
Oxytocin Attenuates Amygdala Reactivity to Fear in Generalized Social Anxiety Disorder
Upload
independentCategory
view
0download
0
of November 20, 2015.This information is current as
Bleomycin-Induced Pulmonary FibrosisAttenuates the Development of
-GalactosylceramideαTreatment with
SekizawaE. Hegab, Tohru Sakamoto, Akihiro Nomura and Kiyohisa
AhmedKazuko Shibuya, Masaru Taniguchi, Mie Mochizuki, Toru Kimura, Yukio Ishii, Yuko Morishima, Akira Shibuya,
http://www.jimmunol.org/content/172/9/5782doi: 10.4049/jimmunol.172.9.5782
2004; 172:5782-5789; ;J Immunol
Referenceshttp://www.jimmunol.org/content/172/9/5782.full#ref-list-1
, 19 of which you can access for free at: cites 39 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 20, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Treatment with �-Galactosylceramide Attenuates theDevelopment of Bleomycin-Induced Pulmonary Fibrosis
Toru Kimura,* Yukio Ishii, 1* Yuko Morishima,* Akira Shibuya, †‡§ Kazuko Shibuya,†‡
Masaru Taniguchi,¶� Mie Mochizuki,* Ahmed E. Hegab,* Tohru Sakamoto,* Akihiro Nomura,*and Kiyohisa Sekizawa*
Pulmonary fibrosis is an end-stage disorder for which efficacious therapeutic options are not readily available. Although itspathogenesis is poorly understood, pulmonary fibrosis occurs as a result of various inflammations. NKT cells modulate inflam-mation because of their ability to produce large amounts of cytokines by stimulation with their glycolipid ligand. In the presentstudy, we investigated the effects of�-galactosylceramide (�-GalCer), a selective NKT cell ligand, on the development of bleo-mycin-induced pulmonary fibrosis. Treatment of mice with�-GalCer prolonged their survival under bleomycin administration byattenuating the development of pulmonary fibrosis. The protective effects of�-GalCer were associated with an increase in thepulmonary level of IFN-� and a decrease in the pulmonary level of fibrogenic cytokines such as TGF-� and connective tissuegrowth factor. The initial pulmonary inflammation caused by bleomycin was also attenuated by�-GalCer with the reduction ofthe macrophage inflammatory protein-2 level. The protective effects of�-GalCer were markedly reduced in mice lacking NKTcells or as a result of treatment with anti-IFN-� Ab. These results suggest that�-GalCer suppresses bleomycin-induced acutepulmonary inflammation and thus attenuates the development of pulmonary fibrosis possibly by regulating several cytokinelevels. The Journal of Immunology, 2004, 172: 5782–5789.
P ulmonary fibrosis is an end-stage, lethal lung disorder forwhich efficacious therapeutic options are not readily avail-able. Evaluation of lung tissue from patients with pulmo-
nary fibrosis shows evidence of inflammation, disordering of lungparenchymal cells, and fibrosis. The pathogenesis of pulmonaryfibrosis, therefore, is thought to be characterized by initial inflam-matory responses followed by fibroproliferation and collagen dep-osition. Because fibrosis is an irreversible change, effective ther-apies for this disorder must be given early in its natural history,before the development of extensive lung destruction and fibrosis.
The mechanisms of the fibrotic process are not fully understood;however, the process appears to result from a complex interactionbetween fibroblasts, lung parenchymal cells, and various inflam-matory cells. Recent studies have shown that the specific cytokinephenotype may provide fundamental mechanisms to regulate thefibrotic process. For example, IL-4 and IL-13 enhance the fibroticprocess by augmenting fibroblast growth and collagen production(1–3). Conversely, IFN-� induces antifibrotic effects by directlysuppressing fibroblast activity such as proliferation and collagenproduction (3–5) or indirectly by attenuating the effects of IL-4 andIL-13 (6). Interestingly, these cytokines are prototypic cytokinesparticipating in a type 1 or type 2 immune response.
NKT cells, typically defined as NK1.1���TCR� cells in mice,are a novel lymphoid lineage distinct from T, B, or NK cells in themouse immune system. Several studies have demonstrated thatNKT cells participate in several immune and inflammatory re-sponses because of their remarkable capacity to produce immuno-regulatory cytokines such as IFN-� and IL-4 after stimulation witheither anti-CD3 (7, 8) or their glycolipid ligand (9, 10).
�-Galactosylceramide (�-GalCer),2 a synthetic glycolipid orig-inally isolated from marine sponges, is specifically recognized in aCD1d-restricted manner by V�14� NKT cells. Exogenously ad-ministered �-GalCer induces several immune responses includingantitumor activity (11), granuloma formation (12), and host resis-tance to Toxoplasma gondii and Cryptococcus neoformans infec-tions (13) by the activation of NKT cells to produce a large amountof IFN-�. Moreover, IFN-� is capable of activating NK cells andT cells to produce IFN-� in a direct manner or by producing IL-12from various APCs (14–16). It is therefore likely that �-GalCerinfluences the natural history of the development of pulmonaryfibrosis.
Bleomycin is an antitumor drug often used as an inducing agentin models of pulmonary fibrosis. Intratracheal administration ofbleomycin in mice induces acute pulmonary inflammation fol-lowed by varying degrees of intraalveolar and interstitial fibrosis(17–19). This fibrotic process resembles that in human pulmonaryfibrosis (18, 20). In the present study, therefore, we investigatedthe effects of �-GalCer on the development of pulmonary fibrosisusing this fibrosis model. The effects of �-GalCer were furthercompared between wild-type mice and NKT cell-deficient mice toevaluate the contribution of NKT cells.
Departments of *Respiratory Medicine, and †Immunology, University of Tsukuba,Tsukuba, Japan; ‡Laboratory for Immune Receptor, RIKEN Research Center for Al-lergy and Immunology, Tsukuba, Japan; §Precursory Research for Embryonic Scienceand Technology Office, Japan Science and Technology, Kawaguchi, Japan; ¶Labo-ratory for Immune Regulation, RIKEN Research Center for Allergy and Immunology,Yokohama, Japan; and �Department of Molecular Immunology, Graduate School ofMedicine, Chiba University, Chiba, Japan
Received for publication April 28, 2003. Accepted for publication February 24, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 Address correspondence and reprint requests to Dr. Yukio Ishii, Department ofPulmonary Medicine, University of Tsukuba, Tennoudai 1-1-1, Tsukuba 305-8575,Japan. E-mail address: [email protected]
2 Abbreviations used in this paper: �-GalCer, �-galactosylceramide; BAL, bron-choalveolar lavage; CTGF, connective tissue growth factor; MIP-2, macrophage in-flammatory protein-2.
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

Materials and MethodsMice
Wild-type C57BL/6 mice were obtained from Charles River Breeding Lab-oratories Japan (Kanagawa, Japan). NKT cell-deficient (J�281�/�) micewere established by specific deletion of the J�281 gene segment with ho-mologous recombination and aggregation chimera techniques (9). All miceused in this study were 6–8 wk of age and were maintained in our animalfacilities under specific pathogen-free conditions. The studies adhered tothe National Institutes of Health guidelines for the experimental use ofanimals. All animal studies were approved by the Institutional ReviewBoard.
Reagents
A synthetic form of �-GalCer, KRN7000, was kindly provided by KirinBrewery (Gunma, Japan). The lyophilized form of �-GalCer was diluted in1 ml of distilled water to make a stock solution. This stock solution wasfurther diluted into an appropriate concentration with saline and used forthe experiments. The vehicle control stock solution was also diluted withsaline and used for all experiments.
Treatment of mice
Mice were i.p. injected with �-GalCer (10 �g/100 �l vehicle) or controlvehicle. One day after the treatment with �-GalCer or vehicle, these micewere administered bleomycin (0.5 mg/100 �l saline; Calbiochem, San Di-ego, CA) or saline intratracheally. We preliminarily examined 0.1 mg, 0.5mg, and 1 mg of bleomycin and concluded that 0.5 mg of bleomycin wasthe best for evaluating both the degree of fibrosis and the survival rate.
Histopathologic assessment
The lungs were removed 28 days after bleomycin or saline administration.Following fixation, the lungs were embedded in paraffin. The sections werethen stained with Masson’s trichrome stain. Histopathologic evaluation ofpulmonary fibrosis was performed using a previously described scoringmethod (21). Briefly, the grade of lung fibrosis was scored on a scale of 0to 8 using the following criteria: grade 0, normal lung; grade 1 to 2, min-
imal fibrous thickening of alveolar or bronchiolar wall; grade 3 to 4, mod-erate thickening of walls without obvious damage to lung architecture;grade 5 to 6, increased fibrosis with definite damage to lung structure;
FIGURE 1. �-GalCer prolongs the survival of wild-type mice followingbleomycin administration. Mice were divided into four groups. The controlgroup (F) was administered saline 1 day after treatment with vehicle (n � 20).The Gal group (f) was administered saline 1 day after �-GalCer treatment(n � 20). The Bleo group (E) was administered bleomycin 1 day after treat-ment with vehicle (n � 20). The Gal�Bleo group (�) were administeredbleomycin 1 day after �-GalCer treatment (n � 20). A, Survival of wild-typeC57BL/6 mice. Differences between the Bleo group (E) and the Gal�Bleogroup (�) were significant (p � 0.05). B, Survival of J�281�/� mice of thesame background. There were no significant differences between the Bleo (E)and Gal�Bleo groups (�).
FIGURE 2. �-GalCer attenuates the development of bleomycin-induced pul-monary fibrosis. Mice were divided into four groups as described in Fig. 1. A,Photomicrograph of lung tissues from wild-type mice 28 days after the adminis-tration of bleomycin or saline is shown. Dense fibrosis was observed in the lungsof the Bleo group, whereas the fibrosis was limited in the Gal�Bleo group. B,Photomicrograph of lung tissues from J�281�/� mice 28 days after the adminis-tration of bleomycin or saline is shown. The degree of fibrosis was similar betweenthe Bleo and Gal�Bleo groups. Masson’s trichrome stain with scale bar � 100�m. C, Quantitative assessment of pulmonary fibrosis 28 days after the adminis-tration of bleomycin or saline using a scoring method as previously described (n �4 in each group). D, Lung hydroxyproline contents 28 days after the administrationof bleomycin or saline (n � 6 in each group). �, Differences between the Bleo andGal�Bleo groups were significant in wild-type mice (p � 0.05), but not inJ�281�/� mice.
5783The Journal of Immunology
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

grade 7 to 8; severe distortion of structure and large fibrous areas. After theexamination of 30 randomly chosen regions in each sample at a magnifi-cation of �100, the mean score of all the fields was taken as the fibrosisscore in each sample.
Assessment of collagen synthesis
Collagen synthesis was assessed using a hydroxyproline assay. The micewere anesthetized and the lungs were removed 28 days after bleomycin orsaline administration. Hydroxyproline content was measured as reportedpreviously (22).
Bronchoalveolar lavage (BAL)
Two days after bleomycin or saline administration, the lungs were lavagedwith six sequential aliquots of 1 ml of PBS. The supernatant of the firstBAL was used to analyze albumin concentration by means of its colorreaction with bromocresol green (Sigma-Aldrich, St. Louis, MO) at 630
nm. The remaining pooled BAL was centrifuged and resuspended in PBS.Cells were counted using a hemocytometer and a differential cell count wasperformed by standard light microscopic techniques based on staining withDiff-Quik (American Scientific Products, McGraw Park, IL).
Flow cytometry
After 2 days and 7 days of bleomycin or saline administration, the lungswere removed, minced, and incubated with RPMI 1640 containing 10%FCS and 75 U/ml collagenase (type 1, Sigma-Aldrich) at 37°C for 90 min.The cells were then filtered through 20-�m nylon mesh. The cell suspen-sions were stained with anti-NK1.1, anti-TCR�, anti-CD4, and anti-CD8Abs (BD PharMingen, San Diego, CA). After staining, the cells were an-alyzed by flow cytometry using the FACSCaliber with CellQuest software(BD Biosciences, San Jose, CA).
Intracellular cytokine analysis
IFN-� production in lung lymphocytes was determined by flow cytometricintracellular cytokine analysis as previously described (23). Briefly, cellswere suspended at 106/ml in RPMI 1640 containing 10% FCS, incubatedwith PMA (50 ng/ml; Sigma-Aldrich) and ionomycin (500 ng/ml; Sigma-Aldrich) for 2 h, and then incubated with brefeldin A (10 �g/ml; Sigma-Aldrich) for 2 h at 37°C. Cells were then washed in PBS and fixed with 2%formaldehyde in PBS for 15 min at room temperature. Fixed cells werewashed in PBS supplemented with 0.5% BSA and 0.02% sodium azide(PBS/BSA/azide). For intracellular cytokine detection, cells were perme-abilized with 0.5% saponin (Sigma-Aldrich) in PBS/BSA/azide, stainedwith anti-IFN-�-PE Ab (BD PharMingen), and analyzed by three-colorFACS analysis.
Measurement of cytokines
Concentration of macrophage inflammatory protein (MIP)-2 was deter-mined in the supernatant of the first BAL 2 days after bleomycin or salineadministration by ELISA (BioSource International, Camarillo, CA). Con-centration of IFN-� was determined in the lung homogenates and serum byELISA (Endogen, Cambridge, MA). A concentration of the active form ofTGF-� was also determined by ELISA (R&D Systems, Minneapolis, MN).To obtain the lung homogenates, the lungs were removed and homogenizedusing Polytron homogenizer (KINEMATICA, Lucerne, Switzerland). Af-ter centrifugation for 1 h at 100,000 � g, the supernatants were recovered.
PCR analysis
Seven days after bleomycin or saline administration, total RNA was ex-tracted from the lung tissues, and RT-PCR was performed using an Ac-cessQuick RT-PCR System (Promega, Madison, WI), according to themanufacturer’s instructions. The sequences of the PCR primers were:TGF-�, 5�-GCCCTGGACACCAACTATTGCT-3� and 5�-CCCACGTAGTACACGATGGG-3�; connective tissue growth factor (CTGF), 5�-CTATGCAGCATGGACCTG-3� and 5�-CTCCGTACATCTTCCTGTAC-3�; andGAPDH, 5�-ACCACAGTCCATGCCATCAC-3� and 5�-TCCACCACCCTGTTGCTGTA-3�. Semiquantitative analysis was performed using theNIH Image software program (Bethesda, MD).
Neutralization of IFN-�
To neutralize IFN-�, mAb to IFN-� (IFN-mAb, 1 mg/kg; BD PharMingen)or PBS was injected into wild-type mice 6 h before intratracheal bleomycinor saline administration. Twenty-eight days after bleomycin or saline ad-ministration, the mice were sacrificed and the lungs were removed for theanalysis.
Statistics
Data were expressed as mean � SEM. Comparisons of data among eachexperimental group were performed using ANOVA and Scheffe’s test. Thesurvival curves were analyzed using the log-rank test. Values of p � 0.05were considered to be statistically significant.
Results�-GalCer prolongs survival after bleomycin administration inwild-type mice
We first examined whether treatment with �-GalCer protects miceagainst bleomycin-induced lung toxicity. Wild-type C57BL/6mice were intratracheally administered bleomycin or saline 1 dayafter �-GalCer or vehicle treatment. Fifty percent of the mice in
FIGURE 3. �-GalCer attenuates bleomycin-induced acute pulmonaryinflammation. Mice were divided into four groups as described in Fig. 1. A,The concentration of albumin in bronchoalveolar lavage fluids 2 days afterthe administration of bleomycin or saline (n � 8 in each group). Thenumbers of macrophages (B), lymphocytes (C), and neutrophils (D) inBAL fluids 2 days after the administration of bleomycin or saline (n � 8in each group). Differences between the control and Bleo groups as shownby asterisks were significant (p � 0.05). Differences between the Bleo andGal�Bleo groups as shown by # were significant in wild-type mice (p �0.05), but not in J�281�/� mice.
5784 PROTECTION FROM PULMONARY FIBROSIS BY �-GalCer
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

the vehicle-treated group died by 28 days after bleomycin admin-istration (Fig. 1A). However, 80% of the mice in the �-GalCer-treated groups survived 28 days after bleomycin administration(Fig. 1A). Treatment with �-GalCer significantly prolonged thesurvival after the bleomycin administration in wild-type mice. Noanimals died in the saline-administered control group regardless of�-GalCer treatment (Fig. 1A).
We also conducted the same experiments using J�281�/� micewith C57BL/6 background. Of the mice in the vehicle-treatedgroup, 65% died by 28 days after bleomycin administration (Fig.1B). Mortality after bleomycin administration was not differentbetween wild-type mice and J�281�/� mice in the vehicle-treatedgroups. The protective effect of �-GalCer treatment was not ob-served in J�281�/� mice: 65% of mice in the �-GalCer-treatedgroup died by 28 days after bleomycin administration (Fig. 1B).No mice died in the saline-administered control group in J�281�/�
mice regardless of �-GalCer treatment (Fig. 1B).
�-GalCer attenuates the development of pulmonary fibrosis
We then assessed the protective effects of �-GalCer on the devel-opment of bleomycin-induced pulmonary fibrosis. Fig. 2A showsthe lung histology 28 days after bleomycin or saline administrationin wild-type mice. Masson’s trichrome stain revealed dense fibro-sis with prominent collagen deposition 28 days after bleomycinadministration (Bleo) in the vehicle-treated group (Fig. 2A). Thedegree of pulmonary fibrosis was markedly decreased in the�-GalCer-treated group (Gal�Bleo), compared with the vehicle-treated group (Fig. 2A). No abnormal alveolar architecture wasobserved after saline administration in either the vehicle-treated(control) or �-GalCer-treated (Gal) groups (Fig. 2A).
In J�281�/� mice, �-GalCer treatment did not prevent the de-velopment of pulmonary fibrosis. Dense fibrosis was observed inboth vehicle-treated and �-GalCer-treated (Bleo and Gal�Bleo)groups 28 days after bleomycin administration (Fig. 2B). No ab-normal alveolar architecture was observed after saline administra-tion in either the vehicle-treated (control) or �-GalCer-treated(Gal) groups (Fig. 2B).
We then assessed the degree of pulmonary fibrosis using a scor-ing method. In wild-type mice, the �-GalCer treatment signifi-cantly decreased the scores of fibrotic lesions 28 days after bleo-mycin administration when compared with the vehicle-treatedgroup (Fig. 2C). In J�281�/� mice, however, there was nosignificant difference in the scores between vehicle-treated and�-GalCer-treated groups 28 days after bleomycin administration(Fig. 2C).
FIGURE 4. �-GalCer increases lung IFN-� level. A, The proportion ofthe lung lymphocyte subset in wild-type mice (upper) and J�281�/� mice(lower) 2 days (left) and 7 days (right) after the administration of bleomy-
cin or saline. Mice were divided into four groups as described in Fig. 1.Representative plots in each group are shown, and the proportion given foreach group is the mean � SEM of four experiments. ¶, Significantly dif-ferent from corresponding saline-administered controls (p � 0.05); †, sig-nificantly different from corresponding vehicle-treated controls (p � 0.05).B, Intracellular production of IFN-� in NKT cells (upper), NK cells (mid-dle), and T cells (lower) in lung lymphocytes 2 days (left) and 7 days(right) after the bleomycin administration in both wild-type and J�281�/�
mice. Mice were divided into four groups as described in Fig. 1. C, Theanalysis of the T cell subset in IFN-�-producing T cells in the Gal�Bleogroup of wild-type mice 7 days after the administration of bleomycin. D,The concentrations of IFN-� in lung homogenates (left) and in serum(right) 7 days after the administration of bleomycin or saline (n � 8 in eachgroup). �, Significant difference (p � 0.05) between the control and Galgroups; #, significant difference (p � 0.05) between the Bleo andGal�Bleo groups.
5785The Journal of Immunology
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

We further assessed the degree of pulmonary fibrosis by mea-suring the lung hydroxyproline content. In wild-type mice, �-Gal-Cer treatment significantly decreased the lung hydroxyproline con-tent 28 days after bleomycin administration when compared withthe vehicle-treated group (Fig. 2D). In J�281�/� mice, however,there was no significant difference in the hydroxyproline contentbetween vehicle-treated and �-GalCer-treated groups 28 days afterbleomycin administration (Fig. 2D).
�-GalCer suppresses acute pulmonary inflammation
We next evaluated the effects of �-GalCer on initial pulmonaryinflammation induced by bleomycin. We preliminarily determinedthat acute pulmonary inflammation indicated by BAL albumincontent and neutrophil number peaked at 2 days after the admin-istration of 0.5 mg of bleomycin.
Significant increases in albumin concentration and in the num-ber of neutrophils were observed in BAL fluid 2 days after bleo-mycin administration in both wild-type and J�281�/� mice, com-pared with saline-administered controls (Fig. 3, A and D). In wild-type mice, BAL albumin concentration was significantly reducedby treatment with �-GalCer from the value of 224.6 � 44.9 mg/mlin vehicle-treated controls 2 days after bleomycin administration(Fig. 3A). Treatment with �-GalCer also significantly decreasedthe number of neutrophils in BAL from the value of 5.4 � 0.5million in vehicle-treated controls at that time (Fig. 3D).
In J�281�/� mice, however, there were no significant differ-ences in BAL albumin concentration or neutrophil number be-tween vehicle-treated and �-GalCer-treated groups 2 days afterbleomycin administration (Fig. 3, A and D). Although the numberof macrophages in BAL fluids increased 2 days after bleomycinadministration in both wild-type and J�281�/� mice, treatmentwith �-GalCer did not affect the number of macrophages in eithertype of mice (Fig. 3B). Few lymphocytes were observed at thattime in both types of mice (Fig. 3C).
�-GalCer increases lung IFN-� levels after bleomycinadministration
To clarify the effect of �-GalCer on NKT cell activation in thelung, we examined the proportion of the lymphocyte subset andIFN-� production by the lung lymphocytes 2 days and 7 days afterthe administration of bleomycin or saline, during acute pulmonaryinflammation and prefibrosis. The proportion of NKT cells, iden-tified by NK1.1 and TCR�, in lung lymphocytes was �1% inwild-type control mice on both day 2 and day 7 (Fig. 4A). Al-
though lung NKT cells were increased significantly on day 2 bybleomycin administration, treatment with �-GalCer did not affectthe proportion of lung NKT cells (Fig. 4A). NKT cells were notdetected in the lungs of J�281�/� mice (Fig. 4A). The proportionof T cells in lung lymphocytes was significantly elevated in bothwild-type and J�281�/� mice at day 2 and day 7 by bleomycinadministration (Fig. 4A). Treatment with �-GalCer did not alter theproportion of lung T cells (Fig. 4A). Conversely, the proportion ofNK cells was significantly increased at day 2 by treatment with�-GalCer in wild-type mice (Fig. 4A).
We then evaluated the intracellular production of IFN-� inNKT, NK, and T cells in the lung 2 days and 7 days after theadministration of bleomycin or saline. At day 2, the proportion ofIFN-�–positive NKT cells in total NKT cells was �1% in eachgroup, and the proportion was not different among the groups inwild-type mice (Fig. 4B, upper left). Similarly, the proportions ofIFN-�-producing NK and T cells among the total cells were both�3%, and there was no difference among the groups (Fig. 4B,middle and lower left). The proportion of IFN-�–positive NK andT cells was not different between wild-type mice and J�281�/�
mice at that time (Fig. 4B, left).At day 7, the proportion of IFN-�-producing NKT cells was not
different among the groups in wild-type mice (Fig. 4B, upperright). In contrast, IFN-�–positive T cells were significantly in-creased by �-GalCer treatment at day 7 in wild-type mice whereasno significant increases were observed in J�281�/� mice (Fig. 4B,lower right). The proportion of IFN-�–positive NK cells did notdiffer among the groups at that time (Fig. 4B, middle right).
The T cell subset was then analyzed in IFN-�-producing T cellsfrom �-GalCer-treated wild-type mice 7 days after bleomycin ad-ministration. Among IFN-�-positive T cells, 72.2 � 2.7% wereCD4-positive and 18.5 � 1.6% were CD8-positive (Fig. 4C).
We further assessed the concentration of IFN-� in lung homog-enates 7 days after bleomycin administration. �-GalCer treatmentsignificantly increased the concentration of IFN-� in wild-typemice at that time (Fig. 4D, left). In J�281�/� mice, �-GalCer didnot alter the concentration of IFN-� in lung homogenates (Fig. 4D,left). Serum IFN-� levels were low, and there were no significantdifferences among the wild-type and J�281�/� mice (Fig. 4D,right).
�-GalCer reduces MIP-2 and fibrogenic cytokine levels
To clarify the molecular mechanism of �-GalCer for protectionagainst bleomycin-induced acute pulmonary inflammation, the
FIGURE 5. �-GalCer inhibited pulmonary expressions of MIP-2 and fibrogenic cytokines in wild-type mice. Mice were divided into four groups asdescribed in Fig. 1. A, Concentration of MIP-2 in BAL fluids of wild-type (Wild) or J�281�/� mice 2 days after the administration of bleomycin or saline(n � 4 per group). B and C, Expression of TGF-� (B) and CTGF (C) in the lungs of wild-type (Wild) or J�281�/� mice treated with �-GalCer or vehicle7 day after the administration of bleomycin or saline (n � 4 per group). Expression was normalized to GAPDH expression. D, Concentration of the activeform of TGF-� in lung homogenates 7 days after the administration of bleomycin or saline (n � 4 per group). �, Significant difference between the controland the Bleo groups (p � 0.05); #, difference between Bleo and Gal�Bleo groups was significant in wild-type mice (p � 0.05), but not in J�281�/� mice.
5786 PROTECTION FROM PULMONARY FIBROSIS BY �-GalCer
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
concentration of MIP-2, a potent neutrophil chemoattractant, wasassessed in BAL fluids 2 days after the administration of bleomy-cin or saline. In both wild-type and J�281�/� mice, the concen-tration of MIP-2 was significantly elevated 2 days after bleomycinadministration (Fig. 5A). In wild-type mice, treatment with �-Gal-Cer significantly reduced the MIP-2 concentration after bleomycinadministration (Fig. 5A). However, in J�281�/� mice, treatment with�-GalCer did not reduce the MIP-2 level at that time (Fig. 5A).
To determine the effects of �-GalCer on the expression of fi-brogenic cytokines, we examined the expression of TGF-� andCTGF in the lungs 7 days after the administration of bleomycin orsaline. Expression of both TGF-� and CTGF mRNAs was signif-icantly up-regulated after bleomycin administration in the lungs ofboth wild-type and J�281�/� mice (Fig. 5, B and C). �-GalCertreatment significantly reduced TGF-� and CTGF mRNA expres-sion in the lungs of wild-type mice but not in J�281�/� mice (Fig.5, B and C).
We further assessed the concentration of the active form ofTGF-� in lung homogenate 7 days after the administration of bleo-mycin or saline. As in the expression of TGF-� mRNA, the proteinlevel of the TGF-� active form was significantly elevated afterbleomycin administration in the lungs of both wild-type andJ�281�/� mice (Fig. 5D). �-GalCer treatment significantly re-duced the concentration of the TGF-� active form in the lungs ofwild-type mice but not in J�281�/� mice (Fig. 5D).
Anti-IFN-� Ab abrogates the effects of �-GalCer
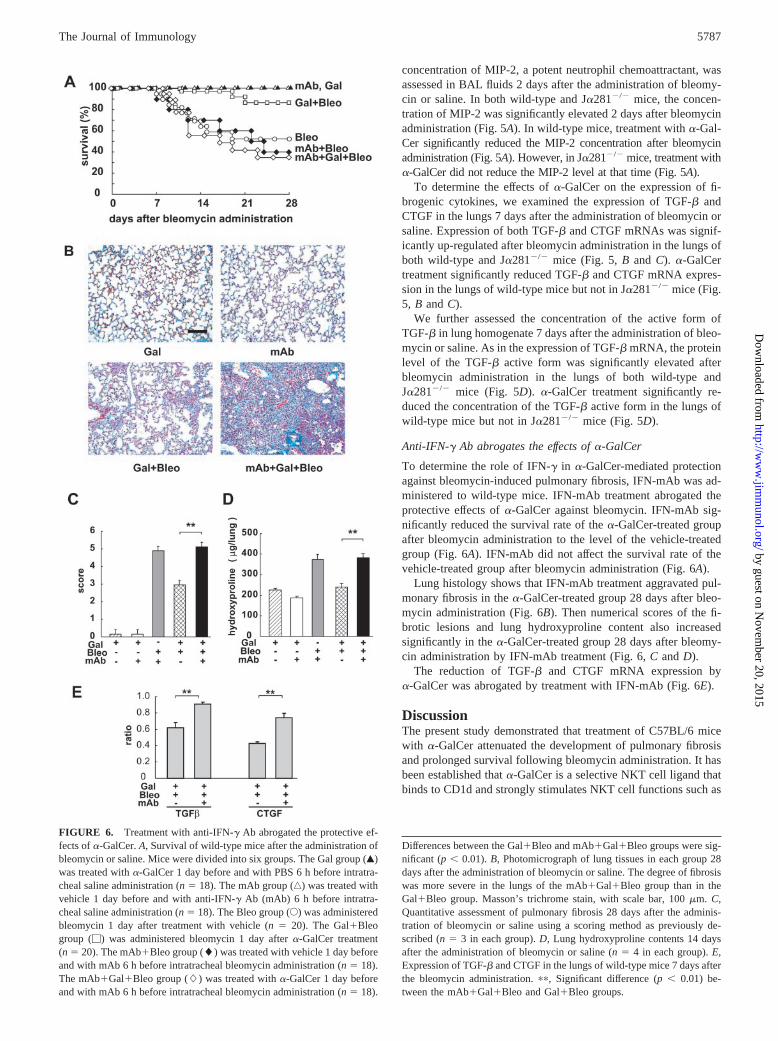
To determine the role of IFN-� in �-GalCer-mediated protectionagainst bleomycin-induced pulmonary fibrosis, IFN-mAb was ad-ministered to wild-type mice. IFN-mAb treatment abrogated theprotective effects of �-GalCer against bleomycin. IFN-mAb sig-nificantly reduced the survival rate of the �-GalCer-treated groupafter bleomycin administration to the level of the vehicle-treatedgroup (Fig. 6A). IFN-mAb did not affect the survival rate of thevehicle-treated group after bleomycin administration (Fig. 6A).
Lung histology shows that IFN-mAb treatment aggravated pul-monary fibrosis in the �-GalCer-treated group 28 days after bleo-mycin administration (Fig. 6B). Then numerical scores of the fi-brotic lesions and lung hydroxyproline content also increasedsignificantly in the �-GalCer-treated group 28 days after bleomy-cin administration by IFN-mAb treatment (Fig. 6, C and D).
The reduction of TGF-� and CTGF mRNA expression by�-GalCer was abrogated by treatment with IFN-mAb (Fig. 6E).
DiscussionThe present study demonstrated that treatment of C57BL/6 micewith �-GalCer attenuated the development of pulmonary fibrosisand prolonged survival following bleomycin administration. It hasbeen established that �-GalCer is a selective NKT cell ligand thatbinds to CD1d and strongly stimulates NKT cell functions such as
FIGURE 6. Treatment with anti-IFN-� Ab abrogated the protective ef-fects of �-GalCer. A, Survival of wild-type mice after the administration ofbleomycin or saline. Mice were divided into six groups. The Gal group (Œ)was treated with �-GalCer 1 day before and with PBS 6 h before intratra-cheal saline administration (n � 18). The mAb group (‚) was treated withvehicle 1 day before and with anti-IFN-� Ab (mAb) 6 h before intratra-cheal saline administration (n � 18). The Bleo group (E) was administeredbleomycin 1 day after treatment with vehicle (n � 20). The Gal�Bleogroup (�) was administered bleomycin 1 day after �-GalCer treatment(n � 20). The mAb�Bleo group (�) was treated with vehicle 1 day beforeand with mAb 6 h before intratracheal bleomycin administration (n � 18).The mAb�Gal�Bleo group (�) was treated with �-GalCer 1 day beforeand with mAb 6 h before intratracheal bleomycin administration (n � 18).
Differences between the Gal�Bleo and mAb�Gal�Bleo groups were sig-nificant (p � 0.01). B, Photomicrograph of lung tissues in each group 28days after the administration of bleomycin or saline. The degree of fibrosiswas more severe in the lungs of the mAb�Gal�Bleo group than in theGal�Bleo group. Masson’s trichrome stain, with scale bar, 100 �m. C,Quantitative assessment of pulmonary fibrosis 28 days after the adminis-tration of bleomycin or saline using a scoring method as previously de-scribed (n � 3 in each group). D, Lung hydroxyproline contents 14 daysafter the administration of bleomycin or saline (n � 4 in each group). E,Expression of TGF-� and CTGF in the lungs of wild-type mice 7 days afterthe bleomycin administration. ��, Significant difference (p � 0.01) be-tween the mAb�Gal�Bleo and Gal�Bleo groups.
5787The Journal of Immunology
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

cytokine production (9, 10). In the present study, the protectiveeffects of �-GalCer were not observed in J�281�/� mice. Thesefindings suggest that activation of NKT cells is required for �-Gal-Cer-mediated protection against the development of pulmonaryfibrosis.
The mechanisms by which �-GalCer attenuates the develop-ment of pulmonary fibrosis are unclear. However, among NKTcell-produced cytokines, IFN-� is known to have antifibrotic ef-fects in experimental animals including bleomycin-induced pul-monary fibrosis model (4, 5). Our study demonstrated that �-Gal-Cer treatment increased the pulmonary IFN-� level in wild-typemice. In addition, neutralization of endogenously synthesizedIFN-� with a specific Ab abrogated the protective effects of �-Gal-Cer in wild-type mice. These findings suggest that the protectiveeffects of �-GalCer against the development of pulmonary fibrosisare, at least in part, mediated by the production of IFN-�. OtherNKT cell-induced host responses, including host resistance to T.gondii and C. neoformans infections (13), granuloma reactioncaused by mycobacterium cell walls (12), and elimination of met-astatic tumors (9, 24) are known to accompany the increase inIFN-� level.
In the analysis of the lymphocyte subset harvested from thelungs, a significant increase in NKT cells was observed 2 days butnot 7 days after bleomycin administration. Treatment with �-Gal-Cer did not affect the number of NKT cells. These results suggestthat NKT cells are recruited into the lung by stimulation with bleo-mycin in the early period of inflammation. However, the propor-tion of IFN-�-producing NKT cells was low and did not differbetween �-GalCer-treated and vehicle-treated mice, suggestingthat NKT cells are not the major cell source of IFN-�. T cells werealso significantly increased in the lungs 2 days and 7 days afterbleomycin administration. On day 7, the proportion of IFN-�-pro-ducing T cells was significantly elevated in wild-type mice as aresult of treatment with �-GalCer, and the elevation paralleled thelung IFN-� level. These results suggest that �-GalCer causes theactivation of T cells to produce IFN-�. NKT cells may participatein the activation of T cells because no increase in the T cell pro-duction of IFN-� was observed in J�281�/� mice. Kawakami etal. (13) demonstrated that a large amount of IFN-� was produced7 days after �-GalCer treatment in mice infected with C. neofor-mans. No such production was noted by this treatment in CD4knockout mice, and CD4� T cells were thus thought to be a majorsource of late-phase IFN-� production induced by �-GalCer. Fur-thermore, in an in vitro experiment, IFN-� synthesis by spleencells obtained from �-GalCer-treated mice on day 7 was com-pletely abrogated both in CD4 knockout mice and NKT-deficientmice (13). These results clearly indicated that CD4�T cells are amajor source of late-phase IFN-� production induced by �-GalCer.Consistently, a majority of IFN-�-producing T cells were shown tobe CD4-positive in the present study.
The contribution of T cells to the pathogenesis of bleomycin-induced pulmonary fibrosis is still controversial. The accumulationof lymphocytes has been reported in bleomycin-induced pneumo-pathy (25, 26). Depletion of lymphocytes by anti-CD4 and anti-CD8 mAbs inhibited pulmonary fibrosis associated with bleomy-cin (27). Bleomycin-induced pulmonary inflammation and fibrosiswere not observed in athymic nude mice (28). These studies sug-gest that T cells participate in the pathophysiology of fibrosis in thebleomycin model. Another study, however, has demonstrated thatbleomycin-induced pulmonary fibrosis occurred in nude mice andSCID mice that lack T cells (29, 30). Although further examina-tions are required to clarify the contribution of T cells to the patho-genesis of fibrosis, the present study first showed that T cells mayact as a negative regulator against the development of pulmonary
fibrosis by producing antifibrotic cytokines through treatment with�-GalCer.
It was interesting that lung NK cells were elevated by the treat-ment with �-GalCer, whereas NKT and T cells were elevated bybleomycin administration in the present study. It has been dem-onstrated that NK cells were the producer of IFN-� after �-GalCertreatment at the early phase of infection with C. neoformansthrough the activation of NKT cells (13). Because the proportionof IFN-�-producing NK cells was low and did not differ betweenthe �-GalCer-treated and vehicle-treated groups, NK cells may notdirectly contribute to the increase in lung IFN-� level in thismodel. The role of NK cells in �-GalCer-mediated protectionagainst pulmonary fibrosis remains to be elucidated.
The administration of bleomycin up-regulates a variety of cy-tokines, including fibrogenic cytokine. Among these cytokines,TGF-� has a role in the development of fibrosis because Absagainst TGF-� markedly inhibit the development of bleomycin-induced pulmonary fibrosis in mice (31). We demonstrated in thisstudy that the treatment with �-GalCer reduced the expression ofTGF-� and CTGF. The reduction of these expressions by �-Gal-Cer was abrogated by treatment with mAb to IFN-�. These find-ings did not contradict the previous finding that treatment withIFN-� markedly reduces the bleomycin-induced increase inTGF-� expression (4). CTGF is a fibrogenic cytokine known to beup-regulated in the lungs of bleomycin-treated mice (32). It wasdemonstrated that CTGF mRNA expression is regulated by TGF-�(33). Therefore, it is reasonable that CTGF expression as well asthat of TGF-� was down-regulated after �-GalCer treatment.Taken together, these findings suggest that the reduction of theexpression of fibrogenic cytokines might be one of the inhibitorymechanisms of �-GalCer against the development of pulmonaryfibrosis.
Although IFN-� shows antifibrotic effects in in vitro and animalstudies, the potential of IFN-� for treating human pulmonary fi-brosis is considered to be limited in view of the recent failures inthe IFN-� treatment clinical trials in the U.S. Pulmonary fibrosis isthought to develop following the initial inflammatory responses inboth human idiopathic pulmonary fibrosis and bleomycin-inducedpulmonary fibrosis. Our observation revealed that �-GalCer sig-nificantly decreased acute pulmonary inflammation without an in-crease in IFN-� production in lung lymphocytes, suggesting that�-GalCer-mediated inhibition of acute pulmonary inflammationoccurs independently with the increase in IFN-�. The mechanismof the anti-inflammatory effects of �-GalCer is unclear; however,we found that the concentration of MIP-2 in BAL fluids, whichwas elevated by bleomycin administration, was significantly re-duced by treatment with �-GalCer. MIP-2 is a CXC chemokineand shows potent chemotactic activity for neutrophils (34). Reg-ulation of neutrophil chemotactic factors is likely to be an addi-tional effect of �-GalCer on the protection against acute pulmonaryinflammation and the subsequent fibrosis. However, NKT cell-me-diated regulation of neutrophilic inflammation is controversial be-cause a recent study demonstrated that monocyte chemoattractantprotein 1-dependent recruitment of V�14-NKT cells induced earlyhost protection against Streptococcus pneumoniae by promotingthe trafficking of neutrophils to the site of infection (35).
We considered that the use of �-GalCer has an advantage overthe systemic administration of anti-fibrotic cytokines such asIFN-� for the treatment of pulmonary fibrosis for the followingreasons: 1) treatment with �-GalCer, even via i.p. injection, in-creased lung IFN-� levels but not the serum level, so that thesystemic side effects of IFN-� can be reduced; 2) effects in additionto the production of IFN-�, such as the regulation of chemokines,are expected to be induced by �-GalCer. However, some problems
5788 PROTECTION FROM PULMONARY FIBROSIS BY �-GalCer
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

have to be solved to use �-GalCer for the treatment of pulmonaryfibrosis. Several studies have demonstrated that �-GalCer inducesnot only IFN-� but also Th2 cytokines such as IL-4 and IL-13under different conditions (36–38). We are currently investigatingthe balance of IFN-� and IL-4 in the lungs using this fibrosismodel with �-GalCer in various amounts and at various adminis-tration schedules to determine the adequate use of �-GalCer for thetreatment of pulmonary fibrosis. The inoculation route of �-GalCeralso should be examined. However, our preliminary examinationshowed that intratracheal administration of �-GalCer did not at-tenuate the development of pulmonary fibrosis by bleomycin (datanot shown).
We demonstrated in the present study that treatment with �-Gal-Cer attenuates the acute pulmonary inflammation and subsequentfibrosis induced by bleomycin. This synthetic glycolipid thereforemay be a candidate agent for the treatment of acute lung injury andpulmonary fibrosis. Furthermore, bleomycin is an antitumor drugcommonly used to treat various types of tumors (39). The mostimportant side effect of bleomycin is pulmonary toxicity, includingpulmonary fibrosis (19). The administration of �-GalCer to suchpatients might be useful for decreasing the side effects ofbleomycin.
References1. Liu, X., T. Kohyama, H. Wang, Y. K. Zhu, F. Q. Wen, H. J. Kim,
D. J. Romberger, and S. I. Rennard. 2002. Th2 cytokine regulation of type Icollagen gel contraction mediated by human lung mesenchymal cells.Am. J. Physiol. Lung Cell Mol. Physiol. 282:L1049.
2. Postlethwaite, A. E., M. A. Holness, H. Katai, and R. Raghow. 1992. Humanfibroblasts synthesize elevated levels of extracellular matrix proteins in responseto interleukin 4. J. Clin. Invest. 90:1479.
3. Sempowski, G. D., S. Derdak, and R. P. Phipps. 1996. Interleukin-4 and inter-feron-� discordantly regulate collagen biosynthesis by functionally distinct lungfibroblast subsets. J. Cell. Physiol. 167:290.
4. Gurujeyalakshmi, G., and S. N. Giri. 1995. Molecular mechanisms of antifibroticeffect of interferon � in bleomycin-mouse model of lung fibrosis: downregulationof TGF-� and procollagen I and III gene expression. Exp. Lung Res. 21:791.
5. Hyde, D. M., T. S. Henderson, S. N. Giri, N. K. Tyler, and M. Y. Stovall. 1988.Effect of murine � interferon on the cellular responses to bleomycin in mice. Exp.Lung Res. 14:687.
6. Saito, A., H. Okazaki, I. Sugawara, K. Yamamoto, and H. Takizawa. 2003. Po-tential action of IL-4 and IL-13 as fibrogenic factors on lung fibroblasts in vitro.Int. Arch. Allergy Immunol. 132:168.
7. Yoshimoto, T., and W. E. Paul. 1994. CD4�, NK1.1� T cells promptly produceinterleukin 4 in response to in vivo challenge with anti-CD3. J. Exp. Med.179:1285.
8. Chen, H., and W. E. Paul. 1997. Cultured NK1.1� CD4� T cells produce largeamounts of IL-4 and IFN-� upon activation by anti-CD3 or CD1. J. Immunol.159:2240.
9. Kawano, T., J. Cui, Y. Koezuka, I. Toura, Y. Kaneko, K. Motoki, H. Ueno,R. Nakagawa, H. Sato, E. Kondo, et al. 1997. CD1d-restricted and TCR-mediatedactivation of V�14 NKT cells by glycosylceramides. Science 278:1626.
10. Hameg, A., I. Apostolou, M. Leite De Moraes, J. M. Gombert, C. Garcia,Y. Koezuka, J. F. Bach, and A. Herbelin. 2000. A subset of NKT cells that lacksthe NK1.1 marker, expresses CD1d molecules, and autopresents the �-galacto-sylceramide antigen. J. Immunol. 165:4917.
11. Smyth, M. J., N. Y. Crowe, Y. Hayakawa, K. Takeda, H. Yagita, andD. I. Godfrey. 2002. NKT cells: conductors of tumor immunity? Curr. Opin.Immunol. 14:165.
12. Apostolou, I., Y. Takahama, C. Belmant, T. Kawano, M. Huerre, G. Marchal,J. Cui, M. Taniguchi, H. Nakauchi, J. J. Fournie, et al. 1999. Murine natural killerT (NKT) cells (correction of natural killer cells) contribute to the granulomatousreaction caused by mycobacterial cell walls. Proc. Natl. Acad. Sci. USA 96:5141.
13. Kawakami, K., Y. Kinjo, S. Yara, Y. Koguchi, K. Uezu, T. Nakayama,M. Taniguchi, and A. Saito. 2001. Activation of V�14� natural killer T cells by�-galactosylceramide results in development of Th1 response and local host re-sistance in mice infected with Cryptococcus neoformans. Infect. Immun. 69:213.
14. Hsieh, C. S., S. E. Macatonia, C. S. Tripp, S. F. Wolf, A. O’Garra, andK. M. Murphy. 1993. Development of TH1 CD4� T cells through IL-12 pro-duced by Listeria-induced macrophages. Science 260:547.
15. Seder, R. A., R. Gazzinelli, A. Sher, and W. E. Paul. 1993. Interleukin 12 actsdirectly on CD4� T cells to enhance priming for interferon � production anddiminishes interleukin 4 inhibition of such priming. Proc. Natl. Acad. Sci. USA90:10188.
16. Guler, M. L., J. D. Gorham, C. S. Hsieh, A. J. Mackey, R. G. Steen,W. F. Dietrich, and K. M. Murphy. 1996. Genetic susceptibility to Leishmania:IL-12 responsiveness in TH1 cell development. Science 271:984.
17. Adamson, I. Y., and D. H. Bowden. 1974. The pathogenesis of bleomycin-in-duced pulmonary fibrosis in mice. Am. J. Pathol. 77:185.
18. Snider, G. L., J. A. Hayes, and A. L. Korthy. 1978. Chronic interstitial pulmonaryfibrosis produced in hamsters by endotracheal bleomycin: pathology and stere-ology. Am. Rev. Respir. Dis. 117:1099.
19. Sleijfer, S. 2001. Bleomycin-induced pneumonitis. Chest 120:617.20. Crystal, R. G., J. D. Fulmer, W. C. Roberts, M. L. Moss, B. R. Line, and
H. Y. Reynolds. 1976. Idiopathic pulmonary fibrosis: clinical, histologic, radio-graphic, physiologic, scintigraphic, cytologic, and biochemical aspects. Ann. In-tern. Med. 85:769.
21. Ashcroft, T., J. M. Simpson, and V. Timbrell. 1988. Simple method of estimatingseverity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 41:467.
22. Woessner, L. F. 1961. The determination of hydroxyproline in tissue and proteinsamples containing small proportions of this amino acid. Arch. Biochem. Biophys.93:440.
23. Murphy, E., K. Shibuya, N. Hosken, P. Openshaw, V. Maino, K. Davis,K. Murphy, and A. O’Garra. 1996. Reversibility of T helper 1 and 2 populationsis lost after long-term stimulation. J. Exp. Med. 183:901.
24. Kawano, T., J. Cui, Y. Koezuka, I. Toura, Y. Kaneko, H. Sato, E. Kondo,M. Harada, H. Koseki, T. Nakayama, et al. 1998. Natural killer-like nonspecifictumor cell lysis mediated by specific ligand-activated V�14 NKT cells. Proc.Natl. Acad. Sci. USA 95:5690.
25. Chandler, D. B., D. M. Hyde, and S. N. Giri. 1983. Morphometric estimates ofinfiltrative cellular changes during the development of bleomycin-induced pul-monary fibrosis in hamsters. Am. J. Pathol. 112:170.
26. Thrall, R. S., and R. W. Barton. 1984. A comparison of lymphocyte populationsin lung tissue and in bronchoalveolar lavage fluid of rats at various times duringthe development of bleomycin-induced pulmonary fibrosis. Am. Rev. Respir. Dis.129:279.
27. Piguet, P. F., M. A. Collart, G. E. Grau, Y. Kapanci, and P. Vassalli. 1989. Tumornecrosis factor/cachectin plays a key role in bleomycin-induced pneumopathyand fibrosis. J. Exp. Med. 170:655.
28. Schrier, D. J., S. H. Phan, and B. M. McGarry. 1983. The effects of the nude(nu/nu) mutation on bleomycin-induced pulmonary fibrosis: a biochemical eval-uation. Am. Rev. Respir. Dis. 127:614.
29. Szapiel, S. V., N. A. Elson, J. D. Fulmer, G. W. Hunninghake, and R. G. Crystal.1979. Bleomycin-induced interstitial pulmonary disease in the nude, athymicmouse. Am. Rev. Respir. Dis. 120:893.
30. Helene, M., V. Lake, Bullock, J. Zhu, H. Hao, D. A. Cohen, and A. M. Kaplan.1999. T cell independence of bleomycin-induced pulmonary fibrosis. J. Leuko-cyte Biol. 65:187.
31. Giri, S. N., D. M. Hyde, and M. A. Hollinger. 1993. Effect of antibody to trans-forming growth factor � on bleomycin induced accumulation of lung collagen inmice. Thorax 48:959.
32. Lasky, J. A., L. A. Ortiz, B. Tonthat, G. W. Hoyle, M. Corti, G. Athas,G. Lungarella, A. Brody, and M. Friedman. 1998. Connective tissue growth fac-tor mRNA expression is upregulated in bleomycin-induced lung fibrosis.Am. J. Physiol. 275:L365.
33. Grotendorst, G. R., H. Okochi, and N. Hayashi. 1996. A novel transforminggrowth factor � response element controls the expression of the connective tissuegrowth factor gene. Cell Growth Differ. 7:469.
34. Driscoll, K. E. 1994. Macrophage inflammatory proteins: biology and role inpulmonary inflammation. Exp. Lung Res. 20:473.
35. Kawakami, K., N. Yamamoto, Y. Kinjo, K. Miyagi, C. Nakasone, K. Uezu,T. Kinjo, T. Nakayama, M. Taniguchi, and A. Saito. 2003. Critical role of V�14�
natural killer T cells in the innate phase of host protection against Streptococcuspneumoniae infection. Eur. J. Immunol. 33:3322.
36. Hong, S., M. T. Wilson, I. Serizawa, L. Wu, N. Singh, O. V. Naidenko, T. Miura,T. Haba, D. C. Scherer, J. Wei, et al. 2001. The natural killer T-cell ligand�-galactosylceramide prevents autoimmune diabetes in non-obese diabetic mice.Nat. Med. 7:1052.
37. Sharif, S., G. A. Arreaza, P. Zucker, Q. S. Mi, J. Sondhi, O. V. Naidenko,M. Kronenberg, Y. Koezuka, T. L. Delovitch, J. M. Gombert, et al. 2001. Acti-vation of natural killer T cells by �-galactosylceramide treatment prevents theonset and recurrence of autoimmune type 1 diabetes. Nat. Med. 7:1057.
38. Singh, N., S. Hong, D. C. Scherer, I. Serizawa, N. Burdin, M. Kronenberg,Y. Koezuka, and L. Van Kaer. 1999. Cutting edge: activation of NK T cells byCD1d and �-galactosylceramide directs conventional T cells to the acquisition ofa Th2 phenotype. J. Immunol. 163:2373.
39. Loehrer, Sr., P. J., D. Johnson, P. Elson, L. H. Einhorn, and D. Trump. 1995.Importance of bleomycin in favorable-prognosis disseminated germ cell tumors:an Eastern Cooperative Oncology Group trial. J. Clin. Oncol. 13:470.
5789The Journal of Immunology
by guest on Novem
ber 20, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















