
Tolerance-induced receptor selection: scope, sensitivity, locus specificity, and relationship to...
12
Tolerance-induced receptor selection: scope, sensitivity, locus specificity, and relationship to lymphocyte- positive selection Djemel Aı ¨t-Azzouzene Patrick Skog Marc Retter Valerie Kouskoff Marc Hertz Julie Lang Jennifer Kench Michael Chumley Doron Melamed Janice Sudaria Amanda Gavin Annica Martensson Laurent Verkoczy Bao Duong Jose ´ Vela David Nemazee Authors’ addresses Djemel Aı ¨t-Azzouzene 1 , Patrick Skog 1 , Marc Retter 1,2 , Valerie, Kouskoff 2 , Marc Hertz 2 , Julie Lang 2 , Jennifer Kench 2 , Michael Chumley 2 , Doron Melamed 2 , Janice Sudaria 1 , Amanda Gavin 1 , Annica Martensson 1 , Laurent Verkoczy 1 , Bao Duong 1,3 , Jose ´ Vela 1,3 , David Nemazee 1 , 1 Department of Immunology, The Scripps Research Institute, La Jolla, CA, USA. 2 National Jewish Medical and Research Center, Denver, CO, USA. 3 Doctoral Program in Chemical and Biological Sciences, The Scripps Research Institute, La Jolla, CA, USA. Correspondence to: David Nemazee Department of Immunology The Scripps Research Institute 10550 North Torrey Pines Road Mail drop IMM-29 La Jolla, CA 92037, USA Tel.: þ1 858 784 9528 Fax: þ1 858 784 9554 E-mail: [email protected] Acknowledgements This research was supported by grants GM44809, AI33608, and AI49940 from the National Institutes of Health. Summary: Receptor editing is a mode of immunological tolerance of B lymphocytes that involves antigen-induced B-cell receptor signaling and consequent secondary immunoglobulin light chain gene recombination. This ongoing rearrangement often changes B-cell specificity for antigen, rendering the cell non-autoreactive and sparing it from deletion. We currently believe that tolerance-induced editing is limited to early stages in B-cell development and that it is a major mechanism of tolerance, with a low-affinity threshold and the potential to take place in virtually every developing B cell. The present review highlights the contributions from our laboratory over several years to elucidate these features. Clonal selection and receptor selection The immune system uses two strikingly different strategies to regulate its antigen receptor repertoire: clonal selection and receptor selection. As it was first proposed by Burnet (1) [and independently by Talmage (2)], clonal selection has been recognized as a major, and perhaps the only, mechanism of antigen-specific regulation of the repertoire. According to the generally accepted model, antigen-specific tolerance and immunity are regulated by the ability of antigen to act on cells, through their clonally restricted antigen receptors, which signal to stimulate cell growth and survival or apoptosis. However, cells that express antigen receptors may also alter their antigenic specificity by modifications of the receptor genes and proteins themselves, a process that we refer to as receptor selection (3). Receptor selection can take place by ongoing V(D)J recombination, which modifies and replaces antigen receptor genes, or by induced somatic mutation, which introduces point mutations (or gene conversions) in antigen receptors. In this review, we focus on studies from our Immunological Reviews 2004 Vol. 197: 219–230 Printed in Denmark. All rights reserved Copyright ß Blackwell Munksgaard 2004 Immunological Reviews 0105-2896 219
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Tolerance-induced receptor selection: scope, sensitivity, locus specificity, and relationship to...

Tolerance-induced receptor selection:
scope, sensitivity, locus specificity,
and relationship to lymphocyte-
positive selection
Djemel Aıt-Azzouzene
Patrick Skog
Marc Retter
Valerie Kouskoff
Marc Hertz
Julie Lang
Jennifer Kench
Michael Chumley
Doron Melamed
Janice Sudaria
Amanda Gavin
Annica Martensson
Laurent Verkoczy
Bao Duong
Jose Vela
David Nemazee
Authors’ addresses
Djemel Aıt-Azzouzene1, Patrick Skog1, Marc Retter1,2,Valerie, Kouskoff2, Marc Hertz2, Julie Lang2, JenniferKench2, Michael Chumley2, Doron Melamed2, JaniceSudaria1, Amanda Gavin1, Annica Martensson1, LaurentVerkoczy1, Bao Duong1,3, Jose Vela1,3, David Nemazee1,1Department of Immunology, The Scripps
Research Institute, La Jolla, CA, USA.2National Jewish Medical and Research Center,
Denver, CO, USA.3Doctoral Program in Chemical and Biological
Sciences, The Scripps Research Institute, La
Jolla, CA, USA.
Correspondence to:
David NemazeeDepartment of Immunology
The Scripps Research Institute
10550 North Torrey Pines Road
Mail drop IMM-29
La Jolla, CA 92037, USA
Tel.: þ1 858 784 9528
Fax: þ1 858 784 9554
E-mail: [email protected]
Acknowledgements
This research was supported by grants GM44809,
AI33608, and AI49940 from the National Institutes of
Health.
Summary: Receptor editing is a mode of immunological tolerance ofB lymphocytes that involves antigen-induced B-cell receptor signaling andconsequent secondary immunoglobulin light chain gene recombination.This ongoing rearrangement often changes B-cell specificity for antigen,rendering the cell non-autoreactive and sparing it from deletion. Wecurrently believe that tolerance-induced editing is limited to early stagesin B-cell development and that it is a major mechanism of tolerance, with alow-affinity threshold and the potential to take place in virtually everydeveloping B cell. The present review highlights the contributions fromour laboratory over several years to elucidate these features.
Clonal selection and receptor selection
The immune system uses two strikingly different strategies to
regulate its antigen receptor repertoire: clonal selection and
receptor selection. As it was first proposed by Burnet (1) [and
independently by Talmage (2)], clonal selection has been
recognized as a major, and perhaps the only, mechanism of
antigen-specific regulation of the repertoire. According to the
generally accepted model, antigen-specific tolerance and
immunity are regulated by the ability of antigen to act on
cells, through their clonally restricted antigen receptors, which
signal to stimulate cell growth and survival or apoptosis.
However, cells that express antigen receptors may also alter
their antigenic specificity by modifications of the receptor
genes and proteins themselves, a process that we refer to as
receptor selection (3). Receptor selection can take place by
ongoing V(D)J recombination, which modifies and replaces
antigen receptor genes, or by induced somatic mutation,
which introduces point mutations (or gene conversions) in
antigen receptors. In this review, we focus on studies from our
Immunological Reviews 2004Vol. 197: 219–230Printed in Denmark. All rights reserved
Copyright � Blackwell Munksgaard 2004
Immunological Reviews0105-2896
219

laboratory examining the mechanism of immune tolerance in
B cells reactive to membrane-expressed autoantigen. In these
studies, we identified V(D)J recombinase-mediated receptor
editing as a major mechanism of tolerance early in B-lymphocyte
development, whereas later at post-bone marrow stages, the
major mechanism of tolerance appeared to be clonal selection
by apoptosis or functional inactivation.
Peripheral and central B-cell tolerance
We first became convinced that editing occurs in a comparison
of peripheral and central tolerance in mice transgenic for the
immunoglobulin (Ig) M and IgD gene forms of antibody 3–83
(4). This antibody is reactive to major histocompatibility
complex (MHC) class I molecules H-2Kk, Kb, and other allo-
forms but is not reactive to H-2d molecules. In the absence of
cognate antigen, B cells in 3–83 transgenic mice retain the
transgene-encoded specificity, populating the mouse with a
virtually monoclonal B-cell population (Fig. 1, middle row). In
the experiment depicted in Fig. 1, these 3–83 Ig transgenic
mice were crossed to mice carrying liver-expressed Kb trans-
gene driven by the albumin promoter, which provides a
model of peripheral B-cell tolerance. As can be seen in the
lower panels, autoreactive B cells are eliminated to a great
extent in the lymph nodes and partially in the spleen, with
little replacement of B cells lacking the transgenic specificity
(detected with an anti-idiotypic antibody). The splenic B cells
are largely short-lived and in part rescuable by Bcl-2 over-
expression (5). Such a pattern of tolerance-induced clonal
elimination conforms to the clonal selection ideal.
In contrast, when 3–83 Ig transgenic mice are bred to H-2k
or H-2b mice, where antigen is present on the whole body, we
found a less profound B-cell loss, with considerable replace-
ment of cells carrying the transgene-encoded specificity by
long-lived B cells retaining the transgenic H-chain, but now
expressing endogenous L-chains. About half of these idiotype-
negative cells carried lL-chains (Fig. 2). A key finding was that
the appearance of these cells was clearly not the result of
outgrowth of pre-existing variants.
As shown in Fig. 3, the emerging lþ cells were specifically
found in 3–83 transgenic mice whose B cells expressed anti-
gen on the whole body, but not in mice deleting transgenic
cells in the periphery (3–83 transgenic�MT-Kb mice), which
were even more B-cell deficient. This finding suggested that
encounter of antigen at a bone marrow stage of development
promoted further L-chain rearrangements.
Editing in tissue culture
The ability of innocuous antigen receptors to suppress V(D)J
recombination and of autoreactive receptors to induce it was
evaluated in a number of ways. A direct ex vivo culture or
modified interleukin-7 (IL-7) culture system was used to
demonstrate that, in the absence of antigen, the 3–83 receptor
could suppress endogenous Ig-k-recombination to approxi-
mately 1–2% of cells, whereas in the presence of antigen,
40–70% underwent endogenous Ig-k recombination (6, 7)
(Figs 4–6). These changes were mirrored in the increase in
recombination-activating gene (RAG) mRNA levels in antigen-
challenged sIgþ bone marrow B cells (6, 7). Importantly, in all
of our studies, VDJ assembly at the H-chain was suppressed, as
determined by the paucity of endogenous H-chain protein
expression and also when monitored with a sensitive polymer-
ase chain reaction method (7) (Fig. 5E).
Bone marrow Spleen Lymph node
WT
3–83 ND
3–83 PD
IgM
0.4% 0.7% 0.2%
31%
0.3%15%27%
3–83
Idio
type
26% 52%
Fig. 1. Self-membrane antigen targeted to the livermediates clonal deletion. In this flow cytometryanalysis, 3–83 transgenic mice on an antigen-freebackground (ND) have abundant B cells carrying thetransgenic receptor idiotype (middle row), whereas3–83 mice expressing the cognate Kb antigen on theliver (lower panels) delete the now autoreactive Bcells in lymph node and spleen, but not in bonemarrow (56). IgM, immunoglobulin M; WT,wildtype; PD, peripherally deleting.
Aıt-Azzouzene et al �Receptor editing
220 Immunological Reviews 197/2004

It should be noted that these in vitro systems were amenable
to the analysis of non-transgenic B cells, and they allowed us
to show that editing could be readily induced in bone marrow
B cells using anti-B-cell receptor (BCR) antibodies (6–8).
Importantly, recirculating mature CD23þIgDþ B cells in
these cultures were refractory to editing, arguing that despite
sharing a similar culture environment to the CD23– cells in the
same dishes, they were editing incompetent (6). Thus, there
appears to be a ‘point of no return’ in preimmune develop-
ment, where B cells become editing incompetent, although
recent work suggests that amongst transitional B cells, editing
competence may be retained under certain conditions (9–11).
Together with considerable data from the anti-DNA trans-
genic models (12–20), in which skewed Jk usage was
observed in cells surviving tolerance, these results led to a
model suggesting that ongoing L-chain recombination could
play a major role in immune tolerance. Indeed, the structure of
the k-locus appeared to be especially suited to such a process. Asshown schematically in Fig. 7, the k-locus permits replacement
of active VJ exons with new ones by nested rearrangements.
Related work studying editing in mice with Ig-k loci tar-
geted with pre-rearranged genes found that such transgenes
suppress rearrangements in some but not all cells (21–23),
a pattern similar to that observed by Storb and colleagues
(24–26) in k-transgenic mice. These results suggested that
either feedback suppression of innocuous receptors is inef-
ficient or tolerance-induced editing is common. For example,
it was found that about one-fourth of the B cells in such
‘knockin’ mice express non-targeted L-chains, rather than the
targeted allele (22).
Editing in a knockin model
How can we explain why normal B cells rarely express two L-
chains on their surface, while functional, pre-rearranged, tar-
geted k-loci often fail to suppress endogenous recombination
Ig-Tg (H-2k)Ig-Tg (H-2d)
Dbl-Tg
Non-Tg
00
00
10
1
1FITC anti-λ
PE
ant
i-IgD
a
10
5.4% 4.8%
0.2%
10
100
100
1000
1000
Days
% la
bele
d B
220h
i B c
ells
20
20
30 40
40
60
80
A
B
H-2b Tg spleen
Fig. 2. Manifestations of receptor editing.
(A) Edited splenic B cells from H-2b 3–83transgenic mouse retain transgenic H-chainas detected with allotype-specificimmunoglobulin D (IgD) antibody, but oftenexpress endogenous l chains (4). (B) EditedB cells in Ig-transgenic H-2k mice have longlifespan as judged by BrdU uptake studiescompared to mice whose B cells encounterantigen in the periphery (Dbl transgenic) oreven non-deleting Ig-transgenic (H-2d) mice(from the doctoral thesis of J. Kench,U. Colorado Health Science Center). FITC,fluorescein isothiocyanate; PE, phycoerythrin;Tg, transgenic.
8
4
2
17
10
7
% Ig
Da /
λ+ c
ells
0H-2d
-H-2b
Whole bobyMT-Kb
LiverAntigenlocation
Fig. 3. Edited lþ B cells are induced by bone marrow antigen but notperipheral antigen. Spleen cells from 3-83 transgenic mice lackingantigen (H-2d) or bearing antigen in the liver (MT-Kb) or whole body(H-2b) were analyzed by flow cytometry for coexpression of transgenicH-chain (IgDa) and endogenous (l) L-chain. Numbers of individual micetested in each group are listed above each bar.
Aıt-Azzouzene et al �Receptor editing
Immunological Reviews 197/2004 221

(Fig. 8). In other words, how can editing be compatible with
the empiric phenomena of allelic and isotypic exclusion? One
possibility is that k-loci are monoallelically expressed (27, 28),
and another is that ongoing rearrangements often occur
because of tolerance-induced editing or because innocuous
H/L pairs often fail to mediate feedback suppression of recombin-
ation. Evidence for the latter two explanations was obtained
using the 3–83 knockin model (29, 30). In this model, hemi-
zygous versions of the targeted H and L transgenes failed to
suppress endogenous IgL rearrangements, despite their lack of
autoreactivity (31). By providing these genes in the homozygous
form, the sIg expression level was elevated to the point where
B-cell development was promoted and endogenous rearrange-
ments suppressed (29, 30). Most importantly, on an antigen-
bearing background, these same cells underwent very efficient
editing, resulting in the generation of splenic B cells in number
essentially similar to that of control mice lacking antigen (Fig. 8).
As in the conventional 3–83 transgenic mice, about half of these
rescued cells expressed l light chains, but in the case of the
knockin mice, there was little overall reduction in B cells. These
studies suggest that tolerance-induced receptor editing is a
remarkably efficient process and that an insufficient level of
receptor expression can lead to alterations in B-cell specificity
that are related to a lack of positive selection. In this respect, the
B cells may to some extent behave like CD4/CD8 double-positive
thymocytes, which in the absence of positive selection, undergo
continuing T-cell receptor � recombinations [to further extend
the analogy, it also seems possible that, like B cells, autoreactive
T cells may be subject to tolerance-induced editing (32–34),
although this is a controversial topic (35)].
Editing in the normal repertoire
Although targeted Ig replacement transgenics represent a more
physiological model than conventional Ig transgenics and can
undergo nested rearrangements with the endogenous receptor
gene elements in cis, they nevertheless distort and accelerate
B-cell development, owing to their premature expression
(Fig. 9). In an attempt to assess the frequency of editing in a
non-transgenic immune system, we assessed the fate of the
Ig-k locus in B cells that expressed l-chains. Because, in Ig-lcells, the k-loci are often deleted by receptor selection (RS)
rearrangement (36–39), we focused on a subset of cells that
retained, but inactivated, the previously rearranged VJk by RS
recombination to the intronic site (Fig. 9). We found that
40–50% of the cells had inactivated in-frame, apparently
functional, rearranged k genes (40). The high frequency,
which was significantly higher than what could be achieved
by random rearrangements, suggested that tolerance often
0.25 60
50
40
30
20
10
0Treatment: – +
0.2
0.15
0.1
0.05
0
% VJκ1-bearing hybridoma in cell mix
% V
Jκ1/
actin
rel
ativ
e in
tens
ity
0
0 1 2 3 4 5 6 7 8 9 10 11
10 20 30 40 50
% B
cel
ls w
ith V
Jκ1
rear
rang
emen
t
0
3–83µ/δ 3–83µ/δ Bcl-2
0.2 2 10 20 40 Mix No tre
atm
ent
No tre
atm
ent
Anti-I
d m
Ab
Anti-I
d m
Ab
VJκ1
Actin
A
B C
% VJκ1 standard curve
Standard curve for quantitation
p = 0.0065
Fig. 4. Induction of Vk-Jk1 joining after
B-cell receptor (BCR) ligation in bone
marrow cultures. (A) Polymerase chainreaction (PCR) analysis of endogenous VJk1rearrangements (top) and control, a-actingene (bottom) after 48 h of culture. Lanes1–6 show data for control mixtures ofvarious proportions of VJk1-positive andVJk1-negative hybridoma cell lines. Lanes8 and 10 show untreated cultures for bonemarrow from a 3–83m/d mouse, and a3–83m/d/Bcl-2 mouse, respectively. Lanes 9and 11 show the anti-idiotype (Id)-treatedcultures for bone marrow from a 3–83m/dmouse and a 3–83m/d/Bcl-2 mouse,respectively. (B) Standard curve derived from(A) relating band intensity to percentage ofVk-Jk1 rearrangement. (C) Estimatedfraction of control and anti-Id-treated B cellsthat bear Vk-Jk1 rearrangement after 48 h oftreatment (6). Reproduced from: Hertz M,Namazee D. BCR ligation induces receptorediting in IgMþIgD� bone marrow cells invitro. Immunity 1997;6:429–436 (also Fig. 6reproduced from this reference).
Aıt-Azzouzene et al �Receptor editing
222 Immunological Reviews 197/2004

promoted RS rearrangement amongst normal B cells. A similar
study in the human system came up with similar results, albeit
with a somewhat lower frequency of in-frame rearrangements
(41).
These experiments were taken a step further by ‘repairing’
an inactivated k-allele that was in-frame but silenced by RS
recombination to the intronic site and expressing it alone or
together with that cell’s H-chain in transgenic mice. The cell
we studied was hybridoma 2H11, which inactivated a Vk-Jk5
gene by RS recombination to the k intron and then went on to
express a l L-chain. We found that, when expressed individu-
ally, the 2H11 mH and repaired k-L-chain transgenics sup-
pressed endogenous rearrangements relatively efficiently. The
mH-chain transgenic mouse suppressed the appearance of
endogenous IgM allotype (Fig. 10A), whereas the k transgenic
prevented (excluded) the surface expression of Ig-l when
bred to the Ig-k–/– background (Fig. 10B, center column of
panels). In contrast, in mice carrying both transgenes, lexpression was not suppressed, indicating that the specificity
of the transgenic H/L pair was stimulating editing, probably
because of autoreactivity (in this particular transgenic model,
the L-chain transgene appears to be expressed on every cell,
possibly because its ectopic chromosomal location and lack
of RS elements prevent its inactivation by editing and force
cells to express additional L-chains to dilute out its func-
tion).
Affinity threshold of editing
Taken at face value, these results suggest that receptor editing
may play a major role in antigen receptor quality control and
40
30
20
10
00 24
Rel
ativ
e R
AG
-2 e
xpre
ssio
n
Rel
ativ
e V
κ-Jκ
1 re
arra
ngem
ents
Time after IL-7 removal (h)48
40
30
20
10
00 24 48
Germline κ
Vκ
VJ23–83 Tg light chain
J1
H3H3
2.8 kb
3.2 kb
3–83 Tgκ+/–
Tg Jκ (3.2 kb)
Germline Jκ (2.8 kb)
% germline Jκ
3–83 Tg 3–83 TgNon-Tg
J2 J3
J3
J4
J4
J5
J5
Cκ
Cκ
B-cells cultureLiver DNA from
No Ab 54.1
96 31 100 217
κ+/– κ+/– κ+/–
S17 S17
1.0 7.1
S17op42
Anti-Id (54.1)
Ant
i-λ
JH1
JH2
JH1
JH2
JH1
JH2
JH2
VHJ558
VH7183
VHQ52
VHGAM3-8
Actin
A B C
D
F
ENormalspleen S17 S17 op42 S17 op42
+IL-7 No IL-73–83 Tg B-cell culture
VDJ
Fig. 5. Editing in interleukin-7 (IL-7)-cultured 3–83 transgenic (Tg)B cells. (A) Antigen-dependent recombination-activating gene (RAG)mRNA induction after IL-7 withdrawal. (B–D) k-Chain gene assemblyand expression. Semi-quantitative Southern blot in (D) using thestrategy shown estimates only a 31% retention of the germline
configuration in anti-idiotype (Id)-treated B cells. (E) Absence ofendogenous H-chain VDJ assembly in editing cells, as assessed bypolymerase chain reaction. The transgenic VH gene is a VHGAM3-8family member. (F) New sIg-l cell-surface expression is associatedwith antigen-induced editing (7).
Aıt-Azzouzene et al �Receptor editing
Immunological Reviews 197/2004 223

immune tolerance. Editing can occur in most, if not all, B cells,
and it appears to occur frequently in a normal immune system,
often as a response to autoreactivity. Various theoretical and
experimental estimates of the precursor frequency of autoreact-
ive B cells in the generating repertoire range from 50 to 90%
(22, 40, 42–45). This frequency raises the question of the
affinity threshold of editing. We have addressed this point
using the MHC alloantigen system and 3–83 transgenic mice,
taking advantage of the fact that the 3–83 antibody recognizes
different H-2K alloforms with a wide range of affinities. Using
readouts for editing, including bone marrow RAG expression
and emergence of Ig-lþ/3–83Hþ cells (Fig. 11), we found that
cell-bound MHC molecules with affinities of <5� 104M�1
for the 3–83 receptor were able to stimulate very efficient
editing (46).
These findings suggest that massive receptor editing stimu-
lated by negative selection in the bone marrow could make a
major impact on the immune repertoire. Because of the likely
massive scope of negative selection along with kinetic delays
and cell losses associated with editing, an initially small non-
autoreactive subset should increase in frequency as develop-
ment progresses. Comparisons between the pre-selected and
post-selected repertoires could therefore easily lead to the
erroneous conclusion that the cells not subject to negative
selection are ‘positively selected’.
Limits to editing and the sparing of a B-1 receptor
Although the low-affinity threshold for editing in our
transgenic system suggests that most autoreactive B cells
should be efficiently eliminated, functional autoreactive
B cells are demonstrable in normal mice, especially those
cells reactive to intracellular antigens (47–49). Some auto-
reactive B cells appear to be actively positively selected, at
least into the B-1 and marginal zone compartments (16,
50–53). Many B-1 cells appear to be polyreactive, and
these cells possibly represent a unique cell lineage or
set of antigenic specificities (54). In addition, B-1 cells
arise predominantly from fetal precursors. We wondered
whether adult bone marrow does not normally generate B-
1 type cells, because bone marrow precursors with B-1
specificity are subject to editing. The work of Michael
Chumley and colleagues (55) addressed this problem in a
novel way, by comparing B cells of transgenic mice carry-
ing the 3–83 receptor with those carrying a VH11/Vk9
B-1-derived receptor, which can bind to liposomes made
2.5
2
1.5
1
0.5
0
0
5
10
* *
*15
% λ
+ B
cel
ls%
λ+ B
cel
ls
20
No
trea
tmen
t
Con
trol
mA
b
Ant
i-Id
mA
b
Ant
i-κ m
Ab
Ant
i-IgD
mA
b
No
trea
tmen
t
Con
trol
mA
b
Ant
i-κ (
1 µg
/ml)
Ant
i-κ (
10 µ
g/m
l)
Ant
i-κ (
100
µg/m
l)
Ant
i-IgD
(10
0 µg
/ml)
A
B
P < 0.0001
P < 0.0001
Fig. 6. Anti-B-cell receptor (BCR)-induced editing in ex vivo bone
marrow cultures as measured by the appearance of lþ cells. Otherexperiments verified that the increases seen were not the result ofselective death of kþ cells or proliferation of lþ cells (6). (A) 3–83transgenic total bone marrow cells were challenged in vitro with 10 mg/mlof the indicated antibodies and the appearance of lþ cells measured. (B)Non-transgenic bone marrow cells were challenged with antibodies asshown and the appearance of lþ cells enumerated.
Aıt-Azzouzene et al �Receptor editing
224 Immunological Reviews 197/2004

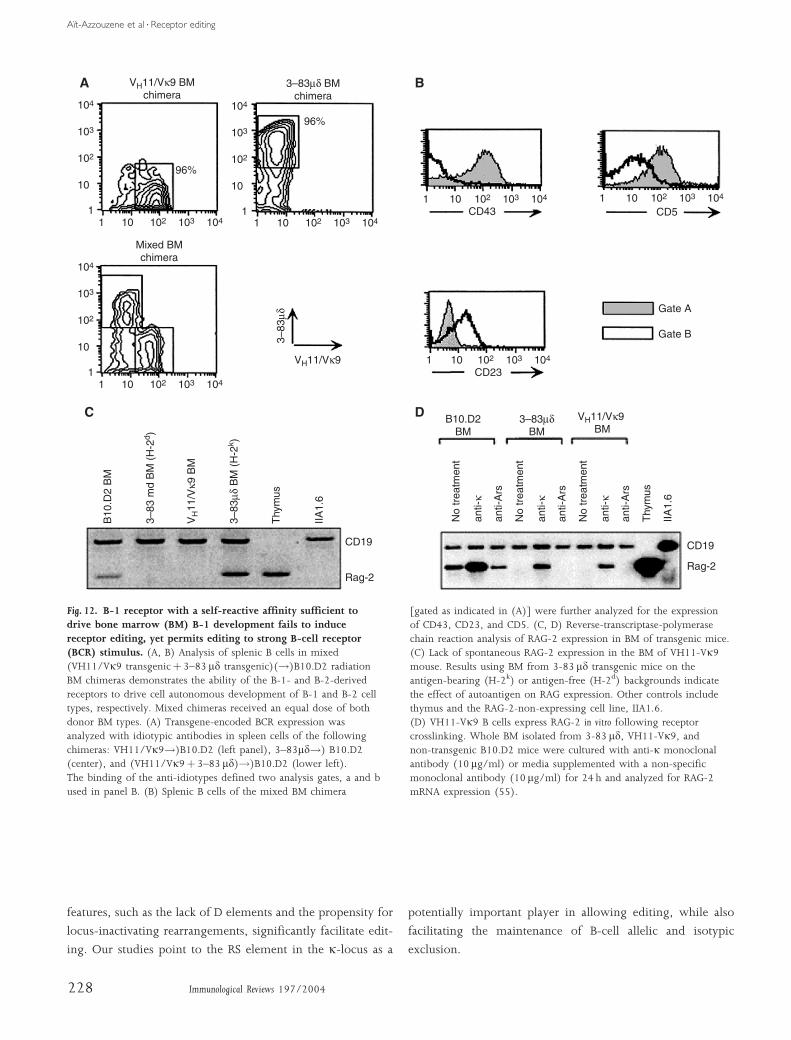
up of phosphatidylcholine, a constituent of the plasma
membrane of all mouse cells. This study had four interest-
ing findings, illustrated in Fig. 12. First, bone marrow from
the particular VH11/Vk9 line used could give rise to B-1 cells
when transferred to irradiated mice, as judged by cell-surface
markers and cell-survival characteristics, whereas mice reconsti-
tuted with 3–83 cells had a B-2 phenotype. This finding sug-
gested that cells constrained by transgenesis to express a B-1
receptor can generate B-1 cells frommouse bone marrow in vivo,
upon reconstitution of lethally irradiated mice. Second, there
was no evidence that VH11/Vk9-expressing cells were
negatively selected. Indeed, the VH11/Vk9 transgenic bone
marrow cells appeared to suppress RAG expression, an indica-
tion that editing was not occurring, and the cells actually
had improved in vivo and in vitro survival compared to 3–83
cells, i.e. B-2 type cells. Third, IL-7-expanded bone marrow B
cells carrying the B-1 receptor rapidly acquired B-1 character-
istics upon in vitro differentiation. Finally, we found that the
VH11/Vk9 transgenic bone marrow B cells were competent
to edit, if challenged with anti-BCR antibody in vitro. These
studies suggest that cells with certain autoreactive specificities
are not negatively selected by editing, possibly because their
affinities are too low or because these specificities are associated
with unique coreceptor signals that drive B-1 type differentia-
tion.
Why should the receptor-editing mechanism exist?
If tolerance can occur by deletion in the periphery, why is
editing necessary? The answer may lie in the high frequency of
autoreactivity in the pre-selected compartment and the con-
sequences that deletion would have on cell loss. Editing may
be a thrifty approach to tolerance. A second reason may be that
peripheral B cells must be ready to respond to antigen much
more quickly than immature B cells, and tolerance in these
cells may need to be correspondingly slower and contingent
on T-cell signals. Therefore, peripheral tolerance must be slow
and perhaps can never be complete.
The macroself antigen approach
A major future direction of research will be into the molecular
mechanisms of editing and B-cell positive selection. Studies of
signaling protein gene knockouts have begun to suggest
important players in B-cell receptor signaling. However, muta-
tions that block the development may not always be easily
interpretable, because, as we have seen, at the immature B-cell
stage, excessive BCR signaling may arrest development and
promote ongoing rearrangements similarly to subnormal
BCR signaling. As a method of screening signaling mutants
quickly for effects on B-cell tolerance, we have recently
V1
V1
V1
V1
V1
V2
V2
V2
V2
V2
V3
V3
V3
V3
V4
V4
Incompatible signal sequences
Compatible signal sequences facilitate receptor editing
V4
V4
D
D
D J
J
J
J
J
J
J
J
J C
C
C
C
C1°
1°
2°
A
B
‘The first chain’ IgH, TCR-γ, TCR-β?
‘The editing locus’ IgL (κ + λ), TCR-α, TCR-γ?
Fig. 7. Role of gene organization in facilitating or inhibiting receptor
editing. V-, D-, and J-coding elements are flanked by differentrecombination signal sequence that constrain the range of possiblerearrangements in cis. (A) In the initially rearranging locus, such asimmunoglobulin H (IgH), the presence of D elements along with Vgenes in the same transcriptional orientation as the J/C cluster forcesdeletional rearrangements. Primary VDJ assembly cannot be replaced by
recombination using conventional signal sequences. (B) In contrast, inloci without D elements, such as the Ig-k locus, sequentialrearrangements are often possible. In this example, a primary (1�) V4-to-J join is replaced by a subsequent secondary (2�) rearrangement betweenV2 and the downstream J. Such secondary rearrangement permits thereplacement of potentially functional V4-to-J joins, i.e. receptor editing.TCR, T-cell receptor.
Aıt-Azzouzene et al �Receptor editing
Immunological Reviews 197/2004 225

developed a macroself antigen approach. As shown in Fig. 13, a
macroself antigen is a synthetic superantigen based on single-
chain antibodies. For the study of central tolerance, we express
an anti-k specificity as a ubiquitous membrane protein. Such
mice lack kþ cells in the peripheral lymphoid organs and
have an increase in the frequency and absolute numbers of
lþ cells. Such macroself antigen-expressing mice are interest-
ing adoptive hosts for bone marrow transfers from mutant
strains of mice, potentially allowing a rapid assessment of
mutations that affect editing and other types of negative selec-
tion.
Conclusion
From this survey of work carried out over a number of
years, the picture emerges that tolerance-induced receptor
editing is a major player in the somatic learning process,
which is a unique feature of the immune system. Editing is
a process that occurs often with great efficiency and min-
imal cell loss. In addition, the affinity threshold of editing
is such that it may eliminate many potentially dangerous
specificities. Editing is also a pathway available in the case
of receptors that fail positive selection, adding to its poten-
tial value as a quality control and repair mechanism and
raising questions about the signals from the antigen recep-
tor that are involved in this process. We still have much to
learn about the parameters of these signals, how they are
regulated, and what might go wrong in disease. For example,
we do not really know what phenotype to predict in a
mutant individual who is unable to carry out editing.
It may well be that such defects contribute to certain auto-
immune conditions. It remains possible that B cells are
heterogeneous with respect to their antigen receptor signal-
ing thresholds for editing, which might underlie the func-
tional and phenotypic heterogeneity of B-cell subsets such as
marginal zone, follicular, and B-1 populations.
An important insight arising from these studies is into the
interplay between antigen receptor gene structure and editing.
Relative to the heavy-chain locus, the k-locus structure pro-
vides only a minor contribution to diversity, but these same
104
103
13.5%
42.1%44.2%
40.9%
102
101
104
103
102
101
0
Donors
Idio
type
0
IgM
Recipients
H-2d H-2b
3–83KI→
3–83KIH→
Fig. 8. Both underexpressed and harmfulB-cell receptors are subject to editing. Datashown are from a bone marrow chimeraexperiment in which donor bone marrowfrom 3-83 knockin heterozygous (3–83KI)or homozygous (3–83KIH) mice developedin the presence (H-2b) or absence (H-2d) of3–83 ligand. 3–83KI B cells only poorlyretained the transgenic specificity on aninnocuous, H-2d background (top left),whereas 3–83KIH B cells express the 3–83receptor well on the H-2d background (lowerleft) and efficiently edit these receptors in thepresence of H-2Kb autoantigen (lower right).Note that tolerance alters receptor specificity,eliminating the autoreactive idiotype, butdoes not alter B-cell numbers in the spleen(29). IgM, immunoglobulin M.
V’s VJ J’s IRS1 iE Cκ RS
RS probeIVS probe
3′E
AVRS VJ-intron-RS
A CB BPCR primers PCR primers
A
B C
Fig. 9. How receptor selection (RS) rearrangements to the Jk-Ckintron inactivate the k-locus and retain VJ joins. A rearranged,potentially functional k-locus (A) can be silenced by two types of RSrecombination (B, C). Type (C) retains the prior Vk-Jk join. (C) RSrecombination eliminates the known enhancers that are critical forefficient rearrangement and expression, thus freezing the locus to furtherV-J recombination. These silenced VJ joins are often in-frame andapparently functional (40). PCR, polymerase chain reaction.
Aıt-Azzouzene et al �Receptor editing
226 Immunological Reviews 197/2004

A
B
104
104
103
103
102
102
101
100
101100 100100
104
104
103
103
102
102
101
100 100 100
100
100100
100100
100
101100 100 100
100 100 100
100 100 100
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101
Spleen
Lymphnode
Bonemarrow
λ
κ
H-only L-only
IgMb
H + L B220+ gatedTransgene
51% 0.5%
1%55%
14%
92%
11% 84%
79%
16%5%
90%
51% 35%
61%
2% 14%
67%
14%
IgMa
Non-transgenic H-chain transgenicFig. 10. Recapitulation of L-chainrearrangements in transgenic mice
carrying antibody H-chain andreconstructed k-chain from recombining
sequence (RS)-edited cell 2H11. (A)H-chain transgene of 2H11 excludesendogenous H-chain expression. Thetransgene fragment was engineered to havethe 2H11 VDJ and flanking genomicsequences upstream of a genomic Cm cassettederived from BALB/c (‘a’ allotype). Thegenetic background of the mice was B10.D2(‘b’ allotype). (B) Allelic exclusion in 2H11L-chain-only transgenic but not in HþL-transgenic mice (arrow) suggests aspecificity-dependent induction of receptorediting. All transgenic mice were crossed tomice lacking endogenous k loci; therefore allk protein is transgenic and all l protein isendogenously encoded. For clarity, cellsanalyzed were gated on B220þ populations.In both 2H11 H-chain only and HþLtransgenics, endogenous H-chain expressionwas suppressed (Fig. 3).
8
6
4
2
0
% Ig
Da
λ+ c
ells
d
H-2 of 3–83 transgenic mice
17
2
107
45
2
5
7 13
k b bm3 bm6 bm8 bm11 Dκ MT-Kb
Non-Tg
10
12
Fig. 11. Low-affinity ligands that mediate efficient central deletion
induce receptor editing as measured by appearance of 3–83-idiotype-negative B cells that coexpress transgenic H-chain (IgDa)and endogenous L-chain (l) (46).
Aıt-Azzouzene et al �Receptor editing
Immunological Reviews 197/2004 227
features, such as the lack of D elements and the propensity for
locus-inactivating rearrangements, significantly facilitate edit-
ing. Our studies point to the RS element in the k-locus as a
potentially important player in allowing editing, while also
facilitating the maintenance of B-cell allelic and isotypic
exclusion.
104
104
103
103
102
102
10
101
1
104
104
103
103
102
102
10
101
1
104103102101
104103102101
104103102101
104
104
103
103
102
102
10
101
1
VH11/Vκ9 BMchimera
Mixed BMchimera
3–83µδ BMchimera
CD43 CD5
CD23
B10
.D2
BM
3–83
md
BM
(H
-2d )
Thy
mus
Thy
mus
IIA1.
6
IIA1.
6
CD19
Rag-2
CD19
Rag-2
No
trea
tmen
t
Gate A
Gate B
anti-
κ
anti-
Ars
No
trea
tmen
t
anti-
κ
anti-
Ars
No
trea
tmen
t
anti-
κ
anti-
Ars
VH11
/Vκ9
BM
3–83
µδ B
M (
H-2
k )
B10.D2BM
3–83µδBM
VH11/Vκ9
3–83
µδ
VH11/Vκ9 BM
A B
DC
96%
96%
Fig. 12. B-1 receptor with a self-reactive affinity sufficient todrive bone marrow (BM) B-1 development fails to induce
receptor editing, yet permits editing to strong B-cell receptor(BCR) stimulus. (A, B) Analysis of splenic B cells in mixed(VH11/Vk9 transgenicþ 3–83 md transgenic)(!)B10.D2 radiationBM chimeras demonstrates the ability of the B-1- and B-2-derivedreceptors to drive cell autonomous development of B-1 and B-2 celltypes, respectively. Mixed chimeras received an equal dose of bothdonor BM types. (A) Transgene-encoded BCR expression wasanalyzed with idiotypic antibodies in spleen cells of the followingchimeras: VH11/Vk9!)B10.D2 (left panel), 3–83md!) B10.D2(center), and (VH11/Vk9þ 3–83 md)!)B10.D2 (lower left).The binding of the anti-idiotypes defined two analysis gates, a and bused in panel B. (B) Splenic B cells of the mixed BM chimera
[gated as indicated in (A)] were further analyzed for the expressionof CD43, CD23, and CD5. (C, D) Reverse-transcriptase-polymerasechain reaction analysis of RAG-2 expression in BM of transgenic mice.(C) Lack of spontaneous RAG-2 expression in the BM of VH11-Vk9mouse. Results using BM from 3-83 md transgenic mice on theantigen-bearing (H-2k) or antigen-free (H-2d) backgrounds indicatethe effect of autoantigen on RAG expression. Other controls includethymus and the RAG-2-non-expressing cell line, IIA1.6.(D) VH11-Vk9 B cells express RAG-2 in vitro following receptorcrosslinking. Whole BM isolated from 3-83 md, VH11-Vk9, andnon-transgenic B10.D2 mice were cultured with anti-k monoclonalantibody (10 mg/ml) or media supplemented with a non-specificmonoclonal antibody (10 mg/ml) for 24 h and analyzed for RAG-2mRNA expression (55).
Aıt-Azzouzene et al �Receptor editing
228 Immunological Reviews 197/2004

104
104
103
6%
25%
26%
47%
>0.2%
>0.2%
103
102
102
101
100
100
100
100
100
100
101
104
104
103
103
102
102
101
101
104
104
103
103
102
102
101
101104103102101
0
20
40
60
80
100
0
20
40
60
Cou
nts
Cou
nts
80
100
104103102101100 100
Rat-IgG1PE Rat-IgG1PEAnti-rat-IgG Fc
Anti-κ Fv
Rat γ1-CH1,2, hinge
H-2Kb Tm/cyto
Igκ macroself Tgline 11
Ig-κ macroself Tgline 11
Transgene surface expression
Igκ macroself Tgline 2
Igκ macroself Tgline 2
Wildtype
A C
B
B220
Tg
Tg
WT
Ig-κ
Fig. 13. Anti-k macroself antigen-transgenic (Tg) mice. (A) Apredesigned ligand that is expressed on cell surfaces and reacts withall immunoglobulin (Ig)-kþ cells. Transgenic mice expressing sucha molecule ubiquitously were generated. (B) Expression pattern of
transgene-encoded protein on blood leukocytes. (C) Elimination of Ig-kcells in the blood of the transgenic mice. A large population of Ig-l cellsis present (not shown), which accounts for the high frequency of B220þ
cells. WT, wildtype.
References
1. Burnet FM. The clonal selection theory of
acquired immunity. Cambridge: The
University Press, 1959.
2. Talmage DW. Immunological specificity:
unique combinations of selected natural
globulins provide an alternative to the classical
concept. Science 1959;129:1643–1648.
3. Nemazee D. Receptor selection in B and T
lymphocytes. Annu Rev Immunol
2000;18:19–51.
4. Tiegs SL, Russell DM, Nemazee D. Receptor
editing in self-reactive bone marrow B cells.
J Exp Med 1993;177:1009–1020.
5. Lang J, et al. Enforced Bcl-2 expression
inhibits antigen-mediated clonal elimination
of peripheral B cells in an antigen dose-
dependent manner and promotes receptor
editing in autoreactive, immature B cells. J Exp
Med 1997;186:1513–1522.
6. Hertz M, Nemazee D. BCR ligation induces
receptor editing in IgMþIgD– bone marrow B
cells in vitro. Immunity 1997;6:429–436.
7. Melamed D, Nemazee D. Self-antigen does not
accelerate immature B cell apoptosis, but
stimulates receptor editing as a consequence
of developmental arrest. Proc Natl Acad Sci
USA 1997;94: 9267–9272.
8. Melamed D, Benschop RJ, Cambier JC,
Nemazee D. Developmental regulation of B
lymphocyte immune tolerance
compartmentalizes clonal selection from
receptor selection. Cell 1998;92:173–182.
9. Sandel PC, Monroe JG. Negative selection of
immature B cells by receptor editing or
deletion is determined by site of antigen
encounter. Immunity 1999;10:289–299.
10. Sandel PC, Gendelman M, Kelsoe G, Monroe
JG. Definition of a novel cellular constituent
of the bone marrow that regulates the
response of immature B cells to B cell antigen
receptor engagement. J Immunol
2001;166:5935–5944.
11. Tze LE, Hippen KL, Behrens TW. Late
immature B cells (IgM (high) IgD (neg))
undergo a light chain receptor editing
response to soluble self-antigen. J Immunol
2003;171:678–682.
12. Chen C, Prak EL, Weigert M. Editing disease-
associated autoantibodies. Immunity
1997;6:97–105.
13. Erikson J, Radic MZ, Camper SA, Hardy RR,
Carmack C, Weigert M. Expression of anti-
DNA immunoglobulin transgenes in non-
autoimmunemice. Nature 1991;349: 331–334.
14. Gay D, Saunders T, Camper S, Weigert M.
Receptor editing: an approach by autoreactive
B cells to escape tolerance. J Exp Med
1993;177:999–1008.
15. Li H, Jiang Y, Prak EL, Radic M, Weigert M.
Editors and editing of anti-DNA receptors.
Immunity 2001;15:947–957.
16. Li Y, Li H, Weigert M. Autoreactive B Cells in
the marginal zone that express dual receptors.
J Exp Med 2002;195:181–188.
Aıt-Azzouzene et al �Receptor editing
Immunological Reviews 197/2004 229

17. Prak EL, Trounstine M, Huszar D, Weigert M.
Light chain editing in kappa-deficient
animals: a potential mechanism of B cell
tolerance. J Exp Med 1994;180:1805–1815.
18. Radic MZ, Erikson J, Litwin S, Weigert M. B
lymphocytes may escape tolerance by revising
their antigen receptors. J Exp Med 1993;177:
1165–1173.
19. Pewzner-Jung Y, Friedmann D, Sonoda E,
Jung S, Rajewsky K, Eilat D. B cell deletion,
anergy, and receptor editing in ‘knock in’
mice targeted with a germline-encoded or
somatically mutated anti-DNA heavy chain. J
Immunol 1998;161:4634–4645.
20. Yachimovich N, Mostoslavsky G, Yarkoni Y,
Verbovetski I, Eilat D. The efficiency of B cell
receptor (BCR) editing is dependent on BCR
light chain rearrangement status. Eur J
Immunol 2002;32:1164–1174.
21. Luning Prak E, Weigert M. Light chain
replacement: a new model for antibody gene
rearrangement. J Exp Med 1995;182:
541–548.
22. Casellas R, et al. Contribution of receptor
editing to the antibody repertoire. Science
2001;291:1541–1544.
23. Pelanda R, Schaal S, Torres RM, Rajewsky K. A
prematurely expressed Ig (kappa) transgene,
but not V (kappa) J (kappa) gene segment
targeted into the Ig (kappa) locus, can rescue
B cell development in lambda5-deficient
mice. Immunity 1996;5:229–239.
24. Gollahon KA, Hagman J, Brinster RL, Storb U.
Ig lambda-producing B cells do not show
feedback inhibition of gene rearrangement. J
Immunol 1988;141:2771–2780.
25. Ritchie KA, Brinster RL, Storb U. Allelic
exclusion and control of endogenous
immunoglobulin gene rearrangement in
kappa transgenic mice. Nature
1984;312:517–520.
26. Storb U. Transgenic mice with
immunoglobulin genes. Annu Rev Immunol
1987;5:151–174.
27. Goldmit M, Schlissel M, Cedar H, Bergman Y.
Differential accessibility at the kappa chain
locus plays a role in allelic exclusion. EMBO J
2002; 21:5255–5261.
28. Mostoslavsky R, et al. Kappa chain monoallelic
demethylation and the establishment of allelic
exclusion. Genes Dev 1998;12:1801–1811.
29. Kouskoff V, Lacaud G, Pape K, Retter M,
Nemazee D. B cell receptor expression level
determines the fate of developing B
lymphocytes: receptor editing versus
selection. Proc Natl Acad Sci USA 2000;97:
7435–7439.
30. Braun U, Rajewsky K, Pelanda R. Different
sensitivity to receptor editing of B cells from
mice hemizygous or homozygous for targeted
Ig transgenes. Proc Natl Acad Sci USA
2000;97:7429–7434.
31. Pelanda R, Schwers S, Sonoda E, Torres RM,
Nemazee D, Rajewsky K. Receptor editing in a
transgenic mouse model: site, efficiency, and
role in B cell tolerance and antibody
diversification. Immunity 1997;7:765–775.
32. McMahan CJ, Fink PJ. RAG reexpression and
DNA recombination at T cell receptor loci in
peripheral CD4þ T cells. Immunity
1998;9:637–647.
33. McGargill MA, Derbinski JM, Hogquist KA.
Receptor editing in developing T cells. Nat
Immunol 2000;1:336–341.
34. Wang F, Huang CY, Kanagawa O. Rapid
deletion of rearranged T cell antigen receptor
(TCR) Valpha-Jalpha segment by secondary
rearrangement in the thymus: role of
continuous rearrangement of TCR alpha chain
gene and positive selection in the T cell
repertoire formation. Proc Natl Acad Sci USA
1998;95:11834–11839.
35. Buch T, Rieux-Laucat F, Forster I, Rajewsky K.
Failure of HY-specific thymocytes to escape
negative selection by receptor editing.
Immunity 2002;16:707–718.
36. Durdik J, Moore MW, Selsing E. Novel kappa
light-chain gene rearrangements in mouse
lambda light chain-producing B lymphocytes.
Nature 1984;307:749–752.
37. Moore MW, Durdik J, Persiani DM, Selsing E.
Deletions of kappa chain constant region
genes in mouse lambda chain-producing B
cells involve intrachromosomal DNA
recombinations similar to V-J joining. Proc
Natl Acad Sci USA 1985;82:6211–6215.
38. Siminovitch KA, Moore MW, Durdik J, Selsing
E. The human kappa deleting element and the
mouse recombining segment share DNA
sequence homology. Nucleic Acids Res
1987;15:2699–2705.
39. Hieter PA, Korsmeyer SJ, Waldmann TA,
Leder P. Human immunoglobulin kappa
light-chain genes are deleted or rearranged in
lambda-producing B cells. Nature
1981;290:368–372.
40. Retter MW, Nemazee D. Receptor editing
occurs frequently during normal B cell
development. J Exp Med 1998;188:
1231–1238.
41. Brauninger A, Goossens T, Rajewsky K,
Kuppers R. Regulation of immunoglobulin
light chain gene rearrangements during early
B cell development in the human. Eur J
Immunol 2001;31:3631–3637.
42. Nemazee D. Antigen receptor ‘capacity’ and
the sensitivity of self-tolerance. Immunol
Today 1996;17:25–29.
43. Nemazee D. Theoretical limits to massive
receptor editing in immature B cells. Curr Top
Microbiol Immunol 1998;229:163–171.
44. Wardemann H, Yurasov S, Schaefer A, Young
JW, Meffre E, Nussenzweig MC. Predominant
autoantibody production by early human B
cell precursors. Science 2003;301:
1374–1377.
45. Louzoun Y, Luning PE, Friedman T, Litwin S,
Weigert M. Comment on Langman and Cohn.
Semin Immunol 2002;14:231–232.
46. Lang J, Jackson M, Teyton L, Brunmark A,
Kane K, Nemazee D. B cells are exquisitely
sensitive to central tolerance and receptor
editing induced by ultralow affinity,
membrane-bound antigen. J Exp Med
1996;184:1685–1697.
47. Dighiero G, et al. Murine hybridomas
secreting natural monoclonal antibodies
reacting with self antigens. J Immunol
1983;131:2267–2272.
48. Rolink AG, Radaszkiewicz T, Melchers F. The
autoantigen-binding B cell repertoires of
normal and of chronically graft-versus-host-
diseased mice. J Exp Med 1987;165:
1675–1687.
49. McHeyzer-Williams MG, Nossal GJ. Clonal
analysis of autoantibody-producing cell
precursors in the preimmune B cell repertoire.
J Immunol 1988;141:4118–4123.
50. Hayakawa K, et al. Positive selection of natural
autoreactive B cells. Science 1999;285:
113–116.
51. Chen X, Kearney JF. Generation and function
of natural self-reactive B lymphocytes. Semin
Immunol 1996;8:19–27.
52. Chen X, Martin F, Forbush KA, Perlmutter
RM, Kearney JF. Evidence for selection of a
population of multi-reactive B cells into the
splenic marginal zone. Int Immunol
1997;9:27–41.
53. Gu H, Tarlinton D, Muller W, Rajewsky K,
Forster I. Most peripheral B cells in mice are
ligand selected. J Exp Med 1991;173:
1357–1371.
54. Hardy RR. Development of VH11þ B cells: a
model for selection of B cells producing
natural autoantibodies. Curr Dir Autoimmun
2003;6:196–211.
55. Chumley MJ, Dal Porto JM, Kawaguchi S,
Cambier JC, Nemazee D, Hardy RR. A VH11V
kappa 9 B cell antigen receptor drives
generation of CD5þ B cells both in vivo and in
vitro. J Immunol 2000;164:4586–4593.
56. Kouskoff V, Lacaud G, Nemazee D. T cell-
independent rescue of B lymphocytes from
peripheral immune tolerance. Science
2000;287:2501–2503.
Aıt-Azzouzene et al �Receptor editing
230 Immunological Reviews 197/2004




















