
A neuroprotective phase precedes striatal degeneration upon nucleolar stress

B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Ultrastructural relationship between the mu opioid receptorand its interacting protein, GPR177, in striatal neurons
Arith-Ruth S. Reyesa, Robert Levensonb, Wade Berrettinic, Elisabeth J. Van Bockstaelea,⁎aDepartment of Neuroscience, Farber Institute for Neurosciences, Thomas Jefferson University, Philadelphia, PA 19107, USAbDepartment of Pharmacology, Penn State College of Medicine, Hershey, PA 17033, USAcDepartment of Psychiatry, Center for Neurobiology and Behavior, University of Pennsylvania School of Medicine, Philadelphia,PA 19104, USA
A R T I C L E I N F O
⁎ Corresponding author. Department of NeuroSuite 400, Philadelphia, PA 19107, USA. Fax:
E-mail address: [email protected] (E.J.
0006-8993/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainres.2010.08.080
A B S T R A C T
Article history:Accepted 24 August 2010Available online 21 September 2010
GPR177, the mammalian ortholog of Drosophila Wntless/Evi/Sprinter, was recentlyidentified as a novel mu-opioid receptor (MOR) interacting protein. GPR177 is a trans-membrane protein pivotal to mediating the secretion of Wnt signaling proteins. Wntproteins, in turn, are essential in regulating neuronal development, a phenomenoninhibited upon chronic exposure to MOR agonists such as morphine and heroin. Wepreviously showed that GPR177 and MOR are co-localized in the mouse dorsolateralstriatum; however, the nature of this interaction was not fully elucidated. Therefore,in the present study, we examined cellular substrates for interactions between GPR177and MOR using a combined immunogold-silver and peroxidase detection approach incoronal sections in the dorsolateral segment of the striatum. Semi-quantitativeanalysis of the ultrastructural distribution of GPR177 and MOR in striatal somata andin dendritic processes showed that, of the somata and dendritic processes exhibitingGPR177, 32% contained MOR immunolabeling while for profiles exhibiting MOR, 37%also contained GPR177 immunoreactivity. GPR177-labeled particles were localizedpredominantly along both the plasma membrane and within the cytoplasm of MOR-labeled dendrites. Somata and dendritic processes that contained both GPR177 andMOR more often received symmetric (inhibitory-type) synapses from unlabeled axonterminals. To further define the phenotype of GPR177 and MOR-containing cellularprofiles, triple immunofluorescence detection showed that GPR177 and MOR arelocalized in neurons containing the opioid peptide, enkephalin, within thedorsolateral striatum. The results provide an anatomical substrate for interactionsbetween MOR and its interacting protein, GPR177, in striatal opioid-containingneurons that may underlie the morphological alterations produced in neurons bychronic opiate use.
© 2010 Elsevier B.V. All rights reserved.
surgery, Farber Institute for Neurosciences, Thomas Jefferson University, 900 Walnut Street,+1 215 503 9238.Van Bockstaele).
er B.V. All rights reserved.
Keywords:GPR177Mu opioid receptorStriatumElectron microscopyConfocal microscopy

72 B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
1. Introduction
Dependence and addiction to opiate drugs (e.g. morphine andheroin) involve the activation of G-protein coupled opioidreceptors (Contet et al., 2004). Specifically, the activation ofmu-opioid receptors (MOR) results in adaptations that oftenlead to tolerance and dependence (Bailey and Connor, 2005).Along with the analgesic and rewarding effects of opiatesprimarily mediated by MOR (Kieffer, 1999), the addictive stateis characterized by inhibitory effects of opiates on axonoutgrowth, dendritic arborization, and neurogenesis in brainregions implicated in reward processing, learning, and mem-ory (Robinson and Kolb, 1999; Eisch et al., 2000). Although therole of MOR in contributing to neuronal changes observed inopioid dependence remains complex, a growing literature onthe multiple interacting proteins involved in the regulation ofMOR appears promising.
GPR177, the mammalian ortholog of Drosophila Wntless/Evi/Sprinter, was recently identified as a MOR interactingprotein that may possibly serve as a substrate underlying thealterations in neuronal structure and synaptic organizationcharacteristics of opioid dependence (Jin et al., 2010). GPR177 isa seven-transmembrane protein essential in mediating thesecretion of Wnt signaling proteins (Banziger et al., 2006;Bartscherer et al., 2006).Wnt proteins are, in turn, extracellularsignaling molecules important for their neurotrophic proper-ties (Ciani and Salinas, 2005; Banziger et al., 2006). Among theirother functions in neuronal development, Wnt proteinsstimulate synapse formation and dendritic morphogenesis,and control axon remodeling and guidance (Ciani and Salinas,2005). GPR177 accompanies these Wnt proteins when itshuttles between the Golgi apparatus and the plasma mem-brane (Franch-Marro et al., 2008). It is precisely in thismechanism of secreting Wnt proteins that the interactionbetween GPR177 and MOR may potentially have an effect (Jinet al., 2010).
Known for its role inmotor control and as a key structure inthe formation of motor-related memories (Albin et al., 1989),the striatum is also involved in cognitive processes includingexecutive function (Kehagia et al., 2010; Watson and Stanton,2009). The striatum receives innervation from many brainregions including dopaminergic fibers from the pars compactaof the substantia nigra and glutamatergic innervation arisingfrom cortical areas conveying sensorimotor, limbic andcognitive information (Graybiel, 1990; Calabresi et al., 1996).The striatal neuronal population is enriched with variouspeptidergic neurons including enkephalinergic, cholinergic,GABAergic and aminergic neurons (Fonnum et al., 1977;Beckstead and Kersey, 1985; Graybiel, 1990; Koos et al., 2004;Taverna et al., 2007).
Our recent studies have demonstrated that GPR177 andMOR are co-localized in striatal perikarya and dendriticprocesses (Jin et al., 2010); thus, we have set out to elucidatethe cellular substrates underlying interactions betweenGPR177 and MOR and define their phenotype in the striatum.In an effort to further characterize the interaction betweenGPR177 and MOR, we semiquantitatively analyzed data fromdual electron microscopic immunocytochemical detection ofGPR177 and MOR. Furthermore, this study used triple light
microscopic immunocytochemical labeling for GPR177, MORand enkephalin (ENK).
2. Results
2.1. GPR177 localization in MOR-containing cells inthe striatum
Using immunofluorescence microscopy, GPR177 immuno-reactivity was localized in striatal somata and in dendriticprocesses (Fig. 1) consistent with our recent report (Jin et al.,2010). Using ultrastructural analysis, GPR177 was visualized asirregularly shaped black deposits (Jin et al., 2010) indicative ofimmunogold-silver particles (Fig. 2). Dual electronmicroscopicimmunolabeling where GPR177 was visualized as immuno-gold-silver particles and MOR was identified by an electron-dense peroxidase reaction product showed that GPR177 waslocalized (1) along the plasma membrane only (Fig. 2A), (2)within the cytoplasmic compartment (Fig. 2B), and (3) alongthe plasma membrane as well as within the cytoplasmiccompartment (Fig. 2C). In many instances, GPR177 waslocalized both within the cytoplasm and along the plasmamembrane (Figs. 2C and 4). Quantification for the distributionof GPR177 immunoreactivity in MOR-labeled dendritesshowed that 37% of these profiles exhibited GPR177 immuno-reactivity within the cytoplasmic compartment only and 11%of these profiles exhibited GPR177 along the plasma mem-brane only. The remaining 52% of the MOR-labeled profilesexhibited GPR177 immunoreactivity both within the cytoplas-mic compartment and along the plasma membrane (Fig. 2E).Of these profiles, however, there were more GPR177 in thecytoplasm as revealed by the higher percentage of immuno-gold-silver particles in the cytoplasm when compared to thetotal number of particles in a profile (66%).
Along with the ratio of cytoplasmic to total immunogold-silver particles, the number of immunogold-silver particlesper micron of plasmalemma and the number of immunogold-silver particles per micron squared of cytoplasm were alsodetermined as presented in Fig. 3. Statistical analysis showedthat these associations did not significantly vary betweenlarge- (>5 μm in perimeter) and small- (<5 μm in perimeter)sized dendrites. Additionally, our results showed that theimmunogold-silver particles were associated in a number oforganelles within the cytoplasmic compartment. For thispurpose, we did not intend to classify the size of dendriticprofiles examined as our earlier results showed that the size ofdendrites did not influence various criteria evaluated. Immu-nogold-silver particles were often associated with endosome-like vesicles (30%) and Golgi apparatus (12%). Some immuno-gold-silver particles were also associated with endoplasmicreticulum and multivesicular bodies while some were associ-ated with organelles that could not be clearly classified.
2.2. GPR177 interactions with MOR in the striatum
The results of the immunofluorescence studieswere confirmedby the ultrastructural results where GPR177 was visualized byimmunogold-silver detection and MOR was identified using anelectron-dense peroxidase reaction product. Results from the
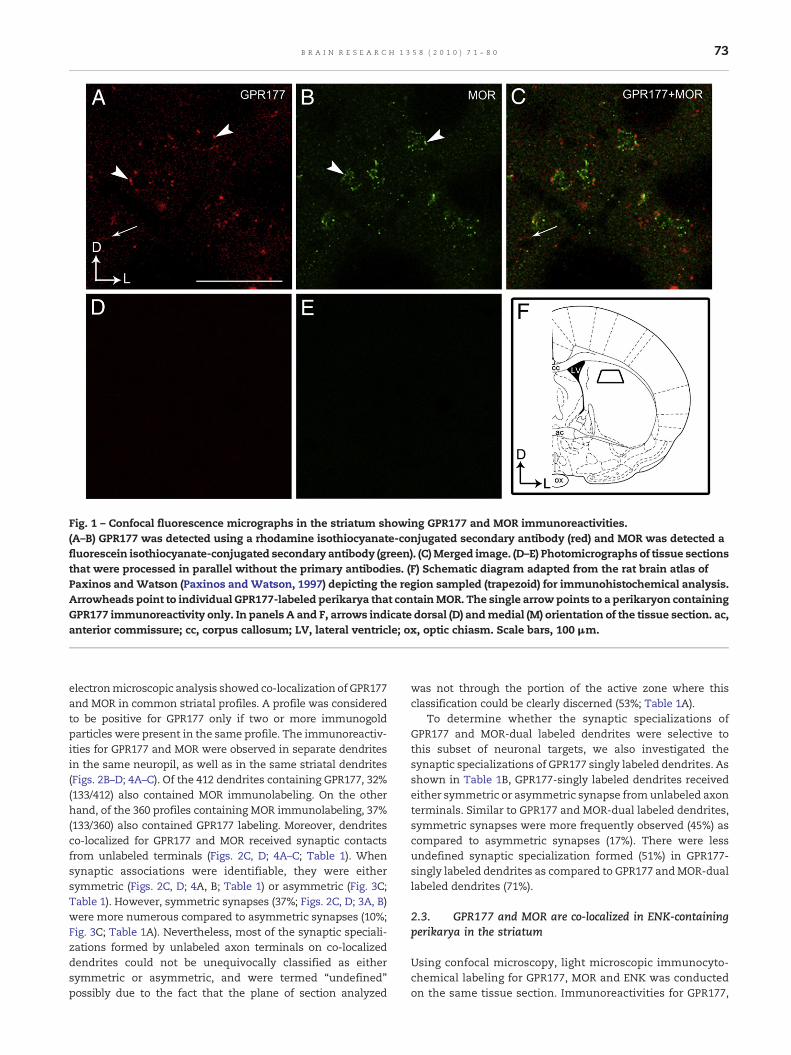
Fig. 1 – Confocal fluorescence micrographs in the striatum showing GPR177 and MOR immunoreactivities.(A–B) GPR177 was detected using a rhodamine isothiocyanate-conjugated secondary antibody (red) and MOR was detected afluorescein isothiocyanate-conjugated secondary antibody (green). (C)Merged image. (D–E) Photomicrographs of tissue sectionsthat were processed in parallel without the primary antibodies. (F) Schematic diagram adapted from the rat brain atlas ofPaxinos and Watson (Paxinos and Watson, 1997) depicting the region sampled (trapezoid) for immunohistochemical analysis.Arrowheads point to individual GPR177-labeled perikarya that containMOR. The single arrow points to a perikaryon containingGPR177 immunoreactivity only. In panels A and F, arrows indicate dorsal (D) andmedial (M) orientation of the tissue section. ac,anterior commissure; cc, corpus callosum; LV, lateral ventricle; ox, optic chiasm. Scale bars, 100 μm.
73B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
electronmicroscopic analysis showed co-localization of GPR177and MOR in common striatal profiles. A profile was consideredto be positive for GPR177 only if two or more immunogoldparticles were present in the same profile. The immunoreactiv-ities for GPR177 and MOR were observed in separate dendritesin the same neuropil, as well as in the same striatal dendrites(Figs. 2B–D; 4A–C). Of the 412 dendrites containing GPR177, 32%(133/412) also contained MOR immunolabeling. On the otherhand, of the 360 profiles containing MOR immunolabeling, 37%(133/360) also contained GPR177 labeling. Moreover, dendritesco-localized for GPR177 and MOR received synaptic contactsfrom unlabeled terminals (Figs. 2C, D; 4A–C; Table 1). Whensynaptic associations were identifiable, they were eithersymmetric (Figs. 2C, D; 4A, B; Table 1) or asymmetric (Fig. 3C;Table 1). However, symmetric synapses (37%; Figs. 2C, D; 3A, B)were more numerous compared to asymmetric synapses (10%;Fig. 3C; Table 1A). Nevertheless, most of the synaptic speciali-zations formed by unlabeled axon terminals on co-localizeddendrites could not be unequivocally classified as eithersymmetric or asymmetric, and were termed “undefined”possibly due to the fact that the plane of section analyzed
was not through the portion of the active zone where thisclassification could be clearly discerned (53%; Table 1A).
To determine whether the synaptic specializations ofGPR177 and MOR-dual labeled dendrites were selective tothis subset of neuronal targets, we also investigated thesynaptic specializations of GPR177 singly labeled dendrites. Asshown in Table 1B, GPR177-singly labeled dendrites receivedeither symmetric or asymmetric synapse fromunlabeled axonterminals. Similar to GPR177 and MOR-dual labeled dendrites,symmetric synapses were more frequently observed (45%) ascompared to asymmetric synapses (17%). There were lessundefined synaptic specialization formed (51%) in GPR177-singly labeled dendrites as compared to GPR177 andMOR-duallabeled dendrites (71%).
2.3. GPR177 and MOR are co-localized in ENK-containingperikarya in the striatum
Using confocal microscopy, light microscopic immunocyto-chemical labeling for GPR177, MOR and ENK was conductedon the same tissue section. Immunoreactivities for GPR177,
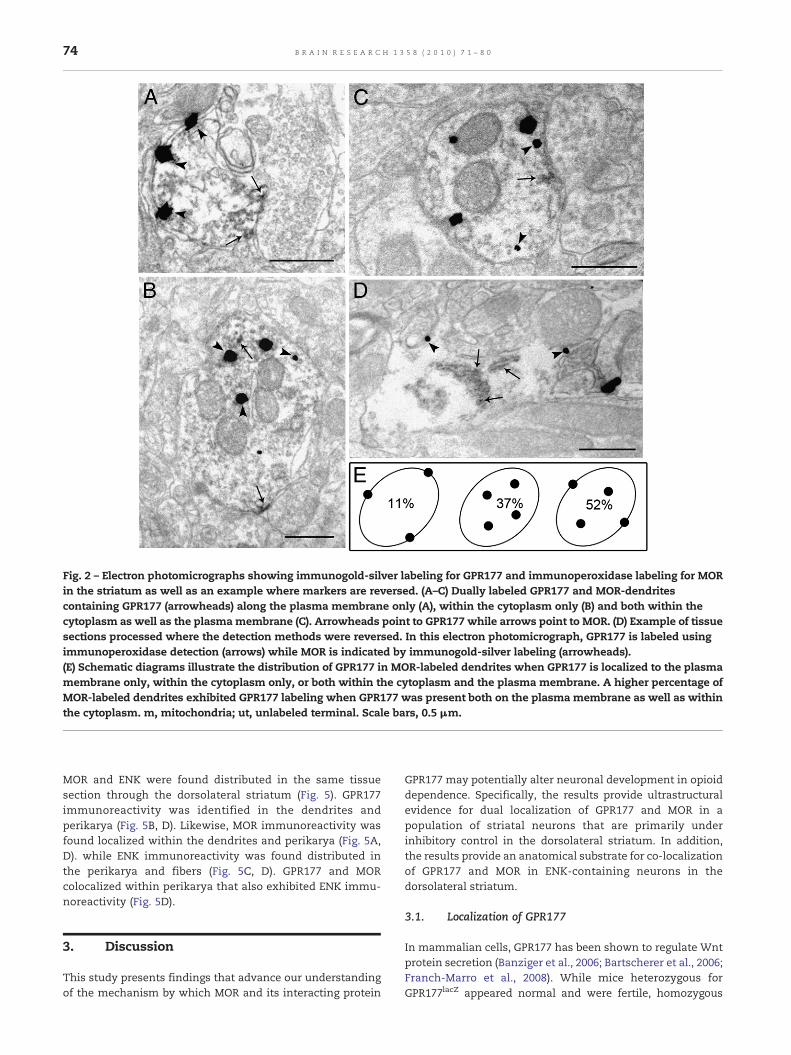
Fig. 2 – Electron photomicrographs showing immunogold-silver labeling for GPR177 and immunoperoxidase labeling for MORin the striatum as well as an example where markers are reversed. (A–C) Dually labeled GPR177 and MOR-dendritescontaining GPR177 (arrowheads) along the plasma membrane only (A), within the cytoplasm only (B) and both within thecytoplasm as well as the plasmamembrane (C). Arrowheads point to GPR177 while arrows point to MOR. (D) Example of tissuesections processed where the detection methods were reversed. In this electron photomicrograph, GPR177 is labeled usingimmunoperoxidase detection (arrows) while MOR is indicated by immunogold-silver labeling (arrowheads).(E) Schematic diagrams illustrate the distribution of GPR177 in MOR-labeled dendrites when GPR177 is localized to the plasmamembrane only, within the cytoplasm only, or both within the cytoplasm and the plasma membrane. A higher percentage ofMOR-labeled dendrites exhibited GPR177 labeling when GPR177 was present both on the plasma membrane as well as withinthe cytoplasm. m, mitochondria; ut, unlabeled terminal. Scale bars, 0.5 μm.
74 B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
MOR and ENK were found distributed in the same tissuesection through the dorsolateral striatum (Fig. 5). GPR177immunoreactivity was identified in the dendrites andperikarya (Fig. 5B, D). Likewise, MOR immunoreactivity wasfound localized within the dendrites and perikarya (Fig. 5A,D). while ENK immunoreactivity was found distributed inthe perikarya and fibers (Fig. 5C, D). GPR177 and MORcolocalized within perikarya that also exhibited ENK immu-noreactivity (Fig. 5D).
3. Discussion
This study presents findings that advance our understandingof the mechanism by which MOR and its interacting protein
GPR177 may potentially alter neuronal development in opioiddependence. Specifically, the results provide ultrastructuralevidence for dual localization of GPR177 and MOR in apopulation of striatal neurons that are primarily underinhibitory control in the dorsolateral striatum. In addition,the results provide an anatomical substrate for co-localizationof GPR177 and MOR in ENK-containing neurons in thedorsolateral striatum.
3.1. Localization of GPR177
In mammalian cells, GPR177 has been shown to regulate Wntprotein secretion (Banziger et al., 2006; Bartscherer et al., 2006;Franch-Marro et al., 2008). While mice heterozygous forGPR177lacZ appeared normal and were fertile, homozygous
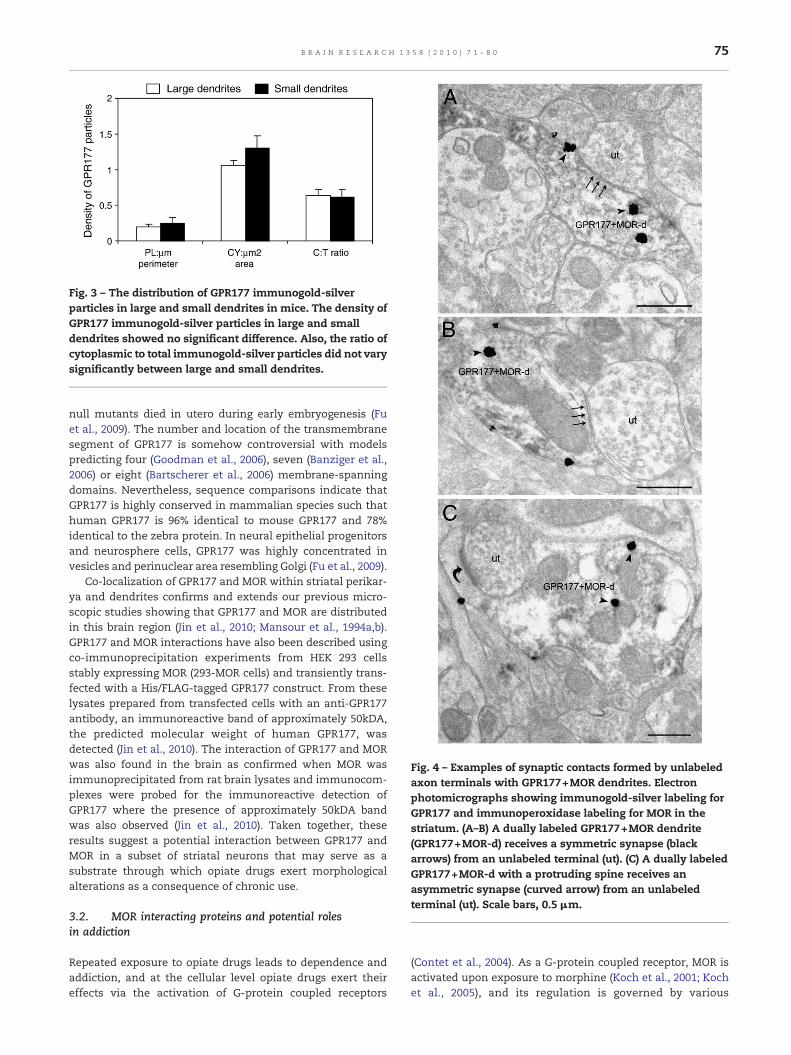
Fig. 3 – The distribution of GPR177 immunogold-silverparticles in large and small dendrites in mice. The density ofGPR177 immunogold-silver particles in large and smalldendrites showed no significant difference. Also, the ratio ofcytoplasmic to total immunogold-silver particles did not varysignificantly between large and small dendrites.
Fig. 4 – Examples of synaptic contacts formed by unlabeledaxon terminals with GPR177+MOR dendrites. Electronphotomicrographs showing immunogold-silver labeling forGPR177 and immunoperoxidase labeling for MOR in thestriatum. (A–B) A dually labeled GPR177+MOR dendrite(GPR177+MOR-d) receives a symmetric synapse (blackarrows) from an unlabeled terminal (ut). (C) A dually labeledGPR177+MOR-d with a protruding spine receives anasymmetric synapse (curved arrow) from an unlabeledterminal (ut). Scale bars, 0.5 μm.
75B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
null mutants died in utero during early embryogenesis (Fuet al., 2009). The number and location of the transmembranesegment of GPR177 is somehow controversial with modelspredicting four (Goodman et al., 2006), seven (Banziger et al.,2006) or eight (Bartscherer et al., 2006) membrane-spanningdomains. Nevertheless, sequence comparisons indicate thatGPR177 is highly conserved in mammalian species such thathuman GPR177 is 96% identical to mouse GPR177 and 78%identical to the zebra protein. In neural epithelial progenitorsand neurosphere cells, GPR177 was highly concentrated invesicles and perinuclear area resembling Golgi (Fu et al., 2009).
Co-localization of GPR177 and MOR within striatal perikar-ya and dendrites confirms and extends our previous micro-scopic studies showing that GPR177 and MOR are distributedin this brain region (Jin et al., 2010; Mansour et al., 1994a,b).GPR177 and MOR interactions have also been described usingco-immunoprecipitation experiments from HEK 293 cellsstably expressing MOR (293-MOR cells) and transiently trans-fected with a His/FLAG-tagged GPR177 construct. From theselysates prepared from transfected cells with an anti-GPR177antibody, an immunoreactive band of approximately 50kDA,the predicted molecular weight of human GPR177, wasdetected (Jin et al., 2010). The interaction of GPR177 and MORwas also found in the brain as confirmed when MOR wasimmunoprecipitated from rat brain lysates and immunocom-plexes were probed for the immunoreactive detection ofGPR177 where the presence of approximately 50kDA bandwas also observed (Jin et al., 2010). Taken together, theseresults suggest a potential interaction between GPR177 andMOR in a subset of striatal neurons that may serve as asubstrate through which opiate drugs exert morphologicalalterations as a consequence of chronic use.
3.2. MOR interacting proteins and potential rolesin addiction
Repeated exposure to opiate drugs leads to dependence andaddiction, and at the cellular level opiate drugs exert theireffects via the activation of G-protein coupled receptors
(Contet et al., 2004). As a G-protein coupled receptor, MOR isactivated upon exposure to morphine (Koch et al., 2001; Kochet al., 2005), and its regulation is governed by various
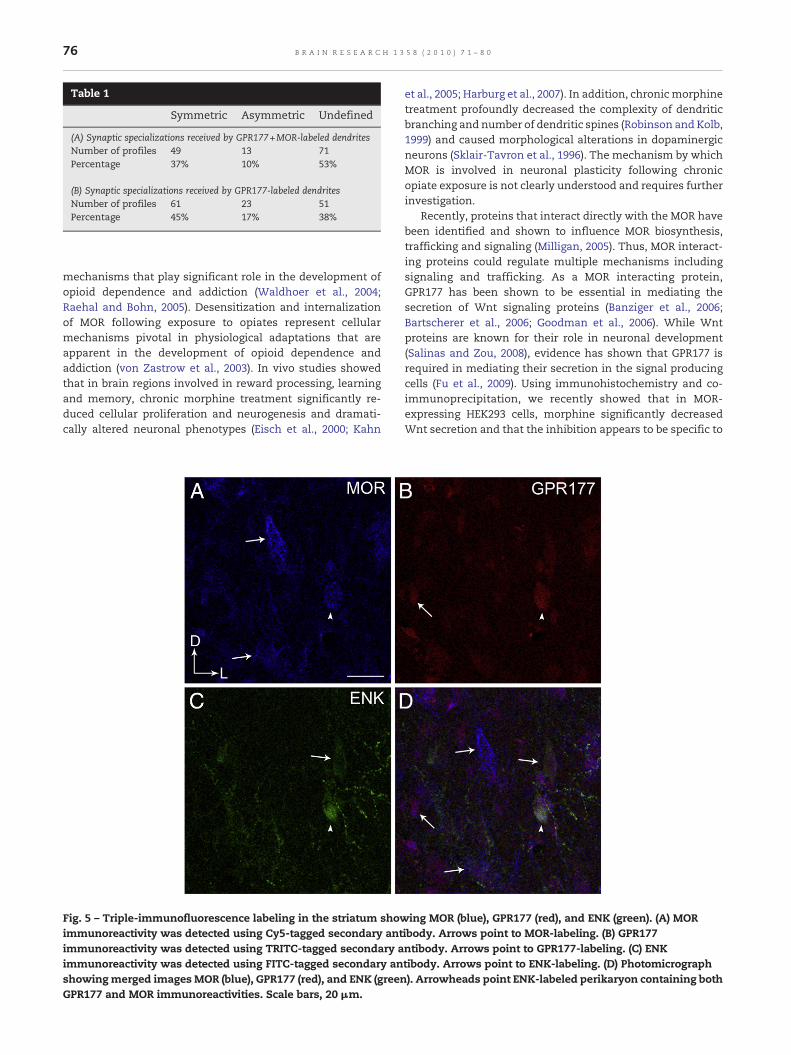
Table 1
Symmetric Asymmetric Undefined
(A) Synaptic specializations received by GPR177+MOR-labeled dendritesNumber of profiles 49 13 71Percentage 37% 10% 53%
(B) Synaptic specializations received by GPR177-labeled dendritesNumber of profiles 61 23 51Percentage 45% 17% 38%
76 B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
mechanisms that play significant role in the development ofopioid dependence and addiction (Waldhoer et al., 2004;Raehal and Bohn, 2005). Desensitization and internalizationof MOR following exposure to opiates represent cellularmechanisms pivotal in physiological adaptations that areapparent in the development of opioid dependence andaddiction (von Zastrow et al., 2003). In vivo studies showedthat in brain regions involved in reward processing, learningand memory, chronic morphine treatment significantly re-duced cellular proliferation and neurogenesis and dramati-cally altered neuronal phenotypes (Eisch et al., 2000; Kahn
Fig. 5 – Triple-immunofluorescence labeling in the striatum showimmunoreactivity was detected using Cy5-tagged secondary antimmunoreactivity was detected using TRITC-tagged secondary aimmunoreactivity was detected using FITC-tagged secondary anshowingmerged imagesMOR (blue), GPR177 (red), and ENK (greenGPR177 and MOR immunoreactivities. Scale bars, 20 μm.
et al., 2005; Harburg et al., 2007). In addition, chronicmorphinetreatment profoundly decreased the complexity of dendriticbranching and number of dendritic spines (Robinson and Kolb,1999) and caused morphological alterations in dopaminergicneurons (Sklair-Tavron et al., 1996). The mechanism by whichMOR is involved in neuronal plasticity following chronicopiate exposure is not clearly understood and requires furtherinvestigation.
Recently, proteins that interact directly with the MOR havebeen identified and shown to influence MOR biosynthesis,trafficking and signaling (Milligan, 2005). Thus, MOR interact-ing proteins could regulate multiple mechanisms includingsignaling and trafficking. As a MOR interacting protein,GPR177 has been shown to be essential in mediating thesecretion of Wnt signaling proteins (Banziger et al., 2006;Bartscherer et al., 2006; Goodman et al., 2006). While Wntproteins are known for their role in neuronal development(Salinas and Zou, 2008), evidence has shown that GPR177 isrequired in mediating their secretion in the signal producingcells (Fu et al., 2009). Using immunohistochemistry and co-immunoprecipitation, we recently showed that in MOR-expressing HEK293 cells, morphine significantly decreasedWnt secretion and that the inhibition appears to be specific to
ing MOR (blue), GPR177 (red), and ENK (green). (A) MORibody. Arrows point to MOR-labeling. (B) GPR177ntibody. Arrows point to GPR177-labeling. (C) ENKtibody. Arrows point to ENK-labeling. (D) Photomicrograph). Arrowheads point ENK-labeled perikaryon containing both

77B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
the activation of MORs considering that DAMGO treatmentdid not affect Wnt secretion from 293-MOR cells (Jin et al.,2010). Interestingly, the morphine-mediated inhibition ofWnt2 secretion was reversed by overexpression of GPR177,indicating that while morphine specifically blocks Wntsecretion from MOR expressing cells, this inhibition can beovercome by overexpression of GPR177 (Jin et al., 2010).
In the present study, dendritic processes that co-localizedGPR177 andMOR exhibited differential patterns of localizationwithin individual profiles and frequently received heteroge-neous synapses from unlabeled axon terminals. GPR177immunoreactivity was localized in MOR-labeled perikarya aswell as in MOR-labeled dendrites (Jin et al., 2010). Withindendrites, GPR177 immunoreactivity was distributed withinthe cytoplasm and along the plasma membrane of MOR-labeled dendrites. A higher percentage of profiles exhibitingMOR and GPR177 showed that the distribution of GPR177 wasmore common within the cytoplasm and along the plasmamembrane as opposed to being restricted to any of theseindividual cellular compartments.
Unlabeled axon terminals formed symmetric, asymmetricor “undefined” synapses (e.g. did not form recognizablejunctions in the plane of section examined) with GPR177 andMOR-labeled dendrites. Semi-quantitative analysis showed apreponderance of symmetric synapses with GPR177 andMOR-labeled dendrites. While asymmetric synapses have beenassociated with excitatory transmission, symmetric synapseshave been associatedwith inhibitory transmission (Gray, 1959;Peters et al., 1991). Electrophysiological studies have shownthat mu selective opioid peptides act to post-synapticallyinhibit striatal neurons (Pickel et al., 1992). The striatumis known to be enriched with GABA-containing neurons(Graveland and DiFiglia, 1985; Gerfen and Wilson, 1996) andthese GABAergic neurons serve as the primary source ofsynaptic inhibition in the striatum (Koos et al., 2004; Tavernaet al., 2007). Likewise, ENK-containing neurons are known tobe localized in the striatal neurons (Pickel et al., 1992). Thesedata suggest that unlabeled axon terminals targeting GPR177and MOR-labeled dendrites may represent inhibitory (ENK-or GABA) axon terminals that may hyperpolarize striatalneuronal activity.
3.3. Functional implications
Since striatal neurons have been shown to contain multiplepeptidergic and aminergic neuromodulators (Fonnum et al.,1977; Graybiel, 1990; Chen et al., 2008), we sought to determinethe phenotype of striatal neurons containing both GPR177 andMOR. Our triple immunofluorescence data show that coloca-lization of GPR177 and MOR is contained in enkephalinergicindividual striatal neurons. Our data extend previous electronmicroscopic studies that GPR177 (Jin et al., 2010), MOR (Wanget al., 1996, 1999) and ENK (Pickel et al., 1992) are contained instriatal neurons that have cortical connections critical forcomplex cognitive functions (Alexander et al., 1986). ENK isknown to regulate functions of striatal neurons such thatperturbations in ENK expression levels reflect alterations instriatal neuronal activity (Steiner and Gerfen, 1999; Parelkarand Wang, 2003). It has been reported that within thestriatum, the cortically derived information is transformed
where representations of motor and cognitive actionsequences can be integrated into more complex behavioralresponses (Graybiel, 1998) emphasizing that in addition to therole of striatum in motor control, the striatum plays anessential role in the function of more complex behaviorsincluding decision making and executive function (Graybiel,2008). It is likely that following exposure to morphine, GPR177and MOR interaction within the enkephalinergic striatalneurons contributes to the mechanism underlying changesin the brain that induce adverse effects on brain functionincluding cognitive impairment, affective disturbances andaddiction (Skaer, 2004).
In summary, using immunofluorescence and immunoe-lectron microscopy, we provide cellular evidence suggestingpotential mechanisms by which GPR177 and MOR mayinteract via enkephalinergic neurons to affect activity ofstriatal neurons following morphine exposure. Interactionsbetween GPR177 and MOR may underlie the morphologicalconsequences accompanying chronic opiate use (e.g., shrunk-en neurons, decreased neurogenesis, loss of dendritic spines)that may result in impaired cognition, affective disturbancesand addiction. Future studies examining whether interferingwith this interaction leads to abrogating morphologicalalterations following chronic opiate use are required toadvance our understanding of the consequences of chronicopiate exposure.
4. Experimental procedures
4.1. Animals
Six three-month old male naive C57/BL/6 wild type mice(20–25 g; Taconic, Albany, NY) were used. The animal proce-dures used in this study were approved by the InstitutionalAnimal Care and Use Committee at Thomas Jefferson Univer-sity and conformed to theNational Institutes of Health Guide for theCare andUse of LaboratoryAnimals.Micewerehoused2–3per cageon a 12-h light schedule (lights on at 0700) in a temperature-controlled (20 °C) colony room. They were allowed ad libitumaccess to standard chow and water. All efforts were made tominimize animal distress and the number of animals used.
4.2. Specificity of antisera
Anti-GPR177 antibody was generated in chickens against apeptide antigen corresponding to theC-terminal 18 amino acids(HVDGPTEIYKLTRKEAQE) of human GPR177 (Gene-Tel Labora-tories, Madison, WI), which is identical to the rat and mousepeptide sequence. Antibodies of the IgY subtypewereharvestedfrom egg yolks and affinity purified prior to use. The character-ization and specificity of the chicken antiserum against GPR177has been previously described (Jin et al., 2010). Western blotanalysis using GPR177 antibody showed expression of theGPR177 in rat brain lysates indicating that GPR177 antibodyrecognized endogenous GPR177 in brain tissue. GPR177 recog-nized single band of proteins with approximate molecularweight of about 50 kDa (Jin et al., 2010). Anti-MOR antibody wasgenerated in rabbit against a glutaraldehyde conjugatecorresponding to the C-terminal 18 amino acids of rat MOR

78 B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
(Surratt et al., 1994; Van Bockstaele et al., 1996a,b). Throughimmunodot-blot analysis, the MOR antiserum was shown tospecifically recognize amino acid sequences of MOR, but notamino acid sequences of either delta- or kappa-opioid receptors(Cheng et al., 1996).
4.3. Immunofluorescence
Three adult mice were deeply anesthetized with sodiumpentobarbital (80 mg/kg) and eachwas transcardially perfusedthrough the ascending aorta with 100 ml of 4% formaldehydein 0.1 M phosphate buffer (PB; pH 7.4). Brains were thenremoved and post fixed in 4% formaldehyde overnight at 4 °C.Sections (40 μm) through the rostrocaudal extent of striatum(Paxinos and Watson, 1997) were cut using a Vibratome(Technical Product International, St Louis, MO, USA) andrinsed extensively in 0.1 M PB and 0.1 M tris-buffered saline(TBS; pH 7.6). Sections were placed for 30 min in 1% sodiumborohydride in 0.1 M PB to reduce amine-aldehyde com-pounds. Tissue sections were rinsed in 0.1 M PB and 0.1 MTBS. Sections were then incubated in 0.5% bovine serumalbumin (BSA) and 0.25% Triton X-100 in 0.1 M TBS for 30 minand rinsed thoroughly in 0.1 M TBS. Following rinses, tissuesections were incubated in a cocktail of chicken anti-GPR177antiserum (1:1000), rabbit anti-MOR (1:2000; Chemicon, Teme-cula, CA) and mouse anti-leucine5-ENK (1:100; FitzgeraldLaboratories, Concord, MA) in 0.5% BSA and 0.25% TritonX-100 in 0.1 M TBS. Sections were incubated at room tempe-rature for 15–18 h in a rotary shaker. Following primaryantibody incubation, sections were washed in 0.1 M TBS andincubated in a secondary antibody cocktail containing tetra-methyl rhodamine isothiocyanate (TRITC) donkey anti-chick-en (1:200; Jackson ImmunoResearch Laboratories Inc., WestGrove, PA, USA), Cy5 donkey anti-rabbit (1:200; JacksonImmunoResearch) and fluorescein isothiocyanate (FITC)donkey-anti-mouse (1:200; Jackson ImmunoResearch) antibo-dies prepared in 0.1% BSA and 0.25% Triton X-100 in 0.1 MTBS for 2 h in the dark on a rotary shaker. The tissue sectionswere washed thoroughly in 0.1 M TBS and 0.1 M PB thenmounted on slides, allowed to dry in the dark. The slides weredehydrated in a series of alcohols, soaked in xyleneand coverslipped using DPX (Sigma-Aldrich Inc., St. Louis,MO, USA). Immunofluorescence labeling on tissue sectionswas visualized using a Zeiss LSM510Meta confocalmicroscope(Carl Zeiss Inc., Thornwood, NY, USA). Digital images ofimmunofluorescent labeling were captured and importedusing the LSM 5 image browser (Carl Zeiss Inc.). Figures wereassembled and adjusted for brightness and contrast in AdobePhotoshop.
4.4. Immunoelectron microscopy
Three adult mice were used for immunoelectronmicroscopy.Methods for perfusion, sectioning, and immunohistochem-ical labeling of tissue sections intended for electron micros-copy have been described previously (Jin et al., 2010; VanBockstaele et al., 1996a,b). Briefly, mice were deeply anes-thetized with sodium pentobarbital (80 mg/kg) and perfusedtranscardially with 10 ml heparinized saline followed with
25 ml of 3.75% acrolein (Electron Microscopy Sciences, FortWashington, PA, USA), and 50 ml of 2% formaldehyde in0.1 M PB (pH 7.4). The brains were removed immediately,sectioned into 1–3 mm coronal slices and postfixed in thesame fixative overnight at 4 °C. Sections (40 μm) wereprocessed for electron microscopic analysis of GPR177 andMOR in the same section. Sections containing the striatumwere processed following the protocol described earlier forimmunofluorescence except that Triton X-100 was not addedto the solution for antibody incubation. Tissue sections wereincubated in a cocktail of chicken anti-GPR177 antiserum(1:1000) and rabbit anti-MOR (1:2000; Chemicon). The follow-ing day, tissue sections were extensively rinsed threetimes in 0.1 M TBS and incubated in biotinylated donkeyanti-rabbit (1:400; Jackson ImmunoResearch, West Grove,PA). Subsequently, a 30-minute incubation of avidin-biotincomplex (Vector Laboratories, Burlingame, CA) followed. Forall incubations and washes, sections were continuouslyagitated with a rotary shaker. MOR was visualized by a4-min reaction in 22 mg of 3,3′-diaminobenzidine (DAB;Sigma-Aldrich Inc., St. Louis, MO) and 10 μl of 30% hydrogenperoxide in 100 ml TBS. Sections were then rinsed threetimes with 0.1 M TBS, followed by rinses with 0.1 MPB and0.01 M phosphate-buffered saline (PBS; pH 7.4). Sections wereincubated in a 0.2% gelatin-PBS and 0.8% BSA buffer for10 min and followed by incubation in ultrasmall (<1 nm)gold-coupled goat anti-chicken (1:100 dilution; ElectronMicroscopy Sciences) at room temperature for 2 h. Sectionswere then rinsed in buffer containing the same concentra-tion of gelatin and BSA as above. Following rinses with0.01 M PBS, sections were incubated in 2% glutaraldehyde(Electron Microscopy Sciences) in 0.01 M PBS for 10 min.This procedure was followed by washes in 0.01 M PBS and0.2 M sodium citrate buffer (pH 7.4). A silver enhancementkit (Amersham Bioscience Corp., Piscataway, NJ) was usedfor silver intensification of the gold particles. The optimaltimes for silver enhancement were determined by empiricalobservation for each experiment and ranged between8 and 10 min. To avoid bias in labeling, some tissue sectionsincubated for GPR177+MOR were reversed labeled such thatGPR177 was labeled for immunoperoxidase and MOR waslabeled with immunogold-silver. Following intensification,tissue sections were rinsed in 0.2 M citrate buffer and0.1 M PB, and incubated in 2% osmium tetroxide (ElectronMicroscopy Sciences) in 0.1 M PB for 1 h, washed in0.1 M PB, dehydrated in an ascending series of ethanolfollowed by propylene oxide, and flat embedded in Epon 812(Electron Microscopy Sciences; Leranth and Pickel, 1989).Thin sections (50–100 nm thick) were then cut witha diamond knife (Diatome-US, Fort Washington, PA, USA)using a Leica Ultracut (Leica Microsystems, Wetzlar,Germany). Captured images of selected sections were com-pared with captured light microscopic images of the blockface before sectioning. Sections were collected on coppermesh grids and examined with an electron microscope(Morgagni, Fei Company, Hillsboro, OR). Digital images weretaken with an AMT advantage HR/HR-B CCD camera system(Advance Microscopy Techniques, Danvers, MA). Figureswere assembled and adjusted for brightness and contrast inAdobe Photoshop.

79B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
4.5. Identification of immunogold-silver labeling in profiles
Profiles exhibiting immunogold-silver labeling were identi-fied by the presence, in single thin sections, of at least twoimmunogold-silver particles within a cellular compartment.A profile containing only one immunogold particle inadjacent thin sections was designated as lacking detectableimmunoreactivity. As observed in lowmagnification electronmicrographs, background labeling in the neuropil, deemedspurious, was not commonly encountered. The criterion oftwo immunogold particles as indicative of GPR177 labeling isconservative and may have led to an underestimation of thenumber of GPR177-labeled profiles. We have previouslyreported GPR177-immunoreactivity in somata and dendriticprocesses in the striatum exhibiting at least two to threeimmunogold particles (Jin et al., 2010). Another factor thatmay have led to the underestimation of labeled profiles is thelimitation of immunocytochemical methods to detect traceamounts of GPR177. Moreover, unbiased stereological meth-ods were not used for counting labeled profiles, and theresults of the numerical analysis can only be considered to bean estimate of the numbers of synapses and labeled profiles.
4.6. Control and data analysis
The quantitative approach used in this study has beendescribed previously (Van Bockstaele et al., 1996a,b). Somesections were processed in parallel with the rest of theprocedures identical, but one of the primary antisera wasomitted. Sections processed in the absence of primaryantibody did not exhibit immunoreactivity. To evaluatecross-reactivity of labeling of the primary antiserum bysecondary antisera, some sections were processed for duallabeling with omission of one of the primary antisera.Semiquantitative analysis was carried out in tissue sectionswith optimal immunohistochemical labeling and preserva-tion of ultrastructural morphology. At least 9 grids contain-ing 5 to 7 thin sections each were collected from at least threeplastic-embedded striatal sections from each animal. Cellu-lar elements were identified based on previous descriptions(Peters et al., 1991). Dendrites were identified by the presenceof endoplasmic reticulum and a diameter of at least 0.4 μm,while axon terminals were identified by the presence ofsynaptic vesicles and a diameter of at least 0.3 μm. Asym-metric and symmetric synapses were also identified basedon previous descriptions (Gray, 1959). Asymmetric synapseswere documented by the presence of thick postsynapticdensities, while symmetric synapses by the presence of thindensities both pre- and postsynaptic. Non-synaptic contacts,or appositions, were defined by closely spaced plasmamembranes of axons, other axon terminals, or dendrites;these lacked recognizable specializations.
GPR177 and MOR immunolabeling was identified as eithercytoplasmic or plasmalemmal. Immunogold-silver grainswere classified as plasmalemmal if they were associatedwith the plasmamembrane, and cytoplasmic if they were notin contact with the plasma membrane. Dendritic profileswere classified as “small” or “large” based on their perimeter.Small dendrites were dendritic profiles with a perimetersmaller than 5 μm and all other profiles more than 5 μm as
large dendrites. The density of GPR177 in the cytoplasm wascalculated as the number of cytoplasmic immunogold-silverparticles per unit area (μm2). Likewise, the density of GPR177on the plasma membrane was calculated as the number ofplasmalemmal immunogold-silver particles per unit perim-eter (μm). All values were obtained per animal and theaverage of the three animals was calculated. The perimeterand area of each dendritic profile was obtained using Image Jsoftware (NIH).
Acknowledgments
This project was supported by the National Institutes ofHealth grants P20 DA 025995 to W.B. and DA 09082 to E.V.B.
R E F E R E N C E S
Albin, R.L., Young, A.B., Penney, J.B., 1989. The functional anatomyof basal ganglia disorders. Trends Neurosci. 12, 366–375.
Alexander, G.E., DeLong, M.R., Strick, P.L., 1986. Parallelorganization of functionally segregated circuits linking basalganglia and cortex. Annu. Rev. Neurosci. 9, 357–381.
Bailey, C.P., Connor, M., 2005. Opioids: cellular mechanisms oftolerance and physical dependence. Curr. Opin. Pharmacol. 5,60–68.
Banziger, C., Soldini, D., Schutt, C., Zipperlen, P., Hausmann, G.,Basler, K., 2006. Wntless, a conserved membrane proteindedicated to the secretion of Wnt proteins from signaling cells.Cell 125, 509–522.
Bartscherer, K., Pelte, N., Ingelfinger, D., Boutros, M., 2006. SecretionofWnt ligands requiresEvi, a conservedtransmembraneprotein.Cell 125, 523–533.
Beckstead, R.M., Kersey, K.S., 1985. Immunohistochemicaldemonstration of differential substance P-, met-enkephalin-,and glutamic-acid-decarboxylase-containing cell body andaxon distributions in the corpus striatum of the cat. J. Comp.Neurol. 232, 481–498.
Calabresi, P., Pisani, A., Mercuri, N.B., Bernardi, G., 1996. Thecorticostriatal projection: from synaptic plasticity todysfunctions of the basal ganglia. Trends Neurosci. 19,19–24.
Chen, T.C., Cheng, Y.Y., Sun, W.Z., Shyu, B.C., 2008. Differentialregulation of morphine antinociceptive effects byendogenous enkephalinergic system in the forebrain of mice.Mol. Pain 4, 41.
Cheng, P.Y., Moriwaki, A., Wang, J.B., Uhl, G.R., Pickel, V.M., 1996.Ultrastructural localization of mu-opioid receptors in thesuperficial layers of the rat cervical spinal cord: extrasynapticlocalization and proximity to Leu5-enkephalin. Brain Res. 731,141–154.
Ciani, L., Salinas, P.C., 2005. WNTs in the vertebrate nervoussystem: from patterning to neuronal connectivity. Nat. Rev.Neurosci. 6, 351–362.
Contet, C., Kieffer, B.L., Befort, K., 2004. Mu opioid receptor:a gateway to drug addiction. Curr. Opin. Neurobiol. 14,370–378.
Eisch, A.J., Barrot, M., Schad, C.A., Self, D.W., Nestler, E.J., 2000.Opiates inhibit neurogenesis in the adult rat hippocampus.Proc. Natl. Acad. Sci. U. S. A. 97, 7579–7584.
Fonnum, F., Walaas, I., Iversen, E., 1977. Localization of GABAergic,cholinergic and aminergic structures in the mesolimbicsystem. J. Neurochem. 29, 221–230.

80 B R A I N R E S E A R C H 1 3 5 8 ( 2 0 1 0 ) 7 1 – 8 0
Franch-Marro, X., Wendler, F., Guidato, S., Griffith, J.,Baena-Lopez, A., Itasaki, N., Maurice, M.M., Vincent, J.P., 2008.Wingless secretion requires endosome-to-Golgi retrieval ofWntless/Evi/Sprinter by the retromer complex. Nat. Cell Biol.10, 170–177.
Fu, J., Jiang, M., Mirando, A.J., Yu, H.M., Hsu, W., 2009. Reciprocalregulation of Wnt and Gpr177/mouse Wntless is required forembryonic axis formation. Proc. Natl. Acad. Sci. U. S. A. 106,18598–18603.
Gerfen,C.R.,Wilson,C.J., 1996. Thebasal ganglia, In: Swanson, L.W.B.A., Hokfelt, T. (Eds.), Handbook of chemical neuroanatomy, 12.Elsevier Science, Amsterdam, pp. 371–468.
Goodman, R.M., Thombre, S., Firtina, Z., Gray, D., Betts, D.,Roebuck, J., Spana, E.P., Selva, E.M., 2006. Sprinter: a noveltransmembrane protein required for Wg secretion andsignaling. Development 133, 4901–4911.
Graveland, G.A., DiFiglia, M., 1985. The frequency and distributionof medium-sized neurons with indented nuclei in the primateand rodent neostriatum. Brain Res. 327, 307–311.
Gray, E.G., 1959. Axosomatic and axo-dendritic synapses of thecerebral cortex: an electron microscopic study. J. Anat. 93,420–433.
Graybiel, A.M., 1990. Neurotransmitters and neuromodulators inthe basal ganglia. Trends Neurosci. 13, 244–254.
Graybiel, A.M., 1998. The basal ganglia and chunking of actionrepertoires. Neurobiol. Learn. Mem. 70, 119–136.
Graybiel, A.M., 2008. Habits, rituals, and the evaluative brain.Annu. Rev. Neurosci. 31, 359–387.
Harburg, G.C., Hall, F.S., Harrist, A.V., Sora, I., Uhl, G.R., Eisch, A.J.,2007. Knockout of the mu opioid receptor enhances thesurvival of adult-generated hippocampal granule cell neurons.Neuroscience 144, 77–87.
Jin, J., Kittanakom, S., Wong, V., Reyes, B.A.S., Van Bockstaele, E.J.,Stagljar, I., Berrettini, W., Levenson, R., Interaction of themu-opioid receptor with GPR177 (Wntless) inhibits Wntsecretion: potential implications for opioid dependence. BMCNeurosci. 11, 33.
Kahn, L., Alonso, G., Normand, E., Manzoni, O.J., 2005. Repeatedmorphine treatment alters polysialylated neural cell adhesionmolecule, glutamate decarboxylase-67 expression and cellproliferation in the adult rat hippocampus. Eur. J. Neurosci. 21,493–500.
Kehagia, A.A., Murray, G.K., Robbins, T.W. Learning and cognitiveflexibility: frontostriatal function and monoaminergicmodulation. Curr. Opin. Neurobiol.
Kieffer, B.L., 1999. Opioids: first lessons from knockout mice.Trends Pharmacol. Sci. 20, 19–26.
Koch, T., Schulz, S., Pfeiffer, M., Klutzny, M., Schroder, H., Kahl, E.,Hollt, V., 2001. C-terminal splice variants of the mousemu-opioid receptor differ in morphine-induced internalizationand receptor resensitization. J. Biol. Chem. 276, 31408–31414.
Koch, T., Widera, A., Bartzsch, K., Schulz, S., Brandenburg, L.O.,Wundrack, N., Beyer, A., Grecksch, G., Hollt, V., 2005. Receptorendocytosis counteracts the development of opioid tolerance.Mol. Pharmacol. 67, 280–287.
Koos, T., Tepper, J.M., Wilson, C.J., 2004. Comparison of IPSCsevoked by spiny and fast-spiking neurons in the neostriatum.J. Neurosci. 24, 7916–7922.
Leranth, C., Pickel, V.M., 1989. Electron microscopic preembeddingdouble-labeling methods, In: Heimer, L., Zaborszky, L. (Eds.),Neuroanatomical tracing methods 2, 1st ed. Plenum Press,New York, pp. 129–172.
Mansour, A., Fox, C.A., Burke, S., Meng, F., Thompson, R.C., Akil, H.,Watson, S.J., 1994a. Mu, delta, and kappa opioid receptormRNAexpression in the rat CNS: an in situ hybridization study.J. Comp. Neurol. 350, 412–438.
Mansour, A., Fox, C.A., Thompson, R.C., Akil, H., Watson, S.J.,1994b. mu-Opioid receptor mRNA expression in the rat CNS:comparison to mu-receptor binding. Brain Res. 643, 245–265.
Milligan, G., 2005. Opioid receptors and their interacting proteins.Neuromolecular Med. 7, 51–59.
Parelkar, N.K.,Wang, J.Q., 2003. PreproenkephalinmRNAexpressionin rat dorsal striatum induced by selective activation ofmetabotropic glutamate receptor subtype-5. Synapse 47,255–261.
Paxinos, G., Watson, C., 1997. The rat brain in stereotaxiccoordinates. Academic Press, New York.
Peters, A., Palay, S.L., Webster, H.d., 1991. The Fine Structure of theNervous System. Oxford University Press, New York.
Pickel, V.M., Chan, J., Sesack, S.R., 1992. Cellular basis forinteractions between catecholaminergic afferents and neuronscontaining Leu-enkephalin-like immunoreactivity in ratcaudate-putamen nuclei. J. Neurosci. Res. 31, 212–230.
Raehal, K.M., Bohn, L.M., 2005. Mu opioid receptor regulation andopiate responsiveness. Aaps J. 7, E587–E591.
Robinson, T.E., Kolb, B., 1999. Morphine alters the structure ofneurons in the nucleus accumbens and neocortex of rats.Synapse 33, 160–162.
Salinas, P.C., Zou, Y., 2008. Wnt signaling in neural circuitassembly. Annu. Rev. Neurosci. 31, 339–358.
Skaer, T.L., 2004. Practice guidelines for transdermal opioids inmalignant pain. Drugs 64, 2629–2638.
Sklair-Tavron, L., Shi, W.X., Lane, S.B., Harris, H.W., Bunney, B.S.,Nestler, E.J., 1996. Chronic morphine induces visible changes inthe morphology of mesolimbic dopamine neurons. Proc. Natl.Acad. Sci. U. S. A. 93, 11202–11207.
Steiner, H., Gerfen, C.R., 1999. Enkephalin regulates acute D2dopamine receptor antagonist-induced immediate-earlygene expression in striatal neurons. Neuroscience 88,795–810.
Surratt, C.K., Johnson, P.S., Moriwaki, A., Seidleck, B.K., Blaschak,C.J., Wang, J.B., Uhl, G.R., 1994. -mu opiate receptor. Chargedtransmembrane domain amino acids are critical for agonistrecognition and intrinsic activity. J. Biol. Chem. 269,20548–20553.
Taverna, S., Canciani, B., Pennartz, C.M., 2007. Membraneproperties and synaptic connectivity of fast-spikinginterneurons in rat ventral striatum. Brain Res. 1152,49–56.
Van Bockstaele, E.J., Colago, E.E., Cheng, P., Moriwaki, A., Uhl, G.R.,Pickel, V.M., 1996a. Ultrastructural evidence for prominentdistribution of the mu-opioid receptor at extrasynaptic siteson noradrenergic dendrites in the rat nucleus locus coeruleus.J. Neurosci. 16, 5037–5048.
Van Bockstaele, E.J., Colago, E.E., Moriwaki, A., Uhl, G.R., 1996b.Mu-opioid receptor is located on the plasma membrane ofdendrites that receive asymmetric synapses from axonterminals containing leucine-enkephalin in the rat nucleuslocus coeruleus. J. Comp. Neurol. 376, 65–74.
von Zastrow, M., Svingos, A., Haberstock-Debic, H., Evans, C., 2003.Regulated endocytosis of opioid receptors: cellularmechanisms and proposed roles in physiological adaptation toopiate drugs. Curr. Opin. Neurobiol. 13, 348–353.
Waldhoer, M., Bartlett, S.E., Whistler, J.L., 2004. Opioid receptors.Annu. Rev. Biochem. 73, 953–990.
Wang, H., Moriwaki, A., Wang, J.B., Uhl, G.R., Pickel, V.M., 1996.Ultrastructural immunocytochemical localization of mu opioidreceptors and Leu5-enkephalin in the patch compartmentof the rat caudate–putamen nucleus. J. Comp. Neurol. 375,659–674.
Wang, H., Gracy, K.N., Pickel, V.M., 1999. Mu-opioid and NMDA-typeglutamate receptors are often colocalized in spiny neuronswithin patches of the caudate–putamen nucleus. J. Comp.Neurol. 412, 132–146.
Watson, D.J., Stanton, M.E., 2009. Spatial discrimination reversallearning in weanling rats is impaired by striatal adminis-tration of an NMDA-receptor antagonist. Learn Mem. 16,564–572.
Copyright © 2022 FDOKUMEN




















