
Localization of putative biochemical messengers during Eisenia foetida (Annelida, Oligochaeta)...
13
TISSUE AND CELL, 1993 25 (3) 311-323 0 1993 Longman Group UK Ltd. CARLA FALUGI* and CAMILLA DAVOLH LOCALIZATION OF PUTATIVE BIOCHEMICAL MESSENGERS DURING E/SE/V/A FOETlDA (ANNELIDA, OLIGOCHAETA) DEVELOPMENT Keywords: Annelids, development, neurogenesis, catecholamines, cholinesterase, neuro- peptides ABSTRACT. This histochemical-immunohistochemical study was performed on the earthworm Eiseniafoetida at different developmental stages, to investigate the presence and distrihutlon of cholinergic molecules (AChE, BuChE, a-hungarotoxin-binding sites), several hiogenic amines (SHT and catecholamines), and some immunologically-related peptides (somatostatin. FMRF-amide, VIP, substance-p, bomhesin). The results showed that the pattern of labelling for the markers is different at different stages. In summary, cholinesterases appeared widely distributed in the early embryonic stages. They then were localized in particular areas of the developing nerve and muscle tissues, whereas in newborn and adult earthworms they were restricted to ventral muscular fibers and to some CNS cells. Biogenic amines were constantly present in the embryonic and adult nervous tissues Immunologically-related peptides were detectable after organogenesis. Our results provide indirect evidence for a role of cholinesterases in regulating early inter- cellular communications, neurogenesis and myogenesis. and support the hypothesis that somr conservative sequences of messenger peptides arose very early in evolution. Introduction In the last few years evidence has been accumulating that signal molecules, including hormones, neurotransmitters and other extracellular chemical messengers, similar to those acting in intercellular communications of vertebrates, appeared early in the course of evolution (Roth ef al., 1982). It has long been known that classical neurotransmitters, such as catecholamines, serotonin, ace- tylcholine (ACh) and ACh-correlated enzymes and receptors, are widely dis- tributed throughout the animal kingdom (for reviews see Gardner and Walker, 1982; Lunt and Olsen, 1988). More recently, immu- nocytochemical, biochemical and molecular biological studies provided substantial evi- ‘Istituto di Anatomia Comparata, Universita di Geneva. Viale Benedetto XV 5, I-16132 Geneva, Italy. tlstituto di Medicina Sperimentale, Consiglio Nazion- ale delle Ricerche (CNR). Viale Marx 15. I-00137 Roma. Italy. Correspondence to Dr Camilla Davoli. Received 20 November 1992 dence that structurally related peptides are found in many different phyla, including annelids (Al-Yousuf, 1990; De Loof and Schoofs, 1990). Furthermore, putative pep- tide messengers have been recognized to occur in the nervous system of all invert- ebrates, even in the most simple, such as coelenterates, in which they probably act as neurotransmitters, local neuromodulators and/or neurohormones (Jesse, 1986: Thorndyke, 1989). In the earthworm Eisenia foetida, neurons containing several vertebrate-type messen- ger peptides and serotonin-like material have been localized and carefully mapped by immunohistochemical studies in the cerebral ganglion (Lkider et al., 1987). Moreover, thyroglobulin(Tg)-like immunoreactive neu- rons have been detected in both brain and ventral nerve cord (Marcheggiano et al.. 1985), raising the possibility that acetyl- cholinesterase (AChE) molecules, showing structural homology with a terminal portion of Tg amino acid sequence (Schumacher et al., 1986) could be involved in the immu- 311
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Localization of putative biochemical messengers during Eisenia foetida (Annelida, Oligochaeta)...

TISSUE AND CELL, 1993 25 (3) 311-323 0 1993 Longman Group UK Ltd.
CARLA FALUGI* and CAMILLA DAVOLH
LOCALIZATION OF PUTATIVE BIOCHEMICAL MESSENGERS DURING E/SE/V/A FOETlDA (ANNELIDA, OLIGOCHAETA) DEVELOPMENT
Keywords: Annelids, development, neurogenesis, catecholamines, cholinesterase, neuro- peptides
ABSTRACT. This histochemical-immunohistochemical study was performed on the earthworm Eiseniafoetida at different developmental stages, to investigate the presence and distrihutlon of cholinergic molecules (AChE, BuChE, a-hungarotoxin-binding sites), several hiogenic amines (SHT and catecholamines), and some immunologically-related peptides (somatostatin. FMRF-amide, VIP, substance-p, bomhesin).
The results showed that the pattern of labelling for the markers is different at different stages. In summary, cholinesterases appeared widely distributed in the early embryonic stages. They then were localized in particular areas of the developing nerve and muscle tissues, whereas in newborn and adult earthworms they were restricted to ventral muscular fibers and to some CNS cells. Biogenic amines were constantly present in the embryonic and adult nervous tissues Immunologically-related peptides were detectable after organogenesis.
Our results provide indirect evidence for a role of cholinesterases in regulating early inter- cellular communications, neurogenesis and myogenesis. and support the hypothesis that somr conservative sequences of messenger peptides arose very early in evolution.
Introduction
In the last few years evidence has been accumulating that signal molecules, including hormones, neurotransmitters and other extracellular chemical messengers, similar to those acting in intercellular communications of vertebrates, appeared early in the course of evolution (Roth ef al., 1982). It has long been known that classical neurotransmitters, such as catecholamines, serotonin, ace- tylcholine (ACh) and ACh-correlated enzymes and receptors, are widely dis- tributed throughout the animal kingdom (for reviews see Gardner and Walker, 1982; Lunt and Olsen, 1988). More recently, immu- nocytochemical, biochemical and molecular biological studies provided substantial evi-
‘Istituto di Anatomia Comparata, Universita di Geneva. Viale Benedetto XV 5, I-16132 Geneva, Italy.
tlstituto di Medicina Sperimentale, Consiglio Nazion- ale delle Ricerche (CNR). Viale Marx 15. I-00137 Roma. Italy.
Correspondence to Dr Camilla Davoli.
Received 20 November 1992
dence that structurally related peptides are found in many different phyla, including annelids (Al-Yousuf, 1990; De Loof and Schoofs, 1990). Furthermore, putative pep- tide messengers have been recognized to occur in the nervous system of all invert- ebrates, even in the most simple, such as coelenterates, in which they probably act as neurotransmitters, local neuromodulators and/or neurohormones (Jesse, 1986: Thorndyke, 1989).
In the earthworm Eisenia foetida, neurons containing several vertebrate-type messen- ger peptides and serotonin-like material have been localized and carefully mapped by immunohistochemical studies in the cerebral ganglion (Lkider et al., 1987). Moreover, thyroglobulin(Tg)-like immunoreactive neu- rons have been detected in both brain and ventral nerve cord (Marcheggiano et al.. 1985), raising the possibility that acetyl- cholinesterase (AChE) molecules, showing structural homology with a terminal portion of Tg amino acid sequence (Schumacher et al., 1986) could be involved in the immu-
311

312
noreaction (Swillens et al., 1986). However, our preliminary attempts at the identification of AChE in the cerebral ganglion of Eisenia foetida have been unsuccessful (Falugi et al., 1987,1988), whereas our recent investigation on iodination activity in the same animal suggested a functional role for Tg-like mol- ecules (Davoli et al., 1989, 1991). On the other hand, ACh in annelids has long been implicated as a neuromuscular transmitter, while a role for this substance as a transmitter in the central nervous system (CNS) is less clear (for review see Gardner and Walker, 1982). Besides the classical function of AChE as an enzyme specifically degrading ACh in nervous cholinergic synapses and endplates, many in vitro and in vivo experiments sug- gested that cholinesterases, both AChE and butyrylcholinesterase (BuChE), act in the regulation of cell proliferation and neurite growth (Layer, 1991). Many authors have also advanced the hypothesis that neuro- transmitters and their correlated enzymes may be involved in the induction processes during morphogenesis (for reviews see Buz- nikov, 1990; Layer, 1991).
Extending our previous research on AChE activity, we report here further findings on the presence and localization of some rep- resentative messenger molecules during dif- ferent developmental stages of Eisenia foetida and discuss their possible functions.
Materials and Methods
Eisenia foetida embryos and newly hatched worms were obtained from cocoons of earth- worms kept under controlled laboratory con- ditions (Davoli et al., 1989). Cocoons were removed every 3 days from adult worms in breeding activity, and raised on horse manure at 25°C. Experiments were carried out using embryos isolated from cocoons at
FALUGI AND DAVOLI
different time intervals from deposition, newborns l-5 days old, and sexually mature earthworms. Embryonic specimens and cephalic and body segments, obtained from newborns and adults sacrificed by exposure to -4O”C, were treated with different fixa- tives according to the various staining methods used.
Histochemical methods
Cholinesterase activity. The specimens were fixed with 1% glutaraldehyde in phos- phate buffered saline (PBS) pH7.4, for 20 min at 4°C and rinsed with several changes of 0.1 M maleate buffer pH 6. Cholinesterase (ChE) activity was detected by direct col- oring thiocholine method of Karnovsky and Roots (1964) using as substrate acetyl- thiocholine iodide (AcTChI). This substrate is hydrolysed mainly by acetylcholinesterase (AChE, E.C. 3.1.1.7.), but also, though with lesser extent, by pseudocholinesterase (butyrylcholinesterase, BuChE, E.C. 3.1.1.8.). For this reason, to check further the specifity of the enzyme reaction, acetyl- P-methylthiocholine iodide (AcMTChI), specifically hydrolysed by AChE, and butyr- ylthiocholine iodide (BuTChI), mainly hydrolysed by BuChE, were also used. The reaction was followed by stereomicroscopy and lasted about 3 hr. All the substrates were obtained from Sigma Chem. Co., USA.
For the determination of enzyme speci- ficity by specific inhibition of the his- tochemical reaction, samples were pre- incubated for 20min in 0.1 mM specific inhibitors in Tyrode solution; then they were incubated in the enzyme reaction mixture, containing the same inhibitors at the same concentration. The inhibitors were: eserine (physostigmine, BDH, England) specific for both AChE and BuChE (useful for eval- uation of the presence of non-specific ester-
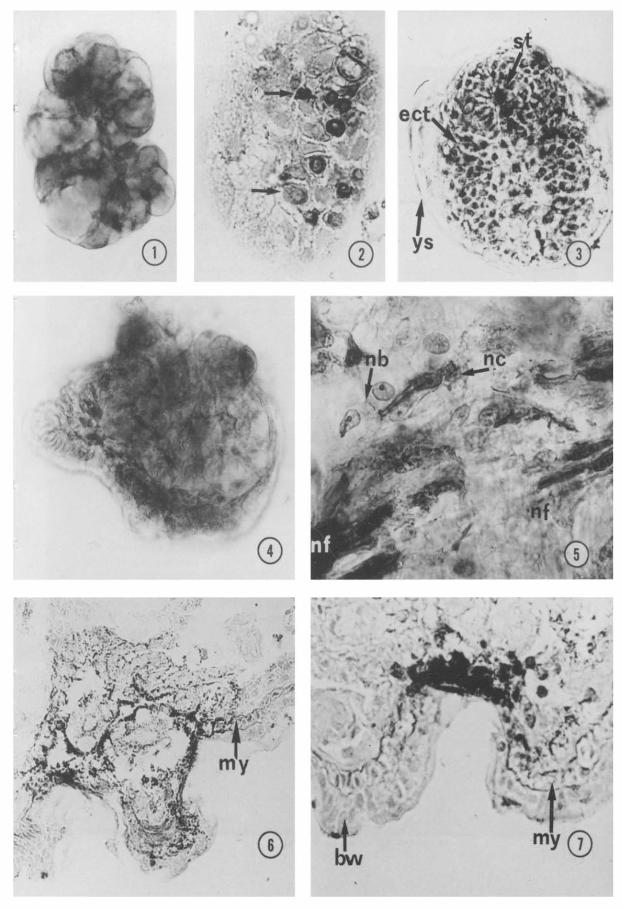
Figs l-4. ChE histochemical staining in early embryo. Diffuse positivity in a planula stage, mounted in fofo (Fig. 1); arrows point to positive teloblasts in a gastrula section (Fig. 2); positive ectoteloblasts in alate gastrula, mounted in tofo (Fig. 3) and in a 4-day embryo. mounted in tofo (Fig. 4). ect, ectoteloblastic band ; St, stomodeum; ys, transient yolk sac. Fig. 1 X800; Fig. 2 XK@; Fig. 3 X600; Fig. 4 X506
Fig. 5. ChE positivity in developing ventral nerve cord of a 5-day embryo. nb, neuroblast border; nc, nerve cell; nf, nerve fibers. x 1OCG.
Figs 6,7. ChE positive cells and fibers in a sense bud (Fig. 6) and a sensorial pit (Fig. 7) of a ‘i-day embryo. bw, body wall; my. myoblasts. Fig. 6 x3MI; Fig. 7 x500.

314
ases) and BW 284c51 (anticholinesterase, Burroughs-Wellcome Labs., USA) specific only for AChE. After the enzyme reaction, samples were washed in distilled water and dehydrated in graded ethanol. Then the smallest embryos were mounted in tuto with DPX, while the other samples were embed- ded in JB4 resin (Polysciences, USA) and cut in 6mn thick sections by a glass knife, using a Pyramitome (LKB, Sweden). His- tochemical staining was examined and pho- tographed on a Zeiss light microscope.
a-bungarotoxin-binding sites labelling. Samples from embryos, newborns and adult worms were fixed with 2% paraformaldehyde in PBS for 1 hr at 4°C embedded in Tech- novit 7100 resin (Kulzer, Germany) and cut with a glass knife, using Pyramitome (LKB, Sweden). The 2 pm thick sections were dried at room temperature for 2 days and then incubated with 10 nM o-bungarotoxin (a- BuTx) conjugated with fluorescein isothio- cyanate (Sigma Chem. Co. USA) for 1 hr at 4°C as previously described (Falugi and Prestipino, 1989). Controls of binding speci- ficity were carried out by pre-incubating materials with 10nM native mu-BuTx or d- tubocurarine chloride (curare). After wash- ing, tissue sections were mounted with 1:l (v/v) glycerol/PBS and observed on a Zeiss microscope equipped for epifluorescence.
Biogenic amines reactivity. Whole embryos, or embryo and adult segments were exposed to formaldehyde vapours at 80°C for 1 hr, and mounted in liquid paraffin (Falk and Owman, 1965). Fluorescent conden- sation product was observed on a Zeiss microscope, equipped for epifluorescence, using two different filter sets (BP 450-490, FT 510, LP 520 and G 365, FT 395, LP 420), in order to detect green or white-blue fluorescence indicative of catecholamines, and yellow fluorescence indicative of indo- lamines (Pearse, 1972; Van Marle, 1990).
Immunohistochemical reactions
The specimens were fixed with 2% para- formaldehyde in PBS for 1 hr at 4°C then embedded in Paraplast paraffin (Monoject,
FALUGI AND DAVOLI
USA) and sectioned with a glass knife. 6 pm thick sections on gelatinated slides were dried at room temperature for 2 days and then dewaxed as usual. The immunoreaction was carried out according to the indirect immu- nofluorescence method of Coons et al. (1955). Primary antisera from rabbit were applied to the tissue sections for 24 hr at 4°C; the sections were then rinsed in PBS and incubated for 1 hr at room temperature with secondary goat anti-rabbit immunoglobulin, conjugated with fluorescein isothiocyanate. Anti-human ChE (Calbiochem, USA) pri- mary antiserum was diluted 1:50 in PBS con- taining bovine serum albumin, whereas 1: 150 dilutions were used for anti-serotonin (5HT), anti-substance P, anti-somatostatin, anti- vasoactive intestinal peptide (VIP), anti- bombesin and anti-FMRFamide. The secon- dary fluoresceinated antibody was diluted 1:50. All antisera were obtained from Tech- nogenetics, Italy. Controls of specificity included: (1) omission of primary antiserum, (2) substitution of primary antiserum with non-immune rabbit Serum or, for 5HT, with serum heated at 60°C for 1 hr, (3)pre-absorp- tion of each primary antiserum with the appropriate antigen for 1 hr at 37°C.
Results
ChE activity and ChE-like immunoreactivity
By histochemical studies, ChE activity was detected during all developing embryonic stages. In particular, during the first week of development the enzymatic reaction product appeared rather ubiquitous in the whole embryo up ‘to the planula stage (Fig. l), whereas during the various phases of gas- trulation the reaction was detected in the ectoteloblastic bands and in the ectodermal ridge surrounding stomodeum (Figs 2, 3). The dorsal ectodermal layer functioning as transient yolk sac was also stained (Fig. 3). During early differentiation stages, ChE activity was mainly located in cells and tissues engaged in morphogenic movements, includ- ing ectoteloblastic strands elongating over
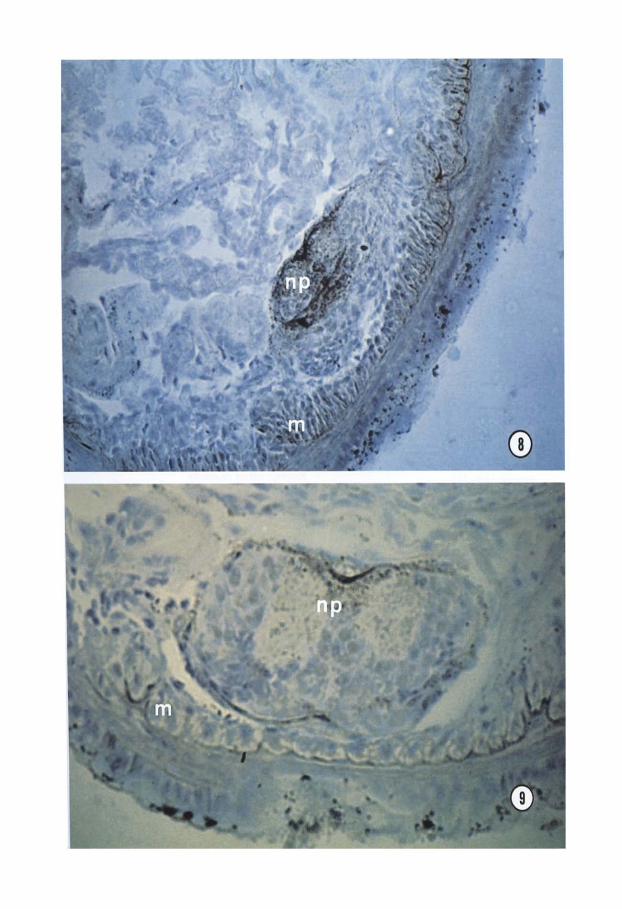
Figs 8, 9. ChE strong positivity in differentiating ventral nerve cord and muscles (Fig. 8). The positivity decreases with further differentiation of the same region (Fig. 9). m, muscle; np, nemopile. Fig. 8 x300; Fig. 9 x600.
316
and sinking under the epithelium to form the CNS (Fig. 4). Successively, positive reaction was found in nerve fibers and bipolar nerve cells around the newly forming CNS and at the border of neuroblasts in contact with them (Fig. 5). Moreover, during devel- opment of epidermal sense buds, the reaction products occurred in nerve fibers running among myoblasts of the body wall (Figs 6,7) and in surface cell clusters (probably nerve cells) (Fig. 7). During the second week, ChE staining was strong in the myoblasts of the ventral body wall and in the neuropile of the differentiating ventral nerve cord (Fig. S), whereas it decreased with further differenti- ation of the same region (Fig. 9). By immu- nohistochemical methods, a similar pattern of ChE distribution was found in all devel- opment stages.
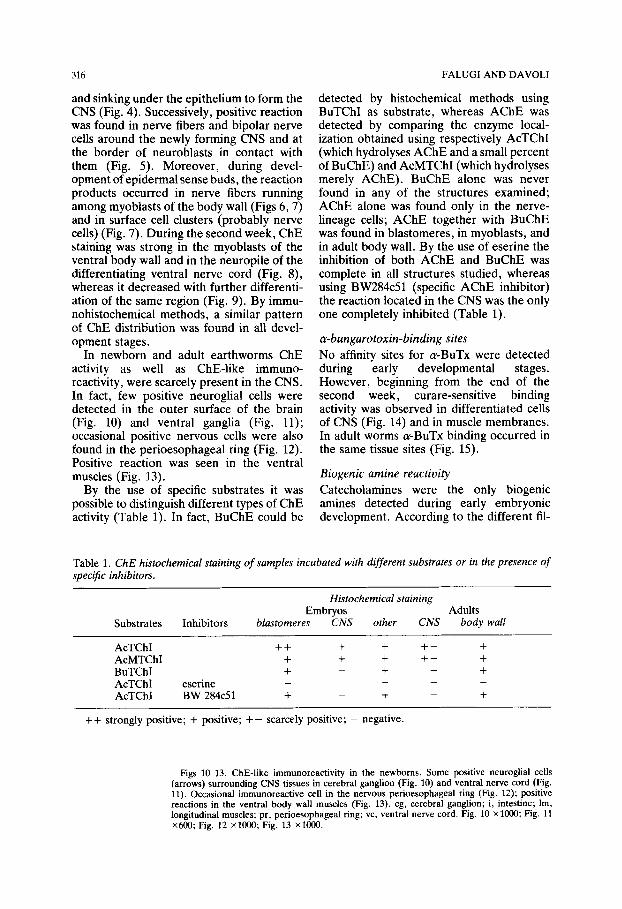
In newborn and adult earthworms ChE activity as well as ChE-like immuno- reactivity, were scarcely present in the CNS. In fact, few positive neuroglial cells were detected in the outer surface of the brain (Fig. 10) and ventral ganglia (Fig. 11); occasional positive nervous cells were also found in the perioesophageal ring (Fig. 12). Positive reaction was seen in the ventral muscles (Fig. 13).
By the use of specific substrates it was possible to distinguish different types of ChE activity (Table 1). In fact, BuChE could be
FALUGI AND DAVOLI
detected by histochemical methods using BuTChI as substrate, whereas AChE was detected by comparing the enzyme local- ization obtained using respectively AcTChI (which hydrolyses AChE and a small percent of BuChE) and AcMTChI (which hydrolyses merely AChE). BuChE alone was never found in any of the structures examined; AChE alone was found only in the nerve- lineage cells; AChE together with BuChE was found in blastomeres, in myoblasts, and in adult body wall. By the use of eserine the inhibition of both AChE and BuChE was complete in all structures studied, whereas using BW284c51 (specific AChE inhibitor) the reaction located in the CNS was the only one completely inhibited (Table 1).
cu-bungarotoxin-binding sites
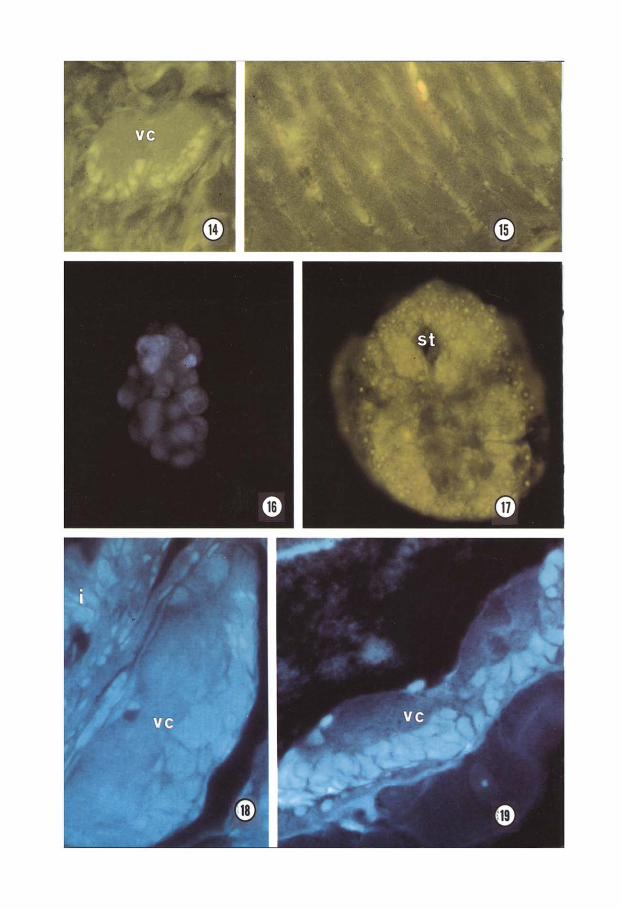
No affinity sites for acBuTx were detected during early developmental stages. However, beginning from the end of the second week, curare-sensitive binding activity was observed in differentiated cells of CNS (Fig. 14) and in muscle membranes. In adult worms LX-BuTx binding occurred in the same tissue sites (Fig. 15).
Biogenic amine reactivity
Catecholamines were the only biogenic amines detected during early embryonic development. According to the different fil-
Table 1. ChE histochemical staining of samples incubated with different substrates or in the presence of specific inhibitors.
Histochemical staining Embryos Adults
Substrates Inhibitors blastomeres CNS other CNS body wait
AcTChI ++ + + +- + AcMTChI + + + +- + BuTChI + _ + _ + AcTChI eserine _ _ _ _ -
AcTChI BW 284~51 + _ + _ +
++ strongly positive; + positive; +- scarcely positive; - negative.
Figs l&13. ChE-like immunoreactivity in the newborns. Some positive neuroglial cells (arrows) surrounding CNS tissues in cerebral ganglion (Fig. 10) and ventral nerve cord (Fig. 11). Occasional immunoreactive cell in the nervous pedoesophageal ring (Fig. 12); positive reactions in the ventral body wall muscles (Fig. 13). cg, cerebral ganglion; i, intestine; lm, longitudinal muscles; pr, perioesophageal ring; vc, ventral nerve cord. Fig. 10 x 1000; Fig. 11 X600; Fig. 12 XlooO; Fig. 13 xlO@l.

. ’ ” .
,.- ‘

MESSENGERS DURING EISENlA DEVELOPMENT
ter sets used, green or white-blue fluor- escence was seen in some ectoblastic cells of planula (Fig. 16), in the ectodermal ridge surrounding stomodeum, and in the ecto- teloblastic strands engaged in morphogenetic and neurogenic movements of gastrula (Fig. 17). During the second week of develop- ment, fluorescence indicating cate- cholamines was observed in the CNS cells, in both the cephalic region (Fig. 18) and the less differentiated caudal region (Fig. 19), and in the gut epithelium (Fig. 18).
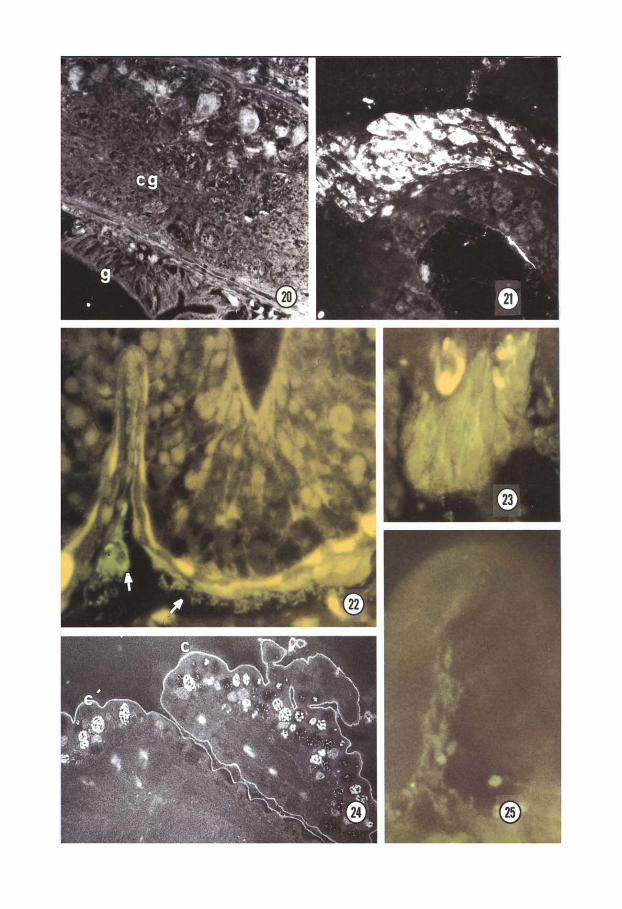
In newborn and adult earthworms, flu- orescent stainings for both catecholamines and indolamines were found in the CNS and in the gut. Accordingly, by the immuno- histochemical method SHT-like reactivity was not detected in embryos, while it was found in large cells located in the CNS and in the gut wall of newborns and adults (Fig. 20).
Peptidergic immunoreactivity
Using different anti-peptides sera, no specific staining was detected during early devel- opment whereas it was generally observed after organogenesis. In fact, at the end of the second week, somatostatin-like immuno- reactivity appeared in the CNS cells (Fig. 21) and in the myoblasts around differentiating gut wall. During the third week other immu- nologically-related peptides began to appear. In fact, FMRFamide-like positivity was observed in peripheral neurons and small nerve bundles surrounding the gut (Fig. 22) as well as in the differentiating cells of the large fold of intestinal wall, so called typhlo- sole (Fig. 23). Substance P-like and VIP-like immunoreactivity was observed in the skin cells (Fig. 24) and in prostomium nerve cells (Fig. 25) respectively. No Bombesin-like pos-
319
itivity was found during the whole period of development and growth.
The specifity of all immunoreactions was confirmed by controls.
Discussion
The results of our investigation show that cholinesterases, both AChE and BuChE, are present in early embryonic stages of Eisenia foetida before any neuromuscular differenti- ation is evident. These findings are similar to those observed during early embryonic development of several invertebrates and vertebrates (Drews, 1975; Fluck, 1978; Mee- de1 and Whittaker, 1979 ; Falugi and Raineri, 198.5). In particular, AChE has been detected in the uncleaved eggs of the sea urchin (Ozaki, 1974), as early as the g-cell stage in the ascidian (Meedel and Whittaker, 1979). and also during the early phase of embryogenesis of the leech (Fitzpatrick- McElligott and Stent, 1981). It has been sug- gested that the appearance of AChE before the differentiation of nerve and muscle tis- sues represents an embryonic feature which plays a role in morphogenetic movements (Drews, 1975) and, in general, in the regu- lation of pre-nervous intercellular com- munications (Buznikov, 1990; Minganti et al., 1981). Moreover, recent studies (Layer, 1991) support the suggestion that ChE mol- ecules may be involved in the regulation of cell proliferation and neurite growth occur- ring in the early morphogenetic period of development (Filogamo and Marchisio, 1971). It is noteworthy that in Eisenia foetida both AChE and BuChE appear widely dis- tributed in early stages of development. whereas they are located in particular areas of differentiating nerve and/or muscle
Figs 14, 15. Fluorescein conjugated cu-BuTx-binding sites in the ventral nerve cord of a 13- day embryo (Fig. 14), and in the muscle membranes of an adult (Fig. 15). vc. ventral nerve cord. Fig. 14 x600; Fig. 15 x600.
Figs 16, 17. Catecholamines fluorescence in early embryos in foto: planula stage observed with G 365, FI 395, LP 420 filter set (Fig. 16), and gastrula stage observed with BP 450.490. FT 510. LP 520 filter set (Fig. 17). st, stomodeum. Fig. 16 x400; Fig. 17 x600.
Fies 18. 19. Catecholamines fluorescence (G 365. FT 395. LP 420 filter set) in transversal Y
sections of a lo-day embryo. Positivity in difierentiated “en&al nerve cord and gut cells (Fig. 18), and in ventral nerve cord of less differentiated caudal region (Fig. 19). i. intestine: vc. ventral nerve cord. Fig. 18 x600; Fig. 19 x600.

MESSENGERS DURING EISENIA DEVELOPMENT
tissues, and are restricted to ventral muscular fibers and to some CNS cells in newborns and adults. According to Layer (1991), the main basis for postulating morphogenetic functions of AChE stems from findings that the expression of AChE can change or dis- appear totally during the course of devel- opment up to the adult state. In this light, since also BuChE activity seems to be involved in the regulation of developmental events in embryonic systems (Layer, 1991; Falugi and Raineri, 198.5), the transient pres- ence of ChE molecules in the early embryos of Eisenia foetida may be regarded as an embryonic feature associated with mor- phogenetic processes of neurogenesis and myogenesis.
acBuTx-binding activity, inhibited by curare, is indicative for the presence of ACh receptors or of their subunit/precursor pro- teins since early differentiation. Its local- ization in the cytoplasm of embryonic and adult nervous cells of Eisenia foerida could be related to the presence of immature forms of receptor molecules or may indicate a rapid receptor turnover. The existence of such receptors on the membranes of muscles, where ChE activity is also expressed, strongly suggests a neuromuscular transmitter role for ACh, even if the actual physiological role of ChE molecules during Eisenia foetida devel- opment and growth remains to be demon- strated. In any case, since ChE histochemical reaction products show that in this animal, as in arthropods (Raineri and Falugi, 1983), during neurogenesis and myogenesis the ner- vous and muscular cells differentiate in a synchronous coordinated pattern, AChE activity may be considered as an early dif-
321
ferentiation marker of these cell types. A similar relationship has been found in chick embryo, in which AChE appears simul- taneously in the early neurons of neural tube and in the myoblasts of myotomes at cor- responding levels. This finding has also been interpreted as the first sign of differentiation (Miki and Mizoguti, 1982) and as a conse- quence, ChE activity has been exploited as a sensitive and early histochemical dif- ferentiation marker in these animal species (Layer, 1991).
In Eisenia foetida, aldehyde-induced his- tofluorescence staining, known to reveal the presence of dopamine, noradrenaline and 5HT (Vaughan, 1988), can be also con- sidered as an early histochemical marker of differentiating nervous cells. In fact, in the embryo, the reaction product appears associ- ated to nerve lineage cells and seems to be related to the presence of catecholamines only, since serotonin-like immunoreactivity was never detected during early develop- ment. It must be pointed out that many his- tochemical, biochemical and physiological findings obtained in adult earthworms sug- gest a neurotransmitter role for primary cat- echolamines in these animals (Gardner and Walker, 1982).
Concerning the immunologically-related neuropeptides, our immunohistochemical results give additional information on the presence of these substances in Eisenia foet- ida. In fact, many neuropeptides have been observed in the adults (Lkhider et al., 1987; Fuji et al., 1989; Al-Yousuf, 1990) while we demonstrated them, also, in the embryos as early as the beginning of the third week. Since embryonic development lasts about 24
Fig. 20. 5HT-like positive cells in the cerebral ganglion and wthin the gut wall of a newborn. cg. cerebral ganglion; g, gut. x600.
Fig. 21. Somatostatin-like immunoreactivity in the cerebral ganglion of a 13-day embryo. X600.
Figs 22, 23. FMRFamide-like immunofluorescence in nerve cell bodies and fibers (arrows) around the gut wall (Fig. 22) and in the gut cells of differentiating typhlosole (Fig. 23). Fig. 22 X 1000; Fig. 23 x 1OOG.
Fig. 24. Substance P-like immunoreactivity in skin secretory cells of a 3-week embryo (the cullcle fluorescence is not specific). c, cuticle. X400.
Fig. 25. VIP-like immunoreactivity in small peripheral neurons in the prostomium of ;1 3. week embryo. x300.

322
days, the immunologically-related peptides appear in rather advanced stages. A similar behaviour has been observed in the embryogenesis of the insect Locusta migratoria, in which somatostatin-like neuro- peptides have been detected 4 days before hatching (at 213 of the embryonic life), and FMRFamide-like substances 1 day later (Remy and Vieillemaringe, 1988). Our data indicate also that somatostatin-like peptides which have been observed in many invert- ebrates, such as planarians, mollusca, cru- stacea, insecta, protochordata (Bautz et al., 1990) and, in particular, in leeches and polychaetes (Al-Youssuf, 1990), are also present in oligochaetes, thus reinforcing the idea of a conservation of these peptidic sequences during phylogenesis.
FALUGI AND DAVOLI
In conclusion, this study provides indirect evidence for a role of ChE molecules during development in regulating early intercellular communications, neurogenesis, and myo- genesis of Eisenia foetida, and gives further support to the hypothesis that some con- servative sequences of messenger neuro- peptides arose very early in evolution.
Acknowledgements
This work was partially supported by grants of the Italian University Research Ministry (MURST grants 60% and 40%).
We extend our appreciation to Professor P. Omodeo for his useful suggestions and to Dr L. Accinni for the critical revision of the manuscript.
References
Al-Yousuf, S. 1990. Neuropeptides in Annelids. In Progress in Comparatiue Endocrinology (eds. A. Epple, C. G.
Scanes, M. H. Stetson), pp. 232-241. Wiley Liss, New York.
Bautz, A., Richoux, J. P. and Schilt, J. 1990. Demonstration by immunocytochemical staining of a somatostatin-2%
(I-14)-like peptide in planarians (Plathyelminthes Turbellaria Tricladida). Gen. Comp. Endocrinol., 78, 469-473.
Buznikov, G. A. 1990. Neurotransmitter in Embryogenesis. Harwood Acad. Publ., Chur.
Coons, A. H., Leduc, E. H. and Connelly, J. M. 1955. Studies on antibody production. I. A method for histochemical
demonstration of specific antibody and its application to a study of the hyperimmune rabbit. J. Exp. Med., 102,
49-59.
Davoli, C., Marcheggiano, A., Serafino, A. and Falugi, C. 1989. Iodine-binding activity in newborn Eisenia foetida. Xith International Symp. Camp. Endocrinol., p. 93.
Davoli, C., Marcheggiano, A., Ravagnan, G., Minti, M., Serafino, A. and Iannoni. C. 1991. Iodination activity in
Eisenia foetida (Annelida Oligochaeta). Cell Tiwue Res. 1 264, 9-14.
De Loof, A. and Schoofs, L. 1990. Homologies between the amino acid sequences of some vertebrate peptide
hormones and peptides isolated from invertebrate sources. Camp. Biochem. Physiol., 95/b, 45%468.
Drew, U. 1975, Cholinesterase in embryonic development. Prog. H&to&em. Cytochem., 7, l-52.
Falck, B. and Owman, C. 1965. A detailed methodological description of the fluorescence method for the cellular
demonstration of biogenic amines. Acta Uniu. Lund. Sect. II, 7, l-23. Falugi, C., DavoIi,C., Iannoni, C. and Marcheggiano, A. 1987. Histochemistry of the nervous system of Eiseniafoetida
(Annelida). Bas. Appl. Histochem., 31, 71. Falugi, C., Marcheggiano, A., Iannoni, C. and Davoli. C. 1988. Localization of cholinesterase activity during the
development of ELenia foetida. J. Histochem. Cytochem.. 36, 949. Falugi, C. and Prestipino, G. 1989. Localization of putative nicotinic cholinoreceptors in the early development of
Paracentrotur lioidus. CeiI Molec. Biol., 3.5, 147-161.
Falugi, C. and Raineri, M. 1985. Acetylcholinesterase (AChE) and pseudocholinesterase (BuChE) activity distribution
pattern in early developing chick limbs. J. Embryol. Exp. Morph., 86, 89-108. Filogamo, G. and Marchisio, P. C. 1971. Acetylcholine system and neural development. Neurosci. Res., 4, 29-64. Fitzpatrick-McElligot, S. and Stent, G. S. 1981. Appearance and localization of acetylcholinesterase in embryo of the
leech Helobdelia triserialk. J. Neurosci.. 1, 901-907.
Flack, R. A. 1978. Acetylcholine and acetylcholinesterase activity in early embryos of the medaka, Oryrias latipes, a
teleost. Deu. Growth Differ., 20, 17-26.
Fuji, K., Ohta, M., Sasaki, T., Sekizawa, Y., Yamada, C. and Kobayashi. H. 1989. FMRFamide immunoreactivity
in the nervous system of the earthworm Eisenia foetida. Zool. Sci.. 4, 951-961.
Gardner, C. R., and Walker, R. _I. 1982. The role of putative neurotransmitters and neuromodulators in annelids and
related invertebrates. Prog. Neurobiol., 18, 81-120. Josse, J. 1986. Neuropeptides: Peripheral and central messengers of the brain. In Comparative Endocrinology:
Deoelopments and Directions (ed. C. L. Ralph), pp. 13-32. Alan R. Liss, New York.

MESSENGERS DURING ELSE/VIA DEVELOPMENT :11 -.
Karnovsky, M. J. and Roots, L. 1964. A ‘direct colouring’ thiocholinc method for cholincstcrasc. J. Hisro&ern
Cytochem., 12, 21e2.21.
Layer, P. G. 1991. Cholinesterase during development of the avian nervous system. Cel[ M&c. N~urobiul. Il. 7-33
Lkhider. M., Marcel, R. and Tramu, G. 1987. Etablissement d’unc carte des newones do cerveau d’Eisenicr f&v&,
(AnnClide, Oligoch&e) contenant des substances immunologiqucment apparent& B des peptidea dc vcrtChr&
Gen. Camp. Endocrinol., 65, 457-468.
Lunt. G. G. and Olsen. R. W. 1988. Comparative Invertebrate Neurochemiq~. Groom Ifelm. London and Sydne!
Marcheggiano. A.. lannooi, C. and Davoli, C. 1985. Thyroglobulin-like lmmunorcactivity in the nervous cystem of
Eisenia foetida (Annelida. Oligochaeta). Cell Tissue Res.. 241, 42%433.
Mecdel. T. H. and Whittakcr, J. R. 1979. Development of acetylcholinestcravz during embryogenrna of the ascidian
Ciona intestinalis. J. Exp. Zool., 210, l-10.
Miki. A. and Mizoguti. H. 1982. Acetylcholinesterase activity m the myotome of the early chick embryo. Cell 71\\w
Ku., 221, 23-40.
Minganti, A., Falugi. C., Raineri, M. and Pestarino. M. 1981. Acethylcholinesterase in the embryonic dcvelopmcnt.
an Invitation to a hypothesis. Acfa Embryo/. Morph. Exp. n.s.. 2, 30.
Ozakl, H. 1974. Localization and multiple forms of acetylcholinesterase in sea urchin embryos. I)ecl Grotuh /Itffrr 16. 267-279.
Pearsc. A. G. E. 1972. Histochemistry. Theorefical and Applied. Churchill Livmgstone, Edinburgh and London.
Rameri. M. and Falugi, C. 1983. Acetylcholinesterase activity in embryonic and larval development of Artemia salina
Leach (Crustacea Phyllopoda). J. Exp. Zool., 227, 229-246.
Remy. C. and Vieillemaringc. J. 1988. Immunocytology of insect peptides and amines. In Neurohormorr~~.s in
Inoerrebrures (eds. M. C. Thorndyke and G. J. Goldsworthy), pp. 43-77 University Press, Cambridge.
Roth. J.. LcRoith. D.. Shiloach, J., Rosenzweig, J. L.. Lesniak, M. A. and Havrankova, J. 1982 The cvolutmnaty
origms of hormones. neurotransmitter. and other cxtraccllular chemical messengers New Engi. J. ,Mrd 306.
52F527.
Schumacher, M., Camp, S., Maulet, Y., Newton. M.. MacPhee-Quigley. K.. Taylor. S. S.. Friedman. T. and Taylor.
P. 19X6. Primary structure of Torpedo colifornicn acetylcholinesterase deduced from its cDNA rcqucncc\ .Yrrrwr.
319, 407-409.
Swillcn?. S.. Ludgatc, M.. Mercken, L.. Dumont. J. E. and Vassart, G. 1986. Analysla of sequence and structure
homologies between thyroglobulin and acetylcholinesterasc: possible functional and clinical significance. Rwclwn
Biophys. Res. Commun., 137, 142-148.
Thorndykc, M. C. 1989. Peptides in invertebrates. In The Compararioe Phvsiology of Keplarorv Peprrde.7 (cd s Holmgrcn), pp. 203-228. Chapman and Hall, London and New York.
Van Marie. J. 1990. Catecholamines, related compounds and nervous system in the tcntaclcs of some Anthozoan\ In
Euoiution of the Firsf Neroous Sysrems (ed. P. A. V. Anderson). pp. 12%140. Plenum Pros. New York and
London.
Vaughan. P. F. T. 1988. Amine transmitters and their associated second messenger systems. In Comparunoe fnoerrebrorc Neurochemistry (eds. G. G. Lunt and R. W. Olsen), pp. 124-174. Groom Helm, London and Sydney




















