
The woody plant poplar has a functionally conserved salt overly sensitive pathway in response to...
14
The woody plant poplar has a functionally conserved salt overly sensitive pathway in response to salinity stress Ren-Jie Tang • Hua Liu • Yan Bao • Qun-Dan Lv • Lei Yang • Hong-Xia Zhang Received: 9 July 2010 / Accepted: 12 August 2010 / Published online: 29 August 2010 Ó Springer Science+Business Media B.V. 2010 Abstract In Arabidopsis thaliana, the salt overly sensi- tive (SOS) pathway plays an essential role in maintaining ion homeostasis and conferring salt tolerance. Here we identified three SOS components in the woody plant Pop- ulus trichocarpa, designated as PtSOS1, PtSOS2 and PtSOS3. These putative SOS genes exhibited an overlap- ping but distinct expression pattern in poplar plants and the transcript levels of SOS1 and SOS2 were responsive to salinity stress. In poplar mesophyll protoplasts, PtSOS1 was specifically localized in the plasma membrane, whereas PtSOS2 was distributed throughout the cell, and PtSOS3 was predominantly targeted to the plasma mem- brane. Heterologous expression of PtSOS1, PtSOS2 and PtSOS3 could rescue salt-sensitive phenotypes of the cor- responding Arabidopsis sos mutants, demonstrating that the Populus SOS proteins are functional homologues of their Arabidopsis counterpart. In addition, PtSOS3 inter- acted with, and recruited PtSOS2 to the plasma membrane in yeast and in planta. Reconstitution of poplar SOS pathway in yeast cells revealed that PtSOS2 and PtSOS3 acted coordinately to activate PtSOS1. Moreover, expres- sion of the constitutively activated form of PtSOS2 par- tially complemented the sos3 mutant but not sos1, suggesting that PtSOS2 functions genetically downstream of SOS3 and upstream of SOS1. These results indicate a strong functional conservation of SOS pathway responsible for salt stress signaling from herbaceous to woody plants. Keywords SOS pathway Salt stress Populus Arabidopsis Functional conservation Introduction Soil salinity has become one of the major worldwide agricultural problems. It is a severe limiting factor that adversely affects plant growth in general and crop pro- ductivity in particular. For most plants, high salt concen- trations impose both an ionic and an osmotic stress (Zhu 2001), coupled with secondary stresses such as oxidative stress (Borsani et al. 2001), genotoxicity (Albinsky et al. 1999) and nutritional disorders (Parida et al. 2004). Sodium (Na ? ) toxicity represents the major ionic stress associated with high salinity. The excessive accumulation of Na ? in the cytoplasm has injurious effects on plant cells in that it prevents uptake of the essential mineral nutrient potassium (K ? ), leading to insufficient cellular K ? amount for enzymatic reactions and osmotic adjustment. Therefore, maintaining an optimal cytosolic K ? /Na ? homeostasis is crucial for plant salt tolerance (Niu et al. 1995; Hasegawa et al. 2000; Zhu 2003; Volkov et al. 2004). In order to preserve a low Na ? concentration in the cytoplasm, plants have evolved several machineries at the cellular level (Blumwald 2000; Horie and Schroeder 2004; Chen et al. 2007). One possible way is to restrict unidirectional Na ? uptake by roots (Rubio et al. 1995; Laurie et al. 2002). Alternatively, Na ? extrusion away from the cytoplasm (Shi et al. 2002) and Na ? compartmentation into the vacuole (Apse et al. 1999) may serve as active means to minimize Electronic supplementary material The online version of this article (doi:10.1007/s11103-010-9680-x) contains supplementary material, which is available to authorized users. R.-J. Tang H. Liu Y. Bao Q.-D. Lv L. Yang H.-X. Zhang (&) National Key Laboratory of Plant Molecular Genetics, Institute of Plant Physiology and Ecology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, 300 Fenglin Road, 200032 Shanghai, China e-mail: [email protected] 123 Plant Mol Biol (2010) 74:367–380 DOI 10.1007/s11103-010-9680-x
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The woody plant poplar has a functionally conserved salt overly sensitive pathway in response to...

The woody plant poplar has a functionally conserved salt overlysensitive pathway in response to salinity stress
Ren-Jie Tang • Hua Liu • Yan Bao •
Qun-Dan Lv • Lei Yang • Hong-Xia Zhang
Received: 9 July 2010 / Accepted: 12 August 2010 / Published online: 29 August 2010
� Springer Science+Business Media B.V. 2010
Abstract In Arabidopsis thaliana, the salt overly sensi-
tive (SOS) pathway plays an essential role in maintaining
ion homeostasis and conferring salt tolerance. Here we
identified three SOS components in the woody plant Pop-
ulus trichocarpa, designated as PtSOS1, PtSOS2 and
PtSOS3. These putative SOS genes exhibited an overlap-
ping but distinct expression pattern in poplar plants and the
transcript levels of SOS1 and SOS2 were responsive to
salinity stress. In poplar mesophyll protoplasts, PtSOS1
was specifically localized in the plasma membrane,
whereas PtSOS2 was distributed throughout the cell, and
PtSOS3 was predominantly targeted to the plasma mem-
brane. Heterologous expression of PtSOS1, PtSOS2 and
PtSOS3 could rescue salt-sensitive phenotypes of the cor-
responding Arabidopsis sos mutants, demonstrating that
the Populus SOS proteins are functional homologues of
their Arabidopsis counterpart. In addition, PtSOS3 inter-
acted with, and recruited PtSOS2 to the plasma membrane
in yeast and in planta. Reconstitution of poplar SOS
pathway in yeast cells revealed that PtSOS2 and PtSOS3
acted coordinately to activate PtSOS1. Moreover, expres-
sion of the constitutively activated form of PtSOS2 par-
tially complemented the sos3 mutant but not sos1,
suggesting that PtSOS2 functions genetically downstream
of SOS3 and upstream of SOS1. These results indicate a
strong functional conservation of SOS pathway responsible
for salt stress signaling from herbaceous to woody plants.
Keywords SOS pathway � Salt stress � Populus �Arabidopsis � Functional conservation
Introduction
Soil salinity has become one of the major worldwide
agricultural problems. It is a severe limiting factor that
adversely affects plant growth in general and crop pro-
ductivity in particular. For most plants, high salt concen-
trations impose both an ionic and an osmotic stress (Zhu
2001), coupled with secondary stresses such as oxidative
stress (Borsani et al. 2001), genotoxicity (Albinsky et al.
1999) and nutritional disorders (Parida et al. 2004). Sodium
(Na?) toxicity represents the major ionic stress associated
with high salinity. The excessive accumulation of Na? in
the cytoplasm has injurious effects on plant cells in that it
prevents uptake of the essential mineral nutrient potassium
(K?), leading to insufficient cellular K? amount for
enzymatic reactions and osmotic adjustment. Therefore,
maintaining an optimal cytosolic K?/Na? homeostasis is
crucial for plant salt tolerance (Niu et al. 1995; Hasegawa
et al. 2000; Zhu 2003; Volkov et al. 2004). In order to
preserve a low Na? concentration in the cytoplasm, plants
have evolved several machineries at the cellular level
(Blumwald 2000; Horie and Schroeder 2004; Chen et al.
2007). One possible way is to restrict unidirectional Na?
uptake by roots (Rubio et al. 1995; Laurie et al. 2002).
Alternatively, Na? extrusion away from the cytoplasm (Shi
et al. 2002) and Na? compartmentation into the vacuole
(Apse et al. 1999) may serve as active means to minimize
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11103-010-9680-x) contains supplementarymaterial, which is available to authorized users.
R.-J. Tang � H. Liu � Y. Bao � Q.-D. Lv � L. Yang �H.-X. Zhang (&)
National Key Laboratory of Plant Molecular Genetics,
Institute of Plant Physiology and Ecology, Shanghai Institutes
for Biological Sciences, Chinese Academy of Sciences,
300 Fenglin Road, 200032 Shanghai, China
e-mail: [email protected]
123
Plant Mol Biol (2010) 74:367–380
DOI 10.1007/s11103-010-9680-x

sodium toxicity in plant cells. These vital Na? transloca-
tion processes are mediated by a battery of ion transporters
(Horie and Schroeder 2004) under the control of delicate
signaling networks that modulate their activities and
affinities (Xiong et al. 2002).
In Arabidopsis, the salt overly sensitive (SOS) pathway
is responsible for Na? homeostasis and salt tolerance under
salt stress. This pathway was established through genetic
identification of several salt overly sensitive (sos) mutants
followed by molecular and biochemical characterization
of the coding SOS proteins (Zhu 2002). A myristoylated
EF-hand calcium-binding protein encoded by SOS3 pre-
sumably senses and interprets the cellular calcium signal
elicited by salt stress (Liu and Zhu 1998; Ishitani et al.
2000). One of the primary downstream targets SOS2, a
serine/threonine protein kinase (Liu et al. 2000), is acti-
vated and recruited to plasma membrane via direct inter-
action with SOS3 (Halfter et al. 2000; Quintero et al.
2002). Subsequently, the SOS2-SOS3 complex phosphor-
ylates the Na?/H? antiporter SOS1 (Shi et al. 2000) to
stimulate its Na?/H? exchange activity at the plasma
membrane (Quintero et al. 2002; Qiu et al. 2002). In yeast,
co-expression of SOS2, SOS3, together with SOS1
increases salt tolerance of a yeast mutant deficient in
sodium transport more dramatically than expression of one
or two SOS proteins, suggesting that full activation of
SOS1 depends on the SOS2-SOS3 kinase complex (Quin-
tero et al. 2002). In Arabidopsis, vesicles of sos1, sos2 and
sos3 plants all display reduced plasma membrane Na?/H?
exchange activity compared to those of wild type plants,
and a constitutively activated SOS2 protein enhance the
Na?/H? exchange activity in a SOS1-dependent and
SOS3-independent manner (Qiu et al. 2002). Accordingly,
overexpression of SOS1 (Shi et al. 2003) or a constitutively
activated form of SOS2 (Guo et al. 2004) or the putative
upstream sensor SOS3 (Yang et al. 2009) all confers
enhanced salt tolerance on transgenic plants. Likewise, the
rice SOS1 homolog OsSOS1 functions as a plasma mem-
brane Na?/H? antiporter and is phosphorylated by the
SOS2–SOS3 kinase complex. Ectopic expression of
OsSOS1 suppressed the growth defects of Arabidopsis sos1
mutant upon salt treatment. The SOS2/SOS3 homolog in
rice, OsCIPK24/OsCBL4, acted coordinately to activate
OsSOS1 in yeast cells, and suppressed salt sensitivity of
Arabidopsis sos2/sos3 mutant, indicating that the SOS
pathway for salt tolerance is also operational in cereals
(Martinez-Atienza et al. 2007).
Although previous studies have highlighted important
roles of SOS pathway in the dicot herbaceous plant Ara-
bidopsis and the monocot plant rice, little is known about
the conservation of this pathway in trees. Populus has
become an important model in woody plants (Jansson
and Douglas 2007), due to its worldwide distribution,
sequenced genome and relative ease of genetic manipula-
tion (Tuskan et al. 2006). Poplars have also received
increasing attention as a renewable source of biomass for
energy and short-fiber pulp for paper making (Luo and
Polle 2009; Pilate et al. 2002). Recently, a putative plasma
membrane Na?/H? antiporter PeSOS1 from Populus eu-
phratica, has been isolated and shown to partially suppress
the salt-sensitivity of the Escherichia coli mutant strain
EP432 (Wu et al. 2007), but analysis of its in planta
function is still lacking. In order to better understand the
molecular mechanism underlying salt tolerance in woody
plants, we isolated three genes coding for key components
in the SOS pathway from the sequenced poplar genotype
Populus trichocarpa. By molecular analysis and functional
characterization of these three PtSOS genes, we show that
the perennial woody plant poplar has a conserved SOS
pathway in response to salinity stress.
Materials and methods
Plant materials, growth conditions and stress treatment
Two poplar genotypes Populus trichocarpa and Populus
alba 9 P. Berolinensis cv. Yin-Zhong were used in this
study. Generally, in vitro grown plants were subcultured
every month by aseptically transferring shoot apices to the
fresh MS medium (Murashige and Skoog 1962) supple-
mented with 0.03 mg/L NAA for rooting. Three-week-old
micro-propagated plantlets were subjected to salt stress by
directly pouring 30 mL of 200 mM NaCl solution into the
tissue-culture containers. Plantlets were also transferred
into individual pots (volume 6 L) after 3-day acclimation
to ex vitro conditions, and grown in the greenhouse under
an 14-h photoperiod comprising natural daylight supple-
mented with lamps (Philips, 600 W), giving a minimum
quantum flux density of 150 lEm-2s-1. The temperature
was kept at about 21–24�C in the daytime and 15–18�C at
night. All the plants were well irrigated according to
evaporation demands during different growth stages and
watered with half strength of Hoagland nutrient solutions
every other week. After 6 months’ growth, the trees would
reach a height of approximately 240 cm and a diameter of
1.2 cm. Various tissues were isolated from vigorously
growing trees including apical buds, young leaves, mature
leaves, elongating internodes, stem segments and roots.
Xylem and phloem tissues were simply separated by
stripping off the bark with a sharp blade. The samples were
frozen in liquid nitrogen immediately and stored at -80�C.
Arabidopsis thaliana plants were grown in the green-
house under long-day conditions (16 h light/8 h dark) at
the temperature 22–23�C. Sterilized seeds were plated on
MS medium solidified with 0.8% agar. All of the sos1, sos2
368 Plant Mol Biol (2010) 74:367–380
123

and sos3 mutants in this study were in the Col-0 gl1
background.
Reverse transcription PCR and quantitative real-time
PCR analyses
Total RNA was extracted with the RNAiso Reagent
(Takara, Japan) from different organs or tissues of two
types of poplar plants at the half-year growth stage in the
green house. Following the manufacturer’s instruction,
RNA samples were obtained with the same reagent from
the salt-treated poplar plantlets and 10-day old Arabidopsis
seedlings. After being treated with DNase I (Promega),
2 lg of total RNA was subjected to reverse transcription
reaction using the reverse transcriptase ReverTra Ace
(TOYOBO, Japan) at 42�C for 1 h. The resulting cDNA
was then used for PCR amplification with the gene-specific
primers listed below. For PtSOS1, the primers S1RT-F
(50-AAAGCCCATGGTTTCCAAGATG-30), and S1RT-R
(50-CCGC-TTCAAATGCTGCAATATC-30) were used;
For PtSOS2, the primers S2RT-F (50-AAAGCCCATGGTT
TCCAAGATG-30), and S2RT-R (50-AGTCATTGTTCGA
AGCAGGCAG-30) were used; For PtSOS3, the primers
S3RT-F (50-TGGAAAAATCGATCCTGACGAGTG-30),and S3RT-R (50-GCTGGTTTAAAATGCACGGATCAC-30)were used; For EF1b, the primers EF1b-F (50-GACAA
GAAGGCAGCGGAGGAGAG-30), and EF1b-R (50-CAA
TGAGGGAATCCACTGACACAAG-30) were used; For
ACTIN2, the primers ACT2-F (50-GGAAGGATCTGTAC
GGTAAC-30) and ACT2-R (50-GGACCTGCCTCATCA
TACT-30) were used.
Quantitative real-time PCR analysis was performed with
the RotorGene 3000 system (Corbett Research) using the
SYBR Green Realtime PCR Master Mix (TOYOBO,
Japan) to monitor double-stranded DNA products. Data
analysis was performed with Rotor-Gene software version
6.0 and relative amounts of mRNA were calculated based
on the comparative threshold cycle method. The relative
expression of each target gene was double-normalized
using the housekeeping gene EF1b and using the control
expression values measured at 0 h.
Subcellular localization of PtSOS1, PtSOS2
and PtSOS3
To determine the subcellular localization of PtSOS pro-
teins, stop-codon-less coding region of PtSOS1, PtSOS2 or
PtSOS3 was in-frame fused upstream to the YFP sequence
in the pA7-YFP vector, respectively. The resulting con-
structs containing PtSOS-YFP translational fusions were
transfected into poplar mesophyll protoplasts, essentially as
described by Sheen and colleges (Sheen 2001; Yoo et al.
2007). Fluorescence of YFP in the transformed protoplasts
was imagined using a confocal laser scanning microscope
(LSM510, Carl Zeiss) after the protoplasts were incubated
at 23�C for 16 h.
Yeast experiments
Yeast two hybrid assays were based on Matchmaker GAL4
Two-Hybrid System 3 (Clontech). The PtSOS2 coding
sequence was excised from the pBluescript II KS construct
using EcoRI-XhoI double digestion and subcloned into to
pGADT7 vector to generate an in-frame fusion with AD.
Similarly, PtSOS3 was inserted into pGBDT7 vector
through EcoRI-SalI sites to generate an in-frame fusion
with BD. To generate PtSOS2 devoid of the FISL motif,
inverse PCR-based mutagenesis was carried out on double-
stranded pKS-PtSOS2 plasmid, resulting in pKS-
PtSOS2DF. Then PtSOS2DF was subcloned into pGADT7
vector. These constructs and empty vector controls were
transformed into yeast strain AH109 by the PEG/LiAc
method. Yeast cells were plated onto SD/-Ade/-His/-Trp/-
Leu medium for stringent screening of the possible interac-
tions. b-galactosidase assays were performed as described in
the Clontech Yeast Protocols Handbook.
For experiments with the yeast Ras recruitment system
(RRS), the p426GPD-RAS vector was used to express RAS
fusion baits and the p425GPD vector (Mumberg et al.
1995) was used to express prey proteins. For p426GPD-
RAS construction, a Ras fragment without the stop codon
was produced by PCR amplification using the pYES-RRS
plasmid (Broder et al. 1998) as a template with the primers
RAS-F (50-CCCACTAGTATGACGGAATATAAGCTGG-30)and RAS-R (50-GGGGGATCC CTTGCAGCTCATGCAG
C-30), and then cloned into the p426GPD vector via SpeI-
BamHI double digestion. Afterwards, the coding sequence
of PtSOS2, PtSOSDF or AtSOS2 was in-frame fused
downstream to RAS in the p426GPD-RAS construct
between BamHI and SalI sites. PtSOS3 or AtSOS3 was
directly inserted into the p425GPD construct. Several
combinations of these plasmids and empty vector controls
were transferred into the yeast strain cdc25-2 (MATa, ade2,
his3, leu2, lys2, trp1, ura3, cdc25-2) and transformed yeast
cells were selected on SD/-Ura/-Leu medium. For inter-
action analysis, serial decimal dilutions of yeast cultures
(stating from OD = 1.0) were spotted onto YPD plates at
the permissive (24�C) or the restrictive (36�C) temperature.
Five randomly selected yeast colonies after transformation
were subjected to this assay.
To express SOS genes in yeast, PtSOS1 or AtSOS1 was
constructed into the p426PMA vector, a derivant from
p426GPD in which the GPD promoter was replaced by the
PMA1 promoter (Capieaux et al. 1989). The yeast vector
p414GPD (Mumberg et al. 1995) was employed to express
PtSOS2 or PtSOS3 alone. To coordinate expression of
Plant Mol Biol (2010) 74:367–380 369
123

PtSOS2 and PtSOS3 within a single plasmid, we con-
structed PtSOS2:CYC1-GPD:PtSOS3 on the backbone of
pBluescript II KS plasmid by successively introducing four
PCR fragments (the PtSOS2 CDS; the CYC1 terminator;
the GPD promoter; the PtSOS3 CDS) into the multiple
cloning site (MCS). The quadruple cassette was then sub-
cloned into the p414GPD plasmid via BamHI-XhoI double
digestion, resulting in the final construct harboring two
expression cassettes in tandem (GPD:PtSOS2:CYC1-
GPD:PtSOS3:CYC1). The Saccharomyces cerevisiae strain
B31 (MATa, ade2, can1, his3, leu2, trp1, ura3, mall0,
Dena1::HIS3::ena4, Dnha1::LEU2, Banuelos et al. 1998)
was transformed with one of or combinations of these
expression constructs. Na? tolerance tests were performed
in arginine-phosphate (AP) medium (8 mM phosphoric
acid, 10 mM L-Arg, 2 mM MgSO4, 0.2 mM CaCl2, 2%
glucose, plus vitamins and trace elements, pH = 6.0).
Aliquots (3 lL) were spotted onto AP plates supplemented
with 0, 120 and 240 mM NaCl in addition to 1 mM KCl,
and grown for 60 h at 30�C.
Bimolecular fluorescence complementation
(BiFC) assays
For generation of the BiFC vectors, the coding region of
PtSOS2 was subcloned via BamHI-SmaI into pSPYNE-
35S, resulting in PtSOS2YFPN; and the coding region of
PtSOS3 was cloned via BamHI-XhoI into pSPYCE-35S,
resulting in PtSOS3YFPC. Infiltration of Nicotiana benth-
amiana leaves was performed as previously described by
Walter et al. (2004). Protoplasts were prepared 3 days after
infiltration by cutting leaf discs into small pieces and
incubating for 3 h in the enzyme solution (0.4 M mannitol,
20 mM MES-K, 10 mM CaCl2, 5 mM b-mercaptoethanol,
0.1% BSA, 1% cellulase R10, 0.3% macerozyme R10, pH
5.7). Fluorescence of YFP in the leaf epidermal cells or
isolated protoplasts was imagined by a confocal laser
scanning microscope (LSM510, Carl Zeiss).
Arabidopsis transformation and complementation test
To prepare constructs for plant transformation, the open
reading frame of PtSOS1, PtSOS2 or PtSOS3 was digested
from the pBluescript II KS vectors with SmaI and SalI,
respectively. To produce the constitutively activated form
of PtSOS2, a Thr169-to-Asp mutation was introduced into
pKS-PtSOS2 by PCR-based Site-Directed Mutagenesis Kit
(Stratagene) using the primers 50-TTGGACTTCTTCATG
ACACATGTGGAACCCCG-30 and 50-CCCCTTCTGCGG
CAACGCACTCAATCC-30. The yielding pKS-PtSOS2TD
plasmid was also digested with BamHI and SalI to release
PtSOS2TD coding sequence. All the fragments were then
respectively inserted into a modified pCAMBIA-2301
vector downstream of the cauliflower mosaic virus 35S
promoter. These plasmids were separately introduced into
the Agrobacterium GV3101 strain for Arabidopsis trans-
formation. Putative transgenic plants were screened on MS
medium containing 50 lg/L kanamycin and transferred to
soil for propagations. Kanamycin-resistant plants of T2
generation were selected and subjected to transgene
expression analyses. Seeds of homozygous T2 lines were
harvested and the T3 generation was employed for com-
plementation tests on MS agar plates supplemented with
NaCl, as indicted for each case.
Results
Isolation of PtSOS1, PtSOS2 and PtSOS3 from Populus
We performed homology-based BLAST searches (Altschul
et al. 1997) to collect candidate gene sequences coding for
poplar SOS pathway components from the Joint Genome
Initiative poplar database (JGI, Populus trichocarpa
genome portal v1.1; http://genome.jgi-psf.org/Poptr
1_1/Poptr1_1.home.html). Two SOS1 (with 89% identity),
SOS2 (with 91% identity) and SOS3 (with 94% identity)
homologues were identified from the whole Populus gen-
ome. Due to high similarity within individual Populus SOS
candidates, we chose only one gene of each as a repre-
sentative, with the closest homology to Arabidopsis SOS
counterpart, for subsequent studies. These three genes were
designated as PtSOS1, PtSOS2 and PtSOS3. Total RNA
was extracted from salt-treated plantlets of Populus
trichocarpa and first-strand cDNA was used to amplify the
coding region of each gene on the basis of public genomic
sequence. The amplified fragments were subcloned to
pBluescript II KS vector and then sequenced independently
three times to confirm proper removal of introns and
fidelity of the polymerase. After manual revision of the
isolated CDS and its coding protein, sequence alignment
was conducted between PtSOS1, 2, 3 and AtSOS1, 2, 3
respectively (Figs. S1, S2, S3). The three putative SOS
proteins in poplar bear significant resemblance to their
Arabidopsis counterparts in size and structure.
The poplar SOS components show overlapping
but distinct expression pattern
As the first step to characterize SOS pathway in woody
plants, expression of poplar SOS genes was analyzed to
clarify whether the three components co-localize and
thereby co-function in planta. We performed RT–PCR to
determine relative transcript abundance in various poplar
370 Plant Mol Biol (2010) 74:367–380
123

tissues from two genotypes in our lab, i.e. the sequenced
genotype Populus trichocarpa and an aspen genotype
Populus alba 9 P. Berolinensis (a common cultivar in the
North of China). The final amount of EF1b products ver-
ified that all samples contained approximately equal
amount of cDNA. In Populus trichocarpa, PtSOS1 and
PtSOS2 exhibited generally high, uniform and ubiquitous
expression in all tissues (Fig. 1a). Strong expression of
PtSOS1 was detected in root, juvenile leaf, stem, xylem
and apex while expression of PtSOS2 might be slightly
higher in young green tissues such as juvenile leaf and apex
than in other organs. The transcript of PtSOS3 was low in
most poplar tissues but relatively abundant in root. In
Populus alba 9 P. Berolinensis, a very similar expression
pattern was observed with regard to PabSOS1 and Pab-
SOS2, while preferential expression of PabSOS3 in root
tissues was even more evident (Fig. 1b). In conclusion, the
poplar SOS components show an overlapping but also a
distinct expression pattern, which may be analogous among
different Populus species.
Expression of SOS genes in the poplar is regulated
by salinity stress
To further examine whether the expression of poplar
SOS components is modified by salinity stress, 3-week-old
micro-propagated poplar plantlets were subjected to
200 mM NaCl for increasing exposure periods and the
transcript level of SOS1, SOS2 and SOS3 was analyzed by
quantitative real-time PCR. Since the sequenced genotype
Populus trichocarpa is extremely susceptible to salinity and
became wilted during our treatment period, we employed
Populus alba 9 P. Berolinensis in this test as our experi-
mental materials. In roots, PabSOS1 was rapidly induced
after the onset of salt stress and increased up to the maximal
level (around fivefold) at the twelfth hour during the NaCl
treatment. Expression of PabSOS2 coordinately increased
within 3 h, reached a maximal threefold to fourfold induc-
tion between 6 and 12 h, and thereafter declined. In contrast,
expression of PabSOS3 in roots was hardly responsive to
salinity stress during our experiment (Fig. 2a). In shoot
tissues (Fig. 2b), transcripts of both PabSOS1 and PabSOS2
began to accumulate within the first 3 h of NaCl treatment
and retained continuously higher expression than basal level
from 6 to 24 h. However, the transcript level of PabSOS3 in
above-ground tissues was reduced almost by half upon 1-h
salt treatment and recovered to the near-basal level after
12 h of salt treatment. These results indicate that SOS1 and
SOS2, but not SOS3, are salt responsive genes in the poplar
SOS pathway.
Subcellular localization of PtSOS1, PtSOS2
and PtSOS3 in poplar cells
Proteins that participate in the same pathway are supposed
to occupy overlapped locations within the cell. In order to
investigate subcellular localization of the Populus SOS
components in plant cells, we fused PtSOS1, PtSOS2 and
PtSOS3 in-frame to the N-terminus of yellow fluorescent
protein (YFP), respectively. The chimeric construct was
delivered into mesophyll protoplasts prepared from poplar
leaf tissues by the polyethylene glycol transformation
method (Sheen 2001; Yoo et al. 2007). Each fusion protein
was transiently expressed under the control of the consti-
tutive 35S promoter and its subcellular localization was
visualized by confocal microscopy. As expected in the case
of the positive control YFP alone, a diffuse green fluores-
cence typical of cytosolic location was observed with a
tendency to accumulate in the nucleus and near the plasma
membrane (Fig. 3, top panels). When fused to YFP,
PtSOS1 was exclusively localized in the plasma membrane
of poplar protoplasts (Fig. 3, second panels from top),
consistent with its identity as a Na?/H? exchanger at the
cell membrane. PtSOS2-YFP, however, was dispersed
Fig. 1 Expression analysis of the SOS1, 2, 3 genes in different
tissues of Populus trichocarpa (a) and Populus alba 9 P. Berolin-ensis (b). cDNA derived from the indicated tissues was used as a
template for PCR with the gene specific primers at different numbers
of PCR cycles. Amplification of the genomic DNA sample as a
control template was also carried out to rule out its possible
contamination in each cDNA sample. PCR products were visualized
by agarose gel electrophoresis and ethidium bromide staining. The
elongation factor gene EF1b was employed as an internal control
Plant Mol Biol (2010) 74:367–380 371
123

throughout the cytoplasm, within the nucleus and also at
the cell periphery, quite similar to YFP alone (Fig. 3, third
panels from top). PtSOS3-YFP was predominantly targeted
to the plasma membrane with leaky intracellular localiza-
tion (Fig. 3, bottom panels). These observations are in
favor of the physical possibility that PtSOS1, PtSOS2 and
PtSOS3 could be involved in the same pathway at the
plasma membrane of poplar cells.
Populus SOS proteins are functional homologues
of their Arabidopsis counterparts
The Arabidopsis mutants in the SOS pathway, i.e. sos1,
sos2 and sos3, are specifically hypersensitive to high
external Na? concentrations. To dissect the biological
function of PtSOS proteins, we constructed transgenic
Arabidopsis lines heterologously expressing each PtSOS
gene in the corresponding sos mutant background for
functional complementation tests. We selected four inde-
pendent complemented lines from rescued sos1, sos2 and
sos3 mutants, respectively. By RT–PCR analysis, we
confirmed the expression of PtSOS1 transgene in sos1,
PtSOS2 transgene in sos2, and PtSOS3 transgene in sos3
(Fig. 4a). Preliminary assays on salt tolerance of these
transgenic lines for each gene showed that they had similar
salt tolerance capacity, and therefore two representative
transgenic lines (CS12, CS16; CS26, CS29; CS35, CS36)
of each gene were chosen for subsequent phenotypic
analyses. In the absence of NaCl treatment, wild type
plants, sos mutants and PtSOS complemented lines dis-
played relatively uniform growth phenotype, although
roots of the seedlings in sos1 background were slightly
shorter (Fig. 4b, top panels). When treated with 60 mM
NaCl, growth of sos1, sos2 and sos3 mutants were all
inhibited compared with wild type plants, albeit to different
extent. Expression of PtSOS1 in sos1, PtSOS2 in sos2 and
PtSOS3 in sos3 substantially alleviate the growth retarda-
tion imposed by moderate salt stress (Fig. 4b, middle
panels). Under 120 mM NaCl treatment, growth inhibition
of sos2 and sos3 mutants was even more pronounced.
Fig. 2 Quantitative real-time
PCR analyses of poplar SOS
components in response to salt
stress. Three-week-old poplar
plantlets grown on MS medium
were treated with 200 mM
NaCl. Samples were collected at
0, 1, 3, 6, 12 and 24 h after the
initiation of treatment. Total
RNA was extracted from roots
a and shoots b respectively, and
real-time PCR analyses were
performed with SOS1, 2 and 3gene specific primers. The
relative expression of each
target gene was double-
normalized using the
housekeeping gene EF1b and
using the control expression
values measured at 0 h. Data
represents the average of three
independent experiments ±SD
372 Plant Mol Biol (2010) 74:367–380
123

Seedlings of sos1 were killed by this high concentration of
salt stress. Nevertheless, the sos1 mutants expressing
PtSOS1 could survive at this lethal NaCl level. The salt-
sensitive phenotype was also greatly suppressed in
PtSOS2- and PtSOS3-complemented lines (Fig. 4b, bottom
panels). A detailed analysis of root length and seedling
fresh weight showed that all the rescued transgenic plants
by PtSOS genes were generally restored to a near-wild-type
level of salt tolerance, although with differential extents
between different lines (Fig. 4c–h). These data indicate the
poplar SOS proteins can substitute for the corresponding
Arabidopsis SOS components, thereby being functional
orthologues of their Arabidopsis counterparts.
PtSOS3 interacts with and recruits PtSOS2
to the plasma membrane
In Arabidopsis, SOS2 interacted with SOS3 to form a
kinase complex for proper biological functions (Halfter
et al. 2000; Gong et al. 2004). Here we determined the
physical interaction between PtSOS2 and PtSOS3 by yeast
two hybrid assays. A combination of the bait plasmid
pGBKT7-PtSOS3 with the prey construct pGADT7-SOS2
resulted in full transcriptional activation of HIS, ADE and
lacZ reporters, whereas co-transformation of either empty
vector with the construct pGBKT7-PtSOS3 or pGADT7-
SOS2 did not activate any of the reporter genes. The FISL
motif (also known as NAF domain) in the SOS2 regulatory
region is necessary and sufficient for SOS3 interaction
(Guo et al. 2001; Albrecht et al. 2001). Deletion of this
conserved FISL motif away from PtSOS2 protein
(PtSOS2DF) abolished its interaction with PtSOS3
(Fig. 5a), suggesting that PtSOS2 also interacts with
PtSOS3 via the FISL motif.
To further address spatial specificity of PtSOS2–PtSOS3
complex formation, we employed the Ras recruitment
system (RRS) to monitor membrane targeting of PtSOS2
by PtSOS3 interaction. This system is based on the ability
of a mammalian Ras variant to be localized to the plasma
membrane through the interaction between two proteins.
Ras membrane localization via protein–protein interaction
results in complementation of the yeast thermo-sensitive
mutant cdc25-2 deficient in the Ras guanyl nucleotide
exchange factor (Broder et al. 1998). As shown in Fig. 6b,
expression of RAS-PtSOS2 fusion alone without PtSOS3
failed to restore yeast growth at the restrictive temperature
(36�C), indicating a cytosolic localization. However, the
RAS-SOS2 fusion was efficiently recruited to the plasma
Fig. 3 Subcellular localization
of PtSOS1-, 2- and 3-YFP
fusion proteins. Confocal
microscopy analysis of YFP
signals from poplar mesophyll
protoplasts transiently
expressing YFP or PtSOS-YFP
fusions as indicated. Scalebar = 5 lm
Plant Mol Biol (2010) 74:367–380 373
123

membrane upon co-expression of PtSOS3, allowing cdc25-
2 strain to grow at 36�C. RAS fusion with the PtSOS2
mutant lacking FISL motif disrupted PtSOS3 interaction
and thus did not restore the thermo-tolerance in the pres-
ence of PtSOS3. In conclusion, PtSOS3 recruited PtSOS2
to the plasma membrane through their physical interaction.
374 Plant Mol Biol (2010) 74:367–380
123
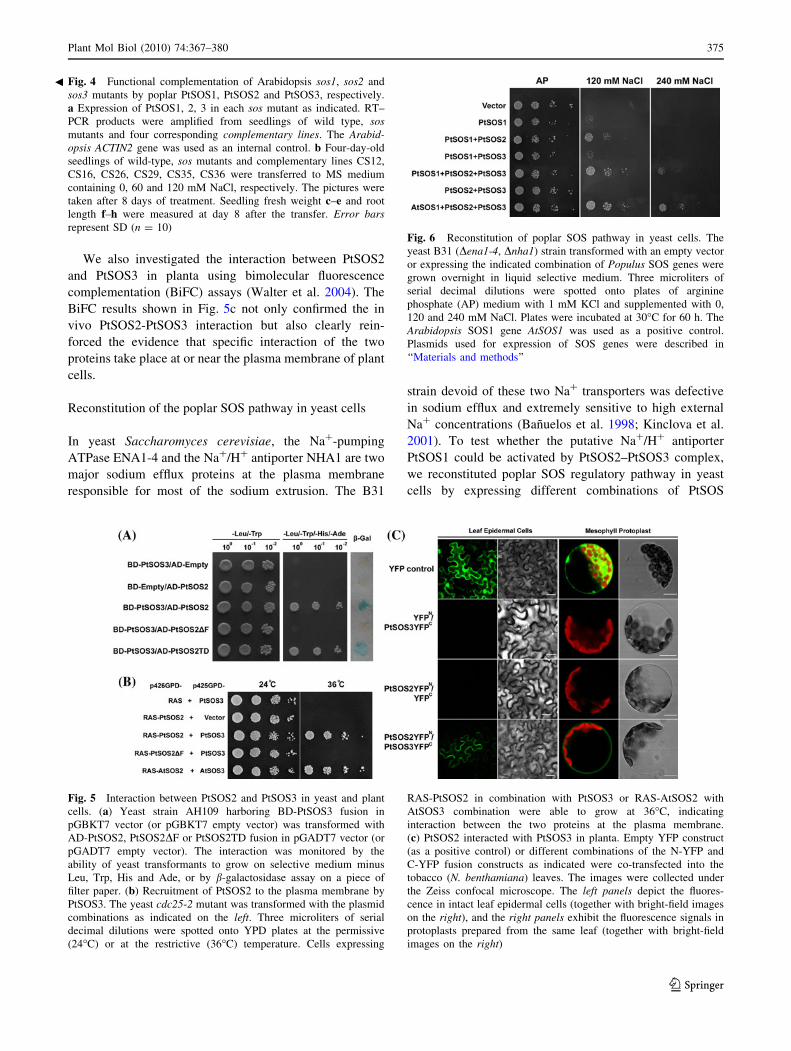
We also investigated the interaction between PtSOS2
and PtSOS3 in planta using bimolecular fluorescence
complementation (BiFC) assays (Walter et al. 2004). The
BiFC results shown in Fig. 5c not only confirmed the in
vivo PtSOS2-PtSOS3 interaction but also clearly rein-
forced the evidence that specific interaction of the two
proteins take place at or near the plasma membrane of plant
cells.
Reconstitution of the poplar SOS pathway in yeast cells
In yeast Saccharomyces cerevisiae, the Na?-pumping
ATPase ENA1-4 and the Na?/H? antiporter NHA1 are two
major sodium efflux proteins at the plasma membrane
responsible for most of the sodium extrusion. The B31
strain devoid of these two Na? transporters was defective
in sodium efflux and extremely sensitive to high external
Na? concentrations (Banuelos et al. 1998; Kinclova et al.
2001). To test whether the putative Na?/H? antiporter
PtSOS1 could be activated by PtSOS2–PtSOS3 complex,
we reconstituted poplar SOS regulatory pathway in yeast
cells by expressing different combinations of PtSOS
Fig. 5 Interaction between PtSOS2 and PtSOS3 in yeast and plant
cells. (a) Yeast strain AH109 harboring BD-PtSOS3 fusion in
pGBKT7 vector (or pGBKT7 empty vector) was transformed with
AD-PtSOS2, PtSOS2DF or PtSOS2TD fusion in pGADT7 vector (or
pGADT7 empty vector). The interaction was monitored by the
ability of yeast transformants to grow on selective medium minus
Leu, Trp, His and Ade, or by b-galactosidase assay on a piece of
filter paper. (b) Recruitment of PtSOS2 to the plasma membrane by
PtSOS3. The yeast cdc25-2 mutant was transformed with the plasmid
combinations as indicated on the left. Three microliters of serial
decimal dilutions were spotted onto YPD plates at the permissive
(24�C) or at the restrictive (36�C) temperature. Cells expressing
RAS-PtSOS2 in combination with PtSOS3 or RAS-AtSOS2 with
AtSOS3 combination were able to grow at 36�C, indicating
interaction between the two proteins at the plasma membrane.
(c) PtSOS2 interacted with PtSOS3 in planta. Empty YFP construct
(as a positive control) or different combinations of the N-YFP and
C-YFP fusion constructs as indicated were co-transfected into the
tobacco (N. benthamiana) leaves. The images were collected under
the Zeiss confocal microscope. The left panels depict the fluores-
cence in intact leaf epidermal cells (together with bright-field images
on the right), and the right panels exhibit the fluorescence signals in
protoplasts prepared from the same leaf (together with bright-field
images on the right)
Fig. 6 Reconstitution of poplar SOS pathway in yeast cells. The
yeast B31 (Dena1-4, Dnha1) strain transformed with an empty vector
or expressing the indicated combination of Populus SOS genes were
grown overnight in liquid selective medium. Three microliters of
serial decimal dilutions were spotted onto plates of arginine
phosphate (AP) medium with 1 mM KCl and supplemented with 0,
120 and 240 mM NaCl. Plates were incubated at 30�C for 60 h. The
Arabidopsis SOS1 gene AtSOS1 was used as a positive control.
Plasmids used for expression of SOS genes were described in
‘‘Materials and methods’’
Fig. 4 Functional complementation of Arabidopsis sos1, sos2 and
sos3 mutants by poplar PtSOS1, PtSOS2 and PtSOS3, respectively.
a Expression of PtSOS1, 2, 3 in each sos mutant as indicated. RT–
PCR products were amplified from seedlings of wild type, sosmutants and four corresponding complementary lines. The Arabid-opsis ACTIN2 gene was used as an internal control. b Four-day-old
seedlings of wild-type, sos mutants and complementary lines CS12,
CS16, CS26, CS29, CS35, CS36 were transferred to MS medium
containing 0, 60 and 120 mM NaCl, respectively. The pictures were
taken after 8 days of treatment. Seedling fresh weight c–e and root
length f–h were measured at day 8 after the transfer. Error barsrepresent SD (n = 10)
b
Plant Mol Biol (2010) 74:367–380 375
123

proteins in the B31 background. As depicted in Fig. 6,
expression of PtSOS1 alone only marginally elevated
halotolerance of yeast cells. Additional expression of
PtSOS2, but not PtSOS3, enabled the yeast strain to survive
better under moderate salinity stress (120 mM NaCl),
above levels imparted by SOS1 alone, but still failed to
afford yeast growth in high saline condition (240 mM
NaCl). Concurrent expression of three poplar SOS com-
ponents PtSOS1, PtSOS2 and PtSOS3 dramatically aug-
mented the salt tolerance of yeast cells, capable to sustain
growth in AP medium containing up to 240 mM NaCl.
This enhancement in salt tolerance was observed only in
the presence of PtSOS1, excluding the possibility that the
PtSOS2–PtSOS3 complex was unmasking endogenous
yeast activity. PtSOS2–PtSOS3 complex was also able to
activate AtSOS1 (as a positive control in our experiment),
with even greater efficacy, suggesting that the regulation of
SOS1 activity might be evolutionally conserved between
woody and herbaceous plant species.
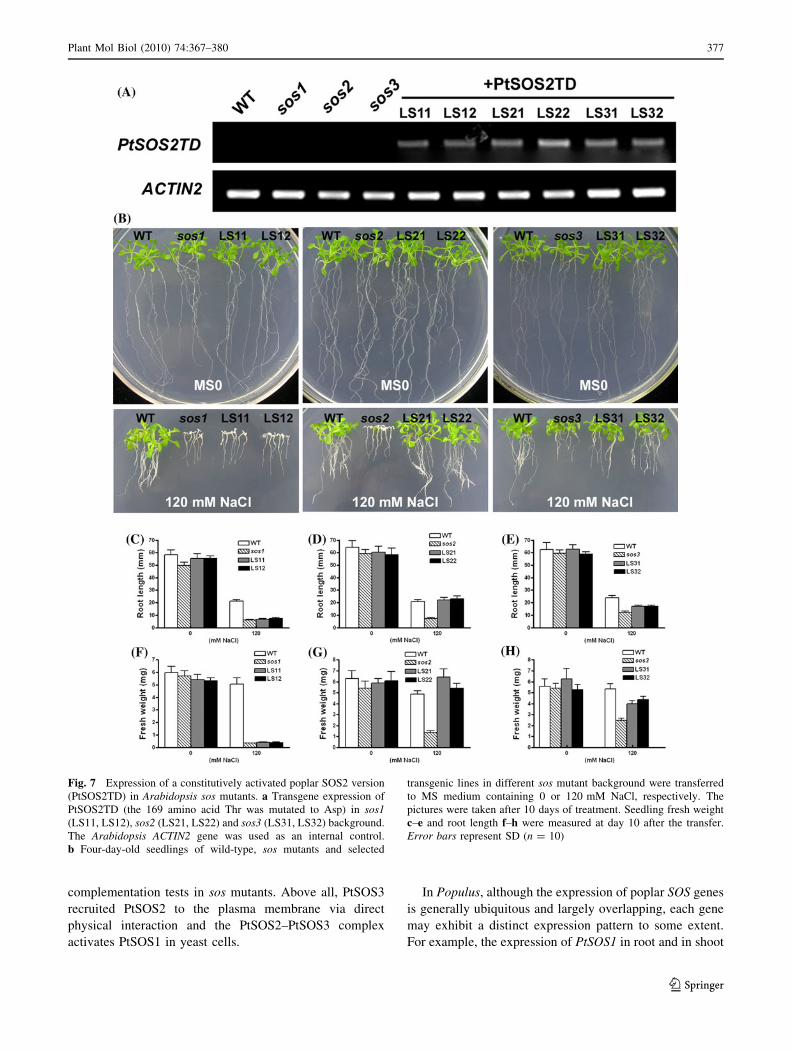
Expression of Constitutively Active PtSOS2 (PtSOS2TD)
complemented salt-sensitive phenotype of sos2 mutant
entirely, and that of sos3 partially, but did not rescue salt
hypersensitivity of sos1 at all.
Physical interaction and yeast reconstitution experi-
ments in this study have demonstrated that the putative
poplar SOS components should be working in the same
pathway as those in Arabidopsis. To further consolidate
this notion in planta, we tested whether PtSOS2 functions
dependent of SOS1 and downstream of SOS3 in salt
tolerance. We generated a constitutively active form of
PtSOS2 (designated as PtSOS2TD) by mutating the 169th
amino acid threonine (T) to aspirate (D) in the activation
loop of this kinase (Fig. S2). This single-point mutation in
SOS2 substantially increased the kinase activity (Guo
et al. 2001) but did not affect its interaction with SOS3
(Fig. 5a), resulting in a constitutively activated and
functional variant of SOS2 both in vitro and in vivo (Guo
et al. 2004). The Arabidopsis sos1, sos2 and sos3 mutants
were transformed with PtSOS2TD under the control of
cauliflower mosaic virus 35S promoter. By RT–PCR
assay using PtSOS2 specific primers, we identified 10–15
transgenic lines overexpressing PtSOS2TD in sos1, sos2
or sos3 mutant background and selected two representa-
tive lines of each exhibiting similar expression levels for
subsequent phenotypic analysis (Fig. 7a). Expression of
PtSOS2TD in sos1 background failed to rescue its salt
overly sensitive phenotype, as is shown in Fig. 7b. The
transgenic sos1 plants and sos1 parental mutants dis-
played equally severe growth defect on MS plates sup-
plemented with 120 mM NaCl (Fig. 7c, f). However,
transgenic sos2 plants harboring PtSOS2TD grew like the
wild-type plants upon high salt treatment (Fig. 7b, middle
panels). A statistical analysis of root length (Fig. 7d) and
fresh weight (Fig. 7g) further indicated that salt hyper-
sensitivity of sos2 was fully rescued by constitutive
expression of the poplar activated form of SOS2. When
expressed in sos3 background, PtSOS2TD partially sup-
pressed the salt-sensitive phenotype of sos3 mutants
(Fig. 7b, right panels). The sos3 transformants harboring
PtSOS2TD grew bigger than sos3 mutants on saline
medium, as revealed by partially recovered fresh weight
(Fig. 7h) and less inhibited root length (Fig. 7e). Taken
together, the enhanced activity or functionality of PtSOS2
is not sufficient for salt tolerance in the absence of SOS1
but somewhat bypass the SOS3 deficiency in planta. By
virtue of strong conservation between Populus and Ara-
bidopsis SOS proteins, the functional complementation
tests here could imply that in poplar trees, PtSOS2 might
function downstream of PtSOS3 and upstream of PtSOS1,
as well.
Discussion
Adaptation to high salinity is one of the important concerns
for woody perennials during their lifespan and over evo-
lutionary timescales. The molecular mechanisms underly-
ing salt tolerance and salt stress signaling are poorly known
in woody plants. With the completion of genome
sequencing in Populus trichocarpa (Tuskan et al. 2006)
and rapidly developing genomic tools, a handful of genes
related to salt stress tolerance (Ottow et al. 2005; Wu et al.
2007; Wang et al. 2008; Martin et al. 2009; Chen et al.
2009; Ye et al. 2009) were isolated and characterized from
the genus Populus that has become the accepted model for
perennial trees (Jansson and Douglas 2007). However,
most of those studies were focused on the biological
function of individual genes, without interpretation of a
gene regulatory network leading to fine-tuning of adaptive
processes. In this report, for the first time, we characterized
the Populus SOS pathway by integrated analysis of three
component genes. Several lines of molecular evidence
support the conclusion that poplar plants have a functional
salt overly sensitive pathway orthologous to the well-doc-
umented Arabidopsis SOS pathway. First, the putative
PtSOS components share high similarity with AtSOS pro-
teins in structure implying their similar biochemical
properties. Second, the overlap in expression and subcel-
lular localization of three poplar SOS elements provides a
prerequisite for their coordinated actions in the same
pathway governing salt stress adaptation. Third, the poplar
SOS orthologues can substitute for the endogenous Ara-
bidopsis counterparts revealed by functional
376 Plant Mol Biol (2010) 74:367–380
123
complementation tests in sos mutants. Above all, PtSOS3
recruited PtSOS2 to the plasma membrane via direct
physical interaction and the PtSOS2–PtSOS3 complex
activates PtSOS1 in yeast cells.
In Populus, although the expression of poplar SOS genes
is generally ubiquitous and largely overlapping, each gene
may exhibit a distinct expression pattern to some extent.
For example, the expression of PtSOS1 in root and in shoot
Fig. 7 Expression of a constitutively activated poplar SOS2 version
(PtSOS2TD) in Arabidopsis sos mutants. a Transgene expression of
PtSOS2TD (the 169 amino acid Thr was mutated to Asp) in sos1(LS11, LS12), sos2 (LS21, LS22) and sos3 (LS31, LS32) background.
The Arabidopsis ACTIN2 gene was used as an internal control.
b Four-day-old seedlings of wild-type, sos mutants and selected
transgenic lines in different sos mutant background were transferred
to MS medium containing 0 or 120 mM NaCl, respectively. The
pictures were taken after 10 days of treatment. Seedling fresh weight
c–e and root length f–h were measured at day 10 after the transfer.
Error bars represent SD (n = 10)
Plant Mol Biol (2010) 74:367–380 377
123

was comparable, whereas PtSOS2 was preferentially
expressed in green tissues rather than roots (Fig. 1). In
Populus alba 9 P. Berolinensis, the expression of Pab-
SOS1 and PabSOS2 was rapidly induced upon salt treat-
ment, with a differential manner in root and in shoot
(Fig. 2). It seemed that the salt-induced up-regulation of
PabSOS1 and PabSOS2 had a long-term effect in shoots
with moderate levels, but was more responsive and tran-
sient in roots. The simultaneous induction of PabSOS1 and
PabSOS2 transcripts by salinity also suggested a fine-tuned
regulation of SOS elements at the gene expression level.
However, the expression of PabSOS3, the upstream com-
ponent of SOS pathway, was not responsive to salt treat-
ment in our test, either in root (Fig. 2a) or in shoot
(Fig. 2b), presumably because SOS3 senses calcium sig-
nals and triggers downstream signaling primarily at protein
modification but not the transcription level. We also
noticed that the transcript abundance of PtSOS3 was pretty
high in root but remarkably diminished in the aerial parts
(Fig. 1). This is consistent with the root-specific expression
of AtSOS3 in Arabidopsis (Quan et al. 2007). On the other
hand, the SOS3-like calcium binding protein 8 (SCaBP8)/
CBL10 was recently identified as a shoot-specific player in
the SOS pathway (Quan et al. 2007). Like SOS3, it
recruited SOS2 to the plasma membrane, but uniquely
underwent phosphorylation by SOS2, leading to stabilized
SCaBP8–SOS2 complex for SOS1 activation (Lin et al.
2009). Interestingly, we found a SCaBP8 homolog existing
in Populus genome to be predominantly expressed in the
above-ground tissues (unpublished data). This fact may
implicate a tissue-differential regulation of salt response in
the poplar SOS pathway.
The sodium efflux protein SOS1 functions as a pivotal
effector molecule in the SOS pathway, and it has been
suggested to fulfill several facets of important roles per-
taining to salt tolerance. To begin with, at the cellular level,
SOS1 would act extruding the excessive Na? to prevent Na?
accumulation in the cytoplasm, as it has been shown in
Arabidopsis (Qiu et al. 2003), rice (Martinez-Atienza et al.
2007), wheat (Xu et al. 2008), Thellungiella (Oh et al. 2009)
and tomato (Olıas et al. 2009). In Populus, the salt-tolerant
genotype Populus euphratica exhibited a higher transcript
abundance of genes related to Na?/H? antiport compared to
the salt-sensitive P. popularis (Ding et al. 2010). In a recent
study based on the scanning ion-selective electrode tech-
nique, it was concluded that Na? extrusion in stressed
Populus euphratica roots was the result of an active Na?/H?
antiport across the plasma membrane (Sun et al. 2009),
which might be mediated by PeSOS1 reported previously
(Wu et al. 2007). In addition, at the whole-plant level, it was
suggested that SOS1 controls long-distance Na? transport
(Shi et al. 2002) and affects Na? partitioning between plant
organs (Olıas et al. 2009). These Na? distribution processes
would be particularly important for woody poplars because
of their big stature and long lifespan. A precise assessment
of possible contributions of PtSOS1 in this aspect awaits the
availability of RNAi lines and/or overexpressors of trans-
genic poplars. Furthermore, SOS1 takes part in the cross-
talk between the ion-homesotasis and oxidative-stress sig-
naling pathways. It was reported that stress-induced SOS1
mRNA stability in Arabidopsis was mediated by reactive
oxygen species (ROS) and SOS1 played negative roles in
tolerance to oxidative stress (Chung et al. 2008). SOS1
interacted with RCD1, a regulator of radical-based signal-
ing, through the long C-terminal cytoplasmic tail, further
suggesting its involvement in regulating oxidative-stress
responses (Katiyar-Agarwal et al. 2006). In NaCl-stressed
Populus euphratica cells, H2O2 production and cytosolic
Ca2? signals were integrated to mediate K?/Na? homeo-
stasis (Sun et al. 2010; Zhang et al. 2007). It needs further
experimentation how the poplar SOS pathway functions in
such signaling cross-talk. Last but not least, by affecting
endocytosis under salinity conditions, SOS1 may also play
dynamic roles in vacuole integrity, membrane trafficking, as
well as pH homeostasis (Oh et al. 2009; Oh et al. 2010).
Further work on poplar SOS genes will be instructed by and
focused on all the above findings so as to elucidate func-
tional significance of SOS pathway in woody species. On the
other hand, engineering PtSOS genes identified in this study
offer good opportunities towards generation of transgenic
tree species with improved salt tolerance, which may be
grown on saline land to prevent erosion and potentially to
ameliorate soils (Singh 1998). At present, genetic transfor-
mation of poplar cultivars by overexpressing the three
PtSOS components is under way in our laboratory.
Acknowledgments We thank Dr. Jorg Kudla (Institut fur Botanik,
Universitat Munster, Germany) for providing the pSPYNE-35S and
pSPYCE-35S plasmids. We are also grateful to Dr. Ami Aronheim for
RRS materials and Dr. Lydie Maresova for the B31 yeast strain.
Special thanks would go to Dr. Rongmin Zhao (University of Tor-
onto, Canada) for his technical guidance on yeast experiments. This
work was supported by the following grants: the National Natural
Science Foundation of China (30872044), the National Basic
Research Program of China (2006CB100106), the National mega
project of GMO crops (2008ZX08001-003; 2008ZX08004-002;
2008ZX08010-004); Shanghai Science & Technology Development
Fund (0853Z111C1; 08DZ2270800).
References
Albinsky D, Masson JE, Bogucki A, Afsar K, Vass I, Nagy F,
Paszkowski J (1999) Plant responses to genotoxic stress are
linked to an ABA/salinity signaling pathway. Plant J 17:73–82
Albrecht V, Ritz O, Linder S, Harter K, Kudla J (2001) The NAF
domain defines a novel protein-protein interaction module
conserved in Ca2?-regulated kinases. EMBO J 20:1051–1063
378 Plant Mol Biol (2010) 74:367–380
123

Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,
Lipman DJ (1997) Gapped BLAST and PSI-BLAST, a new
generation of protein database search programs. Nucleic Acids
Res 25:3389–3402
Apse MP, Aharon GS, Snedden WS, Blumwald E (1999) Salt
tolerance conferred by overexpression of a vacuolar Na?/H?
antiport in Arabidopsis. Science 285:1256–1258
Banuelos MA, Sychrova H, Bleykasten-Grosshans C, Souciet JL,
Potier S (1998) The Nha1 antiporter of Saccharomyces cerevi-siae mediates sodium and potassium efflux. Microbiology
144:2749–2758
Blumwald E (2000) Sodium transport and salt tolerance in plants.
Curr Opin Cell Biol 12:431–434
Borsani O, Valpuesta V, Botella MA (2001) Evidence for a role of
salicylic acid in the oxidative damage generated by NaCl and
osmotic stress in Arabidopsis seedlings. Plant Physiol 126:
1024–1030
Broder YC, Katz S, Aronheim A (1998) The Ras recruitment system,
a novel approach to the study of protein–protein interactions.
Curr Biol 8:1121–1124
Capieaux E, Vignais ML, Setenac A, Goffeau A (1989) The yeast
H?-ATPase gene is controlled by the promoter binding factor
TUF. J Biol Chem 264:7437–7446
Chen Z, Pottosin II, Cuin TA, Fuglsang AT, Tester M, Jha D, Zepeda-
Jazo I, Zhou M, Palmgren MG, Newman IA, Shabala S (2007)
Root plasma membrane transporters controlling K?/Na? homeo-
stasis in salt stressed barley. Plant Physiol 145:1714–1725
Chen J, Xia X, Yin W (2009) Expression profiling and functional
characterization of a DREB2-type gene from Populus euphra-tica. Biochem Biophys Res Commun 378:483–487
Chung JS, Zhu JK, Bressan RA, Hasegawa PM, Shi H (2008)
Reactive oxygen species mediate Na?-induced SOS1 mRNA
stability in Arabidopsis. Plant J 53:554–565
Ding M, Hou P, Shen X, Wang M, Deng S, Sun J, Xiao F, Wang R,
Zhou X, Lu C, Zhang D, Zheng X, Hu Z, Chen S (2010) Salt-
induced expression of genes related to Na?/K? and ROS
homeostasis in leaves of salt-resistant and salt-sensitive poplar
species. Plant Mol Biol 73:251–269
Gong D, Guo Y, Schumaker KS, Zhu JK (2004) The SOS3 family of
calcium sensors and SOS2 family of protein kinases in
Arabidopsis. Plant Physiol 134:919–926
Guo Y, Halfter U, Ishitani M, Zhu JK (2001) Molecular character-
ization of functional domains in the protein kinase SOS2 that is
required for plant salt tolerance. Plant Cell 13:1383–1400
Guo Y, Qiu QS, Quintero FJ, Pardo JM, Ohta M, Zhang C,
Schumaker KS, Zhu JK (2004) Transgenic evaluation of
activated mutant alleles of SOS2 reveals a critical requirement
of its kinase activity and C-terminal regulatory domain for salt
tolerance in Arabidopsis. Plant Cell 16:435–449
Halfter U, Ishitani M, Zhu JK (2000) The Arabidopsis SOS2
protein kinase physically interacts with and is activated by the
calcium-binding protein SOS3. Proc Natl Acad Sci USA 97:
3735–3740
Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ (2000) Plant cellular
and molecular responses to high salinity. Annu Rev Plant Biol
51:463–499
Horie T, Schroeder JI (2004) Sodium transporters in plants. Diverse
genes and physiological functions. Plant Physiol 136:2457–2462
Ishitani M, Liu J, Halfter U, Kim CS, Shi W, Zhu JK (2000) SOS3
function in plant salt tolerance requires N-myristoylation and
calcium binding. Plant Cell 12:1667–1678
Jansson S, Douglas CJ (2007) Populus, a model system for plant
biology. Annu Rev Plant Biol 58:435–458
Katiyar-Agarwal S, Zhu J, Kim K, Agarwal M, Fu X, Huang A, Zhu
JK (2006) The plasma membrane Na?/H? antiporter SOS1
interacts with RCD1 and functions in oxidative stress tolerance
in Arabidopsis. Proc Natl Acad Sci USA 103:18816–18821
Kinclova O, Ramos J, Potier S, Sychrova H (2001) Functional study
of the Saccharomyces cerevisiae Nha1p C-terminus. Mol
Microbiol 40:656–668
Laurie S, Feeney KA, Maathuis FJM, Heard PJ, Brown SJ, Leigh RA
(2002) A role for HKT1 in sodium uptake by wheat roots. Plant J
32:139–149
Lin H, Yang Y, Quan R, Mendoza I, Wu Y, Du W, Zhao S,
Schumaker KS, Pardo JM, Guo Y (2009) Phosphorylation of
SOS3-LIKE CALCIUM BINDING PROTEIN8 by SOS2 protein
kinase stabilizes their protein complex and regulates salt
tolerance in Arabidopsis. Plant Cell 21:1607–1619
Liu J, Zhu JK (1998) A calcium sensor homolog required for plant
salt tolerance. Science 280:1943–1945
Liu J, Ishitani M, Halfter U, Kim CS, Zhu JK (2000) The Arabidopsisthaliana SOS2 gene encodes a protein kinase that is required for
salt tolerance. Proc Natl Acad Sci USA 97:3703–3734
Luo ZB, Polle A (2009) Wood composition and energy content in a
poplar short rotation plantation on fertilized agricultural land in a
future CO2 atmosphere. Glob Change Biol 15:38–47
Martin L, Leblanc-Fournier N, Azri W, Lenne C, Henry C, Coutand
C, Julien JL (2009) Characterization and expression analysis
under bending and other abiotic factors of PtaZFP2, a poplar
gene encoding a Cys2/His2 zinc finger protein. Tree Physiol 29:
125–136
Martinez-Atienza J, Jiang X, Garciadeblas B, Mendoza I, Zhu JK,
Pardo JM, Quintero FJ (2007) Conservation of the salt overly
sensitive pathway in rice. Plant Physiol 143:1001–1012
Mumberg D, Muller R, Funk M (1995) Yeast vectors for the
controlled expression of heterologous proteins in different
genetic backgrounds. Gene 156:119–122
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue cultures. Physiol Plant 15:473–495
Niu X, Bressan RA, Hasegawa PM, Pardo JM (1995) Ion homeostasis
in NaCl stress environments. Plant Physiol 109:735–742
Oh DH, Leidi E, Zhang Q, Hwang SM, Li Y, Quintero FJ, Jiang X,
D’Urzo MP, Lee SY, Zhao Y, Bahk JD, Bressan RA, Yun DJ,
Pardo JM, Bohnert HJ (2009) Loss of halophytism by interfer-
ence with SOS1 expression. Plant Physiol 151:210–222
Oh DH, Lee SY, Bressan RA, Yun DJ, Bohnert HJ (2010)
Intracellular consequences of SOS1 deficiency during salt stress.
J Exp Bot 61:1205–1213
Olıas R, Eljakaoui Z, Li J, De Morales PA, Marın-Manzano MC,
Pardo JM, Belver A (2009) The plasma membrane Na?/H?
antiporter SOS1 is essential for salt tolerance in tomato and
affects the partitioning of Na? between plant organs. Plant Cell
Environ 32:904–916
Ottow EA, Polle A, Brosche M, Kangasjarvi J, Dibrov P, Zorb C,
Teichmann T (2005) Molecular characterization of PeNhaD1,
the first member of the NhaD Na?/H? antiporter family of plant
origin. Plant Mol Biol 58:75–88
Parida A, DAS AB, Mittra B (2004) Effects of salt on growth, ion
accumulation, photosynthesis and leaf anatomy of the mangrove,
Bruguiera parviflora. Trees Struct Funct 108:167–174
Pilate G, Guiney E, Holt K, Petit-Conil M, Lapierre C, Leple JC,
Pollet B, Mila I, Webster EA, Marstorp HG, Hopkins DW,
Jouanin L, Boerjan W, Schuch W, Cornu D, Halpin C (2002)
Field and pulping performances of transgenic trees with altered
lignification. Nat Biotechnol 20:607–612
Qiu QS, Guo Y, Dietrich MA, Schumaker KS, Zhu JK (2002)
Regulation of SOS1, a plasma membrane Na?/H? exchanger in
Arabidopsis thaliana, by SOS2 and SOS3. Proc Natl Acad Sci
USA 99:8436–8441
Plant Mol Biol (2010) 74:367–380 379
123

Qiu QS, Barkla BJ, Vera-Estrella R, Zhu JK, Schumaker KS (2003)
Na?/H? exchange activity in the plasma membrane of Arabid-opsis. Plant Physiol 132:1041–1052
Quan R, Lin H, Mendoza I, Zhang Y, Cao W, Yang Y, Shang M,
Chen S, Pardo JM, Guo Y (2007) SCaBP8/CBL10, a putative
calcium sensor, interacts with the protein kinase SOS2 to protect
Arabidopsis shoots from salt stress. Plant Cell 19:1415–1431
Quintero FJ, Ohta M, Shi H, Zhu JK, Pardo JM (2002) Reconstitution
in yeast of the Arabidopsis SOS signaling pathway for Na?
homeostasis. Proc Natl Acad Sci USA 99:9061–9066
Rubio F, Gassmann W, Schroeder JI (1995) Sodium driven potassium
uptake by the plant potassium transporter HKT1 and mutations
conferring salt tolerance. Science 270:1660–1663
Sheen J (2001) Signal transduction in maize and Arabidopsismesophyll protoplasts. Plant Physiol 127:1466–1475
Shi H, Ishitani M, Kim C, Zhu JK (2000) The Arabidopsis thalianasalt tolerance gene SOS1 encodes a putative Na?/H? antiporter.
Proc Natl Acad Sci USA 97:6896–6901
Shi H, Quintero FJ, Pardo JM, Zhu JK (2002) The putative plasma
membrane Na?/H? antiporter SOS1 controls long-distance Na?
transport in plants. Plant Cell 14:465–477
Shi H, Lee BH, Wu SJ, Zhu JK (2003) Overexpression of a plasma
membrane Na?/H? antiporter gene improves salt tolerance in
Arabidopsis thaliana. Nat Biotechnol 21:81–85
Singh B (1998) Biomass production and nutrient dynamics in three
clones of Populus deltoides planted on Indogangetic plains. Plant
Soil 203:15–26
Sun J, Chen S, Dai S, Wang R, Li N, Shen X, Zhou X, Lu C, Zheng X,
Hu Z, Zhang Z, Song J, Xu Y (2009) NaCl-induced alternations
of cellular and tissue ion fluxes in roots of salt-resistant and salt-
sensitive poplar species. Plant Physiol 149:1141–1153
Sun J, Wang MJ, Ding MQ, Deng SR, Liu MQ, Lu CF, Zhou XY,
Shen X, Zheng XJ, Zhang ZK, Song J, Hu ZM, Xu Y, Chen SL
(2010) H2O2 and cytosolic Ca2? signals triggered by the PM
H?-coupled transport system mediate K?/Na? homeostasis in
NaCl-stressed Populus euphratica cells. Plant Cell Environ
33:943–958
Tuskan GA et al (2006) The genome of black cottonwood, Populustrichocarpa (Torr., Gray). Science 313:1596–1604
Volkov V, Wang B, Dominy PJ, Fricke W, Amtmann A (2004)
Thellungiella halophila, a salt-tolerant relative of Arabidopsisthaliana, possesses effective mechanisms to discriminate
between potassium and sodium. Plant Cell Environ 27:1–14
Walter M, Chaban C, Schutze K, Batistic O, Weckermann K, Nake C,
Blazevic D, Grefen C, Schumacher K, Oecking C, Harter K,
Kudla J (2004) Visualization of protein interactions in living
plant cells using bimolecular fluorescence complementation.
Plant J 40:428–438
Wang JY, Xia XL, Wang JP, Yin WL (2008) Stress responsive zinc-
finger protein gene of Populus euphratica in tobacco enhances
salt tolerance. J Integr Plant Biol 50:56–61
Wu Y, Ding N, Zhao X, Zhao M, Chang Z, Liu J, Zhang L (2007)
Molecular characterization of PeSOS1, the putative Na?/H?
antiporter of Populus euphratica. Plant Mol Biol 65:1–11
Xiong LM, Schumaker KS, Zhu JK (2002) Cell signaling during cold,
drought and salt stress. Plant Cell 14 suppl:S165–S183
Xu H, Jiang X, Zhan K, Cheng X, Chen X, Pardo JM, Cui D (2008)
Functional characterization of a wheat plasma membrane Na?/
H? antiporter in yeast. Arch Biochem Biophys 473:8–15
Yang Q, Chen ZZ, Zhou XF, Yin HB, Li X, Xin XF, Hong XH, Zhu
JK, Gong Z (2009) Overexpression of SOS (salt overly sensitive)
genes increases salt tolerance in transgenic Arabidopsis. Mol
Plant 2:22–31
Ye CY, Zhang HC, Chen JH, Xia XL, Yin WL (2009) Molecular
characterization of putative vacuolar NHX-type Na?/H?
exchanger genes from the salt-resistant tree Populus euphratica.
Physiol Plant 137:166–174
Yoo SD, Cho YH, Sheen J (2007) Arabidopsis mesophyll protoplasts:
a versatile cell system for transient gene expression analysis. Nat
Protoc 2:1565–1572
Zhang F, Wang Y, Yang Y, Wu H, Wang D, Liu J (2007)
Involvement of hydrogen peroxide and nitric oxide in salt
tolerance resistance in the calluses from Populus euphratica.
Plant Cell Environ 30:775–785
Zhu JK (2001) Plant salt tolerance. Trends Plant Sci 6:66–71
Zhu JK (2002) Salt and drought stress signal transduction in plants.
Annu Rev Plant Biol 53:247–273
Zhu JK (2003) Regulation of ion homeostasis under salt stress. Curr
Opin Plant Biol 6:441–445
380 Plant Mol Biol (2010) 74:367–380
123




















