
ELUCIDATING THE INTERACTION BETWEEN Spodoptera frugiperda MULTIPLE NUCLEOPOLYHEDROVIRUS (SfMNPV) AND...
54
[1] TITULO DE LA TESIS DILUCIDANDO LA INTERACCIÓN ENTRE EL NUCLEOPOLIEDROVIRUS MÚLTIPLE DE Spodoptera frugiperda (SfMNPV) Y LA LOMBRIZ TERRESTRE Eisenia fetida: UN ESTUDIO DE LABORATORIO. TESIS QUE PRESENTA DENNIS ADRIÁN INFANTE RODRÍGUEZ PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS
Transcript of ELUCIDATING THE INTERACTION BETWEEN Spodoptera frugiperda MULTIPLE NUCLEOPOLYHEDROVIRUS (SfMNPV) AND...

[1]
TITULO DE LA TESIS
DILUCIDANDO LA INTERACCIÓN ENTRE EL NUCLEOPOLIEDROVIRUS
MÚLTIPLE DE Spodoptera frugiperda (SfMNPV) Y LA LOMBRIZ
TERRESTRE Eisenia fetida: UN ESTUDIO DE LABORATORIO.
TESIS QUE PRESENTA DENNIS ADRIÁN INFANTE RODRÍGUEZ
PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS

[2]

[3]

[4]
DEDICATORIA
A mi madre por enseñarme a perseguir mis sueños.
A mi familia, en especial a mi esposa Nadia Isabel Rodríguez Salazar por acompañarme en esta
aventura de vida.
A todos los amantes de la vida, “bichologos” y a las nuevas mentes inquietas.

[5]
AGRADECIMIENTOS
Al Dr. Trevor Williams por sembrar en mi mente el deseo de ser investigador, encaminarme
hacia la ciencia con gran dedicación y sobre todo con mucha paciencia.
A la Dra. Delia Muñoz Labiano por sus acertados comentarios y observaciones puntuales con
miras de enriquecer este trabajo de investigación.
Al Dr. Jorge E. Valenzuela por todo ese tiempo dedicado, sugerencias acertadas y su experiencia.
Al M.G.A. Gabriel Mercado Vidal por todas las atenciones, por ser mi gran amigo y compañero
en esta aventura.
A la Dra. Isabelle Barois por las sugerencias, su tiempo y la ayuda con la literatura.
A la Dra. Ana Mabel Martínez Castillo por las correcciones y sugerencias en el escrito.
Al CONACYT, por la beca de manutención otorgada.
A mis compañeros de laboratorio Ale, Dulce, por la ayuda y los ánimos durante el desarrollo de
este trabajo.
A mis fieles escuderos, amigos y compañeros de tertulias M.C. Pablo Esau Cruz Domínguez,
M.C. Mariana Robles Pliego.

[6]
ÍNDICE
RESUMEN .............................................................................................................................................. 8
I. INTRODUCCIÓN Y MARCO TEÓRICO.......................................................................................... 10
Spodoptera frugiperda - una plaga importante del maíz. ..................................................................... 10
Baculoviridae ..................................................................................................................................... 12
El SfMNPV como insecticida. ............................................................................................................ 17
Ecología de los baculovirus ................................................................................................................ 18
Dispersión.......................................................................................................................................... 18
Dispersión biótica .............................................................................................................................. 20
Las lombrices de tierra ....................................................................................................................... 20
La biología de Eisenia fetida .............................................................................................................. 23
II. OBJETIVOS ..................................................................................................................................... 25
Objetivo general................................................................................................................................. 25
Objetivos específicos ......................................................................................................................... 25
III. HIPÓTESIS ..................................................................................................................................... 25
IV. MATERIALES Y MÉTODOS ......................................................................................................... 26
Cría de insectos .................................................................................................................................. 26
Amplificación de SfMNPV ................................................................................................................ 26
Mezcla de suelo y estimación de pH ................................................................................................... 27
Estimación de la relación concentración de OBs en suelo-mortalidad ................................................. 28
Estimación del pH intestinal de E. fetida ............................................................................................ 29
Distribución vertical de lombrices en terrarios experimentales ............................................................ 29
Dispersión de OBs en suelo artificial por E. fetida .............................................................................. 31
Estimación de la relación concentración-mortalidad para OBs en suelo .............................................. 34
Estimación del pH intestinal de Eisenia fetida .................................................................................... 35
Supervivencia y descenso en suelo artificial ....................................................................................... 36
Redistribución de OBs en suelo artificial ............................................................................................ 37
Efecto de la presencia de E. fetida en la patogenicidad de OBs en suelo.............................................. 40
VI. DISCUSIÓN .................................................................................................................................... 41
VII. CONCLUSIONES .......................................................................................................................... 45

[7]
VII. REFERENCIAS ............................................................................................................................. 46
FIGURAS Y CUADROS
Figura 1. Estadios de Spodoptera frugiperda………….…………………………………….…………....11
Figura 2. Componentes estructurales de baculovirus ……………….……………...………………….…14
Figura 3. Fenotipos de OBs…………………………………………………....………………………….14
Figura 4. Larva de Spodoptera frugiperda, licuefacción por infección de SfMNPV…...………….…......15
Figura 5. Procesos de infección de los baculovirus………………..…. ….………………… ………….16
Figura 6. Unidades de 30 x 15 x 1 cm con suelo artificia……………………….………….……………..30
Figura 7. Mortalidad de larvas L2 de Spodoptera frugiperda por concentración de OBs/mg… ……..…..35
Figura 8. Disección del intestino de Eisenia fetida…………………….…………...…………… ……….36
Figura 9. Lombrices vivas después de 72 h en suelo estándar ………….……………...……… ……..….36
Figura 10. Promedio de descenso de las lombrices (± EE). ………………………...………………..…...37
Figura 11. Mortalidad de quién ¿por virus con respecto a la profundidad………………..……………..39
Figura 12. Mortalidad observada en los tratamientos con y sin lombriz……… …...…….………………40
Cuadro 1. Peso promedio de las lombrices utilizadas en cada réplica………….……………..…………..31
Cuadro 2. Efecto de las lombrices sobre los OBs……..…………………………………………………..38

[8]
“DILUCIDANDO LA INTERACCIÓN ENTRE EL NUCLEOPOLIEDROVIRUS
MÚLTIPLE DE Spodoptera frugiperda (SfMNPV) Y LA LOMBRIZ TERRESTRE Eisenia
fetida: UN ESTUDIO DE LABORATORIO”
RESUMEN
El suelo es reconocido como un importante reservorio medioambiental de virus
entomopatógenos, en particular de los cuerpos de oclusión (OBs) de los baculovirus (familia
Baculoviridae). El nucleopoliedrovirus múltiple de Spodoptera frugiperda (SfMNPV, género
Alphabaculovirus) ha atraído la atención para ser usado como un insecticida biológico en el
control de S. frugiperda (Lepidoptera: Noctuidae) en cultivos de maíz y sorgo en América. El
objetivo de esta tesis fue evaluar el papel de la lombriz de tierra Eisenia fetida como posible
dispersor de OBs del SfMNPV en un sistema modelo de laboratorio. Para ello, se calibró la
técnica de incorporación de suelos a dieta utilizando un suelo artificial (70 % de arena, 20 % de
caolín, 10 % turba de sphagnum). Los bioensayos se llevaron a cabo en larvas de segundo estadio
de S. frugiperda utilizando mezclas de OBs + suelo artificial incorporado a dieta semisintética
para estimar la concentración letal media (CL50). Concentraciones de 5x104 y 5x105 OBs/g
generaron < 5% de mortalidad por virus, concentraciones de 5x106, 5x107, 5x108 y 5x109 OBs/g
generaron 7, 13, 70 y 96% de mortalidad por virus. El valor de la CL50 se estimó en 2.3 x 108
OBs/g. Los OBs de los baculovirus son muy sensibles a la alcalinidad. Por esta razón, se estimó
el pH intestinal de E. fetida, entre 5.0-6.0 utilizando indicadores de pH. El peso promedio de las
lombrices de tierra utilizadas en los experimentos fue de 345 mg y la profundidad máxima
promedio excavada en el suelo experimental en un período de 72 h fue de 22.5 cm, con un 85 %
de supervivencia de las lombrices de tierra dentro de las unidades experimentales. Se confirmó la
capacidad de las lombrices para redistribuir verticalmente los OBs del SfMNPV en un suelo
artificial bajo condiciones de laboratorio, utilizando bioensayos de incorporación de suelo en
periodos de 1,7 y 15 días. Los OBs se detectaron a profundidades de hasta 22 cm en las unidades
experimentales. La detección de OBs aumentó significativamente con el tiempo (P < 0.001) y
disminuyó significativamente con el aumento de la profundidad del suelo (P < 0.01). La
incubación de las lombrices de tierra en suelo tratado con OBs durante 7 días no afectó
significativamente a la actividad insecticida de los OBs en comparación con el tratamiento de
OBs en el suelo en ausencia de lombrices de tierra (P > 0.05). Concluimos que la incorporación
de suelo artificial contaminado con OBs provocó infecciones per os en larvas de Spodoptera
frugiperda, el pH de E. fetida no es un factor limitante para el paso de OBs de SfMNPV a través
del intestino y E. fetida interactúa con el SfMNPV en el suelo artificial sin causarle inactivación y
puede dispersarlo en condiciones de laboratorio. Esto representa un mecanismo previamente
desconocido de dispersión de baculovirus en el medio ambiente y que puede tener importantes
implicaciones en la persistencia de las poblaciones de OBs en los reservorios del suelo.
Palabras clave: Spodoptera frugiperda, Baculoviridae, suelo artificial, Eisenia fetida.

[9]
ELUCIDATING THE INTERACTION BETWEEN Spodoptera frugiperda MULTIPLE
NUCLEOPOLYHEDROVIRUS (SfMNPV) AND THE EARTHWORM Eisenia fetida: A
LABORATORY STUDY
ABSTRACT
Soil has been recognized as an important environmental reservoir of entomopathogenic viruses,
particularly the occlusion bodies (OBs) of baculoviruses (family Baculoviridae). The multiple
nucleopolyhedrovirus (SfMNPV, genus Alphabaculovirus) of Spodoptera frugiperda
(Lepidoptera: Noctuidae) has attracted attention as a potential biological insecticide for control of
this pest in maize and sorghum crops in the Americas. The aim of this thesis was to evaluate the
potential role of the earthworm Eisenia fetida as a possible disperser of SfMNPV OBs in a model
laboratory system. For this, the soil incorporation bioassay technique was calibrated using an
artificial soil (70% sand, 20% kaolin, 10% sphagnum peat). Bioassays were performed on second
instar larvae of S. frugiperda using OBs + artificial soil mixtures incorporated into semisynthetic
diet to estimate the 50% lethal concentration (LC50). Concentrations of 5x104 and 5x105 OBs/ml
resulted in < 5% virus mortality, whereas concentrations of 5x106, 5x107, 5x108 and 5x109
OBs/ml resulted in 7, 13, 70 and 96% virus mortality. The LC50 value was estimated at 2.3 x 108
OBs/g. Baculovirus OBs are strongly sensitive to alkalinity. For this reason the gut pH of E.
fetida was estimated to be between 5.0-6.0 using pH indicators. The average weight of
earthworms was 345 mg and the average maximum depth burrowed into an experimental soil in a
period of 72 h was 22.5 cm, with 85% survival earthworms in the artificial soil experimental
units. The ability of earthworms to redistribute SfMNPV OBs vertically in artificial soil under
laboratory conditions was confirmed by soil incorporation bioassays over periods of 1, 7 and 15
days. OBs were detected at depths up to 22 cm in experimental units. Fitting generalized linear
models revealed that the detection of OBs increased significantly over time (P <0.001) and
decreased significantly with increasing soil depth (P <0.01). Incubation of earthworms in OB
treated soil for 7 days did not significantly affect the insecticidal activity of the OBs compared to
OBs in soil in the absence of earthworms (P >0.05). We conclude that incorporation of
contaminated OBs artificial soil results in per os infections in S. frugiperda second instar larvae,
that the earthworm’s gut pH is not a limiting factor for OB passage through the intestine and that
E. fetida can effectively disperse SfMNPV OBs under laboratory conditions. This represents a
previously unrecognized mechanism of baculovirus dispersal in the environment that is likely to
have important implications in the persistence of OB populations in soil reservoirs.
Keywords: Spodoptera frugiperda, Baculoviridae, artificial soil, Eisenia fetida.

[10]
I. INTRODUCCIÓN Y MARCO TEÓRICO
Spodoptera frugiperda - una plaga importante del maíz.
El gusano cogollero Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) es una especie
polífaga originaria de América y ampliamente distribuida por regiones tropicales y subtropicales.
En su forma adulta es capaz de migrar grandes distancias durante los meses de verano. Este
insecto tiene un amplio rango de hospederos, sin embargo durante sus estadios larvarios prefiere
alimentarse de plantas de las familias Fabaceae, Poacae y Graminaceae. Dentro de los principales
cultivos que se ven mermados por esta especie se encuentran el maíz, sorgo, algodón, soya, papa,
tomate y pastos forrajeros (Sparks, 1979, Van Huis 1981, Pashley, 1988, Martínez-Castillo,
2003). En México es un serio problema en casi todos los estados con producción de maíz
(Andrews, 1988).
El ciclo de vida de S. frugiperda tiene una duración de 30 días, pero la fenología del cultivo y la
temperatura ambiente son los principales factores que afectan al desarrollo de la larva,
alimentación, longevidad, fecundidad y viabilidad de los huevos. Los huevos se encuentran
cubiertos con escamas de coloración gris-rosada y se ponen en masas de 70 a 300 (Fig. 1). La
eclosión de los huevos se da entre 3 y 5 días y pasan por 5 o 6 estadios larvarios antes de
completar su desarrollo en aproximadamente 20 días. Las larvas pupan en el suelo y este estado
se extiende durante 9 a 13 días. Los adultos o palomillas nocturnas miden entre 28-32 mm, son
de colores café grisáceo o beige, con marcas oscuras y rayas pálidas en la parte central de las
alas, las alas traseras presentan una coloración blanquecina (Jiménez, 2003).

[11]
Figura 1. Estadíos de desarrollo de Spodoptera frugiperda: a) huevos; b)
larvas; c) pupa; d) adulto y larva provocando daños al maíz (e, f)
La mayoría de las aplicaciones de insecticidas químicos en maíz están dirigidas al control de este
insecto. Sin embargo, durante los últimos años se encuentra en desarrollo una alternativa
prometedora y sostenible para su control que consiste en un nucleopoliedrovirus múltiple de S.
frugiperda (SfMNPV, género Alphabaculovirus, familia Baculoviridae), que causa epizootias en
las poblaciones larvarias de este insecto (Williams et al., 1999; Barrera et al., 2011).

[12]
Baculoviridae
La familia Baculoviridae es la más estudiada en lo referente a grupos de virus patógenos de
insectos (Tanada y Kaya, 1993). Dentro del contexto histórico, el estudio de los baculovirus se
encuentra inicialmente relacionado a las mermas ocasionadas por patologías desconocidas en la
cría del gusano de seda, Bombyx mori (Lepidoptera: Bombycidae). Mediante el uso y mejora de
herramientas como el microscopio óptico durante mediados del siglo XIX, se determinó que una
de las principales causas de muerte se caracterizaba por la presencia de cuerpos de oclusión
poliédricos (OBs por sus siglas en inglés) dentro de las células. Posteriormente, a finales de 1940
se determinó la presencia de viriones en forma de bastón mediante microscopía electrónica.
Finalmente esta patología comenzó a denominarse como virus de la poliedrosis nuclear
(posteriormente nucleopoliedrovirus o NPV, por sus siglas en inglés). El término baculovirus fue
sugerido por el Dr. Mauro Martignoni alusivo a la forma de varilla o bastón (baculum=bastón)
que presentaban los viriones (Rohrmann, 2008). Los baculovirus afectan específicamente a
artrópodos. Se han aislado miembros de esta familia en más de 700 especies de insectos, la
mayoría lepidópteros, dípteros e himenópteros. Después de un cambio reciente en su clasificación
(Jehle et al., 2006), la familia consta de cuatro géneros: Alphabaculovirus (NPVs de
lepidópteros), Betabaculovirus (granulovirus [GV] de lepidópteros), Gammabaculovirus (NPVs
de himenópteros) y Deltabaculovirus (NPVs de dípteros). Todos ellos poseen un estrecho
espectro de hospederos, elevada patogenicidad y virulencia, los cuales son atributos ideales para
ser desarrollados como insecticidas biológicos. Además, no representan riesgos ambientales
mayores en términos de residuos tóxicos, efectos a organismos no blanco o riesgos para los
agricultores que aplican los bioinsecticidas basados en baculovirus (Caballero et al., 2009).

[13]
Los NPVs son entidades estructuralmente complejas. Consisten en un genoma empacado dentro
de una nucleocápsida que está envuelta en una membrana de lipoproteínas para formar el virión,
el cual está incluido en una matriz de proteína que representa el poliedro o cuerpo de inclusión.
El genoma de los baculovirus consta de una molécula circular de ADN de doble cadena que
oscila entre 80 y 180 kilopares de bases (kpb) y codifica entre 90 y 180 genes (Van Oers y Vlak,
2007). La unidad genética de los baculovirus es la nucleocápsida, que es una estructura compacta
con un diámetro comprendido entre 30-60 nm y una longitud entre 250-300 nm, cuya función
principal es transportar el genoma viral hacia la célula huésped. Los viriones de los baculovirus
comparten la misma estructura básica y constituyen la unidad morfológica de respuesta para la
infección viral. En la mayoría de los baculovirus existen dos tipos de viriones con idéntico
genotipo y diferente fenotipo. Los viriones ocluidos (ODVs) y viriones brotantes (BVs) cumplen
roles diferentes durante el ciclo biológico del virus (Fig. 2).
La morfología de los OBs es un carácter taxonómico muy importante en los baculovirus. Por
ejemplo, el tamaño y la forma de éstos, permite diferenciar a dos de los géneros más
representativos, los alfabaculovirus (NPVs) y los betabaculovirus (GVs). Los betabaculovirus
presentan OBs pequeños denominados gránulos con un diámetro de 150 nm y 400-600 nm de
longitud, compuestos por una proteína denominada granulina y normalmente un único virion. En
cambio, los OBs de los alfabaculovirus pueden variar entre 0.5 y 15 µm de diámetro: el tamaño
medio oscila entre 0.8 y 2 µm, con formas de cubo, dodecaedro y tetraedro, pero generalmente
suelen tener formas irregulares (Volkman, 2007); aquellos que sólo contienen ODVs de tipo
simple, son denominados SNPVs, y los que contienen ODVs de tipo múltiple, se denominan
MNPVs (Fig. 3) (Francki et al., 1991)

[14]
Figura 2. Componentes estructurales de baculovirus (adaptado de
Rhormann, 2008).
Figura 3. Fenotipos de OBs: los alfabaculovirus (NPV) presentan mayor tamaño
que los betabaculovirus (GV) y contienen viriones derivados de la inclusión
(ODVs). Los NPV pueden ser múltiples (MNPV) o simples (SNPV)
dependiendo del número de nucleocápsides presentes en cada virion (adaptado
de Slack y Arif, 2007).

[15]
Las larvas infectadas por el virus presentan características fácilmente reconocibles que consisten
en modificaciones en la coloración y brillantez del integumento, cambios en el comportamiento
de la larva y ablandamiento del tejido que se vuelve frágil y de fácil ruptura. Poco antes de la
muerte la larva trepa la planta y queda suspendida de los pseudópodos y posteriormente el cuerpo
de la larva se rompe, liberando millones de OBs (Fig. 4).
Figura 4. Cadáver de una larva de Spodoptera
frugiperda presentando licuefacción característica por
infección de SfMNPV.
Proceso de transmisión
El tracto gastrointestinal del insecto es el lugar donde se inicia la infección. Se compone de tres
secciones: intestino anterior, medio y posterior. El intestino anterior se encuentra recubierto por
una cutícula quitinosa y tiene una función de almacenamiento. Una válvula separa el intestino
anterior del intestino medio. En este último se secretan enzimas y ocurre la absorción de los
alimentos. En la entrada y salida del intestino medio el pH es cercano a 7 pero en la región

[16]
central puede variar de 10 a 12 (Dow, 1992). El intestino medio presenta una membrana o matriz
peritrófica, la cual protege la superficie del intestino del material abrasivo de los alimentos y
limita el acceso de microorganismos. Es desgastada por el paso de los alimentos y se regenera a
partir de células epiteliales localizadas cerca a la válvula anterior.
La transmisión horizontal de los baculovirus sucede exclusivamente en los estadios larvarios del
insecto, siendo los primeros estadios los más susceptibles. Ocurre por vía oral o per os cuando los
insectos consumen alimento contaminado con OBs. Los OBs y las partículas de alimento viajan
por el intestino anterior hacia el intestino medio. Los lepidópteros tienen un intestino medio
alcalino (pH 10-11) que provoca la disolución de la poliedrina y, por ende, la liberación de los
viriones ocluidos en el lumen de intestino medio. Estas partículas virales liberadas son los ODVs,
que atraviesan la membrana peritrófica (MP) para, a continuación penetrar en las células del
intestino medio, donde realizan un primer ciclo de replicación (Fig. 5). En estas células brota el
otro tipo de virión conocido como BVs, que penetra la lámina basal con la ayuda de proteasas
(Caballero et al., 2009).
Figura 5. Proceso de infección de los baculovirus (Caballero et al., 2009)

[17]
Los BVs se forman cuando la nucleocápsida abandona la célula en la que ha sido sintetizada y
adquiere una envoltura al atravesar la membrana citoplasmática de la célula huésped por los
puntos donde se han insertado glicoproteínas codificadas por los virus (Slack y Arif, 2007).
También existe la transmisión transovarial y transóvulo, aunque con frecuencia relativa menor.
De manera general, la larva muere a partir de sexto o séptimo día, aunque este periodo es
variable y está dictaminado principalmente por la cepa viral empleada, estado de desarrollo y las
condiciones ambientales (Melo-Molina, 2001). Generalmente, las células afectadas son las de la
matriz traqueal, el cuerpo graso, el tejido muscular, el nervioso, el reproductivo, el glandular, los
hemocitos y las células epiteliales. Poco antes de la muerte, los insectos infectados se vuelven
pálidos, flácidos y frecuentemente suben a las partes apicales de las plantas donde mueren. El
cuerpo del insecto se fragmenta liberando millones de cuerpos de inclusión, que contaminan el
follaje inferior permitiendo la transmisión a otras larvas susceptibles que se alimentan de las
planta y finalmente producirán el siguiente ciclo de infección (Caballero et al., 2001)
El SfMNPV como insecticida.
La estrategia de control de S. frugiperda se basa principalmente en aplicaciones de insecticidas
químicos, particularmente organofosforados y piretroides. Sin embargo, durante los últimos años
se encuentra en desarrollo una alternativa prometedora que consiste en el SfMNPV, que de forma
natural causa epizootias en las poblaciones larvarias de este insecto.
Escribano et al. (1999) caracterizaron diferentes aislados del SfMNPV procedentes de Argentina,
Nicaragua y Estados Unidos, que resultaron ser estructural y genéticamente similares, aunque S.

[18]
frugiperda demostró una mayor susceptibilidad a la cepa de Nicaragua (SfNIC). Esta cepa ha
sido sujeta a diversos estudios enfocados a su desarrollo como un insecticida biológico en
México y Centroamérica.
En Nicaragua durante 1988 y 1989, se realizaron aplicaciones del SfMNPV en cultivos de maíz
logrando un control entre el 70 y 80% en larvas de S. frugiperda, lo cual fue equiparable al uso de
un insecticida estándar (Williams et al., 1999). Por otro lado, experimentos de campo mostraron
que la prevalencia de infección en amplios rangos de aplicación de 6x1012 OBs/ha fue del 40%
con una incidencia añadida del 15-20% de parasitismo por avispas parasíticas (Martínez-Castillo
et al., 2003).
Ecología de los baculovirus
Los baculovirus tienen una función ecológica como reguladores de poblaciones de insectos. Por
varias décadas han sido utilizados como agentes de control biológico, ejemplos de ello son el uso
de los nucleopoliedrovirus en el control de Anticarsia gemmatalis Hübner (Lepidoptera:
Noctuidae), Helicoverpa zea Boddie (Lepidoptera: Noctuidae) y Lymantria dispar Linnaeus
(Lepidoptera: Limantriidae), entre otras especies plaga (Slack y Arif, 2007).
Dispersión
La dispersión de los baculovirus ocurre por medio de factores bióticos y abióticos. La dispersión
abiótica de los baculovirus ocurre principalmente a través del viento y la lluvia y en cambio la
dispersión biótica es promovida por el huésped, parasitoides, depredadores, saprófagos y/o por
efectos de la actividad humana (Caballero et al., 2001; Hosteter y Bell, 1985).

[19]
Dispersión abiótica
Los NPV pueden persistir por décadas en suelos sin disturbios donde pueden ser transportados
abióticamente del insecto hospedero a la planta para iniciar epizootias. En algunos casos los NPV
pueden permanecer viables hasta 41 años (Thomson, et al., 1981). Olofsson (1988) comprobó
que partículas de polvo pueden causar epizootias, aunque el transporte suelo-planta puede
entenderse como un aspecto crítico de la correlación de epizootias con el suelo como reservorio,
este aspecto ha sido estudiado a detalle solo recientemente. Fuxa et al., (2001) clasificó patrones
en la distribución vertical y horizontal en el suelo en parcelas de cultivos. Fuxa y Richter (1996,
1999, 2001, 2006) estudiaron los efectos de manejos agrícolas y la lluvia en relación a la
supresión de poblaciones de insectos por NPV del suelo, incluyendo la rotación de cultivos.
También relacionaron el transporte de NPV del suelo a las plantas para especificar tipos de suelo,
mezclas, velocidades de viento, cantidades de lluvia y altura de la planta encontrando que
concentraciones de 5x108 a 5x106 OBs en las capas superficiales del suelo fueron transportadas a
las plantas por el viento o la lluvia, pero no incluyeron profundidades o distancias como un factor
experimental. Posteriormente, Fuxa et al. (2007) estudiaron las distancias y profundidades de las
cuales los OBs pueden ser transportados del suelo a la planta bajo condiciones conductivas o no
conductivas de transporte abiótico, resaltando que el uso de estos virus como control a largo
plazo depende de predecir epizootias y control de plagas basadas en las concentraciones de NPV
en el suelo y esto podría enormemente mejorar con el entendimiento del transporte suelo-planta.
La disponibilidad de rutas de transmisión de NPV en el ambiente determina en gran medida el
curso de eventos durante epizootias naturales y después de las aplicaciones de virus.

[20]
Dispersión biótica
En la dispersión biótica, el hospedero es lo más importante. Los huéspedes pueden transmitir
infecciones secundarias en la misma generación larvaria así como infecciones primarias en
generaciones tardías. Desde que las larvas son infectadas por la ingestión de follaje contaminado,
la cantidad y distribución del virus en el follaje durante el periodo de alimentación larvario
determina el nivel de infección primaria en una población. Una larva enferma es la fuente de
dispersión del virus más concentrada disponible para su dispersión, las hojas de las plantas
usualmente se vuelven infecciosas para la siguiente generación larvaria. Existen otros agentes
causales secundarios como: Aves, parasitoides, escarabajos, chinches, crisopas, tijerillas entre
otros depredadores que pueden contaminar el follaje con sus heces y estar involucrados en la
dispersión del virus (Castillejos et al., 2001). Los OBs de los baculovirus pasan a través del
intestino de algunos depredadores sin una pérdida significativa de infectividad (Beekman, 1980;
Abbas, 1988; Castillejos et al., 2001). Esto se debe a que la mayoría de los insectos depredadores
tienen un intestino ácido. En cambio, la alta alcalinidad en los intestinos de lepidópteros fitófagos
degrada la matriz proteínica de los OBs (Granados y Lawler, 1981). Para la mayoría de los
baculovirus el principal reservorio de OBs es el suelo donde pueden permanecer activos por
periodos largos de hasta varios años (Fuxa, 2004; Caballero et al., 2009, Peng et al., 1999).
Las lombrices de tierra
Dentro de los principales miembros de la biota edáfica se encuentran las lombrices de tierra
(Annelida: Oligochaeta). Estos animales modifican algunas propiedades físicas del suelo tales

[21]
como la agregación, estabilidad y la porosidad al excavar galerías (Lavelle y Spain, 2001) por lo
que se les ha denominado ingenieros del ecosistema (Lavelle et al., 1997) y también tienen
incidencia sobre las propiedades químicas y biológicas, como la tasa de descomposición de la
materia orgánica, disponibilidad de nutrientes, composición y actividad de microorganismos,
junto con otros invertebrados del suelo (Domínguez et al., 2004).
Han sido estudiadas ampliamente por su papel como organismos modificadores del suelo.
Existen diversos aspectos de su ecología que están aun pobremente entendidos, entre ellos se
encuentran las interacciones entre diferentes especies de lombrices, microorganismos, raíces
vegetales, varias formas de materia orgánica y componentes minerales del suelo y sus
implicaciones para procesos como el ciclo del carbono (Zhang y Hendrix, 1995; Jegou et al.,
1988; Pulleman et al., 2005). Debido a sus hábitos saprófitos generalmente viven en ambientes
en donde coexisten con una gran cantidad y diversidad de microorganismos.
La diversidad de las lombrices de tierra puede ser estudiada bajo dos grandes puntos de vista:
taxonómica y funcional (Fragoso et al., 1997). En el primer caso el número de especies varía en
función de la escala (regional vs. local) y del origen geográfico (nativas vs. exóticas). En el
segundo caso, las especies se separan por categorías ecológicas o gremio funcional (Fragoso,
2001). Las características ecológicas propuestas por Bouche (1972, 1977) y Lavelle (1983) son
las más aceptadas e incluyen tres grandes grupos funcionales de lombrices:
(i) las especies epigeas viven en el horizonte orgánico, habitantes de la hojarasca,
pigmentadas y consumidoras de 100% materia orgánica, (Domínguez y Gómez-
Brandón, 2010).

[22]
(ii) las especies endógeas, habitantes del suelo, no pigmentadas y consumidoras de tierra,
subdivididas en poli, meso y oligohúmicas según el contenido de materia orgánica
(Lavelle, 1983),
(iii) las especies anécicas, habitantes del suelo pero suben a la superficie para alimentarse
Hábitos alimenticios
El análisis de los contenidos del tracto digestivo de las lombrices ha revelado la presencia de una
amplia variedad de material orgánico. Piearce (1978) encontró fragmentos de pasto y otras hojas
vegetales, raíces, células de algas, sedas o quetas de lombrices, semillas, hifas o esporas de
hongos, protozoarios, fragmentos de cutícula de artrópodos, y humus amorfo de una gran
variedad de especies de plantas. Sin embargo, existe una considerable superposición en las dietas
de las diferentes especies. Mientras las lombrices, como grupo, se alimentan de un amplio rango
de materiales, se ha demostrado la existencia de niveles significativos de selección por diferentes
especies (Neilson y Boag, 2003). Incluso el tamaño de partícula del material orgánico tiene
fuerte influencia en el crecimiento y la fecundidad de algunas especies.
Las lombrices asimilan nutrientes de una diversa variedad de materiales, pero su estrategia de
alimentación generalmente consiste en ingerir grandes cantidades ya que presentan una baja
asimilación del material que consumen. En el intestino, la actividad microbiana resulta
estimulada por condiciones favorables en la parte anterior; en el intestino medio se realiza la
digestión de la materia orgánica del suelo, los productos de esta digestión son parcialmente
absorbidos en la parte posterior del intestino. Esto sugiere que este proceso puede resultar en la
asimilación del 3-19% de la materia orgánica en el suelo en un tiempo de tránsito de 30 min a 2-4
horas. (Lavelle y Spain, 2001).

[23]
El tiempo que le toma a la lombriz pasar el alimento por el tracto digestivo depende de la
estrategia de alimentación. El tiempo de tránsito gastrointestinal de Lumbricus terrestris, una
especie anécica, es de 12 a 20 horas (Edwards y Lofty, 1977). Asimismo, para una especie
epigea como L. rubellus, tarda de 6 a 8 horas (Daniel y Anderson, 1992), mientras que para una
especie epigea como E. fetida es de 3 a 4 horas (Edwards y Lofty, 1977).
Las diferentes estrategias de alimentación en las diferentes especies de lombrices de tierra les
permiten coexistir por la explotación de diferentes recursos alimenticios y hábitat (Edwards y
Bohlen, 1992). La mayoría de las comunidades contienen de 3 a 6 especies. De las altas latitudes
hacia el ecuador los grupos dominantes sucesivos son epigeos, anécicos, mesooligohúmicos y
oligohúmicos endógeos (Edwards y Bohlen, 1996; Lavelle, 1983).
Karsten y Drake (1995) encontraron que ciertos microorganismos aeróbicos y anaeróbicos son
más numerosos en los intestinos de lombrices epigeas que las especies endógeas. Los resultados
sugieren que los microorganismos tienen la capacidad de crecer en condiciones anaeróbicas
donde son mucho más abundantes en el intestino de las lombrices que en el suelo del cual fueron
obtenidos, estos resultados concuerdan con los observados por Barois y Laven (1986) en la
lombriz tropical Pontoscolex corethrurus (Glossoscolecidae, Oligochaeta).
La biología de Eisenia fetida
Eisenia fetida (Savigny, 1826) es una especie originaria de Europa pero de amplia distribución,
de fácil manejo y/o cultivo. Es una especie de áreas templadas y ha sido la más utilizada en la
biodegradación de residuos orgánicos o lombricompostaje. Tolera un rango amplio de
temperatura y humedad, lo que la hace resistente y dominante en relación con otras lombrices

[24]
composteras. Esta lombriz se desarrolla bien en ambientes de pH 5-7, a temperaturas de 20 a 28
°C. La temperatura óptima para su crecimiento es de 25°C y el contenido de humedad óptimo es
del 80-85%. En condiciones adecuadas, su ciclo de vida abarca 45-52 días. Los juveniles
alcanzan la madurez en 21-30 días. Las cópulas ocurren cerca de la superficie y las puestas
inician en un entorno posterior a las 48 h después de la cópula. La tasa de producción es de 0.35-
0.5 capullos por día. El pH es uno de los factores determinantes en el adecuado establecimiento
de poblaciones de esta especie que claramente prefiere condiciones que van de neutras a
ligeramente ácidas tolerando valores de pH entre 4 a 9; un pH inferior de 5 o uno superior de 9
causa una alta mortalidad. Asimismo, la producción de capullos es óptima a pH de 5 a 6 pero se
reduce en pH mayores de 7 (Domínguez y Gómez-Brandón 2010).
Algunas especies de lombrices son ampliamente reconocidas como una herramienta de prueba en
evaluaciones de toxicología en suelo. La mayoría de los estudios recientes en lombrices
terrestres se han llevado a cabo enfocándose en metales pesados, bifenilos policlorados (PCBs) e
insecticidas químicos y reguladores de crecimiento. Se considera que las lombrices son de fácil
manejo y crianza, y que son sensibles a los efectos causados por contaminantes en el suelo
(Callahan et al., 1994). Por lo tanto, E. fetida se ha convertido en un invertebrado modelo para
evaluar los efectos de compuestos xenobióticos en suelos (Spurgeon et al., 2002). De hecho la
OECDE (1984) he establecido un protocolo estándar de evaluación. Sin embargo, la interacción
entre OBs y la fauna del suelo ha sido muy poco estudiada, dado la importancia de las lombrices
de tierra en la remoción del suelo y su abundancia en suelos agrícolas. En este trabajo se
consideró pertinente examinar su papel en la dispersión de OBs del NPV de S. frugiperda.

[25]
II. OBJETIVOS
Objetivo general
El objetivo del presente estudio es determinar, bajo condiciones de laboratorio, si una lombriz de
tierra como E. fetida tiene la capacidad de redistribuir los OBs del SfMNPV en un suelo, así
como establecer si existe algún efecto adverso del contacto de los OBs con la lombriz en la
patogenicidad de los mismos.
Objetivos específicos
1. Determinar la relación concentración-mortalidad de OBs en larvas de segundo estadio
de S. frugiperda alimentadas con un suelo estándar mediante la técnica de
incorporación a dieta semisintética.
2. Medir el pH intestinal de E. fetida mediante el uso de indicadores de pH.
3. Determinar si E. fetida tiene la capacidad de redistribuir los OBs verticalmente en un
suelo.
4. Evaluar el efecto de la actividad de E. fetida sobre la patogenicidad de los OBs
presentes en el suelo.
III. HIPÓTESIS
La lombriz de tierra Eisenia fetida tiene la capacidad de redistribuir los OBs de un baculovirus
en el suelo durante su proceso de alimentación y excavación de galerías, sin afectar a su
actividad insecticida.

[26]
IV. MATERIALES Y MÉTODOS
Cría de insectos
Se estableció un pie de cría de S. frugiperda proveniente de la Universidad Autónoma de
Michoacán en los laboratorios de la Red de Manejo Biorracional de Plagas y Vectores del
Instituto de Ecología, A.C. La colonia se mantuvo a temperatura constante (25 ± 2ºC), con un
fotoperiodo de 16:8 h luz/obscuridad. Las larvas fueron criadas sobre una dieta semisintética,
adaptada de Caballero et al., (2001) a base de harina de soya y germen de trigo.
Cria de E. fetida
Se obtuvo un pie de cria de E. fetida del área de lombricompostaje del Instituto de Ecología, A.C.
Las lombrices se mantuvieron en composta con pulpa de café y adicionalmente se agregó
estiércol de vaca para su alimentación durante un periodo de treinta días antes de su uso en los
experimentos de laboratorio.
Amplificación de SfMNPV
Se individualizó un grupo de 100 larvas de tercer estadio en recipientes plásticos con tapa de 15
ml. A cada recipiente se agregó una larva y un trozo de dieta artificial contaminado
superficialmente con 10 µl de una suspensión de 5 x 108 OBs/ml del SfMNPV. Cuatro días
después de la inoculación se cambió la dieta contaminada por dieta no tratada y se revisó la
mortalidad de las larvas durante los 6 días post-inoculación. Las larvas muertas se colectaron y
guardaron a 4°C. Se maceró cada larva infectada y se agregó en un volumen de 500 µl de

[27]
polisorbato 80 (Tween 80) al 0.1% (vol./vol.). Se agitó en un vortex durante 30 s y después se
pasó la suspensión por un filtro de malla plástica para eliminar los restos de tegumento presentes.
La suspensión fue centrifugada a 2319 g durante 5 minutos y el sobrenadante se desechó. A la
pastilla de OBs se le añadieron 500 µl del detergente Tween 80, repitiendo el proceso anterior.
Posterior al segundo centrifugado, se lavaron los restos de Tween 80 con 1 ml de agua destilada y
se realizó un tercer centrifugado a 2319 g. Se resuspendió la pastilla de OBs con 100 µl de agua
destilada y se guardó a 4 ºC.
De una alícuota de la suspensión madre se realizaron tres diluciones de 1/10, 1/100, 1/1000. Se
realizó un conteo de OBs presentes en la suspensión 1/1000 con el microscopio óptico con
contraste de fases a 400x y una cámara de conteo Neubauer, en donde se colocaron 10 µl de la
suspensión de OBs. Cada conteo se realizó por triplicado. Para calcular la concentración de OBs
en la suspensión se utilizó la siguiente formula: Concentración de OBs = (número de OBs
contados × 5 × factor de dilución)/1x10-4. Finalmente se prepararon suspensiones para las
siguientes concentraciones de OBs/ml: 5x104, 5x105, 5x106, 5x107, 5x108, 5x 109.
Mezcla de suelo y estimación de pH
Se elaboró una mezcla de suelo artificial a base de 10% (peso/peso) de turba de esfagno, 20%
caolinita y 70% arena (OECD, 1984). Todos los elementos minerales se cribaron en una malla de
2 mm; el esfagno y la arena fueron lavados, secados a temperatura ambiente en lotes de 1 kg
antes de la preparación del suelo y se revolvieron manualmente en un recipiente plástico durante
15 min.
El pH del suelo se determinó a través del método As-02, medido en agua potenciométricamente
en suspensión sobrenadante de una mezcla de relación suelo:agua 1:2 de acuerdo a la Norma

[28]
Oficial Mexicana (NOM-021-SEMARNAT-2000). El procedimiento consintió en pesar 10g de
suelo en un frasco de plástico de boca ancha al cual se le adicionaron 20 ml de agua destilada al
frasco que contenía suelo, se agitó la mezcla de suelo: agua a intervalos de 5 min. durante 30
min. Se dejó reposar la mezcla de suelo durante 15 min. y finalmente se tomó la medida de pH
con un potenciómetro.
Estimación de la relación concentración de OBs en suelo-mortalidad
Se preparó dieta artificial semisintética y se colocó un volumen de 50 ml en un recipiente plástico
de 250 ml de capacidad. Cuando la dieta alcanzaba una temperatura inferior a 50 °C se
incorporaba una muestra de 10 g de suelo artificial contaminado con 1 ml de suspensión de OBs
de SfMNPV. La mezcla se revolvió vigorosamente con una cuchara plástica para incorporar el
suelo contaminado con OBs en la dieta. Se individualizaron lotes de 30 larvas de segundo estadio
de S. frugiperda y se les colocó un trozo de dieta artificial contaminada durante cuatro días de
exposición. Después de este tiempo, se colocó un trozo de dieta artificial no contaminada y
diariamente se registró la mortalidad por poliedrosis, u otras causas.
Mediante tinción Giemsa se analizó la presencia o no de infección viral en las larvas muertas en
el periodo de prueba de seis días post exposición. Para esto, se tomó una pequeña porción de la
larva muerta con la ayuda de un palillo de madera y se realizó un frotis en un portaobjeto. El
frotis se fijó con alcohol metílico de 3-4 min. Después se lavó suavemente con agua de grifo. Se
agregaron gotas de solución Giemsa y se esperó 30 min. Finalmente el frotis se lavó con
abundante agua para eliminar el exceso de colorante, se secó a temperatura ambiente y se observó
al microscopio a 1000X con una gota de aceite de inmersión para poder observar los OBs
característicos del virus.

[29]
Estimación del pH intestinal de E. fetida
Las lombrices se sacrificaron sumergiéndolas por dos segundos en agua caliente. Se disectaron
140 lombrices adultas cliteladas de E. fetida utilizando grupos de 20 lombrices por cada indicador
de pH. La disección se realizó por la parte ventral. Una vez expuesto el intestino, se agregaron
dos gotas de los siguientes colorantes indicadores de pH para cada individuo: rojo congo a 0.5%
(pH 3.0-5.0); naranja de metilo a 0.1% (pH 3.1-4.4); verde de bromocresol 0.1% (pH 4.0-5.6);
rojo metilo a 0.5% (pH 4.2-6.3); azul de bromotimol 0.1% (pH 6.9-7.6); indigo carmín 0.5% (pH
11.6-14.0). Se realizó otra prueba, en lotes de 10 individuos por indicador, en las cuales se extrajo
el intestino completo y se sumergió en una gota de cada uno de los indicadores; observando si se
presentaba un cambio de coloración durante cinco minutos.
Distribución vertical de lombrices en terrarios experimentales
Se diseñaron terrarios elaborados con placas de vidrio con base y paredes de poliestireno
expandido de 30 x 15 x 1 cm, sujetos con pinzas tipo clip metálicas para determinar la
supervivencia de las lombrices en las unidades experimentales (Fig.6). Para acondicionar las
lombrices de pie de cría a las condiciones de laboratorio, 72 h antes del inicio de las pruebas, se
colocaron las lombrices en el suelo artificial a 25 ±1 °C.

[30]
Figura 6. Terrarios para determinar la
supervivencia de las lombrices en suelo artificial.
Cada terrario se llenó con 250 g (peso seco) de suelo artificial y posteriormente se
añadieron 150 ml de agua destilada para humedecer el suelo a 60% de humedad relativa.
Un grupo de 10 lombrices por cada terrario se lavaron y se pesaron con una balanza
analítica (Cuad.1). Posteriormente se colocaron en la parte superficial de cada unidad
experimental, la cual se incubó a 25 °C, en oscuridad. A las 2, 24, 48, y 72 h posteriores se
observó la actividad de descenso de las lombrices en términos de la máxima profundidad
alcanzada en cada unidad. El experimento se replicó ocho veces.

[31]
Cuadro 1. Peso promedio de E. fetida
utilizadas en cada replica (n=30).
Réplica
Peso promedio ± EE
(mg)
1 376 ± 36
2 354 ± 24
3 308 ± 29
4 316 ± 22
5 318 ± 16
6 312 ± 24
7 396 ± 21
8 381 ± 32
Dispersión de OBs en suelo artificial por E. fetida
Para determinar la habilidad de las lombrices en dispersar los OBs, un total de 45 terrarios se
llenaron con 250 g de suelo estándar cada uno, y posteriormente se añadieron 250 ml de agua
destilada para humedecer a capacidad de campo cada unidad experimental. En cada experimento
Se evaluaron dos tratamientos y un control; el primer tratamiento consistió en colocar sobre la
superficie 1 ml de agua con una suspensión de 1x109 OBs en de agua sin lombrices, otro
consistió en colocar treinta lombrices sobre la superficie por unidad experimental y después se
agregó únicamente 1 ml de suspensión con 1x109 OBs. Para el control se agregó 1 ml de agua
destilada junto con treinta lombrices.
En cada unidad se realizó una toma de muestras de 6 g de suelo húmedo con cucharas plásticas
desechables en cuatro intervalos de profundidades; 0-2, 6-8, 14-16, 22-24 cm y en tres diferentes
tiempos, a los 1, 7 y 15 días después de iniciar el experimento. Para evitar contaminación
cruzada, los controles se muestrearon primero, posteriormente el tratamiento sin lombrices y por
último el tratamiento con lombrices. Cada tratamiento, profundidad de muestreo e intervalo de

[32]
tiempo se replicó en cinco ocasiones. Las muestras de suelo se incorporaron a 50 ml de dieta y se
revolvieron vigorosamente con una cuchara plástica hasta homogeneizar el suelo con la dieta.
Posteriormente se repartieron trozos de la mezcla en vasos plásticos de 15 ml. En cada vaso se
colocó una larva de S. frugiperda en segundo estadio. Las larvas se dejaron 4 días con el trozo de
dieta contaminada con suelo y después se retiró la dieta contaminada y se colocó un trozo de
dieta limpia en cada vaso. Cada muestra de suelo fue repartida entre un grupo de 30 larvas en
total. La mortalidad por virus u otras causas se registró diariamente durante los seis días
posteriores al cambio de la dieta.
Efecto de la actividad de E. fetida en la patogenicidad de OBs en suelo
Antes de realizar el bioensayo, se individualizaron 30 lombrices del pie de cría con un promedio
de peso de 200 mg y se colocaron en cajas Petri con un círculo de papel filtro de 7 cm de
diámetro y 2 ml de agua destilada. Se dejaron durante 4 días sin alimento para vaciar su intestino.
Posteriormente, con estas lombrices pre acondicionadas, se montó un experimento para
determinar el efecto de la presencia de E. fetida en la patogenicidad de OBs en suelo con dos
tratamientos y un control. En el primer tratamiento sin lombriz, en un vaso de plástico de 20 ml
con tapa perforada, se agregaron 10 g de suelo, 10 ml de agua destilada y 1 ml de una suspensión
de 1 x 109 OBs de SfMNPV. El segundo tratamiento fue idéntico al primero pero con la presencia
de una lombriz. El control consistió en 10 g de suelo estándar, 11 ml de agua destilada y una
lombriz de 200 mg de peso, el tiempo de exposición fue de 7 días. Transcurrido este lapso, se
retiraba la lombriz de cada vaso y se individualizaron en cajas Petri cada uno con un círculo de
papel filtro de 7 cm de diámetro y 2 ml de agua destilada. Se dejaron durante 4 días para vaciar su
intestino y corroborar que habían consumido suelo contaminado con OBs, las excretas se secaron
y pesaron. El contenido de suelo en cada vaso se agregó a 50 ml de dieta artificial y se revolvió

[33]
vigorosamente con una cuchara plástica hasta integrarlo a la dieta. Esta mezcla se repartió en 30
vasos de 15 ml con tapa a cada uno se les colocó una larva de segundo estadio de S. frugiperda.
Este experimento fue replicado 15 veces de forma simultánea. Durante 4 días se permitió que las
larvas se alimentaran de esta mezcla con suelo obtenido de los tratamientos descritos
anteriormente. Después de este tiempo se retiró la dieta contaminada, se colocó un trozo de dieta
limpia y se registró la mortalidad diariamente durante los seis días posteriores.
Análisis estadístico
La relación concentración de OBs en suelo y la mortalidad en larvas de S. frugiperda se estimó
mediante modelo logit. El efecto de la presencia E. fetida en la redistribución vertical de OBs se
analizó mediante el ajuste de modelos generales lineales con distribución binomial. Para esto se
utilizó un proceso de simplificación secuencial de modelo hasta identificar el modelo mínimo
significativo (Crawley, 1993). La presencia de sobredispersión en los datos se tomó en cuenta
mediante un procedimiento de ajuste del parámetro de escalamiento de acuerdo al valor de χ2 de
Pearson calculado para el modelo más completo. Los cambios en la desvianza de cada modelo
ajustado se determinaron de acuerdo a los valores críticos de la distribución χ2 con sus
respectivos grados de libertad. Asimismo la actividad insecticida de OBs expuestos o no a las
lombrices se determinó mediante una prueba de t después de haber ajustado los resultados a un
modelo logit con distribución binomial todos los análisis se realizaron utilizando el programa
GLIM 4.0 (Generalised Linear Interactive Modeling, Natural Algorithms Group, 1994)

[34]
V. RESULTADOS
Estimación de la relación concentración-mortalidad para OBs en suelo
Las larvas de segundo estadio aceptaron la dieta + suelo y se presentaron pocas muertes en los
testigos (5%) y ninguna larva murió por virosis, las larvas testigo lograron sobrevivir hasta pupar,
en cambio, larvas tratadas con diferentes concentraciones de SfMNPV presentaron características
de infección típica después del tiempo de exposición de 4 días.
Se observó que en porcentaje de mortalidad causada por virus, en larvas de segundo estadio de
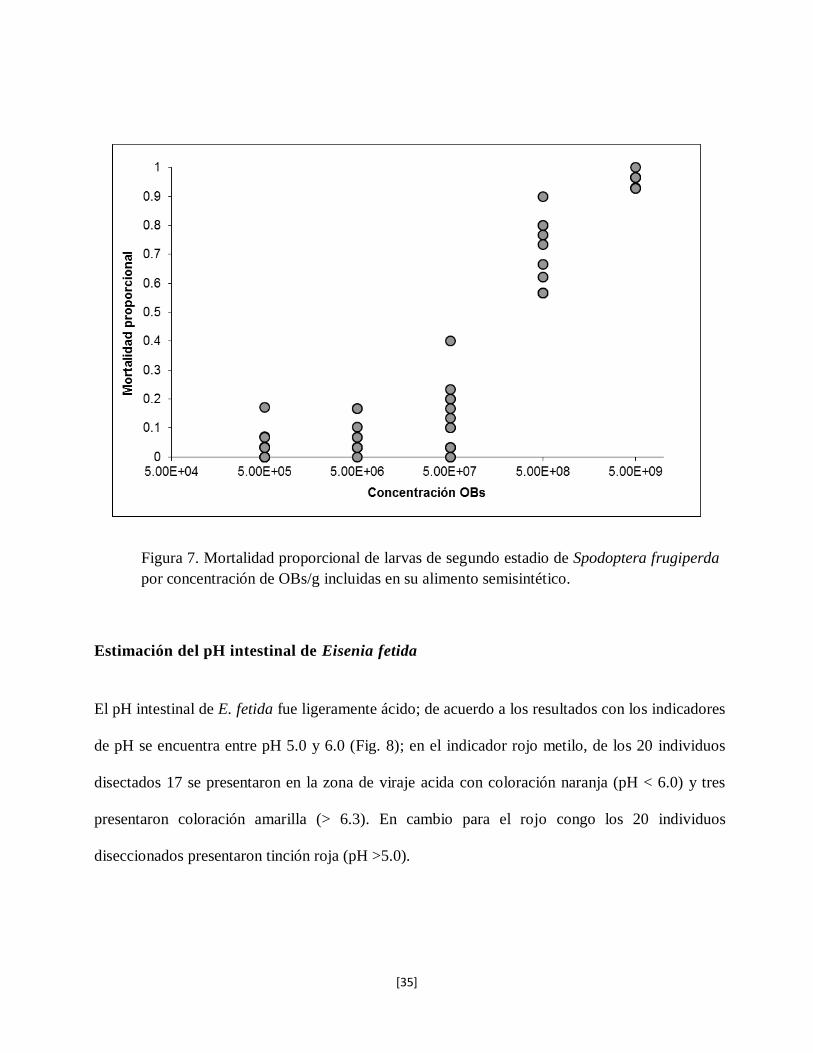
Spodoptera frugiperda varió entre 4 y 92% de acuerdo a la concentración de OBs presentes en la
mezcla de suelo (Figura 7), se fue incrementando la mortalidad proporcional conforme aumentó
la concentración de OBs/g de suelo.
La concentración letal 50% (CL50) se estimó en 2.3 x 108 OBs/g de suelo mediante esta técnica.
La ecuación de la regresión (±EE) fue ln[y] = 0.8639 ±0.0740 x – 16.64 ± 1.409. El parámetro de
escalamiento se ajustó a un valor de 2.8 para tomar en cuenta una ligera sobredispersión en la
distribución de los resultados. Mediante las tinciones Giemsa se confirmó que la muerte de larvas
fue por infección de SfMNPV.
[35]
Figura 7. Mortalidad proporcional de larvas de segundo estadio de Spodoptera frugiperda
por concentración de OBs/g incluidas en su alimento semisintético.
Estimación del pH intestinal de Eisenia fetida
El pH intestinal de E. fetida fue ligeramente ácido; de acuerdo a los resultados con los indicadores
de pH se encuentra entre pH 5.0 y 6.0 (Fig. 8); en el indicador rojo metilo, de los 20 individuos
disectados 17 se presentaron en la zona de viraje acida con coloración naranja (pH < 6.0) y tres
presentaron coloración amarilla (> 6.3). En cambio para el rojo congo los 20 individuos
diseccionados presentaron tinción roja (pH >5.0).

[36]
Figura 8. (a) Disección de intestino, (b) Ejemplo colocación de
colorante sobre el intestino, (c) Intestino disectado sin colorante,
(d) Tinción con rojo congo (pH >5.0), (e) Tinción rojo metilo (pH
< 6.0).
Supervivencia y descenso en suelo artificial
El 81% de los individuos sobrevivió y solo se obtuvo el 3% de mortalidad en los terrarios
con suelo artificial (Fig. 9).
Figura 9. Lombrices vivas después de 72 h en suelo estándar (%), se observó una
alta supervivencia en el suelo artificial

[37]
Se observó el promedio de descensos máximo obtenido dentro de cada intervalo de tiempo. A las
2 h fue de 0 cm, a las 24 h de 7 cm, a las 48 h de 18.5 cm y a las 22 h de 22.5 cm. Se tomó como
registro el máximo descenso de las lombrices observado en cada réplica (Fig. 10).
Figura 10. Promedio de descenso de E. fetida en suelo artificial (± EE). Se tomó
como registro el máximo descenso de las lombrices obtenido en cada réplica
(n=9). Se observó una relación positiva entre el tiempo y la profundidad a partir
de las 24 horas, cuyos valores se incrementaron de 7 (24 h) a 22.5 cm (72 horas)
(8 réplicas).
Redistribución de OBs en suelo artificial
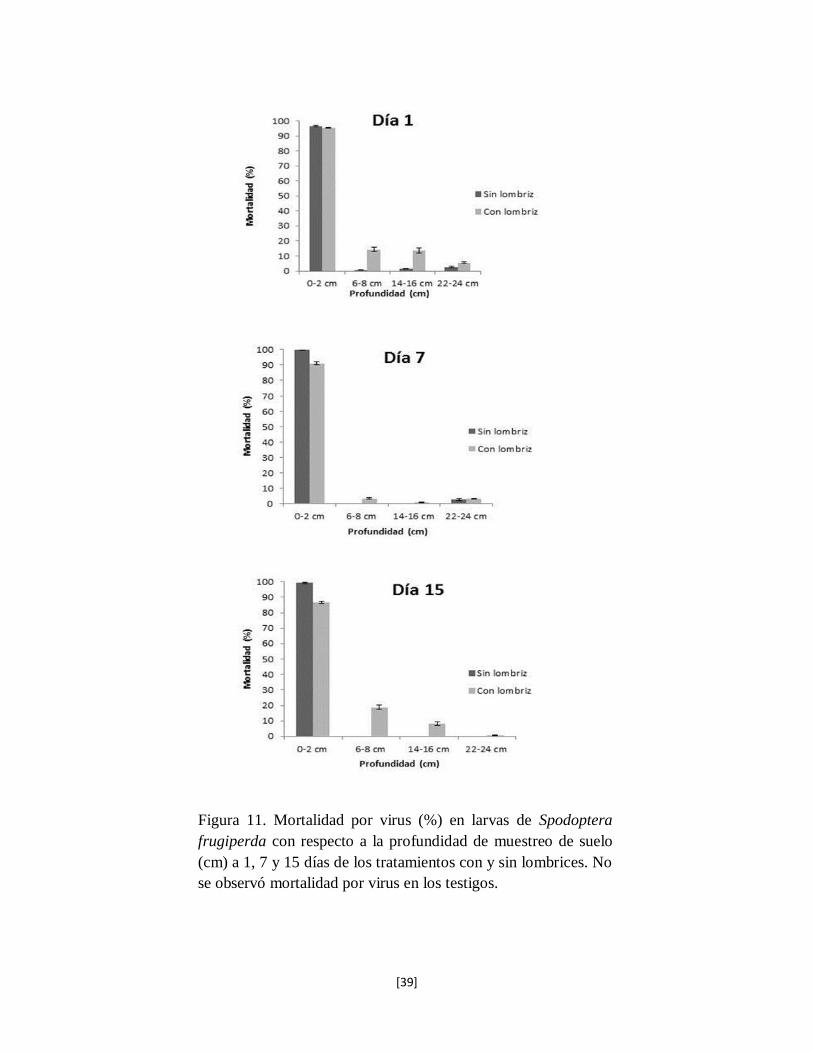
Se observó que las muestras de suelo colectadas en la capa superficial 0-2 cm donde se aplicó el
inoculo, se generaron elevados porcentajes de mortalidad de larvas de S. frugiperda por virus en
los tres diferentes tiempos (1, 7, 15 días); el tratamiento sin lombriz presentó mortalidades del 95,
100 y 99% respectivamente; los tratamientos con lombriz presentaron mortalidades de 95, 91 y
86% para los mismos intervalos de tiempo (Fig. 11). Las capas de suelo de 6-8 cm y 14-16 cm en
el tratamiento sin lombriz presentaron mortalidades inferiores al 1% por virus, en cambio el

[38]
tratamiento con lombriz presentó mortalidades por virosis de 14, 3, 19%, y 14, 1, 8% para las
mismas profundidades. En la última capa de suelo 22-24 cm se presentó una mortalidad inferior
al 3% en los días evaluados en ausencia de lombriz. Para el tratamiento con lombriz se observó
una mortalidad de 6, 3, y 1%. En el tratamiento control no se presentaron muertes por virus y las
larvas sobrevivieron hasta pupar, la mortalidad por otras causas fue del 5%.
Los modelos lineales para lombriz-profundidad, profundidad-tiempo, profundidad, tiempo, y
lombriz, fueron significativos (Cuad.2). La detección de poliedros aumentó significativamente
con el tiempo (P<0.001) y disminuyó significativamente con el aumento de la profundidad del
suelo (P<0.01). Las lombrices aumentaron la dispersión de poliedros.
Cuadro 2. MLG del Efecto de las lombrices, la profundidad y el
tiempo en la distribución de OBs en el suelo experimental.
Factores χ2 Gl P
Lombriz*profundidad*tiempo 4.32 6 0.633(N.S.)
Tiempo*lombriz 1.99 2 0.369(N.S.)
Lombriz*profundidad 47.7 3 <0.001
Profundidad*tiempo 21.1 6 <0.001
Profundidad 1324 3 <0.001
Tiempo 8.85 2 0.012
Lombriz 4.35 1 0.037
[39]
Figura 11. Mortalidad por virus (%) en larvas de Spodoptera
frugiperda con respecto a la profundidad de muestreo de suelo
(cm) a 1, 7 y 15 días de los tratamientos con y sin lombrices. No
se observó mortalidad por virus en los testigos.
[40]
Efecto de la presencia de E. fetida en la patogenicidad de OBs en suelo
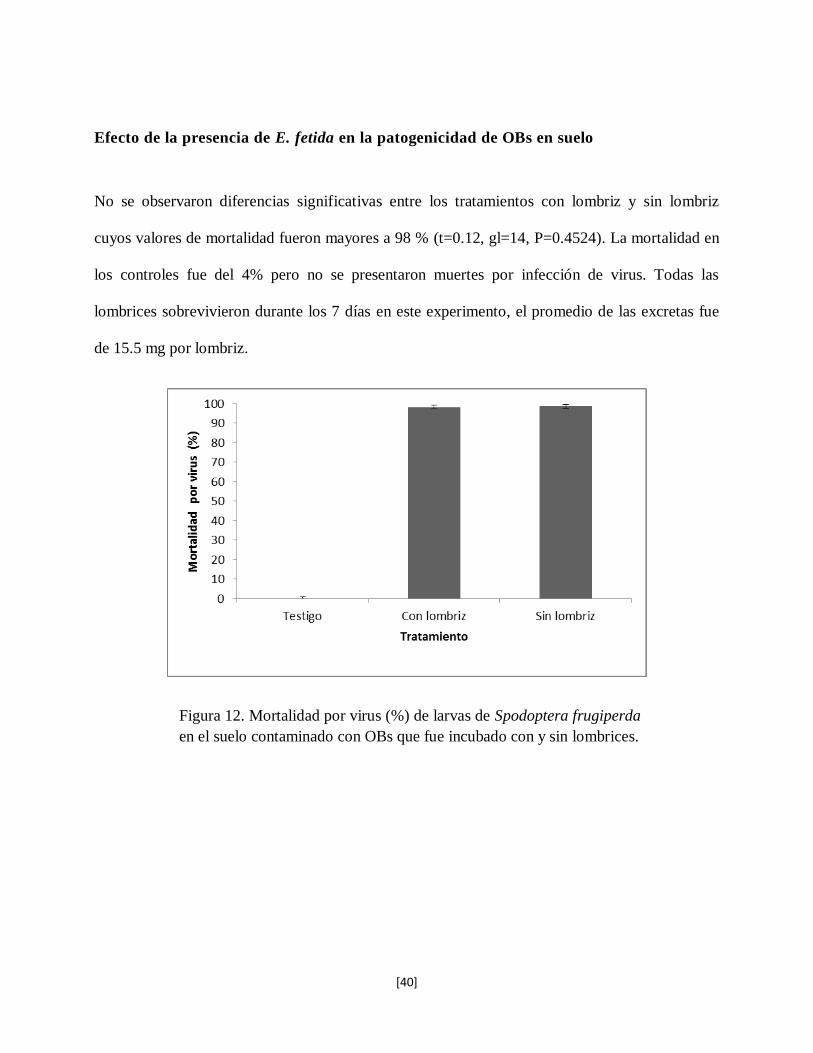
No se observaron diferencias significativas entre los tratamientos con lombriz y sin lombriz
cuyos valores de mortalidad fueron mayores a 98 % (t=0.12, gl=14, P=0.4524). La mortalidad en
los controles fue del 4% pero no se presentaron muertes por infección de virus. Todas las
lombrices sobrevivieron durante los 7 días en este experimento, el promedio de las excretas fue
de 15.5 mg por lombriz.
Figura 12. Mortalidad por virus (%) de larvas de Spodoptera frugiperda
en el suelo contaminado con OBs que fue incubado con y sin lombrices.

[41]
VI. DISCUSIÓN
El objetivo del presente estudio fue determinar bajo condiciones de laboratorio si una lombriz de
tierra, como E. fetida, tiene la capacidad de redistribuir los OBs del SfMNPV en el suelo, así
como establecer si existe algún efecto adverso del contacto de los OBs con la lombriz en la
patogenicidad de los mismos. El hecho de utilizar un suelo estándar facilitó los estudios de
laboratorio y brindó una metodología sencilla pero de gran utilidad para el entendimiento de los
procesos que modulan la interacción OB-suelo. Lo anterior resalta en importancia debido a la alta
variabilidad existente en los suelos naturales y para efectos prácticos, éstos requieren de una
caracterización que resulta difícil de repetir o reproducir de un laboratorio a otro (Fuxa, 2008)
La técnica de incorporación de suelo a la dieta diseñada por Richards y Christian (1999),
adaptada por Murillo et al., (2007), ha demostrado tener más sensibilidad que técnicas basadas en
PCR o procesos como los descritos por Ebling y Holmes (2002), en el aislado de cepas de
baculovirus del suelo. En este trabajo se demostró que la técnica de adicionar un suelo artificial a
la dieta de larvas de segundo estadío de S. frugiperda fue efectiva. La estimación de la CL50 fue
de 2.3 x 108 OBs/g de suelo; superior a lo reportado para la detección del NPV de Helicoverpa
armigera (CL50= 2.15 x 103 OBs/g) en sustratos australianos y OBs de SeMNPV en sustratos de
invernaderos de Almería, España (CL50=2.72 x103 OBs/g de sustrato) (Richards y Christian,
1999; Murillo et al., 2007). Lo anterior, debido a la diferencia en la patogenicidad que presenta el
SfMNPV comparado con las cepas de estos estudios y la susceptibilidad del hospedero.

[42]
Mediante las tinciones Giemsa se confirmó que la muerte de larvas fue por infección de
SfMNPV. Asimismo, entre el quinto y séptimo día posterior a la infección, las larvas mostraban
cambios de coloración, brillo del tegumento, y ablandamiento de tejidos, tornándose frágiles y de
fácil ruptura (Slack y Arif , 2007).
Parece importante evaluar los materiales gelificantes de la dieta, por ejemplo, para este trabajo se
utilizó el agar-agar pero este puede ser substituido por el uso de carragenatos, xantanos o geles de
pectina que podrían mejorar el desempeño de la técnica al gelificar a temperaturas más bajas y
permitir una mejor homogenización del suelo contaminado con OBs y evitar la inactivación del
virus debido a altas temperaturas. La ventilación dentro de los vasos experimentales también fue
clave para evitar el crecimiento de bacterias y hongos sobre la dieta +con suelo y disminuir la
mortalidad de las larvas por otras causas.
A pesar de que el transporte de OBs del suelo-planta es un aspecto crítico para la generación de
nuevos eventos de transmisión, este proceso ha sido estudiado solo recientemente. Existe
evidencia de que algunos baculovirus se adhieren fuertemente a partículas de suelo, donde se dan
interacciones entre minerales en suelos arcillosos con alto contenido de óxidos férricos, atapulgita
y caolín (Christian et al., 2006). La importancia del suelo como reservorio para controlar plagas
de lepidópteros con baculovirus, la aplicación de NPV al suelo para inducir epizootias virales en
plagas de lepidópteros, el efecto de las operaciones agrícolas en el suelo, y el transporte abiótico
del suelo a la planta han sido estudiados por diversos autores (Jaques, 1970, 1974; Young y
Yerian, 1979, 1986; Fuxa y Richter, 1996, 1999, 2007). No obstante, el principal proceso de
dispersión de NPV se debe a la dispersión, distribución y conducta del hospedero, ya que en
estados inmaduros puede redistribuir los baculovirus sobre la planta o el suelo. Por otra parte, la

[43]
transmisión vertical, el canibalismo, la depredación por aves o diversos invertebrados o
parasitoides, también se ha demostrado que dispersan los baculovirus.
Los resultados del presente estudio demuestran que existe un proceso de dispersión que no había
sido reconocido con antelación. Un oligoqueto como E. fetida pudo redistribuir los OBs
verticalmente en las capas intermedias del suelo. Sin embrago, los niveles de dispersión del
SfMNPV en las capas inferiores de suelo fueron bajos; esto se debe principalmente al hábito
conductual de esta especie de permanecer y salir a excretar sobre la superficie. La mortalidad de
larvas de segundo estadío de S. frugiperda fueron más elevadas cuando fueron alimentadas con
suelo contaminado con OBs de la capa de 0-2 cm comparado con suelo de las otras
profundidades, evidentemente porque fue la capa a la cual se aplicó el inóculo. El único estudio
existente al respecto de la interacción de lombrices de tierra y OBs de baculovirus fue realizado
por Olofsson (1989), que alimentó a un cuervo (Turdus merula) con lombrices que consumieron
suelo contaminado con 105 OBs/ml del NPV de Neodipirion sertifer (Hymenoptera:
Diprionidae), demostrando que cuando el ave se alimentaba con lombrices que consumieron
suelo con OBs, las excretas del ave contenían OBs que seguían siendo infectivos para N. sertifer.
En el experimento para verificar si una lombriz podía inactivar o reducir la capacidad insecticida
de los OBs, no se observaron diferencias significativas entre los tratamientos con lombriz y sin
lombriz por lo tanto E. fetida no tuvo un efecto sobre la patogenicidad de los OBs. En este
experimento la lombriz estuvo en contacto externo y consumió suelo contaminado por OBs
durante una semana. El pH del intestino de E. fetida fue ligeramente ácido (pH 5.0-6.0.); por lo
que el pH no debe ser un factor que afecte la patogenicidad de los OBs de SfMNPV durante el
tránsito intestinal. El suelo estándar permitió la supervivencia de las lombrices, el 81% de los
individuos sobrevivió y solo se obtuvo el 3% de mortalidad en las cajas con suelo artificial; este

[44]
suelo artificial ha sido utilizado ampliamente con E. fetida como organismo de prueba en
evaluaciones de toxicología en suelo (OCDE, 1984). A pesar de que se desconoce cómo es la
interacción OBs-suelo - lombriz, se puede extrapolar la cantidad de consumo por el volumen
promedio de suelo excretado por las lombrices al final del experimento, que fue de 15.5 mg por
lombriz. Los resultados de este estudio revelan que las lombrices de tierra tienen la capacidad de
redistribuir los OBs de los NPVs a profundidades importantes en el suelo donde están protegidos
de los efectos nocivos de la radiación UV. Aunque queda por comprobar, es posible que también
pudieran desplazar los OBs de suelos profundos hacia la superficie donde pueden ser
transmitidos a larvas susceptibles que se alimentan de plantas en suelos contaminados por OBs.
Esto representa un nuevo mecanismo de dispersión de OBs y podría tener un papel importante en
la persistencia de poblaciones de virus en suelos agrícolas sujetos a procesos de cultivo como
periodos de arado y barbecho que permiten que las poblaciones de lombrices remuevan
cantidades importantes de suelo entre las capas superiores e interiores del mismo. Sin embargo E.
fetida es una especie epigea que no tiene el hábito conductual de bajar grandes distancias sobre el
perfil de suelo de tal manera sería importante la evaluación de otras especies como P. corethrurus
que es una especie endogea y geofaga.
Para seguir con esta línea de investigación se debe considerar escalar estas pruebas en
invernadero o campo para determinar si otros factores asociados a las prácticas agrícolas tienen
un efecto antagónico o sinérgico con las lombrices y pueden con ello permitir una mejor
dispersión de OBs por parte de estos oligoquetos. Así mismo es pertinente estimular la
alimentación de lombrices terrestres mediante el uso de abonos orgánicos para aumentar los
volúmenes de suelo consumido por lombrices e incrementar con ello la interacción y generando
mayores probabilidades de dispersión de OBs de baculovirus en el suelo, conservar las
poblaciones de lombrices en los suelos agrícolas mediante métodos no invasivos que disminuyan

[45]
las poblaciones y la diversidad de lombrices dentro de los agroecosistemas y con ello mejoren no
solo la dispersión de OBs de baculovirus, sino también las propiedades físicas y químicas del
suelo de cultivo.
VII. CONCLUSIONES
1. El sustrato utilizado en los ensayos de laboratorio funciona como suelo reservorio de los
OBs del SfMNPV, el sustrato no generó efectos adversos en la persistencia y
patogenicidad del inóculo de OBs bajo condiciones de laboratorio.
2. El pH del intestino de E. fetida es ligeramente ácido y no supone un factor limitante para
el paso de OBs de SfMNPV a través del intestino.
3. Eisenia fetida puede interactuar con los OBs de SfMNPV en el suelo artificial sin
inactivarlos durante un periodo de al menos siete días en el laboratorio.
4. Eisenia fetida es capaz de dispersar los OBs verticalmente bajo condiciones de
laboratorio, a profundidades de hasta 22-24 cm en las unidades experimentales utilizadas
en estos estudios.

[46]
VII. REFERENCIAS
Abbas, M.S.T. (1988). Interactions between nuclear polyhedrosis virus, host and predators.
Zeitsch. Pflanzenkrank. Pflanzenschutz 95: 606-610.
Andrews, K. L. (1988). Latin America research on Spodoptera frugiperda (Lepidoptera:
Noctuidae). Fla.Entomol. 71: 630-653.
Barois, I. y Lavelle, P. (1986). Changes in respiration rate and some physicochemical properties
of a tropical soil during transit through Pontoscolex corethrurus (Glossoscolecidae, Oligochaeta).
Soil Biol. Biochem., 18: 539 541.
Barrera, G., Simón, O., Villamizar, L., Williams, T. y Caballero, P. (2011). Spodoptera
frugiperda multiple nucleopolyhedrovirus as a potential biological insecticide: Genetic and
phenotypic comparison of field isolates from Colombia. Biol. Control 58: 113-120.
Barret, J., Brownright, A., Primavera, M., Retnakaran, A. y Palli, S. (1998). Concomitant
primary infection of the midgut epithelial cells and the hemocytes of Trichoplusia ni by
Autographa californica nucleopolyhedrovirus. Tissue Cell 30: 602–616.
Belda, J, Justicia, L., Pascual, F. y Cabello, T. (1994). Distribución espacial de Spodoptera
exigua (Hübner) (Lepidoptera; Noctuidae) en cultivo de pimiento en invernadero. Bol. San. Veg.
Plagas 20: 303-316.
Beekman, A.G. (1980). The infectivity of polyhedral of nuclear polyhedrosis virus (N.P.V.) after
passage through the gut of an insect predator. Experientia 36: 858-859.
Blissard, G.W. y Rohrmann, G. F. (1990). Baculovirus diversity and molecular biology. Annu.
Rev. Entomol. 35: 127-155.

[47]
Bouché, M.B. (1977). Strategies lombriciennes. Ecol. Bull. 25: 122-132.
Bossuyt, H., Six, J. y Hendrix, P.F. (2004). Rapid incorporation of carbon from fresh residues
into newly formed stable microaggregates within earthworm casts. Eur. J. Soil Sci. 55: 393-399.
Caballero, P., Murillo, R., Muñoz, D. y Williams, T. (2009). El nucleopoliedrovirus de
Spodoptera exigua (Lepidoptera: Noctuidae) como bioplaguicida. Rev. Colomb. Entomol. 35:
105-115.
Caballero, P., López–Ferber, M. y Williams, T. (2001). Los baculovirus y sus aplicaciones
como bioinsecticidas en el control biológico de plagas. Phytoma-España, 518pp.
Callahan, C.A., Shiraz, M.A. y Neuhauser, E.F. (1994). Comparative toxicity of chemicals to
earthworms. Environ. Toxicol. Chem. 13: 291-298.
Castillejos, V., García, L., Cisneros, J., Goulson, D., Cave, R.D., Caballero, P. y Williams,
T. (2001). The potential of Chrysoperla rufilabris and Doru taeniatum as agents for dispersal of
Spodoptera frugiperda nucleopolyhedrovirus in maize. Entomol. Exp. Appl. 98: 353-359.
Clavijo, G., Williams, T., Muñoz, D., Caballero, P. y López-Ferber, M. (2010). Mixed
genotype transmission bodies and virions contribute to the maintenance of diversity in an insect
virus. Proc. R. Soc. B. 277: 943–951.
Christian, P., Richards, A. y Williams, T. (2006). Differential adsorption of occluded and
nonoccluded insect-pathogenic virus to soil-forming minerals. Appl. Environ. Microbiol. 72:
4648-4652.
Cory, J. y Myers, J. (2003). The ecology and evolution of insect baculoviruses. Annu. Rev.
Ecol. Syst. 34: 239-272.

[48]
Cuadrado, I.M. y Viñuela, E. (1998). Resistencia a los pesticidas en los cultivos hortícolas.
Fundación para la investigación agraria en la provincia de Almería, La Cañada, Almería, España.
212p.
Daniel, O. y Anderson J. (1992). Microbial biomass and activity in contrasting soil materials
after passage through the gut of the earthworm Lumbricus rubellus Hoffmeister. Soil Biol.
Biochem. 24: 465-470.
Domínguez, J. (2004). State of the art and new perspectives on vermicomposting research. En:
C.A. Edwards (Ed.). Earthworm ecology, 2nd Ed.CRC/Lewis Press, Boca Raton, USA. P.p 401-
424.
Dominguez, J., Aira, M. y Gómez-Brandón, M. (2009). El papel de las lombrices de tierra en
la descomposición de la materia orgánica y el ciclo de nutrientes. Ecosistemas. 18(2):20-31pp.
Dow, J. A. (1992). pH gradients in lepidopteran midgut. J Exp Biol 172, 355-375.
Edwards, C.A. y Lofty, J.R. (1977). Biology of Earthworms. Halsted, New York, USA.
Edwards, C.A. y Bohlen.P.J. (1996). Biology and ecology of earthworms. Chapman and Hall,
London, U.K., 426 pp.
Edwards, C.A. y Bohlen, P.J. (1992). The effects of toxic chemicals on earthworms. Rev.
Environ. Contam. Toxicol. 125: 23-100.
Engelhard, E.K., Kam-Morgan, N.W., Wasburn, J.O. y Volkmann, L.E. (1994). The insect
tracheal system: A conduit for the systematic spread of Autographa californica M nuclear
polyhedrosis virus. Proc. Natl. Acad. Sci. USA. 91: 2324-3227.
Escribano, A., Williams, T., Goulson, D., Cave, R. D., Chapman, J. W., y Caballero, P.
(1999). Selection of a nucleopolyhedrovirus for control of Spodoptera frugiperda (Lepidoptera:

[49]
Noctuidae): structural, genetic, and biological comparision of four isolates from the Americas. J.
Econ.Entomol. 92: 1079-1085.
Fuxa, J.R. (2004). Ecology of insect nucleopolyhedroviruses. Agric. Ecosyst. Environ. 103: 27-
43.
Fuxa, J.R. (2008). Threshold concentrations of nucleopolyhedrovirus in soil to initiate infections
in Heliothis virescens on cotton plants. Microb. Ecol. 55: 530–539.
Fuxa, J.R. y Richter, A.R. (2001). Quantification of soil-to-plant transport of recombinant
nucleopolyhedrovirus: effects of soil type and moisture, air currents, and precipitation. Appl.
Environ. Microbiol. 67: 5566-5170.
Fuxa, J.R. y Richter, A.R. (1999). Classical biological control in an ephemeral habitat with
Anticarsia gemmatalis nucleopolyhedrovirus. Biocontrol 44: 403-419.
Fuxa, J.R. y Richter, A.R. (1996). Effect of agricultural operations and precipitations on vertical
distributions of nuclear polyhedrosis virus in soil. Biol.Control 6: 324-329.
Fragoso, C., J. G.G. Brown, J.C. Patrón, E. Blanchart, P. Lavelle, B. Pashanasi, B.K.
Senapati y T. Kumar. (1997). Agricultural intensification, soil biodiversity and agroecosystem
function in the tropics: the role of earthworms. Appl. Soil Ecol. 6: 17-35.
Francki, R.I.B., Faquet, B.C.M., Kawaoka, Y., Donatelli, I., Guo, Y. y Webster, R.G. (1991).
Classification and nomenclature of viruses. Arch. Virol. 2 (Suppl.), 119.
Granados, R.R. y Lawler, K. (1981). In vivo pathway of infection of Autographa californica
baculovirus invasion and infection. Virology 108: 297-308.

[50]
Hodgson, D., Vanbergen, A. J., Watt, A. D., Hails, R.S. y Cory, J.S. (2001). Phenotypic
variation between naturally co-existing genotypes of a lepidopteran baculovirus. Evol. Ecol. Res.
3: 687–701.
Hostetter, D. L. y Bell, M.R. (1985). Natural dispersal of baculoviruses in the environment, p.
249-284. En: K. Maramorosch y K.E. Sherman (ed.), Viral insecticides for biological control.
Academic Press Inc., Orlando.
Jehle, J.A, Lange, M., Wang, H., Wang, H.Z. y Hauschild, R. (2006). Molecular identification
and phylogenetic analysis of baculoviruses from Lepidoptera. Virology. 346:180-93.
Jegou, D., Cluzeau, D., Balesdent, J. y Trehen, P, (1998). Effects of four ecological categories
of earthworms on carbon transfer in soil. Appl. Soil Ecol. 9: 249-255.
Jiménez Fernández, J.A. (2003). Aislamiento y polimorfismo genético del nucleopoliedrovirus
de Spodoptera frugiperda (Smith) en 9 municipios de la región del Soconusco Chiapas, México.
Tesis de licenciatura. Universidad de Ciencias y Artes de Chiapas. 82 pp.
Karsten G. R. y Drake, H. (1995). Comparative assessment of the aerobic and anaerobic
microfloras of earthworm guts and forest soils. Appl. Environ. Microbiol. 61: 1039-1044.
Kauscheke, E., Mohrig W. y Cooper, E.L. (2007). Coelomic fluid proteins as basic
components of innate immunity in earthworms. Eur. J. Soil Biol. 43: 110-115.
Lasa, R., Ibañes, I., Belda, J.E., Williams, T. y Caballero, P. (2007). Efficacy of Spodoptera
exigua multiple nucleopolyhedrovirus as a biological insecticide for beet armyworm control in
greenhouses of southern Spain. Biocontrol Sci. Technol. 17: 221-232.
Lavelle, P. (1983). The structure of earthworm communities. En Earthworm Ecology, from
Darwin to Vermiculture, (ed. J. E. Satchell), Chapman and Hall, Londres. 449-466 pp.

[51]
Lavelle, P. (1988). Earthworm activities in the soil system. Biol. Fertil. Soils. 6: 237-251.
Lavelle, P. y Spain, A.V. (2001). Soil ecology. Kluwer, Norwell.
Lavelle, P., Bignell, D., Lepage, M., Wolters, V., Roger, P., Ineson, P., Heal, O.W. y
Dhillion, S. (1997). Soil function in a changing world: the role of invertebrate ecosystem
engineers. Eur. J. Soil Biol. 33, 159–193.
Martínez-Castillo, A. (2003). Lethal and sublethal interaccions between optical brighteners and
nucleopolyhedrovirus in Spodoptera frugiperda (J.E.Smith) (Lepidoptera: Noctunidae) Tesis
doctoral. Departamento de Producción Agraria Universidad Pública de Navarrra. 111 pp.
Melo Molina, G. (2001). Desarrollo de una metodología para la detección de baculovirus del
gusano cogollero en muestras de suelo. Tesis de Licenciatura, Instituto Tecnológico de Tuxtla
Gutiérrez, Chiapas. 72 pp.
Muñoz, D., Murillo, R., Krell, P.J., Vlak, J.M. y Caballero, P. (1999). Four genotypic variants
of a Spodoptera exigua nucleopolyhedrovirus (Se-SP2) are distinguishable by a hypervariable
genomic region. Virus Res. 59: 61-74.
Murillo, R., Elvira, S., Muñoz, D., Williams, T. y Caballero, P. (2006). Genetic and
phenotypic variability in Spodoptera exigua nucleopolyhedrovirus isolates from greenhouse soils
in southern Spain. Biol. Control 38: 157-165.
Neilson, R. y Boag, B. (2003). Feeding preferences of some earthworm species common to
upland pastures in Scotland. Pedobiologia 47: 1-8.
Olofsson, E. (1989). Transmission agents of the nuclear polyhedrosis virus of Neodiprion sertifer
(Hym.:Diprionidae). Entomophaga 33:373-380.

[52]
Organization for Economic Cooperation and Development (OECD). (1984). Earthworm,
acute toxicity test. Guidelines for testing of chemicals N°207. OECDE, Paris, Francia, 9 pp.
Pashley, D.P. (1988). Current status of fall armyworm host strain. Fla. Entomol. 71: 227-234.
Peng, F., Fuxa, J.R., Richter, A.R. y Johnson, J. (1999). Effects of heat-sensitive agents. Soil
type, moisture, and leaf surface on persistence of Anticarsia gemmatalis (Lepidoptera:
Noctuidae) nucleopolyhedrovirus. Environ. Entomol. 28: 330-338.
Pierce, T.G. (1978). Gut content of some lumbricid earthworms. Pediobiologia. 18: 153-157.
Pulleman, M.M., Six, J., Uyl, A., Marinissen, J.C.Y. y Jongmans, A.G. (2005). Earthworms
and management affect organic matter incorporation and microaggregate formation in
agricultural soils. Appl. Soil Ecol. 29: 1-15.
Richards, A.R. y Christian, P.D. (1999). A rapid bioassay screen for quantifying
nucleopolyhedroviruses (Baculoviridae) in the environment. J. Virol. Meth. 82: 63-75.
Rohrmann, G. (2008). Introduction to the baculoviruses and their taxonomy.
www.ncbi.nlm.nih.gov/ bookshelf/picrender.fcgi?book=bacvir...pdf.
Slack, J.M. y Arif, B. (2007). Per os infectivity of baculoviruses and the structure of occlusion-
derived virus. Adv. Virus Res. 69: 99–165.
Sparks, A.N. (1979). A review of the biology of the fall armyworm. Fla. Entomol. 62:82-87.
Spurgeon, D.J., Weeks, J.M. y Van Gestel, C.A. (2002). A summary of eleven years progress
in earthworm ecotoxicology. Pedobiologia. 47: 588–606.
Stoltz, D.B., Povan, C. y Dacuhna, A.B. (1973). Nuclear polyhedrosis virus: a possible example
of de novo intra nuclear membrane morphogenesis. J. Gen. Virol. 9: 145-150.

[53]
Simón, O., Williams, T., López-Ferber, M. y Caballero. P. (2004). Virus entry or the primary
infection cycles are not the principal determinants of host specificity of Spodoptera spp.
nucleopolyhedroviruses. J. Gen. Virol. 85: 2845–2855.
Tanada, Y., y Kaya, H.K. (1993). Insec pathology. Academic Press, San Diego, CA.
Thompson G.C., Scott, D.W. and Wickman, B.E. (1981). Long term persistence of the nuclear
polyhedrosis virus of the Douglas-fir tussock moth, Orgyia pseodotsugata (Lepidoptera:
Lymantriidae) in forest soil. Environ. Entomol. 10 : 254-255.
Ukena, K. Oumi, T., Matsushima, O., Ikeda, T., Fujita, T., Minakata, H. y Nomoto, K.
(1995). A novel gut tetradecapeptide isolated from earthworm, Eisenia fetida. Peptides 16: 995-
999.
Van Huis, A. (1981). Integrated pest management in the small farmer’s maize crop in Nicaragua.
Meded. Landbou. Wageningen. 81-6, 221 pp.
Van Oers, M.M. y Vlak, J.M. ( 2007). Baculovirus genomics. Curr. Drug Targets 8: 1051-1068.
Volkman, L.E. (1997). Nucleopolyhedrovirus interactions with their insect host. Adv. Virus.
Res. 48: 313-347.
Volkman, L.E. (2007). Baculovirus infectivity and the actin cytoskeleton. Curr. Drug Targets 8:
1075-1083.
Williams, T., Goulson, D., Caballero, P., Cisneros, J., Martínez, A. M., Chapman, J. W.,
Roman, D.X., y Cave, R.D. (1999). Evaluation of a baculovirus bioinsecticide for small-scale
maize growers in Latin America. Biol. Control. 14: 67-75.

[54]
Young, S.Y. y Yearian, W.C. (1986). Movement of a nuclear polyhedrosis virus from soil to
soybean and transmission in Anticarsia gemmatalis (Hübner) (Lepidoptera: Noctuidae)
populations on soybean. Environ. Entomol. 15: 573-580.
Zhang, Q.L. y Hendrix, P.F. (1995). Earthworm (Lumbricus rubellus and Aporrectodea
caliginosa) effects on carbon flux in soil. Soil Sci. Son. Am. J. 59: 816-823.




















