
Use of replication deficient adenoviruses to investigate the role of G proteins in Ca 2+ signalling...
7
Research Cell Calcium (1998) 24(2). 97-l 03 0 Harcourl Brace &Co. Ltd 1998 Use of replication deficient adenoviruses to investigate the role of G proteins in Ca*+ signalling in epithelial cells P. Poronnikl, L.M. O’Mullanel, E.A. Harding’, R. Greger*, D.I. Cook’ ‘Department of Physiology, University of Sydney, New South Wales, Australia 2Physiologisches Institut, Albert-Ludwigs-Universit&it, Freiburg, Germany Summary Here we report on the feasibility of using replication deficient adenoviruses to modify signal transduction systems in epithelia. We constructed two viruses, one expressing a dominant negative mutant of the a-subunit of G, (Ad-EFl-dnGa,) and the other expressing the wild-type o-subunit of G, (Ad-EFl-wtGa,). We used an adenovirus expressing green fluorescent protein (Ad-EFl-GFP20) to show that infection of cultured cells with an adenovirus results in at least 95% expression of the transgene in both HSG and HT29 cells. We also used an adenovirus that expresses no transgene (Ad-MX17) to demonstrate that adenoviral infection itself does not affect the resting concentration of cytosolic Ca2+ ([Ca”],) or the carbachol responses in these cells. We further show that Ad-EFl-dnGa, inhibits the increase in [Ca2+], produced by muscarinic receptor activation in both the cell lines we studied. This inhibitory effect is not shared by Ad-EFl -v&Go,, which indicates that in both HSG and HT29 cells, the increase in [Ca2+li produced by muscarinic receptor activation is largely mediated by activation of G,. Neither virus affected the resting level of [Ca2+], in these cells. Our findings confirm the feasibility of using replication deficient adenoviruses expressing dominant negative mutants to investigate the role of G proteins in signal transduction systems. INTRODUCTION GTP-binding regulatory proteins (G proteins) are a heterogeneous family of proteins that play a pivotal role in many cellular signalling pathways [l]. One of these pathways links activation of muscarinic receptors by agonists such as acetylcholine and carbachol to an increase in [Ca’+], [2]. In this pathway, the activated muscarinic receptor stimulates phospholipase Cp to release inositol-1,4,5-t&phosphate (IPJ from the Received 7 May 1998 Revised 17 June 1996 Accepted 24 June 1996 Correspondence to: Prof. D.I. Cook, Department of Physiology (F-13), University of Sydney, NSW 2006, Australia Fax: +61 2 9351 9926 E-mail: [email protected] membrane lipid phosphatidylinositol-4,5-bisphosphate. IP, then increases [Ca2+], by activating release of Ca2+ from intracellular stores and by stimulating Caz+ influx from the extracellular medium 121. This inilttx is probably triggered by depletion of the Ca2+ within the intracellular Ca2+stores [3]. The best attested role of G proteins in this pathway is by activating phospholipase Cl3 in response to activation of the muscarinic receptor, which in some tissues appears to be mediated by the pertussis-toxin insensitive G, whereas in others it is believed to be mediated by pertussis toxin-sensitive G proteins of the Gi, class 141.Other roles have been suggested, however, ‘such as modulating release of Ca2+from intracellular stores [5-71. The physiological analysis of the role of G proteins in Ca2+ signalling has relied heavily on the use of antisense techniques to modulate the level of expression of G proteins in cultured cells [6,8], and on the use of permeabilised cell and whole-cell patch-clamp techniques to introduce antibodies and other proteins to the cytosol 97
Transcript of Use of replication deficient adenoviruses to investigate the role of G proteins in Ca 2+ signalling...

Research Cell Calcium (1998) 24(2). 97-l 03 0 Harcourl Brace &Co. Ltd 1998
Use of replication deficient adenoviruses to investigate the role of G proteins in Ca*+ signalling in epithelial cells
P. Poronnikl, L.M. O’Mullanel, E.A. Harding’, R. Greger*, D.I. Cook’ ‘Department of Physiology, University of Sydney, New South Wales, Australia 2Physiologisches Institut, Albert-Ludwigs-Universit&it, Freiburg, Germany
Summary Here we report on the feasibility of using replication deficient adenoviruses to modify signal transduction systems in epithelia. We constructed two viruses, one expressing a dominant negative mutant of the a-subunit of G, (Ad-EFl-dnGa,) and the other expressing the wild-type o-subunit of G, (Ad-EFl-wtGa,). We used an adenovirus expressing green fluorescent protein (Ad-EFl-GFP20) to show that infection of cultured cells with an adenovirus results in at least 95% expression of the transgene in both HSG and HT29 cells. We also used an adenovirus that expresses no transgene (Ad-MX17) to demonstrate that adenoviral infection itself does not affect the resting concentration of cytosolic Ca2+ ([Ca”],) or the carbachol responses in these cells. We further show that Ad-EFl-dnGa, inhibits the increase in [Ca2+], produced by muscarinic receptor activation in both the cell lines we studied. This inhibitory effect is not shared by Ad-EFl -v&Go,, which indicates that in both HSG and HT29 cells, the increase in [Ca2+li produced by muscarinic receptor activation is largely mediated by activation of G,. Neither virus affected the resting level of [Ca2+], in these cells. Our findings confirm the feasibility of using replication deficient adenoviruses expressing dominant negative mutants to investigate the role of G proteins in signal transduction systems.
INTRODUCTION
GTP-binding regulatory proteins (G proteins) are a heterogeneous family of proteins that play a pivotal role in many cellular signalling pathways [l]. One of these pathways links activation of muscarinic receptors by agonists such as acetylcholine and carbachol to an increase in [Ca’+], [2]. In this pathway, the activated muscarinic receptor stimulates phospholipase Cp to release inositol-1,4,5-t&phosphate (IPJ from the
Received 7 May 1998 Revised 17 June 1996 Accepted 24 June 1996
Correspondence to: Prof. D.I. Cook, Department of Physiology (F-13), University of Sydney, NSW 2006, Australia
Fax: +61 2 9351 9926 E-mail: [email protected]
membrane lipid phosphatidylinositol-4,5-bisphosphate. IP, then increases [Ca2+], by activating release of Ca2+ from intracellular stores and by stimulating Caz+ influx from the extracellular medium 121. This inilttx is probably triggered by depletion of the Ca2+ within the intracellular Ca2+ stores [3]. The best attested role of G proteins in this pathway is by activating phospholipase Cl3 in response to activation of the muscarinic receptor, which in some tissues appears to be mediated by the pertussis-toxin insensitive G, whereas in others it is believed to be mediated by pertussis toxin-sensitive G proteins of the Gi, class 141. Other roles have been suggested, however, ‘such as modulating release of Ca2+ from intracellular stores [5-71.
The physiological analysis of the role of G proteins in Ca2+ signalling has relied heavily on the use of antisense techniques to modulate the level of expression of G proteins in cultured cells [6,8], and on the use of permeabilised cell and whole-cell patch-clamp techniques to introduce antibodies and other proteins to the cytosol
97

98 P Poronnik, L M O’Mullane, E A Harding, R Greger, 0 I Cook
[9-l 11. These approaches, however, suffer from significant drawbacks. First, they are d&cult, if not impossible to apply in vivo. Second, the use of whole-cell patch-clamp or permeabilised cell techniques to introduce proteins such as antibodies or dominant negative mutants into the cytosol may itself perturb signalling systems by dialysing out essential mediators or by disturbing the normal ionic composition of the cytosol [lo].
For these reasons we decided to investigate the use of replication deficient adenoviruses as a means for mani- pulating signal transduction systems. The use of adenoviruses to dissect a signal transduction pathway requires that they over-express a protein, such as a dominant negative mutant, which will selectively interfere with a specific component of that signal transduction pathway. This approach is theoretically feasible for G protein a subunits, because a single amino acid mutation in the GTP binding consensus sequence renders the a-
subunit unable to bind GTP [ 121. These mutant (dominant negative) a-subunits are thus refractory to activation by Gprotein-coupled receptors and, in transfected cell lines, have been shown to inhibit the effect of wild-type G protein a-subunits [12-l 51.
In this paper, we show that a replication deficient adenovirus expressing a dominant negative mutant of the a-subunit of G, can be used to inhibit the increase in [Ca2+li produced by muscarinic receptor activation in epithelial cells. We also investigate the impact of various experimental parameters on the magnitude of this inhibition.
MATERIALS AND METHODS
Adenoviral construction
An adenoviral shuttle vector, pAC-EFl, was constructed containing the human elongation factor 1 alpha (EF-la) promoter, a polylinker and a porcine growth hormone poly-A tail. Since EF-la is a constitutive mammalian promoter it gives reliable expression in a variety of cell types [ 161. The EF- 1 a promoter cassette was cut from the plasmid pC-EFl (Dr J.S. Gutkind, NIDR, NIH) as a blunt fragment with Sspl and PvuII. The CMV promoter cassette from the adenoviral shuttle backbone pAC-CMV was excised using Not1 and the adenoviral shuttle backbone blunted by endfilling with Klenow’s reagent. The two DNA fragments were subsequently ligated to yield a new adenoviral shuttle vector, pAC-EFl. This vector was then used to generate the GFP20 and Ga4 expressing viruses.
The cDNAs for the mouse wild-type (wt) a subunit of G, and the dominant negative (dn) mutant of the mouse a subunit of G, were obtained from Dr J.S. Gutkind (NIDR, NII-I). The dominant negative mutant has a single
Cell Calcium (1998) 24(2), 97- 103
ammo acid substitution (G207T) in the GTP-binding consensus sequence which renders it unable to bind GTP and thus unable to be activated [12]. The cDNAs were subcloned as Not1 fragments into the adenoviral shuttle vector, pAC-EFl. Replication deficient human type 5 adenoviruses with gene inserts in the El-deleted region were then produced by homologous recombination between the shuttle vector plasmids and the adenovirus plasmid pJMl7 in 293 cells following calcium phosphate transfection using standard techniques [ 171. Viruses were subsequently plaque-purified and CsCl purified stocks prepared and quantified by plaque assay using standard techniques [ 171.
A virus expressing a humanised mutant of green fluorescent protein (GFP20) under the control of the EF-1 promoter was also constructed. GFP20 was designed by Dr S. Aota (BERI, Osaka, Japan) and contains the mutations, S65A, V68L, S72A, F99S, M153T and V163A. It was excised as a ClaI and HindIII fragment, blunted with Pfu polymerase and was then ligated into the EcoRV site of pAC-EFl. An adenovirus (Ad-EFl-GFP20) was sub- sequently rescued and plaque-purified as described above.
Virus stocks were stored at -80°C in PBS/IO% glycerol + 1 mmol/l MgCl, and were not thawed more than 5 times.
Cell culture and virus infection
Human salivary gland carcinoma cells (HSG) [ 181 were cultured in EMEM and human colonic carcinoma (HT29) cells (ATCC) in Ham’s F 12/DMEM medium. Both media were supplemented with 2 mmol/l glutamine, 10% FCS and antibiotics. Cells were split by trypsinisation every 3-4 days. For fluorimetry, cells were seeded onto 1 cm diameter glass coverslips in 48-well plates (Costar) at a density of 2 x lo4 cells per well, a density which ensured that confluent monolayers were not formed by the time of experimentation. Cells were infected 24 h after plating out by adding the virus in 200 ~1 of culture medium. A further 24 h later, the wells were supplemented with 300 ~1 of fresh medium. We have given the dose of adenovirus as the concentration of virus in the bulk solution since this is the major determinant of uptake of virus by the cells [19]. Using the infection procedures described in this paper a viral concentration of 1 x IO5 pfu/ltl corresponds to a dose of 100 pfi.r/cell applied to a confluent monolayer.
Western blotting
For Western blots, cells were grown to confluence in 60 mm Petri dishes, infected with either 3 x IO4 pfu/l.d or 1 x lo5 pfu/ul of adenovirus and harvested after 3 days. Monolayers were then washed with ice cold PBS, scraped off in 200 ~1 of PBS plus 1% X-Triton with 5 mmol/l EDTA
0 Harcourt Brace & Co. Ltd 1998
Replication deficient adenoviruses in epithelia 99
and 5 @ml each of aprotinin, leupeptin and cz2- macroglobulin. Cells were then disrupted by trituration through a 200 ~1 pipette tip and centrifuged at 14000 rpm at 4°C for 15 min. The Triton X- 100 soluble fraction was then mixed with gel sample buffer and 5% B- mercaptoethanol and heat denatured. 40 ~1 of each sample were then loaded onto a 12% Tris-glycine polyacrylamide minigel (Novex) and electrophoresed. The gel was subsequently blotted onto nitrocellulose (Amersham). The blot was incubated with 1:500 of a rabbit polyclonal anti-Gas antibody (Dr J.S. Gutkind, NIDR, NIH), followed by incubation with a second antibody (1:1500, Amersham) and finally with a streptavidin-horseradish peroxidase conjugate (Amer- sham). The blot was visualised using the standard ECL protocol (Amersham). The X-ray films were then scanned into a computer and the final figure produced using Adobe Photoshop.
Calcium imaging
Cells on the coverslips were loaded by the addition of 75 l.tmol/l Fura-2/AM plus 0.3% of the nonionic detergent Pluronic (to aid in the dispersal of the Fura-2) to the medium and were then incubated on a rocker at 37°C for 45 min. The coverslips were then removed and mounted in a superfusion chamber and placed on the stage of a Zeiss Axiovert fluorescence microscope. A long working distance 20x objective was used to visualise small groups of 20-40 cells. Excitation of the Fura- loaded cells at 340 nm and 380 nm was achieved by using a Sutter filter wheel driven by the Ionvision software package (Improvision, Coventry, UK) running on a Macintosh IIci. Images were acquired using a Videoscope CCD camera and a Pixelgrabber framegrabber board as previously described [20]. The superfusion solutions were heated to 37°C by a water jacket around the perfusion lines. The perfusion solution contained (in mmol/l): NaCl 145.0, KC1 5.5, MgCl, 1.2, NaH,PO, 1.2, Na-HEPES 75, H-HEPES 75, D-glucose 10.0 and CaCl, 1.0 at pH 74.
Confocal microscopy
Confocal microscopy was performed on cells grown on coverslips using a Leica confocal microscope and a 40x NA 1.2 oil immersion objective. GFP20 was visualised using the FITC filter set.
Materials
All restriction enzymes and Klenow’s reagent were from Boehringer. Ligations were performed using the Rapid Ligation Kit from Boehringer. All transformations were performed using Gibco Maximum efficiency DH5a
A
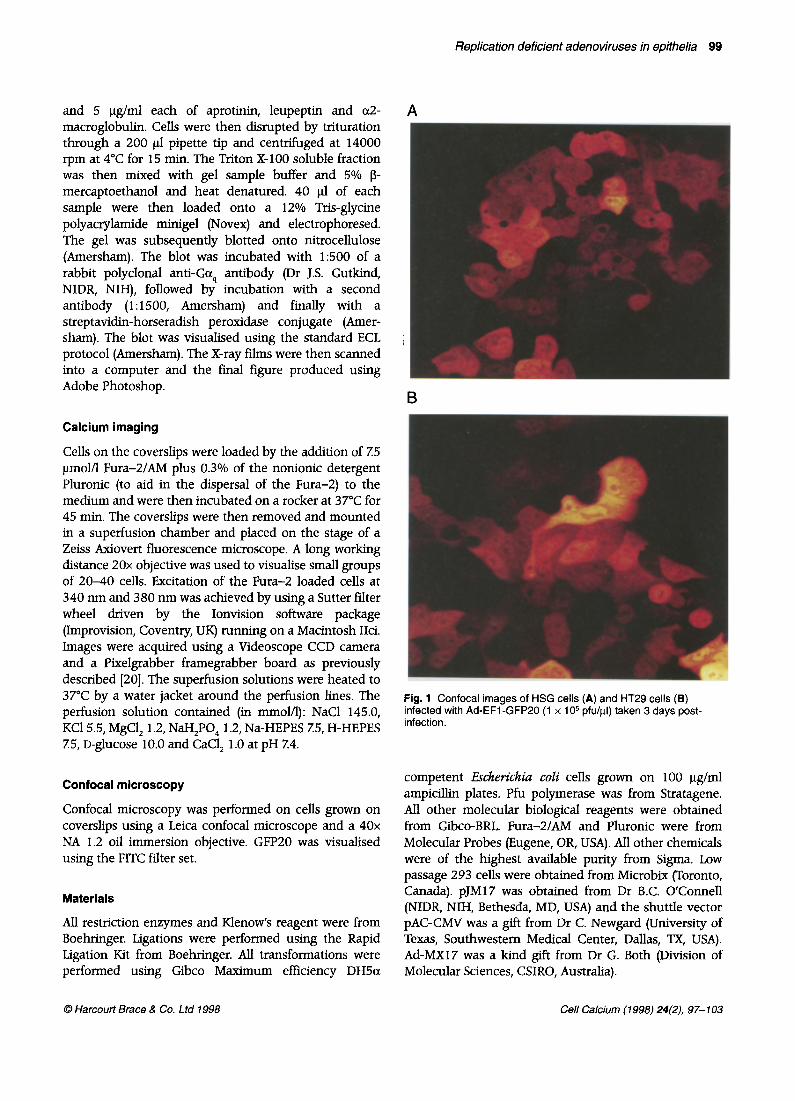
Fig. 1 Confocal images of HSG cells (A) and HT29 cells (B) infected with Ad-EFl-GFP20 (1 x lo5 pfu/pl) taken 3 days post- infection.
competent E.&e&& coli cells grown on 100 pg/ml ampicillin plates. Pfu polymerase was from Stratagene. All other molecular biological reagents were obtained from Gibco-BRL. Fura-21AM and Pluronic were from Molecular Probes (Eugene, OR, USA). All other chemicals were of the highest available purity from Sigma. Low passage 293 cells were obtained from Microbix (Toronto, Canada). pJM17 was obtained from Dr B.C. O’Connell (NIDR, NIH, Bethesda, MD, USA) and the shuttle vector pAC-CMV was a gift from Dr C. Newgard (University of Texas, Southwestern Medical Center, Dallas, TX, USA). Ad-MX17 was a kind gift from Dr G. Both (Division of Molecular Sciences, CSIRO, Australia).
0 Harcourt Brace & Co. L fd 1998 Cell Calcium (1998) 24(2), 97- 103
100 P Poronnik, L M O’Mullane, E A Harding, R Greger, D I Cook
RESULTS
Optimisation of virus infection 60-
We frrst assessed the extent to which a human submandibular gland cell line (HSG) and a human colonic cell line (HT29) can be infected by adenoviruses. To do this we used a virus that expresses a humanised GFP mutant (Ad-EFl-GFP20). We found that exposure of HSG cells (Fig. 1A) and HT29 cells (Fig. 1B) to Ad-EFl-GFP20 at a concentration of 1 x lo5 pfu/fi (equivalent to 100 pfu/cell) for 3 days caused all cells to express GFP20 in the cytoplasm, although there was considerable heterogeneity in the levels of GFP expression from cell to cell.
44-
30-
Fura- loaded HSG and HT29 cells respond to exposure to carbachol with an increase in the Fura- ratio, which represents an increase in [Caz+li (Fig. 2A). Preliminary experiments showed that 3 days after infection with 1 x lo5 pfu/).d of an adenovirus that expresses a dominant negative mutant of the o-subunit of G, (Ad-EFl-dnGa ), the increase in Fura- ratio produced in HSG 41 ce s by 100 qol/l carbachol was markedly reduced. Since infection with 1 x lo5 pfu/pl of a
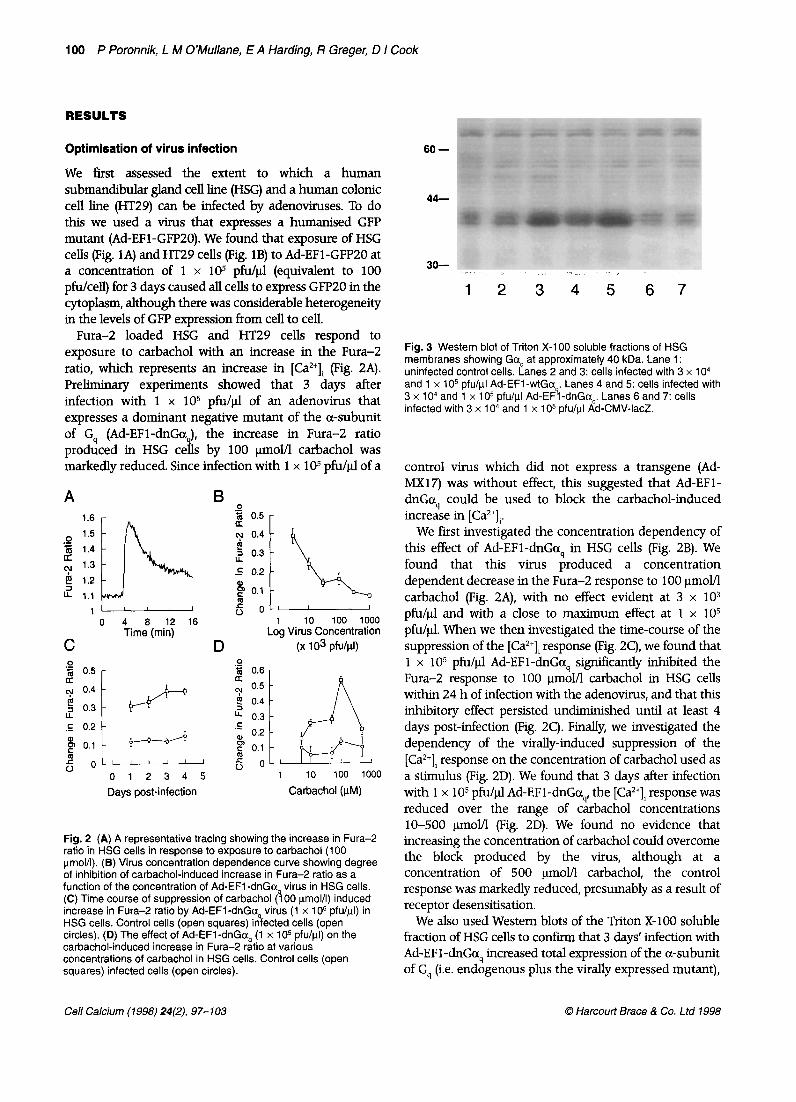
Fig. 3 Western blot of Triton X-l 00 soluble fractions of HSG membranes showing Goq at approximately 40 kDa. Lane 1: uninfected control cells. Lanes 2 and 3: cells infected with 3 x 1 O4 and 1 x 1 O5 pfu/ul Ad-EFl -wtGa,. Lanes 4 and 5: cells infected with 3 x 1 O4 and 1 x 1 O5 pfu/ul Ad-EFl -dnGo,. Lanes 6 and 7: cells infected with 3 x 1 O4 and 1 x 1 O5 pfu/ul Ad-CMV-IacZ.
control virus which did not express a transgene (Ad- MXl7) was without effect, this suggested that Ad-EFl- dnGa, could be used to block the carbachol-induced increase in [Ca”],.
We first investigated the concentration dependency of this effect of Ad-EFl-dnGa, in HSG cells (Fig. 2B). We found that this virus produced a concentration dependent decrease in the Fura- response to 100 ~mol/l carbachol (Fig. 2A), with no effect evident at 3 x lo3 pfu/$ and with a close to maximum effect at 1 x lo5 pfu/t.tl. When we then investigated the time-course of the suppression of the [Ca”], response (Fig. 2C), we found that 1 x lo5 pfu/pl Ad-EFl-dnGc+ significantly inhibited the Fura- response to 100 pmol/l carbachol in HSG cells within 24 h of infection with the adenovirus, and that this inhibitory effect persisted undiminished until at least 4 days post-infection (Fig. 2C). Finally, we investigated the dependency of the virally-induced suppression of the [Ca2+], response on the concentration of carbachol used as a stimulus (Fig. 2D). We found that 3 days after infection with 1 x lo5 pfu/).d Ad-EFl-dnGo,, the [Ca”], response was reduced over the range of carbachol concentrations lo-500 ).mol/l (Fig. 2D). We found no evidence that increasing the concentration of carbachol could overcome the block produced by the virus, although at a concentration of 500 pal/l carbachol, the control response was markedly reduced, presumably as a result of receptor desensitisation.
We also used Western blots of the Triton X-100 soluble fraction of HSG cells to confirm that 3 days’ infection with Ad-EF 1 -dnGaq increased total expression of the a-subunit of G, (i.e. endogenous plus the virally expressed mutant),
,l----- - 6 OLI I I I
0 4 a 12 16 100 1000 Time (min) LJg VirdsYConcentration
C D (x 103 pfu/fll)
5
0
'3 0.6 -
cu 0.5
$ 0.4 -
IL 0.3 -
.c zi
0.2 -
5 0.1 -
6 0-L 1
Days post-infection Carbachol (PM)
Fig. 2 (A) A representative tracing showing the increase in Fura- ratio in HSG cells in response to exposure to carbachol (100 urnoN). (8) Virus concentration dependence curve showing degree of inhibition of carbachol-induced increase in Fura- ratio as a function of the concentration of Ad-EFl -dnGa virus in HSG cells. (C) Time course of suppression of carbachol (100 umol/l) induced increase in Fura- ratio by Ad-EFl -dnGo virus (1 x 1 O5 pfu/ul) in HSG cells. Control cells (open squares) ingfected cells (open circles). (D) The effect of Ad-EFl-dnGa, (1 x lo5 pfu/ul) on the carbachol-induced increase in Fura- ratro at various concentrations of carbachol in HSG cells. Control ceils (open squares) infected cells (open circles).
12 345 67
Cell Calcium (1998) 24(Z), 97-103 0 Harcoutt Brace & Co. ltd 1998

Replication deficient adenoviruses in epithelia 101
0 ‘Z= 1.1 : a 0 4 8 12 16
1.5 D +
I? 1.4
1 .3
1.1 0 4 8 12 16
Time
o 4 8 12 16
NC2
5 1.9
I= 1.8
1.7
1.6
1.5
1.4
D2
1.9 1.8
1.7
1.6
1.5
1.4
1.1 0 4 8 12 16 0 4 8 12 16 0 4 8 12 16
(min)
0 'G
1.2 I 1.2 I 0 4 8 12 16 o 4 8 12 16
Time (min) Fig. 4 Average Fura- responses of HSG cells to 100 umol/l carbachol (all virus concentrations are 1 x 1 O5 pfu/ul). Each curve is the mean of the number of the experiments indicated, with dotted lines representing the standard error of the mean. (A) Uninfected control ceils (n = 28). (B) Cells infected with Ad-MX17 (n = 31). (C) Cells infected with Ad-EFl-dnGa, (n = 31). (D) Cells infected with Ad-EFl -wtGa, (n = 33).
Fig. 5 Average Fura- responses of HT29 cells to 100 kmol/l carbachol (all virus concentrations are 1 x 1 O5 pfu/ul). Each curve is the mean of the number of the experiments indicated, with dotted lines representing the standard error of the mean. (A) Uninfected control cells (n = 15). (B) Cells infected with Ad-MX17 (n = 15). (C) Cells infected with Ad-EFl-dnGaq (n = 17). (D) Cells infected with Ad-EFl-wtGaq (n = 16).
with an increased concentration of adenovirus leading, as expected, to increased expression of GCQ On the other hand, infection with a control virus expressing LacZ was without effect on endogenous levels of Gaq (Fig. 3). Also shown in Figure 3 are the effects on Go4 expression of infection with a virus expressing wild-type a-subunit of Gg (Ad-EFl-wtGc+). It is important to note for the experiments described below that this virus produces similar levels of Gcx expression to those observed following infection with Ad-EF 1-dnGo., (Fig. 3).
Effects of dominant HSG and HT29 cells
negative Gaq on Ca*+ signalling in
In the light of these preliminary studies, we chose a virus concentration of 1 x lo5 pfu/$ (equivalent to 100 pfu/cell) and studied the cells 3 days post-infection. As a control for non-specific effects of adenoviral infection, we used the control adenovirus Ad-MX17 As a control for non-specific effects of over-expression of G protein a- subunits, we used the adenovirus Ad-EFl-wtGa4 which expresses the wild-type o-subunit of G,. The stimulation protocol we used was exposure to 100 l.trnol/l carbachol for 10 min.
Infection with the adenovirus expressing the dominant negative mutant of Gag (Ad-EFl-dnGaJ, however, significantly reduced the magnitude of the peak increase in Fura- ratio from 0.260 f 0.018 (SEM, 1z = 28) to 0.082 f 0.011 (1~ = 31) and the magnitude of the plateau increase in Fura- ratio from 0.061 f 0.007 (n = 28) to 0.016 + 0.006 (n = 31) following stimulation with carbachol (Fig. 4A,C). Infection with the null virus, Ad- MX17, was without effect on these parameters (Fig. 4B), confirming that the inhibitory effect of Ad-EFl-dnGa was not a non-specific consequence of adenovira 9 infection. Similarly, infection with adenovirus expressing the wild-type a subunit of Gs (Ad-EFl-wtGaJ did not have any effect on either the peak or plateau increases in Fura- ratio (Fig. 4D). This result shows that the inhibitory effect of Ad-EFl-dnGc+ is due to interference with the action of endogenous Gg rather than to a non- specific effect of over-expression of G protein a-subunits. Our findings in HT29 cells (Fig. 5) were comparable to those in HSG cells indicating that a similar signalling mechanism operates in these cells.
DISCUSSION
We found that infection with any of these 3 viruses did not alter the resting Fura- ratio in HSG cells (Fig. 4).
In this paper we have shown that adenoviruses can be successfully used to over-express mutant G proteins
0 Harcoud Brace & Co. Ltd 1998 Cell Calcium (1998) 24(2), 97- 103

102 P Poronnik, L M O’Mullane, E A Harding, R Gregec D I Cook
thereby allowing us to elucidate the role of G proteins in signal transduction systems dependent on such proteins. Our data indicate that at virus concentrations of at least up to 1 x lo5 pfu/ul (100 pfu/cell), replication deficient adenovirus infection achieves a high level of transgene expression (Fig. 3), infecting at least 95% of HSG and HT29 cells (Fig. l), but has no significant effect on the carbachol responses in these cells (Figs 4 & 5). We further show that an adenovirus expressing a dominant negative mutant of the a-subunit of G, inhibits the increase in [Ca”], produced by muscarinic receptor activation in both these cell lines (Figs 4 & 5). This inhibitory effect is not shared by an adenovirus expressing the wild-type o- subunit of Gg, indicating that in both HSG and HT29 cells, the increase in [Ca2+], produced by muscarinic receptor activation is largely mediated by activation of Gg.
Previous studies in both HSG and HT29 cells have shown that muscarinic stimulation leads to activation of phospholipase C, with increased intracellular levels of IP, and increased [Ca2+li, due both to release of Ca*+ from intracellular stores and influx of Ca2+ across the plasma membrane [21,22]. It has further been shown that the muscarinic receptor in HT29 cells is of the M3 sub-type [23], and, although the sub-type found in HSG cells is unknown, the predominance of M3 receptors in human salivary tissues [24] makes it seem likely that the receptor in HSG cells is also of the M3 sub-type. M3 receptors have been reported to activate phospholipase Cp by activating G, in a number of tissues including rat parotid cells [25], but in some cell types, M3 receptors have been reported to activate phospholipase CB via the pertussis toxin-sensitive G proteins G, or Go 141. The present findings suggest that muscarinic receptors act via G, rather than Gi, in both HSG and HT29 cells, a conclusion that is ‘consistent with the observation that these responses are insensitive to pertussis toxin (unpublished data).
Although there are several studies using replication deficient adenoviruses to alter cell function by expressing exogenous genes [26-291, the present results are important from the point of view of the use of adenoviruses expressing dominant negative mutants of G protein a-subunits in order to investigate signal transduction systems. To our knowledge, this is the first report studying the use of adenoviruses to analyse G protein linked Ca2+ signalling pathways. Compared to standard transient transfection methods, where the transduction efficiency is typically less than 300/o, the adenoviral technique offers the advantages of rapid and non-disruptive high transduction efficiencies (Figs 1 & 2) and, importantly, the ability to titrate the level of transgene expression (Figs 2B & 3). At the titres used in this study, viral infection also produces few disturbances in cellular function other than those produced by the
Cell Calcium (1998) 24(2), 97- 103
transgene (Figs 4 & 5; [30]). The maximal degree of inhibition obtained using Ad-EF 1 -dnGcc, approximately 60-80% (Figs 4 & 5), compares favourably with the degree of inhibition obtained by antisense methods. It is of interest to note that the inhibitory effect of the Ad- EFl-dnGcrq virus reaches a plateau at approximately 100 pfu/cell (1 x lo5 pfu/ul) and that even a lo-fold further increase in adenoviral concentration does not lead to any significant increase in the degree of inhibition. This probably does not reflect the presence of a component of the activation of phospholipase C being dependent on a G protein other than Gg, since the muscarinic responses in HSG and I-IT29 cells are insensitive to pertussis toxin (data not shown). Rather, it may simply reflect the inability of the cells to produce high enough levels of the mutant protein to inhibit fully the muscarinic response. Some support for this latter possibility comes from the finding that when replication deficient adenoviruses are used to express p-adrenergic receptors in heart muscle, maximal expression and functionality of the transgene, as measured by receptor density and isoproteronol induced CAMP formation, is obtained at 100 pfu/cell, with no increase in activity with higher viral titres [3 11.
It is also noteworthy that the Ad-EFl-wtGag virus did not alter the resting levels of [Ca*+], in either of the cell lines studied (Figs 4 & 5). This is in contrast to a study in which cell lines stably transfected with wild-type Gag have been shown to have increased resting levels [Ca2+], [32]. This difference may reflect lower levels of over- expression produced by adenoviral infections, although it should be noted that clonal selection during the production of stably transfected cell lines often involves the upregulation of other signalling proteins in order to adapt to the over-expression of a single component of a signalling pathway [33].
ACKNOWLEDGEMENTS
This project was supported by the National Health and Medical Research Council of Australia and the Australian Research Council. Dr P.Poronnik is an ARC Postdoctoral Fellow. Prof. D.I. Cook is a Fellow of The Medical Foundation of the University of Sydney. The authors wish to thank Drs B.C. O’Connell, J.S. Gutkind, X.X. Xu and G.W. Both for their essential advice and ongoing support throughout the project and Prof. J.A. Young for helpful discussions.
REFERENCES
1. Hamm H.E. The many faces of G protein signaling. / Biol Chem 1998; 273: 669-672.
2. Clapham D.E. Calcium signaling. Cell 1995; 80: 259-268. 3. Parekh A.B., Penner R. Store depletion and calcium influx.
Physiol Rev 1997; 77: 901-930.
0 Harcourt Brace & Co. Ltd 1998

Replication deficient adenoviruses in epithelia 103
4.
5.
6.
z
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
Exton J.H. Regulation of phosphoinositide phospholipases by hormones, neurotransmitters, and other agonists linked to G proteins. Annu Rev Pharmacol Toxic01 1996; 36: 48 l-509. Berven L.A., Crouch M.F., Katsis F., Kemp B.E., Harland L.M., Bar&t G.J. Evidence that the pertussis toxin-sensitive trimeric GTP-binding protein G,* is required for agonist- and store- activated Ca2+ inflow in hepatocytes. J Biol Chem 1995; 270: 25893-25897. Macrez-Lepretre N., Kalkbrenner F., Schultz G., Mironneau J, Distinct functions of G4 and G,, proteins in coupling a,- adrenoreceptors to Ca*+ release and Ca2+ entry in rat portal vein myocytes. JBiol Chem 1997; 272: 5261-5268. Xu X., Kitamura K., Lau KS., Muallem S., Miller R.T. Differential regulation of Ca*+ release-activated Ca2+ influx by hetero- trimeric G proteins. / Biol Chem 1995; 270: 29169-29175. Dippel E., Kalkbremrer E, Wittig B., Schultz G. A heterotrimeric G protein complex couples the muscaxinic ml receptor to phospho- lipase C-beta. Pmc NatlAcadSci USA 1996; 93: 1391-1396. Barritt G.J., Gregory R.B. An evaluation of strategies available for the identification of GTP-binding proteins required in intracellular signalling pathways. Cell Si’aI 1997; 9: 207-2 18. Komwatana P., Dinudom A., Young J.A., Cook D.I. Cytosolic Na+ controls an epithelial Na+ channel via the G, guanine nucleotide-binding regulatory protein. Proc Nat1 Acad Sci USA 1996; 93: 8107-8111. Zeng W., Xu X., Muallem S. GBy transduces [CaZ+], oscillations and Gaq a sustained response during stimulation of pancreatic acinar cells with [Caz+];mobilizing agonists, / Biol Chem 1996; 271: 18520-18526. Kalinec G., Nazarali AJ., Hermouet S., Xu N., Gutkind J.S. Mutated a subunit of the G4 protein induces malignant transformation in NIH 3T3 cells. Mel Cell Bioll992; 12: 4687-4693. Gordeladze J.O., Hovik K.E., Merendino J.J., Hermouet S., Gutkind S., Accili D. Effect of activating and inactivating mutations of G,- and G,-a protein subunits on growth and differentiation of 3T3- Ll preadipocytes. / Cell Biochem 1997; 64: 242-257 Hermouet S., Aznavoorian S., Spiegel A.M. In vitro and in vivo growth inhibition of murine melanoma K-l 735 cell by a dominant negative mutant a subunit of the G1, protein. Cell Signal 1996; 8: 159-166. Corre I., Hermouet S. Regulation of colony-stimulating factor l- induced proliferation by heterotrimeric G, proteins. Blood 1995; 86: 1776-l 783. Mizushima S., Nagata S. pEF-BOS, a powerful mammalian expression vector. Nucleic Acids Res 1990; 18: 5322. Graham F.L., Prevec L. Manipulation of adenovirus vectors, In: Murray E.J. (ed) Gene Transfer and Expression Protucols, vol. 7 Clifton NJ: Humana, 199 1; 109- 128. Sato M., Yoshida H., Yanagawa T., Yura Y., Urata M. Sensitivity of a neoplastic epithelial duct cell line from a human submandibular salivary gland to human leukocyte interferon as assessed by an in vitro semi-solid agar technique. IntJ Oral surg 1982; 11: 183-189.
19. Mittereder N., March K.L., Trapnell B.C. Evaluation of the concentration and bioactivity of adenovirus vectors for gene therapy. J viral 1996; 70: 7498-7509.
20. Gibb CA., Poronnik P., Day M.L., Cook D.I. Control of cytosolic pH in two-cell mouse embryos: roles of H(‘)-lactate cotransport and Na+/H+ exchange. Am J Physiol1997; 273: C404-C4 19.
21. He X.J., Wu X.Z., Wellner R.B., Baum BJ. Muscarinic receptor regulation of Ca2+ mobilization in a human salivary cell line. @&en Arch 1989; 413: 505-510.
22. Nitschke R., Leipziger J., Greger R. Agonist-induced intracellular Ca2+ transients in HT29 cells. PJlzigeevs Arch 1993; 423: 5 19-526.
23. Kopp R., Mayer P., Pfeiffer A. Agonist-induced desensitization of cholinergically stimulated phosphoinositide breakdown is independent of endogenously activated protein kinase C in HT- 29 human colon carcinoma cells. BiochemJ 1990; 269: 73-78.
24. Vanderheyden P., Gies J.P., Ebinger G., Keyser J.D., Landry Y., Vauquelin G. Human Ml-, M2- and MS-muscarinic cholinergic receptors: binding characteristics of agonists and antagonists, J Neural Sci 1990; 97: 67-80.
25. Sawaki K., Hiramatsu Y., Baum B.J., Ambudkar I.S. Involvement of G aqjll in m3-muscarinic receptor stimulation of phosphatidylinositol4,5 bisphosphate-specific phospholipase C in rat parotid gland membranes. Arch Biochem Biophys 1993; 305: 546-550.
26. Delporte C., O’Connell B.C., He X., Ambudkar IS., Agre P., Baum B J. Adenovirus-mediated expression of aquaporin-5 in epithelial cells. J Biol Chem 1996; 271: 22070-22075.
27 Delporte C., O’Connell B.C., He X., Lancaster H.E., O’Connell AC., Agre P., Baum BJ. Increased fluid secretion after adenovim- mediated transfer of the aquaporin-1 cDNA to irradiated rat salivary glands. Proc Nat1 Acad Sci USA 1997; 94: 3268-3273.
28. Akhter S.A., Skaer C.A., Kypson A.P. et al. Restoration of B- adrenergic signaling in failing cardiac ventricular myocytes via adenoviral-mediated gene transfer. Proc Natl Acad Sci USA 1997; 94: 12100-12105.
29. Rembold CM., Kendall J.M., Campbell A.K. Measurement of changes in sarcoplasmic reticulum [Ca2+] in rat tail artery with targeted apoaequorin delivered by an adenoviral vector. Cell Calcium 1997; 21: 69-79.
30. Caste11 J.V., Hernandez D., Gomez-Foix A.M., Guillen I., Donato T., Gomez-Lechon MJ. Adenovirus-mediated gene transfer into human hepatocytes: analysis of the biochemical functionality of transduced cells. Gene Ther 1997; 4: 455-464.
3 1. Drazner M.H., Peppel KC., Dyer S., Grant A.O., Koch WJ., Lelkowitz RJ. Potentiation of 8-adrenergic signaling by adenoviral-mediated gene transfer in adult rabbit ventricular myocytes. / Clin Invest 1997; 99: 288-296.
32. Kitamura K., Singer W.D., Cano A., Miller R.T. Ga, and Ga,, regulate NHE-1 and intracellular calcium in epithelial cells. Am JPhysioZ 1995; 268: ClOl-CllO.
33. Hermouet S., Murakami T., Spiegel A.M. Stable changes in expression or activation of G protein a, or a4 subunits affect the expression of both Bl and B2 subunits. FEBS Lett 1993; 327: 183-188.
0 Harcout? Brace & Co. Lfd 1998 Cell Calcium (1998) 24(2), 97-103




















