
SuperSAGE: the drought stress-responsive transcriptome of chickpea roots

Arch Microbiol (2012) 194:473–482
DOI 10.1007/s00203-011-0782-xORIGINAL PAPER
Transcriptome analysis of pellicle formation of Shewanella oneidensis
Yili Liang · Haichun Gao · Xue Guo · Jingrong Chen · Guanzhou Qiu · Zhili He · Jizhong Zhou · Xueduan Liu
Received: 16 November 2011 / Revised: 8 December 2011 / Accepted: 12 December 2011 / Published online: 7 January 2012© Springer-Verlag 2012
Abstract Although the pellicle is one of the major growthmodes of microorganisms, the metabolic features of pelliclecells and the determinative factors for pellicle formation arelargely unknown. In recent years, bioWlm development ofShewanella oneidensis, an important model organism forbioremediation studies, has been extensively studied. Inthis paper, a transcriptional proWling of pellicle cells rela-tive to planktonic cells indicated that cells in pellicles weremore metabolically active than the planktonic cells. Mostnotably, up-transcription of general secretion system pro-teins and iron/heme uptake and transport proteins wasobserved in pellicle cells. Unexpectedly, neither the hmuTnor hugA heme transport mutant exhibited a signiWcantdefect in pellicle formation. Expectedly, three type I secre-tion system mutants were severely deWcient in pellicle for-mation, suggesting an essential role of these proteins.
Keywords Pellicle · Shewanella oneidensis · Type I secretion system
Introduction
The air–liquid interface is a selectively advantageous nichefor aerobes due to the accessibility to oxygen. Various spe-cies of aerobes (e.g. Bacillus subtilis and Pseudomonas Xuo-rescens) form a bioWlm-like structure, pellicle, at air–liquidinterface (Kobayashi 2007; Solano et al. 2002; Spiers et al.2003). In recent years, the subject of microbial pellicles hasdrawn a lion share of attention since it is easily studied andadaptable for mutant screens (Guvener and McCarter 2003).
As a special form of bioWlms, pellicles might be com-posed of bacterial cells embedded in a matrix formed byextracellular polymeric substances (EPS), which consist ofproteins, polysaccharide, humic acid, and DNA (Whitchurchet al. 2002). Secreted proteins and exopolysaccharide pro-ductions profoundly inXuenced pellicle development in anumber of organisms (Guvener and McCarter 2003; Kuma-gai et al. 2011). Transposon insertion mutants of Vibrioparahaemolyticus were obtained with defects in genes aVect-ing multiple cell surface characteristics, including extracellu-lar polysaccharide, mannose-sensitive hemagglutinin type 4pili and secreted proteins or transporters of secreted proteins(Enos-Berlage et al. 2005). FLO11 of yeast is essential forunfavorable pellicle formation of wild pellicle-formingyeasts by mechanisms other than increasing cell surfacehydrophobicity (Nakagawa et al. 2011). Pellicle formation byspore-forming bacterium Bacillus subtilis follows a distinctdevelopmental pathway. In addition to the EPS component ofthe matrix, three proteins, encoded in the three-gene operonyqxM-sipW-tasA, were identiWed as involved in matrixassembly in a transposon mutant screen for genes involved in
Communicated by Shuang-Jiang Liu.
Electronic supplementary material The online version of this article (doi:10.1007/s00203-011-0782-x) contains supplementary material, which is available to authorized users.
Y. Liang · X. Guo · G. Qiu · X. Liu (&)School of Minerals Processing and Bioengineering, Central South University, Changsha 410083, Chinae-mail: [email protected]
Y. Liang · H. Gao · J. Chen · Z. He · J. ZhouInstitute for Environmental Genomics and Department of Botany and Microbiology, University of Oklahoma, Norman, OK 73019, USA
H. GaoCollege of Life Sciences, Zhejiang University, Hangzhou, Zhejiang 310029, People’s Republic of China
123

474 Arch Microbiol (2012) 194:473–482
bioWlm formation (Branda et al. 2004). A mutant strain ofPseudomonas aeruginosa deWcient in the production of exo-polysaccharides displayed poor pellicle-forming ability and agrowth disadvantage under static conditions compared withthe wild-type strain (Yamamoto et al. 2011). In the presenceof cellulase, glucose and cellobiose were released from thepellicle of Asaia bogorensis AJ strain, suggesting that it ismade of cellulose (Kumagai et al. 2011).
Shewanella oneidensis MR-1, a facultative gram-negativeanaerobe with a remarkable respiratory versatility, is anatural pellicle former. Because of the potential use of thisorganism for the bioremediation of metal/radionuclide con-taminants in the environment, S. oneidensis MR-1 bioWlmhas been extensively studied, including pellicle development(Bagge et al. 2001; Vriendt et al. 2005; Windt et al. 2006;Teal et al. 2006; Thormann et al. 2004, 2005; Liang et al.2010). In order to characterize features of the pellicle life-style, a microarray analysis on pellicle and planktonic cells ofS. oneidensis was performed. The results revealed that thepellicle cells were more active in aerobic metabolism. Theplanktonic cells appeared to be under anaerobic or microaer-obic conditions once pellicle formation began. Genesinvolved in iron/heme uptake and transport or the generalsecretion pathway were up-regulated in pellicle cells.
Materials and methods
Bacterial strains, plasmids, and culture conditions
Bacterial strains and plasmids used in this study are listedin Table 1. Escherichia coli and S. oneidensis strains wereroutinely grown in Luria–Bertani (LB) broth or on LBplates at 37°C and the room temperature for genetic manip-ulation, respectively. When needed, antibiotics were used atthe following concentrations: ampicillin at 50 �g/ml andgentamycin at 15 �g/ml.
Pellicle formation, measurement of growth, and quantiWcation of pellicles
A fresh colony grown overnight on a LB plate was used toinoculate 50 ml LB and incubated in a shaker (200 rpm) to anOD600 of 0.8 at the room temperature. This culture was thendiluted 500-fold with fresh LB, resulting in the starting cul-tures. Aliquots of 30 ml starting cultures were transferred to50-ml Pyrex beakers. The beakers were kept still for pellicleformation at the room temperature. Cells in pellicle andunderneath were separated based on published paper (Lianget al. 2010). To quantify the pellicles formed by the S. oneid-ensis wild-type and mutant strains, cells from pellicles werecollected, suspended in 30 ml fresh LB, violently vortexed,and applied to the spectrometer at 600 nm.
Microarray experiments, data analysis and validation
For microarray analysis, aliquots of 30 ml starting cultureswere transferred to 50-ml Pyrex beakers and allowed todevelop pellicles at the room temperature. When a com-plete but thin layer of pellicles (young pellicles) at the inter-face were formed (about 30 h), the planktonic cultures andpellicles were separated as described above and applied tocentrifugation at 8,000 rpm for 3 min at room temperature.The resulting pellets were frozen immediately in liquidnitrogen and stored at ¡80°C if not immediately used. Con-struction of whole-genome DNA microarrays, total RNAextraction, cDNA labeling, hybridization, and slide scan-ning were conducted according to the standard procedure(Gao et al. 2004). LOWESS was used to normalize the dataset which subsequently was applied to statistical analysisby analysis of variance (ANOVA) with Benjamini andHochberg False Discovery Rate as multiple testing correc-tion (Gao et al. 2004, 2008). Raw microarray data weredeposited to GEO with the accession number GSE14331.
Quantitative RT-PCR (Q-RT-PCR)
Q-RT-PCR was used to measure the mRNA expressionlevels of four target genes in pellicle and planktonic cells.Primers for chosen genes were listed in Table 1S.
Table 1 Strains and plasmids used in this study
Strain or plasmid
Description Reference or source
E. coli
WM3064 Donor strain for conjugation; �dapA
Lab stock
TOP10 F¡ mcrA � (mrr-hsdRMS-mcrBC) �80lacZ� M15 � lacX74 deoR recA1 araD139
Invitrogen
� (ara-leu)7697 galU galK rpsL (Smr) endA1 nupG
S. oneidensis strains
MR-1 Wild-type Lab stock
JZ3669K hugA deletion mutant derived from MR-1; �hugA
This study
JZ3673K hmuT deletion mutant derived from MR-1; �hmuT
This study
JZ4318K rtxB deletion mutant derived from MR-1; �rtxB
This study
JZ4319K emrA deletion mutant derived from MR-1; �emrA
This study
JZ4320K aggA deletion mutant derived from MR-1; �aggA
Liang et al.2010
Plasmids
pDS3.0 Apr, Gmr, derivative from suicide vector pCVD442
Gao et al. (2006)
123
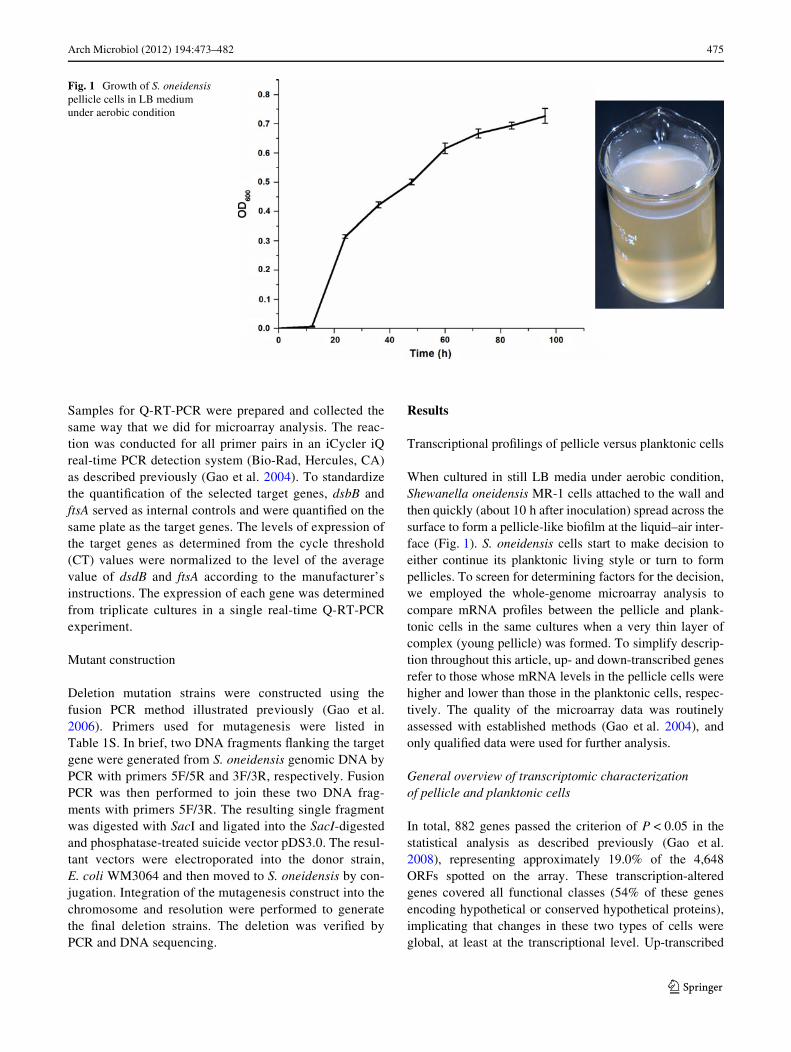
Arch Microbiol (2012) 194:473–482 475
Samples for Q-RT-PCR were prepared and collected thesame way that we did for microarray analysis. The reac-tion was conducted for all primer pairs in an iCycler iQreal-time PCR detection system (Bio-Rad, Hercules, CA)as described previously (Gao et al. 2004). To standardizethe quantiWcation of the selected target genes, dsbB andftsA served as internal controls and were quantiWed on thesame plate as the target genes. The levels of expression ofthe target genes as determined from the cycle threshold(CT) values were normalized to the level of the averagevalue of dsdB and ftsA according to the manufacturer’sinstructions. The expression of each gene was determinedfrom triplicate cultures in a single real-time Q-RT-PCRexperiment.
Mutant construction
Deletion mutation strains were constructed using thefusion PCR method illustrated previously (Gao et al.2006). Primers used for mutagenesis were listed inTable 1S. In brief, two DNA fragments Xanking the targetgene were generated from S. oneidensis genomic DNA byPCR with primers 5F/5R and 3F/3R, respectively. FusionPCR was then performed to join these two DNA frag-ments with primers 5F/3R. The resulting single fragmentwas digested with SacI and ligated into the SacI-digestedand phosphatase-treated suicide vector pDS3.0. The resul-tant vectors were electroporated into the donor strain,E. coli WM3064 and then moved to S. oneidensis by con-jugation. Integration of the mutagenesis construct into thechromosome and resolution were performed to generatethe Wnal deletion strains. The deletion was veriWed byPCR and DNA sequencing.
Results
Transcriptional proWlings of pellicle versus planktonic cells
When cultured in still LB media under aerobic condition,Shewanella oneidensis MR-1 cells attached to the wall andthen quickly (about 10 h after inoculation) spread across thesurface to form a pellicle-like bioWlm at the liquid–air inter-face (Fig. 1). S. oneidensis cells start to make decision toeither continue its planktonic living style or turn to formpellicles. To screen for determining factors for the decision,we employed the whole-genome microarray analysis tocompare mRNA proWles between the pellicle and plank-tonic cells in the same cultures when a very thin layer ofcomplex (young pellicle) was formed. To simplify descrip-tion throughout this article, up- and down-transcribed genesrefer to those whose mRNA levels in the pellicle cells werehigher and lower than those in the planktonic cells, respec-tively. The quality of the microarray data was routinelyassessed with established methods (Gao et al. 2004), andonly qualiWed data were used for further analysis.
General overview of transcriptomic characterization of pellicle and planktonic cells
In total, 882 genes passed the criterion of P < 0.05 in thestatistical analysis as described previously (Gao et al.2008), representing approximately 19.0% of the 4,648ORFs spotted on the array. These transcription-alteredgenes covered all functional classes (54% of these genesencoding hypothetical or conserved hypothetical proteins),implicating that changes in these two types of cells wereglobal, at least at the transcriptional level. Up-transcribed
Fig. 1 Growth of S. oneidensis pellicle cells in LB medium under aerobic condition
123
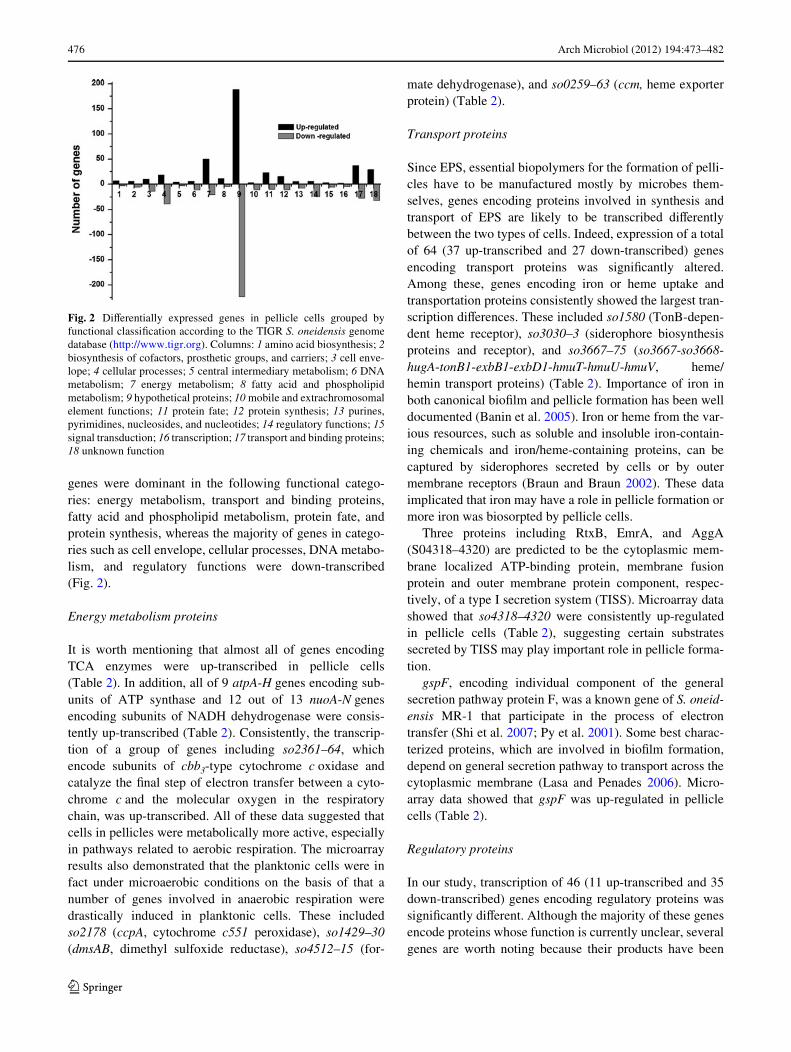
476 Arch Microbiol (2012) 194:473–482
genes were dominant in the following functional catego-ries: energy metabolism, transport and binding proteins,fatty acid and phospholipid metabolism, protein fate, andprotein synthesis, whereas the majority of genes in catego-ries such as cell envelope, cellular processes, DNA metabo-lism, and regulatory functions were down-transcribed(Fig. 2).
Energy metabolism proteins
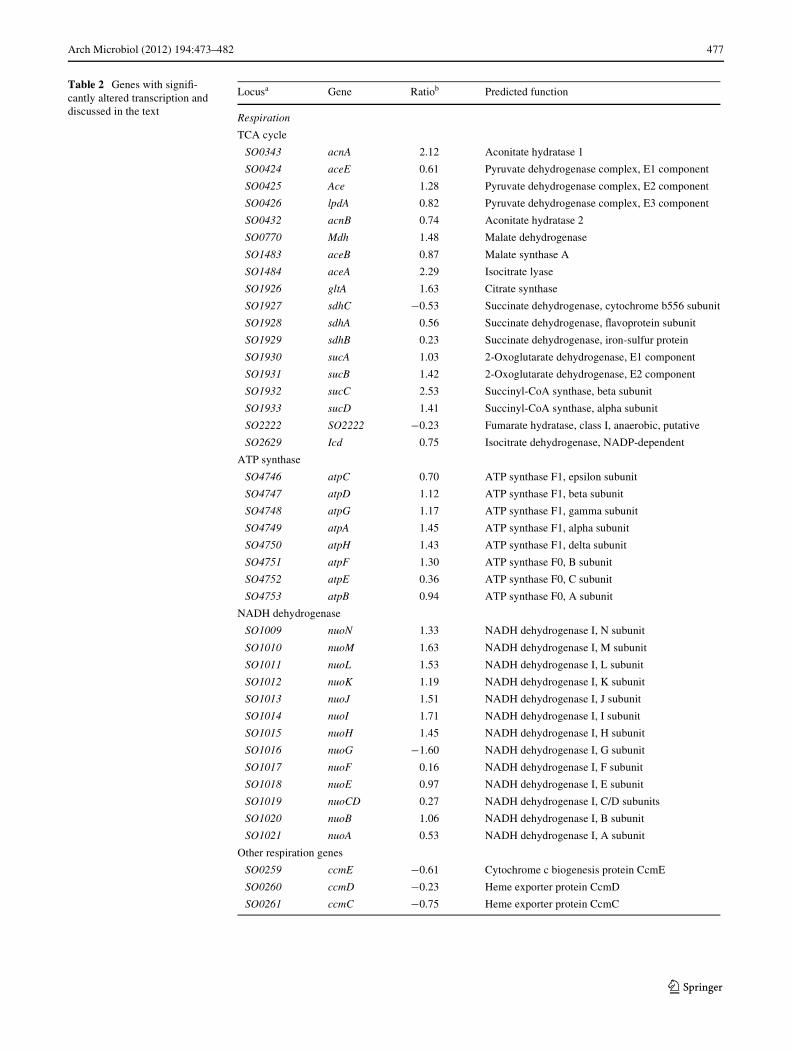
It is worth mentioning that almost all of genes encodingTCA enzymes were up-transcribed in pellicle cells(Table 2). In addition, all of 9 atpA-H genes encoding sub-units of ATP synthase and 12 out of 13 nuoA-N genesencoding subunits of NADH dehydrogenase were consis-tently up-transcribed (Table 2). Consistently, the transcrip-tion of a group of genes including so2361–64, whichencode subunits of cbb3-type cytochrome c oxidase andcatalyze the Wnal step of electron transfer between a cyto-chrome c and the molecular oxygen in the respiratorychain, was up-transcribed. All of these data suggested thatcells in pellicles were metabolically more active, especiallyin pathways related to aerobic respiration. The microarrayresults also demonstrated that the planktonic cells were infact under microaerobic conditions on the basis of that anumber of genes involved in anaerobic respiration weredrastically induced in planktonic cells. These includedso2178 (ccpA, cytochrome c551 peroxidase), so1429–30(dmsAB, dimethyl sulfoxide reductase), so4512–15 (for-
mate dehydrogenase), and so0259–63 (ccm, heme exporterprotein) (Table 2).
Transport proteins
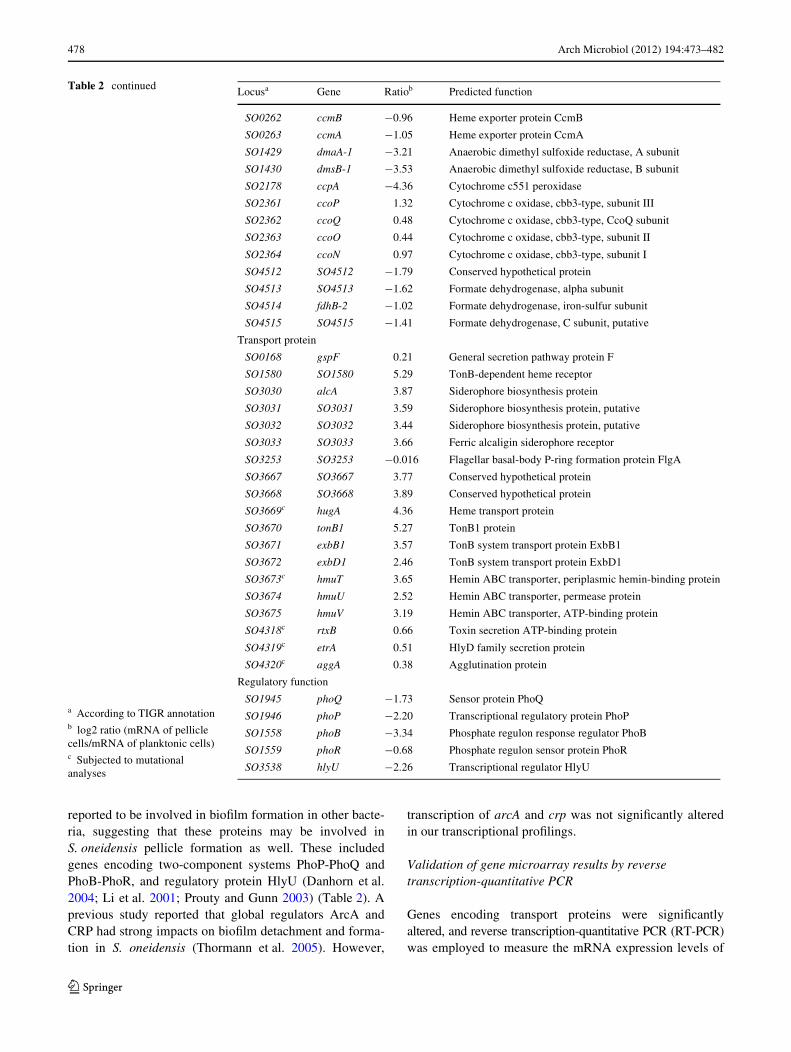
Since EPS, essential biopolymers for the formation of pelli-cles have to be manufactured mostly by microbes them-selves, genes encoding proteins involved in synthesis andtransport of EPS are likely to be transcribed diVerentlybetween the two types of cells. Indeed, expression of a totalof 64 (37 up-transcribed and 27 down-transcribed) genesencoding transport proteins was signiWcantly altered.Among these, genes encoding iron or heme uptake andtransportation proteins consistently showed the largest tran-scription diVerences. These included so1580 (TonB-depen-dent heme receptor), so3030–3 (siderophore biosynthesisproteins and receptor), and so3667–75 (so3667-so3668-hugA-tonB1-exbB1-exbD1-hmuT-hmuU-hmuV, heme/hemin transport proteins) (Table 2). Importance of iron inboth canonical bioWlm and pellicle formation has been welldocumented (Banin et al. 2005). Iron or heme from the var-ious resources, such as soluble and insoluble iron-contain-ing chemicals and iron/heme-containing proteins, can becaptured by siderophores secreted by cells or by outermembrane receptors (Braun and Braun 2002). These dataimplicated that iron may have a role in pellicle formation ormore iron was biosorpted by pellicle cells.
Three proteins including RtxB, EmrA, and AggA(S04318–4320) are predicted to be the cytoplasmic mem-brane localized ATP-binding protein, membrane fusionprotein and outer membrane protein component, respec-tively, of a type I secretion system (TISS). Microarray datashowed that so4318–4320 were consistently up-regulatedin pellicle cells (Table 2), suggesting certain substratessecreted by TISS may play important role in pellicle forma-tion.
gspF, encoding individual component of the generalsecretion pathway protein F, was a known gene of S. oneid-ensis MR-1 that participate in the process of electrontransfer (Shi et al. 2007; Py et al. 2001). Some best charac-terized proteins, which are involved in bioWlm formation,depend on general secretion pathway to transport across thecytoplasmic membrane (Lasa and Penades 2006). Micro-array data showed that gspF was up-regulated in pelliclecells (Table 2).
Regulatory proteins
In our study, transcription of 46 (11 up-transcribed and 35down-transcribed) genes encoding regulatory proteins wassigniWcantly diVerent. Although the majority of these genesencode proteins whose function is currently unclear, severalgenes are worth noting because their products have been
Fig. 2 DiVerentially expressed genes in pellicle cells grouped byfunctional classiWcation according to the TIGR S. oneidensis genomedatabase (http://www.tigr.org). Columns: 1 amino acid biosynthesis; 2biosynthesis of cofactors, prosthetic groups, and carriers; 3 cell enve-lope; 4 cellular processes; 5 central intermediary metabolism; 6 DNAmetabolism; 7 energy metabolism; 8 fatty acid and phospholipidmetabolism; 9 hypothetical proteins; 10 mobile and extrachromosomalelement functions; 11 protein fate; 12 protein synthesis; 13 purines,pyrimidines, nucleosides, and nucleotides; 14 regulatory functions; 15signal transduction; 16 transcription; 17 transport and binding proteins;18 unknown function
123
Arch Microbiol (2012) 194:473–482 477
Table 2 Genes with signiW-cantly altered transcription and discussed in the text
Locusa Gene Ratiob Predicted function
Respiration
TCA cycle
SO0343 acnA 2.12 Aconitate hydratase 1
SO0424 aceE 0.61 Pyruvate dehydrogenase complex, E1 component
SO0425 Ace 1.28 Pyruvate dehydrogenase complex, E2 component
SO0426 lpdA 0.82 Pyruvate dehydrogenase complex, E3 component
SO0432 acnB 0.74 Aconitate hydratase 2
SO0770 Mdh 1.48 Malate dehydrogenase
SO1483 aceB 0.87 Malate synthase A
SO1484 aceA 2.29 Isocitrate lyase
SO1926 gltA 1.63 Citrate synthase
SO1927 sdhC ¡0.53 Succinate dehydrogenase, cytochrome b556 subunit
SO1928 sdhA 0.56 Succinate dehydrogenase, Xavoprotein subunit
SO1929 sdhB 0.23 Succinate dehydrogenase, iron-sulfur protein
SO1930 sucA 1.03 2-Oxoglutarate dehydrogenase, E1 component
SO1931 sucB 1.42 2-Oxoglutarate dehydrogenase, E2 component
SO1932 sucC 2.53 Succinyl-CoA synthase, beta subunit
SO1933 sucD 1.41 Succinyl-CoA synthase, alpha subunit
SO2222 SO2222 ¡0.23 Fumarate hydratase, class I, anaerobic, putative
SO2629 Icd 0.75 Isocitrate dehydrogenase, NADP-dependent
ATP synthase
SO4746 atpC 0.70 ATP synthase F1, epsilon subunit
SO4747 atpD 1.12 ATP synthase F1, beta subunit
SO4748 atpG 1.17 ATP synthase F1, gamma subunit
SO4749 atpA 1.45 ATP synthase F1, alpha subunit
SO4750 atpH 1.43 ATP synthase F1, delta subunit
SO4751 atpF 1.30 ATP synthase F0, B subunit
SO4752 atpE 0.36 ATP synthase F0, C subunit
SO4753 atpB 0.94 ATP synthase F0, A subunit
NADH dehydrogenase
SO1009 nuoN 1.33 NADH dehydrogenase I, N subunit
SO1010 nuoM 1.63 NADH dehydrogenase I, M subunit
SO1011 nuoL 1.53 NADH dehydrogenase I, L subunit
SO1012 nuoK 1.19 NADH dehydrogenase I, K subunit
SO1013 nuoJ 1.51 NADH dehydrogenase I, J subunit
SO1014 nuoI 1.71 NADH dehydrogenase I, I subunit
SO1015 nuoH 1.45 NADH dehydrogenase I, H subunit
SO1016 nuoG ¡1.60 NADH dehydrogenase I, G subunit
SO1017 nuoF 0.16 NADH dehydrogenase I, F subunit
SO1018 nuoE 0.97 NADH dehydrogenase I, E subunit
SO1019 nuoCD 0.27 NADH dehydrogenase I, C/D subunits
SO1020 nuoB 1.06 NADH dehydrogenase I, B subunit
SO1021 nuoA 0.53 NADH dehydrogenase I, A subunit
Other respiration genes
SO0259 ccmE ¡0.61 Cytochrome c biogenesis protein CcmE
SO0260 ccmD ¡0.23 Heme exporter protein CcmD
SO0261 ccmC ¡0.75 Heme exporter protein CcmC
123
478 Arch Microbiol (2012) 194:473–482
reported to be involved in bioWlm formation in other bacte-ria, suggesting that these proteins may be involved inS. oneidensis pellicle formation as well. These includedgenes encoding two-component systems PhoP-PhoQ andPhoB-PhoR, and regulatory protein HlyU (Danhorn et al.2004; Li et al. 2001; Prouty and Gunn 2003) (Table 2). Aprevious study reported that global regulators ArcA andCRP had strong impacts on bioWlm detachment and forma-tion in S. oneidensis (Thormann et al. 2005). However,
transcription of arcA and crp was not signiWcantly alteredin our transcriptional proWlings.
Validation of gene microarray results by reverse transcription-quantitative PCR
Genes encoding transport proteins were signiWcantlyaltered, and reverse transcription-quantitative PCR (RT-PCR)was employed to measure the mRNA expression levels of
Table 2 continued Locusa Gene Ratiob Predicted function
SO0262 ccmB ¡0.96 Heme exporter protein CcmB
SO0263 ccmA ¡1.05 Heme exporter protein CcmA
SO1429 dmaA-1 ¡3.21 Anaerobic dimethyl sulfoxide reductase, A subunit
SO1430 dmsB-1 ¡3.53 Anaerobic dimethyl sulfoxide reductase, B subunit
SO2178 ccpA ¡4.36 Cytochrome c551 peroxidase
SO2361 ccoP 1.32 Cytochrome c oxidase, cbb3-type, subunit III
SO2362 ccoQ 0.48 Cytochrome c oxidase, cbb3-type, CcoQ subunit
SO2363 ccoO 0.44 Cytochrome c oxidase, cbb3-type, subunit II
SO2364 ccoN 0.97 Cytochrome c oxidase, cbb3-type, subunit I
SO4512 SO4512 ¡1.79 Conserved hypothetical protein
SO4513 SO4513 ¡1.62 Formate dehydrogenase, alpha subunit
SO4514 fdhB-2 ¡1.02 Formate dehydrogenase, iron-sulfur subunit
SO4515 SO4515 ¡1.41 Formate dehydrogenase, C subunit, putative
Transport protein
SO0168 gspF 0.21 General secretion pathway protein F
SO1580 SO1580 5.29 TonB-dependent heme receptor
SO3030 alcA 3.87 Siderophore biosynthesis protein
SO3031 SO3031 3.59 Siderophore biosynthesis protein, putative
SO3032 SO3032 3.44 Siderophore biosynthesis protein, putative
SO3033 SO3033 3.66 Ferric alcaligin siderophore receptor
SO3253 SO3253 ¡0.016 Flagellar basal-body P-ring formation protein FlgA
SO3667 SO3667 3.77 Conserved hypothetical protein
SO3668 SO3668 3.89 Conserved hypothetical protein
SO3669c hugA 4.36 Heme transport protein
SO3670 tonB1 5.27 TonB1 protein
SO3671 exbB1 3.57 TonB system transport protein ExbB1
SO3672 exbD1 2.46 TonB system transport protein ExbD1
SO3673c hmuT 3.65 Hemin ABC transporter, periplasmic hemin-binding protein
SO3674 hmuU 2.52 Hemin ABC transporter, permease protein
SO3675 hmuV 3.19 Hemin ABC transporter, ATP-binding protein
SO4318c rtxB 0.66 Toxin secretion ATP-binding protein
SO4319c etrA 0.51 HlyD family secretion protein
SO4320c aggA 0.38 Agglutination protein
Regulatory function
SO1945 phoQ ¡1.73 Sensor protein PhoQ
SO1946 phoP ¡2.20 Transcriptional regulatory protein PhoP
SO1558 phoB ¡3.34 Phosphate regulon response regulator PhoB
SO1559 phoR ¡0.68 Phosphate regulon sensor protein PhoR
SO3538 hlyU ¡2.26 Transcriptional regulator HlyU
a According to TIGR annotationb log2 ratio (mRNA of pellicle cells/mRNA of planktonic cells)c Subjected to mutational analyses
123
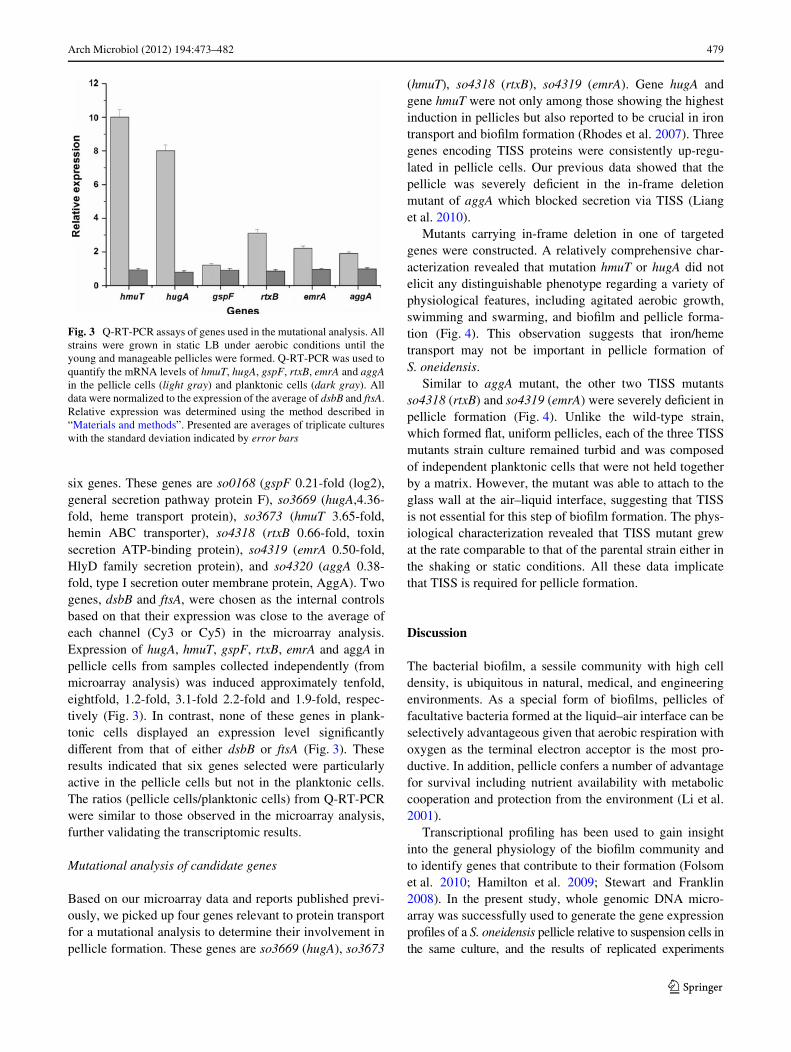
Arch Microbiol (2012) 194:473–482 479
six genes. These genes are so0168 (gspF 0.21-fold (log2),general secretion pathway protein F), so3669 (hugA,4.36-fold, heme transport protein), so3673 (hmuT 3.65-fold,hemin ABC transporter), so4318 (rtxB 0.66-fold, toxinsecretion ATP-binding protein), so4319 (emrA 0.50-fold,HlyD family secretion protein), and so4320 (aggA 0.38-fold, type I secretion outer membrane protein, AggA). Twogenes, dsbB and ftsA, were chosen as the internal controlsbased on that their expression was close to the average ofeach channel (Cy3 or Cy5) in the microarray analysis.Expression of hugA, hmuT, gspF, rtxB, emrA and aggA inpellicle cells from samples collected independently (frommicroarray analysis) was induced approximately tenfold,eightfold, 1.2-fold, 3.1-fold 2.2-fold and 1.9-fold, respec-tively (Fig. 3). In contrast, none of these genes in plank-tonic cells displayed an expression level signiWcantlydiVerent from that of either dsbB or ftsA (Fig. 3). Theseresults indicated that six genes selected were particularlyactive in the pellicle cells but not in the planktonic cells.The ratios (pellicle cells/planktonic cells) from Q-RT-PCRwere similar to those observed in the microarray analysis,further validating the transcriptomic results.
Mutational analysis of candidate genes
Based on our microarray data and reports published previ-ously, we picked up four genes relevant to protein transportfor a mutational analysis to determine their involvement inpellicle formation. These genes are so3669 (hugA), so3673
(hmuT), so4318 (rtxB), so4319 (emrA). Gene hugA andgene hmuT were not only among those showing the highestinduction in pellicles but also reported to be crucial in irontransport and bioWlm formation (Rhodes et al. 2007). Threegenes encoding TISS proteins were consistently up-regu-lated in pellicle cells. Our previous data showed that thepellicle was severely deWcient in the in-frame deletionmutant of aggA which blocked secretion via TISS (Lianget al. 2010).
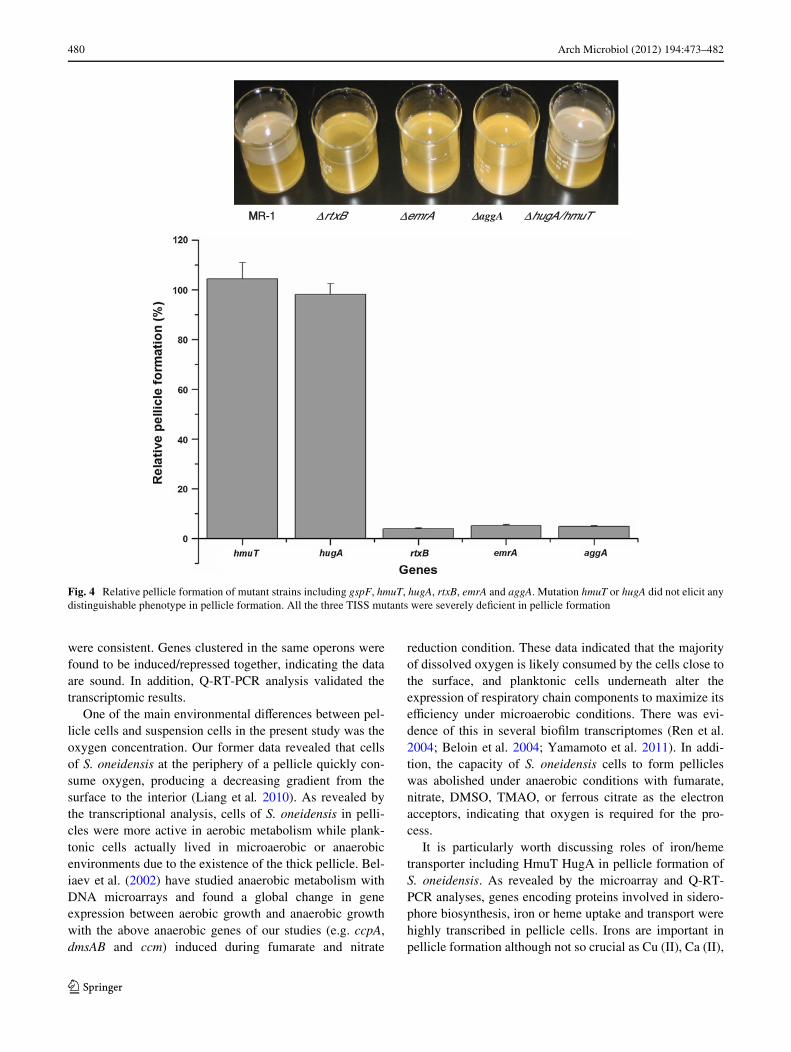
Mutants carrying in-frame deletion in one of targetedgenes were constructed. A relatively comprehensive char-acterization revealed that mutation hmuT or hugA did notelicit any distinguishable phenotype regarding a variety ofphysiological features, including agitated aerobic growth,swimming and swarming, and bioWlm and pellicle forma-tion (Fig. 4). This observation suggests that iron/hemetransport may not be important in pellicle formation ofS. oneidensis.
Similar to aggA mutant, the other two TISS mutantsso4318 (rtxB) and so4319 (emrA) were severely deWcient inpellicle formation (Fig. 4). Unlike the wild-type strain,which formed Xat, uniform pellicles, each of the three TISSmutants strain culture remained turbid and was composedof independent planktonic cells that were not held togetherby a matrix. However, the mutant was able to attach to theglass wall at the air–liquid interface, suggesting that TISSis not essential for this step of bioWlm formation. The phys-iological characterization revealed that TISS mutant grewat the rate comparable to that of the parental strain either inthe shaking or static conditions. All these data implicatethat TISS is required for pellicle formation.
Discussion
The bacterial bioWlm, a sessile community with high celldensity, is ubiquitous in natural, medical, and engineeringenvironments. As a special form of bioWlms, pellicles offacultative bacteria formed at the liquid–air interface can beselectively advantageous given that aerobic respiration withoxygen as the terminal electron acceptor is the most pro-ductive. In addition, pellicle confers a number of advantagefor survival including nutrient availability with metaboliccooperation and protection from the environment (Li et al.2001).
Transcriptional proWling has been used to gain insightinto the general physiology of the bioWlm community andto identify genes that contribute to their formation (Folsomet al. 2010; Hamilton et al. 2009; Stewart and Franklin2008). In the present study, whole genomic DNA micro-array was successfully used to generate the gene expressionproWles of a S. oneidensis pellicle relative to suspension cells inthe same culture, and the results of replicated experiments
Fig. 3 Q-RT-PCR assays of genes used in the mutational analysis. Allstrains were grown in static LB under aerobic conditions until theyoung and manageable pellicles were formed. Q-RT-PCR was used toquantify the mRNA levels of hmuT, hugA, gspF, rtxB, emrA and aggAin the pellicle cells (light gray) and planktonic cells (dark gray). Alldata were normalized to the expression of the average of dsbB and ftsA.Relative expression was determined using the method described in“Materials and methods”. Presented are averages of triplicate cultureswith the standard deviation indicated by error bars
123
480 Arch Microbiol (2012) 194:473–482
were consistent. Genes clustered in the same operons werefound to be induced/repressed together, indicating the dataare sound. In addition, Q-RT-PCR analysis validated thetranscriptomic results.
One of the main environmental diVerences between pel-licle cells and suspension cells in the present study was theoxygen concentration. Our former data revealed that cellsof S. oneidensis at the periphery of a pellicle quickly con-sume oxygen, producing a decreasing gradient from thesurface to the interior (Liang et al. 2010). As revealed bythe transcriptional analysis, cells of S. oneidensis in pelli-cles were more active in aerobic metabolism while plank-tonic cells actually lived in microaerobic or anaerobicenvironments due to the existence of the thick pellicle. Bel-iaev et al. (2002) have studied anaerobic metabolism withDNA microarrays and found a global change in geneexpression between aerobic growth and anaerobic growthwith the above anaerobic genes of our studies (e.g. ccpA,dmsAB and ccm) induced during fumarate and nitrate
reduction condition. These data indicated that the majorityof dissolved oxygen is likely consumed by the cells close tothe surface, and planktonic cells underneath alter theexpression of respiratory chain components to maximize itseYciency under microaerobic conditions. There was evi-dence of this in several bioWlm transcriptomes (Ren et al.2004; Beloin et al. 2004; Yamamoto et al. 2011). In addi-tion, the capacity of S. oneidensis cells to form pellicleswas abolished under anaerobic conditions with fumarate,nitrate, DMSO, TMAO, or ferrous citrate as the electronacceptors, indicating that oxygen is required for the pro-cess.
It is particularly worth discussing roles of iron/hemetransporter including HmuT HugA in pellicle formation ofS. oneidensis. As revealed by the microarray and Q-RT-PCR analyses, genes encoding proteins involved in sidero-phore biosynthesis, iron or heme uptake and transport werehighly transcribed in pellicle cells. Irons are important inpellicle formation although not so crucial as Cu (II), Ca (II),
Fig. 4 Relative pellicle formation of mutant strains including gspF, hmuT, hugA, rtxB, emrA and aggA. Mutation hmuT or hugA did not elicit anydistinguishable phenotype in pellicle formation. All the three TISS mutants were severely deWcient in pellicle formation
MMR-1 ΔrtxB ΔemrA ΔaggA ΔhugA/hmuT
123

Arch Microbiol (2012) 194:473–482 481
Mn (II), and Zn (II) (Liang et al. 2010). However, pellicleformation of S. oneidensis was barely aVected by mutationsin hugA and hmuT. When hugA or hmuT deletion mutantgrew in the LB medium, even if the iron was limited, othercations in LB medium could replace the iron, so these twomutant strains could still form the pellicle. In addition,these two genes reside in a large gene cluster encoding iron/heme uptake and transport proteins, and all these geneshave been found to respond to the environmental changesconsistently (Beliaev et al. 2005; Braun and Braun 2002;Gao et al. 2006). The substantial diVerences in transcriptionof these genes between two diVerent types of cells may beresulted from an imbalance in metabolism. In conclusion,the Wnding that HmuT and HugA have little impacts on pel-licle formation demands a further investigation.
The type I secretion apparatus is a rather simple machinecontaining only three diVerent transport components, two inthe inner membrane, which are speciWc for the passengerprotein, and one that forms a general pore in the outermembrane (Holland et al. 2005). Three genes encodingTISS proteins were consistently up-regulated in S. oneiden-sis MR-1 pellicle cells. At the same time, all the three TISSmutants were severely deWcient in pellicle formation. Inaddition, Theunissen et al. subjected MR-1 to randomtransposon insertion mutagenesis to identify genes contrib-uting to the ability of the organism to form bioWlms onpolystyrene surfaces. RTX, a novel 285-kDa multi-domainprotein, is secreted by TISS to the cell surface, where it is arequisite for solid surface-associated bioWlm development(Theunissen et al. 2010). These data suggested that type Isecretion pathway is directly related to bioWlm formation.
Pellicle formation is a complex process. Many factors,including oxygen, secreted proteins, extracellular organ-elles such as Xagella, and chemical agents supplementedin media such as iron and calcium, played important rolesin bioWlm formation of S. oneidensis MR-1 (Liang et al.2011). The results presented here provided the compre-hensive insights into pellicle formation of S. oneidensis.However, the events in transcriptional regulation duringpellicle formation are incompletely understood. As weknow, the S. oneidensis MR-1 cells formed solid surface-associated (SSA) bioWlms on the Xask wall and anotherkind of suspension cells in LB when aeration of the mediawas provided by shaking, which the dissolved oxygenlevel was as high as the surface growing pellicle cells.Since one of the signiWcant environmental diVerencesbetween pellicle cells and planktonic cells in the presentstudy was the oxygen concentration, the suspension cellsin the agitated medium could be another control. In thefuture, we can expect the combination of genetics, bio-chemistry, and microscopy to yield an ever-increasingunderstanding of the molecular mechanisms of pellicleformation.
Acknowledgments This research was supported by The U.S.Department of Energy under the Genomics: GTL Program throughShewanella Federation, OYce of Biological and EnvironmentalResearch, OYce of Science. This research was also supported by theNational Basic Research Program (973 Program: 2010CB630901),Central South University research startup funding (7602110285) andNational Natural Science Foundation of China (302001124) to YL.
References
Bagge D, Hjelm M, Johansen C, Huber I, Grami L (2001) Shewanellaputrefaciens adhesion and bioWlm formation on food processingsurfaces. Appl Environ Microbiol 67:2319–2325
Banin E, Vasil ML, Greenberg EP (2005) Iron and Pseudomonas aeru-ginosa bioWlm formation. Proc Nat Acad Sci USA 102:11076–11081
Beliaev AS, Thompson DK, Giometti CS, Li GS, Yates J III, NealsonKH, Tiedje JM, Heidelberg JF, Zhou JZ (2002) Gene and proteinexpression proWles of Shewanella oneidensis during anaerobicgrowth with diVerent electron acceptors. Omics J Integr Biol6:39–60
Beliaev AS, Klingeman DM, Klappenbach JA, Wu L, Romine MF,Tiedje JM, Nealson KH, Fredrickson JK, Zhou J (2005) Globaltranscriptome analysis of Shewanella oneidensis MR-1 exposed todiVerent terminal electron acceptors. J Bacteriol 187:7138–7145
Beloin C, Valle J, Latour-Lambert P, Faure P, Kzreminski M, Balestri-no D, Haagensen JA, Molin S, Prensier G, Arbeille B (2004) Glo-bal impact of mature bioWlm lifestyle on Escherichia coli K-12gene expression. Mol Microbiol 51:659–674
Branda SS, Gonzalez-Pastor JE, Dervyn E, Ehrlich SD, Losick R,Kolter R (2004) Genes involved in formation of structured multi-cellular communities by Bacillus subtilis. J Bacteriol 186:3970–3979
Braun V, Braun M (2002) Active transport of iron and siderophoreantibiotics. Curr Opin Microbiol 5:194–201
Danhorn T, Hentzer M, Givskov M, Parsek MR, Fuqua C (2004)Phosphorus limitation enhances bioWlm formation of the plantpathogen Agrobacterium tumefaciens through the PhoR-PhoBregulatory system. J Bacteriol 186:4492–4501
Enos-Berlage JL, Guvener ZT, Keenan CE, McCarter LL (2005) Geneticdeterminants of bioWlm development of opaque and translucentVibrio parahaemolyticus. Mol Microbiol 55(4):1160–1182
Folsom JP, Richards L, Pitts B, Roe F, Ehrlich GD, Parker A, MazurieA, Stewart PS (2010) Physiology of Pseudomonas aeruginosa inbioWlms as revealed by transcriptome analysis. BMC Microbiol-ogy 10:294
Gao HC, Wang Y, Liu XD, Yan TF, Wu LY, Alm E, Arkin A, Thomp-son DK, Zhou JZ (2004) Global transcriptome analysis of the heatshock response of Shewanella oneidensis. J Bacteriol 186:7796–7803
Gao H, Yang ZK, Wu L, Thompson DK, Zhou J (2006) Global tran-scriptome analysis of the cold shock response of Shewanellaoneidensis MR-1 and mutational analysis of its classical coldshock proteins. J Bacteriol 188:4560–4569
Gao HC, Wang XH, Yang ZK, Palzkill T, Zhou JZ (2008) Probing reg-ulon of ArcA in Shewanella oneidensis MR-I by integrated geno-mic analyses. Bmc Genomics 9 (Suppl 1):S11
Guvener ZT, McCarter LL (2003) Multiple regulators control cap-sular polysaccharide production in Vibrio parahaemolyticus.J Bacteriol 185(8):5431–5441
Hamilton S, Bongaerts RJM, Mulholland F, Cochrane B, Porter J, LucchiniS, Lappin-Scott HM, Hinton JCD (2009) The transcriptional pro-gramme of Salmonella enterica serovar Typhimurium reveals a keyrole for tryptophan metabolism in bioWlms. BMC Genomics 10:599
123

482 Arch Microbiol (2012) 194:473–482
Holland IB, Schmitt L, Young J (2005) Type 1 protein secretion in bac-teria, the ABC-transporter dependent pathway (review). MolMembr Biol 22(1–2):29–39
Kobayashi K (2007) Bacillus subtilis pellicle formation proceedsthrough genetically deWned morphological changes. J Bacteriol189:4920–4931
Kumagai A, Mizuno M, Kato N, Nozaki K, Togawa E, Yamanaka S,Okuda K, Saxena IM, Amano Y (2011) UltraWne Cellulose FibersProduced by Asaia bogorensis, an Acetic Acid Bacterium. Bio-macromolecules. doi:10.1021/bm2005615
Lasa I, Penades JR (2006) Bap: a family of surface proteins involvedin bioWlm formation. Res Microbiol 157(2):99–107
Li YH, Lau PCY, Lee JH, Ellen RP, Cvitkovitch DG (2001) Naturalgenetic transformation of Streptococcus mutants growing in bio-Wlms. J Bacteriol 183:897–908
Liang YL, Gao HC, Chen JR, Dong YY, Wu L, He ZL, Liu XD, QiuGZ, Zhou JZ (2010) Pellicle formation in Shewanella oneidensis.BMC Microbiol 10:291
Nakagawa Y, Toda Y, Yamamura H, Hayakawa M, Iimura Y (2011)FLO11 is essential for pellicle formation by wild pellicle-formingyeasts isolated from contaminated wines. J Biosci Bioeng 111(1):7–9
Prouty AM, Gunn JS (2003) Comparative analysis of Salmonella ent-erica serovar typhimurium bioWlm formation on gallstones and onglass. Infect Immun 71:7154–7158
Py B, Loiseau L, Barras F (2001) An inner membrane platform in thetype II secretion machinery of Gram-negative bacteria. Embo Re-ports 2(3):244–248
Ren D, Bedzyk LA, Setlow P, Thomas SM, Ye RW, Wood TK (2004)Gene expression in Bacillus subtilis surface bioWlms with andwithout sporulation and the importance of yveR for bioWlm main-tenance. Biotechnol Bioeng 86:344–364
Rhodes ER, Menke S, Shoemaker C, Tomaras AP, McGillivary G, Ac-tis LA (2007) Iron acquisition in the dental pathogen Actinobacil-lus actinomycetemcomitans: what does it use as a source and howdoes it get this essential metal? Biometals 20:365–377
Shi L, Squier TC, Zachara JM, Fredrickson JK (2007) Respiration ofmetal (hydr)oxides by Shewanella and Geobacter: a key role formultihaem c-type cytochromes. Mol Microbiol 65(1):12–20
Solano C, Garcia B, Valle J, Berasain C, Ghigo JM, Gamazo C, Lasa I(2002) Genetic analysis of Salmonella enteritidis bioWlm forma-tion: critical role of cellulose. Mol Microbiol 43:793–808
Spiers AJ, Bohannon J, Gehrig SM, Rainey PB (2003) BioWlm forma-tion at the air-liquid interface by the Pseudomonas XuorescensSBW25 wrinkly spreader requires an acetylated form of cellulose.Mol Microbiol 50:15–27
Stewart PS, Franklin MJ (2008) Physiological heterogeneity in bio-Wlms. Nat Rev Microbiol 6:199–210
Teal TK, Lies DP, Wold BJ, Newman DK (2006) SpatiometabolicstratiWcation of Shewanella oneidensis bioWlms. Appl EnvironMicrobiol 72:7324–7330
Theunissen S, De Smet L, Dansercoer A, Motte B, Coenye T, Van Bee-umen JJ, Devreese B, Savvides SN, Vergauwen B (2010) The285 kDa Bap/RTX hybrid cell surface protein (SO4317) ofShewanella oneidensis MR-1 is a key mediator of bioWlm forma-tion. Res Microbiol 161:144–152
Thormann KM, Saville RM, Shukla S, Pelletier DA, Spormann AM(2004) Initial phases of bioWlm formation in Shewanella oneiden-sis MR-1. J Bacteriol 186:8096–8104
Thormann KM, Saville RM, Shukla S, Spormann AM (2005) Induc-tion of rapid detachment in Shewanella oneidensis MR-1 bio-Wlms. J Bacteriol 187:1014–1021
Vriendt D, Theunissen KS, Carpentier W, Smet LD, Devreese B, Bee-umen JV (2005) Proteomics of Shewanella oneidensis MR-1 bio-Wlm reveals diVerentially expressed proteins, including AggA andRibB. Proteomics 5:1308–1316
Whitchurch CB, Tolker-Nielsen T, Ragas PC, Mattick JS (2002)Extracellular DNA required for bacterial bioWlm formation. Sci-ence 295:1487
Windt DW, Gao HC, Kromer W, Van Damme P, Dick J, Mast J,Boon N, Zhou JZ, Verstraete W (2006) AggA is required foraggregation and increased bioWlm formation of a hyper-aggre-gating mutant of Shewanella oneidensis MR-1. Microbiology152:721–729
Yamamoto K, Arai H, Ishii M, Igarashi Y (2011) Trade-oV betweenoxygen and iron acquisition in bacterial cells at the air-liquidinterface. FEMS Microbiol Ecol 77(1):83–94
123
Copyright © 2022 FDOKUMEN




















