
Alleviation of nickel toxicity in wheat (Triticum aestivum L.) seedlings ...
Upload
independentCategory
view
0download
0
The transcript composition of egg cells changes significantlyfollowing fertilization in wheat (Triticum aestivum L.)
Stefanie Sprunck1, Ute Baumann2, Keith Edwards3, Peter Langridge2 and Thomas Dresselhaus1,*
1Developmental Biology and Biotechnology, Biocenter Klein Flottbek, University of Hamburg, Ohnhorststrasse 18, D-22609
Hamburg, Germany,2Australian Center for Plant Functional Genomics, University of Adelaide,Waite Campus, Hartley Grove, SA 5064, Australia, and3Department of Biological Sciences, University of Bristol, Woodland Road, Bristol BS8 1UG, UK
Received 1 November 2004; accepted 23 November 2004.*For correspondence (fax þ49 40 42816 229; e-mail [email protected]).
Summary
Here, we report the transcript profile of wheat egg cells and proembryos, just after the first cell division.
Microdissected female gametophytes of wheat were used to isolate eggs and two-celled proembryos to
construct cell type-specific cDNA libraries. In total, 1197 expressed sequence tags (ESTs) were generated.
Analysis of these ESTs revealed numerous novel transcripts. In egg cells, 17.6% of the clustered ESTs
represented novel transcripts, while 11.4% novel clusters were identified in the two-celled proembryo.
Functional classification of sequences with similarity to previously characterized proteins indicates that the
unfertilized egg cell has a higher metabolic activity and protein turnover than previously thought. Transcript
composition of two-celled proembryos was significantly distinct from egg cells, reflecting DNA replication as
well as high transcriptional and translational activity. Several novel transcripts of the egg cell are specific for
this cell. In contrast, some fertilization induced novel mRNAs are abundant also in sporophytic tissues
indicating a more general role in plant growth and development. The potential functions of genes based on
similarity to known genes involved in developmental processes are discussed. Our analysis has identified
numerous genes with potential roles in embryo sac function such as signaling, fertilization or induction of
embryogenesis.
Keywords: egg cell, fertilization, embryogenesis, genomics, wheat.
Introduction
Fertilization leads to a remarkable biologic phenomenon:
the process of embryogenesis. Once activated, the ferti-
lized egg cell starts to divide and progeny cells undergo a
series of specification and differentiation steps, finally
generating a new organism consisting of different cell
types, tissues and organs. The molecular mechanisms of
fertilization and early embryogenesis have been most
intensively studied in animals. In most of the species
studied, maternally stored mRNAs of the egg were iden-
tified as involved in the establishment of embryonic axes,
diversification of cell types and morphological changes
during early embryogenesis (Angerer and Angerer, 2000;
Mohr et al., 2001; Nishida, 1997; Pellettieri and Seydoux,
2002; St Johnston, 1995; Wylie et al., 1996). The transition
from maternal to zygotic/embryonic control of gene
expression (ZGA/EGA) is the first major transition that
occurs following fertilization. Thereafter, transcript profiles
dynamically change throughout embryogenesis (Baugh
et al., 2003; Makabe et al., 2001). Nevertheless, the time
point of transition appears to be species-dependent: ZGA/
EGA occurs at the two-celled stage in mice (Adenot et al.,
1997), the four- to eight-celled stage in cows and humans
and the eight- to 16-celled stage in sheep and rabbits
(Kanka et al., 2003; Nothias et al., 1995). In Caenorhabditis,
the first embryonic transcription is detected in blastomeres
at the four-celled stage (Edgar et al., 1994), while major
embryonic transcription in zebra fish and Xenopus occurs
after 10–12 cell cycles at the beginning of the midblastula
transition (Kane and Kimmel, 1993; Nakakura et al., 1987;
Newport and Kirschner, 1982).
Compared with animals, little is known about transcripts
stored in egg cells of flowering seed plants (angiosperms)
660 ª 2005 Blackwell Publishing Ltd
The Plant Journal (2005) 41, 660–672 doi: 10.1111/j.1365-313X.2005.02332.x

and activation of the zygotic genome after fertilization.
Here, the egg cell is part of the female gametophyte
(embryo sac) which consists of four different cell types: the
egg cell, two synergids, the large central cell and some
antipodal cells (Yadegari and Drews, 2004). During double
fertilization, a phenomenon unique to angiosperms, the
two female gametes (egg and central cell) each fuse with
one sperm cell delivered by the pollen tube. They then
develop into the embryo and endosperm, respectively. As
the embryo sac is typically surrounded by sporophytic
tissues of the ovule and ovary, access to female gametes
and very early embryo and endosperm stages is ham-
pered. Gametic and zygotic transcripts of seed plants are
thus poorly represented in current databases of expressed
sequence tags (ESTs) although some have been generated
through sequencing from cDNA libraries produced from
whole floral tissues. However, the use of these complex
tissues results in under representation of genes expressed
at low levels and in only one or a few cell types. Methods
for the isolation of single cells from the embryo sac of
maize, barley, wheat and rice have been described (Holm
et al., 1994; Kranz et al., 1991; Kumlehn et al., 1998; Zhao
et al., 2000) and the generation of cDNA libraries
from maize and wheat egg cells have been reported
(Dresselhaus et al., 1994; Kumlehn et al., 2001), but our
knowledge of transcript compositions of eggs, zygotes and
proembryos is still limited. Only a few genes have been
reported to be expressed in egg cells of maize and Arabid-
opsis, some of which alter their expression pattern after
fertilization (e.g. Cordts et al., 2001; Dresselhaus et al.,
1999a; Haecker et al., 2004; Heuer et al., 2001; Perry et al.,
1996). Analysis of cell cycle regulatory genes and ribosomal
genes in maize indicated that plant zygotic gene activation
occurs earlier compared with the animal systems investi-
gated so far (Dresselhaus et al., 1999b1 ; Sauter et al., 1998).
To develop a broad picture of the genes involved in the
development and function of female gametophyte cells,
we used a genomics-based approach involving the gen-
eration of ESTs from cell type-specific cDNA libraries.
Applying an experimental technique to microdissect
wheat embryo sac cells (Kumlehn et al., 1998), we isola-
ted single cells of the unfertilized embryo sac as well as
defined stages of zygotes and early embryos. We gener-
ated stage-specific cDNA libraries for the egg cells and
the proembryos, which have completed the first zygotic
cell division. Randomly selected clones from these librar-
ies were sequenced and analyzed. Results obtained from
EST analyses, including functional classification and
comparative studies on the transcript composition of the
egg cell and two-celled proembryo, as well as expression
studies of selected genes are shown. The function of
putative regulators of various processes related to
double fertilization and embryonic genome activation are
discussed.
Results
Isolation and cultivation of single cells from the female
gametophyte of wheat
In order to obtain single gametes and zygotes of defined
developmental stages, an experimental system based on a
microdissectionmethod to isolate embryo sac cells of wheat
(Kumlehn et al., 1998) was established. The isolation pro-
cedure resulted in intact and viable cells free of maternal
tissues (Figure 1c–h). Egg cells and early zygotes were iso-
lated mechanically from ovaries without using cell wall-
degrading enzymes. The isolation procedure is rapid and
helps in minimizing cellular responses triggered by enzy-
matic cell wall degradation of the embryo sac and the sur-
rounding nucellus cells. Wheat ovary tips were dissected at
the micropylar region, either at the time of flowering using
previously emasculated spikelets (Figure 1a), or 4–6 h after
hand-pollination (hap). Ovular tissue containing the egg
apparatus (Figure 1b) was excised in 0.55 M mannitol solu-
tion and egg cells (Figure 1c) or zygotes (Figure 1d) were
released from the ovule tips using fine glass needles. Col-
lected cells were either directly frozen for later mRNA
isolation or kept in mannitol to monitor zygotic embryo
development in vitro. The nucleus of the mature egg cell is
surrounded by dense cytoplasm and typically contains one
large nucleolus (Figure 1c; arrow). Around 13 hap zygotes
were clearly distinguished from unfertilized egg cells by the
occurrence of a second nucleolus (Figure 1d). Around
22 hap the first zygotic cell division was observed (Fig-
ure 1e) and two-celled proembryos were visible 23–24 hap
(Figure 1f). Isolated zygotes autonomously undergo the first
cell divisions, without adding supplements such as plant
hormones, conditionedmedium or feeder cells (Figure 1f,g).
The microdissection technique described here was also
successful for isolating in vivo embryos as early as
2–3 days after fertilization (Figure 1h), giving the possibility
of analyzing gene expression in very young wheat
embryos.
Bioinformatic analysis of ESTs
Unfertilized egg cells and two-celled proembryos were used
to generate cDNA libraries. Subsequently, single-run partial
sequencing of randomly selected cDNA clones (ESTs) was
performed. As cDNAs were cloned in a non-directional
manner, both 5¢ and 3¢ read sequenceswere generated. After
DNA sequencer trace data passed an automated cleanup
pipeline, a total of 1197 ESTs were used for analysis. The
sequences of each library were clustered and assembled
into contigs. Thus, 735 ESTs from the egg cell form 404
independent clusters including 310 singletons (77%), while
462 sequences from the two-celled proembryo generated
298 clusters, containing 236 singletons (79%).
Transcriptome of wheat eggs and proembryos 661
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

The consensus sequences of the clusters were used for
BLASTN and BLASTX searches at the NCBI non-redundant
database (nr), dbEST and SwissProt database. In addition,
we performed BLASTN searches against the TIGR-assembled
wheat genes (Quackenbush et al., 20012 ). BLASTN searches
against the NCBI database category of non-mouse and non-
human ESTs resulted in 629 egg cell ESTs (333 clusters) and
418 two-celled proembryo ESTs (264 clusters) matching
significantly to annotated ESTs mainly generated from
different tissues of wheat, barley or rice (Figure 2, NCBI
dbEST Poaceae). Among these, only 276 ESTs represent
genes present in both, the egg cell and the two-celled
proembryo. Interestingly, 106 egg cell ESTs (71 clusters) and
44 proembryo ESTs (34 clusters) did not match annotated
ESTs and were thus considered as ‘novel’ transcripts. Seven
of these ‘novel’ transcripts were common to the egg cell and
early embryo (Figure 2). In total, a surprisingly small number
of 23.6% of all ESTs (representing 5.4% of all clusters) were
present in both egg cell and two-celled proembryo.
Clusters were further categorized according to their
BLASTX and BLASTN search results into (i) transcripts
encodingproteinsof knownfunction (similar to characterized
proteins), (ii) transcripts of unknown function, present in
dbEST only, (iii) transcripts encoding hypothetical proteins
[similar to uncharacterized genes, such as predicted open-
reading frames (ORFs), unknown proteins, ‘similar to’ and
‘-like’ proteins] showing additional similarity to published
ESTs or cDNAs, (iv) transcripts encoding hypothetical pro-
teins (similar to computer-predictedORFs) without similarity
to any EST or cDNA and (v) novel transcripts of unknown
function (Figure 3). Classification of sequences derived from
the egg cell revealed that 177 of the 404 clusters (43.8%)
represent characterized proteins, while the remaining 56.2%
represent genes of unknown function. Among these, 69
clusters (17.1%) showed a match to annotated ESTs only
(Figure 3a). Significant similarities to hypothetical proteins
predicted from the Arabidopsis or rice genome sequences
were identified for 98 egg cell clusters (24.2%). Among these,
stigma
micropyle
(a) (b) (c) (d)
(e) (f) (g) (h)
Figure 1. Isolation of egg cells and zygotes from the female gametophyte of wheat and the development of zygotes into proembryos in culture.
(a) Mature unpollinated ovary, the arrow indicates direction of section.
(b) Isolated ovule tip showing the egg apparatus consisting of egg cell (arrow) and two flanking synergids (open triangles).
(c) Egg cell containing one nucleolus (arrow).
(d) Two zygotes, 13 hap, each containing two nucleoli (arrows).
(e) Cytokinesis of first zygotic cell division, appearing 20 hap.
(f) Two-celled proembryo, 22 hap.
(g) Four-celled proembryo, 30 hap.
(h) Isolated embryo, 3 days after pollination (bars: 25 lm; hap, hours after pollination).
2-celled proembryoEgg cell
NCBI dbEST Poaceae(>1.5 × 106)
284487
102 41
276
7
Figure 2. Distribution of 735 ESTs derived from egg cells (blue) and 462 ESTs
from the two-celled proembryos (red) among each other and the Poaceae EST
database (yellow), containing more than 1.5 Mio. ESTs (as of March 2004).
662 Stefanie Sprunck et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

11 hypothetical proteins (2.7%) are novel as they represent
computer-predicted ORFs with no match to annotated ESTs
ormRNAsequences.Moreover, 60 clusters (14.9%) represent
completely novel transcripts lacking any similarity to anno-
tated genes, ESTs or proteins. Taken together, 17.6% of all
clusters from the egg cell can be considered as ‘novel.’
The same classificationwas conducted for the 298 clusters
derived from the two-celled proembryo. Here, 163 clusters
(54.7%) significantly matched characterized proteins (Fig-
ure 3b). Moreover, 47 clusters (15.8%)matched to annotated
ESTs only, while 54 clusters (18.1%) of the two-celled
proembryo ESTs showed significant similarities to hypo-
thetical proteins of Arabidopsis or rice. All clusters with
similarity to predicted proteins revealed additional matches
in the Poaceae dbEST. Finally, 34 clusters (11.4%) represent
‘novel’ sequences.
Composition and dynamics of the egg cell and proembryo
transcriptome
Independent clusters derived from egg cells and pro-
embryos were sorted according to decreasing cluster size.
The 20 largest clusters of egg cell and two-celled proembryo
sequences are shown in Table 1a,b, respectively. Largest
egg cell clusters are quite distinct compared with those from
the two-celled proembryo.
Egg cell. The largest egg cell cluster is formed by ESTs
showing sequence similarity to ECA1, a cDNA isolated from
barley microspore cultures (Vrinten et al., 1999). Although
this is the largest cluster in egg cells (6.67% of ESTs;
Table 1a), TaECA1-like ESTs were not identified among the
two-celled proembryo ESTs. The second largest egg cell
cluster shows similarity to cyclophilin A1 (CyP1; Cluster EC-
2). In general, ESTs encoding CyP1 are highly represented in
several wheat cDNA libraries generated from different tissue
types (UniGene Cluster Ta.1078; NCBI). In the egg cell, 4.6%
of ESTs encode CyP1. This number decreases to 1% in the
two-celled proembryo (Table 1b; Cluster 2C-1524). The third
largest cluster of the egg cell is formed by ESTs encoding a
cytosolic ascorbate peroxidase (Cluster EC-3). However,
similar ESTs were not identified among the largest cluster of
the two-celled proembryo. Transcripts encoding histones
and ribosomal proteins were less strongly represented
among the most abundant classes of egg cell ESTs, com-
paredwith the two-celled proembryo. Nevertheless, the core
histones H4 (Cluster EC-6) and H2A (Cluster EC-22) as well as
the histone variant H3.3 (Cluster EC-9) are present (Table 1a).
Several ESTs with similarity to ubiquitin and ubiquitin-like
proteins are found among the most prevalent transcripts
(Clusters EC-5, EC-7, EC-16, EC-17, EC-57). In particular, ESTs
with similarity to rice ubiquitin-like smt3 protein (Cluster EC-
17) and barley ubiquitin-tail fusion protein 1 (Cluster EC-7)
are abundant in egg cells, but missing entirely in ESTs
derived from the two-celled proembryos. The same was
found for ESTs with similarity to small heat shock proteins
(sHSPs), sHSP16.9 and sHSP17.9: transcripts encoding these
sHSPs are abundant in the egg cell (Clusters EC-10, EC-430)
but not in the two-celled proembryo. We also found novel
sequences among the largest clusters of the egg cell: four
clusters show similarity to predicted hypothetical proteins
from rice and Arabidopsis (Clusters EC-4, EC-16, EC-18,
EC-57), while Cluster EC-12 represents a novel gene.
Two-celled proembryo. The largest clusters of the two-cel-
led proembryo are generated by ESTs encoding the histones
H4, H3 and H2B-6 (Table 1b; Clusters 2C-1520, 2C-1521,
2C-1522). Another seven clusters represent various ribo-
somal proteins (Clusters 2C-1523, 2C-1528, 2C-1529,
2C-1531, 2C-1532, 2C-1537, 2C-1539). In particular, tran-
scripts of the ribosomal protein L39 (RPL39; Cluster 2C-1523)
are abundant in the two-celled proembryo. Other transcripts
encode a 12-oxophytodienoic acid reductase involved in
jasmonic acid synthesis (Cluster 2C-1527), and a putative
calmodulin-binding protein (Cluster 2C-1527). Among the
most prevalent transcripts are also four clusters where no
BLASTX hit was found (Clusters 2C-1525, 2C-1526, 2C-1533,
2C-1566).
(a) Egg cell (404 clusters) (b) 2-celled proembryo (298 clusters)
Similar to characterized proteins
Similar to sequences present in dbEST only
Novel transcripts
Similar to hypothetical proteins and ESTs
Similar to hypothetical proteins only
15.8%
54.7%
18.1%
11.4%17.1%
43.8%
21.5%
2.7%
14.9%
Figure 3. Categorization of contigs derived
from the egg cell (a) and two-celled proembryo
(b) cDNA libraries. In the egg cell, 60 contigs
(14.9%) represent novel transcripts. In addition,
11 contigs (2.7%) show similarity to predicted
proteins but no similarity to any expressed
sequence. In the two-celled proembryo, 34 con-
tigs (11.4%) represent novel transcripts, showing
nomatch to published sequences. The portion of
sequences with similarity to characterized pro-
teins is significantly higher in the two-celled
proembryo (54.7%) compared with the egg cell
(43.8%).
Transcriptome of wheat eggs and proembryos 663
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

Comparative functional classification of ESTs encoding
characterized proteins
Sequences encoding characterized proteins were further
classified into functional groups to compare the status of the
unfertilized egg with that of the two-celled proembryo. In
total, 359 clusters (51%) of both cell types containing 753
ESTs revealed significant matches to characterized proteins,
to proteins with putative functions or to proteins with known
functional domains. We defined 11 major functional categ-
ories as shown in Figure 4. However, several proteins can-
not be classified strictly into one of these categories because
their biologic role is also relevant to another functional
group.
Table 1 The 20 largest independent contigs derived from egg cells (a) and two-celled proembryos (b), sorted according to cluster size.Functional annotations of BLASTX results are given where significant similarity to characterized genes or proteins were found
Cluster ID Blast · resulta Accession Organism Number of ESTs
(a) Egg cellEC-1 ECA1-like gene familyb AAF23356 H. vulgare 49EC-2 Cyclophilin A-1 AAK49426 T. aestivum 34EC-3 Ascorbate peroxidase CAA06996 H. vulgare 19EC-6 Histone H4 P59258 T. aestivum 15EC-4 Hypothetical protein BAB44057 O. sativa 14EC-5 Polyubiquitin Q9VZL4 D. melanogaster 13EC-7 Ubiquitin/ribosomal protein CEP52 S33633 O. sativa 13EC-57 Hypothetical proteinc BAC83160 O. sativa 12EC-9 Histone H3.3 P59169 A. thaliana 10EC-10 16.9 kDa class I HSP P12810 T. aestivum 10EC-12 No hit – – 10EC-14 Ribosomal protein L39 CAA64728 Z. mays 8EC-15 Putative ribosomal protein S17 AAN61484 O. sativa 6EC-16 Hypothetical proteinc BAC83160 O. sativa 6EC-17 Ubiquitin-like smt3 protein P55857 O. sativa 6EC-18 Hypothetical proteind BAB07973 O. sativa 6EC-22 Histone H2A CAA64356 T. aestivum 5EC-24 Putative lipid transfer protein AAN05565 O. sativa 5EC-430 17.9 kDa class I HSP AAK54445 O. sativa 5EC-51 Pathogenesis-related protein CAA38223 Zea mays 4
(b) Two-celled proembryo2C-1520 Histone H4 P59258 T. aestivum 392C-1521 Histone H3 CAA25451 T. aestivum 182C-1522 Histone H2B-6 BAA07156 T. aestivum 122C-1523 Ribosomal protein L39 CAA64728 Z. mays 82C-1524 Cyclophilin A-1 AAK49426 T. aestivum 52C-1525 No hite – – 42C-1526 No hit – – 42C-1527 Putative calmodulin-binding proteinf CAE02429 O. sativa 42C-1529 Putative ribosomal protein S12 BAC20920 O. sativa 42C-1537 Ribosomal protein S19 P40978 O. sativa 42C-1528 Ribosomal protein L18a Q943F3 O. sativa 32C-1531 Putative acidic ribosomal protein P2A BAA99431 O. sativa 32C-1532 Putative ribosomal protein S5 BAB64234 O. sativa 32C-1533 No hit – – 32C-1534 Histone H2A AAL40108 T. aestivum 32C-1535 Polyubiquitin S20925 Z. mays 32C-1536 12-oxophytodienoic acid reductase BAC20139 O. sativa 32C-1538 Putative snRNP polypeptide G O82221 A. thaliana 32C-1539 Putative ribosomal protein L37a BAB90039 O. sativa 32C-1566 No hit – – 3
aNCBI database nr.bContig comprises transcripts of at least five different members of a gene family with similarity to ECA1 from barley.cHypothetical protein contains conserved domain COG5227 (NCBI Conserved Domain Summary) of smt3 ubiquitin-like proteins.dHypothetical protein contains conserved domain pfam 00026 of aspartyl protease and is predicted to be extracellular located (SignalP).eCluster shows similarity to members of UniGene Cluster Ta.896 transcribed sequences which have moderate similarity to protein NP_565058(Arabidopsis) containing the domain KOG2315, a predicted translation initiation factor related to eIF-3a.f2C-1527 matches to UniGene Cluster Ta.9469 which has moderate similarity to a calmodulin-binding protein from Arabidopsis (NP_565441).
664 Stefanie Sprunck et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672
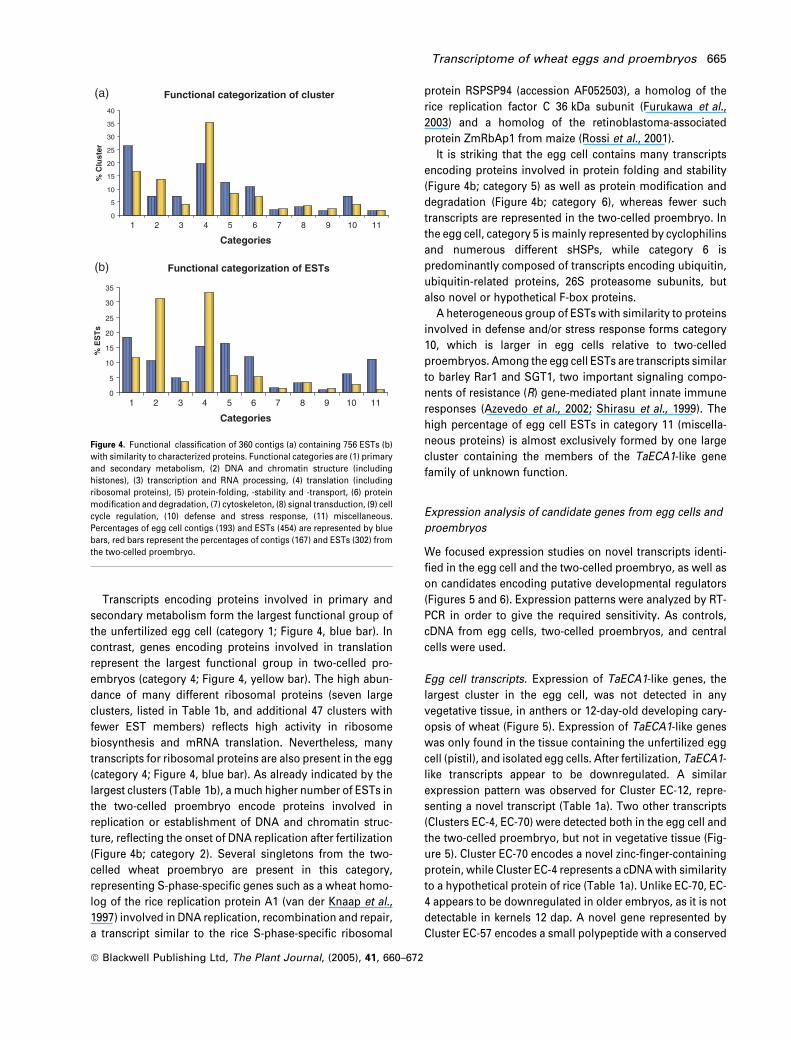
Transcripts encoding proteins involved in primary and
secondary metabolism form the largest functional group of
the unfertilized egg cell (category 1; Figure 4, blue bar). In
contrast, genes encoding proteins involved in translation
represent the largest functional group in two-celled pro-
embryos (category 4; Figure 4, yellow bar). The high abun-
dance of many different ribosomal proteins (seven large
clusters, listed in Table 1b, and additional 47 clusters with
fewer EST members) reflects high activity in ribosome
biosynthesis and mRNA translation. Nevertheless, many
transcripts for ribosomal proteins are also present in the egg
(category 4; Figure 4, blue bar). As already indicated by the
largest clusters (Table 1b), a much higher number of ESTs in
the two-celled proembryo encode proteins involved in
replication or establishment of DNA and chromatin struc-
ture, reflecting the onset of DNA replication after fertilization
(Figure 4b; category 2). Several singletons from the two-
celled wheat proembryo are present in this category,
representing S-phase-specific genes such as a wheat homo-
log of the rice replication protein A1 (van der Knaap et al.,
1997) involved in DNA replication, recombination and repair,
a transcript similar to the rice S-phase-specific ribosomal
protein RSPSP94 (accession AF052503), a homolog of the
rice replication factor C 36 kDa subunit (Furukawa et al.,
20033 ) and a homolog of the retinoblastoma-associated
protein ZmRbAp1 from maize (Rossi et al., 2001).
It is striking that the egg cell contains many transcripts
encoding proteins involved in protein folding and stability
(Figure 4b; category 5) as well as protein modification and
degradation (Figure 4b; category 6), whereas fewer such
transcripts are represented in the two-celled proembryo. In
the egg cell, category 5 ismainly represented by cyclophilins
and numerous different sHSPs, while category 6 is
predominantly composed of transcripts encoding ubiquitin,
ubiquitin-related proteins, 26S proteasome subunits, but
also novel or hypothetical F-box proteins.
A heterogeneous group of ESTs with similarity to proteins
involved in defense and/or stress response forms category
10, which is larger in egg cells relative to two-celled
proembryos. Among the egg cell ESTs are transcripts similar
to barley Rar1 and SGT1, two important signaling compo-
nents of resistance (R) gene-mediated plant innate immune
responses (Azevedo et al., 2002; Shirasu et al., 1999). The
high percentage of egg cell ESTs in category 11 (miscella-
neous proteins) is almost exclusively formed by one large
cluster containing the members of the TaECA1-like gene
family of unknown function.
Expression analysis of candidate genes from egg cells and
proembryos
We focused expression studies on novel transcripts identi-
fied in the egg cell and the two-celled proembryo, as well as
on candidates encoding putative developmental regulators
(Figures 5 and 6). Expression patterns were analyzed by RT-
PCR in order to give the required sensitivity. As controls,
cDNA from egg cells, two-celled proembryos, and central
cells were used.
Egg cell transcripts. Expression of TaECA1-like genes, the
largest cluster in the egg cell, was not detected in any
vegetative tissue, in anthers or 12-day-old developing cary-
opsis of wheat (Figure 5). Expression of TaECA1-like genes
was only found in the tissue containing the unfertilized egg
cell (pistil), and isolated egg cells. After fertilization, TaECA1-
like transcripts appear to be downregulated. A similar
expression pattern was observed for Cluster EC-12, repre-
senting a novel transcript (Table 1a). Two other transcripts
(Clusters EC-4, EC-70) were detected both in the egg cell and
the two-celled proembryo, but not in vegetative tissue (Fig-
ure 5). Cluster EC-70 encodes a novel zinc-finger-containing
protein, while Cluster EC-4 represents a cDNAwith similarity
to a hypothetical protein of rice (Table 1a). Unlike EC-70, EC-
4 appears to be downregulated in older embryos, as it is not
detectable in kernels 12 dap. A novel gene represented by
Cluster EC-57 encodes a small polypeptide with a conserved
Categories
(a) Functional categorization of cluster
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7 8 9 10 11
% C
lust
er
Categories
(b) Functional categorization of ESTs
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7 8 9 10 11
% E
ST
s
Figure 4. Functional classification of 360 contigs (a) containing 756 ESTs (b)
with similarity to characterized proteins. Functional categories are (1) primary
and secondary metabolism, (2) DNA and chromatin structure (including
histones), (3) transcription and RNA processing, (4) translation (including
ribosomal proteins), (5) protein-folding, -stability and -transport, (6) protein
modification and degradation, (7) cytoskeleton, (8) signal transduction, (9) cell
cycle regulation, (10) defense and stress response, (11) miscellaneous.
Percentages of egg cell contigs (193) and ESTs (454) are represented by blue
bars, red bars represent the percentages of contigs (167) and ESTs (302) from
the two-celled proembryo.
Transcriptome of wheat eggs and proembryos 665
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

domain similar to Smt3 ubiquitin-like proteins (Table 1a).
Expression of EC-57 was detected in egg cells, central cells,
proembryos and unfertilized pistils but not in vegetative
tissues, except for a very weak signal in root tissue (Fig-
ure 5). Another group of novel transcripts appeared to be
expressed in male and female reproductive tissues (Fig-
ure 5, Clusters EC-123, EC-50, EC-217, EC-52). With the
exception of EC-52, this group of transcripts is downregu-
lated later in kernel development. While mRNAs of Clusters
EC-217 and EC-52 were detected in the female gametes, the
proembryo and anthers, Cluster EC-123, encoding an
Armadillo repeat (ARM) domain-containing protein, was not
detected in central cells and proembryos. ARM-repeat pro-
teins are characterized by degenerated sequence motifs of
about 42 amino acids originally found in the Drosophila
segment polarity protein armadillo (Nuesslein-Volhard and
Wieschaus, 1980; Riggleman et al., 1989) and regulate a
variety of cellular processes including cell adhesion, intra-
cellular signaling and gene expression (Hatzfeld, 1999).
Candidate genes encoding putative developmental regu-
lators, such as proteins with similarity to an SKP1-related
Fimbriata-associated protein (FAP) from Antirrhinum
(Ingram et al., 1997), a Brassinolide enhanced gene of
unknown function from rice (Yang and Komatsu, 2004), a
putative heterogeneous RNA-binding protein from rice
(accession BAC56813), and a putative lipid transfer protein
(LTP) from rice (accessionAAN05565; Table 1a)were present
in almost all tissues and cells tested (Figure 5; EC-24; EC-75,
EC-91, EC-386). Interestingly, two egg cell ESTs (Cluster
EC-95) encode a protein containing the conserved motif of
the LOB (LATERAL ORGAN BOUNDARIES) domain gene
family. Members of this gene family are thought to play a
role in boundary establishment or communication between
the meristem and initiating lateral organs as well as adaxial–
abaxial polarity establishment (Lin et al., 2003). In addition
to a presence in egg cells, the LOB-like gene transcript was
detected in anthers, pistils, kernels 12 dap, as well as in
coleoptiles, primary leaves and stems (EC-95; Figure 5).
Transcripts of the two-celled proembryo. Several candi-
dates of the two-celled proembryo which potentially encode
proteins involved in development or which match to pub-
lished ESTs of yet unknown function are expressed in almost
all vegetative tissues tested, but their transcripts were not
detected among cDNA from the female gametes before
fertilization (Figure 6). These include Cluster 2C-1570, which
encodes a protein similar to a SET domain protein from rice
(accession BAD05333) and Cluster 2C-1645, which encodes a
protein similar to the WD-repeat protein ZmRbAp1 from
maize (accession AF250047). Corresponding genes appear
to be newly expressed after fertilization. Two other genes
which appear to be constitutively expressed after fertiliza-
tion are represented by Clusters 2C-1696 and 2C-1726.
Cluster 2C-1696 encodes a protein similar to a translationally
controlled tumor protein (TCTP) from wheat (accession
AAM34280). TCTPs are highly conserved cytoplasmic
calcium-binding proteins, present in all eukaryotics and
put. RNA-binding protein
ECA1-like
Novel F-box protein
Hypothetical protein
Novel Zn-finger protein
Novel
Novel, SMT3-like
Hypoth. ARM protein
Novel
LOB domain protein
Lipid transfer protein
BLE-like
Fimbriata assoc. protein
GAP-DH
Novel F-box protein
EC-1
EC-12
EC-4
EC-70
EC-123
EC-50
EC-217
EC-57
EC-52
EC-95
EC-24
EC-386
EC-75
EC-91
Control
Co
leo
pti
le
Pri
mar
y le
af
Ste
m
Ro
ot
w.o
. tip
Eg
g c
ell
Pis
til
An
ther
s
Co
ntr
ol
Ro
ot
tip
Mat
ure
leaf
Ker
nel
, 12d
ap
2-ce
lled
pro
emb
ryo
Cen
tral
cel
l
Figure 5. Expression profiles for transcripts derived from the egg cell cDNA
library (compare also with Table 1a). Expression was examined by RT-PCR
using DNAse-treated total RNA from different tissues of wheat. As controls,
cDNA fromegg cells, central cells and two-celled proembryos have been used.
Unknown protein
Novel
GASA-domain protein
SET-domain protein
RbAP1-like
TCTP
BI-1 like
GAP-DH
Novel
Novel
2C-1566
2C-1563
2C-1533
2C-1526
2C-1807
2C-1570
2C-1645
2C-1696
2C-1726
Control
Co
leo
pti
le
Pri
mar
y le
af
Ste
m
Ro
ot
w.o
. tip
Eg
g c
ell
Pis
til
An
ther
s
Co
ntr
ol
Ro
ot
tip
Mat
ure
leaf
Ker
nel
, 12d
ap
2-ce
lled
pro
emb
ryo
Cen
tral
cel
l
Figure 6. Expression profiles for transcripts derived from the two-celled
proembryo cDNA library (compare also with Table 1b). Expression was
examined by RT-PCR using DNAse-treated total RNA from different tissues of
wheat. As controls, cDNA from egg cells, central cells and two-celled
proembryos have been used.
666 Stefanie Sprunck et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

implicated in cellular processes such as cell cycle progres-
sion, malignant transformation and the protection against
apoptosis (Bommer and Thiele, 2004). Cluster 2C-1726 rep-
resents the transcript of a protein similar to the barley BAX
Inhibitor-1 (BI-1), an intracellular multi-membrane spanning
protein and cell death inhibitor (Huckelhoven et al., 2001).
A small protein of 105 amino acids containing a GASA
domain (pfam02704; present in a gibberellin-regulated pro-
tein family of Arabidopsis) is encoded by Cluster 2C-1807. As
with other candidate genes analyzed by RT-PCR, expression
of 2C-1807 was not detected in the gametes before fertiliza-
tion. After fertilization, 2C-1807 was expressed in all the
tissues tested, except mature leaves (Figure 6). A gene of
unknown function with similarity to barley ESTs derived
from embryo sacs, embryos as well as callus is represented
by Cluster 2C-1563. Expression of 2C-1563 was detected in
unfertilized pistils, developing kernels and very weakly in
stems and root tips, indicating a potential role in cell division
(Figure 6). Two novel clusters of the two-celled proembryo
(2C-1526 and 2C-1533) represent genes expressed weakly in
some, but not all vegetative tissues (Figure 6), while the
novel gene represented by Cluster 2C-1566 is not detectable
in vegetative tissues.
Discussion
Transcriptional activity and protein metabolism seems high
in the unfertilized egg cell
Based on our experiments we conclude that mature egg
cells of plants are not as quiescent as previously thought
(Diboll, 1968; Mogensen, 1982; Taylor and Vasil, 1995). The
high transcriptional activity of wheat egg cells4 was already
indicated by the presence of a large nucleolus (Kumlehn
et al., 1999), the nuclear sub-compartment of rRNA gene
transcription, rRNA processing and ribosome assembly.
This coincides with our identification of numerous egg cell
transcripts encoding ribosomal proteins. Strong transcrip-
tional activity is also indicated by the presence of the histone
variant H3.3 among the largest clusters of the wheat egg cell,
as H3.3 is incorporated into the chromatin of non-dividing
cells and accumulates in regions of actively transcribed
genes in Drosophila (Ahmad and Henikhoff, 2002; McKittrick
et al., 2004), probably by replacing the core histone H3. This
is further supported by the observation that H3.3 was not
present among the ESTs from the two-celled proembryo,
where 3.9% of ESTs encode H3, which is also a marker for
DNA replication (van der Knaap et al., 1997).
We also identified many egg cell transcripts that encode
proteins involved in primary and secondary metabolism,
leading us to the assumption that the unfertilized wheat egg
cell is in a state of high metabolic activity, at least shortly
before anthesis. Furthermore, the egg cell seems to be very
active in mRNA translation and protein turnover, as it
comprises not only a high number of transcripts encoding
ribosomal proteins, but also CyP1, sHSPs, and proteins of
ubiquitin-related protein-conjugation systems. CyP1 is a
peptidyl-prolyl cis-trans isomerase which catalyzes protein
folding by accelerating the slow step of cis-trans isomeriza-
tion of peptidyl-prolyl bonds (Boston et al., 1996). As a
member of the hsp90-based chaperone system, CyP1 is
involved in post-translational modification and protein
turnover. Interestingly, many signaling proteins in animals
(including steroid hormone receptors and kinases) are
regulated by this hsp90-based chaperone system (Buchner,
1999), so this may also be likely for wheat eggs.
Another remarkable finding was the discovery of several
known and novel transcripts encoding ubiquitin, ubiquitin-
related and F-box proteins, indicating that ubiquitination
may be an important post-translational regulatory pathway
for the unfertilized egg cell, both to establish and tomaintain
its functional status. Selection of ubiquitinated target pro-
teins to proteolysis by the 26S proteasome complex is
accomplished by ubiquitin-conjugating enzymes (E2s) and
ubiquitin-protein ligases (E3s). In particular, F-box proteins
linked to the E3 complex are responsible for specific
substrate recognition and binding (Kuroda et al., 2002) and
thus may target individual proteins for proteolysis. Besides
removing abnormal polypeptides, selective degradation of
short-lived regulatory proteins using the ubiquitin/26S pro-
teasome pathway is a powerful regulatory mechanism in a
wide variety of cellular processes in plants including the
regulation of cell cycle, flower development, hormonal
signal transduction, self-incompatibility and pathogen
response (Smalle and Vierstra, 2004; Sullivan et al., 2003).
Moreover, data from yeast and animals indicate other
functions of ubiquitin that do not necessarily involve the
proteasome. Several ubiquitin-like proteins are involved in
ubiquitin-like protein conjugation systems regulating his-
tone modification, protein stabilization, protein localization
and internalization of plasma membrane proteins (Hicke,
2001). It is thus tempting to speculate that specifically
expressed F-box proteins and ubiquitin-related proteins
may be used to prevent the onset of DNA replication and
other cell cycle-related processes in the unfertilized egg cell,
until the sperm fusion triggers zygotic development.
Zygotic gene activation occurs shortly after fertilization in
wheat
Zygotic gene activation in plants has been proposed based
on expression analyses of single genes (Dresselhaus et al.,
1999a; Sauter et al., 1998; Weijers et al., 2001). The appear-
ance of a second nucleolus in zygotes of maize and wheat
has been discussed as a suitable indication for successful
fertilization (Faure et al., 1994; Kumlehn et al., 1998). In
maize, the second nucleolus is derived from the sperm cell
genome (Mol et al., 1994), indicating the activation of
Transcriptome of wheat eggs and proembryos 667
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

paternal rDNA shortly after karyogamy of egg and sperm
nuclei. The present study substantiates the assumption that
ZGA in higher plants occurs earlier than in animals: the
significant changes in transcript composition observed in
proembryos just after the first cell division mirrors the
completed transition towards zygotic gene expression.
Several transcripts derived from the two-celledproembryo
were found to be fertilization-induced, ofwhichmany remain
constantly expressed throughout plant development. Such
transcripts encode, for example, a homolog of the maize
retinoblastoma-associated protein ZmRbAp1, as well as a
SET domain protein. RbAp-like proteins are components of
multiprotein complexes related to chromatin assembly dur-
ing DNA replication, chromatin remodeling and histone
acetylation activities (Ach et al., 1997). Transcripts of ZmR-
bAp1 are abundant during the initial stages of endosperm
formation, and mRNAs are localized in shoot apical meri-
stems and leaf primordia of developing embryos (Rossi
et al., 2001). Our results show that the ZmRbAp1 homolog of
wheat is not expressed in gametes, but in embryos very early
after fertilization aswell as in vegetative tissues, suggesting a
general role in chromatin assembly or modification during
sporophytic development. SET domain proteins are very
important for developmental and epigenetic regulation of
gene expression. Several plant SET domain proteins appear
to control chromatin states via histonemethylation (Springer
et al., 2003). However, transcripts of the fertilization-induced
SET domain protein of wheat proembryos are detected in all
sporophytic tissues and thus appear to have a general
function in plant growth and development.
Genes specifically expressed in egg cells and two-celled
proembryos encode not only novel proteins but also
putative secreted peptides of unknown function
Thesequencingofonly 1197ESTshas led to the identification
of many clusters representing novel transcripts, although
more than 1.5 million Poaceae ESTs are present in the public
EST database. This may reflect the under representation of
gametic transcripts in current databases. Interestingly, the
egg cell containsmorenovel clusters (71), comparedwith the
34 novel clusters of the proembryo. As 11 egg cell clusters
were not similar to any published EST but to annotated hypo-
thetical genesdetected in genomesofArabidopsis and/or rice,
weconclude that thesegenesmight beexclusively expressed
in the gametes of Arabidopsis and/or rice. It seems likely that
some of the 105 novel transcripts encode proteins required
for specific functions of the gamete and the early embryo,
respectively. Expression analyses verified that several novel
transcripts and transcripts with similarity to hypothetical
proteins are abundant in the egg cell prior to fertilization, but
either not or only barely detectable in the two-celled proem-
bryo. The majority of these genes were not expressed in
vegetative tissues, indicating that they might either specify
egg cell identity, or function in processes such as micropylar
pollen tube guidance, fertilization or the onset of embryo-
genesis. Such are transcripts for novel F-box proteins, a zinc-
finger protein, an SMT3-like and an ARM-repeat-containing
protein as well as some completely unknown proteins.
Strikingly, a number of transcripts encode small and
putative secreted proteins. These include transcripts form-
ing the largest cluster of the egg cell with similarity to ECA1
(early culture abundant1), a cDNA of unknown function
previously isolated from androgenic barley microspores
(Vrinten et al., 1999). We compared the 5¢ and 3¢ UTRs of all
wheat ECA1-like (TaECA1-like) ESTs and identified at least
five distinct transcript groups, representing members of a
gene family. All TaECA1-like transcripts encode small pro-
teins of around 151 amino acids, containing six conserved
cysteine residues and a putative signal peptide for extra-
cellular localization. Although the largest cluster in egg cells,
TaECA1-like ESTs were not identified among the two-celled
proembryo ESTs. Interestingly, Cordts et al. (2001) previ-
ously reported about genes encoding cysteine-rich secreted
peptides (ZmES1-4) that are highly and specifically ex-
pressed in cells of the maize female gametophyte and are
downregulated after fertilization. An important role for
ZmES1-4 in the fertilization process can be assumed, as
maize knockout plants are female-sterile5 (S. Amien and
T. Dresselhaus, unpublished data). In general, small extra-
cellular proteins are attractive candidates for signaling
molecules, and may serve as ligands for receptor-like
kinases (Dresselhaus and Sprunck, 2003; Ryan et al., 2002).
Such peptides might mediate interactions between male
and female gametes, communication among the cells of the
female gametophyte, or interactions with maternal tissues.
Physiological and genetic data support the assumption
that cells of the female gametophyte produce diffusible
signals involved in pollen tube attraction and reception
(Higashiyama et al., 2001; Huck et al., 2003; Rotman et al.,
2003; Shimizu and Okada, 2000). Another gene which is
expressed in both the egg cell and the proembryo encodes a
putative LTP. LTPs are secreted hydrophilic peptides which
are characterized by a conserved signature motif of eight
cysteines (Kader, 1997). They are abundant and widespread
in plants, forming a broad family of proteins whose biologic
role is still under discussion, but there is evidence that some
LTPs might act as signaling molecules (Buhot et al., 2001).
Other transcripts found in the egg cell and the proembryo
encode a peptide similar to the pathogenesis-related small
protein PR1 from maize (Casacuberta et al., 1991). This
peptide is secreted and contains six cysteines. A fertilization-
induced transcript encodes a predicted secreted small
protein of 105 amino acids containing a 25-amino acid
signal peptide and a motif of 12 cysteines. Interestingly, this
small protein contains a GASA domain (pfam 02704), found
in a gibberellin-regulated cysteine-rich protein family from
Arabidopsis. The expression of such GASA proteins is
668 Stefanie Sprunck et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

upregulated by the plant hormone gibberellin and most of
these members appear to have a role in developmental
control (Aubert et al., 1998; Herzog et al., 1995).
Conclusions
By generating and analyzing a limited number of ESTs from
cell type-specific cDNA libraries we were able to greatly in-
crease our knowledge about changes in gamete expression
profiles and thus, the transcriptional consequence of fertil-
ization. We have shown that transcriptional changes take
place considerably and soon after fertilization in wheat,
providing now a clear proof that ZGA occurs early in wheat,
and possibly in other cereals. It is also clear now that mature
egg cells of wheat are not quiescent. Especially protein
biosynthesis, protein modification and protein degradation
as well as signaling seem to be crucial biologic functions for
the egg cell before fertilization, while translation, DNA rep-
lication, chromatin remodeling, and other cell cycle-related
processes are characteristic of early zygote/embryo devel-
opment. Especially the identified novel genes which are
specifically expressed in each, the egg cell and the pro-
embryo will be interesting for functional analysis, as they
may be involved in regulating egg cell identity, fertilization
as well as the initiation of embryogenesis.
Experimental procedures
Isolation of wheat embryo sac cells before and after
fertilization
Spikes of Triticum aestivum cv. ‘Florida’ were emasculated
2–4 days before anthesis and covered with bags to prevent
fertilization. Egg cells were isolated mechanically from
microdissected ovules in 0.55 M sterile mannitol using fine-
tipped glass needles and an inverted microscope, as des-
cribed by Kumlehn et al. (1999). Single cells were transferred
into 0.5 ml reaction tubes by using a glass capillary inter-
faced with a hydraulic system to a micropump. Collected
cells were immediately frozen in liquid nitrogen. Wheat
zygotes were isolated 4–6 h after hand-pollination of recep-
tive stigmata by using the same procedure, but transferring
the zygotes into fresh 0.55 M mannitol solution. Defined
stages of zygotes and proembryos were selected using an
inverted microscope. In vivo embryos were isolated using
the similar procedure to that described above.
mRNA isolation and cDNA synthesis
mRNA was isolated from 12 egg cells and 20 proembryos
at the two-celled stage, using the Dynabeads� mRNA
DIRECTTM Micro kit (Dynal, Hamburg, Germany)6 following
themanufacturer’s guidelines with but scaled down to 50 ll.Annealed mRNA was isolated using a magnetic particle
transfer device (PickPenTM; Bio-Nobile, Turku, Finland).7
Subsequently, the SMARTTM PCR cDNA synthesis kit (BD
Biosciences, Heidelberg, Germany)8 was used for cDNA
synthesis. First-strand cDNA, LD-PCR, and determination
of optimal cycle numbers for generating a population
of representative cDNAs was performed according the
manufacturer’s guidelines but using a digoxigenin-11-dUTP
(Roche Applied Science, Mannheim, Germany)9 labeled
fragment of wheat GAPDH as a probe.
Library construction and sequencing
cDNA (150 ll) was used for polishing, according to the
SMARTTM PCR cDNA synthesis kit (BD Biosciences).
Subsequently, 3 lg of EcoRI (NotI) adapters (Invitrogen,
Karlstuhe, Germany)10 were ligated to blunt-end cDNA, using
T4 ligase (New England Biolabs, Frankfurt, Germany).11
Remaining adapters and fragments below 0.3 kb were
removed by electrophoresis in 0.8% low-melting point
agarose (Seaplaque GTG). Afterwards, cDNA was extracted
using b-agarase I (New England Biolabs). After phosphory-
lation of EcoRI cohesive ends (10 U/ll T4 polynucleotide
kinase; New England Biolabs), a second purification step
using ChromaspinTM columns (BD Biosciences) was per-
formed. The cDNAwas then ligated into predigested lambda
ZAP� II/EcoRI/CIAP vector (Stratagene, Amsterdam, the
Netherlands).12 The titer of the unamplified libraries was
1.43 · 106 pfu ml)1 for egg cells and 3.2 · 105 pfu ml)1 for
the two-celled proembryo, respectively. After amplification
and in vivo excision, cloneswere randomly picked from each
library and used to generate ESTs. Insert sizes ranged from
300 to 3000 bp, with an average of 900 bp. The average
readable sequence length of ESTs was about 500 bp. DNA
sequencer trace data subsequently passed an automated
cleanup pipeline including PHRED to call bases and assign
quality values, followed by CROSS_MATCH to align
sequences and to eliminate vector sequences.
Bioinformatics
The sequences of each library were clustered using blast-
clust (NCBI) and assembled into contigs using Vector NTI 8
(Invitrogen). The contig’s consensus sequence or the lon-
gest representative was used for BLASTN searches against
NCBI’s non-redundant (nr) database and the EST database,
and for BLASTX searches against NCBI’s nr database and
SWISSPROT (March 2004). A number of cDNAs resulted in
limited sequence information (100–250 bp) from non-coding
regions. Therefore, BLASTN searches against the TIGRWheat
Gene Index Release 8.0 (Quackenbush et al., 2001) were
performed, using the BLASTN algorithm. If a match with
>95% sequence identity over the total length of the query
sequence was found, the matching sequence was retrieved
and used in subsequent BLASTX searches in place of the
Transcriptome of wheat eggs and proembryos 669
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

original EST. A sequence was considered novel if it did not
show a significant match with a sequence of the NCBI
databases (nr, EST) or to the TIGR-assembled wheat
consensus sequences using the BLASTN algorithm (Altschul
et al., 1997). The significance threshold used for BLASTN
searches were: score >115, expected value <e)25.
For BLASTX searches, the cutoff for a significant match for
all but the short sequences was an e-value of <e)15, score
‡80. Matches to short query sequences (below 260 bp) were
inspected and categorized manually. Clusters encoding
proteins of known function were manually categorized into
broad functional groups using the Munich Information
Centre for Protein Sequences classification as guidance.
Expression analysis by RT-PCR
RNA was isolated from vegetative and generative tissues
using TRIzol� reagent (Invitrogen), essentially following the
manufacturer’s protocol. Starch containing tissues such as
caryopsis were extracted twice, using 3 ml of TRIzol�
reagent per 100 mg of tissue. The quality of the total RNA
preparation was analyzed by denaturating agarose gel elec-
trophoresis. Before RT-PCR, 1 lg of total RNA was digested
with DNAse (RNAse free; Invitrogen) and subsequently used
for first-strand cDNA synthesis using Oligo(dT)23 (Sigma,
Taufkirchen, Germany)13 and Superscript II reverse transcrip-
tase (Invitrogen), following the manufacturer’s protocol but
adding RNAseOUTTM (Invitrogen). Quality and amount of
generated cDNAswas analyzed by PCRwith intron-spanning
primers directed against wheat GAPDH (TaGAP1, 5¢-AGG-
GTGGTGCCAAGAAGGTCA-3¢; TaGAP2, 5¢-TATCCCCACTC-GTTGTCGTA-3¢). Primer pairs directed against selected
sequence clusters were designed according to PrimerSelect
(Lasergene, GATC Biotech AG, Konstanz, Germany),14 result-
ing in amplified products of 120–600 bp length. Sequences
of gene-specific primers are available on request. GAPDH
PCR reactions were carried out for 30 cycles, using 2 ll ofcDNA. PCR reactions for transcript-specific primers of
selected candidates were carried out for 38 cycles, using 2.5
of cDNA as template.
Acknowledgements
This work was funded by the Grains Research DevelopmentCorporation, Australia. We thank Gary Barker and Ian Wilson(University of Bristol, UK) for sequencing and editing the ESTs aspart of the BBSRC Investigating Gene Function program. We aregrateful to Jochen Kumlehn for his excellent introduction to thetechnique of wheat microdissection.
References
Ach, R.A., Taranto, P. and Gruissem, W. (1997) A conserved familyof WD-40 proteins binds to the retinoblastoma protein in bothplants and animals. Plant Cell, 9, 1595–1606.
Adenot, P.G., Mercier, Y., Renard, J.P. and Thompson, E.M. (1997)Differential H4 acetylation of paternal and maternal chromatinprecedes DNA replication and differential transcriptional activityin pronuclei of 1-cell mouse embryos. Development, 124, 4615–4625.
Ahmad, K. and Henikhoff, S. (2002) Histone H3 variants specifymodes of chromatin assembly. Proc. Natl Acad. Sci. USA, 10
(Suppl. 4), 16477–16484.Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z.,
Miller, W. and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs.Nucleic Acids Res. 25, 3389–3402.
Angerer, L.M. and Angerer, R.C. (2000) Animal-vegetal axis pat-terning mechanisms in the early sea urchin embryo. Dev. Biol. 1,1–12.
Aubert, D., Chevillard, M., Dorne, A.M., Arlaud, G. and Herzog, M.
(1998) Expression patterns of GASA genes in Arabidopsis thali-ana: the GASA4 gene is up-regulated by gibberellins in meris-tematic regions. Plant Mol. Biol. 36, 871–883.
Azevedo, C., Sadanandom, A., Kitagawa, K., Freialdenhoven, A.,
Shirasu, K. and Schulze-Lefert, P. (2002) The RAR1 interactorSGT1, an essential component of the R gene triggered diseaseresistance. Science, 295, 2073–2076.
Baugh, L.R., Hill, A.A., Slonim, D.K., Brown, E.L. and Hunter, C.P.
(2003) Composition and dynamics of the Caenorhabditis elegansearly embryonic transcriptome. Development, 130, 889–900.
Bommer, U.A. and Thiele, B.J. (2004) The translationally controlledtumour protein (TCTP). Int. J. Biochem. Cell Biol. 36, 379–385.
Boston, R.S., Viitanen, P.V. and Vierling, E. (1996) Molecular chap-erones and protein folding in plants. Plant Mol. Biol. 32, 191–222.
Buchner, J. (1999) Hsp90 & Co. – a holding for folding. TrendsBiochem. Sci. 24, 136–141.
Buhot, N., Douliez, J.P., Jacquemard, A. et al. (2001) A lipid transferprotein binds to a receptor involved in the control of plant defenceresponses. FEBS Lett. 509, 27–30.
Casacuberta, J.M., Puigdomenech, P. and San Segundo, B. (1991) Agene coding for a basic pathogenesis-related (PR-like) proteinfrom Zea mays. Molecular cloning and induction by a fungus(Fusarium moniliforme) in germinating maize seeds. Plant Mol.Biol. 16, 527–536.
Cordts, S., Bantin, J., Wittich, P.E., Kranz, E., Lorz, H. and Dressel-
haus, T. (2001) ZmES genes encode peptides with structuralhomology to defensins and are specifically expressed in thefemale gametophyte of maize. Plant J. 25, 103–114.
Diboll, A.G. (1968) Fine structural development of the megagamet-ophyte of Zea mays following fertilization. Am. J. Bot. 53, 787–806.
Dresselhaus, T. and Sprunck, S. (2003) Peptide hormone mediatedsignaling in plants and animals reveals mechanistic similarities.In Advances in Plant Physiology, Vol. 6 (Hemantaranjan, A., ed.).Jodhpur: Scientific Publishers, pp. 131–177.15
Dresselhaus, T., Lorz, H. and Kranz, E. (1994) Representative cDNAlibraries from few plant cells. Plant J. 5, 605–610.
Dresselhaus, T., Cordts, S. and Lorz, H. (1999a) A transcript enco-ding translation initiation factor eIF-5A is stored in unfertilizedegg cells of maize. Plant Mol. Biol. 39, 1063–1071.
Dresselhaus, T., Cordts, S., Heuer, S., Sauter, M., Lorz, H. and Kranz,
E. (1999b) Novel ribosomal genes from maize are differentiallyexpressed in the zygotic and somatic cell cycles.Mol. Gen. Genet.261, 416–427.
Edgar, L.G., Wolf, N. and Wood, W.B. (1994) Early transcription inCaenorhabditis elegans embryos. Development, 120, 443–451.
Faure, J.E., Digonnet, C. and Dumas, C. (1994) An in vitro system foradhesion and fusion of maize gametes. Science, 263, 1598–1600.
670 Stefanie Sprunck et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

Furukawa, T., Ishibashi, T., Kimura, S., Tanaka, H., Hashimoto, J.
and Sakaguchi, K. (2003) Characterization of all the subunits ofreplication factor C from a higher plant, rice (Oryza sativa L.).Plant Mol. Biol. 53, 15–25.
Haecker, A., Gross-Hardt, R., Geiges, B., Sarkar, A., Breuninger, H.,
Herrmann, M. and Laux, T. (2004) Expression dynamics of WOXgenes mark cell fate decisions during early embryonic patterningin Arabidopsis thaliana. Development, 131, 657–668.
Hatzfeld, M. (1999) The armadillo family of structural proteins. Int.Rev. Cytol. 186, 179–224.
Herzog, M., Dorne, A.M. and Grellet, F. (1995) GASA, a gibberellin-regulated gene family from Arabidopsis thaliana related to thetomato GAST1 gene. Plant Mol. Biol. 27, 743–752.
Heuer, S., Hansen, S., Bantin, J., Brettschneider, R., Kranz, E., Lorz,
H. and Dresselhaus, T. (2001) The maize MADS box geneZmMADS3 affects node number and spikelet development and isco-expressed with ZmMADS1 during flower development, in eggcells, and early embryogenesis. Plant Physiol. 127, 33–45.
Hicke, L. (2001) Protein regulation by monoubiquitin. Nat. Rev. Mol.Cell Biol. 2, 195–201.
Higashiyama, T., Yabe, S., Sasaki, N., Nishimura, Y., Miyagishima,
S., Kuroiwa, H. and Kuroiwa, T. (2001) Pollen tube attraction bythe synergid cell. Science, 293, 1480–1483.
Holm, P.B., Knudsen, S., Mouritzen, P., Negri, D., Olsen, F.L. and
Roue, C. (1994) Regeneration of fertile barley plants frommechanically isolated protoplasts of the fertilized egg cell. PlantCell, 6, 531–543.
Huck, N., Moore, J.M., Federer, M. and Grossniklaus, U. (2003) TheArabidopsis mutant feronia disrupts the female gametophyticcontrol of pollen tube reception. Development, 130, 2149–2159.
Huckelhoven, R., Dechert, C., Trujillo, M. and Kogel, K.H. (2001)Differential expression of putative cell death regulator genes innear-isogenic, resistant and susceptible barley lines duringinteraction with the powdery mildew fungus. Plant Mol. Biol. 47,739–748.
Ingram, G.C., Doyle, S., Carpenter, R., Schultz, E.A., Simon, R. and
Coen, E.S. (1997) Dual role for fimbriata in regulating floralhomeotic genes and cell division in Antirrhinum. EMBO J. 16,6521–6534.
Kader, J.C. (1997) Lipid-transfer proteins: a puzzling family of plantproteins. Trends Plant Sci. 2, 66–70.
Kane, D.A. and Kimmel, C.B. (1993) The zebrafish midblastulatransition. Development, 119, 447–456.
Kanka, J., Bryova, A., Duranthon, V., Oudin, J.F., Peynot, N. and
Renard, J.P. (2003) Identification of differentially expressedmRNAs in bovine preimplantation embryos. Zygote, 11, 43–52.
van der Knaap, E., Jagoueix, S. and Kende, H. (1997) Expression ofan ortholog of replication protein A1 (RPA1) is induced by gib-berellin in deepwater rice. Proc. Natl Acad. Sci. USA, 94, 9979–9983.
Kranz, E., Bautor, J. and Lorz, H. (1991) In vitro fertilization of single,isolated gametes of maize mediated by electrofusion. Sex. PlantReprod. 4, 12–16.
Kumlehn, J., Lorz, H. and Kranz, E. (1998) Differentiation of isolatedwheat zygotes into embryos and normal plants. Planta, 205, 327–333.
Kumlehn, J., Lorz, H. and Kranz, E. (1999) Monitoring individualdevelopment of isolated wheat zygotes: a novel approach tostudy embryogenesis. Protoplasma, 208, 156–162.
Kumlehn, J., Kirik, V., Czihal, A., Altschmied, L., Matzk, F., Lorz, H.
and Baumlein, H. (2001) Parthenogenetic egg cells of wheat:cellular and molecular studies. Sex. Plant Reprod. 14, 239–243.
Kuroda, H., Takahashi, N., Shimada, H., Seki, M., Shinozaki, K. and
Matsui, M. (2002) Classification and expression analysis of
Arabidopsis F-box-containing protein genes. Plant Cell Physiol.43, 1073–1085.
Lin, W.C., Shuai, B. and Springer, P.S. (2003) The ArabidopsisLATERAL ORGAN BOUNDARIES-domain gene ASYMMETRICLEAVES2 functions in the repression of KNOX gene expressionand in adaxial-abaxial patterning. Plant Cell, 15, 2241–2252.
Makabe, K.W., Kawashima, T., Kawashima, S. et al. (2001) Large-scale cDNA analysis of the maternal genetic information in theegg of Halocynthia roretzi for a gene expression catalog of asci-dian development. Development, 128, 2555–2567.
McKittrick, E., Gafken, P.R., Ahmad, K. and Henikoff, S. (2004) His-tone H3.3 is enriched in covalent modifications associated withactive chromatin. Proc. Natl Acad. Sci. USA, 101, 1525–1530.
Mogensen, H.L. (1982) Double fertilization in barley and the cyto-logical explanation for haploid embryo formation, embryolesscaryopses, and ovule abortion. Carlsberg Res. Com. 47, 313–354.
Mohr, S.E., Dillon, S.T. and Boswell, R.E. (2001) The RNA-bindingprotein Tsunagi interacts with Mago Nashi to establish polarityand localize oskar mRNA during Drosophila oogenesis. GenesDev. 15, 2886–2899.
Mol, R., Matthys-Rochon, E. and Dumas, C. (1994) The kinetics ofcytological events during double fertilization in Zea mays L. PlantJ. 5, 197–206.
Nakakura, N., Miura, T., Yamana, K., Ito, A. and Shiokawa, K. (1987)Synthesis of heterogeneous mRNA-like RNA and low-molecular-weight RNA before the midblastula transition in embryos ofXenopus laevis. Dev. Biol. 123, 421–429.
Newport, J. and Kirschner, M. (1982) A major developmentaltransition in early Xenopus embryos: II. Control of the onset oftranscription. Cell, 30, 687–696.
Nishida, H. (1997) Cell lineage and timing of fate restriction, deter-mination and gene expression in ascidian embryos. Semin. Cell.Dev. Biol. 4, 359–365.
Nothias, J.Y., Majumder, S., Kaneko, K.J. and DePamphilis, M.L.
(1995) Regulation of gene expression at the beginning of mam-malian development. J. Biol. Chem. 270, 22077–22080.
Nuesslein-Volhard, C. and Wieschaus, E. (1980) Mutations affectingsegment number and polarity in Drosophila. Nature, 287, 795–801.
Pellettieri, J. and Seydoux, G. (2002) Anterior-posterior polarity in C.elegans and Drosophila – PARallels and differences. Science, 298,1946–1950.
Perry, S.E., Nichols, K.W. and Fernandez, D.E. (1996) The MADSdomain protein AGL15 localizes to the nucleus during early sta-ges of seed development. Plant Cell, 8, 1977–1989.
Quackenbush, J., Cho, J., Lee, D., Liang, F., Holt, I., Karamycheva,
S., Parvizi, B., Pertea, G., Sultana, R. and White, J. (2001) TheTIGR gene indices: analysis of gene transcript sequences inhighly sampled eukaryotic species. Nucleic Acids Res. 29, 159–164.
Riggleman, B., Wieschaus, E. and Schedl, P. (1989) Molecular ana-lysis of the armadillo locus: uniformly distributed transcripts anda protein with novel internal repeats are associated with a Dro-sophila segment polarity gene. Genes Dev. 3, 96–113.
Rossi, V., Varotto, S., Locatelli, S., Lanzanova, C., Lauria, M., Zan-
otti, E., Hartings, H. and Motto, M. (2001) The maize WD-repeatgene ZmRbAp1 encodes a member of the MSI/RbAp sub-familyand is differentially expressed during endosperm development.Mol. Genet. Genomics, 265, 576–584.
Rotman, N., Rozier, F., Boavida, L., Dumas, C., Berger, F. and Faure,
J.E. (2003) Female control of male gamete delivery during fertil-ization in Arabidopsis thaliana. Curr. Biol. 13, 432–436.
Ryan, C.A., Pearce, G., Scheer, J. and Moura, D.S. (2002) Polypep-tide hormones. Plant Cell, 14 (Suppl.), S251–S264.
Transcriptome of wheat eggs and proembryos 671
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672

Sauter, M., von Wiegen, P., Lorz, H. and Kranz, E. (1998) Cell cycleregulatory genes from maize are differentially controlled duringfertilization and first embryonic cell division. Sex. Plant Reprod.11, 41–48.
Shimizu, K.K. and Okada, K. (2000) Attractive and repulsive inter-actions between female and male gametophytes in Arabidopsispollen tube guidance. Development, 127, 4511–4518.
Shirasu, K., Lahaye, T., Tan, M.W., Zhou, F., Azevedo, C. and
Schulze-Lefert, P. (1999) A novel class of eukaryotic zinc-bindingproteins is required for disease resistance signaling in barley anddevelopment in C. elegans. Cell, 99, 355–366.
Smalle, J. and Vierstra, R.D. (2004) The ubiquitin 26S proteasomeproteolytic pathway. Annu. Rev. Plant Physiol. Plant Mol. Biol. 55,555–590.
Springer, N.M., Napoli, C.A., Selinger, D.A., Pandey, R., Cone, K.C.,
Chandler, V.L., Kaeppler, H.F. and Kaeppler, S.M. (2003) Com-parative analysis of SET domain proteins in maize and Arabid-opsis reveals multiple duplications preceding the divergence ofmonocots and dicots. Plant Physiol. 132, 907–925.
St Johnston, D. (1995) The intracellular localization of messengerRNAs. Cell, 2, 161–170.
Sullivan, J.A., Shirasu, K. and Deng, X.W. (2003) The diverse roles ofubiquitin and the 26S proteasome in the life of plants. Nat. Rev.Genet. 4, 948–958.
Taylor, M.G. and Vasil, I.K. (1995) The ultrastructure of zygotic em-bryo development in pearl millet (Pennisetum glaucum; Poa-ceae). Am. J. Bot. 82, 205–219.
Vrinten, P.L., Nakamura, T., Kasha, K.J. (1999) Characterization ofcDNAs expressed in the early stages of microspore embryo-genesis in barley (Hordeum vulgare L.). Plant Mol. Biol. 41, 455–463.
Weijers, D., Geldner, N., Offringa, R. and Jurgens, G. (2001) Seeddevelopment: early paternal gene activity in Arabidopsis. Nature,414, 709–710.
Wylie, C., Kofron, M., Payne, C., Anderson, R., Hosobuchi, M.,
Joseph, E. and Heasman, J. (1996) Maternal beta-catenin estab-lishes a ‘dorsal signal’ in early Xenopus embryos. Development,122, 2987–2996.
Yadegari, R. and Drews, G.N. (2004) Female gametophyte devel-opment. Plant Cell, 16 (Suppl.), S133–S141.
Yang, G. and Komatsu, S. (2004) Molecular cloning and character-ization of a novel brassinolide enhanced gene OsBLE1 in Oryzasativa seedlings. Plant Physiol. Biochem. 42, 1–6.
Zhao, J., Zhou, C. and Yang, H.Y. (2000) Isolation and in vitro cultureof zygotes and central cells of Oryza sativa L. Plant Cell Rep. 19,321–326.
Accession numbers: The GenBank numbers for the sequences mentioned in this article are AL830671–AL831324, CV 973579–CV 973658 (eggcell) and CV 973119–CV 973578 (proembryo).
672 Stefanie Sprunck et al.
ª Blackwell Publishing Ltd, The Plant Journal, (2005), 41, 660–672
Copyright © 2022 FDOKUMEN




















