
Late season harvest and storage of Rubus berries—Major antioxidant and sugar levels
Upload
independentCategory
view
4download
0
Food Research International 44 (2011) 2666–2673
Contents lists available at ScienceDirect
Food Research International
j ourna l homepage: www.e lsev ie r.com/ locate / foodres
Phytochemicals from berries and grapes inhibited the formation of advancedglycation end‐products by scavenging reactive carbonyls
Wei Wang, Yavuz Yagiz, Timothy J. Buran, Cecilia do Nascimento Nunes, Liwei Gu ⁎Department of Food Science and Human Nutrition, Institute of Food and Agricultural Sciences, University of Florida, Gainesville, Florida 32611, United States
⁎ Corresponding author. Tel.: +1 352 392 1991x210;E-mail address: [email protected] (L. Gu).
0963-9969/$ – see front matter © 2011 Elsevier Ltd. Aldoi:10.1016/j.foodres.2011.05.022
a b s t r a c t
a r t i c l e i n f oArticle history:Received 22 February 2011Accepted 16 May 2011
Keywords:BlueberriesCranberriesCarbonylsAdvanced glycation end‐products
Phenolic phytochemicals were extracted from blueberries, blackberries, strawberries, raspberries, cran-berries, and Noble muscadine grapes. These extracts were purified to remove free sugars. Blueberry extractwas separated into five fractions using a Sephadex LH‐20 column. Berry extracts and fractions significantlyinhibited AGEs generation in (bovine serum albumin) BSA‐fructose, BSA‐methylglyoxal, and arginine‐methylglyoxal models, respectively. Their capacity to scavenge methylglyoxal suggested carbonyl scavengingas a major mechanism of protein glycation inhibition. Procyanidins were detected in all berry extracts andblueberry subfractions and were deduced to be one class of active compounds. (+)‐Catechin, constituent unitof procyanidins, was used as a model compound to react with glyoxal and methylglyoxal. Five catechin‐carbonyl adducts were detected and their structures were tentatively identified using HPLC‐ESI‐MSn. Resultsin this study suggested that sugar‐free phytochemicals extracted from berries were effective carbonylscavengers and protein glycation inhibitors. These phytochemicals could be beneficial to prevent AGE‐relatedchronic diseases.
fax: +1 352 392 9467.
l rights reserved.
© 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Reactive carbonyls are compounds that contain two or morecarbonyl groups. The most common reactive carbonyls aredicarbonyls such as glyoxal and methylglyoxal. Reactive carbonylsreact with proteins to generate advance glycation end products(AGEs). Accumulation of AGEs is a causing factor for diabeticcomplications, aging, and several types of cancers (Ahmed, 2005;Münch, Thome, Foley, Schinzel, & Riederer, 1997). A well-knownAGE in human body is the glycated hemoglobin (HbA1c). Its levelin blood is used to evaluate long term glucose control and risks forcomplications in diabetic patients (American Diabetes Association,2009).
Reactive carbonyls are generated both in foods and the humanbody. Methylglyoxal and glyoxal are found in foods that containlarge amounts of carbohydrates or in foods that are processed athigh temperatures (Nemet, Varga-Defterdarovic, & Turk, 2006).Typical examples include baked meats, soy sauces, and carbonatedsoft drinks (Tan, Wang, Lo, Sang, & Ho, 2008). In the human body,methylglyoxal and glyoxal are generated from carbohydrate metab-olism or auto-oxidation, as well as the oxidation of protein glycationproducts (Hollnagel & Kroh, 1998; O'Brien, Siraki, & Shangari, 2005).
Protein glycation, also known as the Maillard reaction whenoccurring in vitro, is a non-enzymatic reaction between reducingsugars and proteins. During such a reaction, reducing sugars, such asglucose, react with amino groups of proteins to produce Schiff bases.Schiff bases undergo Amadori rearrangement to produce relativelystable compounds called Amadori products. Under low pH oroxidative conditions, Schiff bases or Amadori products degrade togenerate reactive carbonyls including methylglyoxal (Cho, Roman,Yeboah, & Konishi, 2007; O'Brien et al., 2005; Oya et al., 1999).Methylglyoxal is a major reactive carbonyl in vivo. Its concentrationin the blood of diabetic patients was five folds of that in healthyindividuals (Nagaraj et al., 2002). Methylglyoxal glycates proteins ina much faster rate than sugars, causing inter- and intramoleculecross-links of proteins.
Several types of phytochemicals have been suggested to beeffective in scavenging carbonyls and inhibiting the formation ofAGEs. For example, flavan-3-ols from green tea were found toscavenge toxic methylglyoxal under simulated physiological condi-tions (Lo et al., 2006). Procyanidins from cinnamon were shown toinhibit protein glycation (Peng et al., 2008). Pure resveratrolscavenged carbonyls by trapping them and form adducts (Lv et al.,2010). Berries and grapes are rich sources of phenolic nutraceuticals,such as catechins, resveratrol, and anthocyanins (Lo et al., 2006; Lv etal., 2010; Morata, Calderón, González, Gómez-Cordovés, & Suárez,2007; Sang et al., 2007); however, it is not known whetherphytochemicals in berries can scavenge carbonyls and inhibit theformation of AGEs. The objective of present study is to evaluate sucheffect.

2667W. Wang et al. / Food Research International 44 (2011) 2666–2673
2. Materials and methods
2.1. Chemicals and materials
Southern high-bush Blueberries, strawberries, cranberries, rasp-berries, blackberries, and Noble muscadine grapes were purchasedfrom local supermarkets or obtained from local farms in centralFlorida. Only fully ripe fruits were used for experiments. AAPH(2,2′-azotis(2-amidinopropane)) was a product of Wako ChemicalsInc. (Bellwood, RI). Sephadex LH-20, (+)-catechin, and Trolox(6-Hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid) werepurchased from Sigma-Aldrich (St. Louis, MO). Amberlite XAD-7 resinwas a product of Rohm and Haas Co. (Philadelphia, PA). Methylglyoxal(40% solution), glyoxal (40% solution), aminoguanidine, HPLC grademethanol, and other chemicals were purchased from Fisher ScientificCo. (Pittsburg, PA).
2.2. Phytochemical extraction and purification
Frozen berries or muscadine grapes were thawed under roomtemperature. Fruits (200 g) were blended with methanol (200 mlwith 1% formic acid) in a kitchen blender. The mixture was sonicatedin a water-bath sonicater (FS30, Fisher Scientific) for 10 min, thenkept in room temperature for 20 min and filtered through aWhatmanNo.4 filter paper. The extraction was repeated once on the remainingsolid and the methanol extracts were combined. Extracts were driedin a SpeedVac concentrator (Thermo scientific ISS110, Waltham, MA)under ambient temperature.
Dried berry crude extract was re-suspended in 20 ml distilledwater containing 1% formic acid and loaded onto a glass column(2.8×55 cm, 500 ml) packed with Amberlite XAD-7 resin. Aftersample loading, elution was halted for 10 min to facilitate theadsorption of phytochemicals on resin beads. Resin column waseluted with 600 ml of acidified water (containing 1% formic acid) toremove sugars. Phytochemicals absorbed on resin were recoveredwith 250 ml of 80% methanol (with 1% formic acid). Eluents from theresin column were dried on a SpeedVac concentrator. About 0.84,0.73, 1.045, 0.94, 0.50 and 1.32 g of dry extract were obtained fromblueberries, strawberries, cranberries, blackberries, raspberries, andNoble muscadine grapes, respectively.
About 0.8 g of dry blueberry extract obtained after resin absorptionwas homogenized in 10 mL water and loaded into a column(2.8×55 cm, 500 ml) packed with Sephadex LH-20. Five fractionswere collected by eluting the column with 500 ml water, 700 ml of20% methanol, 700 ml of 50% methanol, 700 ml of 70% methanol, and500 ml of 80% acetone. All eluting solvents were acidified with 1%formic acid to stabilize anthocyanins. Solvents in all the fractions wereevaporated on a SpeedVac concentrator and the remaining solidswere weighed. About 0.27, 0.13, 0.08, 0.17 and 0.10 g of dry extractwere obtained for fraction I to fraction V, respectively.
2.3. Chemical analyses of extracts and fractions
The total phenolic contents of berry and extracts were determinedwith Folin-Ciocalteu assay using gallic acid as a standard (Singleton &Rossi, 1965). Total antioxidant capacity was measured with oxygenradical absorbance capacity (ORAC) assay using trolox as a standard(Huang, Ou, Hampsch-Woodill, Flanagan, & Prior, 2002). Totalanthocyanin content was determined with a pH differential assayusing molar absorptivity of cyanidin 3-glucoside of 26,900 (Giusti &Wrolstad, 2001). Total procyanidin content was analyzed with 4-dimethylaminocinnamaldehyde (DMAC) colorimetric method using(+)-catechin as a standard (Prior et al., 2010). An analysis of freesugars was conducted using a Hitachi HPLC system with a refractiveindex detector and a Shodex SP0810 column (300 mm×8 mm, Shodex,Colorado springs, CO) with a SP-G guard column (2 mm×4 mm). An
isocratic solvent delivery of waterwas run at 1.0 mL/min. Purified berryphytochemicals extracts were dissolved in distilled water to aconcentration of 15 mg/ml. Sample injection volume was 5 μL. Sucrose,glucose, fructose, lactose and maltodextrin were used as standards.
2.4. Antiglycation assay in BSA-fructose model
This model evaluates all stages of protein glycation. Fructose(1.5 M, 1 ml) was mixed with berry extracts obtain from Amberliteresin column (0.15 mg/ml, 1 ml) or catechin (0.15 mg/ml, 1 ml) insodium phosphate buffer (50 mM, pH 7.4, with 0.02% sodium azide)in capped test tubes and kept at 37 °C for 2 h. BSA (30 mg/ml, 1 ml)was added in each test tube and the mixtures were incubated at 37 °Cfor six days. Phosphate buffer (1 ml)with no berry extract was used asa blank control. Aminoguanidine (10 mM, final concentration) wasused as a positive control. Fluorescent AGEs were monitored on amicroplate reader (Spectra XMS Gemini, Molecular Device, Sunny-vale, CA) using 340 and 420 nm as the excitation and emissionwavelengths. Experiments were conducted in triplicates. Percentageof the AGE inhibition was calculated by the following equation:
Percentage inhibition = 1− Fluorescent intensity with inhibitorFluorescent intensity without inhibitor
� �× 100%
2.5. Antiglycation assay in BSA-methylglyoxal model
This model evaluates the middle stage of protein glycation.Methylglyoxal (60 mM, 1 ml) was mixed with berry extracts(1.5 mg/ml, 1 ml), catechin (1.5 mg/ml, 1 ml) in sodium phosphatebuffer (50 mM, pH 7.4, with 0.02% sodium azide) and kept at 37 °C for2 h. BSA (30 mg/ml, 1 ml) was added to each test tube and incubatedat 37 °C for six days. Phosphate buffer (1 ml) with no berry extractwas used as a blank control. Aminoguanidine (10 mM, finalconcentration) was used as a positive control. Fluorescent AGEswere monitored on a microplate reader using 340 and 380 nm as theexcitation and emission wavelengths. Experiments were conducted intriplicates. The percentage of the AGE inhibition was calculated usingthe same equation in BSA-fructose model.
2.6. Antiglycation assay in arginine–methylglyoxal model
This model evaluates a major and specific source of AGEproduction. Methylglyoxal (60 mM, 1 ml) was mixed berry extracts(1.5 mg/ml, 1 ml) or catechin (1.5 mg/ml, 1 ml) in sodium phosphatebuffer (50 mM, pH 7.4, with 0.02% sodium azide) and kept at 37 °C for2 h. Arginine (60 mM, 1 ml) was added to each test tube andincubated at 37 °C for six days. Phosphate buffer (1 ml) with noberry extract was used as a blank control. Aminoguanidine (10 mM,final concentration) was used as a positive control. Fluorescent AGEswere monitored on a microplate reader using 340 and 380 nm as theexcitation and emission wavelengths. Experiments were conducted intriplicates. Percentage of the AGE inhibition was calculated using thesame equation as in the BSA-fructose model.
2.7. Methylglyoxal scavenging
Methylglyoxal scavenging was tested using a published methodwith minor modifications (Peng et al., 2008). Methylglyoxal and o-phenylenediamine were dissolved in phosphate buffer (50 mM, pH7.4) to a concentration of 10 mM and 50 mM, respectively. Berry andgrape extracts were dispersed in phosphate buffer to 2.5 mg/ml.Aminoguanidine (10 mM) was used as a positive control. Methyl-glyoxal solution (0.125 ml) was mixed with 0.125 ml of phosphatebuffer (negative control) or berry extracts. The tubes were incubatedat 37 °C. O-phenylenediamine (0.25 ml) was added into each tube at

Table 1Total phenolics, antioxidant capacities, total anthocyanins and total procyanidins of berry extracts and blueberry fractions.
Samples Total phenolics(mg gallic acid equivalent/mg)
Antioxidant capacity(μmol trolox equivalent/mg)
Total anthocyanins(μg cyanidin-3-glucoside equivalent/mg)
Total procyanidins(μg catechin equivalent/mg)
Berry extractsBlueberries 0.54±0.01a 4.9±0.0e 153.5±3.2b 55.2±2.0c
Strawberries 0.40±0.02bc 5.1±0.1d 43.2±0.0d 60.5±1.9c
Cranberries 0.42±0.02bc 5.6±0.0c 51.9±0.2c 75.9±1.4b
Raspberries 0.38±0.01c 5.2±0.1d 20.9±0.2e 12.6±0.45e
Blackberries 0.44±0.01b 6.0±0.1b 163.5±2.2a 36.3±2.4d
Noble grapes 0.57±0.03a 6.9±0.1a 23.6±0.1e 131.8±5.2a
Blueberry fractionsBlueberries 0.54±0.01bc 4.9±0.0c 153.5±3.2b 55.2±2.0a
I 0.42±0.01d 6.0±0.5bc 367.8±4.8a 0.97±0.11c
II 0.57±0.01b 8.2±1.0a 95.1±4.2c 4.1±0.40c
III 0.53±0.01c 6.4±0.5b 68.2±2.2d 38.4±6.6b
IV 0.63±0.02a 8.3±0.4a 46.4±0.7e 31.4±2.7b
V 0.31±0.01e 2.5±0.3d 14.8±0.4f 56.9±3.6a
Values are mean±standard deviation of triplicate assays. Values of the same group in the same column with different letters denoted significantly differences (P≤0.05) from eachother.
2668 W. Wang et al. / Food Research International 44 (2011) 2666–2673
0.5 h, 1 h, 2 h, 4 h and 6 h. The tubes were kept for 30 min for thederivatization reaction between methylglyoxal and o-phenylenedia-mine to complete. HPLC analysis of incubation media was performedon an Agilent 1200 HPLC system (Agilent technologies, Palo Alto, CA).Compound separation was carried out in a Zorbax SB-C18 column(4.6×250 mm, 5 μm, Agilent Technologies, Palo Alto, CA). Mobilephases were composed of 0.1% formic acid in millipore water (mobilephase A) and pure methanol (mobile phase B). The flow rate was1 ml/min and the injection volume was 15 μl. The linear gradient forelution was: 0–3 min, 5–50% B; 3–16 min, 50–50% B; 16–17 min,
Fig. 1. Inhibitory effects of berry extracts (A) and blueberry fractions (B) on AGEformation in BSA-fructosemodel at 37 °C for 6 days. Final concentration of berry extractand catechin was 0.05 mg/mL. Aminoguanidine (10 mM) served as a positive control.Bars with different letters denoted significant differences.
50–90% B; 17–19 min, 90–90% B; 19–19.5 min, 90–5% B; followed by1 min of re-equilibration of the column. Methylglyoxal-o-phenylene-diamine adduct, 1-methylquinoxaline, was detected at 315 nm usinga diode array detector. Its retention time was 12.9 min. The peakarea of 1-methylquinoxaline in each sample was integrated. Thepercentage of remaining methylglyoxal was calculated using thefollowing equation:
Remaining percentage =Peak area at different incubation time
Peak area at time zero× 100%
Fig. 2. Inhibitory effects of berry extracts (A) and blueberry fractions (B) on AGEformation in BSA-methylglyoxal model at 37 °C for 6 days. Final concentration of berryextract and catechin was 0.5 mg/mL. Aminoguanidine (10 mM) served as a positivecontrol. Bars with different letters denoted significant differences.
2669W. Wang et al. / Food Research International 44 (2011) 2666–2673
2.8. HPLC-MSn identification of catechin-carbonyl adducts
(+)-Catechin (10 mM, 1 mL) was mixed with glyoxal (10 mM,1 mL), methylglyoxal (10 mM, 1 mL), or phosphate buffer (50 mM,pH 7.4, 1 ml, control). All chemicals were freshly prepared anddissolved in phosphate buffer (50 mM, pH 7.4). After the mixtureswere incubated at 37 °C for six hours, 200 μl of media was mixed witho-phenylenediamine (50 mM, 200 μl) to terminate the catechin–carbonyl reaction.
Detection of catechin and catechin–carbonyl adductswas performedon an Agilent 1200 HPLC system (Agilent Technologies, Palo Alto, CA)equippedwith a diode array detector, a fluorescence detector and aHCTion trap mass spectrometer (Bruker Daltonics, Billerica, MA). A ZorbaxSB-C18 column (4.6×250 mm, 5 μm, Agilent Technologies, Palo Alto,CA) was used for compound separation. Mobile phases were composedof 0.1% formic acid in Millipore water (mobile phase A) and puremethanol (mobile phase B). The linear gradientwas: 0–5 min, 10–22.5%B; 5–15 min, 22.5–27.5% B; 15–18 min, 27.5–70% B; 18–21min, 70–90%B; 21–23 min, 90–10% B, followed by 2 min of re-equilibration. Theinjection volume was 5 μl and flow rate was 1 ml/min. Fluorescentdetector was set to use 231 nmand 320 nm for excitation and emission,respectively. Amass spectrometerwithelectrospray ionization interfacewas operated at both positive and negative modes using nebulizer50 psi, drying gas 10 L/min and a drying temperature of 300 °C.Retention time for catechin was 12.4 min on a fluorescent chromato-gram. Mass spectra were scanned from 50 to 800 m/z. The mostabundant ions in the full scan spectrumwere isolated and their production spectra (MS2) were recorded.
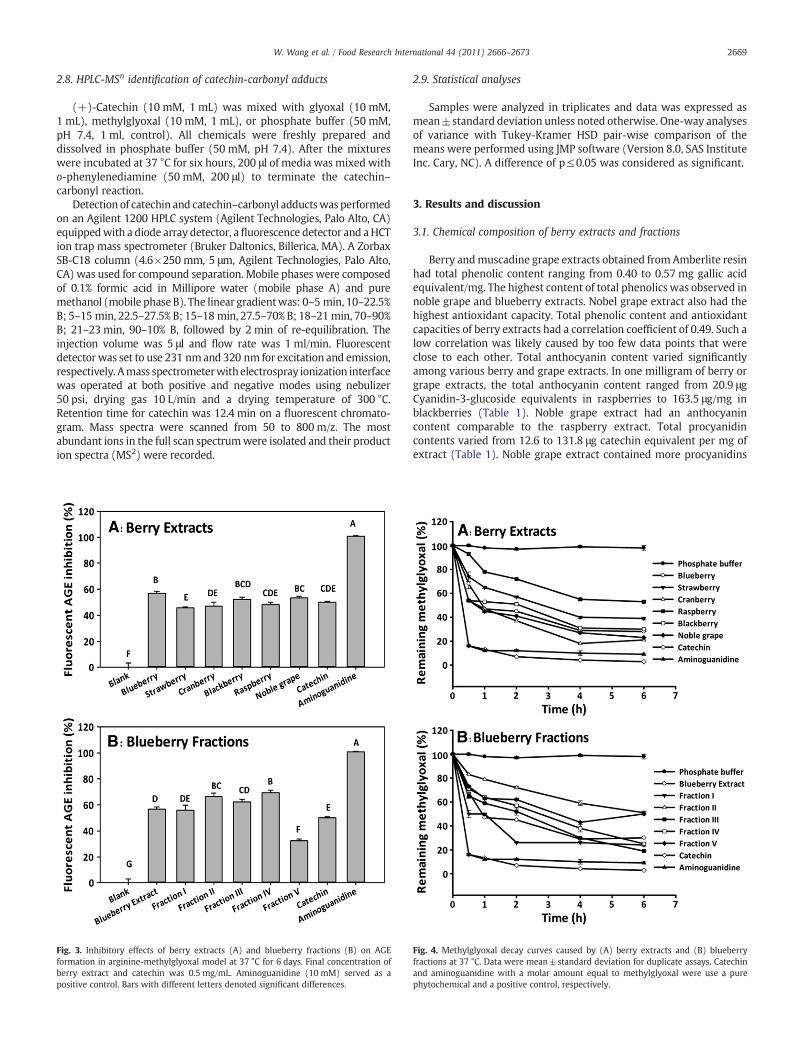
Fig. 3. Inhibitory effects of berry extracts (A) and blueberry fractions (B) on AGEformation in arginine-methylglyoxal model at 37 °C for 6 days. Final concentration ofberry extract and catechin was 0.5 mg/mL. Aminoguanidine (10 mM) served as apositive control. Bars with different letters denoted significant differences.
2.9. Statistical analyses
Samples were analyzed in triplicates and data was expressed asmean±standard deviation unless noted otherwise. One-way analysesof variance with Tukey-Kramer HSD pair-wise comparison of themeans were performed using JMP software (Version 8.0, SAS InstituteInc. Cary, NC). A difference of p≤0.05 was considered as significant.
3. Results and discussion
3.1. Chemical composition of berry extracts and fractions
Berry andmuscadine grape extracts obtained from Amberlite resinhad total phenolic content ranging from 0.40 to 0.57 mg gallic acidequivalent/mg. The highest content of total phenolics was observed innoble grape and blueberry extracts. Nobel grape extract also had thehighest antioxidant capacity. Total phenolic content and antioxidantcapacities of berry extracts had a correlation coefficient of 0.49. Such alow correlation was likely caused by too few data points that wereclose to each other. Total anthocyanin content varied significantlyamong various berry and grape extracts. In one milligram of berry orgrape extracts, the total anthocyanin content ranged from 20.9 μgCyanidin-3-glucoside equivalents in raspberries to 163.5 μg/mg inblackberries (Table 1). Noble grape extract had an anthocyanincontent comparable to the raspberry extract. Total procyanidincontents varied from 12.6 to 131.8 μg catechin equivalent per mg ofextract (Table 1). Noble grape extract contained more procyanidins
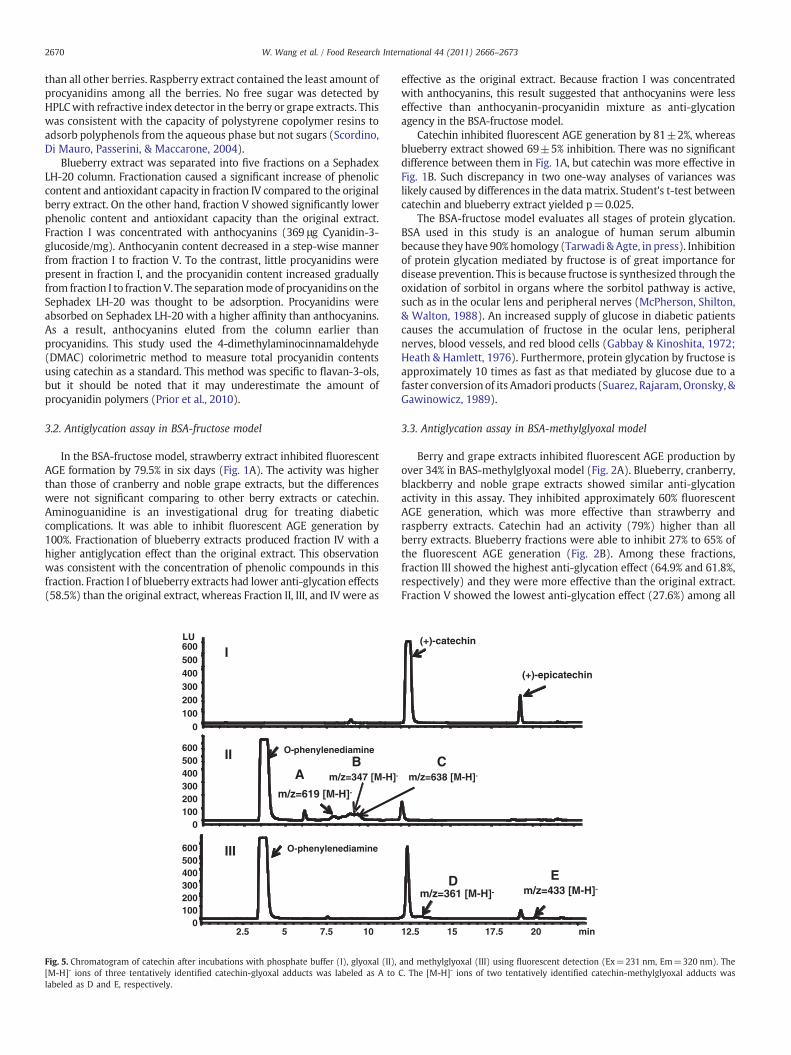
Fig. 4. Methylglyoxal decay curves caused by (A) berry extracts and (B) blueberryfractions at 37 °C. Data were mean±standard deviation for duplicate assays. Catechinand aminoguanidine with a molar amount equal to methylglyoxal were use a purephytochemical and a positive control, respectively.
2670 W. Wang et al. / Food Research International 44 (2011) 2666–2673
than all other berries. Raspberry extract contained the least amount ofprocyanidins among all the berries. No free sugar was detected byHPLCwith refractive index detector in the berry or grape extracts. Thiswas consistent with the capacity of polystyrene copolymer resins toadsorb polyphenols from the aqueous phase but not sugars (Scordino,Di Mauro, Passerini, & Maccarone, 2004).
Blueberry extract was separated into five fractions on a SephadexLH-20 column. Fractionation caused a significant increase of phenoliccontent and antioxidant capacity in fraction IV compared to the originalberry extract. On the other hand, fraction V showed significantly lowerphenolic content and antioxidant capacity than the original extract.Fraction I was concentrated with anthocyanins (369 μg Cyanidin-3-glucoside/mg). Anthocyanin content decreased in a step-wise mannerfrom fraction I to fraction V. To the contrast, little procyanidins werepresent in fraction I, and the procyanidin content increased graduallyfrom fraction I to fractionV. The separationmode of procyanidins on theSephadex LH-20 was thought to be adsorption. Procyanidins wereabsorbed on Sephadex LH-20 with a higher affinity than anthocyanins.As a result, anthocyanins eluted from the column earlier thanprocyanidins. This study used the 4-dimethylaminocinnamaldehyde(DMAC) colorimetric method to measure total procyanidin contentsusing catechin as a standard. This method was specific to flavan-3-ols,but it should be noted that it may underestimate the amount ofprocyanidin polymers (Prior et al., 2010).
3.2. Antiglycation assay in BSA-fructose model
In the BSA-fructose model, strawberry extract inhibited fluorescentAGE formation by 79.5% in six days (Fig. 1A). The activity was higherthan those of cranberry and noble grape extracts, but the differenceswere not significant comparing to other berry extracts or catechin.Aminoguanidine is an investigational drug for treating diabeticcomplications. It was able to inhibit fluorescent AGE generation by100%. Fractionation of blueberry extracts produced fraction IV with ahigher antiglycation effect than the original extract. This observationwas consistent with the concentration of phenolic compounds in thisfraction. Fraction I of blueberry extracts had lower anti-glycation effects(58.5%) than the original extract, whereas Fraction II, III, and IV were as
LU
0100200300400500600
0100200300400500600 O-phenylenediamine
m/z=619 [M-H]-
m/z=347 [M-H]-
I
II
2.5 5 7.5 100
100200300400500600 O-phenylenediamineIII
AB
Fig. 5. Chromatogram of catechin after incubations with phosphate buffer (I), glyoxal (II),[M-H]- ions of three tentatively identified catechin-glyoxal adducts was labeled as A tolabeled as D and E, respectively.
effective as the original extract. Because fraction I was concentratedwith anthocyanins, this result suggested that anthocyanins were lesseffective than anthocyanin-procyanidin mixture as anti-glycationagency in the BSA-fructose model.
Catechin inhibited fluorescent AGE generation by 81±2%, whereasblueberry extract showed 69±5% inhibition. There was no significantdifference between them in Fig. 1A, but catechin was more effective inFig. 1B. Such discrepancy in two one-way analyses of variances waslikely caused by differences in the data matrix. Student's t-test betweencatechin and blueberry extract yielded p=0.025.
The BSA-fructose model evaluates all stages of protein glycation.BSA used in this study is an analogue of human serum albuminbecause they have 90%homology (Tarwadi&Agte, in press). Inhibitionof protein glycation mediated by fructose is of great importance fordisease prevention. This is because fructose is synthesized through theoxidation of sorbitol in organs where the sorbitol pathway is active,such as in the ocular lens and peripheral nerves (McPherson, Shilton,& Walton, 1988). An increased supply of glucose in diabetic patientscauses the accumulation of fructose in the ocular lens, peripheralnerves, blood vessels, and red blood cells (Gabbay & Kinoshita, 1972;Heath & Hamlett, 1976). Furthermore, protein glycation by fructose isapproximately 10 times as fast as that mediated by glucose due to afaster conversion of its Amadori products (Suarez, Rajaram, Oronsky, &Gawinowicz, 1989).
3.3. Antiglycation assay in BSA-methylglyoxal model
Berry and grape extracts inhibited fluorescent AGE production byover 34% in BAS-methylglyoxal model (Fig. 2A). Blueberry, cranberry,blackberry and noble grape extracts showed similar anti-glycationactivity in this assay. They inhibited approximately 60% fluorescentAGE generation, which was more effective than strawberry andraspberry extracts. Catechin had an activity (79%) higher than allberry extracts. Blueberry fractions were able to inhibit 27% to 65% ofthe fluorescent AGE generation (Fig. 2B). Among these fractions,fraction III showed the highest anti-glycation effect (64.9% and 61.8%,respectively) and they were more effective than the original extract.Fraction V showed the lowest anti-glycation effect (27.6%) among all
(+)-epicatechin
m/z=638 [M-H]-
min12.5 15 17.5 20
m/z=361 [M-H]- m/z=433 [M-H]-
C
D E
(+)-catechin
and methylglyoxal (III) using fluorescent detection (Ex=231 nm, Em=320 nm). TheC. The [M-H]- ions of two tentatively identified catechin-methylglyoxal adducts was
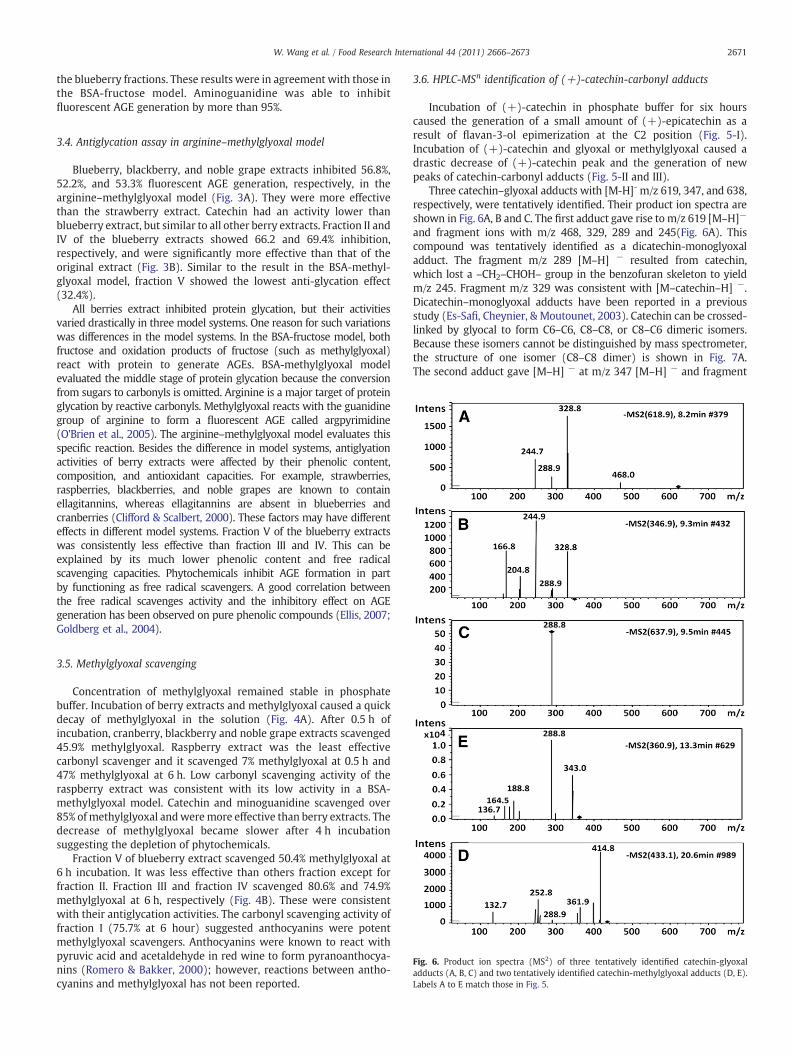
Fig. 6. Product ion spectra (MS2) of three tentatively identified catechin-glyoxaladducts (A, B, C) and two tentatively identified catechin-methylglyoxal adducts (D, E).Labels A to E match those in Fig. 5.
2671W. Wang et al. / Food Research International 44 (2011) 2666–2673
the blueberry fractions. These results were in agreement with those inthe BSA-fructose model. Aminoguanidine was able to inhibitfluorescent AGE generation by more than 95%.
3.4. Antiglycation assay in arginine–methylglyoxal model
Blueberry, blackberry, and noble grape extracts inhibited 56.8%,52.2%, and 53.3% fluorescent AGE generation, respectively, in thearginine–methylglyoxal model (Fig. 3A). They were more effectivethan the strawberry extract. Catechin had an activity lower thanblueberry extract, but similar to all other berry extracts. Fraction II andIV of the blueberry extracts showed 66.2 and 69.4% inhibition,respectively, and were significantly more effective than that of theoriginal extract (Fig. 3B). Similar to the result in the BSA-methyl-glyoxal model, fraction V showed the lowest anti-glycation effect(32.4%).
All berries extract inhibited protein glycation, but their activitiesvaried drastically in three model systems. One reason for such variationswas differences in the model systems. In the BSA-fructose model, bothfructose and oxidation products of fructose (such as methylglyoxal)react with protein to generate AGEs. BSA-methylglyoxal modelevaluated the middle stage of protein glycation because the conversionfrom sugars to carbonyls is omitted. Arginine is a major target of proteinglycation by reactive carbonyls. Methylglyoxal reacts with the guanidinegroup of arginine to form a fluorescent AGE called argpyrimidine(O'Brien et al., 2005). The arginine–methylglyoxal model evaluates thisspecific reaction. Besides the difference in model systems, antiglyationactivities of berry extracts were affected by their phenolic content,composition, and antioxidant capacities. For example, strawberries,raspberries, blackberries, and noble grapes are known to containellagitannins, whereas ellagitannins are absent in blueberries andcranberries (Clifford & Scalbert, 2000). These factors may have differenteffects in different model systems. Fraction V of the blueberry extractswas consistently less effective than fraction III and IV. This can beexplained by its much lower phenolic content and free radicalscavenging capacities. Phytochemicals inhibit AGE formation in partby functioning as free radical scavengers. A good correlation betweenthe free radical scavenges activity and the inhibitory effect on AGEgeneration has been observed on pure phenolic compounds (Ellis, 2007;Goldberg et al., 2004).
3.5. Methylglyoxal scavenging
Concentration of methylglyoxal remained stable in phosphatebuffer. Incubation of berry extracts and methylglyoxal caused a quickdecay of methylglyoxal in the solution (Fig. 4A). After 0.5 h ofincubation, cranberry, blackberry and noble grape extracts scavenged45.9% methylglyoxal. Raspberry extract was the least effectivecarbonyl scavenger and it scavenged 7% methylglyoxal at 0.5 h and47% methylglyoxal at 6 h. Low carbonyl scavenging activity of theraspberry extract was consistent with its low activity in a BSA-methylglyoxal model. Catechin and minoguanidine scavenged over85% ofmethylglyoxal andweremore effective than berry extracts. Thedecrease of methylglyoxal became slower after 4 h incubationsuggesting the depletion of phytochemicals.
Fraction V of blueberry extract scavenged 50.4% methylglyoxal at6 h incubation. It was less effective than others fraction except forfraction II. Fraction III and fraction IV scavenged 80.6% and 74.9%methylglyoxal at 6 h, respectively (Fig. 4B). These were consistentwith their antiglycation activities. The carbonyl scavenging activity offraction I (75.7% at 6 hour) suggested anthocyanins were potentmethylglyoxal scavengers. Anthocyanins were known to react withpyruvic acid and acetaldehyde in red wine to form pyranoanthocya-nins (Romero & Bakker, 2000); however, reactions between antho-cyanins and methylglyoxal has not been reported.
3.6. HPLC-MSn identification of (+)-catechin-carbonyl adducts
Incubation of (+)-catechin in phosphate buffer for six hourscaused the generation of a small amount of (+)-epicatechin as aresult of flavan-3-ol epimerization at the C2 position (Fig. 5-I).Incubation of (+)-catechin and glyoxal or methylglyoxal caused adrastic decrease of (+)-catechin peak and the generation of newpeaks of catechin-carbonyl adducts (Fig. 5-II and III).
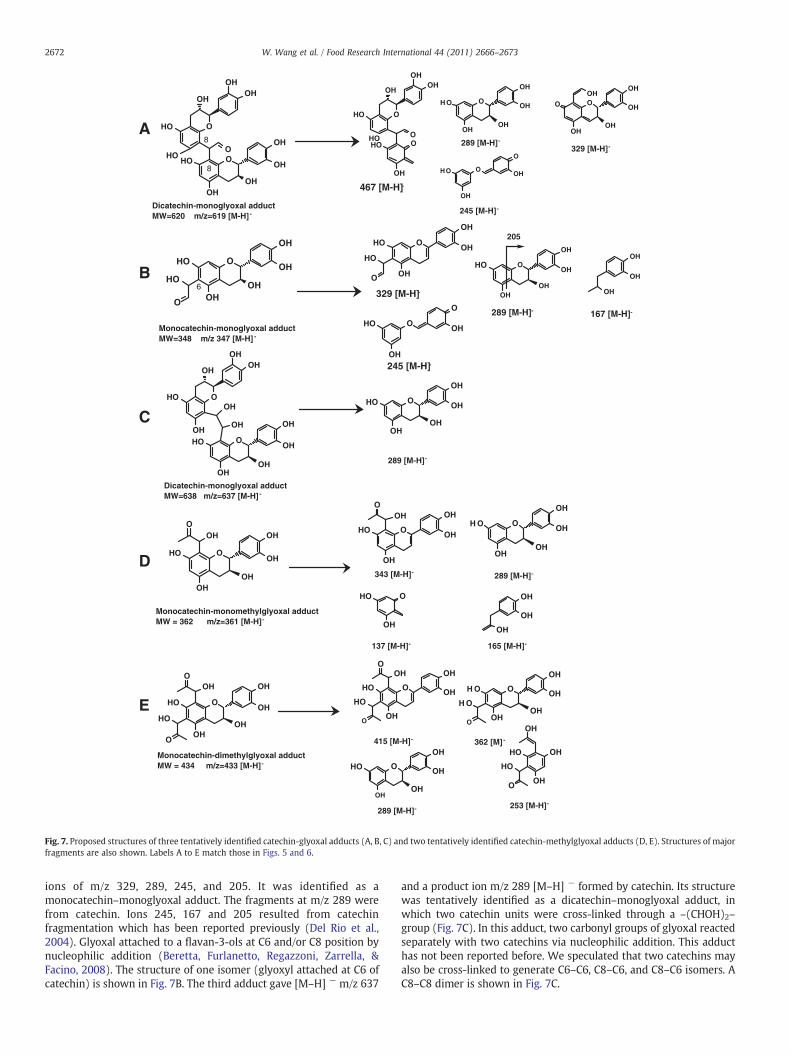
Three catechin–glyoxal adducts with [M-H]- m/z 619, 347, and 638,respectively, were tentatively identified. Their product ion spectra areshown in Fig. 6A, B and C. The first adduct gave rise to m/z 619 [M–H]−
and fragment ions with m/z 468, 329, 289 and 245(Fig. 6A). Thiscompound was tentatively identified as a dicatechin-monoglyoxaladduct. The fragment m/z 289 [M–H] − resulted from catechin,which lost a –CH2–CHOH– group in the benzofuran skeleton to yieldm/z 245. Fragment m/z 329 was consistent with [M–catechin–H] −.Dicatechin–monoglyoxal adducts have been reported in a previousstudy (Es-Safi, Cheynier, & Moutounet, 2003). Catechin can be crossed-linked by glyocal to form C6–C6, C8–C8, or C8–C6 dimeric isomers.Because these isomers cannot be distinguished by mass spectrometer,the structure of one isomer (C8–C8 dimer) is shown in Fig. 7A.The second adduct gave [M–H] − at m/z 347 [M–H] − and fragment
8
Monocatechin-dimethylglyoxal adductMW = 434 m/z=433 [M-H]-
8
6
415 [M-H]- 362 [M]-
289 [M-H]- 253 [M-H]-
Monocatechin-monomethylglyoxal adductMW = 362 m/z=361 [M-H]-
289 [M-H]-
Dicatechin-monoglyoxal adductMW=620 m/z=619 [M-H]-
343 [M-H]-
467 [M-H]-
137 [M-H]-
329 [M-H]-
Monocatechin-monoglyoxal adductMW=348 m/z 347 [M-H] -
329 [M-H]-
289 [M-H]-
165 [M-H]-
167 [M-H]-
245 [M-H]-
205
Dicatechin-monoglyoxal adductMW=638 m/z=637 [M-H] -
289 [M-H]-
289 [M-H]-
245 [M-H]-
OH
O
OH
OH
OH
O
OH
OHOH
HO
HOO
HO
OH
O
O
OH
OHOH
HO
HO OHO
OH
O
OH
OH
OH
OH
OH
O
O
OHOH
OH
O
OH
OH
OH
OH
O
O
OH
OH
OH
OH
HO
O
HO
O
OH
OH
OH
HO
O
HO
O
OH
OH
OH
OH
HO
O
O
OH
OH
HO
OH
OH
OH
HO O
OHOH
OH
HO
OH
O
OH
OH
OH
OH
OH
OH
HO
OH
O
OH
OH
OH
OH
OH
OH
HO O
OH
HO
OHO
O
OH
OH
OH
OH O
OH
OH
OHOH
OH
HO
OHOH
OH
OHO
O
HO
OH
OH
OHO
HOHO O
OH
OH
OHOH
OH O
OH
OH
OHOH
O
OH
HO
OOH
O
OH
OH
OHO
HOHO
OOH
O
OH
OH
OHOH
O
HO
A
B
C
D
E
Fig. 7. Proposed structures of three tentatively identified catechin-glyoxal adducts (A, B, C) and two tentatively identified catechin-methylglyoxal adducts (D, E). Structures of majorfragments are also shown. Labels A to E match those in Figs. 5 and 6.
2672 W. Wang et al. / Food Research International 44 (2011) 2666–2673
ions of m/z 329, 289, 245, and 205. It was identified as amonocatechin–monoglyoxal adduct. The fragments at m/z 289 werefrom catechin. Ions 245, 167 and 205 resulted from catechinfragmentation which has been reported previously (Del Rio et al.,2004). Glyoxal attached to a flavan-3-ols at C6 and/or C8 position bynucleophilic addition (Beretta, Furlanetto, Regazzoni, Zarrella, &Facino, 2008). The structure of one isomer (glyoxyl attached at C6 ofcatechin) is shown in Fig. 7B. The third adduct gave [M–H] − m/z 637
and a product ion m/z 289 [M–H] − formed by catechin. Its structurewas tentatively identified as a dicatechin–monoglyoxal adduct, inwhich two catechin units were cross-linked through a –(CHOH)2–group (Fig. 7C). In this adduct, two carbonyl groups of glyoxal reactedseparately with two catechins via nucleophilic addition. This adducthas not been reported before. We speculated that two catechins mayalso be cross-linked to generate C6–C6, C8–C6, and C8–C6 isomers. AC8–C8 dimer is shown in Fig. 7C.

2673W. Wang et al. / Food Research International 44 (2011) 2666–2673
Two catechin–methylglyoxal adducts were identified. Theirproduct ion spectra are shown in Fig. 6D and E. The first adductyielded m/z 361 [M-H]- and fragments at m/z 343, 289, 165 and 137.Ion m/z 343 [M–H] − was due to water elimination from m/z 361[M–H] −. Ion m/z 289 resulted from catechin. The cleavage of theC-ring of catechin yielded fragmentswithm/z 165 and 137 (Hokkanen,Mattila, Jaakola, Pirttilä, & Tolonen, 2009). This compound wastentatively identified as monocatechin–monomethylglyoxal adduct.The structure of one isomer (methylglyoxal attached at C6 ofcatechin) is shown in Fig. 7D. This structure was similar to amethylglyoxal–epicatechin adduct reported in a previous study(Totlani & Peterson, 2006). The second adduct with m/z 433 [M–H] −
was tentatively identified as a monocatechin–dimethylglyoxaladduct. It yielded m/z 415 after water elimination, m/z 362 afterlosing a methylglyoxal moiety and, m/z 253 after the cleavage ofthe C-ring of catechin. Fig. 7E shows the proposed structure ofthis catechin–dimethylglyoxal adduct. This adduct has not beenidentified before.
4. Conclusions
Our study found that sugar-free phytochemical extracts from berryfruits inhibited protein glycation. Such antiglycation effects wereattributed to their capacity to scavenge reactive carbonyls. Phenoliccompounds, such as catechin, scavenged reactive carbonyls byforming adducts with them. Sugar-free phytochemical extracts fromberries are particularly suitable for individuals with glucose intoler-ance or diabetes. Results from this study suggested these extracts arebeneficial to prevent AGEs associated chronic diseases.
References
Ahmed, N. (2005). Advanced glycation endproducts — role in pathology of diabeticcomplications. Diabetes Research and Clinical Practice, 67(1), 3–21.
American Diabetes Association (2009). Summary of Revisions for the 2009 ClinicalPractice Recommendations. Diabetes Care, 32(Supplement 1), S3–S5.
Beretta, G., Furlanetto, S., Regazzoni, L., Zarrella, M., & Facino, R. M. (2008). Quenchingof [alpha],[beta]-unsaturated aldehydes by green tea polyphenols: HPLC-ESI-MS/MS studies. Journal of Pharmaceutical and Biomedical Analysis, 48(3), 606–611.
Cho, S. J., Roman, G., Yeboah, F., & Konishi, Y. (2007). The road to advanced glycationend products: A mechanistic perspective. Current Medicinal Chemistry, 14(15),1653–1671.
Clifford, M. N., & Scalbert, A. (2000). Ellagitannins — Nature, occurrence and dietaryburden. Journal of the Science of Food and Agriculture, 80(7), 1118–1125.
Del Rio, D., Stewart, A. J., Mullen, W., Burns, J., Lean, M. E. J., Brighenti, F., & Crozier, A.(2004). HPLC-MSn Analysis of Phenolic Compounds and Purine Alkaloids in Greenand Black Tea. Journal of Agricultural and Food Chemistry, 52(10), 2807–2815.
Ellis, E. M. (2007). Reactive carbonyls and oxidative stress: Potential for therapeuticintervention. Pharmacology and Therapeutics, 115(1), 13–24.
Es-Safi, N. -E., Cheynier, V., & Moutounet, M. (2003). Effect of copper on oxidation of(+)-catechin in a model solution system. International Journal of Food Science &Technology, 38(2), 153–163.
Gabbay, K. H., & Kinoshita, J. H. (1972). Mechanism of development and possibleprevention of sugar cataracts. Israel Journal of Medical Sciences, 8(8), 1557–1561.
Giusti, M. M., & Wrolstad, R. E. (2001). Characterization and measurement ofanthocyanins by UV–visible spectroscopy. John Wiley & Sons, Inc.
Goldberg, T., Cai, W., Peppa, M., Dardaine, V., Baliga, B. S., Uribarri, J., et al. (2004).Advanced glycoxidation end products in commonly consumed foods. Journal of theAmerican Dietetic Association, 104(8), 1287–1291.
Heath, H., & Hamlett, Y. C. (1976). The sorbitol pathway: Effect of streptozotocininduced diabetes and the feeding of a sucrose-rich diet on glucose, sorbitol andfructose in the retina, blood and liver of rats. Diabetologia, 12(1), 43–46.
Hokkanen, J., Mattila, S., Jaakola, L., Pirttilä, A. M., & Tolonen, A. (2009). Identification ofphenolic compounds from lingonberry (Vaccinium vitis-idaea L.), bilberry (Vacciniummyrtillus L.) and hybrid bilberry (Vaccinium x intermedium Ruthe L.) leaves. Journal ofAgricultural and Food Chemistry, 57(20), 9437–9447.
Hollnagel, A., & Kroh, L. W. (1998). Formation of α-dicarbonyl fragments from mono- anddisaccharides under caramelization and Maillard reaction conditions. Zeitschrift fürLebensmitteluntersuchung und -Forschung A, 207(1), 50.
Huang, D., Ou, B., Hampsch-Woodill, M., Flanagan, J. A., & Prior, R. L. (2002). High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multi-channel liquid handling system coupled with a microplate fluorescence reader in96-well format. Journal of Agricultural and Food Chemistry, 50(16), 4437–4444.
Lo, C. Y., Li, S., Tan, D., Pan, M. H., Sang, S., & Ho, C. T. (2006). Trapping reactions ofreactive carbonyl species with tea polyphenols in simulated physiologicalconditions. Molecular Nutrition & Food Research, 50(12), 1118–1128.
Lv, L., Shao, X., Wang, L., Huang, D., Ho, C. -T., & Sang, S. (2010). Stilbene glucoside frompolygonummultiflorum thunb: A novel natural inhibitor of advanced glycation endproduct formation by trapping of methylglyoxal. Journal of Agricultural and FoodChemistry, 58(4), 2239–2245.
McPherson, J. D., Shilton, B. H., & Walton, D. J. (1988). Role of fructose in glycation andcross-linking of proteins. Biochemistry, 27(6), 1901–1907.
Morata, A., Calderón, F., González, M. C., Gómez-Cordovés, M. C., & Suárez, J. A. (2007).Formation of the highly stable pyranoanthocyanins (vitisins A and B) in red winesby the addition of pyruvic acid and acetaldehyde. Food Chemistry, 100(3),1144–1152.
Münch, G., Thome, J., Foley, P., Schinzel, R., & Riederer, P. (1997). Advanced glycationendproducts in ageing and Alzheimer's disease. Brain Research Reviews, 23(1–2),134–143.
Nagaraj, R. H., Sarkar, P., Mally, A., Biemel, K. M., Lederer, M. O., & Padayatti, P. S. (2002).Effect of pyridoxamine on chemical modification of proteins by carbonyls indiabetic rats: Characterization of a major product from the reaction of pyridox-amine andmethylglyoxal. Archives of Biochemistry and Biophysics, 402(1), 110–119.
Nemet, I., Varga-Defterdarovic, L., & Turk, Z. (2006). Methylglyoxal in food and livingorganisms. Molecular Nutrition & Food Research, 50(12), 1105–1117.
O'Brien, P. J., Siraki, A. G., & Shangari, N. (2005). Aldehyde sources, metabolism,molecular toxicity mechanisms, and possible effects on human health. CriticalReviews in Toxicology, 35(7), 609–662.
Oya, T., Hattori, N., Mizuno, Y., Miyata, S., Maeda, S., Osawa, T., et al. (1999).Methylglyoxal modification of protein. Chemical and immunochemical character-ization of methylglyoxal–arginine adducts. Journal of Biological Chemistry, 274(26),18492–18502.
Peng, X., Cheng, K. -W., Ma, J., Chen, B., Ho, C. -T., Lo, C., et al. (2008). Cinnamon barkproanthocyanidins as reactive carbonyl scavengers to prevent the formation ofadvanced glycation endproducts. Journal of Agricultural and Food Chemistry, 56(6),1907–1911.
Prior, R. L., Fan, E., Ji, H., Howell, A., Nio, C., Payne, M. J., et al. (2010). Multi-laboratoryvalidation of a standard method for quantifying proanthocyanidins in cranberrypowders. Journal of the Science of Food and Agriculture, 90, 1473–1478.
Romero, C., & Bakker, J. (2000). Effect of storage temperature and pyruvate on kineticsof anthocyanin degradation, vitisin a derivative formation, and color characteristicsof model solutions. Journal of Agricultural and Food Chemistry, 48(6), 2135–2141.
Sang, S., Shao, X., Bai, N., Lo, C. Y., Yang, C. S., & Ho, C. T. (2007). Tea polyphenol (−)-epigallocatechin-3-gallate: A new trapping agent of reactive dicarbonyl species.Chemical Research in Toxicology, 20(12), 1862–1870.
Scordino, M., Di Mauro, A., Passerini, A., & Maccarone, E. (2004). Adsorption offlavonoids on resins: Cyanidin 3-glucoside. Journal of Agricultural and FoodChemistry, 52(7), 1965–1972.
Singleton, V. L., & Rossi, J. A., Jr. (1965). Colorimetry of total phenolics withphosphomolybdic-phosphotungstic acid reagents. American Journal of Enologyand Viticulture, 16(3), 144–158.
Suarez, G., Rajaram, R., Oronsky, A. L., & Gawinowicz, M. A. (1989). Nonenzymaticglycation of bovine serum albumin by fructose (fructation). Comparison with theMaillard reaction initiated by glucose. Journal of Biological Chemistry, 264(7),3674–3679.
Tan, D., Wang, Y., Lo, C. Y., Sang, S., & Ho, C. T. (2008). Methylglyoxal: Its presence inbeverages and potential scavengers. Annals of the New York Academy of Sciences,1126, 72–75.
Tarwadi, K. V., & Agte, V. V. (in press). Effect ofmicronutrients onmethylglyoxal-mediated in vitro glycation of albumin. Biological Trace Element Research,doi:10.1007/s12011-010-8915-7.
Totlani, V. M., & Peterson, D. G. (2006). Epicatechin carbonyl-trapping reactions inaqueous maillard systems: Identification and structural elucidation. Journal ofAgricultural and Food Chemistry, 54(19), 7311–7318.
Copyright © 2022 FDOKUMEN




















