
Functional Characterization of 8-Oxoguanine DNA Glycosylase of Trypanosoma cruzi
Upload
independentCategory
view
0download
0
Infection, Genetics and Evolution 25 (2014) 157–165
Contents lists available at ScienceDirect
Infection, Genetics and Evolution
journal homepage: www.elsevier .com/locate /meegid
Trypanosoma cruzi: Genome characterization of phosphatidylinositolkinase gene family (PIK and PIK-related) and identification of a novelPIK gene
http://dx.doi.org/10.1016/j.meegid.2014.03.0221567-1348/� 2014 Elsevier B.V. All rights reserved.
⇑ Corresponding author at: Departamento de Microbiologia, Imunologia eParasitologia, Escola Paulista de Medicina, UNIFESP, Rua Botucatu 862, 6o andar,04023-062 São Paulo, SP, Brazil. Tel.: +55 11 5576 4532; fax: +55 11 5571 1095x22.
E-mail address: [email protected] (D. Bahia).1 The authors wish it be known that in their opinion the three first authors are joint
first authors.
Priscila Oliveira a,1, Fabio Mitsuo Lima a,1, Mario Costa Cruz a,1, Renata Carmona Ferreira a,Alejandro Sanchez-Flores b, Esteban Maurício Cordero a, Danielle Rodrigues Cortez a,Éden Ramalho Ferreira a, Marcelo Ribeiro da Silva Briones a, Renato Arruda Mortara a,José Franco da Silveira a, Diana Bahia a,c,⇑a Departamento de Microbiologia, Imunologia e Parasitologia, Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo, SP, Brazilb Unidad Universitaria de Apoyo Bioinformático, Instituto de Biotecnología de la UNAM, Mexicoc Departamento de Biologia Geral, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Av. Antonio Carlos 6627, Pampulha, Caixa Postal 486, Belo Horizonte,Minas Gerais CEP 31270-910, Brazil
a r t i c l e i n f o a b s t r a c t
Article history:Received 5 April 2013Received in revised form 17 March 2014Accepted 24 March 2014Available online 13 April 2014
Keywords:Phosphatidylinositol kinasesTrypanosoma cruziTarget of rapamycinFYVE domainFYVE-PIKPIK
Chagas disease is caused by the protozoan Trypanosoma cruzi which affects 10 million people worldwide.Very few kinases have been characterized in this parasite, including the phosphatidylinositol kinases(PIKs) that are at the heart of one of the major pathways of intracellular signal transduction. Recently,we have classified the PIK family in T. cruzi using five different models based on the presence of PIKconserved domains. In this study, we have mapped PIK genes to the chromosomes of two different T. cruzilineages (G and CL Brener) and determined the cellular localization of two PIK members. The kinases havecrucial roles in metabolism and are assumed to be conserved throughout evolution. For this reason, theyshould display a conserved localization within the same eukaryotic species. In spite of this, there is anextensive polymorphism regarding PIK localization at both genomic and cellular levels, among differentT. cruzi isolates and between T. cruzi and Trypanosoma brucei, respectively. We showed in this study thatthe cellular localization of two PIK-related proteins (TOR1 and 2) in the T. cruzi lineage is distinct fromthat previously observed in T. brucei. In addition, we identified a new PIK gene with peculiar feature, thatis, it codes for a FYVE domain at N-terminal position. FYVE-PIK genes are phylogenetically distant fromthe groups containing exclusively the FYVE or PIK domain. The FYVE-PIK architecture is only present intrypanosomatids and in virus such as Acanthamoeba mimivirus, suggesting a horizontal acquisition. OurBayesian phylogenetic inference supports this hypothesis. The exact functions of this FYVE-PIK geneare unknown, but the presence of FYVE domain suggests a role in membranous compartments, such asendosome. Taken together, the data presented here strengthen the possibility that trypanosomatidsare characterized by extensive genomic plasticity that may be considered in designing drugs and vaccinesfor prevention of Chagas disease.
� 2014 Elsevier B.V. All rights reserved.
1. Introduction triatomine hematophagous insects (reviewed in Lima et al., 2010).
Among the parasitic diseases in the world, one of the mostneglected is Chagas disease, caused by Trypanosoma cruzi, aflagellated protozoan transmitted to mammalian hosts by
Chagas disease, also called American trypanosomiasis, is endemicin Latin America, and it has been estimated that 10 million peopleare affected worldwide (see the WHO factsheet on Chagas disease).
Because of the heavy migration of Latin Americans from ende-mic regions, Chagas disease has already reached several countries,including the United States, Canada, Spain, and Japan (Coura andDias, 2009). In the Americas, it is the most important parasitic dis-ease, responsible for a burden of 670,000 disability-adjusted lifeyears (DALYs) (WHO. The World Health Organization report2004. Changing history. Geneva: WHO; 2004).

158 P. Oliveira et al. / Infection, Genetics and Evolution 25 (2014) 157–165
In 2005, the genomic sequences of T. cruzi (El-Sayed et al., 2005)and two other pathogenic trypanosomatids (Trypanosoma bruceiand Leishmania major) were published. Since then, the number ofstudies about the presence, expression and potential interactionsbetween parasite components has increased rapidly. Whereasearlier molecular karyotyping studies generated the originalperceptions of the genetics and evolution of these protozoa andprovided some possible explanations for the parasite’s diversephenotypes – such as drug resistance and pathogenicity – recentsequencing of other genomes has yielded precise information onthe content and architecture of trypanosomatid genomes(Dujardin, 2009).
The phosphatidylinositol kinases (PIKs) comprise a large familyof lipid kinases that includes PI3 and PI4 kinases (conventional)and PI-related kinases (reviewed in Marone et al., 2008). The activ-ity of PIKs has been detected in all eukaryotic cells, and associatedwith several cellular functions and processes such as apoptosis,mitosis, membrane trafficking, cytokinesis and cytoskeletal organi-zation (Marone et al., 2008).
We recently performed a screening for conventional PIK andPIK-related kinases by similarity searches against the five humanpathogenic trypanosomatid genomes available to date (Bahiaet al., 2009a) and also the parasite worm Schistosoma mansoni(Bahia et al., 2009b) genome. We have classified the conventionalPIK and PIK-related subtypes into five different models accordingto their similarity and the presence of domains. In the presentstudy, we determine the genomic organization of PIK andPIK-related genes in two major phylogenetic strains of T. cruzi. Inaddition, we identify a novel PIK gene that encodes a PIK proteincarrying the FYVE domain at the N-terminus, unique to trypanoso-matids. We would like to suggest that the FYVE-PIK proteinswere acquired by horizontal gene transfer from virus totrypanosomatids.
Finally, we also showed the cellular localization ofphosphatidylinositol-related kinase TOR1 and 2 Model 5 in T. cruziepimastigotes and trypomastigotes.
2. Material and methods
2.1. Parasite strains and cultivation; preparation, separation andhybridization of chromosomes
All steps have been previously described by (Cano et al., 1995).Epimastigotes of T. cruzi from G strain (TcI) and clone CL Brener(TcVI) were grown in axenic cultures in liver infusion tryptose(LIT) medium containing 10% fetal bovine serum at 28 �C.Epimastigotes were grown to late logarithmic phase, washedthree times with PBS and collected by centrifugation. An equalvolume of cells was mixed with 2% low melting temperatureagarose prepared in bidistilled water. Aliquots of 100 lL containingapproximately 107 cells were deposited in appropriate molds andleft to solidify at room temperature and processed.
T. cruzi chromosomes were separated by pulsed-field gelelectrophoresis (PFGE) using the Gene Navigator System(Amersham Pharmacia Biotech, NJ, USA) and a hexagonal electrodearray. PFGE was carried out in 1.1% agarose gel in 0.5� TBE (45 mMTris; 45 mM boric acid; 1 mM EDTA, pH 8.3) for 132 h at 13 �C.Chromosomal bands were incubated with 0.25 M HCl for 30 min,denatured with 0.5 M NaOH/1 M NaCl for 20 min, neutralizedwith 1 M Tris-base/0.5 M NaCl for 20 min and transferred to nylonmembranes in 20� SSC using the VacuGene XL system(Pharmacia).
Specific sequences from each of the five models PIK and PIK-related genes were selected by eye (Supplementary Table 1),cloned by PCR and used as probes in Southern-blot hybridizations.
2.2. Domain architecture search
The protein domain architecture of the gene productTc00.1047053506425.80 (TcCLB.506425.80) from the Trypanosomacruzi CL Brener non-Esmeraldo-like genome from TriTrypDB v3.3database was taken as a reference for the domain search. Theprotein domain order and characterization of the reference proteinwas performed using the protein domain database PROSITE (Huloet al., 2006) and the stand-alone ScanProsite tool (Gattiker et al.,2002) with default parameters.
The protein domains (PS50178 and PS50290) found in the initialsurvey were used to search among all annotated proteins in theTriTrypDB v3.3 database to obtain the proteins sharing the samedomains and the same order in other kinetoplastid organisms.Again, the search was performed with the ScanProsite tool usingdefault parameters.
2.3. BLAST searches, multiple sequence alignments and distance trees
Orthology between trypanosomatid proteins found using theScanProsite tools was confirmed using the reciprocal best hit(RBH) criteria (Moreno-Hagelsieb and Latimer, 2008) comparingBLAST hits. To find closely related kinase proteins in other organ-isms, a BLAST search on the NCBI site using these trypanosomatidorthologs http://blast.ncbi.nlm.nih.gov was also performed againstthe RefSeq database, excluding all trypanosomatidae proteins (tax-id:5654). The top 10 hits from each query search were analyzed.
We aligned the proteins and separated domains from all theproteins using MUSCLE v3.8.31 (http://www.drive5.com/muscle)and default parameters. We edited the alignments using Jalviewv2.7 and neighbor-joining distance trees (using the BLOSUM62matrix distance) were generated with the tools inside the sameprogram.
2.4. Phylogenetic analysis
Amino acid sequences were retrieved from TriTrypDB (http://tritrypdb.org) searches using FYVE as word in the Gene TextSearch. 48 ortholog groups resulted from this search. Alignmentof these 48 sequences with the PIK sequences from trypanoso-matid, Homo sapiens, Danio rerio, Drosophila melanogaster andArabidopsis thaliana, totalizing 102 sequences, was performedusing MUSCLE implemented in SEAVIEW software (Edgar,2004a,b; Galtier et al., 1996).
Phylogeny was inferred using Bayesian method as implementedin MrBayes 3.1.2 estimating the amino acid substitution model(Ronquist and Huelsenbeck, 2003). Data were saved every 100generation and run in 4 chains and 2 runs. Bayesian tree wasinferred based on 1 � 107 generations with burnin = 75,000. Theconsensus trees were used to determine the posterior probabilityvalues. The generated consensus tree was visualized in the Figtreev1.3.1 program (http://tree.bio.ed.ac.uk/software/figtree/).
2.5. Antibodies and immunofluorescence assays
Rabbit and rat antisera raised against specific TcTOR1 (Tc00.1047053508231.30; Ac-CSYDTVKFRERSIPV-amide) and TcTOR2peptides (Tc00.1047053510689.40; Ac-IRRIANDPEKRAVC-amide)coupled to KLH were IgG-purified by biochemical affinity column(New England Peptides) (Supplementary data 1). The antibodiesa-BiP and a-DHLADH, used to immunostain the endoplasmicreticulum and mitochondrion, respectively, were a kind gift fromDr. Sergio Schenkman (EPM, UNIFESP).
Epimastigotes were washed in PBS and fixed for 30 minwith 3.5% formaldehyde in PBS. The TcTOR1, TcTOR2, ER andmitochondrion of the parasites were identified by incubation

P. Oliveira et al. / Infection, Genetics and Evolution 25 (2014) 157–165 159
with specific antibodies anti-TcTOR1 and 2 for 12 h, and anti-BiPand anti-DHLADH for 1 h, followed by incubation with secondaryantibodies (Alexa Fluor-488� and 568� conjugated secondary anti-bodies) for 1 h. Samples were stained for 15 min with 100 lg/mL40,6-diamidino-2-phenylindole (DAPI, Invitrogen, Life Technolo-gies) and mounted with 90% glycerol in PBS containing 0.01%p-phenylenediamine.
2.6. Confocal imaging and colocalization analysis
Confocal images of immunostained epimastigotes wereobtained using a Leica TCS SP5 II system. Approximately 15z-stacks of each microscopic field containing parasites wereacquired using a 100� (1.44 NA) oil immersion objective at aresolution of 2048 � 2048 pixels. Images were imported intoImaris v7.5 beta 2 imaging software (Bitplane AG) and renderedusing MIP or Blend image filters. Colocalized voxels (pixels plusz-stacks) having red and green fluorescence were quantified permicroscopic field using the Imaris coloc tool. The colocalization ofeach combination of antibodies studied was performed using thesame red and green colocalization threshold. The number ofcolocalized voxels per field was divided by the number of parasitescounted in the same microscopic field and plotted in a comparativegraph (mean number of colocalized voxels per parasite, S.E.M, n = 5microscopic fields).
3. Results and discussion
3.1. Genomic organization of PIK family
In a previous study, we identified nine complete genesannotated as phosphatidylinositol kinase in clone CL Brener(Bahia et al., 2009a). Here we first analyzed the distribution ofPIK genes in the in silico assembled chromosomes of clone CLBrener. Recently, T. cruzi sequence contigs were assembled into41 scaffolds tentatively named in silico chromosomes (TcChr)(Weatherly et al., 2009). This designation is not accurate given thatsome of these chromosomes could be part of a single chromosome(Souza et al., 2011). Clone CL Brener is a hybrid that displays twohaplotypes. Therefore, the chromosome-sized scaffolds assignedto the Esmeraldo and non-Esmeraldo haplotypes were designatedS and P, respectively.
The PIK genes were assigned to seven in silico chromosomes ofclone CL Brener (Table 1). Three PIK genes were mapped withindistances of 81.6–93.3 Kb on the chromosome TcChr34, indicatingthat they were densely clustered on the genome. Each one ofremaining markers was mapped to a single in silico chromosome
Table 1Location of database completed PIK genes on the CL Brener and G strain chromosomal ba
Model* Locus tag Annotation
1 Tc00.1047053511903.160 PIK2 Tc00.1047053508859.90 PIK3 Tc00.1047053508695.20 PIK4 Tc00.1047053510003.30 PIK5 Tc00.1047053508231.30 TOR1
Tc00.1047053508257.230 TOR-like1Tc00.1047053510689.40 TOR2Tc00.1047053509395.20 ATMTc00.1047053506223.120 ATR
* Classification of the T. cruzi conventional PIK and PIK-related subtypes into five differen2009a).** Chromosome-sized in silico scaffolds assembled by EuPath database (TriTrypDB: htscaffolds assigned to the Esmeraldo and non-Esmeraldo haplotypes were designated S a*** Chromosomal bands are those separated by PFGE and visible after staining with eth
(Table 1). Analysis of the genomic environment of each PIK andof PIK-related genes showed that their neighbor genes display abroad spectrum of biological functions (not shown).
3.2. Chromosomal mapping of PIK gene family
Chromosomal bands separations by PFGE have allowed sizing ofindividual chromosomes and also enabled cloned genes to beassigned to their chromosomes by hybridization with PIK andPIK-related genes (Fig. 1). Model 1 PIK probe hybridized with chro-mosomal bands VII (0.97 Mb) and VI (0.85 Mb) in CL Brener andbands 7 (0.87 Mb) and 6 (0.77 Mb) in the G strain. Model 2 PIKprobe also mapped in two bands for CL Brener (VI and IV) of 0.85and 0.72 Mb, respectively, and in chromosomal band 8 (0.96 Mb)in the G strain. A similar hybridization profile to Model 2 wasobserved with the Model 3 PIK probe, which hybridized with bandsXIX (2.95 Mb) and XV (1.65 Mb) in clone CL Brener and band 9(1.08 Mb) in the G strain. A different hybridization profile wasfound with the Model 4 PIK probe, which mapped to only one chro-mosomal band of 1.43 Mb (XIII) in clone CL Brener and band 11(1.35 Mb) in the G strain.
In addition, PIK-related genes (Model 5) mapped in two chro-mosomal bands in CL Brener and, mostly, in one band in G strain.All five members mapped the chromosomal band XIX (2.95 Mb)in clone CL Brener, whereas the second hybridization signal foreach probe were in bands X (1.20 Mb), XV (1.65 Mb) or XX(3.27 Mb), suggesting that the localization in larger chromosomesis a tendency of PIK-related genes (Fig. 1 and Table 1). In the Gstrain, the Tc00.1047053508231.30 Model 5 (TOR1) PIK-relatedgene was the only one mapped on two chromosomal bands (2.83and 2.00 Mb), whereas the others mapped to one band (Fig. 1and Table 1). Although PIK-related genes Tc00.1047053508231.30 and Tc00.1047053506223.120, annotated as TOR1 and ATR,respectively, hybridized with chromosomal band XIX in clone CLBrener they cannot be assigned to a same chromosomal molecule.The in silico prediction indicates they are located in TcChr37 andTcChr35, respectively, and the second hybridization signal do notmatch with the profile obtained by genes predicted in TcChr34and TcChr1. As previously demonstrated (Souza et al., 2011; Limaet al., 2013), one chromosomal band visualized by PFGE can sheltertwo or more heterologous chromosomes of same size. This mayoccur in CL Brener chromosomal band XIX.
Only one probe (Model 4 PIK) out of nine PIK and PIK-relatedgenes hybridized with one chromosomal band in clone CL Brener,whereas the remaining mapped on two bands. This result couldbe due to the hybrid nature of the CL Brener genome, whichimplies a great number of homologous chromosomes of different
nds.
TcChr** Chr band***
CL Brener G
TcChr26-P VII; VI 7; 6TcChr11-P VI; IV 8TcChr34-P XIX; XV 9TcChr36-P XIII 11TcChr37-S XX; XIX 19; 15TcChr34-P XIX; XV 9TcChr34-P XIX; XV 9TcChr1-P XIX; XV 9TcChr35-S XIX; X 13
t models according to their similarity and presence of specific domains (Bahia et al.,
tp://tritrypdb.org). Clone CL Brener is a hybrid, therefore, the chromosome-sizednd P, respectively.idium bromide.

Fig. 1. Distribution of the PIK family on chromosomal bands of the CL Brener and G strains. Chromosomal bands were fractioned by PFGE and hybridized with PIK and PIK-related genes as probes. Panels A–D show chromosomal mapping of conventional PIK and Mod1–Mod4 genes, respectively. Panels E and F show FYVE-PIK and PIK-relatedgene mapping, respectively. Left, CL Brener (CLB) chromosomal bands recognized by the probe; right, the corresponding chromosomal bands in G strain (G).
160 P. Oliveira et al. / Infection, Genetics and Evolution 25 (2014) 157–165
sizes (Santos et al., 1999). On the other hand, it was possible toobserve in the G strain a different hybridization profile. Sevenout of PIK and PIK-related genes hybridized with only one band.It was also observed that three genes, one from Model 3 and twofrom Model 5, belonging to the TcChr34 showed the samehybridization profile in CL Brener (bands XIX and XV) and G strain(band 9) (Table 1). The same profile was observed for theTc00.1047053509395.20 Model 5 gene that belongs to TcChr1.The fact that markers of chromosome TcChr1 and TcChr34 pro-duced identical hybridization patterns to the chromosomal bandsof different isolates suggests that they are syntenic. Large-scalesynteny is a prominent characteristic that is shared by differenttrypanosomatids (Ghedin et al., 2004) and different T. cruzi lin-eages (Souza et al., 2011; Vargas et al., 2004).
3.3. Identification of a novel phosphatidylinositol kinase gene encodinga FYVE domain at N-terminus unique to trypanosomatids
During the mining step for identifying PIK genes in T. cruzi, wefound a gene that caught our attention (Tc001047053506425.80)in the phylogenetic tree of PIK genes (Fig. 2A). The product of thisgene contains a FYVE domain (PS50178) at the N-terminus followedby the phosphatidylinositol kinase (PS50290) domain (Fig. 2B). Thepresence of FYVE domain accounts for the misalignment of thisgene observed during the construction of the phylogenetic tree of
PIK genes. Searching the literature, we found a protein called PIK-fyve (Rutherford et al., 2006). This protein is not a PI-3-kinase butrather a PI-3-phosphate 5-kinase; therefore it is not related to thisnew PIK model. In addition, there are records of FYVE-PIK with aPI3Ka accessory motif (PF00613) in Polysphondylium pallidum,Dictyostelium purpureum and other organisms. Using the samestrategy employed for PIK and PIK-related genes, we performedthe chromosomal mapping of this T. cruzi FYVE-PIK gene. This probehybridized with bands XIX (2.95 Mb) and XII (1.35 Mb) in CL Brenerand band 12 (1.43 Mb) in G strain (Fig. 1E). This hybridization pat-tern was different from that observed for other PIK family mem-bers, except for CL Brener chromosomal band XIX that harbors sixother PIK genes (Fig. 1E and Table 1).
When searching the FYVE-PIK gene in the available genomes ofother trypanosomatids, we found the corresponding orthologs(Fig. 3) but no paralogs (not shown). The FYVE-PIK genes andproteins of trypanosomatids are phylogenetically distant fromthe group containing exclusively the FYVE or PIK domain (notshown). Moreover, we searched for this particular FYVE-PIKdomain architecture in other non-trypanosomatid organisms byusing BLAST and also in the profile databases PFAM (FYVE[PF01363]; PIK [PF00454]). There is no record in any database ofproteins with this domain combination. Curiously, we found thisarchitecture only in virus species, such as Acanthamoeba polyphagamimivirus, which has been included in our alignments (Figs. 3 and

Fig. 2. Modular architecture and tree based on PI-K in T. cruzi genome. (A) The phylogenetic tree was generated by MEGA 3.1 using the neighbor-joining method withpairwise gap removal and PAM distances from a multiple alignment of the T. cruzi PIK genes generated by CLUSTALW. PI-K, PI-kinase catalytic domain (PS50290); C2, C2-likedomain in PI kinases (PF00792); kA, accessory PIK domain (PF00613); RBD, ras-binding domain (PF08771); FAT and FATC (PF02260). FYVE-PIK is a novel PIK gene in T. cruzithat encodes a FYVE domain (PF01363) at the N-terminus. Models 1, 2, 3, 4, 5 designate different subtypes of PI- and related kinases, based on the tree generated by Bosottiet al. (2000) (Bahia et al., 2009a,b). (B) Primary structure and architecture of the novel T. cruzi PIK with a FYVE domain at N-terminus. A FYVE domain sequence (Fig. 4) isobserved within amino acids 311 to 395. The PIK domain is found from amino acids 757 to 1025, which retains the kinase catalytic activity of the molecule (see alsoSupplementary material Fig. S1).
Fig. 3. Maximum likelihood distance tree of PIK and FYVE-PIK proteins. The tree represents the relationship of trypanosomatid proteins (Trypanosoma genus in red andLeishmania genus in blue) containing FYVE and PIK domains with other human PIK (in black) without a FYVE domain and the Acanthamoeba polyphaga mimivirus protein (inblack) containing both FYVE and PIK domains. Bootstrap values are depicted at the base of each node. (For interpretation of the references to color in this figure legend, thereader is referred to the web version of this article.)
P. Oliveira et al. / Infection, Genetics and Evolution 25 (2014) 157–165 161

Fig. 4. Protein sequence alignment of trypanosomatid FYVE domain. A region of approximately 135 amino acids of the N-terminal FYVE domain of Leishmania spp.,Trypanosoma spp. and Acanthamoeba polyphaga mimivirus was aligned. Boxes depict the most-conserved amino acids (FYVE signature).
162 P. Oliveira et al. / Infection, Genetics and Evolution 25 (2014) 157–165
4 and Supplementary material Fig. S1). However, there is no rela-tionship between the virus gene/protein and the trypanosomatidscontaining the FYVE-PIK architecture.
Fig. 5. Bayesian phylogeny of PIK, FYVE and PIK-FYVE proteins from the trypanosomatigenerations. Tree is shown as midpoint rooted. The arrow indicates the position of Tc00.substitution per site. Letters and numbers represent the TriTrypDB (http://tritrypdb.org
There is an expected divergence between Trypanosoma andLeishmania FYVE-PIK genes/proteins, but the Acanthamoebapolyphaga mimivirus FYVE-PIK is clearly distant from the group of
ds. Tree was inferred with amino-acid sequences and was calculated from 1 � 107
1047053506425.80 (currently TcCLB.506425.80) protein and the bar the number of) accession number.
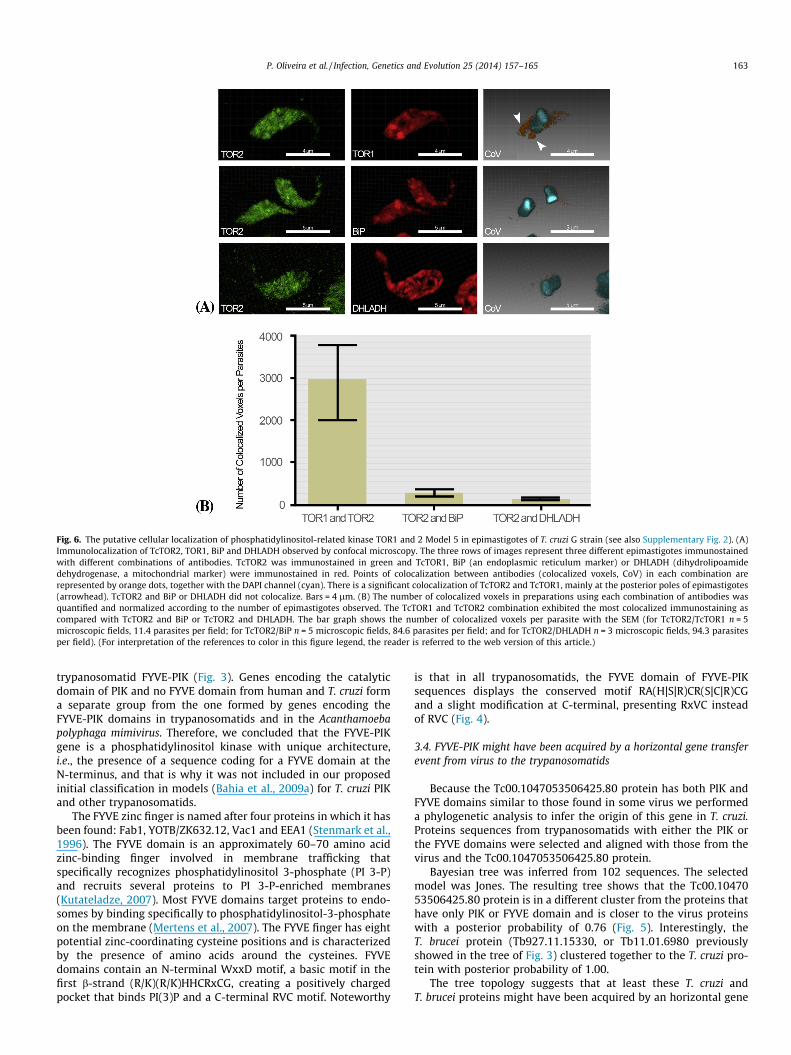
Fig. 6. The putative cellular localization of phosphatidylinositol-related kinase TOR1 and 2 Model 5 in epimastigotes of T. cruzi G strain (see also Supplementary Fig. 2). (A)Immunolocalization of TcTOR2, TOR1, BiP and DHLADH observed by confocal microscopy. The three rows of images represent three different epimastigotes immunostainedwith different combinations of antibodies. TcTOR2 was immunostained in green and TcTOR1, BiP (an endoplasmic reticulum marker) or DHLADH (dihydrolipoamidedehydrogenase, a mitochondrial marker) were immunostained in red. Points of colocalization between antibodies (colocalized voxels, CoV) in each combination arerepresented by orange dots, together with the DAPI channel (cyan). There is a significant colocalization of TcTOR2 and TcTOR1, mainly at the posterior poles of epimastigotes(arrowhead). TcTOR2 and BiP or DHLADH did not colocalize. Bars = 4 lm. (B) The number of colocalized voxels in preparations using each combination of antibodies wasquantified and normalized according to the number of epimastigotes observed. The TcTOR1 and TcTOR2 combination exhibited the most colocalized immunostaining ascompared with TcTOR2 and BiP or TcTOR2 and DHLADH. The bar graph shows the number of colocalized voxels per parasite with the SEM (for TcTOR2/TcTOR1 n = 5microscopic fields, 11.4 parasites per field; for TcTOR2/BiP n = 5 microscopic fields, 84.6 parasites per field; and for TcTOR2/DHLADH n = 3 microscopic fields, 94.3 parasitesper field). (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
P. Oliveira et al. / Infection, Genetics and Evolution 25 (2014) 157–165 163
trypanosomatid FYVE-PIK (Fig. 3). Genes encoding the catalyticdomain of PIK and no FYVE domain from human and T. cruzi forma separate group from the one formed by genes encoding theFYVE-PIK domains in trypanosomatids and in the Acanthamoebapolyphaga mimivirus. Therefore, we concluded that the FYVE-PIKgene is a phosphatidylinositol kinase with unique architecture,i.e., the presence of a sequence coding for a FYVE domain at theN-terminus, and that is why it was not included in our proposedinitial classification in models (Bahia et al., 2009a) for T. cruzi PIKand other trypanosomatids.
The FYVE zinc finger is named after four proteins in which it hasbeen found: Fab1, YOTB/ZK632.12, Vac1 and EEA1 (Stenmark et al.,1996). The FYVE domain is an approximately 60–70 amino acidzinc-binding finger involved in membrane trafficking thatspecifically recognizes phosphatidylinositol 3-phosphate (PI 3-P)and recruits several proteins to PI 3-P-enriched membranes(Kutateladze, 2007). Most FYVE domains target proteins to endo-somes by binding specifically to phosphatidylinositol-3-phosphateon the membrane (Mertens et al., 2007). The FYVE finger has eightpotential zinc-coordinating cysteine positions and is characterizedby the presence of amino acids around the cysteines. FYVEdomains contain an N-terminal WxxD motif, a basic motif in thefirst b-strand (R/K)(R/K)HHCRxCG, creating a positively chargedpocket that binds PI(3)P and a C-terminal RVC motif. Noteworthy
is that in all trypanosomatids, the FYVE domain of FYVE-PIKsequences displays the conserved motif RA(H|S|R)CR(S|C|R)CGand a slight modification at C-terminal, presenting RxVC insteadof RVC (Fig. 4).
3.4. FYVE-PIK might have been acquired by a horizontal gene transferevent from virus to the trypanosomatids
Because the Tc00.1047053506425.80 protein has both PIK andFYVE domains similar to those found in some virus we performeda phylogenetic analysis to infer the origin of this gene in T. cruzi.Proteins sequences from trypanosomatids with either the PIK orthe FYVE domains were selected and aligned with those from thevirus and the Tc00.1047053506425.80 protein.
Bayesian tree was inferred from 102 sequences. The selectedmodel was Jones. The resulting tree shows that the Tc00.1047053506425.80 protein is in a different cluster from the proteins thathave only PIK or FYVE domain and is closer to the virus proteinswith a posterior probability of 0.76 (Fig. 5). Interestingly, theT. brucei protein (Tb927.11.15330, or Tb11.01.6980 previouslyshowed in the tree of Fig. 3) clustered together to the T. cruzi pro-tein with posterior probability of 1.00.
The tree topology suggests that at least these T. cruzi andT. brucei proteins might have been acquired by an horizontal gene

164 P. Oliveira et al. / Infection, Genetics and Evolution 25 (2014) 157–165
transfer (HGT) event from virus to the trypanosomatids. A neigh-bor joining tree with sequences from Fig. 3 added to these 102sequences showed that the Trypanosoma and Leishmania proteinscluster together with the virus sequence; the virus protein is closerto the Trypanosoma proteins than it is to Leishmania proteins (datanot shown). Taken together these data raise two possibilities fromwhere the HGT might have occurred. Assuming that T. cruzi and T.brucei are monophyletic the HGT event could have occurred beforethe separation of Leishmania and Trypanosoma. On the other hand,one can assume that each HGT is independent and occurred indifferent times during the evolution of T. brucei, Leishmania andT. cruzi. We believe that the first hypothesis is more likely to occurbecause it is the most parsimonious involving only one acquisitionof the FYVE-PIK genes. Further analysis has to be made to confirmthis HGT hypothesis.
HGT is an important evolutionary mechanism and is relativelycommon in trypanosomatids (Opperdoes and Michels, 2007). Alarge number of plant-like, bacterial and virus genes were identi-fied in the genome of the trypanosomatids that allowed the adap-tation of these organisms to a vast range of different hosts (Ienneet al., 2012; Opperdoes and Michels, 2007; Rigden et al., 2001;Silva et al., 2013; Steglich and Schaeffer, 2006). This T. cruzi genemight be one which has entered by a HGT event in a trypanoso-matid ancestor that allowed adaptative advantages.
3.5. The cellular localization of phosphatidylinositol-related kinaseTOR1 and 2 Model 5 in T. cruzi is distinct from that in T. brucei
The localization of signaling molecules is key to regulate theirfunction and specificity. We raised specific antibodies to twomembers of Model 5 PIK-related (TcTOR1 and TcTOR2) to verifytheir subcellular localization in epimastigotes of T. cruzi (G strain).TOR (Target of rapamycin) couples nutrient availability toactivation of processes, including metabolism, transcription andtranslation, that leads to cell growth (reviewed in Gough, 2012;Shertz and Cardenas, 2011). There are two TOR complexes (TORC1and 2) with specialized functions (Loewith and Hall, 2011). Theoverall TOR functions may be conserved in early-branchingeukaryotes; however, several new findings have emerged thatdiffer from those described for other eukaryotes. A number of sub-cellular TOR localizations have been reported for different organ-isms (Loewith and Hall, 2011; Sturgill et al., 2008; Tsang andZheng, 2007; Wedaman et al., 2003). In T. brucei TOR, TbTOR1has been shown to be predominantly nuclear and TbTOR2 to beassociated with the endoplasmic reticulum (ER) and mitochondria(Barquilla et al., 2008). The localization of Model 5 PIK-related T.cruzi TOR1 and 2 (TcTOR1 and TcTOR2) in epimastigotes (andtrypomastigotes, Supplementary material Fig. S2) of G strain iscompletely distinct from that previously observed in T. brucei.Unlike TbTOR1, TcTOR1 is excluded from nucleus (Fig 6A: first line,second square) and is concentrated at the posterior portion inpunctuated compartments close to reservosomes. TcTOR2, unlikeTbTOR2, is excluded from ER and mitochondria (Fig. 6A: secondand third lines, Fig. 6B) and is dispersed in the cytoplasm, concen-trating around the TcTOR1 location, in addition to presentingregions of colocalization with TcTOR1 (Fig 6A, arrows, and Fig6B). This unexpected localization of TOR proteins may shed newlight on TOR function in the metabolism, differentiation andinvasion processes of T. cruzi. Because TcTOR1 and TcTOR2 sharea partial localization, it is possible that the different TOR complexes– whether they are actually formed in T. cruzi – act by promotingcross talk to increase their activities. Considering the variety ofprocesses affected by the TOR kinases and that many of them areinfluenced by extracellular events, the endocytic compartmentrepresents a plausible location to anchor TOR-dependent signaling.Considering the roles in cell survival and virulence, inhibition of
the TOR kinase function in trypanosomatids offers an attractivetarget for chemotherapy. However, one study has recently shownthat the use of inhibitors for the TOR pathway in infections of hostcell fibroblasts by infective trypomastigotes has a minimum effectin T. cruzi virulence: no efficacy was observed against T. cruzi inin vivo assays (Diaz-Gonzalez et al., 2011) in contrast to what hasbeen achieved with other trypanosomatids. Given that these TORinhibitors have been effective for T. brucei, it is probably notsurprising that TOR proteins of both parasites are located incompartments distinct from each other and could display distinctsensitivity to these inhibitors.
4. Concluding remarks
The kinases have crucial roles in metabolism and are assumed tobe conserved throughout evolution. For this reason, they shoulddisplay a conserved localization within eukaryotes of the samespecies. However, the PIK family does not follow a pattern ofchromosomal localization between two different T. cruzi lineages,G and CL Brener (or TcI and TcVI clades), nor within the samelineage. It is clear that trypanosomatids exhibit a high genomeplasticity (Dujardin et al., 2000). The intense genomic diversityamong T. cruzi lineages may be originated through different eventsof inter-strain intragenic recombination (Ferreira and Briones,2012).
The unrelated cellular location of TOR1 and 2 in T. cruzi withrespect to T. brucei may also unravel unexpected novel functionsof the TOR complex in this organism. The genomic polymorphismand also the putative distinct localization of PIK proteins withinTrypanosoma genus, such as TOR1 and 2, are issues that may beconsidered in designing drugs and vaccines for prevention ofChagas disease.
Acknowledgments
The authors wish to thank the support of FAPESP (07/50551-2)and CAPES. M.R.B., R.A.M., J.F.S. and D.B. are recipients of CNPq fel-lowships. Anti-BiP and DHLADH were kindly provided by Dr. SergioSchenkman (EPM, UNIFESP). TOR recombinants and unpublisheddata were kindly provided by Drs. Samuel Goldemberg and FabíolaBarbieri Holetz (FIOCRUZ). We thank Dr. Fernando Real (DMIP-UNIFESP) for Confocal acquisitions and immunofluorescence imageedition. Authors also thank BioMed Proofreading (http://www.bio-medproofreading.com/) for English editing services.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.meegid.2014.03.022.
References
Bahia, D., Oliveira, L.M., Lima, F.M., Oliveira, P., Silveira, J.F., Mortara, R.A., Ruiz, J.C.,2009a. The TryPIKinome of five human pathogenic trypanosomatids:Trypanosoma brucei, Trypanosoma cruzi, Leishmania major, Leishmaniabraziliensis and Leishmania infantum – new tools for designing specificinhibitors. Biochem. Biophys. Res. Commun. 390, 963–970.
Bahia, D., Oliveira, L.M., Mortara, R.A., Ruiz, J.C., 2009b. Phosphatidylinositol-andrelated-kinases: a genome-wide survey of classes and subtypes in theSchistosoma mansoni genome for designing subtype-specific inhibitors.Biochem. Biophys. Res. Commun. 380, 525–530.
Barquilla, A., Crespo, J.L., Navarro, M., 2008. Rapamycin inhibits trypanosome cellgrowth by preventing TOR complex 2 formation. Proc. Natl. Acad. Sci. U.S.A. 105,14579–14584.
Bosotti, R., Isacchi, A., Sonnhammer, E.L., 2000. FAT: a novel domain in PIK-relatedkinases. Trends Biochem. Sci. 25, 225–227.
Cano, M.I., Gruber, A., Vazquez, M., Cortés, A., Levin, M.J., González, A., Degrave, W.,Rondinelli, E., Zingales, B., Ramirez, J.L., 1995. Molecular karyotype of clone CL

P. Oliveira et al. / Infection, Genetics and Evolution 25 (2014) 157–165 165
Brener chosen for the Trypanosoma cruzi genome project. Mol. Biochem.Parasitol. 71, 273–278.
Coura, J.R., Dias, J.C., 2009. Epidemiology, control and surveillance of Chagas disease:100 years after its discovery. Mem. Inst. Oswaldo Cruz 104 (Suppl. 1), 31–40.
Diaz-Gonzalez, R., Kuhlmann, F.M., Galan-Rodriguez, C., Madeira da Silva, L.,Saldivia, M., Karver, C.E., Rodriguez, A., Beverley, S.M., Navarro, M., Pollastri,M.P., 2011. The susceptibility of trypanosomatid pathogens to PI3/mTOR kinaseinhibitors affords a new opportunity for drug repurposing. PLoS Negl. Trop. Dis.5, e1297.
Dujardin, J.C., 2009. Structure, dynamics and function of Leishmania genome:resolving the puzzle of infection, genetics and evolution? Infect. Genet. Evol. 9,290–297.
Dujardin, J.C., Henriksson, J., Victoir, K., Brisse, S., Gamboa, D., Arevalo, J., Le Ray, D.,2000. Genomic rearrangements in trypanosomatids: an alternative to the ‘‘onegene’’ evolutionary hypotheses? Mem. Inst. Oswaldo Cruz 95, 527–534.
Edgar, R.C., 2004a. MUSCLE: a multiple sequence alignment method with reducedtime and space complexity. BMC Bioinformatics 5, 113.
Edgar, R.C., 2004b. MUSCLE: multiple sequence alignment with high accuracy andhigh throughput. Nucleic Acids Res. 32, 1792–1797.
El-Sayed, N.M., Myler, P.J., Bartholomeu, D.C., Nilsson, D., Aggarwal, G., Tran, A.N.,Ghedin, E., Worthey, E.A., Delcher, A.L., Blandin, G., Westenberger, S.J., Caler, E.,Cerqueira, G.C., Branche, C., Haas, B., Anupama, A., Arner, E., Aslund, L., Attipoe,P., Bontempi, E., Bringaud, F., Burton, P., Cadag, E., Campbell, D.A., Carrington,M., Crabtree, J., Darban, H., da Silveira, J.F., de Jong, P., Edwards, K., Englund, P.T.,Fazelina, G., Feldblyum, T., Ferella, M., Frasch, A.C., Gull, K., Horn, D., Hou, L.,Huang, Y., Kindlund, E., Klingbeil, M., Kluge, S., Koo, H., Lacerda, D., Levin, M.J.,Lorenzi, H., Louie, T., Machado, C.R., McCulloch, R., McKenna, A., Mizuno, Y.,Mottram, J.C., Nelson, S., Ochaya, S., Osoegawa, K., Pai, G., Parsons, M., Pentony,M., Pettersson, U., Pop, M., Ramirez, J.L., Rinta, J., Robertson, L., Salzberg, S.L.,Sanchez, D.O., Seyler, A., Sharma, R., Shetty, J., Simpson, A.J., Sisk, E., Tammi,M.T., Tarleton, R., Teixeira, S., Van Aken, S., Vogt, C., Ward, P.N., Wickstead, B.,Wortman, J., White, O., Fraser, C.M., Stuart, K.D., Andersson, B., 2005. Thegenome sequence of Trypanosoma cruzi, etiologic agent of Chagas disease.Science 309, 409–415.
Ferreira, R.C., Briones, M.R., 2012. Phylogenetic evidence based on Trypanosomacruzi nuclear gene sequences and information entropy suggest that inter-strainintragenic recombination is a basic mechanism underlying the allele diversityof hybrid strains. Infect. Genet. Evol. 12, 1064–1071.
Galtier, N., Gouy, M., Gautier, C., 1996. SEAVIEW and PHYLO_WIN: two graphic toolsfor sequence alignment and molecular phylogeny. Comput. Appl. Biosci. CABIOS12, 543–548.
Gattiker, A., Gasteiger, E., Bairoch, A., 2002. ScanProsite: a reference implementationof a PROSITE scanning tool. Appl. Bioinformatics 1, 107–108.
Ghedin, E., Bringaud, F., Peterson, J., Myler, P., Berriman, M., Ivens, A., Andersson, B.,Bontempi, E., Eisen, J., Angiuoli, S., Wanless, D., Von Arx, A., Murphy, L., Lennard,N., Salzberg, S., Adams, M.D., White, O., Hall, N., Stuart, K., Fraser, C.M., El-Sayed,N.M., 2004. Gene synteny and evolution of genome architecture intrypanosomatids. Mol. Biochem. Parasitol. 134, 183–191.
Gough, N.R., 2012. TOR signaling, a tale of two complexes. Sci. Signal. 5, eg4.Hulo, N., Bairoch, A., Bulliard, V., Cerutti, L., De Castro, E., Langendijk-Genevaux, P.S.,
Pagni, M., Sigrist, C.J., 2006. The PROSITE database. Nucleic Acids Res. 34, D227–D230.
Ienne, S., Pappas Jr, G., Benabdellah, K., González, A., Zingales, B., 2012. Horizontalgene transfer confers fermentative metabolism in the respiratory-deficientplant trypanosomatid Phytomonas serpens. Infect. Genet. Evol. J. Mol. Epidemiol.Evol. Genet. Infect. Dis. 12, 539–548.
Kutateladze, T.G., 2007. Mechanistic similarities in docking of the FYVE and PXdomains to phosphatidylinositol 3-phosphate containing membranes. Prog.Lipid Res. 46, 315–327.
Lima, F.M., Oliveira, P., Mortara, R.A., Silveira, J.F., Bahia, D., 2010. The challenge ofChagas’ disease: has the human pathogen, Trypanosoma cruzi, learned how tomodulate signaling events to subvert host cells? N. Biotechnol. 27, 837–843.
Lima, F.M., Souza, R.T., Santori, F.R., Santos, M.F., Cortez, D.R., Barros, R.M., Cano, M.I.,Valadares, H.M., Macedo, A.M., Mortara, R.A., da Silveira, J.F., 2013. Interclonalvariations in the molecular karyotype of Trypanosoma cruzi: chromosomerearrangements in a single cell-derived clone of the G strain. PLoS One 8,e63738.
Loewith, R., Hall, M.N., 2011. Target of rapamycin (TOR) in nutrient signaling andgrowth control. Genetics 189, 1177–1201.
Marone, R., Cmiljanovic, V., Giese, B., Wymann, M.P., 2008. Targetingphosphoinositide 3-kinase: moving towards therapy. Biochim. Biophys. Acta1784, 159–185.
Mertens, H.D., Callaghan, J.M., Swarbrick, J.D., McConville, M.J., Gooley, P.R., 2007. Ahigh-resolution solution structure of a trypanosomatid FYVE domain. ProteinSci. 16, 2552–2559.
Moreno-Hagelsieb, G., Latimer, K., 2008. Choosing BLAST options for betterdetection of orthologs as reciprocal best hits. Bioinformatics 24, 319–324.
Opperdoes, F.R., Michels, P.A.M., 2007. Horizontal gene transfer in trypanosomatids.Trends Parasitol. 23, 470–476.
Rigden, D.J., Monteiro, A.C., Grossi de Sá, M.F., 2001. The protease inhibitor chagasinof Trypanosoma cruzi adopts an immunoglobulin-type fold and may have arisenby horizontal gene transfer. FEBS Lett. 504, 41–44.
Ronquist, F., Huelsenbeck, J.P., 2003. MrBayes 3: bayesian phylogenetic inferenceunder mixed models. Bioinform. Oxf. Engl. 19, 1572–1574.
Rutherford, A.C., Traer, C., Wassmer, T., Pattni, K., Bujny, M.V., Carlton, J.G.,Stenmark, H., Cullen, P.J., 2006. The mammalian phosphatidylinositol 3-phosphate 5-kinase (PIKfyve) regulates endosome-to-TGN retrogradetransport. J. Cell Sci. 119, 3944–3957.
Santos, M.R., Lorenzi, H., Porcile, P., Carmo, M.S., Schijman, A., Brandao, A., Araya,J.E., Gomes, H.B., Chiurillo, M.A., Ramirez, J.L., Degrave, W.M., Levin, M.J., daSilveira, J.F., 1999. Physical mapping of a 670-kb region of chromosomes XVIand XVII from the human protozoan parasite Trypanosoma cruzi encompassingthe genes for two immunodominant antigens. Genome Res. 9, 1268–1276.
Shertz, C.A., Cardenas, M.E., 2011. Exploiting and subverting Tor signaling in thepathogenesis of fungi, parasites, and viruses. PLoS Pathog. 7, e1002269.
Silva, D.C.F., Silva, R.C., Ferreira, R.C., Briones, M.R.S., 2013. Examining marginalsequence similarities between bacterial type III secretion system componentsand Trypanosoma cruzi surface proteins: horizontal gene transfer or convergentevolution? Front. Genet. 4, 143.
Souza, R.T., Lima, F.M., Barros, R.M., Cortez, D.R., Santos, M.F., Cordero, E.M., Ruiz,J.C., Goldenberg, S., Teixeira, M.M., da Silveira, J.F., 2011. Genome size, karyotypepolymorphism and chromosomal evolution in Trypanosoma cruzi. PLoS One 6,e23042.
Steglich, C., Schaeffer, S.W., 2006. The ornithine decarboxylase gene of Trypanosomabrucei: evidence for horizontal gene transfer from a vertebrate source. Infect.Genet. Evol. 6, 205–219.
Stenmark, H., Aasland, R., Toh, B.H., D’Arrigo, A., 1996. Endosomal localization of theautoantigen EEA1 is mediated by a zinc-binding FYVE finger. J. Biol. Chem. 271,24048–24054.
Sturgill, T.W., Cohen, A., Diefenbacher, M., Trautwein, M., Martin, D.E., Hall, M.N.,2008. TOR1 and TOR2 have distinct locations in live cells. Eukaryot. Cell 7,1819–1830.
Tsang, C.K., Zheng, X.F., 2007. TOR-in(g) the nucleus. Cell Cycle 6, 25–29.Vargas, N., Pedroso, A., Zingales, B., 2004. Chromosomal polymorphism, gene
synteny and genome size in T. cruzi I and T. cruzi II groups. Mol. Biochem.Parasitol. 138, 131–141.
Weatherly, D.B., Boehlke, C., Tarleton, R.L., 2009. Chromosome level assembly of thehybrid Trypanosoma cruzi genome. BMC Genomics 10, 255.
Wedaman, K.P., Reinke, A., Anderson, S., Yates 3rd, J., McCaffery, J.M., Powers, T.,2003. Tor kinases are in distinct membrane-associated protein complexes inSaccharomyces cerevisiae. Mol. Biol. Cell 14, 1204–1220.
Copyright © 2022 FDOKUMEN




















