
Unveiling fungal zooflagellates as members of freshwater picoeukaryotes: evidence from a molecular...
11
Unveiling fungal zooflagellates as members of freshwater picoeukaryotes: evidence from a molecular diversity study in a deep meromictic lake Emilie Lefèvre, 1 Corinne Bardot, 1 Christophe Noël, 2 Jean-François Carrias, 1 Eric Viscogliosi, 3 Christian Amblard 1 and Télesphore Sime-Ngando 1 * 1 Laboratoire de Biologie des Protistes, UMR CNRS 6023, Université Blaise Pascal (Clermont-Ferrand II), F-63177, Aubière Cedex, France. 2 Institute for Research on Environment and Sustainability, University of Newcastle upon Tyne, NE1 7RU, UK. 3 Institut Pasteur de Lille, BP 245, F-59019, Lille Cedex, France. Summary This study presents an original 18S rRNA PCR survey of the freshwater picoeukaryote community, and was designed to detect unidentified heterotrophic picoflagellates (size range 0.6–5 mm) which are preva- lent throughout the year within the heterotrophic flagellate assemblage in Lake Pavin. Four clone libraries were constructed from samples collected in two contrasting zones in the lake. Computerized sta- tistic tools have suggested that sequence retrieval was representative of the in situ picoplankton diversity. The two sampling zones exhibited similar diversity patterns but shared only about 5% of the operational taxonomic units (OTUs). Phylogenetic analysis clustered our sequences into three taxo- nomic groups: Alveolates (30% of OTUs), Fungi (23%) and Cercozoa (19%). Fungi thus substantially contrib- uted to the detected diversity, as was additionally supported by direct microscopic observations of fungal zoospores and sporangia. A large fraction of the sequences belonged to parasites, including Alveolate sequences affiliated to the genus Perkinsus known as zooparasites, and chytrids that include host-specific parasitic fungi of various freshwater phytoplankton species, primarily diatoms. Phyloge- netic analysis revealed five novel clades that prob- ably include typical freshwater environmental sequences. Overall, from the unsuspected fungal diversity unveiled, we think that fungal zooflagellates have been misidentified as phagotrophic nanoflagel- lates in previous studies. This is in agreement with a recent experimental demonstration that zoospore- producing fungi and parasitic activity may play an important role in aquatic food webs. Introduction Heterotrophic flagellates (HF) are important trophic links in pelagic ecosystems. They are considered to be the primary grazers of bacteria and picoplankton (Fenchel, 1986; Fukami et al., 1996), and represent a prey source for larger microbial grazers and metazoa (Arndt, 1993; Premke and Arndt, 2000). They also are potential sources of nutrients and dissolved organic carbon, which are regenerated as a result of their grazing activities (Eccleston-Parry and Leadbeater, 1995). Heterotrophic flagellates typically occur at concentra- tions of about 10 3 -10 4 cells ml -1 in pelagic ecosystems (Jürgens and Stolpe, 1995; Fukami et al., 1996; Carrias et al., 1998). Since the emergence of the concept of the microbial loop (Azam et al., 1983), the in situ dynamics and grazing activity of aquatic HF have been exclusively investigated by epifluorescence microcopy (Fukami et al., 1996; Carrias et al., 1998; Sime-Ngando et al., 1999). This procedure does not allow an identification of HF at the species-level while although this community belongs to very different phylogenetic groups (Patterson, 1994). Only the largest cells (> 5 mm) belonging to HF can be tentatively identified with this technique. In contrast, the smallest cells (< 5 mm) are usually classified as ‘small unidentified’ due to the lack of conspicuous ultrastructural features (Carrias et al., 1998; Arndt et al., 2000; Cleven and Weisse, 2001). A recent approach that enables simultaneous identifi- cation and counting of specific microorganism taxa without culture is in situ hybridization of whole cells with rRNA-targeted oligonucleotide probes (Lim et al., 1993; 1996; 1999; Amann et al., 1995; Lim, 1996). In order to design probes for specific taxonomic groups, information about their phylogeny is necessary. Over the past several years, the use of 18S rDNA sequence analysis has Received 28 June, 2006; accepted 10 July, 2006. *For corre- spondence. E-mail [email protected]; Tel. (+33) 473 40 7836; Fax (+33) 473 40 7670. Environmental Microbiology (2007) 9(1), 61–71 doi:10.1111/j.1462-2920.2006.01111.x © 2006 The Authors Journal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd
-
Upload
univ-bpclermont -
Category
Documents
-
view
0 -
download
0
Transcript of Unveiling fungal zooflagellates as members of freshwater picoeukaryotes: evidence from a molecular...

Unveiling fungal zooflagellates as members offreshwater picoeukaryotes: evidence from a moleculardiversity study in a deep meromictic lake
Emilie Lefèvre,1 Corinne Bardot,1 Christophe Noël,2
Jean-François Carrias,1 Eric Viscogliosi,3
Christian Amblard1 and Télesphore Sime-Ngando1*1Laboratoire de Biologie des Protistes, UMR CNRS6023, Université Blaise Pascal (Clermont-Ferrand II),F-63177, Aubière Cedex, France.2Institute for Research on Environment andSustainability, University of Newcastle upon Tyne,NE1 7RU, UK.3Institut Pasteur de Lille, BP 245, F-59019, Lille Cedex,France.
Summary
This study presents an original 18S rRNA PCR surveyof the freshwater picoeukaryote community, andwas designed to detect unidentified heterotrophicpicoflagellates (size range 0.6–5 mm) which are preva-lent throughout the year within the heterotrophicflagellate assemblage in Lake Pavin. Four clonelibraries were constructed from samples collected intwo contrasting zones in the lake. Computerized sta-tistic tools have suggested that sequence retrievalwas representative of the in situ picoplanktondiversity. The two sampling zones exhibited similardiversity patterns but shared only about 5% of theoperational taxonomic units (OTUs). Phylogeneticanalysis clustered our sequences into three taxo-nomic groups: Alveolates (30% of OTUs), Fungi (23%)and Cercozoa (19%). Fungi thus substantially contrib-uted to the detected diversity, as was additionallysupported by direct microscopic observations offungal zoospores and sporangia. A large fraction ofthe sequences belonged to parasites, includingAlveolate sequences affiliated to the genus Perkinsusknown as zooparasites, and chytrids that includehost-specific parasitic fungi of various freshwaterphytoplankton species, primarily diatoms. Phyloge-netic analysis revealed five novel clades that prob-ably include typical freshwater environmental
sequences. Overall, from the unsuspected fungaldiversity unveiled, we think that fungal zooflagellateshave been misidentified as phagotrophic nanoflagel-lates in previous studies. This is in agreement with arecent experimental demonstration that zoospore-producing fungi and parasitic activity may play animportant role in aquatic food webs.
Introduction
Heterotrophic flagellates (HF) are important trophic linksin pelagic ecosystems. They are considered to be theprimary grazers of bacteria and picoplankton (Fenchel,1986; Fukami et al., 1996), and represent a prey sourcefor larger microbial grazers and metazoa (Arndt, 1993;Premke and Arndt, 2000). They also are potential sourcesof nutrients and dissolved organic carbon, which areregenerated as a result of their grazing activities(Eccleston-Parry and Leadbeater, 1995).
Heterotrophic flagellates typically occur at concentra-tions of about 103-104 cells ml-1 in pelagic ecosystems(Jürgens and Stolpe, 1995; Fukami et al., 1996; Carriaset al., 1998). Since the emergence of the concept of themicrobial loop (Azam et al., 1983), the in situ dynamicsand grazing activity of aquatic HF have been exclusivelyinvestigated by epifluorescence microcopy (Fukami et al.,1996; Carrias et al., 1998; Sime-Ngando et al., 1999).This procedure does not allow an identification of HF atthe species-level while although this community belongsto very different phylogenetic groups (Patterson, 1994).Only the largest cells (> 5 mm) belonging to HF can betentatively identified with this technique. In contrast, thesmallest cells (< 5 mm) are usually classified as ‘smallunidentified’ due to the lack of conspicuous ultrastructuralfeatures (Carrias et al., 1998; Arndt et al., 2000; Clevenand Weisse, 2001).
A recent approach that enables simultaneous identifi-cation and counting of specific microorganism taxawithout culture is in situ hybridization of whole cells withrRNA-targeted oligonucleotide probes (Lim et al., 1993;1996; 1999; Amann et al., 1995; Lim, 1996). In order todesign probes for specific taxonomic groups, informationabout their phylogeny is necessary. Over the past severalyears, the use of 18S rDNA sequence analysis has
Received 28 June, 2006; accepted 10 July, 2006. *For corre-spondence. E-mail [email protected];Tel. (+33) 473 40 7836; Fax (+33) 473 40 7670.
Environmental Microbiology (2007) 9(1), 61–71 doi:10.1111/j.1462-2920.2006.01111.x
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd
allowed many unexpected evolutionary lineages to bediscovered in natural ecosystems (Moreira and Lopez-Garcia, 2002; Berney et al., 2004). Most of these molecu-lar phylogenetic surveys for eukaryotic microorganismswere conducted in marine ecosystems (Diez et al., 2001;Lopez-Garcia et al., 2001; Moon-van der Staay et al.,2001; Massana et al., 2004; Romari and Vaulot, 2004;Savin et al., 2004; Yuan et al., 2004) and, more recently,in some extreme ecosystems, including anoxic (Dawsonand Pace, 2002; Stoeck and Epstein, 2003; Stoeck et al.,2003; Takishita et al., 2005), acidic and iron-rich environ-ments (Amaral-Zettler et al., 2002), as well as hydrother-mal vents (Edgcomb et al., 2002; Lopez-Garcia et al.,2003) and Antarctic soils (Lawley et al., 2004). Some ofthese studies have revealed that, in marine environments,small HF mostly belong to the stramenopile lineage, i.e.Heterokonta (Massana et al., 2002; 2004). However,despite the increasing number of molecular studies, thediversity of small eukaryotes in lakes is poorly described.To the best of our knowledge, only one molecular studyhas been conducted recently in the dimictic oligome-sotrophic Lake George (New York, USA) by Richards andcolleagues (2005). These authors found that three phyla(Stramenopile, Cryptomonads and Alveolates) dominatedthe detected diversity in summer, and provided evidencethat aspects of eukaryote microbial diversity are specificto certain aquatic environments. Their findings partiallycorroborate microscope studies of lake ecosystems thathave indicated that Stramenopile, Katablepharids andChoanozoa account for more than 90% of the observed
diversity within the small flagellate assemblages (Arndtet al., 2000).
Overall, the diversity, the distribution and the composi-tion of the tiniest members of HF community in freshwaterecosystems remain largely unknown. The aim of thisstudy was to examine the diversity of small eukaryoticcells in the deep meromictic Lake Pavin, using ribosomalRNA gene sequences. Samples were collected from dif-ferent layers (i.e. mixolimnion and oxycline) and during aperiod (i.e. autumn) of high abundances of the so-called‘unidentified small flagellates’ in the study site (Carriaset al., 1998; Colombet et al., 2006). These unknownflagellates, together with Monas-like cells, accounted onaverage for 70% of the total seasonal abundance of HF,and consisted mostly of round-shaped cells with a size< 5 mm (Carrias et al., 1996; 1998).
Results and discussion
Temperature, dissolved oxygen and chlorophyll aconcentrations
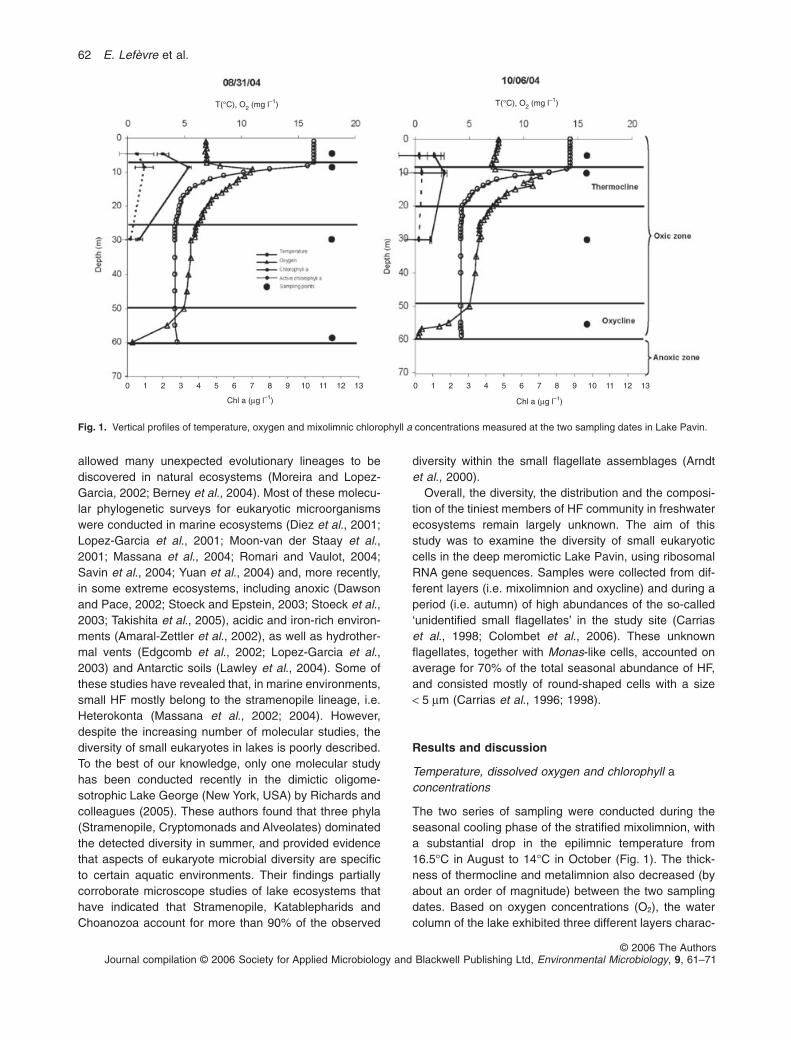
The two series of sampling were conducted during theseasonal cooling phase of the stratified mixolimnion, witha substantial drop in the epilimnic temperature from16.5°C in August to 14°C in October (Fig. 1). The thick-ness of thermocline and metalimnion also decreased (byabout an order of magnitude) between the two samplingdates. Based on oxygen concentrations (O2), the watercolumn of the lake exhibited three different layers charac-
T(°C), O2 (mg l–1) T(°C), O2 (mg l–1)
Chl a (μg l–1) Chl a (μg l–1)
0 1 2 3 4 5 6 7 8 9 10 11 12 13 0 1 2 3 4 5 6 7 8 9 10 11 12 13
Fig. 1. Vertical profiles of temperature, oxygen and mixolimnic chlorophyll a concentrations measured at the two sampling dates in Lake Pavin.
62 E. Lefèvre et al.
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71
terized as oxic (O2 > 6 mg l-1), suboxic (O2 between0.1 and 6 mg l-1), and anoxic, corresponding to mixolim-nion (0–50 m), oxycline (50–60 m) and monimolimnion(> 60 m) respectively. Chlorophyll a concentrations in themixolimnion peaked in the metalimnion together with O2
(Fig. 1), as a consequence of enhanced photosyntheticactivity.
Overall diversity estimates
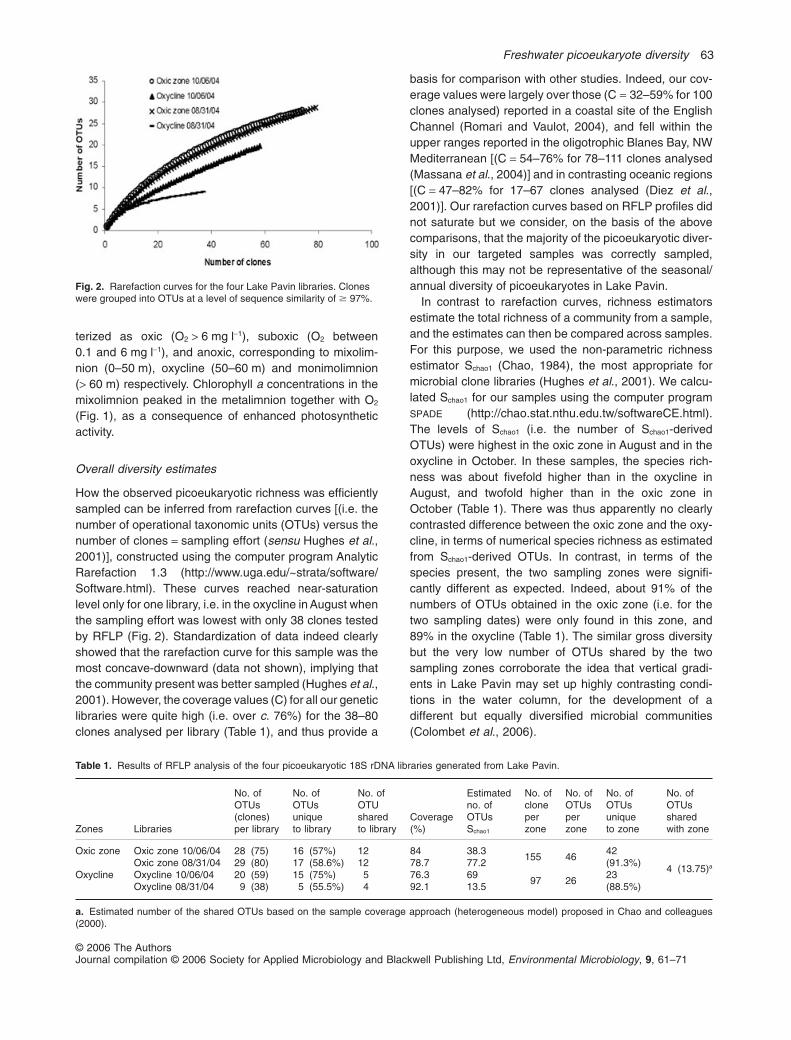
How the observed picoeukaryotic richness was efficientlysampled can be inferred from rarefaction curves [(i.e. thenumber of operational taxonomic units (OTUs) versus thenumber of clones = sampling effort (sensu Hughes et al.,2001)], constructed using the computer program AnalyticRarefaction 1.3 (http://www.uga.edu/~strata/software/Software.html). These curves reached near-saturationlevel only for one library, i.e. in the oxycline in August whenthe sampling effort was lowest with only 38 clones testedby RFLP (Fig. 2). Standardization of data indeed clearlyshowed that the rarefaction curve for this sample was themost concave-downward (data not shown), implying thatthe community present was better sampled (Hughes et al.,2001). However, the coverage values (C) for all our geneticlibraries were quite high (i.e. over c. 76%) for the 38–80clones analysed per library (Table 1), and thus provide a
basis for comparison with other studies. Indeed, our cov-erage values were largely over those (C = 32–59% for 100clones analysed) reported in a coastal site of the EnglishChannel (Romari and Vaulot, 2004), and fell within theupper ranges reported in the oligotrophic Blanes Bay, NWMediterranean [(C = 54–76% for 78–111 clones analysed(Massana et al., 2004)] and in contrasting oceanic regions[(C = 47–82% for 17–67 clones analysed (Diez et al.,2001)]. Our rarefaction curves based on RFLP profiles didnot saturate but we consider, on the basis of the abovecomparisons, that the majority of the picoeukaryotic diver-sity in our targeted samples was correctly sampled,although this may not be representative of the seasonal/annual diversity of picoeukaryotes in Lake Pavin.
In contrast to rarefaction curves, richness estimatorsestimate the total richness of a community from a sample,and the estimates can then be compared across samples.For this purpose, we used the non-parametric richnessestimator Schao1 (Chao, 1984), the most appropriate formicrobial clone libraries (Hughes et al., 2001). We calcu-lated Schao1 for our samples using the computer programSPADE (http://chao.stat.nthu.edu.tw/softwareCE.html).The levels of Schao1 (i.e. the number of Schao1-derivedOTUs) were highest in the oxic zone in August and in theoxycline in October. In these samples, the species rich-ness was about fivefold higher than in the oxycline inAugust, and twofold higher than in the oxic zone inOctober (Table 1). There was thus apparently no clearlycontrasted difference between the oxic zone and the oxy-cline, in terms of numerical species richness as estimatedfrom Schao1-derived OTUs. In contrast, in terms of thespecies present, the two sampling zones were signifi-cantly different as expected. Indeed, about 91% of thenumbers of OTUs obtained in the oxic zone (i.e. for thetwo sampling dates) were only found in this zone, and89% in the oxycline (Table 1). The similar gross diversitybut the very low number of OTUs shared by the twosampling zones corroborate the idea that vertical gradi-ents in Lake Pavin may set up highly contrasting condi-tions in the water column, for the development of adifferent but equally diversified microbial communities(Colombet et al., 2006).
Fig. 2. Rarefaction curves for the four Lake Pavin libraries. Cloneswere grouped into OTUs at a level of sequence similarity of � 97%.
Table 1. Results of RFLP analysis of the four picoeukaryotic 18S rDNA libraries generated from Lake Pavin.
Zones Libraries
No. ofOTUs(clones)per library
No. ofOTUsuniqueto library
No. ofOTUsharedto library
Coverage(%)
Estimatedno. ofOTUsSchao1
No. ofcloneperzone
No. ofOTUsperzone
No. ofOTUsuniqueto zone
No. ofOTUssharedwith zone
Oxic zone Oxic zone 10/06/04 28 (75) 16 (57%) 12 84 38.3155 46
42(91.3%)
4 (13.75)aOxic zone 08/31/04 29 (80) 17 (58.6%) 12 78.7 77.2Oxycline Oxycline 10/06/04 20 (59) 15 (75%) 5 76.3 69
97 2623(88.5%)Oxycline 08/31/04 9 (38) 5 (55.5%) 4 92.1 13.5
a. Estimated number of the shared OTUs based on the sample coverage approach (heterogeneous model) proposed in Chao and colleagues(2000).
Freshwater picoeukaryote diversity 63
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71
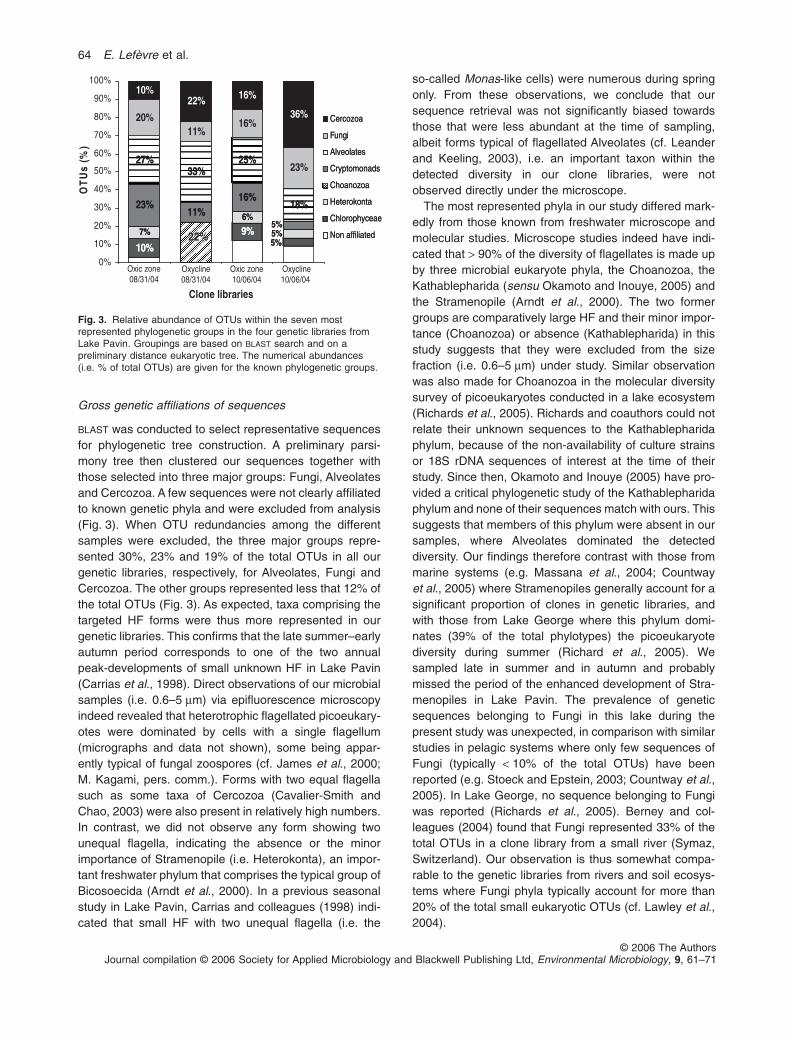
Gross genetic affiliations of sequences
BLAST was conducted to select representative sequencesfor phylogenetic tree construction. A preliminary parsi-mony tree then clustered our sequences together withthose selected into three major groups: Fungi, Alveolatesand Cercozoa. A few sequences were not clearly affiliatedto known genetic phyla and were excluded from analysis(Fig. 3). When OTU redundancies among the differentsamples were excluded, the three major groups repre-sented 30%, 23% and 19% of the total OTUs in all ourgenetic libraries, respectively, for Alveolates, Fungi andCercozoa. The other groups represented less that 12% ofthe total OTUs (Fig. 3). As expected, taxa comprising thetargeted HF forms were thus more represented in ourgenetic libraries. This confirms that the late summer–earlyautumn period corresponds to one of the two annualpeak-developments of small unknown HF in Lake Pavin(Carrias et al., 1998). Direct observations of our microbialsamples (i.e. 0.6–5 mm) via epifluorescence microscopyindeed revealed that heterotrophic flagellated picoeukary-otes were dominated by cells with a single flagellum(micrographs and data not shown), some being appar-ently typical of fungal zoospores (cf. James et al., 2000;M. Kagami, pers. comm.). Forms with two equal flagellasuch as some taxa of Cercozoa (Cavalier-Smith andChao, 2003) were also present in relatively high numbers.In contrast, we did not observe any form showing twounequal flagella, indicating the absence or the minorimportance of Stramenopile (i.e. Heterokonta), an impor-tant freshwater phylum that comprises the typical group ofBicosoecida (Arndt et al., 2000). In a previous seasonalstudy in Lake Pavin, Carrias and colleagues (1998) indi-cated that small HF with two unequal flagella (i.e. the
so-called Monas-like cells) were numerous during springonly. From these observations, we conclude that oursequence retrieval was not significantly biased towardsthose that were less abundant at the time of sampling,albeit forms typical of flagellated Alveolates (cf. Leanderand Keeling, 2003), i.e. an important taxon within thedetected diversity in our clone libraries, were notobserved directly under the microscope.
The most represented phyla in our study differed mark-edly from those known from freshwater microscope andmolecular studies. Microscope studies indeed have indi-cated that > 90% of the diversity of flagellates is made upby three microbial eukaryote phyla, the Choanozoa, theKathablepharida (sensu Okamoto and Inouye, 2005) andthe Stramenopile (Arndt et al., 2000). The two formergroups are comparatively large HF and their minor impor-tance (Choanozoa) or absence (Kathablepharida) in thisstudy suggests that they were excluded from the sizefraction (i.e. 0.6–5 mm) under study. Similar observationwas also made for Choanozoa in the molecular diversitysurvey of picoeukaryotes conducted in a lake ecosystem(Richards et al., 2005). Richards and coauthors could notrelate their unknown sequences to the Kathablepharidaphylum, because of the non-availability of culture strainsor 18S rDNA sequences of interest at the time of theirstudy. Since then, Okamoto and Inouye (2005) have pro-vided a critical phylogenetic study of the Kathablepharidaphylum and none of their sequences match with ours. Thissuggests that members of this phylum were absent in oursamples, where Alveolates dominated the detecteddiversity. Our findings therefore contrast with those frommarine systems (e.g. Massana et al., 2004; Countwayet al., 2005) where Stramenopiles generally account for asignificant proportion of clones in genetic libraries, andwith those from Lake George where this phylum domi-nates (39% of the total phylotypes) the picoeukaryotediversity during summer (Richard et al., 2005). Wesampled late in summer and in autumn and probablymissed the period of the enhanced development of Stra-menopiles in Lake Pavin. The prevalence of geneticsequences belonging to Fungi in this lake during thepresent study was unexpected, in comparison with similarstudies in pelagic systems where only few sequences ofFungi (typically < 10% of the total OTUs) have beenreported (e.g. Stoeck and Epstein, 2003; Countway et al.,2005). In Lake George, no sequence belonging to Fungiwas reported (Richards et al., 2005). Berney and col-leagues (2004) found that Fungi represented 33% of thetotal OTUs in a clone library from a small river (Symaz,Switzerland). Our observation is thus somewhat compa-rable to the genetic libraries from rivers and soil ecosys-tems where Fungi phyla typically account for more than20% of the total small eukaryotic OTUs (cf. Lawley et al.,2004).
23%
27%33%
25%
18%
20%11%
16%
23%
10%22% 16%
36%
5%9%
10%5%
6%
7% 22%5%
11%16%
OT
Us
(%)
Cercozoa
Fungi
Alveolates
Cryptomonads
Choanozoa
Heterokonta
Chlorophyceae
Non affiliated
Oxic zone 08/31/04
Oxycline08/31/04
Oxycline10/06/04
Oxic zone 10/06/04
Clone libraries
23%
27%33%
25%
18%
20%11%
16%
23%
10%22% 16%
36%
5%9%
10%5%
6%
7% 22%5%
11%16%
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Cercozoa
Fungi
Alveolates
Cryptomonads
Choanozoa
Heterokonta
Chlorophyceae
Non affiliated
Fig. 3. Relative abundance of OTUs within the seven mostrepresented phylogenetic groups in the four genetic libraries fromLake Pavin. Groupings are based on BLAST search and on apreliminary distance eukaryotic tree. The numerical abundances(i.e. % of total OTUs) are given for the known phylogenetic groups.
64 E. Lefèvre et al.
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71

During this study, the spatio-temporal changes in therelative importance of the OTUs from the known taxa werecharacterized by: (i) the absence of genetic groups con-taining a majority of autotrophic forms (i.e. Chlorophyceaeand Cryptophyceae) in the oxycline; and (ii) a similardistribution pattern in the oxic zone where Alveolatesdominated the total diversity, followed by Fungi or Crypto-monads, Cercozoa, Chlorophyceae and Heterokonta(Fig. 3). In the oxycline, Choanozoa were present inAugust but not in October, while the dominant Alveolates inAugust were partly replaced by Cercozoa, the dominantgroup in October. The dynamics of phylotypes in the oxy-cline was thus different from that in the oxic zone, inagreement with previous indications on the significance ofvertical gradients in shaping the community composition ofsmall eukaryotes in Lake Pavin (Colombet et al., 2006).
Phylogenetic tree topologies and ecological implications
The preliminary distance tree helped us to focus on themain phylogenetic lineages which are known to compriseheterotrophic flagellated forms, i.e. Alveolates, Cercozoaand Fungi. One of the main pelagic phyla containing smallHF (Heterokonts or Stramenopiles) was weakly repre-sented in our libraries, as discussed above. Indeed, onlyfour of our sequences were scored as Heterokonts, all ofwhich were branched to the heterotrophic basal groups(Massana et al., 2002), including the Bicosoecida andOomycete sequences (data not shown).
Alveolates. The clone libraries constructed yielded 21 dif-ferent Alveolate sequences (Fig. 4), of which 13 belongedto Ciliophora and one to Dinophyceae. Five additionalsequences formed a well-supported novel clade (i.e. cur-rently containing no cultured representatives) essentiallycomposed of sequences found in the oxycline libraries.This clade branched as a sister group to a genus (Perk-insus) known as a bivalve, microeukaryote and fish para-site that can produce free-motile biflagellate zoospores of2–5 mm in diameter (Burreson et al., 2005; Gestal et al.,2006). This genus is being discovered and described atan increasingly rapid rate, almost exclusively withsequence data (cf. Burreson et al., 2005). Our two otherAlveolate sequences, together with the clone LG32-03from Lake George (Richards et al., 2005), were related toColpodella edax, a free-living phagotrophic biflagellate(Leander et al., 2003). Species of the genus Colpodellaare bacterivores or predators of other free-living ‘protists’which, except for their free-living habits, are nearly indis-tinguishable from Perkinsus species based on cell mor-phology (Kuvardina et al., 2002).
Cercozoa. The Bayesian phylogenetic tree yielded 13Lake Pavin phylotypes that grouped with Cercozoa
sequences, with generally high bootstrap supports(Fig. 4). The sequences from the two sampling zones (i.e.oxic zone and oxycline) were generally well separated inthe tree topology. Among them, six grouped into threenovel clades. Two sequences from the Oxycline (October)formed the first novel clade that lacks clear affiliation withany known taxon. These sequences were closely affiliatedto two other sequences known from the dimictic olig-otrophic Lake Georges (Richards et al., 2005), suggestingthat they were typical of freshwater systems. The novelclade II comprised two of our sequences sampled inOctober in the oxic zone and the clone RT5iin4 found in ahigh acidity and heavy metal concentrated river(Amaral-Zettler et al., 2002), supporting former ideasabout the phylogenetic range and the relatively rapidadaptation of microbial eukaryotes in extreme environ-ments (Amaral-Zettler et al., 2002). BLAST results revealed97% of similarity between one of our novel clade IIsequences and the uncultured cercozoan clone4–6.2 (not included in the tree because of the short lengthof the sequence), actually considered as a member ofProteomyxidea, a Class of strict heterotrophic Cercozoa(Bass and Cavalier-Smith, 2004). These two novel cladesoccupy a relatively basal position inside the Cercozoa, i.e.both with Bayesian and Neighbour-joining (NJ) methods.Our environmental libraries have revealed a third novelclade that contains two of our sequences and the uncul-tured clone TAGIRI-3 from Takishita and colleagues(2005), in which no detailed phylogenetic position withinCercozoa is provided.
Six sequences affiliated to known taxa belongedto freshwater Orders, Cercomonadida and Tectofilosida,of which three were tightly affiliated to the genusCercomonas. Cercomonas species are small (10–15 mm)heterotrophic biflagellates which are very common inwater and soil habitats worldwide, where they are knownto play an essential role in natural food chains and bac-terial mortality, although their diversity and taxonomy arestill poorly known (review in Myl’nikov and Karpov, 2004).Our three other known Cercozoa sequences branchedwell with the genus Lecythium, a small (5–10 mm) fresh-water heterotrophic filose amoeba for which floatingspecimens, cysts, plasmodia or zoospores have not beenobserved. Pseudopodia are present constantly in thisgenera and serve both for the capture of prey and forlocomotion (Nikolaev et al., 2003).
Fungi. Of the 16 environmental sequences belonging toFungi, two were putatively identified as the Basidio-mycetous yeast species Rhodosporidium diobovatum andFilobasidium globisporum, and one close to theAscomycetous yeast Taphrina letifera (Fig. 4). The occur-rence of yeasts in lake ecosystems is common and isgenerally associated with allochthonous inputs from
Freshwater picoeukaryote diversity 65
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71
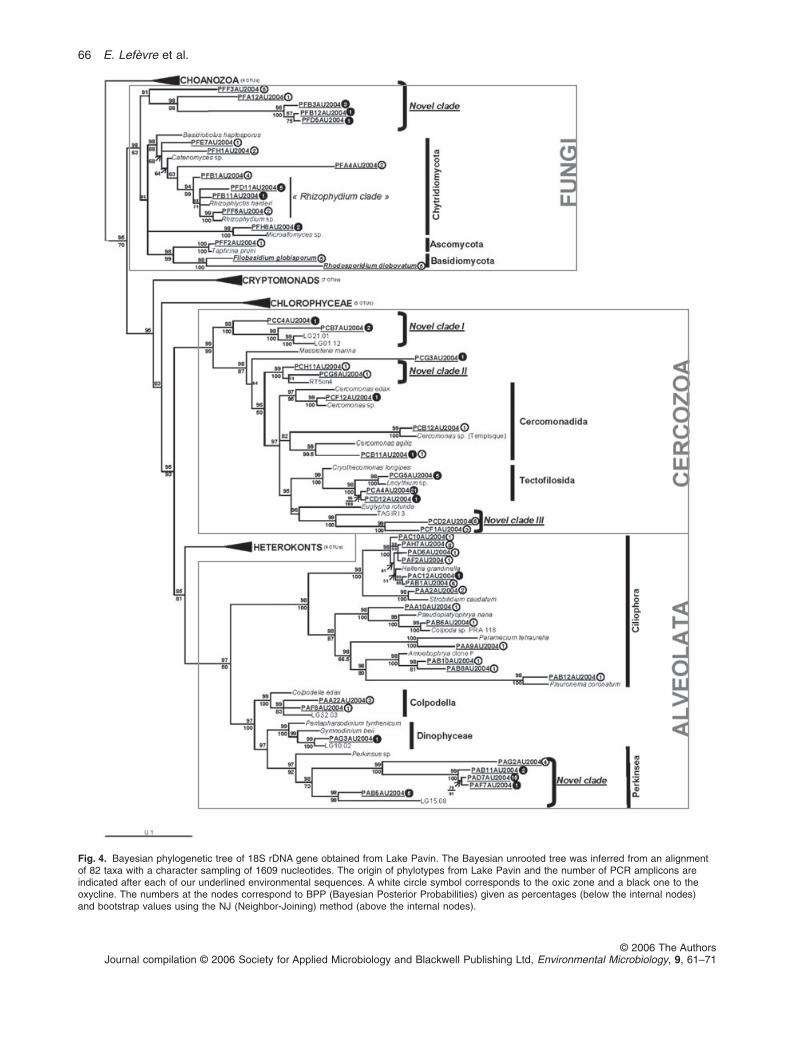
Fig. 4. Bayesian phylogenetic tree of 18S rDNA gene obtained from Lake Pavin. The Bayesian unrooted tree was inferred from an alignmentof 82 taxa with a character sampling of 1609 nucleotides. The origin of phylotypes from Lake Pavin and the number of PCR amplicons areindicated after each of our underlined environmental sequences. A white circle symbol corresponds to the oxic zone and a black one to theoxycline. The numbers at the nodes correspond to BPP (Bayesian Posterior Probabilities) given as percentages (below the internal nodes)and bootstrap values using the NJ (Neighbor-Joining) method (above the internal nodes).
66 E. Lefèvre et al.
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71

saprophytic associations with plant- or animal-derivedorganic compounds (Kurtzman and Fell, 2004). The restof our fungal sequences were unknown. Three of thesesequences showed no clear phylogenetic association withany known eukaryotic groups, and are thus likely to rep-resent previously unsampled groups of Fungi. The otherunknown phylotypes belonged to the Class of Chytridi-omycota (i.e. chytrids), reported as a paraphyletic group(James et al., 2000). This group of Fungi is characterizedby the production of motile reproductive cells that typicallyhave a single, posteriorly directed flagellum (cf. Jameset al., 2000), corresponding to one of the major formsobserved directly via epifluorescence microscopy in oursamples (microphotographs not shown). Indeed, the formof the flagellum is a pertinent identification key and formswith a straight and long single flagellum in our sampleswere tentatively identified as chytrid zoospores (M.Kagami, pers. comm.). All the members of Chytridiomy-cota are considered as ubiquitous of wet soils and fresh-waters, because their unwalled, flagellated zoosporesrequired water for dispersal. These typical microscopicFungi have received little attention by researchers,because only a few of them are of obvious economicimportance (Powell, 1993).
Among the Chytridiomycota sequences, four belongedto Chytridiales, one to Blastocladiales, and two otherscould not be affiliated to any known Order. Most of thediversity of the chytrids lies within the Chytridiales, butstudies on this group are severely limited by the lack ofadequate species descriptions (Shearer et al., 2004).Many Chytridiales have been recorded as parasites offreshwater algae (Gromov et al., 1999). Our Chytridialesequences mostly fell within the ‘Rhizophydium clade’ thatcontains host-specific parasitic fungi of various phy-toplankton species such as diatoms, and are known tooften reduce phytoplankton blooms and to play a crucialrole in phytoplankton succession (Kagami et al., 2004). Inaddition, the free-swimming zoospores of the reproduc-tive stage range in size from 2 to 5 mm (Bruning, 1991),and can represent a food source for zooplankton (Kagamiet al., 2004). The occurrence of these parasitic fungi in oursamples was confirmed from direct light microscopicobservation of their sporangia (i.e. the infective stage)fixed on the diatom Asterionella formosa (Fig. 5). Thesesporangia have probably been miscounted as bacterivo-rous flagellates in previous studies on attached protists inLake Pavin (Carrias et al., 1996; 1998). In addition, alarge part of the small flagellated zoospores certainly cor-respond to the small unidentified HF which were not ableto ingest bacterial food tracers during a seasonal studyconducted in this lake by Carrias and colleagues (1996).Large grazing-resistant diatoms, primarily Aulacoseiraitalica and Asterionella formosa, are important membersin Lake Pavin, where they typically accounted for 50–98%
of the total phytoplankton biomass or production duringthe spring bloom in the whole water column (Amblard andBourdier, 1990). We thus suspect that the parasitic activityof chytrids is an important pathway in Lake Pavin, at leastas one of the potential factors that regulate diatomblooms. This supports the recent experimental demon-stration that zoospore-producing fungi play an importantrole in aquatic food webs.
In addition to Chytridiales, five other chytrid sequencesin our libraries could not be affiliated with any knownsequences. They thus formed an important novel cladelocated at the basal position within the Fungi phylum withhigh bootstrap support.
Conclusions
Overall, this study presents an original 18S rDNA surveyof a lake picoeukaryote community in a site that offers aunique environment with low human influences and highannual reproducibility of seasonal dynamics in the watercolumn. The Bayesian method adopted for the construc-tion of the phylogenetic tree retrieves a number of rela-tionships consistent with hypothesis of eukaryoteevolutionary relationships. In silico statistic tools con-firmed that the targeted diversity was relatively wellsampled, with similar gross diversity but a very lownumber of shared phylotypes between two contrastingzones irrespective of oxygen concentrations in LakePavin. Among the targeted HF diversity, the dominantphyla found in this lake differed markedly from thoseknown from freshwater microscope and molecularstudies, partly because of differences in the sampling
Fig. 5. Picture of the diatom Asterionella formosa with fourattached sporangia of fungi. Inserted picture corresponds to anenlarged sporangium (2¥) for detail of the two typical identificationkeys: the peduncle (P) and the operculum (O).
Freshwater picoeukaryote diversity 67
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71

periods. Indeed, Fungi significantly contributed to thedetected diversity of targeted organisms, both frommolecular data and from direct microscopic observations,indicating that fungal zooflagellates may have been mis-counted as flagellate grazers in previous studies. Withinthe targeted organisms, a large proportion of oursequences (c. 60% of the total) belonged to organismsknown as potential parasites, indicating the possibleimportance of parasitoids in aquatic food webs. Finally, wecould not affiliate several sequences to any known taxo-nomic groups plausibly because they represent new orunsampled taxa, some of which may be typical of fresh-water ecosystems.
Experimental procedures
Study site and sampling
Lake Pavin is a dimictic and oligomesotrophic lake withpartial overturns (i.e. meromixis) situated in the MassifCentral of France (longitude 2°56′E, latitude 45°N). It is adeep (Zmax = 92 m) volcanic mountain lake characterized bya small surface area (44 ¥ 104 m2), a low drainage basin area(50 ¥ 104 m2) and a relatively small human influences. Thissite offers a unique environment with high annual reproduc-ibility of seasonal dynamics in the water column. Sampleswere collected at two different dates during thermal stratifi-cation (31st August 2004 and 6th October 2004), using a 8-LVan Dorn bottle. Sampling depths were at 5 m (epilimnion),9–10 m (metalimnion) and 30 m (hypolimnion) for the oxiclayer of the lake, and at 59–60 m for the oxycline (oxygenconcentration < 0.8 mg l-1). Samples were transferred intoacid clean 10 l plastic carboys previously rinsed thoroughlywith the lake water, and transported within an hour to thelaboratory for immediate processing. Vertical profiles oftemperature and dissolved oxygen were obtained usingan Oxycal-SL 197 multiparameter probe (WWT, Limonest,France). In the laboratory, planktonic organisms of > 5 mm-size were removed from the water by a succession of gravityprefiltrations through 150, 80, 50, 25, 10 and 5 mm pore-sizefilters. Small cells of interest (< 5 mm in size) were collectedon 0.6 mm pore-size polycarbonate filters (25 mm of diam-eter) using a vacuum pump (pressure < 100 mbar). One totwo hundred millilitres of water was filtered until saturation ofthe filter. Then, filters were placed into 2 ml tubes and storedat -80°C until DNA extraction.
Nucleic acid extraction
Filters were treated with 560 ml of Tris-EDTA buffer (TE, 1 mMEDTA and 10 mM Tris-HCl); then, SDS (sodium dodecylsulfate, 1% final concentration) and proteinase K (0.1 mg ml-1
final concentration) were added during 1 h at 37°C for proteindigestion. In order to remove cell wall debris, polysaccha-rides, and the remaining proteins, CTAB (cetyltrimethylam-monium bromide)/NaCl pre-warmed at 65°C and NaClsolution were added (final concentration of 1%/0.07 M and0.7 M respectively) and the mixture incubated at 65°C for10 min. A solution of phenol, chloroform and isoamyl-alcohol
(25:24:1) was then added for nucleic acid extraction. Nucleicacids were precipitated from the aqueous solution with iso-propanol (0.6 volume) at -20°C over night, washed withethanol 70% (stored at -20°C), dried, and suspended in 50 mlTE buffer. The integrity of the native DNA was checked byagarose gel electrophoresis (0.8%) before storage at -20°Cuntil further analysis.
PCR for cloning and sequencing
The complete eukaryotic 18S rRNA gene was amplified fromenvironmental genomic DNA extract using universal eukary-otic primers, 1f (5′-CTGGTTGATCCTGCCAG-3′) and 1520r(5′-CTGCAGGTTCACCTAC-3′). The PCR mixture (50 ml)contained 50 ng of environmental DNA as template, 200 mMof each dNTP, 2 mM of MgCl2, 10 pM of each primer, 2.5 unitsof HotStar Taq DNA polymerase (Qiagen), and the PCRbuffer supplied with the enzyme. The reaction was carried outin an automated thermocycler (PTC 200 MJ Research) withthe following cycle: an initial hot start incubation (15 min at95°C) followed by 29 identical cycles (denaturation at 95°Cfor 1 min, annealing at 57°C for 1 min, and extension at 72°Cfor 1.5 min), and a final extension at 72°C for 10 min.
Clone library construction and sequence analysis
For each sampling date, two genetic libraries were con-structed: one for the oxic zone (obtained by pooling individualPCR products from the three oxic depths sampled), and onefor the oxycline, i.e. a total of four libraries. An aliquot of PCRproducts was cloned using the TOPO-TA cloning kit (Invitro-gen) following the manufacturer’s recommendations. Puta-tive positive colonies were picked and stored for analysis.The presence of the 18S rDNA insert in the colonies waschecked by PCR amplification with the M13 universalprimers. PCR amplification products of the correct size weredigested by the restriction enzyme HaeIII (Invitrogen) for 12 hat 37°C. The digested products were separated by electro-phoresis at 100 V for 4 h in a 2.5% agarose gel. Clonesshowing the same RFLP patterns were grouped and consid-ered to belong to the same OTU.
One representative of each OTU was sequenced fromplasmid products by the MWG Biotech services using theM13 universal primers [M13rev (-29) and M13 uni (-21)].Sequences were submitted to a BLAST search (Altschul et al.,1997) for an approximate phylogenetic affiliation and in orderto select representative taxa for tree construction. A prelimi-nary distance tree was then established using the PHYLIP
package (http://evolution.gs.washington.edu/phylip for anaccurate affiliation of our sequences. This tree containednearly 200 eukaryotic sequences (including about 100 repre-sentatives of all major lineages) that were aligned usingthe alignment tool of the ARB package (http://www.arb-home.de/). The resulting alignment was corrected manuallyand areas of ambiguous alignment were excluded usingBioedit software (http://www.mbio.ncsu.edu/BioEdit/bioedit.html). The preliminary tree highlighted the main groups thatwere emphasized in the final tree, for which sequence align-ments were analysed both by NJ method using the PHYLIP
package and by the Bayesian method using MrBayes3 soft-
68 E. Lefèvre et al.
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71

ware (http://prdownloads.sourceforge.net/mrbayes/mrbayes-3.1.2.zip?download) (Ronquist and Huelsenbeck, 2003).Topologies of the phylogenetic trees generated by the twomethods were fairly similar; therefore, only the tree generatedby the Bayesian method will be shown. This method is moreefficient in recovering accurate support values, i.e. requirefewer data to converge on the correct answer relative to theNJ non-parametric bootstrap (cf. Alfaro et al., 2003). TheMrBayes settings were as follows: (i) the Maximum-Likelihood model employed six substitution types, with basefrequencies estimated from the data; (ii) rate variation acrosssites was modelled using a gamma-shaped rate variationwith a proportion of invariable sites; (iii) the Monte CarloMarkov chain search was run with four chains for 1 000 000generations, with tree being sampled every 1000 genera-tions; and (iv) first trees, with no steady log likelihood values,were discarded to create the consensus tree. Bayesian pos-terior probabilities (BPP) were calculated using a Markovchain Monte Carlo sampling approach (Green, 1995) imple-mented in MrBayes3. Nucleotide sequences obtained in thisstudy have been deposited in GenBank under AccessionNumbers DQ243989–DQ244041.
Acknowledgements
We thank Dr M. Embley for hosting E.L. in his laboratory fortraining and advice regarding phylogenetic method for theaffiliation of eukaryotes. We are grateful to Dr M. Kagami forcomments regarding micrographs of fungi, including that inFig .5, and to Dr R. Massana for advice at the beginning ofthis work. A. Bec, A.H. Lejeune and J.L. Vincenot are alsoacknowledged for image processing and analysing, and C.Portelli for field and technical assistance. E.L. was supportedby a doctoral fellowship from the French ‘Ministère de laRecherche et de la Technologie (MRT)’.
References
Alfaro, M.E., Zoller, S., and Lutzoni, F. (2003) Bayes orbootstrap? A simulation study comparing the performanceof Bayesian Markov chain Monte Carlo sampling and boot-strapping in assessing phylogenetic confidence. Mol BiolEvol 20: 255–266.
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J.,Zhang, Z., Miller, W., and Lipman, D.J. (1997) GappedBLAST and PSI-BLAST: a new generation of protein data-base search programs. Nucleic Acids Res 25: 3389–3402.
Amann, R.I., Ludwig, W., and Schleifer, K.H. (1995) Phyloge-netic identification and in situ detection of individual micro-bial cells without cultivation. Microbiol Rev 59: 143–169.
Amaral-Zettler, L.A., Gomez, F., Zettler, E., Keenan, B.G.,Amils, R., and Sogin, M.L. (2002) Eukaryotic diversity inSpain’s River of Fire. Nature 417: 137.
Amblard, C., and Bourdier, G. (1990) The spring bloom ofthe diatom Melosira italica ssp. subarctica. Lake Pavin:biochemical, energetic and metabolic aspects duringsedimentation. J Plankton Res 12: 645–660.
Arndt, H. (1993) Rotifers as predators on components of themicrobial web (bacteria, heterotrophic flagellates, ciliates)– a review. Hydrobiologia 255/256: 231–246.
Arndt, H., Dietrich, D., Auer, B., Cleven, E.J., Gräfenhan, T.,Weitere, M., and Myl’nikov, A.P. (2000) Functional diversityof heterotrophic flagellates in aquatic ecosystems. In TheFlagellates: Unity, Diversity and Evolution. Leadbeater,B.S.C., and Green, J.C. (eds). London, UK: Taylor &Francis, pp. 240–268.
Azam, F., Fenchel, T., Field, J.G., Gray, J.S., Meyer-Reil,L.A., and Thingstad, F. (1983) The ecological role of water-column microbes in the sea. Mar Ecol Prog Ser 10: 257–263.
Bass, D., and Cavalier-Smith, T. (2004) Phylum-specific envi-ronmental DNA analysis reveals remarkably high globalbiodiversity of Cercozoa (Protozoa). Int J Syst Evol Micro-biol 54: 2393–2404.
Berney, C., Fahrni, J., and Pawlowski, J. (2004) How manynovel eukaryotic ‘kingdoms’? Pitfalls and limitations ofenvironmental DNA surveys. BMC Biol 2: 1–13.
Bruning, K. (1991) Effects of temperature and light on thepopulation dynamics of the Asterionella–Rhizophydiumassociation. J Plankton Res 13: 707–719.
Burreson, E.M., Reece, K.S., and Dungan, C.F. (2005)Molecular, morphological, and experimental evidencesupport the synonymy of Perkinsus chesapeaki and Perk-insus andrewsi. J Eukaryot Microbiol 52: 258–270.
Carrias, J.F., Amblard, C., and Bourdier, G. (1996) Protistanbacterivory in an oligomesotrophic lake: importance ofattached ciliates and flagellates. Microb Ecol 31: 249–268.
Carrias, J.F., Quiblier-Lloberas, C., and Bourdier, G. (1998)Seasonal dynamics of free and attached heterotrophicnanoflagellates in an oligomesotrophic lake. Freshw Biol39: 91–101.
Cavalier-Smith, T., and Chao, E.E. (2003) Phylogeny andclassification of phylum Cercozoa (Protozoa). Protist 154:341–358.
Chao, A. (1984) Nonparametric estimation of the number ofclasses in a population. Scand J Stat 11: 265–270.
Chao, A., Hwang, W.H., Chen, Y.C., and Kuo, C.Y. (2000)Estimating the number of shared species in twocommunities. Statistica Sinica 10: 227–246.
Cleven, E.J., and Weisse, T. (2001) Seasonal successionand taxon-specific bacterial grazing rates of heterotrophicnanoflagellates in Lake Constance. Aquat Microb Ecol 23:147–161.
Colombet, J., Sime-Ngando, T., Cauchie, H.M., Fonty, G.,Hoffmann, L., and Demeure, G. (2006) Depth-related gra-dients of viral activity in Lake Pavin. Appl Environ Microbiol72: 4440–4445.
Countway, P.D., Gast, R.J., Savai, P., and Caron, D.A.(2005) Protistan diversity estimates based on 18S rDNAfrom seawater incubations in the western North Atlantic.J Eukaryot Microbiol 52: 95–106.
Dawson, S.C., and Pace, N.R. (2002) Novel Kingdom-leveleukaryotic diversity in anoxic environments. Proc Natl AcadSci USA 99: 8324–8329.
Diez, B., Pedros-Alio, C., and Massana, R. (2001) Study ofgenetic eukaryotic picoplankton in different oceanicregions by small-subunit rRNA gene cloning andsequencing. Appl Environ Microbiol 67: 2932–2941.
Eccleston-Parry, J.D., and Leadbeater, B.S.C. (1995) Regen-eration of phosphorus and nitrogen by four species ofheterotrophic nanoflagellates feeding on three nutritional
Freshwater picoeukaryote diversity 69
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71

states of a single bacterial strain. Appl Environ Microbiol61: 1033–1038.
Edgcomb, V.P., Kysela, D.T., Teske, A., de Vera Gomez, A.,and Sogin, M.L. (2002) Benthic eukaryotic diversity in theGuaymas basin hydrothermal vent environment. Proc NatlAcad Sci USA 99: 7658–7662.
Fenchel, T. (1986) The ecology of heterotrophicmicroflagellates. In Advances in Microbial Ecology.Marshall, K.C. (ed.). New York, USA: Plenum Press, pp.57–97.
Fukami, K., Murata, N., Morio, Y., and Nishijima, T. (1996)Distribution of heterotrophic nanoflagellates and theirimportance as the bacterial consumer in a eutrophiccoastal seawater. J Oceanogr 52: 399–407.
Gestal, C., Novoa, B., Posada, D., Figueras, A., andAzevedo, C. (2006) Perkinsoide chabelardi n. gen., a pro-tozoan parasite with an intermediate evolutionary position:possible cause of the decrease of sardine fisheries?Environ Microbiol 8: 1105–1114.
Green, P.J. (1995) Reversible jump Markov chain MonteCarlo computation and Bayesian model determination.Biometrika 82: 771–732.
Gromov, B.V., Plujusch, A.V., and Mamkaeva, K.A. (1999)Morphology and possible host range of Rhizophydiumalgavorum sp. nov. (Chytridiales) – an obligate parasite ofalgae. Protistology 1: 62–65.
Hughes, J.B., Hellmann, J.J., Ricketts, T.H., and Bohannan,B.J.M. (2001) Counting the uncountable: statisticalapproaches to estimating microbial diversity. Appl EnvironMicrobiol 67: 4399–4406.
James, T.Y., Porter, D., Leander, C.A., Vilgalys, R., andLongcore, J.E. (2000) Molecular phylogenetics of theChytridiomycota supports the utility of ultrastructural datain chytrid systematics. Can J Bot 78: 336–350.
Jürgens, K., and Stolpe, G. (1995) Seasonal dynamics ofcrustacean zooplankton, heterotrophic nanoflagellates andbacteria in a shallow, eutrophic lake. Freshw Biol 33:27–38.
Kagami, M., Van Donk, E., De Bruin, A., Rijkeboer, M., andIbelings, B.W. (2004) Daphnia can protect diatoms fromfungal parasitism. Limnol Oceanogr 49: 680–685.
Kurtzman, C.P., and Fell, J.W. (2004) Yeast. In Biodiversity ofFungi. Muller, G.M., Bills, G.F., and Foster, M.S. (eds).London, UK: Elsevier Academic Press, pp. 337–342.
Kuvardina, O.N., Leander, B.S., Aleshin, V.V., Myl’nikov,A.P., Keeling, P.J., and Simdyanov, T.G. (2002) The phy-logeny of Colpodellids (Alveolata) using small subunitrRNA gene sequences suggests they are free-living sistergroup to Apicomplexans. J Eukaryot Microbiol 49: 498–504.
Lawley, B., Ripley, S., Bridge, P., and Convey, P. (2004)Molecular analysis of geographic patterns of eukaryoticdiversity in Antarctic soils. Appl Environ Microbiol 70:5963–5972.
Leander, B.S., and Keeling, P.J. (2003) Morphostasis inalveolate evolution. Trends Ecol Evol 18: 395–402.
Leander, B.S., Kuvardina, O.N., Aleshin, V.V., Mylnikov,A.P., and Keeling, P.J. (2003) Molecular phylogeny andsurface morphology of Colpodella edax (Alveolata):insights into the phagotrophic ancestry of Apicomplexans.J Eukaryot Microbiol 50: 334–340.
Lim, E.L. (1996) Molecular identification of nanoplanktonicprotists based on small subunit ribosomal RNA genesequences for ecological studies. J Eukaryot Microbiol 43:101–106.
Lim, E.L., Amaral, L.A., Caron, D.A., and Delong, E.F. (1993)Application of rRNA-based probes for observing marinenanoplanktonic protists. Appl Environ Microbiol 59: 1647–1655.
Lim, E.L., Caron, D.A., and Delong, E.F. (1996) Developmentand field application of a quantitative method for examiningnatural assemblages of protists with oligonucleotideprobes. Appl Environ Microbiol 62: 1416–1423.
Lim, E.L., Dennett, M.R., and Caron, D.A. (1999) The ecologyof Paraphysomonas imperforata based on studies employ-ing oligonucleotide probe identification in coastal watersamples and enrichment cultures. Limnol Oceanogr 44:37–51.
Lopez-Garcia, P., Rodriguez-Valera, F., Pedros-Alio, C., andMoreira, D. (2001) Unexpected diversity of small eukary-otes in deep-sea Antarctic plankton. Nature 409: 603–607.
Lopez-Garcia, P., Philippe, H., Gail, F., and Moreira, D.(2003) Autochthonous eukaryotic diversity in hydrothermalsediment and experimental microcolonizers at the Mid-Atlantic Ridge. Proc Natl Acad Sci USA 100: 697–702.
Massana, R., Guillou, L., Diez, B., and Pedros-Alio, C. (2002)Unveiling the organisms behind novel eukaryotic ribosomalDNA sequences from the ocean. Appl Environ Microbiol68: 4554–4558.
Massana, R., Balagué, V., Guillou, L., and Pedros-alio, C.(2004) Picoeukaryotic diversity in an oligotrophic coastalsite studied by molecular and culturing approaches. FEMSMicrobiol Ecol 50: 231–243.
Moon-van der Staay, S.Y., De Wachter, R., and Vaulot, D.(2001) Oceanic 18S rDNA sequences from picoplanktonreveal unsuspected eukaryotic diversity. Nature 409: 607–610.
Moreira, D., and Lopez-Garcia, P. (2002) The molecularecology of microbial eukaryotes unveils a hidden world.Trends Microbiol 10: 31–38.
Myl’nikov, A.P., and Karpov, S.A. (2004) Review of diversityand taxonomy of cercomonads. Protistology 4: 201–217.
Nikolaev, S.I., Berney, C., Fahrni, J., Myl’nikov, A.P., Aleshin,V.V., Petrov, N.B., and Pawlowski, J. (2003) Gymnophryscometa and Lecythium sp. are core Cercozoa: evolutionaryimplications. Acta Protozoologica 42: 183–190.
Okamoto, N., and Inouye, I. (2005) The Katablepharids are adistant sister group of the Cryptophyta: a proposal for Kat-ablepharidophyta divisio nova/ Kathablepharida phylumnovum based on SSU rDNA and Beta-tubulin phylogeny.Protist 156: 163–179.
Patterson, D.J. (1994) Protozoa, evolution and systematics.In Progress in Protozoology. Hausmann, K., and Hüls-mann, N. (eds). Stuttgart, Germany: G Fisher, pp. 1–14.
Powell, M.J. (1993) Looking at mycology with a Janus face: aglimpse at Chytridiomycetes active in the environment.Mycologia 85: 1–20.
Premke, K., and Arndt, H. (2000) Predation on heterotrophicflagellates by protists: food selectivity determined using alive-staining technique. Arch Hydrobiol 150: 17–28.
70 E. Lefèvre et al.
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71

Richards, T.A., Vepritskiy, A.A., Gouliamova, D.E., andNierzwicki-Bauer, S.A. (2005) The molecular diversity offreshwater picoeukaryotes from oligotrophic lake revealsdiverse, distinctive and globally dispersed lineages.Environ Microbiol 7: 1413–1425.
Romari, K., and Vaulot, D. (2004) Composition and temporalvariability of picoeukaryote communities at a coastal site ofthe English Channel from 18S rDNA sequences. LimnolOceanogr 49: 784–798.
Ronquist, F., and Huelsenbeck, J.P. (2003) MRBAYES 3:Bayesian phylogenetic inference under mixed models.Bioinformatics 19: 1572–1574.
Savin, M.C., Martin, J.L., Giewat, M., and Rooney-Varga, J.(2004) Plankton diversity in the Bay of Fundy as measuredby morphological and molecular methods. Microb Ecol 48:51–65.
Shearer, C.A., Langsam, D.M., and Longcore, J.E. (2004)Fungi in freshwater habitats. In Biodiversity of Fungi.Muller, G.M., Bills, G.F., and Foster, M.S. (eds). London,UK: Elsevier Academic Press, pp. 513–531.
Sime-Ngando, T., Demers, S., and Juniper, S.K. (1999) Pro-tozoan bacterivory in the ice and the water column of a coldtemperate lagoon. Microb Ecol 37: 95–106.
Stoeck, T., and Epstein, S. (2003) Novel eukaryotic lineagesinferred from small-subunit rRNA analyses of oxygen-depleted marine environments. Appl Environ Microbiol 69:2657–2663.
Stoeck, T., Taylor, G., and Epstein, S. (2003) Novel eukary-otes from the permanently anoxic Cariaco basin(Caribbean Sea). Appl Eviron Microbiol 69: 5656–5663.
Takishita, K., Miyake, H., Kawato, M., and Maruyama, T.(2005) Genetic diversity of microbial eukaryotes in anoxicsediment around fumaroles on a submarine caldera floorbased on the small-subunit rDNA phylogeny. Extremo-philes 9: 185–196.
Yuan, J., Chen, M.Y., Shao, P., Zhou, H., Chan, Y.Q., andQu, L.H. (2004) Genetic diversity of small eukaryotes fromthe coastal waters of Nansha Islands in China. FEMSMicrobiol Lett 240: 163–170.
Freshwater picoeukaryote diversity 71
© 2006 The AuthorsJournal compilation © 2006 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 9, 61–71




















