
Twitching Motility Is Essential for Virulence in Dichelobacter nodosus
13
JOURNAL OF BACTERIOLOGY, May 2008, p. 3323–3335 Vol. 190, No. 9 0021-9193/08/$08.000 doi:10.1128/JB.01807-07 Copyright © 2008, American Society for Microbiology. All Rights Reserved. Twitching Motility Is Essential for Virulence in Dichelobacter nodosus † Xiaoyan Han, 1,2 Ruth M. Kennan, 1,2 John K. Davies, 1,2 Leslie A. Reddacliff, 3 Om P. Dhungyel, 4 Richard J. Whittington, 1,4 Lynne Turnbull, 2 Cynthia B. Whitchurch, 2 and Julian I. Rood 1,2 * Australian Research Council Centre of Excellence in Structure and Functional Microbial Genomics 1 and Department of Microbiology, Monash University, 2 Victoria 3800, Australia; Elizabeth Macarthur Agricultural Institute, NSW Department of Primary Industries, PMB 8, Camden, New South Wales 2570, Australia 2 ; and Faculty of Veterinary Science, University of Sydney, PMB 3, Camden, New South Wales 2570, Australia 4 Received 15 November 2007/Accepted 20 February 2008 Type IV fimbriae are essential virulence factors of Dichelobacter nodosus, the principal causative agent of ovine foot rot. The fimA fimbrial subunit gene is required for virulence, but fimA mutants exhibit several phenotypic changes and it is not certain if the effects on virulence result from the loss of type IV fimbria- mediated twitching motility, cell adherence, or reduced protease secretion. We showed that mutation of either the pilT or pilU gene eliminated the ability to carry out twitching motility. However, the pilT mutants displayed decreased adhesion to epithelial cells and reduced protease secretion, whereas the pilU mutants had wild-type levels of extracellular protease secretion and adherence. These data provided evidence that PilT is required for the type IV fimbria-dependent protease secretion pathway in D. nodosus. It was postulated that sufficient fimbrial retraction must occur in the pilU mutants to allow protease secretion, but not twitching motility, to take place. Although no cell movement was detected in a pilU mutant of D. nodosus, aberrant motion was detected in an equivalent mutant of Pseudomonas aeruginosa. These observations explain how in D. nodosus protease secretion can occur in a pilU mutant but not in a pilT mutant. In addition, virulence studies with sheep showed that both the pilT and pilU mutants were avirulent, providing evidence that mutation of the type IV fimbrial system affects virulence by eliminating twitching motility, not by altering cell adherence or protease secretion. Foot rot is a highly contagious and economically significant bacterial disease affecting sheep in most countries. Type IV fimbriae, extracellular proteases, and several genomic islands are potential virulence factors of Dichelobacter nodosus, the principal causative agent of ovine foot rot (3, 23, 43). Type IV fimbriae are major surface antigens of D. nodosus, and more than 30 potential orthologs of type IV fimbrial machinery genes have been identified in the D. nodosus genome (17, 35). By analyzing specific mutants constructed by allelic exchange, we have shown that the expression of the fimbrial subunit gene fimA is mediated by an RpoN-dependent PilS/PilR two-com- ponent signal transduction system (39) and that FimN, FimO, FimP, and PilE are required for fimbrial biogenesis and func- tion (17). Previous studies have shown that the fimA gene is required for virulence in sheep (23). However, fimA mutants of D. nodosus exhibited several phenotypic changes: they were nonpiliated and defective in twitching motility, and they se- creted reduced amounts of extracellular proteases. Therefore, it was not certain that the effects on virulence resulted from reduced fimbria-mediated cell adherence, loss of twitching mo- tility, or reduced protease secretion. Type IV fimbriae mediate twitching motility, a process that enables bacteria to move along solid surfaces (27, 52). Twitch- ing motility involves fimbrial extension and retraction, which reflects fimbrial subunit polymerization and depolymerization events (30, 47) that involve conserved cytosolic hexameric ATPases (21, 25, 31, 45). One of these proteins, PilT, drives fimbrial retraction (33), generating forces of more than 100 pN to power twitching motility (26, 32). By contrast, the FimN protein of D. nodosus (17) or the orthologous PilB protein of Pseudomonas aeruginosa (36) acts as an antagonist of PilT by promoting fimbrial polymerization. A third ATPase, PilU, which has a high level amino acid sequence similarity to PilT, is also required for fimbrial retraction in P. aeruginosa (54). Both pilT and pilU null strains of P. aeruginosa are piliated but are not motile (53, 54). Orthologs of PilT and PilU have been identified in other organisms, such as Neisseria gonorrhoeae and Neisseria menin- gitidis (38, 42, 56), as well as Pseudomonas stutzeri (15). The PilT proteins of N. gonorrhoeae and P. stutzeri are also required for twitching motility. However, in contrast to P. aeruginosa pilU mutants, pilU mutants of N. gonorrhoeae and P. stutzeri still exhibit twitching motility. Mutation of pilT leads to resistance to the fimbria-specific bacteriophage PO4 in P. aeruginosa or to a defect in DNA uptake in N. gonorrhoeae and P. stutzeri, whereas the equivalent pilU mutants still exhibit PO4 phage susceptibility and DNA uptake phenotypes (15, 38, 53, 54). Other studies have shown that P. aeruginosa PilT and PilU are required for corneal infections (60) and for virulence in a mouse model of acute pneumonia (7). Twitching motility me- diated by PilT also appears to be essential for invasion by the plant pathogen Azoarcus sp. (4). In this paper, we describe characterization of two D. nodosus * Corresponding author. Mailing address: Department of Microbiol- ogy, Monash University, Victoria 3800, Australia. Phone: (613) 9905 4825. Fax: (613) 9905 4811. E-mail: [email protected]. † Supplemental material for this article may be found at http://jb .asm.org/. Published ahead of print on 29 February 2008. 3323
Transcript of Twitching Motility Is Essential for Virulence in Dichelobacter nodosus

JOURNAL OF BACTERIOLOGY, May 2008, p. 3323–3335 Vol. 190, No. 90021-9193/08/$08.00�0 doi:10.1128/JB.01807-07Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Twitching Motility Is Essential for Virulence in Dichelobacter nodosus�†Xiaoyan Han,1,2 Ruth M. Kennan,1,2 John K. Davies,1,2 Leslie A. Reddacliff,3 Om P. Dhungyel,4
Richard J. Whittington,1,4 Lynne Turnbull,2 Cynthia B. Whitchurch,2 and Julian I. Rood1,2*Australian Research Council Centre of Excellence in Structure and Functional Microbial Genomics1 and Department of
Microbiology, Monash University,2 Victoria 3800, Australia; Elizabeth Macarthur Agricultural Institute,NSW Department of Primary Industries, PMB 8, Camden, New South Wales 2570, Australia2; and
Faculty of Veterinary Science, University of Sydney, PMB 3, Camden,New South Wales 2570, Australia4
Received 15 November 2007/Accepted 20 February 2008
Type IV fimbriae are essential virulence factors of Dichelobacter nodosus, the principal causative agent ofovine foot rot. The fimA fimbrial subunit gene is required for virulence, but fimA mutants exhibit severalphenotypic changes and it is not certain if the effects on virulence result from the loss of type IV fimbria-mediated twitching motility, cell adherence, or reduced protease secretion. We showed that mutation of eitherthe pilT or pilU gene eliminated the ability to carry out twitching motility. However, the pilT mutants displayeddecreased adhesion to epithelial cells and reduced protease secretion, whereas the pilU mutants had wild-typelevels of extracellular protease secretion and adherence. These data provided evidence that PilT is required forthe type IV fimbria-dependent protease secretion pathway in D. nodosus. It was postulated that sufficientfimbrial retraction must occur in the pilU mutants to allow protease secretion, but not twitching motility, totake place. Although no cell movement was detected in a pilU mutant of D. nodosus, aberrant motion wasdetected in an equivalent mutant of Pseudomonas aeruginosa. These observations explain how in D. nodosusprotease secretion can occur in a pilU mutant but not in a pilT mutant. In addition, virulence studies with sheepshowed that both the pilT and pilU mutants were avirulent, providing evidence that mutation of the type IVfimbrial system affects virulence by eliminating twitching motility, not by altering cell adherence or proteasesecretion.
Foot rot is a highly contagious and economically significantbacterial disease affecting sheep in most countries. Type IVfimbriae, extracellular proteases, and several genomic islandsare potential virulence factors of Dichelobacter nodosus, theprincipal causative agent of ovine foot rot (3, 23, 43). Type IVfimbriae are major surface antigens of D. nodosus, and morethan 30 potential orthologs of type IV fimbrial machinerygenes have been identified in the D. nodosus genome (17, 35).By analyzing specific mutants constructed by allelic exchange,we have shown that the expression of the fimbrial subunit genefimA is mediated by an RpoN-dependent PilS/PilR two-com-ponent signal transduction system (39) and that FimN, FimO,FimP, and PilE are required for fimbrial biogenesis and func-tion (17). Previous studies have shown that the fimA gene isrequired for virulence in sheep (23). However, fimA mutants ofD. nodosus exhibited several phenotypic changes: they werenonpiliated and defective in twitching motility, and they se-creted reduced amounts of extracellular proteases. Therefore,it was not certain that the effects on virulence resulted fromreduced fimbria-mediated cell adherence, loss of twitching mo-tility, or reduced protease secretion.
Type IV fimbriae mediate twitching motility, a process thatenables bacteria to move along solid surfaces (27, 52). Twitch-
ing motility involves fimbrial extension and retraction, whichreflects fimbrial subunit polymerization and depolymerizationevents (30, 47) that involve conserved cytosolic hexamericATPases (21, 25, 31, 45). One of these proteins, PilT, drivesfimbrial retraction (33), generating forces of more than 100 pNto power twitching motility (26, 32). By contrast, the FimNprotein of D. nodosus (17) or the orthologous PilB protein ofPseudomonas aeruginosa (36) acts as an antagonist of PilT bypromoting fimbrial polymerization. A third ATPase, PilU,which has a high level amino acid sequence similarity to PilT,is also required for fimbrial retraction in P. aeruginosa (54).Both pilT and pilU null strains of P. aeruginosa are piliated butare not motile (53, 54).
Orthologs of PilT and PilU have been identified in otherorganisms, such as Neisseria gonorrhoeae and Neisseria menin-gitidis (38, 42, 56), as well as Pseudomonas stutzeri (15). ThePilT proteins of N. gonorrhoeae and P. stutzeri are also requiredfor twitching motility. However, in contrast to P. aeruginosapilU mutants, pilU mutants of N. gonorrhoeae and P. stutzeri stillexhibit twitching motility. Mutation of pilT leads to resistanceto the fimbria-specific bacteriophage PO4 in P. aeruginosa or toa defect in DNA uptake in N. gonorrhoeae and P. stutzeri,whereas the equivalent pilU mutants still exhibit PO4 phagesusceptibility and DNA uptake phenotypes (15, 38, 53, 54).Other studies have shown that P. aeruginosa PilT and PilU arerequired for corneal infections (60) and for virulence in amouse model of acute pneumonia (7). Twitching motility me-diated by PilT also appears to be essential for invasion by theplant pathogen Azoarcus sp. (4).
In this paper, we describe characterization of two D. nodosus
* Corresponding author. Mailing address: Department of Microbiol-ogy, Monash University, Victoria 3800, Australia. Phone: (613) 9905 4825.Fax: (613) 9905 4811. E-mail: [email protected].
† Supplemental material for this article may be found at http://jb.asm.org/.
� Published ahead of print on 29 February 2008.
3323

genes, pilT and pilU, which encode orthologs of the P. aerugi-nosa PilT and PilU proteins. We used natural transformationand homologous recombination to isolate several chromo-somal pilT and pilU mutants, and we describe the effects of themutations in these mutants on twitching motility, natural com-petence, in vitro cell adherence, protease secretion, and viru-lence in sheep. Our results revealed significant differences be-tween D. nodosus and other type IV fimbriate organisms,including Pseudomonas and Neisseria spp. In particular, weshowed that PilT, but not PilU, is required for extracellularprotease secretion and that twitching motility is essential forvirulence.
MATERIALS AND METHODS
Strains, plasmids, and growth conditions. Bacterial strains and plasmids usedin this study are listed in Table 1. Escherichia coli DH5� was used for plasmidpropagation and cloning experiments and was grown at 37°C in 2�YT medium(44). D. nodosus strain VCS1703A and its derivatives were grown in a Coy
anaerobic chamber (Coy Laboratory Products Inc.) as previously described (17,23). D. nodosus strains used in the sheep virulence trial were grown on hoof agar(49). P. aeruginosa strains were grown at 37°C in Luria-Bertani (LB) medium(44).
DNA manipulations and molecular techniques. Unless otherwise stated, stan-dard procedures were used for DNA manipulation and molecular techniques, asdescribed elsewhere (17, 44). Nucleotide and amino acid sequence comparisonswere performed by using the National Center for Biotechnology InformationBLAST server (http://www.ncbi.nlm.nih.gov/BLAST). Amino acid sequenceswere aligned by using the Network Protein Sequence Analysis server provided byPole Bioinformatique lyonnais (http://npsa-pbil.ibcp.fr). Reverse transcriptasePCR (RT-PCR) and quantitative RT-PCR were performed as described previ-ously (17, 39). The sequences of synthetic oligonucleotides are shown in Table S1in the supplemental material.
Mutant construction and confirmation. To construct a pilT suicide vector, a1.5-kb fragment containing the pilT gene was amplified from VCS1703A chro-mosomal DNA with primers JRP1795 and JRP1628 and cloned into the XhoI/BamHI sites of pBluescript II SK� to form pJIR2660. The suicide plasmidpJIR2662 was constructed by inserting a 1.1-kb SmaI/EcoRI fragment that con-tained an erythromycin resistance gene, erm(B), into the T4 polymerase-filledEcoRI site of pJIR2660. In this construct, the erm(B) gene was flanked on each
TABLE 1. Bacterial strains and plasmids
Strain or plasmid Characteristics Source or reference
E. coli DH5� F� endA1 hsdR17(rK� mK
�) thi-1 �� recA1 gyrA96 relA1 rhoA supE44 deoR�80dlacZ�M15 �(lacZYA argF)U169
Invitrogen
D. nodosus strainsVCS1703A Serogroup G, transformable virulent isolate 23JIR3727 VCS1703A fimA �tet(M) 23JIR3770 VCS1703A pilU �erm(B) (pilU1) Natural transformation with
pJIR2581JIR3771 VCS1703A pilU �erm(B) (pilU2) Natural transformation with
pJIR2581JIR3880 VCS1703A pilT �erm(B) (pilT1) Natural transformation with
pJIR2662JIR3881 VCS1703A pilT �erm(B) (pilT2) Natural transformation with
pJIR2662JIR3898 JIR3771 rrnA �pilTU� (pilU2/pilU�) Natural transformation with
pJIR2864JIR3943 JIR3771 rrnA �pilT� Natural transformation with
pJIR3528
P. aeruginosa strainsPAK Wild type D. Bradley, Memorial
University of Newfoundland,St. John’s, Canada
PAKpilA PAK pilA::TcR; previously referred to as AWK 51R364 PAK pilT::Tn5-B21 53S237 PAK pilU::Tn5-B21 54
PlasmidspBluescript IISK�
Apr, lacZ� cloning vector Stratagene
pWSK29 Apr, lacZ� cloning vector 50pUC18K pUC18 SmaI �aphA-3, nonpolar base vector 28pJIR1532 pBluescript II SK� containing tet(M) located between D. nodosus rrnA promoter
and terminator22
pJIR2549 pBluescript II SK� XbaI/BamHI �(1.4-kb PCR product containing pilU�) RecombinantpJIR2581 pJIR2549 T4-filled NarI �erm(B) RecombinantpJIR2582 pUC18K XbaI/BamHI �(650-bp PCR product containing 3 fragment of pilT) RecombinantpJIR2611 pJIR2582 T4-filled EcoRI �(680-bp PCR product containing 5 fragment of
pilT)Recombinant
pJIR2660 pBluescript II SK� XhoI/BamHI �(1.5-kb PCR product containing pilT�) RecombinantpJIR2662 pJIR2660 T4-filled EcoRI �erm(B) RecombinantpJIR2691 pWSK29 containing tet(M) located between D. nodosus rrnA promoter and
terminator17
pJIR2864 pJIR2691 EcoRV �(2.6-kb PCR product containing pilTU operon) RecombinantpJIR3528 pJIR2691 EcoRV �(1.6-kb PCR product containing pilT) Recombinant
3324 HAN ET AL. J. BACTERIOL.

side by 750-bp fragments of the contiguous D. nodosus sequence. To construct anonpolar pilT suicide vector, a 650-bp PCR product that was amplified withprimers JRP1644 and JRP1645 and contained the 3 end of pilT was cloned intoXbaI/BamHI sites of pUC18K, which carries a nonpolar cassette that containsthe kanamycin resistance gene aphA-3 (28). The 680-bp 5 pilT fragment thenwas amplified using primers JRP1642 and JRP1643 and inserted into the T4polymerase-filled EcoRI site of the resultant plasmid, pJIR2582, to obtain thenonpolar pilT suicide vector pJIR2611.
To construct a pilU suicide vector, a 1.4-kb PCR fragment containing the pilUgene was amplified with primers JRP1627 and JRP1629 and cloned into theXbaI/BamHI sites of pBluescript II SK� to construct plasmid pJIR2549. TheSmaI/EcoRI erm(B) cassette was cloned into the T4 polymerase-filled NarI siteof pJIR2549 to form the pilU suicide vector pJIR2581. There was a 750-bpfragment upstream and a 650-bp fragment downstream of erm(B) in pJIR2581.
Mutants (Table 1) were constructed by homologous recombination using nat-ural transformation (23). Transformants were initially screened by determiningresistance to the appropriate antibiotics and then performing a capillary PCRanalysis (17). Potential mutants were further analyzed by PCR using 16S rrnAgene-, relevant antibiotic resistance gene-, and pilT- and pilU-specific primers(see Table S1 in the supplemental material). Southern hybridization analysis andomp1 PCR-restriction fragment length polymorphism analysis were performedas described previously (14, 17, 22).
Natural transformation and complementation of pilU mutants. PlasmidspJIR1532 and pJIR2691 (Table 1), which are suicide vectors containing thetet(M) tetracycline resistance gene flanked by the D. nodosus rrnA promoter andterminator regions, were used to test the abilities of the mutants to undergonatural transformation (17). To complement the pilU mutation, a 2.6-kb PCRproduct produced with primers JRP1795 and JRP1627 and containing the wild-type pilTU region was cloned into the EcoRV site of pJIR2691 to construct thepilU complementation plasmid pJIR2864, which was subsequently introducedinto the pilU mutants JIR3770 and JIR3771 by natural transformation. Comple-mentation derivatives were selected on medium containing both kanamycin andtetracycline and confirmed by PCR analysis using the pilU primers JRP1627 andJRP1629 and the tet(M) primers JRP1138 and JRP1139. Plasmid pJIR3528,which contained the wild-type pilT gene, was constructed by cloning a 1.6-kbPCR product generated with primers JRP1795 and JRP1645 into the EcoRV siteof pJIR2691 and was introduced into the pilU mutant JIR3771 to construct apilT-complemented pilU strain. The resultant derivatives were selected and con-firmed using the methodology described above.
Biological analysis of mutants. Sodium dodecyl sulfate-polyacrylamide gelelectrophoresis and Western immunoblotting analysis were performed withwhole-cell extracts of D. nodosus strains as described previously (17, 23). Twitch-ing motility assays on 1% TAS agar with stab-inoculated cultures were carriedout as described previously (23). Caseinase activity was detected qualitatively onEYE agar plates containing 2% skim milk powder, and elastase activity wasdetected on 0.3% elastin agar (48). Total protease activity was subsequentlydetermined quantitatively using azocasein (Sigma) as the substrate, as previouslydescribed (23). Protease zymogram analyses were carried out as previously de-scribed (24). Transmission electron microscopy was performed using 3-day TASagar cultures as previously described (23). CHO-K1 epithelial cell adherenceassays were carried out in triplicate with at least three independent D. nodosuscultures, using semiconfluent CHO-K1 cells, as previously described (39).CHO-K1 cells, grown in 24-well tissue culture plates, were washed three timeswith 500 l of 37°C prewarmed phosphate-buffered saline (PBS) before additionof 500 l of prewarmed antibiotic-free � minimal essential medium plus 0.5%mannose. D. nodosus cells were incubated for 3 days and harvested from oneEYE or TAS agar plate with 2 ml of prewarmed PBS. To each well, 50 l of aD. nodosus culture (turbidity at 600 nm, 0.4) was added, and the mixture wasincubated for 6 h at 37°C anaerobically. After incubation, the cells were washedfour times with prewarmed PBS and then resuspended in 100 l of 0.1% (wt/vol)digitonin in PBS and incubated for 5 min to lyse the CHO-K1 cells. After serialdilution, the mixture was plated onto EYE agar and incubated for 5 days at 37°Cin the anaerobic chamber. The number of D. nodosus colonies was then com-pared to the viable counts obtained for the original cultures. Adhesion ratioswere calculated by comparison with wild-type levels. To obtain single colonies forstill photography, cells in 3-day D. nodosus cultures were removed from agarplates with 2 ml of PBS, serially diluted, plated onto TAS agar, and incubated for5 days at 37°C anaerobically.
Video microscopy. To examine D. nodosus subsurface colony expansion mi-croscopically, 2.5 ml of TAS agar (containing 0.9% agar [grade A; BDH]) waspoured into 35-mm tissue culture petri dishes (Falcon) so that it formed a thinlayer. D. nodosus strains were stab inoculated into the bottom of the petri dishesand incubated at 37°C anaerobically for 1 day to ensure that the environment in
the petri dishes was anaerobic. The petri dishes then were sealed with Parafilmto maintain the humidity and incubated for 1 day for wild-type strain VCS1703Ato obtain a reasonable-size twitching motility zone (diameter, �1.0 to 1.2 cm)and for 2 days for the pilT and pilU mutants to produce visible growth beneaththe stab inoculum. To maintain anaerobic conditions, the petri dishes, withoutlids, were place into anaerobic incubation bags (AnaerocultP; Merck) and sealedbefore they were removed from the anaerobic chamber.
To examine P. aeruginosa subsurface colony expansion microscopically, 10 mlof LB medium with 1% agar (Oxoid) was poured into standard 90-mm petridishes (BioLab) so that it formed a thin layer. P. aeruginosa strains were stabinoculated into the bottom of the petri dishes and incubated at 37°C overnight.
Bacterial movement at the agar-plastic interface was observed at 37°C with anOlympic IX71 inverted microscope equipped with phase objectives and a heatedstage. Time-lapse images were recorded with either an FViewII or CC12 digitalcamera using AnalySIS Research software (Olympus Soft Imaging Systems). Atleast 10 rate measurements were obtained for each of two time-lapse experi-ments for each strain examined. Time-lapse images were compiled into moviefiles using Quicktime Pro software.
Proteomics. The proteins present in culture supernatants derived from thewild-type and mutant strains were precipitated with trichloroacetic acid, sepa-rated by two-dimensional gel electrophoresis, and analyzed by mass spectrometryas previously described (17).
Sheep pen virulence trials. Standard diagnostic elastase (48) and gelatin gel(37) tests were performed to characterize the strains prior to in vivo virulencetesting. Virulence tests were carried out as previously described (9, 23), exceptthat six groups of eight sheep were challenged blind with the wild-type strainVCS1703A, the pilT mutants JIR3880 and JIR3881, the pilU2 mutant JIR3771,the complemented pilU2/pilU� strain JIR3898, and a negative control that didnot contain any bacteria. All animals were examined and foot rot lesion scoreswere determined at the start of the trial and then at weekly intervals using thestandard method (8, 55). The total weighted foot score, which is the sum of thescores for the four feet after raising scores of 3 and 4 by the power of 2, was usedas an overall disease score for each animal. Blood samples were collected fromthe jugular vein of sheep at the start of the trial and at each time that the feetwere scored. These experiments were carried out in a PC2 containment facilityin accordance with the guidelines of the Australian Government Office of theGene Technology Regulator and the Elizabeth Macarthur Agricultural InstituteAnimal Ethics Committee.
Histopathological examination. The two most severely affected feet from eachsheep that had gross foot rot lesions, two feet from two sheep in the negativecontrol group, and one foot from the sheep with no gross lesions were subjectedto histopathological examination. After the sheep were euthanized, the feet weregently washed in running water to remove surface debris. Each foot was thenremoved at the metacarpal/metatarsal-phalangeal joint with a scalpel and fixedovernight in 10% neutral buffered formalin. The two digits from each fixed footwere separated in the midline of the limb with a scalpel, and a single block oftissue was selected from each digit; this tissue extended from the interdigital skinacross the hard horn of the heel just posterior to the sole in order to encompassthe full extent of the grossly underrun horn in an attempt to include the leadingedge of the lesion. The tissues were processed routinely for histopathologicalexamination and embedded in paraffin. Sections (5 m) were cut and stainedwith hematoxylin and eosin and Gram’s stain. Sections in which bacteria withmorphology suggestive of D. nodosus were present were mounted on Superfrostplus adhesive glass slides and labeled with antibody against D. nodosus.
RESULTS
pilT gene is essential for twitching motility, natural trans-formation, adherence to CHO-K1 cells, and protease secre-tion, whereas pilU is essential only for twitching motility. TheD. nodosus type G strain VCS1703A used in this study wasisolated from an outbreak of virulent ovine foot rot, is naturallytransformable, and produces virulent foot rot in sheep (23).Analysis of the genome sequence of VCS1703A (35) revealedthe presence of two putative genes encoding proteins withsequence identity to PilT (69%) and PilU (38%) from P.aeruginosa. The products of these D. nodosus genes had 31%identity and contained conserved Walker box A and B nucle-otide-binding motifs and the aspartate and histidine boxes thatare conserved in GspE-like ATPases (2). Their C-terminal
VOL. 190, 2008 TWITCHING MOTILITY AND VIRULENCE OF D. NODOSUS 3325

domains contained a GMQTX4LX2LX5I motif, which is con-served in PilT and PilU proteins (38), and the PilT protein alsohad a PilT-specific AIRNLIRE motif (Fig. 1), which is re-quired for PilT function (2). These data suggested that the twogenes from the D. nodosus genome sequence that were iden-tified encoded typical PilT- and PilU-like proteins that werelikely to be part of the type IV fimbrial system.
The D. nodosus pilT and pilU genes were tandemly arranged,with 12 nucleotides between the pilT termination codon andthe pilU initiation codon. RT-PCR carried out using multipleprimer pairs spanning the pilT and pilU region (see Table S1 inthe supplemental material) confirmed that the pilT and pilUgenes comprised an operon (data not shown).
To determine the function of PilT and PilU in D. nodosus,insertionally inactivated double-crossover mutants for eachgene were constructed and confirmed by PCR and Southernhybridization (data not shown). Since disruption of pilT mayhave polarity effects on the downstream pilU gene, both non-polar and polar pilT mutants were constructed.
To see if mutation of pilT had polar effects on pilU expres-sion, RT-PCR was carried out using two different primer pairs(JRP1629/JRP1918 and JRP1917/JRP1918 [see Table S1 inthe supplemental material]). Both products were observed inthe wild-type strain and in both the nonpolar and polar pilTmutants but not in a pilU mutant (data not shown). Therefore,it appears that none of the pilT mutations disrupted the tran-scription of pilU; in the polar mutants pilU must be expressedeither as a result of read-through from the inserted erm(B)gene or from a secondary promoter. In further studies wefocused on two independently derived erythromycin-resistantpilT mutants, JIR3880 (pilT1) and JIR3881 (pilT2).
In a similar manner, transformation of strain VCS1703Awith the pilU suicide vector pJIR2581 led to the production oferythromycin-resistant pilU mutants. PCR analysis and South-ern hybridization confirmed that these mutants resulted from
double-crossover events, and RT-PCR analysis confirmed thatthe two independently derived pilU mutants chosen for furtheranalysis, JIR3770 (pilU1) and JIR3771 (pilU2), did not producepilU-specific transcripts (data not shown).
To determine if the pilT and pilU mutants still produced thefimbrial subunit protein FimA (18, 22), Western immunoblot-ting using antisera raised against serogroup G fimbriae wasperformed using whole-cell lysates of the wild-type strain andthe mutants. The results showed that like the wild-type strain,the pilT and pilU mutants produced the 17-kDa FimA protein(Fig. 2A). Transmission electron microscopy confirmed thattype IV fimbriae were produced by both types of mutants (Fig.2B), indicating that in D. nodosus PilT and PilU are not re-quired for fimbrial biogenesis. These cells did not have in-creased amounts of either surface fimbriae or surface-exposedFimA subunits (data not shown).
Although there was a discernible difference in colony mor-phology between the pilT and pilU mutants, unlike the diffusecolony morphology observed for the wild-type strain, thesemutants had nonspreading colonies with smooth edges (Fig. 3),indicating that they may be defective in twitching motility.Analysis of the mutants using an agar stab twitching motilityassay (23) showed that unlike the wild-type strain, neither thepilT nor pilU mutants exhibited the spreading zones typical oftwitching motility (Fig. 4A). Based on these observations, weconcluded that both the PilT and PilU proteins are essentialfor twitching motility in D. nodosus.
To determine the effect of pilT and pilU mutations on nat-ural transformability, transformation experiments were carriedout with the wild-type strain and the pilT and pilU mutants,using the rrnA-specific D. nodosus suicide vectors pJIR1532and pJIR2691 (Table 1), which carry a tet(M) tetracyclineresistance cassette. Natural transformation of wild-type strainVCS1703A with these plasmids consistently produced lownumbers of tetracycline-resistant colonies (average, ca. 8 � 101
FIG. 1. Amino acid sequence alignment of the conserved regions of PilT and PilU proteins. Identical residues are indicated by a blackbackground, similar residues are indicated by a gray background, and conserved domains are enclosed in boxes. The alignment starts with theWalker A box, spans the Asp box, the Walker B box, the His box, and the AIRNLIRE motif in PilT, and is truncated at the end of theGMQTX4LX2LX5I motif 3 (Pseudomonas putida PilT) to 65 (D. nodosus PilU) amino acid residues from the carboxy termini of the proteins.The sequences of P. aeruginosa PilU (Pa PilU) (accession no. AAG03785), P. stutzeri PilU (Ps PilU) (accession no. CAB56296), N. gonorrhoeaePilU (Ng PilU) (accession no. YP_208935), D. nodosus PilU (Dn PilU) (accession no. CP000513), P. aeruginosa PilT (Pa PilT) (accession no.AAG03784), P. stutzeri PilT (Ps PilT) (accession no. CAB56295), D. nodosus PilT (Dn PilT) (accession no. CP000513), N. gonorrhoeae PilT (NgPilT) (accession no. AAB30824), and P. putida PilT (Pp PilT) (accession no. AAN70658) were aligned by using the CLUSTAL W program at theNPS@ server.
3326 HAN ET AL. J. BACTERIOL.

transformants/g DNA) that resulted from homologous re-combination between the suicide vectors and one of the threechromosomal rrnA operons. However, despite eight attempts,no transformants were obtained with either of the pilT mu-tants. Therefore, we concluded that the PilT protein was es-sential for natural transformation in D. nodosus. By contrast,transformation of the pilU mutants did produce some trans-formants (average, ca. 8 � 10�1 transformant/g DNA).
Natural transformation is the only gene transfer systemavailable for genetic manipulation in D. nodosus; it is not yet
possible to introduce recombinant plasmids by either electro-poration or conjugation. Therefore, since a functional naturaltransformation system is a prerequisite for the complementa-tion of D. nodosus chromosomal mutants (17, 23), it was notpossible to complement the pilT mutants by introducing awild-type copy of the pilT gene into the chromosome. Conse-quently, to rule out the possibility that the phenotypic changesobserved with the pilT mutants were the result of mutations atother sites in the genome, in subsequent studies both of theindependently derived pilT mutants, JIR3880 (pilT1) andJIR3881 (pilT2), were analyzed in all subsequent studies, in-cluding virulence studies with sheep.
An attempt was made to complement a pilU mutation bynatural transformation, even though the transformation fre-quency was at the limit of detection. A 2.6-kb PCR productencompassing the entire pilTU operon, including the potentialpromoter region, was cloned into the EcoRV site of pJIR2691to construct the pilU complementation plasmid pJIR2864. Al-lelic exchange between this plasmid and an rrnA operon wouldintroduce the intact pilTU region into the chromosome andthereby complement the pilU mutation. Transformation of
FIG. 2. Fimbrial analysis of pilT and pilU mutants. (A) Samples (5 l) of whole-cell lysates of wild-type strain VCS1703A (WT), the pilUmutants JIR3770 (pilU1) and JIR3771(pilU2), and the pilT mutants JIR3880 (pilT1) and JIR3881 (pilT2) were separated by 12% sodium dodecylsulfate-polyacrylamide gel electrophoresis, and the production of FimA was determined by Western immunoblotting with fimbrial antisera at a1:1,000 dilution. (B) Transmission electron microscopy of D. nodosus cells. Three-day-old cultures of wild-type strain VCS1703A and the pilU2,pilT1, and pilT2 mutants were removed from TAS agar with PBS, negatively stained, and analyzed by transmission electron microscopy. The profileof the pilU1 mutant appeared to be identical to that of the pilU2 mutant. Bars � 1 m.
FIG. 3. Colony morphology of pilT and pilU mutants. Colonies ofthe wild-type strain VCS1703A (WT), the pilU2 mutant JIR3771(pilU2), and the pilT2 mutant JIR3881 (pilT2) grown on EYE bloodagar plates are shown. Bars � 1 mm.
VOL. 190, 2008 TWITCHING MOTILITY AND VIRULENCE OF D. NODOSUS 3327

FIG. 4. Loss of twitching motility in pilT and pilU mutants. (A) Twitching stab assays were carried out on 1% TAS agar with stab-inoculatedD. nodosus cultures that were incubated for 5 to 7 days under anaerobic conditions or on 1% LB agar with stab-inoculated P. aeruginosa culturesthat were incubated aerobically overnight. Twitching motility was indicated by a large dark zone after staining with Coomassie brilliant blue. Bars �1 cm. (B) Light microscopy of twitching stab zones of D. nodosus and P. aeruginosa strains. D. nodosus cultures were stab inoculated into 1% TASagar and incubated anaerobically at 37°C for 1 to 2 days. P. aeruginosa cultures were stab inoculated into 1% LB agar and incubated aerobicallyovernight. Bars � 100 m. (C) Agar plate morphology of D. nodosus and P. aeruginosa cultures. D. nodosus strains were patched onto TAS agarand incubated anaerobically at 37°C for 3 days. P. aeruginosa strains were patched on LB agar and incubated aerobically at 37°C overnight. Bars �1 cm. The D. nodosus strains used were wild-type strain VCS1703A (WT), the pilU2 mutant JIR3771, the pilU2/pilU� strain JIR3898, the pilT2mutant JIR3881, and a fimA mutant (JIR3727). The results obtained for the D. nodosus pilT1 and pilU1 mutants were not different than the resultsshown here for the pilT2 and pilU2 mutants. The P. aeruginosa strains used were wild-type strain PAK (WT), the pilA mutant PAKpilA, the pilTmutant strain R364, and the pilU mutant S237.
3328 HAN ET AL. J. BACTERIOL.

JIR3770 and JIR3771 with pJIR2864 and selection for eryth-romycin resistance (original pilU mutation) and tetracyclineresistance (complementation cassette) led to the isolation ofseveral independently derived complemented pilU/pilU� deriv-atives after approximately 10 separate experiments. PCR anal-ysis confirmed the predicted genotype of four of these deriva-tives. Colony morphology differences between the pilU mutantsand the pilU/pilU� strains were readily discernible, and thecomplemented strains had diffuse, spreading wild-type colonies(data not shown). Further studies demonstrated that twitchingmotility was restored in the complemented mutants (Fig. 4A).One of the pilU/pilU� strains, JIR3898, which was a derivativeof JIR3771, was chosen for further analysis. To exclude thepossibility that the duplicated pilT gene in JIR3898 may com-plement the pilU mutation, a pilT-complemented pilU strainJIR3943, which contained two copies of pilT, was constructedand characterized. The results showed that JIR3943 had thesame characteristics as JIR3771 (data not shown). Specifically,the twitching motility defect which was observed in the pilUmutant JIR3771 and which was complemented by the wild-typepilTU operon was not complemented by just the wild-type pilTgene.
D. nodosus type IV fimbriae are required for adherence toepithelial CHO-K1 cells (39). In N. meningitidis (42) and N.gonorrhoeae (29), pilT mutants adhere to epithelial cells,whereas N. gonorrhoeae pilU mutants show increased adher-ence (38). By contrast, P. aeruginosa pilT or pilU mutants showreduced adherence to some epithelial cell types (7) but not toother types (60). The results of D. nodosus adherence assaysshowed that both pilT mutants, JIR3880 (pilT1) and JIR3881(pilT2), had a reduced ability to adhere to the CHO-K1 cells invitro (P 0.05, Student’s t test) compared to the wild-typestrain (Fig. 5). The adhesion assays revealed phenotypic dif-ferences between the pilT and pilU mutants; the pilU mutantand the pilU/pilU� strain adhered to CHO-K1 cells at thewild-type level (Fig. 5). Based on these results, we concludedthat notwithstanding the fact that mutation of the pilT geneand mutation of the pilU gene had similar effects on fimbria-mediated twitching motility, unlike PilT, PilU did not play asignificant role in fimbria-mediated epithelial cell adhesion.
Previous studies showed that mutation of the fimA gene(23), the pilR and rpoN genes (39), and the fimN, fimO, fimP,and pilE genes (17) resulted in a reduced ability to secreteextracellular serine proteases. To examine the effect of muta-tion of pilT and pilU on protease secretion, an agar plateelastase test, which has been routinely used as a diagnostic footrot protease test, and caseinase assays were performed initially.The results showed that the wild-type strain and the pilU mu-tants were elastase positive after 7 days of incubation, whereasboth pilT mutants were elastase negative even after 28 days ofincubation (Fig. 6A). Furthermore, the pilT mutants, but not
FIG. 5. Adherence of the pilT and pilU mutants to CHO-K1 epi-thelial cells. The wild-type strain VCS1703A (WT), the pilU2 mutantJIR3771, the pilU2/pilU� strain JIR3898, and the pilT mutants JIR3880(pilT1) and JIR3881 (pilT2) were incubated with CHO-K1 monolayercells for 6 h anaerobically. The monolayers were then washed, andcell-associated bacteria were quantified. Relative adherence is ex-pressed as the ratio of the mutant value to the wild-type value. Meansand standard deviations from at least three independent experimentsare shown.
FIG. 6. Extracellular protease activity of pilT and pilU mutants. Thestrains used were the wild-type strain VCS1703A (WT), the pilU2 mutantJIR3771, the pilU2/pilU� strain JIR3898, and the pilT mutants JIR3880(pilT1) and JIR3881 (pilT2). (A) Qualitative elastase assay on TAS agarcontaining 0.3% (wt/vol) elastin incubated for 28 days. The particulatematter in the agar was the insoluble elastin substrate. Elastase activity wasindicated by a zone of clearing around the colony streak. The zone for thepilT1 mutant JIR3880 was identical to that for JIR3881. (B) Quantitativeprotease activity assay. The protease activities of culture supernatants ofbacteria were determined in triplicate using azocasein as the substrate.The relative protease activity is expressed as the ratio of the mutant valueto the wild-type strain value. Means and standard deviations from at leastthree independent experiments are shown.
VOL. 190, 2008 TWITCHING MOTILITY AND VIRULENCE OF D. NODOSUS 3329

the pilU mutants, exhibited greatly reduced caseinase activityon 1% skim milk agar plates compared to the wild-type strain(data not shown). These results suggested that the pilT mutantsdid not produce wild-type levels of extracellular protease.Quantitative protease assays were subsequently carried outwith culture supernatants, using azocasein as the substrate(23). The results were consistent with the initial observations,with the pilT mutants having significantly lower levels of extra-cellular protease activity (P 0.05, Student’s t test) than thewild-type strain (Fig. 6B) and the pilU mutants having wild-type activity levels. Once more a significant phenotypic differ-ence between the pilT and pilU mutants was demonstrated, andwe concluded that unlike PilT, PilU does not play a significantrole in fimbria-mediated protease secretion.
To determine if the reduction in extracellular protease ac-tivity in the pilT mutants was due to a defect in secretion or todown-regulation of protease gene expression, quantitativereal-time RT-PCR experiments were performed with mRNAmolecules derived from each of three genes encoding theclosely related extracellular proteases AprV2, AprV5, andBprV (17). The results showed that for each gene there was nosignificant difference between the wild-type strain and the pilTmutants (data not shown), suggesting that the pilT mutationshad no effect on protease gene expression and that PilT wasrequired for extracellular protease secretion. Protease zymo-gram analysis was also carried out with culture supernatantsfrom these isogenic strains. The results showed that each of thepilT mutants had greatly reduced AprV2 and AprV5 activities(data not shown). The basic protease BrpV cannot be detectedby this method.
Comparative proteomic analysis coupled with mass spec-trometry was used to identify proteins affected by the pilT andpilU mutations. Analysis of the extracellular proteome re-vealed that the intensities of only six protein spots (Fig. 7 anddata not shown), all of which were variants of AprV2, AprV5,and BprV, were lower in the pilT mutants than in the wild-typestrain, resulting in a profile similar to that of the previouslydescribed fimP mutants (17).
Finally, we observed that after extended passage the cellsurface fimbriae of the pilT mutants were lost and that thesederivatives did not produce FimA. We analyzed two nonfim-briated derivatives of pilT mutants and showed that they hadmutations in either pilR or pilS, which are regulatory genesrequired for FimA expression (39). Note that in all of thephenotypic studies of the pilT mutants reported here, the pilTcultures were confirmed to produce fimbriae and therefore not
to have undergone secondary mutations that affect FimA pro-duction.
Microscopic analysis of twitching motility in pilT and pilUmutants of D. nodosus and P. aeruginosa. Microscopic analysisshowed that in stab-inoculated cultures of the wild-type strainmany individual cells were present at the edge of the twitchingmotility zone, whereas the pilT, pilU, and fimA mutants had noobvious twitching motility zone. With the pilT and fimA mu-tants there were very few or no individual cells past the edge ofthe colony, whereas the pilU mutants produced thick, densesubsurface colonies in which individual cells extruded past thecolony edge more frequently (Fig. 4B). To rule out the possi-bility that these individual motile cells were evidence of sec-ondary mutations, they were subcultured, and the resultantcultures were shown to be identical to the original pilU cultures(data not shown).
Based on these results, the results of the protease secretionexperiments, and previous evidence showing that the type IVfimbrial system of D. nodosus represents the only type II se-cretion system in this bacterium (17, 35), it was postulated thatin the pilU mutants there must be sufficient PilT-mediatedfimbrial retraction to allow subsequent protease secretion withthe next round of fimbrial polymerization but not enough re-traction to observe twitching motility-mediated colony expan-sion. Time-lapse video microscopy showed that within 2 h theleading edge of the wild-type VCS1703A colony, which wascomposed of individual cells, moved away from the originalinoculum at an average rate of 2.6 � 0.2 m per min (seeVideo S1 in the supplemental material). However, no obviouscellular movement was observed for the pilT mutants (seeVideo S2 in the supplemental material) or pilU mutants (seeVideo S3 in the supplemental material) over 4 h. However, oncloser examination, there did appear to be some slight move-ment of individual cells at the periphery of a pilU mutant stabcolony (see Video S3 in the supplemental material). Due to theconstraints involved in maintaining anaerobic conditions underthe microscope, it was not possible to analyze the movementfor a longer time period.
The surface colony morphologies of wild-type D. nodosusand P. aeruginosa strains and mutants with mutations in thepilin subunit genes (fimA or pilA), pilT, and pilU were strikinglysimilar, including the previously reported (6) matte appear-ance of pilU mutants when they were grown on the surface ofnutrient agar compared with the glossy appearance of wild-type, pilin subunit, or pilT mutants (Fig. 4C). Twitching motil-ity stab assays revealed similar gross colony expansion pheno-types at the agar-plastic interface for the D. nodosus and P.aeruginosa wild-type strains and pilT, pilU, or pilin subunitmutants (Fig. 4A). Microscopic analysis of subsurface colonystabs showed that both wild-type strains produced groups ofcells at the leading edge of the expanding colony, followed bya loose network of cells, whereas the nontwitching pilin subunit(fimA or pilA) and pilT mutants produced colonies withsmooth edges (Fig. 4B). In addition, the pilU mutants of bothD. nodosus and P. aeruginosa produced thick subsurface colo-nies with a distinctive fringe and evidence of some migration ofcells beyond the leading edge (Fig. 4B).
Since P. aeruginosa is significantly more motile, we reasonedthat we would be more likely to detect type IV fimbria-medi-ated cellular movements with a P. aeruginosa pilU mutant than
FIG. 7. Reduced secretion of the D. nodosus proteases. Trichloro-acetic acid-precipitated culture supernatants (35 g protein per sam-ple) were analyzed by two-dimensional gel electrophoresis. Sections ofrepresentative gels for the wild-type strain VCS1703A (WT) and thepilT2 mutant JIR3881 are shown. Reduced protein spots for AprV5,AprV2, and BprV are circled.
3330 HAN ET AL. J. BACTERIOL.

with a D. nodosus pilU mutant. We found that in P. aeruginosawild-type strain PAK twitching motility-mediated colony ex-pansion was extremely rapid and occurred at a rate of 10.9 �1.6 m per min (see Video S4 in the supplemental material),which was approximately four times faster than wild-type D.nodosus colony expansion. A P. aeruginosa pilT mutant (R464)exhibited colony expansion through cell division identical tothat observed with a pilA mutant (PAK pilA::TcR) (see VideoS5 in the supplemental material; data not shown) at a rate ofonly 0.11 � 0.03 m per min, which was approximately 100-fold slower than the wild-type strain colony expansion. P.aeruginosa pilU mutant S237 had an overall colony expansionrate that was slightly higher than that of the the pilT or pilAmutants (0.17 � 0.08 m per min). Furthermore, unlike thelatter mutants, this pilU mutant showed numerous cellulartranslocation events, particularly in the region immediatelybehind the outermost edge of the expanding colony (see VideoS6 in the supplemental material). It appears that the P. aerugi-nosa pilU mutant had cellular translocation ability but that thisactivity was uncoordinated and did not result in significantoverall expansion of the colony but rather resulted in piling upof the cells, producing a thick colony edge. These observationsprovide preliminary evidence that limited cellular transloca-tion occurs in pilU mutants of P. aeruginosa, presumablythrough retraction of type IV fimbriae. It has been reportedpreviously that pilU mutants of P. aeruginosa are hyperpiliatedbut are sensitive to type IV fimbria-specific bacteriophages,which is also consistent with the presence of retractile type IVfimbriae in these mutants (54).
Both PilT and PilU are essential for virulence in sheep.Virulence testing with sheep was carried out with the wild-typestrain VCS1703A, the pilT mutants JIR3880 and JIR3881, thepilU2 mutant JIR3771, and the complemented derivativeJIR3898, using a standardized method as previously described(23). Under high-security biocontainment conditions groups ofeight sheep were challenged by applying agar cultures of D.nodosus to the interdigital skin of all four feet. Over a periodof 30 days, each foot was quantitatively scored for foot rotlesions at weekly intervals, and the values were converted to atotal weighted foot score, which represents an objective andquantitative analysis of the severity of the foot rot lesions in asheep (8, 55).
The results (Table 2) showed that only sheep challengedwith the wild-type strain and the complemented pilU2/pilU�
strain JIR3898 contracted virulent foot rot. There was no signof foot rot in the groups challenged with the pilT or pilU
mutants at day 30 postinfection or in the negative controlgroup, which was not challenged with D. nodosus. For ethicalreasons, after 30 days the animals were treated with antibioticsto eliminate the infection and the trial was terminated. Prior tochallenge, foot swabs from the sheep were cultured, and theresults showed that they were all free of D. nodosus. Culturingof foot swabs taken 4 weeks after challenge resulted in D.nodosus isolates only for the sheep challenged with the wild-type and the pilU complemented strains. All 71 isolates ob-tained from infected sheep were identical to the wild-typestrain in diagnostic serogrouping, elastase, gelatin gel, andomp1 PCR-restriction fragment length polymorphism analysistests.
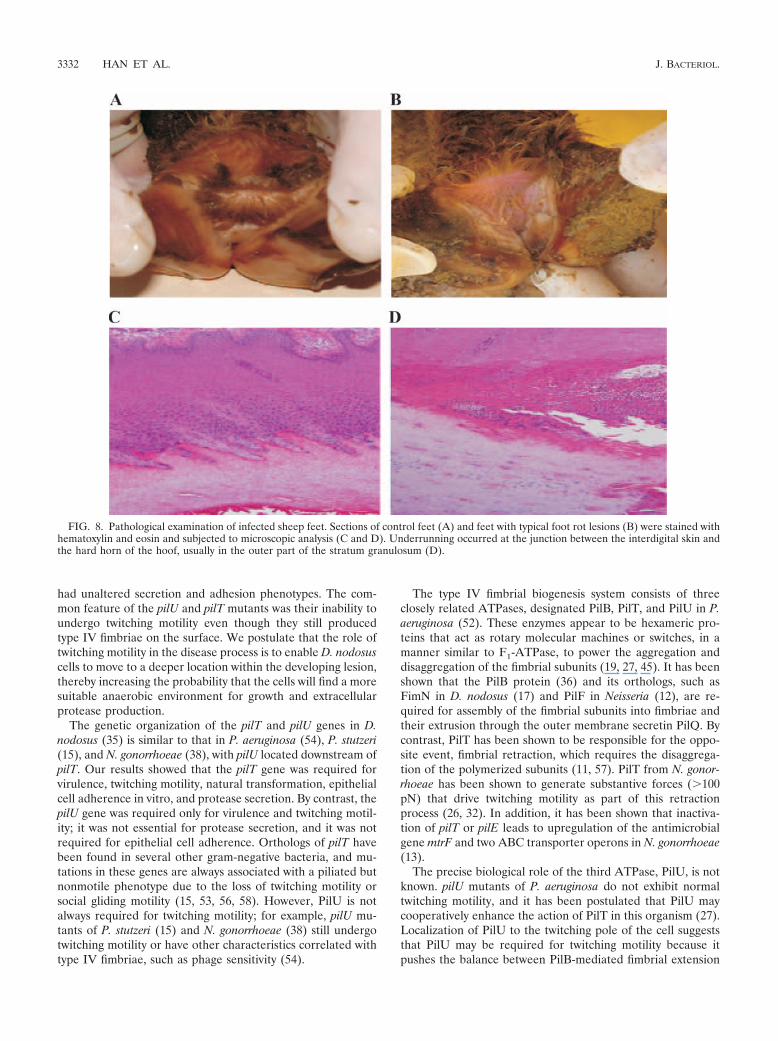
For each sheep that had gross foot rot lesions (Fig. 8B), thetwo most severely affected feet were subjected to histopatho-logical examination. Two feet from two sheep from the nega-tive control group (Fig. 8A) and one foot from each of theother groups (all with no gross lesions) were also examined.Microscopic analysis of hematoxylin- and eosin-stained sec-tions revealed that in the feet with clinical lesions underrun-ning commenced at the junction between the interdigital skinand the hard horn of the hoof, usually in the outer part of thestratum granulosum (Fig. 8D). Sometimes multiple fronts ofunderrunning were present, and the most recent front alwayscommenced at the interdigital area. In affected feet, the kera-tohyalin granules of the stratum granulosum of underrun areaswere smaller and less obvious. At the end of the lesion therewere few bacteria, but sometimes D. nodosus-like organisms(large gram-negative rods, often slightly curved with prominentpoles) were present on the surface or within the adjacent palelystained epithelial cells, singly or in small clumps.
Although surface maceration was a consistent finding for allof the feet that were examined, no histopathological evidenceof disease was apparent in sections from the control animals(Fig. 8C) or the animals infected with the pilT or pilU mutants.Based on both the gross pathological and the detailed his-topathological analyses and on the fact that the effect of thepilU mutation on virulence was reversed by complementationwith the wild-type pilU gene, we concluded that mutation ofeither the pilT or pilU genes eliminated the ability of D. nodo-sus to cause ovine foot rot.
DISCUSSION
D. nodosus is the essential causative pathogen of ovine footrot, and it was previously shown that the type IV fimbrialsubunit protein is essential for virulence, although it was notclear if the effects on virulence resulted from reduced fimbria-mediated cell adherence, loss of twitching motility, or reducedprotease secretion (23). The results presented in this paperprovide evidence that type IV fimbria-mediated twitching mo-tility is essential for D. nodosus cells to cause ovine foot rot.Virulence studies carried out with the native ovine host showedthat two independently derived pilT mutants and a pilU mutantall failed to cause disease, unlike the wild-type strain. Further-more, complementation of the pilU mutant restored the abilityto cause virulent disease. The pilT mutants had reduced abil-ities to secrete extracellular proteases and to adhere to epithe-lial cells, but these parameters could not have been responsiblefor the loss of virulence in these mutants since the pilU mutants
TABLE 2. Virulence of D. nodosus mutants in sheep
Strain Genotype
No. ofinfected
sheep/totalno. onday 30
No. ofinfectedfeet/total
no. onday 30
Totalweightedfoot scoreon day 21
Totalweightedfoot scoreon day 30
None 0/8 0/32 0 0VCS1703A Wild type 6/8 12/32 0.38 23.25JIR3880 pilT1 0/8 0/32 0 0JIR3881 pilT2 0/8 0/32 0 0JIR3771 pilU2 0/8 0/32 0 0JIR3898 pilU2/pilU� 5/8 8/32 0.13 8.63
VOL. 190, 2008 TWITCHING MOTILITY AND VIRULENCE OF D. NODOSUS 3331
had unaltered secretion and adhesion phenotypes. The com-mon feature of the pilU and pilT mutants was their inability toundergo twitching motility even though they still producedtype IV fimbriae on the surface. We postulate that the role oftwitching motility in the disease process is to enable D. nodosuscells to move to a deeper location within the developing lesion,thereby increasing the probability that the cells will find a moresuitable anaerobic environment for growth and extracellularprotease production.
The genetic organization of the pilT and pilU genes in D.nodosus (35) is similar to that in P. aeruginosa (54), P. stutzeri(15), and N. gonorrhoeae (38), with pilU located downstream ofpilT. Our results showed that the pilT gene was required forvirulence, twitching motility, natural transformation, epithelialcell adherence in vitro, and protease secretion. By contrast, thepilU gene was required only for virulence and twitching motil-ity; it was not essential for protease secretion, and it was notrequired for epithelial cell adherence. Orthologs of pilT havebeen found in several other gram-negative bacteria, and mu-tations in these genes are always associated with a piliated butnonmotile phenotype due to the loss of twitching motility orsocial gliding motility (15, 53, 56, 58). However, PilU is notalways required for twitching motility; for example, pilU mu-tants of P. stutzeri (15) and N. gonorrhoeae (38) still undergotwitching motility or have other characteristics correlated withtype IV fimbriae, such as phage sensitivity (54).
The type IV fimbrial biogenesis system consists of threeclosely related ATPases, designated PilB, PilT, and PilU in P.aeruginosa (52). These enzymes appear to be hexameric pro-teins that act as rotary molecular machines or switches, in amanner similar to F1-ATPase, to power the aggregation anddisaggregation of the fimbrial subunits (19, 27, 45). It has beenshown that the PilB protein (36) and its orthologs, such asFimN in D. nodosus (17) and PilF in Neisseria (12), are re-quired for assembly of the fimbrial subunits into fimbriae andtheir extrusion through the outer membrane secretin PilQ. Bycontrast, PilT has been shown to be responsible for the oppo-site event, fimbrial retraction, which requires the disaggrega-tion of the polymerized subunits (11, 57). PilT from N. gonor-rhoeae has been shown to generate substantive forces (�100pN) that drive twitching motility as part of this retractionprocess (26, 32). In addition, it has been shown that inactiva-tion of pilT or pilE leads to upregulation of the antimicrobialgene mtrF and two ABC transporter operons in N. gonorrhoeae(13).
The precise biological role of the third ATPase, PilU, is notknown. pilU mutants of P. aeruginosa do not exhibit normaltwitching motility, and it has been postulated that PilU maycooperatively enhance the action of PilT in this organism (27).Localization of PilU to the twitching pole of the cell suggeststhat PilU may be required for twitching motility because itpushes the balance between PilB-mediated fimbrial extension
FIG. 8. Pathological examination of infected sheep feet. Sections of control feet (A) and feet with typical foot rot lesions (B) were stained withhematoxylin and eosin and subjected to microscopic analysis (C and D). Underrunning occurred at the junction between the interdigital skin andthe hard horn of the hoof, usually in the outer part of the stratum granulosum (D).
3332 HAN ET AL. J. BACTERIOL.

and PilT-mediated retraction toward retraction (5, 6). By con-trast, pilU mutants of N. gonorrhoeae exhibit twitching motility,and it appears that PilU is an antagonist of PilT in this species(38), acting downstream of PilT at the mechanistic level andperhaps competing for a common binding site or complex. Theresults obtained in this study are more consistent with thehypothesis that in D. nodosus PilU acts in concert with PilT,since we showed that although PilU is also essential for twitch-ing motility, mutation of pilU had fewer phenotypic effects thanthe corresponding pilT mutation.
Comparisons of type IV fimbrial biogenesis and type II se-cretion systems have revealed significant similarities (10, 18,40, 41, 46). The current model for type II secretion involves theformation of a pseudopilus, a pilus-like secreton that is formedby pseudopilins with similarity to fimbrial subunits, and theexpulsion of folded proteins through an outer membrane se-cretin channel by cycles of extension and retraction (5, 10, 20,40). Interestingly, no PilT or PilU orthologs have been identi-fied in type II secretion systems, although orthologs of PilB(GspE family), which is postulated to mediate fimbrial elon-gation and therefore to be a functional antagonist of PilT, arepresent (5, 10, 20, 40). It has been postulated (17) that in D.nodosus extracellular proteases primarily are exported fromthe cell by the type IV fimbrial system, which also acts as thesole type II secretion system in this bacterium. This hypothesisis based on the observation that protease secretion requires thetype IV fimbrial subunit protein FimA (23) and the type IVfimbrial biogenesis components FimN, FimO, FimP, and PilE(17). In addition, analysis of the D. nodosus genome sequencedid not reveal any other type II secretion system (17, 35). Inthis paper we provide evidence that PilT is involved in bothtype IV fimbria-mediated twitching motility and protease se-cretion and that pilT mutants have greatly decreased extracel-lular protease activity compared to the wild-type strain, a re-duction that was not due to altered transcription of the threeextracellular protease genes, aprV2, aprV5, and bprV.
To explain the role of PilT in protease secretion in D. no-dosus, we propose that the extracellular protease precursorsbind to the inner surface of the putative PilQ outer membranechannel complex and are secreted when they are pushedthrough PilQ and out of the cell by the growing pseudopilus orpilus (17). It appears that in D. nodosus both PilQ and thefimbrial biogenesis apparatus are required for both fimbrialextrusion and protein secretion. Therefore, pilus or pseudopi-lus retraction mediated by PilT would be important to providenewly synthesized protease precursors continued access toPilQ. We suggest that since retraction does not occur in a pilTmutant, the PilQ complex is blocked by the protruded fimbriae,preventing further protease secretion. This hypothesis wouldexplain the essential role in protease secretion of PilT, which isnot needed for classical type II secretion (5, 10, 20, 40). Thisprocess would also explain our inability to transform the pilTmutants since DNA uptake presumably also requires the PilQouter membrane channel and pilus retraction, as previouslysuggested for P. stutzeri (15). Note that since recent studieshave suggested that PilT-mediated fimbrial retraction may beinvolved in the regulation of glutamine transport genes in N.gonorrhoeae (13), we cannot completely rule out the possibilitythat there is an alternative explanation for the dependence of
protease secretion on the presence of functional type IV fim-briae that undergo PilT-dependent fimbrial retraction.
In contrast to the active role of PilT in protease secretion,PilU was not essential for this process, except for a possibleeffect on BprV secretion. This result is in agreement with thehypothesis that the role of PilU in D. nodosus is simply tofacilitate PilT-mediated retraction. Our model for proteasesecretion inherently implies that although twitching motilitywas not observed in the pilU mutants, some limited amount ofretraction must take place, sufficient for the pili or pseudopilito clear the PilQ complex and to enable a subsequent round ofprotease secretion. The differences in colonial morphology be-tween the pilU and pilT mutants, the observation that moreindividual cells emerged from occasional flares at the growthedge of the pilU mutants, and the observation that these iso-lated cells did not have wild-type-like movement supported thishypothesis. In addition, the evidence obtained from our trans-formation studies, in which a very low level of transformationwas observed in the pilU mutants but not in the pilT mutants,is also in agreement with this hypothesis.
Attempts to use video microscopy to demonstrate cellulartranslocation in a pilU mutant of D. nodosus were unsuccessful,presumably because of technical difficulties inherent in main-taining anaerobic conditions under the microscope for morethan 4 h. However, for the first time, limited motion wasdetected in a pilU mutant of P. aeruginosa, and the movementwas clearly distinct from the movement due to cellular divisionobserved in pilA or pilT mutants of this aerobic bacterium. Thisresult is in agreement with the hypothesis that there is suffi-cient PilT-mediated retraction in a D. nodosus pilU mutant toallow protease secretion and limited DNA uptake through thetype IV fimbrial apparatus.
In N. gonorrhoeae (56) and P. stutzeri (15), disruption of pilTeliminates the ability of these organisms to undergo naturaltransformation, providing evidence that there is a relationshipbetween this process and fimbrial retraction. pilU mutants of P.stutzeri are still transformable, but at a greatly reduced level, asshown here for D. nodosus. Likewise, pilU mutants of P. aerugi-nosa are bacteriophage susceptible, implying that phage DNAcan enter the cell in these mutants (54). Finally, recent studieswith N. meningitidis have shown that DNA binds to the PilQsecretin (1). These observations all support the hypothesis thatin D. nodosus natural transformation requires DNA uptakethat is mediated by PilT-mediated retraction of the type IVfimbriae, a process that is facilitated by PilU and involvestransport through the PilQ secretin. Again, it is necessary topostulate that in a pilU mutant sufficient fimbrial retractionmust occur to allow at least some DNA to enter the cell. Thissuggestion is supported by the protease secretion data and isconsistent with previous suggestions regarding the role of PilUin other organisms (5, 15, 54).
In other bacteria type IV fimbriae mediate the initial adher-ence to host cells (16, 34), which has been reported to beindependent of PilT (29, 42, 60). Moreover, one recent studydemonstrated that PilT negatively regulates the adhesion of N.meningitidis (59). However, in contrast to these results, ourstudy showed that the D. nodosus pilT mutants had reducedadherence to CHO-K1 cells. Similar results were reported forP. aeruginosa, where pilT and pilU mutants showed decreasedadherence to three different epithelial cell lines (7). By con-
VOL. 190, 2008 TWITCHING MOTILITY AND VIRULENCE OF D. NODOSUS 3333

trast, mutation of the pilU gene of D. nodosus had no signifi-cant effect on adherence. These results may be explained byonce more postulating that in such mutants there is enoughfimbrial retraction to allow the extrusion of putative adhesinseither with the fimbriae or onto the cell surface.
In conclusion, we demonstrated that in D. nodosus, type IVfimbria-mediated twitching motility, not adhesion or proteasesecretion, is the key reason why the type IV fimbriae areessential for virulence in sheep, the major host of this bacte-rium. We differentiated the biological effects of the PilT andPilU proteins, two well-conserved ATPases that are both re-quired for twitching motility, and obtained evidence that PilTis also involved in protease secretion and natural transforma-tion in D. nodosus, probably by mediating fimbrial depolymer-ization. These results support the hypothesis that in D. nodo-sus, a relatively slowly growing anaerobic bacterium with asmall genome (ca. 1.4 Mb), reductive evolution has led to asituation where the type IV fimbrial biogenesis system is alsoresponsible for extracellular protein secretion and naturaltransformation.
ACKNOWLEDGMENTS
We sincerely thank Craig Kristo of the Faculty of Veterinary Sci-ence, University of Sydney, and the technical staff of the ElizabethMacarthur Agricultural Institute for invaluable assistance with thesheep virulence trials, Khim Hoe for assistance with electron micros-copy, Ian Smith, David Steer, and Shane Reeve of the Department ofBiochemistry and Molecular Biology, Monash University, for help withproteomics and mass spectrometry analysis, and Steven Morton ofSchool of Physics, Faculty of Science, Monash University, for photo-graphic assistance.
This research was supported by the Australian Research Councilthrough funding of the Australian Research Council Centre of Excel-lence in Structural and Functional Microbial Genomics. Xiaoyan Hanwas supported by a postgraduate scholarship provided by the Centre ofExcellence. Lynne Turnbull and Cynthia Whitchurch were supportedby the National Health and Medical Research Council of Australia.We thank the Rebecca L. Cooper Foundation for a financial contri-bution toward the purchase of the microscope.
REFERENCES
1. Assalkhou, R., S. Balasingham, R. F. Collins, S. A. Frye, T. Davidsen, A. V.Benam, M. Bjoras, J. P. Derrick, and T. Tonjum. 2007. The outer membranesecretin PilQ from Neisseria meningitidis binds DNA. Microbiology 153:1593–1603.
2. Aukema, K., E. M. Kron, T. J. Herdendorf, and K. T. Forest. 2005. Func-tional dissection of a conserved motif within the pilus retraction protein PilT.J. Bacteriol. 187:611–818.
3. Billington, S. J., J. L. Johnston, and J. I. Rood. 1996. Virulence regions andvirulence factors of the ovine footrot pathogen, Dichelobacter nodosus.FEMS Microbiol. Lett. 145:147–156.
4. Bohm, M., T. Hurek, and B. Reinhold-Hurek. 2007. Twitching motility isessential for endophytic rice colonization by the N2-fixing endophyte Azoar-cus sp. strain BH72. Mol. Plant-Microbe Interact. 20:526–533.
5. Burrows, L. I. 2005. Weapons of mass retraction. Mol. Microbiol. 57:878–888.
6. Chiang, P., M. Habash, and L. I. Burrows. 2005. Disparate subcellularlocalization patterns of Pseudomonas aeruginosa type IV pilus ATPases in-volved in twitching motility. J. Bacteriol. 187:829–839.
7. Comolli, J. C., A. R. Hauser, L. Waite, C. B. Whitchurch, J. S. Mattick, andJ. N. Engel. 1999. Pseudomonas aeruginosa gene products PilT and PilU arerequired for cytotoxicity in vitro and virulence in a mouse model of acutepneumonia. Infect. Immun. 67:3625–3630.
8. Egerton, J. R., and D. S. Roberts. 1971. Vaccination against ovine foot-rot.J. Comp. Pathol. 81:179–185.
9. Egerton, J. R., D. S. Roberts, and I. M. Parsonson. 1969. The aetiology andpathogenesis of ovine footrot. I. A histological study of the bacterial inva-sion. J. Comp. Pathol. 79:207–216.
10. Filloux, A. 2004. The underlying mechanisms of type II protein secretion.Biochim. Biophys. Acta 1694:163–179.
11. Forest, K. T., K. A. Satyshur, G. A. Worzalla, J. K. Hansen, and T. J.
Herdendorf. 2004. The pilus-retraction protein PilT: ultrastructure of thebiological assembly. Acta Crystallogr. Sect. D 60:978–982.
12. Freitag, N. E., H. S. Seifert, and M. Koomey. 1995. Characterization of thepilF-pilD pilus-assembly locus of Neisseria gonorrhoeae. Mol. Microbiol. 16:575–586.
13. Friedrich, A., C. G. Arvidson, W. M. Shafer, E. H. Lee, and M. So. 2007. TwoABC transporter operons and the antimicrobial resistance gene mtrF are pilTresponsive in Neisseria gonorrhoeae. J. Bacteriol. 189:5399–5402.
14. Ghimire, S. C., and J. R. Egerton. 1999. PCR-RFLP of outer membraneproteins gene of Dichelobacter nodosus: a new tool in the epidemiology offootrot. Epidemiol. Infect. 122:521–528.
15. Graupner, S., N. Weger, M. Sohni, and W. Wackernagel. 2001. Requirementof novel competence genes pilT and pilU of Pseudomonas stutzeri for naturaltransformation and suppression of pilT deficiency by a hexahistidine tag onthe type IV pilus protein PilAI. J. Bacteriol. 183:4694–4701.
16. Hahn, H. P. 1997. The type-4 pilus is the major virulence-associated adhesinof Pseudomonas aeruginosa—a review. Gene 192:99–108.
17. Han, X., R. M. Kennan, D. Parker, J. K. Davies, and J. I. Rood. 2007. TypeIV fimbrial biogenesis is required for protease secretion and natural trans-formation in Dichelobacter nodosus. J. Bacteriol. 189:5022–5033.
18. Hobbs, M., and J. S. Mattick. 1993. Common components in the assembly oftype 4 fimbriae, DNA transfer systems, filamentous phage and protein-secretion apparatus: a general system for the formation of surface-associatedprotein complexes. Mol. Microbiol. 10:233–243.
19. Hopper, S., B. Vasquez, A. Merz, S. Clary, J. S. Wilbur, and M. So. 2000.Effects of the immunoglobulin A1 protease on Neisseria gonorrhoeae traf-ficking across polarized T84 epithelial monolayers. Infect. Immun. 68:906–911.
20. Johnson, T. L., J. Abendroth, W. G. Hol, and M. Sandkvist. 2006. Type IIsecretion: from structure to function. FEMS Microbiol. Lett. 255:175–186.
21. Kaiser, D. 2000. Bacterial motility: How do pili pull? Curr. Biol. 10:R777–780.
22. Kennan, R. M., S. J. Billington, and J. I. Rood. 1998. Electroporation-mediated transformation of the ovine footrot pathogen Dichelobacter nodo-sus. FEMS Microbiol. Lett. 169:383–389.
23. Kennan, R. M., O. P. Dhungyel, R. J. Whittington, J. R. Egerton, and J. I.Rood. 2001. The type IV fimbrial subunit gene (fimA) of Dichelobacternodosus is essential for virulence, protease secretion, and natural compe-tence. J. Bacteriol. 183:4451–4458.
24. Liu, D., and W. K. Yong. 1993. Use of elastase test, gelatin gel test andelectrophoretic zymogram to determine virulence of Dichelobacter nodosusisolated from ovine foot rot. Res. Vet. Sci. 55:124–129.
25. Maier, B. 2005. Using laser tweezers to measure twitching motility in Neis-seria. Curr. Opin. Microbiol. 8:344–349.
26. Maier, B., L. Potter, M. So, H. S. Seifert, and M. P. Sheetz. 2002. Single pilusmotor forces exceed 100 pN. Proc. Natl. Acad. Sci. USA 99:16012–16017.
27. Mattick, J. S. 2002. Type IV pili and twitching motility. Annu. Rev. Micro-biol. 56:289–314.
28. Menard, R., P. J. Sansonetti, and C. Parsot. 1993. Nonpolar mutagenesis ofthe ipa genes defines IpaB, IpaC, and IpaD as effectors of Shigella flexnerientry into epithelial cells. J. Bacteriol. 175:5899–5906.
29. Merz, A. J., C. A. Enns, and M. So. 1999. Type IV pili of pathogenicNeisseriae elicit cortical plaque formation in epithelial cells. Mol. Microbiol.32:1316–1332.
30. Merz, A. J., and K. T. Forest. 2002. Bacterial surface motility: slime trailsgrappling hooks and nozzles. Curr. Biol. 12:R297–303.
31. Merz, A. J., and M. So. 2000. Interactions of pathogenic Neisseria withepithelial cell membranes. Annu. Rev. Cell Dev. Biol. 16:423–457.
32. Merz, A. J., M. So, and M. P. Sheetz. 2000. Pilus retraction powers bacterialtwitching motility. Nature 407:98–102.
33. Morand, P. C., E. Bille, S. Morelle, E. Eugene, J. L. Beretti, M. Wolfgang,T. F. Meyer, M. Koomey, and X. Nassif. 2004. Type IV pilus retraction inpathogenic Neisseria is regulated by the PilC proteins. EMBO J. 23:2009–2017.
34. Morand, P. C., P. Tattevin, E. Eugene, J. L. Beretti, and X. Nassif. 2001. Theadhesive property of the type IV pilus-associated component PilC1 of patho-genic Neisseria is supported by the conformational structure of the N-termi-nal part of the molecule. Mol. Microbiol. 40:846–856.
35. Myers, G. S., D. Parker, K. Al-Hasani, R. M. Kennan, T. Seemann, Q. Ren,J. H. Badger, J. D. Selengut, R. T. Deboy, H. Tettelin, J. D. Boyce, V. P.McCarl, X. Han, W. C. Nelson, R. Madupu, Y. Mohamoud, T. Holley, N.Fedorova, H. Khouri, S. P. Bottomley, R. J. Whittington, B. Adler, J. G.Songer, J. I. Rood, and I. T. Paulsen. 2007. Genome sequence and identi-fication of candidate vaccine antigens from the animal pathogen Dich-elobacter nodosus. Nat. Biotechnol. 25:569–575.
36. Nunn, D., S. Bergman, and S. Lory. 1990. Products of three accessory genes,pilB, pilC, and pilD, are required for biogenesis of Pseudomonas aeruginosapili. J. Bacteriol. 172:2911–2919.
37. Palmer, M. A. 1993. A gelatin test to detect activity and stability of proteasesproduced by Dichelobacter nodosus. Vet. Microbiol. 36:113–122.
38. Park, H. S. M., M. Wolfgang, and M. Koomey. 2002. Modification of type IV
3334 HAN ET AL. J. BACTERIOL.

pilus-associated epithelial cell adherence and multicellular behavior by thePilU protein of Neisseria gonorrhoeae. Infect. Immun. 70:3891–3903.
39. Parker, D., R. M. Kennan, G. S. Myers, I. T. Paulsen, J. G. Songer, and J. I.Rood. 2006. Regulation of type IV fimbrial biogenesis in Dichelobacter no-dosus. J. Bacteriol. 188:4801–4811.
40. Peabody, C. R., Y. J. Chung, M. Yen, D. Vidal-Ingigliardi, A. P. Pugsley, andM. H. Saier, Jr. 2003. Type II protein secretion and its relationship tobacterial type IV pili and archaeal flagella. Microbiology 149:3051–3072.
41. Pugsley, A. P. 1993. The complete general secretory pathway in gram-neg-ative bacteria. Microbiol. Rev. 57:50–108.
42. Pujol, C., E. Eugene, M. Marceau, and X. Nassif. 1999. The meningococcalPilT protein is required for induction of intimate attachment to epithelialcells following pilus-mediated adhesion. Proc. Natl. Acad. Sci. USA 96:4017–4022.
43. Rood, J. I. 2002. Genomic islands of Dichelobacter nodosus. Curr. Top.Microbiol. Immunol. 264:47–60.
44. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold SpringHarbor, NY.
45. Satyshur, K. A., G. A. Worzalla, L. S. Meyer, E. K. Heiniger, K. G. Aukema,A. M. Misic, and K. T. Forest. 2007. Crystal structures of the pilus retractionmotor PilT suggest large domain movements and subunit cooperation drivemotility. Structure 15:363–376.
46. Sauvonnet, N., G. Vignon, A. P. Pugsley, and P. Gounon. 2000. Pilus forma-tion and protein secretion by the same machinery in Escherichia coli. EMBOJ. 19:2221–2228.
47. Skerker, J. M., and H. Berg, C. 2001. Direct observation of extension andretraction of type IV pili. Proc. Natl. Acad. Sci. USA 98:6901–6904.
48. Stewart, D. J. 1979. The role of elastase in the differentiation of Bacteroidesnodosus infections in sheep and cattle. Res. Vet. Sci. 27:99–105.
49. Thomas, J. H. 1958. A simple medium for the isolation and cultivation ofFusiformis nodosus. Aust. Vet. J. 34:411.
50. Wang, R. F., and S. R. Kushner. 1991. Construction of versatile low-copy-
number vectors for cloning, sequencing and gene expression in Escherichiacoli. Gene 100:195–199.
51. Watson, A. A., J. S. Mattick, and R. A. Alm. 1996. Functional expression ofheterologous type 4 fimbriae in Pseudomonas aeruginosa. Gene 175:143–150.
52. Whitchurch, C. B. 2006. Type IV pili in Pseudomonas species, p. 139–188. InJ. L. Ramos and R. J. Levesque (ed.), Pseudomonas, vol. IV. Kluwer Aca-demic/Plenum Publishers, New York, NY.
53. Whitchurch, C. B., M. Hobbs, S. P. Livingston, V. Krishnapillai, and J. S.Mattick. 1991. Characterisation of a Pseudomonas aeruginosa twitching mo-tility gene and evidence for a specialised protein export system widespread ineubacteria. Gene 101:33–44.
54. Whitchurch, C. B., and J. S. Mattick. 1994. Characterization of a gene, pilU,required for twitching motility but not phage sensitivity in Pseudomonasaeruginosa. Mol. Microbiol. 13:1079–1091.
55. Whittington, R. J., and P. J. Nicholls. 1995. Grading the lesions of ovinefootrot. Res. Vet. Sci. 58:26–34.
56. Wolfgang, M., P. Lauer, H. S. Park, L. Brossay, J. Hebert, and M. Koomey.1998. PilT mutations lead to simultaneous defects in competence for naturaltransformation and twitching motility in piliated Neisseria gonorrhoeae. Mol.Microbiol. 29:321–330.
57. Wolfgang, M., J. P. van Putten, S. F. Hayes, D. Dorward, and M. Koomey.2000. Components and dynamics of fiber formation define a ubiquitousbiogenesis pathway for bacterial pili. EMBO J. 19:6408–6418.
58. Wu, S., J. Wu, and D. Kaiser. 1997. The Myxococcus xanthus pilT locus isrequired for social gliding motility although pili are still produced. Mol.Microbiol. 23:109–121.
59. Yasukawa, K., P. Martin, C. R. Tinsley, and X. Nassif. 2006. Pilus-mediatedadhesion of Neisseria meningitidis is negatively controlled by the pilus-retrac-tion machinery. Mol. Microbiol. 59:579–589.
60. Zolfaghar, I., D. J. Evans, and S. M. Fleiszig. 2003. Twitching motilitycontributes to the role of pili in corneal infection caused by Pseudomonasaeruginosa. Infect. Immun. 71:5389–5393.
VOL. 190, 2008 TWITCHING MOTILITY AND VIRULENCE OF D. NODOSUS 3335




















