
Ventromedial Temporal Lobe Pathology in Dementia, Brain Trauma, and Schizophrenia
20
575 Ventromedial Temporal Lobe Pathology in Dementia, Brain Trauma, and Schizophrenia GARY W. VAN HOESEN, a–c JEAN C. AUGUSTINACK, a AND SARAH J. REDMAN a a Department of Anatomy and Cell Biology and the b Division of Cognitive Neuroscience, Department of Neurology, University of Iowa, Iowa City, Iowa 52242, USA ABSTRACT: The ventromedial temporal area contains numerous anatomical structures collectively or selectively involved in a wide range of neurological and psychiatric disorders. Collective involvement is exemplified best by Alz- heimer’s disease where a host of anatomical structures and a host of cognitive and behavioral changes are manifested. Selective disease of the amygdala can yield deficits in the ability to judge and evaluate emotional expressions. While memory functions are nearly synonymous with the concept of ventromedial temporal area, they overshadow other functions associated with the diverse an- atomical structures in this part of the brain. For example, it could be argued that in addition to output directed toward the hippocampal formation, the out- put of the ventromedial temporal area is equally strong to the ventral striato- pallidal system of the basal forebrain. Denervation of these structures could be associated with the behavioral changes that occur in tandem with the memo- ry-related changes of ventromedial temporal lobe pathology. Here we explore the anatomical and pathological correlates associated with ventromedial tem- poral area pathology and consider how these may impact on ventral striatopal- lidal conceptualizations. We conclude that ventromedial temporal area pathology deprives the basal forebrain of multimodal association information from the endstages of corticocortical sensory processing. This endstage infor- mation carries with it an analysis of real-time sensory awareness, historical- time or past sensory experiences, and decisions from hippocampal output structures regarding relevancy and novelty. In this sense, basal forebrain structures are in a unique position to regulate behavioral responses to a wide range of stimuli and to organize appropriate emotional, motor, autonomic, and endocrine responses to them. INTRODUCTION The ventromedial temporal area is a functionally diverse and anatomically com- plex part of the cerebral hemisphere inserted along the ventral and medial parts of the temporal fossa. 29,83,85 It borders the sphenoid bone and the petrous part of the temporal bone. Portions of the ventromedial temporal area are traversed by the free edge of the tentorium cerebelli prior to its attachment to the petrous apex and the an- terior and posterior clinoid processes. 24,51,63 The paired free edges of the tentorium cerebelli form an aperture through which the upper brain stem passes and the ante- c Author for correspondence: Department of Anatomy and Cell Biology, University of Iowa, Iowa City, IA 52242, USA. Voice: 319-335-7741; fax: 319-335-7198; gary-van- [email protected]
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Ventromedial Temporal Lobe Pathology in Dementia, Brain Trauma, and Schizophrenia

575
Ventromedial Temporal Lobe Pathology in Dementia, Brain Trauma, and Schizophrenia
GARY W. VAN HOESEN,a–c JEAN C. AUGUSTINACK,a AND SARAH J. REDMANa
aDepartment of Anatomy and Cell Biology and the bDivision of Cognitive Neuroscience, Department of Neurology, University of Iowa, Iowa City, Iowa 52242, USA
ABSTRACT: The ventromedial temporal area contains numerous anatomicalstructures collectively or selectively involved in a wide range of neurologicaland psychiatric disorders. Collective involvement is exemplified best by Alz-heimer’s disease where a host of anatomical structures and a host of cognitiveand behavioral changes are manifested. Selective disease of the amygdala canyield deficits in the ability to judge and evaluate emotional expressions. Whilememory functions are nearly synonymous with the concept of ventromedialtemporal area, they overshadow other functions associated with the diverse an-atomical structures in this part of the brain. For example, it could be arguedthat in addition to output directed toward the hippocampal formation, the out-put of the ventromedial temporal area is equally strong to the ventral striato-pallidal system of the basal forebrain. Denervation of these structures could beassociated with the behavioral changes that occur in tandem with the memo-ry-related changes of ventromedial temporal lobe pathology. Here we explorethe anatomical and pathological correlates associated with ventromedial tem-poral area pathology and consider how these may impact on ventral striatopal-lidal conceptualizations. We conclude that ventromedial temporal areapathology deprives the basal forebrain of multimodal association informationfrom the endstages of corticocortical sensory processing. This endstage infor-mation carries with it an analysis of real-time sensory awareness, historical-time or past sensory experiences, and decisions from hippocampal outputstructures regarding relevancy and novelty. In this sense, basal forebrainstructures are in a unique position to regulate behavioral responses to a widerange of stimuli and to organize appropriate emotional, motor, autonomic, andendocrine responses to them.
INTRODUCTION
The ventromedial temporal area is a functionally diverse and anatomically com-plex part of the cerebral hemisphere inserted along the ventral and medial parts ofthe temporal fossa.29,83,85 It borders the sphenoid bone and the petrous part of thetemporal bone. Portions of the ventromedial temporal area are traversed by the freeedge of the tentorium cerebelli prior to its attachment to the petrous apex and the an-terior and posterior clinoid processes.24,51,63 The paired free edges of the tentoriumcerebelli form an aperture through which the upper brain stem passes and the ante-
cAuthor for correspondence: Department of Anatomy and Cell Biology, University of Iowa,Iowa City, IA 52242, USA. Voice: 319-335-7741; fax: 319-335-7198; [email protected]

576 ANNALS NEW YORK ACADEMY OF SCIENCES
rior and medial parts of the ventromedial temporal area protrude into and over theaperture unprotected by dura mater.
The component structures of the ventromedial temporal area include the corticaland subcortical parts of the amygdala, the enfolded allocortical areas rolled into theinferior horn of the lateral ventricle known as the hippocampal formation and thesurface allocortical and periallocortical areas that form the pyriform lobe of the para-hippocampal gyrus. Anteriorly, the latter is formed largely by the primary olfactoryallocortex, whereas posteriorly, the pyriform lobe is formed by the sizable entorhinalperiallocortex, or Brodmann’s area 28. The latter extends posteriorly in all specieswhere it approximates occipital areas. The lateral border of the ventromedial tempo-ral area is the rhinal sulcus in all mammals. It contains the perirhinal cortex, or Brod-mann’s area 35, in its fundus and lateral bank.29,47,71,84 However, in the humanbrain, the rhinal sulcus is normally short, if present at all, and the perirhinal cortexcourses into the collateral sulcus and forms a part of its medial bank. It retains, how-ever, its exact anatomical position sandwiched between the olfactory and entorhinalcortices and the more laterally located temporal isocortices whether the rhinal sulcusis present to any degree or not.83,85
In functional terms, the ventromedial temporal area has been linked historicallyto three major realms of behavior, namely, olfaction, emotion and memory. Olfacto-ry sensation has never been disputed, although great debate over the extent of olfac-tory bulb projections, and hence, the olfactory brain, occupied many decades.Clearly, several brain areas receive direct olfactory bulb projections and these alongwith secondary olfactory projections reach many parts of the ventromedial temporalarea.9,22,27,28 A role in emotion for ventromedial temporal areas was championed byPapez and the “emotive process” was assembled or “built up”, according to his think-ing, in its hippocampal and/or hippocampal-related structures. Although historicalinterest is still given to Papez’s bold proposal,67 and its anatomical underpinnings, itnever received definitive support in the temporal lobe. While it is clear that theamygdala of the ventromedial temporal area plays a role in emotional perception andmemory, and governs some of the key autonomic and endocrine responses pairedwith emotion,9 core elements of affect and feelings are governed by larger neuralsystems in which the amygdala is only one player.25 Ironically, the amygdala wasnot a part of Papez’s circuit.
A definitive role for the ventromedial temporal area in memory is a cornerstoneof neuroscience teaching and the first thought that occurs when this geographic areaof the cerebral hemisphere is mentioned.75 Moreover, the case has been steadilystrengthened and elaborated as time has passed.7,66,80,96 While much of the neuroan-atomical organization of ventromedial temporal area seems geared to subservingmemory-related mechanisms,84 it is easy to lose sight of the fact that many large hip-pocampal output neural systems arise here that contribute to other forms of behavior.These systems may not play a definitive role in memory-related behavior per se, butinstead contribute critical memory-related information to other behaviors.86 One ofthe major non-hippocampal targets of output from the ventromedial temporal areasis the basal forebrain, and particularly, the ventral striatopallidal and magnocellularcell groups that form these areas.3,5,8,39,42,52,56,87,95 These areas have been linkedmore strongly to emotional mechanisms than was ever the case for ventromedialtemporal areas, and it is likely, that it is via these systems, an integrated emotional

577VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
behavior is achieved.41,42 To explore this concept, we will examine several types oftemporal lobe pathology that affects neural systems that contribute powerful input tobasal forebrain structures. The general theme to be considered is that behavioralchanges in these disorders, including schizophrenia, may be understood, at least, inpart, by considering the fact that ventromedial temporal area pathology deafferentsbasal forebrain areas depriving them of real-time, historical-time and memory-relat-ed awareness critical for emotional, motor and homeostatic mechanisms.
ALZHEIMER’S DISEASE
Hippocampal Formation
Hippocampal pathological changes in Alzheimer’s disease (AD) have been ob-served for nearly a century, and documenting them is recommended in all neuro-pathological diagnostic protocols.11,19–21,44–46 In terms of neurofibrillary tangles(NFTs), this signature feature of AD is observed primarily in the subicular and CA 1pyramidal neurons, with the CA 3 pyramids negative, or largely spared. In general,there is typically more subicular/CA 1 pathology in patients who have been ill for along time and less in those with illnesses of shorter duration. NFTs among the varioushilar neurons that form the CA 4 zone of the dentate gyrus are not unusual, but sel-dom total more than 8–10 affected neurons per cross-section, with many cases havingnone. Curiously, CA 1 and subicular NFT pathology fails to have a differential ante-rior-posterior distribution.46 These neurons often have classical NFTs with dense fil-amentous paired helices extending for long distances in the apical and basal dendritesand more delicate filaments surrounding the nucleus in the region of the soma. Many,in long duration cases, form the so-called “extraneuronal or ghost” NFTs, meaningsimply that they are marking the location of a once viable neuron which at death con-tained no stainable cytoplasm. In shorter duration cases, it is not unusual to see sub-icular and CA 1 pyramids with conspicuous tangles in their apical dendrites, butretaining stainable cytoplasm and a still concentric well-defined nucleus and nucle-olus.45 One has to assume that these neurons are altered greatly from a functionalviewpoint, but the apparent integrity of the soma, and presumably, intact axon hillockmight provide an altered abnormal output to areas where they send axons. NFTs areobserved occasionally in the granule cell neurons that form the dentate gyrus, but thisis the exception rather than the rule, and typically, these cases have a lengthy durationof illness.
Neuritic plaques (NPs) are also observed in the hippocampal formation in AD.Unlike the isocortex, where their distribution is often more random across severalcortical laminae, their distribution in the hippocampal formation is more conserva-tive in AD. For example, NPs are often observed in the molecular layer of the dentategyrus where they form a band between the granule cell somas and the hippocampalfissure. In the subiculum and CA 1 zones they are often seen among the somas thatform these fields, but also in the stratum radiatum and stratum laconosum-molecu-lare areas of their apical dendrites. Both in the dentate gyrus, and for parts of the CA1/subicular zones, NPs seem to show a preference for the terminal fields of the per-forant pathway.45,46
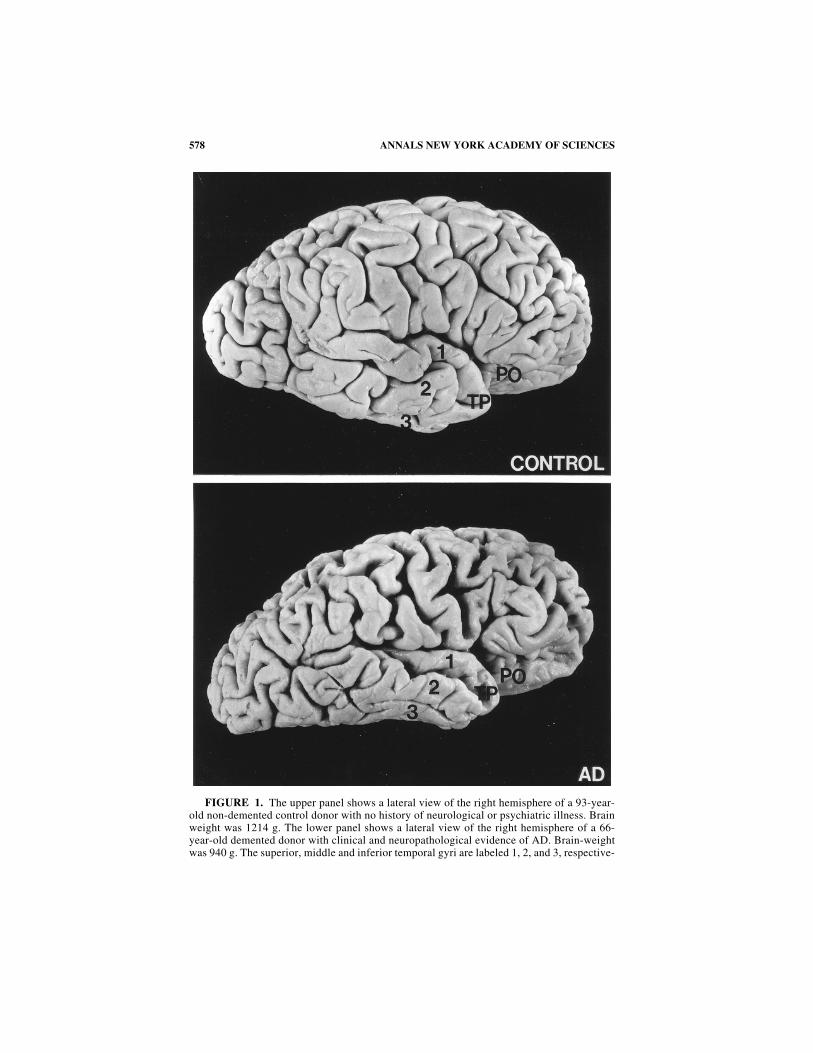
578 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 1. The upper panel shows a lateral view of the right hemisphere of a 93-year-old non-demented control donor with no history of neurological or psychiatric illness. Brainweight was 1214 g. The lower panel shows a lateral view of the right hemisphere of a 66-year-old demented donor with clinical and neuropathological evidence of AD. Brain-weightwas 940 g. The superior, middle and inferior temporal gyri are labeled 1, 2, and 3, respective-

579VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
In summary, hippocampal formation pathology in AD is extensive, affecting py-ramidal projection neurons that in other mammals send axons to widespread parts ofthe telencephalon and diencephalon.70 This would include hippocampal projectionsto the entorhinal, posterior parahippocampal, perirhinal and temporal polar parts ofthe temporal lobe, and to the orbital and medial frontal parts of the frontal lobe (FIG.1).39,70,93,94 Additional input to the anterior and posterior cingulate cortices wouldbe expected to be compromised also. Other non-cortical telencephalic projections al-tered in AD would be CA 1/subicular input to the amygdala, septum, vertical limbof the diagonal band nucleus and nucleus accumbens of the ventral striatum.70,71
Subcortical diencephalic projections to the anterior thalamus and hypothalamuswould no doubt degenerate as well in a severely affected AD case. On the basis ofanimal research, some of the neurons affected by NFTs would be expected to sendaxons directly to cortical areas that in turn send axons to the ventral striatum.40,62,68
The net affect in AD would be a profound disconnection between this part of the ven-tromedial temporal area and the ventral striatum.
Entorhinal Cortex
There is little doubt that the entorhinal (Brodmann’s area 28) and perirhinal(Brodmann’s area 35) cortices are the initial focal points for the formation of NFTsin AD and the entorhinal cortex is the most heavily ravaged by the illness at all pointsin its course (FIG. 2).21,45 Indeed, the normally conspicuous layer II islands of neu-rons can disappear entirely in cell stains in endstage AD;36 however, pathologicalstains, such as thioflavin-S and Bielschowsky, reveal their continued presence, butin the form of extraneuronal NFTs without stainable rough ER and ribosomes. Thesuperficial parts of layer III in entorhinal cortex also contain NFTs as does layer IVdeep to the lamina dissecans. However, the latter is typically a later occurrence inAD after a duration of illness that exceeds three years (FIG. 3). It is not unusual inearly onset familial AD and in sporadic AD with a long duration of illness to seenearly all neurons in all layers of the entorhinal cortex affected by NFTs.
In addition to differential laminar changes, a clear lateral to medial gradient ofNFT formation also occurs, with the first islands affected lying near the collateralsulcus and the last affected lying more medially nearer the periamygdaloid cortex orparasubiculum depending on the anterior-posterior level examined.
In summary, entorhinal cortex pathology in AD greatly compromises the cells oforigin for both the entorhino-dentate and entorhino-hippocampal components of theperforant pathway.88,90,93,94 In addition, neurofibrillary tangles selectively destroymany of the large modified pyramidal neurons that form layer IV. These neurons re-ceive a strong hippocampal output70 and project directly to the ventral stria-tum.38,78,81,93 Moreover, they project to the temporal and frontal cortices54 thatproject strongly to the ventral striatum and the magnocellular parts of the basal fore-brain (FIG. 1).40,62,68,87
ly. In the AD donor, dense NFTs were observed in the subicular/CA 1 zone of the hippocam-pal formation. The pyramids that occupy this zone in non-human primates project to corticalregions such as the temporal polar (TP) and posterior orbital (PO) cortices. Note in the ADbrain that these areas are discolored and atrophic, suggesting that non-direct relayed ventro-medial temporal area input to the ventral striatum and magnocellular cell group of the basalforebrain is also compromised in AD along with direct subicular/CA 1 projections.

580 ANNALS NEW YORK ACADEMY OF SCIENCES
Amygdala
Like the hippocampal formation and entorhinal cortex, the amygdala is damagedheavily, but selectively, in AD and a number of recent investigations have broughtthese changes into sharp focus.30,53,58,65 In so far as NFTs are concerned, the mostheavily damaged amygdaloid nuclei are the accessory basal and cortical nuclei. Thecortical transition area of the posterior amygdala is also damaged heavily, but the lat-eral, laterobasal, mediobasal, central and medial nuclei have a more moderate quan-tity of neurofibrillary changes. Changes in these nuclei can also be variable in apopulation of AD victims, such that it is not unusual to find cases with little or noNFT pathology in the lateral, central and medial amygdaloid nuclei.
A somewhat similar picture emerges when one considers the amygdaloid distri-bution of NPs. For example, these are most dense in the accessory basal nucleus withslightly lesser quantities in the cortical nuclei and the cortical transition area. How-ever, in contrast to the moderate quantity of NFTs, NPs are typically dense in the me-diobasal nucleus equaling or exceeding those of the accessory basal nucleus. Thelateral, laterobasal, central and medial nuclei frequently contain NPs, but their quan-tity is often modest or lacking significant numbers altogether.58
In summary, it is probably accurate to state that the amygdala as a whole is morevariably damaged by the pathology of AD than the hippocampal formation and en-
FIGURE 2. This photograph shows a ventromedial temporal view of the right hemi-sphere in an endstage AD donor. Note the highly atrophic entorhinal cortex (EC), temporalpolar cortex (TP), posterior orbital cortex (PO), and subgenual cortex (SG). All of these ar-eas have been shown in non-human primates to project directly to the ventral striatum andmagnocellular cell groups of the basal forebrain. (Other abbreviations: CC, corpus callo-sum; CS, collateral sulcus; OT, optic tract; RS, rhinal sulcus).

581VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
torhinal cortex parts of the ventromedial temporal area. However, selective nucleisuch as the accessory basal and cortical nuclei are always damaged by dense NFTs,and the former, plus the mediobasal nucleus, contain large quantities of NPs. As ageneral rule, the lateral, central and medial nuclei are consistently the least affected.Caution here though is probably recommended since most AD investigators have fo-cused largely on only NFTs and NPs, and not neuron loss. There is a suggestion inthe literature preceding these more recent studies that neuron loss may be extensivein amygdaloid nuclei that do not contain classic AD pathology.43,91 Irrespective ofthis, it is clear that the basal complex of the amygdala is damaged heavily in AD. Aswith the hippocampal formation and entorhinal cortex, this pathology affects neu-rons that are sources of a powerful afferent input to the ventral striatum and magno-cellular cell groups of the basal forebrain and to frontal and temporal cortices that inturn project to these same basal forebrain targets (FIGS. 4 and 5).3,8,9
TEMPORAL LOBE INJURY
The Tentorial Incisura
Although nearly all components of the ventromedial temporal area are affectedseverely, but selectively, by pathology in AD, and partially so in Pick’s disease, these
FIGURE 3. This is a photomicrograph of a thioflavin-S stained cross-section throughthe entorhinal cortex at endstage AD viewed under fluorescent illumination to reveal neu-rofibrillary tangles. The superficial layer II islands contain dense NFTs as does layer IV nearthe top of the photomicrograph. The latter receives a powerful hippocampal output andprojects directly to the ventral striatum and to temporal and frontal cortices that project tothe basal forebrain.

582 ANNALS NEW YORK ACADEMY OF SCIENCES
conditions form only one class of injury that can affect the temporal lobe. Anotherlarge class of injury relates to the tight encasement and insertion of the ventromedialtemporal area into the temporal fossa adjacent to the irregular boney structure of thispart of the skull. The vulnerability of the temporal lobe from direct forces and forcesgenerated by impact at many points on the skull in head injury is well known.2,37
Likewise, the proximity of ventromedial temporal areas to the inferior horn of thelateral ventricle is of consequence since injury can occur with increased intracranialpressure in the supratentorial space, no matter what its etiology—tumor, abscess, he-matoma, edema, infection or infarction.1,64
Central to both head injury and increased intracranial pressure in the supratento-rial space is the free edge of the tentorium cerebelli which traverses across the ven-
FIGURE 4. This photomicrograph shows a dark field image of terminal axon labeling(white) over the core (c) and shell (s) of the nucleus accumbens (NA) of a rhesus monkeyfollowing an injection of tritiated amino acids into the dorsal portions of the laterobasal andaccessory basal amygdaloid nuclei. (Other abbreviations: AC, anterior commissure; CA,caudate nucleus; GP, globus pallidus; IC, internal capsule; SP, septum; V, anterior horn oflateral ventricle).

583VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
tromedial temporal area of each hemisphere before its attachment to the petrous apexand anterior and posterior clinoid processes (FIG. 6). This arrangement forms an ap-erture through which the upper brain stem passes with a sharp dural incisura sur-rounding the aperture. It is through this aperture that communication between thesupratentorial and infratentorial spaces is achieved.51,63
The anterior part of the parahippocampal gyrus, the so-called gyrus ambiens, liesdirectly in the tentorial aperture unprotected by dura mater.69 The size of the tento-rial aperture varies among humans, but it has been estimated that the free edge of thetentorium cerebelli contracts or grooves the parahippocampal gyrus in 70% of thepopulation, that this groove is visible in neonates, and visible in both the unfixed orfixed brain.24 Retzius69 labeled this groove the inferior rhinal sulcus, but others have
FIGURE 5. This photomicrograph shows a bright field image of dense biotinylated de-tran amine axon labeling in the core (c) of the nucleus accumbens (NA) following an injec-tion into the ventral part of the laterobasal amygdaloid nucleus in the rhesus monkey. (Otherabbreviations: CA, caudate nucleus; DB, vertical limb of diagonal band of Broca; IC, inter-nal capsule; PU, putamen; SP, septum; V, anterior horn of lateral ventricle.)

584 ANNALS NEW YORK ACADEMY OF SCIENCES
labeled it the tentorial notch or groove reflecting, in their estimation, that it repre-sents an indentation and surface marking consistent with the disposition of the freeedge of the tentorium cerebelli.12,29 It is doubtful that it deserves the label of sulcussince it is merely an indentation and bears no relationship to the rhinal sulcus.
Uncal Herniation with Head Injury and Increased Intracranial Pressure
The location of the tentorial incisura and the complications it can create in neu-rological disease and diagnosis has long been appreciated.63 Quite simply, potentialspace in the supratentorial space is not extensive, and when exhausted, supratento-rially located brain structures will herniate across the free edge of the tentorium cer-ebelli into the space of Bichat and infratentorial space. The consequentcompression of the brain stem necessitates emergency efforts to preserve life. Thepathological concept of uncal herniation is an appropriate description for extremeherniation and in autopsy specimens where death occurred. Indeed, in such casesthe uncal hippocampal formation has herniated, and dominates the pathologicalpicture.24 However, in technical terms, it is not the uncal hippocampus that lies inthe tentorial aperture, but instead the entorhinal cortex of the gyrus ambiens of theventromedial temporal area, and it leads the herniation. Thus, partial herniation in-
FIGURE 6. A reproduction of FIGURE 1 from Jefferson’s classic article “The TentorialPressure Cone” showing the course of the free edge of the tentorium cerebelli (TNT) acrossthe parahippocampal gyrus. At the level of the entorhinal cortex (EC) the free edge contactsand notches the cortex. Note that the gyrus ambiens (GA) of the entorhinal cortex bulgesover the free edge of the tentorium cerebelli and sits without dural protection in the tentorialaperture. (Other abbreviations: CS, collateral sulcus).

585VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
jures ventromedial temporal areas that provide input to the ventral striatum and toareas that project to it, and behavior changes in such patients could reflect this sit-
FIGURE 7. A Nissl-stained cross-section of the ventromedial temporal area showinginjury at the tentorial notch (TN) by the free edge of the tentorium cerebelli in a 62-year-olddonor who suffered from agitated depression and psychoses after head injury at age 43. Theasterisks denote layer IV of the entorhinal cortex (EC) which projects to the nucleus accum-bens of the ventral striatum. The unusual, more vertical positioning, of the amygdala (AMG)and hippocampal formation (HP) are no doubt due to the ventromedial temporal area beingforced onto the fixed free edge of the tentorium cerebelli. Gliosis and vascular hypertrophycan still be seen in the amygdala immediately above the tentorial notch. (Other abbrevia-tions: CS, collateral sulcus; PR, perirhinal cortex; RS, rhinal sulcus).

586 ANNALS NEW YORK ACADEMY OF SCIENCES
uation as much as direct entorhinal cortex injury. In this sense, abnormal behaviorfollowing temporal lobe injury and entorhinal herniation could represent a false lo-calizing sign in the same manner as cranial nerve signs do, due to brain stem com-pression, in full uncal herniation.51,63
In a related matter, many patients suffering head trauma recuperate from their in-juries physically, but are left with post-traumatic behavioral changes. In many ofthese cases, it is likely that ventromedial temporal area injury occurred around thefree edge or incisura of the tentorium cerebelli due to the brain being forced onto it(FIG. 7). Thus, post-traumatic behavioral changes in head injury survivors may re-flect distal changes in the basal forebrain equally as much as direct damage to theventromedial temporal area. We suspect that temporal injury around the free edge ofthe tentorium cerebelli, whatever its cause, is more widespread than appreciated anda contributor to poorly understood behavioral changes in humans of all ages. Ana-tomically detailed imaging studies will shed light on such patients.
SCHIZOPHRENIA AND THE VENTROMEDIAL TEMPORAL AREA
The ventromedial temporal area has been of special interest relative to schizo-phrenia research for many years because of clinical observations of psychotic-likebehavior in temporal lobe disease. Some of the disorders discussed above have con-tributed to this interest. For example, in AD, a memory disorder is the hallmark ofthe associated behavioral changes throughout the entire illness, but confusion, delu-sions, aggression, agitation, and hallucinations are not uncommon in many pa-tients.26,34,59–61 These are also common in Pick’s disease and Lewy Body disease,and in fact, may be a more prominent motivator for medical attention than a memoryimpairment. These latter degenerative dementias, however, are less uniquely associ-ated anatomically with the temporal lobe, but nevertheless, involve it to a significantdegree. Pick’s disease, for example, often involves the dentate gyrus and the subic-ular/CA 1 zone of the hippocampal formation (FIG. 8). Other temporal lobe disordersin which psychotic-like behaviors occur include herpes simplex encephalitis86 andtemporal lobe epilepsy.77
The neurobiology of schizophrenia relative to medial temporal structures hasbeen included in recent reviews,13,14,79 and we will not attempt to repeat these veryadequate efforts. Instead, we will comment only on certain issues pertaining to ven-tromedial temporal area cytoarchitecture and neuropathology that have a bearing onthe comments of the text above in the two previous sections.
Postmortem studies in schizophrenia have many difficulties well before theyreach the microscope. Agonal state before death is often problematic as are long his-tories of neuroleptic medication. In many instances, death cannot be anticipated andmedical examiners frequently have agendas and obligations not compatible withshort autolysis times, brain fixation and neuroscience research. Despite this, post-mortem studies have become more common in the past two decades and their resultspoint toward ventromedial temporal area changes, particularly in the hippocampalformation and entorhinal cortex.13,14,79 In the former structure, decreased quantitiesof microtubule-associated proteins have been observed in the subicular/CA 1 partsof the hippocampal formation and entorhinal cortex in schizophrenics.12,72 These

587VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
molecules are expressed strongly during development and are known to play a rolein microtubule cell biology, and particularly, dendritic orientation and elaboration.One might expect on the basis of these results alterations in pyramidal neuron den-dritic morphology, but results on this issue are variable in the hippocampal forma-tion,6,23,55,73,74 but well documented in other limbic cortical areas.15 Smaller neuronsize, however, has been well documented in the subicular/CA 1 zone and entorhinalcortex in schizophrenics, as have volume changes.10,16,31,32
Laminar heterotopy in the entorhinal cortex of schizophrenics has been reportedin several articles12,48–50 and diminished density of neurons in key entorhinal layersis a consistent result.4,32,57 An abnormal spatial arrangement of entorhinal neuronshas also been observed.13 Although the observations of entorhinal laminar heteroto-py have been challenged,4,57 those regarding density and spatial arrangement havenot. Clearly, it is too early to judge entorhinal changes in schizophrenia, but variableresults across patients seems likely. The affirmative or disaffirmative result in thisregard is probably less critical than understanding why some schizophrenics haveentorhinal cellular alterations and others do not. Interactions with abnormalitieselsewhere in the brain would certainly be suspected to contribute to this variability.
Finally, it is of interest that abnormalities around the tentorial notch of the en-torhinal cortex have been reported in some schizophrenics.4,12,48–50,57 Several
FIGURE 8. This photomicrograph is a Bielschowsky-stained cross-section through thesubicular/CA 1 zone of the hippocampal formation in Pick’s disease. Note that nearly everypyramidal neuron contains a large darkly stained Pick body that displaces the nucleus andnucleolus to the periphery of the soma away from their normal central position within thecytoplasm. Abnormal neurites can be seen in the neuropil between neurons as well as rem-nants of organelles and isolated Pick bodies from degenerating neurons. Psychoses and oth-er behavioral disturbances are observed typically in Pick’s disease.

588 ANNALS NEW YORK ACADEMY OF SCIENCES
brains in the Yakovlev Collection have this abnormality (FIG. 9). Those in this col-lection have been questioned because of a prior prefrontal lobotomy.57 However, nu-merous brains in that collection had lobotomies for reasons other than schizophreniaand lacked this abnormality in the ventromedial temporal area. While enthusiasm forabnormalities in the entorhinal cortex caused by the free edge of the tentorium cer-ebelli in some schizophrenics is dampened by similar changes in occasional nor-mals, the key question may be related to timing of injury. If an injury reflected anintra-uterine infection, or was related to birth trauma, it may yield wholly differentlong-term outcomes than those occurring with episodic increased intracranial pres-sure in adulthood. Deep invaginations in the entorhinal cortex around the free edgeof the tentorium cerebelli are not a normal anatomic variation in the human brainsas some authors suggest. Instead, they speak clearly to previous head trauma or ab-normalities in the supratentorial space. Why such changes are over-represented in
FIGURE 9. This is a photograph of a Nissl-stained cross-section through the entorhinalcortex (EC) of a brain from a schizophrenia patient in the Yakovlev Collection at the ArmedForces Institute of Pathology. Note, the deep tentorial notch (TN) and the laminar heterotopyof layer II cell islands at and around the injury. For many reasons, this is regarded as a de-velopmental abnormality. The absence of gliosis is apparent as is the inappropriate migra-tional positioning of layer II neurons. Also, note that layer III pyramidal neurons are sparseat the tentorial notch and for several millimeters medial and lateral to it. Layer IV (arrows)is greatly depleted also at and around the tentorial notch. Both layers III and IV contain pre-dominantly pyramidal neurons whose apical dendrites reach the molecular layer and pial sur-face. Injury or compression of the neuropil would compromise dendritic elaboration and maybe associated with the paucity of neurons seen in these layers. Finally, the normal positioningof the amygdala (AMG) and hippocampal formation (HP) suggest that trauma due to headinjury (see FIG. 8) was probably not a factor in the heterotopy and abnormal tentorial notch.For similar reasons it is unlikely that a previous lobotomy had anything to do with these dra-matic cytoarchitectural changes in the entorhinal cortex of this schizophrenic. The patientwas an adult, well beyond the age of developmental neural migration.

589VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
populations of schizophrenics deserves further study and their presence in occasion-al normals does not diminish this interest.
In summary, ventromedial temporal changes in schizophrenia are commonplaceand the weight of the evidence with all measures employed from molecules12 tobrain and ventricular volumes17,18 adds to the picture. Understanding the variabilitywithin a population of schizophrenics could be key to further isolating the mecha-nisms that cause the disease. As with the other forms of temporal lobe disease high-lighted here, the ventromedial temporal areas that are suspect in schizophrenia areprecisely those that project powerfully to basal forebrain structures or to cortical ar-eas that in turn send axons to this part of the brain. This is the key consideration.
CONCLUDING REMARKS
In the sections above, we have highlighted only selective examples of ventrome-dial temporal area abnormalities that could deafferent the ventral striatopallidal andmagnocellular parts of the basal forebrain. Many others could be added to the list,and they also have behavioral sequela as clinical features. While it is partially arbi-trary to focus only on basal forebrain targets of the ventromedial temporal area ax-ons, it is also important to recognize that the latter structures do not have widespreadfrontal lobe projections.33,35,92 Indeed, ventromedial temporal areas project prima-rily to only those frontal areas that give rise to ventral striatopallidal and magnocel-lular nuclei projections. For example, the subicular/CA 1 zone of the hippocampalformation, the deep layers of the entorhinal cortex and the basal nuclei of theamygdala project powerfully to the anterior cingulate, medial frontal and the orbito-frontal cortices, and only modestly so, if at all, to the dorsolateral and frontal polarcortex—the classical prefrontal granular cortices. Temporal input to these areas ariselargely from more proximal sensory association cortices in the parietal, occipital andtemporal lobes. We are far more impressed with the fact that diseases and injury tothe ventromedial temporal area often target exactly those structures and layers ofcortex that project powerfully to the basal forebrain and to those frontal areas thatalso send axons to the ventral striatum and magnocellular cell groups.
A fundamental principle of mamallian neuroanatomy is that ventromedial tempo-ral areas are at the endstation of corticocortical association systems for allmodalities82,89 and process on line sensory events in real time. These same corticalassociation pathways that converge onto ventromedial temporal areas sequentiallypass through both proximal and distal association cortices that are repositories forpast learning and memory. Thus, the input to ventromedial temporal areas, thatproject to the basal forebrain, has both real-time and historical-time elements asso-ciated with it. Moreover, hippocampal output is adjoined with this for elements ofsignificance, novelty and relevancy.84,89 The same cannot be said for proximal asso-ciation projections to classical dorsolateral and frontal polar prefrontal cortices.
All things considered, we view the basal forebrain in a highly unique positionwhere, by virtue of the digest of its multimodal and hippocampal input, it can orga-nize, or at least predispose, the organism for appropriate emotional, motor, endo-crine and autonomic responses exactly in register with real time perception in thecontext of past history, novelty and its relevance. It is certainly arguable that deficits

590 ANNALS NEW YORK ACADEMY OF SCIENCES
involving psychoses are reducible to deficiencies in appropriate response selectionsto the richly textured real-time, historical-time and memory-related information thatthe basal forebrain receives. As demonstrated recently by Richmond and his collab-orators,76 these are essential elements for a predictable reality, and ventral striatalunits are tuned keenly to these contingencies in primates. A major deficit in schizo-phrenia may lie largely in utilizing real-time, historical-time and memory-related in-formation for predictions about reality. If this is the case, altered ventromedialtemporal areas would be expected to have a large voice in the abnormality.
ACKNOWLEDGMENTS
This research was supported by NIH Grants NS 14944 and PO NS 19632. Wethank Sherry Lohman for library research and typing the manuscript, Paul Reimannfor photography, and Darrell Wilkins for tissue acquisition from the University ofIowa Deeded Body Program.
REFERENCES
1. ADAMS, J.H. 1984. The pathophysiology of raised intracranial pressure. In Green-field’s Neuropathology, 4th edit. J.H. Adams, J.A.N. Corsellis & L. W. Dunchen,Eds. John Wiley & Sons. New York.
2. ADAMS, J.H., D.I. GRAHAM, G. SCOTT, L.S. PARKER & D. DOYLE. 1980. Brain damagein non-missile head injury. J. Clin. Pathol. 33: 1132–1145.
3. AGGLETON, J.P., D.P. FRIEDMAN & M. MISHKIN. 1987. A comparison between the con-nections of the amygdala and hippocampus with the basal forebrain in the macaque.Exp. Brain Res. 67: 556–568.
4. AKIL, M. & D.A. LEWIS. 1997. Cytoarchitecture of the entorhinal cortex in schizo-phrenia. Am. J. Psychiatry 154: 1010–1012.
5. ALHEID, G.F. & L. HEIMER. 1988. New perspectives in basal forebrain organization ofspecial relevance for neuropsychiatric disorders: the striatopallidal, amygdaloid, andcorticopetal components of substantia innominata. Neuroscience 27: 1–39.
6. ALTSCHULER, L.L., A. CONRAD, J.A. KOVELMAN & A. SCHEIBEL. 1987. Hippocampalpyramidal cell orientation in schizophrenia. Arch. Gen. Psychiatry 44: 1094–1098.
7. ALVAREZ, P., S. ZOLA-MORGAN & L.R. SQUIRE. 1995. Damage limited to the hippoc-ampal region produces long-lasting memory impairment in monkeys. J. Neurosci.15(5): 3796–3807.
8. AMARAL, D.G. & J.L. PRICE. 1984. Amygdalo-cortical projections in the monkey. J.Comp. Neurol. 230: 465–496.
9. AMARAL, D.G., J.L. PRICE, A. PITKANEN & S.T. CARMICHAEL. 1992. Anatomical orga-nization of the primate amygdaloid complex. In The Amygdala. J.P. Aggleton, Ed.:1–66. Wiley-Liss. New York.
10. ARNOLD, S.E., B.R. FRANZ, R.C. GUR, R.E. GUR, R.M. SHAPIRO, P.J. MOBERG & J.Q.TROJANOWSKI. 1995. Smaller neuron size in schizophrenia in hippocampal sub-fields that mediate cortical-hippocampal interactions. Am. J. Psychiatry 152: 738–748.
11. ARNOLD, S.E., B.T. HYMAN, J. FLORY, A.R. DAMASIO & G.W. VAN HOESEN. 1991.The topographical and neuroanatomical distribution of neurofibrillary tangles andneuritic plaques in the cerebral cortex of patients with Alzheimer’s disease. Cere-bral Cortex 1(Jan/Feb): 103–116.
12. ARNOLD, S.E., V.M.Y. LEE, R.E. GUR & J.Q. TROJANOWSKI. 1991. Abnormal expres-sion of two microtubule-associated proteins (MAP2 and MAP5) in specific sub-

591VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
fields of the hippocampal formation in schizophrenia. Proc. Natl. Acad. Sci. USA88: 10850–10854.
13. ARNOLD, S.E., D.D. RUSCHEINSKY & L.-Y. HAN. 1997. Further evidence of abnormalcytoarchitecture of the entorhinal cortex in schizophrenia using spatial point pat-tern analyses. Biol. Psychiatry 42(8): 639–647.
14. ARNOLD, S.E. & J.Q. TROJANOWSKI. 1996. Recent advances in defining the neuropa-thology of schizophrenia. Acta Neuropathol. 92: 217–231.
15. BENES, F.M. 1993. Neurobiological investigations in cingulate cortex of schizo-phrenic brain. Schizophrenic Bull. 19: 537–549.
16. BENES, F.M., I. SORENSON & D.E. BIRD. 1991. Reduced neuronal size in posteriorhippocampus of schizophrenic patients. Schizophrenic Bull. 17: 597–608.
17. BOGERTS, B., M. ASHTARI, G. DEGREEF, J.M. ALVIR, R.M. BILDER & J.A. LIEBERMAN.1990. Reduced temporal limbic structure volumes on magnetic resonance imagesin first episode schizophrenia. Psychiatry Res. Neuroimaging 35: 1–13.
18. BOGERTS, B., J.A. LIEBERMAN, M. ASHTARI, R. BILFRT, G. DEGREEF, G. LERNER, C.JOHNS & S. MASIAR. 1993. Hippocampus-amygdala volumes and psychopathologyin chronic schizophrenia. Biol. Psychiatry. 33: 236–246.
19. BRAAK, H. & E. BRAAK. 1992. The human entorhinal cortex: normal morphology andlamina-specific pathology in various diseases. Neurosci. Res. 15: 6–31.
20. BRAAK, H. & E. BRAAK. 1991. Neuropathological staging of Alzheimer-relatedchanges. Acta Neuropathol. 82: 239–259.
21. BRAAK, H. & E. BRAAK. 1985. On areas of transition between entorhinal allocortexand temporal isocortex in the human brain. Normal morphology and lamina-spe-cific pathology in Alzheimer’s Disease. Acta Neuropathol. 68: 325–332.
22. CARMICHAEL, S.T., M.-C. CLUGNET & J.L. PRICE. 1994. Central olfactory connec-tions in the macaque monkey. J. Comp. Neurol. 346: 403–434.
23. CHRISTISON, G.W., M.F. CASANOVA, D.R. WEINBERGER, R. RAWLINGS & J.E. KLEIN-MAN. 1989. A quantitative investigation of hippocampal pyramidal cell size, shape,and variability of orientation in schizophrenia. Arch. Gen. Psychiatry 46: 1027–1032.
24. CORSELLIS, J.A.N. 1958. Individual variation in the size of the tentorial opening. J.Neurol. Neurosurg. Psychiatry 21: 279–283.
25. DAMASIO, A. R. 1994. Descartes’ Error: Emotion, Reason, and the Human Brain.Grosset/Putnam. New York.
26. DAVISON, K. 1983. Schizophrenia-like psychoses associated with organic cerebraldisorders: a review. Psychiatr. Dev. 1: 1–34.
27. DE OLMOS, J.S. 1990. Amygdala. In The Human Nervous System. G. Paxinos, Ed.:583–710. Academic Press, Inc. San Diego.
28. DE OLMOS, J.S., G.F. ALHEID & C.A. BELTRAMINO. 1985. Amygdala. In The Rat Ner-vous System. G. Paximos, Ed.: 223–334. Academic Press. Sydney.
29. DUVERNOY, H.M. 1988. The Human Hippocampus: An Atlas of Applied Anatomy.Bergmann. Munich.
30. ESIRI, M. M., R. C. A. PEARSON, J. E. STEELE, D. M. BOWEN & T. P. S. POWELL.1990. A quantitative study of the neurofibrillary tangles and the choline acetyl-transferase activity in the cerebral cortex and the amygdala in Alzheimer’s disease.J. Neurol. Neurosurg. Psychiatry 53: 161–165.
31. FALKAI, P. & B. BOGERTS. 1986. Cell loss in the hippocampus of schizophrenics.Eur. Arch. Psychiatry in Neurol. Sci. 236: 154–161.
32. FALKAI, P., B. BOGERTS & M. ROZUMEK. 1988. Limbic pathology in schizophrenia:The entorhinal region—a morphometric study. Biol. Psychiatry 24: 515–521.
33. FLETCHER, P. 1998. The missing link: a failure of fronto-hippocampal integration inschizophrenia. Nature Neurosci. 1(4): 266–267.
34. FÖRSTL, H., A. BURNS, R. LEVY & N. CAIRNS. 1994. Neuropathological correlates ofpsychotic phenomena in confirmed Alzheimer’s disease. Br. J. Psychiatry 165: 53–59.
35. GOLDMAN-RAKIC, P.S. 1991. Prefrontal cortical dysfunction in schizophrenia: therelevance of working memory. In Psychopathology and the Brain. B.J. Carroll &J.E. Barrett, Eds.: 1–23. Raven Press. New York.

592 ANNALS NEW YORK ACADEMY OF SCIENCES
36. GÓMEZ-ISLA, T., J.L. PRICE, D.W. MCKEEL, JR., J.C. MORRIS, J.H. GROWDON & B.T.HYMAN. 1996. Profound loss of layer II entorhinal cortex neurons occurs in verymild Alzheimer’s disease. J. Neurosci. 16(14): 4491–5000.
37. GRAHAM, D.I., J.H. ADAMS & T.A. GENNARELLI. 1993. Pathology of brain damage inhead injury. In Head Injury. P. R. Cooper, Ed.: 91–113. Williams and Wilkins.Baltimore, MD.
38. GROENEWEGEN, H.J., P. ROOM, M.P. WITTER & A.H.M. LOHMAN. 1982. Corticalafferents of the nucleus accumbens in the cat, studied with anterograde and retro-grade transport techniques. Neuroscience 7: 977–996.
39. GROENEWEGEN, H.J., E. VERMEULEN-VAN DER ZEE, A. TE KORTSCHOT & M.P. WIT-TER. 1987. Organization of the projections from the subiculum to the ventral stria-tum in the rat. A study using anterograde transport of phaseolus vulgarisleucoagglutinin. Neuroscience 23(1): 103–120.
40. HABER, S. N., K. KUNISHIO, M. MIZOBUCHI & E. LYND-BALTA. 1995. The orbital andmedial prefrontal circuit through the primate basal ganglia. J. Neurosci. 15: 4851–4867.
41. HEIMER, L., G.F. ALHEID, J.S. DE OLMOS, H.J. GROWENEWEGEN, S.N. HABER, R.E.HARLAN & D.S. ZAHM. 1997. The accumbens: beyond the core-shell dichotomy. J.Neuropsychiatry Clin. Neurosci. 9: 354–381.
42. HEIMER, L., J. DE OLMOS, G.F. ALHEID & L. ZABORSKY. 1991. “Perestroika” in thebasal forebrain: opening the border between neurology and psychiatry. Prog. BrainRes. 87: 109–165.
43. HERZOG, A. G. & T. L. KEMPER. 1980. Amygdaloid changes in aging and dementia.Arch. Neurol. 37: 625–629.
44. HYMAN, B.T., L.J. KROMER & G.W. VAN HOESEN. 1988. A direct demonstration ofthe perforant pathway terminal zone in Alzheimer’s disease using the monoclonalantibody Alz-50. Brain Res. 450: 392–397.
45. HYMAN, B.T., G.W. VAN HOESEN, A.R. DAMASIO & C.L. BARNES. 1984. Alzheimer’sdisease: cell-specific pathology isolates the hippocampal formation. Science 225:1168–1170.
46. HYMAN, B.T., G.W. VAN HOESEN, L.J. KROMER & A.R. DAMASIO. 1986. Perforantpathway changes and the memory impairment of Alzheimer’s disease. Ann. Neu-rol. 20(4): 472–481.
47. INSAUSTI, R. 1993. Comparative anatomy of the entorhinal cortex and hippocampusin mammals. Hippocampus 3(Special issue): 19–26.
48. JAKOB, H. & H. BECKMANN. 1994. Circumscribed malformation and nerve cell alter-ations in the entorhinal cortex of schizophrenics. J. Neural Transmission 98: 83–106.
49. JAKOB, H. & H. BECKMANN. 1989. Gross and histological criteria for developmentaldisorders in brains of schizophrenics. J. Royal Soc. Med. 82: 466–469.
50. JAKOB, H. & H. BECKMANN. 1986. Prenatal developmental disturbances in the limbicallocortex in schizophrenics. J. Neural Transmission 65: 303–326.
51. JEFFERSON, G. 1938. The tentorial pressure cone. Arch. Neurol. Psychiatry 40: 857–876.
52. KELLEY, A. E. & V. B. DOMESICK. 1982. The distribution of the projection from thehippocampal formation to the nucleus accumbens in the rat: an anterograde- andretrograde-horseradish peroxidase study. Science 7(10): 2321–2335.
53. KEMPER, T.L. 1983. Organization of the Neuropathology of the amygdala in Alzhe-imer’s disease. In Biological Aspects of Alzheimer’s Disease, Banbury Report 15.R. Katzman, Ed.: 31–35. Cold Spring Harbor Laboratory.
54. KOSEL, K.C., G.W. VAN HOESEN & D.L. ROSENE. 1982. Non-hippocampal corticalprojections from the entorhinal cortex in the rat and rhesus monkey. Brain Res.244: 210–213.
55. KOVELMAN, J. A. & A. B. SCHEIBEL. 1984. A neurohistological correlate of schizo-phrenia. Biol. Psychiatry 19: 1601–1621.
56. KRAYNIAK, P. F., R. C. MEIBACH & A. SIEGEL. 1981. A projection from the entorhi-nal cortex to the nucleus accumbens in the rat. Brain Res. 209: 427–431.

593VAN HOESEN et al.: VENTROMEDIAL TEMPORAL LOBE PATHOLOGY
57. KRIMER, L.S., M.M. HERMAN, R.C. SAUNDERS, J.C. BOYD, T.M. HYDE, J.M. CARTER,J.E. KLEINMAN & D.R. WEINBERGER. 1997. A qualitative and quantitative analysisof the entorhinal cortex in schizophrenia. Cerebral Cortex 7: 732–739.
58. KROMER VOGT, L.J., B.T. HYMAN, G.W. VAN HOESEN & A.R. DAMASIO. 1990. Patho-logical alterations in the amygdala in Alzheimer’s disease. Neuroscience 37(2):377–385.
59. LYKETSOS, C.G., C. STEELE, E. GALIK, A. ROSENBLATT, M. STEINBERG, A. WARREN &J.-M. SHEPPARD. 1999. Physical aggression in dementia patients and its relation-ship to depression. Am. J. Psychiatry 156: 66–71.
60. MCSHANE, R., J. KEENE, C. FAIRBURN, R. JACOBY & T. HOPE. 1998. Psychiatricsymptoms in patients with dementia predict the later development of behavioralabnormalities. Psychol. Med. 28: 1119–1127.
61. MEGA, M.S. & J.L. CUMMINGS. 1994. Frontal-subcortical circuits and neuropsychiat-ric disorders. J. Neuropsychiatry 6: 358–370.
62. MESULAM, M.M. & E.J. MUFSON. 1984. Neural inputs into the nucleus basalis of thesubstantia innominata (Ch4) in the rhesus monkey. Brain 107: 253–274.
63. MEYER, A. 1920. Herniation of the brain. Arch. Neurol. Psychiatry. 4: 387–400.64. MILLER, J.D. & J.H. ADAMS. 1984. The pathophysiology of raised intracranial pres-
sure. In Greenfield’s Neuropathology, 4th edit. J.H. Adams, J.A.N. Corsellis &L.W. Duchen, Eds. John Wiley & Sons. New York.
65. MURPHY, G.M., JR. & W.G. ELLIS. 1991. The amygdala in Down’s syndrome andfamilial Alzheimer disease: four clinicopathological case reports. Biol. Psychiatry30: 92–106.
66. MURRAY, E. A., D. GAFFAN & M. MISHKIN. 1993. Neural substrates of visual stim-ulu-stimulus association in rhesus monkeys. J. Neurosci. 13: 4549–4561.
67. PAPEZ, J.W. 1937. A proposed mechanism of emotion. Arch. Neurol. Psychiatry 38:725–743.
68. PRICE, J.L., S.T. CARMICHAEL & W.C. DREVETS. 1996. Networks related to theorbital and medial prefrontal cortex. In The Emotional Motor System. G. Holstege,R. Bandler & D.B. Saper, Eds.: 461–484. Elsevier Science, Ltd.
69. RETZIUS, G. 1896. Das menschenhirn. Studien in der makroskopischen morphologie.Norstedt & Sohne. Stockholm.
70. ROSENE, D.L. & G.W. VAN HOESEN. 1977. Hippocampal efferents reach widepreadareas of cerebral cortex and amygdala in the rhesus monkey. Science 198: 315–317.
71. ROSENE, D. L. & G. W. VAN HOESEN. 1987. The hippocampal formation of the pri-mate brain: a review of some comparative aspects of cytoarchitecture and connec-tions. In Cerebral Cortex, vol. 6. Plenum Press. New York.
72. ROSOKLIJA, G., M.A. KAUFMAN & D. LIU. 1995. Subicular MAP-2 immunoreactivityin schizophrenia (abstract). Soc. Neurosci. Abstracts 21: 2126.
73. SCHEIBEL, A.B. & J.A. KOVELMAN. 1981. Disorientation of the hippocampal pyrami-dal cells and its processes in the schizophrenic patient. Biol. Psychiatry 16: 101–102.
74. SCHMAJUK, N.A. & M. TYBERG. 1990. The hippocampal lesion model of schizophre-nia. In Neuromethods: Animal Models in Psychiatry, vol. 19. A.A. Boulton, G.B.Baker & M.T.P. Martin-Iverson, Eds. Humana Press. Clifton, NJ.
75. SCOVILLE, W.B. & B. MILNER. 1957. Loss of recent memory after bilateral hippoc-ampal lesions. J. Neurol. Neurosurg. Psychiatry 20: 11.
76. SHIDARA, M., T.G. AIGNER & B.J. RICHMOND. 1998. Neuronal signals in the monkeyventral striatum related to progress through a predictable series of trials. J. Neuro-sci. 18(7): 2613–2625.
77. SLATER, E. & A.W. BEARD. 1963. The schizophrenia-like psychoses of epilepsy. Br.J. Psychiatry 109: 95–150.
78. SORENSEN, K.E. 1985. Projections of the entorhinal area to the striatum, nucleusaccumbens, and cerebral cortex in the guinea pig. J. Comp. Neurol. 238: 308-322.
79. STEVENS, J.R. 1997. Anatomy of schizophrenia revisited. Schizophrenic Bull. 23:373–383.

594 ANNALS NEW YORK ACADEMY OF SCIENCES
80. SUZUKI, W.A., S. ZOLA-MORGAN, L.R. SQUIRE & D.G. AMARAL. 1993. Lesions of theperirhinal and parahippocampal cortices in the monkey produce long-lasting mem-ory impairment in the visual and tactual modalities. J. Neurosci. 13(6): 2430–2451.
81. TOTTERDELL, S. & G.E. MEREDITH. 1997. Topographical organization of projectionsfrom the entorhinal cortex to the striatum of the rat. Neuroscience 78(3): 715–729.
82. VAN ESSEN, D.C., D.J. FELLEMAN, E.A. DEYOE, J. OLAVARRIA & J. KNIERIM. 1990.Modular and hierarchical organization of extrastriate visual cortex in the macaquemonkey. Cold Spring Harbor Symp. Quant. Biol. 3: 679–696.
83. VAN HOESEN, G.W. 1995. Anatomy of the medial temporal lobe. Mag. ResonanceImaging 13(8): 1047–1055.
84. VAN HOESEN, G.W. 1982. The parahippocampal gyrus: new observations regardingits cortical connections in the monkey. Trends Neurosci. 5: 345–350.
85. VAN HOESEN, G.W. 1997. Ventromedial temporal lobe anatomy, with comments onAlzheimer’s disease and temporal injury. J. Neuropsychiatry Clin. Neurosci. 9:331–341.
86. VAN HOESEN, G.W. & A.R. DAMASIO. 1987. Neural correlates of the cognitiveimpairment in Alzheimer’s disease. In Higher Functions of the Nervous System,the Handbook of Physiology. F. Plum, Ed.: 871–898.
87. VAN HOESEN, G.W., M.-M. MESULAM & R. HAAXMA. 1976. Temporal cortical pro-jections to the olfactory tubercle in the rhesus monkey. Brain Res. 109: 375–381.
88. VAN HOESEN, G.W. & D.N. PANDYA. 1975. Some connections of the entorhinal (area28) and perirhinal (area 35) cortices of the rhesus monkey. I. Temporal lobe affer-ents. Brain Res. 95: 1–24.
89. VAN HOESEN, G.W., D.N. PANDYA & N. BUTTERS. 1972. Cortical afferents to theentorhinal cortex of the rhesus monkey. Science 175: 1471–1473.
90. VAN HOESEN, G.W., D.N. PANDYA & N. BUTTERS. 1975. Some connections of theentorhinal (area 28) and perirhinal (area 35) cortices of the rhesus monkey. II.Frontal lobe afferents. Brain Res. 95: 25–38.
91. VEREECKEN, H.L.G., O.J.M. VOGELS & R. NIEUWENHUYS. 1994. Neuron loss andshrinkage in the amygdala in Alzheimer’s disease. Neurobiol. Aging 15(1): 45–54.
92. WEINBERGER, D.R. 1991. Anteromedial temporal-prefrontal connectivity: a func-tional neuroanatomical system implicated in schizophrenia. In Psychopathologyand the Brain. B.J. Carroll & J.E. Barret, Eds.: 25–43. Raven Press. New York.
93. WITTER, M., H.J. GROENEWEGEN, F.H. LOPES DA SILVA & A.H.M. LOHMAN. 1989.Functional organization of the extrinsic and intrinsic circuitry of the parahippoc-ampal region. Prog. Neurobiol. 33: 161–254.
94. WITTER, M.P. 1993. Organization of the entorhinal-hippocampal system: A reviewof current anatomical data. Hippocampus 3: 33–44.
95. WRIGHT, C.I., A.V. BEIJER & H.J. GROENEWEGEN. 1996. Basal amygdaloid complexafferents to the rat nucleus. J. Neurosci. 16(5): 1877–1893.
96. ZOLA-MORGAN, S., L.R. SQUIRE, R.P. CLOWER & N.L. REMPEL. 1993. Damage to theperirhinal cortex exacerbates memory impairment following lesions to the hippoc-ampal formation. J. Neurosci. 13(1): 251–265.




















