
Involvement of the NLRP3 Inflammasome in Innate and Humoral Adaptive Immune Responses to Fungal...
7
Involvement of the NLRP3 Inflammasome in Innate and Humoral Adaptive Immune Responses to Fungal -Glucan 1 Himanshu Kumar, 2 * ‡ Yutaro Kumagai,* ‡§ Tetsuo Tsuchida,* ‡ Paul A. Koenig, 3 * ‡ Takashi Satoh,* ‡ Zijin Guo, † Myoung Ho Jang, † Tatsuya Saitoh,* ‡ Shizuo Akira, 4 * ‡§ and Taro Kawai* ‡ Fungal -glucan, such as curdlan, triggers antifungal innate immune responses as well as shaping adaptive immune responses. In this study, we identified a key pathway that couples curdlan to immune responses. Curdlan promoted the production of the proinflammatory cytokine IL-1 by dendritic cells and macrophages through the NLRP3 inflammasome. Stimulation with Can- dida albicans and Saccharomyces cerevisiae also triggered the NLRP3 inflammasome-mediated IL-1 production. In vivo, NLRP3 was required for efficient Ag-specific Ab production when curdlan was used as an adjuvant, whereas it was dispensable for the induction of Th1 and Th17 cell differentiation. Furthermore, stimulation of purified B cells with curdlan-induced CD69 up- regulation and IgM production while stimulation with other NLRP3 inflammasome activators, such as silica and aluminum salt, did not. Notably, this induction required NLRP3 but was independent of Toll-like receptor and IL-1 receptor family signaling, suggesting the presence of NLRP3-dependent and IL-1 receptor family independent mechanisms in B cells responsible for Ab responses. Collectively, these findings reveal a critical role for the NLRP3 inflammasome in the regulation of antifungal innate immune responses as well as B cell activation. The Journal of Immunology, 2009, 183: 8061– 8067. I nnate immune detection of microbial pathogens such as vi- ruses, bacteria, and fungi is a critical element for their elim- ination as well as establishing Ag-specific adaptive immu- nity. Sentinels in the innate immune system, such as dendritic cells (DC) 5 and macrophages, recognize conserved microbial compo- nents known as pathogen-associated molecular patterns (PAMPs) through germline-encoded pattern-recognition receptors (PRRs), and initiate inflammatory responses. These include membrane- bound TLRs and intracellular proteins such as retinoic acid-induc- ible gene-I-like receptors, cytosolic DNA sensor, and NOD-like receptors (NLR) (1– 4). Of these, NLR constitutes a large family of intracellular PRRs, and several of these are well characterized. Among these NLRs, NLRP3 (also known as NALP3/Cryopyrin/ CIAS1/PYPAF1) responds to multiple stimuli and forms a multi- protein complex with apoptosis-associated speck-like protein con- taining a caspase recruitment domain (ASC) and casapse-1, referred to as the NLRP3 inflammasome, which triggers secretion of an endogenous pyrogen IL-1 (2– 4). Secretion of IL-1 re- quires at least two signals; the first signal is cell priming that re- sults in the synthesis of pro-IL-1 protein within cells, and the second signal is the inflammasome-dependent proteolysis of pro- IL-1 to secretory and bioactive IL-1. The NLRP3 inflamma- some is activated by PAMPs (DNA and RNA), ATP, self-derived molecules (monosodium urate, -amyloid), crystals (silica and asbes- tos), and immune adjuvants (aluminum salt (alum) and biodegradable poly (lactide-co-glycolide) particles) (2– 4). Although it is still unclear how the NLRP3 inflammasome alone responds to diverse stimuli, it has been proposed that the NLRP3 inflammasome is activated by ligand-induced intermediates such as reactive oxygen species (ROS), K efflux, and the lysosome destabilization (2–4). Although mechanisms of the inflammasome on IL-1 produc- tion have been well characterized, its contribution to the generation of adaptive immune responses remains unclear. It was shown that alum adjuvanticity in terms of Ag-specific Ab production and Th2 response is impaired in NLRP3-deficient mice (5–7). However, there is another report showing a dispensable role for NLRP3 in alum-enhanced Ab production (8). Moreover, a dispensable role of NLRP3 on Ab production was reported when biodegradable poly (lactide-co-glycolide) particle adjuvant was used (9). -glucan is the most abundant cell wall component of fungi and yeast, and is also present in bacteria and plants but not in animals, thereby acting as a PAMP. -glucan has diverse immunomodula- tory properties, and has been shown to protect against infection with pathogenic bacteria, viruses, fungi, and protozoa. Further- more, it functions as an adjuvant for the treatment of cancer (10). -glucan is recognized by a C-type lectin, dectin-1, which consists of an extracellular carbohydrate-recognition domain and cytoplas- mic tyrosine-containing domain that is similar to the ITAM motif *Laboratory of Host Defense, † Laboratory of Gastrointestinal Immunology, World Premier International Immunology Frontier Research Center, ‡ Department of Host Defense, Research Institute for Microbial Diseases, and § Global Centers of Excel- lence program, Frontier Biomedical Science Underlying Organelle Network Biology, Osaka University, Japan Received for publication July 29, 2009. Accepted for publication October 19, 2009. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 H.K. was supported by postdoctoral fellowships (P08123) from the Japan Society for Promotion of Science, Japan. P.A.K. was supported by Deutscher Akademischer Austausch Dienst (German Academic Exchange Service). This work was in part sup- ported by grants from National Institutes of Health (PO1 AI070167) and by Grant- in-Aid for Specially Promoted Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan. 2 Current address: Laboratory of Immunology, Indian Institute of Science Education and Research (IISER), Bhopal, Transit campus: ITI (Gas Rahat) Building, Govind- pura, Bhopal, India. 3 Current address: Whitehead Institute for Biomedical Research 9 Cambridge Center, Cambridge, MA 02142. 4 Address correspondence and reprint requests to Prof. Shizuo Akira, 3-1 Yamada- oka, Suita, Osaka, Japan. E-mail address: [email protected] 5 Abbreviations used in this paper: DC, dendritic cell; PAMP, pathogen-associated molecular pattern; PRR, pattern-recognition receptor; NLR, NOD-like receptor; PEC, peritoneal exudate cell; BMDC, GM-CSF-induced bone marrow DC; d, day; WT, wild type; ROS, reactive oxygen species; Syk, spleen tyrosine kinase; ASC, apopto- sis-associated speck-like protein containing a caspase recruitment domain; CARD, caspase recruitment domain. Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00 The Journal of Immunology www.jimmunol.org/cgi/doi/10.4049/jimmunol.0902477
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Involvement of the NLRP3 Inflammasome in Innate and Humoral Adaptive Immune Responses to Fungal...

Involvement of the NLRP3 Inflammasome in Innate andHumoral Adaptive Immune Responses to Fungal �-Glucan1
Himanshu Kumar,2*‡ Yutaro Kumagai,*‡§ Tetsuo Tsuchida,*‡ Paul A. Koenig,3*‡ Takashi Satoh,*‡
Zijin Guo,† Myoung Ho Jang,† Tatsuya Saitoh,*‡ Shizuo Akira,4*‡§ and Taro Kawai*‡
Fungal �-glucan, such as curdlan, triggers antifungal innate immune responses as well as shaping adaptive immune responses. Inthis study, we identified a key pathway that couples curdlan to immune responses. Curdlan promoted the production of theproinflammatory cytokine IL-1� by dendritic cells and macrophages through the NLRP3 inflammasome. Stimulation with Can-dida albicans and Saccharomyces cerevisiae also triggered the NLRP3 inflammasome-mediated IL-1� production. In vivo, NLRP3was required for efficient Ag-specific Ab production when curdlan was used as an adjuvant, whereas it was dispensable for theinduction of Th1 and Th17 cell differentiation. Furthermore, stimulation of purified B cells with curdlan-induced CD69 up-regulation and IgM production while stimulation with other NLRP3 inflammasome activators, such as silica and aluminum salt,did not. Notably, this induction required NLRP3 but was independent of Toll-like receptor and IL-1 receptor family signaling,suggesting the presence of NLRP3-dependent and IL-1 receptor family independent mechanisms in B cells responsible for Abresponses. Collectively, these findings reveal a critical role for the NLRP3 inflammasome in the regulation of antifungal innateimmune responses as well as B cell activation. The Journal of Immunology, 2009, 183: 8061–8067.
I nnate immune detection of microbial pathogens such as vi-ruses, bacteria, and fungi is a critical element for their elim-ination as well as establishing Ag-specific adaptive immu-
nity. Sentinels in the innate immune system, such as dendritic cells(DC)5 and macrophages, recognize conserved microbial compo-nents known as pathogen-associated molecular patterns (PAMPs)through germline-encoded pattern-recognition receptors (PRRs),and initiate inflammatory responses. These include membrane-bound TLRs and intracellular proteins such as retinoic acid-induc-ible gene-I-like receptors, cytosolic DNA sensor, and NOD-likereceptors (NLR) (1–4). Of these, NLR constitutes a large family of
intracellular PRRs, and several of these are well characterized.Among these NLRs, NLRP3 (also known as NALP3/Cryopyrin/CIAS1/PYPAF1) responds to multiple stimuli and forms a multi-protein complex with apoptosis-associated speck-like protein con-taining a caspase recruitment domain (ASC) and casapse-1,referred to as the NLRP3 inflammasome, which triggers secretionof an endogenous pyrogen IL-1� (2–4). Secretion of IL-1� re-quires at least two signals; the first signal is cell priming that re-sults in the synthesis of pro-IL-1� protein within cells, and thesecond signal is the inflammasome-dependent proteolysis of pro-IL-1� to secretory and bioactive IL-1�. The NLRP3 inflamma-some is activated by PAMPs (DNA and RNA), ATP, self-derivedmolecules (monosodium urate, �-amyloid), crystals (silica and asbes-tos), and immune adjuvants (aluminum salt (alum) and biodegradablepoly (lactide-co-glycolide) particles) (2–4). Although it is still unclearhow the NLRP3 inflammasome alone responds to diverse stimuli, ithas been proposed that the NLRP3 inflammasome is activated byligand-induced intermediates such as reactive oxygen species (ROS),K� efflux, and the lysosome destabilization (2–4).
Although mechanisms of the inflammasome on IL-1� produc-tion have been well characterized, its contribution to the generationof adaptive immune responses remains unclear. It was shown thatalum adjuvanticity in terms of Ag-specific Ab production and Th2response is impaired in NLRP3-deficient mice (5–7). However,there is another report showing a dispensable role for NLRP3 inalum-enhanced Ab production (8). Moreover, a dispensable role ofNLRP3 on Ab production was reported when biodegradable poly(lactide-co-glycolide) particle adjuvant was used (9).
�-glucan is the most abundant cell wall component of fungi andyeast, and is also present in bacteria and plants but not in animals,thereby acting as a PAMP. �-glucan has diverse immunomodula-tory properties, and has been shown to protect against infectionwith pathogenic bacteria, viruses, fungi, and protozoa. Further-more, it functions as an adjuvant for the treatment of cancer (10).�-glucan is recognized by a C-type lectin, dectin-1, which consistsof an extracellular carbohydrate-recognition domain and cytoplas-mic tyrosine-containing domain that is similar to the ITAM motif
*Laboratory of Host Defense, †Laboratory of Gastrointestinal Immunology, WorldPremier International Immunology Frontier Research Center, ‡Department of HostDefense, Research Institute for Microbial Diseases, and §Global Centers of Excel-lence program, Frontier Biomedical Science Underlying Organelle Network Biology,Osaka University, Japan
Received for publication July 29, 2009. Accepted for publication October 19, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 H.K. was supported by postdoctoral fellowships (P08123) from the Japan Societyfor Promotion of Science, Japan. P.A.K. was supported by Deutscher AkademischerAustausch Dienst (German Academic Exchange Service). This work was in part sup-ported by grants from National Institutes of Health (PO1 AI070167) and by Grant-in-Aid for Specially Promoted Research from the Ministry of Education, Culture,Sports, Science and Technology of Japan.2 Current address: Laboratory of Immunology, Indian Institute of Science Educationand Research (IISER), Bhopal, Transit campus: ITI (Gas Rahat) Building, Govind-pura, Bhopal, India.3 Current address: Whitehead Institute for Biomedical Research 9 Cambridge Center,Cambridge, MA 02142.4 Address correspondence and reprint requests to Prof. Shizuo Akira, 3-1 Yamada-oka, Suita, Osaka, Japan. E-mail address: [email protected] Abbreviations used in this paper: DC, dendritic cell; PAMP, pathogen-associatedmolecular pattern; PRR, pattern-recognition receptor; NLR, NOD-like receptor; PEC,peritoneal exudate cell; BMDC, GM-CSF-induced bone marrow DC; d, day; WT,wild type; ROS, reactive oxygen species; Syk, spleen tyrosine kinase; ASC, apopto-sis-associated speck-like protein containing a caspase recruitment domain; CARD,caspase recruitment domain.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0902477

(ITAM) (11–14). Engagement of dectin-1 by �-glucan triggers therecruitment of spleen tyrosine kinase (Syk), which activatesNF-�B through the caspase recruitment domain (CARD)9, Bcl10,and MALT1 complex. This results in induction of cytokines suchas IL-10, IL-6, IL-2, IL-23, and TNF-�, and up-regulation of co-stimulatory molecules on DC, which can instruct the differentiation ofboth IFN-�-producing Th1 and IL-17-producing Th17 cells (15–18).Dectin-1 signaling also enhances the effecter responses of cytotoxicCD8 T cells as well as potentiating Ab production (18, 19). Theseresponses are linked to host defense against pathogenic fungi such asCandida albicans, Aspergillus fumigatus, and Pneumocystis carinii,as well as protection against tumors (19–21). Moreover, other c-typelectins, including dectin-2 and mincle, are reported to participate inantifungal responses (22–24). In addition, scavenger receptors such asSCARF1 and CD36 (25), and complement receptors (CR3) areknown to recognize �-glucans (10). Furthermore, lactosylceramide, aglycosphingolipid abundant in the plasma membranes of many cells,is also reported to recognize �-glucans (26).
Curdlan and zymosan are model ligands for �-glucan that areused for the study on antifungal immune responses. Curdlan is apure �-1,3-glucan that was shown to stimulate macrophages andDC independent of TLRs, instead requiring dectin-1. Zymosan isan immunoactive cell-wall extract of Saccharomyces cerevisiaethat is mainly composed of �-glucan with little contamination ofmannans, chitin, protein, and lipids. Recognition of zymosan ismediated by a combination of dectin-1 and TLR2 (13, 14).
In this study, we found that stimulation of DC and macrophageswith zymosan, curdlan, C. albicans, or S. cerevisiae resulted in theproduction of IL-1� through activation of the NLRP3 inflamma-some. This pathway was required for enhancing the effects of curd-lan on B cell-mediated humoral immunity but dispensable for Th1and Th17 cell differentiation. Moreover, B cells were directly ac-tivated in response to curdlan, and this activation required NLRP3but not TLR and IL-1 receptor signaling, suggesting a critical roleof B cell-intrinsic NLRP3 in Ab responses. Thus, the NLRP3 in-flammasome is a critical component for regulating �-glucan-in-duced innate and humoral adaptive immune responses.
Materials and MethodsMice
C57BL/6 mice were obtained from Clea. Mice lacking NLRP3 (NALP3),ASC, NLRC4 (IPAF), and MyD88 are described previously (27–29). All ofthese mice were on the C57BL/6 background. Mice were bred and main-tained in a specific pathogen free facility of the Research Institute forMicrobial Diseases in accordance with the specifications of the Associationfor Assessment and Accreditation of Laboratory Animal Care. Mouse pro-tocols were approved by the Osaka University Animal Care and Use Com-mittee. All mice were sacrificed at 5–9 wk of age.
Reagents
Cells were cultured with RPMI 1640 (Sigma-Aldrich) supplemented withpenicillin (Life Technologies), streptomycin (Life Technologies), 100 mM2-ME (Nacalai), and 10% (vol/vol) heat-inactivated FCS (Life Technolo-gies). Curdlan (Wako) and zymosan A (Wako) were suspended in PBS ata concentration of 10 mg/ml. OVA protein used for immunizations and invitro assays was from Sigma-Aldrich. Anti-IL-1� Ab and the substratereagent pack were from R&D Systems. Anti-CD40 (HM40-3), PE-anti-CD86 (GL1), allophycocyanin-anti-CD11c (HL3), allophycocyanin- orFITC-anti-CD3� (145-2C11), allophycocyanin-anti-CD8 (53-6.7), PerCP-anti-CD4 (GK1.5), FITC or PE-anti-IFN-� (XMG1.2), PE-anti-IL-4(11B11) PE-anti-IL-17 (TC11-18H10.1), PE-anti-CD69 (H1.2F3), strepta-vidin-allophycocyanin, GolgiStop and purified rat anti-mouse CD16/CD32(Mouse Fc-block) were from BD Biosciences Pharmingen. Mouse CD19-microbeads used for the purification of splenic B cells was from MACSMiltenyi Biotec. Alum, silica, PMA, ionomycin, piceatannol, nigericin, andbafilomycin A were from Sigma-Aldrich. FeTPPs were from Calbiochem.Ac-YVAD-CMK was from Peptide Institute. HRP-conjugated anti-mouseIgM, IgG2b, and IgG2c were from Southern Biotechnology Associates. OVA-
specific IgE was measured by using an IgE ELISA quantitation kit (BethylLaboratories) with some modifications. Mouse total IgM, and IgE ELISAquantitation kit was from Bethyl Laboratories. Ab specific for caspase-1 p10(sc-514) was from Santa Cruz Biotechnology. Ultra pure Escherichia coli LPSwas from Invivogen.
Cells
Peritoneal exudate cells (PEC) were isolated as described previously (30).Bone-marrow DC (BMDC) were prepared by cultivating bone-marrowcells in the presence of 10 ng/ml mouse GM-CSF (PeproTech) (30). B cellswere isolated from spleen cells by MACS using anti-CD19 microbeads(positive selection) or a B cell isolation kit (negative selection) from Milte-nyi Biotec as described by the manufacturer.
ELISA
PEC or GM-CSF-induced DC (2–3 � 105 cells/well of 96-well plate) werestimulated in 0.2 ml of medium for 24 h. IL-1� and IL-6 levels weremeasured by ELISA. Spleen cells day 21 from immunized mice (days 0, 7,and 14, described in Ab titer section) (4 � 106 cells/well of 24 wells plate)were cultured with OVA (10 or 50 �g/ml) in 0.5 ml of medium for 72 hand analyzed for IFN-�, IL-17, and IL-4 production by ELISA. ELISA kitsfor IL-1�, IL-6, IL-17, IFN-�, and IL-4 were purchased from R&D Sys-tems, and were performed according to the manufacturer’s instructions.
RT-PCR
Total RNA was isolated from curdlan-stimulated or unstimulated purifiedB cells or unstimulated PEC. cDNA was prepared by RT-PCR as describedpreviously (30). PCR was then done with following primers: NLRP3, 5�-ATGACGAGTGTCCGTTGC-3�and 5�-GATGGTCAGGAGTTCATG-3�and �-actin, 5�-GACATGGAGAAGATCTGGCACCACA-3� and5�-ATCTCCTGCTCGAAGTCTAGAGCAA-3�.
Immunoblotting
Cells were lysed in lysis buffer (1% Nonidet P-40, 50 mM Tris (pH 7.4),150 mM NaCl) supplemented with a complete protease inhibitor mixture(Roche), and mixed with sample buffer. The cell culture supernatants wereprecipitated as described and the protein pellets were resuspended in PBSand mixed with sample buffer (31). The protein samples were resolved,transferred, immunoblotted, and visualized as described previously (31).
Culturing C. albicans and S. cerevisiae
C. albicans Robin Berkhout anamorph (American Type Culture Collectionnumber 18804) and S. cerevisiae AH109 were cultured in Sabouraud Dex-trose (1% peptone, 4% glucose) and YPD medium (BD Biosciences, Clon-tech), respectively. Cells were infected with C. albicans and S. cerevisiaein log phase of growth.
Ab titer
Mice were immunized twice (days 0 and 7) i.p., with OVA (50 �g/mouse)plus curdlan (5 mg/mouse) or OVA (10 �g/mouse) plus silica (50 mg/mouse) in 200 �l PBS. Serum samples were taken at day 14 or day 21 andanalyzed for OVA-specific IgM, total IgG, IgG1, IgG2b, IgG2c, or IgE byELISA. OVA protein (10 �g/ml) in carbonate buffer (pH 9.6) was used tocoat 96-well plates, and incubated overnight at 4°C. Plates were thenwashed with PBS containing 0.05% Tween 20. Serial dilutions of serum inPBS/Tween 20 containing 5% skim milk were applied and incubated for2 h at room temperature. After washing, Abs were detected using HRP-con-jugated goat anti-mouse IgM, IgG1, IgG2b, IgG2c, or IgE (from kit describedabove in the reagents section). After an additional washing step, the plateswere developed with substrate from the substrate reagent pack. The reactionwas stopped by the addition of 1 M H2SO4 and the absorbance was measuredat 450 nm (32, 33). Total IgM in the culture supernatants were determinedusing the mouse IgM quantitation kit from Bethyl Laboratories.
Flow cytometry
Intracellular IFN-� and IL-17, and surface CD86, CD69, CD4, and CD8 ex-pression was analyzed as described previously (30, 34). Intracellular expres-sion of IFN-� in CD4� or CD8� T cells was analyzed using the Cytofix/Cytoperm Kit Plus (with GolgiStop; BD Biosciences) according to themanufacturer’s instructions. In brief, spleen cells were incubated with 50 ng/mlphorbol myristate acetate (Sigma-Aldrich), 5 �M calcium ionophore A23187(Sigma-Aldrich), and GolgiStop in complete medium at 37°C for 5 h. Surfacestaining was performed with corresponding fluorescently labeled Abs for 20min at 4°C; after this, the cells were permeabilized with Cytofix/Cytopermsolution for 20 min at 4°C and intracellular cytokine staining was performed
8062 THE NLRP3 INFLAMMASOME IN FUNGAL IMMUNITY

with fluorescently labeled cytokine Abs for 20 min (30, 34). The data wereacquired using a FACSCalibur (BD Biosciences) and analyzed using FlowJosoftware (Tree Star).
Statistical analysis
Statistical significance between groups was determined using a twotailed Student’s t test. Differences were considered significant when p �0.05, very significant when p � 0.01 and extremely significant whenp � 0.001.
ResultsNLRP3 inflammasome-mediated production of IL-1� by fungal�-glucans
We sought to examine cytokine production in response to �-glucan-containing agents such as curdlan and zymosan. Stimulation of thio-glycolate-elicited peritoneal macrophages (PEC) and BMDC withcurdlan or zymosan resulted in the production of IL-1� as determinedby ELISA (Fig. 1A). We found that stimulation with curdlan for 2 and3 h was not enough to induce IL-1� production while production wasinduced after 24 h of stimulation (Fig. 1B). Because IL-1� productionis regulated by pro-IL-1� synthesis and subsequent processing via theinflammasome, we assessed casapse-1 activation. Treatment ofBMDC with a caspase-1 inhibitor Ac-YVAD-CMK significantly ab-rogated curdlan-induced IL-1� production while it did not suppressIL-6 production (Fig. 1C). The mature form of IL-1� was detected inthe supernatant of BMDC stimulated with curdlan but not LPS (Fig.1D), suggesting that pro-IL-1� was processed to mature IL-1� inresponse to curdlan. Moreover, curdlan stimulation resulted incaspase-1 activation as detected by immunoblotting with an anti-caspase-1 p10 Ab (Fig. 1E).
The findings that curdlan and zymosan trigger IL-1� productionprompted us to investigate whether the NLRP3 inflammasome wasinvolved in this pathway. In this regard, we prepared BMDC fromwild-type (WT) and NLRP3-deficient mice and stimulated them withcurdlan or zymosan. The production of IL-1� after treatment withthese agents was markedly reduced in NLRP3-deficient BMDC,while IL-6 production was unimpaired (Fig. 1F). Similarly, reducedIL-1� production was found in PEC derived from NLRP3-deficientmice (supplementary Fig. 1A).6 In addition, IL-1� production byNaClO-oxidized zymosan, which activates cells through a TLR-inde-pendent, dectin-1/Syk-dependent mechanism, was also reduced inNLRP3-deficient PEC (supplementary Fig. 1B). Curdlan-inducedIL-1� production was also reduced in ASC-deficient cells (supple-mentary Fig. 1C) while the production was intact in NLRC4-deficientcells (supplementary Fig. 1D). Moreover, NLRP3 was dispensable forcurdlan-induced ERK1/2 phosphorylation and I�B� degradation(supplementary Fig. 2). Thus, the NLRP3 inflammasome componentsare required for curdlan- and zymosan-induced pro-IL-1� processingbut not MAPK and NF-�B activation.
As �-glucan is a major component of fungi, we next examinedcytokine production in response to C. albicans and S. cerevisiae. PECderived from WT mice infected with C. albicans or stimulated withS. cerevisiae released IL-1�, and this production was severely im-paired in cells derived from NLRP3-deficient and ASC-deficientmice (Fig. 1G).
6 The online version of this article contains supplemental material.
FIGURE 1. NLRP3 inflammasome-dependent IL-1� production by fungal �-glucans. A, BMDC or PEC from WT mice (n � 3) were stimulated for 24 hwith the indicated concentration of curdlan or zymosan, and the culture supernatants were analyzed for IL-1� production by ELISA. B, PEC from WT mice(n � 2) were stimulated with curdlan (3 mg/ml) for the indicated times (h) and the culture supernatants were analyzed for IL-1� production by ELISA.C, BMDC from WT mice untreated or pretreated with caspase-1 inhibitor, Ac-YVAD-CMK (YVAD) (50 or 100 mM) were stimulated with curdlan for24 h and IL-1� and IL-6 production was measured by ELISA. D and E, BMDC were untreated or stimulated with LPS (1 �g/ml) or curdlan (3 mg/ml)for 24 h. The cell lysates (Lys) and the culture supernatants (Sup) were analyzed by immunoblot analysis using anti-IL-1� Ab (D). The Sup from D werealso analyzed by immunoblot analysis using anti-caspase-1 p10 Ab (E). F, BMDC from NLRP3�/� or NLRP3�/� mice (n � 4) were stimulated with 1or 2 mg/ml curdlan or zymosan for 24 h, and IL-1� and IL-6 production was measured by ELISA. G, PEC from NLRP3�/� or NLRP3�/� mice (n � 4)or ASC�/� or ASC�/� (n � 3) were infected with C. albicans or S. cerevisiae (MOI � 50 or 100) for 24 h and IL-1� production was measured byELISA. N.D., not detected. Results are representative of two to four independent experiments.
8063The Journal of Immunology
Treatment of BMDC with Syk inhibitors abrogated curdlan-in-duced IL-1� mRNA induction and IL-1� production (supplemen-tary Fig. 3), suggesting that curdlan-induced IL-1� production iscontrolled by Syk-mediated synthesis of pro-IL-1� as well as theNLRP3 inflammasome-mediated processing of pro-IL-1�.
Mechanism of the NLRP3 inflammasome activation by curdlan
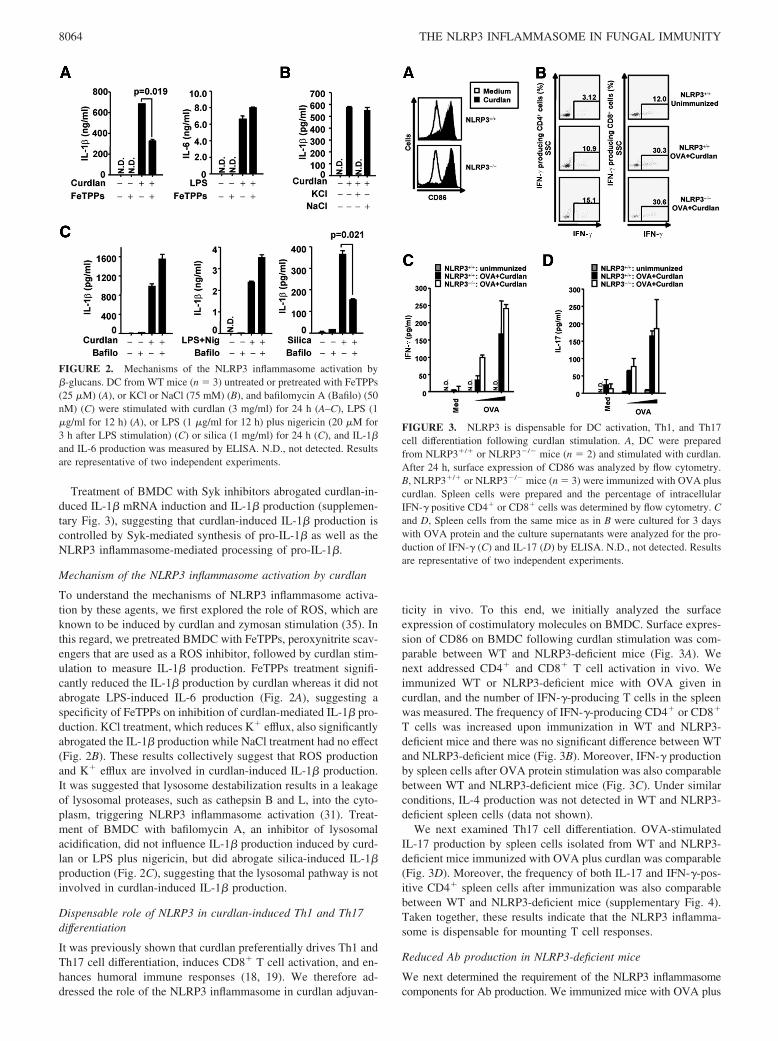
To understand the mechanisms of NLRP3 inflammasome activa-tion by these agents, we first explored the role of ROS, which areknown to be induced by curdlan and zymosan stimulation (35). Inthis regard, we pretreated BMDC with FeTPPs, peroxynitrite scav-engers that are used as a ROS inhibitor, followed by curdlan stim-ulation to measure IL-1� production. FeTPPs treatment signifi-cantly reduced the IL-1� production by curdlan whereas it did notabrogate LPS-induced IL-6 production (Fig. 2A), suggesting aspecificity of FeTPPs on inhibition of curdlan-mediated IL-1� pro-duction. KCl treatment, which reduces K� efflux, also significantlyabrogated the IL-1� production while NaCl treatment had no effect(Fig. 2B). These results collectively suggest that ROS productionand K� efflux are involved in curdlan-induced IL-1� production.It was suggested that lysosome destabilization results in a leakageof lysosomal proteases, such as cathepsin B and L, into the cyto-plasm, triggering NLRP3 inflammasome activation (31). Treat-ment of BMDC with bafilomycin A, an inhibitor of lysosomalacidification, did not influence IL-1� production induced by curd-lan or LPS plus nigericin, but did abrogate silica-induced IL-1�production (Fig. 2C), suggesting that the lysosomal pathway is notinvolved in curdlan-induced IL-1� production.
Dispensable role of NLRP3 in curdlan-induced Th1 and Th17differentiation
It was previously shown that curdlan preferentially drives Th1 andTh17 cell differentiation, induces CD8� T cell activation, and en-hances humoral immune responses (18, 19). We therefore ad-dressed the role of the NLRP3 inflammasome in curdlan adjuvan-
ticity in vivo. To this end, we initially analyzed the surfaceexpression of costimulatory molecules on BMDC. Surface expres-sion of CD86 on BMDC following curdlan stimulation was com-parable between WT and NLRP3-deficient mice (Fig. 3A). Wenext addressed CD4� and CD8� T cell activation in vivo. Weimmunized WT or NLRP3-deficient mice with OVA given incurdlan, and the number of IFN-�-producing T cells in the spleenwas measured. The frequency of IFN-�-producing CD4� or CD8�
T cells was increased upon immunization in WT and NLRP3-deficient mice and there was no significant difference between WTand NLRP3-deficient mice (Fig. 3B). Moreover, IFN-� productionby spleen cells after OVA protein stimulation was also comparablebetween WT and NLRP3-deficient mice (Fig. 3C). Under similarconditions, IL-4 production was not detected in WT and NLRP3-deficient spleen cells (data not shown).
We next examined Th17 cell differentiation. OVA-stimulatedIL-17 production by spleen cells isolated from WT and NLRP3-deficient mice immunized with OVA plus curdlan was comparable(Fig. 3D). Moreover, the frequency of both IL-17 and IFN-�-pos-itive CD4� spleen cells after immunization was also comparablebetween WT and NLRP3-deficient mice (supplementary Fig. 4).Taken together, these results indicate that the NLRP3 inflamma-some is dispensable for mounting T cell responses.
Reduced Ab production in NLRP3-deficient mice
We next determined the requirement of the NLRP3 inflammasomecomponents for Ab production. We immunized mice with OVA plus
FIGURE 2. Mechanisms of the NLRP3 inflammasome activation by�-glucans. DC from WT mice (n � 3) untreated or pretreated with FeTPPs(25 �M) (A), or KCl or NaCl (75 mM) (B), and bafilomycin A (Bafilo) (50nM) (C) were stimulated with curdlan (3 mg/ml) for 24 h (A–C), LPS (1�g/ml for 12 h) (A), or LPS (1 �g/ml for 12 h) plus nigericin (20 �M for3 h after LPS stimulation) (C) or silica (1 mg/ml) for 24 h (C), and IL-1�and IL-6 production was measured by ELISA. N.D., not detected. Resultsare representative of two independent experiments.
FIGURE 3. NLRP3 is dispensable for DC activation, Th1, and Th17cell differentiation following curdlan stimulation. A, DC were preparedfrom NLRP3�/� or NLRP3�/� mice (n � 2) and stimulated with curdlan.After 24 h, surface expression of CD86 was analyzed by flow cytometry.B, NLRP3�/� or NLRP3�/� mice (n � 3) were immunized with OVA pluscurdlan. Spleen cells were prepared and the percentage of intracellularIFN-� positive CD4� or CD8� cells was determined by flow cytometry. Cand D, Spleen cells from the same mice as in B were cultured for 3 dayswith OVA protein and the culture supernatants were analyzed for the pro-duction of IFN-� (C) and IL-17 (D) by ELISA. N.D., not detected. Resultsare representative of two independent experiments.
8064 THE NLRP3 INFLAMMASOME IN FUNGAL IMMUNITY

curdlan, and measured the concentration of serum Abs specific toOVA. Mice receiving OVA alone did not increase the titer of Ag-specific IgM, IgG1, IgG2b, IgG2c, and IgE (data not shown), but thetiters were increased when receiving OVA along with curdlan (Fig.4A). By contrast, NLRP3-deficient mice displayed a significant re-duction in Ab titers (Fig. 4A). Moreover, the titer of IgG1 was alsoconsiderably reduced in ASC-deficient mice (Fig. 4B). These resultsindicate that the NLRP3 inflammasome components are required forthe induction of Ab responses enhanced by curdlan. We next ad-dressed Ab responses in NLRP3-deficient mice that were immunizedwith OVA plus silica. Unlike curdlan, NLRP3-deficient mice receiv-ing silica produced similar amounts of Ag-specific IgG1 as WT mice(Fig. 4C). Furthermore, Ag-specific total IgG and IgG2b was alsocomparable (supplementary Fig. 5). Taken together, these results in-dicate that the NLRP3 inflammasome is required for efficient Ab pro-duction induced by curdlan but not silica, and suggest that curdlan andsilica have different capacities with regard to the induction of Abresponses.
NLRP3-dependent B cell activation in response to curdlan
The finding that Ab production was reduced in NLRP3-deficient micesuggests that curdlan is able to activate B cells. We therefore exam-ined the expression of NLRP3 in B cells. NLRP3 mRNA was de-tected in resting CD19� splenic cells, but it was weak compared withthat in PEC (Fig. 5A) consistent with a previous report (36). However,curdlan stimulation resulted in up-regulation of NLRP3 mRNA to asimilar level as PEC (Fig. 5A), indicating that NLRP3 expression isinducible in B cells. Next, we investigated whether or not curdlanactivates Syk in B cells. We purified splenic B cells and Syk phos-phorylation was examined by immunoblot analysis using an anti-phospho Syk Ab. Curdlan induced Syk phosphorylation in both WTand NLRP3-deficient B cells (Fig. 5B). Moreover, Syk activation wasalso observed in a B cell line (supplementary Fig. 6), excluding thepossibility that the effects were due to contaminating cells in thesplenic B cells.
We next examined the activation of B cells following curdlan stim-ulation. Stimulation of purified B cells with curdlan resulted in up-regulation of CD69 on the cell surface, and this induction was abro-gated by treatment with Syk inhibitors (Fig. 5C). By contrast, alum
and silica stimulation did not enhance CD69 expression on B cells(Fig. 5D). Notably, B cells purified from NLRP3-deficient mice failedto up-regulate CD69 in response to curdlan, whereas they normallyup-regulated CD69 after treatment with LPS (Fig. 5E). Next, we mea-sured Ig production by B cells in WT and NLRP3-deficient mice.When purified B cells were stimulated with curdlan they producedIgM, which was markedly reduced by NLRP3 deficiency (Fig. 5F). Incontrast, IgM production by B cells in response to LPS was compa-rable between WT and NLRP3-deficient mice (Fig. 5F). To under-stand the involvement of IL-1 family cytokines in curdlan-induced Abproduction, we stimulated the purified B cells from WT and MyD88-deficient mice (in which IL-1 receptor signaling is abrogated) withcurdlan. The IgM production was comparable in both WT andMyD88-deficient B cells, whereas LPS-induced IgM production wasabrogated in MyD88-deficient B cells (Fig. 5G), suggesting that IL-1family cytokines are unlikely to be involved in B cell activation.
DiscussionIn this study, we demonstrated a critical role for the NLRP3 inflam-masome in innate and humoral adaptive immune responses inducedby fungus �-glucan. It was reported that curdlan is recognized bydectin-1, activating NF-�B and MAPKs via Syk for the induction ofinflammatory cytokine genes (10, 13, 14). In contrast, zymosan isrecognized by dectin-1 in collaboration with TLR2 (13, 14). Wefound that cells deficient in NLRP3 or ASC abrogated IL-1� produc-tion but induced normal IL-6 production in response to curdlan andzymosan. Therefore, curdlan and zymosan are capable of promotingpro-IL-1� synthesis as well as NLRP3 inflammasome-dependent pro-cessing of pro-IL-1�. Several mechanisms for NLRP3 inflammasomeactivation have been reported. We found that IL-1� production bycurdlan was suppressed in the presence of a ROS inhibitor. Further-more, it was reported that Syk activation is required for curdlan-in-duced ROS production (35), suggesting that Syk controls gene ex-pression as well as NLRP3 inflammasome activation. We also foundthat IL-1� production was suppressed by blocking K� efflux. It wasshown that efflux of K� is promoted by ROS (37). Thus, Syk-depen-dent ROS production may lead to K� efflux, and these intermediatesrobustly activate the NLRP3 inflammasome. Bafilomycin A, a drugused for the inhibition of H�-ATPase in the lysosome, subsequently
FIGURE 4. Reduced Ab produc-tion after curdlan administration inNLRP3- and ASC-deficient mice. Aand B, NLRP3�/� or NLRP3�/�
mice (n � 5) (A) or ASC�/� orASC�/� mice (n � 3) (B) were im-munized twice (on day 0 and 7) withOVA plus curdlan. Seven days afterthe last immunization, sera were col-lected and analyzed for OVA-specificIgM, IgG1, IgG2b, IgG2c, and IgE(A) or IgG1 (B) by ELISA. C,NLRP3�/� or NLRP3�/� mice (n �3) were immunized twice (on day 0and 7) with OVA plus silica. Sevenand fourteen days after the last immu-nization, sera were collected and an-alyzed for OVA-specific IgG1 byELISA. wk, weeks.
8065The Journal of Immunology

inhibiting their acidification, did not suppress curdlan-induced IL-1�production, suggesting that lysosome destabilization is unlikely to beinvolved in �-glucan-induced IL-1� production.
It was reported that curdlan drives Ag-specific Th1 and Th17 celldifferentiation through Syk (18). NLRP3-deficient mice displayednormal Th1 and Th17 cell differentiation when curdlan was receivedalong with OVA. In NLRP3-deficient DC, curdlan-induced surfaceexpression of CD86 and production of inflammatory cytokines, suchas IL-6, are intact, suggesting that these responses are sufficient formounting T cell-mediated immunity. Notably, NLRP3-deficient micehave defects in Ag-specific Ab responses after administration of curd-lan along with OVA. This suggests that curdlan directly influences Bcell activation that is linked to Ab production. We found that stimu-lation of purified B cells with curdlan induced Syk activation thatresulted in CD69 up-regulation and IgM production. It is noted thatthese responses were abrogated by NLRP3 deficiency. Thus, intrinsicNLRP3 may play a critical role in B cell activation and enhancementof Ab responses.
We also found that purified B cells failed to up-regulate CD69when stimulated with other NLRP3 inflammasome activators, such assilica and alum. Furthermore, production of Ag-specific Abs wascomparable in WT and NLRP3-deficient mice when silica was ad-ministrated as an adjuvant. These observations suggest that curdlanand silica have different capacities to mount Ab responses althoughboth can activate NLRP3. In these regards, it is speculated that curd-lan-induced IL-1� participates in B cell activation. Stimulation of Bcells with curdlan induced IL-1� production while stimulation withsilica and alum did not, and this production was markedly suppressedby treatment with Syk inhibitors (supplementary Fig. 7). This sug-gests the possibility that the Syk-NLRP3-IL-1� axis plays a role in Bcell activation. However, it is noteworthy that MyD88-deficient Bcells show normal B cell activation following curdlan treatment, sug-gesting that IL-1 receptor signaling is not required for this activation.
Thus, other cytokines or functional molecules downstream to Syk-NLRP3 are likely to mediate B cell activation by curdlan.
It was previously reported that the production of Ag-specificAbs and Th2 cell priming following immunization of Ag given inalum was abrogated in mice lacking a component of the NLRP3inflammasome, although the requirement of NLRP3 in adjuvanteffects of alum is still controversial (5–8). By contrast, NLRP3 isdispensable for curdlan-induced Th1 and Th17 cell responses, andcurdlan but not alum is able to activate Syk in B cells. Thesefindings strongly suggest that induction mechanisms of adaptiveimmune responses are different between alum and curdlan. It willtherefore be of interest to study these differences in the future.
Recently, three reports have shown that the NLRP3 inflammasomeplays an essential role in host defense against fungal infection (38–40). Gross et al. (38) demonstrated that C. albicans infection resultedin IL-1� production through the NLRP3 inflammasome. Theyshowed that Syk and CARD9 regulate pro-IL-1� synthesis as well asNLRP3 inflammasome activation. ROS and K� efflux, but not lyso-somal destabilization, were involved in NLRP3 inflammasome acti-vation, which was consistent with our findings. Furthermore, theyshowed that NLRP3 was essential in host defense against C. albicansinfection. Hise et al. (39) also showed the importance of the NLRP3inflammasome in C. albicans-induced IL-1� production. They dem-onstrated that dectin-1-medaited C. albicans recognition is requiredfor NLPR3 inflammasome activation. In this study, we provide evi-dence that �-glucan, a major component of C. albicans, promotesNLRP3 inflammasome activation. Altogether, these studies stronglysuggest that fungal �-glucans primarily engage with dectin-1, whichsignals through Syk and CARD9 to induce pro-IL-1� synthesis aswell as NLRP3 inflammasome activation.
More recently, Lamkanfi et al. (41) have shown that zymosanin conjunction with ATP triggers NLRP3-dependent IL-1�production. They showed that treatment of cells with zymosan for
FIGURE 5. Curdlan induces B cell activation through NLRP3. A, Total RNA was isolated from purified CD19� B cells, unstimulated or stimulated withcurdlan (3 mg/ml) for 2 h, or unstimulated PEC were subjected to RT-PCR analysis for NLRP3 and �-actin expression. B, Purified CD19� B cells fromNLRP3�/� or NLRP3�/� mice were stimulated for the indicated times (min) with curdlan (3 mg/ml) and subjected to immunoblot analysis usinganti-phospho Syk Ab (anti-pSyk). C and D, Purified CD19� B cells from WT mice were untreated or pretreated with piceatannol (pice) (2.5 �g/ml) (C)and then stimulated with curdlan (3 mg/ml) (C), alum (1 mg/ml) (D), or silica (1 mg/ml) (D) for 24 h and analyzed for surface expression of CD69 by flowcytometry. E, Purified CD19� B cells from NLRP3�/� or NLRP3�/� mice were stimulated with curdlan (3 mg/ml) or LPS (1 �g/ml) for 24 h and analyzed forsurface expression of CD69 by flow cytometry. F, Purified B cells were stimulated with curdlan (3 mg/ml) or LPS (20 �g/ml) for 3 days and the culturesupernatants were analyzed for total IgM production by ELISA. G, Purified B cells from MyD88�/� and MyD88�/� were stimulated with curdlan or LPS for 3days and the culture supernatants were analyzed for total IgM production by ELISA. N.D., not detected. Results are representative of two independent experiments.
8066 THE NLRP3 INFLAMMASOME IN FUNGAL IMMUNITY

4 h was insufficient to produce IL-1�, but subsequent ATP stim-ulation drove NLRP3 inflammasome-dependent IL-1� production.We demonstrated in this study that stimulation with curdlan or zy-mosan is sufficient to drive the maturation of IL-1�, but this produc-tion required 24 h, not 3 h, stimulation, which contrasted markedly toother NLRP3 inflammasome activators, such as LPS plus ATP, whichinduce IL-1� production within 1 h (27). Thus, zymosan- and curd-lan-induced modifications that occur within 24 h may participate intriggering NLRP3 inflammasome activation.
In conclusion, our results demonstrated that the NLRP3 inflam-masome controls IL-1� production triggered by �-glucan, and thispathway is required for innate responses as well as B cell-mediatedhumoral immunity. Therefore, manipulation of the NLRP3 inflam-masome-mediated pathway is a potential target that could be usedto enhance B cell-mediated immunity.
AcknowledgmentsWe thank Dr. V. M. Dixit (Genentech) for providing NLRP3/cryopyrin/NALP3-, NLRC4/IPAF- and ASC-deficient mice and Drs.K. Maeda and N. Sakaguchi for providing B cell line. We also thank Drs.O. Takeuchi, K.J. Ishii, S. Uematsu, and C. Coban for helpful discussions,C. Funamoto, N. Kitagaki, Y. Fujiwara, and M. Kumagai for technicalassistance and E. Kamada for secretarial assistance.
DisclosuresThe authors have no financial conflict of interest.
References1. Akira, S., S. Uematsu, and O. Takeuchi. 2006. Pathogen recognition and innate
immunity. Cell 124: 783–801.2. Franchi, L., T. Eigenbrod, R. Munoz-Planillo, and G. Nunez. 2009. The inflam-
masome: a caspase-1-activation platform that regulates immune responses anddisease pathogenesis. Nat. Immunol. 10: 241–247.
3. Martinon, F., A. Mayor, and J. Tschopp. 2009. The inflammasomes: guardians ofthe body. Annu. Rev. Immunol. 27: 229–265.
4. Ye, Z., and J. P. Ting. 2008. NLR, the nucleotide-binding domain leucine-richrepeat containing gene family. Curr. Opin. Immunol. 20: 3–9.
5. Eisenbarth, S. C., O. R. Colegio, W. O’Connor, F. S. Sutterwala, andR. A. Flavell. 2008. Crucial role for the Nalp3 inflammasome in the immunos-timulatory properties of aluminium adjuvants. Nature 453: 1122–1126.
6. Kool, M., V. Petrilli, T. De Smedt, A. Rolaz, H. Hammad, M. van Nimwegen,I. M. Bergen, R. Castillo, B. N. Lambrecht, and J. Tschopp. 2008. Cutting edge:alum adjuvant stimulates inflammatory dendritic cells through activation of theNALP3 inflammasome. J. Immunol. 181: 3755–3759.
7. Li, H., S. B. Willingham, J. P. Ting, and F. Re. 2008. Cutting edge: inflamma-some activation by alum and alum’s adjuvant effect are mediated by NLRP3.J. Immunol. 181: 17–21.
8. Franchi, L., and G. Nunez. 2008. The Nlrp3 inflammasome is critical for alumi-nium hydroxide-mediated IL-1� secretion but dispensable for adjuvant activity.Eur. J. Immunol. 38: 2085–2089.
9. Sharp, F. A., D. Ruane, B. Claass, E. Creagh, J. Harris, P. Malyala, M. Singh,D. T. O’Hagan, V. Petrilli, J. Tschopp, et al. 2009. Uptake of particulate vaccineadjuvants by dendritic cells activates the NALP3 inflammasome. Proc. Natl.Acad. Sci. USA 106: 870–875.
10. Tsoni, S. V., and G. D. Brown. 2008. �-glucans and dectin-1. Ann. NY Acad. Sci.1143: 45–60.
11. Ariizumi, K., G. L. Shen, S. Shikano, S. Xu, R. Ritter, 3rd, T. Kumamoto,D. Edelbaum, A. Morita, P. R. Bergstresser, and A. Takashima. 2000. Identifi-cation of a novel, dendritic cell-associated molecule, dectin-1, by subtractivecDNA cloning. J. Biol. Chem. 275: 20157–20167.
12. Brown, G. D., and S. Gordon. 2001. Immune recognition: a new receptor for�-glucans. Nature 413: 36–37.
13. Underhill, D. M. 2007. Collaboration between the innate immune receptors dec-tin-1, TLRs, and Nods. Immunol. Rev. 219: 75–87.
14. Reid, D. M., N. A. Gow, and G. D. Brown. 2009. Pattern recognition: recentinsights from dectin-1. Curr. Opin. Immunol. 21: 30–37.
15. Rogers, N. C., E. C. Slack, A. D. Edwards, M. A. Nolte, O. Schulz,E. Schweighoffer, D. L. Williams, S. Gordon, V. L. Tybulewicz, G. D. Brown,and C. Reis e Sousa. 2005. Syk-dependent cytokine induction by dectin-1 revealsa novel pattern recognition pathway for C type lectins. Immunity 22: 507–517.
16. Hara, H., C. Ishihara, A. Takeuchi, T. Imanishi, L. Xue, S. W. Morris, M. Inui,T. Takai, A. Shibuya, S. Saijo, et al. 2007. The adaptor protein CARD9 is es-sential for the activation of myeloid cells through ITAM-associated and Toll-likereceptors. Nat. Immunol. 8: 619–629.
17. Gross, O., A. Gewies, K. Finger, M. Schafer, T. Sparwasser, C. Peschel,I. Forster, and J. Ruland. 2006. Card9 controls a non-TLR signalling pathway forinnate anti-fungal immunity. Nature 442: 651–656.
18. LeibundGut-Landmann, S., O. Gross, M. J. Robinson, F. Osorio, E. C. Slack,S. V. Tsoni, E. Schweighoffer, V. Tybulewicz, G. D. Brown, J. Ruland, andC. Reis e Sousa. 2007. Syk- and CARD9-dependent coupling of innate immunityto the induction of T helper cells that produce interleukin 17. Nat. Immunol 8:630–638.
19. Leibundgut-Landmann, S., F. Osorio, G. D. Brown, and C. Reis e Sousa. 2008.Stimulation of dendritic cells via the dectin-1/Syk pathway allows priming ofcytotoxic T-cell responses. Blood 112: 4971–4980.
20. Taylor, P. R., S. V. Tsoni, J. A. Willment, K. M. Dennehy, M. Rosas, H. Findon,K. Haynes, C. Steele, M. Botto, S. Gordon, and G. D. Brown. 2007. Dectin-1 isrequired for �-glucan recognition and control of fungal infection. Nat. Immunol. 8:31–38.
21. Saijo, S., N. Fujikado, T. Furuta, S. H. Chung, H. Kotaki, K. Seki, K. Sudo, S. Akira,Y. Adachi, N. Ohno, et al. 2007. Dectin-1 is required for host defense against Pneu-mocystis carinii but not against Candida albicans. Nat. Immunol. 8: 39–46.
22. Sato, K., X. L. Yang, T. Yudate, J. S. Chung, J. Wu, K. Luby-Phelps,R. P. Kimberly, D. Underhill, P. D. Cruz, Jr., and K. Ariizumi. 2006. Dectin-2 isa pattern recognition receptor for fungi that couples with the Fc receptor � chainto induce innate immune responses. J. Biol. Chem. 281: 38854–38866.
23. Wells, C. A., J. A. Salvage-Jones, X. Li, K. Hitchens, S. Butcher, R. Z. Murray,A. G. Beckhouse, Y. L. Lo, S. Manzanero, C. Cobbold, et al. 2008. The mac-rophage-inducible C-type lectin, mincle, is an essential component of the innateimmune response to Candida albicans. J. Immunol. 180: 7404–7413.
24. Yamasaki, S., M. Matsumoto, O. Takeuchi, T. Matsuzawa, E. Ishikawa,M. Sakuma, H. Tateno, J. Uno, J. Hirabayashi, Y. Mikami, et al. 2009. C-typelectin Mincle is an activating receptor for pathogenic fungus, Malassezia. Proc.Natl. Acad. Sci. USA 106: 1897–1902.
25. Means, T. K., E. Mylonakis, E. Tampakakis, R. A. Colvin, E. Seung, L. Puckett,M. F. Tai, C. R. Stewart, R. Pukkila-Worley, S. E. Hickman, et al. 2009. Evo-lutionarily conserved recognition and innate immunity to fungal pathogens by thescavenger receptors SCARF1 and CD36. J. Exp. Med. 206: 637–653.
26. Zimmerman, J. W., J. Lindermuth, P. A. Fish, G. P. Palace, T. T. Stevenson, andD. E. DeMong. 1998. A novel carbohydrate-glycosphingolipid interaction be-tween a �-(1–3)-glucan immunomodulator, PGG-glucan, and lactosylceramide ofhuman leukocytes. J. Biol. Chem. 273: 22014–22020.
27. Mariathasan, S., D. S. Weiss, K. Newton, J. McBride, K. O’Rourke,M. Roose-Girma, W. P. Lee, Y. Weinrauch, D. M. Monack, and V. M. Dixit.2006. Cryopyrin activates the inflammasome in response to toxins and ATP.Nature 440: 228–232.
28. Mariathasan, S., K. Newton, D. M. Monack, D. Vucic, D. M. French, W. P. Lee,M. Roose-Girma, S. Erickson, and V. M. Dixit. 2004. Differential activation ofthe inflammasome by caspase-1 adaptors ASC and Ipaf. Nature 430: 213–218.
29. Adachi, O., T. Kawai, K. Takeda, M. Matsumoto, H. Tsutsui, M. Sakagami,K. Nakanishi, and S. Akira. 1998. Targeted disruption of the MyD88 gene resultsin loss of IL-1- and IL-18-mediated function. Immunity 9: 143–150.
30. Kumar, H., T. Kawai, H. Kato, S. Sato, K. Takahashi, C. Coban, M. Yamamoto,S. Uematsu, K. J. Ishii, O. Takeuchi, and S. Akira. 2006. Essential role of IPS-1in innate immune responses against RNA viruses. J. Exp. Med. 203: 1795–1803.
31. Hornung, V., F. Bauernfeind, A. Halle, E. O. Samstad, H. Kono, K. L. Rock,K. A. Fitzgerald, and E. Latz. 2008. Silica crystals and aluminum salts activate theNALP3 inflammasome through phagosomal destabilization. Nat. Immunol. 9:847–856.
32. Koyama, S., K. J. Ishii, H. Kumar, T. Tanimoto, C. Coban, S. Uematsu, T. Kawai,and S. Akira. 2007. Differential role of TLR- and RLR-signaling in the immuneresponses to influenza A virus infection and vaccination. J. Immunol. 179:4711–4720.
33. Kumar, H., S. Koyama, K. J. Ishii, T. Kawai, and S. Akira. 2008. Cutting edge:cooperation of IPS-1- and TRIF-dependent pathways in poly IC-enhanced anti-body production and cytotoxic T cell responses. J. Immunol. 180: 683–687.
34. Uematsu, S., K. Fujimoto, M. H. Jang, B. G. Yang, Y. J. Jung, M. Nishiyama,S. Sato, T. Tsujimura, M. Yamamoto, Y. Yokota, et al. 2008. Regulation ofhumoral and cellular gut immunity by lamina propria dendritic cells expressingToll-like receptor 5. Nat. Immunol. 9: 769–776.
35. Underhill, D. M., E. Rossnagle, C. A. Lowell, and R. M. Simmons. 2005. Dec-tin-1 activates Syk tyrosine kinase in a dynamic subset of macrophages for re-active oxygen production. Blood 106: 2543–2550.
36. Sutterwala, F. S., Y. Ogura, M. Szczepanik, M. Lara-Tejero, G. S. Lichtenberger,E. P. Grant, J. Bertin, A. J. Coyle, J. E. Galan, P. W. Askenase, and R. A. Flavell.2006. Critical role for NALP3/CIAS1/Cryopyrin in innate and adaptive immunitythrough its regulation of caspase-1. Immunity 24: 317–327.
37. Rada, B. K., M. Geiszt, K. Kaldi, C. Timar, and E. Ligeti. 2004. Dual role ofphagocytic NADPH oxidase in bacterial killing. Blood 104: 2947–2953.
38. Gross, O., H. Poeck, M. Bscheider, C. Dostert, N. Hannesschlager, S. Endres,G. Hartmann, A. Tardivel, E. Schweighoffer, V. Tybulewicz, et al. 2009. Sykkinase signalling couples to the Nlrp3 inflammasome for anti-fungal host de-fence. Nature 459: 433–436.
39. Hise, A. G., J. Tomalka, S. Ganesan, K. Patel, B. A. Hall, G. D. Brown, andK. A. Fitzgerald. 2009. An essential role for the NLRP3 inflammasome in hostdefense against the human fungal pathogen Candida albicans. Cell Host Mi-crobes 5: 487–497.
40. Joly, S., N. Ma, J. J. Sadler, D. R. Soll, S. L. Cassel, and F. S. Sutterwala. 2009.Cutting edge: Candida albicans hyphae formation triggers activation of the Nlrp3inflammasome. J. Immunol. 183: 3578–3581.
41. Lamkanfi, M., R. K. Malireddi, and T. D. Kanneganti. 2009. Fungal zymosan andmannan activate the cryopyrin inflammasome. J. Biol. Chem. 284: 20574–20581.
8067The Journal of Immunology




















