
Ultraviolet B Irradiation Promotes Tumorigenic and Metastatic Properties in Primary Cutaneous...
7
1995;55:3669-3674. Cancer Res Rakesh K. Singh, Mordechai Gutman, Reuven Reich, et al. Interleukin 8 Properties in Primary Cutaneous Melanoma via Induction of Ultraviolet B Irradiation Promotes Tumorigenic and Metastatic Updated version http://cancerres.aacrjournals.org/content/55/16/3669 Access the most recent version of this article at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on October 27, 2013. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on October 27, 2013. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on October 27, 2013. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on October 27, 2013. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on October 27, 2013. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on October 27, 2013. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on October 27, 2013. © 1995 American Association for Cancer cancerres.aacrjournals.org Downloaded from
Transcript of Ultraviolet B Irradiation Promotes Tumorigenic and Metastatic Properties in Primary Cutaneous...

1995;55:3669-3674. Cancer Res Rakesh K. Singh, Mordechai Gutman, Reuven Reich, et al. Interleukin 8Properties in Primary Cutaneous Melanoma via Induction of Ultraviolet B Irradiation Promotes Tumorigenic and Metastatic
Updated version
http://cancerres.aacrjournals.org/content/55/16/3669
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on October 27, 2013. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on October 27, 2013. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on October 27, 2013. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on October 27, 2013. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on October 27, 2013. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on October 27, 2013. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from
Research. on October 27, 2013. © 1995 American Association for Cancercancerres.aacrjournals.org Downloaded from

[CANCER RESEARCH 55. 3669-3674, August 15, 1995]
Ultraviolet B Irradiation Promotes Tumorigenic and Metastatic Properties inPrimary Cutaneous Melanoma via Induction of Interleukin 81
Rakesh K. Singh, Mordechai (intuÃan, Reuven Reich,2 and Menashe Bar-Eli3
Department of Cell Biology, The University of Texas M. D. Anderson Cancer Center, Houston, Texas 77030
ABSTRACT
UV radiation has been shown to play a role in the initiation of humancutaneous melanoma, but its role in the development of malignant melanomato the metastatic state is not very well defined. Although previous studies haveconcentrated on the effect of I V-Ii on the host immune response, the effect ofUV-B on the tumor cells was not elucidated. Here we show that IA -I! caninduce interleukin 8 (IL-8) niKNA and protein secretion in human cutaneousmelanoma with negligible expression of IL-8. I'\ -It-induced IL-8 was con-
stitutively expressed 60 days after irradiation in tumors implanted in mice.Induction of IL-8 was UV-B dose dependent and blocked by cyclohexamide,
indicating that de novo protein synthesis is required for its expression. TheUV-irradiated cells demonstrated enhanced tumorigenicity and metastatic
potential in nude mice. The increase in tumorigenicity and metastatic abilitycould be explained by the increase in .U, 72,000 type IV collagenase activityand angiogenesis attributed to the induction of IL-8 after irradiation. Theacquisition of the metastatic phenotype induced by UV-B could not be attributed to abnormalities in thep53 or MTS-1 (pl6INK4) genes. To the best of our
knowledge, this is the first report to show that UV-B can increase the
aggressiveness of human cutaneous melanoma for growth and metastasis.
INTRODUCTION
The reduction of stratospheric ozone over the past 20 years hasraised concern over the possibility that additional erosion will increasedanger of exposure to UVR4 (1, 2). Epidemiological data suggest that
exposure to UVR plays a major role in the development of at leastsome cutaneous melanoma (3). The role of UV radiation in thedevelopment of human cutaneous melanoma to its metastatic state isnot very well characterized. Earlier studies have demonstrated thatUV irradiation can dramatically enhance the induction of primarycutaneous melanoma in mice (4, 5), and that injection of transplant-able melanoma cells into chronically UV-irradiated skin resulted in an
increased incidence and enhanced spread of melanoma (6, 7). Thesestudies focused mainly on the effect of UV on the host immuneresponse while the effect of UV-irradiation as a promoter agent on the
tumor cells themselves remained unclear. UVR has been shown toproduce marked pathophysiological responses in animal models. It isone of the most potent stimulators of cytokine production fromdifferent cell types in the skin (8, 9). Indeed, after UV irradiation,several cytokines have been shown to be up-regulated in epidermalcells and keratinocytes (10). These cytokines include: IL-1 (11), IL-6(12), IL-8 (9), IL-10 (13, 14), and TNF-a (15). However, the role of
UVR in mediating cutaneous melanoma progression by cytokineproduction has not been well defined.
Malignant melanoma cells produce a variety of cytokines andgrowth factors either constitutively or via induction by other cytokines(16, 17) that support growth and progression directly or indirectly.
Received 4/13/95; accepted 6/14/95.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1Supported in part by NIH Grant CA 41525 (M. B-E.).2 On sabbatical from the Hebrew University, Jerusalem, Israel3 To whom requests for reprints should be addressed, at Department of Cell Biology,
Box 173, University of Texas M. D. Anderson Cancer Center, 1515 Holcombe Boulevard,Houston, TX 77030.
4 The abbreviations used are: UVR, UV radition: IL. interleukin; TNF, tumor necrosis
factor; MTS-1, multiple lumor suppressor 1.
Recently, we showed that the expression of the multifunctional cytokine IL-8 is directly correlated with the metastatic phenotype in
human melanoma cell lines (18). The weakly tumorigenic melanomacells in nude mice did not express detectable levels of IL-8 mRNA
and protein.The purpose of this study was to investigate whether exposure to
UVR can modulate the progression of human cutaneous melanoma tothe tumorigenic and metastatic states. We found UVR exposure toinduce IL-8 mRNA and protein secretion in a human cutaneousmelanoma. UV-irradiated cells demonstrated enhanced tumorigenicity
and metastatic potential in nude mice, which can be attributed to theobserved increase in M, 72,000 type IV collagenase activity andangiogenesis, mediated possibly by the induction of IL-8. Our data
support the notion that UVR maybe involved not only in the initiationof melanomas but in their progression to malignancy.
MATERIALS AND METHODS
Cell Line and Culture Condition. The SB-2 cell line was isolated from
a primary cutaneous lesion (19) and was a gift from Dr. B. Giovanella (St.Joseph's Hospital Cancer Center, Houston, TX). The cells were maintainedin culture as adherent monolayer in Eagle's MEM supplemented with 10%
fetal bovine serum, sodium pyruvate, nonessential amino acids, L-gluta-mine, 2-fold vitamin solution, and penicillin-streptomycin (Flow Labora
tories, Rockville, MD). All cultures were free of mycoplasma and pathogenic murine viruses (assayed by Microbiological Associates, Bethesda,MD). Cultures were maintained for no longer than 4 weeks after recoveryfrom frozen stocks.
Animals. Male athymic BALB/c nude mice were purchased from theAnimal Production Area of the National Cancer Institute, Frederick CancerResearch Facility (Frederick, MD). The mice were housed in laminar flowcabinets under specific pathogen-free conditions and used at 8 weeks of age.
Animals were maintained in facilities approved by the American Associationfor Accreditation of Laboratory Animal Care and in accordance with currentregulations and standards of the United States Department of Agriculture,Department of Health and Human Services, and National Institutes of Health.
UV-B Irradiation. The procedure of Schwarz et al. (20) was used toirradiate the melanoma cells. Five X IO5 cells were plated into 100-mm tissue
culture dishes in 10 ml of complete medium. Medium was removed andmonolayers were washed three times with PBS and overlayed with PBS. Themonolayers were exposed to a radiation source (a single FS-40 sunlight bulbfrom Westinghouse, Bloomfield, NJ) with an output of 1.43 J/m2/s at a
tube-to-target distance of 23 cm. For different doses of irradiation, monolayerswere exposed to different durations. After irradiation, the cells were resus-pended in complete medium. For IL-8 protein estimation, irradiated cells weretrypsinized and plated in a 96-well culture plate. For RNA analysis, irradiated
cells were lysed, and total RNA was isolated as described below.ELISA for Human IL-8. IL-8 levels in cell culture-free supernatants from
different treatments were determined by using an ELISA kit (Quantikine; R&DSystems, Inc., Minneapolis, MN). This assay is a quantitative immunometric"sandwich" enzyme immunoassay. A curve of the absorbance versus the
concentration of IL-8 in the standard wells was plotted. By comparing the
absorbance of the samples to the standard curve, we determined theconcentration of IL-8 in the unknown samples.
Cyclohexamide Treatment. Cyclohexamide was purchased from SigmaChemical Co. (St. Louis. MO). Ten fig/ml (final concentration) of cyclohexamide were added to the culture during the exposure and maintained throughout the duration of incubation.
3669
UV-B ROLF. IN THE PROGRESSION OF MELANOMA
Northern Blot Analysis. Total cellular RNA was isolated by using TRIreagent (Molecular Research, Inc.) according to the manufacturer's
instructions. Twenty /xg of total RNA/lane were electrophoresed on a 1%denaturing formaldehyde/agarose gel, electrotransferred at 0.6 amp to Gene-Screen nylon membrane (DuPont Co., Boston. MA) and UV cross-linked with
120,000 fjJ/cnr by using a UV stratalinker 1800 (Stratagene, La Jolla, CA).Hybridizations were performed as described previously (18). Nylon filterswere washed 3 times at 55—60°Cwith 30 mM NaCl-3 mM sodium citrate (pH
7.2)-().l% SDS (w/v). The cDNA probes used in these analyses were a 1.3-kb
Pstl cDNA fragment corresponding to rat glyceraldehyde 3-phosphate dehy-
drogenase (21), a 0.5-kb EcoRl cDNA fragment corresponding to human IL-8(a gift of Dr. K. Matsushima, Kanazawa, Japan; Ref. 22), and a p53 pR4-2
cDNA probe (23). Each cDNA fragment was purified by agarose gelelectrophoresis, recovered by using GeneClean (BIO 101, Inc., La Jolla, CA),and radiolabeled by using the random primer technique with [a-12P]
deoxyribonucleotide triphosphates.IL-8 mRNA expression was quantitated ¡nthe linear range of the film on a
Personal Densitometer by using the ImageQuant software program (Molecular
Dynamics, Sunnyvale, CA). Each sample measurement was calculated as theratio of area between the 1.8-kb IL-8-specific mRNA transcript and the 1.3-kb
glyceraldehyde 3-phosphate dehydrogenase transcript.
Tumor Cell Injections. To prepare tumor cells for inoculation, cells inexponential growth phase were harvested by brief exposure to 0.25% trypsin-
0.02% EDTA solution (w/v). The flask was sharply tapped to dislodge thecells, and supplemented medium was added. The cell suspension was pipettedto produce a single-cell suspension. The cells were washed and resuspended ¡nCa2+- and Mg2 +-free HBSS to the desired cell concentration. Cell viability
was determined by trypan blue exclusion, and only single-cell suspensions of
>90% viability were used.s.c. tumors were produced by injecting 1 X IO6 tumor cells-0.2 ml HBSS
over the right scapular region. Growth of s.c. tumors was monitored by twiceweekly examination of the mice and measurement of tumors with calipers. Themice were killed 2 months after injection, and tumors were processed for
hematoxylin and eosin staining.For experimental lung metastasis, 1 X 10'' tumor cells-0.2 ml of HBSS were
injected into the lateral tail vein of nude mice. The mice were killed after 60days, and the lungs were removed, washed in water, and fixed with Bouin's
solution for 24 h to facilitate counting of tumor nodules as we described
previously (24, 25). The number of surface tumor nodules was counted byusing a dissecting microscope. Sections of the lungs were stained with hematoxylin and eosin to confirm that the nodules were melanoma and to monitorthe presence of micromctastasis.
Zymography. Collagenolytic activity was determined on substrate- impregnated gels (26), with minor modifications. UV-B-irradiated cells werecultured for 7 and 14 days. Five X IO3 cells/well were replated into a 96-well
culture plate. After 24 h of cell adherence, cultures were washed and refed with
fresh media. Culture supernatants were collected and analyzed for collagenaseactivity. Samples of culture media were separated on gelatin-impregnated (1mg/ml; Difco, Detroit, MI) SDS-8% polyacrylamide gels under nonreducingconditions, followed by 30 min of shaking in 2.5% Triton X-100 (BDH, UnitedKingdom). The gels were then incubated for 16 h at 37°Cin 50 mM Tris, 0.2
M NaCl, 5 mM CaCU, and 0.02% Brij 35 (w/v) at pH 7.6. At the end of theincubation, the gels were stained with 0.5% Coomassie G 250 (Bio-Rad,
Richmond, CA) in methanol:acetic acid:H20 (30:10:60). The intensity of thevarious bands was determined on a computerized densitometer (MolecularDynamics type 300A).
p53 and MTS-1 (p/6INK4 and CDKN2) Gene Expression and Mutation
Analysis. Mutations in the conserved regions of the p53 gene were identifiedby single strand conformation polymorphism analysis of exons 5-9 and veri
fied by direct DNA sequencing of PCR products, as we described previously(27—29).Expression of p53 was analyzed by Northern blot using the PR4-2cDNA probe (23) as we previously described (30). MTS-1 (/>/6INK4) analysis
was performed by PCR amplification of exons 1-2, followed by direct
sequencing.Statistical Analysis. The in vitro data were analyzed for significance by
the Student r test (two-tailed), and the in vivo data were analyzed by theMann-Whitney test.
RESULTS
Induction of IL-8 mRNA by UV-B Irradiation. To assess theeffect of UV-B irradiation on the progression of human primarycutaneous melanoma, we chose to work with the SB-2 cells. SB-2
cells originated from primary cutaneous melanoma (19) and areneither tumorigenic nor metastatic in nude mice (19, 27, 31, 32). Inaddition, SB-2 is the only cell line that exhibited negligible levels ofIL-8 mRNA expression and protein secretion. All of the other 12
human melanoma cell lines analyzed (32) that originated from metastatic lesions produced and secreted IL-8, and the level of IL-8
expression was found to be correlated with their metastatic potentialin vivo (18).
In the first set of experiments, we analyzed the steady state mRNAexpression of IL-8 in UV-irradiated or unirradiated cells. Monolayerswere irradiated with UV-B (200 J/m2) and incubated for different time
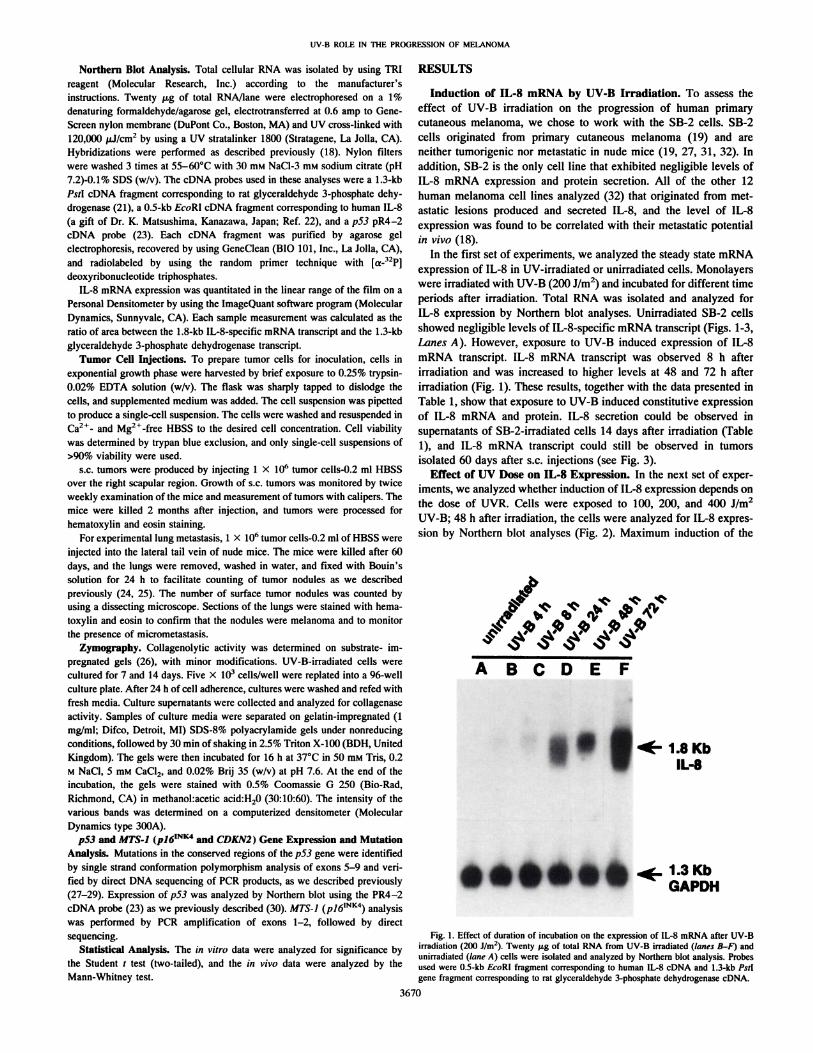
periods after irradiation. Total RNA was isolated and analyzed forIL-8 expression by Northern blot analyses. Unirradiated SB-2 cellsshowed negligible levels of IL-8-specific mRNA transcript (Figs. 1-3,Lanes A). However, exposure to UV-B induced expression of IL-8mRNA transcript. IL-8 mRNA transcript was observed 8 h after
irradiation and was increased to higher levels at 48 and 72 h afterirradiation (Fig. 1). These results, together with the data presented inTable 1, show that exposure to UV-B induced constitutive expressionof IL-8 mRNA and protein. IL-8 secretion could be observed insupernatants of SB-2-irradiated cells 14 days after irradiation (Table1), and IL-8 mRNA transcript could still be observed in tumors
isolated 60 days after s.c. injections (see Fig. 3).Effect of UV Dose on IL-8 Expression. In the next set of exper
iments, we analyzed whether induction of IL-8 expression depends onthe dose of UVR. Cells were exposed to 100, 200, and 400 J/m2
UV-B; 48 h after irradiation, the cells were analyzed for IL-8 expres
sion by Northern blot analyses (Fig. 2). Maximum induction of the
A B C D E F
•* t 1.8KbIL-8
1.3KbGAPDH
Fig. 1. Effect of duration of incubation on the expression of IL-8 mRNA after UV-Birradiation (2(M)J/m2). Twenty jag of total RNA from UV-B irradiated (lanes B-F) and
unirradiated (lane A) cells were isolated and analyzed by Northern blot analysis. Probesused were 0.5-kb EcoRl fragment corresponding to human IL-8 cDNA and 1.3-kb Palgene fragment corresponding to rat glyceraldehyde 3-phosphate dehydrogenase cDNA.
3670

UV-B ROLE IN THE PROGRESSION OF MELANOMA
A B C D
1.8KbIL-8
1.3KbGAPDH
Fig. 2. Effect of different doses of UV-B irradiation on the expression of IL-8 mRNA.Cells were harvested 48 h after irradiation, and 20 fig/lane of total RNA were used forNorthern blot analysis.
v v
A B C D
Induction of IL-8 Secretion by UV-B. The production of IL-8
protein and its secretion to the supernatants was measured by ELISAas described in "Materials and Methods." The dose of UV-B used, 200J/m2, did not induce observable changes in cell viability as assayed by
plasma membrane permeability to trypan blue; the activity in thesupernatants was normalized, therefore, to total cell number. UV-B-irradiated or unirradiated cells (5 X IO3 cells/well) were plated on a
96-well culture plate, and culture supernatants were harvested 24, 48,72, 96 h after irradiation. To analyze the long-term effects of UVR onIL-8 production, irradiated cells were cultured in vitro for 7 and 14days. Twenty-four h before the termination of experiments, cultures
were washed and refed with fresh media. Culture supernatants werecollected and analyzed for IL-8 protein. Unirradiated, continuously
cultured cells were used as control. The unirradiated cells expressedvery low levels of IL-8 protein for the duration of up to 14 days inculture, whereas cells irradiated with UV-B showed induction of IL-8secretion (Table 1). The highest level of IL-8 protein secretion was
observed after 72 h of incubation (Table 1; Fig. 1) and reached aplateau 96 h after irradiation. The cells continued, however, to secreteIL-8 constitutively for at least 14 days after irradiation (Table 1).
Effect of Cyclohexamide Treatment on UV-B-induced IL-8Production. To examine the mechanism of IL-8 induction in SB-2cells, we incubated the UV-B-irradiated cells with or without cyclo-
hexamide (10 ju.g/ml), which blocks cÃenovo protein synthesis. Mostof the UV-B-induced IL-8 protein level was blocked in the presence
of cyclohexamide (Table 2), indicating that de novo protein synthesisis required for enhanced IL-8 production.
Tumor Growth. Next, we analyzed whether induced IL-8 expression in the UV-B- irradiated cells can lead to increased tumorigenic-ity. To that end, 1 X IO6 UV-B-irradiated cells (after 48 h of
incubation) were injected s.c. into BALB/c nude mice. Each mousewas injected with a separate batch of irradiated cells. Tumor growthwas monitored twice a week, and the tumors were harvested toanalyze IL-8 expression when they have reached 1 cm in mean
diameter. The results summarized in Table 3 show that unirradiated
1.8KbIL-8
Table I Expression of IL-8 proietti after UV-B irradiation
1.3KbGAPDH
Fig. 3. Analysis of 3 s.c. tumors for expression of IL-8 mRNA. Twenty /ig of totalRNA/lane were used for Northern blot analysis. Probes used were 0.5-kb £Vr>Rlgenefragment corresponding to human IL-H cDNA, and 1-i-kb-Av/l gene fragment corresponding to rat glyceraldehyde 3-phosphate. Each s.c. tumor (lanes B-D) represents a separate
experiment in which cells were irradiated in vitro, and each irradiated cell population wasinjected into different mice. Lane A, SB-2 parental unirradiated cells grown in culture.
Duration oftreatment
Secretion of IL-8 protein (pg/ml)after UV-B irradiation (2(X) J/m2)"
Unirradiated24 h48 h72h%h
7 days''14 days'*
52.0 ±LO*232.8 ±5.0''
460.9 ±7.2'700.8 ±21.0'
460 ±18.0'338.0 ±6.5'332.0 ±5.7'
" Unirradiated or UV-B-irradiated cells were plated at a concentration of 5 X IO1cells/well in a °6-wellculture plate for different durations as described in "Materials andMethods." Expression of IL-8 protein was analyzed by ELISA. Values are mean nl
pg/ml ±SD of duplicate cultures. This is a representative experiment of two.''This background level in unirradiated cells remained constant up to 14 days in
culture.' P < 0.0(11 compared to unirradiatcd.''UV-B irradiated cells were cultured for 7 and 14 days. 5 X IO3 cells/well were
replated into a 96-well culture plate. After 24 h of cell adherence, cultures were washedand refed with fresh media. Culture supcrnatants were collected and analyzed for IL-8expression as described in "Materials and Methods."
IL-8 gene expression was observed with 400 J/m2 of UV-B (Fig. 2).
With this UV dose, however, we observed about 30% cell death (datanot shown), and increasing the intensity of UV-B above 400 J/m2 did
not result in a further increase in the level of IL-8 gene expression
(data not shown). Therefore, to ensure cell viability, we considered200 J/m2 as the optimum dose and continued to use it throughout our
in vitro and in vivo experiments (vide infra).
Table 2 Effect of C\'clohexatniile on UV-H-inditced IL-8 expression5 X 10" cells were treated with UVR with or without cyclohexamide. and 5 X IO'1cells
were plated into a %-well plate for 24 h. Secretion of IL-8 was analyzed by ELISA.
Values are mean ±SD of duplicate cultures. This is a representative experiment of two.
3671
Treatment IL-8 secreted (pg/ml)
UnirradiatedUVR (200 J/m2)
UVR + CHX" (10 Mg/
88.8 ±6.2237.8 ±7.4
70.8 ±4.6" CHX, cyclohexamide.
UV-B ROLE IN THE PROGRESSION OF MELANOMA
Table 3 Tumor growth and experimental lung metastasis
Tumor growth (s.c.)" Experimental lung metastasis
Treatment Tumorigenicity' Days'' Incidence' Median (range)
UnirradiatedUVR0/10 8/860+110/10 7/100 6(1-10)
1 1 X IO6 cells were injected s.c.' 1 x IO6 cells were injected into the lateral tail vein.
Number of mice with tumor/number of injected mice.' Mean ±SD of days for s.c. tumors to reach I cm (mean diameter).
cells did not form palpable tumors 2 months after injections; theseresults confirmed our data published previously (27, 31, 32). Incontrast, UV-B-irradiated cells grew in all mice that received injec
tions (100% tumor uptake), reaching 1 cm in mean diameter in60 ±11 (SD) days. s.c. tumors from 3 mice that received injectionswere harvested 60 days after injections and assayed for IL-8 gene
expression by Northern blot analysis. Fig. 3 demonstrates that, 60days after injections, s.c. tumors of irradiated SB-2 cells continued toexpress high levels of IL-8-specific mRNA transcript in vivo.
Experimental Lung Metastasis. We have demonstrated previously that expression of IL-8 is directly correlated with metastatic
potential in malignant melanoma cells (18). Next, we analyzed themetastatic potential of UV-B-irradiated cell in an experimental lungmetastasis assay. One X IO6 cells (UV-B irradiated or unirradiated)
were injected into the lateral tail vein of BALB/c nude mice. Separatebatches of irradiated cells were injected into different mice. The micewere killed after 75 days. Lungs were rinsed in water and fixed inBouin's solution to visualize metastatic nodules. Formalin-fixed he-
matoxylin and eosin preparation were also made to analyze microme-
tastasis. We found that unirradiated cells did not metastasize to thelungs (Table 3), whereas UV-irradiated SB-2 cells formed lung métastases in 7 of 10 mice given injections (range of 1-10, median 6;Table 3). These data suggested that UV-B irradiation can lead toenhanced IL-8 production and, hence, contribute to tumor progression
and metastasis.Increase in Afr 72,000 Type IV Collagenase (MMP-2) Activity
after UV-B Irradiation. The metastatic potential of tumor cellsdepends on proper vascularization of the tumor and its ability todegrade type IV collagen. Recently, IL-8 has been shown to beassociated with angiogenesis (33). IL-8 may exert its angiogenic
activity through the induction of type IV collagenase in the tumorcells. Activation of the type IV collagenase may provide a mechanismfor the increase in the metastatic potential of SB-2 cells after UV-B
irradiation. To test this hypothesis, we analyzed the activity of type IVcollagenases (A/r 72,000 and 92,000) in SB-2 cells before and afterUV-B irradiation (200 J/m2). Supernatants from unirradiated and
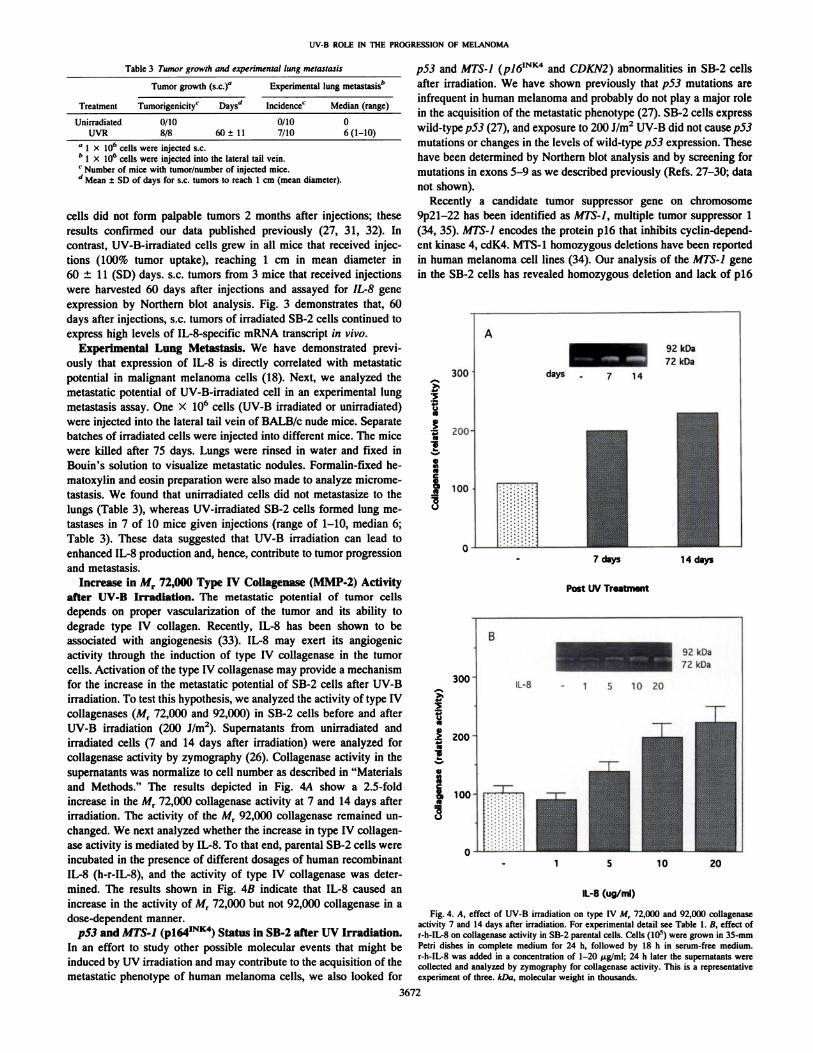
irradiated cells (7 and 14 days after irradiation) were analyzed forcollagenase activity by zymography (26). Collagenase activity in thesupernatants was normalize to cell number as described in "Materialsand Methods." The results depicted in Fig. 4A show a 2.5-fold
increase in the M, 72,000 collagenase activity at 7 and 14 days afterirradiation. The activity of the Mr 92,000 collagenase remained unchanged. We next analyzed whether the increase in type IV collagenase activity is mediated by IL-8. To that end, parental SB-2 cells were
incubated in the presence of different dosages of human recombinantIL-8 (h-r-IL-8), and the activity of type IV collagenase was determined. The results shown in Fig. 4B indicate that IL-8 caused an
increase in the activity of Mr 72,000 but not 92,000 collagenase in adose-dependent manner.
p53 and MTS-1 (P164INK4) Status in SB-2 after UV Irradiation.
In an effort to study other possible molecular events that might beinduced by UV irradiation and may contribute to the acquisition of themetastatic phenotype of human melanoma cells, we also looked for
p53 and MTS-1 (pl6ÕNK4 and CDKN2) abnormalities in SB-2 cells
after irradiation. We have shown previously that p53 mutations areinfrequent in human melanoma and probably do not play a major rolein the acquisition of the metastatic phenotype (27). SB-2 cells expresswild-type p53 (27), and exposure to 200 J/m2 UV-B did not cause p53
mutations or changes in the levels of wild-type p53 expression. These
have been determined by Northern blot analysis and by screening formutations in exons 5-9 as we described previously (Refs. 27-30; data
not shown).Recently a candidate tumor suppressor gene on chromosome
9p21-22 has been identified as MTS-1, multiple tumor suppressor 1(34, 35). MTS-1 encodes the protein pió that inhibits cyclin-depend-ent kinase 4, cdK4. MTS-1 homozygous deletions have been reportedin human melanoma cell lines (34). Our analysis of the MTS-1 genein the SB-2 cells has revealed homozygous deletion and lack of pl6
jj
i
300
200-
100
o-1-7 days
Post UV Treatment
14 days
20
IL-8 (ug/ml)
Fig. 4. A, effect of UV-B irradiation on type IV M, 72,000 and 92,000 collagenaseactivity 7 and 14 days after irradiation. For experimental detail see Table 1. B, effect ofr-h-IL-8 on collagenase activity in SB-2 parental cells. Cells (IO5) were grown in 35-mm
Petri dishes in complete medium for 24 h, followed by 18 h in serum-free medium.r-h-IL-8 was added in a concentration of 1-20 ug/ml; 24 h later the supernatants werecollected and analyzed by zymography for collagenase activity. This is a representativeexperiment of three. kDa, molecular weight in thousands.
3672

UV-B ROLE IN THE PROGRESSION OF MELANOMA
mRNA expression.5 The deletion was observed in the parental SB-2
cells, however, regardless of UV-B irradiation. The observed homozy-gous deletion in the MTS-Ì(CDKN2) gene in parental SB-2 cells,before UV-B irradiation, could not account for the acquisition of themetastatic phenotype induced by UV-B.
DISCUSSION
In this study we demonstrated that SB-2 human melanoma cells,
which are not tumorigenic in nude mice, expressed negligible levels ofIL-8 mRNA and protein. Upon exposure to UV-B, however, IL-8
mRNA and protein production were induced to the constitutive expression state. High levels of IL-8 mRNA transcript could still be
observed in tumors isolated 60 days after s.c. injections (Fig. 3).Induction of IL-8 production in SB-2 cells increased their tumorigenic
and metastatic potential in nude mice.Several possible mechanisms for the induction of IL-8 gene expres
sion in the UV-irradiated SB-2 cells should be considered. Our datasuggested that UV-B irradiation "switched on" the IL-8 gene to be
expressed constitutively, and that inhibition of de novo protein synthesis by cyclohexamide completely blocked IL-8 stimulation, suggesting that new protein synthesis is required for the induction of IL-8
in these cells. Different autocrine and/or paracrine stimulation of othercytokines induced by UV-B may contribute to this up-regulation.Studies have shown that IL-1 or TNF-a can very rapidly induce IL-8in variety of cell types (36-37). In fact, responsive elements for IL-1and TNF-a have been found in the 5 ' flanking region of the IL-8 gene.
The promoter region of the IL-8 gene was also shown to contain anAP-1-binding site (38), which interacts with a dimeric transcriptionalactivator composed of c-jun and c-fos (39). UV-B may induce IL-8expression through the induction of c-jun/c-fos and their binding tothe AP-1 site. This speculation is supported by the fact that UV-Benhanced rapid and transient c-jun expression in mouse skin, as wellas immediate early c-fos expression in mouse epidermal cells (40, 41).The mechanisms regulating induction of IL-8 in SB-2 cells remain tobe elucidated. They might involve one of the above-mentioned mechanisms or be caused by unknown mechanisms regulated by UV-B
irradiation.The enhanced or constitutive expression of IL-8 in melanoma
progression has a major role in pathogenesis because of its multifunctional effects. IL-8 has been shown to be an angiogenic mol
ecule (33), an autocrine growth factor for melanoma cells (18), andan inducer of haptotactic migration in melanoma cells (42). Wehave shown that constitutive expression of IL-8 in different human
melanoma cells correlated with their metastatic potential in nudemice (18). Our data suggested that UV-B-irradiated SB-2 cellsgrew and formed s.c. tumors in all mice injected, and they metas-
tasized to the lungs in 70% of the mice given injections. Thisinduction of tumorigenic and metastatic potential in a humancutaneous melanoma might be attributed to IL-8 expression and itsangiogenic properties. In addition, IL-8 has recently been shown tobe capable of activating phospholipase D in human neutrophils(43). Phospholipase D, in turn, is involved in the intracellularsignal transduction mechanisms that mediate induction of gelatin-ase A (MMP-2) by laminin in tumor cells (44). IL-8, therefore, can
cause an increase in type IV collagenase activity and, hence,contribute to invasion and metastasis. Indeed, our data showed anincrease in MT 72,000 type IV collagenase activity after UV-B
irradiation of the melanoma cells. This increase in collagenaseactivity could be attributed directly to the effect of IL-8 because
activation of type IV collagenase could also be achieved when
' M. Luca, S. Xie, M. Gutman, S. Huang, and M. Bar-Eli, submitted for publication.
human recombinant IL-8 was added to the parental SB-2 cells (Fig.4B). We cannot, however, rule out, the possibility that UV-B
irradiation can cause other genetic changes that may contribute toenhanced tumorigenicity and metastasis. These genetic changes areyet to be defined.
p53 and MTS-I (pl6mtl4 and CDKN2) abnormalities were ruled
out in our studies as a possible mechanism for the acquisition of themetastatic phenotype in SB-2 cells after UV irradiation. Mutations in
p53 were, however, observed to be induced by sunlight during thedevelopment of squamous cell carcinoma of the skin (45). In thismodel, UV-B acted first to induce p53 mutations, and subsequently toselect for clonal expansion of the p53-mu\aied cells. Our study sug
gested that in addition to its role in initiation, UVR maybe involved inthe acquisition of the metastatic phenotype in melanomas by inductionof cytokine production. Understanding the mechanism involved isimportant to providing a means for pharmacological intervention inthe process. We hope that additional studies will elucidate some of therelationships between UV radiation and the progression of humanmelanoma and suggest other means to manipulate the consequence ofexposure to UV irradiation.
ACKNOWLEDGMENTS
We thank Dr. Isaiah J. Fidler and Dr. Margaret L. Kripke for critical reviewand Pathcrine Greenwood for expert preparation of this manuscript.
REFERENCES1. Frederick, J. E., and Snell, H. E. Ultraviolet radiation levels during the Antarctic-
spring. Science (Washington DC). 241: 438-440, 1988.
2. Kerr, R. A. Stratospheric ozone is decreasing. Science (Washington DC). 239:1489-1491, 1988.
3. Holman, C. D. J., Mulroney, C. D., and Armstrong. B. K. Epidemiology of prcinva-sive and invasive malignant melanoma in western Australia. Int. J. Cancer, 25:317-323, 1980.
4. Romerdahl. C. A., Donawho, C., Fidler. I. J.. and Kripke, M. L. Effect of UV-Bradiation on the in viva growth of murine melanoma cells. Cancer Res.,48:4007-4010. 1988.
5. Romerdahl. C. A., Stephens. L. C., Bucana, C. D.. and Kripke. M. L. The role ofultraviolet radiation in the induction of melanocytic skin tumors in inbred mice.Cancer Commun., /: 209-216, 1989.
6. Donawho, C. K.. and Kripke, M. L. Evidence that the local effect of UV radiation onthe growth of murine melanomas is immunologically mediated. Cancer Res., 41:4176-4181. 1991.
7. Donawho, C. K., Wolf, P.. and Kripke, M. L. Enhanced development of murinemelanoma in UV irradiated skin: UV dose response, waveband dependence, andrelation to inflammation. Melanoma Res., 4: 93-100. 1994.
8. Griswold, D. E., Connor, J. R., Dalton, B. J., Lee, J. C., Simon, P., Hillegass, L., Sieg,D. J., and Hanna. N. Activation of the IL-1 gene in UV-irradiatcd mouse skin:
association with inflammatory sequelae and pharmacologie intervention. J. Invest.Dermatol., 97: 1019-1023, 1991.
9. Kondo. S. K., Kono. T.. Sauder, D. N.. and McKcnzie, R. C. IL-8 gene expression andproduction in human keratinocytes and their modulation by UVB. J. Invest. Dermatol., 101: 690-694, 1993.
10. Schwarz, T., and Luger. T. A. Effect of UV irradiation on epidermal cell cytokineproduction. J. Pharmacol. Photobiol., 4: 1-13, 1989.
11. Ansel, J. C., Luger, T. A., and Green, I. The effect of in vitro and in vivo UVirradiation on the production of ETAF activity by human and murine keratinocytes.J. Invest. Dermatol., 81: 519-523, 1983.
12. Urbunski, A., Schwarz, T., Neuner, P.. Krulmunn. J., Kirnbaucr, R., Kock, A., andLuger, T. A. Ultraviolet light induces increased circulating interlcukin-6 in humans.J. Invest. Dermatol., 94: 808-811, 1989.
13. Rivas, J. M., and Ullrich, S. E. Systemic suppression of delayed-type hypersensitivityby supernatants from UV-irradiated keratinocytes. An essential role for keratinocyte-derived IL-10. J. Immunol., /49: 3865-3871, 1992.
14. Grcwe, M., Gyufko, K., and Krutman, J. Interleukin-10 production by cultured human
keratinocytes: regulation by ultraviolet B and ultraviolet Al radiation. J. Invest.Dermatol., 104: 3-6, 1995.
15. Kock, A.. Schwarz, T., Kirnbaucr, R., Urbanski. A., Perry. P., Ansel, J. C., and Luger.T. A. Human keratinocytes are a source of tumor necrosis factor a: evidence forsynthesis and release upon stimulation with endotoxin or ultraviolet light. J. Exp.Med., 172: 1609-1614, 1990.
16. Armstrong, C. A., Tara, D. C., Hart, C. E., Kock, A., Luger, T. A., and Ansel, J. C.Heterogeneity of cytokine production by human malignant melanoma cells. Exp.Dermatol., /: 37-45, 1992.
17. Krasagakis, K., Garbe, C., and (Manos, C. E. Cytokines in human melanoma cells:
3673

UV-B ROLE IN THE PROGRESSION OF MELANOMA
synthesis, autocrine stimulation and regulatory functions-an overview. MelanomaRes., 3: 425-433, 1993.
18. Singh, R. K., Gutman, M., Radinsky, R., Bucana, C. D., and Fidler, I. J. Expressionof interleukin-8 correlates with the metastatic potential of human melanoma cells innude mice. Cancer Res., 54: 3242-3247, 1994.
19. Verschraegen, C. E., Giovanella, B. C., Mendoza, J. T., Kozielsky, A. I., and Stehlin,J. S., Jr. Specific organ metastasis of human melanoma cells injected into the arterialcirculation of nude mice. Anticancer Res., //: 529-536, 1991.
20. Schwarz, T. A., Gschnait, V. F., and Lugar, T. A. Inhibition of the induction ofcontact hypersensitivity by UV-mediated epidermal cytokines. J. Invest. Dermatol.,87: 289-294, 1986.
21. Fort, P., Marty, L., Piechaczyh, M., Sabrouty, S. E., Dani, C., Jeanteur, P., andBlanchard, J. M. Various rat adult tissues express only one major mRNA species fromthe glyceraldehyde-3-phosphate-dehydrogenase multigenic family. Nucleic AcidsRes., 13: 1431-1442, 1985.
22. Matsushima, K., Morishita, K., Yoshimura, T., Lavu, S., Kobayashi, Y., Lew, W.,Appella, E., Kung, H. F., Leonard, E. J., and Oppenheim, J. J. Molecular cloning ofa human monocyte-derived neutrophil chemotactic factor (MDNCF) and the induction of MDNCF mRNA by ¡nterleukin-1and tumor necrosis factor. J. Exp. Med., 167:1883-1893, 1988.
23. Harlow, E., Williamson, R., Ralston, M. R., Helfman, D. M., and Adams, T. E.Molecular cloning and in vitro expression of a cDNA clone for human cellular tumorantigen p53. Mol. Cell Biol., 5: 1601-1610, 1985.
24. Radinsky, R., Fidler, I. J., Price, J. E., Esumi, N., Tsan, R., Petty, C. M., Bucana,C. D., and Bar-Eli, M. Terminal differentiation and apoptosis in experimental lungmétastasesof human osteogenic sarcoma cells by wild type p53. Oncogene, 9:1877-1883, 1994.
25. Huang, S., Singh, R. K., Xie, K., Gutman, M., Berry, K. K., Bucana, C. D., Fidler,I. J., and Bar-Eli, M. Expression of the JE/MCP-I gene suppresses metastatic
potential in murine colon carcinoma cells. Cancer Immunol. Immunother., 39:231-238, 1994.
26. Ballin, M., Gomez, D. E., Sinha, C. C., and Thorgeirsson U. P. ras oncogenemediated activation of a 92 kDa metalloproteinase showing correlation with themalignant phenotype. Biochem. Biophys. Res. Commun., 154: 832-838, 1988.
27. Luca, M., Lenzi, R., Jackson, D. L., Gutman, M., Fidler, I. J., and Bar-Eli, M. p53
mutations are infrequent and do not correlate with the metastatic potential of humanmelanoma cells. Int. J. Oncol., 3: 19-22, 1993.
28. Jackson, D. L., Abbruzzese, J. L., and Bar-Eli, M. Artifactual frame-shift p53mutation at codon 249 detected with the cyclist DNA sequencing method. BioTech-niques, 15: 363-364, 1993.
29. Bar-Eli, M., Abbruzzese, J. L., Jackson, D. L., and Frost, P. p53 gene mutationspectrum in human unknown primary tumors. Anticancer Res. 13: 1619-1624, 1993.
30. Ahuja, H-, Bar-Eli, M., Advani, S. H., Benchimol, S., and Cline, M. J. Alteration inthe p53 gene and clonal evolution of the blast crisis of chronic myelocytic leukemia.Proc. Nati. Acad. Sci. USA, 86: 6787-6787, 1989.
31. Luca, M., Hunt, B., Bucana, C. D., Johnson, J. P., Fidler, I. J., and Bar-Eli. M. Directcorrelation between MUC18 expression and metastatic potential of human melanomacells. Melanoma Res., 3: 35-41, 1993.
32. Gutman, M., Singh, R. K., Radinsky, R., and Bar-Eli, M. Intertumoral heterogeneityof receptor-tyrosine kinases expression in human melanoma cell lines with differentmetastatic capabilities. Anticancer Res., 14: 1759-1766, 1944.
33. Koch, A. E., Polverini, P. J., Kunkel, S. L., Harlow, L. A., DiPietro, L. A., Einer,V. M., Einer, S. G., and Stricter, R. M. Interleukin-8 as a macrophage-derivedmediator of angiogenesis. Science (Washington DC), 258.- 1798-1801, 1992.
34. Kamb, A., Gruis, N. A., Weaver-Feldhaus, J., Liu, Q., Harshman, K., Tavtigian, S. V.,Stocken, E., Day, R. S., Ill, Johnson, B. E., and Skolnick, M. H. A cell cycle regulatorpotentially involved in genesis of many tumor types. Science (Washington DC), 264:436-440, 1994.
35. Nobori, T., Miura, K., Wu, D. J., Lois, A., Takabayashi, K., and Carson, D. A.Deletions of the cyclin-dependent kinase-4 inhibitor gene in multiple human cancers.Nature (Lond.), 368: 753-756, 1994.
36. Barker, J. N. W. N., Sarma, V., Mitra, R. S„Dixit, V. M., and Nickoloff, B. S.Marked synergism between tumor necrosis factor and Interferon gamma in regulationof keratinocyte derived adhesion molecule and chemotactic factors. J. Clin. Invest.,85: 605-608, 1990.
37. Matsushima, K., and Oppenheim, J. J. Interleukin-8 and MCAF: novel inflammatorycytokines inducible by IL-1 and TNF. Cytokine, /: 2-13, 1989.
38. Mukaido, N., Mahe, Y., and Matsushima, K. Cooperative interaction of NF-kB-andC/EBP-like factor binding elements in activating the interleukin-8 gene by proin-flammatory cytokines. J. Biol. Chem., 265.- 21128-21133, 1990.
39. Angel, P., and Karin, M. The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochem. Biophys. Acta, 1072: 129—157,1991.
40. Suzuki, T., Murakami, M., Onai, N., Fukuda, E., Hashimoto, Y., Sonobe, M. H.,Kameda, T., Ichinose, M., Micki, K., and Iba, H. Analysis of AP-1 function in cellulartransformation pathway. J. Virol., 68: 3527-3535, 1994.
41. Shah, G., Ghosh, R., Amstad, P. A., and Cerutti, P. A. Mechanism of induction ofc-fos by ultraviolet B (290-320 nm) in mouse JB6 epidermal cells. Cancer Res., 53:38-45, 1993.
42. Wang, J. M., Taraboletti, G., Matsushima, K., Damme, J. V.. and Mantovani, A.Induction of hapatotactic migration of melanoma cells by neutrophil activatingprotein/IL-8. Biochem. Biophys. Res. Commun., 169: 165-170, 1990.
43. Sozzani, S., Azwu, D. E., Ellenburg, M. D., Locati, M., Rieppi, M., Rojas, A.,Montovani, A., and McPhail, L. C. Activation of phospholipase D by interleukin-8 inhuman neutrophils. Blood, 84: 3895-3901, 1994.
44. Reich, R., Blumenthal, M., and Liscovitch, M. Role of phospholipase D in laminin-induced production of gelatinase A(MMP-2) in metastatic cells. Clin. Exp. Metastasis, 13: 134-140, 1995.
45. Ziegler, A., Jonason, A. S., Leffell, D. J., Simon, A. J., Sharma, H. W., Kimmelman,J., Remington, L., Jacks, T., and Brash, D. E. Sunburn and p53 in the onset of skincancer. Nature, (Lond.), 372: 773-776, 1994.
3674




















