
Theoretical studies on models of lysine-arginine cross-links derived from α-oxoaldehydes: a new...
15
ORIGINAL PAPER Theoretical studies on models of lysine-arginine cross-links derived from α-oxoaldehydes: a new mechanism for glucosepane formation Rasoul Nasiri & Mansour Zahedi & Hélène Jamet & Ali Akbar Moosavi-Movahedi Received: 28 April 2011 /Accepted: 21 June 2011 /Published online: 3 August 2011 # Springer-Verlag 2011 Abstract Availability and high reactivity of α-oxoaldehydes have been approved by experimental techniques not only in vivo systems but also in foodstuffs. In this article we re- examine the mechanism of glucosepane formation by using computational model chemistry. Density functional theory has been applied to propose a new mechanism for glucosepane formation through reaction of α-oxoaldehydes with methyl amine (MA) and methyl guanidine (MGU) models of lysine and arginine residues respectively. This non enzymatic process can be described in three main steps: (1) Schiff base formation from methyl amine, methyl glyoxal (MGO) (2) addition of methyl guanidine and (3) addition of glyceraldehyde. We show that this process is thermodynamically possible and presents a rate-determining step with a reasonable free energy barrier equal to 37.8 kcal mol -1 in water solvent. Comparisons were done with the mechanism formation of GODIC (glyoxal-derived imidazolium cross-link) and MODIC (methyl glyoxal-derived imidazolium cross-link), two other important cross-links in vivo. Keywords α-oxoaldehydes . Cross-link . Density functional theory . Glucosepane . GODIC . MODIC . Physiological glucose levels Introduction The chemical modification and cross-linking of proteins by the Maillard reaction is the results of a cascade of reactions. Condensations, rearrangements, fragmentation and oxida- tion processes of reducing carbohydrates (glucose, ribose, fructose) or α-oxoaldehydes (methyl glyoxal (MGO), glyoxal (GO), 3-deoxyglucosone (3-DG) and glyceralde- hyde (GLA)) occur with amino groups in lysine residues and guanidine groups in arginine residues [5, 7, 9, 25, 29, 33, 34, 46]. The mechanism of the glycation reaction was firstly tested by Maillard at the beginning of the twentieth century by incubation of amino acids with different sugars [37] and can be described at three stages. The early stage of reaction involves initiation of glycation through amino- carbonyl interaction leading to early glycation products, which contain an intact sugar moiety. In the intermediate stage, a variety of carbonyl compounds, which are generally more reactive than the original amino acids, are formed. Finally, successive rearrangements lead irreversibly to the formation of a variety of heterogeneous structures, known as the Advanced Glycation End-products (AGEs). AGEs accumulate in body and have been implicated in the development of many pathophysiologies associated with ageing in general as well as with complications of diabetes, atherosclerosis and renal insufficiency [8, 56]. Electronic supplementary material The online version of this article (doi:10.1007/s00894-011-1161-x) contains supplementary material, which is available to authorized users. R. Nasiri : M. Zahedi (*) Department of Chemistry, Faculty of Sciences, Shahid Beheshti University, G.C., Evin, 19839-63113, Tehran, Iran e-mail: [email protected] e-mail: [email protected] R. Nasiri : H. Jamet Département de Chimie Moléculaire, Chimie Théorique, UMR-5250, Université Joseph Fourier, BP 53, 38041, Grenoble Codex 9, France A. A. Moosavi-Movahedi Institute of Biochemistry and Biophysics, University of Tehran, Tehran, Iran J Mol Model (2012) 18:1645–1659 DOI 10.1007/s00894-011-1161-x
Transcript of Theoretical studies on models of lysine-arginine cross-links derived from α-oxoaldehydes: a new...

ORIGINAL PAPER
Theoretical studies on models of lysine-arginine cross-linksderived from α-oxoaldehydes: a new mechanismfor glucosepane formation
Rasoul Nasiri & Mansour Zahedi & Hélène Jamet &Ali Akbar Moosavi-Movahedi
Received: 28 April 2011 /Accepted: 21 June 2011 /Published online: 3 August 2011# Springer-Verlag 2011
Abstract Availability and high reactivity of α-oxoaldehydeshave been approved by experimental techniques not only invivo systems but also in foodstuffs. In this article we re-examine the mechanism of glucosepane formation by usingcomputational model chemistry. Density functional theory hasbeen applied to propose a new mechanism for glucosepaneformation through reaction of α-oxoaldehydes with methylamine (MA) and methyl guanidine (MGU) models of lysineand arginine residues respectively. This non enzymatic processcan be described in three main steps: (1) Schiff base formationfrom methyl amine, methyl glyoxal (MGO) (2) addition ofmethyl guanidine and (3) addition of glyceraldehyde. Weshow that this process is thermodynamically possible andpresents a rate-determining step with a reasonable free energybarrier equal to 37.8 kcal mol-1 in water solvent. Comparisonswere done with the mechanism formation of GODIC(glyoxal-derived imidazolium cross-link) and MODIC
(methyl glyoxal-derived imidazolium cross-link), two otherimportant cross-links in vivo.
Keywords α-oxoaldehydes . Cross-link . Densityfunctional theory . Glucosepane . GODIC .MODIC .
Physiological glucose levels
Introduction
The chemical modification and cross-linking of proteins bythe Maillard reaction is the results of a cascade of reactions.Condensations, rearrangements, fragmentation and oxida-tion processes of reducing carbohydrates (glucose, ribose,fructose) or α-oxoaldehydes (methyl glyoxal (MGO),glyoxal (GO), 3-deoxyglucosone (3-DG) and glyceralde-hyde (GLA)) occur with amino groups in lysine residuesand guanidine groups in arginine residues [5, 7, 9, 25, 29,33, 34, 46]. The mechanism of the glycation reaction wasfirstly tested by Maillard at the beginning of the twentiethcentury by incubation of amino acids with different sugars[37] and can be described at three stages. The early stage ofreaction involves initiation of glycation through amino-carbonyl interaction leading to early glycation products,which contain an intact sugar moiety. In the intermediatestage, a variety of carbonyl compounds, which aregenerally more reactive than the original amino acids, areformed. Finally, successive rearrangements lead irreversiblyto the formation of a variety of heterogeneous structures,known as the Advanced Glycation End-products (AGEs).
AGEs accumulate in body and have been implicated inthe development of many pathophysiologies associatedwith ageing in general as well as with complications ofdiabetes, atherosclerosis and renal insufficiency [8, 56].
Electronic supplementary material The online version of this article(doi:10.1007/s00894-011-1161-x) contains supplementary material,which is available to authorized users.
R. Nasiri :M. Zahedi (*)Department of Chemistry, Faculty of Sciences,Shahid Beheshti University,G.C., Evin,19839-63113, Tehran, Irane-mail: [email protected]: [email protected]
R. Nasiri :H. JametDépartement de Chimie Moléculaire, Chimie Théorique,UMR-5250, Université Joseph Fourier,BP 53, 38041, Grenoble Codex 9, France
A. A. Moosavi-MovahediInstitute of Biochemistry and Biophysics, University of Tehran,Tehran, Iran
J Mol Model (2012) 18:1645–1659DOI 10.1007/s00894-011-1161-x

They are implicated in pathogenesis of various devastatingdisorders, such as coronary heart diseases [26], cancergrowth [11] and metastasis, insulin resistance [56] and nonalcoholic fatty liver disease [36]. They have also beenshown to have a role in neuro degenerative disease such asAlzheimer's and Parkinson's diseases [22, 48].
According to the type of carbonyl compounds that canbe a reducing agent in glycation reaction, six distinctclasses of AGEs have been found in serum. They areglucose-derived AGEs (GLC-AGEs), MGO-derived AGEs(MGO-AGEs), GO-derived AGEs (GO-AGEs), 3-DG-derived AGEs (3-DG-AGEs), glyceraldehyde-derivedAGEs (Glycer-AGEs) and glycolaldehyde-derived AGEs(Glycol-AGEs) [42, 49]. On the other hand, Ahmed et al.classified AGEs to the first four in aforementioned ones [1,2]. Interestingly, clinical studies have shown that Glycer-AGEs are more important than GLC-AGEs in cardiovas-cular disease (CVD) and nonalcoholic steatohepatitis(NASH) in diabetes [31, 32].
An extense variety of AGEs has been discovered in vitroand in vivo systems; for example, glucosepane, DODIC,GOLD, MOLD, MODIC and GODIC, etc. These cross-links have also been identified in foodstuffs [4]. It is thecase of the glucosepane, recognized as the major cross-linkin vivo under non oxidative conditions [6, 46]. To explainits formation, Biemel et al. proposed a mechanism withlysine linked α-dicarbonyl compound as intermediate [7].This mechanism involves formation of a dideoxyosonefrom glucose and lysine; dideoxyosone then undergoescyclization into aldimine, which then reacts with the sidechain of arginine to yield glucosepane (see Scheme 1).
Experimental studies have previously suggested thatAGEs can arise not only from reducing sugars, but alsofrom α-oxoaldehyde compounds obtained from the degra-dation of sugars [53–55, 58], lipids [3], Schiff baseintermediate or Amadori product [38] in buffered solutionat 37 °C. Decomposition of amadori products has beenillustrated in Scheme 2. The α-oxoaldehyde compoundsfound in vivo may have a dietary origin [27, 35]. In theglycation of human serum albumin (HSA) under physico-logical conditions, separation of glucose from HSA [43]1
and its degradation to the α–oxoaldehydes (such as MGO,GO, 3-DG and GLA) [51], imply another possibility forglucosepane formation. It has been found that there arethree pathways for the in vivo formation of glyceralde-
hydes: namely, glycolytic pathway, polyol pathway, andfructose metabolic pathway [42] and on the other hand, itplays a key role in Maillard cross linking of food proteins[17–19, 57].
According to these results which show availability of theα–oxoaldehydes, a new mechanism for the formation ofglucosepane is re-examined by using computational modelchemistry. This article is devoted to describe a novelmechanism for glucosepane in vivo and food systems.Computational model chemistry will be used to understandand establish proper reaction mechanism. It can givemolecular level insights into a reaction through determina-tion of intermediates and transition states structures andenergies. The mechanism considers reactions between theα-oxoaldehydes MGO and GLA with lysine and arginineamino acids.
We have investigated mechanisms of formation of thecross-links with three main aims: first, to provide insightinto the formation mechanism of glucosepane consistentwith experimental results found from glycation of humanserum albumin under physiological conditions (Scheme 3);second, to study and compare the reaction mechanisms ofthe glucosepane, GODIC, and MODIC; third, to analyzethe changes in the thermodynamic and kinetic propertiescaused by the solvent.
Computational details
The geometries of reactants, products, intermediates, andtransition states involved in the proposed mechanismhave been fully optimized with using the wB97XDfunctional [13]. This functional includes empirical disper-sion and can better treat hydrogen bonding and van derWaals interactions than conventional DFT. The 6-31+G*basis set with both polarized and diffuse function is aneffective choice for more accurate description of hydrogentransfer reactions and hydrogen-bonded species of thisstudy.
For validation of method and basis set, all optimizedstructures in the first step of glucosepane formation processfrom wB97XD level were reoptimized with frozen coremoller-plesset perturbation theory, MP2(FC)/6-31+G*,computational level. Moreover, CCSD(T)/6-31+G* singlepoint calculations have been carried out for the samecompounds to evaluate the reliability of barrier calculated atwB97XD and B3LYP/6-31+G* level. Extension of thebasis set to 6-311++G (3df, 3p) in DFT calculations werealso examined to compare to the same calculations with6-31+G* basis set (see Table 1). This strategy showed thatfirstly, extension of basis set did not have significant effectson free energy diagram while obtained results fromwB97XD/6-31+G* are more reliable than B3LYP/6-31+G
1 In this stage of the reaction, there will be a competition betweenglucose and α-oxoaldehydes in the nucleophilic attack on aminogroups in the proteins. Glucose, because of high stability in its cyclicform, will be less subject to the nucleophilic attacks compared to theα-oxoaldehydes and since α-oxoaldehydes are up to 20,000-fold morereactive than glucose [50]. Therefore, glucosepane can likely bederived through reaction of the α-oxoaldehydes with lysine andarginine amino acids
1646 J Mol Model (2012) 18:1645–1659

ones since former considers non-bonded interactions alongwith long rang correlations better than latter one (comparerelevant CCSD(T) values in Table 1).
The intrinsic reaction calculations (IRC) [20, 21] in thegas phase confirmed that all TSs were in the right reactionpathways unambiguously.
To evaluate the effects of water as a high-permittivitysolvent, on the cross-linking mechanisms we employedpolarizable continuum model (PCM) calculations [52]based on solvent reaction field method (SCRF) to furtheroptimize the geometries of the stationary points in water.Vibrational analyses were performed on all optimizedstructures with the same functional and basis set. The
zero-point energy (ZPE) corrections were also included inthe reported free energies with a scale factor of 0.89 attemperature 298.15 K and 1 atm pressure in water. Allcalculations were performed using the Gaussian 09 soft-ware [16] and the Gauss View program [15] by employingthe software and hardware facilities of the theoretical groupof the DCM, University of Grenoble I Codex, France.
Results and discussion
Density functional theory (DFT) calculations were per-formed on methyl amine (MA) and methyl guanidine
C
C OH
C
C
C
C
OHH
HHO
OHH
OHH
OHH
H
C
C NHH
C
C
C
C
OHH
HHO
OHH
OHH
OHH
H
R1
OH
C
C NH
C
C
C
C
OHH
HHO
OHH
OHH
OHH
H
R1
-H2O
C
CHNH
C
C
C
C
OH
HHO
OHH
OHH
OHH
H
R1
enolization+R1-NH2
CH
HNH
C
C
C
C
HHO
OHH
OHH
OHH
H
R1
C O
a
CH
HNH
C
C
C
C
HHO
OHH
OHH
OHH
H
R1
C O
CH
HNH
C
C
C
C
HO
OHH
OHH
H
R1CH
HN
H
C
C
C
C
HO
OH
OHH
H
R1
C OH
CH
HNH
C
C
C
C
HO
H
OHH
OH
R1
HC OH
-H2O
C O
H H
1,5 H+ shift
tautomerization
CH
HNH
C
C
C
C
HO
OH
OH
R1
HC OH
H
H
keto-enol tautomerization
1,3 H+ shift
tautomerization
CH
HNH
CH
C
C
C
HO
H
OH
OH
R1
HC OH
CH
HNH
CH
CH
C
C
HO
H
O
OH
R1
HC OHketo-enol
tautomerization
enolization
b
CH
HNH
CH
CH
C
C
HO
H
O
OH
R1
HC OH
cyclization
+H+
-H2O
R2N C
NH2
NH -H+
lysine linked alpha-dicarbonyl (dideoxyosone)
R1 = CH2CH2CH2CH2CHNH2COOH
R2 = CH2CH2CH2CHNH2COOH
NH+
R1
HO
OH
O
HON+
R1
HO
OHO
N
R1
OH
OH
H N
N
H
N
R2
H
N
R1
OH
OH
N
NN
R2
H
H
c
aldimine
Scheme 1 Scheme of proposedmechanism for glucosepaneby Biemel, (a) Scheme ofamadori formation step, (b)Scheme of lysine linked alpha-dicarbonyl formation step, (c)Scheme of addition of argininestep (R1 and R2 stand forCH2CH2CH2CH2CHNH2COOH and CH2CH2CH2CHNH2COOH, respectively)
J Mol Model (2012) 18:1645–1659 1647

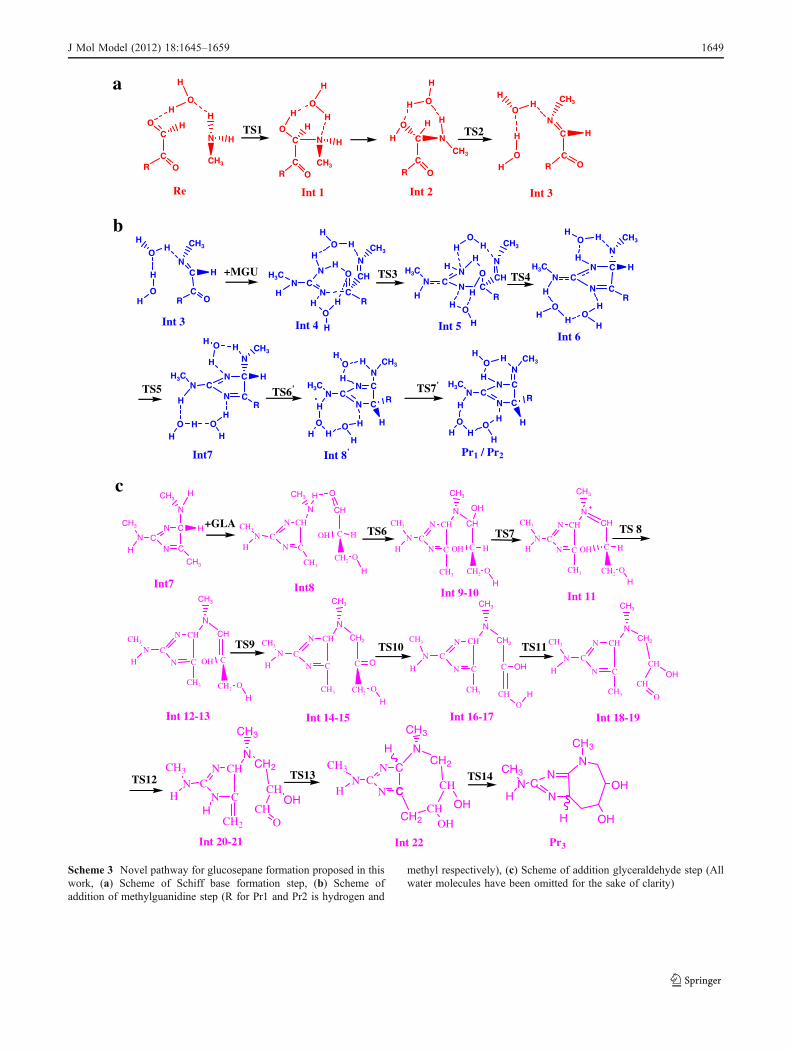
(MGU) which are models for the lysine and arginine aminoacids with α-oxoaldehydes of MGO, GO and GLA todescribe the mechanism of the formation of glucosepane,MODIC and GODIC. Water molecules are added in themechanism, not to simulate the solvent but as reactivespecies. The proposed mechanism (Scheme 3) involvesthree main steps, each one of them involves severalintermediates and transition state structures: (1) Schiff baseformation (Scheme 3a), (2) addition of methyl guanidine(Scheme 3b) (only structures of Int3 to Int7) and (3)addition of glyceraldehyde (Scheme 3c). Because ofsimilarity in new mechanism of Glucosepane with eachone of MODIC and GODIC, we have put their mechanismsin the same schemes (Schemes 3a and b) but note that theyare different-on the two last TS in Schemes 3b.
This section is divided as follows: First, the results anddiscussion of different steps of the glucosepane formationare presented (section A). Then formation mechanism ofGODIC and MODIC is compared with glucosepane insection B. Finally, we discuss the solvent effects overthe reaction mechanisms (section C).
A. Formation mechanism of glucosepane
Schiff base formation
A molecular complex consisting of MGO, MA and onewater molecule was chosen as model compound to studythe initial step of the cross linking process. The free energyprofile in gas phase and solution for the process is depictedin Fig. 1. The electronic energy of the reactant complex isstabilized relative to each of the isolated species while itsfree energy is destabilized. The difference of values of theelectronic and free energies are −11.9 and +9.5 kcal mol-1
respectively. Hydrogen bond interactions between themolecules explain the stabilization of the electronic energyand the destabilization of the free energy reflects thenegative entropic contribution associated to the transforma-tion of three molecules to one specie. Because of instabilityof pre reactive complex, we do not include it in thediscussion.
The process starts with carbinolamine formation bynucleophilic attack of the methyl amine molecule to the
CH
HNH
C
C
C
C
HHO
OHH
OHH
OHH
H
R1
C OCH
HNH
C
C
C
C
OH
OHH
OHH
OHH
H
R1
C OH
-R1-NH2
CH2
C
C
C
C
O
OHH
OHH
OHH
H
C OH
-H2Oenolization
CH3
C
C
C
C
O
OHH
OHH
OHH
H
C O
Amadori Product 1-DG
enolization
C
HNH
C
C
C
C
HHO
OHH
OHH
OHH
H
R1
C OH-R1-NH2
HC
CH2
C
C
C
OHH
OHH
OHH
H
C OCH3
HC
C
C
C
O
OH
OHH
OHH
H
C O
+H+
-H2O
C
HNH
C
C
C
C
H
OHH
OHH
OHH
H
R1
C OH
+H2O
O
retro-aldolMGO
GLA
3-DG
C
C OH
C
C
C
C
OHH
HHO
OHH
OHH
OHH
H
C
C NHH
C
C
C
C
OHH
HHO
OHH
OHH
OHH
H
R1
OH
C
C NH
C
C
C
C
OHH
HHO
OHH
OHH
OHH
H
R1
+H2O
C
CHNH
C
C
C
C
OH
HHO
OHH
OHH
OHH
H
R1
enolization
-R1-NH2Schiff base
Glucose
C
C HO
C
C
C
C
OHH
HHO
HHO
OHH
OHH
HMannose
Scheme 2 The pathways ofamadori products degradation
1648 J Mol Model (2012) 18:1645–1659
R
HC
O
CO
HO
H
H
N
CH3
HC
Int 1 Int 2 Int 3
O
N
CR
TS1 TS2
Re
R
H
CO
CO
HO
H
H
N
CH3
H
R
H
C
O
CO
H
O
H
H
NCH3
H
H
CH3HO
H
H
OH
a
RC
CH
NCH3
O
HO
H
HOH
Int 4 Int 5Int 6
NC
NN
HR
C
C
N
CH3
H
HOH
OH
HO
H
NC
NN
H
H
H
H
H3CN
CN
N
H RC
CH
N
CH3
H
HO
H
HO
H
H3CH3CO
H
H
H
TS3 TS4
NC
NN
H RC
C
NCH3
H
HOH
OH
H OH
H3C
H
H
HC
Int 3
O
N
CR
+MGUH
CH3HO
H
H
OH
Int7 Int 8'
NC
NN
HRC
C
NCH3
H
HOH
OH H
O H
H3C
HPr1 / Pr2
TS6' TS7'
H
NC
NN
HRC
C
NCH3
H
HOH
OH H
OH
H3C
H
H
TS5
NC
N
NH
CH3
C
CHCH3
CH2
COH H
O
Int 9-10 Int 11
+GLATS6
H
NC
NN
HCH3
C
C
N
CH3H
CH3 H
Int7 Int8
N
CH3
CH
OH
NC
N
NH
CH3
C
CHCH3
CH2
COH H
O
H
N
CH3
CH
OH
NC
N
NH
CH3
C
CHCH3
CH2
COH H
O
H
N
CH3
CHTS7
Int 12-13 Int 14-15
TS 8
TS9
N
C
N
N
H
CH3
C
CHCH3
CH2
COH
O
H
N
CH3
CH
TS10
N
C
N
N
H
CH3
C
CHCH3
CH2
C
O
H
N
CH3
CH2
O
b
Int 18-19
TS12 TS13
Int 16-17
N
C
N
N
H
CH3
C
CHCH3
CH
C
OH
N
CH3
CH2
OH
TS11
N
C
N
N
H
CH3
C
CHCH3
CH
CH
O
N
CH3
CH2
OH
Int 22 Pr3
NC
NN
H
CH2
C
CCH3
CH
CH
OH
N
CH3
CH2
OH
N
OH
OH
CH3
N
NCN
CH3
H
H
H
Int 20-21
NC
NN
H
CH2
C
CHCH3
CH
CH
O
N
CH3
CH2
OHH
TS14
c
Scheme 3 Novel pathway for glucosepane formation proposed in thiswork, (a) Scheme of Schiff base formation step, (b) Scheme ofaddition of methylguanidine step (R for Pr1 and Pr2 is hydrogen and
methyl respectively), (c) Scheme of addition glyceraldehyde step (Allwater molecules have been omitted for the sake of clarity)
J Mol Model (2012) 18:1645–1659 1649
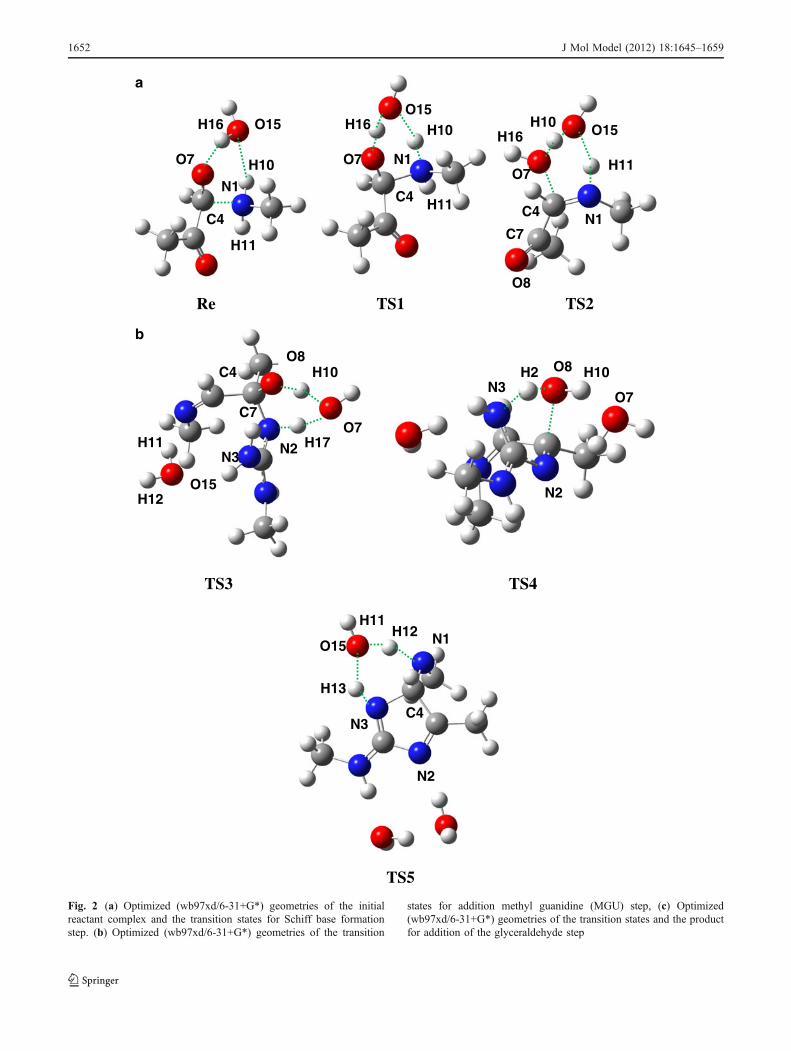
carbonyl carbon atom labeled as C4 in Fig. 2a. This attackis performed via a proton transfer from N1 to O15 with afree energy barrier equals to 15.7 kcal mol-1 (see Fig. 1). Inthis six-membered-ring TS called TS1 in Fig. 2a, the C4-N1
bond is almost formed (1.53 A) and the C4-O7 bond has alength close to that of a single bond (1.35 A). The Schiffbase formation is accomplished with the dehydration ofcarbinolamine to yield a ketoimine. As Int1 cannot bedehydrated directly, it isomerizes to Int2 through two singlebond rotations. The free energy barrier for this transforma-tion is less than 4 kcal without participation of watermolecule and can be done easily. Then Int2 evolves to theSchiff base as Hall and Smith reported this stage viareaction of MA and formaldehyde [24]. The reactionfollows through a concerted proton transfer in which H11
is transferred from N1 to O15 while H10 from O15 to O7 TS2in Fig. 2a illustrates these proton transfers. In this transitionstate, the double bond between N1 and C4 is formed.Finally, a ketoimine compound is created that has twoelectrophilic centers reacts with MGU. As can be seen fromfree energy profile (Fig. 1), the activation free energybarrier is high for the dehydration step and has a valueequal to 33.5 kcal mol-1 in the gas phase which is in goodagreement with high-level ab initio results (33.2 kcal mol-1)on similar reaction [24]. It is worth mentioning here thatthis barrier decreases significantly when more watermolecules are mediated around the reactive center ofreaction, but since water assisted effects are not the goalof this study we do not discuss it. In comparing to otherbarriers of the Schiff base formation (see Fig. 1) the last oneis the rate-limiting step of this reaction in agreement withexperimental kinetic studies [44, 45].
Addition of methyl guanidine (MGU)
In the second step, methyl guanidine was chosen asmolecular model of arginine residue and added to theSchiff base obtained from the first step. Because ofexistence five acidic protons on the guanidino group (seeScheme 4); there are five different neutral methylguanidine tautomers. The guanidino function is usually
assumed to be in the protonated form with an effectivepKa of about 12.0 in aqueous solution at 298 K [40]. Sothe first calculations were performed with this form.However our calculations show the protonated formmoving away from the Schiff base and we did not succeedto optimize an adduct from them. Hence, for this reasonone of the neutral forms was taken into account and couldexplain why rate of the glycation reaction increases withthe basicity [30]. Moreover, the pKa s values in biologicalsystems are not the same as in the solution. A previoustheoretical study shows that all the neutral forms of MGUare in equilibrium to each other [39]. Therefore, weconsidered only one of these tautomers2. Addition ofMGU to the Schiff base leads to Int4 in which guanidinogroup is perpendicular to the ketoimine plan. Two watermolecules assist this step, the water molecule was addedin the Schiff base formation step, and the water moleculeproduced from the last dehydration. Since the NH functionis more nucleophilic than the NH2 one, first N2 attack toC7 of the Schiff base (see Fig. 2b for the notation). Thefirst proton transfer in this step occurs through TS3 with afree energy barrier less than 8.4 kcal mol-1. In thistransition state, a twisted six-membered ring is formed inwhich the proton from NH is being transferred to the watermolecule which in turn has already started to transfer itsproton to O8. The completion of this reaction leads to acarbinolamine (Int5) whose free energy is stabilized about11.1 kcal mol-1 relative to Int4. This stability is due toformation of bond N2-C7 completely in Int5 while distanceof N2-C7 in Int4 is 1.54 Åand N2 has also positive charge. Anew step of dehydration occurs to give Int6. The calculatedfree energy barrier of this dehydration reaction which passesby TS4 (Fig. 1) is also high but 7.5 kcal mol-1 less than
Table 1 Changes of electronic energy for the first step of proposed mechanism in the gas phase (in kcal mol-1)
WB97XDa WB97XDb B3LYPc MP2d CCSD (T)e CCSD (T)f
Re——TS1 5.4 4.2 4.5 5.7 7.2 7.3
TS1——Int1 16.5 17.4 15.1 16.2 18.4 18.7
Int1——Int2 4.8 4.4 4.8 4.7 3.5 4.3
Int2——TS2 32.6 31.7 26.9 28.8 31.0 33.7
TS2———Int3 24.7 26.4 23.9 25.5 26.3 28.7
Superscripts a, b, c, d, e and f indicate calculations at the WB97XD/6-31+G*, WB97XD/6-311++G (3df, 3p), B3LYP/6-31+G*, MP2 (FC)/6-31+G*,CCSD (T)/6-31+G*//WB97XD/6-31+G* and CCSD (T)/6-31+G*//B3LYP/6-31+G* levels, respectively
2 Following the comment of a reviewer, we tested the influence of twodifferent isomers of MGU (see tautomers C and E in Scheme 4) on thereaction profile in second step of cross-linking process. We found thatactivation barriers are to be in different values when tautomers areinvolved in the process (see Figure S1). It is worth noting that isomerF cannot be involved in the process actively since Lederer and Klaiberdefinitely excluded formation of MODIC or GODIC via endocyclicnitrogens of arginine residues (see 34 for more detail)
1650 J Mol Model (2012) 18:1645–1659
similar one in the first step (TS2 in Fig. 1). The NH2 group ofthe MGU acts as a proton transferring carrier, therebylowering the barrier of this second dehydration reactionrelative to the first dehydration reaction. After downhill fromTS4, Int6 can produce a five-membered heterocyclic structurethrough TS5, which involves the concurrent transfer of twoprotons from N3 to N1. The free energy barrier for thisreaction is calculated to be 8.5 kcal mol-1 relative to Int6.From TS5 to Int7, distance of N3-C4 decreases from 1.51 to1.46 Å and the free energy decreases to 35.3 kcal mol-1 whichcould explain the irreversibility of the cross-linking process invivo (see Fig. 1).
Addition of glyceraldehyde (GLA)
Finally, we consider the formation of glucosepane by additionof glyceraldehyde to Int8 obtained from the previous step.Glyceraldehyde (GLA), the simplest reducing sugar, has beenrecognized as one of the most reactive species in the Maillardreaction in vivo and in vitro under physiological conditions[12]. Int8 is the reactant complex due to the addition of theGLA to Int7. N1 in Int8 must be dehydrogenated before itsaddition to the carbonyl function. It is done with the help ofthe water molecule. Thanks to hydrogen bonds with O19 andH11, it acts as a bridge to facilitate the proton transfer througha concerted TS (TS6 in Fig. 2c). This proton transfer is alsosubject to a relatively small barrier (13.5 kcal mol-1, Fig. 1).
In this step, because of the carbonyl mobility alongthe entire GLA backbone and the limited number ofwater molecules available, we had to change theposition of water molecules relative to reactive center
The tautomeric transformation of Int17 to Int18 issimilar to the tautomerization of Int13 to Int14 andtheir free energy barriers are equal to 18.7 and21.6 kcal mol-1 respectively. An enamine (Int19) is
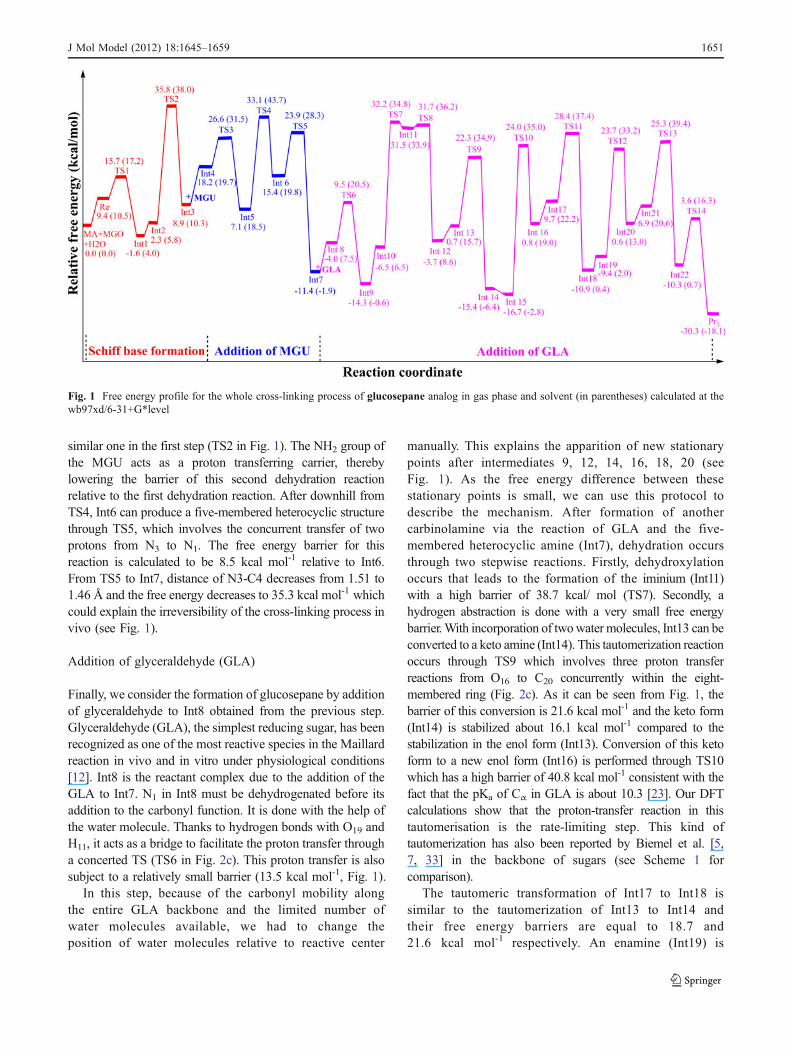
Fig. 1 Free energy profile for the whole cross-linking process of glucosepane analog in gas phase and solvent (in parentheses) calculated at thewb97xd/6-31+G*level
J Mol Model (2012) 18:1645–1659 1651
manually. This explains the apparition of new stationarypoints after intermediates 9, 12, 14, 16, 18, 20 (seeFig. 1). As the free energy difference between thesestationary points is small, we can use this protocol todescribe the mechanism. After formation of anothercarbinolamine via the reaction of GLA and the five-membered heterocyclic amine (Int7), dehydration occursthrough two stepwise reactions. Firstly, dehydroxylationoccurs that leads to the formation of the iminium (Int11)with a high barrier of 38.7 kcal/ mol (TS7). Secondly, ahydrogen abstraction is done with a very small free energybarrier.With incorporation of twowater molecules, Int13 can beconverted to a keto amine (Int14). This tautomerization reactionoccurs through TS9 which involves three proton transferreactions from O16 to C20 concurrently within the eight-membered ring (Fig. 2c). As it can be seen from Fig. 1, thebarrier of this conversion is 21.6 kcal mol-1 and the keto form(Int14) is stabilized about 16.1 kcal mol-1 compared to thestabilization in the enol form (Int13). Conversion of this ketoform to a new enol form (Int16) is performed through TS10which has a high barrier of 40.8 kcal mol-1 consistent with thefact that the pKa of Cα in GLA is about 10.3 [23]. Our DFTcalculations show that the proton-transfer reaction in thistautomerisation is the rate-limiting step. This kind oftautomerization has also been reported by Biemel et al. [5,7, 33] in the backbone of sugars (see Scheme 1 forcomparison).
Re TS1 TS2
TS3 TS4
TS5
C4
N1
H11
O7
H16 O15
H10
H11C4
N1
H10O15
H16
O7 H11
N1
O15H10H16
O7
C4
C7
O8
H11
H12O15
O7
H10O8
C7
C4
N2 H17N3
O7
O8 H10H2N3
N2
H13
O15
H11H12 N1
N3
N2
C4
a
b
Fig. 2 (a) Optimized (wb97xd/6-31+G*) geometries of the initialreactant complex and the transition states for Schiff base formationstep. (b) Optimized (wb97xd/6-31+G*) geometries of the transition
states for addition methyl guanidine (MGU) step, (c) Optimized(wb97xd/6-31+G*) geometries of the transition states and the productfor addition of the glyceraldehyde step
1652 J Mol Model (2012) 18:1645–1659

formed after passing of TS12 which its formation barrieris 33.1 kcal mol-1. In this step of reaction H32, which is anacidic proton, is transferred to N2 (Fig. 2c). The lastcyclization is done through addition of enamine toaldehyde in backbone of glyceraldehyde which is a kindof cross-aldol reaction.
This kind of phenomena has also been found in Morita–Baylis–Hillman reaction and mechanism of direct aldolreactions catalyzed by amino amide derivatives in otherstudies [14, 41]. In this step, H16 is shuttled to O17 throughforming a less strained ten-membered-ring. Four watermolecules assist this cyclization; two of these molecules
serve as a bridge to help the proton transfer and the otherones make the cooperation effect to stabilize the structure viathe formation of a hydrogen bond (Fig. 2c). As can be seenfrom Fig. 1, activation free energy for last cyclization (TS13)is about 18 kcal mol-1. Distance between C23-C33 in TS13reach to 1.7 and it shows that second cyclization throughcarbon-carbon bond formation mechanism can be done.Addition reactions of enamines through new C-C bondformation have been observed experimentally [10]. Thereaction proceeds downhill from TS13 to form a seven-membered heterocyclic ring attached to a five-membered one(Int22) whose stability is 17.2 kcal mol-1 more than Int21.
TS6 TS7
TS8 TS9
TS10 TS12
N1
N2
N3
O8
O7
H11
O15
H13
H12
O19
C20
O7
O8
N2
N3H12
O19 H11H13
O15
C20
N1
O7
O8
N2 N1
N3
H13O15
H22
GLA
O17
O16C21
H26
O16
O17
O15O19
H11
C20N1
H22
H26
O17
O16
H27H12
O19 H26
O15
H30C21 C23
C33H32
O8
H10
O7
H16
N2
c
Fig. 2 (continued)
J Mol Model (2012) 18:1645–1659 1653

The last proton transfer in this process is performed from C8
to C7 (see Fig. 2c TS14) and leads to the final product.
B. Comparison of cross-links
Formation mechanism of GODIC and MODIC(Scheme 3) proceeds through two main steps namely: (1)Schiff base formation (Scheme 3a), (2) addition of methylguanidine (Scheme 3b). All optimized structures for these
cross-links are given in the Supporting information. Theirfree energy surfaces in gas and solvent phases are depictedin Figs. 3 and 4 respectively. When comparing these cross-links (see Scheme 3 along with Figs. 3 and 4), we find thatin the first TS corresponding to the carbinolamineformation, relative free energies as compared to separatedreactants are the same in both cross-links. Moreover,barriers of the first dehydration reaction (TS2) in theformation of GODIC and MODIC are equal to 32.3 and33.5 kcal mol-1 respectively. These barriers have highest
TS11
TS13 TS14
Pr
O15
O19
O17
H27
H13
H22
C21
C23
C33C23
O17
H13
O19O8
H16
N2
H32
C8 C7
H34
d
Fig. 2 (continued)
1654 J Mol Model (2012) 18:1645–1659
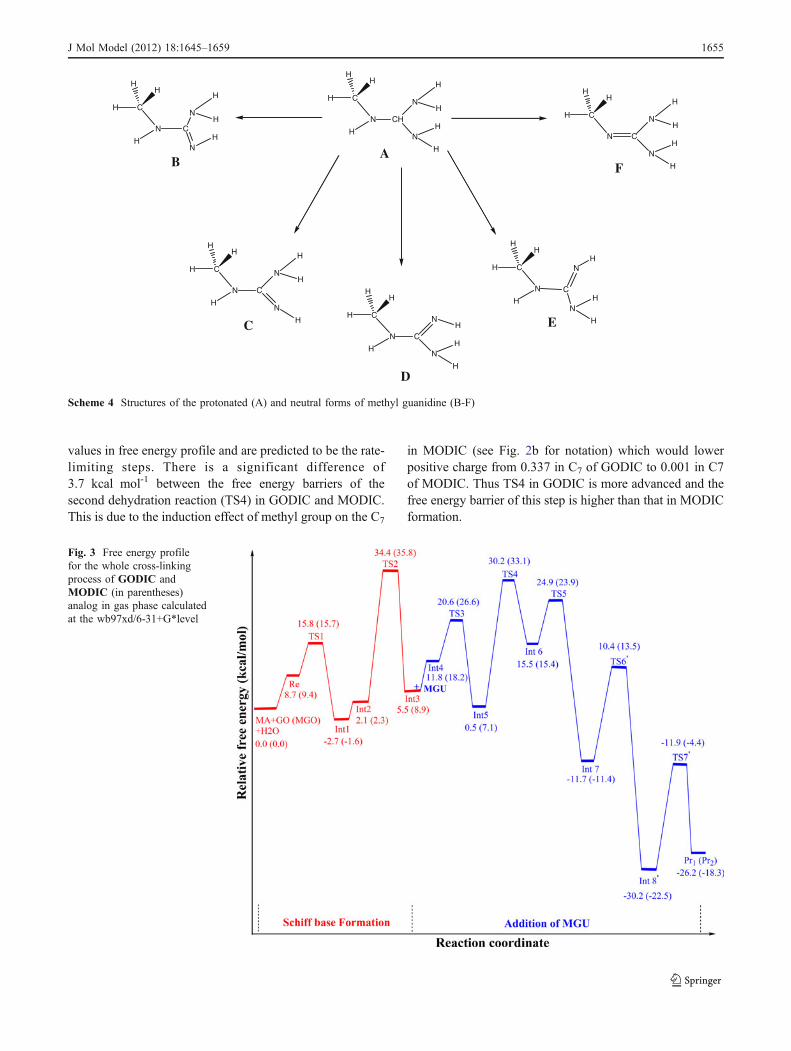
values in free energy profile and are predicted to be the rate-limiting steps. There is a significant difference of3.7 kcal mol-1 between the free energy barriers of thesecond dehydration reaction (TS4) in GODIC and MODIC.This is due to the induction effect of methyl group on the C7
in MODIC (see Fig. 2b for notation) which would lowerpositive charge from 0.337 in C7 of GODIC to 0.001 in C7of MODIC. Thus TS4 in GODIC is more advanced and thefree energy barrier of this step is higher than that in MODICformation.
N
N
C
H
HH
H
H
H
H
H
N
N
C
H
HH
H
H
H
H
N
N
C
H
HH
H
H
H
H N
N
C
H
HH
HH
H
H
N C
N
N
C
H
HH
HH
H
H
N CHN C
N C
N C
N C
N
N
C
HH
H
H
H
H
HA
B
C
D
E
F
Scheme 4 Structures of the protonated (A) and neutral forms of methyl guanidine (B-F)
Fig. 3 Free energy profilefor the whole cross-linkingprocess of GODIC andMODIC (in parentheses)analog in gas phase calculatedat the wb97xd/6-31+G*level
J Mol Model (2012) 18:1645–1659 1655
When we compare Figs. 1 and 3, it is clear that thesecross-links share steps of Schiff base formation andaddition of MGU with glucosepane. As it can be seen fromthe figures, from a thermodynamics point of view, the threeprocesses are exothermic and the glucosepane formation ismore favorable than each of GODIC and MODIC (by ca.4.1 and 12.0 kcal mol-1 respectively). however, the freeenergy barrier of the rate-limiting step in the glucosepaneformation is higher than those in the GODIC and MODICformation (40.8 kcal mol-1 against 32.3 and 33.5 kcal mol-1
respectively). So its formation is slower than others.Table 2 summarizes levels of cross-links in vivo. In the
different matrices of human, concentration of glucosepaneis more important than each of MODIC and GODIC, whichcorroborates with the fact that its formation is highlyexothermic. Moreover experimental results [6, 46] haveconfirmed that time incubation for its formation is higherthan the two others cross-links. So our results are in goodagreement with experimental evidence in vivo.
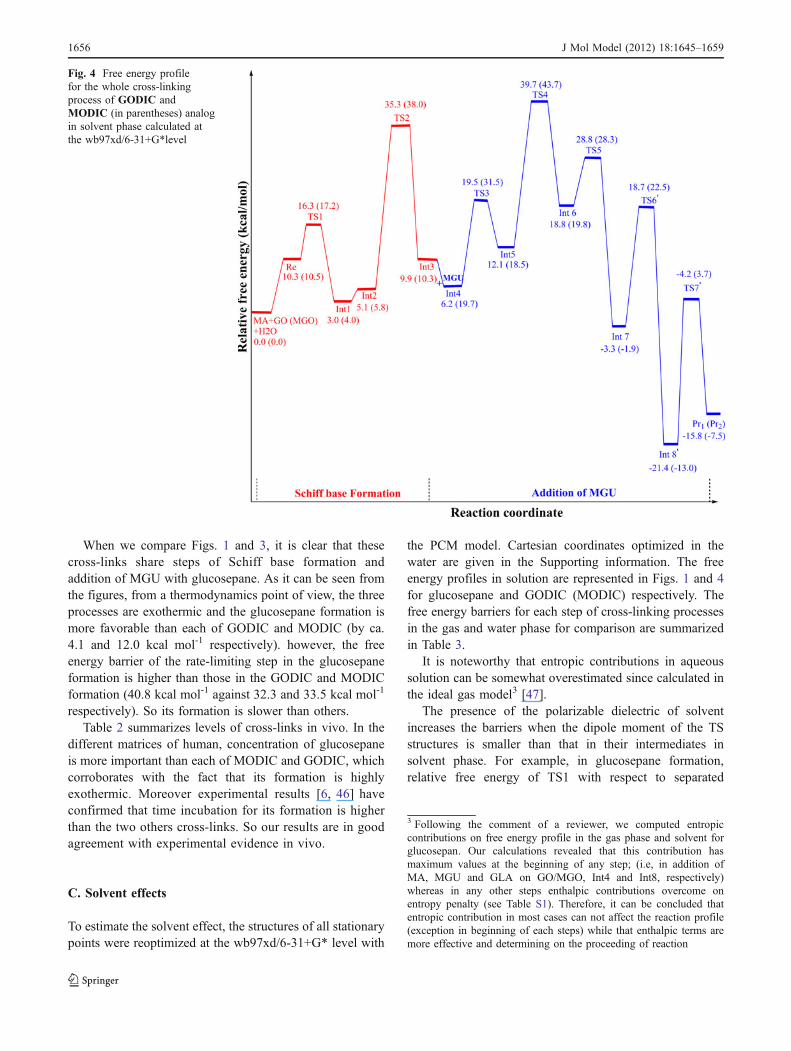
C. Solvent effects
To estimate the solvent effect, the structures of all stationarypoints were reoptimized at the wb97xd/6-31+G* level with
the PCM model. Cartesian coordinates optimized in thewater are given in the Supporting information. The freeenergy profiles in solution are represented in Figs. 1 and 4for glucosepane and GODIC (MODIC) respectively. Thefree energy barriers for each step of cross-linking processesin the gas and water phase for comparison are summarizedin Table 3.
It is noteworthy that entropic contributions in aqueoussolution can be somewhat overestimated since calculated inthe ideal gas model3 [47].
The presence of the polarizable dielectric of solventincreases the barriers when the dipole moment of the TSstructures is smaller than that in their intermediates insolvent phase. For example, in glucosepane formation,relative free energy of TS1 with respect to separated
3 Following the comment of a reviewer, we computed entropiccontributions on free energy profile in the gas phase and solvent forglucosepan. Our calculations revealed that this contribution hasmaximum values at the beginning of any step; (i.e, in addition ofMA, MGU and GLA on GO/MGO, Int4 and Int8, respectively)whereas in any other steps enthalpic contributions overcome onentropy penalty (see Table S1). Therefore, it can be concluded thatentropic contribution in most cases can not affect the reaction profile(exception in beginning of each steps) while that enthalpic terms aremore effective and determining on the proceeding of reaction
Fig. 4 Free energy profilefor the whole cross-linkingprocess of GODIC andMODIC (in parentheses) analogin solvent phase calculated atthe wb97xd/6-31+G*level
1656 J Mol Model (2012) 18:1645–1659
reactants increases by 1.5 kcal mol-1 in solution. It can beattributed to that the dipole moment of TS1 (5.1 Debye) issmaller than that of MA+MGO+H2O separated reactant(5.8 Debye). So the transition state is destabilized in solvent
water by solvation. On the contrary, the dipole moment ofTS7 (13.9 Debye) is larger than that of Int10 (6.2 Debye),and the solvation effect on TS7 is stronger than that onInt11. Therefore, the solvent water stabilizes the transitionstate TS11, leading to a decrease of free energy barrier by10.4 kcal mol-1. It can be conclude that by increasing ofdipole moment of TSs relative to their intermediates(positive values of Δμ (sol) in Table 3), the free energybarriers in solution decrease and inverse too.
Thermodynamics of the whole cross-linking processesfor glucosepane, GODIC and MODIC are disfavored by12.2, 10.4 and 10.8 kcal mol-1 when solvent is taken intoaccount. However these reactions are still favorable (by ca.18.1, 15.8 and 7.5 respectively for mentioned cross-links)(see Table 3 for comparison).
The rate-limiting steps stay the same for all three cross-links formation and the first dehydration reaction (TS2) for theformation of GODIC and MODIC and the last keto-enolconversion on the backbone of glyceraldehyde in the third
Table 2 Levels of the cross-links in different matrices of humana
Glucosepane GODIC MODIC
Human skin collagen 2000 (5000)b 15 (<60) 30 (<60)
Glomerular basementmembrane (GBM)
500 (900) 80 60
Human lens proteins 185.1 21.1 65.5
Human serum albumin(HSA)
17.1 (29.2) N.Dc 4.1 (3.9)
a Data obtained from references 5 and 17b All values are expressed as mmol/mg of protein and in parenthesesare amount of cross-links in diabetic subjectsc N.D stands for "not detected"
Mechanism of GODIC (MODIC)
ΔGgas≠ ΔGsol
≠ ΔGsol≠ - ΔGgas
≠ ΔμSOLa
Re1 0.0 0.0 0.0 0.0
Re1 –– TS1 15.8 (15.7b) 16.3 (17.2) 0.5 (1.5) −1.0 (−0.6)INT2 –– TS2 32.3 (33.5) 30.2 (32.2) −2.1 (−1.2) 2.3 (3.4)
INT4 –– TS3 8.8 (8.4) 13.3 (11.8) 4.5 (3.4) −2.5 (−2.0)INT5 –– TS4 29.7 (26.0) 27.6 (25.2) −2.1 (−0.8) 2.4 (2.1)
INT7 –– TS5 0.4 (0.3) −0.8 (0.9) −1.2 (−1.1) 0.5 (0.7)
INT8 –– TS6 22.0 (25.0) 22.0 (24.4) 0.1 (−0.6) −0.1 (0.1)
INT9 –– TS7 18.3 (18.1) 17.2 (16.7) −1.1 (−1.4) 1.0 (0.7)
Mechanism of glucosepane
Re2 0.0 0.0 0.0 0.0
Re2 –— TS1 15.7 17.2 1.5 −0.6INT2 –— TS2 33.5 32.2 −1.2 3.4
INT4 –— TS3 8.4 11.8 3.4 −2.0INT5 –— TS4 26.0 25.2 −0.8 2.1
INT7 –— TS5 0.3 −0.9 −1.1 0.7
INT9 –— TS6 13.5 13.0 −0.5 1.5
INT11——— TS7 38.8 28.3 −10.4 7.8
INT12——— TS8 0.2 2.3 2.1 −1.3INT14——— TS9 21.5 19.2 −2.4 13.0
INT16——— TS10 40.8 37.8 −3.0 0.2
INT18——— TS11 18.7 15.2 −3.5 0.2
INT20——— TS12 33.1 31.1 −1.9 0.7
INT22——— TS13 18.4 18.8 0.4 −2.4INT23——— TS14 13.8 15.5 1.7 −3.3Thermodynamics of reactions
ΔGgasr ΔGsol
r ΔGsolr - ΔGgas
r ΔμSOLc
Re3 –— Pr1 (Pr2) −26.2 (−18.3) −15.8 (−7.5) 10.4 (10.8) −7.8 (−6.4)Re4 –— Pr3 −30.3 −18.1 12.2 −6.5
Table 3 The reaction barriersand relative free energies re-spect to separated reactantsexpressed in kcal mol-1 for eachreaction step of GODIC(MODIC) and Glucosepanemodels in the gas and solventphase at wb97xd/6-31+G* levelof theory
aΔμSOL=μTS - μINT
b Value in parentheses are forMODIC modelcΔμSOL=μ
Pr - μRe
Re1 is MA+GO (MGO)+H2Oas separated
R2 is MA+MGO+H2O asseparated
Re3 is R1+MGU as separated
Re4 is R2+MGU+GLA asseparated
J Mol Model (2012) 18:1645–1659 1657

step of process for the Glucosepane are still rate-determiningsteps.
Conclusions
In summary, our studies focused upon the cross linkingreactions between α-oxoaldehydes and models of lysine andarginine residues in order to better understand cross-linkingchemistry in vivo and food chemistry. Our results show thatthese reactions are thermodynamically and kinetically favor-able in gas phase and solution with a high stabilization of thefree energy of the glucosepane in respect to the separatedreactants and values of free energy barriers are reasonable.Comparisons with the formation of two other important cross-links, GODIC and MODIC, show that the glucosepaneformation is more exothermic than others while its formationkinetics is slower. These results could illuminate mechanismsof AGE formation in diabetes and aging as well as suggestpotential therapeutic interventions in good agreement withexperimental results in glycation of human serum albuminunder physiological conditions.
Acknowledgments The financial support of Research Council ofShahid Beheshti University is gratefully acknowledged. Rasoul Nasiriwould like to thank the Ministry of Science, Research, andTechnology of Iran for financial support. Many thanks also go toMartin Field and Andrew Greene for their helpful discussions.
References
1. Ahmed N, Argirov OK, Minhas HS, Cordeiro CAA, ThornalleyPJ (2002) Assay of advanced glycation endproducts (AGEs):Surveying AGEs by chromatographic assay with derivatization by6-aminoquinolyl-N-hydroxysuccinimidyl-carbamate and applicationto Nε-carboxymethyl-lysine- and nε-(1-carboxyethyl)lysine-modifiedalbumin. Biochem J 364:1–14
2. Ahmed N, Thornalley PJ (2002) Chromatographic assay ofglycation adducts in human serum albumin glycated in vitro byderivatization with 6-aminoquinolyl-N-hydroxysuccinimidyl-carbamate and intrinsic fluorescence. Biochem J 364:15–24
3. Beisswenger PJ, Szwergold BS, Yeo KT (2001) Glycated proteinsin diabetes. Clin Lab Med 21:53–78
4. Biemel KM, Bühler HP, Reihl O, Lederer MO (2001) Identifica-tion and quantitative evaluation of the lysine-arginine crosslinksGODIC, MODIC, DODIC, and glucosepan in foods. NahrungFood 45:210–214
5. Biemel KM, Conrad J, Lederer MO (2002) Unexpected carbonylmobility in aminoketoses: The key to major maillard crosslinks.Angew Chem Int Ed 41:80–804
6. Biemel KM, Friedl DA, Lederer MO (2002) Identification andquantification of major maillard cross-links in human serumalbumin and lens protein. Evidence for glucosepane as thedominant compound. J Biol Chem 277:24907–24915
7. Biemel KM, Reihl O, Conrad J, Lederer MO (2001) Formationpathways for lysine-arginine cross-links derived from hexoses and
pentoses by maillard processes: Unraveling the structure of apentosidine precursor. J Biol Chem 276:23405–23412
8. Bierhaus A, Schiekofer S, Schwaninger M, Andrassy M, HumpertPM, Chen J, Hong M, Luther HT, Klöting I, Morcos M, HofmannM, Tritschler H, Weigle B, Kasper M, Smith M, Perry G, SchmidtAM, Stern DM, Häring HU, Schleicher E, Nawroth PP (2001)Diabetes-associated sustained activation of the transcription factornuclear factor-κB. Diabetes 50:2792–2808
9. Bose T, Chakraborti AS (2008) Fructose-induced structural andfunctional modifications of hemoglobin: Implication for oxidativestress in diabetes mellitus. Biochim Biophys Acta 1780:800–808
10. Brannock KC, Bell A, Burpitt RD, Kelly CA (1964) Enaminechemistry. IV. Cycloaddition reactions of enamines derived fromaldehydes and acyclic ketones. J Org Chem 29:801–812
11. Bucciarelli LG, Wendt T, Rong Lalla E, Hofmann MA, Goova MT,Taguchi A, Yan S, Yan SD, Stern DM, Schmidt AM (2002) RAGEis a multiligand receptor of the immunoglobulin superfamily:Implications for homeostasis and chronic disease. Cell Mol LifeSci 59:1117–1128
12. Candiano GM, Ghiggeri G, Delfino C, Quierolo C, Cuniberti E,Righetti PG (1985) Reaction of lysine with aldoses. CarbohydrRes 145:99–112
13. Chai JD, Head-Gordon M (2008) Long-range corrected hybriddensity functionals with damped atom-atom dispersion corrections.Phys Chem Chem Phys 10:6615–6620
14. Dong L, Qin S, Su Z, Yang H, Hu C (2010) Computationalinvestigation on the mechanism and the stereoselectivity ofMorita-Baylis-Hillman reaction and the effect of the bifunctionalcatalyst N-methylprolinol. Org Biomol Chem 8:3985–3991
15. Frisch AE, Dennington RD, Keith TA, Neilsen AB, Holder AJ(2003) GaussView, Rev 3.9. Gaussian Inc, Pittsburgh
16. Frisch MJ et al (2009) Gaussian 09, Revision A.01. Gaussian Inc,Wallingford
17. Gerrard JA, Brown PK, Fayle SE (2002) Maillard crosslinkingof food proteins I: The reaction of glutaraldehyde, formalde-hyde and glyceraldehyde with ribonuclease. Food Chem79:343–349
18. Gerrard JA, Brown PK, Fayle SE (2003) Maillard crosslinking offood proteins II: The reactions of glutaraldehyde, formaldehydeand glyceraldehyde with wheat proteins in vitro and in situ. FoodChem 80:35–43
19. Gerrard JA, Brown PK, Fayle SE (2003) Maillard crosslinkingof food proteins III: The effects of glutaraldehyde, formalde-hyde and glyceraldehyde upon bread and croissants. FoodChem 80:45–50
20. Gonzalez C, Schlegel HB (1989) An improved algorithm forreaction path following. J Chem Phys 90:2154–2161
21. Gonzalez C, Schlegel HB (1990) Reaction path following inmass-weighted internal coordinates. J Phys Chem 94:5523–5527
22. Grillo MA, Colombatto S (2008) Advanced glycation end-products (AGEs): Involvement in aging and in neurodegenerativediseases. Amino Acids 35:29–36
23. Guthrie JP, Cossar J (1986) The chlorination of acetone: acomplete kinetic analysis. Can J Chem 64:1250–1266
24. Hall NE, Smith BJ (1998) High-level ab initio molecularorbital calculations of imine formation. J Phys Chem A102:4930–4938
25. Harrington CR, Colaco CA (1994) A glycation connection. Nature370:247–248
26. Hartog JW, Voors AA, Bakker SJ, Smit AJ, van Veldhuisen DJ(2007) Advanced glycation end-products (AGEs) and heartfailure: Pathophysiology and clinical implications. Eur J HeartFail 9:1146–1155
27. Hayashi T, Shibamoto T (1985) Analysis of methyl glyoxal infoods and beverages. J Agric Food Chem 33:1090–1093
1658 J Mol Model (2012) 18:1645–1659

28. Henle T (2005) Protein-bound advanced glycation endproducts(AGEs) as bioactive amino acid derivatives in foods. AminoAcids 29:313–322
29. Hinton DJS, Ames JM (2006) Site specificity of glycation andcarboxymethylation of bovine serum albumin by fructose. AminoAcids 30:425–433
30. Hodge JE (1953) Dehydrated foods: Chemistry of browningreactions in model systems. J Agric Food Chem 1:928–943
31. Kimura Y, Hyogo H, Yamagishi S-I, Takeuchi M, Ishitobi T,Nabeshima Y, Arihiro K, Chayama K (2010) Atorvastatindecreases serum levels of advanced glycation endproducts (AGEs)in nonalcoholic steatohepatitis (NASH) patients with dyslipide-mia: Clinical usefulness of AGEs as a biomarker for theattenuation of NASH. J Gast 45:750–757
32. Kitahara Y, Takeuchi M, Miura K, Mine T, Matsui T, Yamagishi S(2008) Glyceraldehyde-derived advanced glycation end products(AGEs). A novel biomarker of postprandial hyperglycaemia indiabetic rats. Clin Exp Med 8:175–177
33. Lederer MO, Bühler HP (1999) Cross-linking of proteins by maillardprocesses characterization and detection of a lysine-arginine cross-linkderived from D-glucose. Bioorg Med Chem 7:1081–1088
34. Lederer MO, Klaiber RG (1999) Cross-linking of proteins byMaillard processes: Characterization and detection of lysine-arginine cross-links derived from glyoxal and methylglyoxal.Bioorg Med Chem 7:2499–2507
35. Ledl F, Schleicher E (1990) New aspects of the Maillard reaction infoods and in the human body. Angew Chem Int Ed 29:565–594
36. Lim JS, Mietus-Snyder M, Valente A, Schwarz JM, Lustig RH(2010) The role of fructose in the pathogenesis of NAFLD and themetabolic syndrome. Nature 7:251–264
37. Maillard LC (1912) Action des acides amines sur le sucres:Formation des me'lanoidines per voie me'thodique. C R Acad Sci154:66–68
38. Martins SI,Marcelis AT, van BoekelMA (2003) Kineticmodelling ofAmadori N-(1-deoxy-D-fructos-1-yl)-glycine degradation pathways.Part I - Reaction mechanism. Carbohydr Res 338:1651
39. Norberg J, Foloppe N, Nilsson L (2005) Intrinsic relativestabilities of the neutral tautomers of arginine side-chain models.J Chem Theory Comput 1:986–993
40. Nozaki Y, Tanford C (1967) Examination of titration behavior.Methods Enzymol 11:715–734
41. RaoQQ, Luo SW,Gong LZ (2010)Water as dual functional cocatalyst:A theoretical study on the mechanism of direct aldol reaction on watercatalyzed by a leucine derivative. Chin Sci Bull 55:1742–1752
42. Sato T, Iwaki M, Shimogaito N, Wu X, Yamagishi S-I, Takeuchi M(2006) TAGE (Toxic AGEs) theory in diabetic complications. CurrMol Med 6:351–358
43. Sattarahmady N, Moosavi-Movahedi AA, Ahmad F, HakimelahiGH, Habibi-Rezaei M, Saboury AA, Sheibani N (2007) Formation of
the molten globule-like state during prolonged glycation of humanserum albumin. Biochim Biophys Acta 1770:933–942
44. Sayer JM, Jenks WP (1977) Imine-forming elimination reactions.2. Imbalance of charge distribution in the transition state forcarbinolamine dehydration. J Am Chem Soc 99:464–474
45. Sayer JM, Pinsky B, Schonbrunn A, Washtien W (1974)Mechanism of carbinolamine formation. J Am Chem Soc96:7998–8009
46. Sell DR, Biemel KM, Reihl O, Lederer MO, Strauch CM,Monnier VM (2005) Glucosepane is a major protein cross-linkof the senescent human extracellular matrix: Relationship withdiabetes. J Biol Chem 280:12310–12315
47. Strajbl M, Sham YY, Vill-a J, Chu Z-T, Warshel AJ (2000)Calculations of Activation Entropies of Chemical Reactions inSolution. J Phys Chem B 104:4578–4584
48. Takeuchi M, Kikuchi S, Sazaki N, Suzuki T,Watai T, Iwaki M, BucalaR, Yamagishi S (2004) Involvement of advanced glycation end-products (AGEs) in Alzheimer's disease. Curr Alzheimer Res 1:39–46
49. Takeuchi M, Yamagishi S (2008) Possible involvement ofadvanced glycation end-products (AGEs) in the pathogenesis ofAlzheimer's disease. Curr Pharm Des 14:973–978
50. Thornalley PJ (2005) Dicarbonyl intermediates in the Maillardreaction. Ann NY Acad Sci 1043:111–117
51. Thornalley PJ, Langborg A, Minhas HS (1999) Formation ofglyoxal, methylglyoxal and 3-deoxyglucosone in the glycation ofproteins by glucose. Biochem J 344:109–116
52. Tomasi J, Persico M (1994) Molecular interactions in solution: Anoverview of methods based on continuous distributions of thesolvent. Chem Rev 94:2027–2094
53. Wells-Knecht KJ, Zyzak DV, Litchfield JE, Thorpe SR, BaynesJW (1995) Mechanism of autoxidative glycosylation: Identifi-cation of glyoxal and arabinose as intermediates in theautoxidative modification of proteins by glucose. Biochem34:3702–3709
54. Wolff SP, Crabbe MJC, Thornalley PJ (1984) The autoxidation ofglyceraldehyde and other simple monosaccharides. Experientia40:244–246
55. Wolff SP, Dean RT (1987) Glucose autoxidation and proteinmodification. The potential role of "autoxidative glycosylation" indiabetes. Biochem J 245:243–250
56. Yamagishi S-I (2011) Role of advanced glycation end products(AGEs) and receptor for AGEs (RAGE) in vascular damage indiabetes. Exp Gerontology 46:217–224
57. Yaylayan VA (1997) Classification of the Maillard reaction: Aconceptual approach. Trend Food Sci Tech 8:13–18
58. Zyzak DV, Richardson JM, Thorpe SR, Baynes JW (1995)Formation of reactive intermediates from Amadori compoundsunder physiological conditions. Arch Biochem Biophys316:547–554
J Mol Model (2012) 18:1645–1659 1659




















