
Salttolerant rhizobacteria-mediated induced tolerance in wheat ( Triticum aestivum ) and chemical...
10
ORIGINAL PAPER Salt-tolerant rhizobacteria-mediated induced tolerance in wheat (Triticum aestivum) and chemical diversity in rhizosphere enhance plant growth Shweta Tiwari & Pratibha Singh & Rameshwar Tiwari & Kamlesh K. Meena & Mahesh Yandigeri & Dhananjaya P. Singh & Dilip K. Arora Received: 1 January 2011 /Revised: 21 June 2011 /Accepted: 23 June 2011 /Published online: 9 July 2011 # Springer-Verlag 2011 Abstract Salt-tolerant isolates Bacillus pumilus, Pseudo- monas mendocina, Arthrobacter sp., Halomonas sp., and Nitrinicola lacisaponensis isolated from high saline habitats exhibited plant growth-promoting traits like P solubilization and indole acetic acid (IAA), siderophore, and ammonia production. These isolates were inoculated in wheat to assess microbe-mediated responses and plant growth promotion in salt affected soil. Maximum shoot and root length (33.8 and 13.6 cm) and shoot and root biomass (2.73 and 4.48 g dry weight) was recorded in plants inoculated with B. pumilus after 30 days. Total chlorophyll content was maximum in the leaves of the plants treated with Halomonas sp. (24.22 mg g -1 dry weight) followed by B. pumilus (23.41 mg g -1 dry weight) as compared to control (18.21 mg g -1 dry weight) after 30 days. Total protein content was maximum in Arthrobacter sp. inoculated plant leaves (3.19 mg g -1 dry weight) followed by B. pumilus (2.47 mg g -1 dry weight) as compared to control (2.15 mg g -1 dry weight) after 30 days. Total carotenoid content was maximum in plants inoculated with Halomonas sp. (1,075.45 and 1,113.29 μg g -1 dry weight) in comparison to control (837.32 and 885.85 μgg -1 dry weight) after 15 and 30 days. Inoculation of bacterial isolates increased presence of individual phenolics (gallic, caffeic, syringic, vanillic, ferulic, and cinnamic acids) and flavonoid quercetin in the rhizosphere soil. The concentra- tion of IAA in rhizosphere soil and root exudates was also higher in all treatments than in control. Accumulation of phenolics and quercetin in the plants played a cumulative synergistic role that supported enhanced plant growth promotion of wheat in the stressed soil. Keywords Plant growth promotion . Stress tolerance . Bacillus pumilus . Phenolics . Flavonoids Introduction Plants usually face several environmental stresses that can affect crop quality and productivity (Jones 2009). Salinity can decrease plant growth (Amirjani 2010; Bano and Fatima 2009) and in order to improve crop produc- tivity in salt affected areas, suitable biotechnological approaches should be carried out (Zahir et al. 2004). The use of stress-tolerant microbial strains associated with roots of agronomic crops may improve soil fertility and plant resistance towards adverse environmental conditions (Wu et al. 2009). For example, root-colonizing non- pathogenic rhizobacteria can enhance plant resistance towards biotic and abiotic stresses (Yang et al. 2010), and improve plant growth and soil conditions (Scurtz et al. 2000; Sturz and Nowak 2000; Rabie and Almandini 2005). Microorganisms surviving at extreme environmen- tal conditions are suitable for use in different agricultural practices (Egamberdieva and Kucharova 2009). The most intense interactions between microbes and plants that can influence plant growth and plant environ- mental adaptation take place in the rhizosphere soil (Siddikee et al. 2010). Understanding the complexity of microbial adaptations into stressed rhizosphere environment and the effect of these microorganisms on biological, chemical, and physical properties of rhizosphere soil and the plants remains a significant challenge (Yang et al. S. Tiwari : P. Singh : R. Tiwari : K. K. Meena : M. Yandigeri : D. P. Singh (*) : D. K. Arora National Bureau of Agriculturally Important Microorganisms, Kushmaur, Maunath Bhanjan 275101, India e-mail: [email protected] Biol Fertil Soils (2011) 47:907–916 DOI 10.1007/s00374-011-0598-5
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Salttolerant rhizobacteria-mediated induced tolerance in wheat ( Triticum aestivum ) and chemical...

ORIGINAL PAPER
Salt-tolerant rhizobacteria-mediated induced tolerancein wheat (Triticum aestivum) and chemical diversityin rhizosphere enhance plant growth
Shweta Tiwari & Pratibha Singh & Rameshwar Tiwari &Kamlesh K. Meena & Mahesh Yandigeri &Dhananjaya P. Singh & Dilip K. Arora
Received: 1 January 2011 /Revised: 21 June 2011 /Accepted: 23 June 2011 /Published online: 9 July 2011# Springer-Verlag 2011
Abstract Salt-tolerant isolates Bacillus pumilus, Pseudo-monas mendocina, Arthrobacter sp., Halomonas sp., andNitrinicola lacisaponensis isolated from high saline habitatsexhibited plant growth-promoting traits like P solubilizationand indole acetic acid (IAA), siderophore, and ammoniaproduction. These isolates were inoculated in wheat toassess microbe-mediated responses and plant growthpromotion in salt affected soil. Maximum shoot and rootlength (33.8 and 13.6 cm) and shoot and root biomass (2.73and 4.48 g dry weight) was recorded in plants inoculatedwith B. pumilus after 30 days. Total chlorophyll content wasmaximum in the leaves of the plants treated withHalomonas sp. (24.22 mg g−1 dry weight) followed by B.pumilus (23.41 mg g−1 dry weight) as compared to control(18.21 mg g−1 dry weight) after 30 days. Total proteincontent was maximum in Arthrobacter sp. inoculated plantleaves (3.19 mg g−1 dry weight) followed by B. pumilus(2.47 mg g−1 dry weight) as compared to control(2.15 mg g−1 dry weight) after 30 days. Total carotenoidcontent was maximum in plants inoculated with Halomonassp. (1,075.45 and 1,113.29 μg g−1 dry weight) incomparison to control (837.32 and 885.85 μg g−1 dryweight) after 15 and 30 days. Inoculation of bacterialisolates increased presence of individual phenolics (gallic,caffeic, syringic, vanillic, ferulic, and cinnamic acids) andflavonoid quercetin in the rhizosphere soil. The concentra-tion of IAA in rhizosphere soil and root exudates was alsohigher in all treatments than in control. Accumulation ofphenolics and quercetin in the plants played a cumulative
synergistic role that supported enhanced plant growthpromotion of wheat in the stressed soil.
Keywords Plant growth promotion . Stress tolerance .
Bacillus pumilus . Phenolics . Flavonoids
Introduction
Plants usually face several environmental stresses thatcan affect crop quality and productivity (Jones 2009).Salinity can decrease plant growth (Amirjani 2010; Banoand Fatima 2009) and in order to improve crop produc-tivity in salt affected areas, suitable biotechnologicalapproaches should be carried out (Zahir et al. 2004). Theuse of stress-tolerant microbial strains associated withroots of agronomic crops may improve soil fertility andplant resistance towards adverse environmental conditions(Wu et al. 2009). For example, root-colonizing non-pathogenic rhizobacteria can enhance plant resistancetowards biotic and abiotic stresses (Yang et al. 2010),and improve plant growth and soil conditions (Scurtz et al.2000; Sturz and Nowak 2000; Rabie and Almandini2005). Microorganisms surviving at extreme environmen-tal conditions are suitable for use in different agriculturalpractices (Egamberdieva and Kucharova 2009).
The most intense interactions between microbes andplants that can influence plant growth and plant environ-mental adaptation take place in the rhizosphere soil(Siddikee et al. 2010). Understanding the complexity ofmicrobial adaptations into stressed rhizosphere environmentand the effect of these microorganisms on biological,chemical, and physical properties of rhizosphere soil andthe plants remains a significant challenge (Yang et al.
S. Tiwari : P. Singh :R. Tiwari :K. K. Meena :M. Yandigeri :D. P. Singh (*) :D. K. AroraNational Bureau of Agriculturally Important Microorganisms,Kushmaur, Maunath Bhanjan 275101, Indiae-mail: [email protected]
Biol Fertil Soils (2011) 47:907–916DOI 10.1007/s00374-011-0598-5

2008). At present, the greatest interest resides in thedevelopment and application of trait-specific microbialinoculants that can be used for disease management (Whips2001) and stress tolerance in plants (Nadeem et al. 2007;Yang et al. 2008; Neelam and Meenu 2010). Looking intothe perspectives of crop production losses due to theseverity of abiotic stresses, especially salinity, tolerance tostress provided by microbial inoculants becomes moreimportant. In this regard, the microbes inhabiting highsalinity areas can act as potential inoculants for the cropproductivity management. The aim of this work was tostudy the effect of salt-tolerant rhizosphere bacteria ininducing responses to stress tolerance and correspondinglyhigh level of below-ground chemical diversity in the rootrhizosphere associated with plant growth promotion.
The study provides novel concepts for better understandingof mechanism of stress tolerance in plants following microbialinoculation and suggests new avenues for studying practicalmetabolomic parameters of microbe-mediated salinity stresstolerance for enhanced crop productivity.
Material and methods
Isolation and screening for traits
Bacterial strains (AS 121, AS 40, and AS 18) were isolatedfrom the rhizosphere soils collected from Mau (26°N 83°E),
Ghazipur (25.58°N 83.57°E), and Ballia (25.85°N 84.10°E)districts of Uttar Pradesh (India) (Table 1) as described bySmalla et al. (2001). The soil particles attached to roots werecarefully collected after uprooting plants. Other two bacterialstrains (SL 9 and SL 11) were isolated from Sambhar saltlake, Rajasthan (India). Isolation of these strains was doneby serial dilution method using nutrient agar (NA) andKing’s B (KB) as selective media with incubation for 48–72 h. Identification of these bacterial strains was performedby the isolation of genomic DNA as described by Pospiechand Neumann (1995) followed by PCR amplification of 16SrRNA as described by Edwards et al. (1989). The 16S rRNAsequences were identified by comparing the partial 16SrRNA sequences of the isolated strains with those availablein the GenBank database.
Plant growth promotion (PGP) traits such as P solubili-zation and indole acetic acid (IAA), siderophore, andammonia production of the isolates were determinedaccording to Brick et al. (1991), Gaur (1990), Schwynand Neilands (1987), and Dey et al. (2004), respectively.Salt tolerance was screened by growing the isolates for 48 hat 28±2°C in NA medium containing different NaClconcentrations (2–25% w/v) with the estimation of theproline content (Bates et al. 1973), direct reducing sugars(DRS) (Umbrient et al. 1959), polysaccharides as solublesugar (Maness 2010), choline oxidase (Ikuta et al. 1977),and ACC deaminase on DF salt minimal media (Penroseand Glick 2003).
Table 1 Habitats, accession numbers, and properties of the used microbial species
Microbial species Bacillus pumilusAS 121
Pseudomonas mendocinaAS 40
Arthrobactersp. AS 18
Halomonassp. SL 9
Nitrinicola lacisaponensisSL 11
Habitats Wheat rhizosphere,Mau, UP, India
Wheat rhizosphere,Ghazipur, UP, India
Wheat rhizosphere,Ballia, UP, India
Sambhar salt lake,Rajasthan, India
Sambhar salt lake,Rajasthan, India
GenBank accessionnumber
EU 927414 EU 927412 EU 927410 FJ 984532 FJ 973520
PGP traits
IAA (μg mg−1
protein)13.6 30.9 68.8 32.3 43.2
P solubilization + ND ND ND +
Siderophoreproduction
+ ND + + +
Ammonia production + + ND + +
ACC deaminase ND ND + ND ND
Salt tolerance and accumulation of proline and choline oxidase activity
Salt tolerance limit(% NaCl)a
0–12% 0–10% 0–10% 0–22% 0–12%
Proline accumulation(μg mg−1)
6.10 3.68 3.43 ND 5.64
Choline oxidaseactivity (U ml−1)
0.0032 0.0025 0.0014 0.0038 0.0021
ND not detected, + positivea Sodium chloride concentration range on which the microorganisms were grown successfully
908 Biol Fertil Soils (2011) 47:907–916

Plant growth under salt stress
Wheat seeds (cultivar PBW-343) were grown in pre-sterilized plastic pots (diameter 20 cm, 2.5 kg soil)containing heat-sterilized silt loam soil with the followingproperties: pH 8.8, EC 4.6 dS/m, organic C 0.41%, total N0.14%, Olsen P 101 mg per 100 g of soil, sand 28.53%, silt49.6%, and clay 21.87%. Seeds were surface sterilized with0.1% HgCl2 followed by 70% ethanol and were inoculatedwith bacteria suspended in 1% carboxymethyl cellulose(CMC) at a cell concentration of 108 cells/ml. Seeds soakedwith bacterial suspension for 3–4 h were air dried andfinally sown in the plastic pots (10 seeds/pot with fourreplicates). Surface-sterilized seeds soaked with 1% CMCwithout bacteria served as control. The pots were incubatedin a controlled environmental chamber illuminated with350 μmol m−2 s−1 light intensity at 28/18°C day/nighttemperatures and 16/8 h light/dark cycle. After germination,plants were thinned to six plants per pot and were manuallyirrigated with distilled water as required throughout theexperiment. Ten plants were harvested after 15 and 30 daysafter inoculation for analysis.
Analysis
After harvesting, shoot and root length of the plantsalong with their dry weight were recorded. The chloro-phyll content of wheat leaves was quantified by extract-ing the pigment from 0.5 g leaf tissues using methanol/water (9:1, v/v). After centrifugation at 15,000×g for5 min, chlorophyll in the supernatant was quantified interms of A665nm (Ferjani et al. 2003). Total carotenoidcontent was determined as reported by Sadasivam andManikam (1992). Total protein content (TPC) wasextracted as described by Ferjani et al. (2003). One gramof wheat leaves was macerated with 0.5% trichloroaceticacid (5 ml) and the precipitate was separated by centrifu-gation at 15,000×g for 10 min at 4°C. The pellet wasresuspended in 1 N NaOH (5 ml), boiled for 30 min,cooled, and centrifuged at 15,000×g for 5 min. Totalcontent of protein was determined as reported by Lowry etal. (1951) with bovine serum albumin (BSA) as standard.Total soluble phenol (TSP) was estimated spectrophoto-metrically using the Prussian Blue method as modified byGraham (1992) and expressed in terms of gallic acidequivalents by using gallic acid (HiMedia, India) asstandard. Absorbance was read at 700 nm using UV–Visspectrophotometer (Shimadzu Corp., Japan). Analyticalgrade reagents were used throughout the experiments.Total flavonoid content was determined by the aluminumchloride colorimetric method (Kim et al. 2003). Leaftissues (0.5 g) were extracted with methanol (2 ml) and
mixed with 0.1 ml of 10% aluminum chloride hexahy-drate, 0.1 ml of 1 M potassium acetate, and 2.8 ml ofdeionized water. After incubation for 40 min at roomtemperature, the absorbance of the reaction mixture wasdetermined at 415 nm. Total flavonoid content wasexpressed in terms of rutin equivalents.
Extraction of phenolics and phytohormones
Phenolics from wheat leaves were extracted as described bySingh et al. (2003). Briefly, 1 g leaf tissue was macerated ina pestle–mortar with 5 ml of methanol/water (1:1, v/v).Suspended macerate was ultrasonicated for 15 min at roomtemperature followed by centrifugation at 7,500×g for15 min. The clear greenish supernatant was collected andcell debris was again extracted for 4 h. The pooledsupernatant was mixed with charcoal (1 g) to removepigments and the solvent was evaporated under vacuum.Dried samples were re-dissolved in HPLC grade methanolby vortexing and stored at 4°C for analysis.
Phytohormones were extracted from 1 g leaves with80% methanol (10 ml g−1) containing butylated hydrox-ytoluene (BHT, 100 mg) and the homogenates were keptovernight at 4°C in dark. After re-extraction, the super-natant was frozen at −20°C, thawed, and centrifuged at9,000×g for 30 min at 4°C to remove impurities. Thesupernatant was used for the estimation of phytohormones.
Soils attached to plant roots (rhizosphere soil) of eachtreatment and control was dried under vacuum, and 5 gof the soil was extracted twice with methanol/water (1:1,v/v, 5 ml). The supernatants were pooled together andsolvent was evaporated to dryness to obtain rhizospheresoil extract. Roots of the freshly harvested treated andcontrol plants were uprooted and dipped in a vialcontaining water/methanol (80:20, v/v, 10 ml) afterremoving adhered soil particles from the surface. Aftergentle shaking for 30 min on shaker (30 rpm), thesolution was collected and lyophilized. The dried extractswere re-dissolved in methanol (HPLC grade) andsubjected to filtration prior to high performance liquidchromatographic (HPLC) analysis.
HPLC analysis
Extracts of wheat leaves, roots, and rhizospheric soils wereanalyzed using HPLC system (Waters, USA) equipped withbinary Waters 515 reciprocating pumps, variable photodi-ode array (PDA) detector (Waters 2996), and systemcontroller equipped with Waters® Empower™ softwarefor data integration and analysis. Reverse phase analysiswas carried out in isocratic conditions on C-18 column
Biol Fertil Soils (2011) 47:907–916 909

(250×4.6 mm i.d., 5 μm particle size) at 25±1°C at a flowrate of 1 ml/min using mobile phase methanol/0.4% aceticacid in water (76:24, v/v) and detection at 254 and 280 nmfor phenolic acids like gallic, caffeic, syringic, vanillic,ferulic, and cinnamic acids and flavonoid (quercetin).Indole acetic acid was analyzed at a flow rate of 1 ml/minof methanol/1% aqueous acetic acid (60:40, v/v). Sampleswere filtered through a 0.45-μm membrane filter prior toinjection in the sample loop (injection volume 10 μl).HPLC grade solvents and chemicals (E Merck andHiMedia, India) were used throughout the analysis.Qualitative characterization of the compounds in the samplewas done by comparing retention time (Rt) and co-injectionwhile quantitative analysis was performed by comparingpeak areas of the standard compounds obtained from Hi-Media, India.
Statistical analysis
One-way ANOVA was applied to determine the significancebetween different treatments using SPSS for Windows(version 13.0).
Results
On the basis of 16S rRNA homology, the bacterial isolatesAS 121, AS 40, AS 18, SL 9, and SL 11 were identified asBacillus pumilus, Pseudomonas mendocina, Arthrobactersp., Halomonas sp., and Nitrinicola lacisaponensis, respec-tively (Table 1).
The isolates showed different PGP traits like Psolubilization and IAA, siderophore, and ammoniaproduction. Their salt-tolerance capacity varied from2% to 22% NaCl concentration in NA media and,except Halomonas sp., all isolates showed production ofproline and choline oxidase while Arthrobacter sp.exhibited ACC deaminase activity (Table 1). Seed bacte-rization with bacterial suspension supported plant growthafter 15 and 30 days (Fig. 1). Maximum shoot and rootlength (33.9 and 13.6 cm) (Fig. 1a) and biomassaccumulation in terms of shoot and root dry weight(2.73 and 4.48 g) (Fig. 1b) was recorded in plantsinoculated with B. pumilus after 30 days. Total chlorophyllcontent was maximum in the leaves of the plants treatedwith Halomonas sp. (24.22 mg g−1 dry weight) followedby B. pumilus (23.41 mg g−1 dry weight) as compared tocontrol (18.21 mg g−1 dry weight) after 30 days. Totalprotein content was maximum in Arthrobacter sp. inocu-lated plant leaves (3.19 mg g−1 dry weight) followed by B.pumilus (2.47 mg g−1 dry weight) as compared to control(2.15 mg g−1 dry weight) after 30 days (Table 2). Total
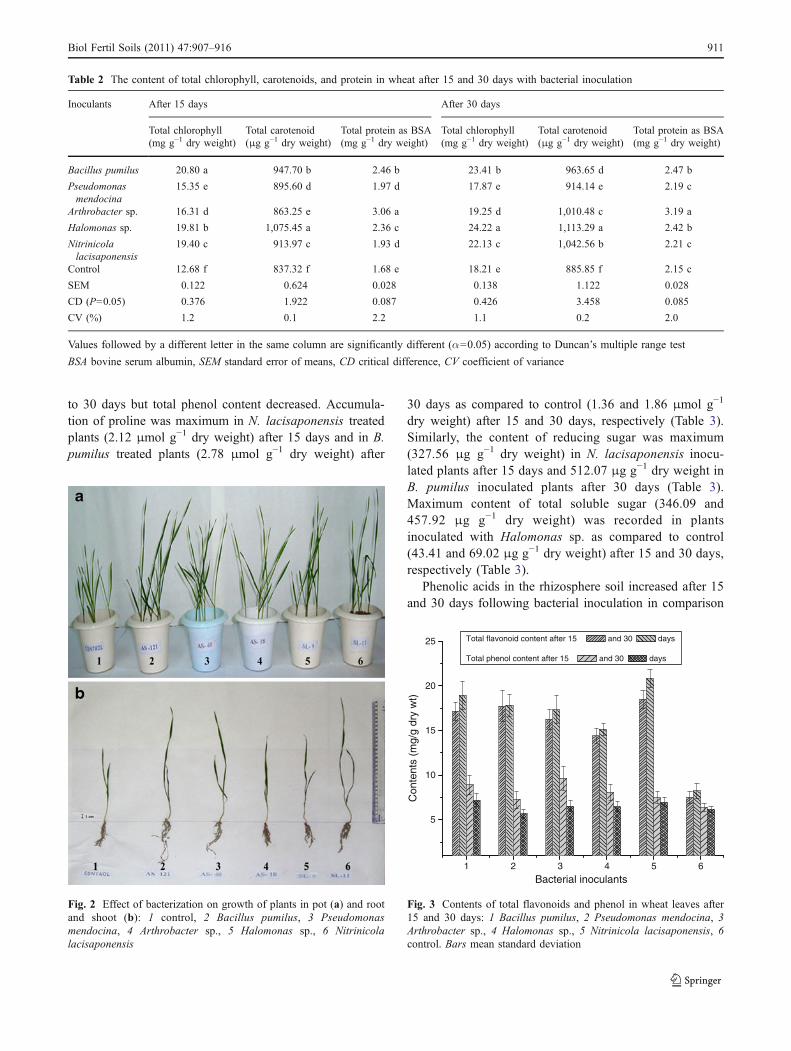
carotenoid content was maximum in plants inoculatedwith Halomonas sp. (1,075.45 and 1,113.29 μg g−1 dryweight) in comparison to control (837.32 and885.85 μg g−1 dry weight) after 15 and 30 days (Table 2).These results were supported by the significantly pro-nounced effect of seed bacterization on growth of theplants in pots (Fig. 2a, b).
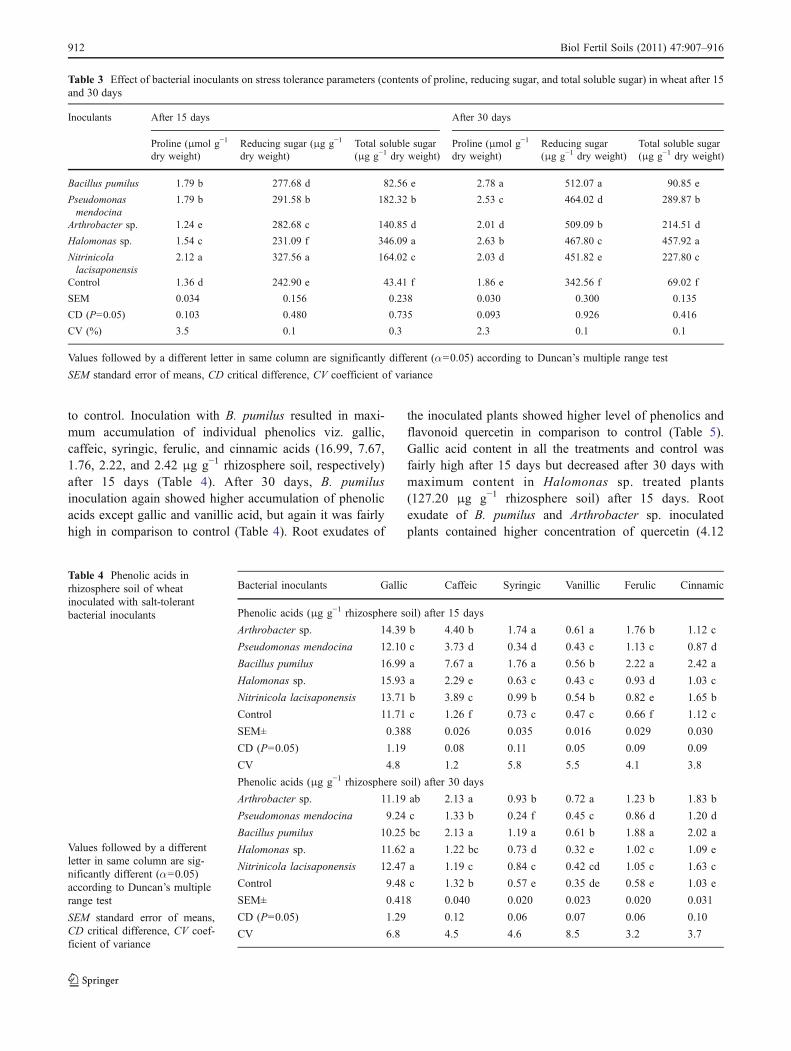
Inoculated bacteria caused differential accumulation oftotal flavonoid and total phenol contents of the leaves oftreated plants (Fig. 3). Maximum content of total flavonoidwas recorded in leaves of the plants inoculated with N.lacisaponensis (20.8 mg g−1 dry weight) after 30 dayswhile total phenol content was highest in Arthrobacter sp.inoculated plant leaves (9.78 mg g−1 dry weight) followedby B. pumilus inoculated plants (9.58 mg g−1 dry weight)after 15 days (Fig. 3). It was interesting to note that thecontent of flavonoids in all the treatments increased from 15
5
10
15
20
25
30
35
40 Root length at 15 days Root length at 30 days Shoot length at 15 days Shoot length at 30 days
Leng
th (
cm)
Bacterial inoculants
1 2 3 4 5 6
1 2 3 4 5 6
1
2
3
4
5
Root dry wt at 15 days Root dry wt at 30 days Shoot dry wt at 15 days Shoot dry wt at 30 days
Dry
Wt.
(g)
Bacterial inoculants
a
b
Fig. 1 Effect of bacterial inoculants on plant growth after 15 and30 days: a root and shoot length, b root and shoot dry weight. 1Bacillus pumilus, 2 Pseudomonas mendocina, 3 Arthrobacter sp., 4Halomonas sp., 5 Nitrinicola lacisaponensis, 6 control. All the datawere significantly different (α=0.05) according to Duncan’s multiplerange test; bars mean standard deviation
910 Biol Fertil Soils (2011) 47:907–916
to 30 days but total phenol content decreased. Accumula-tion of proline was maximum in N. lacisaponensis treatedplants (2.12 μmol g−1 dry weight) after 15 days and in B.pumilus treated plants (2.78 μmol g−1 dry weight) after
30 days as compared to control (1.36 and 1.86 μmol g−1
dry weight) after 15 and 30 days, respectively (Table 3).Similarly, the content of reducing sugar was maximum(327.56 μg g−1 dry weight) in N. lacisaponensis inocu-lated plants after 15 days and 512.07 μg g−1 dry weight inB. pumilus inoculated plants after 30 days (Table 3).Maximum content of total soluble sugar (346.09 and457.92 μg g−1 dry weight) was recorded in plantsinoculated with Halomonas sp. as compared to control(43.41 and 69.02 μg g−1 dry weight) after 15 and 30 days,respectively (Table 3).
Phenolic acids in the rhizosphere soil increased after 15and 30 days following bacterial inoculation in comparison
Fig. 2 Effect of bacterization on growth of plants in pot (a) and rootand shoot (b): 1 control, 2 Bacillus pumilus, 3 Pseudomonasmendocina, 4 Arthrobacter sp., 5 Halomonas sp., 6 Nitrinicolalacisaponensis
Table 2 The content of total chlorophyll, carotenoids, and protein in wheat after 15 and 30 days with bacterial inoculation
Inoculants After 15 days After 30 days
Total chlorophyll(mg g−1 dry weight)
Total carotenoid(μg g−1 dry weight)
Total protein as BSA(mg g−1 dry weight)
Total chlorophyll(mg g−1 dry weight)
Total carotenoid(μg g−1 dry weight)
Total protein as BSA(mg g−1 dry weight)
Bacillus pumilus 20.80 a 947.70 b 2.46 b 23.41 b 963.65 d 2.47 b
Pseudomonasmendocina
15.35 e 895.60 d 1.97 d 17.87 e 914.14 e 2.19 c
Arthrobacter sp. 16.31 d 863.25 e 3.06 a 19.25 d 1,010.48 c 3.19 a
Halomonas sp. 19.81 b 1,075.45 a 2.36 c 24.22 a 1,113.29 a 2.42 b
Nitrinicolalacisaponensis
19.40 c 913.97 c 1.93 d 22.13 c 1,042.56 b 2.21 c
Control 12.68 f 837.32 f 1.68 e 18.21 e 885.85 f 2.15 c
SEM 0.122 0.624 0.028 0.138 1.122 0.028
CD (P=0.05) 0.376 1.922 0.087 0.426 3.458 0.085
CV (%) 1.2 0.1 2.2 1.1 0.2 2.0
Values followed by a different letter in the same column are significantly different (α=0.05) according to Duncan’s multiple range test
BSA bovine serum albumin, SEM standard error of means, CD critical difference, CV coefficient of variance
1 2 3 4 5 6
5
10
15
20
25
Con
tent
s (m
g/g
dry
wt)
Bacterial inoculants
Total flavonoid content after 15 and 30 days
Total phenol content after 15 and 30 days
Fig. 3 Contents of total flavonoids and phenol in wheat leaves after15 and 30 days: 1 Bacillus pumilus, 2 Pseudomonas mendocina, 3Arthrobacter sp., 4 Halomonas sp., 5 Nitrinicola lacisaponensis, 6control. Bars mean standard deviation
Biol Fertil Soils (2011) 47:907–916 911
to control. Inoculation with B. pumilus resulted in maxi-mum accumulation of individual phenolics viz. gallic,caffeic, syringic, ferulic, and cinnamic acids (16.99, 7.67,1.76, 2.22, and 2.42 μg g−1 rhizosphere soil, respectively)after 15 days (Table 4). After 30 days, B. pumilusinoculation again showed higher accumulation of phenolicacids except gallic and vanillic acid, but again it was fairlyhigh in comparison to control (Table 4). Root exudates of
the inoculated plants showed higher level of phenolics andflavonoid quercetin in comparison to control (Table 5).Gallic acid content in all the treatments and control wasfairly high after 15 days but decreased after 30 days withmaximum content in Halomonas sp. treated plants(127.20 μg g−1 rhizosphere soil) after 15 days. Rootexudate of B. pumilus and Arthrobacter sp. inoculatedplants contained higher concentration of quercetin (4.12
Table 3 Effect of bacterial inoculants on stress tolerance parameters (contents of proline, reducing sugar, and total soluble sugar) in wheat after 15and 30 days
Inoculants After 15 days After 30 days
Proline (μmol g−1
dry weight)Reducing sugar (μg g−1
dry weight)Total soluble sugar(μg g−1 dry weight)
Proline (μmol g−1
dry weight)Reducing sugar(μg g−1 dry weight)
Total soluble sugar(μg g−1 dry weight)
Bacillus pumilus 1.79 b 277.68 d 82.56 e 2.78 a 512.07 a 90.85 e
Pseudomonasmendocina
1.79 b 291.58 b 182.32 b 2.53 c 464.02 d 289.87 b
Arthrobacter sp. 1.24 e 282.68 c 140.85 d 2.01 d 509.09 b 214.51 d
Halomonas sp. 1.54 c 231.09 f 346.09 a 2.63 b 467.80 c 457.92 a
Nitrinicolalacisaponensis
2.12 a 327.56 a 164.02 c 2.03 d 451.82 e 227.80 c
Control 1.36 d 242.90 e 43.41 f 1.86 e 342.56 f 69.02 f
SEM 0.034 0.156 0.238 0.030 0.300 0.135
CD (P=0.05) 0.103 0.480 0.735 0.093 0.926 0.416
CV (%) 3.5 0.1 0.3 2.3 0.1 0.1
Values followed by a different letter in same column are significantly different (α=0.05) according to Duncan’s multiple range test
SEM standard error of means, CD critical difference, CV coefficient of variance
Bacterial inoculants Gallic Caffeic Syringic Vanillic Ferulic Cinnamic
Phenolic acids (μg g−1 rhizosphere soil) after 15 days
Arthrobacter sp. 14.39 b 4.40 b 1.74 a 0.61 a 1.76 b 1.12 c
Pseudomonas mendocina 12.10 c 3.73 d 0.34 d 0.43 c 1.13 c 0.87 d
Bacillus pumilus 16.99 a 7.67 a 1.76 a 0.56 b 2.22 a 2.42 a
Halomonas sp. 15.93 a 2.29 e 0.63 c 0.43 c 0.93 d 1.03 c
Nitrinicola lacisaponensis 13.71 b 3.89 c 0.99 b 0.54 b 0.82 e 1.65 b
Control 11.71 c 1.26 f 0.73 c 0.47 c 0.66 f 1.12 c
SEM± 0.388 0.026 0.035 0.016 0.029 0.030
CD (P=0.05) 1.19 0.08 0.11 0.05 0.09 0.09
CV 4.8 1.2 5.8 5.5 4.1 3.8
Phenolic acids (μg g−1 rhizosphere soil) after 30 days
Arthrobacter sp. 11.19 ab 2.13 a 0.93 b 0.72 a 1.23 b 1.83 b
Pseudomonas mendocina 9.24 c 1.33 b 0.24 f 0.45 c 0.86 d 1.20 d
Bacillus pumilus 10.25 bc 2.13 a 1.19 a 0.61 b 1.88 a 2.02 a
Halomonas sp. 11.62 a 1.22 bc 0.73 d 0.32 e 1.02 c 1.09 e
Nitrinicola lacisaponensis 12.47 a 1.19 c 0.84 c 0.42 cd 1.05 c 1.63 c
Control 9.48 c 1.32 b 0.57 e 0.35 de 0.58 e 1.03 e
SEM± 0.418 0.040 0.020 0.023 0.020 0.031
CD (P=0.05) 1.29 0.12 0.06 0.07 0.06 0.10
CV 6.8 4.5 4.6 8.5 3.2 3.7
Table 4 Phenolic acids inrhizosphere soil of wheatinoculated with salt-tolerantbacterial inoculants
Values followed by a differentletter in same column are sig-nificantly different (α=0.05)according to Duncan’s multiplerange test
SEM standard error of means,CD critical difference, CV coef-ficient of variance
912 Biol Fertil Soils (2011) 47:907–916
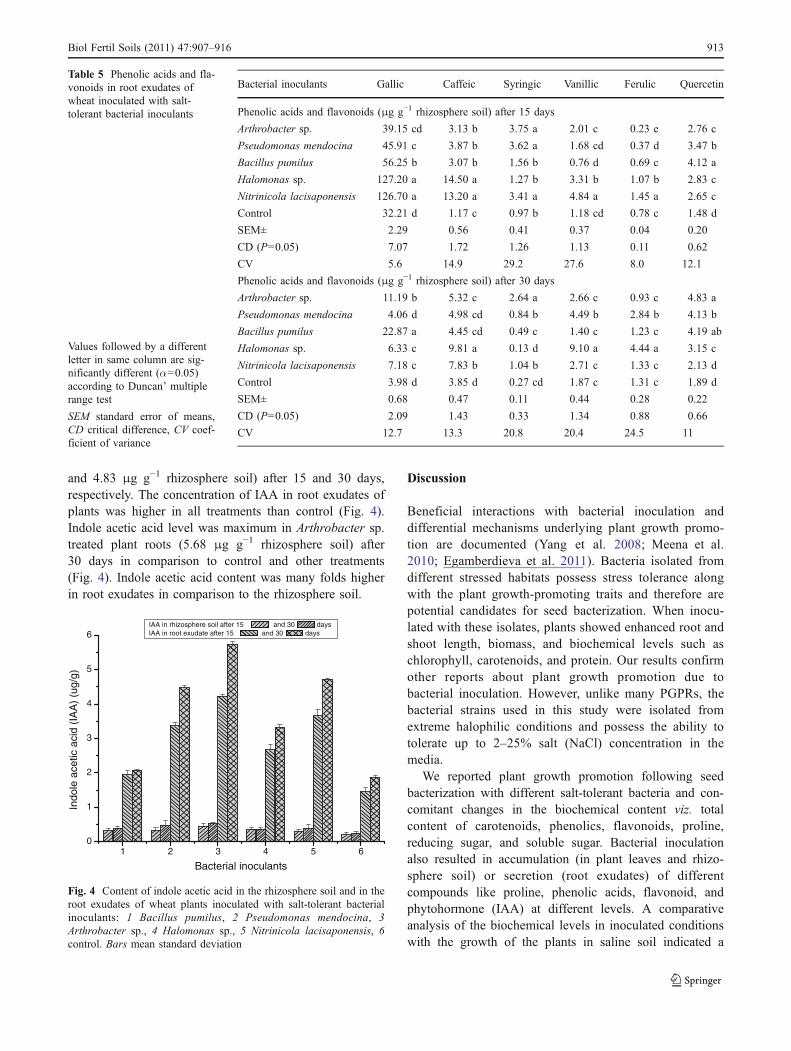
and 4.83 μg g−1 rhizosphere soil) after 15 and 30 days,respectively. The concentration of IAA in root exudates ofplants was higher in all treatments than control (Fig. 4).Indole acetic acid level was maximum in Arthrobacter sp.treated plant roots (5.68 μg g−1 rhizosphere soil) after30 days in comparison to control and other treatments(Fig. 4). Indole acetic acid content was many folds higherin root exudates in comparison to the rhizosphere soil.
Discussion
Beneficial interactions with bacterial inoculation anddifferential mechanisms underlying plant growth promo-tion are documented (Yang et al. 2008; Meena et al.2010; Egamberdieva et al. 2011). Bacteria isolated fromdifferent stressed habitats possess stress tolerance alongwith the plant growth-promoting traits and therefore arepotential candidates for seed bacterization. When inocu-lated with these isolates, plants showed enhanced root andshoot length, biomass, and biochemical levels such aschlorophyll, carotenoids, and protein. Our results confirmother reports about plant growth promotion due tobacterial inoculation. However, unlike many PGPRs, thebacterial strains used in this study were isolated fromextreme halophilic conditions and possess the ability totolerate up to 2–25% salt (NaCl) concentration in themedia.
We reported plant growth promotion following seedbacterization with different salt-tolerant bacteria and con-comitant changes in the biochemical content viz. totalcontent of carotenoids, phenolics, flavonoids, proline,reducing sugar, and soluble sugar. Bacterial inoculationalso resulted in accumulation (in plant leaves and rhizo-sphere soil) or secretion (root exudates) of differentcompounds like proline, phenolic acids, flavonoid, andphytohormone (IAA) at different levels. A comparativeanalysis of the biochemical levels in inoculated conditionswith the growth of the plants in saline soil indicated a
Bacterial inoculants Gallic Caffeic Syringic Vanillic Ferulic Quercetin
Phenolic acids and flavonoids (μg g−1 rhizosphere soil) after 15 days
Arthrobacter sp. 39.15 cd 3.13 b 3.75 a 2.01 c 0.23 e 2.76 c
Pseudomonas mendocina 45.91 c 3.87 b 3.62 a 1.68 cd 0.37 d 3.47 b
Bacillus pumilus 56.25 b 3.07 b 1.56 b 0.76 d 0.69 c 4.12 a
Halomonas sp. 127.20 a 14.50 a 1.27 b 3.31 b 1.07 b 2.83 c
Nitrinicola lacisaponensis 126.70 a 13.20 a 3.41 a 4.84 a 1.45 a 2.65 c
Control 32.21 d 1.17 c 0.97 b 1.18 cd 0.78 c 1.48 d
SEM± 2.29 0.56 0.41 0.37 0.04 0.20
CD (P=0.05) 7.07 1.72 1.26 1.13 0.11 0.62
CV 5.6 14.9 29.2 27.6 8.0 12.1
Phenolic acids and flavonoids (μg g−1 rhizosphere soil) after 30 days
Arthrobacter sp. 11.19 b 5.32 c 2.64 a 2.66 c 0.93 c 4.83 a
Pseudomonas mendocina 4.06 d 4.98 cd 0.84 b 4.49 b 2.84 b 4.13 b
Bacillus pumilus 22.87 a 4.45 cd 0.49 c 1.40 c 1.23 c 4.19 ab
Halomonas sp. 6.33 c 9.81 a 0.13 d 9.10 a 4.44 a 3.15 c
Nitrinicola lacisaponensis 7.18 c 7.83 b 1.04 b 2.71 c 1.33 c 2.13 d
Control 3.98 d 3.85 d 0.27 cd 1.87 c 1.31 c 1.89 d
SEM± 0.68 0.47 0.11 0.44 0.28 0.22
CD (P=0.05) 2.09 1.43 0.33 1.34 0.88 0.66
CV 12.7 13.3 20.8 20.4 24.5 11
Table 5 Phenolic acids and fla-vonoids in root exudates ofwheat inoculated with salt-tolerant bacterial inoculants
Values followed by a differentletter in same column are sig-nificantly different (α=0.05)according to Duncan’ multiplerange test
SEM standard error of means,CD critical difference, CV coef-ficient of variance
1 2 3 4 5 60
1
2
3
4
5
6
Indo
le a
cetic
aci
d (I
AA
) (u
g/g)
Bacterial inoculants
IAA in rhizosphere soil after 15 and 30 days IAA in root exudate after 15 and 30 days
Fig. 4 Content of indole acetic acid in the rhizosphere soil and in theroot exudates of wheat plants inoculated with salt-tolerant bacterialinoculants: 1 Bacillus pumilus, 2 Pseudomonas mendocina, 3Arthrobacter sp., 4 Halomonas sp., 5 Nitrinicola lacisaponensis, 6control. Bars mean standard deviation
Biol Fertil Soils (2011) 47:907–916 913

positive correlation leading to plant growth under stressconditions.
Almost all biochemical changes are related to plantgrowth promotion as well as stress tolerance and, therefore,there was an integrated and cumulative effect of bacterialinoculation. Colonization of roots by PGPRs promotesplant growth and increases accumulation of secondarymetabolites systemically to confer “induced systemictolerance” (IST) as it has been suggested by Yang et al.(2008). Although it has been documented that PGPRs canact as elicitors of tolerance to abiotic stresses (drought, saltor nutrient deficiency or excess) (Yang et al. 2008; Dey etal. 2009), further research is needed since the mechanismsare poorly understood. Our results support the hypothesisthat bacteria induce different physical and biochemicalchanges in plants with enhanced plant growth understressed soil conditions. Probably specific mechanismsare responsible for plant performance under stressedconditions.
Increased levels of carotenoids in plants are directlyrelated to the increased tolerance to light-dependentoxidative damage (Cogdel and Frank 1987; Davison et al.2002) and high chlorophyll and carotenoids contents areclosely associated with antioxidant enzyme systems of theplants (Sairam et al. 1997). During the course of stress,compatible solutes such as proline, polyamines, andcarbohydrates are effective biomarkers, and their accumu-lation in plants represents an indication of stress tolerancemechanism (Tripathi et al. 1998; Kerepesi et al. 1998;Kerepesi and Galiba 2000; Colmer et al. 2005; Barriuso etal. 2008). Evidences for the enhanced levels of carotenoids,chlorophyll, and different compatible solutes in the plantstreated with bacterial inoculants in the present study agreewith the earlier findings and indicate that these biomarkerseffectively play an important role.
An interesting modification caused by bacterial inoc-ulation in the plant is the changes in the composition ofroot exudates, particularly that of small molecules (Baiset al. 2004; Badri and Vivanco 2009). Plant growth anddevelopment requires integration and coordination ofspatial and temporal organization of cellular processesand exchange of signaling molecules, and it is affected byboth biotic and abiotic factors (Ortíz-Castro et al. 2009).Plants exudate a wide array of organic and inorganiccompounds including carbohydrates, amino acids, poly-amines, vitamins, and minerals, which can be nutrients,phytohormones, and signals for microbial communities inthe vicinity of roots (Nelson 2004; Lugtenberg andKamilova 2009; Naz et al. 2009; Sutee et al. 2009).Alternatively, microbes inhabiting the rhizosphere releasephytohormones, small molecules, or volatile compoundswhich can directly or indirectly modulate plant immunityor regulate plant growth and morphogenesis (Costacurta
and Vanderleyden 1995; Castro-Sowinski et al. 2007).There are many evidences of the role of phenolics andflavonoids in plant–microbe interaction and root coloni-zation (O'Callaghan et al. 2000; Kothandaraman et al.2003; Weir et al. 2004; Brencic and Winans 2005;Bhattacharya et al. 2010). It is evident from our work thatlevels of phenolic compounds, the flavonoid quercetin,and phytohormone (IAA) increased in the rhizospherefollowing bacterial inoculation. We, therefore, concludethat bacteria-mediated responses in plant leaves andincreased levels of phenolic acids, quercetin, and IAA inthe root exudates and rhizosphere played a cumulativesynergistic role that enhanced growth of wheat. Anunbiased analysis of whole metabolome in the rhizosphereand at the rhizoplane along with that of the leaf tissuesmay lead to the discovery of certain signal moleculesresponsible for triggering growth promotion or stresstolerance mechanisms in plant.
Acknowledgment Authors are thankful to the Indian Council ofAgricultural Research (ICAR), New Delhi, India for financialassistance under the Network project “Application of Microorganismsin Agriculture and Allied Sectors” (AMAAS).
References
Amirjani MR (2010) Effect of salinity stress on growth, mineralcomposition, proline content, antioxidant enzymes of soybean.Am J Plant Physiol 5:350–360
Badri DV, Vivanco JM (2009) Regulation and function of rootexudates. Plant Cell Environ 32:666–681
Bais HP, Park SW, Weir TL, Callaway RM, Vivanco JM (2004) Howplants communicate using the underground information super-highway? Trends Plant Sci 9:26–32
Bano A, Fatima M (2009) Salt tolerance in Zea mays L. followinginoculation with Rhizobium and Pseudomonas. Biol Fert Soils45:405–413
Barriuso J, Ramos Solano B, Gutiérrez Mañero FJ (2008) Protectionagainst pathogen and salt stress by four plant growth-promotingrhizobacteria isolated from Pinus sp. on Arabidopsis thaliana.Phytopathol 98:666–672
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of freeproline for water stress studies. Plant Soil 39:205–207
Bhattacharya A, Sood P, Citovsky V (2010) The roles of plantphenolics in defence and communication during Agrobacteriumand Rhizobium infection. Mol Plant Pathol 11:705–719
Brencic A, Winans SC (2005) Detection of and response to signalsinvolved in host–microbe interactions by plant-associated bacte-ria. Microbiol Mol Biol Rev 69:155–194
Brick JM, Bostock RM, Silverstone SE (1991) Rapid in-situ assayfor indole acetic acid production by bacteria immobilized onnitrocellulose membrane. Appl Environ Microbiol 57:535–538
Castro-Sowinski S, Herschkovitz Y, Okon Y, Jurkevitch E (2007)Effects of inoculation with plant growth-promoting rhizobacteriaon resident rhizosphere microorganisms. FEMS Microbiol Lett276:1–11
Cogdel RJ, Frank HA (1987) How carotenoids function in photosyn-thetic bacteria. Biochim Biophys Acta 895:63–79
914 Biol Fertil Soils (2011) 47:907–916

Colmer TD, Munns R, Flowers TJ (2005) Improving salt tolerance ofwheat and barley: further prospects. Aust J Exp Agr 45:1425–1443
Costacurta A, Vanderleyden J (1995) Synthesis of phytohormones byplant-associated bacteria. Crit Rev Microbiol 21:1–18
Davison PA, Hunter CN, Horton P (2002) Overexpression of β-carotene hydroxylase enhances stress tolerance in Arabidopsis.Nature 418:203–206
Dey R, Pal KK, Bhatt DM, Chauhan SM (2004) Growth promotionand yield enhancement of peanut (Arachis hypogaea L) byapplication of plant growth promoting rhizobacteria. MicrobiolRes 159:371–394
Dey C, Weinand T, Asch F (2009) Plant–rhizobacteria interactionsalleviate abiotic stress conditions. Plant Cell Environ 32:1682–1694
Edwards U, Rogall TH, Blocker H, Emde M, Bottger EC (1989)Isolation and direct complete nucleotide determination of entiregenes. Characterization of a gene coding for 16S ribosomalRNA. Nucleic Acids Res 17:7843–7853
Egamberdieva D, Kucharova Z (2009) Selection for root colonisingbacteria stimulating wheat growth in saline soils. Biol Fert Soils45:563–571
Egamberdieva D, Kucharova Z, Davranov K, Berg G, Makarova N,Azarova T, Chetobar V, Tikhonovich I, Kamilova F, Validov SZ,Lugtenberg B (2011) Bacteria able to control foot and root rotand to promote growth of cucumber in salinated soils. Biol FertSoils 47:197–205
Ferjani A, Mustardy L, Sulpice R, Marin K, Suzuki I, HagemannM, Murata N (2003) Glucosylglycerol, a compatible solute,sustains cell division under salt stress. Plant Physiol131:1628–1637
Gaur AC (1990) Physiological functions of phosphate solubilizingmicro-organisms. In: Gaur AC (ed) Phosphate solubilizing micro-organisms as biofertilizers. Omega Scientific, New Delhi, pp 16–72
Graham HN (1992) Green tea composition, consumption andpolyphenol chemistry. Prev Med 21:334–350
Ikuta SK, Matuura S, Imamura H, Misaki HY (1977) Oxidativepathway of choline to betaine in the soluble fraction preparedfrom Arthrobacter globiformis. J Biochem 82:157–163
Jones MG (2009) Using resources from the model plant Arabidopsisthaliana to understand effects of abiotic stress. Salinity WaterStress 44:129–132
Kerepesi I, Galiba G (2000) Osmotic and salt stress-induced alterationin soluble carbohydrate content in wheat seedlings. Crop Sci40:482–487
Kerepesi I, Galiba G, Banyai E (1998) Osmotic and salt stressesinduced differential alteration in water-soluble carbohydratecontent in wheat seedlings. J Agr Food Chem 46:5347–5354
Kim D, Jeong SW, Lee CY (2003) Antioxidant capacity of phenolicphytochemicals from various cultivars of plums. Food Chem81:321–326
Kothandaraman N, Chanbasha B, Vladimir BB, Swarup S (2003)Enhancement of plant–microbe interactions using a rhizo-sphere metabolomics-driven approach and its application inthe removal of polychlorinated biphenyls. Plant Physiol132:146–153
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Proteinmeasurement with folin phenol reagent. J Biol Chem 193:265–275
Lugtenberg B, Kamilova F (2009) Plant-growth-promoting rhizobac-teria. Annu Rev Microbiol 63:541–556
Maness N (2010) Extraction and analysis of soluble carbohydrates.Methods Mol Biol 639:341–370
Meena KK, Mesapogu S, Kumar M, Yandigeri MS, Singh G, SaxenaAK (2010) Co-inoculation of the endophytic fungus Piriformo-
spora indica with the P-solubilizing bacteria Pseudomonasstriata affects population dynamics and plant growth in chickpea.Biol Fert Soils 46:169–174
Nadeem SM, Zahir ZA, Naveed M, Arshad M (2007) Preliminaryinvestigations on inducing salt tolerance in maize throughinoculation with rhizobacteria containing ACC deaminase activ-ity. Can J Microbiol 53:1141–1149
Naz I, Bano A, Ul-Hassan T (2009) Isolation of phytohormonesproducing plant growth promoting rhizobacteria from weedsgrowing in Khewra salt range, Pakistan and their implication inproviding salt tolerance to Glycine max. L. Afr J Biotech 8:5762–5766
Neelam T, Meenu S (2010) Salinity-resistant plant growth promotingrhizobacteria ameliorates sodium chloride stress on tomatoplants. J Plant Interact 5:51–58
Nelson EB (2004) Microbial dynamics and interactions in thespermosphere. Annu Rev Phytopathol 42:271–309
O'Callaghan KJ, Stone PJ, Hu X, Griffiths DW, Davey MR, CockingEC (2000) Effects of glucosinolates and flavonoids on coloniza-tion of the roots of Brassica napus by Azorhizobium caulinodansORS571. Appl Environ Microbiol 66:2185–2191
Ortíz-Castro R, Contreras-Cornejo HA, Macías-Rodríguez L, López-Bucio J (2009) The role of microbial signals in plant growth anddevelopment. Plant Signal Behav 4:701–712
Penrose DM, Glick BR (2003) Methods for isolating and character-izing ACC deaminase containing plant growth promotingrhizobacteria. Physiol Plant 118:10–15
Pospiech A, Neumann B (1995) A versatile quick-prep of genomicDNA from gram-positive bacteria. Trends Genet 11:217–218
Rabie GG, Almandini AM (2005) Role of bioinoculants in develop-ment of salt-tolerance of Vicia faba plants under salinity stress.Afr J Biotech 4:210–223
Sadasivam S, Manikam A (1992) Biochemical methods. Wiley Port ofUniversity Grants Commission, New Delhi
Sairam RK, Shukla DS, Saxena DC (1997) Stress induced injury andantioxidant enzymes in relation to drought tolerance in wheatgenotypes. Biol Plant 40:357–364
Schwyn B, Neilands JB (1987) Universal chemical assay for thedetection and determination of siderophore. Anal Biochem160:47–56
Scurtz AV, Christie BR, Nowak J (2000) Bacterial endophytes:potential role in developing sustainable systems of crop produc-tion. Crit Rev Plant Sci 19:1–30
Siddikee MA, Chauhan PS, Anandham R, Han GH, Sa T (2010)Isolation, characterization and use for plant growth promotionunder salt stress, of ACC deaminase-producing halotolerantbacteria derived from coastal soil. J Microbiol Biotechnol20:1577–1584
Singh UP, Sarma BK, Singh DP (2003) Effect of plant growth-promoting rhizobacteria and culture filtrate of Sclerotium rolfsiion phenolic and salicylic acid contents in chickpea (Cicerarietinum). Curr Microbiol 44:131–140
Smalla K, Wieland G, Buchner A, Zock A, Parzy J, Kaiser S, RoskotN, Heuer H, Smalla G (2001) Bulk and rhizosphere soil bacterialcommunities studied by denaturing gradient gel electrophoresis:plant-dependent enrichment and seasonal shift revealed. ApplEnviron Microbiol 67:4742–4751
Sturz AV, Nowak J (2000) Endophytic communities of rhizobacteriaand the strategies required to create yield enhancing associationswith crops. Appl Soil Ecol 15:183–190
Sutee C, Cha-um S, Sompornpailin K (2009) Differential accumu-lations of proline and flavonoids in indica rice varieties againstsalinity. Pak J Bot 41:2497–2506
Tripathi AK, Mishra BM, Tripathi P (1998) Salinity stress responses inthe plant growth promoting rhizobacteria, Azospirillum spp. JBiosci 23:463–471
Biol Fertil Soils (2011) 47:907–916 915

Umbrient WW, Burris RH, Stauffer J, Cohen P, Johnson WJ, LeepxgiGA, Patter VR, Schneider WCI (1959) Monometric Techniques,a manual describing methods applicable to the study of tissuemetabolism. Burgess, Minneapolis
Weir TL, Park SW, Vivanco JM (2004) Biochemical and physiologicalmechanisms mediated by allelochemicals. Curr Opin Plant Biol7:472–479
Whips JM (2001) Microbial interactions and biocontrol in therhizosphere. J Exp Bot 52:487–511
Wu CH, Bernard SM, Anderson GL, Chen W (2009) Developingmicrobe–plant interactions for applications in plant-growth
promotion and disease control, production of useful compounds,remediation and carbon sequestration. Microb Biotechnol 2:428–440
Yang J, Kloepper JW, Ryu C-M (2008) Rhizosphere bacteria helpplants tolerate abiotic stress. Trends Plant Sci 14:1–4
Yang CH, Chai Q, Huang GB (2010) Root distribution and yieldresponses of wheat/maize intercropping to alternate irrigation inthe arid areas of northwest China. Plant Soil Environ 56:253–262
Zahir ZA, Arshad M, Frankenberger WT (2004) Plant growthpromoting rhizobacteria: applications and perspectives in agri-culture. Adv Agron 81:97–168
916 Biol Fertil Soils (2011) 47:907–916




















