
Puromycin–rRNA interaction sites at the peptidyl transferase center
Upload
independentCategory
view
0download
0
Transgenic pigs designed to express humanCD59 and H-transferase to avoid humoralxenograft rejection
Costa C, Zhao L, Burton WV, Rosas C, Bondioli KR, Williams BL,Hoagland TA, Dalmasso AP, Fodor W L. Transgenic pigs designed toexpress human CD59 and H-transferase to avoid humoral xenograftrejection. Xenotransplantation 2002; 9: 45–57. # Munksgaard, 2002
Abstract: Research in pig-to-primate xenotransplantation aims to solvethe increasing shortage of organs for human allotransplantation anddevelop new cell- and tissue-based therapies. Progress towards its clinicalapplication has been hampered by the presence of xenoreactive naturalantibodies that bind to the foreign cell surface and activate complement,causing humoral graft rejection. Genetic engineering of donor cells andanimals to express human complement inhibitors such as hCD59significantly prolonged graft survival. Strategies to decrease thedeposition of natural antibodies were also developed. Expression ofhuman a1,2-fucosyltransferase (H transferase, HT) in pigs modifies thecell-surface carbohydrate phenotype resulting in reduced Gala1,3-Galexpression and decreased antibody binding. We have developedtransgenic pigs that coexpress hCD59 and HT in various cells and tissuesto address both natural antibody binding and complement activation.Functional studies with peripheral blood mononuclear cells and aorticendothelial cells isolated from the double transgenic pigs showed thatcoexpression of hCD59 and HT markedly increased their resistance tohuman serum-mediated lysis. This resistance was greater than with cellstransgenic for either hCD59 or HT alone. Moreover, transgeneexpression was enhanced and protection maintained in pig endothelialcells that were exposed for 24 h to pro-inflammatory cytokines. Thesestudies suggest that engineering donor pigs to express multiple moleculesthat address different humoral components of xenograft rejectionrepresents an important step toward enhancing xenograft survival andimproving the prospect of clinical xenotransplantation.
Cristina Costa,1 Lisa Zhao,1
Willis V. Burton,1 Cristina Rosas,1
Kenneth R. Bondioli,1
Barry L. Williams,1
Thomas A. Hoagland,2
Agustin P. Dalmasso3 andWilliam L. Fodor11Department of Molecular Sciences, AlexionPharmaceuticals Inc, Cheshire, CT, USA,2Department of Animal Sciences, University ofConnecticut, Storrs, CT, USA, and 3Department ofSurgery, University of Minnesota, Minneapolis,MN, USA
Key words: hCD59 – H-transferase – humoral
xenograft rejection – transgenic pigs –
xenotransplantation
Address reprint request to W. L. Fodor, Department
of Molecular Sciences, Alexion Pharmaceuticals Inc,
352 Knotter Drive, Cheshire, CT 06410, USA
(E-mail: [email protected])
Received 12 January 2000;
Accepted 27 March 2001
Introduction
The shortage of human organs for allotransplan-tation has generated interest in using xenotrans-plantation as an alternative approach for patientswith end stage organ failure [1–4]. Moreover,progress in tissue engineering and cell transplan-tation has stimulated the development of newcell- and tissue-based therapies using xenogeneicdonors [5]. Several xenogeneic cell-based thera-pies are currently being tested clinically fordiseases caused by cellular loss or dysfunctionsuch as Parkinson’s disease and Huntington’sdisease [6] or fulminant hepatic failure [7]. The pighas received considerable attention as a source oftissue for xenotransplantation. It exhibits anato-
mical and physiological similarities to humanbeings and it can be genetically engineered toproduce immune-modulating proteins [1–4].Furthermore, transgenic pigs can be propagatedin pathogen-free conditions following protocolsof cesarean derivation. However, several immunebarriers need to be addressed in order to bring pigcells, tissues and organs to clinical use. Manyporcine cell types transplanted into xenogeneicanimal models are rejected by both humoral andcellular-mediated mechanisms [8,9]. Unmodifiedpig organs grafted into primates also suffer avigorous humoral immune attack with massivedeposition of xenoreactive natural antibodies
Xenotransplantation 2002: 9: 45–57
Printed in UK. All rights reserved
Copyright # Munksgaard 2002
XENOTRANSPLANTATIONISSN 0908-665X
45

(XNA) and complement that leads to graft failurewithin minutes to hours [1–4,10]. This rapidrejection process is described as hyperacuterejection (HAR). Several approaches, mainlyfocused on complement inhibition, have beendeveloped to abrogate HAR [1–4]. However, evenin the presence of complement inhibitors andheavy immunosuppression xenografts are rejectedin a period of days to weeks due to delayedxenograft rejection (DXR) [4,11–13]. DXR ismainly constituted by a strong and early antibodyresponse, further complement activation, type IIendothelial activation and acute cellular infiltra-tion [1,11–14]. DXR is now considered the majorhurdle to achieve successful xenotransplantation.Intermittent plasmapheresis treatment may beused to remove XNA, but is not practical forchronic antibody removal and is detrimental tothe recipient [13,14].A general strategy that modifies the donor cell
and organ through transgenic engineering has agreater potential for effectively inhibiting HARand DXR with reduced risk to the patient.Consistent with this idea, we previously generatedtransgenic pigs expressing human CD59 [15].Porcine cells expressing hCD59 were significantlyresistant to human serum cytolysis compared withnon-transgenic control cells [15]. Moreover, kid-neys, hearts and lungs from hCD59-expressingpigs functioned longer than control organs in anex vivo human blood perfusion model [16,17].Other groups obtained similar results with trans-genic animals expressing human complement-regulatory proteins [18–23]. However, theseapproaches did not accomplish complete protec-tion from antibody-mediated rejection [16–23].We therefore pursued the generation of transgenicpigs as donors of cells and organs that express ahuman complement inhibitor and show reducedhuman serum antibody binding.The major xenoepitope recognized by XNA is
the carbohydrate epitope Gala1,3-Gal, which issynthesized by a1,3-galactosyltransferase (a1,3-GT) [24,25]. We developed, with others, anapproach that utilizes the expression of Htransferase to down-regulate Gala1,3-Gal antigenexpression. HT was shown to efficiently competewith a1,3-GT for the same acceptor substrate, N-acetyl lactosamine, impeding the transfer of theterminal galactose residue that gives rise to theGala1,3-Gal antigen [26]. Furthermore, HTgenerates fucosylated residues (H-antigen, the Oblood group antigen) that are universally toler-ated. We, as others, recently applied this strategyto generate transgenic mice and pigs [27–31]. Thereduction in the Gala1,3-Gal epitope in HT-engineered cells resulted in decreased human
antibody reactivity and serum-mediated cytolysis[27–31]. Moreover, transgenic mouse hearts ex-pressing human HT exhibited enhanced survivalwhen challenged with human serum [30].Expression of human complement inhibitors
together with HT or the a1,3-GT knockout (GalKO) in mice has shown an additive effect inprotecting cells and organs against the challengewith human serum [32,33]. Here we present for thefirst time the generation of double transgenic pigsthat coexpress HT and a complement inhibitor inmultiple cells and tissues. The combined expres-sion of HT and hCD59 in peripheral bloodmono-nuclear cells (PBMC) and aortic endothelial cells(PAEC) isolated from the transgenic pigs led tothe highest protection from human serum-medi-ated lysis when compared with controls and singletransgenic cells. Moreover, the double transgeniccells maintained their resistance to XNA andcomplement after a 24-h treatment with porcinepro-inflammatory cytokines. Thus, the combina-torial strategy of HT expression and inhibition ofhuman complement may be critical to preventhumoral rejection of porcine cells and organs.
Materials and methods
Generation and analysis of transgenic pigs
Transgenic pigs expressing either hCD59 or HTwere previously generated [15,31]. Artificialinsemination was employed in hCD59xHT inter-crosses in order to obtain double transgenic off-spring. The animals born were tested for thepresence of the transgenes by PCR and SouthernBlot [34] analysis of genomic DNA followingstandard procedures. F0 to F4 heterozygoustransgenic pigs, as well as control littermates,were used for subsequent experiments.
RNA isolation and RT-PCR analysis
Total RNA was prepared using RNA IsolatorTM
(Genosys Biotechnologies, Inc., The Woodlands,TX, USA) as described by the manufacturer.Expression of the transgenes was evaluated byRT-PCR. First strand cDNA was prepared byreverse transcription using avian myeloblastosisvirus reverse transcriptase (Seikagaku America,Inc, Rockville, MD, USA) and used for polymer-ase chain reaction utilizing Perkin Elmer reagents(Perkin-Elmer, Norwalk, CT, USA). Analysis ofhCD59 expressionwas performedwith the follow-ing primers: (1) 5’-CAGGTTCTGTGGACAAT-CAC and (2) 5’-TCCAGGCTGCCAGAAATG,which are complementary to hCD59 sequencessituated 5’ to the initiation codon and 3’ of thegene, respectively. The expected product is
Costa et al.
46

415 bp. The primers used to detect expression ofHT driven by H2Kb promoter were (3) 5’-GGACAGGAGGCTACACCGTGG and (4) 5’-GGTGACGAAATACCTCAGCG, which corre-spond, respectively, to an internal oligonucleotidein HT and a primer hybridizing to exon 2 of theH2Kb gene 3’ to the HT cDNA. The size of theexpected PCR product from this set of primers is0.4 Kb. To determine expression driven by CMVpromoter, we used primer [5] 5’-ATGTCGGA-GGAGTACGCGG, an HT internal primer, andthe reverse primer (6) 5’-TAAGCTGCAATA-AACAAGTTC, located 5’ to the polyadenylationsite. Positive PCR reactions using these primersgenerate a 717-bp fragment. The b-actin primers(7) 5’-CCAACTGGGACGACATGGAG and (8)5’-AGGTCCAGACGCAGGATGGC were usedas a positive control for each tissue RNA, andproduce a 300-bp PCR product expected from acorrectly processed b-actin mRNA.
Cell isolation and culture
PBMC from transgenic and negative littermatecontrol pigs were purified from whole blood byFicoll gradient centrifugation (Pharmacia Bio-tech, Uppsala, Sweden) and one step ACK lysis(Biofluids, Rockville, MD, USA). Primary cul-tures of PAEC were obtained by scrapping theaorta with a scalpel and culturing in DMEM 10%FCS. To induce endothelial cell activation weobtained porcine cytokine-conditioned media(CM), and cultured PAEC for 24 h with 60%CM. Briefly, CM was derived from culturingporcine PBMC with phytohemagglutinin (PHA,5 mg/ml) for 48 h. Supernatants were subse-quently collected, treated with 10 mM methyla-mannoside and filter-sterilized.
Flow cytometric and immunofluorescence analysis
Direct fluorescence of cell-surface carbohydrateepitopes was performed with fluorescein isothio-cyanate (FITC)-conjugated lectins: IB4 lectinisolated from Griffonia simplicifolia (EY Labor-atories, Inc. San Mateo, CA, USA) that detectsGala-1,3-Gal [35] and UEAI lectin isolated fromUlex europaeus (EY Laboratories) that detects H-substance [36]. Indirect immunofluorescence ofhCD59 was performed with the specific poly-clonal anti-sera ALP3 on PBMC and the specificmousemonoclonal antibody BRA10G (BiodesignInternational, Kennebunk, ME, USA) on PAEC.SLA class I was detected with a murine mono-clonal antibody PT85A (VMRD, Inc., Pullman,WA, USA). Goat anti-rabbit IgG and goat anti-mouse IgG, IgA and IgM (Zymed Laboratories,Inc. San Francisco, CA, USA) FITC-conjugated
anti-sera were used to detect specific antibodybinding. Cell surface expression was then mea-sured by flow cytometry on a Becton DickinsonFACSort.Immunofluorescence was performed on fresh
frozen tissue samples embedded in Tissue-TekOCT compound (Sakura FinetekUSA, Torrance,CA, USA). Sections (5–8 mm) were stained withBRA10G (10 mg/ml) (Biodesign International)and UEAI-FITC (1 : 100 dilution) (EY Labor-atories Inc.).
Human serum-mediated cytolysis assay
Two sets of assays were performed. (1) IsolatedPBMC from control and transgenic pigs werewashed with HBSS 1%BSA and exposed toCalcein AM (Molecular Probes Inc) (10 mm) for20 min. Cells were subsequently incubated at37 uC for 30 minwith increasing concentrations ofhuman serum (from 5 to 100%). Percentage of cellsurvival (calcein loaded cells) and cell death (cellswith reduced fluorescence) was determined byflow cytometry. Relative cell survival and celldeath was determined by normalizing the celldeath of control cells at 100% serum to be 100%.Non-serum-mediated cell death was approxi-mately 5% in all assays. (2) PAEC were trypsin-ized, washed twice with HBSS 1%BSA andexposed to increasing concentrations of humanserum for 2 h at 37 uC in flat-bottom 96-welldishes. Cells were subsequently loaded with cal-cein as described, washed and lyzed with 1%SDS.Cell survival was calculated as the averageamount of fluorescence in duplicate samples,relative to the no-serum-added condition, de-tected with a cytofluor (Perspective Biosystems,Framingham, MA, USA).
Natural antibody and complement deposition assay
Anti-Gal a-1,3-Gal antibody reactivity was de-tected on PAEC using specific purified humanantibodies. Briefly, anti-Gal a-1,3-Gal antibodieswere purified from human serum by columnchromatography utilizing 142a and 142b resins(Glycorex, Lund, Sweden). PAECwere incubatedat 4 uC with the purified antibody (10 mg/ml) for30 min. Goat anti-human IgM and IgG (ZymedLaboratories, Inc. San Francisco, CA, USA)FITC-conjugated anti-sera were used to detectspecific reactivity by flow cytometry.Deposition of the complement components C3
and C4 was evaluated by incubating PAEC withincreasing concentrations of C8 depleted serum(Quidel, San Diego, CA, USA) in HBSS for30 min at 37 uC. Cells were thoroughly washedand stained with FITC-labeled anti-human C3
Transgenic pigs expressing hCD59 and HT
47

and C4 specific antibodies (ICN, Aurora, OH,USA). Mean fluorescence intensity was deter-mined by flow cytometry.
Statistical analysis
All values are expressed as the means ¡ SE.Statistical analysis was carried out using theStudent-Newmann-Keuls test. Differences wereconsidered statistically significant at Pj0.05.
Results
Generation and characterization of hCD59xHT doubletransgenic pigs
Transgenic pigs expressing either hCD59 or H-transferase were previously obtained in ourlaboratory [15,31]. Transgene expression in thetwo HT transgenic lines utilized was designed tobe both constitutive and induced by cytokines bythe use of two different promoters, H2Kb andCMV [31]. In order to produce double transgenicpigs, AT20 and AT21 HT founders were matedto transgenic animals from the hCD59 line.Southern Blot analysis of genomic DNA wasused to assess the presence of the transgenes in thegenome of hCD59xHT offspring (Table 1). Con-sistent with the pattern observed for the HTfounder pigs [31], offspring from HTAT20 line hadabout 10 copies of H2Kb-HT gene and 10 copiesof CMV-HT, while animals from HTAT21 linecontained one copy of each transgene (Table 1). Asingle line of H2Kb-hCD59 transgenic pigs wasderived from founder 153–2 [15]. Upon expand-ing the line, we have observed segregation ofcopies that produces high copy-number trans-genic animals (15–20 copies) and animals withonly one copy of the transgene per haploidgenome. The analysis of piglets from hCD59F23HTAT20 F0 mating determined that a doubletransgenic pig contained a high number of copiesof both HT and hCD59 transgenes. Three out ofthe six transgenic pigs born were also positive forboth sets of HT genes, but had only one copy ofhCD59. Controlled breeding between HTAT21 F1
and hCD59 F2–3 also generated double transgenic
pigs with either one copy or the high-copy numberof the hCD59 transgene (Table 1).Expression of hCD59 andHTwas confirmed by
flow cytometric analysis of PBMC isolated fromsingle and double transgenic pigs (Fig. 1). Controlpig cells showed only background staining for H-
Table 1. Combinations generated of hCD59xHT double transgenic pigsa
CMV-HT H2Kb-HT H2Kb-hCD59
hCD59xHTAT20 10 copies 10 copies 15–20 copies
hCD59lowxHTAT20 10 copies 10 copies 1 copy
hCD59xHTAT21 1 copy 1 copy 15–20 copies
hCD59lowxHTAT21 1 copy 1 copy 1 copy
aValues represent number of copies of each transgene per haploid genome.
Log fluorescence intensity
Rel
ativ
e ce
ll nu
mbe
r
Fig. 1. Flow cytometry analysis of PBMC isolated fromcontrol and transgenic pigs with various levels of HT andhCD59 transgene expression. The following pigs were ana-lyzed: non-transgenic control, AT20 founder (HTAT20), F1and F2 transgenic pigs from HTAT21 line (HTAT21), F3 andF4 transgenic pigs from hCD59 line with high (hCD59) andlow (hCD59low) expression of the transgene and all thehCD59xHT combinations. Unstained cells are shown on theleft of each panel. Staining with UEA-I lectin (thick line) orGS-IB4 lectin (dotted line) was performed to detect on the cellsurface H- and Gala-1,3-Gal epitopes, respectively. Express-ion of hCD59 was assessed with the anti-hCD59 antibodyALP3 (thin line).
Costa et al.
48
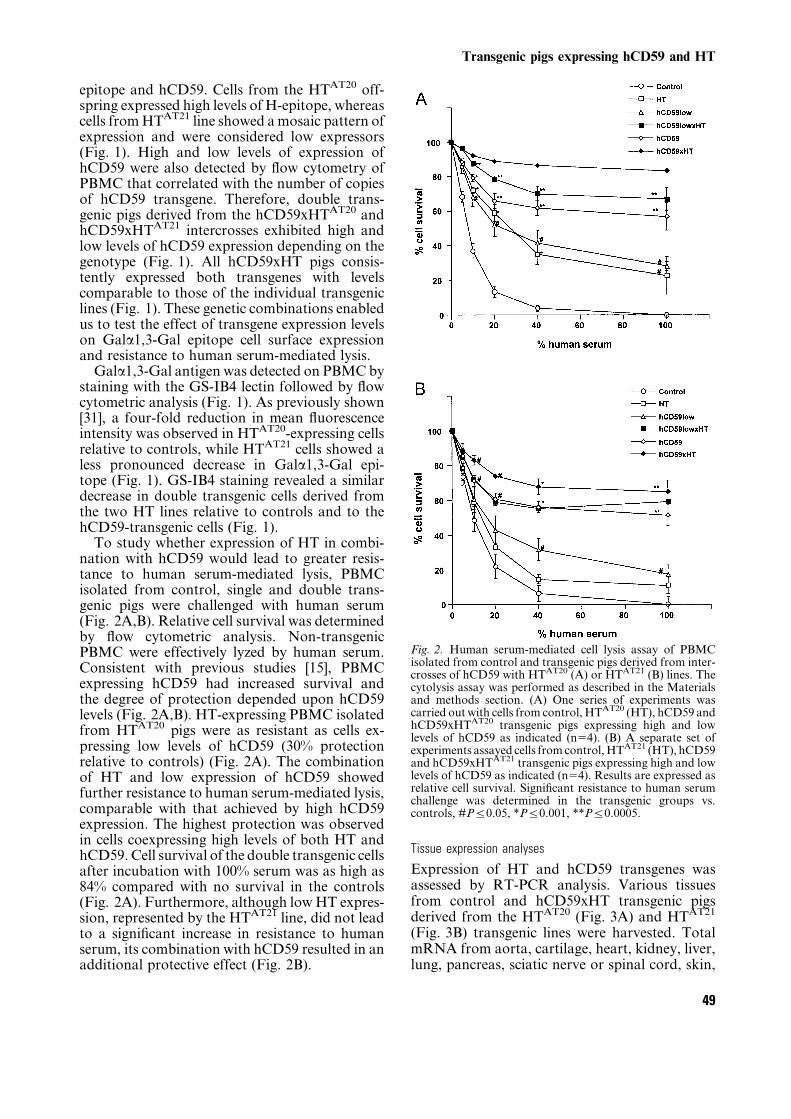
epitope and hCD59. Cells from the HTAT20 off-spring expressed high levels of H-epitope, whereascells fromHTAT21 line showed amosaic pattern ofexpression and were considered low expressors(Fig. 1). High and low levels of expression ofhCD59 were also detected by flow cytometry ofPBMC that correlated with the number of copiesof hCD59 transgene. Therefore, double trans-genic pigs derived from the hCD59xHTAT20 andhCD59xHTAT21 intercrosses exhibited high andlow levels of hCD59 expression depending on thegenotype (Fig. 1). All hCD59xHT pigs consis-tently expressed both transgenes with levelscomparable to those of the individual transgeniclines (Fig. 1). These genetic combinations enabledus to test the effect of transgene expression levelson Gala1,3-Gal epitope cell surface expressionand resistance to human serum-mediated lysis.Gala1,3-Gal antigen was detected on PBMCby
staining with the GS-IB4 lectin followed by flowcytometric analysis (Fig. 1). As previously shown[31], a four-fold reduction in mean fluorescenceintensity was observed in HTAT20-expressing cellsrelative to controls, while HTAT21 cells showed aless pronounced decrease in Gala1,3-Gal epi-tope (Fig. 1). GS-IB4 staining revealed a similardecrease in double transgenic cells derived fromthe two HT lines relative to controls and to thehCD59-transgenic cells (Fig. 1).To study whether expression of HT in combi-
nation with hCD59 would lead to greater resis-tance to human serum-mediated lysis, PBMCisolated from control, single and double trans-genic pigs were challenged with human serum(Fig. 2A,B). Relative cell survival was determinedby flow cytometric analysis. Non-transgenicPBMC were effectively lyzed by human serum.Consistent with previous studies [15], PBMCexpressing hCD59 had increased survival andthe degree of protection depended upon hCD59levels (Fig. 2A,B). HT-expressing PBMC isolatedfrom HTAT20 pigs were as resistant as cells ex-pressing low levels of hCD59 (30% protectionrelative to controls) (Fig. 2A). The combinationof HT and low expression of hCD59 showedfurther resistance to human serum-mediated lysis,comparable with that achieved by high hCD59expression. The highest protection was observedin cells coexpressing high levels of both HT andhCD59. Cell survival of the double transgenic cellsafter incubation with 100% serum was as high as84% compared with no survival in the controls(Fig. 2A). Furthermore, although lowHT expres-sion, represented by the HTAT21 line, did not leadto a significant increase in resistance to humanserum, its combination with hCD59 resulted in anadditional protective effect (Fig. 2B).
Tissue expression analyses
Expression of HT and hCD59 transgenes wasassessed by RT-PCR analysis. Various tissuesfrom control and hCD59xHT transgenic pigsderived from the HTAT20 (Fig. 3A) and HTAT21
(Fig. 3B) transgenic lines were harvested. TotalmRNA from aorta, cartilage, heart, kidney, liver,lung, pancreas, sciatic nerve or spinal cord, skin,
Fig. 2. Human serum-mediated cell lysis assay of PBMCisolated from control and transgenic pigs derived from inter-crosses of hCD59 with HTAT20 (A) or HTAT21 (B) lines. Thecytolysis assay was performed as described in the Materialsand methods section. (A) One series of experiments wascarried out with cells from control, HTAT20 (HT), hCD59 andhCD59xHTAT20 transgenic pigs expressing high and lowlevels of hCD59 as indicated (n54). (B) A separate set ofexperiments assayed cells fromcontrol,HTAT21 (HT), hCD59and hCD59xHTAT21 transgenic pigs expressing high and lowlevels of hCD59 as indicated (n54). Results are expressed asrelative cell survival. Significant resistance to human serumchallenge was determined in the transgenic groups vs.controls, #Pj0.05, *Pj0.001, **Pj0.0005.
Transgenic pigs expressing hCD59 and HT
49

lymph node and spleen was prepared and used tosynthesize first strand cDNA. The RT-PCRanalysis revealed a H2Kb-driven hCD59-specificDNA fragment in all tissues tested from thetransgenic pigs (Fig. 3A,B). HT expression direc-ted by CMV promoter was detected in most RNAsamples isolated from the hCD59xHTAT20 andhCD59xHTAT21 double transgenic pigs. How-ever, expression of HT driven by H2Kb promoterwas confined to a few tissues (Fig. 3A,B). Nospecific PCR products were observed in any of the
samples from the non-transgenic littermate.Endogenously-expressed b-actin was used as acontrol to confirm the integrity of all samplestested by PCR (Fig. 3A,B). The poor quality ofthe RNA isolated from pancreas may account forthe faint or lack of signal in this tissue sample(Fig. 3A,B).Histologic analysis was performed with hearts
and kidneys from two controls and two doubletransgenic pigs from the HTAT21 line. Expressionof hCD59 was confirmed in all vascular structures
Fig. 3. RT-PCR analyses of hCD59 (i) and HT (ii,iii) expression in multiple tissues of hCD59xHT transgenic pigs. (A) Tissueexpression analysis of a hCD59xHTAT20 double transgenic pig (1, 1Kb ladder marker; 2, aorta; 3, cartilage; 4, kidney; 5, liver; 6,lung; 7, pancreas; 8, spinal cord; 9, spleen; 10, control RNA (+); 11, controlRNA (–) and 12, water control). (B) Tissue expressionanalysis of ahCD59xHTAT21double transgenicpig (1, 1Kb laddermarker; 2, aorta; 3, cartilage; 4, heart; 5, kidney; 6, liver; 7, lung;8, pancreas; 9, sciatic nerve; 10, skin; 11, spleen; 12, control RNA (+); 13, control RNA (–) and 14, water control). (A,B) Panels: i.hCD59 expression, driven byH2Kb promoter. ii. HT expression directed by CMV promoter. iii. HT expression driven byH2Kb
promoter. iv. All samples were tested for b-actin mRNA.
Costa et al.
50

Fig. 4. Immunofluorescence analysis in tissue sections from control (left panels) and transgenic pigs (right panels). Staining withanti-hCD59 BRA10G antibody (A,B) and UEA-I lectin (C,D) is shown on heart tissue sections from a control (A,C) and ahCD59xHTAT21 double transgenic pig (B,D). Staining with BRA10G (E,F) andUEA-I lectin (G,H) is shown on kidney sectionsfromacontrol (E,G)andahCD59xHTAT21double transgenicpig (F,H).Results are representativeof twodifferent transgenic andtwo controls examined. Original magnification, X50 (A–H).
Transgenic pigs expressing hCD59 and HT
51

of the ventricular myocardium and the kidneycortex utilizing the specific antibody BRA10G(Fig. 4B,F). No staining was detected in thecontrol tissues (Fig. 4A,E). To detect expressionof H-epitope, the tissue sections were incubatedwith a FITC-conjugated anti-H lectin, UEA-I(Fig. 4C,D,G,H). No staining was detected in the
control heart sections (Fig. 4C), while bothcardiomyocytes and vessels stained for H-epitopein the double transgenic samples (Fig. 4D). In thekidney, H-antigen was strongly expressed in distaltubules of the control samples due to theendogenous HT activity [37], but not in vessels(Fig. 4G). By contrast, arteries, arterioles, veins
Fig. 5. Natural antibody and complement deposition assays by flow cytometry of untreated or CM-treated PAEC isolated fromcontrol, HTAT21, hCD59 and hCD59xHTAT21 transgenic pigs. Anti-Gala-1,3-Gal IgM (A) and IgG (B) reactivity was detectedwith purified specific antibodies. Results are the mean¡ SE of six experiments. C4 (C) and C3 (D) complement deposition wasdetermined as described in the Material and methods section. Results are the mean ¡SE of four experiments. Statisticaldifferences were observed when HTAT21 and hCD59xHTAT21 transgenic PAEC were compared with controls, *P,0.05.
Table 2. Flow cytometric analysis of control and transgenic PAECa
UEA-Ib GS-IB4b BRA10Gb PT85Ac
Control 10.5¡2.9 568.7¡88 0.5¡0.3 100.3¡17.4
Control-CM 15.8¡3.5 713.4¡95 0.7¡0.2 226.4¡37.7*
HTAT21 1113.0¡184** 367.8¡54.7 1.3¡0.4 107.1¡21.4
HTAT21-CM 1525.0¡268** 365.5¡48.5* 0.4¡0.2 265.5¡36.4*
hCD59 18.3¡6.2 824.0¡166 10.6¡1.8** 130.3¡23.2
hCD59-CM 19.0¡5.5 1099.0¡225 42.2¡9** 270.0¡7.40*
hCD59xHTAT21 1018.2¡175** 390.9¡61.2 9.7¡1.3** 110.4¡16.5
hCD59xHTAT21-CM 1183.8¡190** 358.2¡48.7* 28.7¡4.3** 225.4¡30.3*
aAll values are expressed as the mean¡SE of the mean fluorescence intensity (n58). bSignificant differences were detected between transgenic and control cells, *P j 0.01,**Pj0.0005. No differences were observed when comparing HTAT21 and hCD59xHTAT21 cells. cSignificant differences were detected between CM-treated and untreated cells.
Costa et al.
52

and glomerular capillaries stained positive in thetransgenic pig kidney (Fig. 4H).
Functional analyses in endothelial cells
To assess the efficacy of our combinatorial geneticapproach in endothelial cells, we utilized an invitro system based on primary cultured aorticendothelial cells treated or untreated with porcinecytokine-conditioned media (CM). The use ofCM had the purpose of simulating a pro-inflam-matory situation that could induce endothelial cellactivation and thereby mimic one component ofDXR. Primary PAEC cultures from control,HTAT21, hCD59 and hCD59xHTAT21 doubletransgenic pigs were analyzed by flow cytometryto assess transgene expression (Table 2). Express-ion of HT and hCD59 was confirmed in eachof the primary transgenic cell lines. Comparablelevels of hCD59 expression were achieved inhCD59 single and hCD59xHT double transgenicPAEC (Table 2). Gala1,3-Gal epitope expressionwas also determined by staining with GS-IB4lectin (Table 2). A 30–50% reduction in Gala1,3-Gal antigen levels was determined in PAECderived from HTAT21 transgenic pigs relative tocontrols by comparing the mean fluorescenceintensity in all experiments. The hCD59xHTAT21
double transgenic cells showed similar profiles(Table 2).The regulation of the expression of HT and
hCD59 transgenes under pro-inflammatory con-ditions was studied by treating the isolated PAECwith 60% CM for 24 h. Treated and untreatedcells were simultaneously isolated and the expres-sion of hCD59, H- and Gala1,3-Gal epitopes wasassessed by flow cytometry.Up-regulation of SLAclass I (160–200%) was utilized as control for thebiological activity of the CM (Table 2). Thecytokine-mediated inducibility of the H2Kb
promoter directing expression of hCD59 wasconfirmed as hCD59 expression was three- tofour-fold higher in CM-treated cells (Table 2). Anelevation in the levels of H-epitope was alsodemonstrated in the single HT and doubletransgenic cells that probably resulted from theH2Kb-HT regulated expression (Table 2). Eventhough, induced expression of the CMV-HTtransgene cannot be ruled out. While the controland hCD59 transgenic cells showed increasedexpression of Gala1,3-Gal epitope after treatmentwith CM (1.3-fold), the HT and hCD59xHTtransgenic cells maintained the same levels ofexpression as the untreated cells (Table 2). Theseresults indicated that increased H-epitope expres-sion (1.2–1.4-fold) was sufficient to counteract the
Fig. 6. Human serum-mediated cell lysis assay of untreated(A) or CM-treated (B) PAEC isolated from control andtransgenic pigs. The cytolysis assay was performed asdescribed in the Material and methods section. We presenta set of experiments representative of three series that wereperformed with different isolated cells. This set includedprimary cultured PAEC from a control pig (Control), an F1HTAT21 transgenic pig (HT), two high-copy number hCD59transgenic pigs (hCD59) that differed in hCD59 expressionlevels and a hCD59xHTAT21 double transgenic pig(hCD59xHT). Untreated and CM-treated cells were simulta-neously assayed. Results represent mean¡SE of fourindependent experiments. HT PAEC, treated or untreated,were significantly protected at 40% and 100% human serumrelative to controls, *Pj0.02. Only the higher expressorhCD59 transgenic cells achieved significant protection (40%human serum, both conditions), #Pj0.05. The doubletransgenic PAEC, treated or untreated, were significantlyprotected relative to control cells at 40 and 100% humanserum, **Pj0.005. Moreover, hCD59xHT cells were sig-nificantly more protected than hCD59 and HT singletransgenic cells at 100% human serum in the two conditionsanalyzed, Pj0.05.
Transgenic pigs expressing hCD59 and HT
53

up-regulation of Gala1,3-Gal epitope in activatedcells.Natural antibody reactivity and complement
deposition was also assessed on PAEC isolatedfrom the single and double transgenic pigs. Anti-Gala1,3-Gal IgM and IgG reactivity was deter-mined by flow cytometric analysis (Fig. 5A,B).The HTAT21 and hCD59xHTAT21 cells showed acomparable significant reduction (30–40%) inIgM and IgG reactivity relative to control cells.Anti-Gala1,3-Gal antibody binding remained lowafter treatment with CM (Fig. 5A,B). To evaluateclassical complement activation, cell surface C4deposition was determined by incubating the cellswith 20% C8-depleted human serum, followed byincubation with an anti-human C4 specific anti-body. Reactivity was assessed by flow cytometry(Fig. 5C). Control PAEC had a considerableamount of C4 on the cell surface, whereasHTAT21 and hCD59xHTAT21 cells exhibited a40% reduction in C4 deposition (Fig. 5C). Similarresults were obtained at 40% serum with a two-fold overall increase in mean fluorescence inten-sity (data not shown). Deposition of the C3complement component was assayed simulta-neously to C4 (Fig. 5D). We observed that C3reactivity at 20% serum was amplified in relationto C4 deposition in all cells analyzed (Fig. 5D).Although C3 deposition was decreased onHTAT21 transgenic PAEC in parallel to that ofC4 (30–40% reduction), the lowest C3 reactivitycorresponded to the hCD59xHTAT21 combina-tion (60% reduction) (Fig. 5D). Similar resultswere observed for both CM-treated and untreatedcells (Fig. 5D) and at 40% serum (data notshown). Therefore, our data demonstrated asignificant reduction in complement activationin HT and hCD59xHT transgenic PAEC.To assess the functional significance of trans-
gene expression in PAEC, we also performed ahuman serum-mediated cytolysis assay withcontrol and transgenic PAEC untreated or treatedwith CM (Fig. 6A,B). Cell survival was deter-mined by the proportion of dye retained atincreasing concentrations of human serum com-pared with cells with no serum added. Here weshow data representative of three sets of experi-ments that evaluated the percentage of cellsurvival of single and double transgenic cells(HTAT21 line). In all cases, control PAEC showeda marked reduction in cell survival after incuba-tion with 40 and 100% human serum (Fig. 6A).Treatment with CM further decreased cell survi-val (Fig. 6B). Two lines of PAEC that differed intheir level of hCD59 expression were simulta-neously assayed and showed an increased protec-tion at 20 and 40% human serum that correlated
with the level of hCD59 (Fig. 6A,B). However,both hCD59-expressing PAEC were highly sus-ceptible to human serum-mediated lysis at 100%serum. HT single transgenic cells, either CM-treated or untreated, were more resistant to lysisthan controls at all concentrations of serum(Fig. 6A,B). Importantly, the double transgenicPAEC were the most resistant to the challengewith human serum (82% survival at 40% serumand 68% at 100% serum) and the protection wasmaintained after treatment with CM (Fig. 6A).Thus, coexpression of hCD59 and HT markedlyenhances the resistance of endothelial cells tohuman serum-mediated lysis.
Discussion
The use of non-primate animals as donors of cellsand organs for transplantation is complicated byhyperacute rejection of the xenograft mediated bynatural antibody reactivity and complementactivation [1–4,8,9]. The feasibility of delayingHAR by expression of human complementregulatory proteins in the donor organ hashighlighted the need to develop genetic strategiesthat also inhibit the antibody-mediated rejection[16–23]. The deposition of xeno-antibodies notonly plays a role in HAR, but it is an integral partof DXR [12–14]. Wang et al. showed in a primedconcordant rodent model that xenogeneic hearttransplants were rejected at 6 days even withcomplete complement blockade [38]. Rejectionwas predominantly due to the induced antibodyresponse. Immunosuppression only extendedgraft survival temporarily [38]. In the pig-to-primate setting, DXR also occurs in spite ofpotent immunosuppression regimes [4,11–14,22,23] due to T-cell independent responses[14,39]. The Gal a1,3-Gal antigen is the majortarget of the humoral response in HAR and DXR[40]. Anti-Gal a1,3-Gal antibodies constitute themajority of pre-existing natural antibodies in thepig-to-primate combination [24,25]. In addition,induced anti-Gal a1,3-Gal antibody responseslead to increased titers that are refractory tocurrent immunosuppressive protocols [14,39].Attempts to control antibody-induced DXRusing plasmapheresis therapy have not provedefficacious [13,14].The ability to genetically modify the donor
organ to express one or more human complementinhibitors and remodel the carbohydrate cellsurface phenotype may eliminate the need fortreatments that affect global humoral immunityand result in a less immunocompromised recipi-ent. The first studies assessing the efficacy of this
Costa et al.
54

combinatorial approach in engineered animalswere performed in the mouse model [32,33]. Co-expression of HT and hCD59 in splenocytes oftransgenic mice led to enhanced protection whenchallenged with human serum [32]. Tange et al.also showed increased viability ofhCD59-transgenic splenocytes on the Gal KObackground, although the highest concentrationof serum assayed was only 50% [41]. Interestingly,high expression of HT and hCD59was as effectiveas the hCD59xGal KO combination [32]. Here wedemonstrated that coexpression of HT andhCD59 in PBMC and PAEC from transgenicpigs markedly protected these cells from humanserum-mediated cytolysis, even when cells werechallenged with 100% serum. The combination ofthe two engineering components had an additive,or more than additive, protective effect that wasdependent on the levels of transgene expression.This is in accordance with our observations thatthe expression levels of H- and Gala1,3-Galepitopes correlate inversely in different cell types[31]. Furthermore, this level of protection wasmaintained after treatment with cytokine-condi-tioned media, indicating that the combination ofHT and hCD59 provides protection fromhumoral lysis post-cell activation.The human serum-mediated cytolysis assay we
utilized was shown to correlate well with theoutcome of organs in ex vivo perfusion and pig-to-primate transplant setting [15–17,42]. Therefore,our data suggested that this combinatorialapproach is advantageous to the single strategiesdesigned to overcome HAR by expressing eitherHT or complement inhibitors alone. The combi-nation of HT with hCD59 and DAF led toimproved heart performance in a mouse ex vivoheart perfusion model [33]. Thus, we believe thatwe may achieve very high protection to HAR ofpig vascularized organs by expressing high levelsof HT and hCD59 on endothelium. We haveobserved direct correlation between the level oftransgene expression and the level of protection,therefore it is important to attain high expressionlevels by selecting lines of transgenic pigs andutilizing promoters that can direct high geneexpression levels to the donor tissues. A previousattempt to generate double and triple transgenicpigs that combineHTwith complement inhibitorsdid not achieve sufficient levels of HT expressionto evaluate its efficacy [43]. Ourmuch higher levelsof H-epitope expression are probably due to theuse of the CMV promoter. Moreover, futuredevelopments include expansion of the HTAT20
high expressor line, generation of homozygoustransgenic pigs and intercrosses of the two HT
transgenic lines to achieve the highest HTexpression levels in combination with hCD59.The current strategy may also be beneficial in
combating parallel alternative pathway comple-ment activation [44,45], endothelial cell activationand downstream rejection events (DXR). Themembrane attack complex plays a direct role inmediating the activation of endothelial cellsleading to thrombin release and the generationof a pro-coagulant cell surface [12,46]. Kennedyet al. demonstrated that the expression of hCD59blocked this process [47]. Xenoantibodies alsoplay a role in vascular DXR [12–14,46] andantibody-dependent cell-mediated cytotoxicity[48]. Therefore, a reduction in the expression ofthe Gala1,3-Gal epitope may result in decreasedantibody-mediated DXR.The generation of genetically engineered por-
cine endothelial cells highly resistant to inter-species immune responses also opens a new field ofapplications. The recent technical advances in thein vitro production of vessels could clearly benefitfrom our technology. These cells might be utilizedin the development of artificial vessels for bypassgrafting in coronary artery and peripheral vas-cular disease [49]. With regard to other cell types,we have demonstrated expression of both HT andhCD59 in tissues used as the source of cells fortissue engineering and cell transplantation (sciaticnerve, skin, and cartilage). PBMC and PAECexpressing high levels of both transgenes werehighly resistant to the challenge with 100% humanserum. Similarly-engineered cells might elude theeffect of antibody and complement when trans-planted into a primate. In vitro and in vivo studiesevaluating the efficacy of this approach in celltransplantation models are currently focused oncell types to be used in specific indications, such asolfactory ensheathing cells for spinal cord injuryand chondrocytes to repair articular cartilagedefects. Another possible application that wouldbenefit from local complement inhibition ispancreatic islet transplantation, although expres-sion of HT may only be needed in fetal cells [50].Thus, we firmly believe that our combinatorialstrategy will have its major impact in the shortterm in the field of cell transplantation.Although further studies are being pursued to
confirm that this combinatorial strategy mayprevent humoral rejection of pig cells andorgans transplanted in primates, these resultsdemonstrate the efficacy of the strategy.Transgenic pigs expressing HT in combinationwith complement inhibitors may provide a sourceof cells and tissues for clinical transplantation incombination with immunosuppressive regimens.With the advent of pig cloning [51], the possibility
Transgenic pigs expressing hCD59 and HT
55

of generating Gal KO pigs is imminent. Thecombination of HT and complement inhibitorscoupled with the Gal KO pig should continue tobe important. Cells isolated fromGalKOmice arehighly susceptible to human serum-mediated lysis,unless combined with a complement inhibitor[32]. Uncapped epitopes in cells fromGal-deficient mice are probably recognized byhuman natural antibodies. We believe thatexpression of HT may be needed to cover theseepitopes as it produces the universally acceptedblood group O. Further genetic manipulation ofthe donor tissues to fully abrogate cell activationand cellular immune responses promises futuredevelopment of the optimal ‘universal donor’ forclinical xenotransplantation.
Acknowledgments
We thank Y. Shen for excellent technical assis-tance in the histologic analysis and B. A. Bensonfor skilful technical support. The anti-Gal resinswere kindly supplied as a gift from Kurt Nilsson(Glycorex Transplantation AB). This work was inpart supported by a National Institutes of Stand-ards and Technology – Advanced TechnologyProgram grant to W.L.F. A fellowship awardfrom the Spanish Ministry of Education andCulture temporally supported C.C.
References
1. AUCHINCLOSS H, SACHS DH. Xenogeneic transplantation.Annu Rev Immunol 1998; 16: 433.
2. PLATT JL, BACH FH.Discordant xenografting: challengesand controversies. Curr Opin Immun 1991; 3: 735.
3. LU CY, KHAIR-EL-DIN T, DAWIDSON IA et al. Xenotrans-plantation. FASEB J 1994; 8: 1122.
4. LAMBRIGTSD,SACHSDH,COOPERDKC.Discordantorganxenotransplantation in primates. Transplantation 1998;66: 547.
5. EDGE ASB, GOSSEME, DINSMORE J. Xenogeneic cell ther-apy: current progress and future developments in porcinecell transplantation. Cell Transplantation 1998; 7: 525.
6. FINK JS, SCHUMACHER JM, ELLIAS SL et al. Porcine xeno-grafts in Parkinson’s disease and Huntington’s diseasepatients: preliminary results. Cell Transplantation 2000;9: 273.
7. CHEN SC,MULLONC,KAHAKUE et al. Treatment of severeliver failure with a bioartificial liver. Ann N Y Acad Sci1997; 831: 350.
8. BENNETW, SUNBERGB, LUNDGRENT et al. Damage to por-cine isletsofLangerhansafterexposure tohumanblood invitro,orafter intraportal transplantation tocynomologusmonkeys. Transplantation 2000; 69: 711.
9. BARKER RA, RATCLIFFE E, MCLAUGHLIN M, RICHARDS A,DUNNET SB. A role for complement in the rejection ofporcine ventral mesencephalic xenografts in a rat modelof Parkinson’s disease. J Neurosci 2000; 20: 3415.
10. DALMASSOAP,VERCELOTTIGM,FISCHELRJ et al.Mechan-
ism of complement activation in the hyperacute rejectionofporcineorgans inprimate recipients.AmJPathol 1992;140: 1157.
11. KOBAYASHI T, TANIGUCHI S, NEETHLING FA et al. Delayedxenograft rejection of pig-to-baboon cardiac transplantsafter cobra venom factor therapy. Transplantation 1997;64: 1255.
12. PLATT JL, LIN SS,MCGREGORCGA.Acute vascular rejec-tion. Xenotransplantation 1998; 5: 169.
13. LIN SS, WEIDNER BC, BYRNE GW et al. The role of anti-bodies in acute vascular rejection of pig-to-babooncardiac transplants. J Clin Invest 1998; 101: 1745.
14. KOZLOWSKI T, SHIMIZU A, LAMBRIGTS D et al. Porcinekidney and heart transplantation in baboons undergoinga tolerance induction regimen and antibody adsorption.Transplantation 1999; 67: 18.
15. FODORWL,WILLIAMS BL,MATIS LA et al. Expression of afunctional human complement inhibitor in a transgenicpig as a model for the prevention of xenogeneic hyper-acute organ rejection. Proc Natl Acad Sci USA 1994; 91:11153.
16. KROSHUSTJ, BOLMAN IIIRMDALMASSOAP et al. Express-ion of human CD59 in transgenic pig organs enhancesorgan survival in an ex vivo xenogeneic perfusion model.Transplantation 1996; 61: 1513.
17. KULICKDM, SALERNOCT,DALMASSOAP et al. Transgenicswine lungs expressing human CD59 are protected frominjury in a pig-to-human model of xenotransplantation.J Thorac Cardiovasc Surg 2000; 119: 690.
18. MCCURRY KR, KOOYMAN DL, ALVARADO CG et al.Human complement regulatory proteins protect swine-to primate cardiac xenografts from humoral injury. NatMed 1995; 1: 423.
19. DIAMOND LE, MCCURRY KR, MARTIN MJ et al. Charact-erization of transgenic pigs expressing functionally activehuman CD59 on cardiac endothelium. Transplantation1996; 61: 1241.
20. COZZI E, YANNOUTSOS N, LANGFORD GA, PINO-CHAVEZ G,WALLWORK J,WHITEDJG. Effect of transgenic expressionof human decay-accelerating factor on the inhibition ofhyperacute rejection of pig organs. In: COOPER DKC,KEMP E. eds. Xenotransplantation. Springer-Verlag,1997.
21. BYRNEGW,MCCURRYKR,MARTINMJ,MCCLELLANSM,PLATT JL, LOGAN JS. Transgenic pigs expressing humanCD59 and decay accelerating factor produce an intrinsicbarrier to complement-mediated damage. Transplanta-tion 1997; 63: 149.
22. SCHMOECKEL M, BHATTI FNK, ZAIDI A et al. Orthotopicheart transplantation in a transgenic pig-to-primatemodel. Transplantation 1998; 65: 1570.
23. ZAIDI A, SCHMOECKEL M, BHATTI FNK et al. Life-supporting pig-to-primate xenotransplantation usinggenetically modified donors. Transplantation 1998; 65:1584.
24. GALILI U, Evolution and pathophysiology of the humananti-a-galactosyl IgG. (anti-Gal) antibody. SpringerSemin Immunopathol 1993; 15: 155.
25. SANDRIN MS, VAUGHAN HA, DABKOWSKI PL, MCKENZIE
IFC. Anti-pig IgM antibodies in human serum reactspredominantly with Gal (al,3) Gal epitopes. Proc NatlAcad Sci USA 1993; 90: 11391.
26. SANDRINMS, FODORWL,MOUHTOURIS E et al. Enzymaticremodelling of the carbohydrate surface of a xenogeniccell substantially reduces human antibody binding andcomplement-mediated cytolysis. Nature Med 1995; 1:1261.
Costa et al.
56

27. SHARMAA,OKABE J,BIRCHPet al.Reduction in the level ofGala (1,3)Gal in transgenicmiceandpigsbyexpressionofan a (1, 2) fucosyltransferase. Proc Natl Acad Sci USA1996; 93: 7190.
28. KOIKEC,KANNAGIR,TAKUMAYetal. Introductionofa (1,2)–fucosyltransferase and its effect on a–Gal epitopes intransgenic pig. Xenotransplantation 1996; 3: 81.
29. COHNEY S, MCKENZIE IFC, PATTON K et al. Down-regulation of Gala (1,3) Gal expression by a1, 2–fuco-syltransferase. Further characterization of a1BV, 2–fuco-syltransferase transgenic mice. Transplantation 1997; 64:495.
30. CHEN CG, SALVARIS EJ, ROMANELLA M et al. Transgenicexpression of human a1, 2–fucosyltransferase (H–trans-ferase) prolongsmouse heart survival in an ex vivomodelof xenograft rejection. Transplantation 1998; 65: 832.
31. COSTA C, ZHAO L, BURTONWV et al. Expression by a1, 2–fucosyltransferase in transgenic pigs modifies the cellsurface carbohydrate phenotype and confers resistance tohuman serum–mediated cytolysis. FASEB J 1999; 13:1762.
32. COSTA C, ZHAO L, DECESARE S, FODOR WL. Comparativeanalysis of three genetic modifications designed to inhibithuman serum-mediated cytolysis. Xenotransplantation1999; 6: 6.
33. COWAN PJ, CHEN CG, SHINKEL TA et al. Knock-out ofa1,3-galactosyltransferase or expression of a1, 2–fuco-syltransferase further protects CD55–and CD59–expres-sing mouse hearts in an ex vivo model of xenograftrejection. Transplantation 1998; 65: 1599.
34. SOUTHERN EM. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J MolBiol 1975; 98: 503.
35. HAYESCE,GOLDSTEIN IJ.Ana-D-galactosyl-binding lectinfrom Bandeiraea simplicifolia seeds. J Biol Chem 1974;249: 1904.
36. MATSUMOTO I,OSAWAT.Purification and characterizationof an antiH (O) phytohemagglutinin of Ulex europeus.Biochim Biophys Acta 1969; 194: 180.
37. ORIOLR,YONGY,KORENE,COOPERDKC.Carbohydrateantigens of pig tissues reacting with human naturalantibodies as potential targets for hyperacute vascularrejection in pig-to-man organ xenotransplantation.Transplantation 1993; 56: 1433.
38. WANGH,ROLLINSSA,GAOZetal.Complement inhibitionwith an anti-C5 monoclonal antibody prevents hyper-acute rejection in a xenograft heart transplantationmodel. Transplantation 1999; 68: 1643.
39. KOZLOWSKI T, MONROY R, XU Y et al. Anti-Gala1–3Gal
antibody response to porcine bonemarrow inunmodifiedbaboons and baboons conditioned for tolerance induc-tion. Transplantation 1998; 66: 176.
40. XU Y, LORF T, SABLINSKI T et al. Removal of anti-porcinenatural antibodies from human and nonhuman primateplasma in vitro and in vivo by a Gala1–3Galb1–4bGlc-Ximmunoaffinity column. Transplantation 1998; 65: 172.
41. TANGE MJ, SALVARIS EJ, ROMANELLA M et al. Additiveeffects of CD59 expression in Gal knockout mice in vitrobut not in an ex vivomodel.Xenotransplantation 1997; 4:25.
42. TANIGUCHIS,NEETHLINGFA,KORCHAGINAEYetal. Invivoimmunoadsorption of antipig antibodies in baboonsusing a specific Gala1–3Gal column. Transplantation1996; 62: 1379.
43. COWAN PJ, AMINIAN A, BARLOW H et al. Renal xenografsfrom triple-transgenic pigs are not hyperacutely rejectedbut cause coagulopathy in non-immunosuppressedbaboons. Transplantation 2000; 69: 2504.
44. ROMANELLA M, AMINIAN A, ADAM WR, PEARSE MJ,D’APICE AJF. Involvement of both the classical andalternate pathways of complement in an ex vivomodel ofxenograft rejection. Transplantation 1997; 63: 1021.
45. MAKOWKA L, CRAMER DV, HOFFMAN A et al. The use of apig liver xenograft for temporary supportofapatientwithfulminanthepatic failure.Transplantation1995;59:1654.
46. SAADI S, PLATT JL. Transient perturbations of endothelialintegrity induced by natural antibodies and complement.J Exp Med 1995; 181: 21.
47. KENNEDY S, ROLLINS SA, BURTON WV et al. Protectionof porcine aortic endothelial cells from complement-mediated cell lysis and activation by recombinant humanCD59. Transplantation 1994; 57: 1494.
48. SCHAAPHERDER AFM, DAHA MR, TE BULTE M-TJW, VAN
DER WOUDE FJ, GOOSZEN HG. Antibody-dependant cell-mediated cytotoxicity against porcine endotheliuminduced by a majority of human sera. Transplantation1994; 57: 1376.
49. NIKLASON LE, GAO J, ABBOTT WM et al. Functionalarteries grown in vitro. Science 1999; 284: 489.
50. BENNET W, BJORKLAND A, SUNBERG B et al. A comparisonof fetal and adult porcine islets with regard to Gal (1, 3)Gal expression and the role of human immunoglobulinsandcomplement in islet cell cytotoxicity.Transplantation2000; 69: 1711.
51. POLEJAEVA IA, CHEN S-H, VAUGHT TD et al. Cloned pigsproduced by nuclear transfer from adult somatic cells.Nature 2000; 407: 86.
Transgenic pigs expressing hCD59 and HT
57
Copyright © 2022 FDOKUMEN




















