)。
h. Cry1Ac 蛋白对虹鳟鱼的影响
用含有 Cry1Ac 蛋白(剂量为 11.24 mg /kg)的标准鱼食饲喂虹鳟鱼,每天饲
喂一次,共 8 天。设置对照(标准饲料)与处理组各 10 尾,3 次重复。试验结束
后,对照组虹鳟鱼平均体长 35±2 mm,体重 574±116 mg,处理组虹鳟鱼平均体
长 35±2 mm,体重 580±111 mg,无明显差异。另外,每 24 小时观察一次,处理
组与对照组均未发现有死亡或其他不良效应。无害作用量 NOEC ≥11.24 mg/kg,
虹鳟鱼对水体中 Cry1Ac 间接接触的 EEC 值为 107 ng/kg(表 15),得出暴露界限
MOE>11.24÷0.000107≈105,046,该值远大于 10,因此 Cry1Ac 蛋白对水生生物
虹鳟鱼的风险极微小(报告 7A 和报告 7i)。
i. Cry1Ac 蛋白对美洲鹑的影响
给 22 周的美洲鹑强饲法饲喂 Cry1Ac 蛋白,剂量为 14.4 mg/kg。设置对照与
处理组各 10 只(5 只雄性,5 只雌性)。每日观察至少一次,在 0 天、第 3 天、
第 7 天和第 14 天分别测量体重,试验结束后进行大体解剖。14 天后,处理组与
对照组均未发现有死亡或与处理有关的不良效应,体重也无明显的区别,大体解
剖结果也未显示有与处理相关的病变。无害作用量 NOEC >14.4 mg/kg。美洲鹑直
接接触棉籽中 Cry1Ac 的 HEEE 值为 0.69 ng/mg(表 15),得出暴露界限
MOE>14.4÷0.69≈21,由于 MOE 大于 10,Cry1Ac 蛋白对美洲鹑的风险极低(报
告 7A 和报告 7J)。
j. Cry1Ac 蛋白对小鼠的影响
以约7周龄的Crl:CD1(ICR)小鼠为试验动物,灌胃Cry1Ac蛋白质剂量为5050
mg/(kgBW),设置BSA组、空白对照组及处理组,每组16只,雌、雄各半。每日
观察,试验结束后进行大体解剖。14天后,未见毒性反应,未见与处理相关的动
物死亡;在试验期间体重均增长,灌胃Cry1Ac的小鼠平均体重及变化均与灌胃

73
BSA及空白对照组相当,即未见Cry1Ac蛋白质引起小鼠体重异常;整个试验过程
所有动物均表现正常,未观察到与灌胃蛋白质Cry1Ac相关的临床症状;试验结束
时所有小鼠的尸检解剖试验均未发现相关的组织病理学损伤,即Cry1Ac蛋白质
对小鼠生长发育无不良影响。Cry1Ac蛋白质对试验动物的无害作用量NOEC>
5050 mg/(kgBW),小鼠直接接触棉籽中Cry1Ac的HEEE值为0.69 ng/mg(表15),
由此得出暴露界限MOE>5,050÷0.69≈7,319,该值远大于10,因此Cry1Ac蛋白对
哺乳动物小鼠的风险极微小(报告7A和报告8E)。
k. Cry1Ac 蛋白对蜜蜂的影响
给 3-5 天蜜蜂老熟幼虫饲喂 Cry1Ac蛋白,剂量为 11.94 µg/mL(10.85 ng/mg)
与蔗糖溶液适量混合,饲喂后 11 天到 27 天,蜜蜂幼虫羽化成成虫。设置阳性对
照(砷化钾溶液),阴性对照(蔗糖溶液)与处理组各 20 头幼虫,四次重复。统
计第 6 天和第 11 天后幼虫成活率,及第 11 天-27 天羽化成虫的孵化率。生测结
果表明,11 天和 27 天后,阳性对照存活率为 45%,处理组与阴性对照组存活率
均大于 90%,且未观察到其他不利的效应,无害作用量 NOEC >10.85 µg/mg。蜜
蜂直接接触花粉中 Cry1Ac 的 HEEE 值为 2.43 ng/mg(表 15),由此得出暴露界
限 MOE>10.85÷2.43≈4.5, 大于 1。此外,鉴于 Cry1Ac 蛋白仅对少数几种鳞翅目
昆虫幼虫有效的高度作用特异性以及文献报道中也没有关于 Cry 蛋白对蜜蜂的
不良作用(Duan et. al.,2008),因此认为 Cry1Ac 蛋白对蜜蜂的风险极低(报告
7A 和报告 7K)。
l. Cry1Ac 蛋白对帝王斑蝶的影响
2001 年 Hellmich 等人将一龄的帝王斑蝶幼虫暴露于含有 Cry1Ac 蛋白的花
粉中,观察 7 天,设置对照组及处理组。当对照组中 50%的试验昆虫生长影响减
弱时,Cry1Ac 蛋白剂量即 EC50 为 0.9 ng/mL,该 EC50 高于花粉暴露水平的 10
倍(Hellmich et. al.,2001)。因此转基因棉花的 Cry1Ac 蛋白对帝王斑蝶幼虫无
风险(报告 7A)。
m. 对其它非靶标生物的影响
此外,美国渔业和野生动物管理局(U.S. Fish and Wildlife Service)罗列了美
国棉产区相关昆虫种类以及濒临灭绝的鳞翅目昆虫在全美的分布情况(FWS,
1997),见图26,这些濒临灭绝的鳞翅目物种有非常严格的寄主特异性。再结合
Cry1蛋白质的抗虫谱,可知除了已知Kern月见草天蛾(Euproserpinus euterpe)存
在于加利福尼亚州Kern郡以外,其它所有的濒临灭绝鳞翅目物种均不存在棉产区,
而棉花不是Kern月见草天蛾的寄主(FWS, 1984)。
在2004和2009年,美国环保署对转cry1基因棉花注册和重新注册进行了全面
的风险评估,认为该抗虫棉花表达的Cry1蛋白质对指示性物种无毒,其花粉从田
间流失的量及影响极微,与测试物种所喂食的花粉量相比,从棉花植株上漂移到
濒危物种和有益昆虫的摄食花粉上的量极少。濒危和有益的昆虫物种受到Cry1花
74
粉不良影响的可能性极低(USEPA, 2004;USEPA 2009)。
图26. 美国的棉花产区以及相关的濒临灭绝鳞翅目物种的存在区域(FWS,1997)
综上所述,通过对以上一系列具有代表性的非靶标生物进行的生物测定表明,
DAS-21023-5中表达的Cry1Ac蛋白对非靶标生物的影响可以忽略不计。
3.2.9 对生态环境的其他有益或有害作用。
转基因棉花表达的 Cry1Ac 蛋白质在土壤中的半衰期为 1.3 天,不会蓄积;
DAS-21023-5 棉花对田间节肢动物、环境微生物及植物的群落及多样性无负面影
响,在未见对生态环境的其它有益或有害作用。
1)Cry1Ac 蛋白质的环境归趋
在实验室测得植物表达的 Cry1Ac 蛋白在土壤中半衰期为 1.3 天(报告 7A)。
土样为市售土壤,取自密西西比(代表性的棉花种植地区)。取 2 g 烘干土壤,
调整到 1/3 bar 持水力的 75%。土壤中加入 50 mg 冷冻干燥叶片(叶片中 Cry1Ac
含量为 34.1 mg/g),再添加 0.853 mg/kg 的 Cry1Ac,相当于 3.2 倍 EEC 值,即
将转基因棉花整株(落叶后)混合到 15 cm 深土表土中,使其中 Cry1Ac 达到 3.2
倍环境估计浓度(Estimated Environmental Concentrations, EEC)。以非转基因
棉花组织添加到土壤作为阴性对照。将混好的土样在室温保存,定期取样检测目
标蛋白质在土壤中的降解情况,直至第 28 天。采用昆虫生测的方法检测土壤中
杀虫蛋白的活性:将土壤样品和水样琼脂混合,涂到人工饲料的顶部,饲喂烟草
夜蛾新生幼虫,以半数生长抑制剂量 GI50 为评估指标。

75
结果显示 GI50 值随时间延长而升高。最终测得土壤中棉花表达的 Cry1Ac 杀
虫蛋白的半衰期为 1.3 天。因此,棉花表达的 Cry1Ac 不会在土壤中蓄积。
2)转基因棉花对非靶标节肢生物群落及生物多样性的影响
2002年,以PSC355为非转基因对照,在Louisiana和Arizona州调查比较了转基
因棉花对非靶标生物多样性的影响(报告7L)。结果显示,转基因棉花对非靶标
有益节肢动物无明显负面影响,不会降低生物多样性。
实验设计
Louisiana试验点位于典型的烟草夜蛾幼虫、螟蛉虫和棉铃虫防治区。试验排
布按完全随机区组设计,每个处理设3次重复(区组)。每小区约240 m2(15 m×16
行,行距100 cm),各小区间由4行芥菜和藜草分开,用于吸引昆虫的替代寄主或
降低杀虫剂漂散,即降低边界效应。
Arizona试验点的典型害虫为棉红铃虫和麦蛾。Arizona实验点也按完全随机
区组设计,每个处理设2次重复。每个小区至少585 m2(24.4 m ×24 行,行距100
cm)。为了降低杀虫剂漂散,在各个处理小区之间留4行空白缓冲带。
两个实验点均设置以下3种处理:转基因株系不喷施鳞翅目类杀虫剂;非转
基因对照不喷施鳞翅目类杀虫剂;非转基因对照根据当地推荐的农事操作控制所
有害虫,包括烟草夜蛾幼虫和棉铃虫。
捕虫分析
在Louisiana实验点,使用“扫网法”(sweep net)捕虫、“抖动纸片法”(shake
sheet)法捕虫、叶片取样调查、棉蕾取样调查、白花取样调查、棉铃取样调查等
方法取样。在小区中心的8~10行取样,以降低边界效应。“扫网法”在棉株冠部
扫10次为一组,每小区2~3组,合并成一份小区样本;“摇动纸片法”收集各小
区4个位置的节肢动物,合并成一份小区样本。捕虫时期集中在烟草夜蛾幼虫为
害的敏感生长期。
在Arizona实验点,使用“扫网法”和“粘卡陷井法”每周在各小区捕虫,并
对有益节肢动物和棉花害虫的种类和数目进行计数。“扫网法”每小区捕100网,
合并成一个小区样本;“粘卡陷阱法”每个小区24小时内捕6个样本。所有的节肢
动物捕获自每小区中心的16行,以此降低邻近小区昆虫迁移的影响。
调查结果
如果处理间差异显著(α=0.05),则计算最小显著差异来判断处理间的差异
显著性。在 Louisiana 试验点,转基因棉花(不控制鳞翅目害虫)的掠食性半翅
目昆虫如大眼长蝽属、小花蝽属、姬蝽科和螠蝽属、猎蝽科和瓢虫成虫数量显著
高于不控制鳞翅目害虫的非转基因棉花(表 16A 和表 16B)。
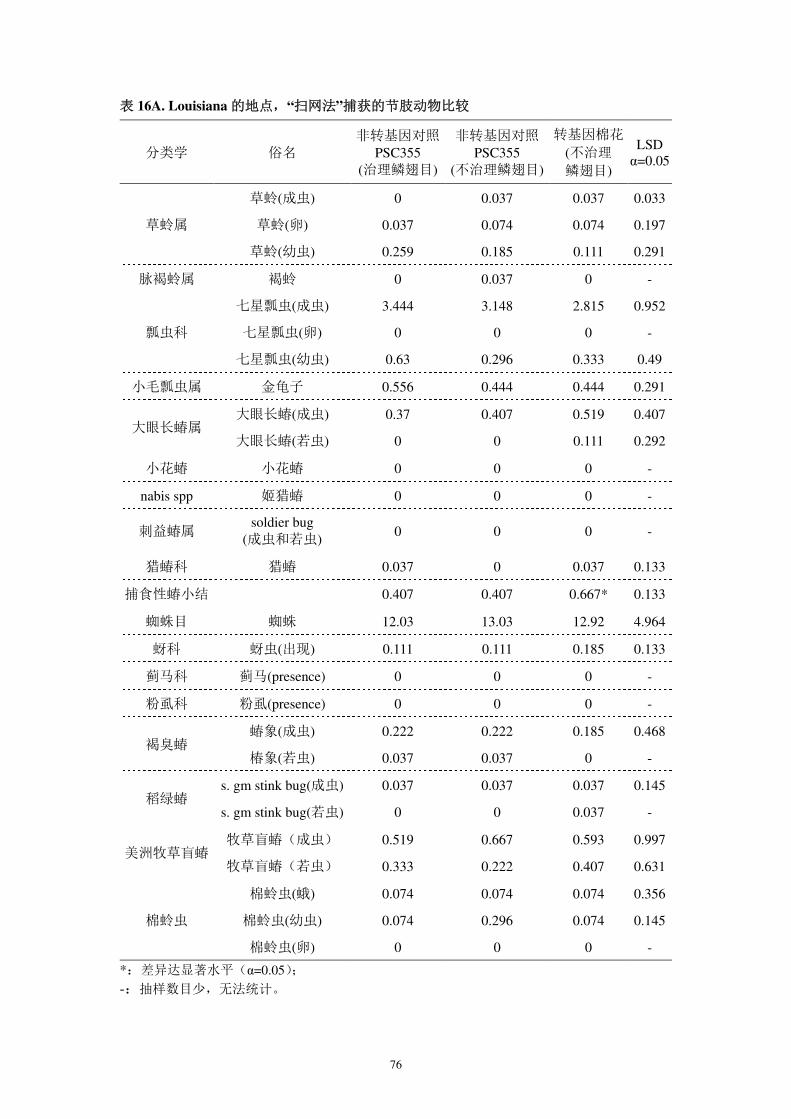
76
表 16A. Louisiana 的地点,“扫网法”捕获的节肢动物比较
分类学 俗名 非转基因对照
PSC355 (治理鳞翅目)
非转基因对照 PSC355
(不治理鳞翅目)
转基因棉花 (不治理 鳞翅目)
LSD α=0.05
草蛉属
草蛉(成虫) 0 0.037 0.037 0.033
草蛉(卵) 0.037 0.074 0.074 0.197
草蛉(幼虫) 0.259 0.185 0.111 0.291
脉褐蛉属 褐蛉 0 0.037 0 -
瓢虫科
七星瓢虫(成虫) 3.444 3.148 2.815 0.952
七星瓢虫(卵) 0 0 0 -
七星瓢虫(幼虫) 0.63 0.296 0.333 0.49
小毛瓢虫属 金龟子 0.556 0.444 0.444 0.291
大眼长蝽属 大眼长蝽(成虫) 0.37 0.407 0.519 0.407
大眼长蝽(若虫) 0 0 0.111 0.292
小花蝽 小花蝽 0 0 0 -
nabis spp 姬猎蝽 0 0 0 -
刺益蝽属 soldier bug
(成虫和若虫) 0 0 0 -
猎蝽科 猎蝽 0.037 0 0.037 0.133
捕食性蝽小结 0.407 0.407 0.667* 0.133
蜘蛛目 蜘蛛 12.03 13.03 12.92 4.964
蚜科 蚜虫(出现) 0.111 0.111 0.185 0.133
蓟马科 蓟马(presence) 0 0 0 -
粉虱科 粉虱(presence) 0 0 0 -
褐臭蝽 蝽象(成虫) 0.222 0.222 0.185 0.468
椿象(若虫) 0.037 0.037 0 -
稻绿蝽 s. gm stink bug(成虫) 0.037 0.037 0.037 0.145
s. gm stink bug(若虫) 0 0 0.037 -
美洲牧草盲蝽 牧草盲蝽(成虫) 0.519 0.667 0.593 0.997
牧草盲蝽(若虫) 0.333 0.222 0.407 0.631
棉蛉虫
棉蛉虫(蛾) 0.074 0.074 0.074 0.356
棉蛉虫(幼虫) 0.074 0.296 0.074 0.145
棉蛉虫(卵) 0 0 0 -
*:差异达显著水平(α=0.05);
-:抽样数目少,无法统计。
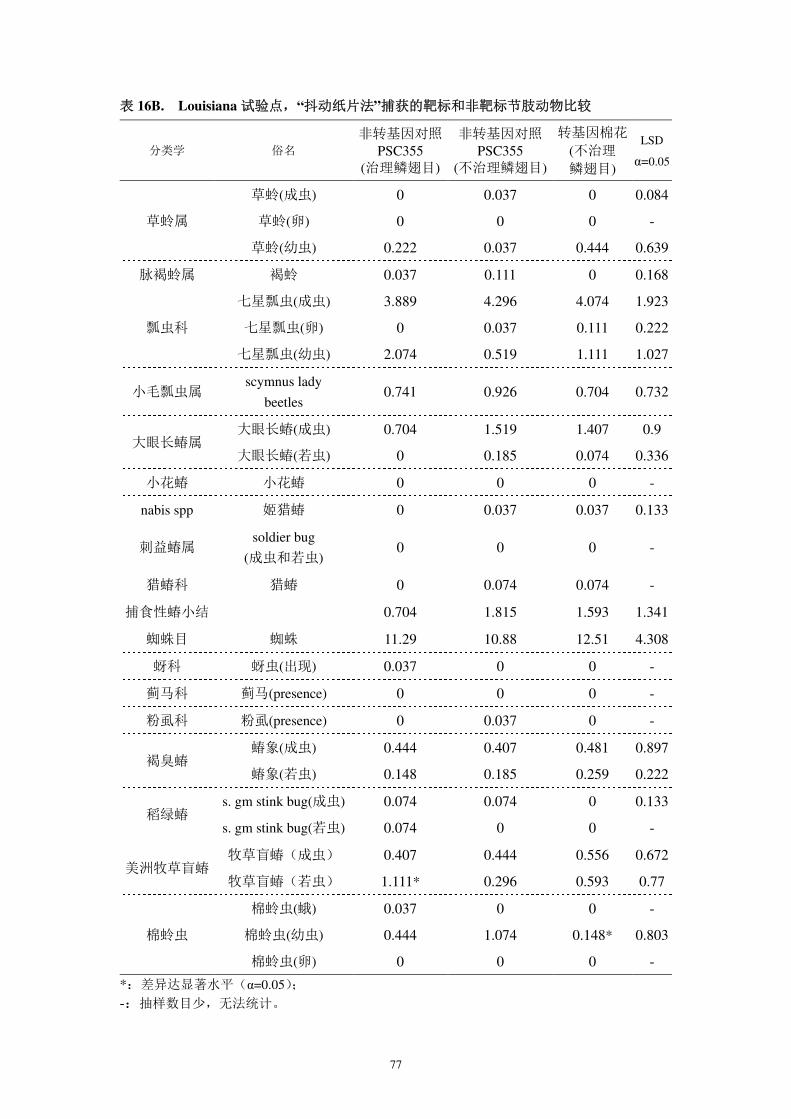
77
表 16B. Louisiana 试验点,“抖动纸片法”捕获的靶标和非靶标节肢动物比较
分类学 俗名 非转基因对照
PSC355 (治理鳞翅目)
非转基因对照 PSC355
(不治理鳞翅目)
转基因棉花 (不治理 鳞翅目)
LSD
α=0.05
草蛉属
草蛉(成虫) 0 0.037 0 0.084
草蛉(卵) 0 0 0 -
草蛉(幼虫) 0.222 0.037 0.444 0.639
脉褐蛉属 褐蛉 0.037 0.111 0 0.168
瓢虫科
七星瓢虫(成虫) 3.889 4.296 4.074 1.923
七星瓢虫(卵) 0 0.037 0.111 0.222
七星瓢虫(幼虫) 2.074 0.519 1.111 1.027
小毛瓢虫属 scymnus lady
beetles 0.741 0.926 0.704 0.732
大眼长蝽属 大眼长蝽(成虫) 0.704 1.519 1.407 0.9
大眼长蝽(若虫) 0 0.185 0.074 0.336
小花蝽 小花蝽 0 0 0 -
nabis spp 姬猎蝽 0 0.037 0.037 0.133
刺益蝽属 soldier bug
(成虫和若虫) 0 0 0 -
猎蝽科 猎蝽 0 0.074 0.074 -
捕食性蝽小结 0.704 1.815 1.593 1.341
蜘蛛目 蜘蛛 11.29 10.88 12.51 4.308
蚜科 蚜虫(出现) 0.037 0 0 -
蓟马科 蓟马(presence) 0 0 0 -
粉虱科 粉虱(presence) 0 0.037 0 -
褐臭蝽 蝽象(成虫) 0.444 0.407 0.481 0.897
蝽象(若虫) 0.148 0.185 0.259 0.222
稻绿蝽 s. gm stink bug(成虫) 0.074 0.074 0 0.133
s. gm stink bug(若虫) 0.074 0 0 -
美洲牧草盲蝽 牧草盲蝽(成虫) 0.407 0.444 0.556 0.672
牧草盲蝽(若虫) 1.111* 0.296 0.593 0.77
棉蛉虫
棉蛉虫(蛾) 0.037 0 0 -
棉蛉虫(幼虫) 0.444 1.074 0.148* 0.803
棉蛉虫(卵) 0 0 0 -
*:差异达显著水平(α=0.05); -:抽样数目少,无法统计。

78
在 Arizona 试验点,使用“扫网法”捕虫,整个生长季节在 16 个取样时间共
布 1600 网,收集了 194 种虫类。使用“扫网法”检测的样品中,转基因棉花对
大约 200 种形态类型的非靶标节肢动物无明显影响。除了 4 种昆虫(棉花盲椿
象、浅条纹跳甲虫、舞虻和绿色草蛉幼虫)在处理间存在显著差异外,其它的虫
类在处理间差异均不显著。绿色草蛉幼虫的数量在转基因棉花(不控制鳞翅目害
虫)小区明显高于对照非转基因棉花(不控制鳞翅目害虫)小区,且这两个处理
的绿色草蛉幼虫数量均比非转基因棉花(控制鳞翅目害虫)的高。
在Arizona试验点使用“粘卡陷井法”捕虫,整个季节每小区设78个陷阱,在
16个捕虫时间点共检测到143个类型的节肢动物。其中28种在所有的小区中均发
生。转基因棉花小区对143种生物类型的非靶标节肢动物无重要影响。
在两个试验地点的整个生长季节,使用多种方法收集转基因棉花及其非转基
因对照小区的非靶标节肢动物。与不喷洒鳞翅目杀虫剂的非转基因对照相比,转
基因棉花对非靶标节肢动物的类型和密度无不良影响;而喷洒鳞翅目杀虫剂的非
转基因对照小区可见明显的负面效应。
3)对微生物群落多样性的影响
DAS-21023-5 棉花不影响土壤微生物的多样性。
本公司以含有转化体 DAS-21023-5 的棉花及其回交受体 PSC355 为试验材
料,于 2003-2004 年在日本进行了土壤微生物比较试验。未见 DAS-21023-5 对土
壤微生物产生负面影响。本试验设 3 次重复,每小区 10 株。在 2003 年播种期
(5 月 6 日)、开花期(7 月 14 日)和收获后(10 月 21 日)分别取 0~15 cm 深
的地表土,保存在 5℃。每小区取 3 点。将土壤烘干称重,稀释后铺于培养皿培
养并计数菌落数,试验方法见土壤微生物研究汇编(1992)新编土壤微生物实验
法养贤堂(东京)。其中,采用肉汁培养基和稀释肉汁培养基培养细菌,采用稀
释肉汁培养基培养放射菌,采用孟加拉红培养基培养丝状细菌。计算每小区 3 次
取样的平均值,采用 t-检验比较转基因棉花和非转基因对照 PSC355 间小区土壤
微生物菌落数。
从表 17 可见,在干燥土壤中,播种期的转基因棉花和其非转基因对照
PSC355 小区间,用稀释肉汁培养基培养的细菌菌落数在二者间差异显著,但数
值仍在自然变异范围内,相应的用肉汁培养基培养的细菌菌落数无显著差异;在
开花期和收获后所测 3 种微生物在 DAS-21023-5 和其非转基因对照 PSC355 间
均未见显著差异。即二者田间相应微生物数量实质等同。
前人研究结果显示 Bt 棉对于微生物群落多样性并无不良影响。土壤微生物
种群变化是衡量 Bt 作物对土壤生态影响的重要指标。张美俊等人在盆栽种植条
件下比较 Bt 棉与对照棉对根际土壤细菌、放线菌、真菌和主要功能类群及多样
性的影响差异,与对照相比,Bt 棉对细菌和真菌生长繁殖有促进作用,对放线
菌、好气固氮菌和钾细菌数量没有显著影响(张美俊等, 2008)。Kapur 等研究
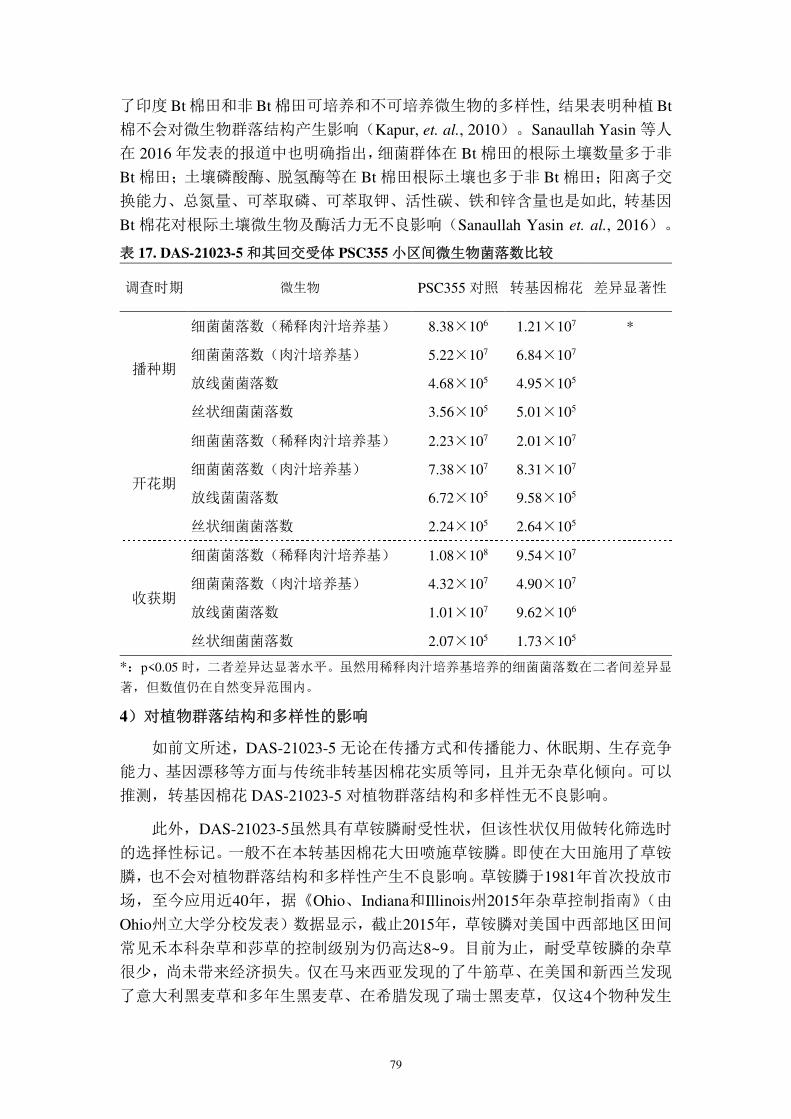
79
了印度 Bt 棉田和非 Bt 棉田可培养和不可培养微生物的多样性, 结果表明种植 Bt
棉不会对微生物群落结构产生影响(Kapur, et. al., 2010)。Sanaullah Yasin 等人
在 2016 年发表的报道中也明确指出,细菌群体在 Bt 棉田的根际土壤数量多于非
Bt 棉田;土壤磷酸酶、脱氢酶等在 Bt 棉田根际土壤也多于非 Bt 棉田;阳离子交
换能力、总氮量、可萃取磷、可萃取钾、活性碳、铁和锌含量也是如此, 转基因
Bt 棉花对根际土壤微生物及酶活力无不良影响(Sanaullah Yasin et. al., 2016)。
表 17. DAS-21023-5 和其回交受体 PSC355 小区间微生物菌落数比较
调查时期 微生物 PSC355 对照 转基因棉花 差异显著性
播种期
细菌菌落数(稀释肉汁培养基) 8.38×106 1.21×107 *
细菌菌落数(肉汁培养基) 5.22×107 6.84×107
放线菌菌落数 4.68×105 4.95×105
丝状细菌菌落数 3.56×105 5.01×105
开花期
细菌菌落数(稀释肉汁培养基) 2.23×107 2.01×107
细菌菌落数(肉汁培养基) 7.38×107 8.31×107
放线菌菌落数 6.72×105 9.58×105
丝状细菌菌落数 2.24×105 2.64×105
收获期
细菌菌落数(稀释肉汁培养基) 1.08×108 9.54×107
细菌菌落数(肉汁培养基) 4.32×107 4.90×107
放线菌菌落数 1.01×107 9.62×106
丝状细菌菌落数 2.07×105 1.73×105
*:p<0.05 时,二者差异达显著水平。虽然用稀释肉汁培养基培养的细菌菌落数在二者间差异显
著,但数值仍在自然变异范围内。
4)对植物群落结构和多样性的影响
如前文所述,DAS-21023-5 无论在传播方式和传播能力、休眠期、生存竞争
能力、基因漂移等方面与传统非转基因棉花实质等同,且并无杂草化倾向。可以
推测,转基因棉花 DAS-21023-5 对植物群落结构和多样性无不良影响。
此外,DAS-21023-5虽然具有草铵膦耐受性状,但该性状仅用做转化筛选时
的选择性标记。一般不在本转基因棉花大田喷施草铵膦。即使在大田施用了草铵
膦,也不会对植物群落结构和多样性产生不良影响。草铵膦于1981年首次投放市
场,至今应用近40年,据《Ohio、Indiana和Illinois州2015年杂草控制指南》(由
Ohio州立大学分校发表)数据显示,截止2015年,草铵膦对美国中西部地区田间
常见禾本科杂草和莎草的控制级别为仍高达8~9。目前为止,耐受草铵膦的杂草
很少,尚未带来经济损失。仅在马来西亚发现的了牛筋草、在美国和新西兰发现
了意大利黑麦草和多年生黑麦草、在希腊发现了瑞士黑麦草,仅这4个物种发生

80
了草铵膦耐受生物型(Heap, I. 2018. www.weedscience.org)。在生产中,加以综
合预防、配合农事操作,不同作用模式的除草剂合理施用,可有效减缓杂草对草
铵膦耐受性演化,并控制风险。
为了避免或减缓耐受性杂草的出现,可以采取如下的抗性监测方案和治理措
施:(1)采用多种作用机制的除草剂,可有效防止产生耐受除草剂的杂草;(2)
采用多样化的杂草管理措施,重点防止生产杂草种子,减少土壤中杂草种子的数
量;(3)在无杂草的地里种植,并保持该田块持续无杂草;(4)播种时,保证种
子中不夹带杂草种子;(5)例行田间检测和调查;(6)了解现有杂草的生物学特
性;(7)施用除草剂时,采用标签所建议的使用量;(8)采用耕作措施,竞争性
抑制杂草的发生;(9)适当时使用机械的和生物的管理方法;(10)预防田块之
间(或田块内)的杂草通过种子扩散,或通过营养繁殖扩散;(11)在收获及收
获后合理管理,以防土壤中草籽积累;(12)合理管理田块边界,以防止杂草进
入。
自 2005 年在美国和 2009 年在巴西商业化种植含有 DAS-21023-5 品系的复
合性状棉花以来,未发现对植物带来负面影响。此长期安全使用历史再次证明转
基因棉花 DAS-21023-5 对植物群落结构和多样性无不良影响。
3.3 转基因植物与受体或亲本植物在对人类健康影响方面的差异:
本公司对 DAS-21023-5 棉花转化体及其表达的外源蛋白质的安全性进行了
综合评价,结果显示本转基因棉花与其非转基因对照对人类健康影响实质等同,
即本转化体及其表达的外源蛋白质对人类健康影响方面无不良影响。
3.3.1 毒性
从以下几个方面对转基因棉花 DAS-21023-5 及其表达的外源蛋白质有无潜
在毒性进行了评价,结果显示:
1) 微生物及转基因棉花表达的外源蛋白质实质等同,微生物表达的外源蛋白
质可以用于安全性评价。
2) cry1Ac 及 pat 基因的供体生物及 Cry1Ac 和 PAT 蛋白质具有安全使用和食用
的历史;
3) Cry1Ac 和 PAT 蛋白质的理化特征清晰;
4) Cry1Ac 和 PAT 蛋白质不属于已知毒性蛋白质和抗营养因子;
5) Cry1Ac 和 PAT 蛋白质受热后免疫活性和功能活性不稳定;
6) Cry1Ac 和 PAT 蛋白质在模拟胃液 SGF 中易消化;
7) Cry1Ac 和 PAT 蛋白质对哺乳动物无经口急性毒性,LD50 均大于 5050 mg/
(kg·BW);
8) Cry1Ac 和 PAT 蛋白质在人群膳食中接触风险极低,接触界限 MoE 分别达
108 和 109 量级。

81
8) 大鼠 90 天喂养试验显示,未见转基因棉花引起哺乳动物亚慢性毒性反应,
未见其引起营养学不良反应,且未见非预期效应;
9) 42 天肉鸡喂养试验显示,未见转基因棉花引起营养学不良反应,且未见非
预期效应;
因此,转基因棉花 DAS-21023-5 及其表达的外源蛋白质 Cry1Ac 和 PAT 对
人、畜无潜在的毒性,详述于下。
1) 微生物及转基因棉花表达的外源蛋白质实质等同,
小鼠经口急性毒性试验、模拟消化试验、热稳定性试验需要大量的 Cry1Ac
和 PAT 蛋白质,但依现有技术,难以从植物转化体 DAS-21023-5 中纯化足够量
的外源蛋白质,本试验从荧光假单胞菌中纯化蛋白质代替植物的相应蛋白质进行
上述试验。评价之前,需要验证两种来源的蛋白质的理化特性、一级结构及生物
活性一致,即实质等同。
DAS-21023-5 棉花和微生物表达的 Cry1Ac 的等同性:1)Cry1Ac 原毒素对
胰蛋白酶敏感,在制备过程中,Cry1Ac 蛋白质很容易被酶裂为截短的、各种中
间形式的片段,其中 65 kDa 的核心蛋白质是稳定的片段。许多文献报道均指出
这类65 kDa的核心蛋白质是Cry1原毒素的完整功能性片断。 SDS-PAGE 显示,
荧光假单胞菌表达的 Cry1Ac 蛋白质有 65 kDa 大小的片段(报告 8F),棉花表达
的 Cry1Ac 亦有 65 kDa 大小的片段(报告 8G);2)Western blot 显示两种来源的
蛋白质均与 Cry1Ac 特异性多克隆抗体发生免疫亲和反应,即二者免疫原性相同
(报告 8F 和 8G);3)两种来源的 Cry1Ac 蛋白质用胰蛋白酶酶解后再以基质辅
助激光解吸/电离飞行时间质谱(MALDI-TOF MS)分析,并使用 MS/MS 测序,
证明了两种来源的 Cry1Ac 的胰蛋白酶肽指纹图谱相同(报告 8F 和 8G);4)采
用 Edman 降解序法测定了荧光假单胞菌来源的 Cry1Ac 蛋白质的 N-末端序列(报
告 8F),相同的方法测定了 DAS-21023-5 棉花来源的 Cry1Ac 蛋白质的 N 末端氨
基酸序列(报告 8G),结果显示两种来源的 Cry1Ac 蛋白质 N-末端测序匹配;5)
对两种来源的 Cry1Ac 进行翻译后修饰分析,采用 GelCode 糖基化染色对微生物
来源的 Cry1Ac 蛋白质染色(报告 8F),对转基因棉花来源的 Cry1Ac 蛋白质染
色(报告 8G),二者均未见与之共价偶联的碳水化合物,即均不被糖基化;6)
以不同活性浓度的 Cry1Ac 饵料处理棉铃虫幼虫、烟草夜蛾幼虫和甜菜夜蛾幼虫,
调查昆虫死亡率和体重,采用 SAS 统计并比较两种来源蛋白质的 LC50(杀死一
半昆虫时的蛋白质浓度)及 GI50(50%昆虫生长受抑制时的蛋白质浓度)。结果
显示,对上述 3 种靶标昆虫的功能效率,两种来源的 Cry1Ac 无显著差异,即二
者生物活性等同(报告 8H)。因此,DAS-21023-5 棉花和微生物表达的 Cry1Ac
实质等同,后者可以代替前者用于生物安全试验分析。
DAS-21023-5 棉花和微生物表达的 PAT 的等同性:微生物来源的 PAT 蛋白
质和 DAS-21023-5 棉花表达的 PAT 蛋白质等同(报告 9F)。 试验显示两种来源
的 PAT 蛋白质具有相同的分子量、相同的免疫活性和生化活性(具体见报告 9F)。

82
同时,也和文献报道的大量的试验数据结论一致,均验证了微生物来源的 PAT 蛋
白质和多种转基因植物来源的 PAT 蛋白质具有等同的生化功能特性(Corinne
Hérouet, 2005)。微生物系统表达的 PAT 蛋白质可代替 DAS-21023-5 棉花表达的
PAT 蛋白质用于各种安全评估试验。
2)cry1Ac 及 pat 基因的供体生物及其表达的 Cry1Ac 和 PAT 具有安全的使用历
史
苏云金芽孢杆菌(cry1Ac 的供体生物)的安全性:苏云金芽孢杆菌是土壤中
天然存在的革兰氏阳性菌。形成孢子时在细胞内产生不同的 δ-内毒素晶体对鳞翅
目、双翅目和鞘翅目害虫分别具有特异性的杀虫活性,因此广泛用于害虫防治。
自 1938 年法国将苏云金芽孢杆菌商业化以来(Kumar et al., 1996),至今以该菌
作为活性成分的注册农药产品已经超过 100 种(EPA, 2011),并广泛应用于粮食、
草料、水果和蔬菜等多种作物。1960 年美国食品药品监督管理局(Food and Drug
Administration)对苏云金芽孢杆菌进行安全评估后,准予豁免其残留要求,并于
1961 年首次在美国环保署(Environmental Protection Agency)注册为农药。至今,
含有苏云金芽孢杆菌活性成分的农药已有数十年的商业化安全应用历史,至今未
有此类微生物对人类、动物或植物有害的报道(Mendelsohn et al., 2003; EPA, 2011;
Sanahuja et al., 2011)。
Cry1Ac 蛋白质的安全应用历史:1986 年美国环境保护署(EPA)首次批准了
种植转 Bt 基因烟草,截止 2017 年 4 月,国际生命科学研究所(International Life
Science Institute,ILSI)网站公布的全球商业化转 Bt 基因作物(单价和双价)达
上百种,涉及棉花、大豆、玉米、番茄和马铃薯等作物。表达 Cry1F 和 Cry1Ac
蛋白质的抗虫棉花复合品系(WideStrike)已在美国和巴西获得审批,商业规模
化种植;并在多个国家批准用于食品和饲料,如澳大利亚、巴西、加拿大、欧盟、
日本、韩国、墨西哥、新西兰和美国(www.biotradestatus.com)。另外,表达 Cry1F
和 Cry1Ac 蛋白质的棉花、玉米和大豆已经在美国和其他国家商业使用十多年
(www.biotradestatus.com)。在其全球商业化应用期间,未见对人类、动物、微生
物和环境造成不良影响,充分表明了 Cry1 蛋白质的安全性。
绿色产色链霉菌(pat 的供体生物)的安全性:pat 基因的供体生物是土壤中
绿色产色链霉菌(Streptomyces viridochromogenes),该微生物广泛存在于自然界
中,不是已知的人、畜病原微生物(Corinne Herouet, 2004)。链霉菌是全球生物
圈中最普遍的组成部分,至今未见该菌对人类,动物或植物有害的报导。转 pat
基因的作物已在全球商业化近二十年,是全球范围内推广的两大除草剂之一,涉
及的作物有油菜、玉米、棉花、大豆和甜菜,约 34 个产品(http://cera-
gmc.org/GMCropDatabase),自应用以来从未对食品和饲料带来不良影响。含 pat
基因的作物获得多个国家的审批,如美国、加拿大、澳大利亚/新西兰、阿根廷、
巴西、南非等(www.biotradestatus.com;EFSA, 2006, 2007a, 2007b, 2008)。尚未
发现该微生物对人类或动物不安全的报道。

83
PAT 蛋白质的安全应用历史: 转 pat 基因的作物已在全球商业化近二十年,
涉及的作物有油菜、玉米、棉花、大豆和甜菜,约 34 个产品(http://cera-
gmc.org/GMCropDatabase),对食品和饲料从未带来不良影响。含 pat 基因的作物
获得美国、加拿大、澳大利亚/新西兰、阿根廷、巴西、南非等多个国家的审批
(www.biotradestatus.com;EFSA, 2006, 2007a, 2007b, 2008)。
3)Cry1Ac 和 PAT 蛋白质的理化特征清晰
Cry1Ac 杀虫蛋白质作用机理: Cry1Ac 被敏感昆虫摄取进入中肠后,被中
肠的蛋白酶酶解,释放具有杀虫活性的核心毒素(65 kDa)。激活后的毒素通过
其结构域 II、III 特定氨基酸与中肠上皮细胞膜的特异性受体结合,其 N-末端随
之形成四聚体并插入细胞膜。毒素四聚体形成跨膜的离子通道或非特异性小孔,
从而导致昆虫中肠上皮细胞裂解和坏死(Bravo et al., 2004;Vadlamudi et al., 1993;
Gahan et al., 2001; Morin et al., 2003; Coates et al., 2005; Yang et al., 2011)。通常,
该类蛋白质仅对少数几种鳞翅目幼虫发挥毒性。
Cry1Ac 的分子和生化特征等信息,包括分子量、氨基酸序列、翻译后的修
饰、功能叙述等资料见转化体与微生物来源的Cry1Ac等同性研究报告(报告8F、
报告 8G 和报告 8F)。
PAT 酶的理化特性和作用机理:草铵膦是植物体中谷氨酸盐合成酶(GS)
的潜在抑制剂,草铵膦对 GS 的抑制作用会引起细胞内氨的快速积累,从而引起
光呼吸终断,最终导致植物细胞死亡。PAT 催化乙酰 CoA 的乙酰基转移至草铵
膦生成乙酰化草铵膦和 CoASH(图 2),从而使草铵膦除草剂失去活性,使植物
体免受伤害。PAT 的表观分子量约为 20 kDa,翻译后不被糖基化(报告 9F),其
氨基酸序列见 2.2.3。
PAT 酶活性影响因素:Josef Vinnemeier 等人通过在 22℃对 PAT 催化乙酰-
CoA 的乙酰基转移至 PPT 生成乙酰化草铵膦和 CoASH,CoASH 可与 DTNB (2-
nitro-5-thiobenzoate)反应生成 TNB [5,5-dithiobis-(2-nitrobenzoic acid)],用酶标仪
检测 TNB 含量,从而确定 PAT 的活性。结果显示,PAT 催化反应最适 pH 值为
8.0。pH 5.5 时失去酶活性,pH 6.5 和 pH 9.5 时,催化效率分别为最大催化效率
的 10%和 30%(Josef Vinnemeier, et al. 1995)。本公司也分别测试了在 pH 3.0,
pH 8.0 和 pH 11.0 条件下 PAT 的催化活性,实验显示,与 Vinnemeier 等人的结论
一致,PAT 活性在 pH8.0 时显示了最大催化效率。当 pH 11.0 时,催化活性急剧
下降,pH 3.0 时,酶活性则完全丧失。
同时 PAT 酶活性受温度影响,本公司于 2013 年以草铵膦为反应底物,以
25℃、37℃、55℃、75℃和 95℃将 PAT 蛋白酶加热 30 分钟,发现 PAT 蛋白酶受
热后溶解度降低,免疫亲合活性和酶活性均随温度升高而急剧降低。当温度超过
55℃时,PAT 丧失至少 99% 酶活性,75℃时检测不到酶活性;ELISA 检测表明,
37℃或更高温度时 PAT 蛋白酶失去≥91%的免疫亲合活性(报告 9C)。
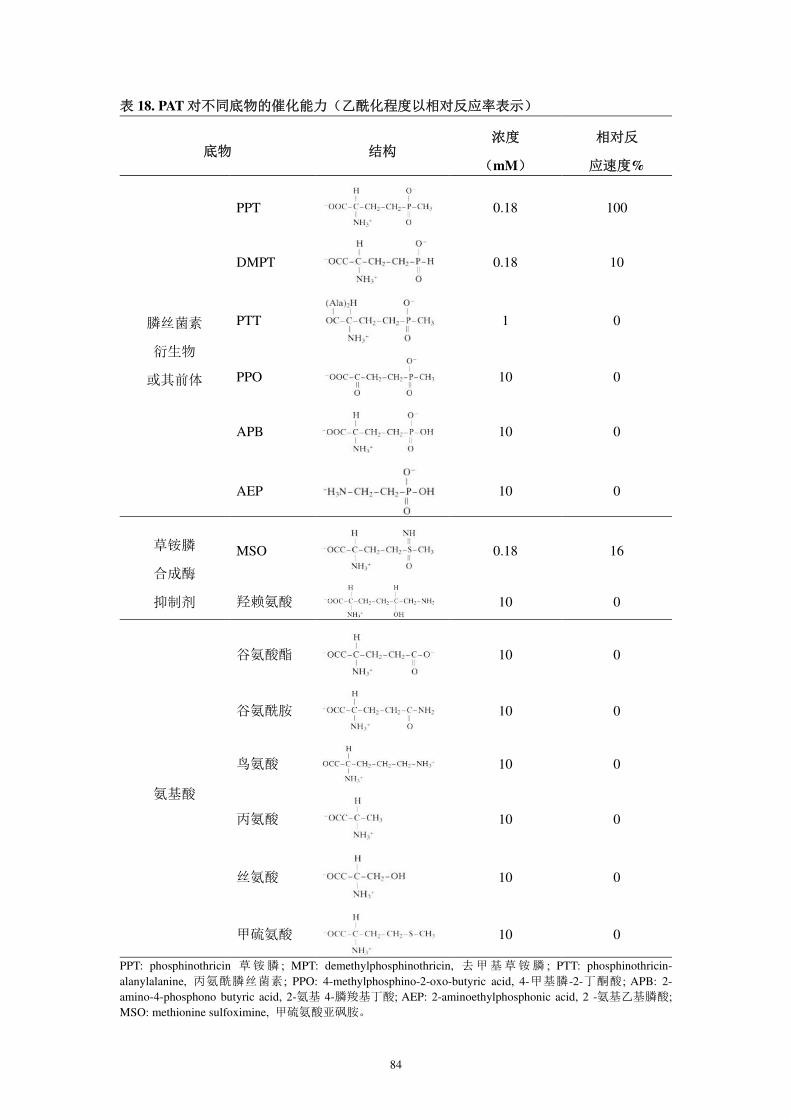
84
表 18. PAT 对不同底物的催化能力(乙酰化程度以相对反应率表示)
底物 结构 浓度
(mM)
相对反
应速度%
膦丝菌素
衍生物
或其前体
PPT
0.18 100
DMPT
0.18 10
PTT
1 0
PPO
10 0
APB
10 0
AEP
10 0
草铵膦
合成酶
抑制剂
MSO
0.18 16
羟赖氨酸
10 0
氨基酸
谷氨酸酯
10 0
谷氨酰胺
10 0
鸟氨酸
10 0
丙氨酸
10 0
丝氨酸
10 0
甲硫氨酸
10 0
PPT: phosphinothricin 草铵膦 ; MPT: demethylphosphinothricin, 去甲基草铵膦 ; PTT: phosphinothricin-alanylalanine, 丙氨酰膦丝菌素; PPO: 4-methylphosphino-2-oxo-butyric acid, 4-甲基膦-2-丁酮酸; APB: 2-amino-4-phosphono butyric acid, 2-氨基 4-膦羧基丁酸; AEP: 2-aminoethylphosphonic acid, 2 -氨基乙基膦酸; MSO: methionine sulfoximine, 甲硫氨酸亚砜胺。

85
PAT 底物特异性:根据 Vinnemeier 等人的文献报道,以草铵膦衍生物或者前
体,如 DMPT(demethylphosphinothricin,去甲基草铵膦)、PTT(phosphinothricin-
alanylalanine, 丙氨酰膦丝菌素)、PPO(4-methylphosphino-2-oxo-butyric acid,4-
甲基膦-2-丁酮酸)、APB(2-amino-4-phosphono butyric acid,2-氨基 4-膦羧基丁
酸)和 AEP(2-aminoethylphosphonic acid,2 -氨基乙基膦酸);两个谷氨酰胺合
成酶抑制剂,如 MSO(methionine sulfoximine,甲硫氨酸亚砜胺)和 δ-羟赖氨酸;
及氨基酸为底物测试 PAT 对底物的特异性,见下表。结果显示,除草铵膦外,仅
DMPT 和 MSO 可以被 PAT 乙酰化,其乙酰化程度分别是草铵膦的 10% 和 16%
(表 18)。该试验显示了 PAT 对底物的特异性,保证了其在植物体内不会作用于
其它物质。国际生命科学研究所环境风险评估中心经审查指出,上述实验结果与
公共领域其它诸多数据均表明,在转基因植物中表达的 PAT 蛋白不会对环境带
来实质性影响(CERA - ILSI Research Foundation 2011)。
反应产物:PAT 可催化乙酰-CoA 的乙酰基转移至草铵膦生成乙酰化草铵膦
和 CoASH,乙酰化草铵膦无杀草活性,使植物体免受伤害。
4)Cry1Ac 和 PAT 与已知毒蛋白质无较高的氨基酸序列相似性。
利用生物信息学分析软件,按国家标准(农业农村 2630 号公告-16-2017)于
2020 年将 Cry1Ac 及 PAT 的氨基酸序列分别与公共蛋白质序列数据库 NCBI-nr
(http://www.ncbi.nlm.nih.gov/)和 UniProtKB (https://www.uniprot.org/blast/)中蛋
白质的序列进行比较。经比对,发现 PAT 和 Cry1Ac 均与已知毒性蛋白质、已知
抗营养因子的氨基酸序列无较高的相似性(报告 8A 和报告 9A)。
比对方法:采用2020年公布的美国卫生研究院国家生物信中心(National
Center for Biotechnology Information,NCBI)非重复蛋白质序列公共数据库
(NCBI-nr)进行分析。该数据库中的序列信息包含了UniProt、PIR、ExPASy
及其它数据库中的蛋白序列。使用BLASTp (protein-protein BLAST;
https://blast.ncbi.nlm.nih.gov/Blast.cgi)程序按默认参数进行搜索,将E值设为
0.01,输出匹配数定为20000。在BLASTp输出序列中分别搜索如下8个关键词
“toxin”、“toxic”、“agglutinin”、 “anti-nutrient”、“antinutrient”、“anti-
nutritional”、“protease inhibitor”和“trypsin inhibitor”。
除了 NCBI,本公司还将 Cry1Ac 和 PAT 与 2020 年公布的公共数据库 UniProt
(https://www.uniprot.org/blast/)中的序列进行了搜索比对。用 BLASTp (protein-
protein BLAST; https://www.uniprot.org/blast/ )程序按默认参数进行搜索,E 值设
置为 0.01,输出匹配数为 500。在 BLASTp 输出序列中分别搜索上述 8 个关键
词。
Cry1Ac 蛋白质比对结果(报告 8A):1)采用 BLASTp 工具将 Cry1Ac 蛋白
质与 NCBI-nr 中的序列进行比对,返回了 2387 个匹配。由于 Cry1Ac 本身属于
杀虫晶体蛋白质,来源于细菌 Bacillus thuringiensis,大部分返回的序列为来自于
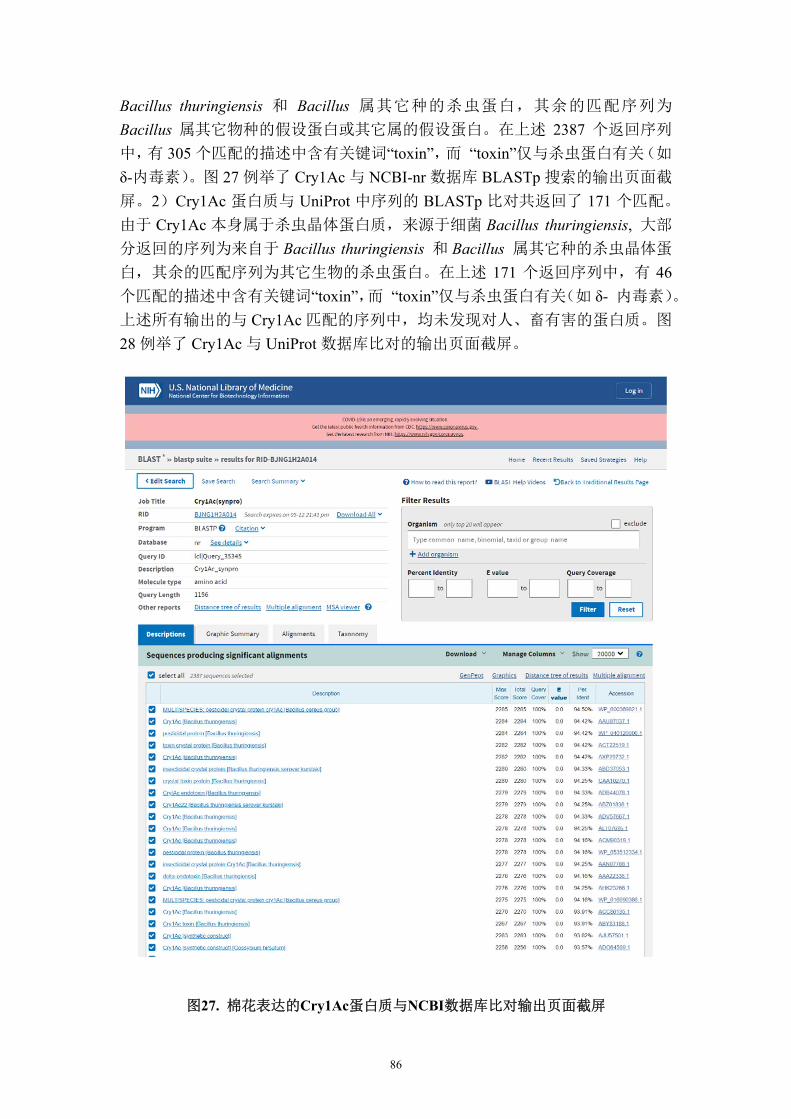
86
Bacillus thuringiensis 和 Bacillus 属其它种的杀虫蛋白,其余的匹配序列为
Bacillus 属其它物种的假设蛋白或其它属的假设蛋白。在上述 2387 个返回序列
中,有 305 个匹配的描述中含有关键词“toxin”,而 “toxin”仅与杀虫蛋白有关(如
δ-内毒素)。图 27 例举了 Cry1Ac 与 NCBI-nr 数据库 BLASTp 搜索的输出页面截
屏。2)Cry1Ac 蛋白质与 UniProt 中序列的 BLASTp 比对共返回了 171 个匹配。
由于 Cry1Ac 本身属于杀虫晶体蛋白质,来源于细菌 Bacillus thuringiensis, 大部
分返回的序列为来自于 Bacillus thuringiensis 和 Bacillus 属其它种的杀虫晶体蛋
白,其余的匹配序列为其它生物的杀虫蛋白。在上述 171 个返回序列中,有 46
个匹配的描述中含有关键词“toxin”,而 “toxin”仅与杀虫蛋白有关(如 δ- 内毒素)。
上述所有输出的与 Cry1Ac 匹配的序列中,均未发现对人、畜有害的蛋白质。图
28 例举了 Cry1Ac 与 UniProt 数据库比对的输出页面截屏。
图27. 棉花表达的Cry1Ac蛋白质与NCBI数据库比对输出页面截屏
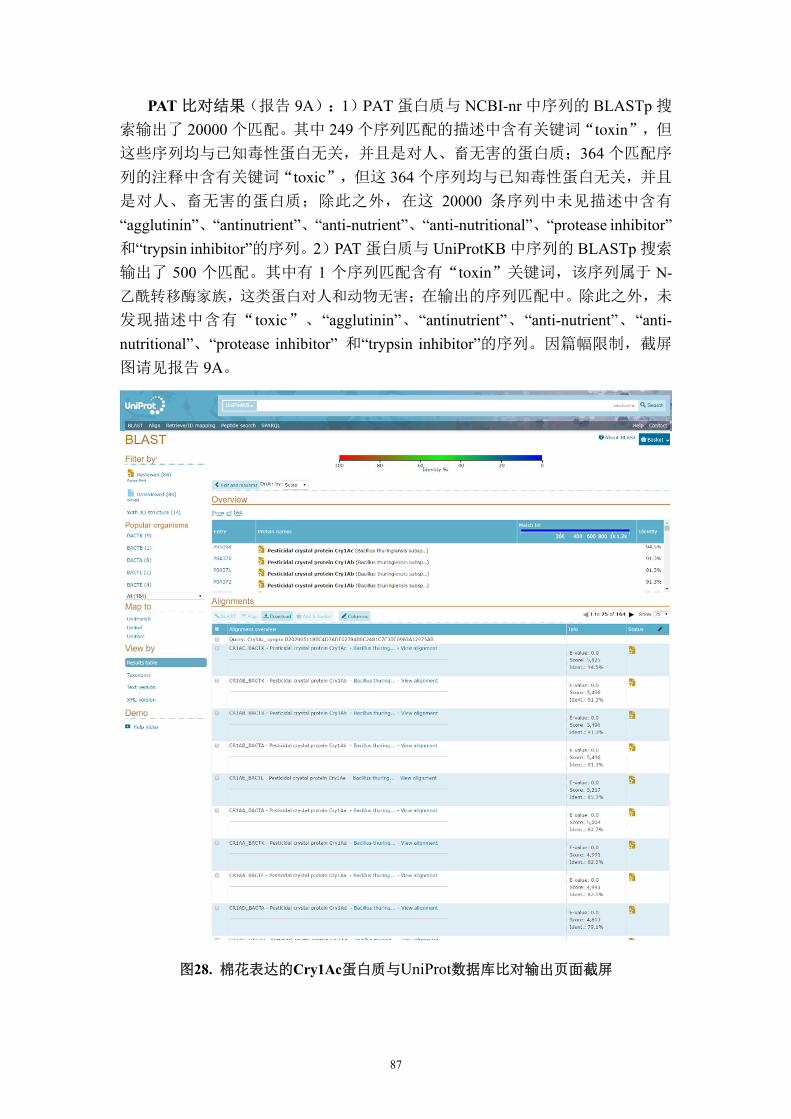
87
PAT 比对结果(报告 9A):1)PAT 蛋白质与 NCBI-nr 中序列的 BLASTp 搜
索输出了 20000 个匹配。其中 249 个序列匹配的描述中含有关键词“toxin”,但
这些序列均与已知毒性蛋白无关,并且是对人、畜无害的蛋白质;364 个匹配序
列的注释中含有关键词“toxic”,但这 364 个序列均与已知毒性蛋白无关,并且
是对人、畜无害的蛋白质;除此之外,在这 20000 条序列中未见描述中含有
“agglutinin”、“antinutrient”、“anti-nutrient”、“anti-nutritional”、“protease inhibitor”
和“trypsin inhibitor”的序列。2)PAT 蛋白质与 UniProtKB 中序列的 BLASTp 搜索
输出了 500 个匹配。其中有 1 个序列匹配含有“toxin”关键词,该序列属于 N-
乙酰转移酶家族,这类蛋白对人和动物无害;在输出的序列匹配中。除此之外,未
发现描述中含有“toxic”、“agglutinin”、“antinutrient”、“anti-nutrient”、“anti-
nutritional”、“protease inhibitor” 和“trypsin inhibitor”的序列。因篇幅限制,截屏
图请见报告 9A。
图28. 棉花表达的Cry1Ac蛋白质与UniProt数据库比对输出页面截屏
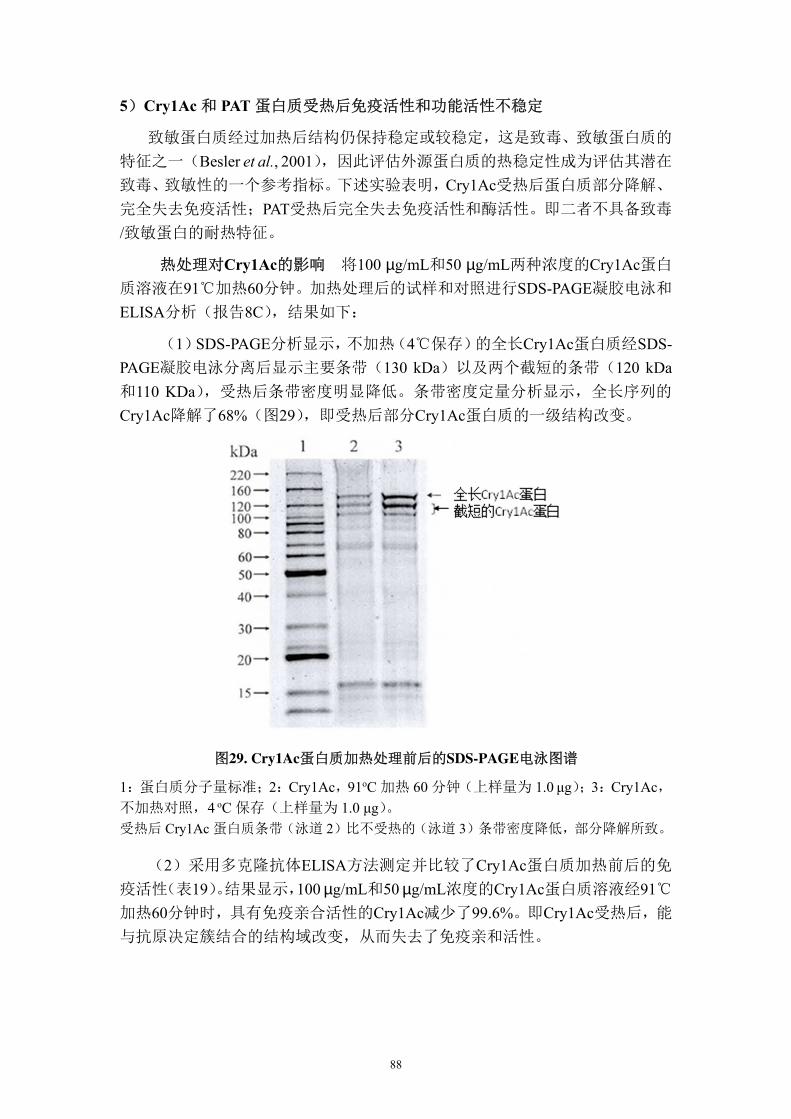
88
5)Cry1Ac 和 PAT 蛋白质受热后免疫活性和功能活性不稳定
致敏蛋白质经过加热后结构仍保持稳定或较稳定,这是致毒、致敏蛋白质的
特征之一(Besler et al., 2001),因此评估外源蛋白质的热稳定性成为评估其潜在
致毒、致敏性的一个参考指标。下述实验表明,Cry1Ac受热后蛋白质部分降解、
完全失去免疫活性;PAT受热后完全失去免疫活性和酶活性。即二者不具备致毒
/致敏蛋白的耐热特征。
热处理对Cry1Ac的影响 将100 µg/mL和50 µg/mL两种浓度的Cry1Ac蛋白
质溶液在91℃加热60分钟。加热处理后的试样和对照进行SDS-PAGE凝胶电泳和
ELISA分析(报告8C),结果如下:
(1)SDS-PAGE分析显示,不加热(4℃保存)的全长Cry1Ac蛋白质经SDS-
PAGE凝胶电泳分离后显示主要条带(130 kDa)以及两个截短的条带(120 kDa
和110 KDa),受热后条带密度明显降低。条带密度定量分析显示,全长序列的
Cry1Ac降解了68%(图29),即受热后部分Cry1Ac蛋白质的一级结构改变。
图29. Cry1Ac蛋白质加热处理前后的SDS-PAGE电泳图谱
1:蛋白质分子量标准;2:Cry1Ac,91oC 加热 60 分钟(上样量为 1.0 μg);3:Cry1Ac,
不加热对照,4 oC 保存(上样量为 1.0 μg)。 受热后 Cry1Ac 蛋白质条带(泳道 2)比不受热的(泳道 3)条带密度降低,部分降解所致。
(2)采用多克隆抗体ELISA方法测定并比较了Cry1Ac蛋白质加热前后的免
疫活性(表19)。结果显示,100 µg/mL和50 µg/mL浓度的Cry1Ac蛋白质溶液经91℃
加热60分钟时,具有免疫亲合活性的Cry1Ac减少了99.6%。即Cry1Ac受热后,能
与抗原决定簇结合的结构域改变,从而失去了免疫亲和活性。

89
表 19. ELISA 测定加热前后具有免疫活性的 Cry1Ac 蛋白质浓度(91oC 加热 60 分钟时)
项目 具有免疫活性的 Cry1Ac 蛋白质浓度
测定浓度(ng/mL) 减少百分比(%)
Cry1Ac 蛋白质加热前的浓度 100 µg/mL 383 99.6
Cry1Ac 蛋白质加热前的浓度 50 µg/mL 182 99.6
浓度减少百分比的均值(%) - 99.6
标准差 - 0
变异系数(CV%) - 0
注:91oC 加热 60 分钟时。
热处理对 PAT 蛋白质的影响 将 PAT 蛋白质溶液在 25℃、37℃、55℃、75℃
和 95℃加热 30 分钟,然后将加热过的以及未加热的(对照)PAT 样品分别采用
SDS-PAGE、多克隆抗体 ELISA 和酶活性测定方法检测加热产物,分析加热对其
稳定性的影响(报告 9C)。结果如下:
(1)如图 30 所示,未经加热的全长 PAT 蛋白质条带在考马斯亮蓝染色的
SDS-PAGE 凝胶上约为 20 kDa 大小,在 25℃、37℃和 55℃加热 30 分钟时,仍
可见 PAT 蛋白质条带,当温度高于 55℃时,在凝胶中检测不到 PAT 蛋白质(因
PAT 变性不溶解所致)。
图 30. 热处理前后 PAT 蛋白质的 SDS-PAGE 电泳图谱
1:分子量标准;2:PAT 不加热(置于冰上);3:25℃加热 30 分钟;4:37℃加热 30分钟;5:55℃加热 30 分钟;6:75℃加热 30 分钟;7:95℃加热 30 分钟;8:分子量
标准。
(2)ELISA 试验表明,PAT 蛋白质受热高于 37℃时丧失了≥91%的免疫亲合
活性,而 55℃或更高温度时几乎检测不到其免疫亲合活性(表 20)。表 20 中极
低的免疫亲合活性(0.8~0.9 %)很可能来源于不依赖于高级结构的多克隆抗体
结合位点。由此可见,PAT 蛋白质受热后,高级结构改变,在 ELISA 中与多克隆
抗体结合的绝大部分抗原表位遭到破坏。
PAT

90
(3) PAT催化乙酰-CoA的乙酰基转移至草铵膦生成乙酰化草铵膦和CoASH。
产物 CoASH 会与 Ellman's 试剂或 DTNB 反应生成 TNB,该产物于 412 nm 波长
处有最大吸光值。基于此原理检测 PAT 的酶活性。结果显示,随着温度升高,酶
活性显著减弱。从表20可见,37℃时酶活性丧失近半,55℃时酶活性丧失>99%,
高于 75℃时检测不到酶活性。因此认为 PAT 蛋白质受热后功能不稳定。
表 20. PAT 经热处理前后免疫亲和活性和酶活性的测定结果 处理温度 免疫亲和活性(%) 酶活性(%)
静置冰上(不加热) 100 100 25℃ 72 100 37℃ 9 53 55℃ 0.9 0.1 75℃ 0.8 0.0 95℃ 0.8 0.0
PAT 蛋白质受热后容易失去免疫亲和活性和功能活性,与前人研究结果一致(OECD, 1999;Hérouet et al., 2005)。
6)Cry1Ac 和 PAT 蛋白质在模拟胃液 SGF 中易消化
通常认为在消化道中不被消化和受热稳定是致敏或致毒物质的特征之一
(Codex, 2003b),在模拟胃液中,Cry1Ac 和 PAT 蛋白质均在 1 分钟内被降解,
依农业农村部 896 号公告-2-1007,表述为:Cry1Ac 和 PAT 在模拟胃液中易消化。
因此不具备一般致毒或致敏蛋白质不易消化的特性。
本试验 SGF 中含胃蛋白酶 0.32%(w/v)(Sigma Aldrich, St. Louis, Missouri),
pH 值为 1.2;设 BSA 为不稳定对照,β-乳球蛋白为稳定对照;消化条件为 37℃
水浴;在不同的时间点终止反应;SGF 消化产物采用 SDS-PAGE 及 Western blot
观察。
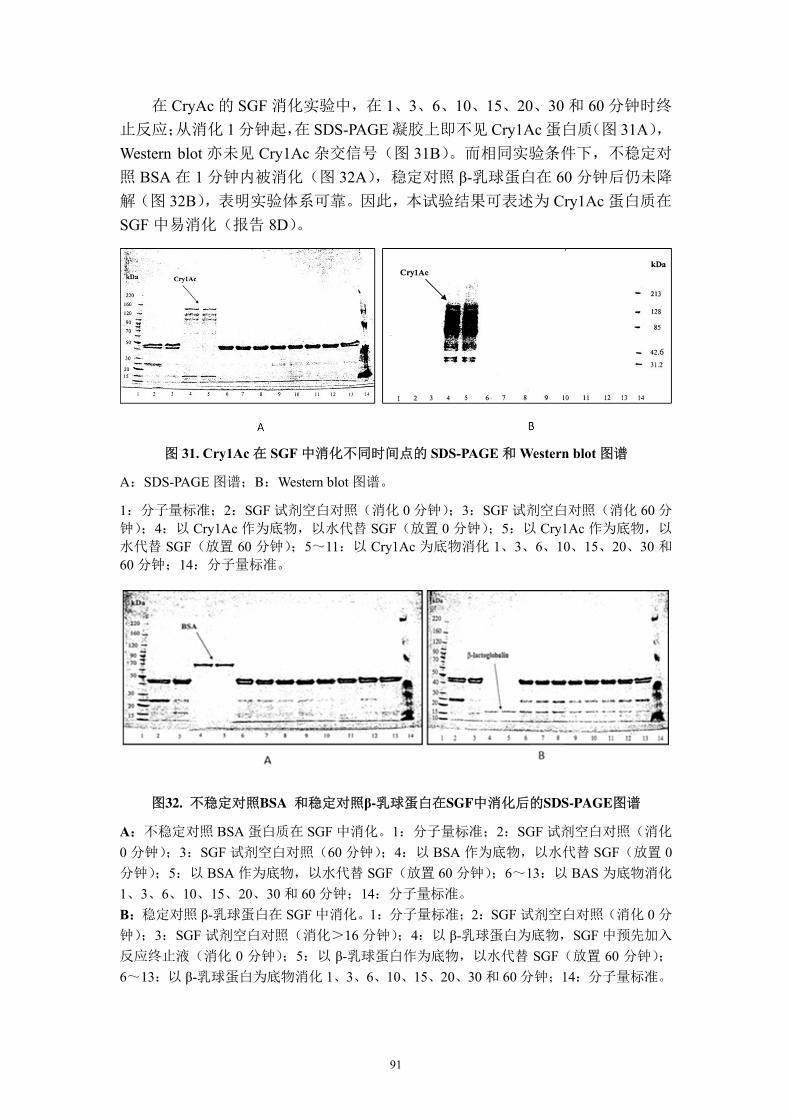
91
在 CryAc 的 SGF 消化实验中,在 1、3、6、10、15、20、30 和 60 分钟时终
止反应;从消化 1 分钟起,在 SDS-PAGE 凝胶上即不见 Cry1Ac 蛋白质(图 31A),
Western blot 亦未见 Cry1Ac 杂交信号(图 31B)。而相同实验条件下,不稳定对
照 BSA 在 1 分钟内被消化(图 32A),稳定对照 β-乳球蛋白在 60 分钟后仍未降
解(图 32B),表明实验体系可靠。因此,本试验结果可表述为 Cry1Ac 蛋白质在
SGF 中易消化(报告 8D)。
图 31. Cry1Ac 在 SGF 中消化不同时间点的 SDS-PAGE 和 Western blot 图谱
A:SDS-PAGE 图谱;B:Western blot 图谱。
1:分子量标准;2:SGF 试剂空白对照(消化 0 分钟);3:SGF 试剂空白对照(消化 60 分
钟);4:以 Cry1Ac 作为底物,以水代替 SGF(放置 0 分钟);5:以 Cry1Ac 作为底物,以
水代替 SGF(放置 60 分钟);5~11:以 Cry1Ac 为底物消化 1、3、6、10、15、20、30 和
60 分钟;14:分子量标准。
图32. 不稳定对照BSA 和稳定对照β-乳球蛋白在SGF中消化后的SDS-PAGE图谱
A:不稳定对照 BSA 蛋白质在 SGF 中消化。1:分子量标准;2:SGF 试剂空白对照(消化
0 分钟);3:SGF 试剂空白对照(60 分钟);4:以 BSA 作为底物,以水代替 SGF(放置 0
分钟);5:以 BSA 作为底物,以水代替 SGF(放置 60 分钟);6~13:以 BAS 为底物消化
1、3、6、10、15、20、30 和 60 分钟;14:分子量标准。
B:稳定对照 β-乳球蛋白在 SGF 中消化。1:分子量标准;2:SGF 试剂空白对照(消化 0 分
钟);3:SGF 试剂空白对照(消化>16 分钟);4:以 β-乳球蛋白为底物,SGF 中预先加入
反应终止液(消化 0 分钟);5:以 β-乳球蛋白作为底物,以水代替 SGF(放置 60 分钟);
6~13:以 β-乳球蛋白为底物消化 1、3、6、10、15、20、30 和 60 分钟;14:分子量标准。
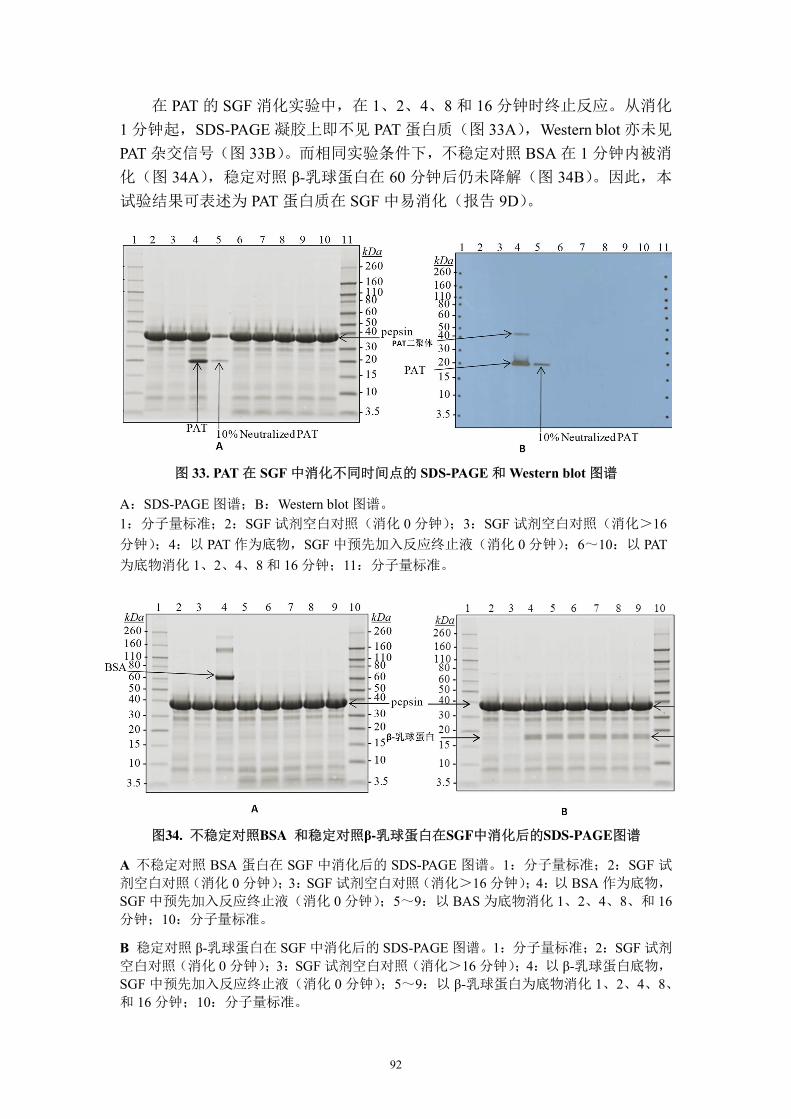
92
在 PAT 的 SGF 消化实验中,在 1、2、4、8 和 16 分钟时终止反应。从消化
1 分钟起,SDS-PAGE 凝胶上即不见 PAT 蛋白质(图 33A),Western blot 亦未见
PAT 杂交信号(图 33B)。而相同实验条件下,不稳定对照 BSA 在 1 分钟内被消
化(图 34A),稳定对照 β-乳球蛋白在 60 分钟后仍未降解(图 34B)。因此,本
试验结果可表述为 PAT 蛋白质在 SGF 中易消化(报告 9D)。
图 33. PAT 在 SGF 中消化不同时间点的 SDS-PAGE 和 Western blot 图谱
A:SDS-PAGE 图谱;B:Western blot 图谱。 1:分子量标准;2:SGF 试剂空白对照(消化 0 分钟);3:SGF 试剂空白对照(消化>16
分钟);4:以 PAT 作为底物,SGF 中预先加入反应终止液(消化 0 分钟);6~10:以 PAT
为底物消化 1、2、4、8 和 16 分钟;11:分子量标准。
图34. 不稳定对照BSA 和稳定对照β-乳球蛋白在SGF中消化后的SDS-PAGE图谱
A 不稳定对照 BSA 蛋白在 SGF 中消化后的 SDS-PAGE 图谱。1:分子量标准;2:SGF 试
剂空白对照(消化 0 分钟);3:SGF 试剂空白对照(消化>16 分钟);4:以 BSA 作为底物,
SGF 中预先加入反应终止液(消化 0 分钟);5~9:以 BAS 为底物消化 1、2、4、8、和 16分钟;10:分子量标准。
B 稳定对照 β-乳球蛋白在 SGF 中消化后的 SDS-PAGE 图谱。1:分子量标准;2:SGF 试剂
空白对照(消化 0 分钟);3:SGF 试剂空白对照(消化>16 分钟);4:以 β-乳球蛋白底物,
SGF 中预先加入反应终止液(消化 0 分钟);5~9:以 β-乳球蛋白为底物消化 1、2、4、8、和 16 分钟;10:分子量标准。

93
7)Cry1Ac 和 PAT 对哺乳动物的经口急性毒性评估
本公司评估了 Cry1Ac 和 PAT 蛋白质的急性毒性(试验设计符合最新中国
农业农村部第 2406 号公告-10-2016《蛋白质经口急性毒性试验》要求)。结果显
示,Cry1Ac 和 PAT 蛋白质对小鼠的经口急性毒性耐受剂量大于 5050 mg/(kgBW),
其 LD50>5050 mg/(kgBW)。实验内容简述于下,详见报告 8E 和报告 9E。
I. Cry1Ac 蛋白质急性毒性试验(报告 8E):
Cry1Ac 急性毒性试验体系及方法
试验动物:以 Crl:CD1(ICR)小鼠为试验动物。约 7 周龄。由 Charles River
实验室供应(Raleigh, North Carolina)。
分组:设试验蛋白 Cry1Ac 组、BSA 组、空白对照组。试验动物随机分组,
每组 16 只,雌、雄各半,三组共 48 只。
缓冲液和溶剂:在甲基纤维素水溶液中加入26.32%的 20 mM环己胺丙磺酸
(Cyclohexylaminopropane Sulfonic Acid, CAPS)制成缓冲液,其中甲基纤维素的
终浓度是0.5%。CAPS有助于蛋白质溶解。当天制备后立即使用。
对照:设 BSA 做为阴性对照,其纯度 100%,缓冲液与受试蛋白相同,灌胃
剂量也为 5050 mg/(kgBW);设 26.32% 20mM CAPS /0.5%甲基纤维素水溶液为
空白对照,灌胃量与蛋白质组相当。
动物准备:在灌胃之前将雌、雄小鼠禁食 4 小时。禁食后,记录小鼠体重,
并进行详细的临床观察。
给药及剂量:以口腔插管方式(灌胃)给药,受试蛋白质溶液总量为 60
mL/(kgBW),分 3 次给药,每次 20 mL/(kgBW), 每两小时给药一次,中间提供
少许饲料。受试蛋白质的剂量为 5050 mg/(kgBW),剂量按禁食后的体重计算。
第三次给药后立刻给动物提供饲料。
每日观察:每日至少进行一次笼边检查,且在相同的时间段检查(通常是在
早晨)。注意是否可观察到明显的临床异常,从而掌握动物的大体健康状况。所
述临床异常包括:活动减少或增加、重复性的行为、发声异常、行动失调或跛行、
受伤、神经肌肉功能异常(自发性收缩痉挛、颤抖和抽搐)、呼吸改变、皮肤和
粘膜发紫或苍白、严重的眼伤(破裂)、粪便稠度改变或粪尿量改变等表现。另
外,每天对发病情况、死亡以及进食和饮水情况进行至少观察两次。
详细的临床观察:在给药之前(即 0 小时),对所有的动物进行详细的临床
观察(DCO),在第一天第一次给药之后隔 0.5 小时、2 小时(第二次给药时)、
2.5 小时、4 小时(第三次给药时)、5 小时和 6 小时观察,以后每天一次。每日
在相同的时间段进行详细的临床观察,包括笼边、掌中和空地观察,记录观察结
果。
体重:所有动物在适应期、试验第 1、2、8 天和最后一天称重。根据试验第

94
一天禁食后的体重计算体重的增量。
解剖病理(尸检):先将试验动物用异氟烷气体和医疗级的氧气麻醉,然后
再用二氧化碳麻醉,暴露并钳住气管,将动物去头处死。尸检包括外表组织和所
有孔窍的检查,将每个角膜放在一片湿润的玻璃片上进行检查,开颅后对大脑、
垂体腺及周边颈部软组织进行检查。去皮暴露胴体,打开胸腔和腹腔对内脏进行
检查。
Cry1Ac 经口急性毒性试验结果
Cry1Ac 蛋白质灌胃剂量为 5050 mg/(kgBW),未见毒性反应,未见动物死
亡,BSA 对照组第 12 天有一只动物卡在饮水设备意外死亡。所有试验小鼠在 14
天的观察期间均存活;在试验期间体重均增长,灌胃 Cry1Ac 的小鼠平均体重及
变化均与灌胃 BSA 及空白对照组相当,即未见 Cry1Ac 蛋白质引起小鼠体重异
常;整个试验过程所有动物均表现正常,未观察到与灌胃蛋白质 Cry1Ac 相关的
临床症状;试验结束时所有小鼠的尸检解剖试验均未发现相关的组织病理学损伤,
即 Cry1Ac 蛋白质对小鼠生长发育无不良影响(报告 8E)。因此,Cry1Ac 对小
鼠的耐受剂量大于 5050 mg/(kgBW),其 LD50>5050 mg/(kgBW)。
II. PAT 蛋白质急性毒性试验(报告 9E):
PAT 急性毒性试验体系及方法
试验动物:以Crl:CD1(ICR)小鼠为试验动物。约7周龄。由Charles River实
验室供应(Kingston, New York)。
分组:设试验蛋白 PAT 组、BSA 组、空白对照组。试验动物随机分组,每
组 20 只,雌、雄各半,共 60 只。
缓冲液和溶剂:测试蛋白质溶于 0.5%甲基纤维素水溶液中。当天准备后立
即使用。
对照:设 BSA 做为阴性对照,其纯度 100%,缓冲液与受试蛋白相同,灌胃
剂量也为 5050 mg/(kgBW),设 0.5%甲基纤维素水溶液为空白对照,灌胃量与蛋
白质组相当。
动物准备:在灌胃之前将雌雄小鼠禁食不禁水 16~18 小时。禁食后,记录
小鼠体重,并进行详细的临床观察。
给药及剂量:以口腔插管(灌胃)方式给药,受试蛋白质溶液总容量为 36
mL/(kgBW),在 24 小时内分两次给药,每次 18 mL/(kgBW)。受试蛋白质的总
剂量为 5050 mg/(kgBW),剂量按禁食后的体重来计算。第二次给药后立刻给动
物提供饲料。
每日观察:每日至少进行一次笼边的检查,且在相同的时间段检查(通常是
在早晨)。注意是否可观察到明显的临床异常,从而掌握动物的大体健康状况。
所述临床异常包括:活动减少或增加、重复性的行为、发声、行动失调或跛行、

95
受伤、神经肌肉功能异常(自发性收缩痉挛、颤抖和抽搐)、呼吸改变、皮肤和
粘膜发紫或苍白、严重的眼伤(破裂)、粪便稠度变化和粪尿量的变化。另外,
每天对发病情况、死亡以及进食饮水情况进行至少观察两次。
详细的临床观察:在第一次/第二次给药之前,对所有的动物进行详细的临
床观察(DCO),在第一次给药之后隔 0.5-1、2(第二次给药)、2.5、4、5、6 和
7 小时(仅限雄性小鼠)观察,以后每天一次。每日在相同的时间段进行详细的
临床观察,包括笼边、掌中和空地观察,记录观察结果。
体重:所有动物在适应期、试验第 1、2、8 和最后一天称重。根据试验第一
天禁食后的体重计算体重的增量。
解剖病理(尸检):先将试验动物用异氟烷气体和医疗级的氧气麻醉,然后
再用二氧化碳麻醉,暴露气管并钳住,去头处死。尸检包括外表组织和所有孔窍
的检查,每个角膜放在一片湿润的玻璃片对眼部进行检查,开颅后对大脑、垂体
腺及周边颈部软组织进行检查。去皮暴露胴体,打开胸腔和腹腔对内脏进行检查。
PAT经口急性毒性试验结果
PAT蛋白质的灌胃剂量为5050 mg/(kgBW)。所有试验小鼠在14天的观察评
估期间均存活;在试验期间体重均增长,灌胃PAT的小鼠平均体重及变化均与灌
胃BSA或者空白对照组相当,即未见试验处理引起小鼠体重差异;整个试验过程
所有动物均表现正常,未观察到与灌胃蛋白PAT相关的临床症状;尸检解剖试验
均未发现组织病理学损伤,即PAT蛋白对小鼠生长发育没有不良影响(报告9E)。
对小鼠的经口急性毒性耐受剂量大于 5050 mg/(kgBW),其LD50 > 5050
mg/(kgBW)。
8)Cry1Ac 和 PAT 在人类膳食中的接触风险评估
评价原理、参数和公式
棉籽(可食部位)的加工产品为棉籽油和棉籽粕,棉籽油经脱酚后可食用,
棉籽粕主要作为饲料。DAS-21023-5中外源蛋白质Cry1Ac和PAT在人类膳食中的
摄入风险是以其在棉籽中的含量、人群棉籽产品消费量以及目标蛋白质的无害作
用量(No Observed Adverse Effect Level,NOAEL)来计算接触界限(Margins of
Exposure,MoE),以MoE做为指标评估该物质对人群的接触风险。MoE计算参数
和方法如下。
NOAEL:NOAEL在数值上与LD50相同,哺乳动物经口急性毒性试验显示,
Cry1Ac和PAT的LD50均大于5050 mg/(kgBW),详见报告8E和报告9E。
棉籽中Cry1Ac和PAT表达量:Cry1Ac和PAT在棉籽中表达量分别为0.57
ng/mg和0.06 ng/mg(报告2)。计算MoE时,保守地假设加工产品(棉籽粕、棉籽
油)中Cry1Ac和PAT蛋白质无损失、无降解、不失活。
96
人群棉产品摄入量:世界卫生组织(WHO)从各国收集并发表了食品消费的
97.5 百分位值数据(97.5th percentile),97.5 百分位值数据取自相应食品最高消
费国家的基于体重的 97.5 百分位值数据
(http://www.who.int/foodsafety/areas_work/chemical-risks/en/ ),其中美国的人群摄入棉
产品水平最高(表 21 和表 22),采用此数据来评估中国人群对棉籽产品的膳食
接触风险更为保守。估算摄入量时,保守地假定中国人群消费的棉籽产品 100%
来自棉花品系 DAS-21023-5。MoE 的计算公式如下:
计算结果
结合上述参数及公式计算MoE,表21列出了参数及计算结果,一般人群对棉
籽和棉籽油中Cry1Ac的MoE分别为1.7×108和0.6×108,儿童对棉籽中Cry1Ac的
MoE为1.7×108(尚无儿童消费棉籽油的数据)。MoE极高,因此人群对转化体DAS-
21023-5中Cry1Ac的接触风险极低。
表 21. 人群对 DAS-21023-5 成熟棉籽中 Cry1Ac 蛋白质的膳食接触界限 MoE
人群 棉籽 产品
消费者 体重
(kg)
每日 棉产品 消费量 (g/人/天)a
棉籽中 Cry1Ac 含量
(ng/mg)
Cry1Ac 接触量 e
mg/kg 体重 /天
NOAELd mg/(kgBW)
接触界限 MoEf
一般人群 全棉籽 65 3.3 0.57 0.00003 5050 >1.7×108
棉籽精炼油 65 9.1 0.57 c 0.00008 5050 >0.6×108
儿童 (1-6 岁)
全棉籽 15 0.8 0.57 0.00003 5050 >1.7×108
棉籽精炼油 15 -b 0.57 c - 5050 - a本数据为 WHO 收集的美国人群消费量,该国摄入棉产品高于中国。 b 尚未见儿童消费棉籽精炼油的数据。 c 假设棉籽表达的 Cry1Ac 蛋白在棉籽油中未损耗。 d NOAEL 的数值与 LD50相同,Cry1Ac 的 LD50>5050 mg/(kgBW)。 e 折合为每人每天每公斤体重接触 Cry1Ac 蛋白质的量。 f 假设所摄入的棉花产品 100%来自 DAS-21023-5。
表22显示普通人群对棉籽和棉籽油中PAT蛋白质的MoE分别为1.7×109和
0.6×109,儿童对棉籽中PAT蛋白质的MoE为1.7×109(尚无儿童消费棉籽油的数据)。
MoE极高,因此人群对转化体DAS-21023-5中PAT的接触风险极低。
NOEAL (mg/kg BW)
蛋白表达量(ng/mg)
接触界限 MoENOAEL
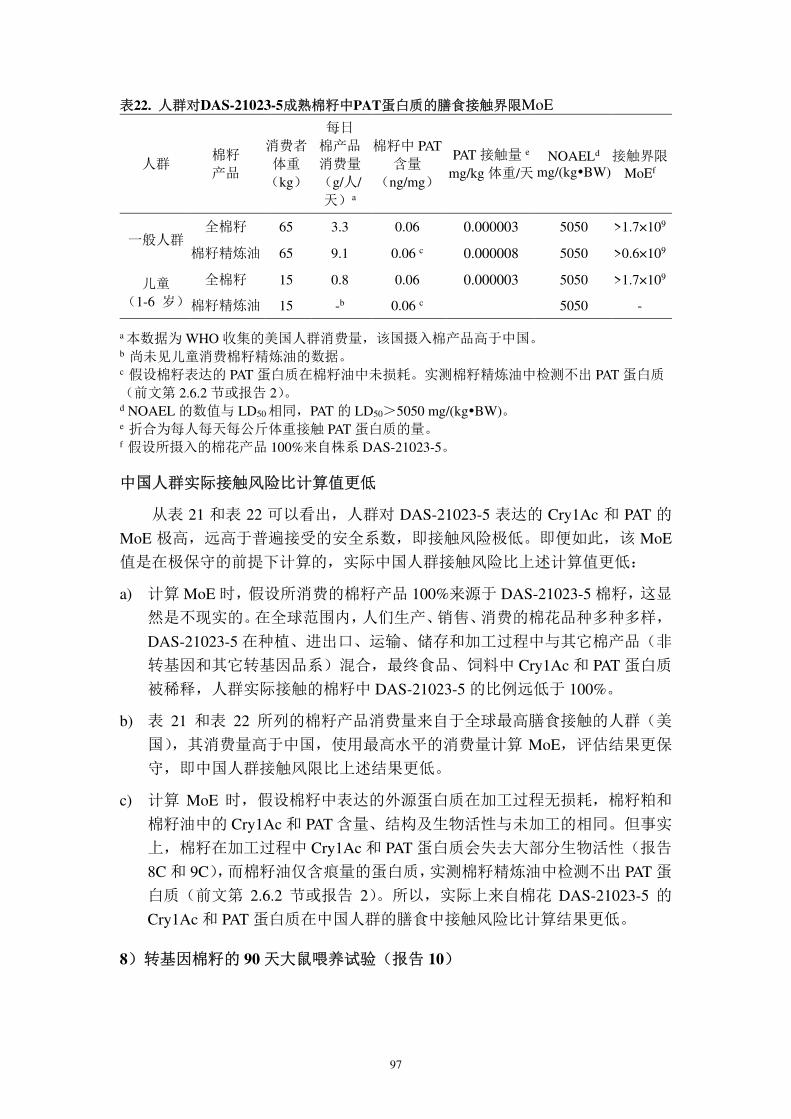
97
表22. 人群对DAS-21023-5成熟棉籽中PAT蛋白质的膳食接触界限MoE
人群 棉籽 产品
消费者 体重
(kg)
每日 棉产品 消费量 (g/人/天)a
棉籽中 PAT 含量
(ng/mg)
PAT 接触量 e mg/kg 体重/天
NOAELd mg/(kgBW)
接触界限 MoEf
一般人群 全棉籽 65 3.3 0.06 0.000003 5050 >1.7×109
棉籽精炼油 65 9.1 0.06 c 0.000008 5050 >0.6×109
儿童 (1-6 岁)
全棉籽 15 0.8 0.06 0.000003 5050 >1.7×109
棉籽精炼油 15 -b 0.06 c 5050 -
a 本数据为 WHO 收集的美国人群消费量,该国摄入棉产品高于中国。 b 尚未见儿童消费棉籽精炼油的数据。 c 假设棉籽表达的 PAT 蛋白质在棉籽油中未损耗。实测棉籽精炼油中检测不出 PAT 蛋白质
(前文第 2.6.2 节或报告 2)。 d NOAEL 的数值与 LD50相同,PAT 的 LD50>5050 mg/(kgBW)。 e 折合为每人每天每公斤体重接触 PAT 蛋白质的量。 f 假设所摄入的棉花产品 100%来自株系 DAS-21023-5。
中国人群实际接触风险比计算值更低
从表 21 和表 22 可以看出,人群对 DAS-21023-5 表达的 Cry1Ac 和 PAT 的
MoE 极高,远高于普遍接受的安全系数,即接触风险极低。即便如此,该 MoE
值是在极保守的前提下计算的,实际中国人群接触风险比上述计算值更低:
a) 计算 MoE 时,假设所消费的棉籽产品 100%来源于 DAS-21023-5 棉籽,这显
然是不现实的。在全球范围内,人们生产、销售、消费的棉花品种多种多样,
DAS-21023-5 在种植、进出口、运输、储存和加工过程中与其它棉产品(非
转基因和其它转基因品系)混合,最终食品、饲料中 Cry1Ac 和 PAT 蛋白质
被稀释,人群实际接触的棉籽中 DAS-21023-5 的比例远低于 100%。
b) 表 21 和表 22 所列的棉籽产品消费量来自于全球最高膳食接触的人群(美
国),其消费量高于中国,使用最高水平的消费量计算 MoE,评估结果更保
守,即中国人群接触风限比上述结果更低。
c) 计算 MoE 时,假设棉籽中表达的外源蛋白质在加工过程无损耗,棉籽粕和
棉籽油中的 Cry1Ac 和 PAT 含量、结构及生物活性与未加工的相同。但事实
上,棉籽在加工过程中 Cry1Ac 和 PAT 蛋白质会失去大部分生物活性(报告
8C 和 9C),而棉籽油仅含痕量的蛋白质,实测棉籽精炼油中检测不出 PAT 蛋
白质(前文第 2.6.2 节或报告 2)。所以,实际上来自棉花 DAS-21023-5 的
Cry1Ac 和 PAT 蛋白质在中国人群的膳食中接触风险比计算结果更低。
8)转基因棉籽的 90 天大鼠喂养试验(报告 10)

98
以含有 DAS-21023-5 转化体的棉籽为受试材料,以棉花 PSC355(与受试转
化体相同的遗传背景)为非转基因对照,以 3 个市售非转基因棉花 PHY72、
PHY78 和 98M-2983 为参照品系。并以 Sprague-Dawley 大鼠品系为试验动物,
每组 24 只,雌、雄各半。在基础饲料中参入 10%棉籽粕。饲喂大鼠 90 天。实
验期间,每天进行一次笼边观察,每周将大鼠从笼中抓取进行临床观察,并记
录动物个体的体重和饲料消耗情况。在饲喂开始之前和和饲喂结束之前 2 周进
行功能性测试和运动行为测试。在饲喂开始之前和和饲喂结束之前进行眼科检
查。除此之外,还需血液学检测、临床化学检测和尿检。对所有实验动物进行
解剖学分析,将指定器官称重。对所有对照组和受试物组实验动物进行全套组
织病理学检测。
试验结果显示,在临床表现、眼科学检测、功能测试、运动行为、体重、
饲料消耗、血液学、凝血时间、临床化学和尿检方面,未见与处理相关的反
应。器官重量、大体组织病理学观察方面,亦未见异常。
因此,DAS-21023-5 棉籽不会引起哺乳动物亚慢性毒性反应,未见实验动
物营养学负面反应,未见非预期效应。
9)转基因棉籽的 42 天肉鸡喂养试验(报告 11)
以含有 DAS-21023-5 转化体的棉籽为受试材料,以非转基因材料为对照组,
以 2 个市售棉花品系为参照组,以 Broiler 肉鸡为试验动物,进行了 42 天的饲喂
试验,比较各组肉鸡表现,从而评价转基因棉花在营养学方面的影响。
结果显示,在试验第 42 天时结束饲喂并称重,处理组与非转基因对照组、
2 个市售参照组间的总体重量指标无显著差异,4 组动物表现等同。在 42 天
内,转基因棉花处理组的饲料系数最低(饲料系数为摄食量/增重),因此在饲
料利用方面表现最佳。体重增重、饲料系数、死亡率指标在转基因棉花组与非
转基因对照组间的差异均不显著。
详见后文“3.3.4 营养成份”一节或报告 11。
3.3.2 过敏性;
通过以下 6 方面的分析,证明了转基因棉花 DAS-21023-5 及其表达两个外源
蛋白质 Cry1Ac 和 PAT 诱发人、畜过敏反应的概率极低:
1) cry1Ac 和 pat 基因及其供体生物具有安全使用历史;
2) Cry1Ac 和 PAT 蛋白质具有安全应用历史;
3) Cry1Ac 和 PAT 与已知的致敏原蛋白质的氨基酸序列无较高的相似性;
4) Cry1Ac 和 PAT 蛋白质受热后免疫活性和功能活性不稳定;
5) Cry1Ac 和 PAT 蛋白质在模拟胃液 SGF 中易消化;
6) Cry1Ac 和 PAT 蛋白质在膳食中接触致敏的风险极低。

99
具体评估如下:
1)Cry1Ac 和 PAT 蛋白质供体生物具有安全使用的历史
如前文第 3.3.1 部分所描述,DAS-21023-5 棉花中外源表达的蛋白质 Cry1Ac
和 PAT 分别来源于广泛存在于自然界的、安全的供体生物苏云金芽孢杆菌和绿
色产色链霉菌。应用多年,至今未见这两类微生物对人类和动物有危害的报道。
这两类微生物也不属于已知的过敏原生物。
2)外源表达蛋白质的安全应用历史
如前文第 3.3.1 部分所描述,Cry1Ac 和 PAT 蛋白质均有长期的安全使用历
史。至今未见这两种蛋白质致敏的报导。
3)外源表达蛋白质与已知蛋白质过敏原序列无较高的相似性(2020 年)
在评估转基因植物中新表达蛋白质的安全性时,会采用证据权重方法,而分
析新表达的蛋白质与已知和假定的致敏原的潜在交叉反应性是证据权重法的关
键之一(Codex Alimentarius Commission, 2003)。本节根据中国国家标准(农业农
村部 1485 号公告-18-2010)和国际标准(Codex Alimentarius Commission, 2003;
FAO/WHO, 2001)对 2 个外源蛋白质的致敏性进行了评估,即采用生物信息学比
对分析 Cry1Ac 和 PAT 的氨基酸序列与已知的及假定的致敏原的潜在交叉反应
性。
评价原则、数据库与工具:蛋白质致敏原资源数据库(COMPARE,
http://comparedatabase.org)是一个公共的、内容广泛的、经同行审定的数据库,
于 2022 年更新,当时含有 2348 个序列,是健康与环境科学研究所(Health and
Environmental Sciences Institute,HESI)蛋白质致敏性、毒性和生物信息学(Protein
Allergens, Toxins, and Bioinformatics,PATB)委员会的合作成果。将目标蛋白质
序列与 COMPARE 数据库中的序列进行了两种独立的搜索:一种是将目标序列
的连续重叠 80 个氨基酸为一个单元(序列窗口),采用 FASTA 程序(v35. 4.4)
在默认参数(Matrix = BLOSUM50; Gap Penalties = -12/-2; ktup = 2,E 值= 10)下
与数据库中的致敏原序列进行比对搜索,然后在输出的匹配中检索同源性>35%
的 序 列 ; 另 一 种比对 分 析 是 采 用 FUZZPRO 程 序 (http://emboss.open-
bio.org/rel/rel6/apps/fuzzpro.html) 搜素是否有连续 8 个氨基酸序列等同。
比对结果:将蛋白质 Cry1Ac 和 PAT 序列分别与 COMPARE 数据库中已知
的和假定的致敏原序列进行比较搜索后,均未发现在≥80 连续氨基酸上同源
性>35%的匹配序列;且均未发现这两个蛋白与致敏原序列有连续 8 个氨基酸的
完全匹配(报告 8B 和 9B)。因此,上述 2 个蛋白质与已知蛋白致敏原没有较高
的氨基酸序列相似性,即生物信息学分析未见其潜在的过敏性。
4)外源表达 Cry1Ac 和 PAT 蛋白质受热不稳定
通常认为受热稳定是一般致敏或致毒物质的特征(Codex, 2003b),Cry1Ac

100
和 PAT 受热不稳定,不具备致敏或致毒物质受热稳定的特征。
100 µg/mL 和 50 µg/mL 浓度的 Cry1Ac在 91℃加热 60 分钟时,降解了 68%,
几乎全部失去免疫亲和活性(报告 8C)。将 PAT 蛋白质在 25℃、37℃、55℃、
75℃和 95℃加热 30 分钟,虽然分子量无变化,在 55℃或更高温度时几乎检测
不到免疫亲合活性,55℃时酶活性丧失>99%(报告 9C)。
详见前文第“3.3.1 毒性”一节。
5)Cry1Ac 和 PAT 蛋白质在模拟胃液 SGF 中易消化
通常认为在消化道中不被消化和加热稳定是一般致敏或致毒物质的特征之
一(Codex, 2003b)。Cry1Ac 和 PAT 蛋白质在模拟胃液(SGF)中 1 分钟内消化
(报告 8D 和报告 9D),即易被模拟胃液消化。因此不具备一般致毒或致敏蛋白
质耐消化的特征。详见前文第“3.3.1 毒性”一节。
6)Cry1Ac 和 PAT 蛋白质在膳食中接触致敏的风险极低
棉籽的加工产品棉籽油和棉籽饼是人、畜食用或饲用的棉籽产品。DAS-
21023-5 中 Cry1Ac 和 PAT 在籽粒中表达量分别为 0.57 ng/mg 和 0.06 ng/mg 组织
干重。保守估计,成人每人每日接触全棉籽中 Cry1Ac 和 PAT 的量分别是为
0.00003 mg/kg·BW(表 21)和 0.000003 mg/kg·BW(表 22),接触量极低,接触
致敏的风险极低。
3.3.3 抗营养因子;
生物信息学分析表明外源蛋白质 Cry1Ac 和 PAT 不属于抗营养因子;成分测
定分析表明转化体 DAS-21023-5 与非转基因对照的抗营养因子实质等同。
1)生物信息学表明外源蛋白质不属于已知的抗营养因子
2020 年,将外源蛋白质 Cry1Ac 和 PAT 氨基酸序列与 NCBI-nr
(http://www.ncbi.nlm.nih.gov/)和 UniProt(https://www.uniprot.org/blast/)公共蛋
白质数据库进行了 BLASTp 搜索,并在输出的匹配中搜索关键词“agglutinin”、
“anti-nutrient”、 “antinutrient”、 “anti-nutritional”、 “protease inhibitor”和 “trpsin
inhibitor”。未见 Cry1Ac 和 PAT 与已知的对人类和动物有害的毒性蛋白质、抗营
养因子存在较高的序列相似性(报告 8A 和报告 9A)。
2)抗营养因子含量测定
2001 年在美国主要棉区的 6 个试验地点取样,检测了 DAS-21023-5 及其对
照棉籽、棉籽加工产品中抗营养因子的含量,并抽取 2 试验点检测了棉花顶端叶
片和棉铃中抗营养因子的含量(报告 2)。试验地点位于美国亚利桑那州、加利福
尼亚州、密西西比州、北卡罗来纳州和得克萨斯州,这些地点代表了美国棉花多
样化的农事操作与生态环境。
对如下抗营养因子含量进行了测定:棉籽中的总棉酚、环丙基类脂肪酸、黄

101
曲霉毒素,棉仁中的总棉酚和游离棉酚。结果显示,上述各项指标的 6 个试验点
平均值在 DAS-21023-5 与非转基因棉籽之间含量相似,且均在文献报道值范围
内(表 23)。
对两个田间试验点抽样分析,测定了棉花顶端叶片和棉铃中总多元酚和总棉
酚的含量(表 24)。比较结果显示,DAS-21023-5 棉花与其非转基因对照间总多
元酚和总棉酚含量相近。
因此,DAS-21023-5 棉花与非转基因棉籽的抗营养因子等同。
表 23. 棉籽和棉仁中抗营养因子含量分析
组织 成分(%) 文献范围a 非转基因对照 DAS-21023-5 棉花
棉籽
总棉酚 0.71~1.24 0.841 0.852
环丙基类脂肪酸
苹婆酸 0.13~0.66 0.311 0.289
锦葵酸 0.17~0.61 1.9 0.384 0.34
二羟基苹婆酸 0.11~0.22 0.213 0.212
黄曲霉毒素(ppb)
AFB1 NA < 1.00 < 1.00 AFB2 NA < 1.00 < 1.00 AFG1 NA < 1.00 < 1.00 AFG2 NA < 1.00 < 1.00
棉仁 总棉酚 NA 0.976 1.15
游离棉酚 NA 0.839 1 a:OECD, 2009。NA:无文献报导。
表 24. 不同组织中抗营养因子含量的分析
组织 取样地点 成分(%) 非转基因对照 DAS-21023-5 棉花
棉花顶端
叶片
北卡试验地点 总多元酚 1.53 1.6
总棉酚 0.031 0.056
德州试验地点 1 总多元酚 0.56 0.73
总棉酚 <0.020 0.087
棉铃
北卡试验地点 总多元酚 0.74 0.7
总棉酚 0.081 0.081
德州试验地点 1 总多元酚) 0.68 0.73
总棉酚 0.094 0.109
3.3.4 营养成份
物质基础等同可反应出转基因棉花与其非转基因对照的安全性等同。一方
面,营养成份测定结果显示 DAS-21023-5 棉花与非转基因棉花营养成份等同
(报告 2),另一方面,动物喂养实验显示转基因棉花不会对哺乳动物和禽类带
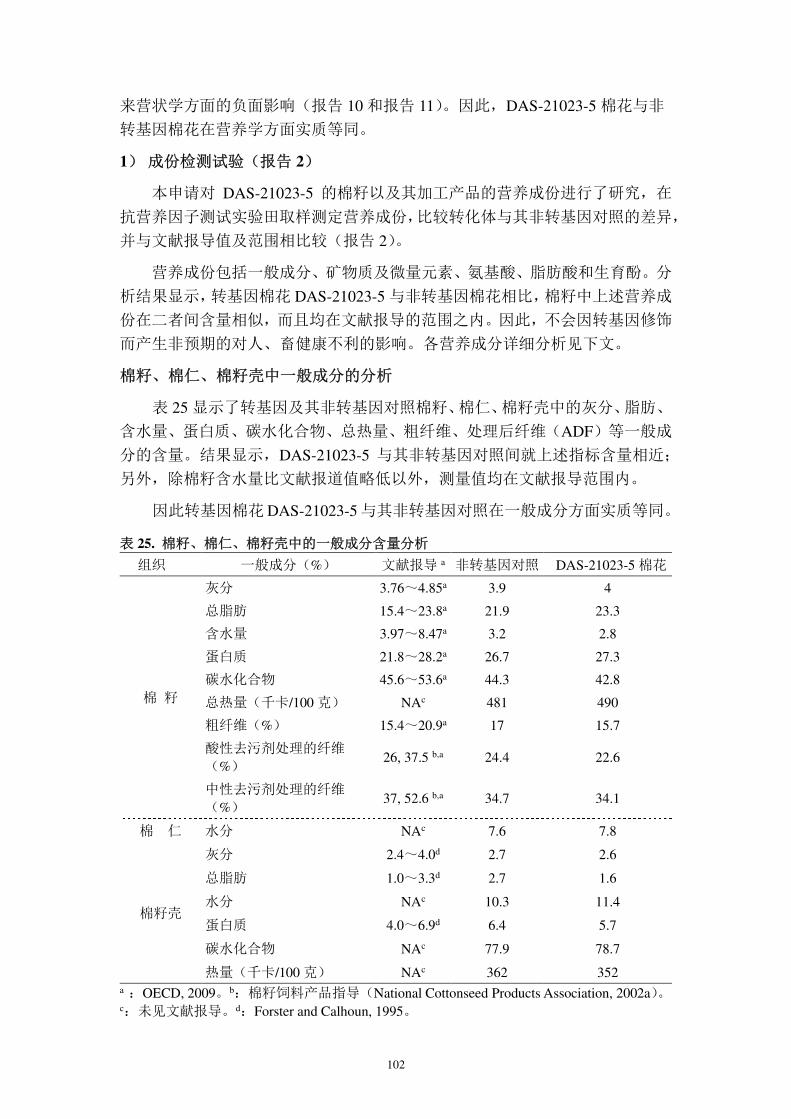
102
来营状学方面的负面影响(报告 10 和报告 11)。因此,DAS-21023-5 棉花与非
转基因棉花在营养学方面实质等同。
1) 成份检测试验(报告 2)
本申请对 DAS-21023-5 的棉籽以及其加工产品的营养成份进行了研究,在
抗营养因子测试实验田取样测定营养成份,比较转化体与其非转基因对照的差异,
并与文献报导值及范围相比较(报告 2)。
营养成份包括一般成分、矿物质及微量元素、氨基酸、脂肪酸和生育酚。分
析结果显示,转基因棉花 DAS-21023-5 与非转基因棉花相比,棉籽中上述营养成
份在二者间含量相似,而且均在文献报导的范围之内。因此,不会因转基因修饰
而产生非预期的对人、畜健康不利的影响。各营养成分详细分析见下文。
棉籽、棉仁、棉籽壳中一般成分的分析
表 25 显示了转基因及其非转基因对照棉籽、棉仁、棉籽壳中的灰分、脂肪、
含水量、蛋白质、碳水化合物、总热量、粗纤维、处理后纤维(ADF)等一般成
分的含量。结果显示,DAS-21023-5 与其非转基因对照间就上述指标含量相近;
另外,除棉籽含水量比文献报道值略低以外,测量值均在文献报导范围内。
因此转基因棉花 DAS-21023-5 与其非转基因对照在一般成分方面实质等同。
表 25. 棉籽、棉仁、棉籽壳中的一般成分含量分析
组织 一般成分(%) 文献报导 a 非转基因对照 DAS-21023-5 棉花
棉 籽
灰分 3.76~4.85a 3.9 4
总脂肪 15.4~23.8a 21.9 23.3
含水量 3.97~8.47a 3.2 2.8
蛋白质 21.8~28.2a 26.7 27.3
碳水化合物 45.6~53.6a 44.3 42.8
总热量(千卡/100 克) NAc 481 490
粗纤维(%) 15.4~20.9a 17 15.7
酸性去污剂处理的纤维
(%) 26, 37.5 b,a 24.4 22.6
中性去污剂处理的纤维
(%) 37, 52.6 b,a 34.7 34.1
棉 仁 水分 NAc 7.6 7.8
棉籽壳
灰分 2.4~4.0d 2.7 2.6
总脂肪 1.0~3.3d 2.7 1.6
水分 NAc 10.3 11.4
蛋白质 4.0~6.9d 6.4 5.7
碳水化合物 NAc 77.9 78.7
热量(千卡/100 克) NAc 362 352 a :OECD, 2009。b:棉籽饲料产品指导(National Cottonseed Products Association, 2002a)。c:未见文献报导。d:Forster and Calhoun, 1995。
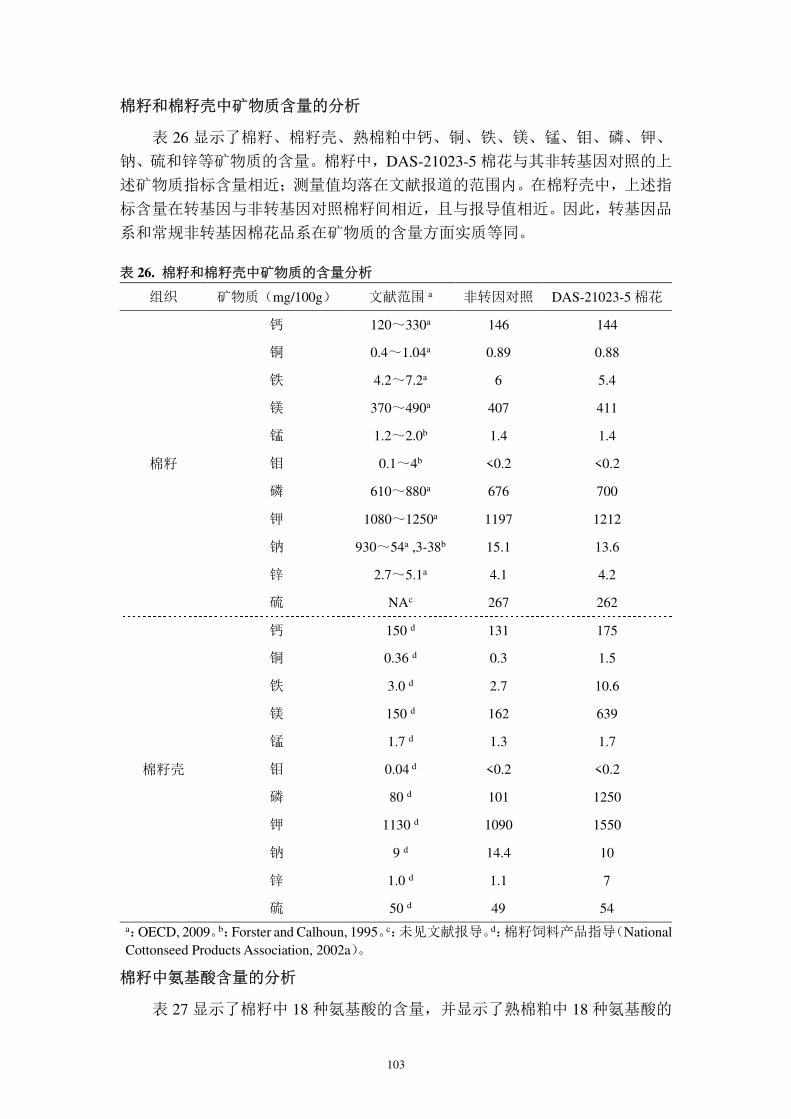
103
棉籽和棉籽壳中矿物质含量的分析
表 26 显示了棉籽、棉籽壳、熟棉粕中钙、铜、铁、镁、锰、钼、磷、钾、
钠、硫和锌等矿物质的含量。棉籽中,DAS-21023-5 棉花与其非转基因对照的上
述矿物质指标含量相近;测量值均落在文献报道的范围内。在棉籽壳中,上述指
标含量在转基因与非转基因对照棉籽间相近,且与报导值相近。因此,转基因品
系和常规非转基因棉花品系在矿物质的含量方面实质等同。
表 26. 棉籽和棉籽壳中矿物质的含量分析
组织 矿物质(mg/100g) 文献范围 a 非转因对照 DAS-21023-5 棉花
棉籽
钙 120~330a 146 144
铜 0.4~1.04a 0.89 0.88
铁 4.2~7.2a 6 5.4
镁 370~490a 407 411
锰 1.2~2.0b 1.4 1.4
钼 0.1~4b <0.2 <0.2
磷 610~880a 676 700
钾 1080~1250a 1197 1212
钠 930~54a ,3-38b 15.1 13.6
锌 2.7~5.1a 4.1 4.2
硫 NAc 267 262
棉籽壳
钙 150 d 131 175
铜 0.36 d 0.3 1.5
铁 3.0 d 2.7 10.6
镁 150 d 162 639
锰 1.7 d 1.3 1.7
钼 0.04 d <0.2 <0.2
磷 80 d 101 1250
钾 1130 d 1090 1550
钠 9 d 14.4 10
锌 1.0 d 1.1 7
硫 50 d 49 54 a:OECD, 2009。b:Forster and Calhoun, 1995。c:未见文献报导。d:棉籽饲料产品指导(National Cottonseed Products Association, 2002a)。
棉籽中氨基酸含量的分析
表 27 显示了棉籽中 18 种氨基酸的含量,并显示了熟棉粕中 18 种氨基酸的
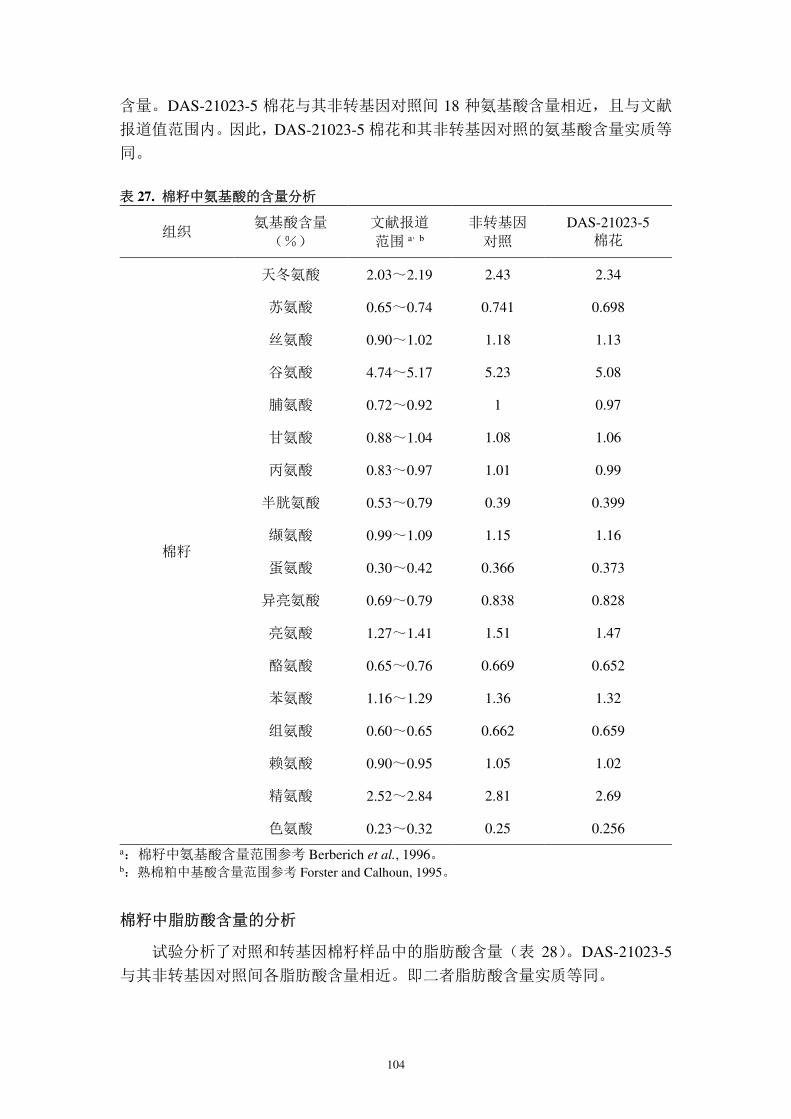
104
含量。DAS-21023-5 棉花与其非转基因对照间 18 种氨基酸含量相近,且与文献
报道值范围内。因此,DAS-21023-5 棉花和其非转基因对照的氨基酸含量实质等
同。
表 27. 棉籽中氨基酸的含量分析
组织 氨基酸含量
(%) 文献报道 范围 a,b
非转基因 对照
DAS-21023-5 棉花
棉籽
天冬氨酸 2.03~2.19 2.43 2.34
苏氨酸 0.65~0.74 0.741 0.698
丝氨酸 0.90~1.02 1.18 1.13
谷氨酸 4.74~5.17 5.23 5.08
脯氨酸 0.72~0.92 1 0.97
甘氨酸 0.88~1.04 1.08 1.06
丙氨酸 0.83~0.97 1.01 0.99
半胱氨酸 0.53~0.79 0.39 0.399
缬氨酸 0.99~1.09 1.15 1.16
蛋氨酸 0.30~0.42 0.366 0.373
异亮氨酸 0.69~0.79 0.838 0.828
亮氨酸 1.27~1.41 1.51 1.47
酪氨酸 0.65~0.76 0.669 0.652
苯氨酸 1.16~1.29 1.36 1.32
组氨酸 0.60~0.65 0.662 0.659
赖氨酸 0.90~0.95 1.05 1.02
精氨酸 2.52~2.84 2.81 2.69
色氨酸 0.23~0.32 0.25 0.256
a:棉籽中氨基酸含量范围参考 Berberich et al., 1996。 b:熟棉粕中基酸含量范围参考 Forster and Calhoun, 1995。
棉籽中脂肪酸含量的分析
试验分析了对照和转基因棉籽样品中的脂肪酸含量(表 28)。DAS-21023-5
与其非转基因对照间各脂肪酸含量相近。即二者脂肪酸含量实质等同。
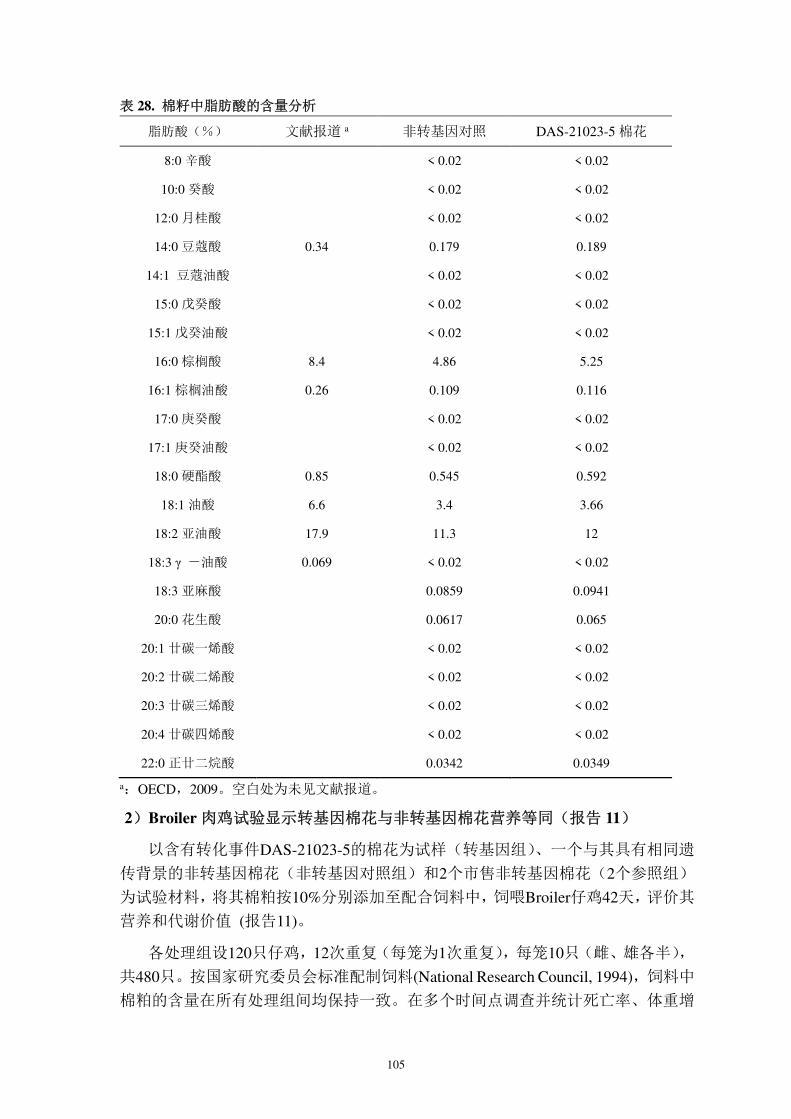
105
表 28. 棉籽中脂肪酸的含量分析
脂肪酸(%) 文献报道 a 非转基因对照 DAS-21023-5 棉花
8:0 辛酸 < 0.02 < 0.02
10:0 癸酸 < 0.02 < 0.02
12:0 月桂酸 < 0.02 < 0.02
14:0 豆蔻酸 0.34 0.179 0.189
14:1 豆蔻油酸 < 0.02 < 0.02
15:0 戊癸酸 < 0.02 < 0.02
15:1 戊癸油酸 < 0.02 < 0.02
16:0 棕榈酸 8.4 4.86 5.25
16:1 棕榈油酸 0.26 0.109 0.116
17:0 庚癸酸 < 0.02 < 0.02
17:1 庚癸油酸 < 0.02 < 0.02
18:0 硬酯酸 0.85 0.545 0.592
18:1 油酸 6.6 3.4 3.66
18:2 亚油酸 17.9 11.3 12
18:3 γ -油酸 0.069 < 0.02 < 0.02
18:3 亚麻酸 0.0859 0.0941
20:0 花生酸 0.0617 0.065
20:1 廿碳一烯酸 < 0.02 < 0.02
20:2 廿碳二烯酸 < 0.02 < 0.02
20:3 廿碳三烯酸 < 0.02 < 0.02
20:4 廿碳四烯酸 < 0.02 < 0.02
22:0 正廿二烷酸 0.0342 0.0349
a:OECD,2009。空白处为未见文献报道。
2)Broiler 肉鸡试验显示转基因棉花与非转基因棉花营养等同(报告 11)
以含有转化事件DAS-21023-5的棉花为试样(转基因组)、一个与其具有相同遗
传背景的非转基因棉花(非转基因对照组)和2个市售非转基因棉花(2个参照组)
为试验材料,将其棉粕按10%分别添加至配合饲料中,饲喂Broiler仔鸡42天,评价其
营养和代谢价值 (报告11)。
各处理组设120只仔鸡,12次重复(每笼为1次重复),每笼10只(雌、雄各半),
共480只。按国家研究委员会标准配制饲料(National Research Council, 1994),饲料中
棉粕的含量在所有处理组间均保持一致。在多个时间点调查并统计死亡率、体重增
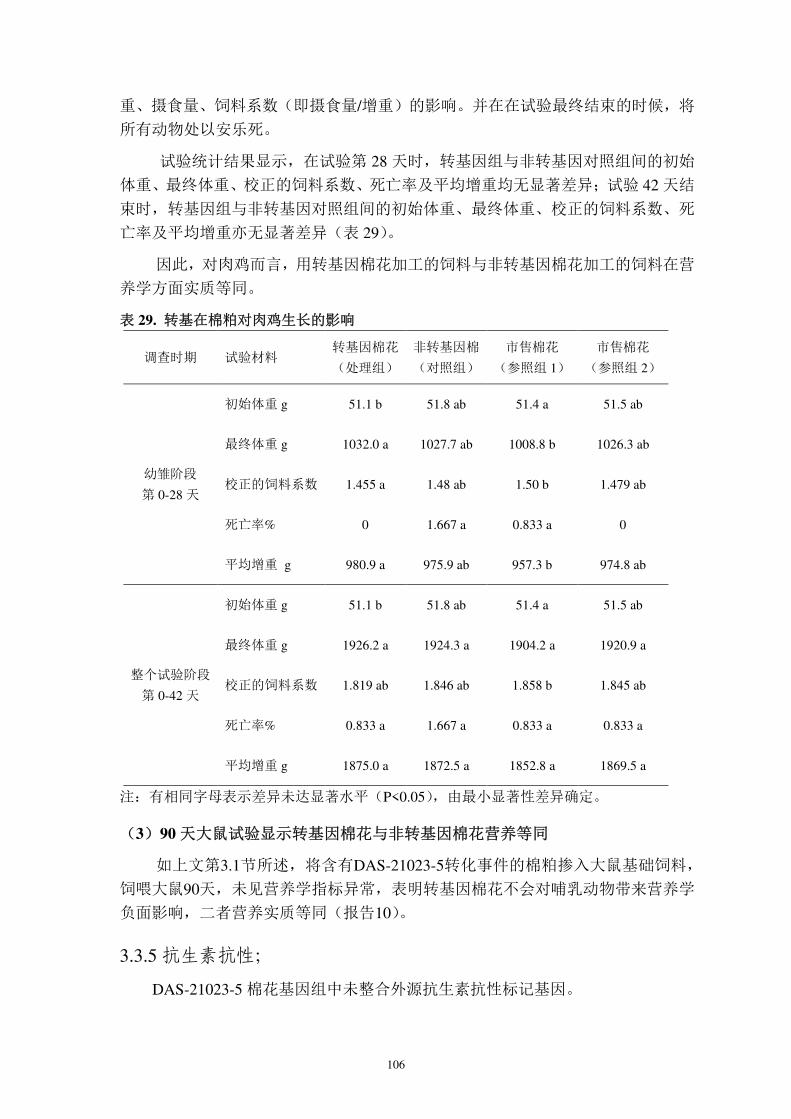
106
重、摄食量、饲料系数(即摄食量/增重)的影响。并在在试验最终结束的时候,将
所有动物处以安乐死。
试验统计结果显示,在试验第 28 天时,转基因组与非转基因对照组间的初始
体重、最终体重、校正的饲料系数、死亡率及平均增重均无显著差异;试验 42 天结
束时,转基因组与非转基因对照组间的初始体重、最终体重、校正的饲料系数、死
亡率及平均增重亦无显著差异(表 29)。
因此,对肉鸡而言,用转基因棉花加工的饲料与非转基因棉花加工的饲料在营
养学方面实质等同。
表 29. 转基在棉粕对肉鸡生长的影响
调查时期 试验材料 转基因棉花
(处理组)
非转基因棉
(对照组)
市售棉花
(参照组 1)
市售棉花
(参照组 2)
幼雏阶段
第 0-28 天
初始体重 g 51.1 b 51.8 ab 51.4 a 51.5 ab
最终体重 g 1032.0 a 1027.7 ab 1008.8 b 1026.3 ab
校正的饲料系数 1.455 a 1.48 ab 1.50 b 1.479 ab
死亡率% 0 1.667 a 0.833 a 0
平均增重 g 980.9 a 975.9 ab 957.3 b 974.8 ab
整个试验阶段
第 0-42 天
初始体重 g 51.1 b 51.8 ab 51.4 a 51.5 ab
最终体重 g 1926.2 a 1924.3 a 1904.2 a 1920.9 a
校正的饲料系数 1.819 ab 1.846 ab 1.858 b 1.845 ab
死亡率% 0.833 a 1.667 a 0.833 a 0.833 a
平均增重 g 1875.0 a 1872.5 a 1852.8 a 1869.5 a
注:有相同字母表示差异未达显著水平(P<0.05),由最小显著性差异确定。
(3)90 天大鼠试验显示转基因棉花与非转基因棉花营养等同
如上文第3.1节所述,将含有DAS-21023-5转化事件的棉粕掺入大鼠基础饲料,
饲喂大鼠90天,未见营养学指标异常,表明转基因棉花不会对哺乳动物带来营养学
负面影响,二者营养实质等同(报告10)。
3.3.5 抗生素抗性;
DAS-21023-5 棉花基因组中未整合外源抗生素抗性标记基因。

107
首先,转化质粒载体 pMYC3006 的 T-DNA 上无抗生素抗性标记基因(图
22)。其次,虽然 pMYC3006 的骨架序列中含有红霉素抗性基因,但不能转化至
棉花受体基因组中。用覆盖载体骨架全序列的探针与 DAS-21023-5 棉花基因组
DNA 进行 Southern blot 时,未见杂交信号,证明该转化体上无载体骨架序列,
更无红霉素抗性基因(详细论述见 2.2.5 部分或报告 1E)。
3.3.6 对人体和食品安全性的其它影响。
未见 DAS-21023-5 对人体和食品安全性的其它影响。另外,本公司对除草剂
残留和真菌毒素情况进行了分析评价。
1)草铵膦残留风险极低
DAS-21023-5 的目标性状是抗特定的鳞翅目害虫。虽然其含有 pat 基因,表
达 PAT 蛋白的植株可耐受除草剂草铵膦,但是此功能仅用做转化筛选时的选择
性标记。此特性一般不用于大田喷洒草铵膦。即便如此,陶氏益农公司于 2013
年在美国 8 个试验点对含有 DAS-21023-5 的转基因棉花进行除草剂残留检测。
分两次喷施,草铵膦总剂量为 1346 g ae/ha。检测结果显示,草铵膦(包括草铵
膦本身及其代谢物 NAG 和 MPPA)在棉籽中的平均残留量为 0.042 µg/g,这一
数值远小于美国 EPA 对棉花最大残留限量 4.0 µg/g(表 30)。
表 30.草铵膦在含有 DAS-21023-5 的转基因棉籽中残留量
分析样品 施药 剂量 a
(g ae/ha)
安全间 隔期 b (天)
样本量
残留量(µg/g)c
最小 最大 中位数 平均值 标准差
未脱绒棉籽 1346 91-136 16 未检出 0.13 0.035 0.042 0.032
a 样品经过两次除草剂的处理,间隔8-14天。 b 安全间隔期:采收田间样本距离最后一次喷施草铵膦的天数。 c包括草铵膦本身及其代谢物NAG和MPPA,以草铵膦酸当量表示。
2)籽粒中真菌毒素风险极低
遗传转化不会提高DAS-21023-5籽粒中真菌毒素的含量。
以大鼠喂养试验所用材料为例,在配制饲料之前,检测了转基因棉籽、非转
基因对照棉籽、商业化参照品系(98M-2983、PHY72和PHY78)棉籽中的真菌毒
素。采用液相色谱-质谱联用(LC-MSMS)的方法对真菌毒素进行测定。测定和
比较结果显示,黄曲酶毒素、玉米赤霉烯酮、DON、各项伏马毒素、T-2毒素、
各项麦角克碱等各指标在转基因棉籽与其非转基因对照间含量一致,且转基因棉
籽与商业化参照品系间含量一致(表31)。表明遗传转化不会提高转基因棉籽中
真菌毒素含量,不会因此带来食用、饲用风险。
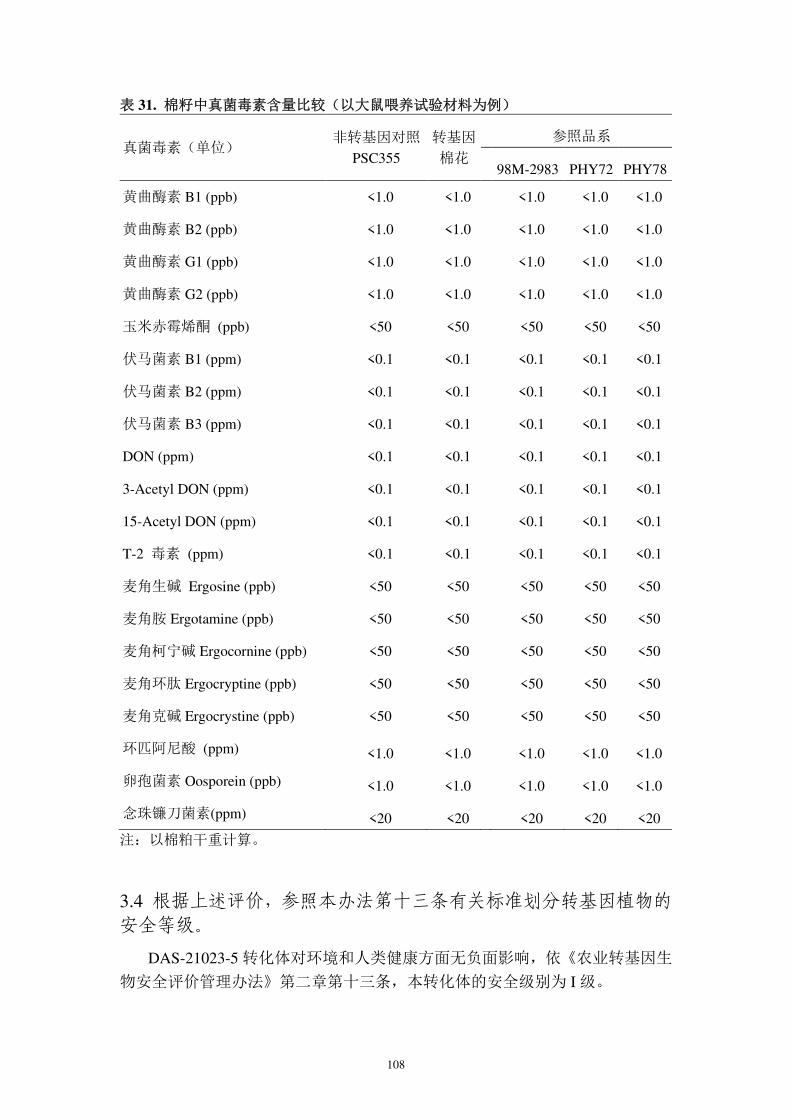
108
表 31. 棉籽中真菌毒素含量比较(以大鼠喂养试验材料为例)
真菌毒素(单位) 非转基因对照
PSC355 转基因 棉花
参照品系
98M-2983 PHY72 PHY78
黄曲酶素 B1 (ppb) <1.0 <1.0 <1.0 <1.0 <1.0
黄曲酶素 B2 (ppb) <1.0 <1.0 <1.0 <1.0 <1.0
黄曲酶素 G1 (ppb) <1.0 <1.0 <1.0 <1.0 <1.0
黄曲酶素 G2 (ppb) <1.0 <1.0 <1.0 <1.0 <1.0
玉米赤霉烯酮 (ppb) <50 <50 <50 <50 <50
伏马菌素 B1 (ppm) <0.1 <0.1 <0.1 <0.1 <0.1
伏马菌素 B2 (ppm) <0.1 <0.1 <0.1 <0.1 <0.1
伏马菌素 B3 (ppm) <0.1 <0.1 <0.1 <0.1 <0.1
DON (ppm) <0.1 <0.1 <0.1 <0.1 <0.1
3-Acetyl DON (ppm) <0.1 <0.1 <0.1 <0.1 <0.1
15-Acetyl DON (ppm) <0.1 <0.1 <0.1 <0.1 <0.1
T-2 毒素 (ppm) <0.1 <0.1 <0.1 <0.1 <0.1
麦角生碱 Ergosine (ppb) <50 <50 <50 <50 <50
麦角胺 Ergotamine (ppb) <50 <50 <50 <50 <50
麦角柯宁碱 Ergocornine (ppb) <50 <50 <50 <50 <50
麦角环肽 Ergocryptine (ppb) <50 <50 <50 <50 <50
麦角克碱 Ergocrystine (ppb) <50 <50 <50 <50 <50
环匹阿尼酸 (ppm) <1.0 <1.0 <1.0 <1.0 <1.0
卵孢菌素 Oosporein (ppb) <1.0 <1.0 <1.0 <1.0 <1.0
念珠镰刀菌素(ppm) <20 <20 <20 <20 <20
注:以棉粕干重计算。
3.4 根据上述评价,参照本办法第十三条有关标准划分转基因植物的
安全等级。
DAS-21023-5 转化体对环境和人类健康方面无负面影响,依《农业转基因生
物安全评价管理办法》第二章第十三条,本转化体的安全级别为 I 级。

109
4 转基因植物产品的安全性评价
4.1 生产、加工活动对转基因植物安全性的影响。
棉花的生产加工活动不影响转基因棉花 DAS-21023-5 及其产品的安全性。
二者加工过程相同
对转基因棉花 DAS-21023-5 与非转基因棉花的加工活动无异,二者不区分处
理。棉花加工主要分为棉纤维加工及棉籽加工两部分,其过程及影响如下。
棉纤维加工过程:收获后的棉花首先进行纤维和棉籽的分离,即直接对籽棉
进行加工,使之成为皮棉、短绒、棉籽的过程。此加工过程采用的相关工艺包括
烘干(或加湿)以及机械压轧等。加热不会增加转基因棉花 DAS-21023-5 及产品
的风险(报告 8C 和报告 9C)。
棉籽的加工过程:籽棉剥离出的棉籽可进一步加工成棉籽油、棉籽粕、棉籽
壳和棉籽绒等(OECD,2009)。加工过程同样采用了物理、机械的加工工艺,尤
其是棉籽油需进一步加工才可以食用。在生产棉籽油过程中,棉籽脱绒、棉籽剥
壳与仁壳分离、棉籽轧胚、蒸炒、棉籽的压榨过程通常采用物理的机械的方式(图
35),蒸炒和压榨加热处理不会增加转基因棉花 DAS-21023-5 及产品的风险(报
告 8C 和报告 9C)。
转基因棉花 DAS-21023-5 与传统非转基因棉花的抗营养成分等同(前文第
3.3.3 节),营养因子含量等同(前文第 3.3.4 节),即二者物质基础等同。且转基
因棉花与非转基因棉花的加工过程相同,理论上,相同的物质基础经过相同的加
工活动所生产的产品的安全性也相同。
实验数据表明,二者加工产品的关键成分实质等同
实验数据也表明,二者加工产品的关键成分实质等同。
2001 年在美国主要棉区的 6 个试验地点取样,不仅测定了转基因与其非转
基因棉籽营养成份和抗营养因子的含量,而且还比较了二者加工产品的(熟棉粕、
精炼棉籽油)营养成分(报告 2)。
表 32 比较了熟棉粕和棉籽油中的抗营养因子,表 33 比较了熟棉粕和棉籽油
中的一般成分,表 34 比较了熟棉粕中的矿物质,表 35 比较了熟棉粕中氨基酸,
表 36 比较了棉籽油中脂肪酸和生育酚。就上述指标,在转基因与非转基因对照
棉花间相近,或与文献报导值相近,或在报导范围内。说明转基因棉花 DAS-
21023-5 加工产品的营养成分与非转基因的等同。

110
图 35. 棉籽加工流程(OECD,2009)
熟棉粕和棉籽油中抗营养因子分析
对如下抗营养因子含量进行了测定:熟棉粕的总棉酚和游离棉酚,棉籽油中
的总棉酚、游离棉酚和环丙基类脂肪酸。结果显示,上述各项指标的 6 个试验点
平均值在 DAS-21023-5 棉花与非转基因之间含量相似,且均在文献报道值范围
内(表 32)。因此DAS-21023-5棉花与非转基因棉花加工产品的抗营养因子等同。
表 32. 棉籽加工产品中抗营养因子含量比较
组织 成分(%) 文献范围 a 非转基因对照 DAS-21023-5 棉花
熟棉粕 总棉酚 0.93~1.43 0.907 1.09
游离棉酚 0.02~0.07 0.044 0.045
棉籽油
总棉酚 0.09 < 0.002 < 0.002
游离棉酚 NA < 0.002 < 0.002
环丙基类脂肪酸
苹婆酸 0.58 0.217 0.223
锦葵花素 0.41 0.272 0.255
羟基苹婆酸 0.22 0.212 0.202 a:OECD, 2009。NA:无文献报导。
熟棉粕和精炼棉籽油中一般成分的分析
表 33 显示了转基因及其非转基因对照熟棉粕和精炼棉籽油中的灰分、脂肪、
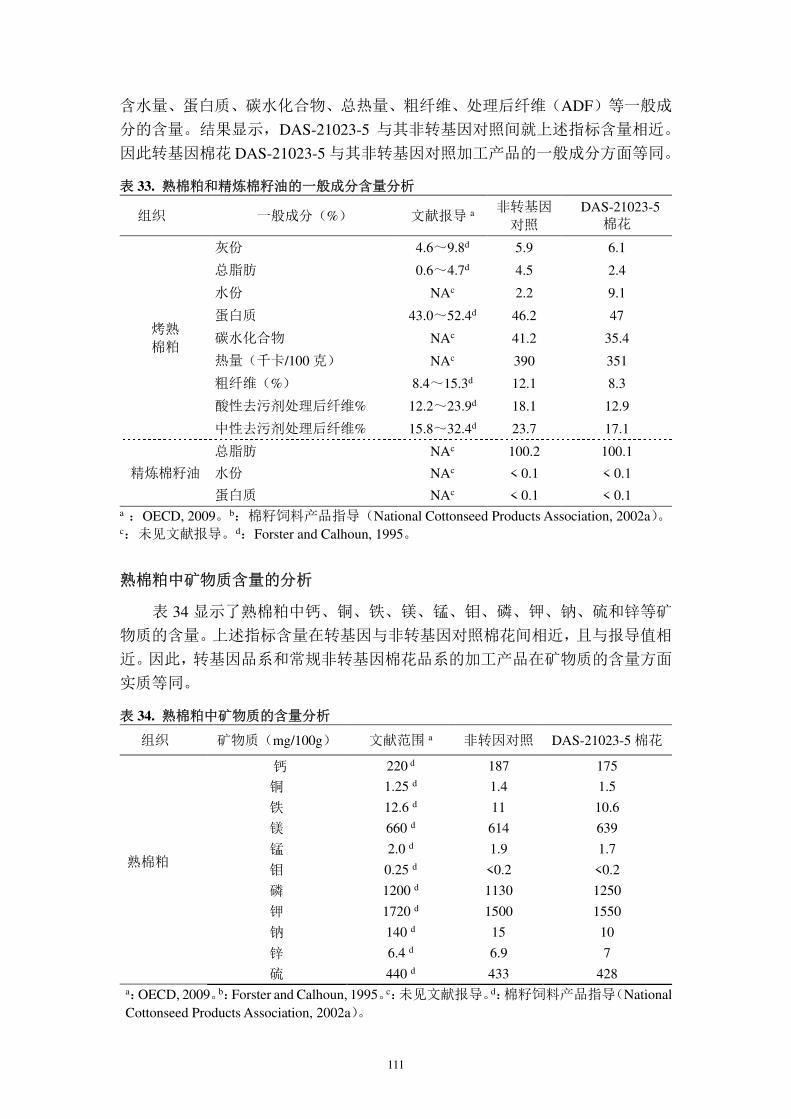
111
含水量、蛋白质、碳水化合物、总热量、粗纤维、处理后纤维(ADF)等一般成
分的含量。结果显示,DAS-21023-5 与其非转基因对照间就上述指标含量相近。
因此转基因棉花 DAS-21023-5 与其非转基因对照加工产品的一般成分方面等同。
表 33. 熟棉粕和精炼棉籽油的一般成分含量分析
组织 一般成分(%) 文献报导 a 非转基因
对照 DAS-21023-5
棉花
烤熟 棉粕
灰份 4.6~9.8d 5.9 6.1
总脂肪 0.6~4.7d 4.5 2.4
水份 NAc 2.2 9.1
蛋白质 43.0~52.4d 46.2 47
碳水化合物 NAc 41.2 35.4
热量(千卡/100 克) NAc 390 351
粗纤维(%) 8.4~15.3d 12.1 8.3
酸性去污剂处理后纤维% 12.2~23.9d 18.1 12.9
中性去污剂处理后纤维% 15.8~32.4d 23.7 17.1
精炼棉籽油
总脂肪 NAc 100.2 100.1
水份 NAc < 0.1 < 0.1
蛋白质 NAc < 0.1 < 0.1 a :OECD, 2009。b:棉籽饲料产品指导(National Cottonseed Products Association, 2002a)。c:未见文献报导。d:Forster and Calhoun, 1995。
熟棉粕中矿物质含量的分析
表 34 显示了熟棉粕中钙、铜、铁、镁、锰、钼、磷、钾、钠、硫和锌等矿
物质的含量。上述指标含量在转基因与非转基因对照棉花间相近,且与报导值相
近。因此,转基因品系和常规非转基因棉花品系的加工产品在矿物质的含量方面
实质等同。
表 34. 熟棉粕中矿物质的含量分析
组织 矿物质(mg/100g) 文献范围 a 非转因对照 DAS-21023-5 棉花
熟棉粕
钙 220 d 187 175
铜 1.25 d 1.4 1.5
铁 12.6 d 11 10.6
镁 660 d 614 639
锰 2.0 d 1.9 1.7
钼 0.25 d <0.2 <0.2
磷 1200 d 1130 1250
钾 1720 d 1500 1550
钠 140 d 15 10
锌 6.4 d 6.9 7
硫 440 d 433 428 a:OECD, 2009。b:Forster and Calhoun, 1995。c:未见文献报导。d:棉籽饲料产品指导(National Cottonseed Products Association, 2002a)。
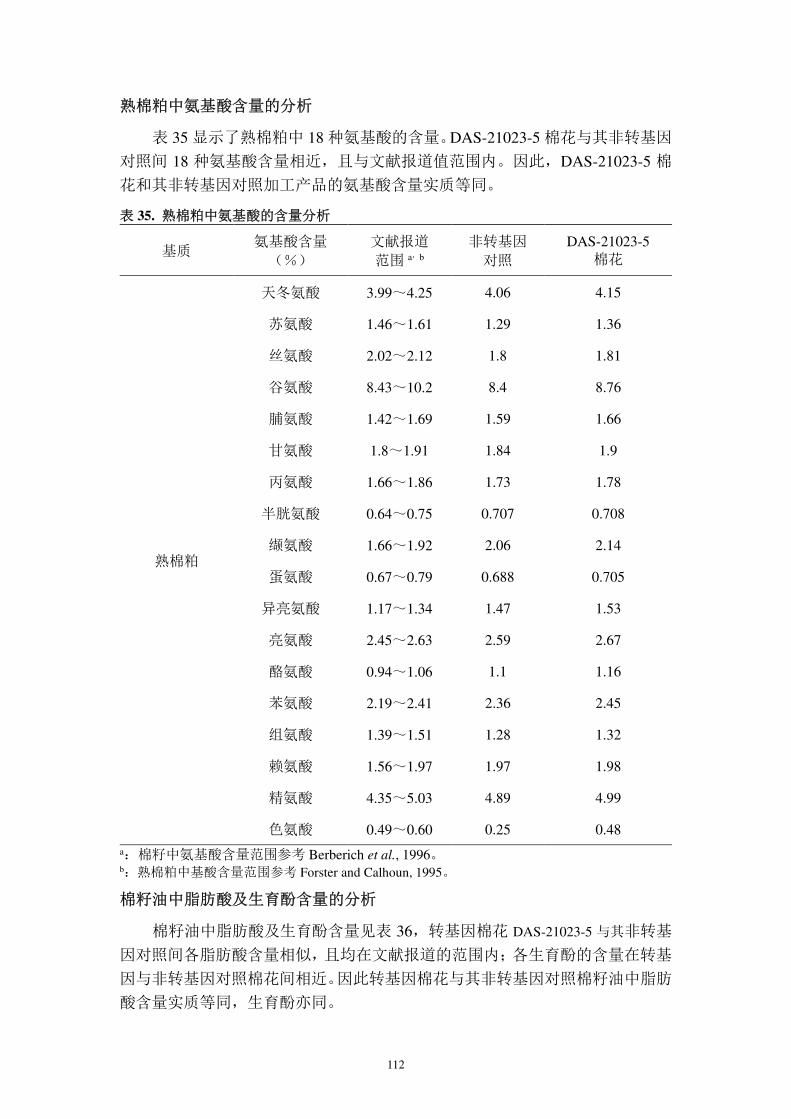
112
熟棉粕中氨基酸含量的分析
表 35 显示了熟棉粕中 18 种氨基酸的含量。DAS-21023-5 棉花与其非转基因
对照间 18 种氨基酸含量相近,且与文献报道值范围内。因此,DAS-21023-5 棉
花和其非转基因对照加工产品的氨基酸含量实质等同。
表 35. 熟棉粕中氨基酸的含量分析
基质 氨基酸含量
(%) 文献报道 范围 a,b
非转基因 对照
DAS-21023-5 棉花
熟棉粕
天冬氨酸 3.99~4.25 4.06 4.15
苏氨酸 1.46~1.61 1.29 1.36
丝氨酸 2.02~2.12 1.8 1.81
谷氨酸 8.43~10.2 8.4 8.76
脯氨酸 1.42~1.69 1.59 1.66
甘氨酸 1.8~1.91 1.84 1.9
丙氨酸 1.66~1.86 1.73 1.78
半胱氨酸 0.64~0.75 0.707 0.708
缬氨酸 1.66~1.92 2.06 2.14
蛋氨酸 0.67~0.79 0.688 0.705
异亮氨酸 1.17~1.34 1.47 1.53
亮氨酸 2.45~2.63 2.59 2.67
酪氨酸 0.94~1.06 1.1 1.16
苯氨酸 2.19~2.41 2.36 2.45
组氨酸 1.39~1.51 1.28 1.32
赖氨酸 1.56~1.97 1.97 1.98
精氨酸 4.35~5.03 4.89 4.99
色氨酸 0.49~0.60 0.25 0.48 a:棉籽中氨基酸含量范围参考 Berberich et al., 1996。 b:熟棉粕中基酸含量范围参考 Forster and Calhoun, 1995。
棉籽油中脂肪酸及生育酚含量的分析
棉籽油中脂肪酸及生育酚含量见表 36,转基因棉花 DAS-21023-5 与其非转基
因对照间各脂肪酸含量相似,且均在文献报道的范围内;各生育酚的含量在转基
因与非转基因对照棉花间相近。因此转基因棉花与其非转基因对照棉籽油中脂肪
酸含量实质等同,生育酚亦同。
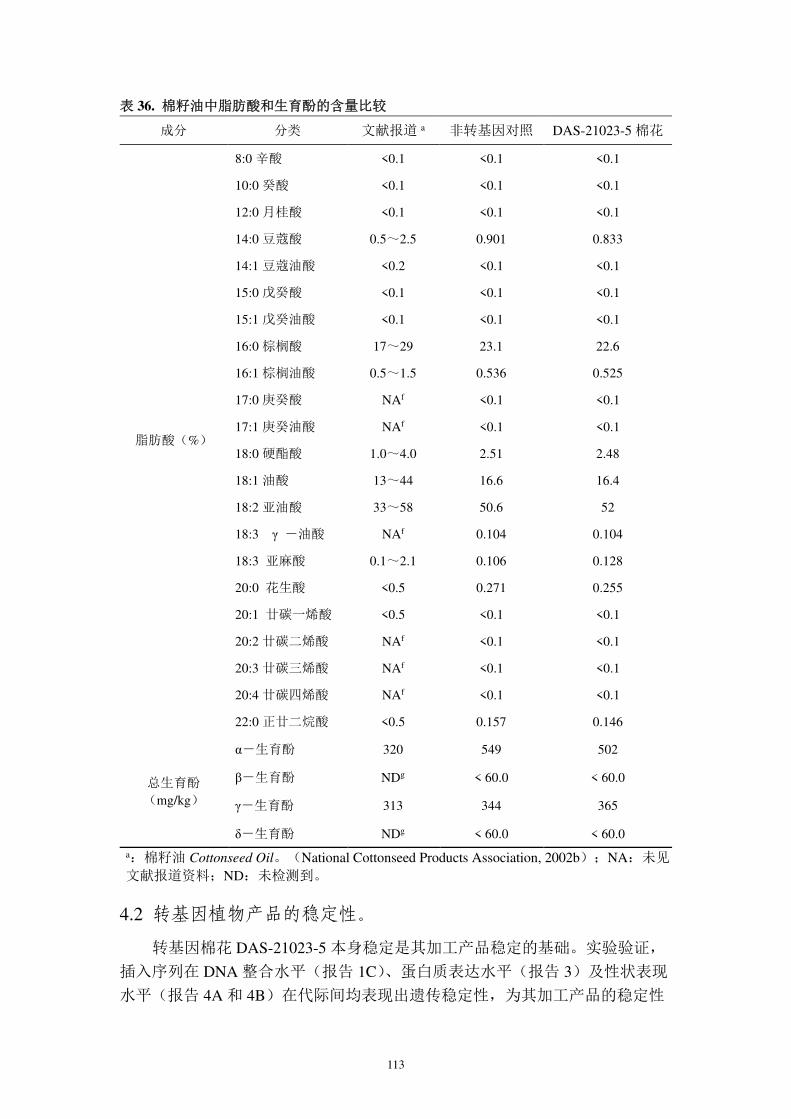
113
表 36. 棉籽油中脂肪酸和生育酚的含量比较
成分 分类 文献报道 a 非转基因对照 DAS-21023-5 棉花
脂肪酸(%)
8:0 辛酸 <0.1 <0.1 <0.1
10:0 癸酸 <0.1 <0.1 <0.1
12:0 月桂酸 <0.1 <0.1 <0.1
14:0 豆蔻酸 0.5~2.5 0.901 0.833
14:1 豆蔻油酸 <0.2 <0.1 <0.1
15:0 戊癸酸 <0.1 <0.1 <0.1
15:1 戊癸油酸 <0.1 <0.1 <0.1
16:0 棕榈酸 17~29 23.1 22.6
16:1 棕榈油酸 0.5~1.5 0.536 0.525
17:0 庚癸酸 NAf <0.1 <0.1
17:1 庚癸油酸 NAf <0.1 <0.1
18:0 硬酯酸 1.0~4.0 2.51 2.48
18:1 油酸 13~44 16.6 16.4
18:2 亚油酸 33~58 50.6 52
18:3 γ -油酸 NAf 0.104 0.104
18:3 亚麻酸 0.1~2.1 0.106 0.128
20:0 花生酸 <0.5 0.271 0.255
20:1 廿碳一烯酸 <0.5 <0.1 <0.1
20:2 廿碳二烯酸 NAf <0.1 <0.1
20:3 廿碳三烯酸 NAf <0.1 <0.1
20:4 廿碳四烯酸 NAf <0.1 <0.1
22:0 正廿二烷酸 <0.5 0.157 0.146
总生育酚 (mg/kg)
α-生育酚 320 549 502
β-生育酚 NDg < 60.0 < 60.0
γ-生育酚 313 344 365
δ-生育酚 NDg < 60.0 < 60.0
a:棉籽油 Cottonseed Oil。(National Cottonseed Products Association, 2002b);NA:未见
文献报道资料;ND:未检测到。
4.2 转基因植物产品的稳定性。
转基因棉花 DAS-21023-5 本身稳定是其加工产品稳定的基础。实验验证,
插入序列在 DNA 整合水平(报告 1C)、蛋白质表达水平(报告 3)及性状表现
水平(报告 4A 和 4B)在代际间均表现出遗传稳定性,为其加工产品的稳定性

114
奠定了基础。
DAS-21023-5 棉花与其非转基因对照之间的加工工艺相同,且测得加工产
品关键成分实质等同(报告 2)。
由于二者具有等同的物质基础与遗传稳定性,加工工艺相同,测得等同的
关键成分,因此 DAS-21023-5 产品具有与其非转基因对照相同的稳定性。
4.3 转基因植物产品与转基因植物在环境安全性方面的差异。
目前,DAS-21023-5 棉花产品包括用于种植栽培的种子和加工产品,与非转
基因传统棉花相比,唯一区别在于引入了表达 Cry1Ac 和选择标记 PAT 的基因,
从而使其抵抗鳞翅目害虫的危害。
首先,未加工的转基因棉花 DAS-21023-5 与其非转基因对照对环境的影响
等同。其生殖方式和生殖率、传播方式和传播能力、休眠期、适应性、生存竞争
能力、遗传物质向其他生物发生转移的可能性、杂草化潜能方面,DAS-21023-5
与其非转基因对照均实质等同(报告 5 和 6)。而外源表达蛋白质仅对特定靶标
害虫有作用(报告 4A 和 4B),对生态环境中的非靶标生物(包括对环境中有益
和有害生物)无不利的影响(报告 7A-7L)。因此,DAS-21023-5 棉花种子和非
转基因传统棉花种子在环境安全性方面是等同的。
其次, DAS-21023-5 棉花与其非转基因对照的加工产品的成分等同(报告
2),没有任何证据显示两者在环境安全性方面会有差异。
各种转 Bt 基因抗虫棉花已在生产上应用近二十年,至今尚未见 Bt 转基因植
物及其产品对环境造成不利影响的报导。另外,本申报书中 DAS-21023-5 棉花仅
在中国用于加工生产,而不是用于栽培种植。
4.4 转基因植物产品与转基因植物在对人类健康影响方面的差异。
DAS-21023-5 棉花中的外源蛋白质无毒性,也不会诱发人畜过敏(报告 8 和
报告 9);DAS-21023-5 棉花与其非转基因对照加工产品营养成分实质等同(报告
2)。因此,DAS-21023-5 与非转基因棉花的加工产品对人类健康影响无差异。
4.5 参照本办法第十四条有关标准划分转基因植物产品的安全等级。
DAS-21023-5 转基因加工产品对环境和人类健康方面无负面影响,依《农业
转基因生物安全评价管理办法》第二章第十四条,安全级别为 I 级。
5 供体生物的安全性评价
5.1 cry1Ac 基因的供体生物具有安全使用的历史

115
cry1Ac 最初从苏云金芽孢杆菌(Bacillus thuringiensis,Bt)中分离而来(
Cardineau et al., 2001; Payne and Sick, 1989)。Bt 菌是一类革兰氏阳性细菌,可
产生孢子,且在环境中普遍存在(Rogoff and Yousten, 1969)。人类和动物常常会
接触到 Bt 菌株产生的孢子和结晶蛋白。Cry 蛋白具有高度的特异性,不同 Cry 蛋
白的活性谱仅限于特定的昆虫目。由于其高度专一性,人们早在 20 世纪 20 年代
开始研究 Bt 微生物杀虫剂(Sanchis, 2011),并在 70 年代早期商业化,目前 Bt 微
生物杀虫剂已在世界各地广泛使用(Bauce et al., 2004; Bernhard, 1992)。自 1938
年法国将苏云金芽孢杆菌商业化以来(Kumar et al., 1996),至今以该菌作为活性
成分的注册农药产品已经超过 100 种(EPA, 2011)。并广泛应用于粮食、草料、
水果和蔬菜等多种作物。人们还分离了各种不同的 Bt 蛋白菌株,用于开发转基
因作物,从而使作物抵抗鳞翅目、鞘翅目和半翅目害虫(Betz et al., 2000; Chougule
and Bonning, 2012; Grover, 2011)。1960 年美国食品药品监督管理局(Food and
Drug Administration)对苏云金芽孢杆菌进行安全评估后,准予豁免其残留要求,
并于 1961 年首次在美国环保署(Environmental Protection Agency)注册为农药。
至今,含有苏云金芽孢杆菌活性成分的农药已有数十年的商业化安全应用历史,
至今未有此类微生物对人类、动物或植物有害的报道(Mendelsohn et al., 2003;
EPA, 2011; Sanahuja et al., 2011)。自 1996 年首例转基因作物商业化以来,尚未
发现在食品、饲料作物上因使用 Bt 微生物产品引起安全问题,亦未见因食用、
饲用含有Bt蛋白质的食品、饲料带来安全问题的报导(ISAAA, 2016; Pardo- López
et al., 2013)。20 多年来,含 Bt 蛋白作物的食品、饲料的商业化和广泛消费展现
了长期的安全使用历史,并增加了转基因作物和 Bt 蛋白质安全性的证据。
5.2 根癌土壤杆菌的安全性
pat 与 cry1Ac 的终止子 AtuORF25/26 3′UTR 以及 pat 的启动子 AtuMas 均
来自自常用的工程菌根癌土壤杆菌(Agrobacterium tumefaciens)。根癌土壤杆菌具
有较长时间的安全应用历史。
根癌土壤杆菌分类学地位为:细菌,根瘤菌科,土壤杆菌属。它广泛分布
于全球土壤和寄主植物中。
根癌土壤杆菌属于革兰氏阴性菌,无芽孢,短杆状,大小 0.6-1.0μm×1.5-
3.0μm,以 1-6 根周生或侧生鞭毛运动,好气性,代谢为呼吸型,最适生长温度
为 25-28℃,最适酸碱度为 pH6.0。菌落通常为圆形、隆起、光滑、白色至灰白
色,半透明。
根癌土壤杆菌寄主范围非常广,有几百个属。主要发生在土表下的根冠处,
也发生于侧根、支根及枝条上,在嫁接处也较为常见。形成大小不一的肿瘤,其
形状、大小质地因寄主不同而异。一般木本寄主的瘤大而硬,木质化;草本寄主
的瘤小而软,肉质。肿瘤初生时圆形、乳白色或略带红色,光滑、柔软。后渐变
为褐色至深褐色,球形或扁球形,木质化而坚硬,表面粗糙凹凸不平。发病植株
由于根部发生癌变,水分和养分的输送严重受阻,使地上部细瘦、叶薄、色黄,

116
严重时干枯死亡。(https://baike.baidu.com/item/根癌土壤杆菌/844336?fr=aladdin )
致瘤质粒(Tumor-inducing,Ti)是在根瘤土壤杆菌细胞中存在的一种染色
体外自主复制的环形双链 DNA 分子,长度约为 200-250 kb。Ti 质粒可以诱导植
物细胞产生不同种类的冠瘿碱,根据产生冠瘿碱的类型,把 Ti 质粒分成了 4 种
类型:章鱼碱型、胭脂碱(nopaline)型、农杆碱(Agropine)型和琥珀碱
(Succinamopine)型。从结构上看,Ti 质粒上有 T-DNA 区、毒性区(Vir 区)
和接合转移区和复制起始区 4 部分。基中 T-DNA 区和毒性区参与了外源基因的
转移和整合,T-DNA 上有与激素合成相关的基因,其表达产物能引起植物细胞
激素紊乱进而导致形成肿瘤。T-DNA 左右两端有 25 bp 重复序列,它们在 T-
DNA 转移及整合至植物基因组的过程中发挥功能。毒性区(Vir 区)处于 T-
DNA 之外,长度介于 30 kb~40 kb 之间。以胭脂碱型 Ti 质粒为例,其 Vir 区有
7 个操纵子,共 22 个基因,这些基因编码的 22 个 Vir 蛋白质在 T-DNA 转移及
整合过程中起辅助作用,其本身不会与植物基因组整合。以 Ti 质粒为基础,
开发了植物基因工程载体,其上剔除了致瘤基因。Ti 质粒是最常用的植物转化
载体。
5.3 绿色产色链霉菌的安全性
pat 基 因 的 供 体 生 物 是 土 壤 中 绿 色 产 色 链 霉 菌 ( Streptomyces
viridochromogenes),该微生物广泛存在于自然界中,不是已知的人、畜病原微生
物(Corinne Herouet, 2004)。链霉菌是全球生物圈中最普遍的组成部分,至今未
见该菌对人类,动物或植物有害的报导。转 pat 基因的作物已在全球商业化近二
十年,是全球范围内推广的两大除草剂之一,涉及的作物有油菜、玉米、棉花、
大豆和甜菜,约 34 个产品(http://cera-gmc.org/GMCropDatabase),自应用以来从
未对食品和饲料带来不良影响。含 pat基因的作物获得多个国家的审批,如美国、
加拿大、澳大利亚/新西兰、阿根廷、巴西、南非等(www.biotradestatus.com;EFSA,
2006, 2007a, 2007b, 2008)。
5.4 供体生物玉米的安全性
cry1Ac 的启 动子 ZmUbi1 来源于玉米(Zea mays L.),玉米有悠久的食用历
史。
栽培玉米 (Zea mays subsp. Mays)是世界上主要的粮食和饲料作物之一。
中国玉米一直保持自给有余,但近几年来,应市场需求旺盛,进口玉米量不断攀
升,2012 年度达 520 万吨,2013 年度达 500 万吨。
玉米是禾本科玉米(Zea)属仅有的一个种 (mays)。目前,mays 中又分为
9 个亚种。原产于拉丁美洲的墨西哥、危地玛拉和秘鲁一带,中国不是玉米的原
产地(崔俊明,2007;OECD,2003)。
玉米不是一种普通的致敏食物 (OECD,2002),但是玉米的花粉中具有与

117
猫尾草(Timonthy grass)花粉致敏原类似的蛋白质 (Petersen ,et al., 2006),
其结构接近于扩张蛋白(Expansin)。因此对猫尾草花粉过敏的患者对玉米花粉也
可能过敏。玉米籽粒中的脂质转移蛋白质(LTP,non-specific lipid transfer protein)、
原丝蛋白(profiline)、扩张蛋白和硫氧还蛋白(Thioredoxins)也会引起过敏
(Weichel, et al., 2006)。对玉米脂质转移蛋白质的致敏性研究表明,玉米脂质转
移蛋白质分子量大约为 9kD, 在加热状态下相对稳定 (Pastorello, et al., 2000;
2003)。现有的研究表明地中海地区的极少人群对玉米籽粒中的非特异性脂质转
移蛋白质过敏,但是在中欧和北欧人群中非常少见(Salcedo, et al., 2004)。
玉米属有性繁殖植物。雌雄同株但花器分离,属单性花,是典型的异花授粉
植物。其雄穗在始花后的第 2-4 天内开花最多,进入盛花期,一天中开花最多的
时间是上午 8-10 时,而此时花粉的生活力也最强;雄穗花期约 7-8 天;雌穗在始
花后的第 2-5 天,处于盛花期,柱头已基本抽齐;花丝生活力可持续 10-15 天;
在田间条件下花粉活力可维持 8-10 小时。玉米花粉主要借风传播异花受精产生
种子繁殖。玉米是 100%异花授粉作物,很容易发生串粉,所以玉米种内杂交,
自交非常普遍。玉米的近缘种绝大多数分布在墨西哥和中美洲地区。玉米和一年
生的大刍草之间存在很高的性亲合性。在墨西哥和危地玛拉地区,玉米和大刍草
若彼此接近便可能自由杂交,异交率可达每 500 株 F1 代玉米中有 1 株杂交种或
大刍草种群的 3-5%为杂交种。尽管种间杂交难度大,但摩擦禾属(Tripsacum)
各物种还是可以与玉米杂交,但是,其杂交种高度不育,且遗传不稳定 (OECD,
2003)。考虑到中国不是玉米的起源地,其与近缘种在自然条件下的异交几乎不
可能发生。此外,玉米不同的杂交品种和自交系之间的异交率取决于各种因素,
包括传媒昆虫的种类和数量、间隔距离、风速和风向、天气条件等。
在自然界中生存繁殖的能力低,越冬性,越夏性能力低。通过长期的人工
选育和栽培化的过程中,玉米获得了很多的农艺性状,同时也失去了在野地中
生存的能力。作为一年生作物,栽培玉米已没有具有杂草化倾向的特征。种子
成为唯一能够在野外生存的器官。玉米种子的生存取决于温度、种子含水量、
基因型、种皮保护物等,只有在合适的气候条件下才能越冬。种子只能在理想
的温度和湿度条件下才能发芽,土壤温度高于 37℃(Riley, et. al.,1981)或低于
10℃不利于种子发芽。玉米是喜光温作物,需要充作的水源。玉米在苗期有一
定抗旱能力,但是在抽穗和生殖生长阶段,对高温和干旱敏感。抽穗和生殖生
长阶段高温干旱会严重降低花粉的活力,影响籽粒的生长发育,进而降低玉米
的结实率。
玉米有稳定的传背景。玉米作为经典的细胞遗传学和数量遗传学的研究对
象由来已久。很多重大的科学发现和突破都是用玉米作研究材料完成的,例如转
座子的发现。采用现代分子生物学手段,也已对玉米进行了详细的研究。用玉米
自交系 B73 的玉米全基因组的测序也已完成。玉米是雌雄异花的开放性授粉植
物,其性状通过有性繁殖稳定地遗传至后代。虽然很久以前人类已开始种植玉米,

118
但是只是从开发授粉的混合群体中选择优良植株的籽粒作为种子。直到 20 世纪
初,G.H. Shull 进行了玉米果穗粒行数的遗传研究,先后于 1908 年和 1909 年发
表了揭示玉米自交导致衰退,杂交产生优势的现象,从而奠定了近代玉米杂交育
种方法基础,并提出了显性学说。自此,通过自花授粉产生纯和自交系,再用自
交系杂交产生杂交种子,是传统的玉米育种方法。而杂交玉米的推广应用显著地
提高了玉米的产量。这一切的基础在于玉米的遗传稳定性。
在自然条件下与其他植物种属进行遗传物质交换的可能性极低。玉米属有
性繁殖植物。因此,在自然条件下与其它植物种属进行遗传物质交换的话,必
然是通过花粉的有性杂交。有关杂交的先决条件是两个亲本必须互相性亲和。
而且这种杂交也依赖于许多环境因素,包括传媒昆虫的种类和数量,风速和风
向,两个亲本间的间隔距离,温度湿度等气候条件。玉米和同种的大刍草之间
存在很高的性亲和性,它们可以产生可育的杂交种。但是这些杂交种的适应性
差,从而防止了玉米基因向大刍草的高频率渗入(OECD,2003)。玉米与摩擦
禾属(Tripsacum)内的物种(T.dactyloides, T. floridanum, T. lanceolatum 和 T.
pilosum)可以杂交,但是难度大,且杂交后代高度不育。有关玉米和摩擦禾属
内物种的杂交研究很多,其主要目的为了将这些野生种的一些有益性状转移到
玉米中来 (OECD,2003)。除此之外,玉米在自然条件下与同其它植物种属
进行遗传物质交换的可能性几乎不存在。
在自然条件下与其它生物(例如微生物)进行遗传物质交换的可能性极
低。如上所述,玉米是有性繁殖植物,与其它植物物种之间进行遗传物质交换
的途径是通过花粉的有性杂交。如果玉米与其它生物(例如微生物)之间发生
遗传物质交换的话,需通过平基因转移(horizontal gene transfer)途径。横向基
因转移虽然是微生物进化的主要力量,但是有关高等植物基因向微生物的水平
转移还没有实例,也没有玉米与微生物通过横向基因转移发生遗传物质交换的
报道。

119
六、相关附件资料
附件 1. 目的基因和标记基因的核苷酸序列及其推导的氨基酸序列;
附件 2. 目的基因和标记基因与载体构建图谱;
附件 3. 目的基因与植物基因组整合及其表达的分子检测或鉴定结果
(PCR 检测、Southern 杂交分析、Northern 或 Western 分析结果、
目的基因产物表达结果);
3.1 DAS-21023-5 转化事件特异性定性 PCR(gel-based PCR)
3.2 DAS-21023-5 转化事件特异性定量 PCR(qPCR)
3.3 Southern blot 检测分子特征
3.4 外源蛋白质的表达量(ELISA)
附件 4. 转基因性状及产物的检测和鉴定技术;
4.1 转基因性状的检测和鉴定技术
4.2 对 DAS-21023-5 棉花中 Cry1Ac 进行定量的 ELISA 方法
4.3 对 DAS-21023-5 棉花中 PAT 进行定量的 ELISA 方法
附件 5. 转基因植物对生态环境安全性的综合评价报告;
附件 6. 食品安全性的综合评价报告;
附件 7. 该类转基因植物国内外生产应用概况;
附件 8. 田间监控方案,包括监控技术、抗性治理措施、长期环境效
应研究方法等;
附件 9. 审查所需的其它相关资料;
附件 10. 转基因生物在输出国家或地区已允许作为相应用途并投放
市场的证明文件;
附件 11. 输出国家或地区经科学试验证明对人类、动植物和生态环境
无害的资料;
附件 12. 转基因生物进口过程中拟采取的安全防范措施。

120
附件 1. 目的基因和标记基因的核苷酸序列及其推导的氨基酸序列
本转化体含目的基因 cry1Ac 和选择标记基因 pat。
目的基因 cry1Ac 的核苷酸序列如下(含终止密码子):
目的基因cry1Ac推导氨基酸序列如下:
标记基因 pat 的核苷酸序列如下(含终止密码子):
标记基因 pat 推导的氨基酸序列如下:
商业保密信息
商业保密信息
商业保密信息
商业保密信息

121
附件 2. 目的基因与载体构建的图谱
转化载体为质粒 pMYC3006(附图 1),全长 15,333 bp,其遗传元件详见附
表 1。质粒 pMYC3006 是通过多个步骤用分子生物学方法构建而成的,基本载体
骨架来源于大肠杆菌,大肠杆菌普遍存在于环境中。
质粒上的 T-DNA 长 8422 bp,其上含有 2 个完整的基因表达盒,即目的基因
cry1Ac 表达盒和标记基因 pat 表达盒,两个表达盒尾-尾相连共用一个双向终止
序列。cry1Ac 基因来自于自然界的土壤细菌苏云金芽孢杆菌,表达杀虫蛋白质
Cry1Ac;pat 基因来自于土壤细菌绿色产色链霉菌,表达膦丝菌素乙酰转移酶 PAT,
二者均有长期安全使用的历史。
骨架序列上有 3 个元件:1)来自于 RK2 质粒的复制起点序列 ori,来源于
革兰氏阴性细菌,2)来自于 RK2 质粒中 TrfA 蛋白质复制序列 trfA,来源于革兰
氏阴性细菌寄,3)抗红霉素的细菌基因 erythromycin,来源于 pAM-beta1 (Martin
et al., 1987)。
质粒 pMYC3006 详细遗传元件信息分析表明,其上无致病性的遗传元件和
DNA 序列,演变为有致病性的可能性极低。
附图 1. 转化载体 pMYC3006 的图谱(示各遗传元件)
AtuORF25/26 3’UTR 片
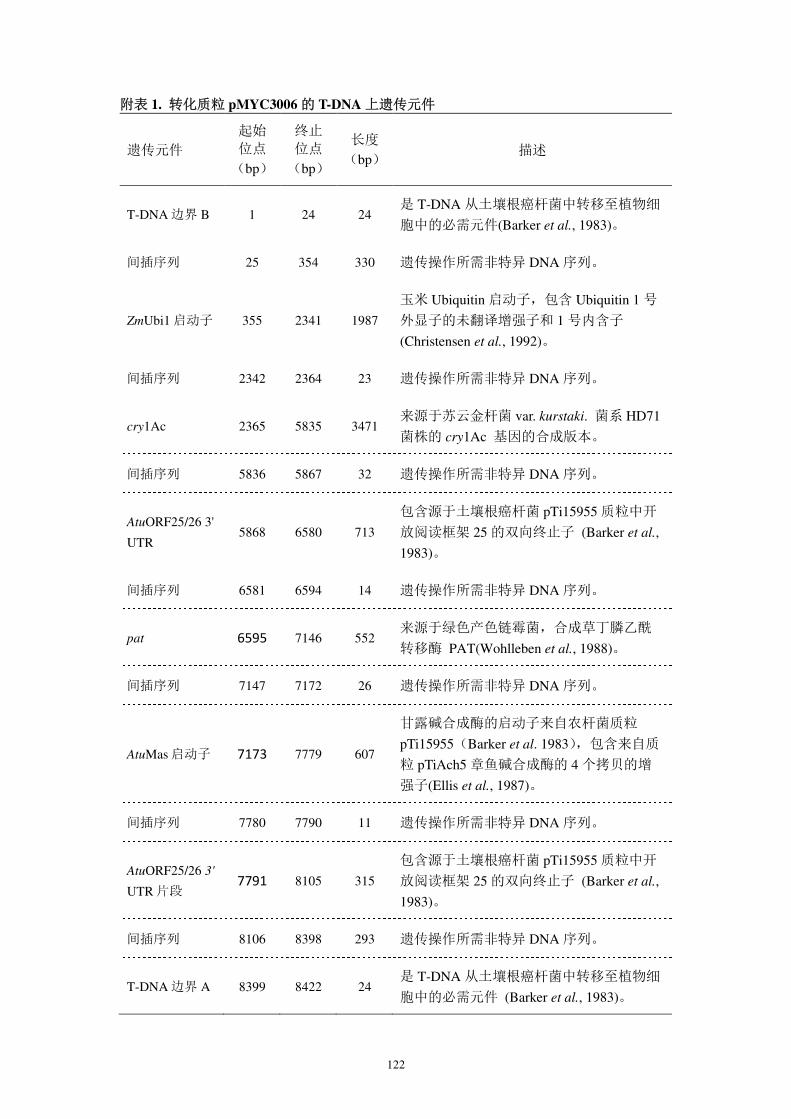
122
附表 1. 转化质粒 pMYC3006 的 T-DNA 上遗传元件
遗传元件
起始 位点
(bp)
终止 位点
(bp)
长度 (bp)
描述
T-DNA 边界 B 1 24 24 是 T-DNA 从土壤根癌杆菌中转移至植物细
胞中的必需元件(Barker et al., 1983)。
间插序列 25 354 330 遗传操作所需非特异 DNA 序列。
ZmUbi1 启动子 355 2341 1987
玉米 Ubiquitin 启动子,包含 Ubiquitin 1 号
外显子的未翻译增强子和 1 号内含子
(Christensen et al., 1992)。
间插序列 2342 2364 23 遗传操作所需非特异 DNA 序列。
cry1Ac 2365 5835 3471 来源于苏云金杆菌 var. kurstaki. 菌系 HD71
菌株的 cry1Ac 基因的合成版本。
间插序列 5836 5867 32 遗传操作所需非特异 DNA 序列。
AtuORF25/26 3'
UTR 5868 6580 713
包含源于土壤根癌杆菌 pTi15955 质粒中开
放阅读框架 25 的双向终止子 (Barker et al.,
1983)。
间插序列 6581 6594 14 遗传操作所需非特异 DNA 序列。
pat 6595 7146 552 来源于绿色产色链霉菌,合成草丁膦乙酰
转移酶 PAT(Wohlleben et al., 1988)。
间插序列 7147 7172 26 遗传操作所需非特异 DNA 序列。
AtuMas 启动子 7173 7779 607
甘露碱合成酶的启动子来自农杆菌质粒
pTi15955(Barker et al. 1983),包含来自质
粒 pTiAch5 章鱼碱合成酶的 4 个拷贝的增
强子(Ellis et al., 1987)。
间插序列 7780 7790 11 遗传操作所需非特异 DNA 序列。
AtuORF25/26 3'
UTR 片段 7791 8105 315
包含源于土壤根癌杆菌 pTi15955 质粒中开
放阅读框架 25 的双向终止子 (Barker et al.,
1983)。
间插序列 8106 8398 293 遗传操作所需非特异 DNA 序列。
T-DNA 边界 A 8399 8422 24 是 T-DNA 从土壤根癌杆菌中转移至植物细
胞中的必需元件 (Barker et al., 1983)。

123
附件 3. 目的基因与植物基因组整合及其表达的分子检测或鉴定结果
(PCR 检测、Southern 杂交分析、Northern 或 Western 分析结果、目
的基因产物表达结果)
3.1 DAS-21023-5 转化事件特异性定性凝胶 PCR 检测方法(报告 1G)
本凝胶 PCR 检测方法仅对棉花转化体 DAS-21023-5 的外源片段 5'端接合区
有效扩增,在其它物种、其它转化事件或非转基因棉花中均不扩增。本方法具
有较高的灵敏度,可检出低至 0.04%(W/W)的 DAS-21023-5 基因组 DNA。
3.1.1 材料
试样为转基因棉花 DAS-21023-5。非转基因对照为其回交受体 PSC355。
14 个参照株系为:转基因棉花 DAS-24236-5、DAS-81807-3 和 DAS-81910-
7;五个商品化的转基因棉花 GHB614(Bayer 公司)、LL25(Bayer 公司)、MON531
(Monsanto 公司)、Bollgard II(Monsanto 公司的 MON 15985-7)和 MON1445
(Monsanto 公司);三个非转基因棉花品种 PSC355、来自 Bayer 的品种、来自
Monsanto 的品种;三个非转基因的其他作物大豆品种 Maverick、玉米品种
SLB01/TCI30 和水稻品种 M202。附表 2 中列出了所有对照样品的详细信息。
附表 2. 受试材料和对照材料
物种及品系 描述
DAS-21023-5 棉花 受试材料
PSC355 棉花 非转基因对照
DAS-24236-5 棉花 陶氏益农内部研发转基因棉花
DAS-81807-3 棉花 转基因棉花,由陶氏益农公司研发
DAS-81910-7 棉花 转基因棉花,由陶氏益农公司研发
转基因棉花 GHB614 商业化转基因棉花,由 Bayer Crop Sciences 研发
非转基因棉花品系 商业化非转基因棉花,由 Bayer Crop Sciences 研发
棉花 LLCotton25 商业化转基因棉花,由 Bayer Crop Sciences 研发
非转基因棉花品系 商业化非转基因棉花,由 Monsanto 公司研发
MON 531 商业化转基因棉花,由 Monsanto 公司研发
Bollgard II 棉花 MON 15985-7 商业化转基因棉花,由 Monsanto 公司研发
MON1445 商业化转基因棉花,由 Monsanto 公司研发
Maverick 大豆 非转基因大豆
SLB01/TCI30 Corn 玉米 非转基因玉米
非转基因水稻 M202 商业化水稻品系 M202
3.1.2 方法
引物信息
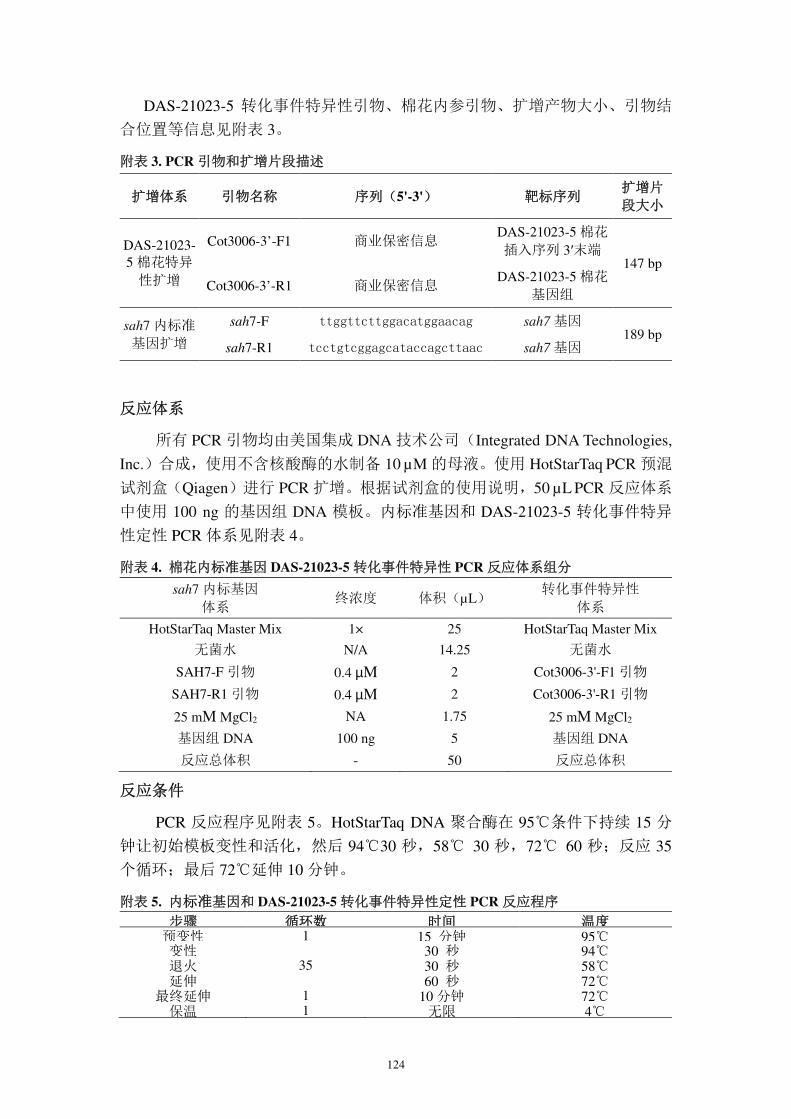
124
DAS-21023-5 转化事件特异性引物、棉花内参引物、扩增产物大小、引物结
合位置等信息见附表 3。
附表 3. PCR 引物和扩增片段描述
反应体系
所有 PCR 引物均由美国集成 DNA 技术公司(Integrated DNA Technologies,
Inc.)合成,使用不含核酸酶的水制备 10 µM 的母液。使用 HotStarTaq PCR 预混
试剂盒(Qiagen)进行 PCR 扩增。根据试剂盒的使用说明,50 µL PCR 反应体系
中使用 100 ng 的基因组 DNA 模板。内标准基因和 DAS-21023-5 转化事件特异
性定性 PCR 体系见附表 4。
附表 4. 棉花内标准基因 DAS-21023-5 转化事件特异性 PCR 反应体系组分
sah7 内标基因 体系
终浓度 体积(µL) 转化事件特异性
体系
HotStarTaq Master Mix 1× 25 HotStarTaq Master Mix
无菌水 N/A 14.25 无菌水
SAH7-F 引物 0.4 µM 2 Cot3006-3'-F1 引物
SAH7-R1 引物 0.4 µM 2 Cot3006-3'-R1 引物
25 mM MgCl2 NA 1.75 25 mM MgCl2
基因组 DNA 100 ng 5 基因组 DNA
反应总体积 - 50 反应总体积
反应条件
PCR 反应程序见附表 5。HotStarTaq DNA 聚合酶在 95℃条件下持续 15 分
钟让初始模板变性和活化,然后 94℃30 秒,58℃ 30 秒,72℃ 60 秒;反应 35
个循环;最后 72℃延伸 10 分钟。
附表 5. 内标准基因和 DAS-21023-5 转化事件特异性定性 PCR 反应程序
步骤 循环数 时间 温度 预变性 1 15 分钟 95℃ 变性 30 秒 94℃ 退火 35 30 秒 58℃ 延伸 60 秒 72℃
最终延伸 1 10 分钟 72℃ 保温 1 无限 4℃
扩增体系 引物名称 序列(5'-3') 靶标序列 扩增片
段大小
DAS-21023-5 棉花特异
性扩增
Cot3006-3’-F1 商业保密信息 DAS-21023-5 棉花
插入序列 3′末端 147 bp
Cot3006-3’-R1 商业保密信息 DAS-21023-5 棉花
基因组
sah7 内标准
基因扩增
sah7-F ttggttcttggacatggaacag sah7 基因 189 bp
sah7-R1 tcctgtcggagcataccagcttaac sah7 基因

125
显象观察
PCR 扩增产物在含有溴化乙锭的 1% 琼脂糖 1×TBE 凝胶中电泳分离,然后
在紫外灯下显影。
3.1.3 结果
特异性
受试株系 DAS-21023-5、非转基因对照及 14 个参照品系的扩增结果显示,
仅在棉花转化体 DAS-21023-5 中观察到了~147 bp 特异性片段(泳道 4),在非
转基因对照棉花、其它物种、其它转化事件或非转基因作物中均不扩增(附图 2)。
在棉花内参扩增体系中,基因 sah7 在所有棉花株系中均有效扩增,表明试验体
系有效。
因此,本扩增方法是 DAS-21023-5 转化事件特异性的。
A. 棉花物种特异性内标准基因 sah7 PCR 扩增
B. DAS-21023-5 转化事件特异性 PCR 扩增
附图 2. 棉花 DAS-21023-5 转化事件特异性 PCR 扩增—示特异性
1:分子量标准;2:无模板对照(H2O);3:非转基因对照 PSC355;4:转基因棉花 DAS-
21023-5 棉花;5:转基因棉花 DAS-24236-5;6:转基因棉花 DAS-81807-3;7:转基因棉
花 DAS-81910-7;8:转基因棉花 GHB614;9:Bayer Crop Sciences 的非转基因棉花。10:
转基因棉花 LL25 ;11:Monsanto 公司的非转基因棉花;12:转基因棉花 MON 531;13:
转基因棉花 Bollgard II;14:转基因棉花 MON1445;15:大豆 Maverick;16:玉米
SLB01/TCI30;17:粳稻 M202。

126
灵敏度
对不同含量(100 ng~0.0122 ng)的 DAS-21023-5 棉花 DNA 模板进行扩增。
在含有 12 pg 棉花基因组 DNA 的系体中,仍可观察到 147 bp 的转化事件特异性
条带(附图 3B)。且内参基因扩增体系中均可见 189 bp 的 sah7 基因扩增片段,
表明扩增体系有效(附图 3A)。本方法可检出低至 12 pg 的 DAS-21023-5 棉花
DNA。
A. 棉花内标准基因 sah7 PCR 扩增
B. DAS-21023-5 转化事件特异性 PCR 扩增
附图 3. 棉花 DAS-21023-5 转化事件特异性定性 PCR 扩增—示灵敏度
1:分子量标准;2 无模板对照(H2O);100 ng 非转基因对照 PSC355;3-17:100 ng、50
ng、25 ng、12.5 ng、6.25 ng、3.13 ng、1.56 ng、0.7 8 ng、0.39 ng、0.195 ng、0.098 ng、
0.05 ng、0.024 ng 和 0.012 ng DAS-21023-5 棉花基因组 DNA。
在以上实验的基础上确定本方法的灵敏度。即在非转基因棉花PSC355基因
组DNA中掺入0.04%的DAS-21023-5基因组DNA(即每100 ng DNA模板含40 pg
DAS-21023-5棉花基因组DNA),来确定该PCR检测方法的灵敏度。
附图4显示了对内标准基因的扩增结果,所有30个重复体系中均扩得了棉花
物种特异性的内标准基因sah7片段;附图5为DAS-21023-5棉花转化事件特异性扩
增图谱,显示30个重复体系中均有目标条带。在含0.04% DAS-21023-5基因组DNA
时,内标准基因体系和转化事件特异性体系均可有效扩增,因此,DAS-21023-5
转化事件特异的定性PCR检测方法的灵敏度是0.04%。

127
附图 4. 棉花内标准基因 sah7 扩增图谱-示灵敏度
1:低分子量标准;2:无模板对照(H2O);3:100 ng PSC355;4:100 ng DAS-21023-5 棉
花基因组 DNA;5~34:30 个含 0.04% DAS-21023-5 基因组 DNA 的反应体系,即 40 pg DAS-21023-5 棉花基因组 DNA 与 99.96 ng PSC355 棉花基因组 DNA 混合。
体系中含 0.04% DAS-21023-5 棉花基因组 DNA,30 个重复反应均扩增了棉花内标准基因
sah7 片段。
附图 5. DAS-21023-5 转化事件特异性扩增图谱-示灵敏度
1:低分子量标准;2:无模板对照(H2O);3:100 ng PSC355;4:100 ng DAS-21023-5 棉
花基因组 DNA;5~34:30 个含 0.04% DAS-21023-5 基因组 DNA 的反应体系,即 40 pg DAS-21023-5 棉花基因组 DNA 与 99.96 ng PSC355 棉花基因组 DNA 混合。
体系中含 0.04% DAS-21023-5 基因组 DNA,30 个重复反应均扩增了 DAS-21023-5 转化事件
特异序列。
3.1.4 小结

128
DAS-21023-5 转化事件特异性定性 PCR 检测方法具有良好的特异性,仅可
对转化事件 DAS-21023-5 有效扩增;且具有较高的灵敏度,可检出低至 0.04%
(W/W)即 17 个拷贝的 DAS-21023-5 基因组 DNA。
本方法可在 3 个实验室重复验证,由于篇幅限制未在此列举,详见报告 1G
《DAS-21023-5 转化事件特异性定性 PCR (gel-based PCR)》。

129
3.2 DAS-21023-5 转化事件特异性定量 PCR 检测方法(报告 1H)
采用两步法 TaqMan 荧光定量 PCR 扩增方法,以非转基因棉花、其它转基
因棉花、其它物种为参照株系,以 sah7 为内标准基因,建立了 DAS-21023-5 转
化事件特异性定量体系:1)本体系仅对棉花转化事件 DAS-21023-5 有效扩增。
2)LOD 为 0.02%,即可以检测低至 8.6 个拷贝的外源基因。3)LOQ 为 0.08%,
即能对低至 36 个拷贝的 DAS-21023-5 棉花转化事件进行定量。
本方法在另外两个实验室循环验证,实验报告见第四册报告 1H。
3.2.1 材料和方法
材料
试验材料为转基因棉花DAS-21023-5。
阴性对照及参照品系包括棉花其它品系(转基因和非转基因)和其它物种的
作物(转基因和非转基因),详见附表 11。
棉花物种特异性内标基因 sah7 扩增体系
本方法采用棉花物种特异性内源基因sah7。sah7基因不仅存在于陆地棉
(Gossypium hirsutum L.)的A染色体组的第8染色体,而且还位于D染色体组的
第20染色体上。位于A染色体组的sah7拷贝与D染色体组上的序列有一些差异,
包括单个或两个核苷酸的替换,微小删除和插入序列。本体系中,采用一套引
物和探针对A染色体组和D染色体组上的sah7进行扩增,扩得的产物大小稍有不
同,扩增片段分别为115 bp和123 bp (Sara Jacchia, et al., 2018)。本实验所使用的
sah7 PCR体系已经通过欧盟转基因作物检测机构验证,参见http://gmo-
crl.jrc.ec.europa.eu /summaries/356043_validated_Method.pdf。
引物序列:
SAH7-uni-f1 5′-AGT TTG TAG GTT TTG ATG TTA CAT TGA G-3′ 28-mer SAH7-uni-r1 5′-GCA TCT TTG AAC CGC CTA CTG-3′ 21-mer
探针序列: SAH7-uni-s1 5′-6FAM-AAA CAT AAA ATA ATG GGA ACA ACC ATG ACA
TGT-TAMRA-3′ 33-mer
扩增片段序列和大小:
扩增片段序列长度 A 染色体亚组为 115 bp,D 染色体亚组为 123 bp(单下划
线代表引物,双下划线代表探针): A 染色体亚组AGTTTGTAGGTTTTGATGTTACATTGAGTGACAGTGAATGAAAGGGTGTGTAAACAT
AAAATAATGGGAACAACCATGACATGTTGGACTGGATCAGTAGGCGGTTCAAAGATG
C
130
D 染色体亚组AGTTTGTAGGTTTTGATGTTACATTGAGTTAAAGTGAATGATAGGGTGTCTAAACAT
AAAATAATGGGAACAACCATGACATGTTGAACTGGAGGTCAAAATCAGTAGGCGGTT
CAAAGATGC
DAS-21023-5 转化事件特异性扩增体系
引物序列: 3006-f3 5′-商业保密信息-3′ 28-mer 3006-r2 5′-商业保密信息-3′ 26-mer
探针序列: 3006-s2(rev) 5′-6FAM -商业保密信息-TAMRA-3′
30-mer
扩增片段序列和大小:
扩增片段序列长度为 90 bp(单下划线代表引物,双下划线代表探针):
反应体系
棉花内标准基因定量 PCR 反应体系见附表 6,棉花 DAS-21023-5 转化事件
特异的定量 PCR 反应体系见附表 7,二者均采用 25 µL 反应体系。
附表 6. 棉花内标准基因定量 PCR 反应体系
化学试剂 母液浓度 最终浓度 体积(μL)
TaqMan® Universal PCR Mix, No AmpErase® UNG 2 × 1 × 12.50
SAH7-uni-f1 10 µM 350 nM 0.88
SAH7-uni-r1 10 µM 250 nM 0.63
SAH7-uni-s1 10 µM 175 nM 0.44
灭菌水 5.56
基因组 DNA 模板 20 ng/µL 5.00
总体积 25.00
附表 7. DAS-21023-5 棉花转化事件特异的定量 PCR 反应体系
化学试剂 母液浓度 最终浓度 体积(μL)
TaqMan® Universal PCR Mix, No AmpErase® UNG 2 × 1 × 12.50
3006-f3 10 µM 400 nM 1.00
3006-r2 10 µM 400 nM 1.00
3006-s2(rev):FAM:TAMRA 10 µM 150 nM 0.38
无菌水 5.12
基因组 DNA 模板 20 ng/µL 5.00
总体积 25.00
商业保密信息
131
反应程序
采用标准两步法 TaqMan 荧光定量 PCR 扩增,内标准体系和 DAS-21023-5
转化事件特异性扩增体系采用相同的反应程序,反应循环条件见附表 8。
附表 8. 内标准基因和转化事件特异性定量扩增反应循环条件
步骤 阶段 温度
(°C) 时间
(秒) 数据收集 循环数
1 初始酶活性激活 95 600 No 1
2 扩增
变性 95 15 No 40
3 退火 & 延伸 60 60 Yes
注:所用设备为 ABI7500 Real-Time PCR System,具有热盖设备;反应在 96 孔板中进行。
标准样品制备
以 10%的比例将转基因棉花 DAS-21023-5 与非转基因棉花 DNA 混合,达到
棉花总 DNA 浓度为 20 ng/μL(每个反应体系含 5 μL,共 100 ng 基因组 DNA),
再依次以 1:5 梯度稀释 3 轮,最终 4 个标准点 1、2、3 和 4 的反应体系中转化体
DNA 含量分别为 10 ng、2 ng、0.4 ng 和 0.08 ng(附表 9)。确保定量极限 LOQ
在标准点之内。
附表 9. 用于绘制标准曲线的样品浓度梯度(calibration samples)
试样 标准浓度 1 标准浓度 2 标准浓度 3 标准浓度 4
PCR 体系中的棉花 DNA 的总量 100 ng 20 ng 4 ng 0.8 ng
PCR 体系中 DAS-21023-5 DNA 的量 10 ng 2 ng 0.4 ng 0.08 ng
参照样品制备方法
鉴定检测极限和定量极限时,DAS-21023-5 棉花 DNA 浓度分别为 5.0%、
2.0%、0.90%、0.50%、0.08%、0.04%和 0.02%(附表 10)。棉花总 DNA 浓度为
20 ng/μL,每个反应含 100 ng。
附表 10. 用于鉴定检测极限与定量极限的已知样品浓度梯度
试样 5.00% 2.00% 0.90% 0.50% 0.08% 0.04% 0.02%
棉花 DNA 的总量 100 ng 100 ng 100 ng 100 ng 100 ng 100 ng 100 ng
DAS-21023-5 DNA 的量 5.0 ng 2.0 ng 0.90 ng 0.50 ng 0.08 ng 0.04 ng 0.02 ng
标准曲线绘制
设定荧光阈值(threshold):组成标准曲线的阈值应该取自于所有扩增曲线
的指数扩增区段。这一区段在转换成对数后是公认的线形区域。不应该将
阈值线取自于扩增平台期或者扩增的初始期。确认阈值线应该位于背景荧

132
光线以上,并且在不同标准样品间扩增曲线明显分离或叉形效应的水平之
上。
将基线(baseline)设置在“Auto baseline”。
保存所有的设置。
以同样的程序设置另一个 PCR 体系的荧光阈值和基线。
保存设置,并将数据作为微软 Excel 文件输出以备后续计算。
棉花内标准基因体系和 DAS-21023-5 转化体标准曲线的绘制:根据标准
样品平均 Ct 值与其相应的靶标 DNA 含量的对数值绘制而成。
定量计算
用已知 DAS-21023-5 棉花基因组 DNA 含量混入已知非转基因棉花基因组
DNA 中,并梯度稀释得到标准浓度样品系列。根据棉花内标准基因(sah7)体系
和 DAS-21023-5 转化事件特异性反应体系的实验所得值与其相应的标准浓度含
量(总 DNA 或 DAS-21023-5 转化事件 DNA)的对数值分别绘制棉花内标准基
因(sah7)体系和 DAS-21023-5 转化事件特异性的标准曲线。棉花内标准基因
sah7 标准曲线实际上相当于总棉花基因组 DNA 标准曲线,与内参基因 sah7 拷
贝数无关。未知样品的转基因 DNA 绝对含量通过与 DAS-21023-5 转化事件特异
性的标准曲线的比对而推算,其相对含量(%)= [DAS-21023-5 含量平均值/总棉
花基因组 DNA (sah7)含量平均值] ×100 推算得出,所以各表中的转基因含量值
与内标准基因是否为双拷贝并无关系。因此两个拷贝的内源基因(sah7)对本检
测方法的浓度和拷贝数计算没有影响。内源基因仅作为一种棉花检测方法以及总
DNA 含量检测上的参考,只要此方法可以稳定地定量反应总棉花基因组 DNA 含
量,结果与内参基因拷贝数无关。测试样品的实测 Ct 值须在标准曲线所涵盖的
Ct值范围之内。另,2.33 pg棉花DNA=1个基因组拷贝(Arumuganathan and Earle,
1991)。
质量控制、数据分析和结果评估
a) 标准曲线扩增效率接受标准
标准曲线的平均斜率≥-3.6 且≤-3.1 时接受评估结果。
标准曲线的决定系数 R2≥0.98 时,接受评估结果。
检测极限对照有典型扩增曲线,且 Ct 值≤38。
所有阴性对照样品在循环结束前均无典型扩增曲线。
b) LOQ 准确度及精确度接受标准
定量偏差 Bias≤25%。Bias 反映了定量的准确度。
RSDr≤25%。RSDr(Relative Standard Deviation of repeatability)为实验室内平均值
的相对标准差,反映了实验室内定量的重复性的精确度。
RSDR<35%。RSDR(Relative Standard Deviation of reproducibility)为实验室间平均
值的相对标准差,反映了实验室间定量再现性的精确度。

133
检测目的基因和内标准基因时,进行2次或3次重复。因此可以得到2个或3个
转化体/内标基因拷贝数的结果,计算转化体含量平均值。
虽然尚无确切的公式来计算随机变量比率的平均值和方差,但是可以获得近
似值。E值代表平均值,独立随机变量比率的方差可以通过下面的公式计算近似
值:
公式中 �̅是转基因目的DNA片段拷贝数的算术平均值,是内标准基因DNA
拷贝数的算术平均值。
上 述 近 似 值 以 假 定 在 X 和 Y 之 间 无 相 互 关 联 为 前 提 , 由 公 式
sd[X/Y]=SQRT[Var(X/Y)]计算得到标准差,再通过标准差计算相对标准差RSDr
(Relative repeatability standard deviation)。
计算方法参见欧盟转基因作物检测机构 EUR 24790 EN-2011 指南附录 4。
c) 无模板对照 (NTCs)
所有的无模板对照均须显示阴性结果(即Ct值≥40时仍未出现典型扩增曲
线)。
3.2.2 结果
特异性
用已经获准商业化种植的转基因棉花和其它转基因作物的基因组 DNA 为模
板,测定了 DAS-21023-5 特异性定量 PCR 的特异性。结果表明,本体系仅对 DAS-
21023-5 棉花 DNA 扩增,在阴性对照、其它的转基因棉花品系或者其它转基因
作物及非转基因作物中均不扩增(附表 11),表明该体系仅对 DAS-21023-5 具有
特异性。
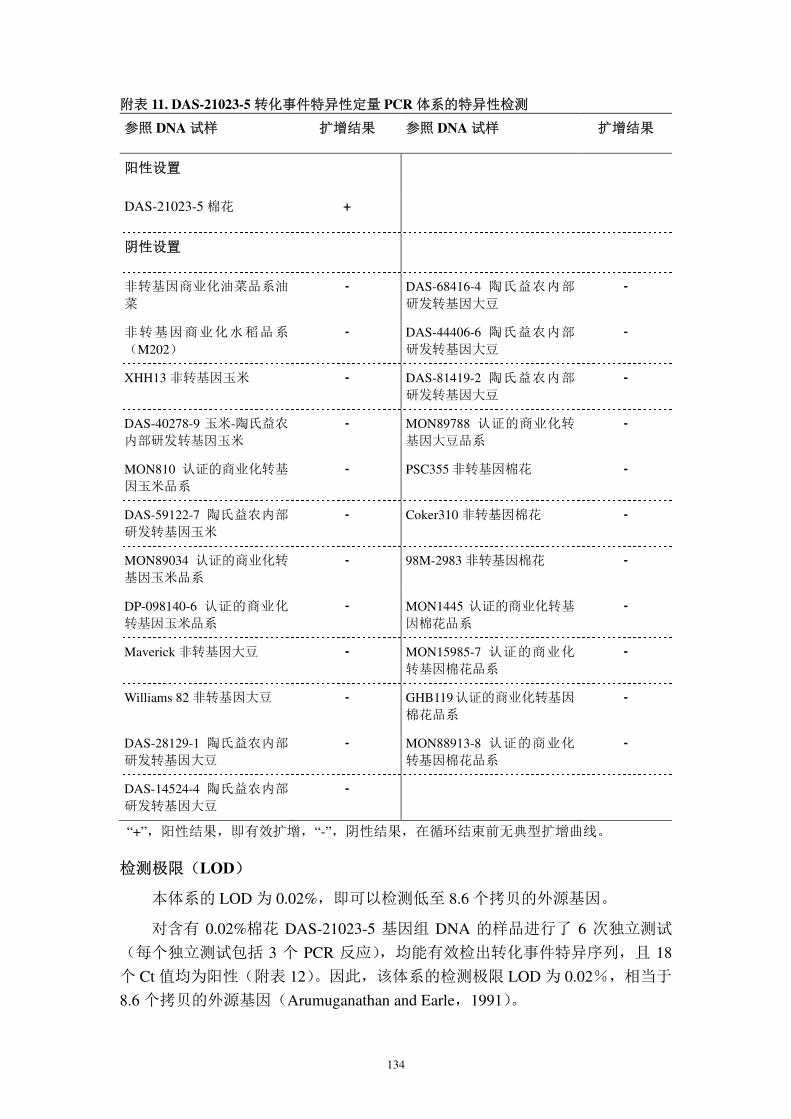
134
附表 11. DAS-21023-5 转化事件特异性定量 PCR 体系的特异性检测
参照 DNA 试样 扩增结果 参照 DNA 试样 扩增结果
阳性设置
DAS-21023-5 棉花 +
阴性设置
非转基因商业化油菜品系油
菜 - DAS-68416-4 陶氏益农内部
研发转基因大豆 -
非转基因商业化水稻品系
(M202) - DAS-44406-6 陶氏益农内部
研发转基因大豆 -
XHH13 非转基因玉米 - DAS-81419-2 陶氏益农内部
研发转基因大豆 -
DAS-40278-9 玉米-陶氏益农
内部研发转基因玉米 - MON89788 认证的商业化转
基因大豆品系 -
MON810 认证的商业化转基
因玉米品系 - PSC355 非转基因棉花 -
DAS-59122-7 陶氏益农内部
研发转基因玉米 - Coker310 非转基因棉花 -
MON89034 认证的商业化转
基因玉米品系 - 98M-2983 非转基因棉花 -
DP-098140-6 认证的商业化
转基因玉米品系 - MON1445 认证的商业化转基
因棉花品系 -
Maverick 非转基因大豆 - MON15985-7 认证的商业化
转基因棉花品系 -
Williams 82 非转基因大豆 - GHB119认证的商业化转基因
棉花品系 -
DAS-28129-1 陶氏益农内部
研发转基因大豆 - MON88913-8 认证的商业化
转基因棉花品系 -
DAS-14524-4 陶氏益农内部
研发转基因大豆 -
“+”,阳性结果,即有效扩增,“-”,阴性结果,在循环结束前无典型扩增曲线。
检测极限(LOD)
本体系的 LOD 为 0.02%,即可以检测低至 8.6 个拷贝的外源基因。
对含有 0.02%棉花 DAS-21023-5 基因组 DNA 的样品进行了 6 次独立测试
(每个独立测试包括 3 个 PCR 反应),均能有效检出转化事件特异序列,且 18
个 Ct 值均为阳性(附表 12)。因此,该体系的检测极限 LOD 为 0.02%,相当于
8.6 个拷贝的外源基因(Arumuganathan and Earle,1991)。
135
附表 12. 含 0.02% DAS-21023-5 棉花 DNA 的 6 次独立测试的 Ct 值-实验室 1
GM 浓度 反应
结果 1 2 3 4 5 6
0.02% (LOD)
37.20 37.75 37.44 37.16 37.67 37.56 18 个 PCR 反应
结果均为阳性 37.49 37.73 37.07 37.78 37.16 37.44
37.11 37.74 37.70 37.56 37.24 36.46
LOQ及定量的准确度和精确度
定量极限指在可接受的精确度和准确度下,能够定量检出的样品中目标成份
的最低量浓度(或最低量)。本体系的定量极限为 0.08%,即可对低至 36 个拷贝
的转化事件进行定量。
附表 13 显示了对 DAS-21023-5 棉花 5 个浓度(5.00%~0.08%)的参照样品
进行测量的平均值(Mean)、定量偏差(预期值与实测值的相对偏差 Bias)、标准
差(STDEV)和实验室内相对标准差(RSDr)。浓度为 0.08%的 DAS-21023-5 棉
花 DNA 的样品测试结果显示,检测平均值为 0.069%, |Bias|=13.24%,
RSDr=24.73%,因此在可接受的精确度和准确度下(Bias≤25%,RSDr≤25%),DAS-
21023-5 特异性定量 PCR 体系的定量极限为 0.08%。
附表13. DAS-21023-5转化事件6次独立试验的定量极限
转化事件 已知浓度
%
DAS-21023-5 转化事件浓度实测值% Mean Bias STDEV RSDr
扩增 1 扩增 2 扩增 3 扩增 4 扩增 5 扩增 6
5.00 6.31% 5.95% 4.72% 4.89% 4.10% 4.21%
4.88% -2.50% 0.77 15.87% 5.76% 5.24% 4.70% 4.30% 4.07% 4.25%
2.00 1.93% 2.63% 1.94% 1.93% 1.85% 1.99%
2.06% 3.02% 0.24 11.52% 1.94% 2.43% 1.98% 2.14% 1.89% 2.06%
0.90 0.65% 0.86% 0.73% 0.65% 0.71% 0.85%
0.72% -19.71% 0.10 13.32% 0.62% 0.86% 0.65% 0.63% 0.65% 0.80%
0.50 0.40% 0.57% 0.39% 0.34% 0.42% 0.54%
0.45% -9.76% 0.080 17.75% 0.45% 0.56% 0.44% 0.33% 0.46% 0.51%
0.08
(LOQ)
0.065% 0.086% 0.064% 0.053% 0.064% 0.10%
0.069% -13.24% 0.017 24.73%
0.068% 0.092% 0.059% 0.042% 0.058% 0.081%
注:扩增1-扩增6表示在不同的PCR板上扩增,分2次独立的测试,表中输出数据(run1-run6)均为3次PCR反应平均值。Bias为定量误差,指示定量的准确性;RSDr为实验室内重复性参
数,代表精确度。

136
扩增效率
附表 14 显示了本实验室的 PCR 体系标准曲线数据,从表中可以看出无论
棉花内标准基因还是 DAS-21023-5 转化事件特异的 PCR 标准曲线,其斜率均满
足≥-3.6 且≤-3.1,且决定系数 R2>0.98,故本实验室的标准曲线数据均符合可
接收的标准。
附表 14. PCR 扩增体系标准曲线数据
扩增体系 斜率 R2 扩增效率
棉花内体系
-3.41 0.996 0.96 -3.52 0.993 0.92 -3.39 0.998 0.97 -3.35 0.999 0.99 -3.32 0.995 1.00 -3.40 0.995 0.97
DAS-21023-5 转化 事件特异性体系
-3.58 0.998 0.90 -3.65 1.000 0.88 -3.26 0.997 1.03 -3.23 0.992 1.04 -3.31 0.996 1.00 -3.48 0.998 0.94
3.2.3 小结
1)本体系仅对棉花转化事件DAS-21023-5有效扩增。
2)本方法的LOD为0.02%,能检测出低至8.6个拷贝的DAS-21023-5棉花转化
事件。
3)本方法的LOQ 为0.08%,能对低至36个拷贝DAS-21023-5棉花转化事件进
行定量。
另,本方法可在 3 个实验室重复验证,由于篇幅限制未列举,详见报告 1H
《DAS-21023-5 转化事件特异性定量 PCR (qPCR)》。

137
3.3 Southern blot 检测分子特征(分析外源基因拷贝数、完整性、转化体上载
体骨架残留情况)
Southern blot 显示 DAS-21023-5 棉花的外源 DNA 片段中仅含单一插入位点
的、单拷贝的、结构完整的 cry1Ac 和 pat 基因表达盒,该转化体基因组中未整
合载体骨架序列,且在 BC1F3、BC3F3、BC3F4、BC4F1 四代间稳定整合遗传。
详见上文“2.2.5 插入序列的拷贝数”部分,或详见报告 1E。
3.4 ELISA 分析测量插入序列所表达的 Cry1Ac 和 PAT 蛋白质量
2001 年经美国农业部的批准在美国主要棉花产区的六个试验地点种植了
DAS-21023-5 棉花,在不同时期取不同组织采用 ELISA 方法定量检测 Cry1Ac 和
PAT 蛋白质的表达量(报告 2)。同时,也测定了棉籽和棉籽加工产品(包括棉
仁、外壳、棉粕和棉籽油)中外源蛋白质的含量。
Cry1Ac 蛋白质表达量:在可食部位棉籽中 0.57 ng/mg,在蜜腺中检测不到
Cry1Ac 蛋白质;在幼叶和花组织中含量最高,总平均值为 1.92 ng/mg 组织干重;
其他组织器官(顶端叶、棉蕾、棉铃、花粉、棉籽)中的均低于 1.84 ng/mg 组织
干重,在根中低于 0.2 ng/mg 组织干重。
PAT 蛋白质表达量:PAT 蛋白质在 DAS-21023-5 各组织器官中表达量很低
或检测不到。在可食部位棉籽中 0.06 ng/mg(低于定量极限),全株(收获前)
表达为 0.11 ng/mg。其他组织器官(花、棉蕾、棉铃、开花授粉期的全株)几乎
检测不到。
加工产品中外源蛋白质含量:将田间试验收获的 DAS-21023-5 棉籽进行常
规加工处理,得到棉仁、棉籽壳、熟棉粕和精炼棉籽油,采用 ELISA 方法对其
中的 Cry1Ac 和 PAT 蛋白质时行定量。结果显示,Cry1Ac 和 PAT 蛋白质在棉籽
加工后的棉仁和棉籽壳中很低,而在熟棉粕和精炼棉籽油中则检测不到了。
详见上文“2.6.2 插入序列的表达量及其分析方法”部分,或详见报告 2。

138
附件 4. 转基因性状及产物的检测和鉴定技术
本节提供了转基因棉花 DAS-21023-5 对靶标昆虫抗性的鉴定技术和外源蛋
白质的检测技术。
4.1 转基因性状的检测和鉴定技术
DAS-21023-5 棉花的抗虫性状,即对靶标昆虫的控制效果是通过田间试验和
实验室生物测定来鉴定的。检测结果如本申请第 3.2.8 部分所描述, 转基因抗虫
棉花 DAS-21023-5 可有效控制鳞翅目害虫,对以叶片为食的害虫种类(如甜菜夜
蛾、草地贪夜蛾、粉纹夜蛾、大豆尺蠖、南方粘虫等)有较优的抗虫效果(报告
4A 和 4B)。

139
4.2 ELISA 定量检测转基因棉花中 Cry1Ac 的试验方法
适用范围:本方法适用于从转 cry1Ac 和 cry1F 基因抗虫棉中提取并检测
Cry1Ac 的含量。
原理:本 ELISA 体系为“三明治式”ELISA,结合显色酶底物来定量检测棉
花样品中提取的 Cry1Ac 蛋白。以特异性的单克隆抗体包被在 96 孔滴定板上,
用另一个与辣根过氧化酶偶联的单克隆抗体作为检测抗体。棉花的提取液在特异
性的抗体包被的 96 孔滴定板孔中与辣根过氧化酶偶联的抗体共同孵育,如果棉
花的提取液中含有 Cry1Ac 蛋白,孔中包被的抗体和辣根过氧化酶标记的抗体将
与其结合。经洗涤后去除游离的多余抗体。这样,包被抗体─Cry1Ac 蛋白─检测
抗体在孔中结合在一起形成“三明治式”复合物。最后,经洗涤后加入辣根过氧
化酶的显色底物,显色后加反应终止液并用酶标仪读取吸光值。显色的强度与孔
中加入的 Cry1Ac 蛋白质的量是成正相关,棉花提取液中 Cry1Ac 蛋白质的浓度
可由已知浓度的标准品蛋白质建立的标准曲线来推算。
4.2.1 一般要求
1)材料
1/8" 号铬钢株,货号 BS-0125-C,购自 Small Parts Inc.公司;
2.0 mL 锥管,货号 02-681-361, Fisher Scientific 或类似公司产品;
保温设备,精度高,货号 51221087,Jouan, Inc 或类似公司产品;
12 头移液器(排枪),10~300 µL;
不同大小的移液管;
反应板盖或类似产品;
试剂 reservoirs,不要求无菌;
单头移液器,10 µL~1.0 mL;
5 mL 聚丙烯试管;
15 mL 聚丙烯离心管,带盖;
2.0 mL 锥型小型离心管,货号 02-681-344,购自 Fisher Scientific 公司;
96 孔滴定板,货号 35-3918,BD Falcon 或类似公司产品。
2) 设备
离心机,能放 2 mL Eppendorf 离心管;
-20℃低温冰箱;
酶标仪,可读取 450 nm波长处的吸光度,型号 0200-2018,Molecular Devices;
2~8℃冰箱;
涡旋仪,货号 12-812, Fisher Scientific 公司;
振荡研磨仪,型号 2000-115,Certiprep 公司,New Jersey;
96 孔板洗板仪,Model Elx 405,Bio-Tek Instruments, Inc.公司。

140
4.2.2 试剂
1)Cry1Ac 微量滴定板 ELISA 试剂盒,货号 7140220,购自 Strategic Diagnostics,
Inc.公司。 Newark, DE 19702。于 2~8℃保存。试剂盒中有:
抗体包被的 96 孔滴定板;
酶标记的 Cry1Ac 抗体溶液;
Cry1Ac 抗体偶联物 (浓缩液) ;
显色底物(120 mL);
反应终止液溶液(120 mL);
提取缓冲液(PBS Tween, Sigma P-3563)。
2)PBST 缓冲液,pH 7.4,货号 P-3563,购自 Sigma 公司,2~8℃保存。
3) 磷酸盐缓冲盐水缓冲液加入 0.05% Tween-20 (PBST),pH 7.4,于 2~8 °C 保
存最长时期为 6 个月。PBST 干粉可用于制备缓冲液,货号为 P-3563,购自 Sigma
公司。
4)PVP,分子量为 40,000,货号 PVP-40,购自 Sigma 公司。
5)胰蛋白酶片剂,货号为 T-7168,购自 Sigma 公司,-20℃保存。.
6)对甲苯磺酰氟(PMSF),分子量为 174.2,货号为 23,479-6,购自 Aldrich Chemical
公司。
7)Cry1Ac 标准样品:截短的 Cry1Ac 核心蛋白质,由 Dow AgroSciences LLC 公
司提供,Zionsville 路 9330 号,Indianapolis, IN 46268。
8)PBST+1% PVP (w/v) (PBST/PVP),于 2~8℃保存最长时期为 6 个月。干粉
可用于制备缓冲液,货号为 P-3563,购自 Sigma 公司。
在 1 L PBST 中加入 10 mg PVP; 在溶液中放入搅拌子,容器放在搅拌器上,搅拌至完全溶解,于 2~8℃保存
最长时期为 3 个月。如见任可污染需重新配制。 9)胰蛋白酶溶液 在 1L 去离子水中加入一片胰蛋白酶片剂;
37℃ 温育 15 分钟,使其溶解;
制备好之后,在冰上保存 2 小时。
10)配制 PMSF 溶液,10 mM。
称取 8.78 mg PMSF,溶于 5 mL 异丙醇;
混合后于 2~8℃保存最多 1 个月。
11)配制 Cry1Ac 储存液,1000 ng/mL。
用冻干的 Cry1Ac 浓缩液制备 Cry1Ac 标样溶液,-20℃冻存;
用 PBST/PVP 配制 1000 ng/mL 储存液,依标样要求,调至不同浓度,置冰
141
上,2 小时内使用。如见任可污染需重新配制。没用完的 Cry1Ac 储存液不
留存,弃去。
4.2.3 试验流程
1)试验前 30 分钟从冰箱取出 ELISA 试剂盒,回暖至 20~25℃。
2)试剂瓶中加入 6 mL of PBST 缓冲液,再加入 22 L 与 Cry1Ac 抗体偶联物浓缩
液,充分混匀,此为 50 倍抗体偶联物稀释液。依反应孔的数目按需调整体积。
标记配制日期和失效日期,Cry1Ac 抗体偶联于配制日起 15 天失效。
3)在 5 mL 聚丙烯离心管中配制 Cry1Ac 标样:在进行下一轮稀释之前涡旋数秒,
将离心管置冰上;标样在试验之前现用现配制,配制比例见附表 15。
附表 15. 标样配制方法
母液的 浓度 ng/mL
母液等分体
积 µL 起始缓冲液 体积 µL
反应终 体积 µL
最终标样 浓度 ng/mL
等分之后 体积 a µL
1000 15 1485 1500 10.00 540
10.00 960 240 1200 8.00 450
8.00 750 250 1000 6.00 450
6.00 550 330 880 3.75 480
3.75 400 350 750 2.00 430
2.00 320 320 640 1.00 440
1.00 200 200 400 0.50 400
0 0 500 500 0 500 a:最终的体积是取出用于稀释下一梯浓度标样后的剩余的体积。
据标样与相应吸光度绘制标准曲线(附图6)。
附图6. ELISA检测Cry1Ac蛋白质的标准曲线

142
4)试样准备:
称取 15 mg 棉花组织冻干粉,分配至 2 mL 聚丙烯管,每管加入 2~3 个金
属珠,加入 1.5 mL 提取缓冲液,盖紧盖子;
使用 Geno/Grinder 自动振荡研磨仪提取试样中的蛋白液,dial 设为 500,切
换开关设为 1×,即每分钟振荡 1500 下 ,每循环 5 分钟;
≥3,000 rpm 离心 5 分钟,或直至上清液清晰分离。将上清液移至另一离心
管。置冰上,3 小时内使用。
5)试样处理
将 950 µL 提取液移至 2.0 mL 离心管;
每 950 µL 提取液中加入 30 μL 胰蛋白酶溶液,混合,在 37℃温育 1 小时;
加入 20 µL 10 mM PMSF 终止反应,置冰上,2 小时内使用。
6)将 Cry1Ac 试样加入 ELISA 反应孔
将 ELISA 标准样品稀释,加样至 96 孔 U 底微量滴定板,每孔 100 μL。每板
标准样品设 3 次重复。
准备试样稀释液,加样到 96 孔微滴定板,每孔 100 μL。记录样品位置。板
上有标准样品,
取 50 µL Cry1Ac 抗体偶联物,加到孔壁包被了抗体的反应孔,然后加 50 µL ELISA 标准样品溶液时(也包被了抗体),反应板与加入试样的位置和方向
保持一致。
7)将 96 孔板加盖,轻柔地旋转或在振荡仪上振荡 5 分钟,使试剂混匀。室温下
存放 1 小时(± 5 分钟)。
8)用洗板机洗板 3~5 次,反应孔中加入 PBST 洗液,每次甩干洗液。
9)每反应孔加入 100 µL TMB 底物,加盖轻柔温匀,室温黑暗保存 15±2 分钟。
10) 加入 100 µL 反应终止液,轻柔温匀,以终止反应。使用 MAXline Vmax 酶
标仪读取 450 nm 波长处的吸光值。
11) 保存读数,存档,分析。
4.2.4 数据分析和统计
1)标准曲线:
用标准样品的吸光值绘制标准曲线。使用 SOFTmax PRO 软件或 Microsoft
Excel 进行分析和计算。依标准样品已知浓度及其对应的吸光度的二次回归关系,
绘制 Cry1Ac 的 ELISA 标准曲线。

143
2)方程拟合的标准曲线上的最佳抛物线方程式:
y = A + Bx + Cx2
其中:
y = 平均吸光度 (OD) x = 参考标准样品的浓度
3)计算试样中 Cry1Ac 的量
使用 SOFTmax PRO 或 Microsoft Excel 计算 Cry1Ac 每一反应孔中的浓度。
将未被测样品的吸光值代入二次方程来计算浓度。
预期的浓度(未知的试样浓度) = ( )
C2
OD-A*4C - B B2
+−
判断 Cry1Ac 提取液试样的浓度时,还要将预期浓度乘以稀释倍数和方法参
数。
方法参数 (MF) = 组织重量 (mg) 抽提体积 (mL)
× 0.95
0.95 表示每 mL 终样品(加入抑制剂后)含 0.95 mL 提取物。
4.2.5 分析物的接受标准
每批次的分析应满足试验接受标准(附表 16)。参照下表的参数决定是否接
受每个 ELISA 板上获得的数据。如果不能满足如下标准,需重复试验。
附表 16. 棉花中 Cry1Ac 特异性 ELISA 试验接受标准
参数 标准
缓冲液空白对照 450 nm 波长处的吸光度<0.100
0 ng/mL 标准样品 450 nm 波长处的吸光度<0.100
1000 ng/mL 标准样品 450 nm 波长处的吸光度 ≥ 1.00
标准曲线 R2 (决定系数)>0.990
所有阳性样品重复 a 三次重复的变异系数 CV≤ 15%
已知浓度的或质量控制样品、溶液 OD 变异系数 CV ≤ 20%
质量控制样品、溶液(如可能) 测量值 ≤ ±20% 真实值 a:指吸光度大于定量极限(4 ng/mL)吸光度的样品,包括阳性对照和待测样品。

144
4.3 ELISA 定量检测转基因棉花中 PAT 的试验方法
适用范围:本ELISA方法适用于转基因棉花叶、根、花、铃、花粉、整株、
棉籽和及其加工产品(棉籽壳、棉籽仁、熟棉籽粕和棉籽油)中PAT蛋白质的定
量检测。检测范围为0.25 ng/mL~6.0 ng/mL。棉籽油中PAT的检测极限LOD为0.30
ng/g,其它组织和产品的LOD为0.25 ng/mg。
原理:本 ELISA 体系为“三明治式”ELISA,结合显色酶底物来定量检测棉
花样品中提取的 PAT 蛋白。以特异性的多克隆抗体包被在 96 孔滴定板上,用与
辣根过氧化酶偶联的单克隆抗体作为检测抗体。棉花的提取液在特异性的多克隆
抗体包被的 96 孔滴定板孔中与辣根过氧化酶偶联的单克隆抗体共同孵育,如果
棉花的提取液中含有 PAT 蛋白,孔中包被的抗体和辣根过氧化酶标记的抗体将
与其结合。经洗涤后去除游离的多余抗体。这样,包被抗体─PAT─检测抗体在孔
中结合在一起形成“三明治式”复合物。最后,经洗涤后加入辣根过氧化酶的显
色底物,显色后加反应终止液并用酶标仪读取吸光值。显色的强度与孔中加入的
PAT 蛋白的量是成正相关,棉花提取液中 PAT 蛋白质的浓度可由已知浓度的标
准品蛋白质建立的标准曲线来推算。
4.3.1 试剂和标准样品
1)试剂
i. PAT 微量滴定板ELISA试剂盒,货号AP-014-V10,购自Envirologix Inc.公司;
于2~8℃保存。试剂盒中有如下试剂:
抗体包被的96孔滴定板, PAT抗体偶联物(1×),
底物溶液(1×),
反应终止液(1N HCl);
ii. PVP,分子量为40,000,货号PVP-40,购自Sigma公司;
iii. PBST,pH 7.4,货号P-3563,购自Sigma公司。
2)标准样品
标样来自Dow AgroSciences公司;
如需要,Dow AgroSciences公司法规科学部门还提供阳性对照和阴性对
照。
3)试剂和标样的配制
i. 配制磷酸盐缓冲盐水加入0.05% Tween 20 (PBST),pH 7.4
1 L去离子水中加一包溶质,
容器中放入搅拌子,放在搅拌器上搅拌,
在20~25℃可保存2~3个月,2~8℃可保存6个月;
ii. 配制磷酸盐缓冲盐水加入0.05% Tween 20和1% PVP (w/v),即(PBST/PVP)
145
将10 g PVP加入1 L PBST,
容器中放入搅拌子,放在搅拌器上搅拌,
2~8℃可保存3个月;
iii. 配制3000 ng/mL和100 ng/mL PAT 母液
PAT起始贮存液可为溶液,也可为冻干粉。如300 μg/mL溶液,
如使用300 μg/mL的溶液为母液,用之前涡旋混匀;移取10 μL至990 μL PBST/PVP,充分混匀,制成3000 ng/mL母液;从中移取40 μL至1160 μL PBST/PVP中,配成100 ng/mL母液。冰上保存,2小时内使用;
iv. 在15 mL试管中,按附表17稀释样品:
附表17. 试样配制方法
超始贮备液浓度
(ng/mL) 加样体系
(mL) 缓冲液体
(mL) 终体积
(mL) 工作浓度
(ng/mL) 相当于样品浓度
(ng/mL)
100 0.48 9.52 10.00 4.80 0.48 100 0.36 9.64 10.00 3.60 0.36 100 0.12 9.88 10.00 1.20 0.12 100 0.06 9.94 10.00 0.60 0.06 0 0 10.00 -- 0 0
4.3.2 提取蛋白质
1)对于棉籽油以外的样品,称取15 mg棉花组织冻干粉,分配至2 mL 聚丙烯试
管,每管加入2~3个金属珠,加入1.5 mL提取缓冲液,盖紧盖子;以缓冲液作
为提取空白对照;
2)使用Geno/Grinder自动振荡研磨仪提取试样中的蛋白液,dial设为500,切换
开关设为1×,即每分钟振荡1500下 ,每循环3分钟;
3)≥3,000 rpm离心5分钟,或直至上清液清晰分离。将上清液移至另一离心
管。置冰上,4小时内使用;
4)从油中提取测试物
量取800 mg棉籽油,分配至5 mL聚丙烯管,加入1.5 mL提取缓冲液,设
阴性对照,设提取空白对照(提取过程以缓冲液代替试样),
盖紧盖子,用Geno/Grinder自动振荡研磨仪提取试样中的蛋白液,dial设
为500,切换开关设为1×,即每分钟振荡1500下,每循环为3分钟。涡
旋,每循环为3分钟,
≥3,000 rpm离心至少5分钟,直至上清液清晰分离,
将上清液移至另一试管,置冰上保存,4小时内使用。
4.3.3 标准曲线建立
1)准备PAT 特异性ELISA试剂盒
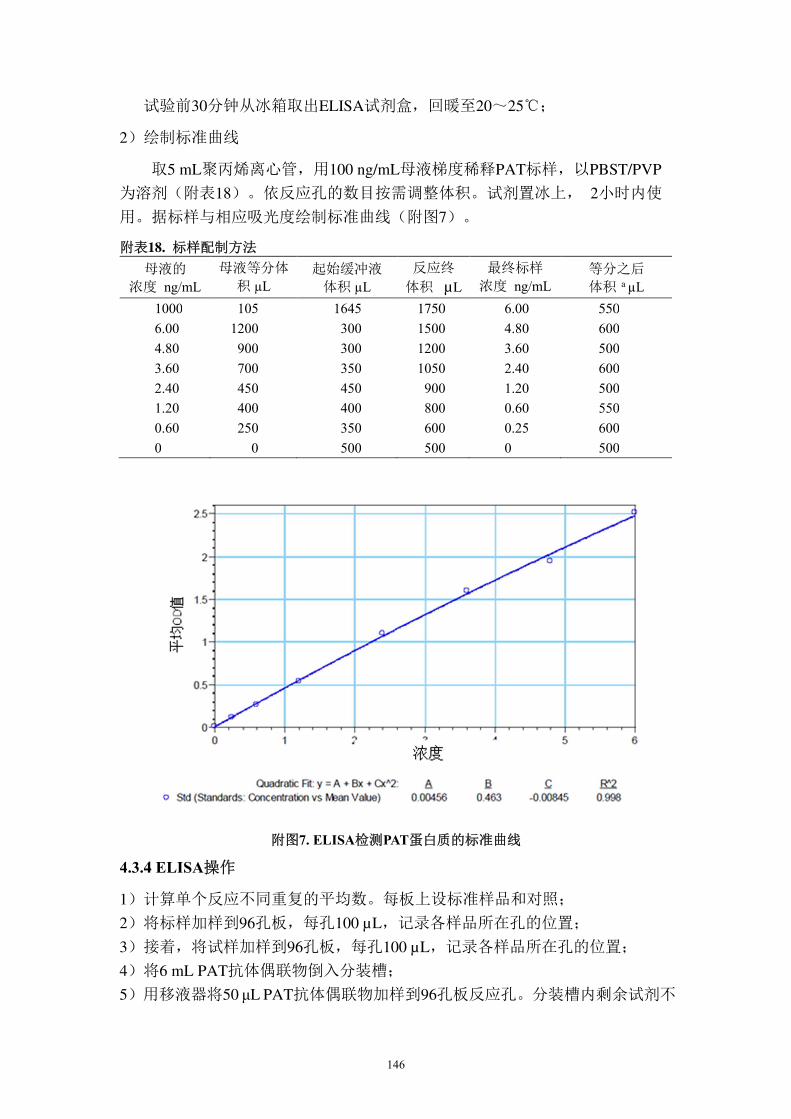
146
试验前30分钟从冰箱取出ELISA试剂盒,回暖至20~25℃;
2)绘制标准曲线
取5 mL聚丙烯离心管,用100 ng/mL母液梯度稀释PAT标样,以PBST/PVP
为溶剂(附表18)。依反应孔的数目按需调整体积。试剂置冰上, 2小时内使
用。据标样与相应吸光度绘制标准曲线(附图7)。
附表18. 标样配制方法
母液的 浓度 ng/mL
母液等分体
积 µL 起始缓冲液 体积 µL
反应终 体积µL
最终标样 浓度 ng/mL
等分之后 体积 a µL
1000 105 1645 1750 6.00 550 6.00 1200 300 1500 4.80 600 4.80 900 300 1200 3.60 500 3.60 700 350 1050 2.40 600 2.40 450 450 900 1.20 500 1.20 400 400 800 0.60 550 0.60 250 350 600 0.25 600 0 0 500 500 0 500
附图7. ELISA检测PAT蛋白质的标准曲线
4.3.4 ELISA操作
1)计算单个反应不同重复的平均数。每板上设标准样品和对照;
2)将标样加样到96孔板,每孔100 µL,记录各样品所在孔的位置;
3)接着,将试样加样到96孔板,每孔100 µL,记录各样品所在孔的位置;
4)将6 mL PAT抗体偶联物倒入分装槽;
5)用移液器将50 μL PAT抗体偶联物加样到96孔板反应孔。分装槽内剩余试剂不

147
存留;
6)将50 μL ELISA不同浓度的标样溶液加样至反应孔;
7)封盖。轻柔混匀,在20~30℃温育60分钟;
8)每孔使用350 μL PBST洗板,用纸巾吸干PBST。共洗板5次;
9)将12 mL 底物溶液倒入分装槽;
10)用移液器将100 μL 底物溶液加样到96孔板反应孔。分装槽内剩余试剂不存留;
11)封盖。轻柔混匀,在20~30℃温育15分钟;
12)将12 mL反应终止液倒入分装槽;
13)用移液器将100 μL反应终止液加样到96孔板反应孔。轻柔混匀。此步聚需避
光操作;
14)读取450 nm波长的吸光值,减去650 nm波长的背景吸光值,在终止反应后30
分钟内完成读取。
4.3.5 数据分析和计算
1)使用SOFTmax PRO 软件或Microsoft Excel进行分析和计算。依标准样品已
知浓度及其对应的吸光度的二次回归关系,绘制PAT的ELISA标准曲线。
2)方程拟合的标准曲线上的最佳抛物线方程式:
y = A + Bx + Cx2
其中:
y = 平均吸光度 (OD)
x = 参考标准样品的浓度
3)计算待测样品中PAT的浓度
使用SOFTmax PRO或Microsoft Excel计算PAT每一反应孔中的浓度。将未被
测样品的吸光值代入二次方程来计算浓度。SOFTmax PRO计算其平均数、标准
差、变异系数。
4)计算待测试样中PAT的浓度
预估浓度是平均错误百分率的基础。预估浓度由曲线和吸光度的系数决
定,使用SOFTmax PRO或Microsoft Excel计算PAT每一反应孔中的浓度。将未被
测样品的吸光值代入二次方程来计算浓度。
待测的浓度(未知的试样浓度) = ( )
C2
OD-A*4C - B B2
+−
4.3.6 分析物的接受标准
参照附表 19 中的参数决定是否接受每个 ELISA 试验数据。

148
附表 19. 棉花中 PAT 特异性 ELISA 试验接受标准
参数 可接受标准
空白对照 吸光度(450 nm~650 nm)<0.100
0 ng/mL 标准样品 吸光度(450 nm~650 nm)<0.100
6 ng/mL 标准样品 吸光度(450 nm~650 nm)≥1.500
标准曲线 R2 (Correlation of determination) >0.990
阳性样品 变异系数百分值≤15%
标准未知样品 变异系数百分值≤20%
质量控制样品、溶液(如适用) 测量值≤ ±20% 真实值

149
附件 5.转基因植物对生态环境安全性的综合评价报告
《转基因植物对生态环境和食品安全性的综合评价报告》
一、摘要
DAS-21023-5 棉花表达杀虫晶体蛋白质 CrylAc 和 PAT(PAT 仅用做选择标
记),从而使转基因棉花免受特定鳞翅目害虫的危害。
自 1999 年起对 DAS-21023-5 棉花分子特征、遗传稳定性、环境和食品安全
性进行了大量的试验研究并做出了综合的安全性评估。评估结果显示:1)外源
基因在转基因棉花中有单一的插入位点、单拷贝、结构完整,且转化体不含载体
骨架序列,转化方法安全,外源片段整合至第 8 染体体,外源片段全序列及旁侧
序列明确,表达量及表达部位、表达时期明确。2)遗传稳定性的鉴定证明外源
基因在代际间稳定整合、稳定表达且目标性状稳定表现。3)对环境的影响及相
互作用方面与非转基因棉花对照等同,未表现出任何生存竞争力增强、杂草化倾
向或基因漂移的风险,对非靶标生物以及生物多样性没有不利的影响。4)DAS-
21023-5 外源表达的蛋白质无毒性、无致敏性,转基因棉花及其产品的营养成分
和抗营养因子与非转基因对照等同,在人类膳食中的接触风险极低,不含抗生素
抗性基因,生产加工不会增加其食用风险,也未观察到其它的不利影响。
二、背景介绍
棉花是世界上最主要的经济作物之一。Bt棉花在全球十数年的商业化推广
带来了巨大的经济效益、环境效益和生态效益。在此期间,人们从未停止对其
生物安全性的研究和评价,目前尚未发现一例可证的对人类、动物、植物、微
生物和环境有害的事例。
转化受体经遗传转化后,将转化位点转育至回交受体PSC355中,选育出了
转基因株系DAS-21023-5(见附图8)。DAS-21023-5(含cry1Ac基因)棉花与
DAS-24236-5棉花(含cry1F基因)杂交,选育出复合的商业化品系DAS-24236-
5×DAS-21023-5。从1999年以来进行了大量的安全性试验研究,自2004起获得
美国、加拿大、巴西、欧盟、日本、南韩、澳大利亚和新西兰、哥伦比亚、中
国台湾等多个国家或地区的批准并以相应用途投放市场。
三、受体生物学特性
转基因棉花 DAS-21023-5 的转化受体生物为锦葵科棉属陆地棉种
(Malvaceae Gossypium hirsutum)的栽培品种 GC510。早在 5000 年前人类就开始
使用棉花。陆地棉为四倍体棉种,最初起源于墨西哥。我国主要种植陆地棉,棉
区广阔,北起新疆北部的玛纳斯河流域,南至海南岛,西起新疆的喀什地区,东
抵长江三角洲的沿海地带和东北的辽河流域均种植棉花。根据不同地区积温多寡、
纬度高低、降水量多少等自然生态条件,将我国植棉区由南而北、从东向西依次
划分为华南棉区、长江流域棉区、黄河流域棉区、北部特早熟棉区和西北内陆棉
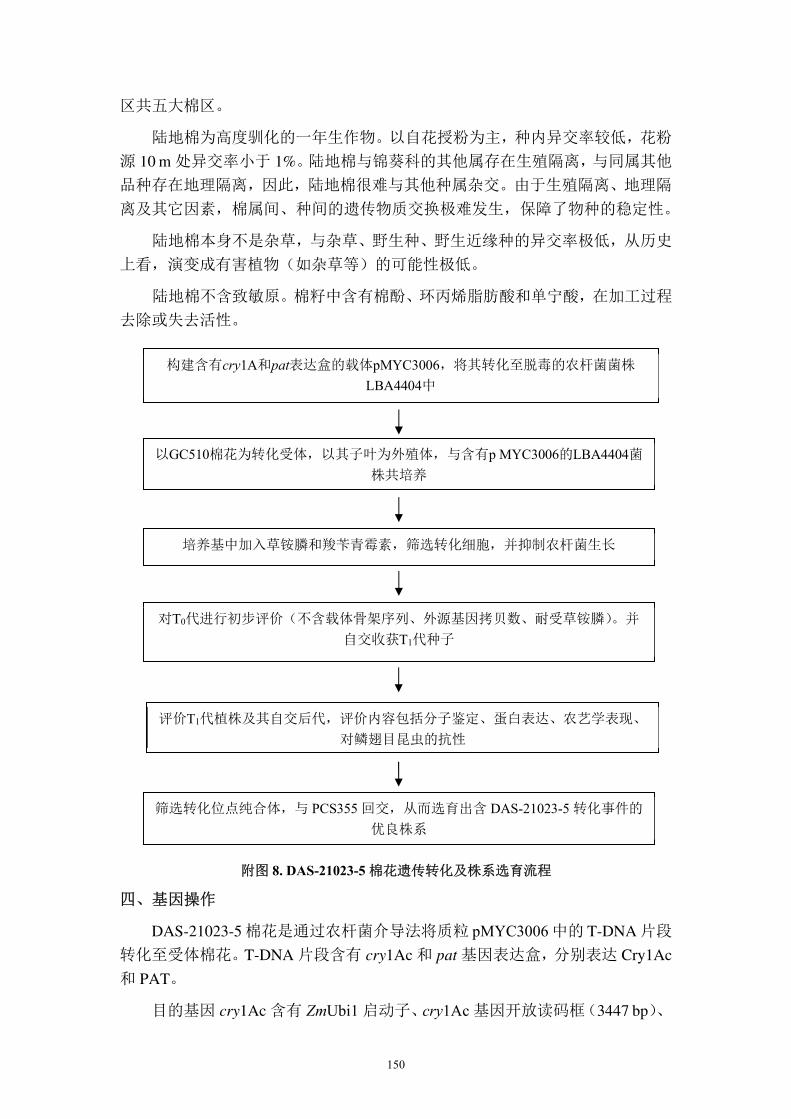
150
区共五大棉区。
陆地棉为高度驯化的一年生作物。以自花授粉为主,种内异交率较低,花粉
源 10 m 处异交率小于 1%。陆地棉与锦葵科的其他属存在生殖隔离,与同属其他
品种存在地理隔离,因此,陆地棉很难与其他种属杂交。由于生殖隔离、地理隔
离及其它因素,棉属间、种间的遗传物质交换极难发生,保障了物种的稳定性。
陆地棉本身不是杂草,与杂草、野生种、野生近缘种的异交率极低,从历史
上看,演变成有害植物(如杂草等)的可能性极低。
陆地棉不含致敏原。棉籽中含有棉酚、环丙烯脂肪酸和单宁酸,在加工过程
去除或失去活性。
附图 8. DAS-21023-5 棉花遗传转化及株系选育流程
四、基因操作
DAS-21023-5 棉花是通过农杆菌介导法将质粒 pMYC3006 中的 T-DNA 片段
转化至受体棉花。T-DNA 片段含有 cry1Ac 和 pat 基因表达盒,分别表达 Cry1Ac
和 PAT。
目的基因 cry1Ac 含有 ZmUbi1 启动子、cry1Ac 基因开放读码框(3447 bp)、
构建含有cry1A和pat表达盒的载体pMYC3006,将其转化至脱毒的农杆菌菌株
LBA4404中
以GC510棉花为转化受体,以其子叶为外殖体,与含有p MYC3006的LBA4404菌
株共培养
培养基中加入草铵膦和羧苄青霉素,筛选转化细胞,并抑制农杆菌生长
评价T1代植株及其自交后代,评价内容包括分子鉴定、蛋白表达、农艺学表现、
对鳞翅目昆虫的抗性
筛选转化位点纯合体,与 PCS355 回交,从而选育出含 DAS-21023-5 转化事件的
优良株系
对T0代进行初步评价(不含载体骨架序列、外源基因拷贝数、耐受草铵膦)。并
自交收获T1代种子

151
AtuORF25/26 3′UTR 终止序列。cry1Ac 来源于苏云金芽孢杆菌,所表达的 Cry1Ac
可以保护植物免受多种鳞翅目害虫的侵害。
选择标记基因 pat表达盒中含有AtuMas启动子、pat基因开放读码框(522bp)
和上述双向 AtuORF25/26 3′UTR 终止序列(与 cry1Ac 共用同一终止子),pat 基
因来源于土壤细菌绿色产色链霉菌,表达的 PAT 可使转化体耐受草铵膦,在转
基因棉花 DAS-21023-5 开发过程中,pat 作为选择标记基因。DAS-21023-5 棉花
的转化、筛选过程见附图 8。
五、遗传稳定性
1 同一世代内个体间的遗传稳定性
使用 Cry1Ac 蛋白质特异性的侧流试纸条以及转化体特异性 PCR 方法,检
测了 DAS-21023-5 的 BC4F2 群体中 Cry1Ac 蛋白质的表达和外源基因整合情况。
结果显示,在 121 个测试植株中,Cry1Ac 蛋白质表达和 PCR 阳性的为 91 株,
其余 30 株不表达 Cry1Ac 蛋白质且 PCR 阴性。转化体的表型分离和遗传分离的
比例一致,均符合单一遗传位点孟德尔分离比例 3:1,详见报告 1F。
2 代际间外源基因整合的稳定性
Southern blot 显示,转基因棉花 DAS-21023-5 四个世代 BC1F3、BC3F3、BC3F4
和 BC4F1 的植株中均在同一个位点含有单一拷贝的、结构完整的片段,且转化体
均不含架体骨架序列(报告 1E)。
3 代际间外源基因表达水平的稳定性
跟踪检测了 DAS-21023-5 三个世代(BC4F2 、F2 和 BC3F6)外源蛋白质时、
空表达的情况。取样时期和部位分别为 BBCH 15 期的叶片、根和成熟籽粒,采
用 ELISA 方法测定 Cry1Ac 和 PAT 蛋白质的含量。三代之间不同时期的组织中,
无论是 Cry1Ac 还是 PAT 的表达量均相对一致。表明转化体 DAS-21023-5 代际
间在外源蛋白质表达水平相对稳定(报告 3)。
4 代际间外源基因性状表现水平的稳定性
在 2001~2004 年,对目标性状进行了多年、多世代、多地点的田间试验和
实验室研究,结果均显示,DAS-21023-5 棉花对鳞翅目昆虫具有稳定的防治效果
(报告 4A 和 4B)。
六、环境安全评价
经过多年多点田间试验,在对生存竞争能力、基因漂移和对非靶标生物的影
响方面 DAS-21023-5 棉花与非转基因对照等同,对环境无负面的影响。
1 生存竞争能力
2002 年在美国棉区内的 20 个试验地点,以 DAS-21023-5 棉花材料进行了大
规模的田间试验,其种子活力、种子休眠、生长发育、生殖率及产量等各项农艺

152
学性状均与传统棉花等同,即目标性状的导入没有改变其生存竞争能力,亦未赋
予其杂草化倾向和侵蚀自然生态环境的能力(报告 5)。
2 基因漂移
理论上,DAS-21023-5 棉花的遗传物质向其它生物漂移的概率极低;实验数
据表明,该转化体与其非基因对照的遗传物质向其它物种漂移的概率等同。
棉花以自花授粉为主,可与同种内其它品种授、受粉,距花粉源 1 m 处异
交率约为 1%(Van Deynze et al., 2005)。另外,由于遗传隔离、生殖隔离及地理
隔离限制了转基因棉花 DAS-21023-5 的外源基因向其它属、种的漂移。仅在实验
条件下,棉花才能和锦葵科的其他属异交产生可育杂种植株(Mehetre et al., 1980),
但是自发的属间杂交的概率极低。而遗传物质从植物向其他物种如微生物、动物
或人类的水平转移概率同样微小(Nielsen et al., 2000;Bertolla and Simonet, 1999)。
尚未见有棉花的遗传物质向其它动物、微生物转移的报导(FAO/WHO, 2000;
Conner et al., 2003)。
通过实验比较了 DAS-21023-5 棉花及其非转基因对照花粉的活力和形状大
小,结果显示二者的花粉特征无显著差异(报告 6),因此二者通过花粉传播的概
率等同。
总之,DAS-21023-5 棉花的遗传物质向其它生物漂移的可能性极低,漂移概
率与其非转基因对照等同,对环境的影响等同。
3 对靶标与非靶标生物的影响
对靶标害虫的影响
转基因棉花 DAS-21023-5 的靶标生物是鳞翅目昆虫。本公司于 2001 年和
2002 年在超过十个地点的田间试验数据均显示,DAS-21023-5 棉花可有效控制
鳞翅目害虫,对以叶片为食的害虫种类(如甜菜夜蛾、草地贪夜蛾、粉纹夜蛾、
大豆尺蠖及南方粘虫等)有较优的抗虫效果(报告 4A 和 4B)。
对非靶标生物和生物多样性的影响
以微生物来源的 Cry1Ac 蛋白为测试物对植食性昆虫蚜虫、天敌昆虫草蛉、
瓢虫、寄生蜂、土壤腐生昆虫蚯蚓、跳虫、水生动物大型溞、虹鳟鱼、禽类美洲
鹑、哺乳动物小鼠、经济传粉昆虫蜜蜂及保护昆虫帝王斑蝶等 12 种非靶标生物
进行了一系列生物测定进行毒性评估,由接触途径、外源蛋白质在相应组织中表
达量或在土壤、水中的归趋来判断直接或间接接触水平,再结合外源蛋白质对非
靶标生物的喂食试验计算 MoE,从而判断接触风险的大小。由于所有非靶标生
物对 Cry1Ac 的 MoE 极高或较高,表明 Cry1Ac 对非靶标生物无不良影响(报告
7A-7K)。
转基因棉花对非靶标节肢生物群落无影响(报告 7L):2002 年,以 PSC355
为非转基因对照,采用扫网法、陷阱法等多种取样方法,在 Louisiana 和 Arizona

153
收集、计数、比较非靶标生物(有益昆虫)。结果显示,非靶标节肢生物的群落
和数量在转基因棉田中并不低于其非转基因对照;转基因 Bt 棉花对微生物群落
多样性及植物群落和结果多样性均无负面影响。转基因棉花表达的 Cry1Ac 蛋白
质在土壤中的半衰期为 1.3 天,不会蓄积。
七、食用安全评价
转基因抗虫 DAS-21023-5 棉花与非转基因对照棉花的营养成分、抗营养因
子含量等同,且二者加工产品中营养成分、抗营养因子含量也等同,外源蛋白质
无潜在的毒性与致敏性,转化体上无抗生素抗性标记基因,未观察到其它对食用
安全的不利影响,因此,DAS-21023-5 棉花与其非转基因对照棉花的食用风险等
同。
1 营养学评价
DAS-21023-5 棉花与其非转基因对照的抗营养因子和关键营养成分含量等
同。
对 DAS-21023-5 棉花和非转基因对照的植株、棉籽及其加工产品(棉仁、棉
籽壳、熟棉粕和精炼棉籽油)的抗营养因子、营养成分如一般成分、矿物质、氨
基酸和脂肪酸进行了取样分析,结果显示,两者的各项指标等同(报告 2)。同
时,也测定了其中的抗营养因子和天然毒素,包括萜烯醛类化合物、环丙烯类脂
肪酸(如锦葵酸、苹婆酸和二氢苹婆酸等)、总棉酚、游离棉酚及黄曲霉毒素等,
结果同样显示二者的棉籽及其加工产品中抗营养因子和天然毒素的含量相似(报
告 2)。并且,测定值与文献报道值近似或是在文献报道值范围内,因此,转基因
棉花在营养成分和抗营养因子方面与其非转基因对照实质等同。在包含此棉花的
复合性状转基因棉籽中也没有检测到超过域值的除草剂残留。另一方面,90 天
大鼠喂养和 42 天肉鸡喂养试验均未见全食品对实验动物带来营养学负面影响。
2 毒理学评价
DAS-21023-5 棉花及期表达的外源蛋白质 Cry1Ac 和 PAT 无潜在的毒性。在
毒性。
首先,外源基因均来自安全的供体生物并有长期的安全应用历史;Cry1Ac 及
PAT 蛋白质有较安全的应用历史;生物信息学表明,外源蛋白质与已知毒性蛋白
质氨基酸和抗营养因子序列无较高的相似性(报告 8A 和 9A);外源蛋白质受热
不稳定(报告 8C 和报告 9C),易被模拟胃液消化(报告 8D 和报告 9D);以小
鼠为实验动物,显示 Cry1Ac 和 PAT 对哺乳动物无经口急性毒性,LD50 均大于
5050 mg/(kgBW)(报告 8E 和报告 9E);Cry1Ac 和 PAT 在人类膳食中接触风险
极低,MoE 分别达 108和 109 数量级。90 天大鼠喂养试验均未见全食品对实验动
物带来毒理学负面影响。

154
3 致敏性评价
棉花本身不是致敏生物,转基因抗虫棉花 DAS-21023-5 中,外源基因 cry1Ac
和 pat 的供体生物不是致敏生物,且有长期安全的使用历史;Cry1Ac 和 PAT 本
身也有安全的应用历史;Cry1Ac 和 PAT 与已知致敏原氨基酸序列无较高的相似
性(报告 8B 和报告 9B),受热后不稳定(报告 8C 和报告 9C),在模拟胃液中迅
速被降解(报告 8D 和报告 9d),即不具有致敏性蛋白质受热稳定和不易消化的
特征,且翻译后不被糖基化(报告 8D、报告 9D)。Cry1Ac 和 PAT 在人类膳食
中接触量极少。因此,DAS-21023-5 表达的外源蛋白质诱发人、畜过敏的可能性
极低。
4 抗生素抗性
DAS-21023-5 棉花基因组中未引入抗生素抗性基因。首先,转化质粒
pMYC3006 上的 T-DNA 上不含抗生素抗性标记。其次,质粒 pMYC3006 骨架上
虽有红霉素抗性基因,但不能整合至受体基因组中,经多世代的 Southern blot 分
析证明,转基因抗虫棉花基因组中未整合载体骨架序列,即不含骨架上的抗生素
抗性标记基因(报告 1E)。
5 除草剂列留及真菌素素
DAS-21023-5 的目标性状是抗特定的鳞翅目害虫。虽然其含有 pat 基因,但
是此功能仅用做遗传转化的选择性标记。一般不用于大田喷洒草铵膦。即便如此,
本公司仍对含有 DAS-21023-5 的转基因棉花进行除草剂残留检测。检测结果显
示,草铵膦及其代谢物在棉籽中的残留量为 0.042 µg/g,该值远低于美国 EPA 界
定的草铵膦在棉花最大残留限量 4.0 µg/g,即本转化体不会带来农药残留危害。
以大鼠喂养试验所用材料为例,在配制饲料之前,采用液相色谱-质谱联用
(LC-MS/MS)技术检测并比较了转基因棉花和其非转基因对照中真菌毒素含量。
结果显示,黄曲酶毒素、玉米赤霉烯酮、DON、各项伏马毒素、T-2 毒素等各指
标均在转基因棉花与其非转基因对照间含量一致。表明遗传转化不会提高本转基
因棉花籽粒中真菌毒素含量,不会因此带来食用、饲用风险。
八、非预期效应
本公司对 DAS-21023-5 棉花进行的大量的分子特征鉴定、遗传稳定性检测、
环境安全和食用安全性评估试验,尚未观察到非预期效应。
九、生产前景及安全监控措施
在转基因抗虫棉花品系 DAS-21023-5 抗鳞翅目昆虫,于 2004 年获得美国农
业部批准后在美国及其它国家广泛种植,带来了可观的经济效益和生态效益。
本公司在中国对 DAS-21023-5 棉花的申请仅限于进口用作食品和饲料
的加工原料,不涉及在中国境内的商业化生产和种植活动。
作为转基因抗虫棉花 DAS-21023-5 的研发商,本公司在获得农业农村部

155
颁发的转基因生物安全证书后,将要求贸易商严格遵守《农业转基因生物安全管
理条例》和其它相关法律法规的要求,确保 DAS-21023-5 棉化在进口加工过
程中用途不被改变、包装牢固、标识清楚、运输环节严格操作。在必要情况下,
本公司会向贸易商提供技术支持和指导,确保 DAS-21023-5 棉化的相关贸易符
合农业行政主管部门的要求。
十、结论
对转基因抗虫棉花 DAS-21023-5 进行了大量的分子特征、遗传稳定性、环境
安全性和食用安全评估后,认为该棉花在环境和食品方面的风险与传统非转基因
棉花等同,不会对人类、动物、微生物和环境带来负面影响。

156
附件 6.转基因植物对食品安全性的综合评价报告
转基因植物对食品安全性的综合评价报告请参见附件 5:《转基因植物对生
态环境和食品安全性的综合评价报告》。

157
附件 7. 该类转基因植物国内外生产应用概况
全球转基因作物应用概况
自 1996 年转基因作物大规模商业化以来,转基因作物的种植已经超过了二
十年。有 26 个国家种植了转基因作物,44 个国家进口转基因作物。1996 至 2018
年间,全球转基因作物累计种植面积达到 25 亿公顷。二十多年的商业化证明,
转基因作物为农民乃至为全社会带来了农业、环境、经济、健康和社会效益
(ISAAA, 2018)。
根据 ISAAA报告(ISAAA, 2018),2018年全球转基因作物的种植面积达 1.917
亿公顷。美国仍是全球转基因作物的领先者,2018 年美国生物技术作物的种植
面积达到 7500 万公顷 (占全球种植面积的 39%),其中大豆 3408 万公顷、玉米
3317 万公顷、棉花 506 万公顷、油菜 90 万公顷、甜菜 49.1 万公顷及苜蓿 126 万
公顷。第二大转基因作物种植国巴西 2018 年的种植面积为 5130 万公顷,比 2017
年增加了 100 万公顷,占全球种植面积的 27%,其中大豆 3486 万公顷、玉米 1538
万公顷、棉花 100 万公顷,另外还有 400 公顷甘蔗。阿根廷 2018 年的转基因作
物种植面积排在第三名,为 2390 万公顷,转基因技术采用率接近 100%,其中包
括 1800 万公顷大豆、550 万公顷玉米、37 万公顷棉花。加拿大排名第四,为 1275
万公顷,其中包括 2400 万公顷大豆、160 万公顷玉米和 870 万公顷油菜、1.5 万
公顷甜菜、400 公顷苜蓿和 65 公顷马铃薯。印度的种植面积排名第五,有 1160
万公顷的 Bt 棉花,采用率为 95%。2015 年前五大种植国的转基因作物种植面积
都超过了 1000 万公顷。
从性状类型来看,抗虫性状和除草剂耐受性状和为重要的性状;其他类型的
性状也在增加,如耐储藏性状、营养改良性状等;复合性状转基因作物呈现出不
断增长趋势,复合性状作物的面积增加了 4%,占全球转基因作物种植总面积的
42%。
国际、国内 Bt 抗虫棉花的应用概况
自 1996 年以来,带有抗虫性状的转基因棉花已在美国及全球广泛种植,Bt
抗虫棉的种植面积迅速增加,1997 年占棉花总种植面积的 15%,1999 年占 32%,
2001 年占 37%,2010 年达 73%。2015 年以后,转基因棉花种植面积达 94%以
上。促使美国棉农种植 Bt 抗虫棉的两大主要因素是提高虫害的防治效率和降低
农药施用成本(Fernandez-Cornejo 和 McBride,2012),从而提高产出和生产环境
安全性。
中国是棉花种植和消费大国。中国农业农村部于 1997 年批准国产抗虫棉商
业化,从此抗虫棉在中国种植面积不断扩大,2008 年,中国转基因棉花生产面
积增加了 30 万公顷,达到 380 万公顷,占中国棉花种植面积的 70%;2009 年
中国转基因棉花的种植面积已占总面积的 90%;2015 年中国种植了 380 万公顷
转基因作物,其中主要为抗虫棉,转基因棉花在中国的采用率达 96%。

158
Bt 抗虫棉的推广应用产生了巨大的社会和经济效益。据调查,棉农种植抗
虫棉每公顷增加收益 7650 元,其中包括:每公顷增产皮棉 375 公斤,价值
4500 元;增产棉籽 600 公斤,价值 750 元;减少农药费用 900 元;节约劳力
1500 元(贾士荣等,2001)。种植转基因抗虫棉后,每年化学农药的使用量减
少 1 万~1.5 万吨,相当于中国化学杀虫剂年生产总量 7.5%左右;棉农的劳动
强度和防治成本显著下降,棉农中毒事件降低了 70%~80%,棉田生态环境得
到明显改善。到目前为止,转基因 Bt 抗虫棉已在中国黄河和长江棉区广泛种
植。
转基因抗虫棉花品系 DAS-21023-5 的审批应用
转基因抗虫棉花 DAS-21023-5 表达来自苏云菌芽孢杆菌 CrylAc 杀虫晶体蛋
白质。大量的试验证明,DAS-21023-5 棉花能抵抗棉铃虫、红铃虫和烟草夜蛾幼
虫等多种棉花害虫,能显著减少杀虫剂的施用。DAS-21023-5 棉花与本公司开发
的另一个转基因品系 DAS-21023-5 聚合育成的双价抗虫棉,从而扩大了抗虫范
围和时间,延长其使用年限(目标昆虫抗性减缓)。本公司于 2004 年起 DAS-
21023-5 棉花陆续获得美国农业部(USDA)、美国环境保护署(EPA)、美国食
品与药物管理局(FDA)、巴西国家生物安全技术委员会(CTNBio)、加拿大卫
生部 (Health Canada)、加拿大食品监督局(CFIA)、欧盟委员会(EC)、澳
大利亚和新西兰食品标准局(FSANZ)、日本厚生劳动省(MHLW)、日本农林
水产省(MAFF)、韩国食品药品管理局(MFDS) 、韩国农村振兴厅(KRDA)、
哥伦比亚农业研究所(ICA)、哥伦比亚卫生和社会保护部(INVIMA)、南非农
林渔业部(DAFF)和中国台湾地区(TFDA)等机构的批准,并以相应用途投放
市场。迄今未见其对人类、动物、植物、微生物和环境带来任何不利影响的报导。

159
附件 8. 田间监控方案,包括监控技术、抗性治理措施、长期环境效
应的研究方法等
本公司进口转基因抗虫棉花 DAS-21023-5 用于加工原料,不作任何栽培、种
植和生产的用途。
在获得 DAS-21023-5 的安全证书以后,本公司在日后进口贸易过程中将按
照农业农村部《转基因生物安全评价管理办法》、《转基因生物安全管理条例》及
《农业转基因生物进口安全管理办法》所述原则和要求,严格使用和管理安全证
书,并要求贸易相关方清楚掌握安全证书的应用范围和有效期,要求贸易相关方
在有效期范围内向科迪华农业科技有限责任公司申请 DAS-21023-5 进口安全证
书的复印件。

160
附件 9. 审查所需的其它相关资料
本公司针对含有转化体 DAS-21023-5 的棉花进行了大量研究,包括分子生
物学特性分析、遗传稳定性分析、环境安全性评估及食品安全性评估,并提供了
本转化体特异性定性和定量检测方法,相关技术报告详见后文附件资料(报告
1~报告 11,英文原文和中文译文)。
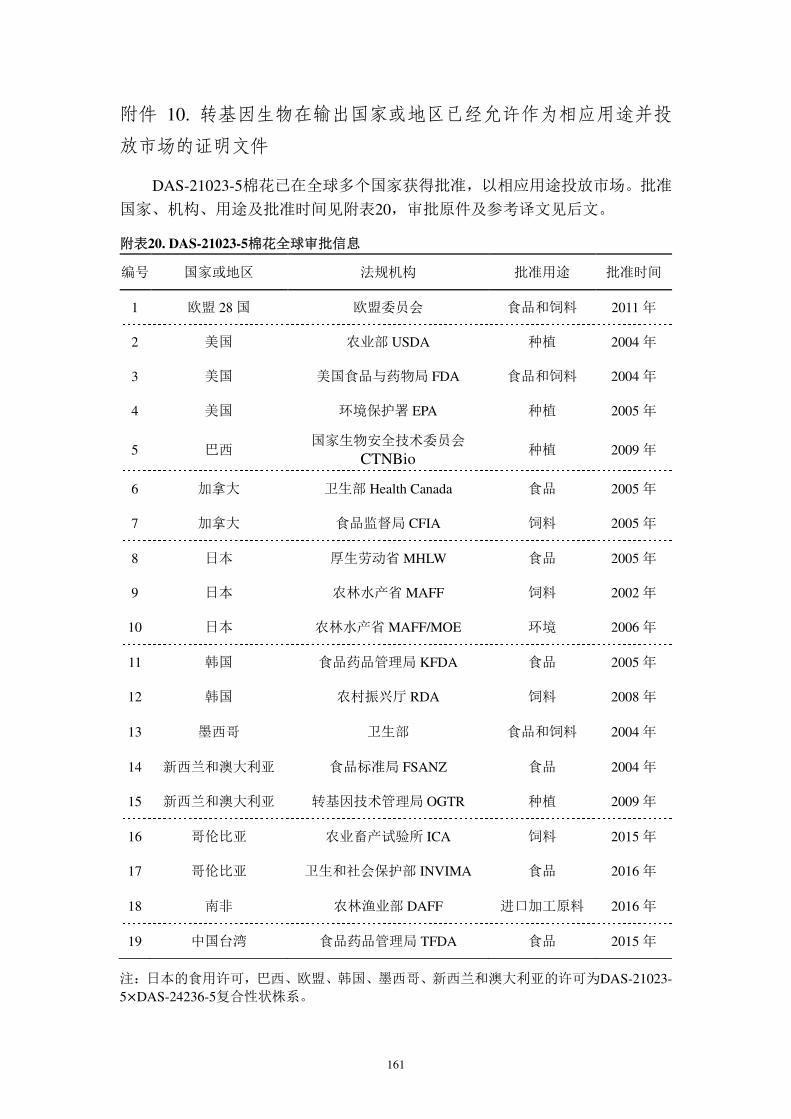
161
附件 10. 转基因生物在输出国家或地区已经允许作为相应用途并投
放市场的证明文件
DAS-21023-5棉花已在全球多个国家获得批准,以相应用途投放市场。批准
国家、机构、用途及批准时间见附表20,审批原件及参考译文见后文。
附表20. DAS-21023-5棉花全球审批信息
编号 国家或地区 法规机构 批准用途 批准时间
1 欧盟 28 国 欧盟委员会 食品和饲料 2011 年
2 美国 农业部 USDA 种植 2004 年
3 美国 美国食品与药物局 FDA 食品和饲料 2004 年
4 美国 环境保护署 EPA 种植 2005 年
5 巴西 国家生物安全技术委员会
CTNBio 种植 2009 年
6 加拿大 卫生部 Health Canada 食品 2005 年
7 加拿大 食品监督局 CFIA 饲料 2005 年
8 日本 厚生劳动省 MHLW 食品 2005 年
9 日本 农林水产省 MAFF 饲料 2002 年
10 日本 农林水产省 MAFF/MOE 环境 2006 年
11 韩国 食品药品管理局 KFDA 食品 2005 年
12 韩国 农村振兴厅 RDA 饲料 2008 年
13 墨西哥 卫生部 食品和饲料 2004 年
14 新西兰和澳大利亚 食品标准局 FSANZ 食品 2004 年
15 新西兰和澳大利亚 转基因技术管理局 OGTR 种植 2009 年
16 哥伦比亚 农业畜产试验所 ICA 饲料 2015 年
17 哥伦比亚 卫生和社会保护部 INVIMA 食品 2016 年
18 南非 农林渔业部 DAFF 进口加工原料 2016 年
19 中国台湾 食品药品管理局 TFDA 食品 2015 年
注:日本的食用许可,巴西、欧盟、韩国、墨西哥、新西兰和澳大利亚的许可为DAS-21023-5×DAS-24236-5复合性状株系。

162
1. 欧盟批准证明文件
欧盟委员会实施决议
2011年12月22日
根据欧洲议会和欧盟理事会第1829/2003条例(EC),批准含有转基因棉花281-
24-236×3006-210-23(DAS-24236-5×DAS-21023-5)的产品或者由其组成和生
产的产品投放市场。
(公布文件号C(2011)9532) (仅英文文件为官方文件)
(本文件限欧洲经济区有效) (2011/89/EU)
欧盟,
依据欧洲执行条约:
依据欧洲议会和欧盟理事会2003年9月22日颁布的有关转基因食品和饲料的
第1829/2003条例,特别是其中的第7(3)和19(3)条款:
鉴于
(1) 在2005年6月22日,陶氏益农(欧洲)公司依据第1829/2003条例(EC)
第5和17条的要求,向荷兰主管部门提交一份申请,申请将含有转基因棉花281-
24-236×3006-210-23或由其组成和生产的食品、食品成分和饲料推向市场
(“申请书”)。
(2) 该申请书还申请将包含或由281-24-236×3006-210-23棉花组成的产品除
了食品和饲料之外,与其它棉花相同的用途投放市场(栽培种植除外)。因
此,根据1829/2003条例(EC)的第5(5)和17(5)条款,申请书包括欧洲议
会第2001/18/EC号指令的附件III和IV和欧盟理事会议2001年3月12日所颁布的转
基因生物有目的环境释放, 第90/220/EEC号指令废止所要求的数据和资料,以
及依据第2001/18/EC号指令附件II进行的安全性评价资料和结论。它还包括第
2001/18/EC号指令附件VII要求的环境效应监控计划。
(3) 2010年6月15日,参照第1829/2003号条例(EC)第6和第18条款,欧盟食
品安全局(Europen Food Safety Authority, EFSA)批准陶氏益农的申请,认为棉
花281-24-236×3006-210-23与非转基因对照品系一样,对环境以及人类和动物
的健康是安全的。因此认为,含有转基因棉花281-24-236×3006-210-23(DAS-
24236-5×DAS-21023-5)或由其组成和生产的产品投放市场后,在申请书中所
述的用途背景下不会对环境或者人类和动物的健康造成任何不良影响。
(4) 在本决议中,EFSA依据第1829/2003号条例(EC)第6(4)和18(4)条
款,考虑了国家主管部门协商会议中成员国提出的所有具体问题和考虑。
(5) 在此决议中,EFSA得出结论申请人提交的包括常规监控方案在内的环境
监控计划与提出的产品用途是一致的。但是,鉴于棉籽的物理特性及其运输方

163
式,EFSA建议除了常规监控方案,需采取具体措施对种子可能溢漏及植物可能
生长地区的野生棉花进行有效监控。
(6) 为了更好的描述监控要求和遵从EFSA的建议,对申请人提交的监管计划
进行了修改。引入了一些具体的措施来限制损失,溢漏以及避免其他外来棉花
群体意外混杂。
(7) 综合上述考虑,权威部门决定批准该产品。
(8) 依据2004年1月14日欧盟委员会颁发的第65/2004条例中建立转基因生物产
品专一识别编码的产生和分配体系,每一个转基因生物(GMO)必须分配一个专
一识别编码。
(9) 依据EFSA的意见,除了第1829/2003号条例(EC)第13(1)和25(2)条
款所要求的标识以外,含有或来源于281-24-236×3006-210-23棉花的食品,食品
材料和饲料不需要其它特别的标识。但为了保证产品用途符合本决议规定,含有
转基因生物的饲料以及除食品或饲料外的其他含转基因生物的产品标识上必须
明确注明上述产品不得用于栽培。
(10) 欧洲议会和欧盟理事会议在2003年9月22号颁布的关于转基因生物的可追
踪性和标识以及生产的转基因生物的食品和饲料的可追踪性的第1830/2003条例
(EC)及第2001/18/EC条修正指令,在条款4(6)中明确规定了含有生物产品的
标识要求。该条例中条款4第1-5条规定了含有或包括生物的产品的可追踪性要求,
条款5规定了生产的转基因生物的食品或饲料的可追踪性要求。
(11) 许可持有人应提供每年环境效应监测计划的实施和执行结果报告。这些
结果提交时应依据2009年10月13日2009/7700/EC欧盟委员会决议中建立的转基
因生物有目的环境释放的标准报告格式以及欧洲议会和欧洲理事会颁布的
2001/18/EC中对转基因产品市场投放的指令。参照第1829/2003号条例(EC)第6
(5)和18(5)条款要点(e),EFSA的意见并非强加该产品投放市场的特殊条
件或限制,和/或使用及加工的特殊条件或限制,包括食品和饲料使用的监测要
求或系统/环境和/或地带保护的特定要求。
(12) 根据第1829/2003条例(EC),所有和产品批准相关的信息必须在欧盟转
基因食品和饲料注册委员会进行立案。
(13) 根据欧洲议会和欧盟理事会在2003年7月15颁布的有关于转基因生物的越
境活动的第1946/2003号条例(EC)第9(1)和15(2)(C)条,这个决定将会
经由生物安全情报交换所(Biosafety Clearing-House)通知至卡塔赫纳生物多样
性生物安全议定书成员(parties to the Cartagena Protocol on Biosafety to the
convention on Biological Diversity)。
(14) 已经就此决议中提到的措施征询申请人意见。
(15) 在食物供应链和动物健康常务委员会主席规定时间内,食物供应链和动

164
物健康常务委员会没有提出异议时,委员会需要提交一份相应的实施方案。
(16) 因为在2011年12月15日的会议中,理事会无法达成绝大多数或者同意或
者反对这项提议的决议,理事会表示对这份文件的议程已有结论,这些措施将被
联盟采用。
已经同意本决议:
第1条
转基因生物和特殊识别号
根据本决议的附件细则(b)和第65/2004号条例(EC),转基因棉花281-24-
236×3006-210-23的专一识别号为:DAS-24236-5×DAS-21Ø23-5。
第2条
批准
为满足第1829/2003号条例第4(2)和16(2)条款(EC),基于决议中所列
情况,批准以下产品:
(a) 含有、包括或者由DAS-24236-5×DAS-21Ø23-5棉花产生的食品或食品原
料;
(b) 含有、包括或者由DAS-24236-5×DAS-21Ø23-5棉花产生的饲料;
(c) 除食品、饲料外,其他含有、包括DAS-24236-5×DAS-21Ø23-5棉花的产
品除了栽培以外具有和其他棉花相同的用途。
第3条
标识
1. 为了满足条例1829/2003(EC)第13(1)和第25(2)条款以及第
1830/2003号条例(EC)第4条(6)条款的标识要求,生物的名称应为“棉花”。
2. 根据条例2(b)和(c),含有或者包括DAS-24236-5×DAS-21 Ø 23-5棉花
产品的标签及附带文件上必须有“不能用于栽培种植”的标识。
第4条
监测环境影响
1. 根据附件中细目(h),许可持有者应该确保环境效应监测计划的落实和实
施。
2. 根据决议2009/770/EC,许可持有者应向委员会提交关于监测计划实施和执
行结果的年度报告。
第5条
欧盟登记
根据第1829/2003条例第28条款(EC),本决议附件中提到的资料必须在欧

165
盟转基因食品和饲料注册委员会进行项目登记。
第6条
审批持有人
本许可持有人为陶氏益农欧洲英国分公司, 代表美国Mycogen种业。
第7条
有效期
本决议在通知日后的十年内有效。
第8条
地址
此决议发送至陶氏益农欧洲分公司地址:European Development Centre, 3
Milton Park, Abingdon, Oxon OX14 4RN, United Kingdom.
2011年12月22日于布鲁塞尔完成。
致欧盟委员会
John DALLI
欧盟委员会成员

166

167

168

169

170
2. 美国农业部(USDA)批准证明文件
Irene Gatti博士
法规事物主管-生物技术
Dow AgroSciences LLC
9330 Zionsville Road
Indianapolis, IN 46268
收到日期:2004年7月16日
登记日期:2004年7月20日
尊敬的Gatti博士:
您编号分别为03-036-01p和03-036-02p,要求确定转基因棉花DAS-24236-5和
3006-210-23非监管状态的申请于2004年7月15日得到批准。附件中是签字的环境
评估(Environmental Assessment, EA)和没有显著影响裁决(Finding of No
Significant Impact)的副本,还有对环境评估和申请意见的总结和回复,并附有
涵盖两个申请的监管决定声明。这些文件很快也将在生物技术法规部
( Biotechnology Regulatory Service, BRS ) 网 页
(http://www.aphis.usda.gov/brs/ea_pubs.html)上公布。如果有任何疑问,请联系
生物技术法规部监管分部的秘书Terry Hampton女士[(301)734-5715]。由于Kay
Peterson女士已经接受了美国农业部动植物卫生检验局(The Animal and Plant
Health Inspection Service, APHIS)的新职位,因此她不再处理与申请相关的文件。
如果申请人注意到任何与这些申请书中描述的信息以及与我们的环境分析
有实质不同的信息,必须在5日内用书面形式告知美国农业部动植物卫生检验局。
我们已经起草了一份将在联邦登记(Federal Register)公布的公告,告知公众这
些转基因棉花转化体及其后代不再受7 CFR 340法规监管的决议。届时将会通知
您具体公布日期。
您真诚的
Cindy Smith
副执行长
生物技术法规部
美国农业部动植物卫生检验局
收到日期:2004年7月26日
登记日期:2004 年 7 月 26 日

171
关于Mycogen /Dow公司编号为03-036-01p和03-036-02p要求确定转基因抗虫棉
281-24-236和3006-210-23(分别表达合成的B.t. Cry 1F和Cry1Ac)非监管状态
申请的审批
环境评估(EA)以及无显著影响的裁决
2004年7月
美国农业部动植物卫生检验局(Animal and Plant Health Inspection Service,
APHIS)根据法规7 CFR 第340部分的条款,在Mycogen Seeds c/Dow AgroSciences
LLC(Mycogen /Dow)公司编号为03-036-01p和03-036-02p要求确定非监管状态
的申请获得批准之前,美国农业部动植物卫生检验局已经准备了一份环境评估报
告。这两个申请涉及的是通过遗传改造在转基因棉花转化体281-24-236和3006-
210-23中分别表达合成的杀虫蛋白Cry1F和Cry1Ac,从而对某些鳞翅目害虫产生
抗性;同时也通过遗传改造表达了用于转化的选择标记蛋白膦丝菌素乙酰转移酶
(PAT),从而对除草剂草铵膦产生抗性。在2004年3月9日发表在《联邦登记公
报》的公告中(69 FR 10972-10973, Docket No. 04-010-1),APHIS公布了陶氏益
农公司的申请书和环境评估来征集公众的意见和评论。在规定的60天的评阅期内,
农业部动植物卫生检验局收到了6份评阅意见。农业部动植物卫生检验局对这些
意见的分析和回复见本裁决的附件。根据环境评估的分析以及我们对评阅意见的
回复,农业部动植物卫生检验局作出这两个转基因品系及其后代(包括这些品系
的杂交后代)对环境没有显著影响的裁决,并且不再处于监管状态。本裁决作为
附件E附于环境评估报告。
Cindy Smith 副执行长 生物技术法规部 美国农业部动植物卫生检验局 2004 年 7 月 15 日

172

173

174
3. 美国环保署(EPA)批准证明文件

175

176

177

178
4. 美国食品与药物管理局(FDA)批准证明文件
尊敬的Gatti博士:
本函旨在对陶氏益农公司(DAS)旗下Mycogen Seeds 公司寻求美国食品与
药品管理局(FDA)(食品安全和应用营养中心以及兽医药中心)就其基因修饰
的转基因棉花DAS-21Ø23-5咨询进行的回复。根据DAS公司提交的资料,3006-210-23转基因棉花通过基因改造表达了两个新蛋白,即Cry1Ac和草丁膦乙酰转移
酶(PAT)。Cry1Ac蛋白赋予植物对鳞翅目害虫的抗性,PAT蛋白赋予植物对化
学合成的草丁膦产品(如草铵膦)的耐受性,并被用作筛选标记。DAS陈述有意
将Cry1Ac转基因棉花3006-210-23,与Cry1F转基因棉花281-24-236(另外提交一
份单独文件给FDA,被指定为BNF0085)以复合性状产品销售。所有与本通告相
关的材料均归档于编号BNF0092的文档中。该文档将由食品添加剂安全办公室保
存。 作为对该产品咨询作出结论的一部分,DAS于2003年3月18日提交了该基因
改造棉花安全性和营养评估的总结报告,并分别于2003年5月30日、6月27日和9月25日提交了额外的说明材料。这些信息交流让FDA 了解到DAS为确保本产品
符合FDA 管辖范围内法律、法规的要求而采取的步骤措施。根据DAS进行的安
全性和营养评估,我们的理解是:DAS总结该新品种衍生的棉籽、棉仁、棉粕、
棉籽油和棉籽壳与目前市售的棉籽、棉仁、棉粕、棉籽油以及棉籽壳在成份、安
全性和其他相关参数上没有实质差异,而且基因改造的转基因棉花3006-210-23没有带来任何问题需要FDA在其上市前进行审核或批准。
因为美国环境保护署(EPA)监管杀虫剂的物质和非活性成分,FDA不对
Cry1Ac和PAT蛋白的安全性信息进行评价。DAS有责任在3006-210-23棉花衍生
的食品或饲料上市前,获得所有相关的批准,包括EPA和美国农业部的批准。 根据DAS提交给FDA的资料,目前我们对转基因棉花3006-210-23衍生的棉
籽、棉仁、棉粕、棉籽油以及棉籽壳没有其它问题。不过,诚如贵公司所知,DAS仍有持续责任确保公司在市场上销售的产品是安全且健康的,并且符合所有相关
法律法规的要求。 您真诚的 Laura M. Taratino博士 主管 食品安全和应用营养中心 食品添加剂安全办公室

179

180
5. 巴西国家生物安全技术委员会批准证明文件
巴西国家生物安全技术委员会关于转基因WideStrike棉花的商业化技术意见
(1757/2009号)
技术意见号:1757/2009 申请编号: 01200.005322/2006-55 申请人:陶氏益农工业有限公司。 CNPJ : 47.180.625/0001-46 地址:Rua Alexandre Dumas, 1671, 1º Andar, Ala A, 04717-903 圣保罗省圣保罗市。 事宜:转基因棉花的商业化。 前次纪要:749 /2006发表于2006年9月29日。 会议:2009年3月19日第121 CTNBio定期会议。 决定:批准。
巴西国家生物安全技术委员会(CTNBio)审查了以下有关转基因抗虫和耐
草铵膦棉花商业化的技术申请,该棉花包括转化体281-24-236/3006-210-23的后
代及其与非转基因品种杂交产生的后代,以及带有这两个转化体的群体,根据技
术意见的各项条款决定予以批准。
陶氏益农工业有限公司请求CTNBio对转基因抗虫和耐除草剂棉花
WideSTrike(Gossypium hirsutum)的安全性提出技术意见。其目的是为了在符合
巴西境内棉属栽培品系均适用的所有其它规定和要求的前提下,使该转基因棉花
能够自由注册,并应用于环境、人类和动物消费、销售和工业用途以及,与该转
基因棉花相关衍生品系、栽培品种和副产品的其它用途。WideStrike棉花是由两
个转化体281-24-236和3006-210-23通过基因叠加技术培育而成的,转化体281-24-
236含有编码杀虫毒素Cry1F蛋白的cry1F基因,且转化体3006-210-23中含有表达
杀虫毒素Cr1Ac蛋白的cry1Ac基因。这两者均为杀虫结晶蛋白,也被称为D-内毒
素,分别来源于苏云金芽孢杆菌aizawai亚种 菌系PS811和亚种菌系HD73 。除了
由基因cry1F和Cry1Ac赋予的抗虫特性外,WideStrike棉花还具有耐草铵膦除草剂
的特性,这是因为含有两个拷贝的编码膦丝菌素乙酰转移酶(PAT) pat基因。
pat基因是依据链霉菌viridochromogenes中的自然pat基因合成的,该细菌是土壤
中广泛存在的一种非致病菌。使用含有的Pat基因可以进行植株筛选。 (技术资料部分略去)
Walter Colli 巴西国家生物安全技术委员会主任

181

182

183
6. 加拿大卫生部(Health Canada)食用批准证明文件
加拿大卫生部(Health Canada)
保健产品和食品管理局 Tunney’s Pasture
Ottawa, Ontario K1A 0L2
2005年7月29日 Deborah Straw 女士 法规事物主管 陶氏益农加拿大公司 1144-29- Avenue N.E Suite 201 Calgary, Alberta T2E 7P1
尊敬的Straw女士,
本函内容是关于抗虫棉花Bt Cry1Ac (转基因棉花3006-210-23)和Bt Cry1F(转
基因棉花281-24-236)的新食品申请。保健产品和食品管理局官员已经审查了由陶
氏益农加拿大公司提供的资料,有关含转基因棉花3006-210-23和281-24-236的食
品在加拿大以人类食品销售的可行性评估。
根据提交的资料,转基因棉花品系3006-210-23和281-24-236的研发程序涉及
两组不同基因表达盒的导入。转基因棉花3006-210-23通过导入编码Cry1Ac蛋白
的基因以及编码草丁膦乙酰转移酶蛋白(PAT)的基因获得。Cry1Ac蛋白的表达
能使棉花抵抗鳞翅目害虫,如烟草夜蛾、棉铃虫及红铃虫等。由于转基因操作的
结果,转基因棉花品系3006-210-23含有以下的新成分:
1) 一个拷贝的人工优化合成的cry1Ac基因(cry1Ac(synpro));
2) 由cry1Ac(synpro)编码序列表达的Cry1Ac蛋白;
3) 一个拷贝的人工优化合成的pat基因,基于绿色产色链霉菌基因(pat)的序列;
以及
4) pat编码序列表达的PAT蛋白。
转基因棉花品系281-24-236是通过导入Cry1F蛋白的编码基因以及草丁膦乙
酰转移酶蛋白(PAT)的编码基因获得。Cry1F蛋白的表达能使棉花抵抗鳞翅目
害虫,如烟草夜蛾等。由于转基因操作的结果,转基因棉花281-24-236含有以下
的新成分:
1) 一个拷贝的人工优化合成的cry1F基因(cry1F(synpro));
2) 由cry1F(synpro)编码序列表达的Cry1F蛋白;

184
3) 一个拷贝的人工优化合成的pat基因,基于绿色产色链霉菌基因(pat)的序列;
以及
4) pat编码序列表达的PAT蛋白。
基于对所提交资料的评估,我们对在加拿大销售含有转基因棉花3006-21-023
和281-24-236的食品没有异议。
需要注意的是,本批准意见仅基于销售含有转基因棉花3006-21-023和281-24-
236的商业品系作为人类食品的适合性。陶氏益农加拿大公司有持续的责任确保
公司的产品符合所有适用的法令和法规要求。如获得任何对健康和安全有潜在影
响的新信息,应当汇报给加拿大卫生部考虑,以确保加拿大市场上所有新食品有
持续的安全性和可靠性。销售对消费者健康有危害的食品将违反《食品和药品法》
的规定。
请注意我们将提供本涵复本给加拿大食品监督局(CFIA)的同事,因该部门负
责品种登记、动物饲料、环境释放和标识等事宜。我们也提供本涵复本给害虫
防治与管理监管局(PMRA)的同事,以供该部门参考。 您真诚的 (签名) Paul R. Mayers 总监 加拿大食品管理局 抄送:G. Orriss 先生, CFIA K. Dodds 博士, PMRA

185

186

187

188
7. 加拿大食品监督局(CFIA)批准证明文件
动物健康和生产处 加拿大食品监督局(CFIA) 59 Camelot Dr., Ottawa, Ontario K1A 0Y9 Deborah Straw 法规事物主管-植物遗传 陶氏益农加拿大公司 201, 1144 29th Avenue NE Calgary, Alberta T2E 7P1 2005 年 7 月 28 日 尊敬的 Straw 女士:
饲料部 (Feed Section)已经审阅了陶氏益农加拿大公司关于转基因棉花 3006-
210-23 作为牲畜饲料用途的申请。转基因棉花 3006-210-23 因为介导入来自苏云
菌芽孢杆菌的 cry1Ac 合成基因而赋予抗虫性。
遵照《饲料管理法规》(Feeds Regulations)的相关规定, 陶氏益农加拿大公司
依据管理条例 Dir95-03 “植物来源的新饲料评估的指导方案”提交了相关资料。评
估后认为该棉花品系 3006-210-23 与常规棉花品系具有等同的营养性和安全性;
因此棉花品系 3006-210-23 及其产品符合现今营养成分定义的内容,批准其在加
拿大作为畜牧饲料成分之用。
遵循法规监管部门间批准程序的协调,CFIA 获知加拿大卫生部已总结棉花品
系 3006-210-23 的衍生产品可供人类安全食用。
经《饲料管理法规》第 4.3 部分(Feeds Regulations,section 4.3)授权, CFIA
批准棉花品系 3006-210-23 及其任何后代品系可在加拿大作为牲畜饲料之用,若
满足以下条件:
不得进行种间杂交;
用途与所申请的内容类似;
通过周密的特性鉴定已知植物不显示新增加的性状,并且就潜在环境影响和
牲畜饲料安全性方面与目前用于牲畜饲料的棉花具有实质等同性;以及
应CIFA要求提供支持棉花品系3006-210-23衍生品系具实质等同性的分析数
据。
陶氏益农加拿大公司已向CFIA提交检测及鉴定测试方法。基于提交信息的审
核,该方法对棉籽中Cry1F蛋白的检测似乎可行。陶氏益农加拿大公司已经同意
提供CFIA合适的参照材料,应要求将与CFIA合作以帮助这些方法能够在CFIA实

189
验室达到完整功能。
在任何时候,如果陶氏益农加拿大公司获知任何有关棉花品系3006-210-23
或其衍生品种的释放造成环境、牲畜或人类健康风险的新信息,按照《饲料管理
法规》第4.4(1)部分的规定,陶氏益农加拿大公司必须立即向CFIA提供这些信息。
CFIA将根据这些新的信息对棉花品系3006-210-23用作牲畜饲料及其对环境及人
类健康可能产生的影响和风险重新进行评估。CFIA可能会在牲畜饲料的使用或
释放情况方面增加附加条件,也可能会改变批准的条件,或是吊销批准并要求陶
氏益农加拿大公司停止其释放及牲畜饲料之用,并采取相应的必要措施以消除或
降低风险。
一份解释棉花品系3006-210-23批准依据的判决文件将会在批准后30天公布。
CFIA也会将此批准决定通知其他联邦政府部门和省级机构。 您诚挚的, (签名) Linda Morrison 饲料部全国经理 抄送:Dr. Stephen Yarrow, 加拿大食品监督局 (Plant Biosafety Office,PBO植物
生物安全 办公室) Dr. William Yan, 加拿大卫生部 (新食品部) Dr. Richard Aucoin, 害虫防治与管理监管局 Dr. Jim Louter, 加拿大环保局(生物技术部) Dr. Liz Foster, 加拿大食品监督局(实验室总监) 各省联系人:Marney James (British Columbia省) Dale Armstrong (Alberta省) Bill Greuel (Saskatchewan省) Lynn Manaigre (Manitoba省) Bill Ingratta (Ontario省) Bruno Maltais (Quebec省) Gerald Chevrier (New Brunswick省) Tony Sturz (Prince Edward Island省) Claude Caldwell (Nova Scotia省) Goldie Porter (Newfoundland 省)

190

191
192
8. 日本厚生劳动省(MHLW)批准证明文件
生物已通过重组DNA技术来源的食品和食品添加剂安全性评价程序的公告
日本厚生劳动省(MHLW)公告第393号
依据重组DNA技术来源的食品和食品添加剂安全评价程序第3-4条规定
(2000年厚生省公告第233号)对下述使用重组DNA技术生产的生物已经,通
过食品与食品添加剂第1A-3号标准(1959年厚生省370号公告)规定的安全性评
价审查程序予以公告。 2005年9月5日 厚生劳动省部长
已通过重组DNA技术来源的食品和食品添加剂安全性评价程序的生物: 作物 名称 申请人 棉花 棉花281 陶氏化学日本 棉花 棉花 3006 陶氏化学日本

193

194
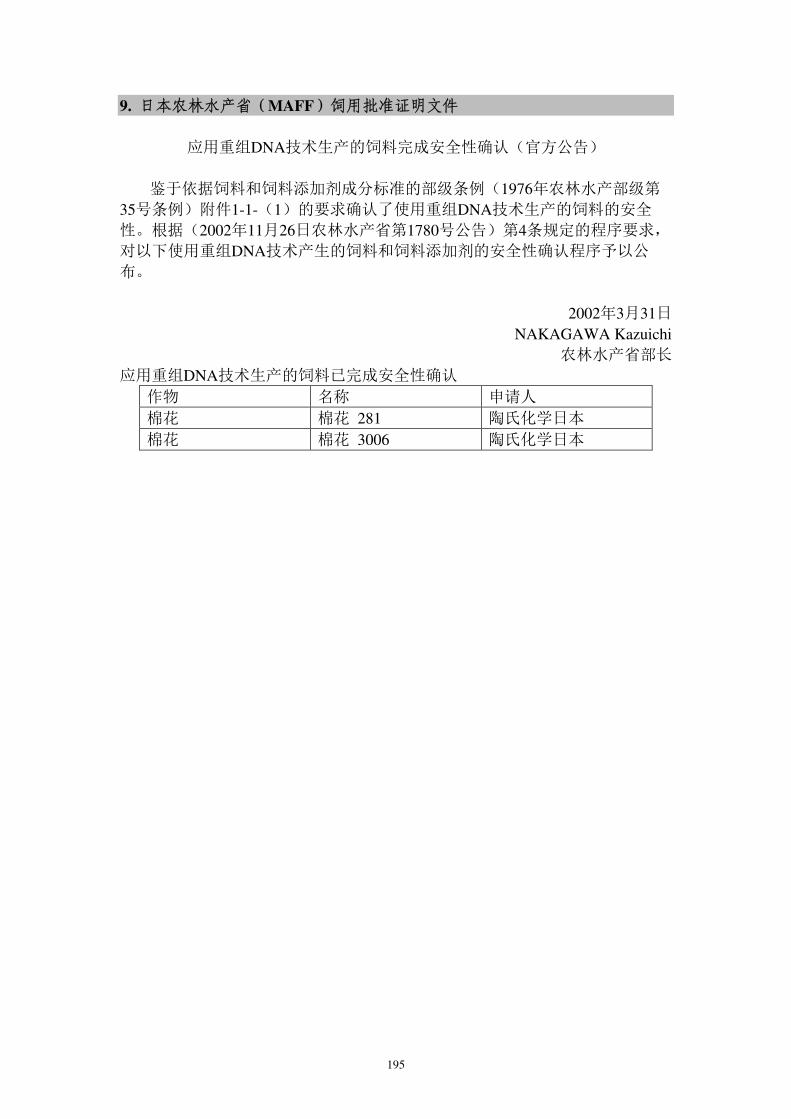
195
9. 日本农林水产省(MAFF)饲用批准证明文件
应用重组DNA技术生产的饲料完成安全性确认(官方公告)
鉴于依据饲料和饲料添加剂成分标准的部级条例(1976年农林水产部级第
35号条例)附件1-1-(1)的要求确认了使用重组DNA技术生产的饲料的安全
性。根据(2002年11月26日农林水产省第1780号公告)第4条规定的程序要求,
对以下使用重组DNA技术产生的饲料和饲料添加剂的安全性确认程序予以公
布。
2002年3月31日 NAKAGAWA Kazuichi
农林水产省部长 应用重组DNA技术生产的饲料已完成安全性确认
作物 名称 申请人 棉花 棉花 281 陶氏化学日本 棉花 棉花 3006 陶氏化学日本

196

197
10. 日本农林水产省环境法令
2004 消费者产品安全法第459 号
环境法令060410001 2006 年 4 月 10 日
陶氏益农日本 商业主管Monty Bayer
农林水产省部长NAKAGAWA Kazuchi 环境代理临时代理部长 NIKAI Toshihiro
关于生物技术改良生物法以保持生物多样性, 针对批准一个品种受法规监管使用规则条款 根据法律条款(97 号法案,2003),该条款适用于生物技术改良生物以保
持生物多样性,批准附件中所描述的一个转化事件品种受监管使用的申请,批
准于2004 年12 月10 日生效。 附件 批准号 06-46P-0013

198

199
11. 韩国食品药品管理局(KFDA)批准证明文件
韩国食品药品管理局
收件人:陶氏益农公司
主题:转基因棉花281/3006审查结果通知
1.这是关于编号3申请(2004年2月16日)的回复。
2.贵公司要求审查转基因棉花281/3006是否适用于 “转基因食品安全性评价指
南(公告第2003-37号)”第4条第一段所涵盖的安全评估。
3.转基因食品安全评价委员会已经证实转基因棉花281/3006作为食品与传统棉
花相比并未表现出任何改变。
附件:转基因食物安全性评价审查结果通知报告
委员,韩国食品药品管理局 (官印)
负责人:Soon Ho Lee 研究员:Sun Hee Park 营养评价组组长:Hye Kyung
Park
发件人:营养评价组-201(2006年2月2日)
地址:#5 Nokbun-dong, Eunpyung-Gu, Seoul 122-704 /www.kfda.go.kr
电话:02-380-1678 传真02-380-1358

200
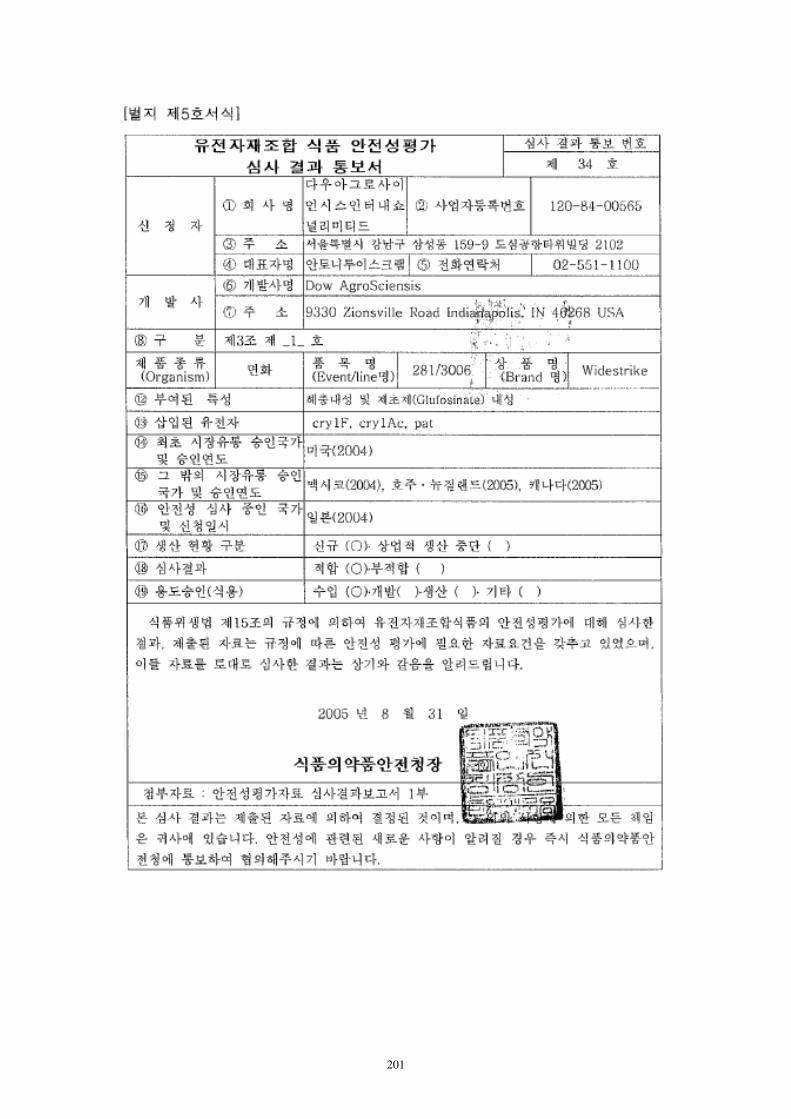
201
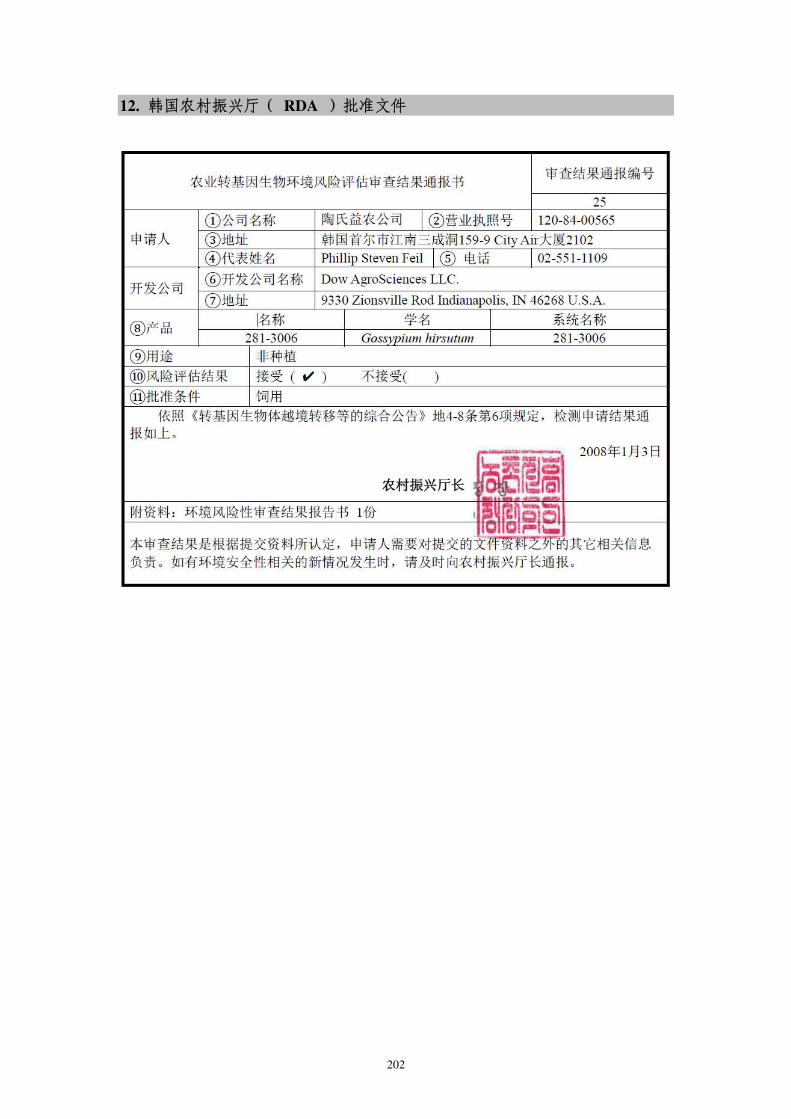
202
12. 韩国农村振兴厅( RDA )批准文件

203

204
13. 墨西哥卫生部批准文件

205
14. 澳大利亚和新西兰食品标准局(FSANZ)批准文件
Matthew Cahill 博士 陶氏益农公司 Locked Bag No. 502 P.O.Frechs Forest N.S.W. 1640
尊敬的Cahill博士
A518号申请-抗虫耐除草剂棉花产品MXB-13来源食品
澳大利亚新西兰食品标准管理局(FSANZ)通知您如下事项:
澳大利亚和新西兰食品法规部际联席会议(the Council)决定,FSANZ完成
了对抗虫耐除草剂棉花产品MXB-13来源食品草案的审查,并同意批准A518号申
请;
作为A518号申请的批准结果,FSANZ将于2005年4月28日公布相应的食品
标准草案变更(第77号修订)。
《食品标准公告》的复印件可从FSANZ的网站下载:
http://www.foodstandards.gov.au/standardsdevelopment/gazettenotices/index.cfm.
如您对上述问题有任何疑问,请联系标准管理官员Cathie Humphries女士。
联系电话:+61 2 6271 2280;电子邮箱:[email protected].
您诚挚的, MELANIE FISHER 食品标准主管(堪培拉) (《食品标准公告》翻译略)

206

207
15. 澳大利亚转基因技术管理办公室(OGTR)批准证明文件
2009年11月25日 决议书
签发陶氏益农公司关于商业化种植转基因棉花的许可DIR091
转基因技术管理办公室决定签发关于陶氏益农澳大利亚公司(陶氏)申请
DIR091的许可。申请者已获得允许商业化种植转基因抗虫棉WideStrike™的批
准。此许可准许在南纬22o以南现有所有植棉区种植转基因棉;以及在整个澳大
利亚,包括澳大利亚北部采取措施限制转基因生物潜在的扩散及存留的地区使
用转基因棉籽做动物饲料。
签发此许可的决议是根据2000年转基因技术法案和国家和地方法律的要
求,就风险评估和风险管理计划与公众,国家和地方政府,澳大利亚政府机
构,环境,遗产和艺术部,转基因技术科学顾问委员会以及所有相关地方议会
充分的商议后做出的。
最终的风险评估和风险管理计划和签发此许可的决定考虑到了商议过程中
提出的与人类健康和安全以及环境保护方面的问题。
管理概要,技术概要,最终的风险评估和风险管理计划,以及对此决定的
一系列问答和此许可,可通过转基因技术管理办公室的网站或通过以下联系方
式获得。
转基因技术法规办公室 MDP 54 GPO 信箱 9848 CANBERRA ACT 2601
电话:1800 181 030, 传真 02 6271 4202, 网址:http://www.ogtu.gov.au

208

209
16. 哥伦比亚农业研究所(ICA)批准文件
第002664号决议号
2015年7月27日 关于批准陶氏益农公司3006-210-23(DAS-21023-5)棉花用于动物饲料及其加
工原材料 哥伦比价农业研究所所长, ICA
行使1992年第2141号政令, 1994年1840号政令,2005年第4525号政令,2008年第4765号政令所赋予的法定特别权利,
鉴于:
国家政府,根据2002年第740号法律,颁布2005年第4525号政令,授权农业与
农村发展部通过哥伦比亚农业研究所(ICA)审批改性活体生物(LMOs)的越境转
移、运输、处理和使用,以用于农业、畜牧业、渔业、商业森林种植和农副业用
途,因为这些产品可能对于生物多样性的保护和利用造成负面影响。
2005年第4525号政令根据2002年第740号法律所规定的程序制订了改性活体
生物(LMOs)的监管框架,并成立专门针对农业、畜牧业、渔业、商业森林种植和
农副业用途的LMOs的国家生物技术委员会,其职能包括在行政文件的签发中向
ICA所长提供建议,以审批与改性活体生物相关的活动。
2006年4月17日实施的946号决议确立了ICA办理申请解除应用于农业、畜牧
业、渔业、商业森林种植和农副业用途的改性活体生物限制的流程。
Dow Agrosciences de Colombia S.A.公司, 依法向ICA申请批准应用3006-210-
23(DAS-21023-5)棉花作为动物饲料及其加工原材料。
3006-210-23 (DAS-21023-5)棉花通过基因工程方法, 经农杆菌质粒插入
cry1Ac基因及pat基因。
经试验与非转基因对照相比,3006-210-23棉花在营养成分上未表现明显差异。
3006-210-23棉花的营养品质与其非转基因对照和其他商业品种等同。
无试验证据表明新表达的蛋白会对人类或动物产生致毒或者致敏的影响。
3006-210-23棉花表达Cry1Ac和PAT蛋白,使其能够抵御一些害虫的侵害并耐
受除草剂草铵膦, 且已有安全使用的历史。
Cry1Ac和PAT蛋白已被详尽地研究,且无任何证据表明这些蛋白对动物或人
类致毒。
3006-210-23棉花表达的Cry1Ac和PAT蛋白对人类或动物健康几乎不可能产
生致敏或不利影响。
转基因技术或表达Cry1Ac和PAT蛋白没有改变通常在棉花植物体中的抗营
养成分。
3006-210-23棉花已在其他国家批准共人类和动物食用,并未发现有害的结果。

210
2015年5月26日召开的全国生物安全技术委员会(CTNBio)第二十四次会议
中,由环境和可持续发展部、卫生与社会保护部、农业与农村发展部、ICA以及
科技和创新管理部参加,对风险评估结果经协商一致认为推荐ICA批准应用3006-
210-23(DAS-21023-5)用作动物饲料及其加工原材料。
根据上述情况:
决议:
条款 1-批准 Dow AgroSciences de Colombia S.A.公司申请应用 3006-210-23(DAS-
21023-5)用作动物饲料及其加工原材料。
条款 2-出于生物安全性方面的考量,ICA 可以在认为需要的情况下取消本决议,
为此无需征求其他方面意见,其他方面也无权提出任何赔偿。
条款 3- 应用 3006-210-23(DAS-21023-5)用作动物饲料及其加工原材料的公司必
须遵守如下义务:
1. 遵守 2005 年 4525 号政令及其他现有相关标准
2. 如此决议强调的,3006-210-23 棉花只能用作动物饲料及其加工原材料,其他
用途必须经相关部门批准。
3. 允许 ICA 核查、监督、监控和提取必要的样品,以履行其职责。
4. 如发现在生物安全方面存在现实的或危机的风险或危害,应及时向 ICA 报告。
5. 在出现紧急情况时应及时有效的采取必要的缓解措施。
6. 满足农业产品方面的其他现行法规。
条款 4 -违反本决议的规定将会受到 ICA 根据相应法律作出的行政处罚,不影响
ICA 行使生物安全性的其他职权。
条款 5 – 本决议将根据 2005 年第 4525 号政令的规定公示在哥伦比亚农业研究
所 ICA 的网站上:
www.ica.gov.co.
条款 6 – 按照行政程序和行政诉讼法(2011 年第 1437 号法律)第 67-69 条的规
定将本行政文件作出通告。
条款 7 – 如针对本决议申请再审,需根据行政程序和行政诉讼法(2011 年第 1437
号法律)第 76 条的规定,在本通知后的十(10)个工作日内提出申请。
条款 8 – 本决议自签发之日起开始生效。
特此通告并遵守。 签发于波哥大,2015 年 7 月 27 日
JUAN ANDRES ANGULO MOSQUERA 所长

211

212
17. 哥伦比亚卫生和社会保护部(INVIMA)批准文件
卫生和社会保护部
2016 年第 005853 号决议
( )
授权棉花 3006-210-23(DAS-21ø23-5)用作食品或者食品加工原料供人类消费
卫生和社会保护部
行使监管权力,尤其是 2015 年第 1071 号令第 2.13.7.3.5 条赋予的权力,以及
涉及到的
《联合国生物多样性公约》,该公约采纳于 1992 年 6 月 5 日,并由哥伦比亚
政府通过 1994 年颁布的第 165 部法律批准,由宪法法院通过 1994 年的 C-519 决
议强制执行。
《生物多样性生物技术安全公约卡塔赫纳议定书》,该议定书采纳于 2000 年
1 月 29 日,在哥伦比亚通过 2002 年政府颁布的第 740 部法律批准,由宪法法院
通过 2003 年的 C-071 决议强制执行。
2005 年第 4525 号令(包含在 2015 年第 1071 号令),建立了活体转基因生物
(LMO)的监管框架。农业、渔业和农村发展部门的独有法规,包括修改的 2015 年
的第 1298、1449、1565、1648、1780、1934、2020、2179 和 2537 号令,2016 年
的第 13、440 和 947 号令,在第二册第 13 编第 3 章第 7 节里,对上述监管框架
作了编辑。
上述文件通过社会保护部(Social Protection Ministry)发布的 2007 年第 227 条
决议成立,社会保护部的前身是活体转基因生物(LMO)国家生物安全技术委员会,
这里的活体转基因生物专指应用于健康和人类消费的活体转基因生物。从此以后,
CTNSalud 由社会保护部、现在的卫生和社会保护部或其代表、国家食品药品监
督局(INVIMA)或其代表以及 Colciencias 主任或其代表整合而形成。
CTNSalud 的工作职能之一,是对向该办公室提议的活体转基因生物(LMO)
发展活动发放行政授权书。
位于波哥大特区,识别号为 T.I.N. 800,087.795-2 的哥伦比亚陶氏益农股份公
司 (DOW AGROSCIENCES DE COLOMBIA S.A.)的法定代表人的 José Cooz
Sánchez 博士,在与国家食品药品监督局(INVIMA)的通信(文件编号 12103782,
2012 年 12 月 26 日)中,请求授权转抗虫基因 B.t. Cry1Ac 的抗虫棉 3006-210-23
(DAS-21ø23-5)用作食品或者食品加工原料供人类消费(FIs. 19,37)。
请求授权转抗鳞翅目昆虫基因的棉花 3006-210-23(DAS-21ø23-5)用作食品
或者食品加工原料供人类消费的上述社会团体,提供了有关的风险评估和食品安

213
全的支持性文件,并作了分析说明,CTNSalud 又作了进一步的分析说明(FIs. 13-
17),详见下述相关部分:
1. 据在 2013 年 8 月 20 日召开的国家生物安全技术委员会的特别会议上第
4 部法案(FIs. 23-29)记录,对上述社会团体提交的申请作了分析说明,并阐述了
附加信息的要求,认为:
a. 为了支持提交的摘要文件的全部研究,包括:
i. 新表达蛋白的致敏性。
ii. 新表达蛋白的毒性。
iii. 胃和肠液中的体外消化率。
iv. 对受试动物急性经口毒性。
v. 新表达蛋白的稳定性。
vi. 受试动物饲喂。
vii. 温度对转化体中新表达蛋白的影响(FIs. 27 和 28)。
b. 通过国家食品药品监督局(INVIMA),在国际检测和监测转基因生物中心
实验室对所请求的该特定转化体的适用性和有效的检测方法(FI. 28)。
c. 当国际检测和监测转基因生物中心实验室通过国家食品药品监督局
(INVIMA)提出要求时,该社会团体可以提供该转化体的证明材料(500 g 纯度为
100%的转基因生物样本),对此提供了相关的说明性文件。
2. 据在 2015 年 12 月 4 日召开的 CTNSalud 特别会议上第 4 部法案(FIs. 31-
36)记录,分析了请求方通过 2015 年 10 月 22 日与国家食品药品监督局(INVIMA)
的通信提出的信息,即通信相关文件编号为 15112351(FIs. 21 和 22),以及哥伦
比亚陶氏益农股份公司对棉花转化体 3006-210-23 (DAS-21ø23-5)做出的风险评
估结果,从中发现:
a. 棉花转化体 3006-210-23 (DAS-21ø23-5),是由被称为 GC510 的 Acala 棉花种
质系转基因改造而来,GC510 是 Germain’s Agribusiness Inc.于 1984 年在美国发
布的(FI. 14)。
b. 基因转化是运用携带 cry1Ac 和 pat 基因的卸甲质粒 pMYC3006 通过农杆菌
(Agrobacterium tumefaciens)转化法完成的。cry1Ac基因是从苏云金芽孢杆菌(B.t.)
kurstaki 亚种的 HD-73 菌株中分离而来,pat 基因(用作可选标记)是从绿色产
色链霉菌(Streptomyces viridochromogenes)中分离而来(FI. 14)。
c. 合成原毒素 cry1Ac(synpro)是一种杀虫蛋白晶体(ICP:也称做“δ-内毒素”),其
核心毒素最初是在苏云金芽孢杆菌 kurstaki 亚种的 HD-73 菌株中发现的。引入该
基因的目的是为农民提供新的控制棉花鳞翅目有害昆虫的途径(FI. 14)。

214
d. Pat 基因作为选择标记,表达 PAT 蛋白,该蛋白具有抗化学合成的、含膦基化
合物的性能,例如草铵膦铵盐(FI. 15)。
e. Southern blot 证实了棉花转化体 3006-210-23(DAS-21ø23-5)中 cry1Ac 和 pat 基
因稳定地插入进植物基因组中,在棉花植株的组织中表达,并且世代稳定遗传。
f. Cry1Ac 以及 PAT 蛋白的供体生物,分别是苏云金芽孢杆菌和绿色产色链霉,
都不是已知的过敏原(FI. 15)。
g. 总之,对于棉花转化体 3006-210-23 (DAS-21ø23-5)中表达的 Cry1Ac 以及 PAT
蛋白的多种分析表明,这两种蛋白在食品中不存在任何潜在的致敏性(FI. 15)。
h. 为了评估来自用于食品消费的棉花转化体 3006-210-23 (DAS-21ø23-5)的
Cry1Ac 蛋白潜在的毒性风险,在动物上做了急性经口毒性试验。结果表明,所
有试验动物都没有出现病变。这也证明 Cry1Ac 蛋白对哺乳动物无毒。该结论也
可被消化研究中该蛋白被快速消化的结果所支持(FI. 16)。
i. 为了证明转 Cry1Ac 棉花,转化体 3006-210-23 (DAS-21ø23-5)与对应的非转基
因棉花等同,对包括矿物质、氨基酸、脂肪酸和抗营养因子等在内的组分进行了
分析(FI. 16)。
k. 棉籽组分分析结果显示:在转化体 3006-210-23 (DAS-21ø23-5)的棉花植株中
插入 cry1Ac 基因,表达产生 Cry1Ac 蛋白,与非转基因棉花相比,矿物质、氨基
酸、脂肪酸和抗营养因子等未发生改变。
国际食品法典委员会依据 2002 年第 740 部法律、2015 年第 1071 号令及其
修改和指南(CAC/GL 44-2003 和 CAC/GL 45-2003,分别于 2008 年和 2011 年
修订),同时考虑到授权使用用途,对该转基因棉进行了评估。
基于上述信息,CTNSalud 在 2015 年 12 月 4 日的特别会议上(法令 4)做
出以下决定:......建议卫生和社会保护部签发行政许可,授权转化体 DAS-21ø23-
5 可以直接用作食品或食品加工原料供人类消费(FIs. 31 和 33)。
上诉信息
得到的结论
第一条,授权给位于波哥大特区的哥伦比亚陶氏益农股份公司的申请,其识别号
为 T.I.N. 800,087.795-2,法人代表 MIRANDA DANIELLA FERRAZ DE SOUZA
博士,外侨登记号码为 491118,将棉花 3006-210-23 (DAS-21ø23-5)用作食品或食
品加工原料供人类消费。
该授权有效期为自本决议执行之日起 5 年,在全国范围内有效,当事人在授
权到期前至少提前 3 个月提出续申请,有效期可以再延长同等期限。但是,必须
认可《生物多样性生物技术安全公约卡塔赫纳议定书》附件 II 所需的文件,该议
定书通过 2002 年第 740 部法律获得通过和批准,也应遵守其修订或者替代的标
准。

215
第二条,如果出现由于使用哥伦比亚陶氏益农股份公司的棉花 3006-210-23
(DAS-21ø23-5)技术,对人类健康产生不良反应的报道,该社会团体就要改进递
交给 CTNSalud 的风险管理的各个阶段的文件。这样做的目的,是为了防止、避
免、减轻以及控制有可能发生的危害人类健康的不良反应的出现。
第三条,进口作为食品或食品加工原料用于人类消费的棉花 3006-210-23 (DAS-
21ø23-5)技术的进口商,必须遵守 2002 年第 740 部法律批准的卡塔赫纳议定书
第 18 条序号 2 段落 a)中的规定,以及 2011 年第 4254 号决议第 7 条的规定。
这一点是由当时的社会保护部发布,或来自于其修改或替代的标准。
第四条,依照当时的社会保护部的 2007 年第 1122 部法律第 34 条段落 b)和 c)
中的条款,以及 2011 年第 4254 号决议的第 9 条,国家食品药品监督局(INVIMA)
以及国土健康董事会(Territorial Directorates of Health),行使对作为食品或食品加
工原料供人类消费的棉花 3006-210-23(DAS-21ø23-5)技术检查、监督和控制职能。
根据社会保护部 1979 年第 9 部法律以及 2013 年第 1229 号决议,或者修改或替
代的标准,社会保护部有权采取安全措施,以及实施相应的制裁。
第五条,将这一决议的内容通知到 MIRANDA DANIELLA FERRAZ DE SOUZA
博士本人,哥伦比亚陶氏益农股份公司的法人代表,外侨登记号码为 491118,或
者依据《行政程序和行政诉讼法》第 67 和 68 条规定,告知被授权的人。
段落,依据《行政程序和行政诉讼法》第 69 条规定,如果无法通知到本人,应
发布公告。
第六条,依据《行政程序和行政诉讼法》第 74 条规定,推迟关于针对本项决议
反对动议的复议行为。
第七条,本决议自发布之日起实施,自执行之日起生效。
发布,通知和遵守
于波哥大特区的
ALEJANDRA GAVIRIA URIBE
卫生和社会保护部部长

216

217

218

219

220

221
18. 南非农林渔业部(DAFF)批准文件
农林渔业部遗传资源理事会
专用邮袋:X973,Pretoria 001
询问人: Refilwe Ngoepe·电话: 012 319 6364·传真: 012 319 6298·邮箱: [email protected]·相关
信息: 39.4.2/陶氏益农-14/015
K Snyman 先生 Dow AgroSciences Southern Africa (Pty) Ltd. Baobab House Eastwood Office Park 2988 Liz John West Street Lynnwood Ridge Pretoria 0081
传真:(012)361 8126
尊敬的 Snyman 先生:
转基因生物商品许可
依据 1997 年颁布的《转基因生物法》(1997 年第 15 部法案),您申请的转
基因棉花商品已获批准并签发许可,许可证号为 39.4(6/16/584)。
基于转基因法案授予的任何许可在任何条件下无论何时如果违反了 1997 转
基因生物法案条款(1997 法案第 15 项),你将被以 1997 转基因法案 21 款起
诉。
此致!
NL Mkhonza 女士

222
注册员:《转基因生物法》(1997 年第 15 部法案)
转基因生物商品授权许可证:棉花转化体 DAS-24236-5 × DAS-21023-5
许可证号:39.4(6/16/584)
根据 1997 年《转基因生物法》(1997 年第 15 部法案)第 2 条规定,以及下
列条款规定的条件,特此授权
进口方/进口商(更多信息的联络人): K Snyman 先生 Dow AgroSciences Southern Africa (Pty) Ltd Baobab House Eastwood Office Park 2988 Liz John West Street Lynwood Ridge Pretoria 0081 电话:+27(12)361 8120
使用下列转基因生物
通用名:棉花
学名:Gossypium hirsutum L. (陆地棉)
转化体:DAS-24236-5 × DAS-21023-5
基因:cry1F, cry1Ac 以及 pat
引入性状:抗虫性
唯一标识符:DAS-24236-5 × DAS-21Ø23-5
申请人须遵守签发许可证所附的所有条件。
时间: (签章) Mkhonza(签名)
注册员

223
以下承诺书由许可证持有人或授权代理人填写完成,并且须在收到许可证后的
一周内交回注册员办公室:
我,签署人……………………作为……………………为……………………(机构),
注册号……………………………………………………………………
单位地
址:………………………………………………………………………………………
………………………….
…………………………………………………………………………………………
…………………………………………………………………………………………
………………………………邮编: …………………………………………
完全知悉上述条件,同时明白如果违反或未准守上述任何条件,根据 1997 年
《转基因生物法》(1997 年第 15 部法案)规定,我将属于犯罪的情形并将受到起
诉。
签名:
时间:
许可证号码: 39.4(6/16/584)
交回注册员办公室:
联系人:Ms R Ngoepe
电话:+27 12 319 6364
传真:+27 12 319 6298
可以接受承诺书的传真件,同时应邮寄承诺书原件。
请注意:所有的许可证承诺书须送至我单位,否则所有新的许可证请求将不被
批准。

224

225

226

227

228

229
19. 中国台湾食品药品管理局(TFDA)批准文件

230
附件 11. 输出国家或地区经过科学试验证明对人类、动植物和生态环
境无害的资料
本公司针对 DAS-21023-5 棉花进行了大量研究,包括分子生物学特性分析、
遗传稳定性分析、环境安全性评估及食品安全性评估,检测报告英文原文和中文
译文见技术资料附件(报告 1~报告 11,第二~四册)。

231
附件 12. 转基因生物进口过程中拟采取的安全防范措施
在转基因抗虫棉花品系 DAS-21023-5 进口安全证书的使用方面,本公司将
采取下述安全防范措施来保证进口过程的安全。
根据《农业转基因生物安全管理条例》,农业农村部针对转基因生物进口的安全
管理制定了相应的进口安全管理办法,适用于在中国境内从事农业转基因生物进口
活动的安全管理。对于进口转基因生物材料,农业农村部按照用于研究和试验、用
于生产及用作加工原料三种途径实行管理。
本申请目的是为 DAS-21023-5 棉花获得进口用作加工原料的安全证书。为了
确保 DAS-21023-5 棉花获得安全证书以后,在实际粮食进出口贸易中符合中国法
规要求,不作规定范围外的其他使用、不扩散进入环境中,避免生物安全风险,本
公司将严格遵守中国相关法规及农业农村部对安全证书的要求。在向谷物贸易商提
供安全证书时,我公司有严格的证书申请流程并要求贸易商签署证书使用协议,
要求其严格遵守协议要求,在加工经营活动中不得改变用途并采取严格的安全控
制措施,确保不会对环境和人类健康带来任何风险。
(1) 向贸易商发放安全证书的安全管理
贸易商向本公司申请安全证书复印件时,我们将要求贸易商承诺遵守中国法律
并履行所签订的使用协议内容。
- 严格遵守中国粮食进出口贸易及安全证书相关的法律法规;进出口过程
中积极配合农业主管部门及相关行政机构的审查。
- 必须确保安全证书仅用于相关转化体进口到中国作为食物和饲料加工原
料使用。
- 贸易商必须采取合理措施,确保相关转化体进口到中国不被用于种植、
培育、繁殖、作物生产及研究目的。
- 贸易商必须采取合理安全管理措施,确保相关转化体进口到中国不会被
释放到环境中,或其他可能影响生物多样性或人体健康的行为。
- 如发现贸易商未遵守本协议,本公司保留收回安全证书使用的权利。
(2) 贸易商在进口过程中的安全防范措施
在实际的粮食进口过程中,本公司要求贸易商采取以下安全防范措施:
① 遵守中国法律法规:在进口过程中,严格遵守中国及出口国家的相关法
律和法规,这些要求包括检验检疫要求,以及中国政府在进口安全证书中提出的
要求和条件。
② 包装和标识:严格遵守中国及出口国家的相关法律和法规,如谷物包装
与标识有关的法律法规。

232
③ 运输环节:
i. 产地国内陆运输:本公司要求贸易商严格遵守中国及出口国家的相关
法律和法规,如根据法律法规要求,使用特定运输工具用以装载转基
因产品。
ii. 出口终端:本公司要求贸易商严格遵守中国及出口国家的相关法律和
法规, 如对指定的筒仓进行清洗,以避免与其他转基因生物的混杂。在
确认其出口装船之前,接受并积极配合出口国相关部门的检验。
iii. 装船:本公司要求贸易商严格遵守中国及出口国家的相关法律和法规,
如船只到达装货港之前进行清仓,对船仓和船上各作业区进行彻底地清
洗,请专门机构进行监督和检查,出具可证明船只清洁且安全的证书。
装货过程中有独立的检测机构负责监装,以确保装货过程在独立和公
正的检测下完成。
iv. 海运:运输全程中本公司要求贸易商严格遵守中国及出口国家的相关法
律和法规,如确保舱盖密闭,船方定期对仓内货物进行检查,规定船方
不得随意靠港,特别是可能存在疫情的港口。
v. 卸货:当船只到达中国口岸时,要求中国进口商严格遵守中国及出口国
家的相关法律和法规,执行转基因安全管理条例、农业转基因生物安全
评价管理办法以及中国农业农村部所制定的各项规法规与公告,避免在
港口卸货和中转的转基因产品流出。
④ 监测和报告:本公司要求贸易商严格遵守中国及出口国家的相关法律和法
规, 如果发现了任何违反规定操作和意外事件,将立即向当地农业行政主管部门和
农业农村部门报告,采取有效的措施来控制影响。
⑤ 技术指导和支持:在必要时,我公司会向贸易商提供技术指导和支持。

233
七、本单位农业转基因生物安全小组审查意见
转基因棉花 DAS-21023-5 中含有目标基因 cry1Ac 和标记基因 pat,cry1Ac
编码的杀虫蛋白质 Cry1Ac 使 DAS-21023-5 对主要害虫鳞翅目昆虫表现出杀虫
活性。自 2004 年以后,DAS-21023-5 棉花在美国、加拿大、日本、韩国、巴
西、欧盟、哥伦比亚、南非和中国台湾等国家或地区获得批准,以相应用途投
放市场。
本公司对转化受体植物、基因操作、DAS-21023-5 转化体本身、DAS-21023-
5 的加工过程及加工产品进行了大量的研究和评价,结果表明,该转基因棉花对
人体健康、植物、微生物安全以及生态环境无不良影响:
1)DAS-21023-5 的外源序列以单拷贝、单位点整合至转化受体基因组第 8 染
色体,且转化体不含载体骨架序列,转化方法安全,外源片段全序列及旁侧序列
明确,表达量及表达部位、表达时期明确,接合区未产生潜在致毒、致敏的读码
框;2)外源基因在 DNA 整合水平、蛋白质表达水平及性状表现水平均在代际间
稳定遗传;3)DAS-21023-5 与非转基因对照在生存竞争能力、基因漂移及对生
物多样性影响方面实质等同;4)DAS-21023-5 所表达的外源基因不会引起人、
畜中毒或过敏,转化体中无抗生素抗性基因、DAS-21023-5 的籽粒及加工产品营
养成分和抗营养因子与非转基因对照等同。因此,该转化体与其非转基因对照同
样安全。
鉴于以上科学依据及其它国家审批经历,本公司生物安全负责部门同意转
基因棉花 DAS-21023-5 在中国申请进口用作加工原料。
科迪华农业科技有限责任公司
2021 年 10 月

234
八、本单位审查意见
鉴于详实的科学依据及其它国家审批经历,同意申报书《转 cry1Ac 基因抗
虫棉花 DAS-21023-5 进口用作加工原料的安全证书》递交中国农业农村部。
科迪华农业科技有限责任公司
2021 年 10 月

235
参考文献
1. 崔俊明,2007。新编玉米育种学。中国农业科学技术出版社。
2. 崔荣荣, 韦颖, 孟攀潘, 马去龙, 金媛, 李秋然, 强胜, 戴伟民, 宋小玲, 2012. 抗草铵膦转基因水稻明恢 86b 杂草化潜力评价. 中国水稻科学 26, 467-475.
3. 冯献, 詹玲, 2012. 中国棉花消费分析及未来展望. 农业展望 8, 56-58.
4. 刘洪艳, 弭晓菊, 崔继哲, 2007. bar 基因、PAT 蛋白和草丁膦的特性与
安全性[J]. 生态学杂志, 26(6): 938-942.
5. 贾士荣, 2000. 我国棉花基因工程的研究进展. 中国农业科技导报. 2, 18-20.
6. 刘谦, 朱鑫泉, 2002. 生物安全. 科学出版社, 北京, pp. 67-73.
7. 强胜, 2000. 杂草学. 中国农业出版社, pp. 8-14.
8. 田逢秀, 1996. 棉花病害及防治. 新疆农业科技 1.
9. 晓婷, 2010. 棉花的历史. 中国纤检 7, 015.
10. 叶国祥, 2011. 中国陆地棉品种演化的遗传学分析. 南京农业大学.
11. 喻树迅, 2013. 我国棉花生产现状与发展趋势. 中国工程科学 15, 9-13.
12. 张海清, 2004. 中国棉花产区比较优势研究. 中国农业科学院.
13. 张立祯, 曹卫星, 2003. 基于生理发育时间的棉花生育期模拟模型. 棉花
学报 15, 97-103.
14. 赵琦, 2007. 轧花工艺对机采棉和手摘棉性能影响的探究. 新疆大学.
15.中华人民共和国农业农村部 1485 号公告-18-2010,《转基因生物及其产
品食用安全检测 外源蛋白质过敏性生物信息学分析方法》。中华人民
共和国农业农村部.
16.中华人民共和国农业农村部 2630 号公告-16-2017,《转基因生物及其产
品食用安全检测 外源蛋白质与毒性蛋白质和抗营养因子的氨基酸序列
相似性生物信息学分析方法》。
17. Addison, S.J., Farrell, T., Roberts, G.N., Rogers, D.J., 2007. Roadside surveys support predictions of negligible naturalisation potential for cotton (gossypium hirsutum) in north‐east australia. Weed Research 47, 192-201.
18. Alexander, M. P. (1969). Differential staining of aborted and non-aborted pollen. Stain Technology. 44:117-122.
19. Alexander, M. P. (1980). A versatile stain for pollen, fungi, yeast, and bacteria. Stain Technology. 55:13-18.
20. Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic local alignment search tool. Journal of Molecular Biology 215, 403 - 410.
21. Atlagić, J., Terzić, S., Marjanović-Jeromela, A., 2012. Staining and

236
fluorescent microscopy methods for pollen viability determination in sunflower and other plant species. Industrial Crops and Products 35, 88 - 91.
22. Arumuganathan, K., Earle, E., 1991. Nuclear DNA content of some important plant species. Plant Molecular Biology Reporter 9, 208-218.
23. Bakkeren, GZ., Koukolikova N., Grimsley N., 1989. Recovery of Agrobacterium tumfaciens T-DNA molecules from whole plants early after transfer. Cell. 57(5):847-857.
24. Barker, R.F., Idler, K.B., Thompson, D.V., Kemp, J.D., 1983. Nucleotide-sequence of the t-DNA region from the agrobacterium tumefaciens octopine ti plasmid pti15955. Plant Molecular Biology 2, 335 - 350.
25. Bates, S.L., Zhao, J.-Z., Roush, R.T., Shelton, A.M., 2005. Insect resistance management in gm crops: Past, present and future. Nature Biotechnology 23, 57-62.
26. Batista, C.E.d.A., Ciampi, A.Y., Hoffman, L.V., Barrosa, P.A.V., 2005. Conservação, diversidade e estrutura genética de populações de gossipium
mustelinum presente no semi-árido nordestino. V Congresso Brasileiro de Algodão.
27. Bauce É, Carisey N, Dupont A, van Frankenhuyzen K, 2004. Bacillus
thuringiensis subsp. kurstaki Aerial Spray Prescriptions for Balsam Fir Stand Protection Against Spruce Budworm (Lepidoptera: Tortricidae). Journal of Economic Entomology 97: 1624-1634
28. Bayer, E., Gugel, K.H., Hagele, K., Hagenmaier, H., Jessipow, S., König, W.A. and Zähner, H. 1972. Stofwechproduckte von Mikroorganismen.Phosphinothricin und hosphinothricyl-alanyl-alanin. Helvetica Chimica Acta. 55:224-239.
29. Berberich, S.A., Ream, J.E., Jackson, T.L., Wood, R., Stipanovic, R., Harvey, P., Patzer, S., Fuchs, R., L, 1996. The composition of insect-protected cottonseed is equivalent to that of conventional cottonseed. Journal of Agricultural and Food Chemistry 44, 365 - 371.
30. Bernhard K, 1992. betQuantitative determination of δ-endotoxin contents in spray-dried preparations of Bacillus thuringiensis strain GC-91. World Journal of Microbiology and Biotechnology 8: 24-29
31. Bertolla, F., Simonet, P., 1999. Horizontal gene transfers in the environment: Natural transformation as a putative process for gene transfers between transgenic plants and microorganisms. Research in Microbiology 150, 375-384.
32. Besler, M., Steinhart, H., Paschke, A., 2001. Stability of food allergens and allergenicity of processed foods. Journal of Chromatography B 756, 207-228.
33. Betz FS, Hammond BG, Fuchs RL, 2000. Safety and Advantages of Bacillus
thuringiensis-Protected Plants to Control Insect Pests. Regulatory Toxicology and Pharmacology 32: 156-173
34. Bonhomme, S., Grelon, M., Guerche, P., Horlow, C., and Vezon, D. EMBO Course: Practical Course on Genetic and Molecular Analysis of Arabidopsis, Module 1, Arabidopsis gametogenesis.

237
ftp://ftp.arabidopsis.org/Protocols/EMBOmanual/ch1.pdf
35. Bravo, A., 1997. Phylogenetic relationships of bacillus thuringiensis -endotoxin family proteins and their functional domains. Journal of Bacteriology 179, 2793 - 2801.
36. Bravo, A., Gómez, I., Conde, J., Muñoz-Garay, C., Sánchez, J., Miranda, R., Zhuang, M., Gill, S.S., Soberón, M., 2004. Oligomerization triggers binding of a bacillus thuringiensis cry1ab pore-forming toxin to aminopeptidase n receptor leading to insertion into membrane microdomains. Biochimica et Biophysica Acta 1667, 38 - 46.
37. Brubaker, C.L., Bourland, F.M., Wendel, J.F., 1999. The origin and domestication of cotton. In: Smith, C.W., Cothren, J.T. (Eds.), Cotton: Origin, history, technology, and production. John Wiley & Sons, New York, pp. 3-31.
38. Brubaker, C.L., Wendel, J.F., 1994. Reevaluating the origin of domesticated cotton (gossypium hirsutum; malvaceae) using nuclear restriction fragment length polymorphisms (rflps). American Journal of Botany 81, 1309-1326.
39. Cardineau, G.A., Stelman, S.J., Narva, K.E., 2001. Plant-optimized genes encoding pesticidal toxins. U.S. Patent No. 6,218,188 B1.
40. Carstens, K., Anderson, J., Bachman, P., De Schrijver, A., Dively, G., Federici, B., Hamer, M., Gielkens, M., Jensen, P., Lamp, W., Rauschen, S., Ridley, G., Romeis, J., Waggoner, A., 2011. Genetically modified crops and aquatic ecosystems: Considerations for environmental risk assessment and non-target organism testing. Transgenic Research 21, 813 - 842.
41. CERA - ILSI Research Foundation. 2011. "A Review of the Environmental Safety of the PAT Protein." Environmental Biosafety Research 10: 73-101.
42. Chauhan, D.S., Singh, P., Singh, P., 1983. Isolation distance in upland cotton under nagpur conditions. . Cotton Development 13, 23.
43. Chougule NP, Bonning BC, 2012. Toxins for Transgenic Resistance to Hemipteran Pests. Toxins 4: 405-429
44. Christensen, A.H., Sharrock, R.A., Quail, P.H., 1992. Maize polyubiquitin genes: Structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Molecular Biology 18, 675 - 689.
45. Coates, B.S., Sumerford, D.V., Hellmich, R.L., Lewis, L.C., 2005. Sequence variation in the cadherin gene of ostrinia nubilalis: A tool for field monitoring. Insect Biochemistry and Molecular Biology 35, 129-139.
46. Codex, 2003a. Principles for the risk analysis of foods derived from modern biotechnology. Codex of Alimentarius, CAC/GL 44-2003.
47. Codex, 2003b. Guideline for the conduct of food safety assessment of foods derived from recombinant-dna plants. Codex of Alimentarius. CAC/GL 45-2003.
48. Conner, A.J., Glare, T.R., Nap, J.P., 2003. The release of genetically modified crops into the environment. Part ii. Overview of ecological risk assessment. The Plant Journal 33, 19-46.

238
49. Corinne Hérouet, David J. Esdaile, Bryan A. Mallyon, Eric Debruyne, Arno Schulz, Thomas Currier, Koen Hendrickx, Robert-Jan van der Klis, Dominique Rouan, 2005. Safety evaluation of the phosphinothricin acetyltransferase proteins encoded by the pat and bar sequences that confer tolerance to glufosinate-ammonium herbicide in transgenic plants. Regulatory Toxicology and Pharmacology 41: 134–149.
50. Coutinho, E.M., 2002. Gossypol: A contraceptive for men. Contraception 65, 259-263.
51. Demanèche S, Sanguin H, Poté J, Navarro E, Bernillon D, Mavingui P, Wildi W, Vogel TM and Simonet P, 2008. Antibiotic-resistant soil bacteria in transgenic plant fields. Proceedings of the National Academy of Sciences of the USA, 105, 3957-3962.
52. Devine, M., Duke, S.O. and Fedtke, C. 1993. Physiology of Herbicide Action. Prentice Hall, Englewood Cliffs, NJ, pp. 251-294.
53. Dobrindt U, Hochhut B, Hentschel U, Hacker J, 2004. Genomic islands in pathogenic and environmental microorganisms. Nature Reviews Microbiology 2, 414-424.
54. Duan, J.J., Lundgren, J.G., Naranjo, S., Marvier, M., 2009. Extrapolating non-target risk of bt crops from laboratory to field. Biology Letters 6, 74-77.
55. Duan, J.J., Marvier, M., Huesing, J., Dively, G., Huang, Z.Y., 2008. A meta-analysis of effects of bt crops on honey bees (hymenoptera: Apidae). PLoS ONE 3, e1415.
56. Duke, J.A., 1983. Gossypium hirsutum l. . In: Duke, J.A. (Ed.), Handbook of energy crops. Purdue University, Center for New Crops & Plant Products, West Lafayette, Indiana.
57. Duke, S.O., 1996. Herbicide-resistant crops: Agricultural, environmental, economic, regulatory, and technical aspects. CRC Press, Inc., Boca Raton.
58. EFSA, 2005. Opinion of the scientific panel on genetically modified organisms on a request from the commission related to notification (reference c/es/01/01) for the placing on the market of insect-tolerant genetically modified maize 1507, for import, feed and industrial processing and cultivation under part c of directive 2001/18/ec form pioneer hi-bred international/mycogen seeds. The EFSA Journal 181, 1-33.
59. EFSA, 2006. Opinion of the scientific panel on genetically modified organisms on an application (reference efsa-gmo-nl-2005-13) for the placing on the market of glufosinate-tolerant genetically modified llcotton25, for food and feed uses, and import and processing under regulation (ec) no 1829/2003 from bayer cropscience. EFSA Journal 429, 1-19.
60. EFSA, 2007a. Opinion of the scientific panel on genetically modified organisms on an application (reference efsa-gmo-nl-2005-18) for the placing on the market of the glufosinate tolerant soybean a2704-12, for food and feed uses, import and processing under regulation (ec) no 1829/2003 from bayer cropscience. EFSA Journal 524, 1-22.
61. EFSA, 2007b. Opinion of the scientific panel on genetically modified organisms on an application (reference efsa-gmo-uk-2004-04) for the placing

239
on the market of glufosinate tolerant genetically modified rice llrice62 for food and feed uses, import and processing, under regulation (ec) no 1829/2003 from bayer cropscience gmbh. EFSA Journal 588, 1-25.
62. EFSA, 2008. Opinion of the scientific panel on genetically modified organisms on applications (references efsa-gmo-uk-2005-25 and efsa-gmo-rx-t45) for the placing on the market of the glufosinate-tolerant genetically modified oilseed rape t45, for food and feed uses, import and processing and for renewal of the authorisation of oilseed rape t45 as existing product, both under regulation (ec) no 1829/2003 from bayer cropscience. EFSA Journal 635, 1-22.
63. EFSA, 2009. Consolidated presentation of the joint scientific opinion of the GMO and BIOHAZ panels on the "Use of antibiotic resistance genes as marker genes in genetically modified plants" and the scientific opinion of the GMO panel on "Consequences of the opinion on the use of antibiotic resistance genes as marker genes in genetically modified plants on previous EFSA assessments of individidual GM plants". The EFSA Journal 1108, 1-8.
64. Ellis, J.G., Llewellyn, D.J., Walker, J.C., Dennis, E.S., Peacock, W.J., 1987. The ocs element: A 16 base pair palindrome essential for activity of the octopine synthase enhancer. The EMBO Journal 6, 3203-3208.
65. Embrey, S. K. (2013). Influences of pH on Phosphinothricin Acetyltransferase (PAT) protein. Indianapolis, IN, Dow AgroSciences LLC.
66. EPA, 1998. Guidelines for ecological risk assessment. EPA/630/R-95/002F. www.epa.gov/raf/publications/pdfs/ECOTXTBX.PDF.
67. EPA, 2001a. Sap report no. 2000-07 fifra scientific advisory panel meeting, october 18 - 20, 2000; final report: "Sets of scientific issues being considered by the environmental protection agency regarding: Bt plant-pesticides risk and benefit assessments".
68. EPA, 2001b. Sap report no. 2001-09 fifra scientific advisory panel meeting, july 17 - 18, 2001; a set of scientific issues being considered by the environmental protection agency regarding: Assessment of additional scientific information concerning starlink corn. 39.
69. EPA, 2004. Sap report no. 2004-05 fifra scientific advisory panel meeting, june 8 - 10, 2004; product characterization, human health risk, ecological risk, and insect resistance management for bacillus thuringiensis (bt) cotton products. 84.
70. EPA, 2007. White paper on tier-based testing for the effects of proteinaceous insecticidal plant-incorporated protectants on non-target arthropods for regulatory risk assessments.
71. EPA, 2011. Bacillus thuringiensis: Preliminary work plan and summary document for registration review. http://www.regulations.gov/#!documentDetail;D=EPA-HQ-OPP-2011-0705-0002.
72. FAO, 2000. Safety aspects of genetically modified foods of plant origin.
73. Fernandez-Cornejo, J., Hallahan, C., Nehring, R., Wechsler, S., Grube, A., 2012. Conservation tillage, herbicide use, and genetically engineered crops in the united states: The case of soybeans. AgBioForum 15, 231-241.

240
74. Forster, L.A., Calhoun, M.C., 1995. Nutrient values for cottonseed products deserve new look. Feedstuffs 67, 105.
75. Freire, E.C., Moreira, J.d.A.N., dos Santos, J.W., Pereira de Andrade, F., 1998. Relações taxonômicas entre os algodoeiros mocó e gossypium
mustelinum do nordeste brasileiro. Pesquisa Agropecuária Brasileira 33, 1555-1561.
76. Fryxell, P.A., 1979. The nature history of the cotton tribe (malvaceae, tribe gossypieae). Texas A&M University Press, College Station and London.
77. Fryxell, P.A., 1984. Taxonomy and germplasm resources. In: Kohel, R.J., Lewis, C.F. (Eds.), Cotton ASA-CSSA-SSSA Madison, WI, pp. 27-57.
78. FWS. (1984). Kern primrose sphinx moth. U.S. Fish & Wildlife Service, Endangered Species Div., Sacramento Fish & Wildlife Office, Sacramento, CA Accessed electronically, 9 Sep 02. http://sacramento.fws.gov/es/animalsppacct/kernprimrosesphinxmoth.htm.
79. FWS., 1997. Endangered Species Database. U.S. Fish & Wildlife Service, Washington, DC.
80. Gahan, L.J., Gould, F., Heckel, D.G., 2001. Identification of a gene associated with bt resistance in heliothis virescens. Science 293, 857-860.
81. Glare, T., O'Callaghan, M., 2000. Bacillus thuringiensis: Biology, ecology, and safety. Wiley, New York.
82. Gogarten JP, Townsend JP, 2005. Horizontal gene transfer, genome innovation and evolution. Nature Reviews Microbiology 3, 679-687.
83. Gouffon, C., Van Vliet, A., Van Rie, J., Jansens, S., Jurat-Fuentes, J.L., 2011. Binding sites for bacillus thuringiensis cry2ae toxin on heliothine brush border membrane vesicles are not shared with cry1a, cry1f, or vip3a toxin. Applied and Environmental Microbiology 77, 3182-3188.
84. Grover LM, 2011.Genetically engineered crops : biotechnology, biosafety, and benefits. Nova Science, Hauppauge, N.Y., xi, 234 p. pp
85. Hellrnich, R. L.; Siegfried, B. D.; Sears, M. K.; Stanley-Horn, D. E.; Daniels, M. 1.; Mattila, H.R; Spencer, T.; Bidne, K. G.; Lewis, L. C.,2001. Monarch larvae sensitivity to Bacillus thuringiensis-purified proteins and pollen. Proc. Nat. Acad. Sci. 98 (21): 11925-11930.
86. Hérouet, C., Esdaile, D.J., Mallyon, B.A., Debruyne, E., Schulz, A., Currier, T., Hendrickx, K., van der Klis, R.-J., Rouan, D., 2005. Safety evaluation of the phosphinothricin acetyltransferase proteins encoded by the pat and bar sequences that confer tolerance to glufosinate-ammonium herbicide in transgenic plants. Regulatory Toxicology and Pharmacology 41, 134-149.
87. Holm, L.G., Doll, J., Eric, H., Pancho, J.V., Herberger, J.P., 1997. World weeds: Natural histories and distribution. John Wiley & Sons, New York.
88. Holm, L.G., Pancho, J.V., Herberger, J.P., Plucknett, D.L., 1979. A geographical atlas of world weeds. John Wiley & Sons, New York.
89. Huang, J., Hu, R., Rozelle, S., Qiao, F., Pray, C.E., 2002. Transgenic varieties and productivity of smallholder cotton farmers in china. Australian Journal of Agricultural and Resource Economics 46, 367-387.

241
90. ISAAA Briefs, BRIEF 53, Global Status of Commercialized Biotech/GM Crops: 2018. Ithaca, NY, USA: International Service for the Acquisition of Agri-biotech Applications (ISAAA).
91. Jones, L.A., Wedegaertner, T.C., 1985. Advances in feeding cottonseed products. American Oil Chemists’ Society.
92. Josef Vinnemeier, Wolfgang Dröge-Laser, Elfriede K. Pistorius and Inge Broer. 1995. Purification and Partial Characterization of the Streptomyces viridochromogenes TÜ494 Phosphinothricin-N-acetyltransferase Mediating Resistance to the Herbicide Phosphinothricin in Transgenic Plants. Z. Naturforsch. 50c, 796-805 (1995).
93. Kapur M, Bhatia R, Pandey G, Pandey J, Paul D, Jain RK., 2010. A case study for assessment of microbial community dynamics in genetically modified Bt cotton crop fields. Currt Microbiol, 61(2): 118–124.
94. Kay E, Vogel TM, Bertolla F, Nalin R and Simonet P. 2002. In situ transfer of antibioticresistance genes from transgenic (transplastomic) tobacco plants in bacteria. Applied and Environmental Microbiology, 68, 3345-3351.
95. Kebede, H., Burow, G., Dani, R.G., Allen, R.D., 2007 A-genome cotton as a source of genetic variability for upland cotton (gossypium hirsutum). Genetic Resources and Crop Evolution 54, 885-895.
96. Keese P, 2008. Risks from GMOs due to horizontal gene transfer. Environmental Biosafety Research 7, 123-149
97. Kim SE, Moon JS, Kim JK, Yoo RH, Choi WS, Lee EN, Lee SH and Kim SU, 2010. Monitoring of possible horizontal gene transfer from transgenic potatoes to soil microorganisms in the potato fields and the emergence of variants in Phytophthora infestans. Journal of Microbiology and Biotechnology, 20, 1027-1031.
98. Kishore, G.M. and Shah, D.M. 1988. Amino acid biosynthesis inhibitors as herbicides. Annual Review of Biochemistry. 57:627-663.
99. Kumar, P.A., Sharma, R.P., Malik, V.S., 1996. The insecticidal proteins of bacillus thuringiensis. Advanced Applied Microbiology 42, 1-12, 12A, 13-43.
100.Kung SC, Retchless AC, Kwan JY, Almeida PP. 2013. Effects of DNA Size on Transformation and Recombination Efficiencies in Xylella fastidiosa. Applied Environmental Microbiology, 79, 1712-1717
101.Lacape, J.M., Dessauw, D., Rajab, M., Noyer, J.L., Hau, B., 2007. Microsatellite diversity in tetraploid gossypium germplasm: Assembling a highly informative genotyping set of cotton ssrs. Molecular Breeding 19, 45-48.
102.Lee, M.K., Miles, P., Chen, J.-S., 2006. Brush border membrane binding properties of bacillus thuringiensis vip3a toxin to heliothis virescens and helicoverpa zea midguts. Biochemical and Biophysical Research Communications 339, 1043-1047.
103.Liu, J.-G., Yang, A.-Z., Shen, X.-H., Hua, B.-G., Shi, G.-L., 2011. Specific binding of activated vip3aa10 to helicoverpa armigera brush border

242
membrane vesicles results in pore formation. Journal of Invertebrate Pathology 108, 92-97.
104.Liu, R.J., 1995. Effect of vesicular-arbuscular mycorrhizal fungi on verticillium wilt of cotton. Mycorrhiza 5, 293-297.
105.Llewellyn, D., Fitt, G., 1996. Pollen dispersal from two field trials of transgenic cotton in the namoi valley, australia. Molecular Breeding 2, 157-166.
106.Lu Y, Wu K, Jiang Y, et al. Mirid bug outbreaks in multiple crops correlated with1151-1154.Martin, B., Alloing, G., Méjean, V., Claverys, J.-P., 1987. Constitutive expression of erythromycin resistance mediated by the ermam determinant of plasmid pamβ1 results from deletion of 5′ leader peptide sequences. Plasmid 18, 250-253.
107.Ma BL, Blackshaw RE, Roy J and He T, 2011. Investigation on gene transfer from genetically modified corn (Zea mays L.) plants to soil bacteria. Journal of Environmental Science and Health. Part. B, Pesticides, food contaminants, and agricultural wastes, 46, 590-599.
108.Majewski J, Zawadzki P, Pickerill P, Cohan FM, Dowson CG. 2000. Barriers to genetic exchange between bacterial species: Streptococcus pneumoniae transformation. Journal of Bacteriology 182: 1016–1023.
109.Martin, B., Alloing, G., Méjean, V., Claverys, J.-P., 1987. Constitutive expression of erythromycin resistance mediated by the ermam determinant of plasmid pamβ1 results from deletion of 5′ leader peptide sequences. Plasmid 18, 250-253.
110.Marvier, M., McCreedy, C., Regetz, J., Kareiva, P., 2007. A meta-analysis of effects of bt cotton and maize on nontarget invertebrates. Science 316, 1475-1477.
111.McClintock, J.T., Schaffer, C.R., Sjoblad, R.D., 1995. A comparative review of the mammalian toxicity of bacillus thuringiensis-based pesticides. Pesticide Science 45, 95-105.
112.McGregor, S.E. 1976. Insect Pollination of Cultivated Crop Plants. Agriculture Handbook No. 496. U.S. Department of Agriculture, Agricultural Research Service. GPO, Washington, D.C. 411 pp.
113.Mehetre, S.S., Thombre, M.V., Tyyab, M.A., 1980. Cytomorphological studies in an intergeneric hybrid between gossypium hirsutum l.(2n= 52) and hibiscus panduraeformis burm. Euphytica 29, 323-330.
114.Mendelsohn, M., Kough, J., Vaituzis, Z., Matthews, K., 2003. Are bt crops safe? Nature Biotechnology 21, 1003-1009.
115.Merkus, H. G., 2009. Particle Size Measurements: Fundamentals, Practice,
Quality. Springer, pp 15.
116.Morin, S., Biggs, R.W., Sisterson, M.S., Shriver, L., Ellers-Kirk, C., Higginson, D., Holley, D., Gahan, L.J., Heckel, D.G., Carrière, Y., Dennehy, T.J., Brown, J.K., Tabashnik, B.E., 2003. Three cadherin alleles associated with resistance to bacillus thuringiensis in pink bollworm. Proceedings of the National Academy of Sciences 100, 5004-5009.

243
117.Naranjo, S.E., 2005. Long-term assessment of the effects of transgenic bt cotton on the abundance of nontarget arthropod natural enemies. Environmental Entomology 34, 1193 - 1210.
118.Nehl, D.B., Allen, S.J., Mondal, A.H., Lonergan, P.A., 2004. Black root rot: A pandemic in australian cotton. Australasian Plant Pathology 33, 87-95.
119.Nielsen KM, Bones AM, Smalla K and van Elsas JD, 1998. Horizontal gene transfer from transgenic plants to terrestrial bacteria - a rare event? FEMS Microbiology Reviews, 22, 79-103.
120.Nielsen, K.M., van Elsas, J.D., Smalla, K., 2000. Transformation of acinetobacter sp. Strain bd413 (pfg4δnptii) with transgenic plant DNA in soil microcosms and effects of kanamycin on selection of transformants. Applied and Environmental Microbiology 66, 1237-1242.
121.OECD, 1999. Consensus document on general information concerning the genes and their enzymes that confer tolerance to phosphinothricin herbicide. 11 (ENV/JM/MONO(99)13): 1-26. www.oecd.org/dataoecd/16/52/46815628.pdf.
122.OECD, 2003. Consensus document on the biology of Zea mays subs mays (Maize).
123.OECD, 2007. Consensus document on safety information on transgenic plants expressing bacillus thuringiensis-derived insect control proteins. ENV/JM/MONO(2007)14.
124.OECD, 2008. Consensus document on the biology of cotton (gossypium
spp.). 64. ENV/JM/MONO(2008)33.
125.OECD, 2009. Consensus document on compositional considerations for new varieties of cotton (gossypium hirsutum and gossypium barbadense): Key food and feed nutrients and anti-nutrients. 32. ENV/JM/MONO(2004)16.
126.Pabst, W., and Gregorová, E., 2007. Characterization of particles and particle systems. ICT. Vol 602.
127.Pastorello, EA. et al. 2000. The maize major allergen, which is responsible for food-induced allergic ractions, is a lipid transfer protein. J.Allergy Clin Immunol 106:744-751.
128.Pastorello, EA. et al. 2003. Lipid-transfer protein is the major maize allergen maintaining IgE-binding activity after cooking at 100℃, as demonstrated in anaphylactic patients and patients with positive double-blind, placebo-copntrolled food challenge results. J.Allergy Clin Immunol 112:775-783.
129.Paul, J.H., 1999. Microbial gene transfer: An ecological perspective. Journal of Molecular Microbiology and Biotechnology 1, 45-50.
130.Payne J, Sick AJ, inventors. Jun. 27, 1989. Bacillus thuringiensis isolate active against lepidopteran pests, and genes encoding novel lepidopteran-active toxins. US Patent, Application No. 371,955
131.Pearson, W., 2000. Flexible sequence similarity searching with the fasta3 program package. Methods in Molecular Biology 132, 185-219.
132.Pearson, W.R., 1999. Flexible sequence similarity searching with the fasta3 program package. In: Misener, S., Krawetz, S.A. (Eds.), Bioinformatics

244
methods and protocols. Humana Press, pp. 185-219.
133.Petersen, A, Dresselhaus, T.,Grobe, K., Becker, W. 2006. Proteome analysis of maize pollen for allergy-relavent components. Proteomics, 6:6317-6325.
134.Randall, R.P., 2002. A global compendium of weeds. RG and FJ Richardson, Melbourne, Australia.
135.Rang, C., Bergvingson, D., Bohorova, N., Hoisington, D., Frutos, R., 2004. Competition of bacillus thuringiensis cry1 toxins for midgut binding sites: A basis for the development and management of transgenic tropical maize resistant to several stemborers. Current Microbiology 49, 22-27.
136.Raps, A., Kehr, J., Cugerli, P., Moar, W.J., Bigler, F., Hilbeck, A., 2001. Immunological analysis of phloem sap of bacillus thuringiensis corn and of the nontarge herbivore rhopalosiphum padi (homoptera: Aphididae) for the presence of cry1ab. Molecular Ecology 10, 525 - 533.
137.Reddy, K.R., Hodges, H.F., Reddy, V.R., 1992. Temperature effects on cotton fruit retention. Agronomy Journal 84 26-30.
138.Riley, Grahm J.P.1981. Effects of high temperature on the germination of maize (Zea mays L.). Planta, 151:68-74.
139.Rogoff MH, Yousten AA, 1969. Bacillus Thuringiensis: Microbiological Considerations. Annual Review of Microbiology 23: 357-386
140.Romeis, J., Meissle, M., Bigler, F., 2006. Transgenic crops expressing bacillus thuringiensis toxins and biological control. Nature Biotechnology 24, 63-71.
141.Salcedo, G., Sanchez-Monger, R., Diaz-Perales, A., Garcia-Casado, G., Barber,D. 2004. Plant non-specific lipid transfer proteins as food and pollen allergens. Clinical & Experimental Allergy, 34:1336-1341.
142.Sanahuja, G., Banakar, R., Twyman, R.M., Capell, T., Christou, P., 2011. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnology Journal 9, 283-300.
143.Sanaullah Yasin, Hafiz Naeem Asghar, Fiaz Ahmad, Zahir Ahmad Zahir & Ejaz Ahmad Waraich. 2016. Impact of Bt-cotton on soil microbiological and biochemical attributes. Plant Production Science. 19:4, 458-467.
144.Sanchis V, 2011. From microbial sprays to insect-resistant transgenic plants: history of the biospesticide Bacillus thuringiensis. A review. Agronomy for Sustainable Development 31: 217-231
145.Santos, J.E.P., Villaseňor, M., DePeters, E.J., Robinson, P.H., Baldwin Jr, B.C., 2002. Type of cottonseed and level of gossypol in diets of lactating dairy cows: Effects on lactation performance and plasma gossypol. Journal of Dairy Science 85, 1491-1501.
146.Sanvido, O., Romeis, J., Bigler, F., 2007. Ecological impacts of genetically modified crops: Ten years of field research and commercial cultivation. Advances in Biochemical Engineering/Biotechnology 107, 235 - 278.
147.Schlüter K, Futterer J, Potrykus I, 1995. "Horizontal" gene transfer from a transgenic potato line to a bacterial pathogen (Erwinia chrysanthemi) occurs -

245
if at all - at an extremely low frequency. Nature Biotechnology 13, 1094-1098.
148.Schnepf, E., Crickmore, N., Van Rie, J., Lereclus, D., Baum, J., Feitelson, J., Zeigler, D.R., Dean, D.H., 1998. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiology and Molecular Biology Reviews 62, 775-806.
149.Senchina, D.S., Alvarez, I., Cronn, R.C., Liu, B., Rong, J., Noyes, R.D., Paterson, A.H., Wing, R.A., Wilkins, T.A., Wendel, J.F., 2003. Rate variation among nuclear genes and the age of polyploidy in gossypium. Molecular Biology and Evolution 20, 633-643.
150.Shah, D., Horsch, R., Klee, H., Kishore, G., Winter, J., Tumer, N., Hironaka, C., Sanders, P., Gasser, C., Aykent, S., Siegel, N., Rogers, S. and Fraley, R. 1986. Engineering herbicide tolerance in transgenic plants. Science. 233:478-481.
151.Stalker, D.M., Thomas, C.M., Helinski, D.R., 1981. Nucleotide sequence of the region of the origin of replication of the broad host range plasmid rk2. Molecular and General Genetics 181, 8-12.
152.Tepfer D, Garcia-Gonzales R, Mansouri H, Seruga M, Message B, Leach F and Perica MC, 2003. Homology-dependent DNA transfer from plants to a soil bacterium under laboratory conditions: implications in evolution and horizontal gene transfer. Transgenic Research, 12, 423-437.
153.Umbeck, P.B., Kenneth A, Norheim, E.V., McCarty, J.C., Parrot, W.L., Jenkins, J.N., 1991. Degree of pollen dispersal by insects from a field test of genetically engineered cotton. Journal of Economic Entomology 84, 1943-1950.
154.USDA, 2000. Genetically engineered crops for pest management in u.S. Agriculture. 28. Agricultural Economic Report No. (AER-786). http://www.ers.usda.gov/publications/aer-agricultural-economic-report/aer786.aspx.
155.USEPA, 2001b. Biopesticides Registration Action Document: Bacillus thuringiensis Plant-Incorporated Protect ants, 16 Oct 2001. USEPA, OPP, BPPD, Washington, DC. Accessed electronically 3 Jul 2002. http://www.epa.gov/pesticides/biopesticides/reds/brad_bt_pip2.htm.
156.Vadlamudi, R.K., Ji, T.H., Bulla, L.A., 1993. A specific binding protein from manduca sexta for the insecticidal toxin of bacillus thuringiensis subsp. Berliner. The Journal of Biological Chemistry 268, 12334 - 12340.
157.Weber, E., 2003. Invasive plant species of the world: A reference guide to environmental weeds. CABI Publishing, Wallingford, England.
158.Weichel,M.,Vergoossen, NJ, Bonomi, S., Scibilia, J., Ortolani, C., Ballmer-Weber, B.K., Pastorello, EA, Cramen, R. 2006. Screening the allergenic repertories of wheat and amize with sear from double-blind, placebo-controlled food allergen challenge positive patients. Allergy, 61: 128-135.
159.Wendel, J.F., Brubaker, C.L., Percival, A.E., 1992. Genetic diversity in gossypium hirsutum and the origin of upland cotton. American Journal of Botany 79, 1291-1310.

246
160.Wendel, J.F., Cronn, R.C., 2003. Polyploidy and the evolutionary history of cotton. Advances in Agronomy 78, 139-186.
161.Wendel, J.F., Percy, R.G., 1990. Allozyme diversity and introgression in the galapagos islands endemic gossypium darwinii and its relationship to continental g. Barbadense. Biochemical Systematics and Ecology 18, 517-528.
162.Wohlleben, W., Arnold, W., Broer, I., Hillemann, D., Strauch, E., Punier, A., 1988. Nucleotide sequence of the phosphinothricin n-acetyltransferase gene from streptomyces viridochromogenes tü494 and its expression in nicotiana
tabacum. Gene 70, 25-37.
163.Wolfenbarger, L.L., Naranjo, S.E., Lundgren, J.G., Bitzer, R.J., Watrud, L.S., 2008. Bt crop effects on functional guilds of non-target arthropods: A meta-analysis. PLoS ONE 3, e2118.
164.Wolt, J.D., Keese, P., Raybould, A., Fitzpatrick, J.W., Burachik, M., Gray, A., Olin, S.S., Schiemann, J., Sears, M., Wu, F., 2010. Problem formulation in the environmental risk assessment for genetically modified plants. Transgenic Research 19, 425-436.
165.Yang, Y., Zhu, Y.C., Ottea, J., Husseneder, C., Leonard, B.R., Abel, C., Luttrell, R., Huang, F., 2011. Down regulation of a gene for cadherin, but not alkaline phosphatase, associated with cry1ab resistance in the sugarcane borer diatraea saccharalis. PLoS ONE 6, e25783.
166.Youssef, Y.A., Mankarios, A.T., 1974. Studies on the rhizosphere mycoflora of broad bean and cotton. Mycopathologia et Mycologia applicata 54, 173-180.
167.Yu, H.L., Li, Y.H., Wu, K.M., 2011. Risk assessment and ecological effects of transgenic bacillus thuringiensis crops on non-target organisms. Journal of Integrative Plant Biology 53, 520-538.
168.Zhang, Y.J, Xie, M., Peng D.L., Zhao, J.J., Zhang Z.R. Dynamics of microbial population size in rhizosphere soil of Monsanto’s Cry1Ac cotton. Plant Soil Environ. Vol. 62, 2016, No. 2: 92–97.
169.Vélez A.M., Spencer T.A., Alves A.P., Moellenbeck D., Meagher R.L., Chirakkal H. and Siegfried B.D., 2013. Inheritance of Cry1F resistance, crossresistance and frequency of resistant alleles in Spodoptera frugiperda. Bulletin of Entomological Research, Page 1 of 14.
170.Bravo, A., Gill, S.S., Soberón, M., 2007. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 49, 423 - 435.
171.Storer, N.P., Thompson, G.D., Head, G.P., 2012. Application of Pyramided Traits Against Lepidoptera in Insect Resistance Management for Bt Crops. GM Crops and Food: Biotechnology in Agriculture and the Food Chain 3, 154-162.
172.Jurat-Fuentes, J.L., Adang, M.J., 2006. The Heliothis virescens cadherin protein expressed in Drosophila S2 cells functions as a receptor for Bacillus
thuringiensis Cry1A but not Cry1Fa toxins. Biochemistry 45, 9688-9695.
173.Storer, N.P., Thompson, G.D., Head, G.P., 2012. Application of Pyramided

247
Traits Against Lepidoptera in Insect Resistance Management for Bt Crops. GM Crops and Food: Biotechnology in Agriculture and the Food Chain 3, 154-162.
174.Hernández, C.S., Ferré, J., 2005. Common Receptor of Bacillus thuringiensis Toxins Cry1Ac, Cry1Fa, and Cry1Ja in Helicoverpa armigera, Helicoverpa zea, and Spodoptera exigua. Applied and Environmental Microbiology 71, 5627-5629.
175.Pereira, E.J.G., Lang, B.A., Storer, N.P., Siegfried, B.D., 2008. Selection for Cry1F resistance in the European corn borer and cross-resistance to other Cry toxins. Entomologia Experimentalis et Applicata 126, 115-121.
176.Zhang Tiantao, He Mingxia, Angharad M. R. Gatehouse, Wang Zhenying, Martin G. Edwards, Li Qing and He Kanglai., 2014. Inheritance Patterns, Dominance and Cross-Resistance of Cry1Ab- and Cry1Ac-Selected Ostrinia furnacalis (Guenée). Toxins, 6, 2694-2707.




















