
U-V-fluorescence studies on thein vitro intracellular accumulation of carcinogenic hydrocarbon
27
U-%Fluorescence Studies on the in vifro Intracellular Accumulation of Carcinogenic Hydrocarbon' KENNETH M. RICHTER AND VIRENDER K. SAINI DepaThnent of Anatomy, University of Oklahoma Medical Center, Oklahoma City, Oklahoma, and Department of Anatomy, Dalhousie University, Halifax, Nova Scotia, Canada Carcinogenic hydrocarbons (CH) when activated by u-v light peaking at a wave- length of 365 millimicra emit a strong, characteristic blue fluorescence (Miller and Baumann, '43a;b). They can be detected visually by u-v-fluorescence microscopy in dilutions as great as 1 : 40 million to 1 : 1 bil- lion (Graffi, '39, '40b; Simpson and Cramer, '43a; Peacock, '36). However u-v-fluores- cence microscopic studies on the intra- cellular solubilization and distribution of the hydrocarbons are exceedingly limited (Peacock, '36, '40; Grafli, '39, 40a,b; Gunther, '41; Doniach, Mottram and Wei- gert, '43; Simpson and Cramer, '43a,b, '45a; Ahlstrom, '49; Setiila and Ekwall, '50a,b; Richter, '52a, '55). Studies by GrafIi ('39, '40a,b), Gunther ('41), and Richter ('51, '52a,b, '55) have provided the only direct objective data presently avail- able on the penetration, accumulation and distribution of these substances within liv- ing cells. Even though the total information in this area is &ted and somewhat confused by differences in experimental approaches; it is evident that intracellular penetration, accumulation and distribution of carcino- genic hydrocarbons are not the same for all living cell types and special proto- plasmic systems, and that these dissimi- larities are constant and characteristic from one cell type to another. It seems essential, therefore, to our general and specific thinking in the area of carcino- genic mechanisms and in the area of the influences of these substances on cellular biology generally that much more precise comparative cytologic data is needed than we now have on these fundamental phenomena. This report describes the in- tracellular penetration, accumulation and distribution of certain carcinogenic hydro- carbons in several functionally and mor- phologically diverse normal and neoplastic cell types in vitro and presents a brief review of the literature bearing on these phenomena. MATERIALS AND METHODS Materials. The carcinogenic hydrocar- bons used included : dibenzanthracene (DBA), benzpyrene (BP), and methyl- cholanthrene (MC) all from Hoffman- LaRoche. The cell types studied include: (a) seminiferous epithelial cells and inter- stitial cells of the adult mouse (Carworth Farms Strain no. 1 ), (b) human peripheral blood granular leucocytes, platelets and erythrocytes, mouse and fowl erythrocytes and fowl leucocytes and thrombocytes, ( c ) embryonal chick heart fibroblasts, (d) Walker rat sarcoma, and (e) the HeLa cell strain. Methods. The experimental approach followed is based on in vitro methods used previously in similar studies (Richter, '51a, '52a,b, '55; Lewis, '35). a. Suspensions of living testicular tissue cells were prepared by mincing testes un- der aseptic conditions in Tyrode and hu- man ascitic fluid. In experimental prep- arations, the cell suspension was mounted on sterile, physiologically clean slides on which a field of microcrystalline benz- pyrene had been deposited by evaporation from a saturated ethyl-alcoholic solution. Coverslips were added immediately and sealed with sterile vaseline. Control prep arations were set-up the same way, but benzpyrene was omitted. b. In the case of peripheral blood cells, Supported in part by U.S.P.H.S. research grant C-2214 from the National Cancer Institute and CP-30 from the American Cancer Society. 209
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of U-V-fluorescence studies on thein vitro intracellular accumulation of carcinogenic hydrocarbon

U-%Fluorescence Studies on the in vifro Intracellular Accumulation of Carcinogenic Hydrocarbon'
KENNETH M. RICHTER AND VIRENDER K. SAINI DepaThnent of Anatomy, University of Oklahoma Medical Center, Oklahoma City, Oklahoma, and Department of Anatomy, Dalhousie University, Halifax, Nova Scotia, Canada
Carcinogenic hydrocarbons (CH) when activated by u-v light peaking at a wave- length of 365 millimicra emit a strong, characteristic blue fluorescence (Miller and Baumann, '43a;b). They can be detected visually by u-v-fluorescence microscopy in dilutions as great as 1 : 40 million to 1 : 1 bil- lion (Graffi, '39, '40b; Simpson and Cramer, '43a; Peacock, '36). However u-v-fluores- cence microscopic studies on the intra- cellular solubilization and distribution of the hydrocarbons are exceedingly limited (Peacock, '36, '40; Grafli, '39, 40a,b; Gunther, '41; Doniach, Mottram and Wei- gert, '43; Simpson and Cramer, '43a,b, '45a; Ahlstrom, '49; Setiila and Ekwall, '50a,b; Richter, '52a, '55). Studies by GrafIi ('39, '40a,b), Gunther ('41), and Richter ('51, '52a,b, '55) have provided the only direct objective data presently avail- able on the penetration, accumulation and distribution of these substances within liv- ing cells.
Even though the total information in this area is &ted and somewhat confused by differences in experimental approaches; it is evident that intracellular penetration, accumulation and distribution of carcino- genic hydrocarbons are not the same for all living cell types and special proto- plasmic systems, and that these dissimi- larities are constant and characteristic from one cell type to another. It seems essential, therefore, to our general and specific thinking in the area of carcino- genic mechanisms and in the area of the influences of these substances on cellular biology generally that much more precise comparative cytologic data is needed than we now have on these fundamental phenomena. This report describes the in- tracellular penetration, accumulation and distribution of certain carcinogenic hydro-
carbons in several functionally and mor- phologically diverse normal and neoplastic cell types in vitro and presents a brief review of the literature bearing on these phenomena.
MATERIALS AND METHODS
Materials. The carcinogenic hydrocar- bons used included : dibenzanthracene (DBA), benzpyrene (BP), and methyl- cholanthrene (MC) all from Hoffman- LaRoche.
The cell types studied include: (a) seminiferous epithelial cells and inter- stitial cells of the adult mouse (Carworth Farms Strain no. 1 ), (b) human peripheral blood granular leucocytes, platelets and erythrocytes, mouse and fowl erythrocytes and fowl leucocytes and thrombocytes, (c) embryonal chick heart fibroblasts, (d) Walker rat sarcoma, and (e) the HeLa cell strain.
Methods. The experimental approach followed is based on in vitro methods used previously in similar studies (Richter, '51a, '52a,b, '55; Lewis, '35).
a. Suspensions of living testicular tissue cells were prepared by mincing testes un- der aseptic conditions in Tyrode and hu- man ascitic fluid. In experimental prep- arations, the cell suspension was mounted on sterile, physiologically clean slides on which a field of microcrystalline benz- pyrene had been deposited by evaporation from a saturated ethyl-alcoholic solution. Coverslips were added immediately and sealed with sterile vaseline. Control prep arations were set-up the same way, but benzpyrene was omitted.
b. In the case of peripheral blood cells, Supported in part by U.S.P.H.S. research grant
C-2214 from the National Cancer Institute and CP-30 from the American Cancer Society.
209

210 KENNETH M. RICHTER AND VIRENDER K. SAINI
whole fresh blood obtained by puncture was placed on physiologically clean and sterile slides which had been seeded with one of the CH. Coverslips were added and sealed with sterile vaseline. Control prep- arations were set-up the same way, but without carcinogenic hydrocarbon.
c. In the case of chick heart fibroblasts, Walker rat sarcoma cells, and HeLa cells, the cells were cultured by conventional methods in completely fluid nutrient media (mixtures of Eagle's medium, serum, hu- man ascitic fluid, chick embryo extract, balanced saline) on glass coverslips on which carcinogenic hydrocarbon had been deposited by evaporation from an ethyl alcholic solution. For critical microscopic examination the coverslips bearing the cul- tures were mounted aseptically on stand- ard slides and sealed with vaseline. Con- trol preparations were cultured and set-up the same way but without carcinogenic hydrocarbon. In some instances the cul- tured cells were not exposed to the carcino- gen until moments before critical examina- tion was begun, and then, the coverslips bearing the cultured cells were mounted and sealed on slides seeded with crystal- line carcinogen.
This experimental situation is equivalent to those used in the in vitro studies of the effects of CH on tissues in culture by Lewis ('35), Earle and Voegtlin ('38), and Earle ('43). By this method, (1) no special CH solvent is introduced into the extra- cellular medium, and (2) the aqueous extracellular medium is saturated with solubilized carcinogen. It is estimated on the basis of Earle's measurements on rather equivalent Mandler candle filtrates of serum suspensions of CH, that the extra- cellular saturated concentration of solu- bilized CH in these preparations is prob- ably in the neighborhood of 1.0 y/cm*, and Peacock ('40) has reported that serum will dissolve 1.5 Y of CH per cm3.
Control and experimental preparations were studied by integrated ordinary, dark- contrast-medium phase, and u-v-fluores- cence microscopic methods. The u-v-fluo- rescence microscopic set-up included : ( 1 ) quartz substage optics, (2) American Op- tical ordinary and reflecting objectives, (3) u-v absorption filters of two types: the Corning no. 3389 filter, and 5% aqueous
sodium-nitrite, (4) infrared absorption fil- ters of two types: the Corning no. 3966 filter, and aqueous copper-sulfate, (5 ) the Corning u-v transmitting filter (300- 425 mu) no. 5840, and (6) the Osram HBO-200 mercury arc lamp.
Precautions were taken to avoid second- ary influences of cell injury by u-v, in- frared, and high intensity light (Lewis, '35; Earle, '43) on the intracellular pene- tration and distribution of CH. These in- cluded: (1) the utilization of neutral den- sity, infrared, and u-v absorption filters at all times during phase-contrast and ordi- nary light microscopic study, (2) the limit- ing of u-v-fluorescence study of each prep- aration to a period of under 20 minutes, and ( 3 ) the correlative study of control preparations.
In practice, the in vitro preparations were first studied in detail and specific cells selected, for subsequent u-v-fluorescence study, with the ordinary or phase micro- scope. The preparations were then trans- ferred to the u-v-fluorescence microscope set for transmission of low intensity ordi- nary light (with infrared and u-v filtered out). When the cell or cells initially selected under phase or ordinary light microscopy had been located and all opti- cal adjustments had been made, the u-v- fluorescence microscope was then set for u-v transmission only and the select cell or preparation was then examined for no more than 20 minutes.
OBSERVATIONS
General solubility of CH in aqueous media and living cells
Fluorescence intensity varies somewhat directly with concentration of fluorescent substance, but suitable methods based on fluorescence intensity measurements have not been devised for their quantitative estimation in media, tissues or cells (Eriinko, '55). It is thus only possible to demonstrate solubilization of the CH by virtue of their fluorescence property (Miller and Baumann, '43a,b) and to express concentration levels in broad terms relative to demonstrable degrees of fluores- cence intensity.
In experimental preparations, the char- acteristic blue fluorescence of the CH spreads from the region of the crystalline

INTRACELLULAR CARCINOGENIC HYDROCARBON 211
material (fig. 53) in diminishing intensity gradients through all of the types of media used. This reflects clearly the progressive solubilization of crystalline CH and its diffusion in the several aqueous media (fig. 53). In the immediate surround of the crystalline bed the medium is saturated and toward the perimeter of the gradient zone and beyond, the concentration of solu- bilized CH diminishes progressively (fig. 53). The solubilization of CH and the establishment of concentration gradients progress rapidly and are demonstrable by u-v-fluorescence microscopy within seconds and moments after bringing the crystalline material and the media together.
The time-sequence of acquisition of the fluorescence of the CH by living cells varies in accordance with their relative positions in the fluorescence-intensity (concentra- tion-gradient ) zone, and shows the progres- sive movement of solubilized substance into the living cell (fig. 53). In general, intracellular penetration and accumula- tion of CH proceed rapidly, so that it is demonstrable in the living cells within seconds and moments after bringing the crystalline CH and the cell suspension to- gether. As to the amount of solubilized CH which may be accumulated intracellu- larly, it can only be demonstrated that it is accumulated within specific sites in the living cells at concentrations which greatly exceed those attainable in solubilized form in the extracellular medium (figs. 1-40,45, 48, 50, 52, 53). As the relatively low fluo- rescence intensity shown by the extracellu- lar medium represents at most a saturated concentration level of 1.0-1.5 y/cms, the contrasting high fluorescence intensities within the cell indicate the general proto- plasmic concentration of solubilized CH to be several times greater than the saturated extracellular concentration, or X times 1.0- 1.5 y of CH per cm3 of protoplasm (figs. 1-40, 44, 45, 48, 50, 52, 53).
In all control preparations, the various media used and the several cell types stud- ied showed no intrinsic fluorescence at the activating u-v wave-lengths employed (fig. 53). No demonstrable cytomorphk differ- ences between the control and the carcino- gen-treated cells were noted during the various in vitro experimental periods. (See figure legends.) This finding agrees with
previous observations and those of others on living cells in vitro maintained under basically identical conditions (Lewis, '35; Earle, '43; Richter, '51, '52a,b, '55). Be- cause of these joint circumstances, and in view of the specific objective of this study, further observations on the control prep- arations as such will be omitted. The nor- malcy of the experimentally-treated cells is evident in all illustrations (figs. 1-53).
Specific intracellulat distribution of CH Germinal cells. Inasmuch as the gen-
eral objective of this study is one of deter- mining the specific site or sites within liv- ing cells in which CH may be accumulated, it is particularly helpful that the detailed cytology of mammalian spermatogenesis is well-known and that the identities and transformations of the mitochondrial, Golgi, general cytoplasmic and nuclear components during spermatogenesis are well-established and enjoy an essentially uncontroversial status (Papanicolaou and Stockard, '18; Gatenby and Woodger, '21; Gatenby, '29, '31, '35; Gresson and Zlotnik, '45; Clermont, Glegg and Leblond, '55; Burgos and Fawcett, '55; Clermont and Leblond, '53, '55).
In this category of cells BP is differ- entially accumulated and distributed in specific protoplasmic sites in characteristic patterns and in relative concentrations which are constant for each spermatogenic cell type or phase.
The primary spermatocyte has a spher- ical nucleus and well-defined nuclear membrane (fig. 1). The mitochondria are granular to short rod-like units distributed in a peri- or juxtanuclear position. The Golgi structure at certain phases is a rela- tively large spherical unit situated charac- teristically in a juxtanuclear position (fig. 1). Benzpyrene is demonstrable in this cell type in increasing amounts respectively in the cytoplasmic ground-substance, the plasma and nuclear membranes, and the Golgi structure (figs. 1, 2). There is no demonstrable BP in the intranuclear com- ponents, i.e., chromatin, karyolymph etc., or in the mitochondria (figs. 1 ,2 ) . Multi- nucleate spermatocytes are of normal oc- currence, and such cells possess corre- spondingly increased numbers of Golgi structures etc. (fig. 7). In these cells, too,

212 KENNETH M. RICHTER AND VIRENDER K. SAINI
the carcinogen accumulates differentially, and in amounts characteristic of those in the mono-nucleate spermatocyte, i.e., in the cytoplasmic ground-substance, the plasma and nuclear membranes, and the multiple Golgi structures (figs. 7,8); there is no demonstrable BP in the mitochondria and in the intranuclear area (fig. 8). In some mono-nucleate spennatocytes, but most commonly in the multinucleate ones, small vesicular aggregates occur which are of uncertain identity but of normal oc- currence (fig. 7). Benzpyrene accumulates in them in relatively high amounts (fig. 8).
The spermatocyte at the metaphase stage of meiosis shows a plate of discrete chro- mosomes, an achromatic spindle, a Golgi structure at each spindle pole, and mito- chondria disposed about the spindle (fig. 3). Spermatocytes with multipolar spin- dles are of normal occurrence and differ from the mono-nucleate cell in that more than one spindle, metaphase ,chromosomal plate and more than two polar Golgi struc- tures are present (fig. 5) . Benzpyrene ac- cumulates in these cells in increased amounts respectively in the cytoplasmic ground-substance, the plasma membrane, nuclear membrane remnants (?), and the polar Golgi structures (figs. 3-6). The chromosomes, and mitochondria show no demonstrable hydrocarbon; and if the spin- dle shows any at all, it must be present in exceedingly low amount (figs. 3-6).
Spermatid transformations have been grouped by Leblond and Clermont ('52) into 4 descriptive phases; ( 1 ) the Golgi phase, (2) the cap phase, (3) the acro- some phase, and (4) the maturation phase.
The early Golgi phase spermatid is a small, rounded cell with a spherical ec- centric nucleus and a single prominent nu- cleolus (fig. 9). The Golgi structure (acroblast) is a relatively large, spherical unit, similar to that of the spermatocyte, situated in a characteristic juxtanuclear position. The mitochondria are small gran- ular units irregularly dispersed in the cyto- plasm (fig. 9). Benzpyrene is demon- strable in this spermatid in increased amounts respectively in the cytoplasmic ground-substance, the plasma and nuclear membranes, and the acroblast Golgi struc- ture (fig. 10). Benzpyrene is not demon- strable in any intranuclear component, i.e.,
nucleolus, chromatin and karyolymph, or in the mitochondria (fig. 10).
In slightly later Golgi phase spennatids, the acroblast (Golgi structure) segregates an acrosomal vacuole which consists struc- turally of a limiting membrane, i.e., the wall of the acrosomal vacuole (fig. ll), and an internal clear filling substance (fig. 11) in which the acrosomal granule later forms (fig. 13). The mitochondria of this phase are short rods irregularly disposed about the nucleus (figs. 11, 13). In these phases benzpyrene is demonstrable in in- creasing amounts respectively in the cyto- plasmic ground-substance, the nuclear and plasma membranes, the wall of the acroso- mal vacuole, and the acroblast (figs. 11- 14). The intranuclear substance, mito- chondria, acrosomal granule and the clear filling substance of the acrosomal vacuole show no demonstrable benzpyrene accumu- lations (figs. 11-14).
In spermatids of the cap and acrosome phases (Leblond and Clermont, '52), the acroblast separates from the acrosomal complex and moves into the prospective tail cytoplasm (figs. 13,15). Concurrently, the nucleus becomes slightly elongated. The acrosomal complex (which includes the wall of the acrosomal vacuole, the internal clear filling vacuolar substance, and the acrosomal granule) enlarges and is closely fitted over the anterior end of the nucleus as the head cap (fig. 15). In this process, the acrosomal granule remains stationed at the anterior tip of the nucleus and the remainder of the complex is re- flected caudally over the nucleus. That portion of the complex containing the acrosomal granule forms a short append- age at the tip of the nucleus or prospective tip of the mature sperm (fig. 15). In cells at these phases benzpyrene is accumulated in increased amounts respectively in the cytoplasmic ground-substance, the plasma and nuclear membranes, the wall of the acrosomal vacuole, and the acroblast or Golgi remnant (fig. 16). There is no demon- strable accumulation of BP in the intra- nuclear substance, the acrosomal granule, the clear filling substance of the acrosomal vacuole, and the mitochondria (fig. 16).
Spermatids of the final or maturation phase (Leblond and Clermont, '52) are characterized by (1) a sloughing of excess

INTRACELLULAR CARCINOGENIC HYDROCARBON 213
tail cytoplasm containing the Golgi rem- nant, (2) the formation of the axial fda- ment of the tail, (3 ) the formation of the mitochondrial spiral sheath of the mid- piece, and (4) the conversion of the acroso- mal complex into the definitive sickle- shaped apical-piece of the sperm head (figs. 17, 19). In spermatids of this phase BP is demonstrable in increased amounts respectively in the cytoplasmic ground-sub- stance, wall of the acrosomal vacuole of the apical-piece, and the Golgi remnant (figs. 17-20). Benzpyrene is not demon- strable in the intranuclear substance, the nuclear membrane, the plasma membrane (if present), the mitochondrial spiral sheath, and the axial filament (figs. 17- 20).
Graffi ('40) examined mouse cells iden- tified as spermatogonia but his observa- tions were exceedingly limited and vague and cannot be correlated with the observa- tions recorded here.
The detailed cytology of the Sertoli cell is incompletely established and all cytoplasmic elements can not be identified with surety at this time. They generally are quite severely injured during the course of preparing the cell suspension and intact normal cells have not yet been observed. However, in tissue fragments containing both germinal cells and a large portion of a Sertoli cell, the latter shows two general types of cytoplasmic inclusion: ( a ) a short rod or filamentous component, and (b) a small granule component (fig. 21). In such cell fragments, BP is demon- strable in increased amounts respectively in the cytoplasmic ground-substance and in the small granule component (figs. 21, 22). There is no demonstrable BP in the intranuclear substance, the nuclear and plasma membranes, and the rod or fila- mentous cytoplasmic component (figs. 21, 22).
Interstitial cells. The interstitial cell has abundant cytoplasm and a relatively small, spherical nucleus. The cytoplasm shows (a ) an abundant widely dispersed small granule component, and (b) a lipid granule component present in variable numbers and sizes. The former is of un- certain identity, but appears comparable to the osmiophilic pigment deposits re- ported by Burgos and Fawcett ('55). The
Sertoli cells.
lipid granule component is identified with the lipid droplets reported by others (Fawcett and Burgos, '56) (figs. 23, 25). Benzpyrene is accumulated within these cells in increased amounts respectively in the cytoplasmic ground-substance, the small granule component, and the lipid granules (figs. 23-26). Benzpyrene is not demonstrable in the intranuclear substance and the nuclear and plasma membranes (figs. 23-26). The mitochondrial compon- ent was not identified. Graffi's findings ('40) while incomplete are similar to those recorded here.
Chick heart fibroblasts. The general cytology of the chick heart fibroblast in cul- ture is well known; however, the precise identities of all the cytoplasmic structural elements are not (Richter, '56, '52a; Palay, '58). These cells normally show an abun- dance of filamentous mitochondria which are of clear-cut status. They generally show, also, numerous characteristic peri- nuclear small granules which are osmio- philic and of duplex character but whose status is uncertain. The identity of the Golgi component in these cells is uncer- tain. Fat droplets frequently are present and usually put in an appearance in as- sociation with the small granule compo- nent (fig. 33). The carcinogenic hydro- carbons, BP, MC, and DBA, are accumu- lated in this cell type in increased amounts respectively in the cytoplasmic ground substance, the perinuclear small granule component (Graffi, '39), and the fat drop- lets when present. The nuclear and plasma membranes, the intranuclear components (karyolymph, nucleolus, chromatin) and mitochondria show no demonstrable accu- mulations of these substances (figs. 31- 34) (Richter, '52a).
Walker rat sarcoma cells in culture show rather un- controversial filamentous mitochondria and an abundance of small cytoplasmic granules of uncertain identity (figs. 27, 29). What comprises the Golgi complex is not clearly established. Carcinogenic hydrocarbons, BP, MC, and DBA, are ac- cumulated in increased amounts respec- tively in the cytoplasmic ground-substance the perinuclear small granule component (Graffi, '39), and the fat droplets when
Walker rat sarcoma cells.

214 KENNETH M. RICHTER AND VIRENDER K. SAINI
present. The nuclear and plasma mem- branes, the intranuclear components (karyolymph, nucleolus, chromatin) and mitochondria show no demonstrable ac- cumulations of these substances (figs. 27- 30) (Richter, '52a).
HeLa cervical carcinoma cells. The gen- eral cytologic features of the HeLa cell are well known, but the status of certain cyto- plasmic inclusions are not clearly estab- lished (Gey, Shapras and Borysko, '54; Gey, Bang and Gey, '54; Rose, '57, '60a). It shows an extensive system of character- istic slender filamentous mitochondria which are readily recognized by all (figs. 35, 37, 39). A second prominent category of inclusion consists of an abundance of relatively small granules usually referred to as a juxtanuclear aggregate (Rose, '57, '60a). Some of them are small and uni- formly dark; others are dark with light centers (figs. 35, 37). Their distribution and association with each other are vari- able; sometimes they are clumped, or widely separated; they may be dispersed or aggregated near the nucleus or about the cytocentrum area (figs. 35, 37, 39). Oc- casional cells show large juxtanuclear granules which are similar to variant pino- cytic granules (Rose, '57) (fig. 39). Solu- bilized BP is accumulated rapidly within the living HeLa cell (figs. 35-38). It is accumulated in increased amounts respec- tively in the cytoplasmic ground-substance (and this includes the ectoplasmic bulbous processes) (figs. 35, 36), the cytocentrum area, and the small granule (juxtanuclear aggregate) component (figs. 35-40, 48). Granules similar to variant pinocytic gran- ules show accumulations of BP which are only slightly in excess of that in the cyto- plasmic ground-substance (figs. 39-40). The associated satellites (Rose, '57) show a slightly higher accumulation than the V-P granule proper (figs. 39-40). The nu- clear and plasma membranes, the nucleoli, chromatin, karyolymph and mitochondria show no demonstrable accumulations of hydrocarbon. The mitochondria sometimes appear as dark (negative) filaments (figs.
Peripheral blood cells; granular leuco- cytes. The detailed descriptive cytology of the granular leucocytes is well known; however, the origin and relationships of the
35-38).
characterizing cytoplasmic granules are not clearly established (Richter, '42, '55; Low and Freeman, '58; Bessis, '56). The granular leucocytes both in viuo and in uitro normally show numerous specific granules widely dispersed in the cytoplasm (figs. 42-44) and plasmocrine droplets containing variable amounts of protein which are produced cyclically and extruded from the cell (figs. 42-43) (Richter, '52a, '55, '57). Mitochondria are present as dis- persed short rods which are not readily distinguishable (figs. 42-43). An osmio- phile Golgi component is present as a dif- fuse area in the region of the nuclear %of" or perinuclearly (Richter, '42, '55; Low and Freeman, '58) (figs. 42-43). In gen- eral, solubilized CH are accumulated in increased amounts respectively in the cyto- plasmic ground-substance, the specific granules, and "toxic" granules (figs. 44, 45,51,52). The nuclear and plasma mem- branes, intranuclear components, and plasmocrine droplets show no demon- strable accumulation of CH (compare figs. 42-44 with 45). Dark (negatively fluores- cent) rod-shaped structures having dimen- sions similar to those of mitochondria are seen in some u-v fluorescence micrographs (fig. 45). Graffi ('39) in a cursory study of mouse marrow reported vaguely the accumulation of CH in granules, taken to be mitochondria, in the immature myeloid cells. Peacock ('40) with an inadequate method reported that CH were not ac- cumulated in peripheral blood leucocytes.
Human neutrophilic and eosinophilic granular leucocytes and those of certain sub-mammalian species aggregate on micro-piles of crystalline CH (fig. 49) and frequently phagocytose crystals of these substances (figs. 42, 43, 46, 47) (Richter, '51, '52b); although this was reported by Peacock ('36) not to occur. Specific living cells containing one to several crystals of MC were followed for more than 48 hours or until natural cell death intervened. It is known that granular leucocytes in cul- tures of buffy coat normally die within 24-48 hours. In control preparations in this study they do not survive beyond 48- 64 hours. Cell death in CH-treated prep- arations thus cannot soundly be attributed to a toxic CH effect and is consistent with observations of others (Simpson, Carru-

INTRACELLULAR CARCINOGENIC HYDROCARBON 215
thers and Cramer, '45; Simpson and Cramer, '45a). Phagocytosed crystals re- mained virtually unchanged for as long as they were observed in both living cells and in dead ones (Richter, '51). This persist- ence of crystalline form is not unexpected in view of the fact that the experimental situation maintains saturated concentra- tion levels of solubilized CH both in the extracellular medium and in the internum of the cell (figs. 41-47, 53).
Peripheral blood cells; erythrocytes. Peripheral blood erythrocytes of mammals generally are non-nucleate protoplasmic units and ordinarily show no special formed cytoplasmic organelles; although classical cytologic literature is variable on this point (Jordan, '38). Submammalian erythrocytes for the most part are nucleate cells and appear to possess rather conven- tional Golgi and mitochondrial components (Jordan, '38). In these studies, neither the non-nucleate human and mouse periph- eral blood erythrocytes nor the nucleate erythrocytes of the fowl show demonstrable accumulations of CH (figs. 51,52) (Richter, '52a, '55). Hemolyzed human erythro- cytes, also, show no intracellular accumu- lation of CH (figs. 44-45). Nucleate fowl erythrocytes when damaged by u-v radia- tion sometimes show a limited cytoplasmic accumulation of CH.
Peripheral blood cells; platelets. Hu- man peripheral blood platelets normally represent non-nucleate cytoplasmic frag- ments of megakaryocytes and character- istically possess a few small granules of unknown genic relation (Jordan, '38; Low and Freeman, '58). Carcinogenic hydro- carbon accumulates in the platelet in in- creased amounts respectively in the cyto- plasmic ground-substance and in the spe- cific granulations (fig. 50). There is no demonstrable accumulation in the vacu- olar or plasmocrine droplet (fig. 50).
Submammalian vertebrate species are reported to have both nucleate and non- nucleate thrombocytes (Jordan, '38). The most common form in the fowl is the nu- cleate thrombocyte. It shows ordinarily a single, dark juxtanuclear body and several dispersed smaller granules or rods which are interpreted here as being respectively a Golgi structure and mitochondria (Jor- dan, '38). Carcinogenic hydrocarbons ac-
cumulate in this cell in increased amounts respectively in the cytoplasmic ground- substance and the juxtanuclear Golgi body (? ) (figs. 51, 52). There is no special accumulation in the nuclear and plasma membranes, the small granular component (mitochondria?) and the intranuclear area (figs. 51-52). Because of inadequate methods, Peacock ('40) was led to report that platelets and thrombocytes do not ac- cumulate CH.
DISCUSSION
Solubility of Ch in aqueous systems Information on the solubility of the
carcinogenic hydrocarbons in aqueous sys- tems is of fundamental concern in all studies relating to general cellular influ- ences and the mechanism of action of these substances.
The perspective that the CH are insolu- ble in aqueous systems has had rather pro- found and diverse influences on the think- ing, the design, and the complexity of the majority of studies employing these substances experimentally both in vivo and in vitro. Many studies have employed them directly in crystalline form (Schlum- berger, '48-'49; Lewis, '35; Earle, '43; Earle and Voegtlin, '38; Rieck, '60; Murray and Kopeck, '53) in approaches which imply a tacit assumption that these substances through solubilization phenomena some- how become biologically effective in molec- ular or dissolved form. The majority of studies, however, have utilized special solvent vehicles (oils, benzene, alcohol, glycerine, paraffi, lard, lanolin, carbowax) association colloids, organic acid deriva- tives etc. to introduce the CH into both in vitro and in vivo biologic systems (Murray and Kopeck, '53; Gr&, '39, 40; Haddow, '47; Simpson and Cramer, '43a,b, '45a,b; Stowell and Cramer, '42; Ahlstrom, '49; Gunther, '41; Setala and Ekwell, '50a,b, '49). Behind these approaches is the perspective that the CH are insoluble or are not effectively soluble in aqueous systems, This perspective also underlies studies in which glass fragments have been used as control substitutes for crystalline CH (Rieck, '60). It i s not always recog- nized that upon the addition to water of solutions of CH taken up in solvents which are miscible with water (e.g., glycerine,

216 KENNETH M. RICHTER AND VIRENDER K. SAINI
alcohol) that the bulk of the CH separates out immediately in microcrystalline form, so that such solvents are in large measure superfluous. The possibility that chylo- micra, hematoconia, protoplasmic particles or cells normally present in body fluids may function as CH vehicles is relatively neglected (Peacock, '36, '40); it is clear that CH accumulates in relatively high amounts in chylomicra, hematoconia, blood platelets (fig. 50) and leucocytes (Setiila and Ermda, '51; Richter, '52a, '51). The involvement in diverse ways of the many CH vehicles in experimental carcinogenesis and in the induction of specific cellular changes has been empha- sized frequently (Peacock, '36; Earle and Voegtlin, '38; Simpson and Cramer, '43b, '45a,b; Strait, Hrenoff and DeOme, '48; Berenblum and Shubik, '47). The points emphasized by these examples of the in- fluence of the "concept of the insolubility of CH in aqueous systems" are simply (1) that this perspective is prone to contribute to faulty experimental design and conse- quent interpretations relating to specific effects of CH per se, and (2) that special CH vehicles or others are additional experi- mental complications whose essentiality in the introduction of CH into protoplasmic or biologic systems is not beyond questioning.
Quantitative data on the solubility of CH in aqueous systems is vague and limited because of technical difficulties; and most information which is available is objective in nature (Peacock, '36, '40; Doniach, Mot- tram and Weigert, '43a,b; Richter, '52a, '55). It has been reported that the CH are insoluble in aqueous systems (Mueller and Rusch, '43) or virtually so (Haddow, '47). Lange ('56) lists no solubility data on MC or BP, and DBA is given as being only slightly soluble in acetic acid. Attempts made here to obtain consistent spectro- photometric data on their solubility in water have been uniformly unsuccessful; although single microcrystals of CH iso- lated by micromanipulative methods have been observed directly by polarizing micros- copy to go into solution in water and sera. Peacock ('40) although not giving any de- tails reported them to be soluble in serum to the extent of 1.5 yjcm3. On the basis of present data, it would appear to be firmly established that they are only
slightly soluble in water, serum and plasma etc. and at levels which neighbor on 1 y/cm3 which ( 1) may or may not be measurable by conventional methods as reported (Haddow, '47; Mueller and Rusch, '43; Earle, '43; Peacock, '40), but which (2) are readily demonstrable objectively by the more delicate u-v-fluorescence micro- scopic methods, although these have not yet been put on a precise quantitating basis (Peacock, '36, '40; Doniach, Mottram and Weigert, '43a,b; Richter, '52a, '55; Eranku, '60).
General intracellular penetration and accumulation of CH
The physical state of CH in aqueous ex- tracellular systems has a special bearing on their partitioning among living cells and tissues (Peacock, '40). It is clear from direct observations presented here and previously (Doniach, Mottram, and Wei- gert, '43a; Richter, '52a, '55) and from indirect ones (Peacock, '36, '40; Doniach, Mottram and Weigert, '43a,b) that CH solubilized in aqueous media without spe- cial solvents are available to or have access to living cells and that they are soluble in living protoplasmic systems. As demon- strated by u-v-fluorescence microscopy, they are shown to be accumulated intra- cellularly in most cases from aqueous me- dia in solubilized form in amounts which may exceed even saturated solubilized con- centrations of 1.0-1.5 y/cm3 present in the extracellular medium (figs. 1-40, 44, 45, 48, 50, 53). This one-sided partitioning of CH is due in part to the richness of proto- plasm generally in masked and unmasked lipids, but apparently more than simple solubilization in such protoplasmic con- stituents is involved (Doniach, Mottram and Weigert, '43a,b).
In some cell types, they are accumulated intracellularly in both their solubilized and crystalline forms, for granular leucocytes of several vertebrate species phagocytose crystalline CH in addition to that which is accumulated in high amount in solubilized form (figs. 41-47) (Richter, '51, '52b). Paramecia, also, ingest crystalline CH as reported by Doniach, Mottram and Weigert ('43a) and confirmed independently in this laboratory. In these instances, it is appar- ent that the total amount of intracellular

INTRACELLULAR CARCINOGENIC HYDROCARBON 217
CH (solubilized and crystalline) per cubic centimeter of protoplasm may be rather phenomenally high relative to that per cubic centimeter of extracellular medium. This type of partitioning of CH is referable clearly to the special cytophysiologic capa- city of these cells to phagocytose. It sug- gests that special functional attributes of other kinds characterizing other cell types may play equally significant roles in the intracellular penetration and accumulation of these substances in living cells. This appears to be borne out by peripheral blood erythrocytes and erythroplasts gen- erally, but in the opposite direction, i.e., toward rejection of CH, in that these living cells normally show no demonstrable ac- cumulation of CH (Peacock, '40; Richter, '52a, '55) (figs. 44, 45, 51, 52). This is not due to a masking effect by hemoglobin (Peacock, '40), for naturally hemolysed cells also show no demonstrable CH in either their interna or plasma membranes (figs. 44, 45). If, however, the living cells are modified by prolonged u-v-irradiation, by air-drying (unpublished observation) or by hemolysis with distilled water (Peacock, '40), CH may be demonstrably accumu- lated in them. The general negative pene- tration of CH into living red cells is surely a reflection of special cytophysiologic factors.
It would appear (1) that there is little justification for the use of special vehicles in the introduction of CH into the proto- plasmic system, and (2) that the degree of intracellular penetration and accumulation of CH reflects jointly the specific composi- tional, structural and cytophysiological aspects of the living cells themselves.
Intracellular distribution of CH Influence of methods. All u-v-fluores-
cence microscopic observations so far re- ported and bearing specifically on the intra- cellular distribution of the hydrocarbons have been based on rather different experi- mental and observational approaches. The observations reported by most were based on fixed and/or frozen sectioned tissues following the in vivo administration of the hydrocarbons dissolved in conventional vehicles (Simpson and Cramer, '43a,b, '45a; Setala and Ekwall, '50a,b; Ahlstrom, '49), or as crystalline suspensions (Doniach,
Mottram and Weigert, '43; Peacock, '36). Gunther ('41) studied mesenterial spreads directly following the intraperitoneal in- jection of BP dissolved in olive oil or glycerine. Those of GrafFi ('39, '40) were based on the direct examination of living cells in vitro bathed in media to which hydrocarbon had been added as a glycerine solution. His experimental media in gen- eral contained from 0.04% to 20% glycer- ine. The use of glycerine in the prepara- tion of dead cell models is well known (Hoffmann-Berling, '58). The approaches used here differ from those of others in that artifactual intracellular distributions of CH due to possible secondary effects of special solvent vehicles, fixation and sec- tioning methods were avoided by introduc- ing crystalline CH directly into the cellular medium and by observing living cells directly under as near optimal in vitro conditions as possible.
The precise influences of CH vehicles, and of fixation and/or sectioning methods on the particular intracellular distributions of CH which have been reported by others cannot be evaluated adequately. It is known that some of the CH vehicles used have cytophysiologic effects (Ahlstrom, '49; Simpson and Cramer, '43a,b, '45a,b; Graffi, '39, '40). The possibility that sec- ondary intracellular distributions might result as a consequence of formalin fixa- tion was considered only by Gunther ('41) in a study on Paramecium. His finding, however, that there was no redistribution of CH following treatment with formalin is somewhat questionable because his cells already were partially glycerinated at the time of fixation as a consequence of having added the CH to the experimental medium as a glycerine solution. As none of the workers studying formalin and/or frozen sectioned preparations have reported the CH to be more precisely distributed than as simply being in (a) the cytoplasmic area and (b) in cytoplasmic lipid droplets; it would seem that more precise distribu- tions could not be made out clearly, or were not preserved, or were not looked for. In view of the precise intracellular dis- tributions obtained here in living cells with- out special CH solvents, it would seem that special solvents, fixation and sectioning methods are not above suspicion and, as

218 KENNETH M. RICHTER AND VIRENDER K. SAINI
potential complications, might better be avoided, if possible, just on general prin- ciples.
With one exception (Graffi, ’40) there is uniform agreement that the hydrocarbons as such, do not accumulate in demonstrable amount within the intranuclear area, i.e., exclusive of the nuclear membrane. Graffi (’40) re- ported the occasional accumulation of BP in the nucleoli of slightly glycerinated mouse ascites carcinoma cells. This unique exception is probably an artifact referable to an effect of glycerination or over u-v- irradiation. With this exception, the CH have not been demonstrable in any intranu- clear component (i.e., nucleolus, chro- matin, karyolymph, etc.) of any inter- kinetic or post-mitotic cell type reported on so far and quite irrespective of the experi- mental approach or preparatory method employed. In addition, the CH are not demonstrable in special nuclear derivatives such as the chromosomes and possibly the spindle of dividing cells (figs. 3-6) (Ahl- strom, ’49). The observations reported here and previously (Richter, ’52a) and those of Lewis (’35) and Earle (’43) on equivalent experimental cultural situa- tions, and those of Graffi (’39, 40) as well on low glycerine containing cultures indi- cate that intracellular CH does not inter- fere measurably with cell division in short- term cultures, i.e., of 2-6 days duration. Cell growth appears to be inhibited only after some weeks and months of continu- ous exposure to virtually maximal amounts of solubilized CH when introduced in ex- cess in crystalline form (Earle, ’43).
An explanation for the uniform lack of accumulation of CH in the intranuclear substance based on our present level of information is not possible. In view of the demonstrated solubility of CH in aqueous systems generally, and in the cytoplasm, and that the nucleus is an aqueous system, too, it would appear that the mechanism must be referable to something more elab- orate and discrete than simple solubiliza- tion phenomena. It merits much further critical consideration than it has received so far. (Negative evidence is always diffi- cult to evaluate. The uniform absence of typical CH-fluorescence from the intranu- clear components could be more apparent
Relative to the intranuclear area.
than real. Such would be the case if CH accumulated there in a form which did not fluoresce or whose fluorescence was masked or in some manner quenched. The same “ifs” can be applied to the general negative findings on the accumulation of CH in the erythrocytes as a group, or in plasmocrine droplets, or mitochondria etc. As there appears to be no positive informa- tion on hand in support of these condi- tioned possibilities, it seems necessary to view all negative u-v-fluorescence observa- tions as indicating simply that CH per se is not present in sufficient concentration to be detected.) At best it can only be sug- gested that the absence of measurable ac- cumulations of CH in the intranuclear components must be referable to funda- mental structural and compositional differ- ences between the nuclear and cytoplasmic areas generally.
Relative to nuclear and plasma mem- brunes. The carcinogenic hydrocarbons accumulate in relatively high amounts in the nuclear and plasma membranes of some cell types but not in those of others. Another significant point is that CH always is either (a ) accumulated in both the nu- clear and plasma membranes of a cell or (b) is accumulated in neither of them. Thus all cell types so far reported on in detail fall into one of two categories: (1) cells in which hydrocarbons localize in both the nuclear and plasma membranes (e.g., mouse ascites carcinoma cells, and spermatogenic cells through the cap phase), and (2) cells in which hydrocar- bons localize in neither the nuclear nor plasma membranes (e.g., granular and non-granular leucocytes, thrombocytes, fibroblasts, Walker rat sarcoma cells, HeLa cells, Sertoli cells, and interstitial cells). There appear to be no exceptions to this “tandem rule.” Reversal of accumulation in these membranes may occur temporar- ily, as during cell division (figs. 3-6), or permanently, as during the differential acrosome and maturation phases of the spermatid (figs, 15-20). The precise basis for the “tandem rule” response is not known. It is not surprising though, if, as has been suggested, the outer nuclear mem- brane is a special modification of cyto- plasmic endoplasmic reticulum and plasma membrane (Novikoff, ’59; Watson, ‘55).

INTRACELLULAR CARCINOGENIC HYDROCARBON 219
Even so, it is not evident what the basis is for the accumulation of CH in these mem- branes of some cell types and not in those of others. Information at present indicates simply that CH localization or lack of it in the nuclear and plasma membranes must be referable to fundamental structural, compositional and cytophysiologic differ- ences between the living cells themselves.
Relative to cytoplasmic ground-sub- stance. As indicated previously, there is uniform agreement among all workers that the hydrocarbons accumulate in demon- strable amounts only in the cytoplasmic area. The detailed observations of GrafFi ('39, '40) and Richter ('52a, '55) and those reported here establish that CH are de- finitely accumulated in the cytoplasmic ground-substance, and that it is present here in lowest amounts relative to other specific cytoplasmic components having demonstrable accumulations. The amount though low, however, always exceeds that solubilized in the extracellular medium.
Relative to mitochondria2 and Golgi com- plexes. Interpretations have varied as to the distribution and localization of CH in the mitochondrial and Golgi structures. Ahlstrom ('49) studying frozen sections of epidermal cells identified perinuclear lipid granules as mitochondria and reported that while the hydrocarbons were primarily con- centrated in the mitochondria, there was possibly some accumulation, also, in the Golgi component. His lipid droplets, how- ever, appear to agree more closely with the perinuclear Golgi structures of these cells as described by Montagna ('52) than with the mitochondrial component. Graffi ('39, '40) studying partially glycerinated cells, reported their accumulation in mitochon- dria (and lipochondria) which were identi- fied by Janus green staining. However, it is known that other components of uncon- troversial identities (Golgi acroblast) may be stained by Janus green (Richter, '40). Some of the cells reported on by Graffi have typical, filamentous mitochondrial com- ponents which are recognized by all in addition to his Janus green staining juxta- nuclear granules. He is not clear on what the lipochondria are versus mitochondria, but he described special accumulations in certain structures which he identified as possibly Golgi apparatus. Richter ('52a,
'55), in comparative studies on blood leuco- cytes and fibroblasts by the methods out- lined here, reported the hydrocarbons not to be demonstrable in mitochondria. They were found to accumulate in high concen- trations in the typical Golgi structures of lymphoid cells (figs. 51, 52) and in the cytoplasmic grandations of the granular leucocytes (figs. 44, 45) which were con- sidered to be special Golgi structures (Richter, '42, '55, '56). Fibroblasts, HeLa cells and Walker rat sarcoma cells have characteristic filamentous mitochondria; yet as shown here, CH appear not to ac- cumulate in them but rather in the juxta- nuclear granular aggregates (figs. 27-40). Immature male germ cells having mito- chondrial and Golgi structures of uncon- troversial identities, also, show the hydro- carbons to be concentrated in the Golgi structures and not in the mitochondrial components (figs. 1-14).
While there have been misinterpreta- tions of the identities of mitochondrial and Golgi structures in some instances, and while identities of the Golgi and mitochon- drial components in some cell types have not been firmly established, it would seem that the bulk of the data points to the special accumulation of CH in the Golgi component and certain special derivatives in living cells rather than in the mitochon- drial system.
Relative to secretion products. Little direct information is available on the intra- cellular incorporation of CH in the secre- tory products of living cells. It is the usual finding that secretion products put in their appearance in association with the Golgi complex (Bowen, '26a,b,c,d; Richter, '40, '55, '56; Rose, '60b; Palay, '58). Descrip- tively, there are three types of secretion products : ( 1 ) the plasmocrine type which is essentially a clear homogeneous aqueous droplet (Renaut, '07; Richter, '55, '56), (2) the rhagiocrine type which is essen- tially a clear aqueous droplet in which grains of particulate substance accumulate (Gatenby, '31; Renaut, '07; Richter, '55; Palade, '59), and (3) the lipocrine type which is essentially a discrete lipid droplet reported variously to be formed in associa- tion with either mitochondrial or Golgi structures (Renaut, '07; Palade, '59; Bowen,

220 KENNETH M. RICHTER AND VIRENDER K. SAINI
'26b); although which is not yet definitely established.
So far the only plasmocrine products which have been studied in this connection are those elaborated by eosinophilic and neutrophilic granulocytes and monocytes (Richter, '51, '52a,b, '54, '55). Carcino- genic hydrocarbons do not accumulate in these secretion products, even though the celI is saturated with solubilized and crys- talline CH and the secretion droplets may be associated with adherent carcinogen crystals for many minutes in living cells (Richter, '51a). The vacuolar droplets within human peripheral blood platelets may, also, be equivalent to plasmocrine secretion products; these, too, show no accumulation of CH (fig. 50).
The acrosomal complex has been con- sidered exemplary of a rhagiocrine type of secretion product (figs. 11, 13, 15) (Richter, '55; Fawcett and Burgos, '56; Burgos and Fawcett, '55; Bowen, '24, '25). As shown here, carcinogenic hydrocarbon accumulates in high concentration in the wall only of the acrosomal vacuole (figs. 11-20), not in the main mass of the elabo- rated complex.
Lipid droplets form in practically all kinds of cells and under many assorted cytophysiologic circumstances (Murray and Kopeck, '53) and it is not readily apparent whether they represent true lipid synthesis or an accumulation of lipid from some ex- tracellular source (Palade, '59). Fibro- blasts in culture commonly show lipid droplets and it has been shown that CH accumulate in them in maximal concentra- tions (Graffi, '39, '40; Richter, '52a). The same is true of the lipid inclusions of the interstitial cells of Leydig which are gen- erally thought to be secretion products, al- though not clearly established (Fawcett and Burgos, '56). The lipid inclusions of seba- ceous gland cells, fat cells, and liver cells, as determined by examination of fixed sections, also, show high accumulations of CH (Peacock, '36, '40; Simpson and Cramer, '43a,b; Giinther, '41; Setala and Ekwall, '50b). Doniach, Mottram and Weigert ('43) reported the presence of CH in milk secreted by mice following the i-v-injection of crystalline CH.
At the present limited level of infornia- tion, it seems clear that the incorporation
of CH in the secretion products of living cells, both in vivo and in vitro, varies with the type of secretion product. Their incor- poration is greatest in the lipocrine and lipid-rich secretions generally and minimal in the rhagiocrine type of product; there appears to be no incorporation of CH in the plasmocrine type, but more information on typical glands of all types is needed. With the exception of the observations of Simp- son and Cramer ('43) that lanolin protects the skin against MC carcinogenesis, nothing further seems to have been re- ported on the biologic or physiologic con- sequences of the possible incorporation of CH in secretion products.
SUMMARY By integrated ordinary, phase- and u-v-
fluorescence microscopy, observations have been made directly on (1) the solubiliza- tion of crystalline carcinogenic hydrocar- bons (BP, DBA, MC) in balanced saline, human ascitic fluid, blood plasma, and assorted tissue culture media, and (2) on their penetration, intracellular accumula- tion and specific localization in the various protoplasmic structures of diverse living cell types in vitro which include: male germinal epithelial cells and interstitial cells of the mouse, chick fibroblasts and peripheral blood cells of the adult fowl, human peripheral blood cells, Walker rat sarcoma cells, and HeLa cervical carci- noma cells.
By u-v-fluorescence microscopy it has been demonstrated that crystalline CH are soluble in all aqueous media studied, even though the concentrations may be too low to be measured by conventional methods.
Solubilization, intracellular penetration and accumulation of CH proceed rapidly and solubilized hydrocarbon can be demon- strated intracellularly within seconds and moments following the introduction of crystalline CH into the aqueous extracellu- lar medium.
Non-nucleate peripheral blood erythro- cytes of man and mouse, and the nucleate peripheral blood erythrocytes of the fowl do not show a demonstrable penetration or accumulation of solubilized CH.
With the exception of the peripheral blood erythrocytes, all other living cell types (including human platelets) which have been studied so far show intracellular

INTRACELLULAR CARCINOGENIC HYDROCARBON 22 1
accumulations of solubilized CH which are greatly in excess of even saturated aque- ous concentration levels in the extracellular media. Some cell types may in addition accumulate crystalline CH by phagocytosis (i.e., the granular leucocytes, and Para- mecium).
In all instances in which CH are accu- mulated intracellularly, the hydrocarbons are not demonstrably accumulated within any intranuclear component (exclusive of the nuclear membrane), such as the karyo- lymph, chromatin, nucleolus, or chrome somes of dividing cells.
Carcinogenic hydrocarbons are demon- strable only in the cytoplasmic area where they are distributed differentially in specific sites and in relative concentrations which may exceed by several times 1.0 y/cm3 of protoplasm, and which are constant for each cell type or cytophysiologic phase. The general pattern is as follows:
1, Lipid cytoplasmic inclusions when present and lipocrine secretion products show the highest relative accumulations of CH.
2. In cell types having Golgi structures of uncontroversial identities, this organelle uniformly shows a high accumulation of CH. A special Golgi derivative, i.e., the wall of the acrosomal vacuole, shows a high accumulation. The characterizing granular components of uncertain status of granular leucocytes, platelets, HeLa, Walker rat sarcoma and chick fibroblast cells show equally high accumulations of CH.
3. While varying from one cell type to another, the CH are accumulated in inter- mediate amounts (1) in both the nuclear and plasma membranes or (2) in neither of them.
4. The CH are accumulated in the low- est relative amount in the cytoplasmic matrix, but in an amount which always exceeds that solubilized in the extracellular medium.
5. In cell types having mitochondrial systems of uncontroversial identity, this cytoplasmic component does not show de- monstrable accumulations of CH.
6. Plasmocrine secretion products and vacuolar components show no accumula- tions of CH.
Relative to ( 1 ) intracellular penetration of CH (or lack of it) and (2) to differ- ential intracellular distribution and accu- mulation of CH, present information war- rants only the broad suggestion that specific variations from one cell type to another would appear to be attributable to fundamental compositional, structural and cytophysiologic differences among the liv- ing cells themselves.
LITERATURE CITED Ahlstrom, C. G. 1949 Localization of the car-
cinogenic hydrocarbons within the epithelium of the skin. Exp. Cell Res., 1: 160-162.
Berenblum, I., and P. Shubik 1947 Role of croton oil applications associated with single paintings of carcinogen in tumor production in mouse skin. Brit. J. Canc., 1: 379-382.
Bessis, M. 1956 Cytology of the Blood and Blood-forming Organs. Grune and Stratton, N. Y., pp. 1-629.
Bowen, R. H. 1924 Acrosome of the animal sperm. Anat. Rec., 28: 1-13.
Further notes on the acrosome of animal sperm. Ibid., 31: 201-232.
Studies on the Golgi apparatus in gland cells. I. Glands of alimentary tract. Quart. J. Micr. Sci., 70: 76-112.
Studies on the Golgi apparatus in gland cells. 11. Skin glands. Ibid., 70: 193- 215.
Studies on the Golgi apparatus in gland cells. 111. Lachrymal glands and repro- ductive glands. Ibid., 70: 395-418.
Studies on the Golgi apparatus in gland cells. IV. Critique of topography, structure etc. Ibid., 70: 410-449.
Burgos, M. H., and D. W. Fawcett 1955 Fine structure of the testis. J. Biophys. Biochem.
Clermont, Y., R. E. Glegg and C. Leblond 1955 Presence of carbohydrates in the acrosome of the guinea pig spermatozoid. Exp. Cell Res., 8: 453-457.
Clermont, Y., and C. P. Leblond 1953 Renewal of spermatogonia in the rat. Am. J. Anat., 93: 475-501.
1955 Spermatogenesis of man, monkey, rat, and other mammals as shown by periodic acid-Schiff technique. Ibid., 96: 229-254.
1943a The fluorescence of 3:4-benzpyrene in vivo. I. The distribution of fluorescence at various sites, especially the skin of mice. Brit. J. Exp. Path.,
1943b The fluorescence of 3:4-benz- pyrene in vivo. Part 11: The inter-relationships of the derivatives formed in various sites. Ibid.,
Earle, W. R., and C. Voegtlin 1938 The mode of action of methylcholanthrene on cultures of normal tissues. Am. J. Canc., 34: 373-390.
Earle, W. R. 1943 Production of malignancy in vitro. IV. The mouse fibroblast cultures and changes seen in the living cells. J. Nat. Canc. Inst., 4: 165-212.
1925
1926a
1926b
1926c
1926d
Cytol., 1: 287-300.
Doniach, I., J. C. Mottram and F. Weigert
24: 1-9.
24: 9-14.

222 HENNETH M. RICHTER AND VIRENDER K. SAINI
Erkko, 0. 1955 Quantitative Methods in Histo- logy and Microscopic Histochemistry. Little, Brown and Company, Boston, pp. 1-160.
Fawcett, D. W., and M. H. Burgos 1956 Ob- servations on the cytomorphosis of the germinal and interstitial cells of the human testis. Ciba Foundation Coloquia on Aging. G. E. W. Wolstenholme and E. C. P. Millar, ed. Little, Brown and Co., Boston, 2: 86-99.
Gatenby, J. B., and J. H. Woodger 1921 The cytoplasmic inclusions of the germ cells. Part IX, on the origin of Golgi apparatus on the middle piece of ripe sperm of cavia and the development of acrosomes. Quart. J. Micr. Sci.,
Gatenby, J. B. 1929 Study of Golgi apparatus vacuolar system of Cavia, Helix and Abraxas by intravital methods. Proc. Roy. Soc., B., 104: 302-321.
1931 Note on human spermatid cells supravitally stained in neutral red. Anat. Rec.,
- 1935 The cytoplasmic inclusions in the spermatogenesis of man. Quart. J. Micr. Sci.,
Gey, G. O., P. Shapras and E. Borysko 1954 Ac- tivities and responses of living cells and their components as recorded by cinephase micros- copy and electron microscopy. Ann. N. Y. Acad. Sci., 58: 1089-1095.
Gey, G. O., F. B. Bang, M. K. Gey 1954 Re- sponses of a variety of normal and malignant cells to continuous cultivation, and some prac- tical applications of these responses to problems in the biology of disease. Ibid., 58: 976-999.
Graffi, A. 1939 Zellulare Speicherung cancer- ogener Kohlenwasserstoffe. Ztschr. Krebs- forsch., 49: 477495.
1940a Penetration of cytoplasm by carcinogenic hydrocarbons. (Abstract) Am. J. Cancer, 39: 255.
1940b Intracellulare Benzpyrenspeiche- rung in lebenden Normal und Tumorzellen. Ztschr. Krebsforsch., 50: 196-219.
Gresson, R. A. R., and 1. Zlotnik 1945 A com- parative study of the cytoplasmic components of the male germ cells of certain mammals. Proc. Roy. SOC. Edin., B., 62: 157-170.
Giinther, W. H. 1941 Vber den histologischen Nachweiss des Benzpyrens. Ztschr. Krebs- forch., 52: 57-60.
Haddow, A. 1947 Mode of action of chemical carcinogens. Growth, 1 I: 339-358.
Hoffmann-Berling, H. 1958 The role of cell structures in cell movements. In: Cell, Organ- ism and Milieu, D. Rudnick, ed. Ronald Press
Jordan, H. E. 1938 Comparative hematology. In: The Handbook of Hematology, H. Downey, ed. P. B. Hoeber, Inc., N. Y., vol. 2: pp. 703- 862.
Lange, N. A. 1956 Lange’s Handbook of Chem- istry. Handbook Publishers, Inc., Sandusky, Ohio, 9th ed., pp. 1-1969.
Leblond, C. P., and Y. Clermont 1952 Spermio- genesis of rat, mouse, hamster, and guinea pig as revealed by “periodic acid fuchsin sulfurous acid” technique. Am. J. Anat., 90: 167-216.
65: 265-292.
48: 121-129.
78: 1-29.
CO., N. Y., pp. 45-62.
Lewis, M. R. 1935 The photosensitivity of chick embryo cells growing in media containing cer- tain carcinogenic substances. Am. J. Canc.,
Low, F. N., and J. A. Freeman 1958 Electron Microscopic Atlas of Normal and Leukemic Human Blood. McGraw-Hill Book Comnanv.
25: 305-309.
- -. Jnc., N. Y., pp. 1-347.
Miller, J. A,, and C. A. Baumann 1943a Effect of naphthacene on the fluorescence of hydro- carbons. Canc. Res., 3: 217-222. - 19431, Factors that alter the fluores- cence of certain carcinogens. Ibid., 3: 223-229.
Montagna, W. 1952 The cytology of mammal- ian epidermis and sebaceous glands. Internat. Rev. Cytol., 1: 265-304.
Mueller, G. C., and H. P. Rusch 1943 Irradiated benzpyrene and inhibition of urease activity. Cancer Res., 3: 113-116.
Murray, M. R., and G. Kopeck 1953 A Bibliog- raphy of the Research in Tissue Culture 1884-1950. Academic Press, Inc., N. Y.
Novikoff, A. B. 1959 Approaches to the in vivo function of subcellular varticles. In: Subcellu- lar Particles, T. Hayashi, ed. Ronald Press Co.,
Palade, G. E. 1959 Functional changes in the N. Y., pp. 1-22.
structure of cell components. In: Ibid., pp.
Palay, S. L. 1958 The morphology of secretion. In: Frontiers of Cytology, S. L. Palay, ed. Yale Univ. Press, New Haven, pp. 305-342.
Papanicolaou, G. N., and C. R. Stockard 1918 The development of the idiosome in the germ cell of the male guinea pig. Am. J. Anat., 24: 37-70.
Peacock, P. R. 1936 Evidence regarding the mechanism of elimination of 1:2 benzpyrene, 1 : 2 : 5 : 6-dibenzanthracene and anthracene from the blood stream of injected animals. Brit. J. Exp. Path., 17: 164-172.
1940 Biophysical factors influencing the absorption and distribution of benzpyrene, and their bearing on the mechanism of carcino- genesis. Am. J. Canc., 40: 251-254.
Renaut, J. 1907 Les cellules connectives Rhagi- ocrine. Arch. Anat. Microscop., 9: 495-606.
Richter, K. M. 1940 A study of the cytoplasmic structures in the male germinal cells of several species of Notonecta, with special reference to the Golgi system. J. Morph., 67: 489-521. - 1942 An experimental study of the cytology of human peripheral blood neutro- philes and lymphocytes. Ibid., 71 : 53-75.
The reaction of peripheral blood leucocytes in vitro to certain carcinogen crys- tals. Anat. Rec., 109: 82.
U-V fluorescence studies on the solubility and intracellular localization of cer- tain carcinogens in human blood cells in vitro and in cells of embryonic chick heart cultures. Ibid., 112: 381.
19521, A cinematographic record of the reaction pattern of human peripheral blood leucocytes in vitro to crystalline methylcho- lanthrene. Ibid., 112: 474.
1955 Studies on leukocytic secretory activity. Ann. N. Y. Acad. Sci., 59: 863-895.
64-83.
1951
1952a

INTRACELLULAR CARCINOGENIC HYDROCARBON 223
1956 Studies on the individual and joint effects of histamine, and an antihistaminic on growth, contractility and plasmocrine activ- ity in cultures of embryonic chick heart. J. Cell. Comp. Physiol., 48: 147-166.
A preliminary note on the general chemical characterization of plasmocrine prod- ucts elaborated by human peripheral blood neutrophiles in vitro by microinterferometry. Anat. Rec., 128: 606-607.
Rieck, A. F. 1960 Inhibition of development and regeneration in the forelimb of Amblys- toma, resulting from implants of certain carcin- ogenic crystals. Ibid., 136: (suppl. 2) 265.
Rose, G. G. 1957 Microkinetospheres and VP satellites of pinocytic cells observed in tissue cultures of Gey’s strain HeLa with phase con- trast cinematographic techniques. J. Biophys. Biochem. Cytol., 3: 697-704.
1960a Evidence for an intercellular exchange of cytoplasmic components between associated cells in tissue culture. Tex. Rept. Biol. Med., 18: 103-115.
1960b The Golgi complex in living cells. Presented at the 18th Annual Meeting, Electron Microscope Society of America. To appear in: J. Appl. Physics.
Schlumberger, H. G. 1948-49 Effect of methyl- cholanthrene on frog’s kidney. J. Natl. Cancer Inst., 9: 111-118.
Setala, K., and P. Ekwall 1949 Skin tumors produced in mice by carcinogenic hydrocarbons solubilized in aqueous solutions of association colloids. 11. Acta Path. Microbiol. Scand., 26:
1957
804-808.
- 1950a Penetration of benzpyrene into stomach wall of mouse. Science, 112: 229-231.
1950b Penetration of benzpyrene through the intact skin of newborn mice. Nature, 166: 188-189.
Setaa, K., and P. Ermda 1951 Chylomicrons as carriers for carcinogenic hydrocarbons. Science, 114: 151-152.
Simpson, W. L., C. Carruthers and W. Cramer 1945 Loss of carcinogenic activity when methylcholanthrene is dissolved in anhydrous lanolin. Canc. Res., 5: 1-4.
Simpson, W. L., and W. Cramer 1943a Fluo- rescence studies on carcinogens in skin. I. Histological localization of 20-methylcholan- threne in mouse skin after a single application. Ibid., 3: 363469.
1943b Sebaceous glands and experi- mental skin carcinogenesis in mice. Ibid., 3: 51 5-5 18.
1945d Fluorescence studies of carcino- gens in skin. 11. Mouse skin after single and multiple applications of 20-methylcholanthrene. Ibid., 5: 449-463.
Sensitization of skin by carcino- genically inactive methylcholanthrene to sub- sequent carcinogens. Ibid., 5: 5-10.
Stowell, R. E., and W. Cramer 1942 Effects of solvents in methylcholanthrene epidermal car- cinogenesis. Ibid., 2: 193-197.
Strait, L. A., M. K. Hrenoff and K. B. DeOme 1948 Influence of solvents upon the effective- ness of carcinogenic agents. Ibid., 8: 231-240.
Watson, M. L. 1955 The nuclear envelope. J. Biophys. Biochem. Cytol., I: 257-270.
1945b

All figures are un-retouched, ordinary light (ol), dark-contrast-medium phase (dmp) and u-v fluorescence (uvf) photomicrographs of living cells taken with the 35-mm Leica camera equipped with the Micro-Ibso attachment having a Y3x conical extension tube, or as specified with the 16-mm Cine-Special Camera.
PLATE 1
EXPLANATION OF FIGURES
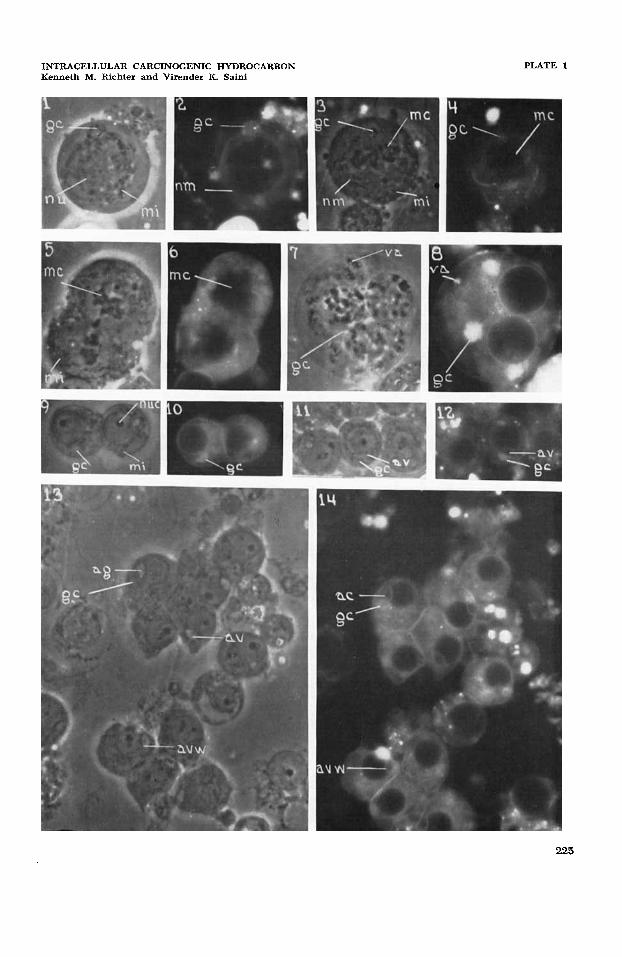
1
2
Primary spermatocyte showing a juxtanuclear Golgi complex (gc), perinuclear mito- chondria (mi), and nucleus (nu); dmp, x 1041. Same cell as in figure 3 showing the differential accumulation of fluorescing benzpyrene in the Golgi complex (gc), and lesser amounts in the nuclear and plasma membranes and in the cytoplasmic ground-substance. There is no intranuclear or mitochondrial fluorescence; uvf, X 1023.
3 Primary spermatocyte in hrst meiotic division showing metaphase plate chromosomes (mc), polar Golgi complexes (gc), dispersed mitochondria (mi), and possibly portion of nuclear membrane (nm); dmp, X 1041.
Same cell as in figure 3 showing a high accumulation of benzpyrene in the polar Golgi complexes (gc), remnant of the nuclear membrane, and low accumulation in the cyto- plasmic ground substance. Metaphase plate chromosomes and spindle, and mitochondria show no fluorescence; uvf, X 1023.
5 Incompletely cleaved multipolar spermatocyte showing two metaphase plate sets of chromosomes (mc), granular to filamentous mitochondria (mi), and multiple polar Golgi structures (gc); dmp, x 1041.
4
6
7
8
9
10
11
12
13
14
Same cell as in figure 5 showing high accumulation of benzpyrene in the polar Golgi structures and remnants of the nuclear membrane, and low accumulation in the cyto- plasmic ground-substance. Chromosomes (mc), spindle, and mitochondria show no fluorescence; uvf, x 1023. Multinucleate spermatocyte showing aggregates of vesicular structures of uncertain identity (va) and several juxtanuclear Golgi structures (gc); dmp, x 1041. Same cell as in figure 7 showing a high accumulation of benzpyrene in the juxtanuclear Golgi structures (gc), nuclear and plasma membranes, lower accumulation in the vesicular aggregates (va), and lowest accumulation in the cytoplasmic ground-substance. Intranuclear substance and mitochondria show no fluorescence; uvf, x 1023. Young Golgi phase spermatids showing mitochondria (mi), acroblast Golgi structure (gc). A nucleolus (nuc) is prominent in each spermatid; dmp, x 1041. Same cells as in figure 9 showing a high accumulation of benzpyrene in the Golgi structure (gc), slightly lower accumulations in the cytoplasmic ground-substance. Nucleoli and other intranuclear substance, and mitochondria show no fluorescence; uvf, x 1023. Slightly older Golgi phase spermatids in an early phase of formation of the acrosomal vacuole (av) by the acroblast Golgi structure (gc); dmp, X 1041.
Same cell as in figure 11 showing highest accumulation of benzypyrene in the acroblast Golgi structure (gc), intermediate accumulations in the wall of the acrosomal vacuole (avw) and the nuclear and plasma membranes, and lowest accumulation in the cyto- plasmic ground-substance. The intranuclear substance, the filling substance of the acrosomal vacuole, and the mitochondria show no benzpyrene fluorescence; uvf, x 1023. Spermatids in a phase of acrosomal complex formation showing the acrosomal granule (ag), acrosomal vacuole (av), the wall of the acrosomal vacuole (avw), the acroblast Golgi structure (gc), and mitochondrial rods and granules; dmp, x 1041. Same cells as in figure 13 showing highest accumulation of fluorescent benzpyrene in the Golgi structures, next highest accumulations in the nuclear and plasma membranes and wall of the acrosomal vacuole (avw), and lowest accumulation in the cytoplasmic ground-substance. Intranuclear substance, acrosomal granule and vacuole, and mito- chondria show no fluorescence; uvf, X 1023.
224
INTRACELLULAR CARCINOGENIC HYDROCARBON Kenneth M. Richter and Virender K. Saini
PLATE 1
225

PLATE 2 EXPLANATION OF FIGURES
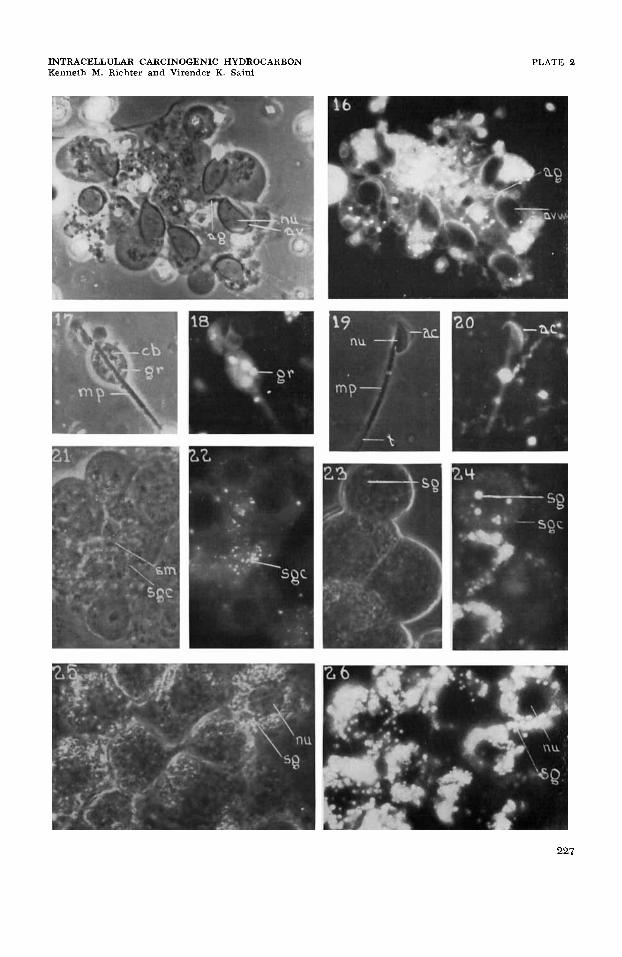
15
16
17
18
19
20
21
22
23
24
25
26
Spermatids in cap and acrosome phases showing nuclear elongation (nu) , spread of acrosomal vacuole (head cap) (av) over nucleus and the terminally situated acrosomal granule (ag), and formation of granular cytoplasmic remnant; dmp, x 1041. Same cells as in figure 15 showing high accumulations of fluorescent benzpyrene in the wall of the acrosomal vacuole (avw), and the cytoplasmic remnant. There is no fluo- rescence in the intranuclear area, the acrosomal vacuolar substance, the acrosomal granule (ag), nuclear and plasma membranes; uvf, X 1023.
Maturation phase spermatid showing the cytoplasmic remnant or hleb (cb) containing the Golgi remnants (gr), and the mitochondria1 spiral-mid-piece sheath (mp); dmp, X 1041.
Same cell as in figure 17 showing high accumulations of benzpyrene in the cytoplasmic remnant and Golgi remnant (gr), and the acrosomal vacuolar wall, and reduced accumu- lation in the cytoplasmic ground substance in the region of the mid-piece; uvf, x 1023. Mature sperm showing the nucleus (nu) , acrosomal complex (ac), neck, mitochondrial sheath (mp), and tail ( t ) ; dmp, X 1041.
Same cell as in figure 19 showing a high accumulation of benzpyrene in the wall of the acrosomal vacuole (avw) and a low accumulation in the cytoplasmic ground-substance of the mid-piece area. There is no fluorescence in the intranuclear area, the acrosomal vacuolar substance and acrosomal granule, mitochondrial sheath, or axial filament; uvf, X 1023.
Sertoli cell mass (sm) and associated group of spermatids. The Sertoli cell mass shows cytoplasmic rods and filaments and small granule component (sgc); the former is thought to be mitochondrial, the latter to be Golgi complex; dmp, x 1041. Same cell group as in figure 21 showing a high accumulation of benzpyrene in the small granule component, and low accumulations in the cytoplasmic ground-substance. The intranuclear area, the nuclear and plasma membranes, and the filamentous cytoplasmic component show no fluorescence; uvf, X 1023.
A group of interstitial cells in an early phase showing nuclei, scattered lipid secretion granules (sg), and small cytoplasmic granule component; dmp, x 1041. Same cell group as in figure 23 showing very high accumulations of benzpyrene in the secretion granules (sg), intermediate accumulations in the small granule component (sgc), and very low accumulations in the cytoplasmic ground substance. The intranu- clear area, nuclear and plasma membranes show no fluorescence; uvf, x 1023. Interstitial cell group in advanced secretory phase showing nuclei (nu) , and massive accumulations of lipid secretion granules (sg); dmp, x 1041. Same cell group as in figure 25 showing a high accumulation of benzpyrene in the secretion granules ( sg ) and very low accumulations in the cytoplasmic ground-substance. The intranuclear area, and the nuclear and plasma membranes show no fluorescence; uvf, x 1023.
226
INTRACELLULAR CARCINOGENIC HYDROCARBON Kenneth M. Richter and Virender K. Saiiii
PLATE 2
227

PLATE 3
EXPLANATION OF FIGURES
27
28
29
30
31
32
33
34
Growth zone of a living 4-day culture of Walker rat sarcoma grown on a bed of crystal- line methylcholanthrene; 01, x 190.
Same cell group as in figure 27 showing a high accumulation of fluorescing methylcho- lanthrene within the living cells relative to the extracellular medium and its localization in the general cytoplasmic area and cytoplasmic granulations; uvf, x 190.
Two living Walker rat sarcoma cells from a 4-day culture grown on a bed of crystalline methylcholanthrene; 01, x 1023.
Same cells as in figure 29 showing a high accumulation of methylcholanthrene in the cytoplasmic granulations and a very low accumulation in the cytoplasmic ground sub- stance. The nucleus and its contents show no fluorescence; uvf, x 1023.
Growth zone of living two-day culture of embryonal chick heart fibroblasts grown on a crystalline bed of benzpyrene; dmp, x 190.
Same cell group as in figure 31 showing a high accumulation of solubilized benzpyrene in the cells relative to the extracellular medium, and its accumulation in the cytoplasmic area generally and in the cytoplasmic granules; uvf, x 190.
Living chick heart fibroblasts from a two-day culture grown on a bed of crystalline benzpyrene showing nuclear detail, fine juxtanuclear granules, some fat droplets, and filamentous mitochondria; dmp, x 1041.
Same cell group as in figure 33 showing a high accumulation of benzpyrene in the fine cytoplasmic granules and lipid droplets and a low accumulation in the cytoplasmic ground substance. The nucleus and its contents and the mitochondria show no Auores- cence; uvf, x 1023.
228
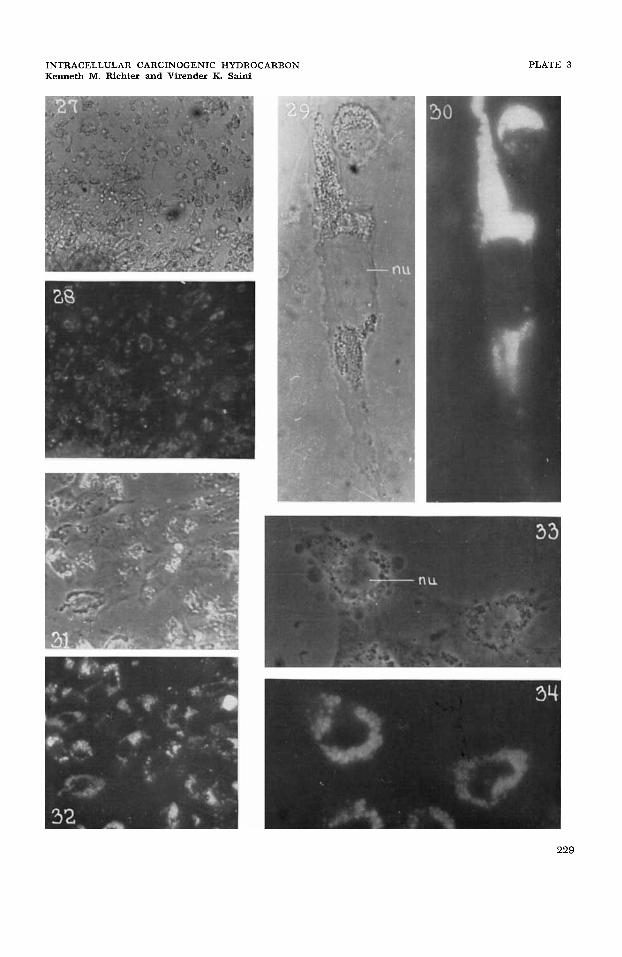
INTRACELLULAR CARCINOGENIC HYDROCARBON Kenneth M. Richter and Virender K. Saini
PLATE 3
229

PLATE 4
EXPLANATION OF FIGURES
35
36
37
38
39
40
41
42
43
A living HeLa cell in a culture to which crystalline benzpyrene had been added 15 minutes previously showing the nuclear detail (nu) , the cytocentrum area (gz) (Golgi zone of some workers), filamentous mitochondria (mi) , juxtanuclear cytoplasmic granules and ectoplasmic granules (eg) or bulbous cytoplasmic processes; dmp, X 1041.
Same cell as in figure 35 showing a high accumulation of benzpyrene in the cytoplasmic juxtanuclear granules, an intermediate accumulation in the area of the cytocentrum (gz) and a low accumulation in the cytoplasmic ground substance. Nuclear components and mitochondria show no fluoresence; uvf, x 1023. Living HeLa cells in a culture to which crystalline benzpyrene had been added 15 minutes previously showing ( a ) all structural features seen in figure 35 and 36 except the ectoplasmic granules, and ( b ) delicate intercellular cytoplasmic strands (cs); dmp, x 1041. Same cells as in figure 37 showing high accumulations of benzpyrene in the dispersed fine cytoplasmic granules, a n intermediate accumulation in the cytocentrum area, and a low accumulation in the cytoplasmic ground substance and intercellular cytoplasmic strands. Nuclear components, mitochondria, and nuclear and plasma membranes show no accumulation; uvf, x 1023. Living HeLa cells from a 6-day monolayer culture grown on a bed of crystalline benz- pyrene showing typical cellular detail; dmp, x 1041. Same cells as in figure 39 showing the differential accumulation of benzpyrene in high amounts in the dispersed fine cytoplasmic granules and in low amounts in the cyto- plasmic ground substance. Nuclear components, mitochondria, and the nuclear and plasma membranes show no accumulation; uvf, x 1023. A frame from a motion picture sequence of a living human neutrophilic granular leucocyte in whole peripheral blood which has phagocytosed 6-8 crystals of methylcho- lanthrene (mc) in vitro; dmp, x 2107. A frame from a motion pitcure sequence of a living human neutrophilic granular leucocyte which has phagocytosed a compound, cross-shaped crystal of methylcho- lanthrene in vitro; dmp, x 2107. Same cell as in figure 42 but from a frame approximately 10 seconds later showing a partial rotation of the methylcholanthrene crystal within the cell cytoplasm; dmp, X 2107.
230

INTRACELLULAR CARCINOGENIC HYDROCARBON Kenneth M. Richter and Virender K. Saini
PLATE 4
231

PLATE 5
EXPLANATION OF FIGURES
44 A spread of fresh whole human peripheral blood after 8 hours in vitro over a bed of crystalline methylcholanthrene showing three living neutrophilic granular leucocytes (ng) , several non-hemolysed erythrocytes ( rc) and numerous hemolysed erythrocytes (hrc); 01, x 1170.
Same cells as in figure 44 showing ( a ) high intracellular accumulation of solubilized methylcholanthrene in the cytoplasmic area and specific granules of the granular leuco- cytes relative to that in the plasma, and ( b ) no intracellular accumulation of methylcho- lanthrene in either (1) the residues of the hemolysed erythrocytes (hrc) or (2) the non-hemolysed erythrocytes ( rc) ; uvf, x 1170.
Two frames about 10 seconds apart from a motion picture sequence of whole human peripheral blood set-up over a bed of crystalline methylcholanthrene (mc) show- ing a phagocytosed methlycholanthrene crystal being carried along by a neutrophilic granular leucocyte. A fixed, extracellular crystal (x) is a reference point; dmp, X 2107.
48 A portion of a monolayer 6-day culture of HeLa cells grown on a bed of crystalline benzpyrene showing a high accumulation of solubilized benzpyrene in the cytoplasmic granules and cytoplasmic area relative to that in the extracellular medium; uvf, x 190.
45
46 and 47
232
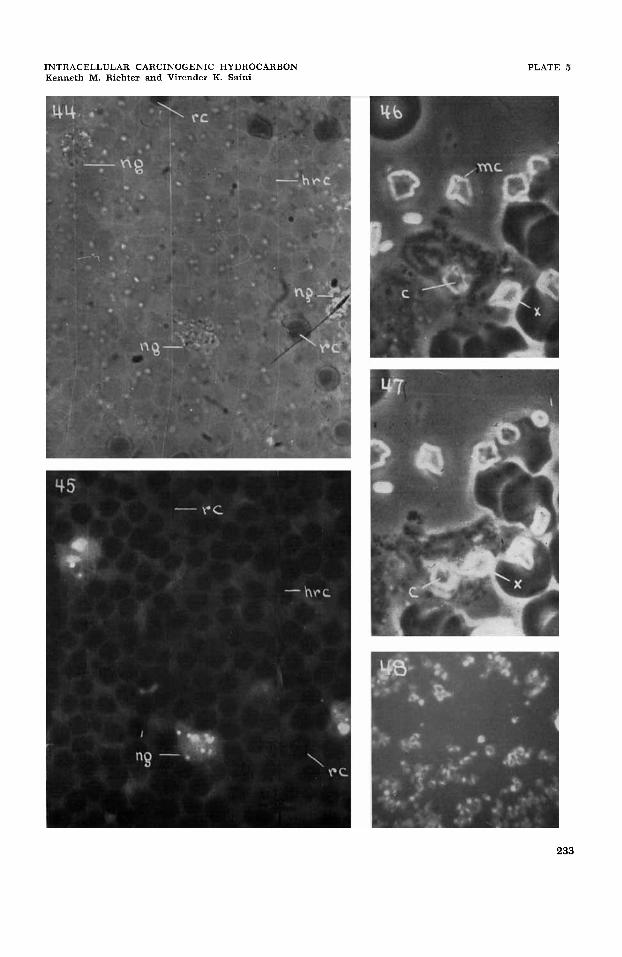
INTRACELLULAR CARCINOGENIC HYDROCARBON Kenneth M. Richter and Virender K. Saini
PLATE 5
233

PLATE 6
EXPLANATION O F FIGURES
49 Frame from a motion picture sequence showing the early aggregation of granular leuco- cytes, neutrophilic (ng) and eosinophilic (eg), i n a microcrystalline pile of methycholan- threne (mc); dmp, x 2107.
Human peripheral blood showing relatively low and high accumulations of BP respec- tively in the cytoplasmic ground-substance and the specific granulations of blood platelets (pl). The characteristic vacuolar or plasmocrine droplets (pd) in the platelets show no accumulation of BP; uvf, x 1023.
51 Fresh domestic fowl peripheral blood showing ( 1 ) nucleate erythrocytes (rc), ( 2 ) nucleate thrombocytes ( thr) , ( 3 ) a lymphoid cell (lc) with filamentous mitochondria and juxtanuclear Golgi bodies, and (4 ) granular leucocytes (gl); dmp, x 1041.
Same cell group as in figure 51 showing the presence of solubilized BP in the blood plasma and its general differential accumulation in relatively high amounts in the cytoplasmic area and inclusions of the lymphoid cell, the thrombocytes, and the granu- lar leucocytes. There is no accumulation of BP in the erythrocytes; uvf, x 1023.
A film of fresh human peripheral blood near a bed of amorphic crystalline BP (vp) show- ing a typical u-v-fluorescence intensity-gradient zone or concentration-gradient zone (arrow) established within a few minutes by the progressive solubilization and diffusion of BP into the plasma (p) . At zone “s” the plasma contains a saturated or maximal amount of solubilized BP; and at zone “0” it contains virtually no solubilized BP. The time-sequence of intracellular accumulation of solubilized BP is evidenced by several neutrophilic granular leucocytes (ng) and platelets (pl) ; uvf, x 518.
50
52
53
234
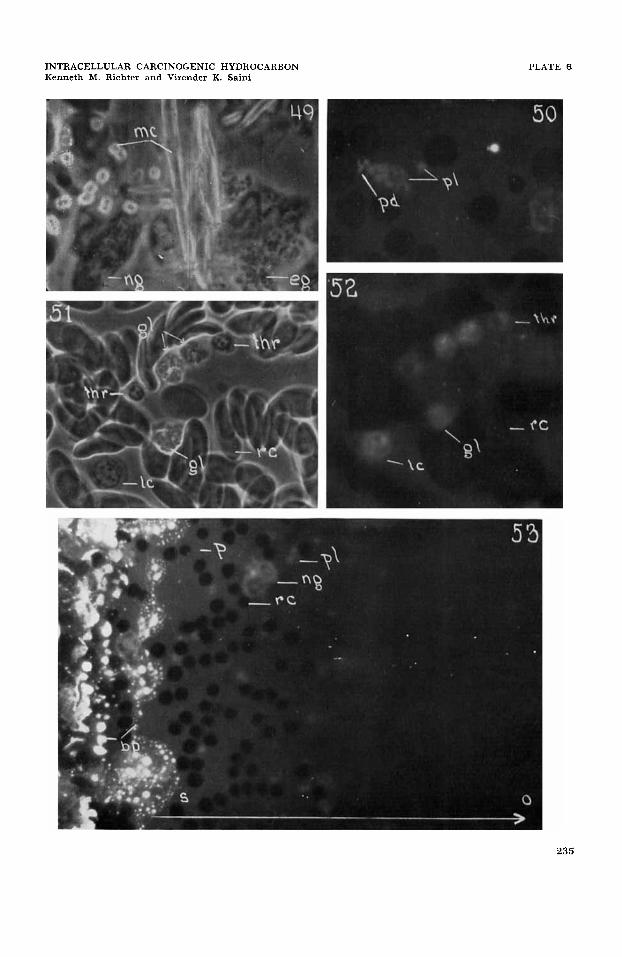
INTRACELLULAR CARCINOGENIC HYDROCARBON Kenneth M. Richter and Virender K. Saini
PLATE 6
235




















