
TNF/IL1/NIK/NF-κB transduction pathway: a comparative study in normal and pathological human...
11
TNF/IL-1/NIK/NF-jB transduction pathway: a comparative study in normal and pathological human prostate (benign hyperplasia and carcinoma) C Nun ˜ ez, 1 J R Cansino, 2 F Bethencourt, 2 M Pe ´rez-Utrilla, 2 B Fraile, 1 P Martı ´nez-Onsurbe, 3 G Olmedilla, 3 R Paniagua 1 & M Royuela 1 1 Department of Cell Biology and Genetics, University of Alcala ´, 2 Department of Urology, ‘‘La Paz’’ Hospital, Madrid and 3 Department of Pathology, Prı ´ncipe de Asturias Hospital, Alcala ´ de Henares, Madrid, Spain Date of submission 4 November 2007 Accepted for publication 25 February 2008 Nun ˜ ez C, Cansino J R, Bethencourt F, Pe ´rez-Utrilla M, Fraile B, Martı ´nez-Onsurbe P, Olmedilla G, Paniagua R & Royuela M (2008) Histopathology 53, 166–176 TNF/IL-1/NIK/NF-jB transduction pathway: a comparative study in normal and pathological human prostate (benign hyperplasia and carcinoma) Aims: Tumour necrosis factor (TNF)-a induces death or cell proliferation by activation of nuclear factor (NF)- jB, also activated by interleukin (IL)-1a. The aim was to investigate upstream and downstream components of NIK transduction pathway in normal (NP), benign prostatic hyperplasia (BPH), prostatic intraepithelial neoplasia (PIN) and prostatic carcinoma (PC). Methods and results: Immunohistochemistry and Wes- tern blotting were performed. In NP, the cytoplasm of epithelial cells was intensely immunoreactive to IL-1 receptor-associated kinase (IRAK), TNF receptor-asso- ciated factor (TRAF)-6, NF-jB inducing kinase (NIK), Ijja ⁄ b,IjBa and p-IjB; weakly to NF-jB-p50; and negative to NF-jB-p65. BPH samples were intensely immunoreactive to IRAK, TRAF-6, NIK, Ijja ⁄ b,IjBa, p-IjB; weakly to NF-jB-p50 and NF-jB-p65. Whereas low-grade PIN showed intermediate results between NP and BPH, results in high-grade PIN were similar to those found in PC (low Gleason). In PC, immunoreac- tivity was intense for IRAK, TRAF-6, NIK, Ijja ⁄ b (increasing with Gleason), IjBa, p-IjB (decreasing with Gleason); weak for NF-jB-p50 and NF-jB-p65 (dec- reasing with Gleason). Nuclear NF-jB was observed in PC. Conclusions: NF-jB enhances cell proliferation, but also ATF-2 or Elk-1. Since IL-1 and TNF-a are related to inflammation and their immunoexpression increases in PC, inhibition of these cytokines might be a possible target for PC treatment, because they decrease the activity of all transduction pathway members that activate transcription factors such as NF-jB, Elk-1 or ATF-2. Keywords: IjB, Ijja ⁄ b, IRAK-1, NF-jB (p50), NF-jB (p65), p-IjB, prostatic cancer, TRAF-6 Abbreviations: BPH, benign prostatic hyperplasia; IL, interleukin; IRAK, IL-1 receptor-associated kinase; NF, nuclear factor; NIK, NF-jB inducing kinase; NP, normal human prostate; PC, prostatic carcinoma; PIN, prostatic intraepithelial neoplasia; TNF, tumour necrosis factor; TRAF, TNF receptor-associated factor Introduction Inflammation has been suggested as a causal factor in several human tumours, including prostatic cancer. This process causes cell and genomic damage, pro- motes cellular replacement and creates a cytokine-rich tissue microenvironment that can enhance cell repli- cation, angiogenesis and tissue repair. 1,2 In this way, the role of proinflammatory cytokines, such as inter- leukin (IL)-1 and tumour necrosis factor (TNF)-a that trigger activation of nuclear factor (NF)-jB (a common Address for correspondence: M Royuela, Department of Cell Biology and Genetics, University of Alcala ´, E-28871 Alcala ´ de Henares (Madrid), Spain. e-mail: [email protected] Ó 2008 The Authors. Journal compilation Ó 2008 Blackwell Publishing Limited. Histopathology 2008, 53, 166–176. DOI: 10.1111/j.1365-2559.2008.03092.x
Transcript of TNF/IL1/NIK/NF-κB transduction pathway: a comparative study in normal and pathological human...

TNF/IL-1/NIK/NF-jB transduction pathway: a comparativestudy in normal and pathological human prostate (benignhyperplasia and carcinoma)
C Nunez,1 J R Cansino,2 F Bethencourt,2 M Perez-Utrilla,2 B Fraile,1 P Martınez-Onsurbe,3
G Olmedilla,3 R Paniagua1 & M Royuela1
1Department of Cell Biology and Genetics, University of Alcala, 2Department of Urology, ‘‘La Paz’’ Hospital, Madrid and3Department of Pathology, Prıncipe de Asturias Hospital, Alcala de Henares, Madrid, Spain
Date of submission 4 November 2007Accepted for publication 25 February 2008
Nunez C, Cansino J R, Bethencourt F, Perez-Utrilla M, Fraile B, Martınez-Onsurbe P, Olmedilla G, Paniagua R &Royuela M
(2008) Histopathology 53, 166–176
TNF/IL-1/NIK/NF-jB transduction pathway: a comparative study in normal and pathologicalhuman prostate (benign hyperplasia and carcinoma)
Aims: Tumour necrosis factor (TNF)-a induces deathor cell proliferation by activation of nuclear factor (NF)-jB, also activated by interleukin (IL)-1a. The aim wasto investigate upstream and downstream componentsof NIK transduction pathway in normal (NP), benignprostatic hyperplasia (BPH), prostatic intraepithelialneoplasia (PIN) and prostatic carcinoma (PC).Methods and results: Immunohistochemistry and Wes-tern blotting were performed. In NP, the cytoplasm ofepithelial cells was intensely immunoreactive to IL-1receptor-associated kinase (IRAK), TNF receptor-asso-ciated factor (TRAF)-6, NF-jB inducing kinase (NIK),Ijja ⁄ b, IjBa and p-IjB; weakly to NF-jB-p50; andnegative to NF-jB-p65. BPH samples were intenselyimmunoreactive to IRAK, TRAF-6, NIK, Ijja ⁄ b, IjBa,p-IjB; weakly to NF-jB-p50 and NF-jB-p65. Whereas
low-grade PIN showed intermediate results between NPand BPH, results in high-grade PIN were similar tothose found in PC (low Gleason). In PC, immunoreac-tivity was intense for IRAK, TRAF-6, NIK, Ijja ⁄ b(increasing with Gleason), IjBa, p-IjB (decreasing withGleason); weak for NF-jB-p50 and NF-jB-p65 (dec-reasing with Gleason). Nuclear NF-jB was observedin PC.Conclusions: NF-jB enhances cell proliferation, but alsoATF-2 or Elk-1. Since IL-1 and TNF-a are related toinflammation and their immunoexpression increases inPC, inhibition of these cytokines might be a possibletarget for PC treatment, because they decrease theactivity of all transduction pathway members thatactivate transcription factors such as NF-jB, Elk-1 orATF-2.
Keywords: IjB, Ijja ⁄ b, IRAK-1, NF-jB (p50), NF-jB (p65), p-IjB, prostatic cancer, TRAF-6
Abbreviations: BPH, benign prostatic hyperplasia; IL, interleukin; IRAK, IL-1 receptor-associated kinase; NF,nuclear factor; NIK, NF-jB inducing kinase; NP, normal human prostate; PC, prostatic carcinoma; PIN, prostaticintraepithelial neoplasia; TNF, tumour necrosis factor; TRAF, TNF receptor-associated factor
Introduction
Inflammation has been suggested as a causal factor inseveral human tumours, including prostatic cancer.
This process causes cell and genomic damage, pro-motes cellular replacement and creates a cytokine-richtissue microenvironment that can enhance cell repli-cation, angiogenesis and tissue repair.1,2 In this way,the role of proinflammatory cytokines, such as inter-leukin (IL)-1 and tumour necrosis factor (TNF)-a thattrigger activation of nuclear factor (NF)-jB (a common
Address for correspondence: M Royuela, Department of Cell Biology
and Genetics, University of Alcala, E-28871 Alcala de Henares
(Madrid), Spain. e-mail: [email protected]
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Limited.
Histopathology 2008, 53, 166–176. DOI: 10.1111/j.1365-2559.2008.03092.x

transcription factor), has been well established inprostatic cancer.
IL-1 plays a crucial role in inflammation, stress, anddisease. IL-1 comprises two ligands (IL-1a and IL-1b),which bind to two types of transmembrane receptors(IL-1RI and IL-1RII) and there is also a specific receptorantagonist (IL-1Ra).3,4 IL-1a and IL-1Ra present highaffinity to IL-1RII.5 IL-1a and IL-1b bound to theirreceptors (RI and RII) promote the association of IL-1receptor-associated kinase (IRAK). In humans, fourdifferent IRAK molecules have been described: IRAK-1,-2, -M, and -4.6,7 They all comprise four domains: aN-terminal death domain, a linker region, a centrallylocated classical serine ⁄ threonine kinase domain and aC-terminal domain. Whereas IRAK-1 and IRAK-4 havebeen identified in all tissues, IRAK-2 and IRAK-M havea narrower cellular distribution.7,8 Genotyping meth-ods have shown that IRAK-1 and IRAK-4 are associ-ated with prostatic cancer risk.9 IRAK-1, the mostdescribed member of the IRAK family, is a membraneproximal serine-threonine kinase that, when phosphor-ylated, forms a complex with TNF receptor-associatedfactor (TRAF)-6.6 TRAF-6 interacts with the C-termi-nus of IRAK-1, which contains three TRAF-6 bindingsites, leading to the activation of several downstreamsignalling pathways, such as the activation of NF-jBinducing kinase (NIK), which is a MAP3K-relatedkinase that activates the IKK complex composed ofIKK-a and IKK-b.10 NIK stimulate IKK-b, whichinduces IKK-a degradation. The IKK complex phospho-rylates IjB, following its ubiquitination and rapiddegradation, causing the nuclear translocation ofNF-jB (Figure 1), which, in turn, activates targetgenes involved in carcinogenesis: tumour initiation,
malignant transformation and metastasis.11 Using theprostatic carcinoma cell lines LNCaP, DU45 and PC3,Gasparian et al.12 found that increased IKK activationleads to the activation of NF-jB. A potential role ofNF-jB in the development of such different tumoursas breast,11,13 melanoma,14 colon,15 pancreas,16 thy-roid17 and prostate18–20 have been reported.
NF-jB represents a group of structurally related andevolutionarily conserved proteins, with five members inmammals: Rel (c-Rel), RelA (p65), RelB, NF-jB1 (p50and its precursor p105), and NF-jB2 (p52 and itsprecursor p100).21 Among the various NF-jB dimers,the complex consisting of p65 and p50 is the mostabundant in most cell types.22 Several authors havedescribed cytoplasmic and nuclear immunoexpressionof NF-jB (p50 and p65) in human benign hyperplasticand carcinomatous prostate.18–20,22
TNF-a induces apoptosis when bound to its receptorTNFRI, which binds in turn to the TNF receptor-associated death domain molecules. These activate aprotein cascade (TRAF-2, ASK-1, MEK-4 and JNK) thatends with the activation of the activator protein-1(Ap-1)23 (Figure 1). However, the TNF-a signal maydiverge to NF-jB because TRAF-2 may also phosphor-ylate and activate NIK.24
The activation of NIK pathway in vivo has not yetbeen adequately addressed. We have previously repor-ted the immunohistochemical expression and localiza-tion of IL-125 and TNF-a26 in normal prostate, benignprostatic hyperplasia (BPH) and prostatic carcinoma(PC). At present, immunohistochemical studies of thedifferent components of IL-1 ⁄ NIK ⁄ NF-jB in prostatictissue are scanty and confused. To our knowledge,although some studies of Ijja ⁄ b, IjBa and p-IjBhave been reported,12,18 there have been no studies ofTRAF-6, IRAK-1 or NIK, and studies on NF-jB arecontradictory.18–20 Prostatic diseases are chronic con-ditions that develop over a long period. Simple micro-nodular hyperplasia needs a long period for evolutionto macroscopic volume enlargement and clinicalexpression. The evolution of PC needs a long period,probably due to the development of early and laterprecancerous modifications and then clinical PC.27 Forthis reason, several clinical studies have suggested thatprostatic intraepithelial neoplasia (PIN) precedes car-cinoma by 10 years or more, with low-grade PIN firstemerging in men in the third decade of life.27–29 Theaim of the present study was to investigate immuno-histochemically the upstream (TRAF-6, IRAK-1) anddownstream (Ijja ⁄ b, IjBa, p-IjB and NF-jB) compo-nents of NIK transduction pathway in normal humanprostate (NP), their modification and evolution inbenign disease (BPH), premalignant lesions (PIN) and
TNF
TNFR1 Plasma membrane
TRADD
RIP
ASK1
MKK4
JNK
AP1 (death)
(Nucleus)
Cytoplasm
IRAK/TRAF-6
IRAK
IL-1
IL-1r1/IL-1RAcP
NIK
IKK-α/IKK-β
Iκβ/NF-κβ
NF-κβ (survival)
TRAF-2
Figure 1. NIK, JNK and p38 transduction pathways triggered by
tumour necrosis factor (TNF)-a and interleukin (IL)-1.
NIK transduction pathway in prostatic cancer 167
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.

PC, as well as their possible involvement in apoptosis orproliferation via TNF-a and IL-1 stimulation.
Materials and methods
Prostatic tissue was obtained from: (i) transurethralresections from 35 men (age 53–88 years) withclinically and histopathologically diagnosed BPH;(ii) radical prostatectomies from 27 men (age 20–59 years) diagnosed with low-grade PIN (n = 12) orhigh-grade PIN (n = 15); (iii) radical prostatectomiesfrom 93 men (age 54–69 years) with PC of low (grades1–6, n = 21), medium (grade 7, n = 47) and high(grade ‡ 8, n = 25) Gleason grades, with and withoutmetastasis or lymph node infiltration at the time ofsurgery; and (iv) histologically normal prostates (NP)obtained at autopsy (8–10 h after death) from 20 men(age 20–38 years) without histories of reproductive,endocrine or related diseases. Each diagnosed samplewas divided into two portions; one was immediatelyprocessed for immunohistochemistry, the other wasfrozen in liquid nitrogen and maintained at )80�Cfor Western blotting analysis. For the latter portion,cryostat sections were stained with toluidine blue toconfirm the histopathological diagnosis. Since PINreplaces synonymous terms including intraductal dys-plasia, marked atypia, hyperplasia with malignantchange or large acinals atypical hyperplasia, reproduc-ibility of the diagnosis of low-grade PIN is lacking. Allpathological, clinical or personal data were anony-mized and separated from any personal identifiers. Thisstudy was made with the consent of the patients’relatives or their family in autopsy cases. All proce-dures were examined and approved by the Principe deAsturias Hospital Ethics Committee (reference numberSAF2007-61928) and were in accordance with theethical standards of the Committee for Human Exper-imentation, with the Helsinki Declaration of 1975(revised in 1983) and the Committee on PublicationEthics (COPE) guidelines.
The primary antibodies used were: rabbit antihumanTNF-a, IL-1 a, IL-1b, IL-1Ra, IL-1RI, Ijja ⁄ b, IRAK-1and NF-jB (p-50); mouse antihuman IL-1RII, TRAF-2,TRAF-6, NIK, IjBa, p-IjB (phosphorylated form of IjB)and NF-jB (p-65); and goat antihuman TNF-RI (SantaCruz Biotechnology, Santa Cruz, CA, USA) diluted1:100 (IjBa), 1:150 (IL-1a, IL-1b, IL-1Ra, IL-1RI),1:200 (TNF-a, TNF-RI, NIK, IRAK, TRAF-6, RIK),1:250 [NF-jB (p50), NF-jB (p65), p-IjB, Ijja ⁄ b,TRAF-2], 1:500 (IL1-RII) and 1:10 000 chicken anti-a-actin (Amersham, Little Chalfont, UK) for Westernblotting; or 1:50 (IL-1a, IL-1b, IL-1Ra, IL-1RI, TRAF-2,Ijja ⁄ b, IjBa, p-IjB and NF-jB ⁄ p65), 1:100 (IL-1RII,
TNF-a, TNF-RI, IRAK, NIK and NF-jB ⁄ p50) and 1:200(TRAF-6) for immunohistochemistry.
Immunochemical procedure specificity was checkedusing negative and positive controls. For negativecontrols, tissues of each type (normal, BPH, PIN andPC) were incubated with blocking peptides (Santa CruzBiotechnology) or preimmune serum at the sameimmunoglobulin concentration used for each antibody.As positive controls, homogenates (for Western blot)and histological sections (for immunohistochemistry) ofhuman skin, thymus or tonsils were incubated with thesame antibodies.
Comparative histological quantification of immuno-labelling among the different types of prostates wasperformed for each antibody. From each prostate, sixhistological sections were selected at random. In eachsection, the intensity of immunoreactivity (opticaldensity) per unit surface area was measured with anautomatic image analyser (MIP4 version 4.4; Consult-ing Image Digital, Barcelona, Spain) in five lightmicroscopic fields per section, using the ·40 objective.Delineation of surface areas was carried out manuallyusing the mouse of the image analyser. For eachimmunopositive section, one negative control section(the next in a series of consecutive sections) was alsoused and the optical density of this control section wassubtracted from that of the immunoreactive section.From the average values obtained (by the automaticimage analyser) for each prostate, the means ± SDfor each prostate type (NP, BPH, PIN and PC) werecalculated. The results were corroborated by twodifferent observers. The statistical significance betweenmeans of the different prostate group samples wasassessed by one-way anova at P £ 0.05, by multiplepairwise comparisons (GraphPad PRISMA 3.0 com-puter program; GraphPad, San Diego, CA, USA).
To determine whether the source of material (sur-gery or autopsy) could be responsible for changes in theimmunohistochemical pattern, five prostatic biopsyspecimens (taken because of the suspicion of prostaticdisease but histological examination revealed a normalpattern) were processed for immunohistochemistry.The results of the quantitative immunohistochemicalstudy in these specimens were compared with those ofautopsy prostates.
Results
western blot analysis
Western blot analysis showed a single band for all theantibodies studied at the corresponding molecularweight in BPH and PC. In normal prostates (NP), only
168 C Nunez et al.
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.

NF-jB ⁄ p65 was not detected, whereas immunoreac-tivity for the other antibodies was found at thecorresponding molecular weight (data not shown).
Comparison of optical densities revealed significantdifferences between the three prostate groups (NP,BPH, PC) (Figures 2 and 3).
immunohistochemistry
No immunoreactivity was observed in the negativecontrols incubated with preimmune serum, or using
the antibodies preabsorbed with an excess of purifiedantigens.
No significant histological or quantitative immuno-histochemical differences between the two subgroupsof normal prostates (biopsies and autopsies) wereobserved.
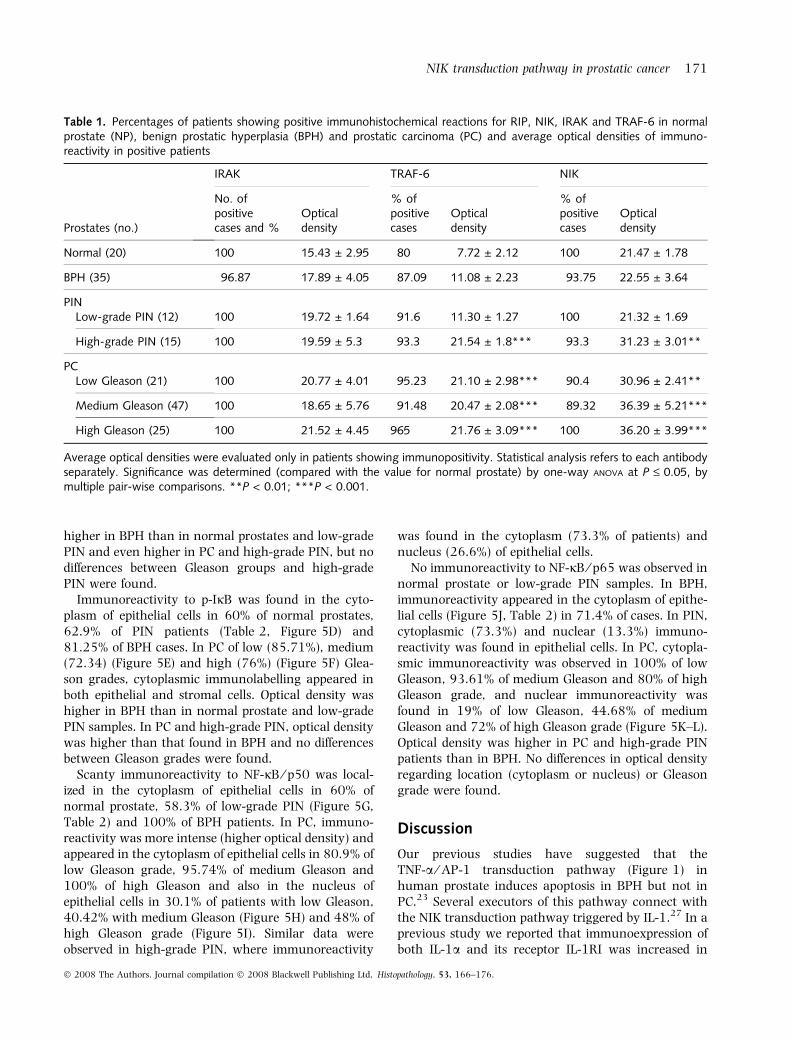
Immunopositivity for IRAK was observed in thecytoplasm of epithelial cells in all normal prostates, in96.87% of patients with BPH (Figure 4A, Table 1) andin all PIN and PC patients (Figure 4B,C and Table 1).Optical density to IRAK was similar in four groups (NP,BPH, PIN and PC). In PC no differences were observedbetween the three Gleason groups. No differences werefound between low- and high-grade PIN.
Immunoreactivity for TRAF-6 appeared in the cyto-plasm of epithelial cells in 80% of normal prostates,87.09% of patients with BPH (Figure 4D, Table 1),92.59% of patients with PIN (Figure 4E) and 90% ofPC patients (Figure 4F and Table 1). Optical densitywas similar in normal, BPH and low-grade PIN. Thehighest optical density was found in PC and high-gradePIN, but no differences were observed between high-grade PIN and the three Gleason groups.
NIK consistently showed immunoreactivity in thecytoplasm of epithelial cells in all normal prostates(Figure 4G, Table 1), 96.29% of PIN samples, 93.75%of BPH patients, 90.4% of cancer patients with lowGleason grade, 89.32% of cancer patients with mediumGleason grade (Figure 4H) and all cancer patients withhigh Gleason grade (Figure 4I). Optical density washigher in PC samples than in normal prostate, low-grade PIN and BPH. No differences were observedbetween PC with low Gleason and high-grade PIN. Thehighest density was found in PC with medium and highGleason grades.
Immunoreactivity to Ijja ⁄ b was present in basalepithelial cells in 80% of normal prostates (Figure 4J,Table 2), 75% of low-grade PIN samples, 53.3% ofhigh-grade PIN, 54.3% of BPH patients, 52.4% of PCwith low Gleason grade, 72.34% of PC with mediumGleason grade and 68% of PC with high Gleason grade.The lowest optical density was found in normalprostate and low-grade PIN, the highest in PC withlow Gleason grade (Figure 4K). No differences wereobserved between PIN (high grade), BPH and PC(medium and high Gleason grades) (Figure 4L).
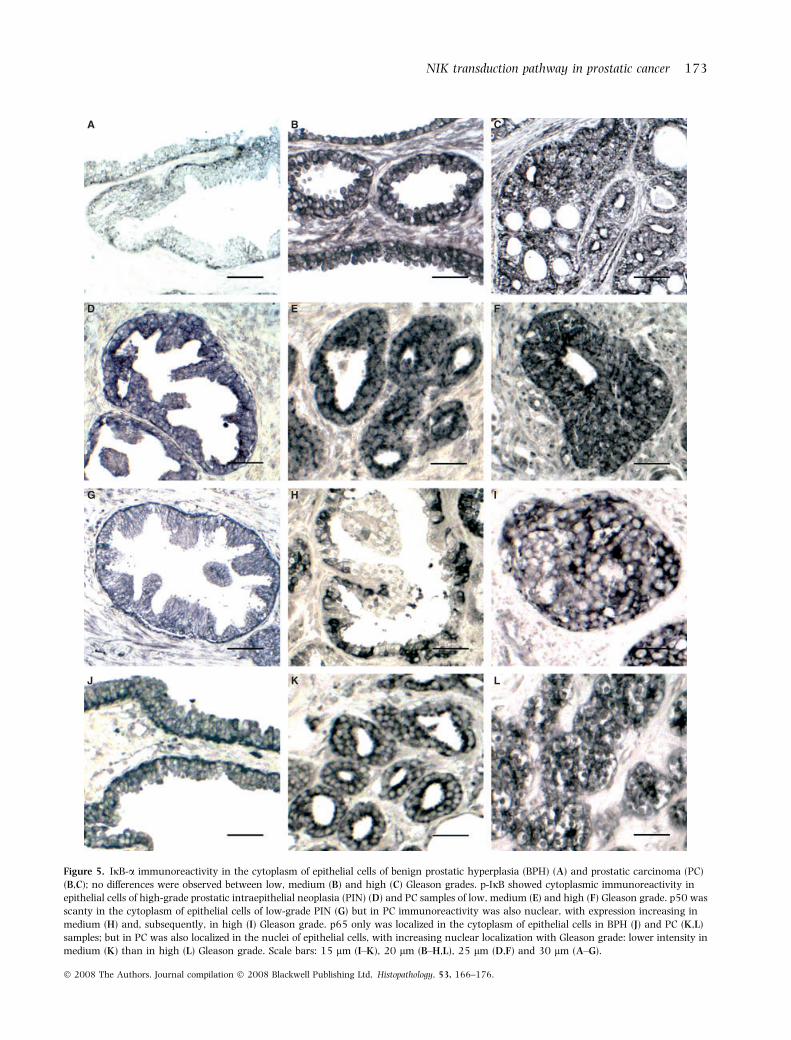
Immunoreactivity to IjBa was observed in thecytoplasm of epithelial cells in 40% of normal prostates(Table 2), 55.5% of PIN samples and 68.57% of BPHpatients (Figure 5A). In PC, the percentage of positivecases increased with the malignancy: 71.4% in lowGleason grade, 78.72% in medium grade (Figure 5B)and 92% in high grade (Figure 5C). Optical density was
IRAK TRAF-6 NIK Actin0
25
50
75
100NP
BPH
PC
Opt
ical
den
sity
Figure 2. Comparison of the intensity of immunoreactivity
(measured as average optical density ± SD) in Western blot analysis
in normal prostate (NP), benign prostatic hyperplasia (BPH) and
prostatic carcinoma (PC). Average optical densities were evaluated
only in patients showing immunopositivity. Statistical analysis refers
to each antibody separately. Significance was determined by one-way
ANOVA test at P £ 0.05, by multiple pair-wise comparisons.
IKK IkBα p-Ikβ p50 p65 Actin 0
25
50
75 PN
BPH
PC
Opt
ical
den
sity
Figure 3. Comparison of the intensity of immunoreactivity
(measured as average optical density ± SD) in Western blot analysis
in normal prostate (NP), benign prostatic hyperplasia (BPH) and
prostatic carcinoma (PC). Average optical densities were evaluated
only in patients showing immunopositivity. Statistical analysis refers
to each antibody separately. Significance was determined by one-way
ANOVA at P £ 0.05, by multiple pair-wise comparisons.
NIK transduction pathway in prostatic cancer 169
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.

A B C
D E F
G H I
J K L
Figure 4. IRAK immunoreactivity in the epithelial cells of benign prostatic hyperplasia (BPH) (A) and prostatic carcinoma (PC) (B,C); no
differences were observed between low, medium (B) and high (C) Gleason grades. The cytoplasm of epithelial cells showed immunopositivity for
TRAF-6 in BPH (D), low-grade prostatic intraepithelial neoplasia (PIN) (E) and PC with medium Gleason grade (F). NIK immunoreactivity in
epithelial cells of normal prostate (G) presents a lower optical density than in PC samples (H,I), but no differences were found between low,
medium (H) and high (I) Gleason grades. Ijj was present in basal epithelial cells in normal prostate (J) but in all cytoplasm of secretory cells in
BPH and PC (K,L); the highest optical density was found in low Gleason (K) and no differences were observed between BPH, medium and high (L)
Gleason. Scale bars: 20 lm (D,F–I,L) and 25 lm (A–C,E,J,K).
170 C Nunez et al.
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.
higher in BPH than in normal prostates and low-gradePIN and even higher in PC and high-grade PIN, but nodifferences between Gleason groups and high-gradePIN were found.
Immunoreactivity to p-IjB was found in the cyto-plasm of epithelial cells in 60% of normal prostates,62.9% of PIN patients (Table 2, Figure 5D) and81.25% of BPH cases. In PC of low (85.71%), medium(72.34) (Figure 5E) and high (76%) (Figure 5F) Glea-son grades, cytoplasmic immunolabelling appeared inboth epithelial and stromal cells. Optical density washigher in BPH than in normal prostate and low-gradePIN samples. In PC and high-grade PIN, optical densitywas higher than that found in BPH and no differencesbetween Gleason grades were found.
Scanty immunoreactivity to NF-jB ⁄ p50 was local-ized in the cytoplasm of epithelial cells in 60% ofnormal prostate, 58.3% of low-grade PIN (Figure 5G,Table 2) and 100% of BPH patients. In PC, immuno-reactivity was more intense (higher optical density) andappeared in the cytoplasm of epithelial cells in 80.9% oflow Gleason grade, 95.74% of medium Gleason and100% of high Gleason and also in the nucleus ofepithelial cells in 30.1% of patients with low Gleason,40.42% with medium Gleason (Figure 5H) and 48% ofhigh Gleason grade (Figure 5I). Similar data wereobserved in high-grade PIN, where immunoreactivity
was found in the cytoplasm (73.3% of patients) andnucleus (26.6%) of epithelial cells.
No immunoreactivity to NF-jB ⁄ p65 was observed innormal prostate or low-grade PIN samples. In BPH,immunoreactivity appeared in the cytoplasm of epithe-lial cells (Figure 5J, Table 2) in 71.4% of cases. In PIN,cytoplasmic (73.3%) and nuclear (13.3%) immuno-reactivity was found in epithelial cells. In PC, cytopla-smic immunoreactivity was observed in 100% of lowGleason, 93.61% of medium Gleason and 80% of highGleason grade, and nuclear immunoreactivity wasfound in 19% of low Gleason, 44.68% of mediumGleason and 72% of high Gleason grade (Figure 5K–L).Optical density was higher in PC and high-grade PINpatients than in BPH. No differences in optical densityregarding location (cytoplasm or nucleus) or Gleasongrade were found.
Discussion
Our previous studies have suggested that theTNF-a ⁄ AP-1 transduction pathway (Figure 1) inhuman prostate induces apoptosis in BPH but not inPC.23 Several executors of this pathway connect withthe NIK transduction pathway triggered by IL-1.27 In aprevious study we reported that immunoexpression ofboth IL-1a and its receptor IL-1RI was increased in
Table 1. Percentages of patients showing positive immunohistochemical reactions for RIP, NIK, IRAK and TRAF-6 in normalprostate (NP), benign prostatic hyperplasia (BPH) and prostatic carcinoma (PC) and average optical densities of immuno-reactivity in positive patients
Prostates (no.)
IRAK TRAF-6 NIK
No. ofpositivecases and %
Opticaldensity
% ofpositivecases
Opticaldensity
% ofpositivecases
Opticaldensity
Normal (20) 100 15.43 ± 2.95 80 7.72 ± 2.12 100 21.47 ± 1.78
BPH (35) 96.87 17.89 ± 4.05 87.09 11.08 ± 2.23 93.75 22.55 ± 3.64
PINLow-grade PIN (12) 100 19.72 ± 1.64 91.6 11.30 ± 1.27 100 21.32 ± 1.69
High-grade PIN (15) 100 19.59 ± 5.3 93.3 21.54 ± 1.8*** 93.3 31.23 ± 3.01**
PCLow Gleason (21) 100 20.77 ± 4.01 95.23 21.10 ± 2.98*** 90.4 30.96 ± 2.41**
Medium Gleason (47) 100 18.65 ± 5.76 91.48 20.47 ± 2.08*** 89.32 36.39 ± 5.21***
High Gleason (25) 100 21.52 ± 4.45 965 21.76 ± 3.09*** 100 36.20 ± 3.99***
Average optical densities were evaluated only in patients showing immunopositivity. Statistical analysis refers to each antibodyseparately. Significance was determined (compared with the value for normal prostate) by one-way ANOVA at P £ 0.05, bymultiple pair-wise comparisons. **P < 0.01; ***P < 0.001.
NIK transduction pathway in prostatic cancer 171
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.

Tab
le2.
Per
centa
ges
of
pat
ients
show
ing
posi
tive
imm
unohis
toch
emic
alre
actions
for
IKK
,p-IjB,
IjB,
p50
and
p65
innorm
alpro
stat
e(N
P),
ben
ign
pro
stat
ichyp
erpla
sia
(BPH
)an
dpro
stat
icca
rcin
om
a(P
C)
and
aver
age
optica
lden
sities
of
imm
unore
activi
tyin
posi
tive
pat
ients
Pro
stat
es(n
o.)
IKK
IjB
p-I
jBp50
p65
No.
of
posi
tive
case
san
d%
Optica
lden
sity
No.
of
posi
tive
case
san
d%
Optica
lden
sity
%of
posi
tive
case
sO
ptica
lden
sity
No.
of
posi
tive
case
san
d%
Optica
lden
sity
%of
posi
tive
case
sO
ptica
lden
sity
Norm
al(2
0)
80
10.2
9±
1.3
140
8.8
8±
2.3
460
8.7
1±
1.8
860
7.8
8±
2.4
0–
BPH
(35)
54.3
15.0
7±
2.4
0*
68.5
716.6
±2.9
5*
81.2
514.3
1±
4.9
3*
100
18.6
±1.7
2*
71.4
9.0
8±
3.2
3
PIN Lo
w-g
rade
PIN
(12)
75
13.6
3±
1.8
150
10.3
1±
1.6
358.3
11.9
5±
1.9
158.3
15.3
2±
2.0
0*
0–
Hig
h-g
rade
PIN
(15)
Cyt
opla
sm53.3
18.6
8±
1.0
5**
60
21.3
4±
2.1
0***
66.6
26.3
6±
1.1
4***
73.3
27.5
1±
2.7
9**
73.3
30.1
7±
3.0
8**
Nucl
eus
26.6
28.7
6±
3.3
113.3
29.7
7±
2.1
3
PC Lo
wG
leas
on
(21)
Cyt
opla
sm52.4
22.8
9±
1.1
4***
71.4
22.3
6±
3.0
3***
85.7
127.8
3±
4.6
0***
80.9
28.0
1±
2.2
5**
100
23.8
±2.9
3*
Nucl
eus
30.1
32.0
8±
2.9
619
28.3
4±
2.0
4
Med
ium
Gle
ason
(47)
Cyt
opla
sm72.3
417.4
3±
2.3
4**
78.7
221.1
8±
2.1
0***
72.3
426.5
2±
3.6
8***
95.7
429.1
7±
1.2
2**
93.6
129.0
9±
1.9
0**
Nucl
eus
40.4
233.8
2±
4.0
744.6
834.9
1±
4.3
2
Hig
hG
leas
on
(25)
Cyt
opla
sm68
16.7
4±
2.5
4**
92
23.7
7±
2.3
6***
76
26.2
1±
3.8
6***
100
39.3
7±
4.1
5***
80
36.7
1±
2.1
8***
Nucl
eus
48
41.2
6±
2.3
1***
72
46.5
6±
1.2
0***
Ave
rage
optica
lden
sities
wer
eev
aluat
edonly
inpat
ients
show
ing
imm
unoposi
tivi
ty.
Stat
istica
lan
alys
isre
fers
toea
chan
tibody
separ
atel
y.In
each
sam
ple
,cy
topla
smic
and
nucl
ear
imm
unore
activi
tyw
ere
separ
ated
.Si
gnifi
cance
was
det
erm
ined
(com
par
edw
ith
the
valu
efo
rnorm
alpro
stat
e)by
one-
way
AN
OV
Aat
P£
0.0
5,
by
multip
lepai
r-w
ise
com
par
isons.
*P
<0.0
5;
**P
<0.0
1;
***P
<0.0
01.
172 C Nunez et al.
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.
A B C
D E F
G H I
J K L
Figure 5. IjB-a immunoreactivity in the cytoplasm of epithelial cells of benign prostatic hyperplasia (BPH) (A) and prostatic carcinoma (PC)
(B,C); no differences were observed between low, medium (B) and high (C) Gleason grades. p-IjB showed cytoplasmic immunoreactivity in
epithelial cells of high-grade prostatic intraepithelial neoplasia (PIN) (D) and PC samples of low, medium (E) and high (F) Gleason grade. p50 was
scanty in the cytoplasm of epithelial cells of low-grade PIN (G) but in PC immunoreactivity was also nuclear, with expression increasing in
medium (H) and, subsequently, in high (I) Gleason grade. p65 only was localized in the cytoplasm of epithelial cells in BPH (J) and PC (K,L)
samples; but in PC was also localized in the nuclei of epithelial cells, with increasing nuclear localization with Gleason grade: lower intensity in
medium (K) than in high (L) Gleason grade. Scale bars: 15 lm (I–K), 20 lm (B–H,L), 25 lm (D,F) and 30 lm (A–G).
NIK transduction pathway in prostatic cancer 173
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.

PC.25 The present study provides information about therole of TNF-a and IL-1 in the proliferation ⁄ apoptosisequilibrium in the human prostate.
In normal prostate, immunoexpression of TRAF-2,IRAK, TRAF-6, NIK, Ijja ⁄ b, p-IjB and NF-jB ⁄ p50was present in the cytoplasm of epithelial cells. NIKseems to be triggered by TNF ⁄ TRAF-2 or IL-1 ⁄ IRAK ⁄TRAF-6, because in addition to the presence of TNF,TNFRI and TRAF-2,23,26 immunoexpression of IL-1family members has been demonstrated.25 Activationof NF-jB requires the successive action of NIK, IKKcomplex and IjB. The transduction pathway from NIKto NF-jB seems to be inactive because immunoreac-tivity to IKK, IjB and NF-jB ⁄ p50 was scanty and noimmunoreactivity to NF-jB ⁄ p65 was found. Shuklaet al.30 have described weak immunoreactivity to IjBaand NF-jB ⁄ p65 in BPH. The scanty expression of NF-jB ⁄ p50 was localized in the cytoplasm of epithelialcells, because the low expression of p-IjB is unable toactivate NF-jB ⁄ p50. Since in low-grade PIN weobtained similar results to those described in normalprostate samples, we think that this transductionpathway could also be inactivated.
In BPH, immunoreactivity to IRAK, TRAF-6 and NIKwas similar to that described in low-grade PIN andnormal prostate, but decreased the expression of TRAF-2. This agrees with the high levels of IL-1a and IL-1RIreported in the same patients used in this study.25 Atthe same time, immunoexpression of Ijj, IjB (phos-phorylated and non-phosphorylated) and NF-jB (p50and p65) was increased. Both NF-jBs were localized inthe cytoplasm of epithelial cells, perhaps because theincrease in p-IjB immunoexpression is insufficientto activate the degradation to IjB-a required forthe translocation of NF-jB to the nucleus. Severalauthors31,32 have reported that inflammation might bea possible condition of hyperplastic proliferation ofprostatic tissue with risk of BPH progression. In theseconditions, a tissue microenvironment rich in cyto-kines, such as TNF-a and IL-1, would occur.33 Usingthe same samples as in this study, increased levels ofTNFa ⁄ Ap-1 transduction pathway components havealso been reported,23 and it has been suggested thatthis apoptotic pathway is activated in an attempt toinhibit uncontrolled cell proliferation (typical of BPHdiseases), although this attempt is insufficient and iscounteracted by other anti-apoptotic signals such asBcl-2,34 p2135 or transcription factors such as Elk-1and p-ATF-2. For this reason, Ricote et al.36 havesuggested that the overexpression of these transcriptionfactors is involved in cell proliferation and survival.
In PC, conversely to what occurs in BPH, the pro-apoptotic effect of TNF-a ⁄ AP-1 pathway decreases,
diverging towards the p38 pathway.23 However,TRAF-2 might be involved in the NIK activationpathway, although immunoexpression of TRAF-2 wasdetected in a low number of cases (decrease withGleason grade). All PC patients who were negative toTRAF-2 were positive for NF-jB ⁄ p50, and only onepatient (high Gleason group) who was negative forTRAF-2 was also negative for NF-jB ⁄ p65. Only fourPC patients were negative for TRAF-2 and NIK at thesame time. The percentages of PC and PIN (high-grade)patients who were positive for NIK were similar tothose found in the BPH group; but optical density washigher in PC and PIN (high-grade) groups. These data,in addition to the elevated immunoexpression of IL-1,IRAK and TRAF-6 (no variations between the Gleasongrade groups), suggest that in PC NIK is stimulatedby IL-1.
The percentages of samples positive for IKK, IjB-aand p-IjB were similar in PC with low Gleason, BPHand high-grade PIN groups. On the other hand, theoptical density was higher in PC and high-grade PIN,but no differences were found between these groups.The increase in the expression of IjB-a and p-IjBin PC (low Gleason) suggests that phosphorylation top-IjB and the degradation to IjBa increases with thesubsequent nuclear translocation of NF-jB. Thesefindings agree with other results supporting thenotion that NF-jB (p50 and p65) changes its locationfrom the cytoplasm to the nucleus in PC patients,although Ross et al.18 in PC have described a decreasein IjBa expression correlated with increase in bothnuclear NF-jB expression and tumour grade. Shuklaet al.30 have described in human prostate, a progres-sive increase in the expression of NF-jB ⁄ p65 (but notof NF-jB ⁄ p50) in PC compared with benign samplesand this increase correlated with increasing levels ofIjBa and its phosphorylation. When NF-jB is locatedin the nucleus (the active form) it may promote cellgrowth and proliferation in prostatic cancer cells byregulating the expression of c-myc, cyclin D1 orIL-6;37,38 but also by up-regulation of the expressionof several anti-apoptotic proteins, including Bcl-xL,39
the inhibitor of apoptosis protein40 or TRAF-1and 2.41 When we analysed our data obtained inhigh-grade PIN, it was observed that these data aresimilar to those obtained in PC with Gleason low-grade. In this respect, we agree with other authors42
who have described PIN as the most likely precursorof prostatic adenocarcinoma.
In PC with medium or high Gleason grades, thepercentage of cases positive for IKK was higher thanthat found in PC (low Gleason) and PIN (high-grade)groups, but the optical density was lower. In turn, the
174 C Nunez et al.
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.

percentage of cases positive for IjB was higher in PCwith medium or high Gleason grades than in the lowGleason group, whereas the percentage of casespositive for p-IjB decreased, without differences inoptical density between the two Gleason groups. Thepercentage of cases positive for NF-jB (p50 and p65)with nuclear location increased with Gleason grade;the highest optical density appeared in the highGleason grade and did not differ between mediumand low Gleason grades. Nuclear location of NF-jB ⁄p65 has been described by several authors,18–20,22
although their results are discrepant. WhereasDomingo-Domenech et al.19 have indicated thatnuclear localization of NF-jB is an independent prog-nostic factor in PC, other authors18,20 have concludedthat NF-jB is a new predictive marker of prostaticcancer and several have proposed the use of NF-jBinhibitors as therapeutic agents, either alone or com-bined with other agents. In this regard, Andreakoset al.43 consider NIK as a potent adjuvant strategy thatoffers great potential for genetic vaccine developmentin mouse myeloid cells. NF-jB is involved in cell pro-liferation triggered by IL-1 ⁄ NIK, but also by TNF ⁄ NIK.
TNF ⁄ TRAF-2 pathway may diverge to NIK but alsotowards p38, which activates transcription factorssuch as NF-jB, ATF-2 and Elk-1 (related to enhancedcell proliferation and survival). p38 activation is alsostimulated by IL-1. There is accumulating evidencethat strongly suggests that chronic inflammation is akey factor in tumorigenesis,33 and that the inflamma-tory process includes expression of cytokines such asTNF-a or IL-1,44 required for the activation of tran-scription factors related to survival and cell prolifera-tion (NF-jB, ATF-2 and Elk-1); it can be deduced thatinflammation might be implicated in the developmentof prostatic carcinogenesis.45
Translocation of NF-jB to the nucleus in PC mightbe due to overexpression of several components of theIL-1 ⁄ NIK ⁄ NF-jB or TNF ⁄ NF-jB (NIK or p38) path-ways. Since nuclear localization of NF-jB was observedonly in PC patients, it may be considered as a markerfor predicting PC. In order to search for a dominanttarget for therapy, it should be taken into account thatPC is a heterogeneous disease, in which multipletransduction pathways may interact in uncontrolledapoptosis ⁄ cell proliferation. Since IL-1a or TNF-a arethe most common cytokines related to inflammationand these cytokines are increased in PC,25,26 inhibitionof these cytokines might be a possible target for PCtreatment; such inhibition would affect all the trans-duction pathway members that activate several tran-scription factors related to survival and proliferationsuch as NF-jB, Elk-1 and ATF-2.
Acknowledgement
Supported by grants from the ‘Ministerio de Educaciony Ciencia’, Spain (SAF2007-61928).
References
1. Cerutti PA, Trump BF. Inflammation and oxidative stress in
carcinogenesis. Cancer Cells 1991; 3; 1–7.
2. Hussain SP, Hofseth LJ, Harris CC. Radical causes of cancer.
Nat. Rev. Cancer 2003; 3; 276–285.
3. Dinarello CA. The interleukin-1 family: 10 years of discovery.
FASEB J. 1994; 8; 1314–1325.
4. Sims JE, Dower SK. Interleukin-1 receptors. Eur. Cytokine Netw.
1994; 5; 539–546.
5. Boraschi D, Bossu P, Macchia G, Ruggiero P, Tagliabue A.
Structure–function relationship in the IL-1 family. Front. Biosci.
1996; 1; d270–d308.
6. Cao Z, Henzel WJ, Gao X. IRAK: a kinase associated with the
interleukin-1 receptor. Science 1996; 271; 1128–1131.
7. Li S, Strelow A, Fontana EJ, Wesche H. IRAK-4: a novel mem-
ber of the IRAK family with the properties of an IRAKkinase.
Proc. Natl Acad. Sci. U.S.A. 2002; 99; 5567–5572.
8. Rosati O, Martin MU. Identification and characterization of
murine IRAK-2. Biochem. Biophys. Res. Commun. 2002; 297; 52–
58.
9. Sun J, Wiklund F, Hsu FC et al. Interactions of sequence variants
in interleukin-1 receptor-associated kinase4 and the toll-like
receptor 6-1-10 gene cluster increase prostate cancer risk. Cancer
Epidemiol. Biomarkers Prev. 2006; 15; 480–485.
10. DiDonato JA, Hayakawa M, Rothwarf DM, Zandi E, Karin M. A
cytokine-responsive IkappaB kinase that activates the transcrip-
tion factor NF-kappaB. Nature 1997; 388; 548–554.
11. Wu JT, Kral JG. The NF-kappaB ⁄ IkappaB signaling system: a
molecular target in breast cancer therapy. J. Surg. Res. 2005; 23;
158–169.
12. Gasparian AV, Yao YJ, Kowalczyk D et al. The role of IKK in
constitutive activation of NF-jB transcription factor in prostate
carcinoma cells. J. Cell Sci. 2002; 115; 141–151.
13. Sovak MA, Bellas RE, Kim DW et al. Aberrant nuclear factor-
kappaB ⁄ Rel expression and the pathogenesis of breast cancer.
J. Clin. Invest. 1997; 100; 2952–2960.
14. Devalaraja MN, Wang DZ, Ballard DW, Richmond A. Elevated
constitutive IiB kinase activity and IiB-a phosphorylation in
Hs294T melanoma cells lead to increased basal MGSA ⁄ GRO-a
transcription. Cancer Res. 1999; 59; 1372–1377.
15. Dejardin E, Deregowski V, Chapelier M et al. Regulation of NF-iB
activity by Ikappa-B related proteins in adenocarcinoma cells.
Oncogene 1999; 18; 2567–2577.
16. Wang W, Abbruzzese JL, Evans DB, Larry L, Cleary KR, Chiao PJ.
The nuclear factor-IB transcription factor is constitutively active
in human pancreatic adenocarcinoma cells. Clin. Cancer Res.
1999; 5; 119–127.
17. Visconti R, Cerutti J, Battista S et al. Expression of the
neoplastic phenotype by human thyroid carcinoma cell lines
requires NF-iB p65 protein expression. Oncogene 1997; 15;
1987–1994.
18. Ross JS, Kallakury BV, Sheehan CE et al. Expression of nuclear
factor-kappa B and I kappa B alpha proteins in prostatic
adenocarcinomas: correlation of nuclear factor-kappa B immuno-
reactivity with disease recurrence. Clin. Cancer Res. 2004; 10;
2466–2472.
NIK transduction pathway in prostatic cancer 175
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.

19. Domingo-Domenech J, Mellado B, Ferrer B et al. Activation of
nuclear factor-kappaB in human prostate carcinogenesis and asso-
ciation to biochemical relapse. Br. J. Cancer 2005; 93; 1285–1294.
20. Lessard L, Karakiewicz PI, Bellon-Gagnon P et al. Nuclear
localization of nuclear factor-kappaB p65 in primary prostate
tumors is highly predictive of pelvic lymph node metastases.
Clin. Cancer Res. 2006; 12; 5741–5745.
21. Ghosh S, May MJ, Kopp EB. NF-kB and Rel proteins: evolution-
arily conserved mediators of immune responses. Annu. Rev.
Immunol. 1998; 16; 225–260.
22. Suh J, Payvandi F, Edelstein LC et al. Mechanisms of constitutive
NF-kappaB activation in human prostate cancer cells. Prostate
2002; 52; 183–200.
23. Ricote M, Royuela M, Garcia-Tunon I, Paniagua R, Fraile B.
Pro-apoptotic tumor necrosis factor-alpha transduction pathway
in normal prostate, benign prostatic hyperplasia and prostatic
carcinoma. J. Urol. 2003; 170; 787–790.
24. Ozes ON, Mayo LD, Gustin JA, Pfeffer SR, Pfeffer LM, Donner DB.
NF-kappaB activation by tumour necrosis factor requires the Akt
serine-threonine kinase. Nature 1999; 401; 82–85.
25. Ricote M, Garcia-Tunon I, Bethencourt FR, Fraile B, Paniagua R,
Royuela M. Interleukin-1 (IL-1alpha and IL-1beta) and its
receptors (IL-1RI, IL-1RII, and IL-1Ra) in prostate carcinoma.
Cancer 2004; 100; 1388–1396.
26. de Miguel MP, Royuela M, Bethencourt FR, Santamaria L, Fraile
B, Paniagua R. Immunoexpression of tumour necrosis factor-aand its receptors 1 and 2 correlates with proliferation ⁄ apoptosis
equilibrium in normal, hyperplasic and prostatic carcinomatous
human prostate. Cytokine 2000; 12; 535–538.
27. Raingeaud J, Gupta S, Rogers JS et al. Proinflammatory cytokines
and environmental stress cause p38 mitogen activated protein
kinase activation by dual phosphorylation on tyrosine and
threonine. J. Biol. Chem. 1995; 270; 7420–7426.
28. Sciarra A, Mariotti G, Salciccia S et al. Prostate growth and
inflammation. J. Steroid Biochem. Mol. Biol. 2008; 108; 254–260.
29. Sakr WA, Haas GP, Cassin BF, Pontes JE, Crissman JD. The
frequency of carcinoma and intraepithelial neoplasia of the
prostate in young male patients. J. Urol. 1993; 150; 379–385.
30. Shukla S, MacLennan GT, Fu P et al. Nuclear factor-kappaB ⁄ p65
(Rel A) is constitutively activated in human prostate adenocar-
cinoma and correlates with disease progression. Neoplasia 2004;
6; 390–400.
31. Crawford ED, DeAntoni EP, Ross CA. The role of prostate-specific
antigen in the chemoprevention of prostate cancer. J. Cell.
Biochem. 1996; 25; 149–155.
32. Roehrborn CG. Definition of at-risk patients: baseline variables.
BJU Int. 2006; 97; 7–11.
33. Mantovani A, Locati M, Vecchi A, Sozzani S, Allavena P. Decoy
receptors: a strategy to regulate inflammatory cytokines and
chemokines. Trends Immunol. 2001; 22; 328–336.
34. Huang SP, Wu WJ, Chang WS et al. p53 Codon 72 and p21
codon 31 polymorphisms in prostate cancer. Cancer Epidemiol.
Biomarkers Prev. 2004; 13; 2217–2224.
35. Aaltomaa S, Lipponen P, Eskelinen M, Ala-Opas M, Kosma VM.
Prognostic value and expression of p21 (waf1 ⁄ cip1) protein in
prostate cancer. Prostate 1999; 39; 8–15.
36. Ricote M, Garcia-Tunon I, Bethencourt F et al. The p38
transduction pathway in prostatic neoplasia. J. Pathol. 2006;
208; 401–407.
37. Joyce D, Albanese C, Steer J, Fu M, Bouzahzah B, Pestell RG. NF-
kappaB and cell-cycle regulation: the cyclin connection. Cytokine
Growth Factor Rev. 2001; 12; 73–90.
38. Suh J, Rabson AB. NF-kappaB activation in human prostate
cancer: important mediator or epiphenomenon? J. Cell. Biochem.
2004; 91; 100–117.
39. Chen CD, Sawyers CL. NF-kappa B activates prostate-specific
antigen expression and is upregulated in androgen-indepen-
dent prostate cancer. Mol. Cell. Biol. 2002; 22; 2862–
2870.
40. Stehlik C, de Martin R, Binder BR, Lipp J. Cytokine induced
expression of porcine inhibitor of apoptosis protein (iap) family
member is regulated by NF-kappa B. Biochem. Biophys. Res.
Commun. 1998; 243; 827–832.
41. Wang CY, Mayo MW, Korneluk RG, Goeddel DV, Baldwin AS Jr.
NF-kappaB antiapoptosis: induction of TRAF1 and TRAF2 and
c-IAP1 and c-IAP2 to suppress caspase-8 activation. Science
1998; 281; 1680–1683.
42. Graham SD Jr, Bostwick DG, Hoisaeter A et al. Report of the
committee on staging and pathology. Cancer 1992; 70; 359–
361.
43. Andreakos E, Williams RO, Wales J, Foxwell BM, Feldmann M.
Activation of NF-kappaB by the intracellular expression of NF-
kappaB-inducing kinase acts as a powerful vaccine adjuvant.
Proc. Natl Acad. Sci. U.S.A. 2006; 103; 14459–14464.
44. Sica A, Rubino L, Mancino A et al. Targeting tumour-
associated macrophages. Expert Opin. Ther. Targets 2007; 11;
1219–1229.
45. Palapattu GS, Sutcliffe S, Bastian PJ et al. Prostate carcinogenesis
and inflammation: emerging insights. Carcinogenesis 2004; 26;
1170–1181.
176 C Nunez et al.
� 2008 The Authors. Journal compilation � 2008 Blackwell Publishing Ltd, Histopathology, 53, 166–176.




















