
The Xerophyta viscosa Aldose Reductase (ALDRXV4) Confers Enhanced Drought and Salinity Tolerance to...
14
1 23 Molecular Biotechnology Part B of Applied Biochemistry and Biotechnology ISSN 1073-6085 Mol Biotechnol DOI 10.1007/s12033-012-9567-y The Xerophyta viscosa Aldose Reductase (ALDRXV4) Confers Enhanced Drought and Salinity Tolerance to Transgenic Tobacco Plants by Scavenging Methylglyoxal and Reducing the Membrane Damage Deepak Kumar, Preeti Singh, Mohd Aslam Yusuf, Chandrama Prakash Upadhyaya, Suchandra Deb Roy, Thomas Hohn & Neera Bhalla Sarin
-
Upload
mobilelandus -
Category
Documents
-
view
0 -
download
0
Transcript of The Xerophyta viscosa Aldose Reductase (ALDRXV4) Confers Enhanced Drought and Salinity Tolerance to...

1 23
Molecular BiotechnologyPart B of Applied Biochemistry andBiotechnology ISSN 1073-6085 Mol BiotechnolDOI 10.1007/s12033-012-9567-y
The Xerophyta viscosa Aldose Reductase(ALDRXV4) Confers EnhancedDrought and Salinity Tolerance toTransgenic Tobacco Plants by ScavengingMethylglyoxal and Reducing theMembrane DamageDeepak Kumar, Preeti Singh, MohdAslam Yusuf, Chandrama PrakashUpadhyaya, Suchandra Deb Roy,Thomas Hohn & Neera Bhalla Sarin

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media, LLC. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

RESEARCH
The Xerophyta viscosa Aldose Reductase (ALDRXV4)Confers Enhanced Drought and Salinity Tolerance to TransgenicTobacco Plants by Scavenging Methylglyoxal and Reducingthe Membrane Damage
Deepak Kumar • Preeti Singh • Mohd Aslam Yusuf •
Chandrama Prakash Upadhyaya • Suchandra Deb Roy •
Thomas Hohn • Neera Bhalla Sarin
� Springer Science+Business Media, LLC 2012
Abstract We report the efficacy of an aldose reductase
(ALDRXV4) enzyme from Xerophyta viscosa Baker in
enhancing the prospects of plant’s survival under abiotic
stress. Transgenic tobacco plants overexpressing ALD-
RXV4 cDNA showed alleviation of NaCl and mannitol-
induced abiotic stress. The transgenic plants survived
longer periods of water deficiency and salinity stress and
exhibited improved recovery after rehydration as compared
to the wild type plants. The increased synthesis of aldose
reductase in transgenic plants correlated with reduced
methylglyoxal and malondialdehyde accumulation and an
elevated level of sorbitol under stress conditions. In addi-
tion, the transgenic lines showed better photosynthetic
efficiency, less electrolyte damage, greater water retention,
higher proline accumulation, and favorable ionic balance
under stress conditions. Together, these findings suggest
the potential of engineering aldose reductase levels for
better performance of crop plants growing under drought
and salt stress conditions.
Keywords Abiotic stress � Aldose reductase �Methylglyoxal � Transgenics � Xerophyta viscosa
Abbreviations
FW Fresh weight
MDA Malondialdehyde
MG Methylglyoxal
ROS Reactive oxygen species
RWC Relative water content
TW Turgid weight
WT Wild type
Introduction
Different environmental stresses imposed on plants may
result in similar responses at the cellular and molecular
levels. This is due to the fact that diverse environmental
stresses often activate similar cell signaling pathways and
cellular processes, such as the production of stress proteins,
up-regulation of antioxidants, and accumulation of com-
patible solutes [1, 2]. It has been demonstrated that the
level of methylglyoxal (MG), a cytotoxic compound
increases upon exposure of plants to various abiotic stres-
ses [3] as does the content of reactive oxygen species
(ROS) [4]. At higher concentrations, MG is harmful to the
system as it is a potent cytotoxin and reacts with the major
macromolecules like RNA, DNA, and proteins leading to
cell death [5]. MG is mainly catabolized by two major
enzymatic pathways—the glyoxalase pathway [6] and an
alternate pathway involving aldose/aldehyde reductases
(ALR) that convert MG into acetol in an NADPH-depen-
dent two-step reaction [7].
Deepak Kumar and Preeti Singh contributed equally to this study.
D. Kumar � P. Singh � M. A. Yusuf � C. P. Upadhyaya �S. D. Roy � N. B. Sarin (&)
School of Life Sciences, Jawaharlal Nehru University,
New Delhi 110067, India
e-mail: [email protected]
Present Address:C. P. Upadhyaya
Department of Botany, Guru Ghasidas Central University,
Bilaspur 495009, India
T. Hohn
Institute of Botany, University of Basel,
Schonbeinstrasse 6, 4056 Basel, Switzerland
Mol Biotechnol
DOI 10.1007/s12033-012-9567-y
Author's personal copy

ALR (EC.1.1.1.21) belongs to the well-conserved aldo–
keto reductase super family of enzymes in plants and ani-
mals. They are monomeric, cytosolic proteins that catalyze
the NADPH-dependent reduction of a variety of carbonyl
metabolites including the aldehyde form of glucose which
is subsequently reduced to the corresponding sugar alcohol,
sorbitol, that is associated with maintenance of osmotic
balance of the cytoplasm and protection of macromolecules
in both animals and plant systems, under stress [8, 9].
The aldose reductase homologs have been shown to
have osmoprotective function and linked to the acquisition
of desiccation and freezing tolerance, as well as mainte-
nance of seed dormancy and longevity [10, 11]. The
recently characterized rice aldo–keto reductase (OsAKR1)
protected transgenic tobacco plants against oxidative and
heat stress [12]. An aldose reductase (ALDRXV4) gene was
cloned from the desiccation-tolerant plant Xerophyta vis-
cosa Baker, and identified because it was able to confer
tolerance to severe water loss in a desiccation-intolerant
mutant Escherichia coli strain [13].
In this study, we functionally validated the efficacy of
overexpression of ALDRXV4 gene in the model plant
tobacco in enhancing the tolerance of transgenic plants to
drought and salinity stress as compared to wild type (WT)
plants. We also investigated the plausible mechanism for the
protection and found that increased detoxification of MG,
decreased lipid peroxidation, and an increase in the content
of osmolytes—sorbitol and proline contribute to the
enhanced tolerance of these transgenic plants against stress.
Materials and Methods
Vector Construction and Tobacco Transformation
The full-length cDNA of ALDRXV4 (accession no.
AF133841) cloned under CaMV 35S promoter in pGEMT
easy cloning vector (Promega, WI, USA) was provided by
Prof. Jennifer A. Thomson (University of Cape Town, South
Africa). The cDNA with CaMV 35S promoter was amplified
with primers containing NheI and BstEII restriction sites
nested within the forward (50AAAGGCTAGCGTTGAAG
ATGCCTCTGCCGAC30) and reverse (50GTAAGGTGACC
TTGCAATTACTTTAGACTTCACCG-30) primers, respec-
tively. The binary vector pCAMBIA 2301 (CAMBIA,
Canberra, Australia) used in this study contains a 2.8 kb
GUS expression cassette and neomycin phosphotransferase
(nptII) gene driven by the CaMV 35S promoter. The GUS
gene along with its upstream CaMV 35S promoter and lacZ awas removed by double digestion with restriction enzymes
XbaI (site present in the pUC18 MCS of pC2301) and BstEII
(site present upstream of the nos poly A in pC2301). The
PCR-amplified CaMV 35S-ALDRXV4 cDNA described
above was then cloned in this vector between the XbaI and
BstEII sites. The NheI and XbaI sites were, however, not
restored after this ligation. The construct, thus formed, was
named pCAM-ALDRXV4. It contained the neomycin phos-
photransferase gene (nptII) as the plant selectable marker.
The construct was mobilized into Agrobacterium tumefac-
iens strain GV3101 by freeze–thaw method [14]. Agrobac-
terium-mediated transformation of tobacco (Nicotiana
tabacum cv. Xanthium) leaf disks was performed according
to the standard protocol of Horsch et al. [15]. Regenerated
shoots were separated from the calli and transferred onto
Murashige and Skoog’s (MS) medium, with 100 mg/l
kanamycin for selection of the transformants. The plantlets
were subsequently hardened for 2 weeks and transferred to
soil for flower and seed setting.
Genomic DNA PCR and Southern Blot Analysis
Total genomic DNA was extracted from leaves of the
putative transformed and WT plants using the CTAB
method [16] and quantitated spectrophotometrically. The
presence of the transgene (ALDRXV4) in the T0 transgenic
lines was confirmed by PCR analysis using the same
primers as were used in the cloning of ALDRXV4 cDNA.
The amplified products were separated by electrophoresis
on a 0.8 % agarose gel.
Southern blot analysis was performed according to
standard procedures [17]. Genomic DNA (10 lg/lane)
from WT and PCR positive plants was digested with BstEII
and separated by electrophoresis on a 0.8 % agarose gel
and blotted onto a nylon membrane (Amersham Pharmacia,
USA). [a32P] dCTP-labeled probe (amplified from pCAM-
ALDRXV4 by PCR using the primers described above) was
prepared using Random Primer Labeling Kit (Amersham
Pharmacia Biotech, UK) and used for hybridization of the
membrane at 65 �C using standard procedures [17].
RT-PCR Analysis
Total RNA was isolated from leaves of both transgenic and
WT plants using TRIzol reagent (Invitrogen, CA, USA)
and treated with RNase-free DNase I in order to remove
any contaminating genomic DNA. RT-PCR amplification
was conducted using an RT-PCR kit (AccuScript, Strata-
gene, USA) in accordance with the manufacturer’s
instructions. Total RNA (1 lg) was used for generation of
first-strand cDNA using MMLV reverse transcriptase. The
gene specific primers for amplification of ALDRXV4 cDNA
were 50-AAAGGCTAGCGTTGAAGATGCCTCTGCCGA
C-30 and 50-GTAAGGTGACCTTGCAATTACTTTAGAC
TTCACCG-30. The primers for tobacco actin (as internal
control) were 50-AGTAAGGTGACCTTGCAATTACTTT
AGACTTCACCG-30 and 50-AAAGGCTAGCGTTGAAG
Mol Biotechnol
Author's personal copy

ATGCCTCTGCCGAC-30. The RT-PCR products were
visualized by electrophoresis on 1.2 % agarose gel.
Western Blot Analysis
To check the expression levels of ALDRXV4, soluble protein
was extracted from transgenic and WT tobacco plants. Young
leaves (100 mg fresh weight [FW] from transgenic as well as
untransformed control plants were ground in 200 ll extrac-
tion buffer (20 % glycerol, 62.5 mM Tris–HCl, pH 6.8, 5 %
2-mercaptoethanol, and 0.1 % SDS). The extract was centri-
fuged at 13,000 rpm for 20 min at 4 �C and the supernatant
was used for ALR enzyme assays and for western blot anal-
ysis. Protein concentration was estimated by the method of
Bradford [18]. Protein extracts were fractionated using 12 %
SDS-PAGE and transferred to nitrocellulose membrane
(Sigma-Aldrich) using the semi-dry transfer system (Owl
Separation Systems, UK). Transfer of proteins onto the
membrane was checked by staining with Ponceau S stain [19].
The membrane was briefly rinsed in Tris Buffered Saline with
Tween-20 (TBST) and incubated in blocking solution (2 %
BSA in TBST) for 1 h with gentle shaking at room tempera-
ture. The membrane was given brief washes (5 min each)
thrice with TBST and incubated for 1 h with maize aldose
reductase rabbit antiserum (1:2,000 dilution). The membrane
was washed thrice with TBST (5 min/wash), after which it
was incubated with secondary goat anti-rabbit IgG, conju-
gated with horse-radish peroxidase (Sigma-Aldrich; 1:5,000
dilution), for 1 h. The membrane was incubated with diam-
inobenzidine (DAB) prepared in TBS, till the color developed.
All the dilutions of primary and secondary antibodies were
made in TBST containing 0.3 % BSA.
Seed Germination and Seedling Growth
Seed germination was studied in three sets of *100 seeds
each of WT and T1 progeny of the transgenic lines. The
seeds were surface sterilized and placed on MS agar plates
with or without mannitol (200 mM) and NaCl (200 mM),
incubated at 25 ± 2 �C under 16 h light/8 h dark photo-
period with the light intensity of 50 lmol/m s-1. Germi-
nation was scored daily and percentage germination was
calculated after 7 days.
T1 seedlings were also tested for growth on different mannitol
and NaCl concentrations. The 7-day-old WT and kanamycin-
positive T1 seedlings were transferred to MS medium supple-
mented with 100, 200, and 300 mM mannitol or 100, 200, and
300 mM NaCl. Growth was visually followed up to day 15.
Leaf Disk Senescence Assay
Healthy and fully expanded leaves from the transgenic as
well as WT plants were briefly washed with sterile distilled
water and leaf disks of 1 cm diameter were punched out
and floated in a 10 ml solution of mannitol (200, 400, 600,
and 800 mM, 3–4 days), NaCl (200, 400, 600, and
800 mM, 3–4 days), or MG (5 and 10 mM, 2 days),
respectively. In all the above treatments, leaf disks floated
on distilled water served as experimental control. The
chlorophyll content in the leaf was estimated according to
the procedure of Arnon [20].
Stress Tolerance Experiments
For drought and salt stress treatments 6-day-old seedlings of
WT grown on MS plates and transgenic lines grown on MS
plates supplemented with 100 mg/l kanamycin were trans-
ferred into plastic pots containing agropeat and vermiculite
(3:1 v/v) in a growth chamber at 25 �C, 60 % relative
humidity, and 16 h light/8 h dark photoperiod with light
supplied at an intensity of 150 lmol/m2 s-1. For drought
stress WT and transgenic plants were left unwatered for
14 days and then rewatered for 6 days for recovery.
For salt stress treatment, the plants were irrigated with a
200 mM NaCl solution for 14 days and then with water for
6 days. For tolerance of the plants under abiotic stress
throughout their life cycle, seedlings selected on kanamycin
were transferred to earthen pots and grown in a glasshouse
(16 h light/8 h dark, 25 ± 2 �C). Starting 2 weeks after
transfer, the plants were watered biweekly with mannitol
(200 mM) or NaCl (200 mM) solution or water. These plants
were used for performing different experiments.
Measurements of Chlorophyll Fluorescence
Photosynthetic activity was measured as photochemical
yield (Fv/Fm), which represents the maximum quantum
yield of photosystem II, by recording the chlorophyll
fluorescence using a portable chlorophyll fluorescence
meter (Handy-PEA, Hansatech, UK) after 30 min of dark
adaptation of the leaves. Measurements were conducted at
room temperature (25 �C) using saturating light pulse of
white light (8,000 lmol/m2 s-1 for 0.8 s).
Measurement of relative water content (RWC)
For determination of RWC, fresh leaves were detached and
weighed immediately to record FW, followed by dipping
them in distilled water for 12 h. The leaves were then
blotted to wipe off excess water, weighed to record turgid
weight (TW), and subjected to oven drying at 70 �C for
24 h to record dry weight (DW). The RWC was determined
by the equation:
RWC ¼ ðFW� DWÞ � 100=ðTW� DWÞ:
Mol Biotechnol
Author's personal copy

Measurement of Electrolyte Leakage
Electrolyte leakage was determined according to the
method described by Sairam and Srivastava [21]. Leaves
were collected and washed with deionized water for three
times in order to remove surface-adhered electrolytes. They
were then placed in test tubes and immersed in 10 ml of
deionized water and the electrical conductivity (EC1) was
measured. After incubation at 55 �C for 30 min the EC2
was determined again. The samples were heated in boiling
water for 1 h before the total conductivity was measured in
the solution (EC3). Relative ion leakage was expressed as a
percentage of the total conductivity:
Relative Electrolyte Leakage¼ ½ðEC2�EC1Þ=EC3� � 100:
Measurement of Aldose Reductase Activity
The ALR enzyme activity was determined by monitoring
the decrease in OD at 340 nm due to NADPH utilization
[22]. The reaction mixture made in 0.1 M potassium
phosphate buffer (pH 6.2) contained 0.15 mM of NADPH,
20 mM MG as substrate, and the crude protein extract in a
final volume of 1 ml. The reference cuvette (blank) con-
tained everything except the substrate.
Estimation of Sorbitol
Sugar alcohols were estimated as described by Pommerr-
enig et al. [23]. Sugar alcohols were extracted from the
frozen powdered material by incubation in 80 % ethanol at
80 �C for 4 h. After centrifugation at 2,00,0009g for
15 min, 800 ll of the ethanolic supernatant was transferred
into a new tube and the ethanol was completely evaporated
under vacuum. The dry residue was dissolved in distilled
water, and the sorbitol content was determined enzymati-
cally with the use of a D-sorbitol colorimetric assay kit
(BioVision Inc., CA, USA). D-Sorbitol was oxidized by
NAD?-dependent sorbitol dehydrogenase to D-fructose and
the reduced NADH was consumed in a diaphorase reaction
for the reduction of iodonitrotetrazolium. The absorbance
of formazan product, thus formed, was measured at 560 nm
and the sorbitol content was estimated as per the manu-
facturer’s instructions.
Estimation of Methylglyoxal
MG was estimated according to the method of Yadav et al.
[3]. About 0.5 g leaf tissue was homogenized in 3 ml of
0.5 M perchloric acid. After incubating for 15 min on ice,
the mixture was centrifuged at 4 �C for 10 min at
11,0009g. The supernatant was decolorized by adding
charcoal (10 mg/ml), kept for 15 min at room temperature,
and centrifuged at 11,0009g for 10 min. Before using this
supernatant for MG assay, it was neutralized by incubating
with saturated solution of potassium carbonate for 15 min
at room temperature and centrifugation at 11,0009g for
10 min. The neutralized supernatant was used for MG
assay. In a total volume of 1 ml, 250 ll of 7.2 mM 1,2-
diaminobenzene, 100 ll of 5 M perchloric acid, and 650 ll
of the neutralized supernatant were added in that order. The
absorbance at 335 nm of the derivatized MG was read after
25 min in a Hitachi U-2000 spectrophotometer (Hitachi,
Japan). The final concentration of MG was calculated from
the standard curve and expressed in terms of lmol/g FW.
Measurement of MDA Content
The tobacco leaves were ground to a fine powder under
liquid nitrogen. 3 ml of 10 % trichloroacetic acid was
added to 0.2 g of the powder and left at 4 �C overnight.
After centrifugation at 1,0009g for 20 min, the supernatant
was transferred to a new tube for measurements. 2 ml of
0.6 % thiobarbituric acid (TBA) was added to 2 ml of the
supernatant. The mixture was vortexed thoroughly, heated
in boiling water for 15 min, cooled immediately and cen-
trifuged. Absorbance values of the supernatant were
detected at wavelengths of 532 and 450 nm using water as
blank. The formula for the calculation of MDA content
was:
MDA content (lmol/l) ¼ 6:45� OD532 � 0:56� OD450
Estimation of Proline
Proline content was estimated using the standard protocol
[24]. 100 mg of leaf tissue was crushed in liquid N2 and
homogenized in 10 ml of 3 % sulfosalicylic acid. The
homogenate was centrifuged at 15,000 rpm and to 2 ml of
the upper aqueous phase, 2 ml of 0.2 % ninhydrin and 2 ml
of glacial acetic acid were added. After incubation at
100 �C for 1 h, reaction was terminated on ice. Proline was
extracted from the mixture with 4 ml of toluene. The
optical density of the upper aqueous phase was measured at
520 nm and proline concentration was determined from the
standard graph made using the purified L-proline (Sigma,
MO, USA) and calculated on a FW basis (lmol/g).
Estimation of Endogenous Ion Content
For determination of endogenous Na? and K? contents,
100 mg leaf tissue (unstressed and salt-treated) of WT and
transgenic plants was taken and digested in 0.1 % HNO3.
Ions were extracted in distilled water by boiling for 30 min
twice. The filtered extract, thus obtained, was used to
measure specific ions using a flame photometer.
Mol Biotechnol
Author's personal copy

Statistical Analysis
All experiments were performed at least three times inde-
pendently. Results were assessed by Student’s t test. Sig-
nificance was defined as P \ 0.05.
Results
Generation and Analysis of Transgenic Tobacco Plants
Overexpressing ALDRXV4 cDNA
Six different tobacco transgenic lines (X1, X2, X3, X4, X5,
and X6) were generated by overexpressing ALDRXV4 gene
under the control of constitutive CaMV 35S promoter using
the construct shown in Fig. 1a. Integration of the ALD-
RXV4 transgene into T0 transgenic tobacco plants was
confirmed by PCR using the gene specific primers (data not
shown) and Southern blot analysis using CaMV 35S-
ALDRXV4 specific fragment as probe (Fig. 1a). While a
single-copy transgene insertion was seen in the lines X1,
X2, X3, X4, and X6, two-copies of the transgene were
found to be integrated into the genome of the transgenic
line X5 (Fig. 1b). The RT-PCR analysis showed that
mRNA of ALDRXV4 was successfully expressed in the six
transgenic lines (Fig. 1c), whereas it was absent in the WT
plant. Western blot analysis of representative lines using
polyclonal antibodies raised against maize ALR showed
the presence of a single prominent band of expected size
(36 kDa) confirming the expression of the transgene at
protein level (Fig. 1d). The transgenic plants grew nor-
mally in glass house and showed no morphological dif-
ferences when compared to the WT plants, thus, indicating
that expression of ALDRXV4 had no deleterious effects on
the morphological growth of these plants.
Overexpression of ALDRXV4 Enhances Drought
and Salt Stress Tolerance During Germination
and Early Seedling Development
To study the effect of overproduction of ALDRXV4 on the
germination of transgenic tobacco seeds exposed to
drought and salt stress, seeds of WT and T1 transgenic lines
(X1–X6) were germinated on MS medium (without kana-
mycin selection), supplemented with 200 mM mannitol
and 200 mM NaCl to simulate drought and salt stress,
respectively. The germination rates of WT and transgenic
seeds under non-stressed (MS medium) conditions were
similar and no phenotypic difference was observed in the
developing seedlings. However, germination of WT seeds
was significantly impaired under stress conditions when
compared to transgenic seeds. The germination of WT
seeds on medium supplemented with 200 mM mannitol
and 200 mM NaCl was 40 and 38 %, respectively, whereas
WT X1 X2 X3 X4 X5 X6B
ALDRXV4
CWT X1 X2 X3 X4 X5 X6
Actin
DWT X1 X2 X3 X4 X5 X6
36 KDa
Ponceau
A
probe ~ 1.3 kb
35ST 35SP lox
RBLB
lox 35SPnptII nosTXV4
BstEII
Fig. 1 Transformation of tobacco plants for overexpression of
ALDRXV4. a Schematic representation of the pCAM-ALDRXV4construct used for tobacco transformation. The BstEII restriction site
used in the Southern analysis is marked. The region for amplification
of the probe used in the Southern analysis is also marked. b Southern
blot analysis showing the integration and copy number of the
transgene. c RT-PCR analysis confirming expression of ALDRXV4 in
young fully expanded leaves of transgenic tobacco plants. Actin gene
was used as an internal control. d Western blot analysis of total
protein extract from leaves of WT and transgenic tobacco lines.
Lower panel shows the Ponceau stained membrane after transfer for
checking the equal loading of proteins. WT tobacco control. (X1, X2,
X3, X4, X5, and X6) independently transformed T1 transgenic lines
of tobacco
Mol Biotechnol
Author's personal copy
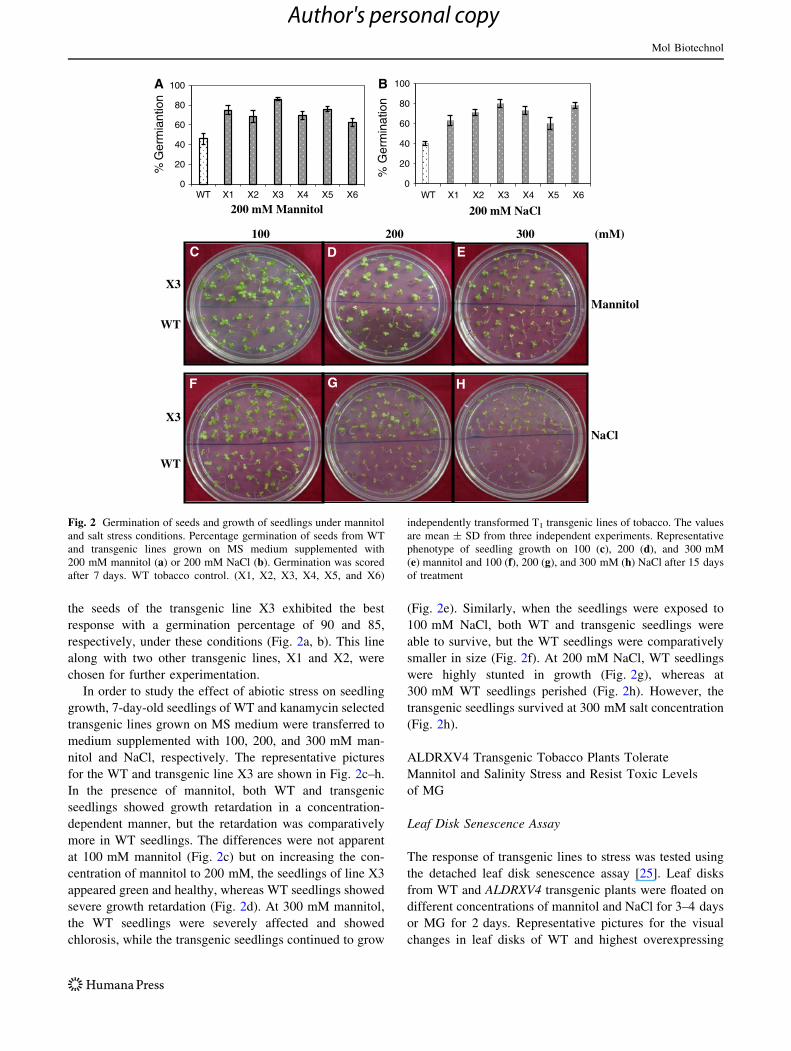
the seeds of the transgenic line X3 exhibited the best
response with a germination percentage of 90 and 85,
respectively, under these conditions (Fig. 2a, b). This line
along with two other transgenic lines, X1 and X2, were
chosen for further experimentation.
In order to study the effect of abiotic stress on seedling
growth, 7-day-old seedlings of WT and kanamycin selected
transgenic lines grown on MS medium were transferred to
medium supplemented with 100, 200, and 300 mM man-
nitol and NaCl, respectively. The representative pictures
for the WT and transgenic line X3 are shown in Fig. 2c–h.
In the presence of mannitol, both WT and transgenic
seedlings showed growth retardation in a concentration-
dependent manner, but the retardation was comparatively
more in WT seedlings. The differences were not apparent
at 100 mM mannitol (Fig. 2c) but on increasing the con-
centration of mannitol to 200 mM, the seedlings of line X3
appeared green and healthy, whereas WT seedlings showed
severe growth retardation (Fig. 2d). At 300 mM mannitol,
the WT seedlings were severely affected and showed
chlorosis, while the transgenic seedlings continued to grow
(Fig. 2e). Similarly, when the seedlings were exposed to
100 mM NaCl, both WT and transgenic seedlings were
able to survive, but the WT seedlings were comparatively
smaller in size (Fig. 2f). At 200 mM NaCl, WT seedlings
were highly stunted in growth (Fig. 2g), whereas at
300 mM WT seedlings perished (Fig. 2h). However, the
transgenic seedlings survived at 300 mM salt concentration
(Fig. 2h).
ALDRXV4 Transgenic Tobacco Plants Tolerate
Mannitol and Salinity Stress and Resist Toxic Levels
of MG
Leaf Disk Senescence Assay
The response of transgenic lines to stress was tested using
the detached leaf disk senescence assay [25]. Leaf disks
from WT and ALDRXV4 transgenic plants were floated on
different concentrations of mannitol and NaCl for 3–4 days
or MG for 2 days. Representative pictures for the visual
changes in leaf disks of WT and highest overexpressing
200 mM Mannitol
0
20
40
60
80
100
WT X1 X2 X3 X4 X5 X6
% G
erm
iant
ion
A
0
20
40
60
80
100
WT X1 X2 X3 X4 X5 X6
% G
erm
inat
ion
200 mM NaCl
B
C D E
F G H
X3
WT
X3
WT
Mannitol
100 200 300 (mM)
NaCl
Fig. 2 Germination of seeds and growth of seedlings under mannitol
and salt stress conditions. Percentage germination of seeds from WT
and transgenic lines grown on MS medium supplemented with
200 mM mannitol (a) or 200 mM NaCl (b). Germination was scored
after 7 days. WT tobacco control. (X1, X2, X3, X4, X5, and X6)
independently transformed T1 transgenic lines of tobacco. The values
are mean ± SD from three independent experiments. Representative
phenotype of seedling growth on 100 (c), 200 (d), and 300 mM
(e) mannitol and 100 (f), 200 (g), and 300 mM (h) NaCl after 15 days
of treatment
Mol Biotechnol
Author's personal copy
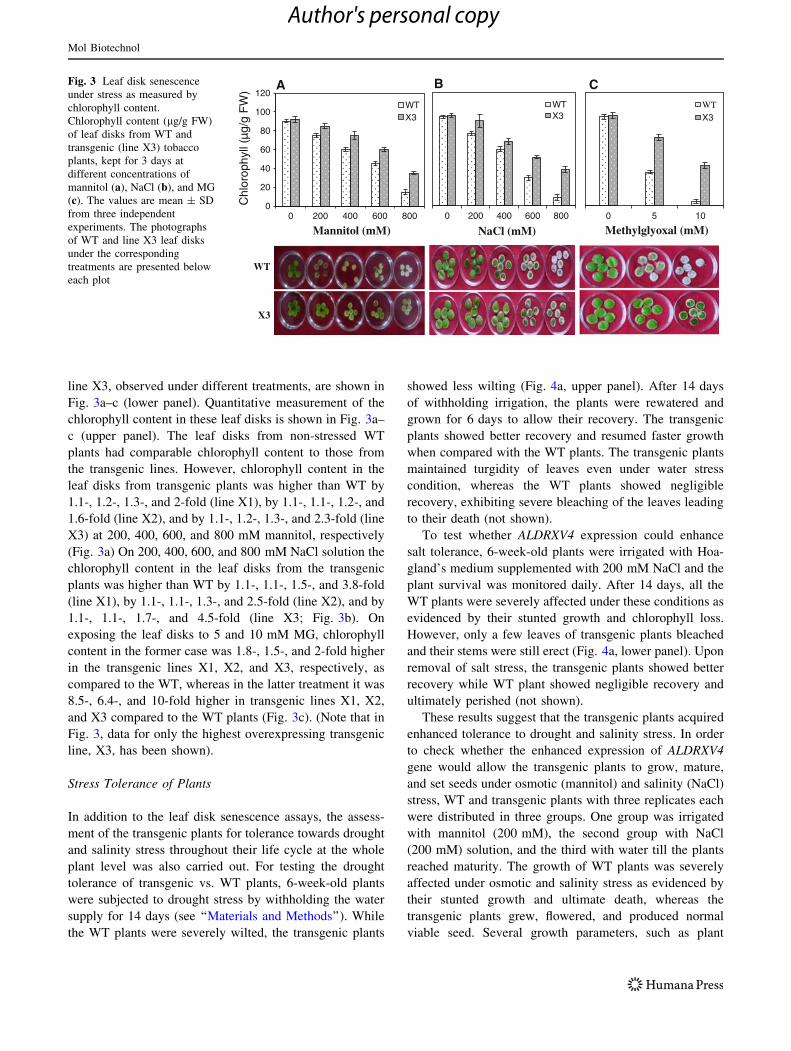
line X3, observed under different treatments, are shown in
Fig. 3a–c (lower panel). Quantitative measurement of the
chlorophyll content in these leaf disks is shown in Fig. 3a–
c (upper panel). The leaf disks from non-stressed WT
plants had comparable chlorophyll content to those from
the transgenic lines. However, chlorophyll content in the
leaf disks from transgenic plants was higher than WT by
1.1-, 1.2-, 1.3-, and 2-fold (line X1), by 1.1-, 1.1-, 1.2-, and
1.6-fold (line X2), and by 1.1-, 1.2-, 1.3-, and 2.3-fold (line
X3) at 200, 400, 600, and 800 mM mannitol, respectively
(Fig. 3a) On 200, 400, 600, and 800 mM NaCl solution the
chlorophyll content in the leaf disks from the transgenic
plants was higher than WT by 1.1-, 1.1-, 1.5-, and 3.8-fold
(line X1), by 1.1-, 1.1-, 1.3-, and 2.5-fold (line X2), and by
1.1-, 1.1-, 1.7-, and 4.5-fold (line X3; Fig. 3b). On
exposing the leaf disks to 5 and 10 mM MG, chlorophyll
content in the former case was 1.8-, 1.5-, and 2-fold higher
in the transgenic lines X1, X2, and X3, respectively, as
compared to the WT, whereas in the latter treatment it was
8.5-, 6.4-, and 10-fold higher in transgenic lines X1, X2,
and X3 compared to the WT plants (Fig. 3c). (Note that in
Fig. 3, data for only the highest overexpressing transgenic
line, X3, has been shown).
Stress Tolerance of Plants
In addition to the leaf disk senescence assays, the assess-
ment of the transgenic plants for tolerance towards drought
and salinity stress throughout their life cycle at the whole
plant level was also carried out. For testing the drought
tolerance of transgenic vs. WT plants, 6-week-old plants
were subjected to drought stress by withholding the water
supply for 14 days (see ‘‘Materials and Methods’’). While
the WT plants were severely wilted, the transgenic plants
showed less wilting (Fig. 4a, upper panel). After 14 days
of withholding irrigation, the plants were rewatered and
grown for 6 days to allow their recovery. The transgenic
plants showed better recovery and resumed faster growth
when compared with the WT plants. The transgenic plants
maintained turgidity of leaves even under water stress
condition, whereas the WT plants showed negligible
recovery, exhibiting severe bleaching of the leaves leading
to their death (not shown).
To test whether ALDRXV4 expression could enhance
salt tolerance, 6-week-old plants were irrigated with Hoa-
gland’s medium supplemented with 200 mM NaCl and the
plant survival was monitored daily. After 14 days, all the
WT plants were severely affected under these conditions as
evidenced by their stunted growth and chlorophyll loss.
However, only a few leaves of transgenic plants bleached
and their stems were still erect (Fig. 4a, lower panel). Upon
removal of salt stress, the transgenic plants showed better
recovery while WT plant showed negligible recovery and
ultimately perished (not shown).
These results suggest that the transgenic plants acquired
enhanced tolerance to drought and salinity stress. In order
to check whether the enhanced expression of ALDRXV4
gene would allow the transgenic plants to grow, mature,
and set seeds under osmotic (mannitol) and salinity (NaCl)
stress, WT and transgenic plants with three replicates each
were distributed in three groups. One group was irrigated
with mannitol (200 mM), the second group with NaCl
(200 mM) solution, and the third with water till the plants
reached maturity. The growth of WT plants was severely
affected under osmotic and salinity stress as evidenced by
their stunted growth and ultimate death, whereas the
transgenic plants grew, flowered, and produced normal
viable seed. Several growth parameters, such as plant
WT
X3
0
20
40
60
80
100
120
0 200 400 600 800
Chl
orop
hyll
(µg/
g F
W)
WTX3
A
0 200 400 600 800
WTX3
B
0 5 10
WT
X3
C
NaCl (mM) Methylglyoxal (mM)Mannitol (mM)
Fig. 3 Leaf disk senescence
under stress as measured by
chlorophyll content.
Chlorophyll content (lg/g FW)
of leaf disks from WT and
transgenic (line X3) tobacco
plants, kept for 3 days at
different concentrations of
mannitol (a), NaCl (b), and MG
(c). The values are mean ± SD
from three independent
experiments. The photographs
of WT and line X3 leaf disks
under the corresponding
treatments are presented below
each plot
Mol Biotechnol
Author's personal copy
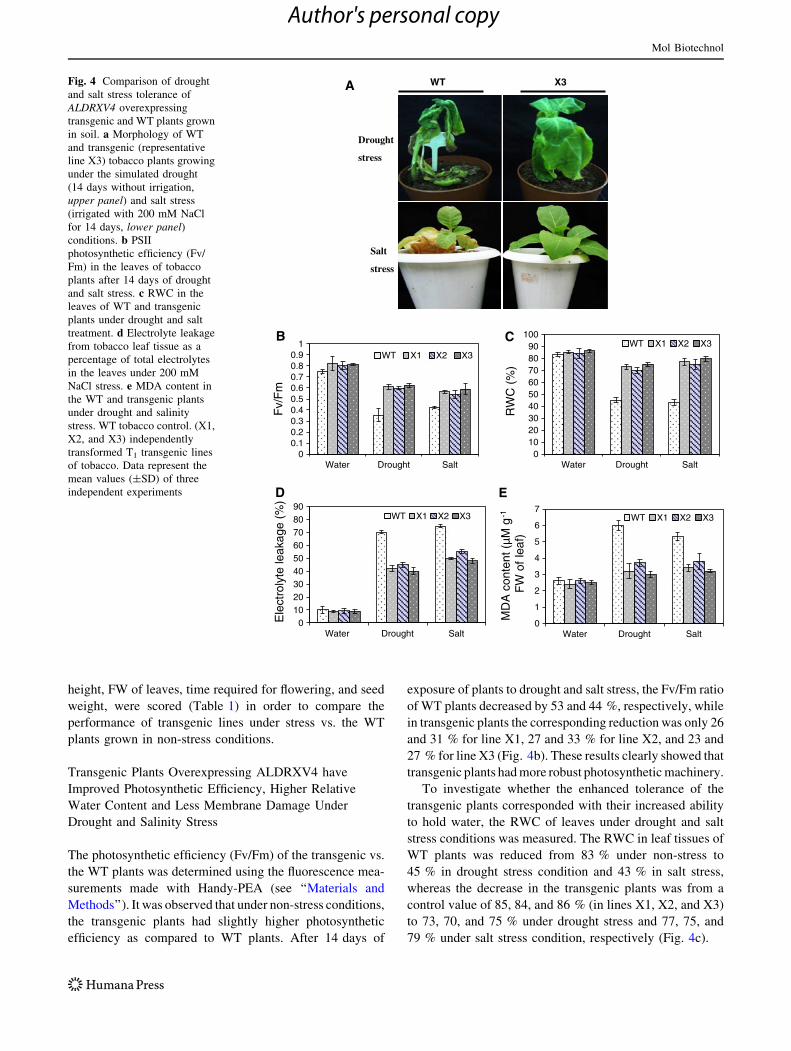
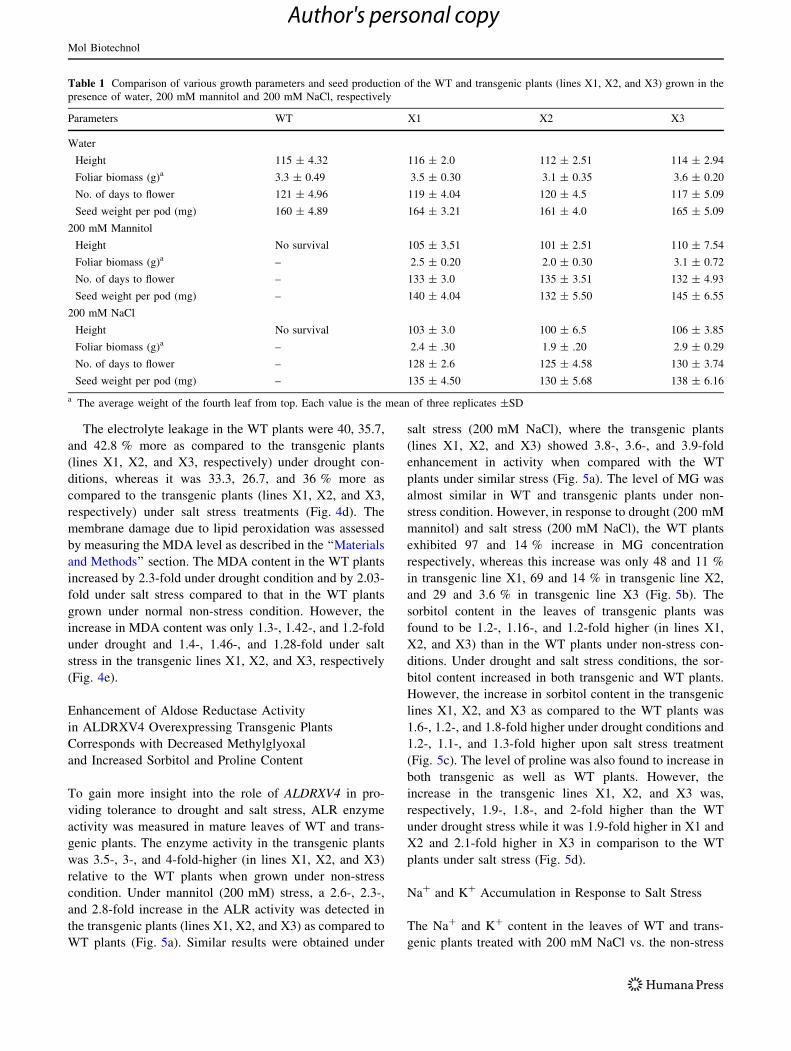
height, FW of leaves, time required for flowering, and seed
weight, were scored (Table 1) in order to compare the
performance of transgenic lines under stress vs. the WT
plants grown in non-stress conditions.
Transgenic Plants Overexpressing ALDRXV4 have
Improved Photosynthetic Efficiency, Higher Relative
Water Content and Less Membrane Damage Under
Drought and Salinity Stress
The photosynthetic efficiency (Fv/Fm) of the transgenic vs.
the WT plants was determined using the fluorescence mea-
surements made with Handy-PEA (see ‘‘Materials and
Methods’’). It was observed that under non-stress conditions,
the transgenic plants had slightly higher photosynthetic
efficiency as compared to WT plants. After 14 days of
exposure of plants to drought and salt stress, the Fv/Fm ratio
of WT plants decreased by 53 and 44 %, respectively, while
in transgenic plants the corresponding reduction was only 26
and 31 % for line X1, 27 and 33 % for line X2, and 23 and
27 % for line X3 (Fig. 4b). These results clearly showed that
transgenic plants had more robust photosynthetic machinery.
To investigate whether the enhanced tolerance of the
transgenic plants corresponded with their increased ability
to hold water, the RWC of leaves under drought and salt
stress conditions was measured. The RWC in leaf tissues of
WT plants was reduced from 83 % under non-stress to
45 % in drought stress condition and 43 % in salt stress,
whereas the decrease in the transgenic plants was from a
control value of 85, 84, and 86 % (in lines X1, X2, and X3)
to 73, 70, and 75 % under drought stress and 77, 75, and
79 % under salt stress condition, respectively (Fig. 4c).
Drought
stress
Salt
stress
WT X3A
0
1
2
3
4
5
6
7
Water Drought Salt
MD
A c
onte
nt (
µM g
-1
FW
of
leaf
)WT X1 X2 X3
E
0102030405060708090
Water Drought Salt
Ele
ctro
lyte
leak
age
(%)
WT X1 X2 X3
D
0102030405060708090
100
Water Drought Salt
RW
C (
%)
WT X1 X2 X3C
00.10.20.30.40.50.60.70.80.9
1
Water Drought Salt
Fv/
Fm
WT X1 X2 X3
B
Fig. 4 Comparison of drought
and salt stress tolerance of
ALDRXV4 overexpressing
transgenic and WT plants grown
in soil. a Morphology of WT
and transgenic (representative
line X3) tobacco plants growing
under the simulated drought
(14 days without irrigation,
upper panel) and salt stress
(irrigated with 200 mM NaCl
for 14 days, lower panel)conditions. b PSII
photosynthetic efficiency (Fv/
Fm) in the leaves of tobacco
plants after 14 days of drought
and salt stress. c RWC in the
leaves of WT and transgenic
plants under drought and salt
treatment. d Electrolyte leakage
from tobacco leaf tissue as a
percentage of total electrolytes
in the leaves under 200 mM
NaCl stress. e MDA content in
the WT and transgenic plants
under drought and salinity
stress. WT tobacco control. (X1,
X2, and X3) independently
transformed T1 transgenic lines
of tobacco. Data represent the
mean values (±SD) of three
independent experiments
Mol Biotechnol
Author's personal copy
The electrolyte leakage in the WT plants were 40, 35.7,
and 42.8 % more as compared to the transgenic plants
(lines X1, X2, and X3, respectively) under drought con-
ditions, whereas it was 33.3, 26.7, and 36 % more as
compared to the transgenic plants (lines X1, X2, and X3,
respectively) under salt stress treatments (Fig. 4d). The
membrane damage due to lipid peroxidation was assessed
by measuring the MDA level as described in the ‘‘Materials
and Methods’’ section. The MDA content in the WT plants
increased by 2.3-fold under drought condition and by 2.03-
fold under salt stress compared to that in the WT plants
grown under normal non-stress condition. However, the
increase in MDA content was only 1.3-, 1.42-, and 1.2-fold
under drought and 1.4-, 1.46-, and 1.28-fold under salt
stress in the transgenic lines X1, X2, and X3, respectively
(Fig. 4e).
Enhancement of Aldose Reductase Activity
in ALDRXV4 Overexpressing Transgenic Plants
Corresponds with Decreased Methylglyoxal
and Increased Sorbitol and Proline Content
To gain more insight into the role of ALDRXV4 in pro-
viding tolerance to drought and salt stress, ALR enzyme
activity was measured in mature leaves of WT and trans-
genic plants. The enzyme activity in the transgenic plants
was 3.5-, 3-, and 4-fold-higher (in lines X1, X2, and X3)
relative to the WT plants when grown under non-stress
condition. Under mannitol (200 mM) stress, a 2.6-, 2.3-,
and 2.8-fold increase in the ALR activity was detected in
the transgenic plants (lines X1, X2, and X3) as compared to
WT plants (Fig. 5a). Similar results were obtained under
salt stress (200 mM NaCl), where the transgenic plants
(lines X1, X2, and X3) showed 3.8-, 3.6-, and 3.9-fold
enhancement in activity when compared with the WT
plants under similar stress (Fig. 5a). The level of MG was
almost similar in WT and transgenic plants under non-
stress condition. However, in response to drought (200 mM
mannitol) and salt stress (200 mM NaCl), the WT plants
exhibited 97 and 14 % increase in MG concentration
respectively, whereas this increase was only 48 and 11 %
in transgenic line X1, 69 and 14 % in transgenic line X2,
and 29 and 3.6 % in transgenic line X3 (Fig. 5b). The
sorbitol content in the leaves of transgenic plants was
found to be 1.2-, 1.16-, and 1.2-fold higher (in lines X1,
X2, and X3) than in the WT plants under non-stress con-
ditions. Under drought and salt stress conditions, the sor-
bitol content increased in both transgenic and WT plants.
However, the increase in sorbitol content in the transgenic
lines X1, X2, and X3 as compared to the WT plants was
1.6-, 1.2-, and 1.8-fold higher under drought conditions and
1.2-, 1.1-, and 1.3-fold higher upon salt stress treatment
(Fig. 5c). The level of proline was also found to increase in
both transgenic as well as WT plants. However, the
increase in the transgenic lines X1, X2, and X3 was,
respectively, 1.9-, 1.8-, and 2-fold higher than the WT
under drought stress while it was 1.9-fold higher in X1 and
X2 and 2.1-fold higher in X3 in comparison to the WT
plants under salt stress (Fig. 5d).
Na? and K? Accumulation in Response to Salt Stress
The Na? and K? content in the leaves of WT and trans-
genic plants treated with 200 mM NaCl vs. the non-stress
Table 1 Comparison of various growth parameters and seed production of the WT and transgenic plants (lines X1, X2, and X3) grown in the
presence of water, 200 mM mannitol and 200 mM NaCl, respectively
Parameters WT X1 X2 X3
Water
Height 115 ± 4.32 116 ± 2.0 112 ± 2.51 114 ± 2.94
Foliar biomass (g)a 3.3 ± 0.49 3.5 ± 0.30 3.1 ± 0.35 3.6 ± 0.20
No. of days to flower 121 ± 4.96 119 ± 4.04 120 ± 4.5 117 ± 5.09
Seed weight per pod (mg) 160 ± 4.89 164 ± 3.21 161 ± 4.0 165 ± 5.09
200 mM Mannitol
Height No survival 105 ± 3.51 101 ± 2.51 110 ± 7.54
Foliar biomass (g)a – 2.5 ± 0.20 2.0 ± 0.30 3.1 ± 0.72
No. of days to flower – 133 ± 3.0 135 ± 3.51 132 ± 4.93
Seed weight per pod (mg) – 140 ± 4.04 132 ± 5.50 145 ± 6.55
200 mM NaCl
Height No survival 103 ± 3.0 100 ± 6.5 106 ± 3.85
Foliar biomass (g)a – 2.4 ± .30 1.9 ± .20 2.9 ± 0.29
No. of days to flower – 128 ± 2.6 125 ± 4.58 130 ± 3.74
Seed weight per pod (mg) – 135 ± 4.50 130 ± 5.68 138 ± 6.16
a The average weight of the fourth leaf from top. Each value is the mean of three replicates ±SD
Mol Biotechnol
Author's personal copy
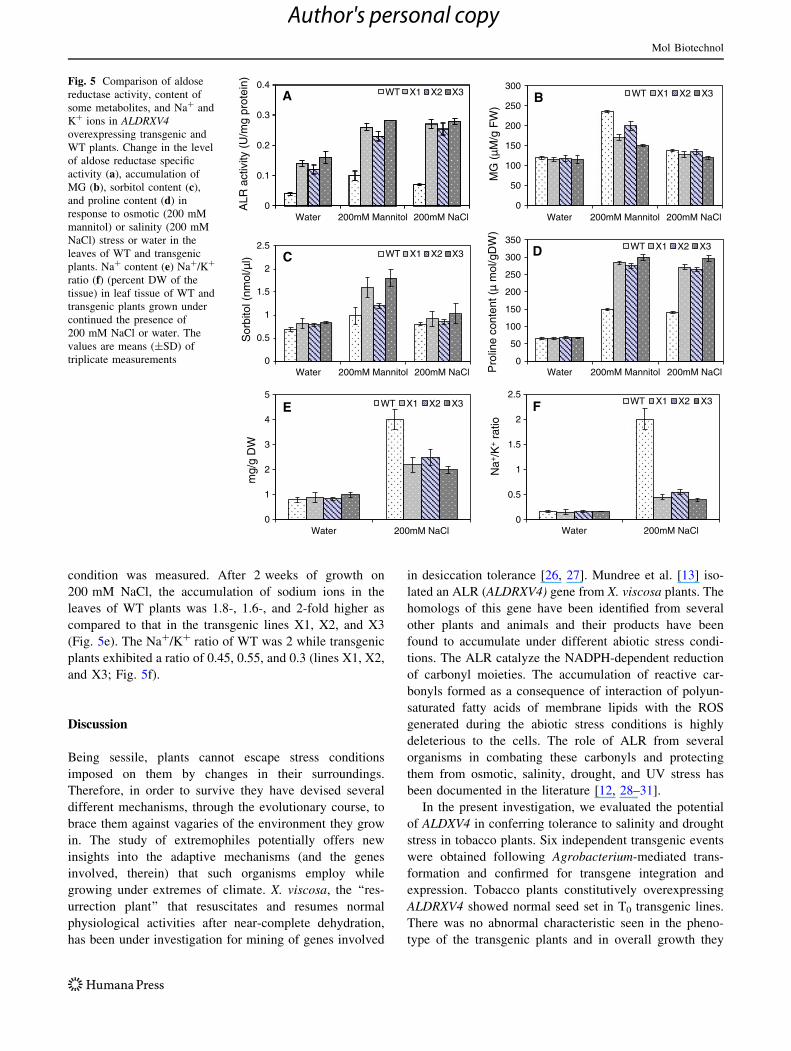
condition was measured. After 2 weeks of growth on
200 mM NaCl, the accumulation of sodium ions in the
leaves of WT plants was 1.8-, 1.6-, and 2-fold higher as
compared to that in the transgenic lines X1, X2, and X3
(Fig. 5e). The Na?/K? ratio of WT was 2 while transgenic
plants exhibited a ratio of 0.45, 0.55, and 0.3 (lines X1, X2,
and X3; Fig. 5f).
Discussion
Being sessile, plants cannot escape stress conditions
imposed on them by changes in their surroundings.
Therefore, in order to survive they have devised several
different mechanisms, through the evolutionary course, to
brace them against vagaries of the environment they grow
in. The study of extremophiles potentially offers new
insights into the adaptive mechanisms (and the genes
involved, therein) that such organisms employ while
growing under extremes of climate. X. viscosa, the ‘‘res-
urrection plant’’ that resuscitates and resumes normal
physiological activities after near-complete dehydration,
has been under investigation for mining of genes involved
in desiccation tolerance [26, 27]. Mundree et al. [13] iso-
lated an ALR (ALDRXV4) gene from X. viscosa plants. The
homologs of this gene have been identified from several
other plants and animals and their products have been
found to accumulate under different abiotic stress condi-
tions. The ALR catalyze the NADPH-dependent reduction
of carbonyl moieties. The accumulation of reactive car-
bonyls formed as a consequence of interaction of polyun-
saturated fatty acids of membrane lipids with the ROS
generated during the abiotic stress conditions is highly
deleterious to the cells. The role of ALR from several
organisms in combating these carbonyls and protecting
them from osmotic, salinity, drought, and UV stress has
been documented in the literature [12, 28–31].
In the present investigation, we evaluated the potential
of ALDXV4 in conferring tolerance to salinity and drought
stress in tobacco plants. Six independent transgenic events
were obtained following Agrobacterium-mediated trans-
formation and confirmed for transgene integration and
expression. Tobacco plants constitutively overexpressing
ALDRXV4 showed normal seed set in T0 transgenic lines.
There was no abnormal characteristic seen in the pheno-
type of the transgenic plants and in overall growth they
0
0.1
0.2
0.3
0.4
Water 200mM Mannitol 200mM NaCl
ALR
act
ivity
(U
/mg
prot
ein)
0
50
100
150
200
250
300
Water 200mM Mannitol 200mM NaCl
MG
(µM
/g F
W)
WT X1 X2 X3B
0
0.5
1
1.5
2
2.5
Water 200mM Mannitol 200mM NaCl
Sor
bito
l (nm
ol/µ
l) WT X1 X2 X3C
0
50
100
150
200
250
300
350
Water 200mM Mannitol 200mM NaClPro
line
cont
ent (
µ m
ol/g
DW
)
WT X1 X2 X3D
0
1
2
3
4
5
Water 200mM NaCl
mg/
g D
W
WT X1 X2 X3E
0
0.5
1
1.5
2
2.5
Water 200mM NaCl
Na+
/K+
ratio
WT X1 X2 X3F
A WT X1 X2 X3Fig. 5 Comparison of aldose
reductase activity, content of
some metabolites, and Na? and
K? ions in ALDRXV4overexpressing transgenic and
WT plants. Change in the level
of aldose reductase specific
activity (a), accumulation of
MG (b), sorbitol content (c),
and proline content (d) in
response to osmotic (200 mM
mannitol) or salinity (200 mM
NaCl) stress or water in the
leaves of WT and transgenic
plants. Na? content (e) Na?/K?
ratio (f) (percent DW of the
tissue) in leaf tissue of WT and
transgenic plants grown under
continued the presence of
200 mM NaCl or water. The
values are means (±SD) of
triplicate measurements
Mol Biotechnol
Author's personal copy

were similar to WT plants growing under normal condi-
tions. The T1 transgenic plants showed increased tolerance
to mannitol and salinity (NaCl) stress as assessed by seed
germination and growth of the seedlings.
The tolerance was reflected not only at the germination
level but also during subsequent plant growth. After
14 days exposure to drought and salt stress, transgenic
plants grew normally whereas the WT plants perished.
Significant tolerance against stress conditions was also
observed in the leaf disk senescence assays. The leaf disks
from transgenic plants survived better during stress treat-
ments (mannitol, NaCl, and MG) by retaining higher
chlorophyll levels when compared to the WT, which
exhibited complete senescence of leaf disks due to reduced
level of chlorophyll. The transgenic plants showed a clear
advantage in overcoming the deleterious effect brought in
by MG toxicity in a concentration-dependent manner, thus,
implicating a role of ALDRXV4 overexpression in miti-
gating stress-induced damage to the photosynthetic appa-
ratus. MG is a glycolysis-derived toxic aldehyde and is a
known substrate for aldehyde reductases in organisms
including E. coli and yeasts [12, 32, 33]. High accumula-
tion of MG inhibits cell proliferation, degrades proteins by
modifying several amino acid residues and is, thus, toxic to
the cells [3, 34–36]. Glyoxalase pathway enzymes have
also been shown to detoxify MG in transgenic plants [36].
This signifies the existence of parallel pathways to bolster
the cellular defense against unwanted metabolites.
The damage caused by increased peroxidation of lipids
in the membrane bilayers as assessed by the MDA content
was found to be less in the transgenic tobacco plants
exposed to the two different stresses in comparison to the
WT plants. A pertinent consequence of the decreased MDA
levels was a better photosynthetic performance of the
transgenic plants compared to the WT plants. Accumula-
tion of MDA beyond a threshold level has been reported to
cause irreparable damage to the photosynthetic machinery
[37]. The ratio, Fv/Fm, is a measure of the proportion of
light absorbed by chlorophyll molecules associated with
PSII in the photochemical reaction and is, thus, an indi-
cation of actual photosynthetic efficiency [38]. The ability
of aldose reductase to modulate photosynthetic efficiency
has been demonstrated in tobacco plants expressing alfalfa
aldose reductase [30]. Under simulated abiotic stress con-
ditions (salinity and water stress) transgenic plants were
less sensitive to photoinhibition as compared to WT plants.
The build-up of various osmolytes, such as proline and
betaines has been positively correlated with stress tolerance
in many studies [39]. High proline content provides pro-
tection against ROS-induced disruption of photosystems
[40]. Proline is known to contribute to membrane stability
[39, 41] and has been widely documented as a parameter
for selection for salt stress tolerance. In this study,
significantly higher proline content was observed to accu-
mulate under stress conditions in the transgenic plants as
compared to WT plants. The mechanistic relation, if any,
between the ALR and proline content in the cells would,
however, require further experimentation.
The Na?/K? ratio was found to be relatively stable in
the leaves of the transgenic plants grown either under the
normal conditions or under drought and salt stress. This
revelation is in agreement with the generally accepted
understanding that the maintenance of Na?/K? homeosta-
sis is an important aspect of salt tolerance [42].
In conclusion, transgenic tobacco plants overexpressing
ALDRXV4 under the control of constitutive CaMV 35S
promoter have been generated. Our results, unequivocally,
establish the efficacy of aldose reductase gene from
X. viscosa in conferring tolerance against salt and drought
stress through detoxification of MG and reducing mem-
brane damage. Experiments are in progress to prove that
the results obtained in this study on a model plant translate
well for the crop plants.
Acknowledgments The authors thank Prof. Jennifer A. Thomson,
University of Cape Town, South Africa for ALDRXV4 cDNA and Dr.
Sylvia de Sousa, Universidade Federal de Sao Joao del-Rei, Brasil for
aldose reductase antibodies. Dr. Mikail Pooggin’s help in training
some members of NBS’ group during their visits to FMI and Uni-
versity of Basel, Switzerland, supported by Indo-Swiss Collaboration
in Biotechnology, is duly acknowledged. Dr. S.G. Mundree is
acknowledged for discussions related to this manuscript. DK and PS
are thankful to CSIR and UGC, India, respectively, for fellowships.
MAY is a UGC-Dr. D.S. Kothari Postdoctoral Fellow. This work was
funded by DST, India (Grant No. SR/SO/BB-37/2008). The research
in the lab of NBS is funded by grants from UGC-CAS, DST-FIST,
and DST-PURSE.
References
1. Knight, H., & Knight, M. R. (2001). Abiotic stress signalling
pathways: Specificity and cross-talk. Trends in Plant Science, 6,
262–267.
2. Cushman, J. C., & Bohnert, H. J. (2000). Genomic approaches to
plant stress tolerance. Current Opinion in Plant Biology, 3,
117–124.
3. Yadav, S. K., Singla-Pareek, S. L., Ray, M., Reddy, M. K., &
Sopory, S. K. (2005). Methylglyoxal levels in plants under
salinity stress are dependent on glyoxalase I and glutathione.
Biochemical and Biophysical Research Communications, 337,
61–67.
4. Pitzschke, A., Forzani, C., & Hirt, H. (2006). Reactive oxygen
species signaling in plants. Antioxidants & Redox Signaling, 8,
1757–1764.
5. Kalapos, M. P. (2008). The tandem of free radicals and methyl-
glyoxal. Chemico-Biological Interactions, 171, 251–271.
6. Thornalley, P. J. (1990). The glyoxalase system: New develop-
ments towards functional characterization of a metabolic pathway
fundamental to biological life. Biochemical Journal, 269, 1–11.
7. Vander Jagt, D. L., & Hunsaker, L. A. (2003). Methylglyoxal
metabolism and diabetic complications: Roles of aldose reduc-
tase, glyoxalase-I, betaine aldehyde dehydro-genase and
Mol Biotechnol
Author's personal copy

2-oxoaldehyde dehydrogenase. Chemico-Biological Interactions,143, 341–351.
8. Bartels, D., & Nelson, D. (1994). Approaches to improve stress
tolerance using molecular genetics. Plant, Cell and Environment,17, 659–667.
9. Jin, Y., & Penning, T. M. (2007). Aldo–keto reductases and
bioactivation/detoxification. Annual Review of Pharmacologyand Toxicology, 47, 263–292.
10. Lee, S. P., & Chen, T. H. H. (1993). Molecular cloning of
abscisic acid-responsive mRNAs expressed during the induction
of freezing tolerance in bromegrass (Bromus inermis Leyss)
suspension culture. Plant Physiology, 101, 1089–1096.
11. Li, B., & Foley, M. E. (1995). Cloning and characterization of
differentially expressed genes in imbibed dormant and after rip-
ened Avena fatua embryos. Plant Molecular Biology, 29, 823–831.
12. Turoczy, Z., Kis, P., Torok, K., Cserhati, M., Lendvai, A., Dudits,
D., et al. (2011). Overproduction of a rice aldo–keto reductase
increases oxidative and heat stress tolerance by malondialdehyde
and methylglyoxal detoxification. Plant Molecular Biology, 75,
399–412.
13. Mundree, S. G., Whittaker, A., Thomson, J. A., & Farrant, L. M.
(2000). An aldose reductase homolog from the resurrection plant
Xerophyta viscosa Baker. Planta, 211, 693–700.
14. Hofgen, R., & Willmitzer, L. (1988). Storage of competent cells
for Agrobacterium transformation. Nucleic Acids Research, 16,
9877.
15. Horsch, R. B., Fry, J. E., Hoffmann, N. L., Eichholtz, D., Rogers,
S. G., & Fraley, R. T. (1985). A simple and general method for
transferring genes into plants. Science, 227, 1229–1231.
16. Rogers, S. O., & Bendich, A. J. (1994). Extraction of total cel-
lular DNA from plants, algae and fungi. In S. B. Gelvin &
R. A. Schilperoort (Eds.), Plant molecular biology manual, D1(pp. 1–8). Dordrecht: Kluwer Academic Press.
17. Sambrook, J., Fritsch, E. F., & Maniatis, T. (1989). Molecularcloning. A laboratory manual. New York: Cold Spring Harbor.
18. Bradford, M. (1976). A rapid and sensitive method for the quan-
titation of microgram quantities of protein utilizing the principle of
protein-dye binding. Analytical Biochemistry, 72, 248–254.
19. Towbin, H., Staehelin, T., & Gordon, J. (1979). Electrophoretic
transfer of proteins from polyacrylamide gels to nitrocellulose
sheets: Procedure and some applications. Proceedings of theNational academy of Sciences of the United States of America,76, 4350–4354.
20. Arnon, D. J. (1949). Copper enzymes in isolated chloroplasts:
Polyphenoloxidase in Beta vulgaris. Plant Physiology, 24, 1–15.
21. Sairam, R. K., & Srivastava, G. C. (2002). Changes in antioxidant
activity in subcellular fractions of tolerant and susceptible wheat
genotypes in response to long term salt stress. Plant Science, 162,
897–904.
22. Hayman, S., & Kinoshita, J. H. (1965). Isolation and properties of
lens aldose reductase. Journal of Biological Chemistry, 240,
877–882.
23. Pommerrenig, B., Papini-Terzi, F. S., & Sauer, N. (2007). Dif-
ferential regulation of sorbitol and sucrose loading into the
phloem of Plantago major in response to salt stress. PlantPhysiology, 144, 1029–1038.
24. Bates, L. S., Waldren, R. P., & Teare, I. D. (1973). Rapid
determination of free proline for water stress studies. Plant andSoil, 39, 205–207.
25. Fan, L., Zheng, S., & Wang, X. (1997). Antisense suppression ofphospholipase D alpha retards abscisic acid- and ethylene-pro-
moted senescence of postharvest Arabidopsis leaves. Plant Cell, 9,
2183–2196.
26. Sherwin, H., & Farrant, J. M. (1996). Differences in rehydration
of three desiccation-tolerant angiosperm species. Annals of Bot-any, 78, 703–710.
27. Oliver, M. J., & Bewley, J. D. (1997). Desiccation-tolerance of
plant tissues: A mechanistic overview. Horticultural Reviews, 18,
171–213.
28. Kolb, N. S., Hunsaker, L. A., & Vander Jagt, D. L. (1994). Aldose
reductase-catalyzed reduction of acrolein: Implications in cyclo-
phosphamide toxicity. Molecular Pharmacology, 45, 797–801.
29. Colrat, S., Latche, A., Guis, M., Pech, J. C., Bouzayen, M., Fallot,
J., et al. (1999). Purification and characterization of a NADPH-
dependent aldehyde reductase from mung bean that detoxifies eu-
typine, a toxin from Eutypa lata. Plant Physiology, 119, 621–626.
30. Oberschall, A., Deak, M., Torok, K., Sass, L., Vass, I., Kovacs, I.,
et al. (2000). A novel aldose/aldehyde reductase protects trans-
genic plants against lipid peroxidation under chemical and
drought stresses. Plant Journal, 24, 437–446.
31. Hideg, E., Nagy, T., Oberschall, A., Dudits, D., & Vass, I. (2003).
Detoxification function of aldose/aldehyde reductase during
drought and ultraviolet-B (280–320 nm) stresses. Plant, Cell andEnvironment, 26, 513–522.
32. Vander Jagt, D. L., Hunsaker, L. A., Vander Jagt, T. J., Gomez,
M. S., Gonzales, D. M., Deck, L. M., et al. (1997). Inactivation of
glutathione reductase by 4-hydroxynonenal and other endogenous
aldehydes. Biochemical Pharmacology, 53, 1133–1140.
33. Hegedus, A., Erdei, S., Janda, T., Toth, E., Horvath, G. V., &
Dudits, D. (2004). Transgenic tobacco plants overproducing
alfalfa aldose/aldehyde reductase show higher tolerance to low
temperature and cadmium stress. Plant Science, 166, 1329–1333.
34. Abordo, E. A., Minhas, H. S., & Thornalley, P. J. (1999).
Accumulation of alpha-oxoaldehydes during oxidative stress: A
role in cytotoxicity. Biochemical Pharmacology, 58, 641–648.
35. Martins, A. M., Cordeiro, C. A., & Ponces Freire, A. M. (2001).
In situ analysis of methylglyoxal metabolism in Saccharomycescerevisiae. FEBS Letters, 499, 41–44.
36. Singla-Pareek, S. L., Yadav, S. K., Pareek, A., Reddy, M. K., &
Sopory, S. K. (2006). Transgenic tobacco overexpressing gly-
oxalase pathway enzymes grow and set viable seeds in zinc-
spiked soils. Plant Physiology, 140, 613–623.
37. Mishra, R. K., & Singhal, G. S. (1992). Function of photosyn-
thetic apparatus of intact wheat leaves under high light and heat
stress and its relationship with peroxidation of thylakoid lipids.
Plant Physiology, 98, 1–6.
38. Harbinson, J., Genty, B., & Baker, N. R. (1989). Relationship
between the quantum efficiencies of photosystems I and II in pea
leaves. Plant Physiology, 90, 1029–1034.
39. Ashraf, M., & Harris, P. J. C. (2004). Potential biochemical
indicators of salinity tolerance in plants. Plant Science, 166,
3–16.
40. Hare, P. D., Cress, W. A., & Van Staden, J. (1998). Dissecting the
roles of osmolyte accumulation during stress. Plant, Cell andEnvironment, 21, 535–553.
41. Hanson, A. D., & Burnet, M. (1994). Evolution and metabolic
engineering of osmoprotectant accumulation in higher plants. In
J. H. Cherry (Ed.), Cell biology: Biochemical and cellularmechanism of stress tolerance in plants (pp. 291–302). Berlin:
Springer.
42. Rus, A., Yokoi, S., Sharkhuu, A., Reddy, M., Lee, B. H., Mat-
sumoto, T. K., et al. (2001). AtHKT1 is a salt tolerance deter-
minant that controls Na? entry into plant roots. Proceedings ofthe National academy of Sciences of the United States of Amer-ica, 98, 14150–14155.
Mol Biotechnol
Author's personal copy




















